-
Bezugnahme auf verwandte
Anmeldungen
-
Die
vorliegende Erfindung nimmt die Priorität der US-Provisional Patentanmeldung
Nr. 60/112,723, eingereicht am 18. Dezember 1998 und der US-Provisional
Patentanmeldung mit dem Titel „Production
of Cyclic Peptides and Proteins In Vivo", eingereicht am 7. Oktober 1999 in
Anspruch, die beide durch diese Bezugnahme hierin mitaufgenommen
sind.
-
Bemerkungen
bezüglich öffentlich
geförderter
Forschung
-
Diese
Erfindung wurde mit Unterstützung
der Regierung mit der Beihilfe GM13306 und GM19891 vorgenommen,
verliehen von den National Institutes of Health. Die Regierung hat
bestimmte Rechte an der Erfindung.
-
Gebiet der
Erfindung
-
Die
Erfindung betrifft das Gebiet der Biochemie. Die Erfindung betrifft
insbesondere zyklische Peptide, Verfahren zur Herstellung von zyklischen
Peptiden und Verfahren zum Screenen von zyklischen Peptiden bezüglich spezieller
Eigenschaften.
-
Hintergrund der Erfindung
-
Kleine
lineare Peptide sind zur Untersuchung verschiedener physiologischer
Phänomene
von Nutzen, weil sie einen breiten Bereich biologischer Aktivitäten zeigen
und in beinahe unbegrenzt variablen Sequenzen unter Verwendung herkömmlicher
Techniken in einer Festphasensynthese und kombinatorischen Chemie
synthetisiert werden können.
Diese Qualitäten
machen kleine lineare Peptide ebenfalls zum Identifizieren und Entwickeln
neuer Arzneistoffe nützlich.
Beispielsweise können
große
Bibliotheken bzw. Banken unzähliger
unterschiedlicher kleiner linearer Peptide synthetisch hergestellt
und danach bezüglich
spezieller Eigenschaften in verschiedenen biologischen Assays gescreent
werden. Siehe beispielswiese Scott, J. K. und G. P. Smith, Science
249:386, 1990; Devlin, J. J., et al., Science 24:404, 1990; Furka,
A. et al., Int. J. Pept. Protein Res. 37:487, 1991; Lam, K. S.,
et al., Nature 354:82, 1991. Solche Peptide innerhalb der Bank,
die die spezielle Eigenschaft zeigen, können dann als Kandidaten für weitere
Untersuchungen isoliert werden. Eine Mikrosequenzierung oder andere
chemische Analysen können
dann dazu verwendet werden, ausgewählte Peptide beispielsweise durch
Aminosäuresequenz
zu charakterisieren. Trotz dieser Vorteile wurde nur eine Hand voll
kleiner linearer Peptide zu weithin verwendeten pharmazeutischen
Arzneistoffen entwickelt. Ein Grund hierfür besteht darin, dass kleine
lineare Peptide üblicherweise
zu rasch aus dem Körper
entfernt werden, um einen therapeutischen Nutzen zu besitzen.
-
Der
Ringschluss oder die Zyklisierung können die Geschwindigkeit reduzieren,
in der Peptide in vivo abgebaut werden und verbessern deswegen dramatisch
ihre pharmakokinetischen Eigenschaften. Der große Teil der zyklischen Peptide,
die einen therapeutischen Wert besitzen, wurden nach Isolierung
aus natürlichen Quellen
identifiziert (beispielsweise Caclitonine, Oxytocin und Vasopressin).
Leider ist der Pool natürlich
existierender zyklischer Peptide, der bezüglich einer speziellen biologischen
Eigenschaft gescreent werden kann, inhärent beschränkt. Und darüber hinaus
machen die beschwerlichen Schritte, die zum Isolieren und Aufreinigen
zyklischer Peptide aus natürlichen
Quellen erforderlich sind, solche Screenings kostenintensiv und
unpraktisch. Somit würden
Syntheseverfahren zur Erzeugung großer Anzahlen unterschiedlicher
Peptide von unbegrenzt variablen Aminosäuresequenzen das Identifizieren
spezieller zyklischer Peptide als Kandidaten für neue Arzneistoffe in großem Maße erleichtern.
-
Verschiedene
Verfahren zum Erzeugen zyklischer Peptide wurden beschrieben. Beispielsweise
wurden chemische Reaktionsvorschriften entwickelt, wie beispielsweise
solche, die in den US-Patenten Nr. 4,033,940 und 4,102,877 beschrieben
wurden, um zirkularisierte Peptide zu erzeugen. In anderen Techniken werden
biologische und chemische Verfahren kombiniert, um zyklische Peptide
zu erzeugen. Diese letzteren Verfahren schließen zunächst das Exprimieren linearer
Präkursoren
zyklischer Peptide in Zellen (beispielsweise Bakterien) ein, um
lineare Vorläufer
bzw. Präkursoren
zyklischer Peptide zu erzeugen und danach ein exogenes Mittel zuzusetzen,
wie beispielsweise eine Protease oder ein nukleophiles Reagenz,
um diese linearen Präkursoren
chemisch zu zyklischen Peptiden umzuwandeln. Siehe beispielsweise
Camerero, J. A., und Muir, T. W., J. Am. Chem. Societey. 121:5597
(1999); Wu, H. et al, Proc. Natl. Acad. Sci. USA, 95:9226 (1998).
-
Wenn
sie einmal hergestellt sind, können
solche zyklischen Peptide bezüglich
ihrer pharmakologischen Aktivität
gescreent werden. Beispielsweise kann eine Bibliothek hergestellt
werden, die eine große
Anzahl unterschiedlicher zyklischer Peptide enthält und kann danach bezüglich spezieller
Eigenschaften gescreent werden, wie beispielsweise der Fähigkeit,
an einen spezifischen Target bzw. Ziel-Liganden zu binden. Die Bibliothek
wird mit dem Ziel-Liganden vermischt und solche Elemente der Bibliothek,
die an den Ziel-Liganden
binden, können
isoliert und durch Aminosäuresequenzierung
identifiziert werden. In ähnlicher
Weise können
Bibliotheken für
zyklische Peptide zu Assays bezüglich
einer spezifischen biologischen Aktivität zugesetzt werden. Solche
zyklischen Peptide, die die biologische Aktivität modulieren, können dann
isoliert und durch Sequenzieren identifiziert werden.
-
Leider
können
diese Screeningassays, weil der Schritt des Identifizierens der
aktiven Peptide schwierig sein kann, sich als mühsam und zeitaufwendig erweisen.
Beispielsweise erfordern Screeningassays üblicherweise einen Umkehr-Kartierungsschritt,
weil die tatsächliche
Menge an zyklischem Peptid, die an einen Target-Liganden bindet
oder eine biologische Aktivität
moduliert, üblicherweise
so gering ist, dass sie nicht direkt sequenziert werden kann. Um
dieses Problem zu vermeiden, kann eine Karte, die den physischen
Ort der verschiedenen zyklischen Peptide, die die Bibliothek ausmachen,
anzeigt, hergestellt werden. Teilmengen an zyklischen Peptiden aus
den unterschiedlichen Orten werden dann auf entsprechende Orte innerhalb
des Screeningassays übertragen;
und solche Areale im Assay, die die gescreente Aktivität zeigen
(beispielsweise eine Bindung oder Modulierung einer biologischen
Aktivität)
werden dann zu ihrer entsprechenden Lokalisierung in der Bibliothek
bzw. Bank zurückdatiert.
Die zyklischen Peptide in diesem Areal der Bibliothek können dann
isoliert und sequenziert werden. Schwierigkeiten, die sich aus dem
Bedarf einer räumlichen
Auftrennung und den Beschränkungen
ergeben, die durch die Probenhandhabung auferlegt werden, begrenzen
die Anzahl der Kandidatenpeptide, die in einer gegebenen Zeitspanne
gescreent werden können.
-
Die
Zahl der Peptide, die in einem Assay gescreent werden kann, kann
dramatisch erhöht
werden, indem Zellen verwendet werden, die die Peptide exprimieren.
Beispielsweise können
Bakterien, die zum Exprimieren einer Bibliothek linearer Peptide
entwickelt wurden, einem Screeningassay zugesetzt werden, und solche
Bakterien, die die Gescreenten als charakteristisch exprimieren
können
direkt aus dem Assay aufgesammelt werden. Die aufgesammelten Bakterien
können
dann in großen
Zahlen derart reproduziert werden, dass die ausgewählten linearen
Peptide in großen
Mengen hergestellt werden können,
um ihre Identifizierung (beispielsweise durch Sequenzierung) und
Produktion zu erleichtern. Die Herstellung und Screening kleiner
linearer Peptidbibliotheken in vivo hat jedoch sich als etwas schwierig
erwiesen, weil kleine lineare Peptide rasch durch normale zelluläre Stoffwechselprozesse
abgebaut werden. Die Zyklisierung der Peptide kann dieses Problem
umgehen, indem es die Peptide innerhalb einer Zelle stabil macht.
-
Trotz
dem war diese intrazelluläre
Produktion großer
Bibliotheken zyklischer Peptide bis jetzt nicht möglich, weil
im allgemeinen leicht durchzuführende
Verfahren zum Zyklisieren von Peptiden in vivo nicht verfügbar waren.
Beispielsweise verwendet ein bekanntes Verfahren zur Erzeugung zyklischer
Peptide in vivo nicht-ribosomale Peptidsynthetase (NRPS)-Komplexe
(Cane et al, Science 282:63, 1998). Derartige NRSP Komplexe jedoch
sind weder leicht handzuhaben noch im allgemeinen zur Herstellung
von mehr als einem einzigen zyklischen Peptid zu einem Zeitpunkt
von Nutzen. Überdies
ist anders als bei der ribosomalen Peptidsynthese, bei der die lineare
Sequenz von Monomeren (Aminosäuren)
durch die lineare Basensequenz im Nukleinsäuremolekül, das dieses codiert, vorgegeben
ist, die lineare Sequenz der Monomere in einem Peptid, das durch
das NRPS-Verfahren hergestellt wurde, durch die Untereinheitsorganisation
des NRPS-Komplexes vorgegeben. Die Veränderung der Sequenz eines zyklischen
Peptides, das durch NRPS hergestellt wurde, umfasst das Klonieren
der Untereinheit(en), die die erwünschten Monomere einbeziehen
und die Einführung der
Untereinheit(en) in Wirtszellen, die bereits alle notwendigen Untereinheiten
beherbergen. Die Herstellung einer Bibliothek unter Verwendung dieser
Techniken würde
die Einbringung von Kombinationen (sowohl in der Zusammensetzung
als auch in der Reihenfolge) von NRPS Untereinheiten in Wirtszellen
und das Entwickeln eines Verfahrens zur Sicherstellung erfordern,
dass die Untereinheiten sich in korrekten supramolekularen Strukturen
zusammenfügen.
-
HOLFORD
M. ET AL.: ,Adding ,splice' to
protein engineering' STRUCTURE,
Band 6, l5 August 1998 (1998-08-15), Seiten 951-956, XP000864581
GB betrifft die Einführung
eines „Spleißens" in die Proteintechnik.
-
Zusammenfassung
der Erfindung
-
Es
wurde ein allgemeines Verfahren zur in vivo Produktion und Screening
zyklischer Peptidbibliotheken entdeckt. Bei diesem Verfahren ist
ein Nukleinsäuremolekül derart
konstruiert, dass eine Nukleotidsequenz, die das zu zyklisierende
Peptid codiert am einen Ende mit einer Nukleotidsequenz flankiert
ist, die den carboxyterminalen Anteil eines getrennten (= split)
(oder trans) Inteins (C-Intein oder I
C)
und an einem anderen Ende mit einer Nukleotidsequenz flankiert ist,
die den aminoterminalen Anteil eines getrennten Inteins (N-Intein oder
I
N) codiert. Die Expression des Konstrukts
innerhalb eines Wirtssystems wie beispielsweise einer bakteriellen
oder eukaryontischen Zelle hat die Produktion eines Fusionsproteins
zur Folge. Die beiden getrennten Inteinbestandteile (d. h. I
C und I
N) des Fusionsproteins
assemblieren dann um ein aktives Enzym zu bilden, das die Amino-
und Carboxytermini zusammenspleißt, um ein Grundgerüst für ein zyklisches
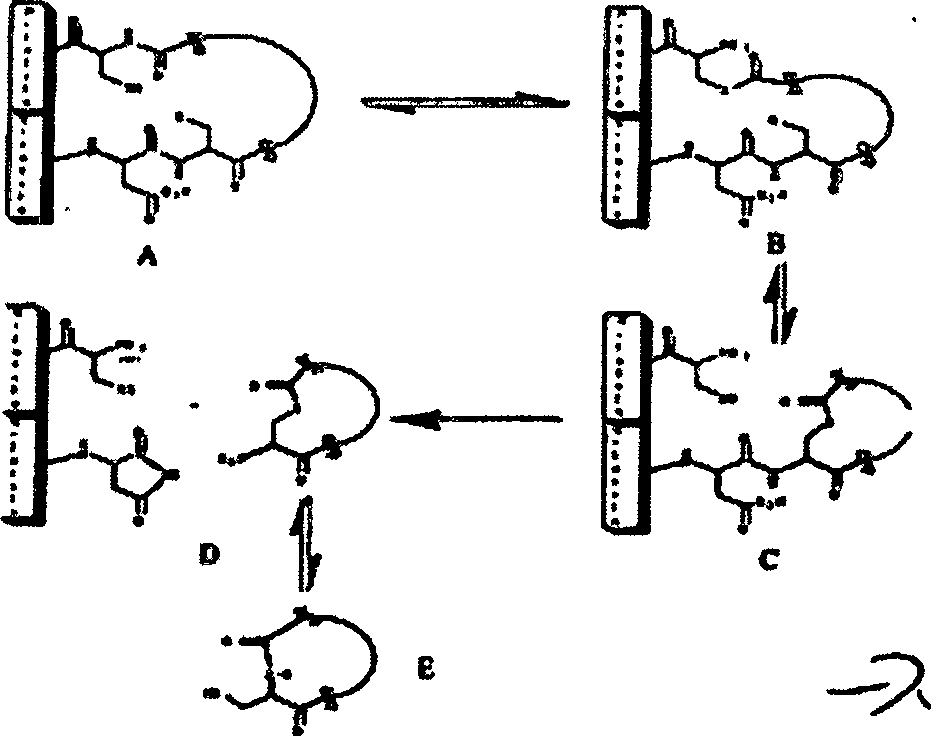
Peptid zu erzeugen. Die chemische Reaktion ist unten dargestellt. Mechanismus
der inteinvermittelten Zyklisierung
-
Die
Bildung des aktiven Inteins aus den amino- und carboxyterminalen
Fragmenten stabilisiert das Esterisomer einer Aminosäure an der
Kontaktstelle bzw. Kreuzung zwischen dem N-Intein und dem zu zyklisierenden
Peptid (in B oben, X=S oder 0). Wenn R=XH wird das Heteroatom aus
dem C-Intein im Gleichgewicht gehalten, um den Ester anzugreifen
und um ein zyklisches Ester-Zwischenprodukt (C) zu erzeugen. Die
Inteinkatalysierte Aminosuccinimidbildung (D) setzt das zyklische
Peptid (in der Lactonform) frei, das sich spontan umgruppiert, so
dass das thermodynamisch vorteilhafte Grundgerüst (Lactamform) zyklische Peptidprodukt (E)
gebildet wird. Dieses Verfahren kann angepasst werden, um die Auswahl
oder das Screening zyklischer Peptide mit vorher bestimmten Eigenschaften
zu erleichtern.
-
Demgemäß beansprucht
die vorliegende Erfindung ein nicht natürlich vorkommendes Proteinsäuremolekül, das ein
Polypeptid mit einem ersten Anteil eines getrennten Inteins, einen
zweiten Anteil eines getrennten (split) Inteins und ein Zielpeptid
codiert, das zwischen dem ersten Anteil eines getrennten Inteins
und dem zweiten Anteil eines getrennten Inteins eingebracht ist.
Die Expression des Nukleinsäuremoleküls in einem
Wirtssystem erzeugt ein Polypeptid, das im Wirtssystem spontan spleißt, so dass
sich eine zyklisierte Form des Zielproteins ergibt oder ein Spleißzwischenprodukt
einer zyklisierten Form des Zielpeptids, wie beispielsweise ein
aktives Inteinzwischenprodukt, ein Thioesterzwischenprodukt oder
ein Lariat-Zwischensprodukt.
-
Sowohl
der erste Anteil eines getrennten (Split-) Inteins als auch der
zweite Anteil eines Splitinteins können aus einem natürlich vorkommenden
Splitintein, wie beispielsweise Ssp DnaE gewonnen werden. Bei verschiedenen
Variationen können
ein oder beide Splitinteinanteile von nicht natürlich vorkommenden Splitinteinen,
wie beispielsweise solchen gewonnen werden, die von RecA, DnaB,
Psp Pol-I und Pfu-Inteinen abgeleitet sind.
