ES2695125T3 - Métodos y organismos para la producción de 1,4-butanodiol acoplada al crecimiento - Google Patents
Métodos y organismos para la producción de 1,4-butanodiol acoplada al crecimiento Download PDFInfo
- Publication number
- ES2695125T3 ES2695125T3 ES13001197T ES13001197T ES2695125T3 ES 2695125 T3 ES2695125 T3 ES 2695125T3 ES 13001197 T ES13001197 T ES 13001197T ES 13001197 T ES13001197 T ES 13001197T ES 2695125 T3 ES2695125 T3 ES 2695125T3
- Authority
- ES
- Spain
- Prior art keywords
- dehydrogenase
- butanediol
- bdo
- coa
- production
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- WERYXYBDKMZEQL-UHFFFAOYSA-N butane-1,4-diol Chemical compound OCCCCO WERYXYBDKMZEQL-UHFFFAOYSA-N 0.000 title claims abstract description 224
- 238000004519 manufacturing process Methods 0.000 title claims abstract description 161
- 238000000034 method Methods 0.000 title claims abstract description 140
- 230000012010 growth Effects 0.000 title claims description 106
- SJZRECIVHVDYJC-UHFFFAOYSA-M 4-hydroxybutyrate Chemical compound OCCCC([O-])=O SJZRECIVHVDYJC-UHFFFAOYSA-M 0.000 claims abstract description 291
- 244000005700 microbiome Species 0.000 claims abstract description 99
- 101710088194 Dehydrogenase Proteins 0.000 claims abstract description 96
- 102000004190 Enzymes Human genes 0.000 claims abstract description 76
- 108090000790 Enzymes Proteins 0.000 claims abstract description 76
- SJZRECIVHVDYJC-UHFFFAOYSA-N 4-hydroxybutyric acid Chemical compound OCCCC(O)=O SJZRECIVHVDYJC-UHFFFAOYSA-N 0.000 claims abstract description 55
- 108020004707 nucleic acids Proteins 0.000 claims abstract description 52
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 52
- 102000039446 nucleic acids Human genes 0.000 claims abstract description 52
- 230000001419 dependent effect Effects 0.000 claims abstract description 51
- 101150014383 adhE gene Proteins 0.000 claims abstract description 48
- 108020002663 Aldehyde Dehydrogenase Proteins 0.000 claims abstract description 22
- 102000005369 Aldehyde Dehydrogenase Human genes 0.000 claims abstract description 22
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 claims abstract description 22
- 229930195712 glutamate Natural products 0.000 claims abstract description 22
- 102000007698 Alcohol dehydrogenase Human genes 0.000 claims abstract description 18
- 230000003698 anagen phase Effects 0.000 claims abstract description 7
- 235000015097 nutrients Nutrition 0.000 claims abstract description 6
- YEJRWHAVMIAJKC-UHFFFAOYSA-N 4-Butyrolactone Chemical group O=C1CCCO1 YEJRWHAVMIAJKC-UHFFFAOYSA-N 0.000 claims description 147
- WYURNTSHIVDZCO-UHFFFAOYSA-N Tetrahydrofuran Chemical group C1CCOC1 WYURNTSHIVDZCO-UHFFFAOYSA-N 0.000 claims description 107
- 238000000855 fermentation Methods 0.000 claims description 65
- 230000004151 fermentation Effects 0.000 claims description 63
- YLQBMQCUIZJEEH-UHFFFAOYSA-N tetrahydrofuran Natural products C=1C=COC=1 YLQBMQCUIZJEEH-UHFFFAOYSA-N 0.000 claims description 52
- 230000001851 biosynthetic effect Effects 0.000 claims description 33
- 102000011929 Succinate-CoA Ligases Human genes 0.000 claims description 21
- 108010075728 Succinate-CoA Ligases Proteins 0.000 claims description 21
- 108091022930 Glutamate decarboxylase Proteins 0.000 claims description 19
- 102000008214 Glutamate decarboxylase Human genes 0.000 claims description 19
- 102000003929 Transaminases Human genes 0.000 claims description 18
- 108090000340 Transaminases Proteins 0.000 claims description 18
- 108010021809 Alcohol dehydrogenase Proteins 0.000 claims description 14
- 102000004357 Transferases Human genes 0.000 claims description 10
- 108090000992 Transferases Proteins 0.000 claims description 10
- 238000004821 distillation Methods 0.000 claims description 6
- HNJBEVLQSNELDL-UHFFFAOYSA-N pyrrolidin-2-one Chemical group O=C1CCCN1 HNJBEVLQSNELDL-UHFFFAOYSA-N 0.000 claims description 3
- KDYFGRWQOYBRFD-UHFFFAOYSA-L succinate(2-) Chemical compound [O-]C(=O)CCC([O-])=O KDYFGRWQOYBRFD-UHFFFAOYSA-L 0.000 abstract description 58
- 102000019259 Succinate Dehydrogenase Human genes 0.000 abstract description 33
- 108010012901 Succinate Dehydrogenase Proteins 0.000 abstract description 33
- 230000006696 biosynthetic metabolic pathway Effects 0.000 abstract description 18
- 101100398785 Streptococcus agalactiae serotype V (strain ATCC BAA-611 / 2603 V/R) ldhD gene Proteins 0.000 abstract description 14
- 101100386830 Zymomonas mobilis subsp. mobilis (strain ATCC 31821 / ZM4 / CP4) ddh gene Proteins 0.000 abstract description 14
- 101150026107 ldh1 gene Proteins 0.000 abstract description 14
- 101150041530 ldha gene Proteins 0.000 abstract description 14
- 101710181816 Pyruvate-formate-lyase deactivase Proteins 0.000 abstract description 4
- KDYFGRWQOYBRFD-UHFFFAOYSA-N Succinic acid Natural products OC(=O)CCC(O)=O KDYFGRWQOYBRFD-UHFFFAOYSA-N 0.000 abstract 6
- KDYFGRWQOYBRFD-NUQCWPJISA-N butanedioic acid Chemical compound O[14C](=O)CC[14C](O)=O KDYFGRWQOYBRFD-NUQCWPJISA-N 0.000 abstract 4
- 239000001384 succinic acid Substances 0.000 abstract 1
- CDQSJQSWAWPGKG-UHFFFAOYSA-N butane-1,1-diol Chemical compound CCCC(O)O CDQSJQSWAWPGKG-UHFFFAOYSA-N 0.000 description 207
- 108090000623 proteins and genes Proteins 0.000 description 147
- 238000006243 chemical reaction Methods 0.000 description 136
- 230000002503 metabolic effect Effects 0.000 description 104
- 239000000047 product Substances 0.000 description 99
- 230000000813 microbial effect Effects 0.000 description 97
- 241000588724 Escherichia coli Species 0.000 description 91
- 230000015572 biosynthetic process Effects 0.000 description 79
- 230000037430 deletion Effects 0.000 description 75
- 230000004048 modification Effects 0.000 description 75
- 238000012986 modification Methods 0.000 description 75
- 238000012217 deletion Methods 0.000 description 74
- 210000004027 cell Anatomy 0.000 description 49
- 238000013461 design Methods 0.000 description 49
- 101100456369 Aquifex aeolicus (strain VF5) mdh1 gene Proteins 0.000 description 43
- 101150058595 MDH gene Proteins 0.000 description 43
- 101100384788 Methanocaldococcus jannaschii (strain ATCC 43067 / DSM 2661 / JAL-1 / JCM 10045 / NBRC 100440) comC gene Proteins 0.000 description 43
- 101100020705 Mycoplasma gallisepticum (strain R(low / passage 15 / clone 2)) ldh gene Proteins 0.000 description 43
- 101100290490 Rattus norvegicus Mdh1 gene Proteins 0.000 description 43
- 239000000243 solution Substances 0.000 description 43
- 230000000694 effects Effects 0.000 description 42
- 150000001875 compounds Chemical class 0.000 description 40
- 241000894007 species Species 0.000 description 40
- 238000000926 separation method Methods 0.000 description 39
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 35
- 108090000472 Phosphoenolpyruvate carboxykinase (ATP) Proteins 0.000 description 35
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 35
- 239000008103 glucose Substances 0.000 description 35
- 230000009849 deactivation Effects 0.000 description 32
- 230000014509 gene expression Effects 0.000 description 30
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 29
- 230000037361 pathway Effects 0.000 description 29
- 101150015622 pyk gene Proteins 0.000 description 28
- VNOYUJKHFWYWIR-ITIYDSSPSA-N succinyl-CoA Chemical compound O[C@@H]1[C@H](OP(O)(O)=O)[C@@H](COP(O)(=O)OP(O)(=O)OCC(C)(C)[C@@H](O)C(=O)NCCC(=O)NCCSC(=O)CCC(O)=O)O[C@H]1N1C2=NC=NC(N)=C2N=C1 VNOYUJKHFWYWIR-ITIYDSSPSA-N 0.000 description 28
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 description 27
- 229930027945 nicotinamide-adenine dinucleotide Natural products 0.000 description 26
- -1 polybutylene terephthalate Polymers 0.000 description 25
- 239000000203 mixture Substances 0.000 description 23
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 21
- 108010026217 Malate Dehydrogenase Proteins 0.000 description 21
- 102000013460 Malate Dehydrogenase Human genes 0.000 description 21
- XJLXINKUBYWONI-DQQFMEOOSA-N [[(2r,3r,4r,5r)-5-(6-aminopurin-9-yl)-3-hydroxy-4-phosphonooxyoxolan-2-yl]methoxy-hydroxyphosphoryl] [(2s,3r,4s,5s)-5-(3-carbamoylpyridin-1-ium-1-yl)-3,4-dihydroxyoxolan-2-yl]methyl phosphate Chemical compound NC(=O)C1=CC=C[N+]([C@@H]2[C@H]([C@@H](O)[C@H](COP([O-])(=O)OP(O)(=O)OC[C@@H]3[C@H]([C@@H](OP(O)(O)=O)[C@@H](O3)N3C4=NC=NC(N)=C4N=C3)O)O2)O)=C1 XJLXINKUBYWONI-DQQFMEOOSA-N 0.000 description 21
- 230000002068 genetic effect Effects 0.000 description 21
- 238000005984 hydrogenation reaction Methods 0.000 description 21
- 239000000126 substance Substances 0.000 description 21
- 239000000758 substrate Substances 0.000 description 21
- 238000006722 reduction reaction Methods 0.000 description 20
- 238000003786 synthesis reaction Methods 0.000 description 20
- 239000002609 medium Substances 0.000 description 19
- 241000894006 Bacteria Species 0.000 description 18
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 18
- YXFVVABEGXRONW-UHFFFAOYSA-N Toluene Chemical compound CC1=CC=CC=C1 YXFVVABEGXRONW-UHFFFAOYSA-N 0.000 description 18
- 230000009467 reduction Effects 0.000 description 17
- 239000002028 Biomass Substances 0.000 description 16
- 101100282733 Escherichia coli (strain K12) ghrA gene Proteins 0.000 description 16
- 101100014624 Escherichia coli (strain K12) ghrB gene Proteins 0.000 description 16
- 101100182965 Escherichia coli (strain K12) maeA gene Proteins 0.000 description 16
- 238000004458 analytical method Methods 0.000 description 16
- 108010008221 formate C-acetyltransferase Proteins 0.000 description 16
- 230000006870 function Effects 0.000 description 16
- 238000000126 in silico method Methods 0.000 description 16
- 230000003834 intracellular effect Effects 0.000 description 16
- 101150007902 ASPA gene Proteins 0.000 description 15
- 101100378784 Emericella nidulans (strain FGSC A4 / ATCC 38163 / CBS 112.46 / NRRL 194 / M139) aldA gene Proteins 0.000 description 15
- 101100057016 Talaromyces wortmannii astA gene Proteins 0.000 description 15
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 15
- 230000008878 coupling Effects 0.000 description 15
- 238000010168 coupling process Methods 0.000 description 15
- 238000005859 coupling reaction Methods 0.000 description 15
- 229910052760 oxygen Inorganic materials 0.000 description 15
- 239000001301 oxygen Substances 0.000 description 15
- 101100442929 Bacillus licheniformis (strain ATCC 14580 / DSM 13 / JCM 2505 / CCUG 7422 / NBRC 12200 / NCIMB 9375 / NCTC 10341 / NRRL NRS-1264 / Gibson 46) deoC2 gene Proteins 0.000 description 14
- 101100310802 Dictyostelium discoideum splA gene Proteins 0.000 description 14
- 229910002651 NO3 Inorganic materials 0.000 description 14
- NHNBFGGVMKEFGY-UHFFFAOYSA-N Nitrate Chemical compound [O-][N+]([O-])=O NHNBFGGVMKEFGY-UHFFFAOYSA-N 0.000 description 14
- 101150107914 Ubr5 gene Proteins 0.000 description 14
- 101150013644 deoC gene Proteins 0.000 description 14
- 101150090240 edd gene Proteins 0.000 description 14
- 230000008030 elimination Effects 0.000 description 14
- 238000003379 elimination reaction Methods 0.000 description 14
- 239000000543 intermediate Substances 0.000 description 14
- 101150108859 maeB gene Proteins 0.000 description 14
- BOPGDPNILDQYTO-NNYOXOHSSA-N nicotinamide-adenine dinucleotide Chemical compound C1=CCC(C(=O)N)=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](COP(O)(=O)OP(O)(=O)OC[C@@H]2[C@H]([C@@H](O)[C@@H](O2)N2C3=NC=NC(N)=C3N=C2)O)O1 BOPGDPNILDQYTO-NNYOXOHSSA-N 0.000 description 14
- KHPXUQMNIQBQEV-UHFFFAOYSA-N oxaloacetic acid Chemical compound OC(=O)CC(=O)C(O)=O KHPXUQMNIQBQEV-UHFFFAOYSA-N 0.000 description 14
- 230000008569 process Effects 0.000 description 14
- 101150100525 pykA gene Proteins 0.000 description 14
- 101150053304 pykF gene Proteins 0.000 description 14
- 238000007792 addition Methods 0.000 description 13
- DTBNBXWJWCWCIK-UHFFFAOYSA-K phosphonatoenolpyruvate Chemical compound [O-]C(=O)C(=C)OP([O-])([O-])=O DTBNBXWJWCWCIK-UHFFFAOYSA-K 0.000 description 13
- ZSLZBFCDCINBPY-ZSJPKINUSA-N Acetyl-CoA Natural products O[C@@H]1[C@H](OP(O)(O)=O)[C@@H](COP(O)(=O)OP(O)(=O)OCC(C)(C)[C@@H](O)C(=O)NCCC(=O)NCCSC(=O)C)O[C@H]1N1C2=NC=NC(N)=C2N=C1 ZSLZBFCDCINBPY-ZSJPKINUSA-N 0.000 description 12
- 238000004364 calculation method Methods 0.000 description 12
- 239000007788 liquid Substances 0.000 description 12
- 238000006241 metabolic reaction Methods 0.000 description 12
- 238000005457 optimization Methods 0.000 description 12
- 239000003960 organic solvent Substances 0.000 description 12
- 229930029653 phosphoenolpyruvate Natural products 0.000 description 12
- 230000001105 regulatory effect Effects 0.000 description 12
- VQUVCIYMYJAJIZ-UHFFFAOYSA-N 2-[n-ethyl-4-[2-(1-methylpyridin-1-ium-4-yl)ethenyl]anilino]ethanol;tetraphenylboranuide Chemical compound C1=CC(N(CCO)CC)=CC=C1\C=C\C1=CC=[N+](C)C=C1.C1=CC=CC=C1[B-](C=1C=CC=CC=1)(C=1C=CC=CC=1)C1=CC=CC=C1 VQUVCIYMYJAJIZ-UHFFFAOYSA-N 0.000 description 11
- 239000002253 acid Substances 0.000 description 11
- 101150097574 gsk gene Proteins 0.000 description 11
- 101150029104 prpC gene Proteins 0.000 description 11
- 230000002441 reversible effect Effects 0.000 description 11
- KPGXRSRHYNQIFN-UHFFFAOYSA-L 2-oxoglutarate(2-) Chemical compound [O-]C(=O)CCC(=O)C([O-])=O KPGXRSRHYNQIFN-UHFFFAOYSA-L 0.000 description 10
- LCTONWCANYUPML-UHFFFAOYSA-M Pyruvate Chemical compound CC(=O)C([O-])=O LCTONWCANYUPML-UHFFFAOYSA-M 0.000 description 10
- 239000003054 catalyst Substances 0.000 description 10
- 230000010261 cell growth Effects 0.000 description 10
- 230000007423 decrease Effects 0.000 description 10
- 230000002255 enzymatic effect Effects 0.000 description 10
- 239000002207 metabolite Substances 0.000 description 10
- 238000004088 simulation Methods 0.000 description 10
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 9
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 9
- 230000003044 adaptive effect Effects 0.000 description 9
- 229910002092 carbon dioxide Inorganic materials 0.000 description 9
- 230000004077 genetic alteration Effects 0.000 description 9
- 231100000118 genetic alteration Toxicity 0.000 description 9
- 230000006680 metabolic alteration Effects 0.000 description 9
- 102000004169 proteins and genes Human genes 0.000 description 9
- 230000002829 reductive effect Effects 0.000 description 9
- 241000186570 Clostridium kluyveri Species 0.000 description 8
- 241000233866 Fungi Species 0.000 description 8
- 238000010353 genetic engineering Methods 0.000 description 8
- 230000001939 inductive effect Effects 0.000 description 8
- 230000002427 irreversible effect Effects 0.000 description 8
- 230000004060 metabolic process Effects 0.000 description 8
- 101150094267 mqo gene Proteins 0.000 description 8
- 239000007787 solid Substances 0.000 description 8
- 241000193401 Clostridium acetobutylicum Species 0.000 description 7
- 101150099894 GDHA gene Proteins 0.000 description 7
- 101100277701 Halobacterium salinarum gdhX gene Proteins 0.000 description 7
- 101100392454 Picrophilus torridus (strain ATCC 700027 / DSM 9790 / JCM 10055 / NBRC 100828) gdh2 gene Proteins 0.000 description 7
- 101100116769 Saccharolobus solfataricus (strain ATCC 35092 / DSM 1617 / JCM 11322 / P2) gdhA-2 gene Proteins 0.000 description 7
- 238000010276 construction Methods 0.000 description 7
- 238000012423 maintenance Methods 0.000 description 7
- 230000037353 metabolic pathway Effects 0.000 description 7
- 230000003647 oxidation Effects 0.000 description 7
- 238000007254 oxidation reaction Methods 0.000 description 7
- 101150000475 pntAB gene Proteins 0.000 description 7
- NLXLAEXVIDQMFP-UHFFFAOYSA-N Ammonia chloride Chemical compound [NH4+].[Cl-] NLXLAEXVIDQMFP-UHFFFAOYSA-N 0.000 description 6
- 108020002908 Epoxide hydrolase Proteins 0.000 description 6
- 241000029538 [Mannheimia] succiniciproducens Species 0.000 description 6
- 238000013459 approach Methods 0.000 description 6
- 239000007864 aqueous solution Substances 0.000 description 6
- 239000003153 chemical reaction reagent Substances 0.000 description 6
- 239000012526 feed medium Substances 0.000 description 6
- 239000007791 liquid phase Substances 0.000 description 6
- 238000000622 liquid--liquid extraction Methods 0.000 description 6
- VNWKTOKETHGBQD-UHFFFAOYSA-N methane Chemical compound C VNWKTOKETHGBQD-UHFFFAOYSA-N 0.000 description 6
- 239000012071 phase Substances 0.000 description 6
- 108090000765 processed proteins & peptides Proteins 0.000 description 6
- 230000029058 respiratory gaseous exchange Effects 0.000 description 6
- 230000028327 secretion Effects 0.000 description 6
- 238000000638 solvent extraction Methods 0.000 description 6
- 241000948980 Actinobacillus succinogenes Species 0.000 description 5
- 241000722954 Anaerobiospirillum succiniciproducens Species 0.000 description 5
- 241000228245 Aspergillus niger Species 0.000 description 5
- 241001465318 Aspergillus terreus Species 0.000 description 5
- 235000014469 Bacillus subtilis Nutrition 0.000 description 5
- 241000186226 Corynebacterium glutamicum Species 0.000 description 5
- 241000252867 Cupriavidus metallidurans Species 0.000 description 5
- 102000005486 Epoxide hydrolase Human genes 0.000 description 5
- 241000589232 Gluconobacter oxydans Species 0.000 description 5
- 241000588749 Klebsiella oxytoca Species 0.000 description 5
- 241001138401 Kluyveromyces lactis Species 0.000 description 5
- 241000235058 Komagataella pastoris Species 0.000 description 5
- 240000006024 Lactobacillus plantarum Species 0.000 description 5
- 241000589540 Pseudomonas fluorescens Species 0.000 description 5
- 241000589776 Pseudomonas putida Species 0.000 description 5
- 241001148115 Rhizobium etli Species 0.000 description 5
- 244000253911 Saccharomyces fragilis Species 0.000 description 5
- 244000057717 Streptococcus lactis Species 0.000 description 5
- 241000588902 Zymomonas mobilis Species 0.000 description 5
- 238000009825 accumulation Methods 0.000 description 5
- 230000008859 change Effects 0.000 description 5
- 239000003795 chemical substances by application Substances 0.000 description 5
- 238000006073 displacement reaction Methods 0.000 description 5
- 230000002349 favourable effect Effects 0.000 description 5
- 239000007789 gas Substances 0.000 description 5
- 230000001976 improved effect Effects 0.000 description 5
- 230000002779 inactivation Effects 0.000 description 5
- 238000002955 isolation Methods 0.000 description 5
- 230000000670 limiting effect Effects 0.000 description 5
- 229940049920 malate Drugs 0.000 description 5
- BJEPYKJPYRNKOW-UHFFFAOYSA-L malate(2-) Chemical compound [O-]C(=O)C(O)CC([O-])=O BJEPYKJPYRNKOW-UHFFFAOYSA-L 0.000 description 5
- 230000002018 overexpression Effects 0.000 description 5
- 239000013612 plasmid Substances 0.000 description 5
- 229920000642 polymer Polymers 0.000 description 5
- 229920001184 polypeptide Polymers 0.000 description 5
- 102000004196 processed proteins & peptides Human genes 0.000 description 5
- 239000000376 reactant Substances 0.000 description 5
- LWIHDJKSTIGBAC-UHFFFAOYSA-K tripotassium phosphate Chemical compound [K+].[K+].[K+].[O-]P([O-])([O-])=O LWIHDJKSTIGBAC-UHFFFAOYSA-K 0.000 description 5
- 108010030844 2-methylcitrate synthase Proteins 0.000 description 4
- KPGXRSRHYNQIFN-UHFFFAOYSA-N 2-oxoglutaric acid Chemical compound OC(=O)CCC(=O)C(O)=O KPGXRSRHYNQIFN-UHFFFAOYSA-N 0.000 description 4
- 108010009924 Aconitate hydratase Proteins 0.000 description 4
- 102000009836 Aconitate hydratase Human genes 0.000 description 4
- QGZKDVFQNNGYKY-UHFFFAOYSA-N Ammonia Chemical compound N QGZKDVFQNNGYKY-UHFFFAOYSA-N 0.000 description 4
- 108010071536 Citrate (Si)-synthase Proteins 0.000 description 4
- 102000006732 Citrate synthase Human genes 0.000 description 4
- RGJOEKWQDUBAIZ-IBOSZNHHSA-N CoASH Chemical compound O[C@@H]1[C@H](OP(O)(O)=O)[C@@H](COP(O)(=O)OP(O)(=O)OCC(C)(C)[C@@H](O)C(=O)NCCC(=O)NCCS)O[C@H]1N1C2=NC=NC(N)=C2N=C1 RGJOEKWQDUBAIZ-IBOSZNHHSA-N 0.000 description 4
- BDAGIHXWWSANSR-UHFFFAOYSA-M Formate Chemical compound [O-]C=O BDAGIHXWWSANSR-UHFFFAOYSA-M 0.000 description 4
- 108090000698 Formate Dehydrogenases Proteins 0.000 description 4
- 102000016901 Glutamate dehydrogenase Human genes 0.000 description 4
- 108010075869 Isocitrate Dehydrogenase Proteins 0.000 description 4
- 102000012011 Isocitrate Dehydrogenase Human genes 0.000 description 4
- JVTAAEKCZFNVCJ-UHFFFAOYSA-M Lactate Chemical compound CC(O)C([O-])=O JVTAAEKCZFNVCJ-UHFFFAOYSA-M 0.000 description 4
- CSNNHWWHGAXBCP-UHFFFAOYSA-L Magnesium sulfate Chemical compound [Mg+2].[O-][S+2]([O-])([O-])[O-] CSNNHWWHGAXBCP-UHFFFAOYSA-L 0.000 description 4
- SECXISVLQFMRJM-UHFFFAOYSA-N N-Methylpyrrolidone Chemical compound CN1CCCC1=O SECXISVLQFMRJM-UHFFFAOYSA-N 0.000 description 4
- 241000187432 Streptomyces coelicolor Species 0.000 description 4
- 150000001413 amino acids Chemical group 0.000 description 4
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 4
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 4
- 230000001580 bacterial effect Effects 0.000 description 4
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 4
- 239000011942 biocatalyst Substances 0.000 description 4
- 230000008238 biochemical pathway Effects 0.000 description 4
- 238000009835 boiling Methods 0.000 description 4
- 239000001569 carbon dioxide Substances 0.000 description 4
- 230000001413 cellular effect Effects 0.000 description 4
- 239000003638 chemical reducing agent Substances 0.000 description 4
- 238000005520 cutting process Methods 0.000 description 4
- 230000002950 deficient Effects 0.000 description 4
- 238000010586 diagram Methods 0.000 description 4
- 230000004907 flux Effects 0.000 description 4
- 238000012224 gene deletion Methods 0.000 description 4
- 238000012239 gene modification Methods 0.000 description 4
- 230000005017 genetic modification Effects 0.000 description 4
- 235000013617 genetically modified food Nutrition 0.000 description 4
- 230000006872 improvement Effects 0.000 description 4
- 238000010348 incorporation Methods 0.000 description 4
- 238000005259 measurement Methods 0.000 description 4
- 230000007935 neutral effect Effects 0.000 description 4
- 230000036961 partial effect Effects 0.000 description 4
- 239000002243 precursor Substances 0.000 description 4
- 239000002994 raw material Substances 0.000 description 4
- 150000003839 salts Chemical group 0.000 description 4
- 230000002269 spontaneous effect Effects 0.000 description 4
- 230000004102 tricarboxylic acid cycle Effects 0.000 description 4
- 241000203069 Archaea Species 0.000 description 3
- 244000063299 Bacillus subtilis Species 0.000 description 3
- 108020005199 Dehydrogenases Proteins 0.000 description 3
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 3
- 108010036781 Fumarate Hydratase Proteins 0.000 description 3
- 102100036160 Fumarate hydratase, mitochondrial Human genes 0.000 description 3
- 235000014663 Kluyveromyces fragilis Nutrition 0.000 description 3
- 235000013965 Lactobacillus plantarum Nutrition 0.000 description 3
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 3
- 102000016387 Pancreatic elastase Human genes 0.000 description 3
- 108010067372 Pancreatic elastase Proteins 0.000 description 3
- 235000018368 Saccharomyces fragilis Nutrition 0.000 description 3
- 241000235347 Schizosaccharomyces pombe Species 0.000 description 3
- 235000014897 Streptococcus lactis Nutrition 0.000 description 3
- 240000008042 Zea mays Species 0.000 description 3
- 235000016383 Zea mays subsp huehuetenangensis Nutrition 0.000 description 3
- 235000002017 Zea mays subsp mays Nutrition 0.000 description 3
- 230000002378 acidificating effect Effects 0.000 description 3
- 230000002411 adverse Effects 0.000 description 3
- HSFWRNGVRCDJHI-UHFFFAOYSA-N alpha-acetylene Natural products C#C HSFWRNGVRCDJHI-UHFFFAOYSA-N 0.000 description 3
- 235000019270 ammonium chloride Nutrition 0.000 description 3
- 230000009604 anaerobic growth Effects 0.000 description 3
- 239000006227 byproduct Substances 0.000 description 3
- 238000004422 calculation algorithm Methods 0.000 description 3
- 238000001944 continuous distillation Methods 0.000 description 3
- 230000007812 deficiency Effects 0.000 description 3
- 238000011161 development Methods 0.000 description 3
- 230000018109 developmental process Effects 0.000 description 3
- 125000002534 ethynyl group Chemical group [H]C#C* 0.000 description 3
- 230000003203 everyday effect Effects 0.000 description 3
- 239000012467 final product Substances 0.000 description 3
- 238000009472 formulation Methods 0.000 description 3
- 230000004190 glucose uptake Effects 0.000 description 3
- 230000034659 glycolysis Effects 0.000 description 3
- 238000009904 heterogeneous catalytic hydrogenation reaction Methods 0.000 description 3
- 238000004128 high performance liquid chromatography Methods 0.000 description 3
- 239000001257 hydrogen Substances 0.000 description 3
- 229910052739 hydrogen Inorganic materials 0.000 description 3
- 125000004435 hydrogen atom Chemical class [H]* 0.000 description 3
- 238000001727 in vivo Methods 0.000 description 3
- 230000001965 increasing effect Effects 0.000 description 3
- 229940031154 kluyveromyces marxianus Drugs 0.000 description 3
- 229940072205 lactobacillus plantarum Drugs 0.000 description 3
- 150000002596 lactones Chemical class 0.000 description 3
- 235000009973 maize Nutrition 0.000 description 3
- 230000007246 mechanism Effects 0.000 description 3
- 238000012269 metabolic engineering Methods 0.000 description 3
- 230000035772 mutation Effects 0.000 description 3
- 150000007524 organic acids Chemical class 0.000 description 3
- 235000005985 organic acids Nutrition 0.000 description 3
- 239000004033 plastic Substances 0.000 description 3
- 229920003023 plastic Polymers 0.000 description 3
- 239000005014 poly(hydroxyalkanoate) Substances 0.000 description 3
- 150000004040 pyrrolidinones Chemical class 0.000 description 3
- 239000002904 solvent Substances 0.000 description 3
- 239000007858 starting material Substances 0.000 description 3
- UIUJIQZEACWQSV-UHFFFAOYSA-N succinic semialdehyde Chemical compound OC(=O)CCC=O UIUJIQZEACWQSV-UHFFFAOYSA-N 0.000 description 3
- 230000032258 transport Effects 0.000 description 3
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 3
- PKAUICCNAWQPAU-UHFFFAOYSA-N 2-(4-chloro-2-methylphenoxy)acetic acid;n-methylmethanamine Chemical compound CNC.CC1=CC(Cl)=CC=C1OCC(O)=O PKAUICCNAWQPAU-UHFFFAOYSA-N 0.000 description 2
- 108090000124 3-hydroxybutyrate dehydrogenases Proteins 0.000 description 2
- 102000034279 3-hydroxybutyrate dehydrogenases Human genes 0.000 description 2
- SXIFAEWFOJETOA-UHFFFAOYSA-N 4-hydroxy-butyl Chemical group [CH2]CCCO SXIFAEWFOJETOA-UHFFFAOYSA-N 0.000 description 2
- 108010093796 4-hydroxybutyrate dehydrogenase Proteins 0.000 description 2
- 102100039702 Alcohol dehydrogenase class-3 Human genes 0.000 description 2
- 108030004524 Aryl-aldehyde dehydrogenases Proteins 0.000 description 2
- 101000950981 Bacillus subtilis (strain 168) Catabolic NAD-specific glutamate dehydrogenase RocG Proteins 0.000 description 2
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 2
- 108091026890 Coding region Proteins 0.000 description 2
- 102000005870 Coenzyme A Ligases Human genes 0.000 description 2
- 108010064744 D-lactaldehyde dehydrogenase Proteins 0.000 description 2
- 241000196324 Embryophyta Species 0.000 description 2
- 241000206602 Eukaryota Species 0.000 description 2
- 108060002716 Exonuclease Proteins 0.000 description 2
- 101000892220 Geobacillus thermodenitrificans (strain NG80-2) Long-chain-alcohol dehydrogenase 1 Proteins 0.000 description 2
- 108700023156 Glutamate dehydrogenases Proteins 0.000 description 2
- 108010000445 Glycerate dehydrogenase Proteins 0.000 description 2
- KFZMGEQAYNKOFK-UHFFFAOYSA-N Isopropanol Chemical compound CC(C)O KFZMGEQAYNKOFK-UHFFFAOYSA-N 0.000 description 2
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 2
- 102000003855 L-lactate dehydrogenase Human genes 0.000 description 2
- 108700023483 L-lactate dehydrogenases Proteins 0.000 description 2
- 102000003960 Ligases Human genes 0.000 description 2
- 108090000364 Ligases Proteins 0.000 description 2
- 108010011449 Long-chain-fatty-acid-CoA ligase Proteins 0.000 description 2
- 101150026369 MT-ND6 gene Proteins 0.000 description 2
- OFOBLEOULBTSOW-UHFFFAOYSA-L Malonate Chemical compound [O-]C(=O)CC([O-])=O OFOBLEOULBTSOW-UHFFFAOYSA-L 0.000 description 2
- 108020000290 Mannitol dehydrogenase Proteins 0.000 description 2
- LRHPLDYGYMQRHN-UHFFFAOYSA-N N-Butanol Chemical compound CCCCO LRHPLDYGYMQRHN-UHFFFAOYSA-N 0.000 description 2
- 102100028386 NADH-ubiquinone oxidoreductase chain 6 Human genes 0.000 description 2
- 101150090932 ND6 gene Proteins 0.000 description 2
- IOVCWXUNBOPUCH-UHFFFAOYSA-M Nitrite anion Chemical compound [O-]N=O IOVCWXUNBOPUCH-UHFFFAOYSA-M 0.000 description 2
- 102000013566 Plasminogen Human genes 0.000 description 2
- 108010051456 Plasminogen Proteins 0.000 description 2
- 102000012479 Serine Proteases Human genes 0.000 description 2
- 108010022999 Serine Proteases Proteins 0.000 description 2
- 102100026974 Sorbitol dehydrogenase Human genes 0.000 description 2
- 229920002334 Spandex Polymers 0.000 description 2
- QAOWNCQODCNURD-UHFFFAOYSA-N Sulfuric acid Chemical compound OS(O)(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-N 0.000 description 2
- XLOMVQKBTHCTTD-UHFFFAOYSA-N Zinc monoxide Chemical compound [Zn]=O XLOMVQKBTHCTTD-UHFFFAOYSA-N 0.000 description 2
- 230000000397 acetylating effect Effects 0.000 description 2
- 239000000654 additive Substances 0.000 description 2
- 230000004075 alteration Effects 0.000 description 2
- 229910021529 ammonia Inorganic materials 0.000 description 2
- 230000003466 anti-cipated effect Effects 0.000 description 2
- 239000012298 atmosphere Substances 0.000 description 2
- 230000006399 behavior Effects 0.000 description 2
- 230000003542 behavioural effect Effects 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 238000005842 biochemical reaction Methods 0.000 description 2
- 239000001273 butane Substances 0.000 description 2
- 229910052799 carbon Inorganic materials 0.000 description 2
- 238000009903 catalytic hydrogenation reaction Methods 0.000 description 2
- 239000012707 chemical precursor Substances 0.000 description 2
- RNCXCPUOPDATPU-UHFFFAOYSA-N chromium(3+) oxocopper oxygen(2-) Chemical compound [O-2].[Cr+3].[Cu]=O.[O-2].[O-2].[Cr+3] RNCXCPUOPDATPU-UHFFFAOYSA-N 0.000 description 2
- 210000000349 chromosome Anatomy 0.000 description 2
- RGJOEKWQDUBAIZ-UHFFFAOYSA-N coenzime A Natural products OC1C(OP(O)(O)=O)C(COP(O)(=O)OP(O)(=O)OCC(C)(C)C(O)C(=O)NCCC(=O)NCCS)OC1N1C2=NC=NC(N)=C2N=C1 RGJOEKWQDUBAIZ-UHFFFAOYSA-N 0.000 description 2
- 239000005516 coenzyme A Substances 0.000 description 2
- 229940093530 coenzyme a Drugs 0.000 description 2
- 229920001577 copolymer Polymers 0.000 description 2
- 238000012136 culture method Methods 0.000 description 2
- 238000012258 culturing Methods 0.000 description 2
- KDTSHFARGAKYJN-UHFFFAOYSA-N dephosphocoenzyme A Natural products OC1C(O)C(COP(O)(=O)OP(O)(=O)OCC(C)(C)C(O)C(=O)NCCC(=O)NCCS)OC1N1C2=NC=NC(N)=C2N=C1 KDTSHFARGAKYJN-UHFFFAOYSA-N 0.000 description 2
- 230000001627 detrimental effect Effects 0.000 description 2
- 238000009792 diffusion process Methods 0.000 description 2
- 230000007613 environmental effect Effects 0.000 description 2
- 102000013165 exonuclease Human genes 0.000 description 2
- 238000000605 extraction Methods 0.000 description 2
- 239000012634 fragment Substances 0.000 description 2
- BTCSSZJGUNDROE-UHFFFAOYSA-N gamma-aminobutyric acid Chemical compound NCCCC(O)=O BTCSSZJGUNDROE-UHFFFAOYSA-N 0.000 description 2
- 108010051015 glutathione-independent formaldehyde dehydrogenase Proteins 0.000 description 2
- HHLFWLYXYJOTON-UHFFFAOYSA-N glyoxylic acid Chemical compound OC(=O)C=O HHLFWLYXYJOTON-UHFFFAOYSA-N 0.000 description 2
- 238000009905 homogeneous catalytic hydrogenation reaction Methods 0.000 description 2
- 150000004678 hydrides Chemical class 0.000 description 2
- 238000007327 hydrogenolysis reaction Methods 0.000 description 2
- 229910052816 inorganic phosphate Inorganic materials 0.000 description 2
- 229910052943 magnesium sulfate Inorganic materials 0.000 description 2
- 235000019341 magnesium sulphate Nutrition 0.000 description 2
- 125000000325 methylidene group Chemical group [H]C([H])=* 0.000 description 2
- 238000002703 mutagenesis Methods 0.000 description 2
- 231100000350 mutagenesis Toxicity 0.000 description 2
- OFBQJSOFQDEBGM-UHFFFAOYSA-N n-pentane Natural products CCCCC OFBQJSOFQDEBGM-UHFFFAOYSA-N 0.000 description 2
- 230000010627 oxidative phosphorylation Effects 0.000 description 2
- 230000000737 periodic effect Effects 0.000 description 2
- 239000003208 petroleum Substances 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 230000004962 physiological condition Effects 0.000 description 2
- 108010010718 poly(3-hydroxyalkanoic acid) synthase Proteins 0.000 description 2
- 229920000070 poly-3-hydroxybutyrate Polymers 0.000 description 2
- 229920001707 polybutylene terephthalate Polymers 0.000 description 2
- 229910000160 potassium phosphate Inorganic materials 0.000 description 2
- 235000011009 potassium phosphates Nutrition 0.000 description 2
- 230000002265 prevention Effects 0.000 description 2
- 230000017854 proteolysis Effects 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 230000000284 resting effect Effects 0.000 description 2
- 239000004759 spandex Substances 0.000 description 2
- 238000012360 testing method Methods 0.000 description 2
- 239000012808 vapor phase Substances 0.000 description 2
- 108030006959 (+)-borneol dehydrogenases Proteins 0.000 description 1
- 108030006853 (-)-menthol dehydrogenases Proteins 0.000 description 1
- ZRHONLCTYUYMIQ-TZFCGSKZSA-N (2Z,4E)-2-aminomuconic acid Chemical compound OC(=O)C(/N)=C/C=C/C(O)=O ZRHONLCTYUYMIQ-TZFCGSKZSA-N 0.000 description 1
- PDGVYDMEIVBXAS-YFKPBYRVSA-N (8s)-8-amino-1,6-dioxacycloundecane-2,5,7,11-tetrone Chemical compound N[C@H]1CCC(=O)OC(=O)CCC(=O)OC1=O PDGVYDMEIVBXAS-YFKPBYRVSA-N 0.000 description 1
- 108030006845 (R)-4-hydroxyphenyllactate dehydrogenases Proteins 0.000 description 1
- 108030005055 (R)-aminopropanol dehydrogenases Proteins 0.000 description 1
- 108030005903 (R,R)-butanediol dehydrogenases Proteins 0.000 description 1
- 108030005057 (S,S)-butanediol dehydrogenases Proteins 0.000 description 1
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 1
- 108010011958 1,3-propanediol dehydrogenase Proteins 0.000 description 1
- 102100038794 17-beta-hydroxysteroid dehydrogenase type 6 Human genes 0.000 description 1
- GOCCREQJUBABAL-UHFFFAOYSA-N 2,2-dihydroxyacetic acid Chemical compound OC(O)C(O)=O GOCCREQJUBABAL-UHFFFAOYSA-N 0.000 description 1
- 108010015450 2,5-dioxovalerate dehydrogenase Proteins 0.000 description 1
- 108090000168 2-Oxoisovalerate Dehydrogenase (Acylating) Proteins 0.000 description 1
- 108030006988 2-alkyn-1-ol dehydrogenases Proteins 0.000 description 1
- 108700015926 2-hydroxy-3-oxopropionate reductases Proteins 0.000 description 1
- 108010027355 2-hydroxy-4-carboxymucconate-6-semialdehyde dehydrogenase Proteins 0.000 description 1
- AFENDNXGAFYKQO-UHFFFAOYSA-N 2-hydroxybutyric acid Chemical compound CCC(O)C(O)=O AFENDNXGAFYKQO-UHFFFAOYSA-N 0.000 description 1
- 108010025408 2-oxoaldehyde dehydrogenase (NAD+) Proteins 0.000 description 1
- 108010067707 2-oxoaldehyde dehydrogenase (NADP+) Proteins 0.000 description 1
- 108010021680 2-oxoglutarate decarboxylase Proteins 0.000 description 1
- 102000052553 3-Hydroxyacyl CoA Dehydrogenase Human genes 0.000 description 1
- 108700020831 3-Hydroxyacyl-CoA Dehydrogenase Proteins 0.000 description 1
- 125000004080 3-carboxypropanoyl group Chemical group O=C([*])C([H])([H])C([H])([H])C(O[H])=O 0.000 description 1
- 108030006979 3-hydroxy-2-methylbutyryl-CoA dehydrogenases Proteins 0.000 description 1
- 102100039358 3-hydroxyacyl-CoA dehydrogenase type-2 Human genes 0.000 description 1
- 108010039779 3-hydroxybenzyl-alcohol dehydrogenase Proteins 0.000 description 1
- 108010055682 3-hydroxybutyryl-CoA dehydrogenase Proteins 0.000 description 1
- 108030006717 3-hydroxypimeloyl-CoA dehydrogenases Proteins 0.000 description 1
- 108030001569 3-hydroxypropionate dehydrogenases Proteins 0.000 description 1
- 108010039636 3-isopropylmalate dehydrogenase Proteins 0.000 description 1
- 108030006351 3-methylbutanal reductases Proteins 0.000 description 1
- 108030006716 4-(hydroxymethyl)benzenesulfonate dehydrogenases Proteins 0.000 description 1
- 108030004443 4-formylbenzenesulfonate dehydrogenases Proteins 0.000 description 1
- 108010068231 4-hydroxybenzaldehyde dehydrogenase Proteins 0.000 description 1
- BAMBWCGEVIAQBF-CITAKDKDSA-N 4-hydroxybutyryl-CoA Chemical compound O[C@@H]1[C@H](OP(O)(O)=O)[C@@H](COP(O)(=O)OP(O)(=O)OCC(C)(C)[C@@H](O)C(=O)NCCC(=O)NCCSC(=O)CCCO)O[C@H]1N1C2=NC=NC(N)=C2N=C1 BAMBWCGEVIAQBF-CITAKDKDSA-N 0.000 description 1
- 108030006833 4-hydroxycyclohexanecarboxylate dehydrogenases Proteins 0.000 description 1
- 108030004477 4-hydroxyphenylacetaldehyde dehydrogenases Proteins 0.000 description 1
- 108030003557 4-oxoproline reductases Proteins 0.000 description 1
- 108030006715 6-hydroxyhexanoate dehydrogenases Proteins 0.000 description 1
- 108010016173 6-oxohexanoate dehydrogenase Proteins 0.000 description 1
- 108010092060 Acetate kinase Proteins 0.000 description 1
- 101100536799 Acinetobacter baylyi (strain ATCC 33305 / BD413 / ADP1) tgnE gene Proteins 0.000 description 1
- 229920001817 Agar Polymers 0.000 description 1
- 108010053754 Aldehyde reductase Proteins 0.000 description 1
- 102100027265 Aldo-keto reductase family 1 member B1 Human genes 0.000 description 1
- 102100026452 Aldo-keto reductase family 1 member B15 Human genes 0.000 description 1
- 108030001557 Allyl-alcohol dehydrogenases Proteins 0.000 description 1
- 102000006589 Alpha-ketoglutarate dehydrogenase Human genes 0.000 description 1
- 108020004306 Alpha-ketoglutarate dehydrogenase Proteins 0.000 description 1
- 108010065763 Aminobutyraldehyde dehydrogenase Proteins 0.000 description 1
- 108010006591 Apoenzymes Proteins 0.000 description 1
- 108010043325 Aryl-alcohol dehydrogenase Proteins 0.000 description 1
- 102000004625 Aspartate Aminotransferases Human genes 0.000 description 1
- 108010003415 Aspartate Aminotransferases Proteins 0.000 description 1
- 108010063172 Aspartate dehydrogenase Proteins 0.000 description 1
- 241000193830 Bacillus <bacterium> Species 0.000 description 1
- 108010045347 Benzaldehyde Dehydrogenase (NADP+) Proteins 0.000 description 1
- 108030006818 Benzyl-2-methyl-hydroxybutyrate dehydrogenases Proteins 0.000 description 1
- BVKZGUZCCUSVTD-UHFFFAOYSA-M Bicarbonate Chemical compound OC([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-M 0.000 description 1
- 108030004471 Butanal dehydrogenases Proteins 0.000 description 1
- FERIUCNNQQJTOY-UHFFFAOYSA-N Butyric acid Chemical compound CCCC(O)=O FERIUCNNQQJTOY-UHFFFAOYSA-N 0.000 description 1
- 101100351264 Candida albicans (strain SC5314 / ATCC MYA-2876) PDC11 gene Proteins 0.000 description 1
- 239000004215 Carbon black (E152) Substances 0.000 description 1
- 208000032544 Cicatrix Diseases 0.000 description 1
- 108010061190 Cinnamyl-alcohol dehydrogenase Proteins 0.000 description 1
- 102100034229 Citramalyl-CoA lyase, mitochondrial Human genes 0.000 description 1
- 241001112696 Clostridia Species 0.000 description 1
- 108020004705 Codon Proteins 0.000 description 1
- 108010024679 Coniferyl-alcohol dehydrogenase Proteins 0.000 description 1
- 229910020521 Co—Zn Inorganic materials 0.000 description 1
- 108030006973 Cyclohexane-1,2-diol dehydrogenases Proteins 0.000 description 1
- 108030006766 Cyclohexanol dehydrogenases Proteins 0.000 description 1
- 108030006984 Cyclopentanol dehydrogenases Proteins 0.000 description 1
- 102100037579 D-3-phosphoglycerate dehydrogenase Human genes 0.000 description 1
- 108030006712 D-arabinitol 2-dehydrogenases Proteins 0.000 description 1
- 108030001701 D-arabinitol 4-dehydrogenases Proteins 0.000 description 1
- 108010090581 D-arabinitol dehydrogenase Proteins 0.000 description 1
- 108010010795 D-iditol 2-dehydrogenase Proteins 0.000 description 1
- 108010001539 D-lactate dehydrogenase Proteins 0.000 description 1
- 108090000603 D-malate dehydrogenase (decarboxylating) Proteins 0.000 description 1
- 108030007033 D-pinitol dehydrogenases Proteins 0.000 description 1
- 102000007528 DNA Polymerase III Human genes 0.000 description 1
- 108010071146 DNA Polymerase III Proteins 0.000 description 1
- 238000000018 DNA microarray Methods 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 108030006776 Diethyl 2-methyl-3-oxosuccinate reductases Proteins 0.000 description 1
- 102100023319 Dihydrolipoyl dehydrogenase, mitochondrial Human genes 0.000 description 1
- 206010059866 Drug resistance Diseases 0.000 description 1
- 108700036530 EC 1.1.1.190 Proteins 0.000 description 1
- 108700036531 EC 1.1.1.191 Proteins 0.000 description 1
- 108700033362 EC 1.1.1.55 Proteins 0.000 description 1
- 108700035452 EC 1.2.1.58 Proteins 0.000 description 1
- 241001646716 Escherichia coli K-12 Species 0.000 description 1
- 108700039887 Essential Genes Proteins 0.000 description 1
- 241000195619 Euglena gracilis Species 0.000 description 1
- 241000874889 Euphilotes enoptes Species 0.000 description 1
- 102100029075 Exonuclease 1 Human genes 0.000 description 1
- 101150096236 FDH2 gene Proteins 0.000 description 1
- 108010042891 Farnesol dehydrogenase Proteins 0.000 description 1
- 206010016275 Fear Diseases 0.000 description 1
- 108030006724 Fluoren-9-ol dehydrogenases Proteins 0.000 description 1
- 108030004434 Fluoroacetaldehyde dehydrogenases Proteins 0.000 description 1
- 108030006714 Galactitol-1-phosphate 5-dehydrogenases Proteins 0.000 description 1
- 108010070600 Glucose-6-phosphate isomerase Proteins 0.000 description 1
- 108010035824 Glyceraldehyde 3-Phosphate Dehydrogenase (NADP+) Proteins 0.000 description 1
- 108010056007 Glyceraldehyde-3-Phosphate Dehydrogenase (NADP+)(Phosphorylating) Proteins 0.000 description 1
- 108090000094 Glyceraldehyde-3-Phosphate Dehydrogenase (Phosphorylating) Proteins 0.000 description 1
- 102000003679 Glyceraldehyde-3-Phosphate Dehydrogenase (Phosphorylating) Human genes 0.000 description 1
- 101710195364 Glycerol-1-phosphate dehydrogenase [NAD(P)+] Proteins 0.000 description 1
- 108010090367 Glycerol-3-Phosphate Dehydrogenase (NAD+) Proteins 0.000 description 1
- 101710114313 Glycerol-3-phosphate dehydrogenase [NAD(P)+] Proteins 0.000 description 1
- 108010068673 Glycolaldehyde dehydrogenase Proteins 0.000 description 1
- 108010038519 Glyoxylate reductase Proteins 0.000 description 1
- 241000606768 Haemophilus influenzae Species 0.000 description 1
- 102100039894 Hemoglobin subunit delta Human genes 0.000 description 1
- 108030006983 Hexadecanol dehydrogenases Proteins 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 101000918264 Homo sapiens Exonuclease 1 Proteins 0.000 description 1
- UFHFLCQGNIYNRP-UHFFFAOYSA-N Hydrogen Chemical compound [H][H] UFHFLCQGNIYNRP-UHFFFAOYSA-N 0.000 description 1
- 108030006987 Hydroxycyclohexanecarboxylate dehydrogenases Proteins 0.000 description 1
- 102000004286 Hydroxymethylglutaryl CoA Reductases Human genes 0.000 description 1
- 108090000895 Hydroxymethylglutaryl CoA Reductases Proteins 0.000 description 1
- 108030006790 Hydroxyphenylpyruvate reductases Proteins 0.000 description 1
- 108010059247 Hydroxypyruvate reductase Proteins 0.000 description 1
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 1
- 102100030764 Inactive L-threonine 3-dehydrogenase, mitochondrial Human genes 0.000 description 1
- 102100034343 Integrase Human genes 0.000 description 1
- 108020003285 Isocitrate lyase Proteins 0.000 description 1
- 108030006269 Isocitrate-homoisocitrate dehydrogenases Proteins 0.000 description 1
- 108010068679 Isopiperitenol dehydrogenase Proteins 0.000 description 1
- OYIFNHCXNCRBQI-BYPYZUCNSA-M L-2-aminoadipate(1-) Chemical compound [O-]C(=O)[C@@H]([NH3+])CCCC([O-])=O OYIFNHCXNCRBQI-BYPYZUCNSA-M 0.000 description 1
- 108010009384 L-Iditol 2-Dehydrogenase Proteins 0.000 description 1
- 108030001699 L-arabinitol 2-dehydrogenases Proteins 0.000 description 1
- 108010064700 L-arabinitol 4-dehydrogenase Proteins 0.000 description 1
- KABXUUFDPUOJMW-BYPYZUCNSA-N L-glutamic 5-semialdehyde Chemical compound OC(=O)[C@@H](N)CCC=O KABXUUFDPUOJMW-BYPYZUCNSA-N 0.000 description 1
- 108010081263 L-glycol dehydrogenase Proteins 0.000 description 1
- 108030003446 L-threonate 3-dehydrogenases Proteins 0.000 description 1
- 108010043075 L-threonine 3-dehydrogenase Proteins 0.000 description 1
- 108030003529 L-xylose 1-dehydrogenases Proteins 0.000 description 1
- 108010057617 Lactaldehyde dehydrogenase Proteins 0.000 description 1
- 102100030931 Ladinin-1 Human genes 0.000 description 1
- 102100025357 Lipid-phosphate phosphatase Human genes 0.000 description 1
- 108010011927 Long-chain-alcohol dehydrogenase Proteins 0.000 description 1
- 108010058996 Long-chain-aldehyde dehydrogenase Proteins 0.000 description 1
- 108060001084 Luciferase Proteins 0.000 description 1
- 239000005089 Luciferase Substances 0.000 description 1
- 108010042544 Malate Dehydrogenase (NADP+) Proteins 0.000 description 1
- 108020004687 Malate Synthase Proteins 0.000 description 1
- 101710155796 Malate:quinone oxidoreductase Proteins 0.000 description 1
- 241001529626 Malica Species 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 108090000428 Mannitol-1-phosphate 5-dehydrogenases Proteins 0.000 description 1
- 241001465754 Metazoa Species 0.000 description 1
- 241000204031 Mycoplasma Species 0.000 description 1
- WHNWPMSKXPGLAX-UHFFFAOYSA-N N-Vinyl-2-pyrrolidone Chemical compound C=CN1CCCC1=O WHNWPMSKXPGLAX-UHFFFAOYSA-N 0.000 description 1
- 230000004988 N-glycosylation Effects 0.000 description 1
- 108010024592 Octanol dehydrogenase Proteins 0.000 description 1
- 108030005095 Omega-hydroxydecanoate dehydrogenases Proteins 0.000 description 1
- 102000004316 Oxidoreductases Human genes 0.000 description 1
- 108090000854 Oxidoreductases Proteins 0.000 description 1
- 101150050255 PDC1 gene Proteins 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 108010055471 Phenylacetaldehyde dehydrogenase Proteins 0.000 description 1
- 108700023175 Phosphate acetyltransferases Proteins 0.000 description 1
- 108010038555 Phosphoglycerate dehydrogenase Proteins 0.000 description 1
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 1
- 102100032709 Potassium-transporting ATPase alpha chain 2 Human genes 0.000 description 1
- 108030005906 Propanediol-phosphate dehydrogenases Proteins 0.000 description 1
- 108010026552 Proteome Proteins 0.000 description 1
- 108010083204 Proton Pumps Proteins 0.000 description 1
- 108010053763 Pyruvate Carboxylase Proteins 0.000 description 1
- 108010092799 RNA-directed DNA polymerase Proteins 0.000 description 1
- 102000018120 Recombinases Human genes 0.000 description 1
- 108010091086 Recombinases Proteins 0.000 description 1
- 238000005602 Reppe reaction Methods 0.000 description 1
- 102000018822 Retinal Dehydrogenase Human genes 0.000 description 1
- 108010027691 Retinal dehydrogenase Proteins 0.000 description 1
- 108030003457 Ribitol-5-phosphate 2-dehydrogenases Proteins 0.000 description 1
- 241000282849 Ruminantia Species 0.000 description 1
- 101100101631 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) UGA2 gene Proteins 0.000 description 1
- 108010064340 Salicylaldehyde dehydrogenase Proteins 0.000 description 1
- 241000607142 Salmonella Species 0.000 description 1
- 108030007031 Sorbitol-6-phosphate 2-dehydrogenases Proteins 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- 108010084086 Succinate-Semialdehyde Dehydrogenase Proteins 0.000 description 1
- 102000005566 Succinate-Semialdehyde Dehydrogenase Human genes 0.000 description 1
- 239000004433 Thermoplastic polyurethane Substances 0.000 description 1
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 description 1
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 description 1
- 108010044234 Vanillin dehydrogenase Proteins 0.000 description 1
- CIGVKKBXYHSHDK-UHFFFAOYSA-N [Na].[Cu]=O Chemical compound [Na].[Cu]=O CIGVKKBXYHSHDK-UHFFFAOYSA-N 0.000 description 1
- ODXRWHGACRVUPD-UHFFFAOYSA-N [O-2].[Th+4].[Co+2].[Ni+2].[O-2].[O-2].[O-2] Chemical compound [O-2].[Th+4].[Co+2].[Ni+2].[O-2].[O-2].[O-2] ODXRWHGACRVUPD-UHFFFAOYSA-N 0.000 description 1
- GFCDJPPBUCXJSC-UHFFFAOYSA-N [O-2].[Zn+2].[Cu]=O Chemical compound [O-2].[Zn+2].[Cu]=O GFCDJPPBUCXJSC-UHFFFAOYSA-N 0.000 description 1
- 108091000039 acetoacetyl-CoA reductase Proteins 0.000 description 1
- 125000002777 acetyl group Chemical group [H]C([H])([H])C(*)=O 0.000 description 1
- 230000021736 acetylation Effects 0.000 description 1
- 238000006640 acetylation reaction Methods 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 125000002252 acyl group Chemical group 0.000 description 1
- 108010092860 acylglycerone-phosphate reductase Proteins 0.000 description 1
- 230000000996 additive effect Effects 0.000 description 1
- 210000001789 adipocyte Anatomy 0.000 description 1
- 238000005377 adsorption chromatography Methods 0.000 description 1
- 230000004103 aerobic respiration Effects 0.000 description 1
- 239000008272 agar Substances 0.000 description 1
- 108010058033 alcohol dehydrogenase (NADP+) Proteins 0.000 description 1
- 108010048916 alcohol dehydrogenase (acceptor) Proteins 0.000 description 1
- 150000001298 alcohols Chemical class 0.000 description 1
- 229940024606 amino acid Drugs 0.000 description 1
- 108010066436 aryl-alcohol dehydrogenase (NADP+) Proteins 0.000 description 1
- 229940009098 aspartate Drugs 0.000 description 1
- 230000004888 barrier function Effects 0.000 description 1
- 108010060169 benzaldehyde dehydrogenase (NAD+) Proteins 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 230000036983 biotransformation Effects 0.000 description 1
- 230000005587 bubbling Effects 0.000 description 1
- 239000000872 buffer Substances 0.000 description 1
- 235000019437 butane-1,3-diol Nutrition 0.000 description 1
- 108010057307 butanol dehydrogenase Proteins 0.000 description 1
- 229930188620 butyrolactone Natural products 0.000 description 1
- 125000004432 carbon atom Chemical group C* 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 238000010531 catalytic reduction reaction Methods 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 230000019522 cellular metabolic process Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- PSJHTNRVTMXYER-UHFFFAOYSA-N chromium;copper;oxygen(2-) Chemical compound [O-2].[Cr].[Cu] PSJHTNRVTMXYER-UHFFFAOYSA-N 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 230000001427 coherent effect Effects 0.000 description 1
- 239000003086 colorant Substances 0.000 description 1
- 230000000295 complement effect Effects 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 239000000470 constituent Substances 0.000 description 1
- 238000010924 continuous production Methods 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 238000002425 crystallisation Methods 0.000 description 1
- 230000008025 crystallization Effects 0.000 description 1
- 238000012786 cultivation procedure Methods 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 210000000172 cytosol Anatomy 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 230000003413 degradative effect Effects 0.000 description 1
- 238000009795 derivation Methods 0.000 description 1
- 238000001212 derivatisation Methods 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 235000014113 dietary fatty acids Nutrition 0.000 description 1
- BEPAFCGSDWSTEL-UHFFFAOYSA-N dimethyl malonate Chemical compound COC(=O)CC(=O)OC BEPAFCGSDWSTEL-UHFFFAOYSA-N 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 238000000909 electrodialysis Methods 0.000 description 1
- 238000010828 elution Methods 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 230000009483 enzymatic pathway Effects 0.000 description 1
- 238000001952 enzyme assay Methods 0.000 description 1
- 150000002118 epoxides Chemical class 0.000 description 1
- 230000032050 esterification Effects 0.000 description 1
- 238000005886 esterification reaction Methods 0.000 description 1