-
In
einem weiteren Aspekt beansprucht die Erfindung ein nicht natürlich vorkommendes
Nukleinsäuremolekül, das ein
Polypeptid mit einem ersten Anteil eines Splitinteins, einem zweiten
Anteil eines Splitinteins, einem dritten Teil eines Splitinteins
und einem vierten Anteil eines Splitinteins codiert. Dieses Molekül kann ein erstes
Zielpeptid aufweisen, dass zwischen dem ersten Anteil eines Splitinteins
und dem zweiten Anteil eines Splitinteins angeordnet ist, und ein
zweites Zielpeptid wird zwischen dem dritten Anteil eines Splitinteins
und dem vierten Anteil eines Splitinteins eingebracht. Der erste
Anteil eines Splitinteins kann zum dritten Anteil eines Splitinteins
komplementär
sein, jedoch nicht zum zweiten Anteil eines Splitinteins, und der
zweite Anteil eines Splitinteins kann zum vierten Anteil eines Splitinteins
komplementär
sein, jedoch nicht komplementär zum
dritten Anteil eines Splitinteins.
-
Ebenfalls
im Umfang der Erfindung ist ein Expressionsvektor, der ein Nukleinsäuremolekül innerhalb der
Erfindung umfasst. Die Expression des Vektors in einem Wirtssystem
erzeugt ein Polypeptid, das im Wirtssystem spontan spleißt, so dass
sich ein zyklisches Peptid oder ein Spleißzwischenprodukt ergibt. Der
Expressionsvektor der Erfindung kann ebenfalls eine regulatorische
Sequenz enthalten, die die Expression des Polypeptids im Wirtssystem
erleichtert. Das Nukleinsäuremolekül des Vektors
kann eine Nukleotidsequenz einschließen, die ein Peptid codiert,
dass das Screening der zyklisierten Form des Zielpeptides bezüglich einer speziellen
Eigenschaft erleichtert und/oder einer Nukleotidsequenz, die ein
Peptid codiert, dass das Aufreinigen der zyklisierten Form des Zielpeptides
aus dem Wirtssystem erleichtert. Der Expressionsvektor kann ebenfalls
induzierbar sein.
-
Gemäß eines
weiteren Aspektes umfasst die vorliegende Erfindung einen Expressionsvektor,
der ein Polypeptid mit einem Zielpeptid codiert, das ein erstes
Ende fusioniert an einen ersten Anteil eines Splitinteins und ein
zweites Ende, fusioniert an einen zweiten Anteil eines Splitinteins
aufweist. Die Expressionsvektoren der Erfindung können ein
Plasmid, eine Bakteriophage, ein Virus, ein lineares Nukleinsäuremolekül oder eine andere
Art von Vektor sein.
-
Die
Erfindung umfasst zusätzlich
ein im wesentlichen reines Polypeptid mit einem ersten Anteil eines Splitinteins,
einem zweiten Anteil eines Splitinteins und ein Zielpeptid, das
sich zwischen dem ersten Anteil eines Splitinteins und einem zweiten
Anteil eines Splitinteins befindet. Das Polypeptid kann alles sein,
das spontan im Wirtssystem spleißt, um eine zyklisierte Form
des Zielpeptids zu ergeben oder es kann ein Spleißintermediat
sein.
-
Ebenfalls
innerhalb der Erfindung liegt ein Wirtssystem, das ein Nukleinsäuremolekül der Erfindung beherbergt.
Das Wirtssystem kann ein Prokaryont, wie beispielsweise ein Bakterium,
ein Archaebakterium, eine Eukaryonte wie beispielsweise eine Hefe
oder eine Säugetierzelle,
eine Pflanzenzelle, ein in vitro Transkriptions/Translationssystem
oder ein Zelllysat sein.
-
In
einem weiteren Aspekt umfasst die vorliegende Erfindung ein Verfahren
zur Herstellung eines Peptidmoleküls. Dieses Verfahren schließt die folgenden
Schritte ein: Bereitstellung eines isolierten Nukleinsäuremoleküls der Erfindung;
Bereitstellen eines Wirtssystems; Einbringen des isolierten Nukleinsäuremoleküls in das
Wirtssystem; und Exprimieren des isolierten Nukleinsäuremoleküls. In einer
Variation hat der Schritt des Exprimierens des isolierten Nukleinsäuremoleküls die Produktion
eines Polypeptids zur Folge, das spontan im Wirtssystem spleißt, um die
zyklisierte Form des Zielpeptids zu ergeben. Dieses Verfahren kann
ebenfalls den Schritt umfassen, die zyklisierte Form des Zielpeptides
aus dem Wirtssystem aufzureinigen.
-
Gemäß einer
weiteren Variation dieses Verfahrens hat der Schritt des Exprimierens
des isolierten Nukleinsäuremoleküls die Produktion
eines Spleißzwischenproduktes
einer zyklisierten Form des Zielpeptids zur Folge. Dieses Verfahren
kann ebenfalls den Schritt der Aufreinigung des Spleißzwischenproduktes
einer zyklisierten Form des Zielpeptides aus dem Wirtssystem umfassen.
Eine noch weitere Variation dieses Verfahrens schließt den Schritt
ein, das zyklische Peptid aus dem Spleißzwischenprodukt zu bilden.
-
Gemäß eines
weiteren Aspektes dieser Erfindung wird das Zielpeptid in einer
zyklisierten Form im Wirtssystem in Abwesenheit eines exogen zugesetzten
Mitteils wie beispielsweise einer Protease oder eines Thiols erzeugt.
-
Ein
weiterer Aspekt der Erfindung ist ein Verfahren zur Herstellung
einer Bibliothek aus Peptidmolekülen.
Dieses Verfahren involviert die Schritte der Bereitstellung einer
Vielzahl von Nukleinsäuremolekülen, die eine
Vielzahl von Zielpeptiden codieren, die heterogene Aminosäuresequenzen
aufweisen; den Einbau jeweils der Vielzahl von Nukleinsäuremolekülen in ein
Expressionsvektor zur Bildung einer Vielzahl von Expressionsvektoren
und die Expression der Expressionsvektoren im Wirtssystem. Die Vielzahl
von Nukleinsäuremolekülen ist
zwischen ein Nukleinsäuremolekül, das einen
ersten Anteil eines Splitinteins codiert und einem Nukleinsäuremolekül, das einen
zweiten Anteils eines Splitinteins codiert, eingebracht, in jeder
der gebildeten Expressionsvektoren derart, dass die Expression der
Expressionsvektoren in einem Wirtssystem die Erzeugung einer Vielzahl
von Peptidmolekülen,
wie beispielsweise Polypeptiden zur Folge hat, die spontan im Wirtssystem
spleißen,
um zyklisierte Formen der Zielpeptide zu ergeben oder das Spleißen von
Zwischenprodukten der zyklisierten Formen der Zielpeptide.
-
Und
gemäß eines
noch weiteren Aspektes schließt
die vorliegende Erfindung ein Verfahren zum Screening eines Peptidmoleküls nach
einer vorherbestimmten Eigenschaft ein. Dieses Verfahren schließt die folgenden
Schritte ein: Bereitstellen eines Nukleinsäuremoleküls, das ein Polypeptid codiert,
das einen ersten Anteil eines Splitinteins umfasst, einen zweiten
Anteil eines Splitinteins umfasst und ein Zielpeptid umfasst, dass
sich zwischen dem ersten Anteil eines Splitinteins und dem zweiten
Anteil eines Splitinteins befindet; Bereitstellen des Wirtssystems;
Einbringen des isolierten Nukleinsäuremoleküls in das Wirtssystem; Anordnen des
Wirtssystems unter Bedingungen, die die Erzeugung des Peptidmoleküls verursachen;
und Testen des Peptidmoleküls
bezüglich
vorherbestimmter Eigenschaften. Die Expression des Nukleinsäuremoleküls in einem
Wirtssystem erzeugt entweder eine zyklisierte Form des Zielpeptids,
die sich aus einem spontanen Spleißen des Polypeptids im Wirtssystems
ergibt oder ein Spleißzwischenprodukt
einer zyklisierten Form des Zielpeptides.
-
Gemäß einer
Variation dieses Verfahrens schließt die vorherbestimmte Eigenschaft
die Fähigkeit
ein, spezifisch ein Zielmolekül
zu binden und der Schritt des Testens des Peptidmoleküls nach
vorherbestimmten Eigenschaften schließt die Schritte ein (a) das
Peptidmolekül
mit dem Zielmolekül
in Berührung
zu bringen und (b) zu bestimmen, ob das Peptidmolekül an das
Zielmolekül
bindet. In einer weiteren Variation besteht die vorherbestimmte
Eigenschaft darin, eine biochemische Reaktion zu modulieren und
der Schritt des Testens des Peptidmoleküls nach vorherbestimmten Eigenschaften
umfasst den Schritt (a) das Peptidmolekül mit einem System in Berührung zu
bringen, dass die biochemische Reaktion enthält und (b) zu bestimmen, ob
das Peptidmolekül
die biochemische Reaktion moduliert. Der Schritt der Bestimmung,
ob das Peptidmolekül
an einem Zielmolekül
bindet oder eine biochemische Reaktion moduliert, kann durch Beobachten
einer Farbveränderung,
eines Fluoreszenzsignals, durch Analysieren des Zellzyklus oder
der Reproduktion eines Organismus gemessen werden.
-
Das
Zielmolekül
in diesem Verfahren kann ein zellassoziiertes Molekül sein,
wie beispielsweise ein membranassoziiertes Molekül oder ein intrazelluläres Molekül (beispielsweise
ein Kernmolekül
oder ein oder mehrere Organellen, wie beispielsweise Mitochondrien,
Lysosomen, endoplasmatische Reticula, Chloroplasten, Golgi und Periplasma).
Es kann ebenfalls ein extrazelluläres Molekül sein.
-
Die
biochemische Reaktion kann ein zellassoziierter Prozess sein, wie
beispielsweise ein intrazellulärer
Stoffwechselvorgang, ein membranassoziierter Vorgang, ein kernassoziierter
Vorgang. Es kann ebenfalls eine extrazelluläre Reaktion sein.
-
In
diesem Verfahren kann der Schritt des Testens des Peptidmoleküls bezüglich einer
vorherbestimmten Eigenschaft unter Verwendung eines Hybridsystems
durchgeführt
werden und/oder des Schrittes einer Immobilisierung des Peptidmoleküls an einem
Festphasenträger.
-
Die
Erfindung umfasst ebenfalls ein Verfahren zum Aufreinigen eines
zyklischen Peptides aus einem Gemisch. Dieses Verfahren schließt die folgenden
Schritte ein: Bereitstellen eines Gemisches, das ein Spleiß-Zwischenprodukt
enthält,
konjugiert mit einem Affinitäts-Marker; Vermischen
des konjugierten Spleiß-Zwischenproduktes
mit einem Festphasenträger,
der daran einen Liganden aufweist, der spezifischen an den Affinitätsmarker
derart bindet, dass der Träger
spezifisch mit dem Spleißzwischenprodukt
gebunden wird; Waschen des Trägers
zur Entfernung und spezifisch gebundenen Materials vom Träger; Zusetzen
eines Reagenzes zum Träger,
das ein zyklisches Peptid aus dem Spleißzwischenprodukt herstellt;
und Eluieren des zyklischen Peptids vom Träger.
-
In
einer Variation des vorhergehenden schließt die Erfindung ebenfalls
ein Verfahren zum Aufreinigen eines zyklischen Peptides aus einem
Gemisch ein, das die folgenden Schritte einschließt: Bereitstellen
eines Gemisches, das ein Spleißzwischenprodukt
konjugiert mit einem Affinitätsmarker
aufweist; Vermischen des konjugierten Spleißintermediats bzw. Zwischenproduktes
mit einem Festphasenträger,
der daran einen Liganden aufweist, der spezifisch den Affinitätsmarker
derartig bindet, dass der Träger
spezifisch mit dem Spleißzwischenprodukt
gebunden wird; Waschen des Trägers
zur Entfernung von unspezifisch gebundenen Material vom Träger; Eluieren
des Spleißzwischenproduktes
vom Träger;
und Zusetzen eines Reagenzes zum eluierten Spleißzwischenprodukt, das ein zyklisches
Peptid auf dem Spleißzwischenprodukt
herstellt.
-
Zusätzlich in
der Erfindung eingeschlossen ist ein Verfahren zur Aufreinigung
eines C-Moleküls, das ein
Spleißzwischenprodukt
aus einem Gemisch bindet. Dieses Verfahren schließt die folgenden
Schritte ein: Bereitstellung eines Festphasenträgers, der daran spezifisch
gebunden das Spleißzwischenprodukt
aufweist; Inberührungbringen
des Trägers
mit einem C-Molekül
im Gemisch; Waschen des Trägers
zur Entfernung und spezifisch gebundenen Materials vom Träger; und
Eluieren des Zielmoleküls
vom Träger.
-
Wie
hierin verwendet bedeutet der Satz „nicht natürlich vorkommend" direkt oder indirekt
hergestellt oder so hergestellt, dass eine humane Wirkung verursacht
wird. Somit ist ein nicht natürlich
vorkommendes Nukleinsäuremolekül eines,
das durch menschliche Manipulation erzeugt wurde und nicht durch
natürliche evolutionäre Prozesse.
-
Der
Begriff „Nukleinsäuremolekül" bedeutet irgendeine
Kette von zwei oder mehr Nukleotiden, die in Folge gebunden sind.
Beispielsweise kann ein Nukleinsäuremolekül eine DNA
oder eine RNA sein.
-
Wie
hierin verwendet bedeutet der Begriff „Peptid" eine Kette von zwei oder mehreren Aminosäuren, die
in Folge gebunden sind und schließt Polypeptide und Proteine
ein. Der Begriff „Polypeptid" bedeutet ein Polymer,
das aus zwei oder mehreren Peptiden besteht, unabhängig von
der Länge
oder von einer post-translationalen Modifikation. Der Begriff „Protein" bedeutet irgendeine
Kette von Aminosäuren
und schließt
Peptide, Polypeptide, Proteine und modifizierte Proteine, wie beispielsweise
Glycoproteine, Lipoproteine, Phosphoproteine, Metalloproteine und
dergleichen ein.
-
Ein „lineares
Peptid" ist ein
Peptid, dass nicht in ringförmiger
Form vorliegt und weist im allgemeinen sowohl eine carboxyterminale
Aminsosäure
mit einem freien Carboxy-Terminus
als auch eine aminoterminale Aminosäure mit einem freien Amino-Terminus
ein.
-
Im
Vergleich ist ein „zyklisches
Peptid" ein Peptid,
das „zyklisiert" wurde. Der Begriff „zyklisch" bedeutet, dass Atome
von Bestandteilen vorliegen, die einen Ring bilden. Wenn ein Peptid
bezeichnet wird, soll der Begriff „Zyklisieren" bedeuten, dass das
Peptid in eine zyklische oder „zyklisierte" Form ungewandelt
wird. Somit ist beispielsweise ein lineares Peptid „zyklisiert", wenn sein freier
Aminoterminius kovalent an seinen freien Carboxyterminus (d. h.
Kopf an Schwanz Format) gebunden ist, so dass keine freien Carboxy-
oder Aminoterminii im Peptid verbleiben.
-
Wie
hierin verwendet, ist ein „Spleißzwischenprodukt" ein Polypeptid,
dass während
der Intein-vermittelten Zyklisierungsreaktion erzeugt wird, die
oben dargestellt ist, vor der Bildung des freigesetzten zyklischen Peptidproduktes.
Spleißzwischenprodukte
schließen „aktive
Intein-Zwischenprodukte" (d.
h. solche mit einer chemischen Struktur, die dem in der obigen Abbildung
als „A" markierten Polypeptid ähnlich sind), „Thioesterzwischenprodukte" (d. h. solche mit
einer chemischen Struktur, die den in der obigen Abbildung als „B" markierten Polypeptid ähnlich ist)
und „Lariatzwischenprodukte" (d. h. solche mit
einer chemischen Struktur, die dem in der obigen Abbildung als „C" markierten Polypeptid ähnlich sind)
ein.
-
Der
Begriff „Zielpeptid" soll ein Peptid
bedeuten, dass zyklisiert oder in einem Spleißzwischenprodukt dargeboten
werden soll. Beispielsweise wäre
ein Peptid, dass sich zwischen einem carboxyterminalen Anteil eines
Splitinteins und einem aminoterminalen Anteil eines Splitinteins
in einem Vorläuferprotein
befindet ein Zielpeptid, wenn das Peptid nach Spleißen des
Vorläuferproteins
zyklisiert wird oder ein Teil eines Spleißzwischenproduktes nach Verarbeitung
bzw. Prozessierung (beispielsweise Faltung) des Vorläuferproteins
wird.
-
Wie
hierin verwendet bedeutet das Wort „Intein" eine natürlich vorkommende oder künstlich
konstruierte Polypeptidsequenz, die innerhalb eines Vorläuferproteins
eingebettet ist, die eine Spleißreaktion
während der
post-translationalen Prozessierung des Proteins katalysieren kann.