- 150000002170 ethers Chemical class 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 238000002270 exclusion chromatography Methods 0.000 description 1
- 239000000284 extract Substances 0.000 description 1
- 229930195729 fatty acid Natural products 0.000 description 1
- 239000000194 fatty acid Substances 0.000 description 1
- 150000004665 fatty acids Chemical class 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 239000012530 fluid Substances 0.000 description 1
- 235000013305 food Nutrition 0.000 description 1
- 108010045736 formate dehydrogenase (NADP+) Proteins 0.000 description 1
- 101150043302 gabD gene Proteins 0.000 description 1
- 108010029645 galactitol 2-dehydrogenase Proteins 0.000 description 1
- 229940065734 gamma-aminobutyrate Drugs 0.000 description 1
- 238000004817 gas chromatography Methods 0.000 description 1
- 239000000499 gel Substances 0.000 description 1
- 238000012246 gene addition Methods 0.000 description 1
- 238000003208 gene overexpression Methods 0.000 description 1
- JFCQEDHGNNZCLN-UHFFFAOYSA-N glutaric acid Chemical compound OC(=O)CCCC(O)=O JFCQEDHGNNZCLN-UHFFFAOYSA-N 0.000 description 1
- 108020004445 glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 description 1
- 102000006602 glyceraldehyde-3-phosphate dehydrogenase Human genes 0.000 description 1
- 108010000361 glycerol 2-dehydrogenase (NADP+) Proteins 0.000 description 1
- 150000002334 glycols Chemical class 0.000 description 1
- 108090000471 glyoxylate dehydrogenase (acylating) Proteins 0.000 description 1
- 108010091048 glyoxylate reductase (NADP+) Proteins 0.000 description 1
- 108090001018 hexadecanal dehydrogenase (acylating) Proteins 0.000 description 1
- 108010017999 homoisocitrate dehydrogenase Proteins 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 230000006801 homologous recombination Effects 0.000 description 1
- 229930195733 hydrocarbon Natural products 0.000 description 1
- 150000002430 hydrocarbons Chemical class 0.000 description 1
- 230000007062 hydrolysis Effects 0.000 description 1
- 238000006460 hydrolysis reaction Methods 0.000 description 1
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 1
- 239000012535 impurity Substances 0.000 description 1
- 108010093674 indanol dehydrogenase Proteins 0.000 description 1
- 239000000976 ink Substances 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 238000011835 investigation Methods 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 108010029918 isocitrate dehydrogenase (NADP+) Proteins 0.000 description 1
- 108091022889 lactaldehyde reductase Proteins 0.000 description 1
- 238000009630 liquid culture Methods 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 108090000286 malate dehydrogenase (decarboxylating) Proteins 0.000 description 1
- FPYJFEHAWHCUMM-UHFFFAOYSA-N maleic anhydride Chemical compound O=C1OC(=O)C=C1 FPYJFEHAWHCUMM-UHFFFAOYSA-N 0.000 description 1
- 229940099596 manganese sulfate Drugs 0.000 description 1
- 239000011702 manganese sulphate Substances 0.000 description 1
- 235000007079 manganese sulphate Nutrition 0.000 description 1
- SQQMAOCOWKFBNP-UHFFFAOYSA-L manganese(II) sulfate Chemical compound [Mn+2].[O-]S([O-])(=O)=O SQQMAOCOWKFBNP-UHFFFAOYSA-L 0.000 description 1
- 108010007922 mannitol 2-dehydrogenase (NADP) Proteins 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 238000005374 membrane filtration Methods 0.000 description 1
- 125000005395 methacrylic acid group Chemical group 0.000 description 1
- ZIYVHBGGAOATLY-UHFFFAOYSA-N methylmalonic acid Chemical compound OC(=O)C(C)C(O)=O ZIYVHBGGAOATLY-UHFFFAOYSA-N 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- MQBCDKMPXVYCGO-FQBKTPCVSA-N mycothiol Chemical compound CC(=O)N[C@@H](CS)C(=O)N[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@H](O)[C@@H](O)[C@H](O)[C@H]1O MQBCDKMPXVYCGO-FQBKTPCVSA-N 0.000 description 1
- 108010074581 mycothiol Proteins 0.000 description 1
- MQBCDKMPXVYCGO-UHFFFAOYSA-N mycothiol Natural products CC(=O)NC(CS)C(=O)NC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(O)C1O MQBCDKMPXVYCGO-UHFFFAOYSA-N 0.000 description 1
- 108010050450 myo-inositol 2-dehydrogenase Proteins 0.000 description 1
- IJDNQMDRQITEOD-UHFFFAOYSA-N n-butane Chemical compound CCCC IJDNQMDRQITEOD-UHFFFAOYSA-N 0.000 description 1
- 239000003345 natural gas Substances 0.000 description 1
- 239000002773 nucleotide Substances 0.000 description 1
- 125000003729 nucleotide group Chemical group 0.000 description 1
- 150000002894 organic compounds Chemical class 0.000 description 1
- 230000001151 other effect Effects 0.000 description 1
- 238000012261 overproduction Methods 0.000 description 1
- 238000006213 oxygenation reaction Methods 0.000 description 1
- 239000003973 paint Substances 0.000 description 1
- 238000005373 pervaporation Methods 0.000 description 1
- 239000008363 phosphate buffer Substances 0.000 description 1
- 108010085336 phosphoribosyl-AMP cyclohydrolase Proteins 0.000 description 1
- 230000000865 phosphorylative effect Effects 0.000 description 1
- 230000008560 physiological behavior Effects 0.000 description 1
- 230000035790 physiological processes and functions Effects 0.000 description 1
- 229920000903 polyhydroxyalkanoate Polymers 0.000 description 1
- 229920000909 polytetrahydrofuran Polymers 0.000 description 1
- 239000011591 potassium Substances 0.000 description 1
- 229910052700 potassium Inorganic materials 0.000 description 1
- 239000000843 powder Substances 0.000 description 1
- 230000019525 primary metabolic process Effects 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 235000019833 protease Nutrition 0.000 description 1
- 108020001775 protein parts Proteins 0.000 description 1
- 108010052174 pyruvate dehydrogenase (NADP+) Proteins 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 230000001172 regenerating effect Effects 0.000 description 1
- 102000037983 regulatory factors Human genes 0.000 description 1
- 108091008025 regulatory factors Proteins 0.000 description 1
- 230000008672 reprogramming Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 229920005989 resin Polymers 0.000 description 1
- 239000011347 resin Substances 0.000 description 1
- 108010035291 retinol dehydrogenase Proteins 0.000 description 1
- 238000001223 reverse osmosis Methods 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 108010020957 ribitol 2-dehydrogenase Proteins 0.000 description 1
- 238000007363 ring formation reaction Methods 0.000 description 1
- 238000005096 rolling process Methods 0.000 description 1
- 231100000241 scar Toxicity 0.000 description 1
- 230000037387 scars Effects 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 239000011734 sodium Substances 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- 159000000000 sodium salts Chemical class 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 238000006467 substitution reaction Methods 0.000 description 1
- 230000008093 supporting effect Effects 0.000 description 1
- 230000001629 suppression Effects 0.000 description 1
- 230000004083 survival effect Effects 0.000 description 1
- 230000009897 systematic effect Effects 0.000 description 1
- 230000009885 systemic effect Effects 0.000 description 1
- 125000003718 tetrahydrofuranyl group Chemical group 0.000 description 1
- 229920006344 thermoplastic copolyester Polymers 0.000 description 1
- 229920002803 thermoplastic polyurethane Polymers 0.000 description 1
- 229960000187 tissue plasminogen activator Drugs 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 238000011282 treatment Methods 0.000 description 1
- 238000000108 ultra-filtration Methods 0.000 description 1
- 239000002699 waste material Substances 0.000 description 1
- 239000011787 zinc oxide Substances 0.000 description 1
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/02—Preparation of oxygen-containing organic compounds containing a hydroxy group
- C12P7/04—Preparation of oxygen-containing organic compounds containing a hydroxy group acyclic
- C12P7/18—Preparation of oxygen-containing organic compounds containing a hydroxy group acyclic polyhydric
-
- B—PERFORMING OPERATIONS; TRANSPORTING
- B01—PHYSICAL OR CHEMICAL PROCESSES OR APPARATUS IN GENERAL
- B01D—SEPARATION
- B01D3/00—Distillation or related exchange processes in which liquids are contacted with gaseous media, e.g. stripping
- B01D3/001—Processes specially adapted for distillation or rectification of fermented solutions
- B01D3/002—Processes specially adapted for distillation or rectification of fermented solutions by continuous methods
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0006—Oxidoreductases (1.) acting on CH-OH groups as donors (1.1)
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02E—REDUCTION OF GREENHOUSE GAS [GHG] EMISSIONS, RELATED TO ENERGY GENERATION, TRANSMISSION OR DISTRIBUTION
- Y02E50/00—Technologies for the production of fuel of non-fossil origin
- Y02E50/10—Biofuels, e.g. bio-diesel
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Health & Medical Sciences (AREA)
- Biotechnology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Biomedical Technology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Método para fabricar un producto posterior de 4-hidroxibutirato o 1,4-butanodiol, que comprende: (a) cultivar en fase de crecimiento exponencial en una cantidad suficiente de nutrientes y medios un microorganismo no natural que comprende perturbación de los genes adhE, ldhA, pflAB y mdh, en el que dicho microorganismo expresa al menos un ácido nucleico exógeno que codifica para una enzima en una ruta biosintética de 4-hidroxibutirato o 1,4-butanodiol en cantidades suficientes para producir 4- hidroxibutirato o 1,4-butanodiol, codificando dicho al menos un ácido nucleico exógeno para 4- hidroxibutanoato deshidrogenasa, semialdehído succínico deshidrogenasa independiente de CoA, succinil- CoA sintetasa, semialdehído succínico deshidrogenasa dependiente de CoA, 4-hidroxibutirato:CoA transferasa, glutamato:semialdehído succínico transaminasa, glutamato descarboxilasa, aldehído deshidrogenasa independiente de CoA, aldehído deshidrogenasa dependiente de CoA o alcohol deshidrogenasa, y en el que dicho microorganismo produce 4-hidroxibutirato o 1,4-butanodiol; (b) aislar opcionalmente 4-hidroxibutirato o 1,4-butanodiol; y (c) convertir químicamente 4-hidroxibutirato o 1,4-butanodiol en el producto posterior.
Description
DESCRIPCION
Metodos y organismos para la produccion de 1,4-butanodiol acoplada al crecimiento
Antecedentes de la invencion
La presente invencion se refiere generalmente a metodos para fabricar productos posteriores de 4-hidroxibutirato o 1.4- butanodiol.
1.4- Butanodiol (BDO) es un dialcohol de cuatro atomos de carbono que actualmente se fabrica exclusivamente a traves de varias rutas petroqmmicas. El BDO forma parte de una familia de disolventes e intermedios polimericos de gran volumen que incluye gamma-butirolactona (GBL), tetrahidrofurano (THF), pirrolidona, N-metilpirrolidona (NMP), y N-vinil-pirrolidona. La oportunidad global de mercado para esta familia excede los 4.000 millones de $.
Se producen globalmente al ano aproximadamente 2.500 millones de libras de BDO con un crecimiento anual del 4 5% con un precio reciente de venta que vana de 1,00 a 1,20 $/lb. La demanda de BDO se debe en gran parte a su uso como compuesto intermedio de las resinas plasticas de tereftalato de polibutileno (PBT), poliuretanos termoplasticos y eteres de copoliester. BDO sirve tambien como precursor principal de THF, que se emplea como compuesto intermedio para los copolfmeros de poli(tetrametilenglicol) PTMEG necesarios para la produccion de lycra y spandex. Se producen globalmente aproximadamente 700 millones de lb de THF por ano con una tasa anual de crecimiento de aproximadamente el 6%. Un porcentaje significativo del crecimiento (>30%) de BDO y THF se produce en Asia (China e India). GBL es actualmente un producto de un volumen mas pequeno (400 millones de lb/ano) que tiene numerosas aplicaciones como disolvente, como un aditivo para tintas, pinturas, y colorantes, asf como el precursor primario de derivados de pirrolidona tales como NMP.
Los procesos convencionales para la smtesis de BDO usan materias primas petroqmmicas como sus materiales de partida. Por ejemplo, se hace reaccionar acetileno con 2 moleculas de formaldehndo en la reaccion de smtesis de Reppe (Kroschwitz y Grant, Encyclopedia of Chem. Tech., John Wiley and Sons, Inc., Nueva York (1999)), seguido por hidrogenacion catalttica para formar 1,4-butanodiol. Se ha estimado que el 90% del acetileno producido en los Estados Unidos se consume en la produccion de butanodiol. Como alternativa, se puede formar por esterificacion e hidrogenacion catalttica de anhndrido maleico, que se deriva de butano. Posteriormente, el butanodiol se puede transformar adicionalmente; por ejemplo, mediante oxidacion de la Y-butirolactona, que se puede convertir adicionalmente en pirrolidona y N-metil-pirrolidona, o hidrogenolisis del tetrahidrofurano (figura 1). Estos compuestos tienen usos variados como compuestos intermedios, disolventes, y aditivos polimericos, y tienen un mercado combinado de casi 2000 millones de lb/ano.
Las soluciones convencionales basadas en materias primas de hidrocarburos utilizan metano para producir formaldetndo. Por tanto, un gran porcentaje de la produccion comercial de BDO se basa en metano como material de partida. La produccion de acetileno se basa tambien en materiales de partida basados en petroleo (vease la figura 1). Por tanto, los costes de produccion de BDO fluctuan con el precio del petroleo y el gas natural.
El documento JPS62285779 (A) se refiere a una cepa bacteriana que pertenece al genero Bacillus (cepa C-105 (FERM P-8788)) y a la produccion de 1,4-butanodiol usando la misma.
Es deseable desarrollar un metodo para la produccion de estos productos qmmicos por medios alternativos que sustituyan no solo materias primas basadas en petroleo por renovables, y usen tambien menos energfa y procesos de menor coste de inmovilizado. El Departamento de energfa de Estados Unidos ha propuesto los 1,4-diacidos, y particularmente el acido succmico, como compuestos intermedios clave producidos biologicamente para la fabricacion de la familia de productos del butanodiol (DOE Report, "Top Value-Added Chemicals from Biomass” , 2004). Sin embargo, el acido succmico es muy costoso de aislar y purificar y requiere elevadas temperaturas y presiones para la reduccion catalftica a butanodiol.
Por tanto, existe una necesidad de medios alternativos para producir eficazmente cantidades comerciales de 1,4-butanodiol y sus precursores qmmicos. La presente invencion satisface esta necesidad y proporciona tambien ventajas relacionadas.
Sumario de la invencion
La invencion se define mediante las reivindicaciones.
Breve descripcion de los dibujos
El archivo de patente o de solicitud contiene al menos un dibujo ejecutado a color. La Oficina proporcionara copias de esta patente o publicacion de solicitud de patente con dibujos a color, previa peticion y pago de la tasa necesaria.
La Figura 1 es un diagrama esquematico que muestra un punto de entrada del acido 4-hidroxibutanoico (4-HB) en la lmea de producto de la familia de productos qmmicos del 1,4-butanodiol (BDO), y la comparacion con las rutas de smtesis qmmica de las materias primas petroqmmicas. Las flechas negras solidas muestran las rutas de smtesis qmmica; las flechas azules punteadas muestran una ruta biosintetica de 4-HB y las posteriores etapas de conversion de la familia de productos qmmicos de BDO.
La Figura 2 es un diagrama esquematico que muestra las rutas bioqmmicas de produccion de 4-hidroxibutirato (4-HB) y Y-butirolactona (GBL). Los enzimas que catalizan las reacciones biosinteticas de 4-HB son: (1) semialdelmdo succmico deshidrogenasa independiente de CoA; (2) succinil-CoA sintetasa; (3) semialdelmdo succmico deshidrogenasa dependiente de CoA; (4) glutamato: semialdelmdo succmico transaminasa; (5) glutamato decarboxilasa; (6) 4-hidroxibutanoato deshidrogenasa. La conversion (7) corresponde a una reaccion no enzimatica espontanea, que convierte 4-HB en GBL.
La Figura 3 es un diagrama esquematico que muestra la smtesis qmmica de 1,4-butanodiol (BDO) y los productos posteriores Y-butirolactona (GBL), tetrahidrofurano (THF) y algunas pirrolidonas.
La Figura 4 es un diagrama de flujo de proceso esquematico de los bioprocesos de produccion de Y-butirolactona. El panel (a) ilustra la fermentacion con alimentacion discontinua con separacion discontinua y el panel (b) ilustra la fermentacion con alimentacion discontinua con separacion continua.
La Figura 5 muestra la envolvente de produccion teorica de una cepa disenada mediante OptKnock comparada con una cepa tfpica de produccion no acoplada al crecimiento. Senalar que las trayectorias evolutivas potenciales de la cepa OptKnock son fundamentalmente diferentes en que conduce a un fenotipo de alta produccion.
La Figura 6 muestra las rutas bioqmmicas que conducen hasta el 1,4-butanodiol. 1) semialdelmdo succmico deshidrogenasa independiente de CoA; 2) succinil-CoA sintetasa; 3) semialdelmdo succmico deshidrogenasa dependiente de CoA; 4) glutamato: semialdehfdo succmico transaminasa; 5) glutamato decarboxilasa; 6) 4-hidroxibutanoato deshidrogenasa; 7) 4-hidroxibutiril CoA:acetil-CoA transferasa; 8) aldehfdo deshidrogenasa; 9) alcohol deshidrogenasa.
La Figura 7 muestra los lfmites de solucion de la tasa de crecimiento anaerobio frente al rendimiento de BDO para una cepa de E. coli que tiene las rutas de produccion de BDO mostradas y las desactivaciones genicas previstas por OptKnock en la Figura 5 suponiendo a) la irreversibilidad de la PEP carboxiquinasa y b) la reversibilidad de la PEP carboxiquinasa. Se supuso una tasa de captacion de glucosa basa de 20 mmol/gDW/h junto con un requerimiento de mantenimiento de ATP no asociado al crecimiento de 7,6 mmol/gDW/h.
La Figura 8 muestra una representacion grafica del metabolismo central de E. coli.
La Figura 9 muestra los lfmites de solucion de la tasa de crecimiento anaerobio frente al rendimiento de BDO para una cepa de E. coli que tiene las rutas de produccion de BDO mostradas y las desactivaciones genicas previstas por OptKnock en la Figura 5 suponiendo la reversibilidad de la PEP carboxiquinasa. Se supuso una tasa de captacion de glucosa basa de 20 mmol/gDW/h junto con un requerimiento de mantenimiento de ATP no asociado al crecimiento de 7,6 mmol/gDW/h.
Descripcion detallada de la invencion
Esta invencion se refiere al diseno y a la produccion de microorganismos que no se producen naturalmente que tienen capacidades de produccion biosinteticas para el acido 4-hidroxibutanoico (4-HB), Y-butirolactona y 1,4-butanodiol. La invencion utiliza modelos estequiometricos in silico del metabolismo de Escherichia coli que identifican disenos metabolicos para la produccion biosintetica de acido 4-hidroxibutanoico (4-HB) y 1,4-butanodiol (BDO). Los resultados descritos en el presente documento indican que se pueden disenar y conseguir mediante ingeniena genetica recombinante rutas metabolicas para conseguir la biosmtesis de 4-HB y de los productos posteriores tales como 1,4-butanodiol en Escherichia coli y otras celulas u organismos. La produccion biosintetica de 4-HB, por ejemplo, para los disenos in silico se puede confirmar mediante la construccion de cepas que tienen el genotipo metabolico disenado. Estas celulas u organismos disenados metabolicamente mediante ingeniena genetica se pueden someter a evolucion adaptativa para aumentar adicionalmente la biosmtesis de 4-HB, incluyendo en condiciones que se aproximan al crecimiento maximo teorico.
La invencion se refiere ademas a estrategias de diseno mediante ingeniena metabolica para obtener rendimientos elevados de 1,4-butanodiol (BDO) en Escherichia coli (veanse los Ejemplos V-VII). Como se divulga en el presente documento, se empleo un modelo estequiometrico a escala genomica del metabolismo de E. coli utilizando la infraestructura informatica de optimizacion de dos niveles OptKnock para identificar estrategias informaticas con multiples inactivaciones genicas. Las deleciones se introducen de tal manera que se reduce la redundancia en la red con el efecto en ultima instancia de acoplar el crecimiento a la produccion de BDO en la red. Las caractensticas de produccion de BDO acopladas al crecimiento de las cepas disenadas las convierten en geneticamente estables y adecuadas para bioprocesos continuos. Se identificaron estrategias de diseno de cepas suponiendo la adicion de
capacidades de reaccion no naturales en E. coli que conducen a una ruta metabolica desde el succinato semialdetndo a BDO. De los cientos de estrategias identificadas por OptKnock, surgio un diseno que satisface multiples criterios. Este diseno, que utiliza la eliminacion de adhE, IdhA, mdh, aspA, y pflAB, 1) conduce a un elevado rendimiento de BDO previsto a crecimiento maximo, 2) requirio un numero de desactivaciones razonable, 3) no tuvo un efecto perjudicial sobre el rendimiento teorico maximo de BDO, 4) consiguio un acoplamiento fuerte entre la produccion de b Do y el crecimiento celular, y 5) era solido con respecto a la supuesta irreversibilidad o reversibilidad de la PEP carboxiquinasa. Se divulgan tambien en el presente documento metodos para el ensayo experimental de disenos de cepas y su evolucion hacia el crecimiento maximo teorico.
En determinadas realizaciones, las caractensticas de la biosmtesis de 4-HB de las cepas disenadas las convierten en geneticamente estables y adecuadas para bioprocesos continuos. Se identificaron estrategias de diseno de cepas independientes con la incorporacion de capacidades de reaccion no naturales o heterologas diferente en E. coli que conducen a rutas metabolicas productoras de 4-HB y 1,4-butanodiol derivados de la semialdetndo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa y semialdetndo succmico deshidrogenasa dependiente de CoA, o glutamato:semialdetndo succmico transaminasa. Se identificaron disenos metabolicos in silico que dieron como resultado la biosmtesis de 4-HB en especies de E. coli y levadura procedentes de cada una de estas rutas metabolicas. El compuesto intermedio de 1,4-butanodiol, Y-butirolactona, se puede generar en cultivo mediante ciclacion espontanea en condiciones de pH<7,5, particularmente en condiciones acidas, tales como por debajo de pH 5,5, por ejemplo, pH<7, pH<6,5, pH<6, y particularmente a pH<5,5 o inferior.
Las cepas identificadas mediante el componente de calculo de la plataforma pueden llevarse a la produccion real genomanipulando cualquiera de las alteraciones metabolicas previstas que conducen a la produccion biosintetica de 4-HB, 1,4-butanodiol u otros compuestos intermedios y/o productos posteriores. En otra realizacion mas, las cepas que presentan produccion biosintetica de estos compuestos pueden someterse adicionalmente a evolucion adaptativa para aumentar ademas la biosmtesis del producto. Los niveles de rendimiento de la biosmtesis del producto despues de la evolucion adaptativa tambien se pueden predecir mediante el componente de calculo del sistema.
Tal como se usa en el presente documento, se pretende que el termino "que no se produce naturalmente", cuando se usa en referencia a un organismo microbiano o microorganismo de la divulgacion signifique que el organismo microbiano tiene al menos una alteracion genetica que no se encuentra normalmente en una cepa que se produce naturalmente de las especies citadas, incluyendo las cepas silvestres de las especies citadas. Las alteraciones geneticas incluyen, por ejemplo, modificaciones que introducen acidos nucleicos expresables que codifican polipeptidos metabolicos, otras adiciones de acidos nucleicos, deleciones de acidos nucleicos y/u otras perturbaciones funcionales del material genetico microbiano. Dicha modificacion incluye, por ejemplo, regiones codificantes y fragmentos funcionales de las mismas, de polipeptidos heterologos, homologos o heterologos y homologos de las especies citadas. Las modificaciones adicionales incluyen, por ejemplo, regiones reguladoras no codificantes en las que las modificaciones alteran la expresion de un gen u operon. Los polipeptidos metabolicos ilustrativos incluyen enzimas comprendidas en la ruta biosintetica de 4-HB y enzimas comprendidas en la ruta biosintetica de la familia de compuestos de BDO.
Una modificacion metabolica se refiere a una reaccion bioqmmica que esta alterada a partir de su estado que se produce naturalmente. Por tanto, los microorganismos que no se producen naturalmente tienen modificaciones geneticas en acidos nucleicos que codifican polipeptidos metabolicos o, fragmentos funcionales de los mismos. Se describen modificaciones metabolicas ilustrativas adicionalmente a continuacion para E. coli y organismos microbianos de levaduras.
Tal como se usa en el presente documento, se pretende que el termino "aislado", cuando se usa en referencia a un organismo microbiano, signifique un organismo que esta sustancialmente exento de al menos un componente que posee el organismo microbiano natural citado. El termino incluye un organismo microbiano al que se eliminan alguno o todos los componentes que se encuentran en su entorno natural. El termino incluye tambien un organismo microbiano al que se eliminan alguno o todos los componentes que el organismo microbiano tiene en entornos que no se producen naturalmente. Por tanto, un organismo microbiano aislado esta parcial o completamente separado de otras sustancias que se encuentran en la naturaleza o a medida que crece, se almacena o subsiste en entornos que no se producen naturalmente. Los ejemplos espedficos de organismos microbianos aislados incluyen microbios parcialmente puros, microbios sustancialmente puros y microbios cultivados en un medio que no se produce naturalmente.
Tal como se usa en el presente documento, se pretende que los terminos "microbiano", "organismo microbiano" o "microorganismo" signifiquen cualquier organismo que exista como una celula microscopica que este incluido en los dominios de archaea, bacteria o eukarya. Por tanto, se pretende que el termino abarque celulas u organismos procariotas o eucariotas que tengan un tamano microscopico e incluye bacterias, archaeas y eubacterias de todas las especies, asf como microorganismos eucariotas tales como levaduras y hongos. El termino incluye tambien cultivos celulares de cualquier especie que se pueda cultivar para la produccion de un producto bioqmmico.
Tal como se usa en el presente documento, se pretende que el termino "acido 4-hidroxibutanoico" signifique un
derivado 4-hidroxi de acido butmco que tiene la formula qmmica C4H8O3 y una masa molecular de 104,11 g/mol (126,09 g/mol para su sal de sodio). El compuesto qmmico acido 4-hidroxibutanoico se conoce tambien en la tecnica como 4-HB, 4-hidroxibutirato, acido gamma-hidroxibutmco o GHB. Se pretende que el termino tal como se usa en el presente documento incluya cualquiera de las diversas formas salinas de los compuestos e incluye, por ejemplo, 4-hidroxibutanoato y 4-hidroxibutirato. Los ejemplos espedficos de formas salinas de 4-HB incluyen sodio 4-HB y potasio 4-HB. Por tanto, los terminos acido 4-hidroxibutanoico, 4-HB, 4-hidroxibutirato, 4-hidroxibutanoato, acido gamma-hidroxibutmco y GBH asf como otros nombres reconocidos en la tecnica se usan como sinonimos en el presente documento.
Tal como se usa en el presente documento, se pretende que el termino "monomerico" cuando se usa en referencia a 4-HB signifique 4-HB en forma no polimerica o sin derivatizar. Los ejemplos espedficos de 4-HB polimerico incluyen acido poli-4-hidroxibutanoico y los copolfmeros de, por ejemplo, 4-Hb y 3-HB. Un ejemplo espedfico de una forma derivatizada de 4-HB es 4-HB-CoA. Se conocen tambien en la materia otras formas de 4-HB polimericas y otras formas derivatizadas de 4-HB.
Tal como se usa en el presente documento, se pretende que el termino ‘Vbutirolactona” signifique una lactona que tiene la formula qmmica C4H6O2 y una masa molecular de 86,089 g/mol. El compuesto qmmico Y-butirolactona se conoce tambien en la tecnica como GBL, butirolactona, 1,4-lactona, 4-butirolactona, acido 4-hidroxibutmco lactona, y acido gamma-hidroxibutmco lactona. Se pretende que el termino, tal como se usa en el presente documento, incluya cualquiera de las diversas formas salinas de los compuestos.
Tal como se usa en el presente documento, se pretende que el termino "1-4 butanodiol" signifique un derivado de alcohol del alcano butano, que transporta dos grupos hidroxilo que tienen la formula qmmica C4H10O2 y una masa molecular de 90,12 g/mol. El compuesto qmmico 1,4 butanodiol se conoce tambien en la tecnica como BDO y es un compuesto intermedio o precursor qmmico de una familia de compuestos denominada en el presente documento como familia BDO de compuestos, algunos de los cuales se ilustran en la figura 1.
Tal como se usa en el presente documento, se pretende que el termino "tetrahidrofurano" signifique un compuesto organico heterodclico que corresponde al analogo completamente hidrogenado del compuesto aromatico furano que tiene la formula qmmica C4H8O y una masa molecular de 72,11 g/mol. El compuesto qmmico tetrahidrofurano se conoce tambien en la tecnica como THF, tetrahidrofurano, 1,4-epoxibutano, oxido de butileno, oxido de ciclotetrametileno, oxaciclopentano, oxido de dietileno, oxolano, furanidina, hidrofurano, tetraoxido de metileno. Se pretende que el termino, tal como se usa en el presente documento, incluya cualquiera de las diversas formas salinas de los compuestos.
Tal como se usa en el presente documento, se pretende que el termino "CoA" o "coenzima A" signifique un cofactor organico o grupo prostetico (parte no proteica de un enzima) cuya presencia se requiere para la actividad de muchas enzimas (la apoenzima) para formar un sistema enzimatico activo. La coenzima A funciona en determinadas enzimas condensadoras, actua en la transferencia del acetilo u otros grupos acilo y en la smtesis y oxidacion de acidos grasos, la oxidacion del piruvato y en otras acetilaciones.
Tal como se usa en el presente documento, se pretende que el termino "sustancialmente anaerobio", cuando se usa en referencia a un cultivo o condicion de crecimiento, signifique que la cantidad de oxfgeno es menos de aproximadamente 10% de saturacion para el oxfgeno disuelto en un medio lfquido. Se pretende tambien que el termino incluya camaras precintadas de medio lfquido o solido mantenidas con una atmosfera de menos de aproximadamente un 1% de oxfgeno.
Los organismos microbianos no naturales tal como se divulgan pueden contener alteraciones geneticas estables, que se refieren a microorganismos que se pueden cultivar durante mas de cinco generaciones sin perder la alteracion. En general, las alteraciones geneticas estables incluyen modificaciones que persisten mas de 10 generaciones, las modificaciones particularmente estables persistiran mas de aproximadamente 25 generaciones, y mas especialmente, las modificaciones geneticas estables seran superiores a 50 generaciones, incluyendo de forma indefinida.
Los expertos en la materia entenderan que las alteraciones geneticas, incluyendo las modificaciones metabolicas ilustradas en el presente documento, se describen con referencia a genes de E. coli y levadura y sus correspondientes reacciones metabolicas. Sin embargo, dada la secuenciacion completa del genoma de una amplia variedad de organismos y el alto nivel de capacidad en el campo de la genomica, los expertos en la materia seran facilmente capaces de aplicar las ensenanzas y las directrices proporcionadas en el presente documento esencialmente a todos los diferentes organismos. Por ejemplo, las alteraciones metabolicas de E. coli ilustradas en el presente documento pueden aplicarse facilmente a otras especies incorporando los mismos acidos nucleicos codificantes o sus analogos de especies diferentes de las especies citadas. Dichas alteraciones geneticas incluyen, por ejemplo, alteraciones geneticas de especies homologas, por lo general, y en particular, ortologas, desplazamientos de genes paralogos o no ortologos.
Un ortologo es un gen o genes que estan relacionados por una descendencia vertical y son responsables de funciones sustancialmente iguales o identicas en diferentes organismos. Por ejemplo, la epoxido hidrolasa de raton y la epoxido hidrolasa de ser humano se pueden considerar ortologos para la funcion biologica de hidrolisis de los epoxidos. Los genes estan relacionados por una descendencia vertical cuando, por ejemplo, comparten una similitud de secuencias en cantidad suficiente para indicar que son homologos, o estan relacionados evolutivamente a partir de un ancestro comun. Los genes se pueden considerar tambien ortologos si comparten la estructura tridimensional pero no necesariamente la similitud de la secuencia, de una cantidad suficiente para indicar que han evolucionado a partir de un ancestro comun en la medida en la que la similitud de la secuencia primaria no sea identificable. Los genes que son ortologos pueden codificar protemas con una similitud de secuencias de aproximadamente 25% a 100% de identidad en la secuencia de aminoacidos. Puede considerarse que los genes que codifican protemas que comparten una similitud de aminoacidos menor del 25% proceden de descendencia vertical si su estructura tridimensional muestra tambien similitudes. Los miembros de la familia de enzimas de las serina proteasas, incluyendo el activador del plasminogeno tisular y la elastasa, se consideran procedentes de una descendencia vertical a partir de un ancestro comun.
Los ortologos incluyen genes o sus productos genicos codificados que mediante, por ejemplo, evolucion, han divergido en estructura o actividad global. Por ejemplo, donde una especie codifica un producto genico que presenta dos funciones y donde dichas funciones se han separado en distintos genes en una segunda especie, los tres genes y sus correspondientes productos se considera ortologos. Para la produccion acoplada al crecimiento de un producto bioqmmico, los expertos en la materia entenderan que el gen ortologo que hospeda la actividad metabolica que se va a perturbar es el que se va a seleccionar para la construccion del microorganismo no natural. Un ejemplo de ortologos que presentan actividades separables es cuando distintas actividades se han separado en distintos productos genicos entre dos o mas especies o en una unica especie. Un ejemplo espedfico es la separacion de la proteolisis de la elastasa y la proteolisis del plasminogeno, dos tipos de actividad serina proteasa, en moleculas distintas como activador del plasminogeno y la elastasa. Un segundo ejemplo es la separacion de la actividad de la 5'-3' exonucleasa de micoplasma y la actividad de la ADN polimerasa III. La ADN polimerasa de la primera especie se puede considerar un ortologo para cualquiera o ambas exonucleasa o polimerasa de la segunda especie y viceversa.
Por el contrario, los paralogos son homologos relacionados por, por ejemplo, duplicacion seguida de divergencia evolutiva y que tienen funciones similares o comunes, pero no funciones identicas. Los paralogos pueden originarse o derivarse de, por ejemplo, la misma especie o de una especie diferente. Por ejemplo, la epoxido hidrolasa microsomial (epoxido hidrolasa I) y la epoxido hidrolasa soluble (epoxido hidrolasa II) se pueden considerar paralogos por que representan dos enzimas distintas, que han evolucionado simultaneamente a partir de un ancestro comun, que catalizan diferentes reacciones y que tiene funciones distintas en la misma especie. Los paralogos son protemas de la misma especie con una similitud de secuencias significativa entre sf sugiriendo que son homologas, o relacionadas a traves de evolucion simultanea a partir de un ancestro comun. Los grupos de familias de protemas paralogas incluyen los homologos de HipA, genes de la luciferasa, peptidasas, y otros.
El desplazamiento de un gen no ortologo es un gen no ortologo de una especie que se puede sustituir por una funcion genica de una especie diferente a la que se hace referencia. La sustitucion incluye, por ejemplo, ser capaz de llevar a cabo sustancialmente la misma funcion o una funcion similar en la especie de origen en comparacion con la funcion a la que se hace referencia en la especie diferente. Aunque generalmente, el desplazamiento de un gen no ortologo se podra identificar como estructuralmente relacionado con un gen conocido que codifica la funcion a la que se hace referencia, los genes menos estructuralmente relacionados, pero funcionalmente similares, y sus correspondientes productos genomicos estaran comprendidos, sin embargo, en el significado del termino tal como se usa en el presente documento. La similitud funcional requiere, por ejemplo, al menos algo de similitud estructural en el sitio activo o la region de union de un gen no ortologo en comparacion con un gen que codifica la funcion que se desea sustituir. Por tanto, un gen no ortologo incluye, por ejemplo, un paralogo o un gen no relacionado.
Por tanto, en la identificacion y construccion de los organismos microbianos no naturales que tienen capacidad de biosmtesis de 4-HB, GBL y/o BDO, los expertos en la materia entenderan que, aplicando las ensenanzas y directrices proporcionadas en el presente documento para una especie concreta, la identificacion de las modificaciones metabolicas puede incluir la identificacion y la inclusion o la inactivacion de ortologos. En la medida en la que los desplazamientos de los genes paralogos y/o no ortologos estan presentes en el microorganismo citado que codifica un enzima que cataliza una reaccion metabolica similar o sustancialmente similar, los expertos en la materia tambien pueden utilizar estos genes relacionados evolutivamente.
Los desplazamientos de los genes ortologos, paralogos y no ortologos pueden determinarse mediante metodos bien conocidos por los expertos en la materia. Por ejemplo, el examen de las secuencias de acidos nucleicos o aminoacidos de dos polipeptidos revelara la identidad y las similitudes de secuencia entre las secuencias comparadas. Basandose en dichas similitudes, un experto en la materia puede determinar si la similitud es suficientemente alta para indicar que las protemas estan relacionadas evolutivamente a partir de un ancestro comun. Algoritmos bien conocidos por expertos en la materia, tales como Align, BLAST, Clustal W y otros comparan y determinan la similitud o identidad de la secuencia en bruto, y determinan tambien la presencia o significancia de huecos en la secuencia a los que se puede asignar a un peso o puntuacion. Dichos algoritmos tambien son bien
conocidos en la tecnica y son analogamente aplicables para determinar la similitud o identidad de la secuencia de nucleotidos. Los parametros de similitud suficiente para determinar el parentesco se calculan besandose en metodos bien conocidos para calcular la similitud estadfstica, o la posibilidad de encontrar una correspondencia similar en un peptido aleatorio, y la significancia de la correspondencia determinada. Una comparacion informatica de dos o mas secuencias puede, si se desea, optimizarse tambien visualmente por los expertos en la materia. Puede esperarse que los productos o protemas genicas relacionadas tengan una similitud elevada, por ejemplo, 25% a l0o% de identidad de secuencias. Las protemas que no estan relacionadas pueden tener una identidad que es esencialmente la misma que la que se esperana que se produjera por azar, si se escaneara una base de datos de suficiente tamano (aproximadamente 5%). Las secuencias entre 5% y 24% pueden representar o no suficiente homologfa para concluir que las secuencias comparadas estan relacionadas. El analisis estadfstico adicional para determinar la significancia de dichas correspondencias dado el tamano del conjunto de datos se puede llevar a cabo para determinar la relevancia de estas secuencias.
Los parametros ilustrativos para determinar el parentesco de dos o mas secuencias usando el algoritmo BLAST, por ejemplo, pueden ser como se muestra a continuacion. En resumen, se pueden llevar a cabo las alineaciones de la secuencia de aminoacidos utilizando BLASTP version 2.0.8 (5 de enero de 1999) y los siguientes parametros: Matriz: 0: BLOSUM62; apertura de hueco: 11; extension de hueco: 1; x_desechar: 50; esperado: 10,0; tamano de palabra: 3; filtro: on. Se pueden llevar a cabo alineaciones de acidos nucleicos utilizando BLASTN version 2.0.6 (16 de septiembre de 1998) y los siguientes parametros: Correspondencia: 1; correspondencia incorrecta: -2; apertura de hueco: 5; extension de hueco: 2; x_desechar: 50; esperado: 10,0; tamano de palabra: 11; filtro: off Los expertos en la materia sabran que modificaciones se pueden hacer a los anteriores parametros para aumentar o disminuir el rigor de la comparacion, por ejemplo, y determinar el parentesco de dos o mas secuencias.
La divulgacion proporciona un biocatalizador microbiano que no se produce naturalmente que incluye un organismo microbiano que tiene una ruta biosintetica del acido 4-hidroxibutanoico (4-HB) que incluye al menos un acido nucleico exogeno que codifica la 4-hidroxibutanoato deshidrogenasa, semialdehfdo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdehndo succmico deshidrogenasa dependiente de CoA, glutamato: semialdetndo succmico transaminasa o glutamato decarboxilasa, en el que el acido nucleico exogeno se expresa en cantidades suficientes para producir acido 4-hidroxibutanoico monomerico (4-HB). La succinil-CoA sintetasa se denomina tambien succinil-CoA sintasa o succinil CoA ligasa.
Los biocatalizadores microbianos que no se producen naturalmente de la divulgacion incluyen organismos microbianos que emplean combinaciones de reacciones metabolicas para producir biosinteticamente los compuestos de la divulgacion. Los compuestos ilustrativos producidos por los microorganismos no naturales incluyen, por ejemplo, acido 4-hidroxibutanoico, 1,4-butanodiol y Y-butirolactona. En la figura 1 se ilustran las relaciones de estos compuestos ilustrativos con respecto a la smtesis qrnmica o biosmtesis.
En una realizacion, se disena mediante ingeniena genetica un organismo microbiano no natural para producir 4-HB. Este compuesto es un punto de entrada util en la familia de compuestos de 1,4-butanodiol. Las reacciones bioqmmicas para la formacion de 4-HB a partir de succinato, a partir de succinato a traves de succinil-CoA o a partir de a-cetoglutarato se muestran en la figura 2.
La invencion se describe en el presente documento con referencia general a la reaccion metabolica, el reactivo o el producto de los mismos, o con referencia espedfica a uno o mas acidos nucleicos o genes que codifican un enzima asociada con, o que cataliza, la reaccion metabolica, el reactivo o el producto al que se hace referencia. Salvo que se indique de forma expresa lo contrario en el presente documento, los expertos en la materia entenderan que la referencia a una reaccion constituye tambien la referencia a los reactivos y productos de la reaccion. De manera similar, salvo que se indique de forma expresa lo contrario en el presente documento, la referencia a un reactivo o producto hace referencia tambien a la reaccion y que la referencia a cualquiera de esta reaccion metabolica constituye tambien una referencia al gen o genes que codifican los enzimas que catalizan la reaccion, el reactivo o el producto al que se hace referencia. Del mismo modo, dados los campos bien conocidos de la bioqmmica, enzimologfa y genomica metabolicas, la referencia hecha en el presente documento a un gen o un acido nucleico codificante constituye tambien una referencia a la correspondiente enzima codificada y a la reaccion que cataliza, asf como a los reactivos y productos de la reaccion.
La produccion de 4-HB por medios biosinteticos utilizando los organismos microbianos de la divulgacion es especialmente util debido ya que da como resultado 4-HB monomerico. Los organismos microbianos no naturales de la divulgacion y su biosmtesis de compuestos de la familia de 4-HB y BDO es tambien especialmente util ya que el producto 4-HB (1) se secreta; (2) esta desprovisto de cualesquiera derivatizaciones tales como Coenzima A; (3) evita los cambios termodinamicos durante la biosmtesis, y (4) permite la conversion qrnmica espontanea de 4-HB a Y-butirolactona (GBL) en un medio con pH acido. Esta caractenstica se ilustra posteriormente como etapa 7 en la figura 2 y es tambien especialmente util para una smtesis o biosmtesis qrnmica eficaz de compuestos de la familia BDO tales como 1,4-butanodiol y/o tetrahidrofurano (THF), por ejemplo.
Los organismos microbianos carecen generalmente de la capacidad de sintetizar 4-HB y, por tanto, cualquiera de los compuestos que se muestran en la figura 1 son conocidos por estar comprendidos en la familia de compuestos del
1.4- butanodiol o conocidos por los expertos en la materia por estar comprendidos en la familia de compuestos del 1.4- butanodiol. Ademas, los organismos que tienen todas las capacidades enzimaticas metabolicas necesarias no son conocidos por producir 4-HB a partir de los enzimas descritos y las rutas bioqmmicas ilustradas en el presente documento. Mas bien, con la posible excepcion de unos pocos microorganismos anaerobios descritos adicionalmente a continuacion, los microorganismos que tienen la capacidad enzimatica utilizan 4-HB como sustrato para producir, por ejemplo, succinato. Por el contrario, los organismos microbianos no naturales de la divulgacion generan 4-HB como producto. Tal como se ha descrito anteriormente, la biosmtesis de 4-HB en su forma monomerica no es solo particularmente util en la smtesis de la familia de compuestos de BDO, tambien permite la biosmtesis adicional de compuestos de la familia de BDO y evita en conjunto los procedimientos de smtesis qmmica.
Los organismos microbianos no naturales de la divulgacion que pueden producir 4-HB monomerico se producen asegurando que un organismo hospedador microbiano incluye capacidades funcionales para la smtesis bioqmmica completa de al menos una ruta biosintetica de 4-HB de la divulgacion. Asegurar al menos que una ruta biosintetica de 4-HB obligatoria confiere capacidad de biosmtesis de 4-HB sobre el organismo microbiano hospedador.
Se ilustran en el presente documento tres requisitos de las rutas biosinteticas de 4-HB que se muestran a fines ilustrativos en la figura 2. Un requisito de la ruta biosintetica de 4-HB incluye la biosmtesis de 4-HB a partir de succinato. Los enzimas que participan en esta ruta de 4-HB incluyen la semialdehndo succmico deshidrogenasa independiente de CoA y la 4-hidroxibutanoato deshidrogenasa. Otro requisito de la ruta biosintetica de 4-HB incluye la biosmtesis del succinato a traves de la succinil-CoA. Los enzimas que participan en esta ruta de 4-HB incluyen la succinil-CoA sintetasa, la semialdehndo succmico deshidrogenasa dependiente de CoA y la 4-hidroxibutanoato deshidrogenasa. Un tercer requisito de la ruta biosintetica de 4-HB incluye la biosmtesis de 4-HB a partir de acetoglutarato. Los enzimas que participan en esta ruta biosintetica de 4-HB incluyen glutamato:semialdehndo succmico transaminasa, glutamato decarboxilasa y 4-hidroxibutanoato deshidrogenasa. Cada una de estas rutas biosinteticas de 4-HB, sus sustratos, reactivos y productos se describen ademas adicionalmente en los Ejemplos.
Los organismos microbianos no naturales de la divulgacion se pueden producir introduciendo acidos nucleicos expresables que codifican una o mas de los enzimas que participan en una o mas rutas biosinteticas de 4-HB. Dependiendo del organismo microbiano hospedador seleccionado para la biosmtesis, se pueden expresar los acidos nucleicos para alguna o todas las rutas biosinteticas de 4-HB concretas. Por ejemplo, si un hospedador seleccionado es deficiente en ambas enzimas de la ruta del succinato a 4-HB (la ruta del succinato), y esta ruta se selecciona para la biosmtesis de 4-HB, entonces se introducen acidos nucleicos expresables de la semialdetndo succmico deshidrogenasa independiente de CoA y la 4-hidroxibutanoato deshidrogenasa en el hospedador para la posterior expresion exogena. Como alternativa, si el hospedador seleccionado es deficiente en 4-hidroxibutanoato deshidrogenasa, entonces se necesita un acido nucleico codificante de esta enzima para conseguir la biosmtesis de 4-HB.
De manera analoga, cuando se selecciona la biosmtesis de 4-HB para producirse a traves de la ruta del succinato a succinil-CoA (la ruta de la succinil-CoA), los acidos nucleicos que codifican las deficiencias del hospedador respecto a los enzimas succinil CoA sintetasa, semialdehndo succmico deshidrogenasa dependiente de CoA y/o 4-hidroxibutanoato deshidrogenasa deben expresarse exogenamente en el hospedador receptor. La seleccion de la biosmtesis de 4-HB a traves de la ruta del a-cetoglutarato al semialdehndo succmico (la ruta del a-cetoglutarato) utilizara la expresion exogena para cubrir las deficiencias del hospedador respecto a una o mas de los enzimas glutamato: semialdehndo succmico transaminasa, glutamato decarboxilasa y/o 4-hidroxibutanoato deshidrogenasa.
Dependiendo de los constituyentes de la ruta biosintetica de 4-HB de un organismo microbiano hospedador seleccionado, los biocatalizadores de 4-HB microbianos no naturales de la invencion incluiran al menos un acido nucleico que codifica una ruta de 4-HB expresada exogenamente y hasta los acidos nucleicos que codifican las seis rutas de 4-HB. Por ejemplo, se puede establecer la biosmtesis de 4-HB a partir de las tres rutas en un hospedador deficiente en 4-hidroxibutanoato deshidrogenasa mediante la expresion exogena de un acido nucleico que codifica una 4-hidroxibutanoato deshidrogenasa. Por el contrario, se puede establecer la biosmtesis de 4-HB a partir de las tres rutas en un hospedador deficiente en las seis enzimas mediante la expresion exogena de las seis semialdehndo succmico deshidrogenasas independientes de CoA, succinil-CoA sintetasa, semialdehndo succmico deshidrogenasa dependiente de CoA, glutamato: semialdehndo succmico transaminasa, glutamato decarboxilasa y 4-hidroxibutanoato deshidrogenasa.
Con las ensenanzas y directrices que se proporcionan en el presente documento, los expertos en la materia entenderan que el numero de acidos nucleicos codificantes que se introducen en una forma expresable seran equivalentes a las deficiencias de la ruta de 4-HB del organismo microbiano hospedador seleccionado. Por tanto, un organismo microbiano no natural de la divulgacion puede tener uno, dos, tres, cuatro, cinco o seis acidos nucleicos codificantes que codifican las anteriores enzimas que constituyen las rutas biosinteticas de 4-HB. En algunas realizaciones, los organismos microbianos no naturales pueden incluir tambien otras modificaciones geneticas que facilitan u optimizan la biosmtesis de 4-HB o que transmiten otras funciones utiles para el organismo microbiano hospedador. Una de dichas funcionalidades adicionales puede incluir, por ejemplo, el aumento de la smtesis de uno o mas de los precursores de la ruta de 4-HB tal como succinato, succinil-CoA y/o a-cetoglutarato.
En algunas realizaciones, un organismo microbiano no natural de la divulgacion se genera a partir de un hospedador que contiene la capacidad enzimatica de sintetizar 4-HB. En esta realizacion espedfica puede ser util aumentar la smtesis o la acumulacion de un producto de la ruta de 4-HB hasta, por ejemplo, impulsar las reacciones de la ruta de 4-HB hacia la produccion de 4-HB. La smtesis aumentada o acumulacion se puede llevar a cabo mediante, por ejemplo, la sobreexpresion de acidos nucleicos que codifican una o mas de los enzimas de la ruta de 4-HB anteriormente descrita. Se puede producir la sobreexpresion del enzima o enzimas de la ruta de 4-HB, por ejemplo, mediante la expresion exogena del gen o genes endogenos, o mediante la expresion exogena del gen o genes heterologos. Por tanto, los organismos que se producen naturalmente pueden generarse facilmente para convertirse en organismos microbianos no naturales productores de 4-HB de la invencion mediante la sobreexpresion de uno, dos, tres, cuatro, cinco o los seis acidos nucleicos que codifican los enzimas de la ruta biosintetica de 4-HB. Ademas, se puede generar un organismo no natural mediante mutagenesis de un gen endogeno que da como resultado un aumento en la actividad de un enzima en la ruta biosintetica de 4-HB.