Eine Liste bekannter Inteine ist unter http://neb.com/inteins.html
veröffentlicht.
Ein „Splitintein" ist ein Intein,
dass zwei oder mehrere getrennte Bestandteile aufweist, die nicht
aneinander fusioniert sind.
-
Wie
hierin verwendet, bedeutet der Begriff „dazwischen angeordnet" sich dazwischen
befinden. Somit ist einem Polypeptid, das ein erstes Peptid angeordnet
zwischen einem zweiten und einem dritten Peptid aufweist, die Kette
der Aminosäuren,
die das erste Peptid ausmachen, physisch zwischen der Kette der
Aminosäuren,
die das zweite Peptid ausmachen und der Kette der Aminosäuren, die
das dritte Peptid ausmachen, angeordnet.
-
Eine
Vielzahl von Peptiden mit „heterogenen
Aminosäuresequenzen" bedeutet, dass die
Vielzahl von Peptiden aus zumindest zwei, jedoch im allgemeinen
einer größeren Anzahl
von unterschiedlichen Peptiden unterschiedlicher Aminosäuresequenzen
zusammengesetzt ist.
-
Wie
hierin verwendet, betrifft der Begriff „Wirtssystem" irgendein Medium
oder Träger,
bei dem ein Nukleinsäuremolekül transkribiert,
repliziert und/oder translatiert werden kann; und/oder irgendein
Medium oder Träger,
in den ein Polypeptid gespleißt
oder in anderer Weise post-translational prozessiert werden kann.
-
Wie
hierin verwendet, bedeutet das Wort „spontan", dass die beschriebene Wirkung ohne
Zusatz einer exogenen Substanz eintritt. Beispielsweise spleißt ein Vorläuferpolypeptid
innerhalb eines Wirtssystems spontan im Wirtssystem, um ein zyklisches
Peptid zu ergeben, wenn nichts dem Wirtssystem zugesetzt wird, als das
Vorläuferpolypeptid
oder ein Nukleinsäuremolekül, dass
das Vorläuferpolypeptid
codiert. Im Vergleich spleißt
ein Vorläuferpolypeptid
innerhalb eines Wirtssystems nicht spontan im Wirtssystem, wenn
ein Mittel von außen
dem Wirtssystem zugeführt
werden muss, um das zyklische Peptid zu erzeugen.
-
Wie
hierin verwendet, bedeutet der Begriff „Spleiß" oder „Spleißen", einen zentralen Anteil des Polypetids
zur Bildung von zwei oder mehreren kleinen Polypeptidmolekülen auszuschneiden.
In einigen Fällen schließt das Spleißen ebenfalls
den Schritt ein, zwei oder mehrere kleinere Polypeptide zusammen
zu fusionieren, um ein neues Polypeptid zu bilden.
-
Wie
hierin verwendet, bedeutet das Wort „abgeleitet", direkt oder indirekt
gewonnen von, isoliert von, aufgereinigt aus, abstammend von oder
in anderer Weise sich aus etwas ergebend.
-
Wie
hierin verwendet, bedeutet der Satz „Expressionsvektor" einen Träger, der
die Transkription und/oder Translation eines Nukleinsäuremoleküls in einem
Wirtssystem erleichtert. Ein Expressionsvektor ist „induzierbar", wenn eine exogene
Substanz einem Wirtssystem zugesetzt wird, das den Expressionsvektor enthält und verursacht,
dass der Vektor exprimiert wird (beispielsweise verursacht es, dass
ein Nukleinsäuremolekül innerhalb
des Vektors in mRNA transkribiert wird).
-
Der
Begriff „Expression
von" einer Nukleinsäure bedeutet,
dass die Nukleinsäure
zu einem Polypeptid transkribiert und/oder translatiert und/oder
repliziert wird.
-
Wie
hierin verwendet, bedeutet der Begriff „regulatorische Sequenz" eine Nukleotidsequenz,
die die Expression „beispielsweise
Transkription" eines
Nukleinsäuremoleküls moduliert.
Beispielsweise sind Promotoren und Enhancer regulatorische Sequenzen.
-
Der
Begriff „fusioniert" bedeutet kovalent
gebunden. Beispielsweise wird ein erstes Peptid an ein zweites Peptid
fusioniert, wenn die beiden Peptide aneinander kovalent gebunden
werden (beispielsweise über eine
Peptidbindung).
-
Wie
hierin verwendet, ist eine „isolierte" oder „im wesentlichen
reinen" Substanz
eine, die aus Bestandteilen abgetrennt wurde, die diese natürlicherweise
begleiten. Typischerweise ist ein Polypeptid im wesentlichen rein,
wenn es zumindest 50% (beispielsweise 60%, 70%, 80%, 90%, 95% und
99%) nach Gewicht frei von den anderen Proteinen und natürlich vorkommenden
organischen Molekülen
ist, mit denen sie natürlicherweise
assoziiert ist.
-
Eine „Vorläufer-DNA" ist insbesondere
Desoxyribonukleinsäure,
aus der Mutationen hergestellt werden oder auf diesen basieren.
-
Der
Begriff „Zielmolekül" bedeutet jedes Molekül, das verwendet
wird, um die Bindung oder funktionellen Eigenschaften eines anderen
Moleküls
zu bestimmen.
-
Hierin
bedeutet „binden" oder „bindet", dass ein Molekül ein anderes
Molekül
in einer Probe erkennt und an diesem anhaftet bzw. sich an dieses
bindet, jedoch im wesentlichen nicht andere Moleküle in der
Probe erkennt oder an diese bindet. Ein Molekül „bindet spezifisch" ein weiteres Molekül, wenn
es eine Bindungsaffinität
von mehr als 105 bis 106 Liter/Mol
für das
andere Molekül
aufweist.
-
Ein „zellassoziierter
Prozess" ist einer,
der innerhalb einer Zelle oder in großer Nähe der Zelle stattfindet.
-
Ein „membranassoziiertes
Ereignis" ist ein
zellassoziierter Prozess, der auf der Plasmamembran einer Zelle
stattfindet.
-
Ein „Kernereignis" ist ein zellassoziierter
Prozess, der im Kern einer Zelle stattfindet.
-
Im
Vergleich zu einem zellassoziierten Ereignis ist eine „extrazelluläre Reaktion" eine, die nicht
innerhalb einer Zelle stattfindet.
-
Der
Begriff „Hybridsystem" bedeutet zwei Hybridsysteme,
reverse Zwei-Hybridsysteme, Einhybridsysteme, Split-Hybridsysteme,
kleine Moleküle
bzw. small molecule Hybridsysteme und alle ähnlichen Systeme zum Identifizieren
von Interaktionen zwischen Peptiden und anderen Molekülen (beispielsweise
Proteinen und Nukleinsäuremolekülen). Bezüglich eines Überblicks
beispielhafter Hybridsysteme, siehe Vidal und Legrain, Nucleic Acids
Res. 27:919, 1999.
-
Soweit
nichts anderes definiert, weisen alle hierin genannten technischen
Begriff dieselbe Bedeutung auf, wie sie üblicherweise vom Durchschnittsfachmann
auf dem Gebiet verstanden werden, den die Erfindung betrifft. Obwohl
Verfahren und Materialien verwendet werden können, die den hierin beschriebenen ähnlich oder äquivalent
sind, um die vorliegende Erfindung in der Praxis auszuführen oder
zu testen, sind geeignete Verfahren und Materialien unten beschrieben.
Alle Veröffentlichungen,
Patentanmeldungen, Patente und andere hierin erwähnten Bezugnahmen, sind durch
Bezugnahme in ihrer Gesamtheit mit aufgenommen. Im Falle eines Konfliktes
entscheidet die vorliegende Beschreibung einschließlich der
Definitionen. Zusätzlich
sind spezielle Ausführungsformen,
die unten diskutiert werden, lediglich illustrativ und soll en nicht
als Einschränkung gesehen
werden.
-
Weitere
Merkmale und Vorteile der Erfindung ergeben sich aus der folgenden
ausführlichen
Beschreibung und aus den Ansprüchen.
-
Kurze Beschreibung
der Zeichnungen
-
Die
vorliegende Erfindung wird genau in den beiden beigefügten Ansprüchen dargelegt.
Die obigen und weiteren Vorteile dieser Erfindung sind besser verständlich,
indem auf die nachfolgende Beschreibung in Verbindung mit den begleitenden
Zeichnungen Bezug genommen wird, in denen:
-
1 eine
schematische Darstellung eines Überblickes
einer allgemeinen Zyklisierungsreaktion innerhalb der Erfindung
ist.
-
2 eine
schematische Darstellung einer Reihe von chemischen Reaktionsschritten
ist, die in einem Peptidzyklisierungsverfahren der Erfindung auftreten.
-
3 ist
eine Genkarte eines (a) Plasmides pARCB, (b) Plasmid pARCP-DHFR,
(c), Plasmid pARCPAH-DHFR, (d) eines modifizierten Vektors, der
ein Cystein (TGY) oder Serin (TCN) Codon aufweist, erzeugt durch
Klonieren in den Mfel Ort (N repräsentiert eine Nucleobase, S
repräsentiert
C oder G und Y repräsentiert Pyrmindine),
(e) Plasmid pARCP-p und (f) Plasmid pARCBD-p.
-
4 ist
eine fotographische Darstellung einer Natriumdodecylsulfatpolyacrylamidgelelectrophorose (SDS-PAGE)-Analyse
einer Dehydrofolatreductase (DHFR) Zyklisierung auf einen 10-20%
Gradienten, Tris/Glycin-Fertiggel (Biorad).
-
5 ist
eine graphische Darstellung einer DHFR Aktivität von Wildtyp (Dreiecke) und
zyklischem DHFR (Rauten) Aktivität
nach Präinkubation
bei 365°C.
-
6 ist
eine schematische Darstellung der erwarteten Endoproteinase Lys-C
Verdauungsmuster für lineare
und zyklische DHFR.
-
7 ist
eine Fotographie von FeCuY Platten, die in einem in vivo Effekt
verwendet werden, um die Tyrosinasehemmung durch Pseudostellarin
F nachzuweisen.
-
8 ist
eine schematische Darstellung eines Verfahrens zur Aufreinigung
zyklischer Peptide im Umfang der vorliegenden Erfindung.
-
9 ist
eine schematische Darstellung eines weiteren Verfahrens zur Aufreinigung
zyklischer Peptide im Umfang der vorliegenden Erfindung.
-
10 ist
eine schematische Darstellung eines Festphasenträger/Affinitätschromatographie basierten
Verfahrens zum Identifizieren/Aufreinigen von Molekülen, die
spezifischen an ein Spleiß-Zwischenprodukt binden.
-
11 ist
eine schematische Darstellung eines weiteren Festphasenträger/Affinitätschromatographie basierten
Verfahrens zum Identifizieren/Aufreinigen von Molekülen, die
spezifisch an ein Spleißzwischenprodukt
binden.
-
12 ist
eine schematische Darstellung eines weiteren Festphasenträger/Affinitätschromatographie basierten
Verfahrens zum Identifizieren/Aufreinigen von Molekülen, die
spezifisch ein Spleißzwischenprodukt binden.
-
13 ist
eine schematische Darstellung eines weiteren Festphasenträger/Affinitätschromatographie basierten
Verfahrens zum Identifizieren/Aufreinigen von Molekülen, die
spezifisch ein Spleißzwischenprodukt binden.
-
14 ist
eine schematische Darstellung der Verwendung von Aptamergerüsten in
der Erfindung.
-
15 ist
eine schematische Darstellung von zwei Reaktionen zur Herstellung
von Aptameren im Umfang der vorliegenden Erfindung.
-
16 ist
eine schematische Darstellung eines Verfahrens zum Screening innerhalb
der Erfindung.
-
17 ist
eine schematische Darstellung eines weiteren Verfahrens zum Screening
innerhalb des Umfangs der vorliegenden Erfindung.
-
18 ist
eine schematische Darstellung eines weiteren Verfahrens zum Screening
innerhalb des Umfangs der vorliegenden Erfindung.
-
19 ist
eine schematische Darstellung eines weiteren Verfahrens zum Screening
innerhalb des Umfangs der vorliegenden Erfindung.
-
Ausführliche Beschreibung
-
Die
Trans-Spleißfähigkeit
von Splitinteinen wurde ausgebeutet, um ein allgemeines Verfahren
zur Herstellung zyklischer Peptide und von Spleißzwischenprodukten zu entwickeln,
die Peptide in einer Loop- bzw. Schlaufen-Konformation zeigen. In
diesem Verfahren ist ein Zielprotein zwischen zwei Anteilen eines
Splitinteins in einem Vorläuferpolypeptid
angeordnet. In einem geeigneten Wirtssystem kommen die beiden Anteile des
Splitinteins physisch zusammen und bilden ein aktives Intein in
einer Konformation, die auch das Zielpeptid in eine Loop-Konfiguration
zwingt. In dieser Konfiguration wird das Esterisomer der Aminosäure an der Grenze
zwischen den einen Inteinanteilen (beispielsweise IN)
und dem Zielpeptid derart stabilisiert, dass ein Heteroatom vom
anderen Anteil des Inteins (beispielsweise IC)
dann mit dem Ester reagieren kann, um ein zyklisches Esterzwischenprodukt
zu bilden. Das aktive Intein katalysiert dann die Bildung eines
Aminosuccinimids, das eine zyklisierte Form des Zielpeptids freisetzt
(d. h. eine Lactonform), die sich dann spontan umordnet, um das
thermodynamisch günstige
Grundgerüst
des zyklischen Peptidproduktes zu bilden (d. h. die Lactamform).
Durch Anhalten der Reaktion an gegebenen Punkten vor Freisetzung
des zyklischen Peptides können
Spleißzwischenprodukte,
die das Zielpeptid in einer Loop-Konfiguration
tragen, erzeugt werden. Um derartige Peptide zu erzeugen, können Nukleinsäuremoleküle, die
ein Polypeptid mit der Zielpeptidsequenz zwischen den beiden Inteinanteilen
eingefügt,
konstruiert werden. Die Einbringung dieser Konstrukte in einen Expressionsvektor
stellt ein Verfahren zur Erzeugung des Polypeptids in einem Wirtssystem
bereit, wobei das Polypeptid zu einem zyklischen Peptid oder einem
Spleißzwischenprodukt
gespleißt
werden kann. Unter Anwendung dieses Verfahrens können mehrere unterschiedliche
zyklische Peptide oder Spleißzwischenprodukte hergestellt
werden, um eine Bibliothek von zyklisierten oder teilweise zyklisierten
Peptiden zu erzeugen, die für
spezielle Eigenschaften gescreent werden können.
-
Unter
Bezugnahme auf 1 schließt ein Überblick über eine Ausführungsform
der Erfindung ein Verfahren zur Herstellung eines zyklischen Peptides
aus einem Nukleinsäuremolekül ein. In
diesem Verfahren wird ein Nukleinsäuremolekül so hergestellt, dass seine
Nukleotidsequenz ein Polypeptid codiert, dass eine aufeinanderfolgende
Reihenfolge eines ersten Anteils eines Splitinteins (beispielsweise
IC), eines zu zyklisierenden Peptids (d.
h. ein Zielpeptid) und einen zweiten Anteil eines Splitinteins (beispielsweise
IN) codiert. Das Nukleinsäuremolekül kann in
einen Expressionsvektor eingebracht werden, um seine Expression
in einem Wirtssystem zu erleichtern, worin die Nukleinsäure transkribiert
und in ein Vorläuferpolypeptid
translatiert werden kann, dass das zu zyklisierende Peptid zwischen
zwei Splitinteinanteilen eingefügt
aufweist. Durch die obigen Schritte kommen die beiden Anteile des
Splitinteins zusammen und ordnen das Vorläuferprotein in einer Konformation
an, die chemische Reaktionen beginnt, die letztendlich ein zyklisches
Peptid zur Folge haben (siehe 2).
-
Nukleinsäuremoleküle
-
Nukleinsäuremoleküle innerhalb
der Erfindung schließen
solche ein, die ein Polypeptid mit einem ersten Anteil eines Splitinteins,
einen zweiten Anteil eines Splitinteins und einem Zielpeptid codieren,
das sich zwischen dem ersten Anteil eines Splitinteins und dem zweiten
Anteil eines Splitinteins befindet. In einer Ausführungsform
der Erfindung hat die Expression des Nukleinsäuremmoleküls in einem Wirtssystem ein
Polypeptid zur Folge, das spontan im Wirtssystem spleißt, so dass
sich eine zyklisierte Form des Zielpeptids ergibt. In einer weiteren
Ausführungsform
der Erfindung hat die Expression des Nukleinsäuremoleküls in einem Wirtssystem ein
Polypeptid zur Folge, das ein Spleiß-Zwischenprodukt einer zyklisierten Form
des Zielpeptids ist. Die Nukleinsäuren der Erfindung können gemäß der Verfahren
hergestellt werden, die hierin beschrieben sind und können ebenfalls
unter Verwendung der hierin vorgesehenen Anleitung in Verbindung
mit Verfahren zur Herstellung und Manipulierung von Nukleinsäuremolekülen hergestellt
werden, die allgemein in der Technik bekannt sind (siehe beispielsweise
Ausubel et al., Herausgeber, Current Protocols in Molecular Biology,
New York: John Wiley & Sons,
1997; Sambrook et al., Molecular Cloning: A laboratory Manual (2nd Edition), Cold Spring Harbor Press, 1998).
Beispielsweise kann ein Nukleinsäuremolekül innerhalb
des Umfangs der Erfindung hergestellt werden, indem separat ein
Polynukleotid hergestellt wird, dass den ersten Anteil eines Splitinteins
codiert, ein Polynukleotid, das den zweiten Anteil eines Splitinteins
codiert und ein Polynukleotid, das das Zielpeptid codiert. Die drei
Polynukleotide können
zusammen ligiert werden, um ein Nukleinsäuremolekül zu bilden, das ein Polypeptid
codiert, dass das Zielpeptid zwischen dem ersten Anteil eines Splitinteins
und dem zweiten Anteil eines Splitinteins eingebracht aufweist.
-
Nukleinsäuren, die
Inteine codieren
-
Nukleotidsequenzen,
die den ersten Anteil eines Splitinteins und den zweiten Anteil
eines Splitinteins der Nukleinsäuremoleküle innerhalb
der Erfindung codieren, können
aus bekannten Inteinen abgeleitet werden. Eine reichhaltige und
beschreibende Liste derartiger Inteine wurde durch das New England
Biolabs unter http//wvvw.neb.com/intein/int_reg.html veröffentlicht.
Jedes dieser bekannten Intein kann verwendet werden, solang sie
mit der Erfindung kompartibel sind.
-
Nukleotidsequenzen,
die entweder natürlich
vorkommende oder künstlich
produzierte Splitinteine codieren, können verwendet werden, um die
Inteinanteile von Nukleinsäuremolekülen innerhalb
der Erfindung zu erzeugen. Natürlich
vorkommende Splitinteine werden in der Natur als zwei getrennte
Komponenten expremiert, die aneinander binden um ein aktives Spleißmittel
zu bilden. Die Nukleinsäuremoleküle, die
diese natürlich
vorkommenden Komponenten codieren, können somit in der Erfindung
verwendet werden. Ein Beispiel eines natürlich vorkommenden Splitinteins,
das verwendet werden kann, ist Ssp DnaE (Wu et al, Proc. Natl. Acad.
Sci. USA 95:9226,1998).
-
Inteine,
die in ihrem natürlichen
Zustand (d. h. solche, die in einer kontinuierlichen Kette von Aminosäuren existieren),
nicht getrennt bzw. gespalten bzw. gespleißt sind, können künstlich unter Verwendung bekannter
Techniken getrennt werden. Beispielsweise können zwei oder mehr Nukleinsäuremoleküle, die
unterschiedliche Anteile solcher Inteine codieren, hergestellt werden,
so dass ihre Expression zwei oder mehr künstliche Splitinteinebestandteile
mit sich bringt. Siehe Beispielsweise Evans et al, J. Biol. Chem.
274:18359, 1999; Mills et al, Proc. Natl. Acad. Sci. USA 95:3543,
1998. Die Nukleinsäuren,
die derartige nicht natürlich
vorkommende Inteinbestandteile (Anteile) codieren, können in
der Erfindung verwendet werden. Solche Nukleinsäuremoleküle, die nicht natürlich vorkommende
Splitinteinanteile codieren, die effizient am selben Vorläuferpolypeptid
interagieren, um zyklische Peptide oder Spleißzwischenprodukte zu ergeben,
werden bevorzugt. Beispiele für
nicht natürlich
vorkommende Splitinteine, von denen solche Nukleinsäuremoleküle abgeleitet
werden können,
schließen
Psp Pol-1 (Southworth, M.W., et al., The EMBO J. 17:918, 1998),
Mycobacterium tuberculosis RecA Intein, (Lew, B.M., et al, J. Biol.
Chem. 273:15887, 1998; Shingledecker, K., et al, Gene 207:187, 1998;
Mills, K.V., et al, Proc. Natl. Acad. Sci. USA 95:3543, 1998) Ssp
DnaB/Mxe GyrA (Evans, T.C. et al, J. Biol. Chem. 274:18359, 1999),
und Pfu (Otomo et al, Biochemistry 38:16040, 1999; Yamazaki et al,
J. Am. Chem. Soc. 120:5591, 1998).
-
Nukleinsäuren die
Zielpeptide oder Peptide codieren die in Spleißzwischenprodukten gezeigt
werden.
-
Zahlreiche
Verfahren zur Herstellung von Nukleinsäuren, die Peptide einer bekannten
oder eine zufallsbedingten Sequenz codieren, sind in der Technik
bekannt. Beispielsweise können
Polynukleotide, die eine vorher bestimmte oder eine zufallsbedingte
Sequenz aufweisen, chemisch durch Festphasensynthese unter Verwendung
kommerziell erhältlicher
Ausrüstung
und Reagenzien hergestellt werden. Eine polymerase Kettenreaktion
kann ebenfalls dazu verwendet werden, Polynukleotide bekannter oder
zufallsbedingter Sequenzen herzustellen. Siehe beispielsweise Ausubel
et al, oben. Als weiteres Beispiel können Restriktionsendonukleasen
dazu verwendet werden, ein größeres Nukleinsäuremolekül oder sogar
ganze chromosomale DNA in eine Vielzahl kleinerer Polynukleotidfragmenten
enzymatisch aufzuspalten bzw. zu verdauen, die dazu verwendet werden
können,
Nukleinsäuremoleküle der vorliegenden
Erfindung herzustellen.
-
Polynukleotide,
die Peptidsequenzen codieren, die zyklisiert werden sollen, werden
vorzugsweise so hergestellt, dass ein Terminus des Polynukleotides
einen Asparagin-, Serin-, Cystein- oder Threoninrest codiert, um
die Zyklisierungsreaktion zu vereinfachen. Aus demselben Grund werden
Polynukleotide, die Peptidsequenzen zur Herstellung von Spleißzwischenprodukten
codieren, vorzugsweise so hergestellt, dass der Terminus eine andere
Aminosäure
als Asparagin-, Serin-, Cystein oder Threonin-Reste codiert, so
dass die Zyklisierungsreaktion vermieden wird.
-
Ligierung von Polynukleotiden
die Inteinproteine und Zielpeptide oder Peptide codieren die Spleißzwischenprodukte
zeigen
-
Wenn
sie einmal etabliert sind, können
die herkömmlichen
Verfahren verwendet werden, Nukleinsäuremoleküle, die Inteinproteine codieren,
an ein Nukleinsäuremolekül zu ligieren,
das ein Zielpeptid (oder ein Peptid in einem Spleißzwischenprodukt)
codiert, um ein größeres Nukleinsäuremolekül zu bilden,
das ein Polypeptid codiert, das die erste Inteinanteil-Zielpeptid-zweite
Inteinanteil-Reihenfolge aufweist. Siehe beispielsweise Ausubel
et al, oben.
-
Nukleinsäuremoleküle, die
vielfache Splitinteine und multiple Peptide codieren
-
Unter
Verwendung von Techniken, die den oben beschriebenen ähnlich sind,
könnte
der Fachmann auf dem Gebiet ebenfalls Nukleinsäure herstellen, die mehr als
einen Satz von zwei Anteilen eines Splitinteins codieren, in dem
Peptide eingebracht sind. Beispielsweise schließt die Erfindung Nukleinsäuremoleküle ein, die
ein Vorläuferpolypeptidmolekül codieren,
das N Polypeptide (N = eine ganze Zahl größer als oder gleich 1) umfassen
und die N-Zielpeptide zwischen 2N Inteinproteinen eingefügt aufweisen,
derart, dass irgendein Zielpeptid i (i = eine ganze Zahl größer als
1 und repräsentiert
die Position eines Zielpeptids im Vorläuferpeptid) zwischen Inteinanteil
2i-1 und 2i eingefügt
ist (beispielsweise befindet sich Zielpeptid 1 zwischen Inteinanteil
1 und 2, Zielpeptid 2 zwischen Inteinanteilen 3 und 4 etc.). Solange
die Inteinproteine 2i-1 und 2i nicht komplementär sind (d. h. dazu in der Lage
sind, physisch Wechsel zu wirken, um ein Spleißereignis zu katalysieren) kann
das Zielpeptid i nicht zyklisieren. Wenn jedoch der Inteinanteil
2i mit dem Inteinanteil 2i+1 komplementär ist und der Inteinanteil
2n mit dem Inteinanteil 1 komplementär ist, kann die Gesamtanordnung
von N Polypeptiden N-1 Trans-Spleißungen (zwischen 2 Polypeptiden)
und eine Cisspleißung
(Ligierung der beiden Enden aneinander) durchführen, um ein Produkt entstehen
zu lassen, bei dem 1-N Zielpeptide covalent aneinander in einem
zyklischen Peptid/Protein gebunden sind (beispielsweise Inteinanteile
2 & 3 transspleißen die
Zielpeptide 1 und 2; die Inteinanteile vier & fünf transspleißen die
Zielpeptide 2 & 3;
die Inteinanteile 2N-2 & 2N-1 transspleißen die
Zielpeptide N-1 & N;
und die Inteinanteile N & 1
cis-spleißen
um das zyklische Produkt zu schließen, das die N-Zielsequenzen
enthält).
Die Reihenfolge der Trans-/Cisspleißereignisse ist irrelevant.
Die am langsamsten spleißende
Spezies (gleichgültig,
ob sie der komplementäre
Inteinanteil 2N&1,
2&3 oder 80&81 ist) wird bei
Fehlen das Cis-Spleißen
durchführen.
-
Somit
können
Nukleinsäurekonstrukte
hergestellt werden, die zwei oder mehrere Polypeptide exprimieren,
die jeweils aus einem Zielpeptid zusammengesetzt sind, das sich
zwischen zwei Anteilen eines Splitinteins befindet, wobei die Inteinbestandteile
nicht komplementär
sind (d. h. nicht vom selben Intein abgeleitet sind oder in anderer
Weise kooperieren, um irgendeine der Zyklisierungsreaktionen zu
katalysieren). In solchen Konstrukten kann kein Polypeptid zyklisiert
werden, bis es in Gegenwart eines zweiten Polypeptids exprimiert
wird, das den geeigneten komplementären Inteinbestandteil aufweist.
Konstrukte solcher Nukleinsäuren
innerhalb des Umfangs der Erfindung können nur eine Polypeptid pro
Konstrukt oder mehr als ein Polypeptid pro Konstrukt codieren (beispielsweise
ein bifunktionelles Plasmid).
-
Expressionsvektoren
-
Die
Expressionsvektoren der vorliegenden Erfindung können durch Einfügen von
Polynukleotiden, die ein Zielpeptid codieren, in einen geeigneten
Expressionsvektor hergestellt werden, der die Expression des Polynukleotids
in einem Wirtssystem erleichtert. Derartige geeignete Vektoren schließen Plasmide,
Bacteriophagen und virale Vektoren ein. Eine große Anzahl von diesen ist in
der Technik bekannt und viele sind kommerziell erhältlich oder
aus der wissenschaftlichen Wissenschaftsgemeinschaft erhältlich.
Der Fachmann auf dem Gebiet kann geeignete Vektoren zur Verwendung
in einer speziellen Anwendung auf Grundlage beispielsweise der Art
des ausgewählten
Wirtssystems (beispielsweise in vitro Systeme, prokaryontische Zellen,
wie beispielsweise Bakterien und eukaryontische Zellen, wie beispielsweise
Hefe- oder Säugetierzellen)
und den ausgewählten
Expressionsbedingungen auswählen.
-
Expressionsvektoren
im Umfang der vorliegenden Erfindung können eine Strecke von Nukleotiden einschließen, die
ein Zielpolypeptid codieren und eine Strecke von Nukleotiden, die
als regulatorische Domäne funktionieren,
die Expression moduliert oder kontrolliert (beispielsweise die Transkription)
von Nukleotidsequenzen innerhalb des Vektors. Beispielsweise kann
die regulatorische Domäne
ein Promotor oder ein Enhancer bzw. Verstärker sein.
-
Expressionsvektoren
innerhalb der Erfindung können
Nukleotidsequenzen einschließen,
die ein Peptid codieren, dass das Screening der zyklisierten Form
des Zielpeptids oder das Spleißen
eines Zwischenproduktes bezüglich
einer speziellen Eigenschaft (beispielsweise ein Affinitätsmarker,
wie beispielsweise eine Chitinbindungsdomäne oder eine Biotinmarkierung;
eine farb- oder lichtemittierende Markierung; eine radioaktive Markierung
etc.) oder die Aufreinigung der zyklisierten Form des Zielpeptides
oder des Spleißzwischenproduktes
aus einem Wirtssystem (beispielsweise eine Affinitätsmarkierung,
wie beispielsweise eine Chitinbindungsdomäne, eine Biotinmarkierung,
eine farb- oder lichtemittierende Markierung, eine radioaktive Markierung
etc.) erleichtert.
-
In
bevorzugten Ausführungsformen
werden die Expressionsvektoren im Rahmen der vorliegenden Erfindung
mit Restriktionsorten sowohl zwischen als auch innerhalb der Nukleinsäuresequenzen
hergestellt, die die Splitinteinanteile codieren, um die Klonierung
einer breiten Vielzahl von Zyklisierungstargets oder Spleißzwischenprodukten
zu ermöglichen.
In einigen Ausführungsformen
kann ein Expressionvektor der vorliegenden Erfindung ein induzierbarer
Expressionsvektor, beispielsweise ein Arabinose induzierbarer Vektor
sein. Solche Vektoren können
zur Kontrolle der Expression von Zyklisierungsvorläufern oder
Spleißzwischenprodukten
innerhalb eines Wirtssystems verwendet werden. Weitere Vektoren
können
zur Verwendung in der Erfindung auf Grundlage Ihrer Kompatibilität mit bekannten
bakteriellen Expressionsstämmen
und Hybridsystemen ausgewählt
werden. Siehe beispielsweise Zhang et al, Curr. Biol. 9:417, 1999;
Pellitier et al, Nat. Biotechnol. 17:683, 1999; Karimova et al,
Proc. Natl. Acad. Sci. USA 95:5752, 1998; Dmitrova et al, Mol. Gen.
Genet., 257:205, 1998; Xu et al, Proc. Natl. Acad. Sci. USA 96:151,
1999, Rossi et al, Proc. Natl. Acad. Sci. USA 94:8405, 1997.
-
Polypeptide
-
Polypeptide
im Umfang der Erfindung schließen
irgendwelche Polypeptide ein, die durch Expression einer Nukleinsäure der
Erfindung hergestellt werden können.
Beispielsweise ist ein im wesentlichen reines Vorläuferpolypeptid,
das ein Zielpeptid aufweist (oder ein Peptid, das von einem Spleißzwischenprodukt
dargeboten wird) zwischen dem ersten Anteil eines Splitinteins und
dem zweiten Anteils eines Splitinteins eingefügt ist, in der vorliegenden
Erfindung eingeschlossen. In einigen Ausführungsformen des Vorläuferpolypeptids kann
das Zielpeptid direkt an den ersten und zweiten Inteinanteilen fusioniert
werden. Das Vorläuferpolypeptid spleißt spontan
im Wirtssystem, so dass sich eine zyklisierte Form des Zielpeptids
ergibt (oder ein Spleißzwischenprodukt,
das ein Peptid zeigt).
-
Zyklisierte
Formen von Zielpeptiden und Spleißzwischenprodukte, die Peptide
darbieten, liegen ebenfalls innerhalb der vorliegenden Erfindung.
Vorzugsweise werden diese durch Spleißen bzw. Spalten eines Vorläuferpeptids
der Erfindung produziert. Die zyklisierten Formen der Zielpeptide
können
irgendeine Aminosäuresequenz
aufweisen, die durch die Verfahren der Erfindung zyklisiert werden
können.
Das Spleißzwischenprodukt
kann ein aktives Inteinzwischenprodukt, eine Thioesterzwischenprodukt
oder ein Lariatzwischenprodukt sein und kann ein Peptid irgendeiner
kompatiblen Aminosäuresequenz
zeigen.
-
Wirtssysteme
-
Wirtssysteme,
die in der Erfindung verwendet werden können, schließen irgendwelche
Systeme ein, die die Transkription, Translation und/oder Replikation
eines Nukleinsäuremoleküls der Erfindung
unterstützen;
oder die eine posttranslationale Modifikation (beispielsweise Spleißen) eines
Polypeptids oder Proteins der Erfindung unterstützen. Zahlreiche derartige
Wirtssysteme sind bekannt. Beispielsweise kann das Wirtssystem in
der Erfindung insbesondere wenn es wünschenswert ist, Artefakte
oder Störungen
zu vermeiden, die durch lebende Wirtssysteme verursacht sind, die
Form eines in vitro Transkriptions-/Translationssystems einnehmen.
Derartige Systeme können
im Labor gemäß veröffentlichter
Techniken hergestellt werden oder können kommerziell bezogen werden.