En realizaciones especialmente utiles, se emplea la expresion exogena. La expresion exogena confiere la capacidad de personalizar la expresion y/ los elementos reguladores del hospedador y la aplicacion para conseguir un nivel de expresion deseado que controla el usuario. Sin embargo, se puede utilizar tambien la expresion endogena en otras realizaciones tales como mediante la eliminacion de un efector regulador negativo o la induccion del promotor del gen cuando se une a un promotor inducible u otro elemento regulador. Por tanto, un gen endogeno que tiene un promotor inducible que se produce naturalmente puede regularse por exceso proporcionado el agente inductor adecuado, o la region reguladora de un gen endogeno se puede genomanipular para incorporar un elemento regulador inducible, permitiendo por tanto la regulacion de la expresion aumentada de un gen endogeno en el tiempo deseado. De manera similar, se puede incluir un promotor inducible como un elemento regulador de un gen exogeno introducido en un organismo microbiano no natural (vease el Ejemplo II).
Se pretende que "exogeno", tal como se usa en el presente documento, signifique que la molecula a la que se hace referencia o la actividad a la que se hace referencia se introduce en el organismo microbiano hospedador. Por tanto, el termino tal como se usa en referencia a la expresion de un acido nucleico codificante se refiere a la introduccion del acido nucleico codificante en una forma expresable en el organismo microbiano. Cuando se usa en referencia a una actividad biosintetica, el termino se refiere a una actividad que se introduce en el organismo hospedador de referencia. La fuente puede ser, por ejemplo, acidos nucleicos homologos o heterologos codificantes que expresan la actividad a la que se hace referencia tras su introduccion en el organismo microbiano hospedador. Por tanto, el termino "endogeno" se refiere a una molecula o actividad a la que se hace referencia que esta presente en el hospedador. De manera similar, el termino, cuando se usa en referencia a la expresion de un acido nucleico codificante, se refiere a la expresion de un acido nucleico codificante contenido en el organismo microbiano El termino "heterologo" se refiere a una molecula o actividad derivada de una fuente diferente de la de la especie a la que se hace referencia mientras que "homologo" se refiere a una molecula o actividad derivada del organismo microbiano hospedador. De acuerdo con ello, la expresion exogena de un acido nucleico codificante puede utilizar un acido nucleico homologo o heterologo codificante.
Las fuentes de acidos nucleicos codificantes de un enzima de la ruta de 4-HB pueden incluir, por ejemplo, cualquier especie donde el producto genico codificado es capaz de catalizar la reaccion a la que se hace referencia. Dichas especies incluyen organismos procariotas y eucariotas que incluyen, aunque no de forma limitativa, bacterias, incluyendo archaeas y eubacterias, y eucariotas, incluyendo levaduras, plantas, insectos, animales, y mairnferos, incluyendo seres humanos. Por ejemplo, los organismos microbianos que tienen produccion biosintetica de 4-HB se ilustran en el presente documento con referencia a E. coli y levaduras hospedadoras. Sin embargo, con la secuencia genomica completa disponible ahora para mas de 550 especies (con mas de la mitad de estas disponibles en bases de datos publicas tales como la NCBI), que incluyen 395 genomas de microorganismos y una variedad de genomas de levaduras, hongos, plantas, y marnfferos, la identificacion de los genes que codifican la actividad biosintetica de 4-HB necesaria para uno o mas genes en especies relacionadas o distantes, incluyendo por ejemplo, desplazamientos de genes homologos, ortologos, paralogos y no ortologos de genes conocidos, y el intercambio de alteraciones geneticas entre organismos es rutinario y bien conocido en la tecnica. De acuerdo con ello, las alteraciones metabolicas que permiten la biosmtesis de 4-HB y otros compuestos descritos en el presente documento con referencia a un organismo concreto tal como E. coli o levadura pueden aplicarse facilmente a otros microorganismos, incluyendo organismos procariotas y eucariotas analogos. Con las ensenanzas y directrices que se proporcionan en el presente documento, los expertos en la materia sabran que una alteracion metabolica ilustrada en un organismo se puede aplicar igualmente a otros organismos.
En algunos casos, tales como cuando existe una ruta biosintetica de 4-HB alternativa en una especie no relacionada, se puede conferir la biosmtesis de 4-HB a la especie hospedadora mediante, por ejemplo, la expresion exogena de un paralogo o paralogos de la especie no relacionada que cataliza una reaccion metabolica similar pero no identica para sustituir a la reaccion a la que se hace referencia. Como existen determinadas diferencias entre redes metabolicas entre diferentes organismos, los expertos en la materia entenderan que puede diferir la utilizacion de genes reales entre diferentes organismos. Sin embargo, con las ensenanzas y directrices que se proporcionan en el presente documento, los expertos en la materia entenderan tambien que las ensenanzas y metodos de la invencion se pueden aplicar a todos los organismos microbianos que utilizan alteraciones metabolicas analogas a las ilustradas en el presente documento para construir un organismo microbiano en una especie de interes que
sintetizara el 4-HB monomerico.
Se pueden seleccionar organismos microbianos hospedadores a partir de, y los organismos microbianos no naturales generarse en, por ejemplo, bacterias, levaduras, hongos o cualquiera de una variedad de diferentes microorganismos aplicable a los procesos de fermentacion. Las bacterias ilustrativas incluyen especies seleccionadas entre E. coli, Klebsiella oxytoca, Anaerobiospirillum succiniciproducens, Actinobacillus succinogenes, Mannheimia succiniciproducens, Rhizobium etli, Bacillus subtilis, Corynebacterium glutamicum, Gluconobacter oxydans, Zymomonas mobilis, Lactococcus lactis, Lactobacillus plantarum, Streptomyces coelicolor, Clostridium acetobutylicum, Pseudomonas fluorescens, y Pseudomonas putida. Las levaduras u hongos ilustrativos incluyen especies seleccionadas entre Saccharomyces cerevisiae, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces marxianus, Aspergillus terreus, Aspergillus niger y Pichia pastoris.
Se pueden llevar a cabo metodos para construir y ensayar los niveles de expresion de un hospedador productor de 4-HB que no se produce naturalmente, por ejemplo, por metodos recombinantes y de deteccion bien conocidos en la materia. Se pueden encontrar descritos dichos metodos en, por ejemplo, Sambrook y col., Molecular Cloning: A Laboratory Manual, Tercera Ed., Cold Spring Harbor Laboratory, Nueva York (2001); Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Md. (1999). 4-HB y GBL se pueden separar mediante, por ejemplo, HPLC usando una columna Spherisorb 5 ODS 1 y una fase movil de tampon fosfato 10 mM al 70% (pH=7) y metanol al 30%, y detectarse con un detector UV a 215 nm (Hennessy et al. J. Forensic Sci.
46(6):1-9 (2004)). BDO se detecta mediante cromatograffa gaseosa o mediante HPLC y un detector de mdice de refraccion utilizando una columna Aminex HPX-87H y una fase movil de acido sulfurico 0,5 mM (Gonzalez-Pajuelo et al., Met. Eng. 7:329-336 (2005)).
Los organismos microbianos no naturales de la divulgacion se construyen utilizando metodos bien conocidos en la tecnica como se ilustra anteriormente para expresar de forma exogena al menos un acido nucleico que codifica un enzima de una ruta de 4-HB en cantidades suficientes para producir 4-HB monomerico. Los niveles ilustrativos de expresion de enzimas 4-HB en cada ruta se describen adicionalmente a continuacion en los Ejemplos. Siguiendo las ensenanzas y directrices proporcionadas en el presente documento, los organismos microbianos no naturales de la divulgacion pueden conseguir la biosmtesis del 4-HB monomerico dando como resultado concentraciones intracelulares entre aproximadamente 0,1-25 mM o mas. En general, la concentracion intracelular de 4-HB monomerico esta entre aproximadamente 3-20 mM, particularmente entre aproximadamente 5-15 mM y mas particularmente entre aproximadamente 8-12 mM, incluyendo aproximadamente 10 mM o mas. Las concentraciones intracelulares entre y por encima de cada uno de estos intervalos ilustrativos puede conseguirse tambien a partir de los organismos microbianos que no se producen naturalmente de la divulgacion.
Como se describe adicionalmente a continuacion, una condicion de crecimiento ilustrativa para conseguir la biosmtesis de 4-HB incluye condiciones de cultivo o fermentacion anaerobias. En determinadas realizaciones, los organismos microbianos que no se producen naturalmente de la divulgacion se pueden mantener, cultivar o fermentar en condiciones de cultivo anaerobias o sustancialmente anaerobias. En resumen, las condiciones anaerobias se refieren a un entorno desprovisto de oxfgeno. Las condiciones sustancialmente anaerobias incluyen, por ejemplo, un cultivo, una fermentacion discontinua o continua de tal manera que la concentracion de oxfgeno disuelto en el medio permanezca entre 0 y 10% de saturacion. Las condiciones sustancialmente anaerobias incluyen tambien celulas en crecimiento o en reposo en un medio lfquido o en agar solido en el interior de una camara precintada mantenida con una atmosfera de menos de un 1% de oxfgeno. El porcentaje de oxfgeno puede mantenerse mediante, por ejemplo, burbujeo en el cultivo una mezcla de N2/CO2 u otro gas o gases no de oxfgeno adecuados.
La divulgacion tambien proporciona un biocatalizador microbiano que no se produce naturalmente que incluye un organismo microbiano que tiene rutas biosintetica del acido 4-hidroxibutanoico (4-HB) y 1,4-butanodiol (BDO) que incluye al menos un acido nucleico exogeno que codifica la 4-hidroxibutanoato deshidrogenasa, semialdehndo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdetndo succmico deshidrogenasa dependiente de CoA, 4-hidroxibutirato:CoA transferasa, glutamato: semialdehndo succmico transaminasa, glutamato descarboxilasa, aldehndo deshidrogenasa independiente de CoA, aldehndo deshidrogenasa dependiente de CoA o alcohol deshidrogenasa, en el que el acido nucleico exogeno se expresa en cantidad suficiente para producir 1,4-butanodiol (BDO).
Se pueden generar tambien organismos microbianos que no se produce naturalmente que biosintetizan BDO. Siguiendo las ensenanzas y directrices proporcionadas anteriormente para la construccion de organismos microbianos que sintetizan 4-HB, se pueden incorporar rutas de BDO adicionales en los organismos microbianos productores de 4-HB para generar organismos que sintetizan tambien BDO y otros compuestos de la familia BDO. En la figura 3 se ilustra la smtesis qrnmica de BDO y sus productos posteriores. Los organismos microbianos no naturales de la divulgacion capaces de la biosmtesis de BDO evitan esta smtesis qrnmica utilizando 4-HB como punto de entrada como se ilustra en la figura 2. Como se describe adicionalmente a continuacion, se pueden usar productores de 4-HB para convertir qmmicamente 4-HB en GBL y a continuacion, en BDO o THF, por ejemplo. Como alternativa, se pueden modificar adicionalmente los productores de 4-HB para incluir capacidades biosinteticas para la conversion de 4-HB y/o GBL en BDO.
Las rutas de BDO adicionales a introducir en productores de 4-HB incluyen, por ejemplo, la expresion exogena en un fondo de hospedador deficiente o la sobreexpresion de una aldehndo deshidrogenasa independiente de CoA, aldehndo deshidrogenasa dependiente de CoA o alcohol deshidrogenasa. En ausencia de la acil-CoA sintetasa endogena capaz de modificar 4-HB, los organismos microbianos productores de BDO que no se producen naturalmente pueden incluir ademas una acil-CoA sintetasa exogena selectiva para 4-HB. En la Tabla 1 a continuacion se relacionan alcohol y aldehfdo deshidrogenasas ilustrativos que se pueden usar para estas conversiones in vivo de 4-HB a BDO.
Tabla 1. Alcohol y aldehido deshidrogenasas para la conversion de 4-HB en BDO.
ALCOHOL DESHIDROGENASAS
ec: 1.1.1.1 alcohol deshidrogenasa
ec: 1.1.1.2 alcohol deshidrogenasa (NADP+)
ec: 1.1.1.4 (R,R)-butanodiol deshidrogenasa
ec: 1.1.1.5 acetoma deshidrogenasa
ec: 1.1.1.6 glicerol deshidrogenasa
ec: 1.1.1.7 propanodiol-fosfato deshidrogenasa
ec: 1.1.1.8 glicerol-3-fosfato deshidrogenasa (NAD+)
ec: 1.1.1.11 D-arabinitol 4-deshidrogenasa
ec: 1.1.1.12 L-arabinitol 4-deshidrogenasa
ec: 1.1.1.13 L-arabinitol 2-deshidrogenasa
ec: 1.1.1.14 L-iditol 2-deshidrogenasa
ec: 1.1.1.15 D-iditol 2-deshidrogenasa
ec: 1.1.1.16 galactitol 2-deshidrogenasa
ec: 1.1.1.17 manitol-1-fosfato 5-deshidrogenasa
ec: 1.1.1.18 inositol 2-deshidrogenasa
ec: 1.1.1.21 aldehfdo reductasa
ec: 1.1.1.23 histidinol deshidrogenasa
ec: 1.1.1.26 glioxilato reductasa
ec: 1.1.1.27 L-lactato deshidrogenasa
ec: 1.1.1.28 D-lactato deshidrogenasa
ec: 1.1.1.29 glicerato deshidrogenasa
ec: 1.1.1.30 3-hidroxibutirato deshidrogenasa
ec: 1.1.1.31 3-hidroxibutirato deshidrogenasa
ec: 1.1.1.35 3-hidroxiacil-CoA deshidrogenasa
ec: 1.1.1.36 acetoacetil-CoA reductasa
ec: 1.1.1.37 malato deshidrogenasa
ec: 1.1.1.38 malato deshidrogenasa (oxaloacetato-descarboxilante)
ec: 1.1.1.39 malato deshidrogenasa (descarboxilante)
ec: 1.1.1.40 malato deshidrogenasa (oxaloacetato-descarboxilante) (NADP+)
ec: 1.1.1.41 isocitrato deshidrogenasa (NAD+)
ec: 1.1.1.42 isocitrato deshidrogenasa (NADP+)
ec: 1.1.1.54 alil-alcohol deshidrogenasa
ec: 1.1.1.55 lactaldehfdo reductasa (NADPH)
ec: 1.1.1.56 ribitol 2-deshidrogenasa
ec: 1.1.1.59 3-hidroxipropionato deshidrogenasa
ec: 1.1.1.60 2-hidroxi-3-oxopropionato reductasa
ec: 1.1.1.61 4-hidroxibutirato deshidrogenasa
ec: 1.1.1.66 omega-hidroxidecanoato deshidrogenasa
ec: 1.1.1.67 manitol 2-deshidrogenasa
ec: 1.1.1.71 alcohol deshidrogenasa [NAD(P)+]
ec: 1.1.1.72 glicerol deshidrogenasa (NADP+)
ec: 1.1.1.73 octanol deshidrogenasa
ec: 1.1.1.75 (R)-aminopropanol deshidrogenasa
ec: 1.1.1.76 (S,S)-butanodiol deshidrogenasa
ec: 1.1.1.77 lactaldehfdo reductasa
ec: 1.1.1.78 metilglioxal reductasa (dependiente de NADH)
ec: 1.1.1.79 glioxilato reductasa (NADP+)
ec: 1.1.1.80 isopropanol deshidrogenasa (NADP+)
ec: 1.1.1.81 hidroxipiruvato reductasa
ec: 1.1.1.82 malato deshidrogenasa (NADP+)
ec: 1.1.1.83 D-malato deshidrogenasa (descarboxilante)
ec: 1.1.1.84 dimetilmalato deshidrogenasa
ec: 1.1.1.85 3-isopropilmalato deshidrogenasa
ec: 1.1.1.86 cetol-acido reductoisomerasa
ec: 1.1.1.87 homoisocitrato deshidrogenasa
ec: 1.1.1.88 hidroximetilglutaril-CoA reductasa
ec: 1.1.1.90 aril-alcohol deshidrogenasa
ec: 1.1.1.91 aril-alcohol deshidrogenasa (NADP+)
ec: 1.1.1.92 oxaloglicolato reductasa (descarboxilante)
ec: 1.1.1.94 glicerol-3-fosfato deshidrogenasa [NAD(P)+]
ec: 1.1.1.95 fosfoglicerato deshidrogenasa
ec: 1.1.1.97 3-hidroxibencil-alcohol deshidrogenasa
ec: 1.1.1.101 acilglicerona-fosfato reductasa
ec: 1.1.1.103 L-treonina 3-deshidrogenasa
ec: 1.1.1.104 4-oxoprolina reductasa
ec: 1.1.1.105 retinol deshidrogenasa
ec: 1.1.1.110 indololactato deshidrogenasa
ec: 1.1.1.112 indanol deshidrogenasa
ec: 1.1.1.113 L-xilosa 1-deshidrogenasa
ec: 1.1.1.129 L-treonato 3-deshidrogenasa
ec: 1.1.1.137 ribitol-5-fosfato 2-deshidrogenasa
ec: 1.1.1.138 manitol 2-deshidrogenasa (NADP+)
ec: 1.1.1.140 sorbitol-6-fosfato 2- deshidrogenasa
ec: 1.1.1.142 D-pinitol deshidrogenasa
ec: 1.1.1.143 secuoyitol deshidrogenasa
ec: 1.1.1.144 perilil-alcohol deshidrogenasa
ec: 1.1.1.156 glicerol 2-deshidrogenasa (NADP+)
ec: 1.1.1.157 3-hidroxibutiril-CoA deshidrogenasa
ec: 1.1.1.163 ciclopentanol deshidrogenasa
ec: 1.1.1.164 hexadecanol deshidrogenasa
ec: 1.1.1.165 2-alquin-1-ol deshidrogenasa
ec: 1.1.1.166 hidroxiciclohexanocarboxilato deshidrogenasa
ec: 1.1.1.167 hidroximalonato deshidrogenasa
ec: 1.1.1.174 ciclohexano-1,2-diol deshidrogenasa
ec: 1.1.1.177 glicerol-3-fosfato-1-deshidrogenasa (NADP+)
ec: 1.1.1.178 3-hidroxi-2-metilbutiril-CoA deshidrogenasa
ec: 1.1.1.185 L-glicol deshidrogenasa
ec: 1.1.1.190 indol-3-acetaldehfdo reductasa (NADH)
ec: 1.1.1.191 indol-3-acetaldehfdo reductasa (NADPH)
ec: 1.1.1.192 alcohol deshidrogenasa de cadena larga
ec: 1.1.1.194 coniferil-alcohol deshidrogenasa
ec: 1.1.1.195 cinnamil-alcohol deshidrogenasa
ec: 1.1.1.198 (+)-borneol deshidrogenasa
ec: 1.1.1.202 1,3-propanodiol deshidrogenasa
ec: 1.1.1.207 (-)-mentol deshidrogenasa
ec: 1.1.1.208 (+)-neomentol deshidrogenasa
ec: 1.1.1.216 farnesol deshidrogenasa
ec: 1.1.1.217 bencil-2-metil-hidroxibutirato deshidrogenasa
ec: 1.1.1.222 (R)-4-hidroxifenillactato deshidrogenasa
ec: 1.1.1.223 isopiperitenol deshidrogenasa
ec: 1.1.1.226 4-hidroxiciclohexanocarboxilato deshidrogenasa
ec: 1.1.1.229 dietil 2-metil-3-oxosuccinato reductasa
ec: 1.1.1.237 hidroxifenilpiruvato reductasa
ec: 1.1.1.244 metanol deshidrogenasa
ec: 1.1.1.245 ciclohexanol deshidrogenasa
ec: 1.1.1.250 D-arabinitol 2-deshidrogenasa
ec: 1.1.1.251 galactitol 1-fosfato 5- deshidrogenasa
ec: 1.1.1.255 manitol deshidrogenasa
ec: 1.1.1.256 fluoren-9-ol deshidrogenasa
ec: 1.1.1.257 4-(hidroximetil)bencenosulfonato deshidrogenasa ec: 1.1.1.258 6-hidroxihexanoato deshidrogenasa
ec: 1.1.1.259 3-hidroxipimeloil-CoA deshidrogenasa
ec: 1.1.1.261 glicerol-1-fosfato deshidrogenasa [NAD(P)+]
ec: 1.1.1.265 3-metilbutanal reductasa
ec: 1.1.1.283 metilglioxal reductasa (dependiente de NADPH) ec: 1.1.1.286 isocitrato-homoisocitrato deshidrogenasa
ec: 1.1.1.287 D-arabinitol deshidrogenasa (NADP+) butanol deshidrogenasa
ALDEHDO DESHIDROGENASAS
ec: 1.2.1.2 formiato deshidrogenasa
ec: 1.2.1.3 aldehndo deshidrogenasa (NAD+)
ec: 1.2.1.4 aldehndo deshidrogenasa (NADP+)
ec: 1.2.1.5 aldehndo deshidrogenasa [NAD(P)+]
ec: 1.2.1.7 benzaldehndo deshidrogenasa (NADP+)
ec: 1.2.1.8 betarna-aldehndo deshidrogenasa
ec: 1.2.1.9 gliceraldehndo-3-fosfato deshidrogenasa (NADP+)
ec: 1.2.1.10 acetaldehndo deshidrogenasa (acetilante)
ec: 1.2.1.11 semialdehndo de aspartato deshidrogenasa
ec: 1.2.1.12 gliceraldehndo-3-fosfato deshidrogenasa (fosforilante)
ec: 1.2.1.13 gliceraldehndo-3-fosfato deshidrogenasa (NADP+) (fosforilante)
ec: 1.2.1.15 semialdehndo de malonato deshidrogenasa
ec: 1.2.1.16 semialdehndo de succinato deshidrogenasa [NAD(P)+]
ec: 1.2.1.17 glioxilato deshidrogenasa (acilante)
ec: 1.2.1.18 semialdehndo de malonato-deshidrogenasa (acetilante)
ec: 1.2.1.19 aminobutiraldehndo deshidrogenasa
ec: 1.2.1.20 semialdehndo de glutarato-deshidrogenasa
ec: 1.2.1.21 glicolaldehfdo deshidrogenasa
ec: 1.2.1.22 lactaldehfdo deshidrogenasa
ec: 1.2.1.23 2-oxoaldehfdo deshidrogenasa (NAD+)
ec: 1.2.1.24 semialdehndo de succinato-deshidrogenasa
ec: 1.2.1.25 2-oxoisovalerato deshidrogenasa (acilante)
ec: 1.2.1.26 2,5-dioxovalerato deshidrogenasa
ec: 1.2.1.27 semialdehndo de metilmalonato deshidrogenasa (acilante)
ec: 1.2.1.28 benzaldehfdo deshidrogenasa (NAD+)
ec: 1.2.1.29 aril-aldehndo deshidrogenasa
ec: 1.2.1.30 aril-aldehndo deshidrogenasa (NADP+)
ec: 1.2.1.31 semialdehndo de L-aminoadipato deshidrogenasa
ec: 1.2.1.32 semialdehndo de aminomuconato deshidrogenasa
ec: 1.2.1.36 retinal deshidrogenasa
ec: 1.2.1.39 fenilacetaldehndo deshidrogenasa
ec: 1.2.1.41 glutamato-5-semialdel'ndo deshidrogenasa
ec: 1.2.1.42 hexadecanal deshidrogenasa (acilante)
ec: 1.2.1.43 formiato deshidrogenasa (NADP+)
ec: 1.2.1.45 4-carboxi-2-hidroximuconato-6- semialdehndo deshidrogenasa
ec: 1.2.1.46 formaldehndo deshidrogenasa
ec: 1.2.1.47 4-trimetilamoniobutiraldel'ndo deshidrogenasa
ec: 1.2.1.48 aldehfdo deshidrogenasa de cadena larga
ec: 1.2.1.49 2-oxoaldehfdo deshidrogenasa (NADP+)
ec: 1.2.1.51 piruvato deshidrogenasa (NADP+)
ec: 1.2.1.52 oxoglutarato deshidrogenasa (NADP+)
ec: 1.2.1.53 4-hidroxifenilacetaldel'ndo deshidrogenasa
ec: 1.2.1.57 butanal deshidrogenasa
ec: 1.2.1.58 fenilglioxilato deshidrogenasa (acilante)
ec: 1.2.1.59 gliceraldehndo-3-fosfato deshidrogenasa (NAD(P)+) (fosforilante)
ec: 1.2.1.62 4-formilbencenosulfonato deshidrogenasa
ec: 1.2.1.63 6-oxohexanoato deshidrogenasa
ec: 1.2.1.64 4-hidroxibenzaldehndo deshidrogenasa
ec: 1.2.1.65 salicilaldehndo deshidrogenasa
ec: 1.2.1.66 formaldehndo deshidrogenasa dependiente de micotiol
ec: 1.2.1.67 vanillin deshidrogenasa
ec: 1.2.1.68 coniferil-aldel'ndo deshidrogenasa
ec: 1.2.1.69 fluoroacetaldehndo deshidrogenasa
ec: 1.2.1.71 semialdehndo de succinilglutamato deshidrogenasa
Por tanto, ademas de cualquiera de las diversas modificaciones ilustradas previamente para establecer la biosmtesis de 4-HB en un hospedador seleccionado, los organismos microbianos productores de BDO pueden incluir cualquiera de las combinaciones y permutaciones anteriores de las modificaciones metabolicas de la ruta de 4-HB asf como cualquier combinacion de expresion para la aldehfdo deshidrogenasa independiente de CoA, la aldehfdo deshidrogenasa dependiente de CoA o una alcohol deshidrogenasa para generar rutas biosinteticas para BDO. Por tanto, los productores de BDO de la divulgacion pueden tener la expresion exogena de, por ejemplo, uno, dos, tres, cuatro, cinco, seis, siete, ocho, nueve o las 10 enzimas de cualquiera de las seis rutas de 4-HB y/o cualquiera de los enzimas de las 4 rutas de BDO.
El diseno y la construccion de los organismos microbianos geneticamente modificados se lleva a cabo usando metodos bien conocidos en la tecnica para conseguir suficientes cantidades de expresion para producir BDO. En
particular, los organismos bacterianos no naturales de la divulgacion pueden conseguir la biosmtesis de BDO dando como resultado concentraciones intracelulares entre aproximadamente 0,1-25 mM o mas. En general, la concentracion intracelular de BDO esta entre aproximadamente 3-20 mM, particularmente entre aproximadamente 5 15 mM y mas particularmente entre aproximadamente 8-12 mM, incluyendo aproximadamente 10 mM o mas. Las concentraciones intracelulares entre y por encima de cada uno de estos intervalos ilustrativos pueden conseguirse tambien a partir de los organismos microbianos no naturales de la divulgacion. Como con los productores de 4-HB, los productores de BDO tambien se pueden mantener, cultivar o fermentar en condiciones anaerobias.
La divulgacion proporciona ademas un metodo para la produccion de 4-HB. El metodo incluye cultivar un organismo microbiano que no se produce naturalmente que tiene una ruta biosintetica del acido 4-hidroxibutanoico (4-HB) que comprende al menos un acido nucleico exogeno que codifica la 4-hidroxibutanoato deshidrogenasa, semialdehndo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdehndo succmico deshidrogenasa dependiente de CoA, glutamato: semialdehndo succmico transaminasa o glutamato decarboxilasa en condiciones sustancialmente anaerobias durante un periodo de tiempo suficiente para producir acido 4-hidroxibutanoico monomerico (4-HB). El metodo puede incluir adicionalmente la conversion qrnmica de 4-HB a GBL y de BDO o THF, por ejemplo.
Se entiende que, en los metodos de la divulgacion, cualquiera del uno o mas acidos nucleicos exogenos se puede combinar en un organismo microbiano no naturales de la divulgacion siempre que se produzca el producto deseado, por ejemplo, 4-HB, BDO, THF o GBL. Por ejemplo, un organismo microbiano no natural que tiene una ruta biosintetica de 4-HB puede comprender al menos dos acidos nucleicos exogenos que codifican los enzimas deseadas, de tal manera que la combinacion de 4-hidroxibutanoato deshidrogenasa y la semialdehndo succmico deshidrogenasa independiente de CoA; 4-hidroxibutanoato deshidrogenasa y la semialdehndo succmico deshidrogenasa dependiente de CoA; la semialdehndo succmico deshidrogenasa dependiente de CoA y la succinil-CoA sintetasa; succinil-CoA sintetasa y glutamato decarboxilasa, y similares. Por tanto, se entiende que cualquier combinacion de dos o mas enzimas de una ruta biosintetica puede estar incluida en un organismo microbiano no natural de la divulgacion. De manera similar, se entiende que cualquier combinacion de tres o mas enzimas de una ruta biosintetica puede estar incluida en un organismo microbiano no natural de la divulgacion, por ejemplo, 4-hidroxibutanoato deshidrogenasa, semialdehndo succmico deshidrogenasa independiente de CoA y succinil-CoA sintetasa; 4-hidroxibutanoato deshidrogenasa, semialdehndo succmico deshidrogenasa dependiente de CoA y glutamato: semialdehndo succmico transaminasa, y asf sucesivamente, segun se desee, siempre que la combinacion de enzimas de la ruta biosintetica deseada de como resultado la produccion del correspondiente producto deseado. Cualquiera de los organismos microbianos que no se producen naturalmente descritos anteriormente puede cultivarse para producir los productos biosinteticos de la divulgacion. Por ejemplo, los productores de 4-HB pueden cultivarse para la produccion biosintetica de 4-HB. El 4-HB puede estar aislado o tratarse como se describe a continuacion para generar GBL, THF y/o BDO. De manera similar, los productores de BDO pueden cultivarse para la produccion biosintetica de BDO. El BDO se puede aislar o someterse a tratamientos adicionales para la smtesis qrnmica de la familia de compuestos de BDO tales como los compuestos posteriores ilustrados en la figura 3.
En algunas realizaciones, las condiciones de cultivo incluyen crecimiento anaerobio o sustancialmente anaerobio o condiciones de mantenimiento. Se han descrito las condiciones anaerobias ilustrativas anteriormente y son bien conocidas en la tecnica. Se describen a continuacion en los Ejemplos las condiciones anaerobias ilustrativas para los procesos de fermentacion. Puede emplearse cualquiera de estas condiciones con los organismos que no se producen naturalmente, asf como otras condiciones anaerobias bien conocidas en la tecnica. En dichas condiciones anaerobias, los productores de 4-HB y BDO pueden sintetizar 4-HB y BDO monomerico, respectivamente, a concentraciones intracelulares de 5-10 mM o mas, asf como las otras concentraciones ilustradas anteriormente. Pueden generarse tambien numerosos compuestos posteriores para organismos bacterianos no naturales productores de 4-HB y BDO de la divulgacion Con respecto a los organismos microbianos productores de 4-HB de la divulgacion, 4-HB y GBL monomericos se encuentran en equilibrio en el medio de cultivo. La conversion de 4-HB a GBL puede llevarse a cabo eficazmente, por ejemplo, cultivando los organismos microbianos en medio a pH acido. A pH menor o igual a 7,5, en particular a o por debajo de pH 5,5, 4-HB se convierte espontaneamente en GBL, como se ilustra en la figura 1.
El GBL resultante se puede separar de 4-HB y otros componentes del cultivo usando una variedad de metodos bien conocidos en la tecnica. Dichos metodos de separacion incluyen, por ejemplo, los procedimientos de extraccion ilustrados en los Ejemplos, asf como los metodos que incluyen la extraccion lfquido-lfquido continua, pervaporacion, filtracion con membrana, separacion con membrana, osmosis inversa, electrodialisis, destilacion, cristalizacion, centrifugacion, filtracion extractiva, cromatograffa de intercambio ionico, cromatograffa de exclusion por tamano, cromatograffa de adsorcion, y ultrafiltracion. Todos los metodos anteriores son bien conocidos en la materia. El GBL separado se puede purificar mediante, por ejemplo, destilacion.
Otro compuesto posterior que se puede producir a partir de organismos microbianos no naturales productores de 4-HB de la divulgacion incluye, por ejemplo, BDO. Este compuesto se puede sintetizar mediante, por ejemplo, hidrogenacion qrnmica de GBL. Las reacciones de hidrogenacion qrnmica son bien conocidas en la tecnica. Un
procedimiento ilustrativo incluye la reduccion qmmica de 4-HB y/o GBL o una mezcla de estos dos componentes que se derivan del cultivo usando un catalizador de hidrogenacion heterogeneo u homogeneo junto con hidrogeno, o un agente reductor de tipo hidruro usado estequiometrica o catalfticamente, para producir 1,4-butanodiol.
Son igualmente aplicables otros procedimientos bien conocidos en la tecnica para la anterior reaccion qmmica e incluye, por ejemplo, el documento WO N° 82/03854 (Bradley, et al.), que describe la hidrogenolisis de la gammabutirolactona en fase vapor sobre catalizador de oxido de cobre y oxido de cinc. La patente britanica n° 1.230.276, que describe la hidrogenacion de la gamma-butirolactona usando un catalizador de oxido de cobre-oxido de cromo. La hidrogenacion se lleva a cabo en fase lfquida. Se ilustran tambien reacciones discontinuas que tienen presiones totales de reactor elevadas. Las presiones parciales de reactivos y productos en los reactores estan tambien por encima de los respectivos puntos de rodo. La patente britanica n° 1.314.126, que describe la hidrogenacion de la gamma-butirolactona en fase lfquida sobre un catalizador de oxido de mquel-cobalto-torio. Se ilustran reacciones discontinuas teniendo presiones totales elevadas y presiones parciales de componentes tambien por encima de los respectivos puntos de rodo de los componentes. La patente britanica n° 1.344.557, que describe la hidrogenacion de la gamma-butirolactona en fase lfquida sobre un catalizador de oxido de cobre-oxido de cromo. Una fase vapor o una fase mixta que contiene vapor se indica como adecuada en algunos casos. Se ilustra un reactor tubular de flujo continuo utilizando presiones totales del reactor elevadas. La patente britanica n° 1.512.751, que describe la hidrogenacion de la gamma-butirolactona a 1,4-butanodiol en fase lfquida sobre un catalizador de oxido de cobreoxido de cromo. Se ilustran reacciones discontinuas que tienen presiones totales de reactor elevadas y, cuando sea determinable, las presiones parciales de reactivos y productos estaran tambien por encima de los respectivos puntos de rodo. La patente de Estados Unidos n° 4.301.077, que describe la hidrogenacion a 1,4-butanodiol de la gammabutirolactona sobre un catalizador de Ru--NI--Co--Zn. La reaccion se puede llevar a cabo en fase lfquida o fase gaseosa o en una fase lfquido-gas mixta. Se ilustran reacciones en fase lfquida de flujo continuo a presiones totales del reactor elevadas y productividades de reactor relativamente bajas. La patente de Estados Unidos n° 4.048.196, que describe la produccion de 1,4-butanodiol mediante la hidrogenacion en fase lfquida de gamma-butirolactona sobre un catalizador de oxido de cobre-oxido de cinc. Se ilustra ademas un reactor tubular de flujo continuo que funciona a presiones totales de reactor elevadas y a presiones parciales de reactivos y productos elevadas. Y la patente de Estados Unidos n° 4.652.685, que describe la hidrogenacion de lactonas a glicoles.
Un compuesto posterior adicional que se puede producir a partir de organismos microbianos no naturales productores de 4-HB de la divulgacion incluye, por ejemplo, THF. Este compuesto se puede sintetizar mediante, por ejemplo, hidrogenacion qmmica de GBL. Un procedimiento ilustrativo bien conocido en la tecnica aplicable para la conversion de GBL a THF incluye, por ejemplo, la reduccion qmmica de 4-HB y/o GBL o una mezcla de estos dos componentes que se derivan del cultivo usando un catalizador de hidrogenacion heterogeneo u homogeneo junto con hidrogeno, o un agente reductor de tipo hidruro usado estequiometrica o catalfticamente, para producir tetrahidrofurano. Son igualmente aplicables otros procedimientos bien conocidos en la tecnica para la anterior reaccion qmmica e incluyen, por ejemplo, la patente de Estados Unidos n° 6.686.310, que describe una ruta sol-gel de area superficial elevada preparada para catalizadores de hidrogenacion. Se describen tambien procesos para la reduccion de gamma butirolactona a tetrahidrofurano y 1,4-butanodiol.
Las condiciones de cultivo pueden incluir, por ejemplo, procedimientos de cultivo ftquido, asf como fermentacion y otros procedimientos de cultivo a gran escala. Como se describe adicionalmente a continuacion en los Ejemplos, se pueden obtener rendimientos particularmente utiles de los productos biosinteticos de la divulgacion en condiciones de cultivo anaerobias o sustancialmente anaerobias.
La divulgacion proporciona ademas un metodo de fabricacion de 4-HB. El metodo incluye fermentar un organismo microbiano que no se produce naturalmente que tiene una ruta biosintetica del acido 4-hidroxibutanoico (4-HB) que comprende al menos un acido nucleico exogeno que codifica la 4-hidroxibutanoato deshidrogenasa, semialdehfdo sucdnico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdeddo sucdnico deshidrogenasa dependiente de CoA, glutamato: semialdeddo succmico transaminasa o glutamato decarboxilasa en condiciones sustancialmente anaerobias durante un periodo de tiempo suficiente para producir acido 4-hidroxibutanoico monomerico (4-HB), comprendiendo el proceso una fermentacion de alimentacion discontinua y una separacion discontinua; fermentacion de alimentacion discontinua y separacion continua, o fermentacion continua y separacion continua.
El cultivo y las hidrogenaciones qmmicas descritas anteriormente se pueden escalar hasta y crecer de forma continua para fabricar 4-HB, GBL, BDO y/o THF. Los procedimientos de crecimiento ilustrativos incluyen, por ejemplo, fermentacion de alimentacion discontinua y separacion discontinua; fermentacion de alimentacion discontinua y separacion continua, o fermentacion continua y separacion continua. Todos estos procesos son bien conocidos en la tecnica. Emplear los productores de 4-HB permite la biosmtesis simultanea de 4-Hb y la conversion qmmica en GBL, BDO y/o t Hf empleando los anteriores procedimientos de hidrogenacion simultaneos con metodos de cultivos continuos tales como fermentacion. Son tambien conocidos en la tecnica otros procedimientos de hidrogenacion y se pueden aplicar igualmente a los metodos de la divulgacion.
Los procedimientos de fermentacion son particularmente utiles para la produccion biosintetica de cantidades comerciales de 4-HB y/o BDO. En general, y como con procedimientos de cultivo no continuos, la produccion
continua y/o casi continua de 4-HB o BDO incluira cultivar un organismo productor de 4-HB o BDO no natural de la invencion en nutrientes y medio suficientes para sostener y/o sostener casi el crecimiento en fase exponencial. El cultivo continuo en dichas condiciones puede incluir, por ejemplo, 1 d^ a, 2, 3, 4, 5, 6 o 7 dfas o mas. Ademas, el cultivo continuo puede incluir 1 semana, 2, 3, 4 o 5 o mas semanas y hasta varios meses. Como alternativa, los microorganismos de la divulgacion pueden cultivarse durante horas, si resulta adecuado para una aplicacion concreta. Debe entenderse que las condiciones de cultivo continuas y/o casi continuas pueden incluir tambien todos los intervalos de tiempo entre estos periodos ilustrativos.
Son bien conocidos en la tecnica los procedimientos de fermentacion. En resumen, la fermentacion para la produccion biosintetica de 4-HB, BDO u otros productos derivados de 4-HB de la divulgacion se puede utilizar en, por ejemplo, fermentacion de alimentacion discontinua y separacion discontinua; fermentacion de alimentacion discontinua y separacion continua, o fermentacion continua y separacion continua. Los ejemplos de procedimientos de fermentacion discontinua y continua bien conocidos en la tecnica se ilustran adicionalmente a continuacion en los Ejemplos.
Ademas, con respecto a los procedimientos de fermentacion anteriores utilizando los productores de 4-HB o BDO de la divulgacion para la produccion continua de cantidades sustanciales de 4-HB y BDO monomericos, respectivamente, los productores de 4-HB tambien pueden, por ejemplo, someterse simultaneamente a procedimientos de smtesis qmmica como se ha descrito anteriormente para la conversion qmmica del 4-HB monomerico en, por ejemplo, GBL, BDO y/o THF. Los productores de BDO pueden, de manera similar, por ejemplo, someterse simultaneamente a procedimientos de smtesis qmmica como se ha descrito anteriormente para la conversion qmmica de BDO en, por ejemplo, THF, GBL, pirrolidonas y/u otros compuestos de la familia BDO. Ademas, los productos de los productores de 4-HB y BDO pueden separarse del cultivo de fermentacion y someterse secuencialmente a conversion qmmica, como se divulga en el presente documento.
En resumen, se puede llevar a cabo la hidrogenacion de GBL en el caldo de fermentacion como se describe en Frost et al., Biotechnology Progress 18: 201-211 (2002). Otro procedimiento para la hidrogenacion durante la fermentacion incluye, por ejemplo, los metodos descritos en, por ejemplo, la patente de Estados Unidos N° 5.478.952. Este metodo se ilustra ademas en los ejemplos siguientes.
Por tanto, la invencion proporciona adicionalmente un metodo de fabricacion de Y-butirolactona (GBL), tetrahidrofurano (THF) o 1,4-butanodiol (BDO). El metodo incluye fermentar un organismo microbiano que no se produce naturalmente que tiene las rutas biosinteticas del acido 4-hidroxibutanoico (4-HB) y 1,4-butanodiol (BDO), las rutas comprenden al menos un acido nucleico exogeno que codifica la 4-hidroxibutanoato deshidrogenasa, semialdelmdo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdehndo succmico deshidrogenasa dependiente de CoA, 4-hidroxibutirato:CoA transferasa, glutamato: semialdehndo succmico transaminasa, glutamato descarboxilasa, 1,4-butanodiol semialdehndo deshidrogenasa independiente de CoA, 1,4-butanodiol semialdehndo deshidrogenasa dependiente de CoA, 1,4-butanodiol alcohol deshidrogenasa, tanto dependiente de CoA como independiente de CoA, en condiciones sustancialmente anaerobias durante un periodo de tiempo suficiente para producir 1,4-butanodiol (BDO), GBL o THF, comprendiendo la fermentacion una fermentacion de alimentacion discontinua y separacion discontinua; fermentacion de alimentacion discontinua y separacion continua, o fermentacion continua y separacion continua.
Ademas de la biosmtesis de 4-HB, BDO y otros productos de la divulgacion como se ha descrito en el presente documento, se pueden utilizar tambien los organismos microbianos no naturales y los metodos de la invencion en diversas combinaciones entre sf y con otros organismos microbianos y metodos bien conocidos en la tecnica para conseguir la biosmtesis del producto mediante otras rutas. Por ejemplo, una alternativa para producir BDO diferente que usan los productores de 4-HB y las etapas qmmicas o diferentes que usan los productores de BDO es directamente mediante la adicion de otro organismo microbiano capaz de convertir 4-HB o un producto de 4-HB ilustrado en el presente documento en BDO.
Uno de dichos procedimientos incluye, por ejemplo, la fermentacion de un organismo microbiano productor de 4-HB de la divulgacion para producir 4-HB, como se describe anteriormente y a continuacion. A continuacion, se puede usar el 4-HB como un sustrato para un segundo organismo microbiano que convierte 4-HB en, por ejemplo, BDO, GBL y/o THF. El 4-HB se puede anadir directamente a otro cultivo del segundo organismo, o el cultivo original de los productores de 4-HB se puede agotar de estos organismos microbianos por, por ejemplo, separacion celular, y despues, la adicion posterior del segundo organismo del caldo de fermentacion se puede utilizar para producir el producto final sin etapas de purificacion intermedias. Un segundo organismo ilustrativo que tiene la capacidad de utilizar bioqmmicamente 4-HB como sustrato para la conversion de BDO, por ejemplo, es Clostridium acetobutylicum (vease, por ejemplo, Jewell et al., Current Microbiology, 13:215-19 (1986)).
En otras divulgaciones, los organismos microbianos no naturales y los metodos de la divulgacion se pueden ensamblar en una amplia variedad de subrutas para conseguir la biosmtesis de, por ejemplo, 4-HB y/o BDO como se ha descrito. En estas divulgaciones, se pueden segregar rutas biosinteticas para un producto deseado en diferentes organismos microbianos y los diferentes organismos microbianos se pueden cultivar simultaneamente para producir el producto final. En dicho esquema biosintetico, el producto de un organismo microbiano es el sustrato de un
segundo organismo microbiano hasta que se sintetiza el producto final. Por ejemplo, se puede llevar a cabo la biosmtesis de BDO como se ha descrito anteriormente para construir un organismo microbiano que contiene las rutas biosinteticas para la conversion de un sustrato tal como succinato endogeno mediante 4-HB en el producto final BDO. Como alternativa, tambien puede producirse BDO biosinteticamente a partir de organismos microbianos mediante cultivo simultaneo o fermentacion simultanea usando dos organismos en el mismo recipiente. Siendo un primer organismo microbiano un productor de 4-HB con genes para producir 4-HB a partir de acido succmico, y siendo un segundo organismo microbiano un productor de BDO con genes para convertir 4-HB en BDO.
Con las ensenanzas y directrices que se proporcionan en el presente documento, los expertos en la materia entenderan que existe una amplia variedad de combinaciones y permutaciones de organismos microbianos que no se producen naturalmente y los metodos de la divulgacion junto con los otros organismos microbianos, con el cultivo simultaneo de otros organismos microbianos que no se producen naturalmente que tienen subrutas y con combinaciones de otros procedimientos qmmicos y/o bioqmmicos bien conocidos en la tecnica para producir productos de 4-HB, BDO, GBL y THF.
Un metodo informatico para identificar y disenar alteraciones metabolicas que favorecen la biosmtesis de un producto es la infraestructura informatica OptKnock, Burgard et al., Biotechnol Bioeng, 84: 647-57 (2003). OptKnock es un programa de modelacion y simulacion metabolica que sugiere estrategias de delecion de genes que dan como resultado microorganismos geneticamente estables que producen un exceso del producto diana. Espedficamente, la infraestructura examina la totalidad de la red metabolica y/o bioqmmica de un microorganismo con el fin de sugerir manipulaciones geneticas que fuercen la bioqmmica deseada a ser un subproducto obligatorio del crecimiento celular. Al acoplar la produccion bioqmmica con el crecimiento de la celula a traves de deleciones genicas u otras perturbaciones de los genes funcionales estrategicamente situadas, las presiones de seleccion sobre el crecimiento impuestas por las cepas disenadas por ingeniena genetica despues de largos penodos de tiempo en un biorreactor conducen a mejoras en el rendimiento como resultado de la produccion bioqmmica acoplada al crecimiento obligatoria. Finalmente, cuando se realizan las deleciones genicas, existe una posibilidad poco importante de que las cepas disenas reviertan a su estado original porque los genes seleccionados por OptKnock se eliminan completamente del genoma. Por tanto, esta metodologfa informatica se puede utilizar bien para identificar rutas alternativas que conducen a la biosmtesis de 4-HB y/o BDO o bien utilizarse en relacion con organismos microbianos no naturales para optimizacion adicional de la biosmtesis de 4-HB y/o BDO.
En resumen, OptKnock es un termino que se utiliza en el presente documento para referirse a un metodo y sistema informatico para modelar el metabolismo celular. El programa OptKnock se refiere a una infraestructura de modelos y metodos que incorporan restricciones concretas a los modelos de analisis del balance de flujo (FBA). Estas restricciones incluyen, por ejemplo, informacion cinetica cualitativa, informacion cualitativa reguladora, y/o datos experimentales de micromatrices de ADN. OptKnock tambien calcula soluciones a diversos problemas metabolicos mediante, por ejemplo, estrechar los lfmites de flujo derivados de los modelos de balance de flujo y, posteriormente, sondear los lfmites de rendimiento de las redes metabolicas en la presencia de adiciones o deleciones de genes. La infraestructura informatica OptKnock permite la construccion de formulaciones modelo que permiten una consulta efectiva de los lfmites de rendimiento de las redes metabolicas, y proporciona metodos para resolver los problemas de programacion lineal entera mixta. Los metodos de modelacion y simulacion metabolica citados en el presente documento como OptKnock se describen en, por ejemplo, la solicitud de patente estadounidense con numero de serie 10/043.440, presentada el 10 de enero de 2002, y en la patente internacional n° PCT/US02/00660, presentada el 10 de enero de 2002.
Otro metodo informatico para identificar y disenar alteraciones metabolicas que favorecen la produccion biosintetica de un producto es un sistema de modelacion y simulacion metabolica denominado SimPheny®. Este sistema y metodo informatico se describe en, por ejemplo, la solicitud de patente estadounidense con numero de serie 10/173.547, presentada el 14 de junio de 2002, y en la solicitud de patente internacional n° PCT/US03/18838, presentada el 13 de junio de 2003.
SimPheny es un sistema informatico que se puede utilizar para producir un modelo de red in silico y para simular el flujo de masa, energfa o carga a traves de las reacciones qmmicas de un sistema biologico para definir un espacio de soluciones que contiene todas y cada una de las posibles funcionalidades de las reacciones qmmicas del sistema, determinando de esta forma una gama de actividades permitidas por el sistema biologico. Este enfoque se denomina como modelacion basada en restricciones, porque el espacio de soluciones esta definido por restricciones tales como la estequiometna conocida de las reacciones incluidas, asf como por la termodinamica de las reacciones y las restricciones de capacidad asociadas con los flujos maximos a traves de las reacciones. El espacio definido por estas restricciones se puede interrogar para determinar las capacidades fenotfpica y conductual del sistema biologico o de sus componentes bioqmmicos. Los metodos de analisis tales como el analisis convexo, la programacion lineal y el calculo de rutas extremas descritos, por ejemplo, en Schilling et al., J. Theor. Biol. 203:229 248 (2000); Schilling et al., Biotech. Bioeng. 71:286-306 (2000) y Schilling et al., Biotech. Prog. 15:288-295 (1999), se pueden utilizar para determinar dichas capacidades fenotfpicas. Como se describe en los siguientes Ejemplos, esta metodologfa de calculo se utilizo para identificar y analizar las rutas biosinteticas de 4-HB, tanto posibles como optimas, en organismos microbianos no productores de 4-HB.
Tal como se ha descrito anteriormente, un metodo basado en restricciones utilizado en los programas informaticos aplicables a la invencion es el analisis del balance de flujo. En analisis del balance de flujo se basa en equilibrar los flujos en estado estacionario, y se puede implementar como se describe en, por ejemplo, Varma y Palsson, Biotech. Bioeng. 12:994-998 (1994). El enfoque del balance de flujo se ha aplicado a redes de reacciones para simular o predecir propiedades sistemicas de, por ejemplo, el metabolismo de los adipocitos tal y como se describe en Fell y Small, J. Biochem. 138:781-786 (1986), secrecion de acetato desde E. coli en condiciones de maximizacion de ATP como se describe en Majewski y Domach, Biotech. Bioeng. 35:732-738 (1990) o la secrecion de etanol por levaduras como se describe en Vanrolleghem et al., Biotech. Prog. 12:434-448 (1996). Ademas, este enfoque se puede utilizar para predecir o simular el crecimiento de E. coli en una variedad de fuentes de carbono unico, asf como el metabolismo de H. influenzae como se describe en Edwards y Palsson, Proc. Natl. Acad. Sci. USA 97:5528-5533 (2000) , Edwards y Palsson, J. Biol. Chem. 274:17410-17416 (1999) y Edwards et al., Nature Biotech. 19:125-130 (2001) .
Una vez que se ha definido el espacio de soluciones, se puede analizar para determinar posibles soluciones en diferentes condiciones. Este enfoque informatico es coherente con la realidad biologica porque los sistemas biologicos son flexibles y pueden alcanzar el mismo resultado de muchas formas diferentes. Los sistemas biologicos estan disenados mediante mecanismos evolutivos que han sido restringidos por restricciones fundamentales que deben afrontar todos los sistemas vivos. Por tanto, la estrategia de modelado basado en restricciones abarca estas realidades generales. Ademas, la capacidad de imponer continuamente restricciones adicionales sobre un modelo de red mediante el endurecimiento de las restricciones da como resultado una reduccion en el tamano del espacio de soluciones, mejorando de esta forma la precision, por lo que se puede predecir el comportamiento fisiologico o el fenotipo.
Con las ensenanzas y directrices que se proporcionan en el presente documento, un experto en la tecnica podra aplicar diferentes infraestructuras informaticas para la modelacion y simulacion metabolica para disenar e implementar la biosmtesis de 4-HB, BDO, GBL, THF y otros compuestos de la familia BDO en organismos hospedadores microbianos diferentes a E. coli y levadura. Dichos metodos de modelacion y simulacion metabolica incluyen, por ejemplo, los sistemas informaticos anteriormente ilustrados como SimPheny® y OptKnock. Para ilustracion de la invencion, algunos metodos se han descrito en el presente documento con referencia a la infraestructura informatica OptKnock para modelacion y simulacion. Los expertos en la materia sabran como aplicar la identificacion, diseno e implementacion de las alteraciones metabolicas usando OptKnock a cualquiera otra de estas infraestructuras informaticas de modelacion y simulacion y metodos bien conocidos en la tecnica.
La capacidad de una celula u organismo para producir un producto bioqmmico de forma biosintetica se puede ilustrar en el contexto de los lfmites de produccion bioqmmica de una red metabolica tfpica calculada usando un modelo in silico. Estos lfmites se obtienen fijando la tasa o tasas de recaptacion del sustrato o sustratos limitantes a sus valores experimentalmente determinados, y calculando las tasas maximas y mmimas de produccion bioqmmica para cada nivel de crecimiento que se puede alcanzar. La produccion de un producto bioqmmico deseado generalmente esta en competencia directa con la formacion de recursos intracelulares. En estas circunstancias, una velocidad de produccion bioqmmica mejorada necesariamente dara como resultado tasas de crecimiento inferiores a las maximas. Las desactivaciones sugeridas mediante los programas de modelacion y simulacion metabolica tales como OptKnock estan disenadas para restringir los lfmites de la solucion permitidos, forzando un cambio en el comportamiento metabolico de la cepa natural. Aunque los lfmites reales de solucion para una cepa dada se ampliaran o se restringiran a medida que la tasa o tasas de recaptacion de sustrato aumenten o disminuyan, cada punto experimental estara dentro de los lfmites de la solucion calculada. Las representaciones graficas, tales como las que permiten predicciones exactas de lo cerca que estan las cepas disenadas de sus lfmites de comportamiento tambien indican todo lo que se puede mejorar.
La infraestructura matematica OptKnock se ilustra en el presente documento para detectar deleciones de genes que llevan a la biosmtesis de productos y, particularmente, a la biosmtesis de productos acoplada con el crecimiento. El procedimiento construye un modelo metabolico basado en restricciones que estrecha la gama de posibles fenotipos que puede mostrar un sistema celular mediante la imposicion sucesiva de restricciones fisicoqmmicas de control, Price et al., Nat. Rev. Microbiol., 2: 886-97 (2004). Tal como se ha descrito anteriormente, los modelos y simulaciones basados en restricciones son bien conocidos en la tecnica y por lo general implican la optimizacion de una diana celular espedfica, sometida a la estequiometna de la red, para sugerir una distribucion de flujo probable.
En resumen, la maximizacion de un objetivo celular cuantificado como flujo de reaccion agregado para una red metabolica estacionaria que comprende un conjunto N={1, . . . , N} de metabolitos y un conjunto M={1, . . . , M} de reacciones metabolicas se expresa matematicamente de la siguiente forma:
m a x im iz a r sometido a 
Vsustrato _ Vsustrato_captaci6n mmol/gD^V'h