Beispielsweise ist STP2-T7 (Katalo Nr. 69950-3) und STP-SP6 (Katalog Nr.
69997-3) von Novagen (Madison, WI) erhältlich. Promega (Madison, WI)
vertreibt ebenfalls derartige Systeme (beispielsweise Katalog Nrn.
L1170, L2080, L4600, L4610, L4130, L4140, L1130, L1020 und L1030,
genausow wie Stratagene (La Jolla, CA), das ein System mit dem Markennamen
IN VITRO EXPRESS (Katalog Nr. 200360) vermarktet. Nicht lebende
Wirtssysteme zur Verwendung in der Erfindung können ebenfalls von einem lebenden
Organismus gewonnen werden. Beispielsweise kann ein Zelllysat, wie
beispielsweise ein Reticulocytlysat in einigen Anwendungen verwendet
werden.
-
Wirtssysteme
können
ebenfalls die Form lebender Organismen einnehmen. Lebende Organismen werden
für Wirtssysteme
bevorzugt, weil diese üblicherweise
in zahlreichen Kopien reproduziert werden können, wodurch eine kontinuierliche,
leicht expandierbare und leicht manipulierbare Quelle ausgewählter Nukleinsäuremoleküle bereitgestellt
wird. Lebende Organismen, die als Wirtssysteme im Umfang der vorliegenden Erfindung
verwendet werden können,
schließen
Prokaryonten, wie beispielsweise Bakterien (beispielsweise Escherichia
coli) und Eukaryonten, wie beispielsweise Hefe- und Säugetier-
(beispielsweise Menschen, Maus, Rind, Schaf, Schwein etc.) Zellen
ein. Archaebakterien, Pflanzenzellen und andere Organismen, die
zur Verwendung mit den Verfahren der Erfindung geeignet sind, können ebenfalls
als Wirtssystem dienen.
-
Das
spezielle Wirtssystem, das für
eine spezielle Anwendung am besten geeignet ist, variiert abhängig von
den vielen unterschiedlichen Faktoren. Der Fachmann auf dem Gebiet
jedoch sollte dazu in der Lage sein, ein geeignetes Wirtssystem
für eine
spezielle Anwendung auf Grundlage bekannter Anwendungen der unterschiedlichen
Wirtssysteme auszuwählen.
Wenn beispielsweise die Produktion eines zyklischen Peptids in großem Maßstab erwünscht wird,
wäre ein
bakterieller Wirt oder ein Insektenwirt geeignet. Wo es als weiteres Beispiel
erwünscht
ist, die Interaktion humaner Zellbestandteile zu analysieren, wäre die Anwendung
einer humanen Zelle als Wirtssystem wahrscheinlich geeigneter als
die Verwendung eines bakteriellen Systems.
-
Verfahren zur Herstellung
eines Polypeptids, zyklischen Peptids oder eines Spleißzwischenproduktes
-
Die
Polypeptide der vorliegenden Erfindung können durch konventionelle Verfahren
der Herstellung von Polypeptiden einer bekannten Aminosäuresequenz
hergestellt werden. Beispielsweise können Polypeptide innerhalb
der Erfindung durch Festphasensynthese unter Verwendung kommerziell
erhältlicher
Geräte
und Reagenzien hergestellt werden. Bekannte in vitro Verfahren zur
Herstellung zyklischer Peptide können
ebenfalls dazu verwendet werden, zyklische Peptide herzustellen.
In vielen Fällen
jedoch werden die Polypeptide der vorliegenden Erfindung vorzugsweise
durch Exprimieren von Nukleinsäuremolekülen hergestellt,
die dies in einem Wirtssystem codieren. Beispielsweise können Nukleinsäuremoleküle innerhalb
der Erfindung in einen Expressionvektor eingebaut und dann in ein
Wirtssystem eingebracht werden. Das Wirtssystem kann dann unter
Bedingungen angeordnet werden, die die Expression des Vektors verursachen,
was die Bildung eines Vorläuferpeptids
und anschließend
einer zyklisierten Form des Zielpeptids oder eines Spleißzwischenprodukts, das
das Peptid zeigt, resultiert.
-
Ein
bevorzugtes Verfahren zur Herstellung eines zyklischen Peptides
oder eines Spleißzwischenproduktes
schließt
die folgenden Schritte ein: (a) Bereitstellen eines isolierten Nukleinsäuremoleküls, das
ein Polypeptid mit einem Zielpeptid zwischen dem ersten Anteil eines
Splitinteins und dem zweiten Anteil eines Splitinteins eingebracht
aufweist; (b) Bereitstellen eines Wirtssystems; (c) Einbringen des
isolierten Nukleinsäuremoleküls in das
Wirtssystem; und (d) Exprimieren des isolierten Nukleinsäuremoleküls. Die
Expression des Nukleinsäuremoleküls im Wirtssystem
erzeugt das Peptidmolekül
in Form eines Spleißzwischenproduktes
einer zyklisierten Form des Zielpeptides oder ein Polypeptid, das
spontan spleißt,
um eine zyklisierte Form des Zielpeptids zu ergeben.
-
In
bevorzugten Ausführungsformen
dieses Verfahrens findet die Produktion des Polypeptids, der zyklischen
Peptide oder der Spleißzwischenprodukte
in vivo statt (beispielsweise mit einem lebenden Wirtssystem) und
in Abwesenheit irgendeines exogen zugesetzten Mittels, wie beispielsweise
eines Mittels zum katalysieren der Zyklisierung eines Peptids (beispielsweise
einer Protease oder eines Thiols).
-
Die
Herstellung von Polypeptiden, zyklischen Peptiden oder Spleißzwischenprodukten
kann unter Verwendung von Standardtechniken zur Charakterisierung
von Proteinen überwacht
werden. Siehe beispielswiese Sambrook et al, oben. Beispielhafte
Techniken, die dazu verwendet werden können, schließen herkömmliche
Chromatographie, HPLC, FPLC und dergleichen ein. Eine Elektrophorese,
wie beispielsweise Natriumdodecylsulfat-Polyacrylamidgeleletrophorese (SDS/PAGE),
2-dimensionale Gelectrophorese, elektromagnetische strahlungsbasierte
Spektroskopie, Massenspektroskopie; Analyse von enzymatischen Verdauungsprodukten;
Thermostabilitätsassays;
etc.
-
Die Aufreinigung von Polypeptiden
zyklischen Peptiden oder Spleißzwischenprodukten
der Erfindung
-
Herkömmliche
Verfahren zur Aufreinigung von Proteinen können angepasst werden, um die
Polypeptide, zyklischen Peptide und Spleißzwischenprodukte der Erfindung
aufzureinigen. Die Erfindung schließt ebenfalls ein bevorzugtes
Verfahren zur Aufreinigung eines zyklischen Peptids aus einem Gemisch
ein. In diesem Verfahren ist ein Affinitätsmarker am zyklischen Peptid
angebracht, um dessen Aufreinigung zu unterstützen. Dieses Verfahren schließt die folgenden
Schritte ein: (a) Bereitstellen eines Gemisches, das ein zyklisches
Peptid mit einem Affnitätsmarker
konjugiert, enthält;
(b) Mischen des konjugierten zyklischen Peptids mit einem Festphasenträger, der
daran einen Liganden aufweist, der spezifisch den Affinitätsmarker
so bindet, dass der Träger
spezifisch mit dem zyklischen Peptid verbunden wird; (c) Waschen
des Trägers
zur Entfernung von nicht spezifisch gebundenem Material; und (d)
Eluieren des zyklischen Peptids von dem Träger.
-
In
diesen Verfahren kann der Affinitätsmarker irgendein Molekül sein,
dass einen Liganden an einen Festphasenträger bindet. Beispielsweise
kann der Affinitätsmarker
eine Chitinbindungsdomäne
sein, wobei der Ligand Chitin ist (siehe den Abschnitt Beispiele
unten) oder kann eine Biotinmarkierung sein, wobei der Ligand Streptavidin
ist. Viele andere Affinitätsmarker-Ligandenpaare
sind bekannt und können
in der Erfindung verwendet werden. Weil der Affinitätsmarker
spezifisch den Liganden am Festphasenträger bindet, werden die zyklischen
Peptide (mit dem gebundenen Affnitätsmarker) spezifisch den Träger binden.
Der Träger
kann dann mit einem Puffer gewaschen werden (beispielsweise salzreichen,
saurem oder alkalischen Puffer), der Material im Gemisch entfernt,
das nicht am Träger
spezifisch gebunden ist. Das affinitätsmarkierte zyklische Peptid kann
dann vom Festphasenträger
unter Verwendung eines Puffers eluiert werden, der eine Substanz
enthält, die
die Markierung vom Liganden trennt (beispielsweise einen kompetitiven
Inhibitor, wie beispielsweise überschüssigen unkonjugierten
Affinitätsmarker;
oder Denaturierungsmittel) oder ein Enzym oder chemische Reaktionspartner,
den das zyklische Peptid vom Affinitätsmarker abspaltet.
-
In
einer analogen Weise können
Spleißzwischenprodukte
eher als zyklische Peptide aufgereinigt werden. Zyklische Peptide
können
ebenfalls aus einem Gemisch unter Verwendung von Spleißzwischenprodukten aufgereingt
werden. Beispielsweise schließt
ein Verfahren zur Aufreinigung eines zyklischen Peptids aus einem
Gemisch die folgenden Schritte ein: (a) Bereitstellen eines Gemisches,
das ein Spleißzwischenprodukt konjugiert mit
einem Affinitätsmarker
enthält;
(b) Mischen des konjugierten Spleißzwischenproduktes mit einem
Festphasenträger,
der daran einen Liganden aufweist, der spezifisch den Affinitätsmarker
derart bindet, dass der Träger
spezifisch mit dem Spleißzwischenprodukt
gebunden wird; (c) Waschen des Trägers zur Entfernung von unspezifisch
gebundenem Material; (d) Zusetzen eines Reagenzes zum Träger, der
ein zyklisches Peptid aus dem Spleißzwischenprodukt herstellt;
und (e) Eluieren des zyklischen Peptids vom Träger. Bei einer Variation des
vorhergehenden werden die Schritte (d) und (e) umgekehrt, so dass
Schritt (d) das Eluieren des Spleißzwischenproduktes vom Träger und
Schritt (e) das Zusetzen eines Reagenzes zum eluierten Spleißzwischenprodukt
ist, das ein zyklisches Peptid aus dem Spleißzwischenprodukt herstellt.
Reagenzien, die zugesetzt werden können, um ein zyklisches Peptid
aus einem Spleißzwischenprodukt
herzustellen, schließen
Thiole, Proteasen und andere Substanzen ein, die die Zyklisierung
des Spleißzwischenproduktes
katalysieren können.
-
Als
spezielles Beispiel kann durch Fusionieren von IC an
einen Affinitätsmarker
und durch Entfernung des essentiellen Asparaginrestes (siehe d in 3)
ein zyklischer Ester an einer Affinitätssäule immobilisiert werden. Die
sich ergebende zyklische Peptidsäule
kann zur Affinitätsaufreinigung
des zyklischen Peptids selbst verwendet werden. Ein breiter Bereich
von proteolytischen Verfahren kann verwendet werden, um den zyklischen
Ester aus dem Affnitätsmarker
und IC abhängig von der Sequenz des zyklischen
Peptidproduktes freizusetzen.
-
Unter
Bezugnahme auf 8 ist ein Verfahren zur Aufreinigung
zyklischer Peptide dargestellt. In diesem Verfahren wird ein aktives
Inteinzwischenprodukt (Spezies 1) mutagenisiert, um das katalytische
Asparagin (Schritt A) durch eine nicht-katalytische Aminosäure (Y)
zu ersetzen, und um einen Affinitätsmarker stromabwärts von
IN (Schritt B) einzubringen, um Spezies
2 zu ergeben. Die intein-vermittelte Zyklisierungsreaktion wird
fortschreiten, bis das Lariatzwischenprodukt gebildet ist (Schritt
C). Dieses Molekül
wird dann durch eine Affinitätssäule (Schritt
D) mit einem Festphasenträger
mit einem Liganden daran passiert, der spezifisch den Affinitätsmarker
bindet und somit die Retention und Aufreinigung des IN/IC nicht-kovalenten Komplexes (Spezies 3)
ermöglicht.
Die IN/IC-Reaktion
wird dann gestört,
um ein Lariatzwischenprodukt (Spezies 4) zu ergeben, die aus der
Affinitätssäule eluiert
werden kann. Eine proteolytische oder chemische Spaltung an Aminosäure Y (Schritt
F) setzt das Lactonzwischenprodukt (Spezies 5) frei. Acyl-an-N Umordnung
(Schritt G) hat das thermodynamisch bevorzugte zyklische Amidprodukt
zur Folge (Spezies 6).
-
Unter
Bezugnahme auf 9 ist eine Variation des vorhergehenden
Verfahrens zur Aufreinigung zyklischer Peptide dargestellt. In diesem
Verfahren wird ein aktives Inteinzwischenprodukt (Spezies 1) mutagenisiert,
um das katalytische Asparagin (Schritt A) durch eine nicht-katalytische
Aminosäure
(Y) zu ersetzen und einen Affinitätsmarker stromaufwärts von
IC (Schritt B) einzubringen, um Spezies
2 zu ergeben. Die inteinvermittelte Zyklisierungsreaktion wird fortschreiten,
bis sich ein Lariatzwischenprodukt gebildet hat (Schritt C). Dieses
Molekül
wird dann durch eine Affinitätssäule (Schritt
D) passiert, die einen Festphasenträger mit einem Liganden daran
aufweist, der spezifisch das Affinitätsmarker/IC-Zwischenprodukt
bindet (Spezies 3). Die Auftrennung des Affinitätsmarkers (Schritt E) aus dem
Liganden (beispielsweise unter Verwendung eines Moleküls, das
kompetitiv die Marker-Liganden-Wechselwirkung hemmt, unter Verwendung
eines salzreichen Puffers oder Denaturierungsmittels oder unter
Verwendung eines chemischen Mittels oder einer Protease zur Spaltung
des Markers) ermöglicht
die Widergewinnung des Lariatzwischenproduktes (Spezies 4). Eine
proteolytische oder chemische Spaltung an Aminosäure Y (Schritt F) setzt das
Lactonzwischenprodukt (Spezies 5) frei. Eine Acyl-an-N Umordnung
(Schritt G) ergibt das thermodynamisch bevorzugte zyklische Amidprodukt (Spezies
6).
-
Verfahren
zur Herstellung einer Bibliothek zyklischer Peptide und Spleißzwischenprodukte
-
Zahlreiche
Verfahren zur Herstellung linearer Peptidbibliotheken sind in der
Technik bekannt.
-
Modifikationen
solcher bekannter Verfahren können
mit den Verfahren zur Herstellung zyklischer Peptide und Spleißzwischenprodukte,
die hierin gelehrt werden, verwendet werden, um Bibliotheken zyklischer Peptide
und Spleißzwischenprodukte
zu erzeugen. Im allgemeinen schließt ein Verfahren zur Herstellung
einer Bibliothek zyklischer Peptide und/oder von Spleißzwischenprodukten
die folgenden Schritte ein: (a) Bereitstellen einer Vielzahl von
Nukleinsäuremolekülen, die
eine Vielzahl von Zielpeptiden mit heterogenen Aminosäuresequenzen
codieren; (b) Einbauen jeweils der Vielzahl von Nukleinsäuremolekülen in einen
Expressionsvektor zur Bildung einer Vielzahl von Expressionsvektoren,
wobei jeder der Vielzahl von Nukleinsäuremolekülen zwischen einem Nukleinsäuremolekül, das einen
ersten Anteil eines Splitinteins codiert und einem Nukleinsäuremolekül, das einen
zweiten Anteil eines Splitinteins codiert, angeordnet wird, in jedem
der gebildeten Expressionsvektoren, so dass die Expression der Expressionsvektoren
in einem Wirtssystem die Erzeugung einer Vielzahl von Spleißzwischenprodukten
zyklisierter Formen der Zielpeptide oder Polypeptide zur Folge hat,
die spontan im Wirtssystem spleißen, so dass sich zyklisierte
Formen der Zielpeptide ergeben; und (c) Expremieren der Expressionsvektoren
im Wirtssystem.
-
Als
spezifischere Beispiele können
die in Childs et al, in Sequence Specificity in Transcription and Translation
(Alan R. Liss, Inc., 1985) beschriebenen Verfahren und das Doppelstrangligationsverfahren
beschrieben in Schumacher et al, Science 271:1854, 1996, zur Verwendung
in der vorliegenden Erfindung modifiziert werden. Bekannte PCR-basierte Verfahren
können
ebenfalls dazu verwendet werden, Polynukleotide zu erzeugen, die
Peptide mit zufallsbedingten Sequenzen codieren, die zirkularisiert
oder als Spleißzwischenprodukt
in der Erfindung expremiert werden können. Siehe beispielsweise
Caldwell und dJoyce, PCR Methods Appl. 2:22, 1992; Ostermeier et
al, Proc. Natl. Acad. Sci. USA 96:3562, 1999; Nested Deletion Protocol
and Reagents from Promega; und Stemmer, W.P. Nature 370:389, 1994
(DNA shuffling). Die Vielzahl von Polynukleotiden, die Peptide mit
heterogenen Sequenzen codieren, können das Zielpeptid (oder das
in einem Spleißzwischenprodukt
darzustellende Peptid) in die Nukleinsäuremoleküle und Expressionsvektoren
der Erfindung wie oben beschrieben eingebaut werden und dann in
einem Wirtssystem zur Herstellung einer Bibliothek zyklischer Peptide
oder Spleißzwischenprodukte
expremiert werden.
-
Verfahren zum Screenin
eines zyklischen Peptides nach vorherbestimmten Eigenschaften
-
Es
existieren zahllose Techniken zum Screening von kleinen Molekülen bzw.
small molecules nach speziellen Eigenschaften. Siehe beispielsweise
Fernades, P., Current Opin. Chem. Biol. 2:597, 1998, Science 286:1759,
1999; U.S. Patente Nrn. 5,585,277 und 5,989,814. Insbesondere sind
ebenfalls Verfahren zur Bestimmung bekannt, welches Peptid in einer
kombinatorischen Peptidbibliothek spezifisch an ein Zielprotein
bindet.
-
Beispielsweise
U.S. Patent Nr. 5,834,318. Viele dieser Verfahren können adaptiert
werden, um zyklische Peptide und/oder Spleißzwischenprodukte, die mit
den Verfahren der Erfindung hergestellt wurden, nach speziellen
Eigenschaften zu screenen.
-
Ein
allgemeines Verfahren zum Screening eines Peptidmoleküls nach
einer vorher bestimmten Eigenschaft schließt die folgenden Schritte ein:
(a) Bereitstellen eines Nukleinsäuremoleküls, das
ein Polypeptid mit einem Zielpeptid zwischen einem ersten Anteil
eines Splitinteins und einem zweiten Anteil eines Splitinteins eingebracht
aufweist, codiert, derart, dass die Expression des Nukleinsäuremoleküls in einem
Wirtssystem das Peptidmolekül
entweder in zyklisierter Form des Zielpeptids (als Ergebnis eines
spontanten Spleißens
des Polypeptids im Wirtssystem) oder ein Spleißzwischenprodukt in einer zyklisierten
Form des Zielpeptids erzeugt; (b) Bereitstellen des Wirtssystems;
(c) Einbringen des isolierten Nukleinsäuremoleküls ins Wirtssystem; (d) Anordnen
des Wirtssystems unter Bedingungen, die die Produktion des Peptidmoleküls verursachen;
und (e) Testen des Peptidmoleküls
bezüglich
der vorher bestimmten Eigenschaft.
-
Schritt
(a) kann wie anderswo beschrieben durchgeführt werden, beispielsweise
unter Verwendung von molekularbiologischen Techniken (siehe Ausubel
et al und Sambrook et al, siehe oben), um Polynukleotide zu erzeugen,
die das Zielpeptid, den ersten Anteil eines Inteins und den zweiten
Anteil eines Inteins codieren. Die sich ergebenden drei Polynukleotide
können
dann fusioniert werden (beispielsweise ligiert), um ein Nukleinsäuremolekül zu bilden.
Das bereitgestellte Wirtssystem kann irgendeines von denen sein,
die hierin beschrieben wurden, bei denen das Nukleinsäuremoleküls (beispielsweise
Bakterien, Hefe, Säugetierzelle)
exprimiert werden kann. Die Nukleinsäure kann in ein Wirtssystem
durch bekannte Verfahren abhängig
von der Form des Nukleinsäuremoleküls und des
verwendeten Wirtssystems eingebracht werden. Beispielsweise können die
Nukleinsäuremoleküle in eine
Zelle durch Elektroporation, Lipofection, unter Verwendung einer
kalziumchloridvermittelten Transformation, unter Verwendung einer
Gen"kanone" unter Verwendung
eines Bacteriophagenvektors (wenn das Wirtssystem ein Bakterium
ist) unter Verwendung eines Plasmidkonstruktes, unter Verwendung
eines viralen Vektors etc. eingebracht werden.
-
Das
Wirtssystem kann unter Bedingungen angeordnet werden, die verursachen,
dass das Peptidmolekül
durch Einstellen der Bedingungen gemäß der speziellen Form des Nukleinsäuremoleküls und des
verwendeten Wirtssystems erzeugt wird. Für ein humanes Zell-Wirtssystem
kann dies das Anordnen der Zelle in einem geeigneten nährstoffreichen
Medium und das Kultivieren der Zelle in einem 37°C befeuchteten 5-10% CO2 Inkubator bedeuten. Für induzierbare Expressionsvektoren
kann dies den Zusatz der Substanz zum Wirtssystem bedeuten, das
die Expression des Nukleinsäuremoleküls im Vektor
induziert. Beispielsweise kann dieser Schritt, wenn ein arabinoseinduzierbarer
Expressionsvektor verwendet wird, den Zusatz von Arabinose zum Wirtssystem
einschließen.
-
Das
Testen des Peptidmoleküls
bezüglich
vorherbestimmter Eigenschaften kann durch eine große Vielzahl
unterschiedlicher Verfahren durchgeführt werden, beispielsweise
durch Messen der Bindung des Peptidmoleküls an einen bekannten Liganden
und Analysieren der Fähigkeit
des Peptidmoleküls
zu modulieren (d. h. Erhöhen
oder Senken der Geschwindigkeit der (biochemischen) Reaktion. Bezüglich einer
Beschreibung verschiedener Verfahren, die zum Testen von Peptiden
nach vorherbestimmten Eigenschaften verwendet werden können, siehe
Fernades, P., siehe oben.
-
Speziellere
beispielhafte Verfahren, die dazu verwendet werden können, zyklische
Peptide oder Spleißzwischenprodukte
nach speziellen Eigenschaften zu screenen, schließen die
Verwendung eines Festphasenträgers
und einer Affinitätschromatographie
zum Identifizieren von Molekülen
ein, die spezifisch zyklische Peptide oder Spleißzwischenprodukte binden; die
Verwendung der Phage Display Technologie; und die Verwendung von
Aptamerpeptidfusionskonstrukten und/oder Hybridsystemen zum Identifizieren
zyklischer Peptide oder von Spleißzwischenprodukten, die eine
spezifische biochemische Reaktion oder ein intrazelluläres Ereignis
modulieren können.
-
Festphasenträger/Affinitätschromatographie
zum Identifizieren von Molekülen
die mit zyklischen Peptiden und/oder Spleißzwischenprodukten interagieren
-
Zyklische
Peptide oder Spleißzwischenprodukte
können
an einem Festphasenträger
immobilisiert werden, um die Identifizierung und/oder Aufreinigung
von Molekülen
zu erleichtern, die spezifisch eingegebenes zyklisches Peptid oder
Spleißzwischenprodukt
binden. Bezüglich
eines allgemeinen Überblicks
von Peptidaffinitätssäulen zur
Aufreinigung siehe Bumbach, G. A. und D. J. Hammon, Biopharm, 5:24,
1992. Unten sind Beispiele angegeben, wie dies im Rahmen der Erfindung
durchgeführt
werden kann.
-
Nunmehr
bezugnehmend auf 10 ist ein Verfahren zum Identifizieren/Aufreinigen
von Molekülen, die
spezifisch ein gegebenes Spleißzwischenprodukt
binden, dargestellt. In diesem Verfahren wird ein aktives Inteinintermediat
bzw. Zwischenprodukt (Spezies 1) mutagenisiert, so dass das katalytische
Asparagin (Schritt A) durch eine nichtkatalytische Aminosäure (Y)
ersetzt wird und einen Affinitätsmarker
stromaufwärts
von IC (Schritt B) einzubringen, um Spezies
2 zu erzeugen. Die Intein-vermittelte Zyklisierungsreaktion wird
fortschreiten, bis ein Lariatzwischenprodukt gebildet ist (Schritt
C). Dieses Molekül
wird dann durch eine Affinitätssäule (Schritt
D) mit einem Festphasenträger
mit einem Liganden daran passiert, der spezifisch den Affinitätsmarker/IC-Zwischenprodukt (Spezies 3) bindet. Eine
Lösung,
die Zielmoleküle
enthält
(d. h. Kandidaten zur Bindung des Spleißzwischenproduktes) wird dann
durch die Säule
passiert (Schritt E). Zielmoleküle,
die spezifisch das Spleißzwischenprodukt
binden, werden selektiv innerhalb der Säule zurückgehalten. Diese Zielmoleküle können aus
der Säule
entfernt und biochemisch analysiert (beispielsweise sequenziert)
werden.
-
Nunmehr
unter Bezugnahme auf 11 ist ein weiteres Verfahren
zum Identifizieren/Aufreinigen von Molekülen, die spezifisch ein gegebenes
Spleißzwischenprodukt
binden, dargestellt. In diesem Verfahren wird ein aktives Inteinzwischenprodukt
(Spezies 1) mutagenisiert, so dass das katalytische Asparagin (Schritt
A) durch eine nichtkatalytische Aminosäure (Y) ersetzt wird und um
ein Affinitätsmarker
stromabwärts
von IN (Schritt B) einzubringen, um Spezies
2 zu ergeben. Die inteinvermittelte Zyklisierungsreaktion wird fortschreiten,
bis sich ein Lariatzwischenprodukt gebildet hat (Schritt C). Dieses
Molekül
wird dann durch eine Affinitätssäule (Schritt
D) passiert, die einen Festphasenträger mit einem Liganden daran
aufweist, der spezifisch den Affinitätsmarker bindet, um die Zurückhaltung
bzw. Retention und Aufreinigung des IN/IC nicht-kovalenten Komplexes (Spezies 3)
zu ermöglichen.
Die Spaltung des Affinitätsmarkers
(Schritt E) ermöglicht
die Wiedergewinnung des Lariatzwischenproduktes (Spezies 4). Eine
Lösung,
die Zielmoleküle
enthält
(d. h. Kandidaten zur Bindung der Spleißzwischenprodukte) wird dann
durch die Säule
passiert (Schritt E). Zielmoleküle,
die spezifisch das Spleißzwischenprodukt
binden, werden selektiv innerhalb der Säule zurückgehalten.
-
Nunmehr
bezugnehmend auf 12 ist ein weiteres Verfahren
zum Identifizieren/Aufreinigen von Molekülen dargestellt, die spezifisch
ein vorgegebenes Spleißzwischenprodukt
binden. In diesem Verfahren wird ein aktives Inteinzwischenprodukt
(Spezies 1) mutagenisiert, um das IN Nucleophil
(Schritt A) durch eine nicht-katalytische Aminosäure (Z) zu ersetzen und einen
Affinitätsmarker
stromabwärts
von IN (Schritt B) einzubringen, um ein
IC-Peptid-IN-Markierungsfusionsprotein
(Spezies 2) herzustellen. Die Intein-vermittelte Zyklisierungsreaktion
erzeugt das Fusionsprotein (Schritt C). Dieses Protein wird dann
durch eine Affinitätssäule (Schritt
D) passiert, die einen Festphasenträger aufweist mit einem Liganden
daran, der spezifisch den Affinitätsmarker bindet, um die Retention
und Aufreinigung des Proteinkomplexes zu ermöglichen (Spezies 3). Eine Lösung, die
Zielmoleküle
enthält
(d. h. Kandidaten zur Bindung der Spleißzwischenprodukte) wird dann
durch die Säule
(Schritt E) passiert. Zielmoleküle,
die spezifisch das Spleißzwischenprodukt
binden, werden selektiv innerhalb der Säule zurückgehalten.
-
Nunmehr
bezugnehmend auf 13 ist ein noch weiteres Verfahren
zum Identifizieren/Aufreinigen von Molekülen dargestellt, die spezifisch
eingegebenes Spleißzwischenprodukt
binden. In diesem Verfahren wird ein aktives Inteinzwischenprodukt
(Spezies 1) mutagenisiert, so dass das IN-Nucleophil
(Schritt A) durch eine nicht-katalytische Aminosäure (Z) ersetzt wird und ein
Affinitätsmarker
stromaufwärts
von IC (Schritt B) eingebracht wird, um
ein Markierungs-IC-Peptid-IN-Fusionsprotein (Spezies
2) zu ergeben. Die inteinvermittelte Zyklisierungsreaktion erzeugt
das Fusionsprotein (Schritt C). Dieses Protein wird dann durch eine
Affinitätssäule (Schritt
D) passiert, die einen Festphasenträger mit einem Liganden daran
aufweist, der spezifisch den Affinitätsmarker bindet und so die
Retention und Aufreinigung des Proteinkomplexes ermöglicht (Spezies
3). Eine Lösung,
die Zielmoleküle
enthält
(d. h. Kandidaten zur Bindung der Spleißzwischenprodukte) wird dann durch
die Säule
passiert (Schritt E). Zielmoleküle,
die spezifisch das Spleißzwischenprodukt
binden, werden selektiv innerhalb der Säule zurückgehalten.
-
Phage Display
-
Verfahren
zum Screening von Molekülen
unter Verwendung von Phage Display liegen ebenfalls innerhalb der
vorliegenden Erfindung. Konventionelle Verfahren unter Verwendung
von Phage Display können
modifiziert werden, indem der Phage verwendet wird, um die zyklischen
Peptide und/oder Spleißzwischenprodukte
der Erfindung darzubieten. Wenn beispielsweise Z in 1 ein
Phagenhüllprotein
ist, und XH=H in 2 ist, wird die Spleißreaktion
nicht über
das erste Esterzwischenprodukt hinaus fortschreiten, wodurch sich
ergeben wird, dass das Zielprotein als Schleife dargeboten wird.
Auf diese Weise können
Bibliotheken, die Phagenteilchen umfassen, die Schleifenzielpeptide
zeigen, hergestellt und dazu verwendet werden, Moleküle zu isolieren,
die die dargestellte Schleife binden. Beispielsweise kann ein Zielmolekül an einem
Festphasenträger immobilisiert
werden. Phagenbibliotheken, die unterschiedliche Schleifen-Peptide
zeigen, können
dann mit dem Träger
vermischt werden. Solche Phagen, die Schleifen-Peptide zeigen, die
das Zielmolekül
binden, würden
am Träger
selektiv zurückgehalten.
Nach Elution vom Träger
(beispielsweise durch Spaltung der Esterbindung der durch den Phagen
dargebotenen Schleifen bzw. Loop-Peptide mit hohen Konzentrationen
eines wirkungsvollen Nucleophils) können die Aminosäuresequenzen
der Loop-Peptide durch standardmolekularbiologische Verfahren bestimmt
werden.
-
Aptamere
-
Peptidaptamere
sind Polypeptide, die eine konformationell eingeengte Zielpeptidregion
einer variablen Sequenz enthalten, die von einem Gerüst dargeboten
wird. Weil zyklische Peptide oder Spleißzwischenprodukte als Aptamere
dienen können,
können
bekannte Verfahren zum Analysieren von Aptameren modifiziert werden,
um bei der Identifizierung spezieller Eigenschaften der zyklischen
Peptide oder Spleißzwischenprodukte
zu assistieren. Siehe beispielsweise Geyer et al, Proc. Natl. Acad.
Sci. USA 96:8567, 1999; Caponigro et al, Proc. Natl. Acad. Sci.
USA 95:7508, 1998: Mikhail et al, Proc. Natl. Acad. Sci. USA 95:14266,
1998; Norman et al, Science 285:591 1999.
-
Beispielsweise,
bezugnehmend auf 14 können zyklische Proteine als
Apatamergerüste
in einer Technik verwendet werden, die es Elementen einer Peptidbibliothek
erlaubt, als eingeengte Schleifen bzw. Schlaufen zwischen dem N-Terminus
und C-Terminus des zyklischen Proteingerüstes dargeboten zu werden. Wie
in 14 dargestellt, kann eine Aptamerbibliothek als
ein IC-Gerüst-IN Fusionsprotein
(Spezies 1) exprimiert werden. Die Prozessierung der intein-vermittelten
Zyklisierungsreaktion in vivo (Schritt A) ergibt ein IN, ein
IC und ein zyklisches Gerüstprotein
(Spezies 2). Die Aptamerbibliothek wird in der Linkerregion zwischen dem
N-Terminus und dem C-Terminus dargeboten. In 14 repräsentiert
N irgendeine Aminosäure
und das tiefgestellte n und m sind ganze Zahlen gleich oder größer als
0, und X repräsentiert
Serin, Threonin oder Cystein.