Vj =20 W ■ _
V / t {reaccion irrev.}
donde Sy es el coeficiente estequiometrico del metabolite i en la reaccion j, Vj es el flujo de la reaccion j, Vsustrato_captacion representa la tasa o tasas de recaptacion teoricas o medidas del sustrato o sustratos limitantes, y Vatp_principal es el ATP de mantenimiento necesario no asociado al crecimiento. El vector v incluye tanto los flujos internos como los externos. En este estudio, se considera frecuentemente que el objetivo celular es la obtencion de precursores biosinteticos en las proporciones necesarias para la formacion de biomasa, Neidhardt, F. C. et al., Escherichia coli and Salmonella: Cellular and Molecular Biology, 2a ed. 1996, Washington, D.C.: ASM Press. 2 v. (xx, 2822, lxxvi). Los flujos por lo general se indican por 1 gDWh (gramo de peso seco multiplicado por hora) de tal manera que la formacion de la biomasa se expresa como g de biomasa producida/gDWh o 1/h.
El modelado de las deleciones genicas, y dicha eliminacion de reacciones, utiliza en primer lugar la incorporacion de variables binarias a la estructura teorica basada en restricciones, Burgard et al., Biotechnol. Bioeng., 74: 364-375 (2001), Burgard et al., Biotechnol. Prog., 17: 791-797 (2001). Estas variables binarias

suponen un valor de 1 si la reaccion j esta activa y un valor de 0 si esta inactiva. La siguiente restriccion,

garantiza que el flujo de la reaccion, Vj, se ajusta a cero solamente si la variable yj es igual a cero. Como alternativa, cuando yj es igual a uno, Vj puede tomar cualquier valor entre un lfmite inferior Vjmin y un lfmite superior Vjmax. En el presente documento, Vjmin y Vjmax se identifican minimizando y maximizando, respectivamente, cada flujo de reaccion sometido a las restricciones de la red anteriormente descritas, Mahadevan et al., Metab. Eng., 5: 264-76 (2003). Las desactivaciones de genes/reacciones optimas se identifican resolviendo un problema de optimizacion de dos niveles que selecciona el conjunto de reacciones activas (yj=1) de tal forma que una solucion de crecimiento optimo de la red resultante produce un exceso del producto qmmico de interes. Desde el punto de vista matematico, este problema de optimizacion de dos niveles se expresa en forma del siguiente problema de optimizacion entero mixto de dos niveles:
Maximizar Vquimica (OptKnock) Vj
sometido a maximizar Vbiomasa
Vj
M
sometido a 
Z /=i V / = f t Vsustrato =
V sustrato_captacion V / £ {sustrato(s)  limitante(s)} Vatp — Vatp_principal J VBiomasa — Vbiomasa
limitante(s)} Vatp — Vatp_principal J VBiomasa — Vbiomasa
donde Vquimico es la produccion del producto diana deseado, por ejemplo, succinato u otro producto bioqufmico, y K es el numero de desactivaciones permisibles. Destacar que poner el valor de K en cero devuelve la solucion de biomasa maxima de la red completa, mientras que poner el valor de K en uno identifica la unica desactivacion de gen/reaccion (y =0) de tal forma que la red resultante implica la maxima sobreproduccion dado su maximo rendimiento de biomasa. La restriccion final garantiza que la red resultante cumple un mfnimo rendimiento de biomasa. Burgard et al., Biotechnol. Bioeng., 84: 647-57 (2003), proporcionan una descripcion mas detallada de la formulacion del modelo y del procedimiento de resolucion. Los problemas que contienen cientos de variables binarias se pueden resolver en unos pocos minutos o en varias horas usando CPLEX 8.0, GAMS: The Solver Manuals. 2003: GAMS Development Corporation, que se puede acceder a traves de GAMS, Brooke et al., GAMS Development Corporation (1998), un entorno de modelado, en una estacion de trabajo IBM RS6000-270. La infraestructura OptKnock ya ha sido capaz de identificar estrategias de delecion de genes prometedoras para la produccion bioqufmica en exceso, Burgard et al., Biotechnol. Bioeng., 84: 647-57 (2003), Pharkya et al., Biotechnol. Bioeng., 84: 887-899 (2003), y establece un marco sistematico que abarca de forma natural futuras mejoras en marcos de modelizacion metabolica y reguladora.
Cualquier solucion del problema OptKnock de dos niveles anteriormente descrito proporcionara un conjunto de reacciones metabolicas a perturbar. La eliminacion de cada reaccion dentro del conjunto o la modificacion metabolica puede dar como resultado 4-HB o BDO como producto obligado durante la fase de crecimiento del organismo. Como las reacciones son conocidas, una solucion del problema OptKnock de dos niveles tambien proporcionara el gen o genes asociados que codifican una o mas enzimas que catalizan cada reaccion dentro del conjunto de reacciones. La identificacion de un conjunto de reacciones y sus correspondientes genes que codifican los enzimas que participan en cada reaccion es generalmente un proceso automatizado, que se realiza mediante la correlacion de las reacciones con una base de datos de reacciones que incluye relaciones entre enzimas y genes codificantes.
Una vez identificado, el conjunto de reacciones a perturbar para obtener la produccion de 4-HB o BDO se implementa en la celula u organismo diana mediante la perturbacion funcional de al menos un gen que codifica cada reaccion metabolica dentro del conjunto. Un medio especialmente util para conseguir la perturbacion funcional del conjunto de reaccion es mediante la delecion de cada gen de codificacion. Sin embargo, en algunos casos, puede ser beneficioso perturbar la reaccion mediante otras anomalfas geneticas entre las que se incluyen, por ejemplo, mutacion, delecion de regiones reguladoras tales como promotores o sitios de union a cis de los factores reguladores, o mediante truncamiento de la secuencia de codificacion en cualquiera de numerosas ubicaciones. Estas ultimas anomalfas, que dan como resultado la delecion completa del conjunto genico pueden ser utiles, por ejemplo, cuando se desea una evaluacion rapida del acoplamiento del succinato o cuando es menos probable que ocurra la reversion genica.
Para identificar soluciones productivas adicionales para el problema OptKnock de dos niveles anteriormente descrito que proporcione conjuntos adicionales de reacciones a perturbar o modificaciones metabolicas que puedan dar como resultado una biosfntesis, incluyendo la biosfntesis de 4-HB u otro producto bioqufmico acoplada al crecimiento, se puede implementar otro metodo de optimizacion, denominado de cortes enteros. Este metodo progresa resolviendo el problema OptKnock anteriormente ilustrado de forma iterativa con la incorporacion de una restriccion adicional que se denomina corte entero en cada iteracion. La restriccion de corte entero evita eficazmente que el procedimiento de resolucion seleccione exactamente el mismo conjunto de reacciones identificado en cualquier iteracion anterior que obligatoriamente acopla la biosfntesis del producto con el crecimiento. Por ejemplo, si una modificacion metabolica acoplada con el crecimiento anteriormente identificada especifica que las reacciones 1, 2, y 3 se deber perturbar, entonces, la siguiente restriccion impide las mismas reacciones se consideren simultaneamente en posteriores soluciones: yi+y2+y3 > 1. El metodo de corte entero es bien conocido en la tecnica y se puede encontrar descrito en, por ejemplo, la referencia, Burgard et al., Biotechnol. Prog., 17: 791-797 (2001). Analogamente a todos los metodos descritos en el presente documento con referencia a su uso combinado con la infraestructura informatica OptKnock para modelizacion y simulacion metabolica, el metodo de corte entero para reducir la redundancia en el analisis informatico iterativo tambien se puede aplicar a otras infraestructuras informaticas bien conocidas en la materia incluyendo, por ejemplo, SimPheny®.
Las restricciones del tipo anterior impiden la identificacion de conjuntos de reaccion mas grandes que incluyen conjuntos ya identificados. Por ejemplo, el uso del metodo de optimizacion de corte entero anterior en una iteracion posterior impedirfa la identificacion de un cuadruple conjunto de reacciones que especifique que las reacciones 1, 2, y 3 se deben perturbar, ya que estas reacciones se han identificado con anterioridad. Para garantizar la identificacion de todos los posibles conjuntos de reacciones que conducen a la produccion biosintetica de un producto, se puede utilizar una modificacion del metodo de corte entero.
En resumen, el procedimiento de corte entero modificado comienza con la iteracion 'cero' que calcula la produccion maxima del producto bioqufmico deseado para el crecimiento optimo con una red natural. Este calculo corresponde
a una solucion OptKnock con K igual a 0.
A continuacion, se consideran desactivaciones simples y se introducen dos conjuntos de parametros, objstoreiter e ystoreiter,j, para guardar la funcion objetivo (vquimico) e informacion se activacion-desactivacion de la reaccion (yj), respectivamente, en cada iteracion, iter. A continuacion, las siguientes restricciones se anaden sucesivamente a la formulacion OptKnock en cada iteracion.