-
Andere
Beispiele der Verwendung von Aptameren liegen ebenfalls innerhalb
der vorliegenden Erfindung. Unter Bezugnahme auf 15 sind
zwei derartige Verfahren beschrieben. In Reaktion I wird ein aktives Inteinzwischenprodukt
mutagenisiert, um die nukleophile Aminosäure von IN (Schritt
A) mit einer nicht-katalytischen Aminosäure (Z) zu ersetzen. Diese
Prozesse inaktivieren die Spleißreaktion,
um Spezies 2 zu ergeben. Wegen der starken Wechselwirkung zwischen
IC und IN ermöglicht diese
Technik den Elementen eine Peptidbibliothek als eingeengte Schlaufe
in der Linkerregion zwischen den beiden Inteinproteinen (Target)
dargeboten zu werden. In Reaktion II wird ein aktives Inteinintermediat
mutagenisiert, um das katalytische Asparagin (Schritt A) durch eine
nichtkatalytische Aminosäure
(Y) zu ersetzen, so dass sich Spezies 2 ergibt. Das Fortschreiten
der intein-vermittelten Zyklisierungsreaktion schreitet in vivo
(Schritt B) fort und stoppt auf der Stufe des Lariatzwischenproduktes
(Spezies 3) und ermöglicht
den Elementen einer Peptidbibliothek als eingeengtes Lacton dargeboten
zu werden.
-
Hybridsysteme
-
Das
Hefe Zwei-Hybridsystem ist ein wohl untersuchtes Verfahren zur Analysierung
von in vivo Protein-Proteinwechselwirkungen. Fields, S. und O. Song,
Nature (London) 340:245, 1989. Dieses und Variationen hiervon, wie
beispielsweise Ein-Hybridsysteme, Drei-Hybridsysteme, reverse Zwei-Hybridsysteme,
Split-Hybridsysteme, alternative N-Hybridsysteme, Small Molecule basierte
Hybridsysteme können
dazu verwendet werden, die Eigenschaften zyklischer Peptide zu untersuchen
und/oder von Spleißzwischenprodukten
zu untersuchen, in dem bekannte Verfahren angepasst werden. Siehe
beispielsweise Drees, B. L., Current Opin. Chem. Biol., 3:64, 1999;
Vidal, M., and P. Legrain, Nucleic Acids Research, 27:919, 1999;
Current Protocols in Molecular Biology, Ausg., Ausubel, F. m., et
al, Wiley, New York, 1996; Huang, j. und S.L. Schreiber, Proc. Natl. Acad.
Sci. (USA) 94:13396, 1997; Yang, M., et al, Nucleic Acids Res. 23:
1152, 1995; Colas, P., et al, Nature (London), 380:548, 1996; Xu,
C. W., et al, Proc. Natl. Acad. Sci. (USA) 94:12473, 1997.
-
Beispielsweise
liegt ein Verfahren zum Identifizieren eines Zielproteins, unter
Bezugnahme auf 16, das mit einem Spleißzwischenprodukt
interagiert, im Rahmen der vorliegenden Erfindung. In diesem Verfahren
wird ein aktives Inteinzwischenprodukt (Spezies 1) mutagenisiert,
um das katalytische Asparagin (Schritt A) durch eine nichtkatalytische
Aminosäure
(Y) zu ersetzen und eine DNA-Bindungsdomäne stromabwärts von IN (Schritt
B) einzubringen, um Spezies 2 zu ergeben. Die inteinvermittelte
Zyklisierungsreaktion wird fortschreiten, bis das Lariatzwischenprodukt
(Spezies 3) gebildet ist (Schritt C). IN und
IC bilden einen starken nicht kovalenten
Komplex. Das sich ergebende Lariatzwischenprodukt wird dann mit
einem Zielprotein koexprimiert, das an eine DNA Bindungsdomäne gebunden
ist (Schritt D). Eine Interaktion des Lariatzwischenproduktes mit
dem Zielprotein (Spezies 4) verursacht eine Aktivierung einer Promotorregion
(Schritt E), was zur Expression des Reportergens (*) führt. Dieses
Verfahren ermöglicht
die Identifizierung von Zielmolekülen, die dazu in der Lage sind,
das Lariatzwischenprodukt zu binden. Dieses Verfahren kann derart
modifiziert werden, dass ein bekanntes Molekül (anstelle eines unbekannten
Zielproteins) an eine DNA Bindungsdomäne gebunden wird, so dass Lariatzwischenprodukte
identifiziert werden können,
die ein Loop-Peptid darbieten, das das bekannte Molekül bindet.
Unter Bezugnahme auf 14 ist ein weiteres Verfahren
zum Identifizieren eines Zielproteins beschrieben, das mit einem
Spleißzwischenprodukt
wechselwirkt. In diesem Verfahren wird ein aktives Inteinzwischenprodukt
(Spezies 1) mutagenisiert, um das katalytische Asparagin (Schritt
A) durch eine nichtkatalytische Aminosäure (Y) zu ersetzen und eine
DNA Bindungsdomäne
stromaufwärts
von IC (Schritt B) einzubringen, um Spezies
2 zu ergeben. Die inteinvermittelte Zyklisierungsreaktion wird fortschreiten,
bis das Lariatzwischenprodukt (Spezies 3) gebildet ist (Schritt
C). Dieses Molekül
wird dann mit einem Zielprotein koexprimiert, das an eine DNA-Bindungsdomäne (Schritt
D) gebunden ist. Eine Interaktion des Lariatzwischenproduktes mit
dem Zielprotein (Spezies 4) verursacht eine Aktivierung einer Promotorregion
(Schritt E), was zur Expression des Reportergens (*) führt. Dieses
Verfahren ermöglicht
die Identifizierung eines Zielmoleküls, das dazu in der Lage ist,
das Lariatzwischenprodukt zu binden. Dieses Verfahren kann derart
modifiziert werden, dass ein bekanntes Molekül (anstelle eines unbekannten
Zielproteins) an eine DNA-Bindungsdomäne gebunden wird, so dass Lariatzwischenprodukte
identifiziert werden können,
die ein Loop-Peptid zeigen, das das bekannte Molekül bindet.
-
Unter
Bezugnahme auf 18 ist ein weiteres Verfahren
zum Identifizieren eines Zielproteins beschrieben, das mit einem
Spleißzwischenprodukt
wechselwirkt. In diesem Verfahren wird ein aktives Inteinzwischenprodukt
(Spezies 1) mutagenisiert, so dass das IN-Nukleophil
(Schritt A) durch eine nicht-katalytische Aminosäure (Z) ersetzt wird und das
eine DNA-Bindungsdomäne
(DBD) stromabwärts
von IN eingebracht wird (Schritt B). Diese
Verfahren werden die Spleißreaktion
inaktivieren und werden ein IC-Peptid-IN-DBD Fusionsprotein (Spezies 2) erzeugen.
Dieses Molekül
wird dann mit einem Zielprotein koexprimiert werden, das an eine
DNA-Bindungsdomäne
gebunden ist (Schritt C). Die Interaktion des Fusionsproteins mit
dem Zielprotein (Spezies 3) verursacht eine Aktivierung einer Promotorregion
(Schritt D), was zur Expression des Reportergens (*) führt. Dieses
Verfahren ermöglicht
die Identifizierung von Zielmolekülen, die dazu in der Lage sind, das
Fusionsprotein zu binden. Dieses Verfahren kann derart modifiziert
werden, das ein bekanntes Molekül (anstelle
eines unbekannten Zielproteins) an eine DNA-Bindungsdomäne so gebunden ist, dass Fusionsproteine
identifiziert werden können,
die ein Loop-Peptid darbieten, das das bekannte Molekül bindet.
-
Unter
Bezugnahme auf 19 wird ein noch weiteres Verfahren
zum Identifizieren eines Zielproteins beschrieben, das mit einem
Spleißzwischenprodukt
interagiert. Bei diesem Verfahren wird ein aktives Inteinzwischenprodukt
(Spezies 1) mutagenisiert, so dass das IN-Nukleophil
(Schritt A) durch eine nicht-katalytische Aminosäure (Z) mutagenisiert wird
und so dass eine DNA-Bindungsdomäne
stromaufwärts
von IC (Schritt B) eingebracht wird. Diese
Verfahren werden die Spleißreaktion
inaktivieren und werden ein DBD-IC-Peptid-IN-Fusionsprotein
erzeugen (Spezies 2). Dieses Molekül wird dann mit einem Zielprotein
koexprimiert, das an eine DNA-Bindungsdomäne gebunden ist (Schritt C).
Eine Interaktion des Fusionsproteins mit dem Zielprotein (Spezies
3) in Schritt D verursacht eine Aktivierung einer Promotorregion
(Schritt E), was zur Expression des Reportergens (*) führt. Dieses
Verfahren ermöglicht
die Identifizierung von Zielmolekülen, die dazu in der Lage sind,
das Fusionsprotein zu binden. Dieses Verfahren kann derart modifiziert
werden, dass ein bekanntes Molekül
(anstelle eines unbekannten Zielproteins) an eine DNA-Bindungsdomäne so gebunden
wird, dass Fusionsproteine identifiziert werden können, die
ein Loop-Peptid darbieten, das das bekannte Molekül bindet.
-
Targeting zyklischer Peptide
und Spleißzwischenrpodukte
In Vivo
-
Die
zyklischen Peptide und Spleißzwischenprodukte
im Umfang der Erfindung können
speziell auf besondere Zellorte oder auf extrazelluläre Sekretion
ausgerichtet werden, indem Modifikationen von Targetingverfahren
angewendet werden, die in der Technik bekannt sind. Siehe beispielsweise
Wilkinson et al, J. Membrane Biol., 155:189, 1997; Komiya et al,
The EMBO J., 17:3886, 1998; Kouranov and Schnell, J. Biol. Chem. 271:31009,
1996; Bhagwat et al, J. Biol. Chem, 274:24014, 1999; Adam, S.A.,
Current Opin. Cell Biol. 11:402-406, 1999; Gorlich, D., Current
Opin. Cell Biol. 9:412, 1997; Pemberton et al, Current Opin. Cell
Biol. 10:292, 1998; Sakaguchi, M., Current Opin. Cell Biol. 8:595,
1997; Folsch et al., The EMBO J. 17:6508, 1998. Beispielsweise können verschiedene
Signalpeptide an die zyklischen Peptide oder Spleißzwischenprodukte der
Erfindung gebunden werden, um diese an vorher bestimmte Zellkompartimente
zu lokalisieren oder dass diese in den extrazellulären Raum
nach der Translation abgesondert werden. In dieser Weise können die
zyklischen Peptide oder Spleißzwischenprodukte
der Erfindung so auf zelluläre
Orte, wie beispielsweise Mitochondrien, Lysosomen, endoplasmatische
Reticula, Chloroplasten, Golgi, Periplasma, Kern, Plasmamembran, ausgerichtet
werden. Dieses Verfahren zum Targeting kann ebenfalls in den Verfahren
zur Erzeugung von Peptidbibliotheken und in den Verfahren zum Screening
derartiger Bibliotheken verwendet werden, wo es wünschenswert
ist, Moleküle
zu identifizieren, die mit einer Aktivität an vorherbestimmten Zellorten
interagieren oder diese zeigen.
-
Beispiele
-
Herstellung zyklischer
Dihydrofolatreductase (DHFR und zyklischem Pseudostellarin F Materialien und Methoden
-
Vectorkonstruktion
-
Das
Gen für
das Ssp DnaE N-Intein (IN) wurde aus einer
Ssp 6803 genomischen DNA mit Taq-Polymerase und Primern amplifiziert,
die 5'-BglII und
NsiI und 3'-PstI
Restriktionsstellen einbrachten. Das Ssp DnaE IC-Gen
wurde in ähnlicher
Weise mit Primern amplifiziert, so dass 5'-NcI und 3'-NdeI und SacI Restriktionsstellen eingebracht
wurden. Plasmid pDIMCP ergab sich aus dem individuellen Klonieren
der Inteinfragmente in pDIMC7 (identisch zu pDIMC6 (siehe Ostermeier
et al., Proc. Natl. Acad. Sci. USA 96:3562, 1999) außer der
Konversion der BamHI Restriktionsstelle in BgIII). Eine Alinin zu
Histidinmutation im IC-Gen (A35H) wurde
eine Quick-Change Mutagenese (Stratagene) bewirkt, die pDIMCPAH
zur Folge hatte. Ein Ausschneiden der Inteinfragmente als ein NcoI/PstI
Digest und eine Ligation in pAR4 (abgeleitet von pAR3 (Perez-Perez, J.
und J. Gutierrez, Gene, 158:141, 1995; American Type Culture Collection
(ATCC), 87026) mit einer einmal vorkommenden NcoI Stelle in der
multiplen Klonierungsstelle) erzeugte pARCP (a in 3)
und pARCPAH. E. coli DHFR wurde aus pET22b-DHFR (Miller, G.P. und
S. J. Benkovic, Biochemistry 37:6327, 1998) mit Primern amplifiziert,
die einen 5'-NdelI
Ort einbrachten, gefolgt von (CAC)6 (das
sechs Histidinreste codierte) und einer 3'-PstI Stelle, digeriert mit NdeI/PstI
und ligiert in NdeI/NsiI, digeriertes pARCP oder pARCPAH, um pARCP-DHFR
(b in 3) und pARCPAH-DHFR (c in 3) zu erzeugen.
Eine Polyhistidinsequenz wurde synthetisch mit NdeI, NsiI und BspMI
Stellen hergestellt und in pARCPAH ligiert, um das Plasmid pARCP2-6H
zu erzeugen, das Cyclo-[CHMHHHHHHGAGAA] codiert. Plasmid-pARCP-p wurde in drei Schritte
aus pDIMCPAH erzeugt: (1) Quick-Change Mutagenese brachte eine AfIII
Stelle in IN ein, und erzeugte pDIMCPMA;
(2) das Pseudostellarin F Gen wurde synthetisch hergestellt und
in MfeI/AfIII digeriertes pDIMCPMA eingebracht, um pDIMCP-p zu erzeugen;
und (3) das Fusionskonstrukt wurde aus pDIMCP-p als ein NcoI/PstI
Fragement ausgeschnitten und in NcoI/PstI digeriertes pAR4 ligiert,
um pARCP-p (e in 3)
zu erzeugen. Um das Plasmid pARCBD-p zu erzeugen, wurde eineKpnI
Stelle am Carboxyterminus des IN-Gens von
pARCP-p durch Quick-Change Mutagenese eingebracht, um pARCPpK zu
erzeugen. Das Gen, das die Chitinbindungsdomäne codierte, wurde aus Plasmid
pCYBI (New England Biolabs, Inc., Beverly, Massachusetts) mit Primern
amplifiziert, die eine 5'KpnI
Stelle und eine 3'HindII
Stelle einbrachten. Sowohl das PCR Produkt, als auch pARCPpK wurden
mit KpnI und HindIII digeriert und zusammen ligiert, um pARCBD-p
(f in 3) zu erzeugen. Alle Enzyme waren von Promega
oder New England Biolabs sowie nichts anderes erwähnt ist.
-
DHFR Aufreinigung
-
XL-1
Blue Zellen, die entweder pARCP-DHFR oder pARCPAH-DFHR beherbergten,
wurden in LB Medium plus 50 μg/ml
Chloramhenicol bei 37°C
gezüchtet,
bis die Kultur eine OD600 von 0,7 erreichte.
Die Kultur wurde mit L-(+) Arabinose bis zu einer Endkonzentration
von 0,5% induziert, und bei 28°C
für 24
Stunden gezüchtet.
Die Zellen wurden durch Zentrifugation (7000 × g, 10 Minuten) geerntet und
im flüssigen
Stickstoff tiefgefroren. Die Zellen wurden lysiert und die DHFR
enthaltenden Proteine wurden wie beschrieben aufgereinigt (Miller
und Benkovic, id). Das zyklische Produkt wurde von anderen DHFR-enthaltenden
Intermediaten durch FLPC unter Verwendung einer Mono-Q Säule (Amersham
Pharmacia) aufgetrennt, eluiert mit einem Gradienten von 0-1 M NaCL
in 50 mM Tris-HCl über
30 Minuten. Ein Westernblotting wurde mit Anti-His (Qiagen) durchgeführt und
mit Ziegen anti-maus-alkalische phosphatase-konkugierten Antikörpern (Pierce)
gemäß den Anweisungen
des Herstellers.
-
Endoproteinase Lys-C Digestion
-
Wildtyp
oder zyklisches DHFR (50 μg)
wurde mit 0,5 μg
Endoproteinase Lys-C in 0,5 M NH4HCO3 bei 37°C
behandelt. Die Proben wurden bei 6 und 24 Stunden entnommen, auf
einem SDS/16% PAGE Gel visualisiert und einer matrixunterstützten, Laserdisorptionsionisierung
(MALDI) time-of-flight mass Spektrometrie (Moore, W.T., Methods
Enzymol, 289:520, 1997) unterworfen.
-
DHFR Assays
-
Die
Thermostabilität
wurde durch Präinkubation
von 100 nM Wildtyp oder zyklischem DHFR bei entweder 25°C oder 65°C in MTEN
Puffer (50 mM 2-(N-Morpholino)ethansulfonsäure (MES),
25 mm Tris(hydroxymethyl)aminomethan (Tris), 25 mM Ethanolamin und
l00 mM NaCl) untersucht. Teilmengen wurden bei verschiedenen Zeitpunkten
entnommen und bei Raumtemperatur für fünf Minuten in Gegenwart von
100 μm 7,8-Diyhdrofolat
equilibriert. Die Aktivitätsassays
wurden mit einem reduzierten Nikotinamid Adenindinukleotid (NADHPH),
wie kürzlich
beschrieben, begonnen (Miller und Benkovic, siehe oben).