En la ecuacion anterior, £ y M son numeros pequenos y grandes, respectivamente. En general, £ se puede ajustar a aproximadamente 0,01 y M se puede ajustar a aproximadamente 1000. Sin embargo, tambien se pueden utilizar numeros mas grandes y/o mas pequenos que los citados. M garantiza que la restriccion solamente puede ser vinculante para estrategias de desactivacion anteriormente identificadas, mientras que £ garantiza que la adicion de desactivaciones a una estrategia previamente identificada debe llevar a un aumento de al menos £ en la produccion bioqufmica durante el crecimiento optimo. Esta estrategia pasa a deleciones dobles cuando una estrategia de delecion unica no consigue mejorar la cepa natural. Despues se consideran las deleciones triples cuando ninguna estrategia de delecion doble mejora la cepa natural, y asf sucesivamente. El resultado final es una lista ordenada, representada por la produccion bioqufmica deseada en el crecimiento optimo, de diferentes estrategias de delecion que se diferencian entre si como mfnimo en una inactivacion. Este procedimiento de optimizacion, asf como la identificacion de una amplia variedad de conjuntos de reacciones que, cuando se perturban, conducen a la biosfntesis, incluyendo la produccion acoplada con el crecimiento, de un producto bioqufmico. Con las ensenanzas y directrices que se proporcionan en el presente documento, los expertos en la materia entenderan que los metodos y disenos de la ingenierfa metabolica ilustrada en el presente documento se pueden aplicar analogamente para identificar nuevas rutas biosinteticas y/o al acoplamiento obligatorio entre el crecimiento de la celula o el microorganismo y cualquier producto bioqufmico.
Los metodos anteriormente ilustrados e ilustrados adicionalmente en los Ejemplos siguientes permiten la construccion de las celulas y organismos que producen de forma biosintetica, incluyendo el acoplamiento obligado entre la produccion de un producto bioqufmico deseado y el crecimiento de la celula u organismo disenados para contener las alteraciones geneticas deseadas. En este sentido, se han identificado alteraciones geneticas que dan como resultado la biosfntesis de 4-HB y 1,4-butanodiol. Las cepas de microorganismos construidas con las alteraciones metabolicas identificadas producen niveles elevados de 4-HB o BDO en comparacion con los microorganismos sin modificar. Estas cepas se pueden utilizar provechosamente para la produccion comercial de 4-HB, BDO, THF y GBL, por ejemplo, en un proceso de fermentacion continuo sin estar sometidas a presiones de seleccion negativa.
Por tanto, los metodos informaticos descritos en el presente documento permiten la identificacion e implementacion de modificaciones metabolicas que se identifican mediante un metodo in silico seleccionado para OptKnock o SimPheny. El conjunto de modificaciones metabolicas puede incluir, por ejemplo, la adicion de una o mas rutas enzimaticas biosinteticas y/o la perturbacion funcional de una o mas reacciones metabolicas entre las que se incluyen, por ejemplo, la perturbacion por delecion del gen.
La aplicacion del metodo OptKnock para identificar rutas adecuadas para la produccion del 1,4-butanodiol (BDO) acoplada al crecimiento se describe en los Ejemplos V-VII. Tal como se ha descrito anteriormente, la metodologfa OptKnock se desarrollo sobre la base de que las redes microbianas mutantes pueden evolucionar hacia sus fenotipos de crecimiento maximo previstos por el modelo cuando se someten a periodos prolongados se seleccion por crecimiento. En otras palabras, la estrategia aprovecha la capacidad del organismo para autooptimizarse bajo presion selectiva. La infraestructura OptKnock permite la enumeracion exhaustiva de combinaciones de deleciones de genes que fuerzan un acoplamiento entre la produccion bioqufmica y el crecimiento de la celula en funcion de la estequiometrfa de la red. La identificacion de desactivaciones de genes/reacciones optimas requiere la resolucion de un problema de optimizacion de dos niveles que selecciona el conjunto de reacciones activas de tal forma que una solucion de crecimiento optimo de la red resultante produce un exceso del producto bioqufmico de interes (Burgard et al. Biotechnol. Bioeng.84:647-657 (2003)).
La produccion bioqufmica acoplada con el crecimiento se puede visualizar en el contexto de los lfmites de produccion bioqufmica de una red metabolica tfpica. Estos lfmites se obtienen a partir de un modelo in silico fijando la tasa o tasas de recaptacion del sustrato o sustratos limitantes a sus valores experimentalmente determinados, y calculando las tasas maximas y mfnimas de produccion bioqufmica para cada nivel de crecimiento que se puede alcanzar. La produccion implica basicamente todo lo que es posible abarcando todos los fenotipos biologicos potenciales disponibles para una cepa dada. Aunque se producen excepciones, tfpicamente, la produccion de un producto bioqufmico deseado esta en competencia directa con la formacion de recursos intracelulares (vease la figura 5, region gris). Asf, un rendimiento bioqufmico mejorado necesariamente dara como resultado un crecimiento inferior al maximo. Adicionalmente, la aplicacion de presiones de seleccion por crecimiento puede impulsar el rendimiento de produccion no acoplada al crecimiento de una cepa hacia un genotipo de baja produccion (punto A,
figura 5), independientemente de su punto de partida inicial. La desactivacion sugerida por OptKnock esta disenada para restringir el Kmite admisible de la solucion, forzando un cambio en el comportamiento metabolico como se representa en la figura 5 (region cian). La estrategia de la ingeniena de evolucion se puede aplicar para impulsar el rendimiento de una cepa disenada mediante OptKnock hacia el Kmite situado mas a la derecha. Esto dara como resultado una cepa de produccion elevada que debena ser intrmsecamente estable para presiones de seleccion por crecimiento adicionales (punto B, figura 5).
Se uso un modelo estequiometrico in silico del metabolismo de E. coli para identificar los genes esenciales de las rutas metabolicas, como se ha ilustrado y descrito anteriormente en, por ejemplo, las publicaciones de patente de Estados Unidos US 2002/0012939, US 2003/0224363, US 2004/0029149, US 2004/0072723, US 2003/0059792, US 2002/0168654 y US 2004/0009466, y en la patente de Estados Unidos. n° 7.127.379. Como se divulga en el presente documento, la infraestructura matematica OptKnock se aplico para detectar deleciones de genes que condudan a una produccion de BDO acoplada al crecimiento (veanse los Ejemplos V-VII). Se determinaron inicialmente estrategias mediante un modelo reducido de la red metabolica de E. coli. Este modelo “pequeno” incluye las funcionalidades metabolicas clave de la red, eliminando de esta forma la redundancia asociada con las redes metabolicas a escala genomica. Se presto atencion a garantizar que el modelo no se habfa reducido hasta el punto de no incluir rutas potencialmente activas que teman dianas viables. En su conjunto, el modelo reducido contema 262 reacciones y su implementacion redujo el tiempo de CPU de OptKnock CPU en aproximadamente diez veces en comparacion con la aplicacion de OptKnock al modelo a escala genomica de E. coli (Reed et al., Genome Biol. 4:R54 (2003)).
Ademas, la solucion del problema OptKnock de dos niveles proporciona solamente un conjunto de deleciones. Para enumerar todas las soluciones significativas, es decir, todos los conjuntos de desactivaciones que conducen a una formacion de la produccion acoplada al crecimiento, se puede implementar una tecnica de optimizacion denominada de cortes enteros. Esto conlleva resolver el problema OptKnock de forma iterativa con la incorporacion de una restriccion adicional que se denomina corte entero en cada iteracion, tal como se ha descrito anteriormente.
Para las rutas bioqmmicas que conducen al 1,4-butanodiol, se espera que la conversion de glucosa en BDO en E. coli derive de un total de tres etapas de reduccion intracelular a partir del semialdetndo succmico (vease la figura 6). E. coli produce de forma natural el semialdetndo succmico mediante el ciclo intermedio TCA, alfa-cetoglutarato, mediante la accion de dos enzimas, la glutamato:semialdehndo succmico transaminasa y glutamato descarboxilasa. Una ruta alternativa, utilizada por el anaerobio estricto Clostridium kluyveri para degradar el succinato, activa el succinato a succinil-CoA, y posteriormente convierte el succinil-CoA en semialdetndo succmico usando una semialdetndo succmico deshidrogenasa dependiente de CoA (Sohling y Gottschalk, Eur. J. Biochem. 212:121-127 (1993)). La conversion de succinato semialdetndo en BDO requiere en primer lugar la actividad de la 4-hidroxibutanoato (4-HB) deshidrogenasa, un enzima no natural para E. coli o levaduras, pero que se encuentra en diversas bacterias tales como C. kluyveri y Ralstonia eutropha (Lutke-Eversloh y Steinbuchel, FEMS Microbiol. Lett.
181:63-71 (1999); Sohling y Gottschalk, J. Bacteriol. 178:871-880 (1996); Valentin et al., Eur. J. Biochem.227:43-60 (1995); Wolff y Kenealy, Protein Expr. Purif. 6:206-212 (1995)). El precedente de la conversion de 4-HB en BDO se ha demostrado en el anaerobio estricto, Clostridium acetobutylicum, que cuando se alimento con 4-HB, demostro producir cuantitativamente BDO (Jewell et al., Current Microbiology 13:215-219 (1986)). Las biotransformaciones necesarias de 4-HB a BDO se suponen similares a las de la conversion de acido butmico en butanol habitual de las especies de Clostridia, que transcurre mediante un derivado de CoA (Girbal et al., FEMS Microbiology Reviews 17:287-297 (1995)).
Se divulga adicionalmente un microorganismo no natural que comprende una o mas perturbaciones genicas, las una o mas perturbaciones genicas se producen en genes que codifican un enzima obligatorio para acoplar la produccion de 1,4-butanodiol al crecimiento del microorganismo cuando la perturbacion del gen reduce la actividad de un enzima, en donde la una o mas perturbaciones genicas transmiten una produccion de 1,4-butanodiol acoplada al crecimiento estable en el microorganismo no natural. La una o mas perturbaciones genicas pueden ser, por ejemplo, las descritas en la Tabla 6 o 7. La una o mas perturbaciones genicas pueden comprender perturbaciones que comprenden una delecion del uno o mas genes, tales como los genes divulgados en la Tabla 6 o 7.
Como se divulga en el presente documento, el microorganismo no natural puede ser una bacteria, levadura u hongo. Por ejemplo, el microorganismo no natural puede ser una bacteria tal como Escherichia coli, Klebsiella oxytoca, Anaerobiospirillum succiniciproducens, Actinobacillus succinogenes, Mannheimia succiniciproducens, Rhizobium etli, Bacillus subtilis, Corynebacterium glutamicum, Gluconobacter oxydans, Zymomonas mobilis, Lactococcus lactis, Lactobacillus plantarum, Streptomyces coelicolor, Clostridium acetobutylicum, Pseudomonas fluorescens, y Pseudomonas putida. El microorganismo no natural tambien puede ser una levadura tal como Saccharomyces cerevisiae, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces marxianus, Aspergillus terreus, Aspergillus niger, y Pichia pastoris.
Un microorganismo no natural de la divulgacion puede comprender un conjunto de modificaciones metabolicas que acoplan obligatoriamente la produccion de 1,4-butanodiol al crecimiento del microorganismo, comprendiendo el conjunto de modificaciones metabolicas la perturbacion de uno o mas genes seleccionados entre el conjunto de
genes que comprenden adhE, IdhA, pflAB; adhE, IdhA, pflAB, mdh; adhE, IdhA, pflAB, mdh, mqo; adhE, IdhA, pflAB, mdh, aspA; adhE, mdh, IdhA, pflAB, sfcA; adhE, mdh, IdhA, pflAB, maeB; adhE, mdh, IdhA, pflAB, sfcA, maeB; adhE, IdhA, pflAB, mdh, pntAB; adhE, IdhA, pflAB, mdh, gdhA; adhE, IdhA, pflAB, mdh, pykA, pykF, dhaKLM, deoC, edd, yiaE, ycdW; y adhE, IdhA, pflAB, mdh, pykA, pykF, dhaKLM, deoC, edd, yiaE, ycdW, prpC, gsk, o un ortologo del mismo, en el que el microorganismo muestra una produccion de 1,4-butanodiol acoplada al crecimiento estable.
La divulgacion proporciona un microorganismo no natural que comprende un conjunto de modificaciones metabolicas obligadas para acoplar la produccion de 1,4-butanodiol al crecimiento del microorganismo, comprendiendo el conjunto de modificaciones metabolicas la perturbacion de uno o mas genes, o un ortologo del mismo, en el que el conjunto de modificaciones metabolicas comprende la perturbacion de adhE y IdhA, en el que el microorganismo muestra una produccion de 1,4-butanodiol acoplada al crecimiento estable. El conjunto de modificaciones metabolicas puede comprender ademas la perturbacion de mdh. El conjunto de modificaciones metabolicas puede comprender ademas la perturbacion de uno o mas genes seleccionados entre el conjunto de genes que comprenden mqo, aspA, sfcA, maeB, pntAB, y gdhA y puede incluir, por ejemplo, la perturbacion de sfcA y maeB. El conjunto de modificaciones metabolicas puede comprender ademas la perturbacion de uno o mas genes seleccionados entre el conjunto de genes que comprenden pykA, pykF, dhaKLM, deoC, edd, yiaE, ycdW, prpC, y gsk, incluyendo la perturbacion de todos los pykA, pykF, dhaKLM, deoC, edd, yiaE y ycdW, y puede comprender ademas la perturbacion de prpC y gsk. Cualquiera de los conjuntos de modificaciones metabolicas anteriormente descritas puede comprender ademas la perturbacion de pflAB. En una divulgacion particular, el conjunto de modificaciones metabolicas comprende la perturbacion de uno o mas genes seleccionados entre el conjunto de genes que comprenden adhE, IdhA, pflAB, mdh, y aspA, incluyendo hasta la totalidad de los genes adhE, IdhA, pflAB, mdh, y aspA.
Un microorganismo no natural de la divulgacion puede comprender ademas una ruta biosintetica 1,4-butanodiol (BDO) que comprende al menos un acido nucleico exogeno que codifica la 4-hidroxibutanoato deshidrogenasa, semialdehndo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdehndo succmico deshidrogenasa dependiente de CoA, 4-hidroxibutirato:CoA transferasa, glutamato: semialdetndo succmico transaminasa, glutamato descarboxilasa, aldehndo deshidrogenasa independiente de CoA, aldehndo deshidrogenasa dependiente de CoA o alcohol deshidrogenasa, en el que el acido nucleico exogeno se expresa en cantidad suficiente para producir 1,4-butanodiol (BDO), como se divulga en el presente documento.
La divuglacion proporciona adicionalmente un metodo para producir un microorganismo no natural que tiene una produccion de 1,4-butanodiol acoplada al crecimiento estable identificando in silico un conjunto de modificaciones metabolicas que requieren la produccion de 1,4-butanodiol durante el crecimiento exponencial bajo un conjunto definido de condiciones, y modificar geneticamente un microorganismo para que contenga el conjunto de modificaciones metabolicas que requieren la produccion de 1,4-butanodiol. Dichos metodos pueden incluir adicionalmente la adicion de genes exogenos que expresan las actividades enzimaticas deseadas en un microorganismo. Dicho conjunto de modificaciones metabolicas se puede identificar por un metodo in silico seleccionado entre OptKnock o SimPheny (vease anteriormente y los Ejemplos V-VII).
Ademas, la invencion proporciona un metodo para producir 1,4-butanodiol acoplado al crecimiento de un microorganismo. El metodo puede incluir las etapas de cultivar en la fase de crecimiento exponencial, en cantidad suficiente de nutrientes y medio, un microorganismo no natural que comprende un conjunto de modificaciones metabolicas que acoplan obligatoriamente la produccion de 1,4-butanodiol al crecimiento del microorganismo, en el que el microorganismo muestra una produccion de 1,4-butanodiol acoplada al crecimiento estable, y aislar el 1,4-butanodiol producido a partir del microorganismo no natural. El conjunto de modificaciones metabolicas que comprende la perturbacion de uno o mas genes se puede seleccionar entre el conjunto de genes que comprenden adhE, IdhA, pflAB; adhE, IdhA, pflAB, mdh; adhE, IdhA, pflAB, mdh, mqo; adhE, IdhA, pflAB, mdh, aspA; adhE, mdh, IdhA, pflAB, sfcA; adhE, mdh, IdhA, pflAB, maeB; adhE, mdh, IdhA, pflAB, sfcA, maeB; adhE, IdhA, pflAB, mdh, pntAB; adhE, IdhA, pflAB, mdh, gdhA; adhE, IdhA, pflAB, mdh, pykA, pykF, dhaKLM, deoC, edd, yiaE, ycdW; y adhE, IdhA, pflAB, mdh, pykA, pykF, dhaKLM, deoC, edd, yiaE, ycdW, prpC, gsk, o un ortologo del mismo.
En una realizacion, la divulgacion proporciona un metodo para producir 1,4-butanodiol acoplado al crecimiento de un microorganismo. El metodo puede incluir las etapas de cultivar en la fase de crecimiento exponencial, en cantidad suficiente de nutrientes y medio, un microorganismo no natural que comprende un conjunto de modificaciones metabolicas que acoplan obligatoriamente la produccion de 1,4-butanodiol al crecimiento del microorganismo, comprendiendo el conjunto de modificaciones metabolicas la perturbacion de uno o mas genes, o un ortologo del mismo, en el que el conjunto de modificaciones metabolicas comprende la perturbacion de adhE y IdhA, en el que el microorganismo muestra una produccion de 1,4-butanodiol acoplada al crecimiento estable; y aislar el 1,4-butanodiol producido a partir del microorganismo no natural. En una divulgacion adicional, el conjunto de modificaciones metabolicas puede comprender ademas la perturbacion de mdh. En otra divulgacion, el conjunto de modificaciones metabolicas puede comprender ademas la perturbacion de uno o mas genes seleccionados entre el conjunto de genes que comprenden mqo, aspA, sfcA, maeB, pntAB, y gdhA y puede incluir, por ejemplo, la perturbacion de sfcA y maeB. En otra divulgacion mas, el conjunto de modificaciones metabolicas puede comprender ademas la perturbacion de uno o mas genes seleccionados entre el conjunto de genes que comprenden pykA, pykF, dhaKLM, deoC, edd, yiaE, ycdW, prpC, y gsk, incluyendo la perturbacion de todos los pykA, pykF, dhaKLM, deoC, edd, yiaE y
ycdW, y puede comprender ademas la perturbacion de prpC y gsk. Cualquiera de los conjuntos de modificaciones metabolicas anteriormente descritas puede comprender ademas la perturbacion de pflAB.
En un metodo para producir BDO, el microorganismo no natural puede comprender ademas una ruta biosintetica 1,4-butanodiol (BDO) que comprende al menos un acido nucleico exogeno que codifica la 4-hidroxibutanoato deshidrogenasa, semialdel'ndo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdel'ndo succmico deshidrogenasa dependiente de CoA, 4-hidroxibutirato:CoA transferasa, glutamato: semialdel'ndo succmico transaminasa, glutamato descarboxilasa, aldehndo deshidrogenasa independiente de CoA, aldehndo deshidrogenasa dependiente de CoA o alcohol deshidrogenasa, en el que el acido nucleico exogeno se expresa en cantidad suficiente para producir 1,4-butanodiol (BDO).
Se entiende que tambien se incluyen modificaciones que no afectan sustancialmente a la actividad de las diversas realizaciones de esta invencion dentro de la definicion de la invencion proporcionada en el presente documento. Por consiguiente, se pretende que los siguientes ejemplos ilustren, pero no limiten, la presente invencion.
EJEMPLO I
Biosintesis del acido 4-hidroxibutanoico
Este Ejemplo describe las rutas bioqmmicas para la produccion de 4-HB.
Los informes anteriores sobre la smtesis de 4-HB en microbios se han centrado en este compuesto como un compuesto intermedio en la produccion del plastico biodegradable poli-hidroxialcanoato (PHA) (patente de Estados Unidos n° 6.117.658). El uso de copolfmeros 4-HB/3-HB puede ser ventajoso comparado con el polfmero de poli-3-hidroxibutirato (PHB) mas habitual porque el plastico resultante es menos quebradizo (Saito y Doi, Intl. J. Biol. Macromol. 16:99-104 (1994)). La produccion del 4-HB monomerico descrito en el presente documento es fundamentalmente un proceso diferente por varios motivos: 1). El producto se secreta, a diferencia del PHA, que se produce intracelularmente, y permanece en la celula; 2) para los organismos que producen polfmeros de hidroxibutanoato, no se produce 4-HB libre, sino en su lugar el derivado de Coenzima A, que utiliza la polihidroxialcanoato sintasa; 3) en el caso del polfmero, la formacion de productos granulares altera la termodinamica; y 4) el pH extracelular no es un problema para la produccion del polfmero, mientras que resultara alterado si 4-HB esta presente en forma de acido libre o como base conjugada, y tambien el equilibrio entre 4-HB y GBL.
4-HB se puede producir por dos etapas de reduccion desde succinato, un metabolito central del ciclo TCA, con el semialdetndo succmico como el compuesto intermedio (figura 2). La primera de estas enzimas, la semialdetndo succmico deshidrogenasa, es natural para muchos organismos incluyendo E. coli, en el que se han encontrado las dos enzimas dependientes de NADH y NADPH (Donnelly y Cooper, Eur. J. Biochem.113:555-561 (1981); Donnelly y Cooper, J. Bacteriol. 145:1425-1427 (1981); Marek y Henson, J. Bacteriol. 170:991-994 (1988)). Existe tambien evidencia que respalda la actividad de la semialdel'ndo succmico deshidrogenasa en S. cerevisiae (Ramos et al., Eur. J. Biochem. 149:401-404 (1985)), y se ha identificado un posible gen por homologfa de secuencia. Sin embargo, la mayona de los informes indican que esta enzima progresa en la direccion de la smtesis de succinato, en oposicion a lo que se muestra en la figura 2 (Donnelly y Cooper, mas arriba; Lutke-Eversloh y Steinbuchel, FEMS Microbiol. Lett. 181:63-71 (1999)), participando en la ruta degradativa de of 4-HB y gamma-aminobutirato. Una ruta alternativa, utilizada por el anaerobio estricto Clostridium kluyveri para degradar el succinato, activa el succinato a succinil-CoA, y posteriormente convierte el succinil-CoA en semialdel'ndo succmico usando una semialdel'ndo succmico deshidrogenasa alternativa que se sabe que actua en esta direccion (Sohling y Gottschalk, Eur. J. Biochem.212:121-127 (1993)). Sin embargo, esta ruta tiene el coste energetico del ATP necesario para convertir el succinato a succinil-CoA.
La segunda enzima de la ruta, 4-hidroxibutanoato deshidrogenasa, no es natural para E. coli o levaduras, pero que se encuentra en diversas bacterias tales como C. kluyveri y Ralstonia eutropha (Lutke-Eversloh y Steinbuchel, mas arriba; Sohling y Gottschalk, J. Bacteriol. 178:871-880 (1996); Valentin et al., Eur. J. Biochem. 227:43-60 (1995); Wolff y Kenealy, Protein Expr. Purif. 6:206-212 (1995)). Se sabe que estas enzimas son dependientes de NADH, con lo que se supone que pueden existir formas dependientes de NADPH. Una ruta adicional hacia 4-HB desde alfacetoglutarato se ha demostrado en E. coli dando como resultado la acumulacion de poli(acido 4-hidroxibutmco) (Song et al., Wei Sheng Wu Xue. Bao. 45:382-386 (2005)). La cepa recombinante necesita la sobreexpresion de tres genes heterologos, PHA sintasa (R. eutropha), 4-hidroxibutirato deshidrogenasa (R. eutropha) y 4-hidroxibutirato:CoA transferasa (C. kluyveri), junto con dos genes naturales de E. coli: glutamatensemialdelmdo succmico transaminasa y glutamato descarboxilasa. Las etapas 4 y 5 de la figura 2 pueden llevarse a cabo alternativamente mediante una alfa-cetoglutarato descarboxilasa tal como la identificada en Euglena gracilis (Shigeoka et al., Biochem. J. 282(Pt2):319-323 (1992); Shigeoka y Nakano, Arch. Biochem. Biophys. 288:22-28 (1991); Shigeoka y Nakano, Biochem J.292(Pt 2):463-467 (1993)). Sin embargo, esta enzima aun no se ha aplicado para alterar la produccion de 4-HB o polfmeros relacionados en ningun organismo.
La direccionalidad notificada de la semialdel'ndo succmico deshidrogenasa condujo a la investigacion de la termodinamica del metabolismo de 4-HB. Espedficamente, este estudio investigo si las reacciones implicadas en la
conversion del succinato o succinil-CoA en 4-HB eran termodinamicamente favorables, o no lo eran, (es dedr, AGt<0) en las tipicas condiciones fisiologicas presentes en E. coli y S. cerevisiae. Se supone que todas las reacciones de oxidacion/reduccion utilizan NADH, aunque los resultados para la supuesta utilizacion de NADPH senan similares. Se calcularon las energfas libres de Gibbs estandar de formacion (AGf°) para cada compuesto de la figura 2 basandose en el metodo de contribucion de grupos (Mavrovouniotis, M. L., J. Biol. Chem. 266:14440-14445 (1991)). Cada energfa de Gibbs estandar de formacion se transformo a continuacion para obtener un criterio de cambio espontaneo a la presion, temperatura, pH y fuerza ionica especificados (Alberty, R. A., Biochem. Biophys. Acta 1207:1-11 (1994)) (ecuacion 1).
r - T1 - N
AG A l,p H ) = AG"
( / = 0) N„ RT In( 1 O'”" ) - 2,915V / - ----- ~f=- (1)
f 1
i
nji
Donde AGfo es la ene^a de Gibbs estandar de formacion, Nh es el numero de atomos de hidrogeno en el compuesto, R es la constante universal de los gases, T es constante a 298 K, z es la carga de la molecula al pH de interes, I es la fuerza ionica en M, y B es una constante igual a 1,6 L0,5/mol0,5.
La Ecuacion 1 revela que tanto el pH intracelular como la fuerza ionica tienen un papel para determinar la factibilidad termodinamica. Normalmente, el pH intracelular de las celulas esta muy bien regulado, incluso cuando hay grandes variaciones en el pH del cultivo. El pH intracelular de E. coli y S. cerevisiae se han notificado en la bibliografia. E. coli mantiene un pH intracelular de 7,4-7,7 durante las condiciones de crecimiento tfpicas en tampones neutros, pero puede disminuir a 7,2 en un medio a pH 6, e incluso bajar hasta 6,9 para un pH externo de 5 (Riondet et al., Biotechnology Tech. 11:735-738 (1997)). Sin embargo, el crecimiento de E. coli queda fuertemente inhibido a un pH externo inferior a 6. El pH de las levaduras muestra una variacion mayor. Durante la fase de crecimiento exponencial, el pH interno de S. cerevisiae se ha medido en el intervalo de 6,7-7,0 para un pH externo controlado a 5,0 (Dombek e Ingram, Appl. Environ. Microbiol. 53:1286-1291 (1987)). Por otra parte, en celulas en reposo, el pH interno baja por debajo de 6 cuando el pH externo es 6 o inferior (Imai y Ohno, J. Biotechnol. 38:165-172 (1995)). Este analisis supone un pH intracelular de 7,4 para E. coli y 6,8 para S. cerevisiae. Tambien se supone una fuerza ionica de 0,15 (Valenti et al., mas arriba).
Las energfas de Gibbs de formacion transformadas se calcularon en el estado estandar (pH=7,0, I=0) y para los estados fisiologicos de E. coli (pH=7,4, I=0,15) y S. cerevisiae (pH=6,8, I=0,15). Las energfas de Gibbs de reaccion (AGr) transformadas se calcularon a continuacion calculando la diferencia de AGf entre productos y reactivos. Las energfas de Gibbs de las reacciones transformadas necesarias para convertir succinato o succinil-CoA en 4-HB se proporcionan en la Tabla 2. A resaltar que el error estandar, Uf,est, para la AGf calculada mediante la teona de contribucion de grupos es de 4 kcal/mol. Esta incertidumbre sobre el valor de AGr, Ur,est, se puede calcular como la norma euclidea de la incertidumbre de AGf para cada compuesto (ecuacion).

Donde n es el coeficiente estequiometrico e i es el compuesto. Para las reacciones examinadas, esta incertidumbre es de aproximadamente 8 kcal/mol.
Tabla 2. Energfa libre de Gibbs de reaccion (kcal/mol) para diferentes valores de pH y fuerza ionica. La primera columna esta en condiciones estandar, mientras que las demas se han ajustado de acuerdo con la ecuacion 1. La temperatura es constante a 298 K. Las barras de error de estos valores son de aproximadamente 8 kcal/mol, calculado mediante la ecuacion 2. Abreviaturas: suc, succinato; sucsa, semialdehndo succrnico; succoa, succinil-CoA; Pi, fosfato inorganico.

Despues de considerar la posible incertidumbre en los calculos de los inventores, la Tabla 2 revela que es mas probable que la reaccion de la semialdetndo succrnico deshidrogenasa (etapa 1 de la figura 2) encuentre una barrera termodinamica. Tambien se estudio si esta reaccion se puede acercar a la factibilidad termodinamica variando las concentraciones supuestas de los metabolitos participates. Por ejemplo, las energfas de Gibbs
estandar suponen concentraciones de 1 M para todos los compuestos participates (excepto el agua). En un entorno anaerobio, NADH estara presente en una concentracion varias veces superior a la de NAD. Suponiendo [NADH]=5x[NAD], los inventores calcularon el efecto sobre AGt usando la ecuacion

Este cambio dio como resultado una diferencia de solo aproximadamente 1 kcal/mol en los valores de delta G para la semialdetdo succmico deshidrogenasa. Tambien se utilizo la ecuacion 3 para calcular otros efectos sobre AGt, tales como una concentracion de succinato elevada para impulsar las reacciones. Una diferencia de 1000 veces en las concentraciones de succinato y semialdetndo succmico contribuyo a aproximadamente 5 kcal/mol a delta G. Tomado conjuntamente con una incertidumbre supuesta de 8 kcal/mol, la posibilidad de que la semialdetndo succmico deshidrogenasa actue en la direccion del semialdetndo succmico en algun conjunto de condiciones fisiologicas no se puede eliminar. Asf, los autores siguen considerando la ruta directa desde succinato hasta 4-HB en su analisis teorico posterior.
Se analizaron en dos microbios las capacidades de produccion microbiana de 4-hidroxibutirato, Escherichia coli y Saccharomyces cerevisiae, usando los modelos metabolicos in silico de cada organismo. Las rutas potenciales para dar 4-HB progresan mediante un compuesto de succinato, succinil-CoA, o alfa-cetoglutarato como se muestra en la figura 2.
La primera etapa en la ruta de produccion de 4-HB desde succinato implica la conversion de succinato en semialdetndo succmico mediante un semialdetndo succmico deshidrogenasa dependiente de NADH o NADPH. En E. coli, gabD es una semialdetndo succmico deshidrogenasa dependiente de NADP que forma parte de una agrupacion de genes implicada en la captacion y degradacion del 4-aminobutirato (Niegemann et al., Arch. Microbiol.
160:454-460 (1993); Schneider et al., J. Bacteriol. 184:6976-6986 (2002)). Se cree que sad codifica el enzima para la actividad de la semialdetndo succmico deshidrogenasa dependiente de NAD (Marek and Henson, mas arriba). S. cerevisiae contiene solamente la semialdetndo succmico deshidrogenasa dependiente de NADPH, teoricamente asignada a UGA2, que se encuentra en el citosol (Huh et al., Nature 425:686-691 (2003)). Los calculos del rendimiento maximo suponiendo la ruta del succinato para dar 4-HB tanto en E. coli como en S. cerevisiae requiere solamente la asuncion de que se ha anadido una 4-HB deshidrogenasa no natural a sus redes metabolicas.
La ruta desde succinil-CoA hasta 4-hidroxibutirato se describe en la patente de Estados Unidos n° 6.117.658 como parte de un proceso para fabricar polihidroxialcanoatos que comprenden unidades monomericas de 4-hidroxibutirato. Clostridium kluyveri es un ejemplo de organismo conocido que tiene actividad semialdetndo succmico deshidrogenasa dependiente de CoA (Sohling y Gottschalk, mas arriba; Sohling y Gottschalk, mas arriba). En este estudio, se supone que dicha enzima, derivada de C. kluyveri u otro organismo, se expresa en E. coli o S. cerevisiae junto con una 4-HB deshidrogenasa no natural o heterologa para completar la ruta desde succinil-CoA hasta 4-HB. La ruta desde alfa-cetoglutarato hasta 4-HB se ha demostrado en E. coli dando como resultado la acumulacion de poli(acido 4-hidroxibutmco) hasta un 30% del peso celular seco (Song et al., mas arriba). Como E. coli y S. cerevisiae tienen de forma natural o endogena glutamato:semialdetndo succmico transaminasa y glutamato descarboxilasa (Coleman et al., J. Biol. Chem. 276:244-250 (2001)), la ruta desde AKG hasta 4-HB se puede completar en ambos organismos suponiendo solamente que esta presente una 4-HB deshidrogenasa no natural.
EJEMPLO II
Produccion de acido 4-hidroxibutanoico en E. coli
Este Ejemplo describe los rendimientos biosinteticos del acido 4-hidroxibutanoico resultantes de cada ruta bioqmmica.
En esta seccion, se calcularon los rendimientos teoricos maximos de 4-HB procedente de glucosa suponiendo que cada una de las tres rutas metabolicas representadas graficamente en la figura 2 son funcionales en E. coli. Un modelo metabolico a escala genomica de E. coli, similar al descrito en Reed et al., Genome Biol. 4:R54 (2003), se uso como base para el analisis. Se calculo la ganancia energetica, en terminos de moleculas de ATP producidas, de cada ruta de rendimiento maximo suponiendo condiciones anaerobias, salvo que se indique otra cosa. Se supone que el 4-hidroxibutirato sale de E. coli mediante el mecanismo de cotransporte paralelo de protones, como es el caso de la mayona de los acidos organicos. Es tambien posible que GBL se secrete por difusion simple, y, en este caso, el rendimiento energetico sena mas favorable que en el caso considerado aqrn. Tambien se investigo el impacto de la especificidad del cofactor (es decir, la dependencia de NADH o NADPH) de los enzimas participates sobre el rendimiento maximo y el rendimiento energetico de cada ruta.
En las Tablas 3 A-C se muestran los resultados del analisis. Desde un punto de vista energetico y de rendimiento, la ruta del succinato a 4-HB es la mas prometedora con la condicion que se puedan superar las preocupaciones
termodinamicas planteadas en el Ejemplo I. Espedficamente, los calculos revelan que el rendimiento maximo teorico de 4-HB a partir de glucosa es de 1,33 mol/mol (0,77 g/g; 0,89 moles C/moles C) suponiendo que la ruta de 4-HB a succinato es funcional. Ademas, la produccion anaerobia de 4-HB mediante succinato dana como resultado una produccion neta de 1,8, 1,5, o 1,1 mol de ATP por glucosa dependiendo de la especificidad del cofactor supuesta de los enzimas participates. Estos rendimientos energeticos son comparables con los 2,0 ATP por glucosa que se pueden obtener mediante fosforilacion del sustrato debido a la produccion de etanol o lactato sugiriendo el potencial de la produccion anaerobia de homo-4-HB en E. coli.
La ruta de la succinil-CoA de 4-HB es la segunda ruta mas prometedora cuando se considera el rendimiento maximo y el rendimiento energetico. Se puede conseguir en E. coli un rendimiento de 1,33 mol/mol de 4-HB si al menos una de las etapas de la ruta se supone dependiente de NADH. Sin embargo, como esta ruta requiere la formacion de succinil-CoA, su rendimiento energetico es considerablemente menor que el de la ruta del succinato. Se anticipa un requerimiento de oxfgeno a altos rendimientos de 4-HB si se supone que las etapas de la semialdetndo sucdnico deshidrogenasa dependiente de CoA y de la 4-HB deshidrogenasa dependen de NADPH. En este caso, la produccion de 4-HB en el rendimiento maximo no dana como resultado una ganancia neta de ATP y no podna soportar las demandas de mantenimiento energetico necesarias para la supervivencia de E. coli. Por tanto, se podna originar algo de energfa de la fosforilacion oxidativa para permitir la produccion homofermentativa de 4-HB. La ruta del alfa-cetoglutarato hacia 4-HB es la menos favorable de las tres rutas potenciales con un rendimiento maximo conseguible de 1,0 mol de 4-HB por mol de glucosa. Ademas del rendimiento maximo menor, esta ruta requiere la utilizacion de 1,5 moles de oxfgeno por mol de glucosa convertido en 4-HB. El rendimiento energetico de esta ruta no se ve afectado por la supuesta especificidad del cofactor de la 4-HB deshidrogenasa.
Tabla 3. La estequiometna global de conversion del sustrato a 4-HB suponiendo que las rutas de produccion de A) succinato, B) succinil-CoA, o C) alfa-cetoglutarato son funcionales en E. coli. La glucosa y el oxfgeno se captan mientras que el resto de moleculas se producen.
A) Ruta del succinato

B) Ruta de la succinil-CoA

C) Ruta del alfa-cetoglutarato

A fin de corroborar las predicciones de calculo propuestas en este informe, las cepas que expresan una ruta completa hacia 4-HB se pueden construir y someter a ensayo. La corroboracion se lleva a cabo con E. coli (Ejemplo
II) y S. cerevisiae (Ejemplo III). En E. coli, los genes relevantes se expresan en un operon sintetico tras un promotor inducible en un plasmido con un numero de copias medio o alto; por ejemplo, el promotor PBAD que se induce por arabinosa, en un plasmido de la serie pBAD (Guzman et al., J. Bacteriol. 177:4121-4130 (1995)). En S. cerevisiae, los genes se integran en el cromosoma tras del promotor PDC1, sustituyendo el gen natural de la piruvato carboxilasa. Se ha notificado que esto da como resultado una mayor expresion de los genes extranos que desde un plasmido (Ishida et al., Appl. Environ. Microbiol. 71:1964-1970 (2005)), y garantizaran tambien la expresion durante las condiciones anaerobias.
Las celulas que contienen construcciones relevantes se hacen crecer en medios mmimos que contienen glucosa, con adicion de arabinosa en el caso de E. coli que contiene genes expresados con el promotor PBAD. Se toman muestras periodicas para evaluar la expresion genica y un analisis de actividad enzimatica. Se llevaron a cabo ensayos de actividad enzimatica en extractos de celulas brutas utilizando procedimientos bien conocidos en la materia. Como alternativa, se pueden utilizar ensayos basados en la oxidacion de NAD(P)H, que se producen en todas las etapas de reaccion de las deshidrogenasas y se pueden detectar mediante espectrofotometna. Ademas, se pueden usar anticuerpos para detectar el nivel de enzimas concretas. En lugar de, o ademas de, medidas de actividad enzimatica, se puede aislar el ARN de muestras paralelas y el transcripto del gen de interes se puede medir mediante PCR con transcriptasa inversa. Cualesquiera de las construcciones que caredan de expresion del transcripto detectable se volvieron a analizar para asegurar que los acidos nucleicos codificables se habfan incorporado en una forma expresable. Cuando se detectan los transcriptos, este resultado indica cualquiera de una carencia de traduccion o la produccion de un enzima inactiva. Se puede emplear adicionalmente una variedad de metodos bien conocidos en la tecnica, tales como la optimizacion de codones, la genomanipulacion de un sitio de union a ribosoma fuerte, el uso de un gen procedente de una especie diferente, y la prevencion de la N-glicosilacion (para la expresion de enzimas bacterianas en levadura) mediante conversion de restos Asn en Asp. Una vez que se detectaron todas las actividades enzimaticas requeridas, la siguiente etapa es medir la produccion de 4-HB in vivo. Se hacen crecer cultivos en matraces de cultivo por triplicado tanto anaerobia como microaerobicamente, dependiendo de las condiciones requeridas (vease anteriormente), y se toman muestras periodicas. Los acidos organicos presentes en los sobrenadantes del cultivo se analizaron mediante HPLC utilizando la columna Aminex AH-87X. El tiempo de elucion de 4-HB se determinara usando un patron adquirido de un proveedor de sustancias qmmicas.
Se puede implementar la ruta independiente de CoA y someterse a ensayo para corroboracion. En este caso, los genes expresados en exceso son los de la semialdetdo sucdnico deshidrogenasa natural de cada organismo, y de la 4-hidroxibutanoato deshidrogenasa de Ralstonia eutropha. Una vez detectadas las actividades enzimaticas como se ha descrito anteriormente, las cepas se someten a ensayo para determinar la produccion de 4-HB. Se puede obtener tambien la corroboracion implementando la ruta dependiente de CoA. La semialdetdo sucdnico deshidrogenasa dependiente de CoA y la 4-hidroxibutanoato deshidrogenasa procedente de Clostridium kluyveri se expresan como se ha descrito anteriormente. Ademas, se puede llevar tambien a cabo la sobreexpresion de succinil-CoA sintetasa natural, para canalizar mas succinato hacia la ruta heterologa. Finalmente, si la produccion de 4-HB es desfavorable, se pueden probar diferentes condiciones de cultivo, tales como un cambio en el estado de la oxigenacion que pueda manipular la proporcion NAD(P)H/NAD(P).
EJEMPLO III
Produccion de acido 4-hidroxibutanoico en levaduras
Este Ejemplo describe los rendimientos biosinteticos del acido 4-hidroxibutanoico resultantes de cada ruta bioqmmica en S. cerevisiae.
En esta seccion, se calcularon los rendimientos maximos teoricos de 4-HB a partir de glucosa suponiendo que cada una de estas tres rutas metabolicas representadas graficamente en la figura 2 son funcionales en S. cerevisiae. Se uso como base del analisis un modelo metabolico a escala genomica de S. cerevisiae, similar al descrito en Forster et al. (Genome Res. 13:244-253 (2003)). Se calculo la ganancia energetica de cada ruta de rendimiento maximo suponiendo condiciones anaerobias salvo que se indique lo contrario. Se supone que el 4-hidroxibutirato sale de S. cerevisiae mediante el mecanismo de cotransporte paralelo de protones, como es el caso de la mayona de los acidos organicos. Tambien se investigo el impacto de la especificidad del cofactor (es decir, la dependencia de NADH o NADPH) de los enzimas participates sobre el rendimiento maximo y el rendimiento energetico de cada ruta.
En las Tablas 4 A-C se muestran los resultados del analisis. Analogamente a E. coli, la ruta del succinato a 4-HB es la mas prometedora con la condicion que se puedan superar las preocupaciones termodinamicas planteadas en el Ejemplo I. Los calculos revelan que el rendimiento maximo teorico de 4-HB a partir de glucosa es de 1,33 mol/mol (0,77 g/g; 0,89 mol C/mol C) en S. cerevisiae. Ademas, la produccion anaerobia de 4-HB mediante succinato dana como resultado una produccion neta de 1,4, 1,1, o 0,5 mol de ATP por glucosa dependiendo de la especificidad del cofactor supuesto de los enzimas participates.
La ruta de la succinil-CoA a 4-HB es la segunda ruta mas favorable. Se puede conseguir un rendimiento maximo de
1,33 mol de 4-HB/mol de glucosa en S. cerevisiae sin tener en cuenta la especificidad del cofactor. Sin embargo, la generacion de energfa neta en el rendimiento maximo teorico es posible solo si se supone que las etapas de la semialdehndo succmico deshidrogenasa dependiente de CoA y la 4-HB deshidrogenasa son dependientes de NADPH. Si alguna etapa es dependiente de NADPH, no se obtendra ATP neto de la produccion de 4-HB anaerobia y se requerina una fuente de energfa alternativa (por ejemplo, la fosforilacion oxidativa) para soportar el crecimiento y el mantenimiento celular. La ruta del alfa-cetoglutarato hacia 4-HB es la menos favorable de las tres rutas potenciales en S. cerevisiae aunque el rendimiento maximo de 1,1-1,2 mol de 4-HB por mol de glucosa es ligeramente mayor que el que se encontro en E. coli. Sin embargo, esta ruta requiere una captacion de oxfgeno de 0,8-0,9 mol de oxfgeno por mol de glucosa para convertirse en energeticamente neutra.
Tabla 4. La estequiometna global de conversion del sustrato a 4-HB en S. cerevisiae, suponiendo las rutas de produccion de A) succinato, B) succinil-CoA, o C) alfa-cetoglutarato son funcionales en S. cerevisiae. La glucosa y el oxfgeno se captan mientras que el resto de moleculas se producen.
A) Ruta del succinato