-
Synthese von Cyclo-(Ser-Gly-Gly-Tyr-Leu-Pro-Pro-Leu)
-
Einer
Lösung
von 3,5 mg (4 μmol)
NH2-Ser-Gly-Gly-Tyr-Leu-Pro-Pro-Leu-Co2H und 1,8 mg (16 μmol) N-Hydroxysuccinimid in
20 ml Dimethylformamid wurden 3,0 mg (16 μmol) 1-Ethyl-3-(3-dimethylaminopropyl)carbodiimid
(EDC) zugesetzt. Die Reaktion wurde für 10 Stunden bei 25°C gerührt. Zusätzliche
3,0 mg EDC wurden dann zugesetzt und das Rühren wurde bei 25°C für weitere
10 Stunden fortgesetzt. Das Lösungsmittel wurde
durch Rotationsverdampfen entfernt und der Rückstand wurde in 2 ml Wasser
zur Aufreinigung durch Umkehrphasen HPLC auf einer Whatman Partisil
10 ODS-3 9,4-mM X 50-cm Säule
gelöst,
eluiert mit einem linearen Gradienten von 0-50% (V/V) Acetonitril
in 0,1% Trifluoressigsäure/Wasser über 30 Minuten.
Die geeigneten Fraktionen wurde lyophilisiert, so dass sich 2,8
mg (80%) eines weißen
Feststoffes ergaben [m/z 785 (MH+)]. 1H-NMR und UV-vis Spektren des synthetisch
hergestellten Materials waren mit veröffentlichten Spektren für das isolierte
Naturprodukt konsistent (Morita, H., et al, Tetrahedron 50:9975,
1994).
-
Pseudostellarin
F Aufreinigung
-
Die
E. coli Stämme
XL1-Blue, DH5a oder B121-DE3, die pARCP-p beherbergten, wurden gezüchtet, und
wie zur DHFR Aufreinigungen beschrieben, geerntet. Die Medien (500
ml) wurden dreimal mit 1-Butanol (3 × 100 ml) extrahiert. Die Extrakte
wurden kombiniert und abgedampft und der feste Rückstand wurde in 2 ml 0,1 M
K2HPO4 (pH 8,0,
Lysepuffer) resuspendiert. Die Zellen wurden in 10 ml Lysepuffer
resuspendiert, beschallt und durch Zentrifugation geklärt (20 000 × g, 20
Minuten). Das Lysat wurde extrahiert (3 × 5 ml n-Butanol) und die Extrakte
wurden kombiniert, abgedampft und in 500 μl Lysepuffer resuspendiert.
Das rekombinante Produkt wurde aus einem Lysat und Medienextrakten
durch HPLC wie oben beschrieben, aufgereinigt. Die Lyophilisation
der geeigneten Fraktionen aus dem Lysat und den Medienextraktionen
ergab einen öligen
Rückstand,
m/z 785,47 (MH+), 807,43 (MNa+)
und 823,44 (MK+). 1H-NMR
und UV-vis Spektren des rekombinanten Materials waren mit veröffentlichten
Spektren für
das isolierte Naturprodukt konsistent (Morita et al, siehe oben).
Proteine, die an die Chitinbindungsdomäne fusioniert waren, wurden
wie oben beschrieben durch Erzeugung des geklärten Lysates hergestellt. Das
Lysat wurde über
eine Chitinsäule
(New England Biolabs, Inc.) passiert, die mit Lyse-Puffer equilibriert
war. Die Säule
wurde isokratische eluiert und Fraktionen, die Spleißzwischenprodukte
enthielten, wurden gepoolt und einer MALDI Massenspektralanalyse
unterworfen.
-
Tyrosinaseklonierung
-
Das
Tyrosinasegen (einschließlich
ORF 438) aus Streptomyces antibioticus (Bernan et al, Gene 37:101,
1985) wurde mit Vent-Polymerase von piJ702 (ATCC Nr. 35287) mit Primern
amplifiziert, die 5'NdeI und
3'EcoRI Restriktionsstellen
einbrachten. Das PCR Produkt wurde mit NdeI und EcoRi digeriert
und in in ähnlicher
Weise digeriertem pDIMN2 (Ostermeier et al., siehe oben) ligiert,
um pDIMN-Y zu erzeugen. Transformierte Ligationsgemische wurden
bei Umgebungstemperatur für
5 Tage gezüchtet
und Kolonien, die Tyrosinase exprimierten, wurden durch Pigmentbildung
auf FeCuY Platten identifiziert [LB Agarplatten, die Ampicillin
(200 μg/ml),
FeCl3·6H2O (0,2 mM), CuSO4·5H2O (0,2 mM), L-Tyrosin (0,3 mg/ml, Y) und
Isopropyl-B-D-Galactosid (1mM)] (Della-Cioppa, G. et al., Biotechnology
8:634, 1990).
-
Ergebnisse
-
Design der Genkonstrukte
-
Die
Gene, die Ssp IC und IN codierten,
wurden aus genomischer Ssp Gen DNA durch molekularbiologische Standardverfahren
amplifiziert (Sambrook et al, siehe oben) und in pDIMC7 seriell
ligiert. Das sich ergebende Zyklisierungsvorläufer(CP)-Fragment wurde aus
pDIMC7 ausgeschnitten und dem AraB Promotor von pAR3 benachtbart
kloniert, um die pARCP Vektorreihe (3) zu erzeugen.
Diese Vektoren aktivieren die Expression von zyklischen Vorläufern in
Gegenwart von Arabinose. Das E. coli DHFR-Gen wurde zwischen den
NdeI und NsiI Stellen von pARCP kloniert, um eine In-Frame-Fusion
mit jedem der Splitinteingene (b und c in 3) zu erzeugen.
Der PCR-Primer, der zum Amplifizieren von DHFR verwendet wurde,
brachte ebenfalls eine Sequenz ein, die einen Sechs-Histidin-Marker
am 5'-Ende des DHFR
Gens codierte, um eine Immundetektion der zu zyklisierenden Region
zu ermöglichen.
Zwei DHFR Konstrukte wurden assembliert, um die Rolle des vorletzten
Restes von IC in der Säure/Base-Katalyse einer Asparaginseitenkettenzyklisierung
zu untersuchen. Das Plasmid pARCP-DHFR (b in 3) codiert
Wildtyp IC, das einen Alaninrest benachbart
dem terminalen Asparagin aufweist. Plasmid pARCPAH-DHFR (c in 3)
incorporiert eine Alanin-an-Histidin-Mutation an der vorletzten
Position im IC-Gen. Um Pseudostellarin-F
(Cyclo-[SGGYLPPL]) zu erzeugen, wurde der Vektor durch stille Mutation
modifiziert, um eine AfIII Stelle am 5'-Ende
des IN-Gens (d in 3) zu erzeugen.
Eine MfeI Stelle trat natürlicherweise
am 3'-Ende des Wildtyp-IC-Gens auf. Die Ligation eines synthetisch hergestellten,
doppelsträngigen
Insertes, dass Pseudostellarin F codierte, in dem modifizierten
Vektor, erzeugte das Plasmid pARCP-p (e in 3). Eine
KpnI Stelle wurde am 3'-Ende
des IN- Gens
eingebracht, um das Gen für
die Chitingbindungsdomäne
an das Pseudostellarin erzeugende Konstrukt (f in 3)
zu fusionieren.
-
Herstellung und Charakterisierung
von zyklischem DHFR)
-
Die
DHFR Zyklisierung war leicht durch SDS-PAGE nach Arabinoseinduktion
von pARCP-DHFR, wie in 4 dargestellt, erkennbar (F:
IC-DHFR-IN Fusionsprotein.
T: IC-DHFR-IN Fusionsthioesterzwischenprodukt. R: IC-DHFR Lariatzwischenprodukt. L: lineares
DHFR. O: zyklischesDHFR. IN: N-Intein. IC: C-Intein. Bahn 1: nicht induziertes XL1-Blue/pARCP-DHFR.
Bahn 2: arabinoseinduziertes XLI-Blue/pARCP-DHFR. Bahn 3: arabinoseinduziertes
XLIBlue/pARCPAH-DHFR. Bahn 4: Bahn 3-Rohlysat nach Methotrexatagarose.
Bahn 5: Bahn 4-Material nach FPLC. Bahn 6: Wildtyp DHFR).
-
Banden
mit Apparentmolekulargewichten bzw. Molekülmassen entsprechend den linearen
(L, 23 kDa) und zyklischen (0,21 kDA) DHFR Produkten, entsprechend
den Fusionsproteinen (F, 37 kDa), IN (14
kDa) und IC (4 kDa) waren klar sichtbar,
genauso wie Banden die provisorisch als Thioester (T, 36 kDa) und
Lariatzwischenprodukte (R, 26 kDa) bezeichnet wurden (4,
Bahn 2). Die Mutation des vorletzten Restes von IC (A35)
von Alanin zu Histidin (c in 3) verbesserte
die Ausbeute von zyklischem DHFR ( 4, Bahn
3). Eine Methotrexat-Agarose-Affinitätschromatographie des Rohlysates
(4, Bahn 4) bestätigte, dass die Mehrzahl der
induzierten Banden korrekt gefaltetes DHFR enthielt. Obwohl IN nicht kovalent an DHFR gebunden ist, wurde
es auf der Methotexatsäule
vermutlich aufgrund einer nicht-kovalenten Komplexbildung mit dem IC-DHFR
Lariatzwischenprodukt (R) zurückgehalten.
Der Methotrexat-Agaroseeluant wurde durch FPLC fraktioniert und
ermöglichte
die Aufreinigung von 5 mg des zyklischen Produktes pro Liter Kultur
(4, Bahn 5). Ein Westernblotting (nicht dargestellt)
mit einem Anti-His-Antikörper
demonstriert das Vorhandensein der Polyhistidinlinkersequenz (d
in 3) im FPLC aufgereinigten Protein. Das Protein
migrierte rascher in STS/PAGE Analysen als rekombinantes DHFR (4,
Bahn 6) trotz der zusätzlichen
11-Aminosäurelinkersequenz
(b in 3), was eine zusätzliche topologische Beschränkung bedeutet.
Darüber
hinaus wurde keine Reaktion nachgewiesen, wenn das FPLC-aufgereinigte Protein
mit Phenylisothiocyanat umgesetzt wurde (Edman, P., Acta Chem. Scand.,
4:283, 1950), was nahe legt, dass der Aminoterminus nicht verfügbar war.
-
Zyklisches
DHFR wies Steady-State kinetische Parameter und Substrat, Cofaktor-
und Methotrexatdissoziationskonstanten auf, die vom Wildtypenzym
bei 25°C
nicht unterscheidbar waren. Die Aktivitätsassays, die nach 65°C Präinkubation
von Wildtyp und zyklischem DHFR durchgeführt wurden, zeigten, dass die
Zyklisierung die Thermostabilität
des Enzyms verbesserte (5). Die Endoproteinase Lys-C-Digestion
wurde dazu verwendet, unzweifelhaft zu demonstrieren, dass das FPLC
aufgereinigte Protein zyklisches DHFR war. Die Digestion des Wildtypenzyms
erzeugte aminoterminale (4,4 kDa) und carboxyterminale (6,3 kDa)
Fragmente; in einem zyklischen Protein wären diese beiden Fragmente
verbunden, was ein 10,7 kDa Digestionsprodukt zur Folge hätte (7).
Das FPLC aufgereinigte Material war gegenüber einer Proteolyse im Vergleich zu
dem Wildtypenzym resistent und Massenspektralanalysen der Digestionsgemische
identifizierten eine 10,7 kDa Peak im Produkt, das sich aus dem
zyklischen Protein ergab, das im Wildtypenzym (Daten nicht dargestellt)
abwesend war.
-
Produktion und Charakterisierung
von Pseudostellarin F
-
Die
Pseudostellarin F Produktion wurde leicht in vivo durch Hemmung
der rekombinanten Streptomyces antibioticus Tyrosinase (7)
nachgewiesen. In dem in 7 dargestellten Experiment waren XLI-Blue-Zellen
mit pDIM-NY und entweder pARCP2-6H (a & b) oder pARCP-p (c & d) co-transformiert.
Die Zellen wurden auf FeCuY Platten mit Chlroamphenicol (50 μg/ml) entweder
ohne (a & c)
oder mit (b & d) L-(+)Arabinose
(0,5%) ausplatiert.
-
Eine
Co-Expression von Pseudostellarin F in Tyrosinase exprimierenden
Zellen reduziert die Pigmentbildung (d in 7) dramatisch.
Die Expression eines unverwandten zyklischen Peptides aus pARCP2-6H hemmte
Tyrosinase nicht (a und b in 7) und die
Hemmung erforderte die Arabinoseinduktion absolut (vgl. c und d
in 7). Eine SDS/PAGE-Analyse von Arabinose-induziertem
pARCP-p in mehreren bakteriellen Stämmen (BL21-DE3, DH5a, und XLI-Blue)
ermöglichten
die Visualisierung von Banden, die dem Fusionsprotein (F), dem Thioesterzwischenprodukt
(T) und IN entsprachen. Eine intensive niedermolekulare
Gewichtsbande war ebenfalls sichtbar, jedoch war die Auflösung nicht
ausreichend, um das Lariatzwischenprodukt (R) und IC (Daten
nicht dargestellt) aufzutrennen. Obwohl Pseudostellarin F zu klein
war, um durch SDS/PAGE visualisiert zu werden, zeigte die Massenspektralanalyse
sein Vorhandensein sowohl im Rohzelllysat als auch Medien. Ungefähr 30 μg des rekombinanten
zyklischen Peptides wurden aus dem Zelllysatprogramm feuchter Zellmasse
isoliert. Pseudostellarin F wurde ebenfalls aus den Medien durch
1-Butanolextraktion gefolgt von HPLC isoliert, mit einer Ausbeute,
die zwischen 2 mg/Liter (XL I Blue) und 20 mg/Liter (BL21-DE3) variierte, abhängig vom
Expressionsstamm. Das NMR-Spektrum des rekombinanten Materials war
mit demjenigen konsistent, das für
das Naturprodukt berichtet wurde (Morita et al., siehe oben) und
die Retentionszeit des bakteriell expremierten zyklischen Peptides
war mit einem synthetisch hergestellten Standard identisch. Das
rekombinante Material versagte dabei mit Ninhydrin zu reagieren
(siehe Gordon, A.J., und R.A. Ford, The Chemist's Companion: A Handbook of Practical
Data, Techniques, and References, Wiley Interscience, New York,
1972), was auf ein zyklisches Peptidgrundgerüst (Lactam) eher als auf ein
Lactonprodukt hinweist. Weder HPLC noch Massenspektralanalyse ergab
irgendeinen Hinweis auf die Produktion des linearen Stammpeptides.
-
Eine
Chitinbindungsdomäne
wurde an den Carboxy-Terminus von I
N fusioniert,
um die Zwischenprodukte der Intein-vermittelten Ligationsreaktionsaffinität aufzureinigen
und um diese durch MALDI Massenspektometrie (Tabelle 1) zu charakterisieren
Tabelle
1: Massenspektrencharakterisierung von Pseudostellarin F Zyklisierungs-Zwischenprodukten
-
Alle
Zwischenprodukte der Spleißreaktion,
einschließlich
IC, wurden zurückgehalten, wenn das Rohzelllysat
aus Arabinose-induzierten pARCBD-p in XL1-Blue über eine Chitinaffinitätssäule passiert
wurde. Pseudostellarin F wurde aus dem nicht zurückgehaltenen Material durch
1-Butanolextraktion wiedergewonnen. Die beobachteten Molekularmassen
für das
Fusionsprotein (F), das Thioesterzwischenprodukt (T) und IN wiesen eine ausgezeichnete Übereinstimmung
mit den Werten auf, die aus der Gensequenz vorhergesagt wurden.
Die Masse von IC war mit der Asparagin-zyklisierten
Form konsistent, wie es aus dem vorgeschlagenen Mechanismus der
Produktfreisetzung vorhergesagt wurde. Die Molekularmasse des Lariatzwischenproduktes
(R) war mit dem linearen IC-Pseudostellarin
F-Fusionsprodukt konsistenter als das verzweigte Lactonprodukt,
das aus der Trans-Veresterungsreaktion erwartet wurde.
-
Weitere Ausführungsformen
-
Während die
obige Beschreibung viele spezielle Beispiele enthält, sollten
diese nicht als Beschränkung
des Umfangs der Erfindung angesehen werden, sondern eher als Beispiele
bevorzugter Ausführungsformen.
Viele andere Variationen sind möglich.
Demgemäß sollte
der Umfang der Erfindung nicht durch die dargestellten Ausführungsformen,
sondern durch die beigefügten
Ansprüche
und ihren Äquivalenzbereich
bestimmt werden. SEQUENCE
LISTING