B) Ruta de la succinil-CoA

C) Ruta del alfa-cetoglutarato

EJEMPLO IV
Biosintesis del acido 4-hidroxibutanoico. v-butirolactona y 1,4-butanodiol
Este Ejemplo describe la produccion biosintetica de acido 4-hidroxibutanoico, Y-butirolactona y 1,4-butanodiol usando fermentacion y otros bioprocesos.
Se describen a continuacion los metodos para integrar de la etapa de fermentacion de 4-HB en un proceso completo para la produccion de GBL, 1,4-butanodiol (BDO) y tetrahidrofurano (THF) purificados. Como 4-HB y GBL estan en equilibrio, el caldo de fermentacion contendra ambos compuestos. A un pH bajo, este equilibrio se desplaza para favorecer GBL. Por tanto, la fermentacion puede funcionar a pH 7,5 o menos. Tras la retirada de la biomasa, la corriente de producto entra en una etapa de separacion en la que se retira el GBL y la corriente restante enriquecida en 4-HB se recircula. Finalmente, GBL se destila para eliminar cualquier impureza. El proceso funciona en uno de tres modos: 1) fermentacion de alimentacion discontinua y separacion discontinua; 2) fermentacion de alimentacion
discontinua y separacion continua; 3) fermentacion continua y separacion continua. En la figura 4 se muestran esquematicamente los dos primeros de estos modos. Los procedimientos de fermentacion integrada descritos a continuacion se usan tambien para las celulas productoras de BDO de la invencion para la biosmtesis de BDO y los productos de la familia de BDO posteriores.
Protocolo de fermentacion para producir 4-HB/GBL (discontinua): El organismo de produccion se hace crecer en un biorreactor de 10 l en el que se burbujea una mezcla de N2/CO2, utilizando 5 l de caldo que contiene 5 g/l de fosfato de potasio, 2,5 g/l de cloruro de amonio, 0,5 g/l de sulfato de manganeso, y 30 g/l de solucion de mafz macerado, y una concentracion inicial de glucosa de 20 g/l. A medida que las celulas crecen y utilizan la glucosa, se alimenta mas glucosa al 70% al biorreactor a una tasa aproximadamente igual al consumo de glucosa en equilibrio. La temperatura del biorreactor se mantiene a 30 grados C. El crecimiento continua durante aproximadamente 24 horas, hasta que 4-HB alcanza una concentracion de entre 20-200 g/l, siendo la densidad celular entre 5 y 10 g/l. El pH no esta controlado, y disminuira normalmente a pH 3-6 al final del ciclo. Tras la finalizacion del periodo de cultivo, el contenido del fermentador se pasa a traves de una unidad de separacion celular (por ejemplo, una centnfuga) para eliminar las celulas y los desechos celulares, y el caldo de fermentacion se transfiere a una unidad de separacion del producto. El aislamiento de 4-HB y/o GBL tendna lugar mediante procedimientos de separacion convencionales empleados en la tecnica para separar los productos organicos de soluciones acuosas diluidas, tales como la extraccion lfquido-lfquido utilizando un disolvente organico inmiscible en agua (por ejemplo, tolueno) para proporcionar una solucion organica de 4-HB/GBL. La solucion resultante se somete a continuacion a metodos de destilacion convencionales para eliminar y recircular el disolvente organico y para proporcionar GBL (punto de ebullicion 204-205° C) que se afsla como un lfquido purificado.
Protocolo de fermentacion para producir 4-HB/GBL (completamente continuo): El organismo de produccion se hace crecer en primer lugar en modo discontinuo utilizando el equipo y la composicion del medio descrito anteriormente, excepto que la concentracion inicial de glucosa es de 30-50 g/l. Cuando la glucosa se agoto, se suministro de forma continua medio de alimentacion de la misma composicion a una velocidad de entre 0,5 l/h y 1 l/h, y el lfquido se retiro a la misma velocidad. La concentracion de 4-HB en el biorreactor sigue siendo constante a 30-40 g/l, y la densidad celular permanece constante entre 3-5 g/l. La temperatura se mantuvo a 30 grados C, y el pH se mantuvo a 4,5 utilizando NaOH y HCl concentrado, segun sea necesario. El biorreactor se hace funcionar de forma continua durante un mes, con muestras tomadas cada dfa para asegurar la consistencia de la concentracion de 4-HB. En modo continuo, el contenido del fermentador se retiro constantemente a medida que se suministraba medio de alimentacion nuevo. Se sometio a continuacion la corriente de salida, que contema celulas, medio, y productos de 4-HB y/o GBL, a procedimientos de separacion de productos en continuo, con o sin retirada de celulas y desechos celulares, y tendna lugar mediante metodos convencionales de separacion continua empleados en la tecnica para separar los productos organicos de soluciones acuosas diluidas, tales como la extraccion continua lfquido-lfquido utilizando un disolvente organico inmiscible en agua (por ejemplo, tolueno) para proporcionar una solucion organica de 4-HB/GBL. La solucion resultante se somete posteriormente a metodos de destilacion continua convencionales para eliminar y recircular el disolvente organico y para proporcionar GBL (punto de ebullicion 204 205° C) que se afsla como un lfquido purificado.
Protocolo de reduccion de g BL: Una vez que se aislo y purifico GBL tal como se ha descrito anteriormente, se sometio a continuacion a protocolos de reduccion tales como aquellos bien conocidos en la tecnica (referencias citadas) para producir 1,4-butanodiol o tetrahidrofurano (THF) o una mezcla de los mismos. Los catalizadores de hidrogenacion heterogeneos u homogeneos combinados con GBL bajo presion de hidrogeno son bien conocidos por proporcionar los productos 1,4-butanodiol o tetrahidrofurano (THF) o una mezcla de los mismos. Es importante senalar que la mezcla de producto de 4-HB/GBL que se separo del caldo de fermentacion, como se ha descrito anteriormente, puede someterse directamente, antes del aislamiento y la purificacion de GBL, a estos mismos protocolos de reduccion para proporcionar los productos 1,4-butanodiol o tetrahidrofurano o una mezcla de los mismos. Los productos resultantes, 1,4-butanodiol y THF, se aislaron y purificaron a continuacion mediante procedimientos bien conocidos en la materia.
Protocolo de fermentacion e hidrogenacion para producir BDO o THF directamente (discontinuo): Se hacen crecer las celulas en un biorreactor de 10 l en el que se burbujea una mezcla de N2/CO2, utilizando 5 l de caldo que contiene 5 g/l de fosfato de potasio, 2,5 g/l de cloruro de amonio, 0,5 g/l de sulfato de magnesio, y 30 g/l de solucion de mafz macerado, y una concentracion inicial de glucosa de 20 g/l. A medida que las celulas crecen y utilizan la glucosa, se alimenta mas glucosa al 70% al biorreactor a una tasa aproximadamente igual al consumo de glucosa en equilibrio. La temperatura del biorreactor se mantiene a 30 grados C. El crecimiento continua durante aproximadamente 24 horas, hasta que 4-HB alcanza una concentracion de entre 20-200 g/l, siendo la densidad celular entre 5 y 10 g/l. El pH no esta controlado, y disminuira normalmente a pH 3-6 al final del ciclo. Tras la finalizacion del periodo de cultivo, el contenido del fermentador se pasa a traves de una unidad de separacion celular (por ejemplo, una centnfuga) para eliminar las celulas y los desechos celulares, y el caldo de fermentacion se transfiere a una unidad de reduccion (por ejemplo, recipiente de hidrogenacion), donde la mezcla de 4-HB/GBL se redujo directamente a cualquiera de 1,4-butanodiol o THF o una mezcla de los mismos. Tras finalizar el procedimiento de reduccion, el contenido del reactor se transfiere a una unidad de separacion del producto. El aislamiento de 1,4-butanodiol y/o THF tendna lugar mediante procedimientos de separacion convencionales empleados en la tecnica para separar los productos organicos de soluciones acuosas diluidas, tales como la extraccion lfquido-lfquido utilizando un disolvente organico inmiscible en agua (por ejemplo, tolueno) para
proporcionar una solucion organica de 1,4-butanodiol y/o THF. La solucion resultante se sometio a continuacion a metodos de destilacion convencionales para eliminar y recircular el disolvente organico y para proporcionar 1,4-butanodiol y/o THF, que se aislaron como lfquidos purificados.
Protocolo de fermentacion e hidrogenacion para producir BDO o THF directamente (completamente continuo): Las celulas se hacen crecer en primer lugar en modo discontinuo utilizando el equipo y la composicion del medio descritos anteriormente, excepto que la concentracion inicial de glucosa es de 30-50 g/l. Cuando la glucosa se agoto, se suministro de forma continua medio de alimentacion de la misma composicion a una velocidad de entre 0,5 l/h y 1 l/h, y el lfquido se retiro a la misma velocidad. La concentracion de 4-HB en el biorreactor sigue siendo constante a 30-40 g/l, y la densidad celular permanece constante entre 3-5 g/l. La temperatura se mantuvo a 30 grados C, y el pH se mantuvo a 4,5 utilizando NaOH y HCl concentrado, segun sea necesario. El biorreactor se hace funcionar de forma continua durante un mes, con muestras tomadas cada dfa para asegurar la consistencia de la concentracion de 4-HB. En modo continuo, el contenido del fermentador se retiro constantemente a medida que se suministraba medio de alimentacion nuevo. La corriente de salida, que contema celulas, medio, y productos de 4-HB y/o GBL, se paso a continuacion a traves de una unidad de separacion celular (por ejemplo, una centnfuga) para eliminar las celulas y los desechos celulares, y el caldo se fermentacion se transfirio a una unidad de reduccion continua (por ejemplo, recipiente de hidrogenacion), donde la mezcla de 4-HB/GBL se redujo directamente a cualquiera de 1,4-butanodiol o THF o una mezcla de los mismos. Tras finalizar el procedimiento de reduccion, el contenido del reactor se transfiere a una unidad de separacion continua del producto. El aislamiento de 1,4-butanodiol y/o THF tendna lugar mediante procedimientos de separacion continua convencionales empleados en la tecnica para separar los productos organicos de soluciones acuosas diluidas, tales como la extraccion lfquido-lfquido utilizando un disolvente organico inmiscible en agua (por ejemplo, tolueno) para proporcionar una solucion organica de 1,4-butanodiol y/o THF. La solucion resultante se sometio a continuacion a metodos de destilacion continua convencionales para eliminar y recircular el disolvente organico y para proporcionar 1,4-butanodiol y/o THF, que se aislaron como lfquidos purificados.
Protocolo de fermentacion para producir BDO directamente (discontinuo): El organismo de produccion se hace crecer en un biorreactor de 10 l en el que se burbujea una mezcla de N2/CO2, utilizando 5 l de caldo que contiene 5 g/l de fosfato de potasio, 2,5 g/l de cloruro de amonio, 0,5 g/l de sulfato de magnesio, y 30 g/l de solucion de mafz macerado, y una concentracion inicial de glucosa de 20 g/l. A medida que las celulas crecen y utilizan la glucosa, se alimenta mas glucosa al 70% al biorreactor a una tasa aproximadamente igual al consumo de glucosa en equilibrio. La temperatura del biorreactor se mantiene a 30 grados C. El crecimiento continua durante aproximadamente 24 horas, hasta que BDO alcanza una concentracion de entre 20-200 g/l, siendo la densidad celular generalmente entre 5 y 10 g/l. Tras la finalizacion del periodo de cultivo, el contenido del fermentador se pasa a traves de una unidad de separacion celular (por ejemplo, una centnfuga) para eliminar las celulas y los desechos celulares, y el caldo de fermentacion se transfiere a una unidad de separacion del producto. El aislamiento de BDO tendna lugar mediante procedimientos de separacion convencionales empleados en la tecnica para separar los productos organicos de soluciones acuosas diluidas, tales como la extraccion lfquido-lfquido utilizando un disolvente organico inmiscible en agua (por ejemplo, tolueno) para proporcionar una solucion organica de BDO. La solucion resultante se somete a continuacion a metodos de destilacion convencionales para eliminar y recircular el disolvente organico y para proporcionar GBL (punto de ebullicion 228-229° C) que se afsla como un lfquido purificado.
Protocolo de fermentacion para producir BDO directamente (completamente continuo): El organismo de produccion se hace crecer en primer lugar en modo discontinuo utilizando el equipo y la composicion del medio descrito anteriormente, excepto que la concentracion inicial de glucosa es de 30-50 g/l. Cuando la glucosa se agoto, se suministro de forma continua medio de alimentacion de la misma composicion a una velocidad de entre 0,5 l/h y 1 l/h, y el lfquido se retiro a la misma velocidad. La concentracion de BDO en el biorreactor sigue siendo constante a 30-40 g/l, y la densidad celular permanece constante entre 3-5 g/l. La temperatura se mantuvo a 30 grados C, y el pH se mantuvo a 4,5 utilizando NaOH y HCl concentrado, segun sea necesario. El biorreactor se hace funcionar de forma continua durante un mes, con muestras tomadas cada dfa para asegurar la consistencia de la concentracion de BDO. En modo continuo, el contenido del fermentador se retiro constantemente a medida que se suministraba medio de alimentacion nuevo. Se sometio a continuacion la corriente de salida, que contema celulas, medio, y el producto de BDO, a procedimientos de separacion de productos en continuo, con o sin retirada de celulas y desechos celulares, y tendna lugar mediante metodos convencionales de separacion continua empleados en la tecnica para separar los productos organicos de soluciones acuosas diluidas, tales como la extraccion continua lfquido-lfquido utilizando un disolvente organico inmiscible en agua (por ejemplo, tolueno) para proporcionar una solucion organica de BDO. La solucion resultante se somete posteriormente a metodos de destilacion continua convencionales para eliminar y recircular el disolvente organico y para proporcionar BDO (punto de ebullicion 228 229° C) que se afsla como un lfquido purificado (pf 20° C).
EJEMPLOV
Estrategias de desactivacion procedentes de modelos in silico para la produccion de 1,4-butanodiol acoplada al crecimiento de Escherichia coli
Este ejemplo describe el diseno in silico de estrategias de desactivacion para generar la produccion de 1,4butanodiol acoplada al crecimiento de E. coli.
La estrategia para generar la produccion de BDO acoplada al crecimiento implica en primer lugar la demostracion de una ruta funcional que posteriormente se optimiza mediante la evolucion adaptiva e inserciones, deleciones, y sobreexpresiones dirigidas de genes. A continuacion se describen mas detalladamente estrategias de genomanipulacion de cepas identificadas mediante OptKnock para generar la produccion de BDO acoplada al crecimiento en cepas de Escherichia coli. Todas las implementaciones de OptKnock suponen que estan disponibles las actividades de todas los enzimas resenadas en la figura 6 para E. coli en todas las condiciones. Ademas, todas las etapas de reduccion de esta ruta se suponen dependientes de NADH, aunque se espera que algunos de los disenos sean de aplicacion independientemente de la especificidad del cofactor.
El rendimiento potencial de BDO de las rutas bioqmmicas para dar 1,4-butanodiol en E. coli se detalla a continuacion. Se utilizaron tres condiciones, anaerobias, anaerobias adicion de nitrato, y aerobias. Los rendimientos teoricos maximos de cada escenario, suponiendo que la produccion de BDO debe ser energeticamente neutra, se muestran en la Tabla 5. La Tabla 5 muestra los rendimientos teoricos maximos de 1,4-butanodiol (BDO) para cinco conjuntos de condiciones ambientales: 1) respiracion aerobia, 2) fermentacion anaerobia con produccion simultanea de acetato, 3) fermentacion anaerobia con produccion simultanea de etanol, 4) respiracion de nitrato que conduce a la formacion de nitrito, y 5) respiracion de nitrato que conduce a la formacion de amoniaco. Los valores negativos indican captacion del metabolito; los valores positivos indican secrecion del metabolito. Se suponen unidades molares, junto con la neutralidad energetica de la ruta. El rendimiento teorico maximo sin suponer neutralidad energetica es de 1,091 mol/mol (0,545 g/g) de glucosa para todos los casos. El mayor rendimiento se obtiene en condiciones aerobias, cuando se puede generar suficiente ATP para mantener la ruta energeticamente neutra con perdida minima de carbono mediante la respiracion. En el caso anaerobio, el rendimiento disminuye a medida que el ATP se fabrica mediante fosforilacion del sustrato, y se produce como subproducto acetato o etanol. La adicion controlada de nitrato puede proporcionar practicamente el mismo rendimiento que el oxfgeno, dependiendo la cantidad exacta de si se produce una reduccion adicional del nitrito a amoniaco.
Tabla 5. Rendimientos teoricos maximos de 1,4-butanodiol (BDO) para cinco conjuntos de condiciones ambientales.

Si se supone que fosfoenolpiruvato (PEP) carboxiquinasa funciona en la direccion desde el fosfoenolpiruvato hasta el oxaloacetato, la produccion de BDO se convierte en un generador de energfa, y el rendimiento teorico maximo para todos los escenarios pasa a ser de 0,545 g/g con el CO2 como unico subproducto. Se sabe que PEP carboxiquinasa produce oxaloacetato a partir de PEP en bacterias de rumiantes tales como Mannheimia succiniciproducens (Hong et al., Nat. Biotechnol. 22:1275-1281 (2004)). Sin embargo, se cree que el papel de la PEP carboxiquinasa en la produccion de oxaloacetato en E. coli es minoritario en comparacion con PEP carboxilasa, posiblemente debido a la mayor Km del bicarbonato de la PEP carboxiquinasa (Kim et al., Appl. Environ. Microbiol.
70:1238-1241 (2004)). Sin embargo, la actividad de la PEP carboxiquinasa natural de E. coli desde PEP en direccion al oxaloacetato se ha demostrado recientemente en mutantes ppc de K-12 (Kwon et al., J. Microbiol. Biotechnol.
16:1448-1452 (2006)).
En mas detalle, los disenos identificados para aumentar la produccion de BDO en E. coli se describen a continuacion. Se supone un requisito de mantenimiento energetico no asociado con el crecimiento de 7,6 mmol/gDW/h junto con una tasa de captacion de glucosa maxima espedfica de 20 mmol/gDW/h. Se supone que BDO se exporta mediante difusion. Se identificaron estrategias de desactivacion suponiendo que 1) PEP carboxiquinasa es irreversible y solamente funciona para convertir oxaloacetato en PEP y 2) PEP carboxiquinasa es reversible. Para ambos casos, el codigo de OptKnock descrito en Burgard et al. (Biotechnol. Bioeng. 84:647-657 (2003)) se modifico para permitir una respiracion limitada, no limitada, o ninguna, de nitrato. Anadir la posibilidad de respiracion de nitrato sirve para aumentar el rendimiento teorico maximo de BDO cuando se supone que la PEP carboxiquinasa es irreversible y tambien, en algunos casos, permite la seleccion de estrategias de desactivacion que
no se seleccionanan de otra manera en condiciones anaerobias debido al rendimiento energetico desfavorable. Espedficamente, se anadieron seis reacciones de captacion de nitrato a la red de E. coli con Ifmites inferiores de 0, -2, -5, -10, -20, o -1000 mmol/gDW/h (el valor negativo significa captacion del metabolito en el modelo estequiometrico). Las restricciones dobles de OptKnock se ajustaron de acuerdo con ello, y se anadio una restriccion adicional que permitia activar solamente una reaccion de captacion de nitrato cada vez. OptKnock selecciona la cantidad de respiracion de nitrato optima para cada estrategia de desactivacion identificada. Finalmente, todas las simulaciones suponen que la reaccion catalizada por adhE en E. coli (esto es, acetil-CoA+NADH^-etanol+NAD) se ha eliminado. Como el acetil-CoA es un compuesto intermedio necesario para la produccion de BDO, casi todos los disenos OptKnock incluinan esta delecion, de todos modos, para evitar que la produccion de etanol compita con la produccion de BDO. La inclusion de esta delecion se realizo a priori para disminuir el tiempo de CPU en el procedimiento informatico.
Las estrategias de desactivacion derivadas mediante OptKnock se proporcionan en las Tablas 6 y 7 suponiendo que PEP carboxiquinasa es irreversible y reversible, respectivamente. En la presente memoria descriptiva, las estrategias de desactivacion se relacionan mediante abreviaturas de reaccion en mayusculas por razones de simplicidad. Los correspondientes genes que se debenan desactivar para evitar que se produzca una reaccion concreta en E. coli se proporcionan en la Tabla 8 junto con la estequiometna de la reaccion. Los nombres de los metabolitos correspondientes a la Tabla 8 se relacionan en la Tabla 9.
Tabla 6. Estra teg ias de desactivac ion derivadas m ed ian te O ptK nock, supon iendo que PEP carboxiqu inasa es irreversib le .





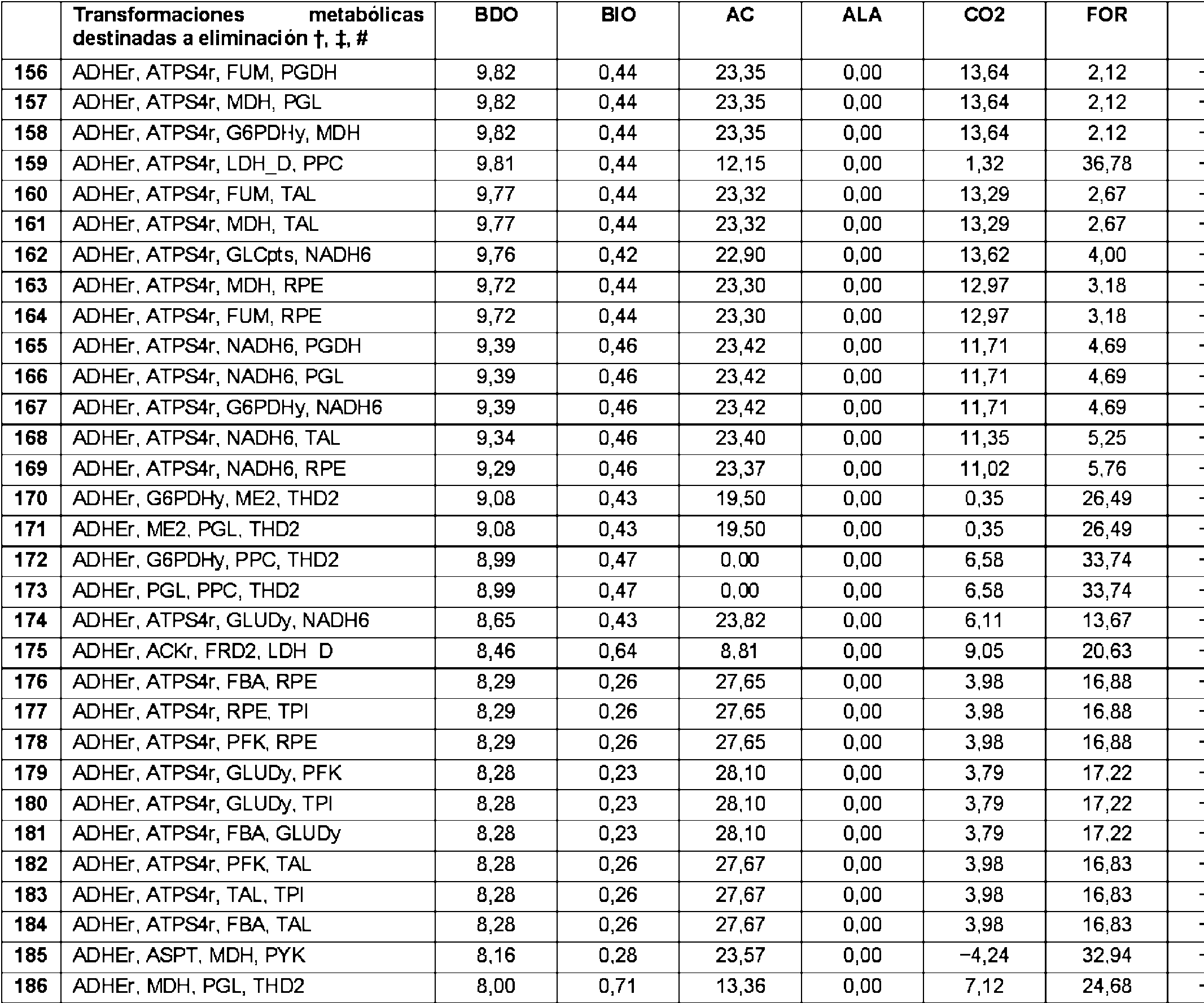





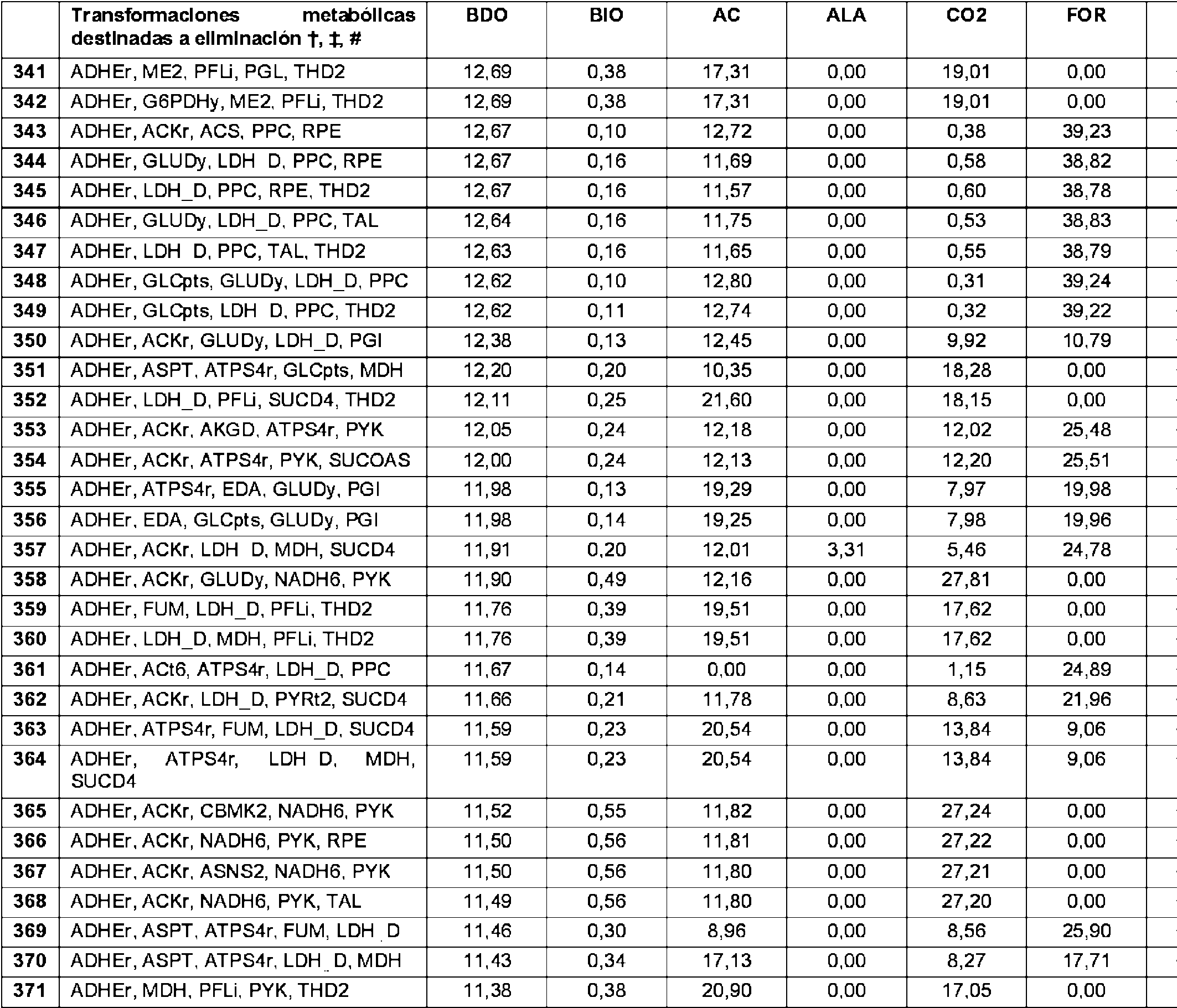


Tabla 6 (cont.)















G)
ro
Tabla 7. Estrategias de desactivacidn derivadas mediante OptKnock, suponiendo que PEP carboxiquinasa es reversible.
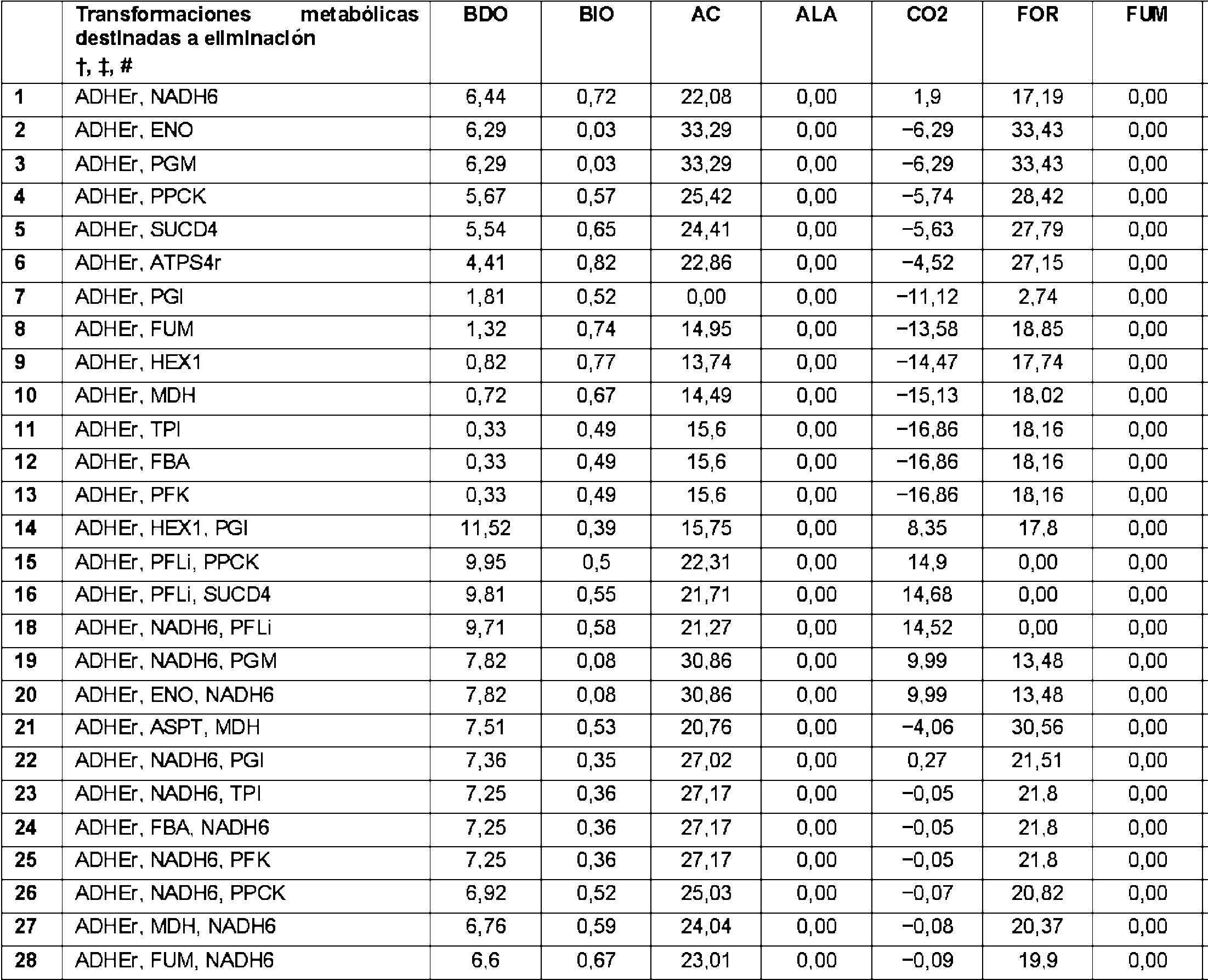

















Tabla 7 (Cont.)




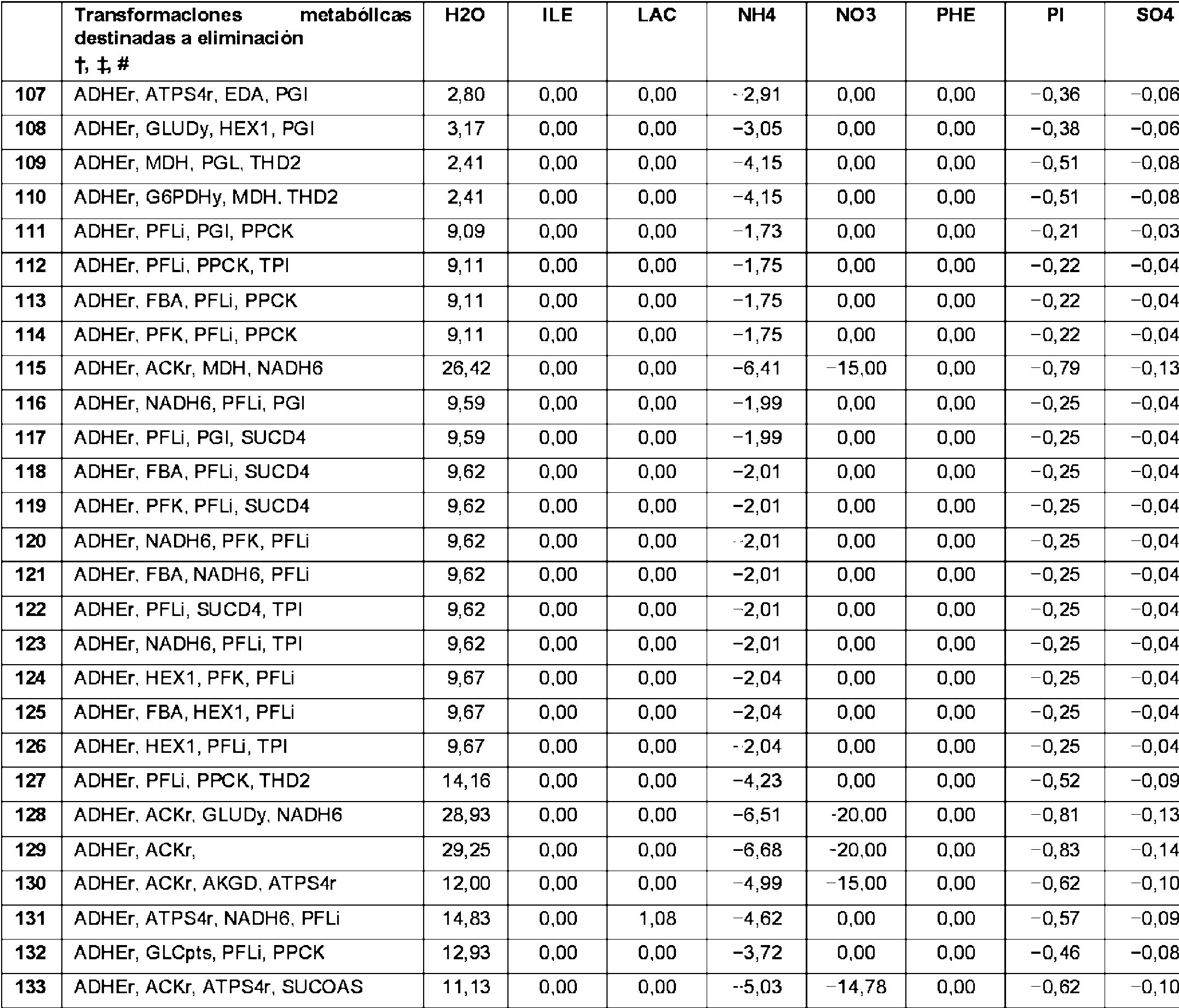














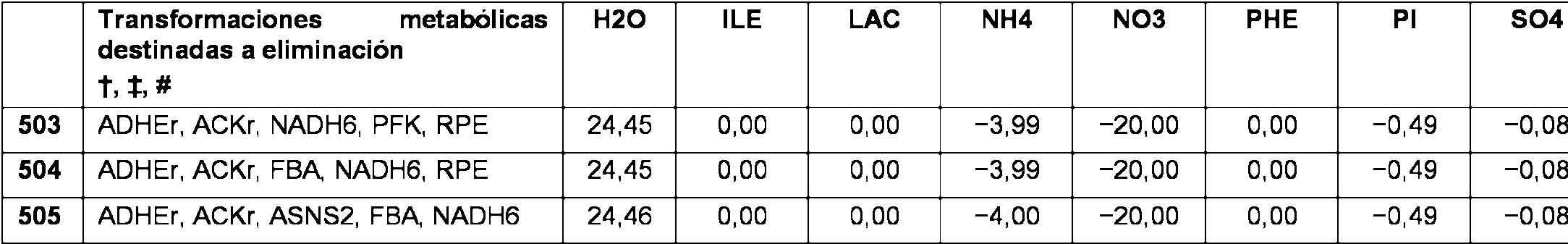
Tabla 8. Genes correspondientes a desactivar para evitar que se produzca una reaccion concreta en E. coli.
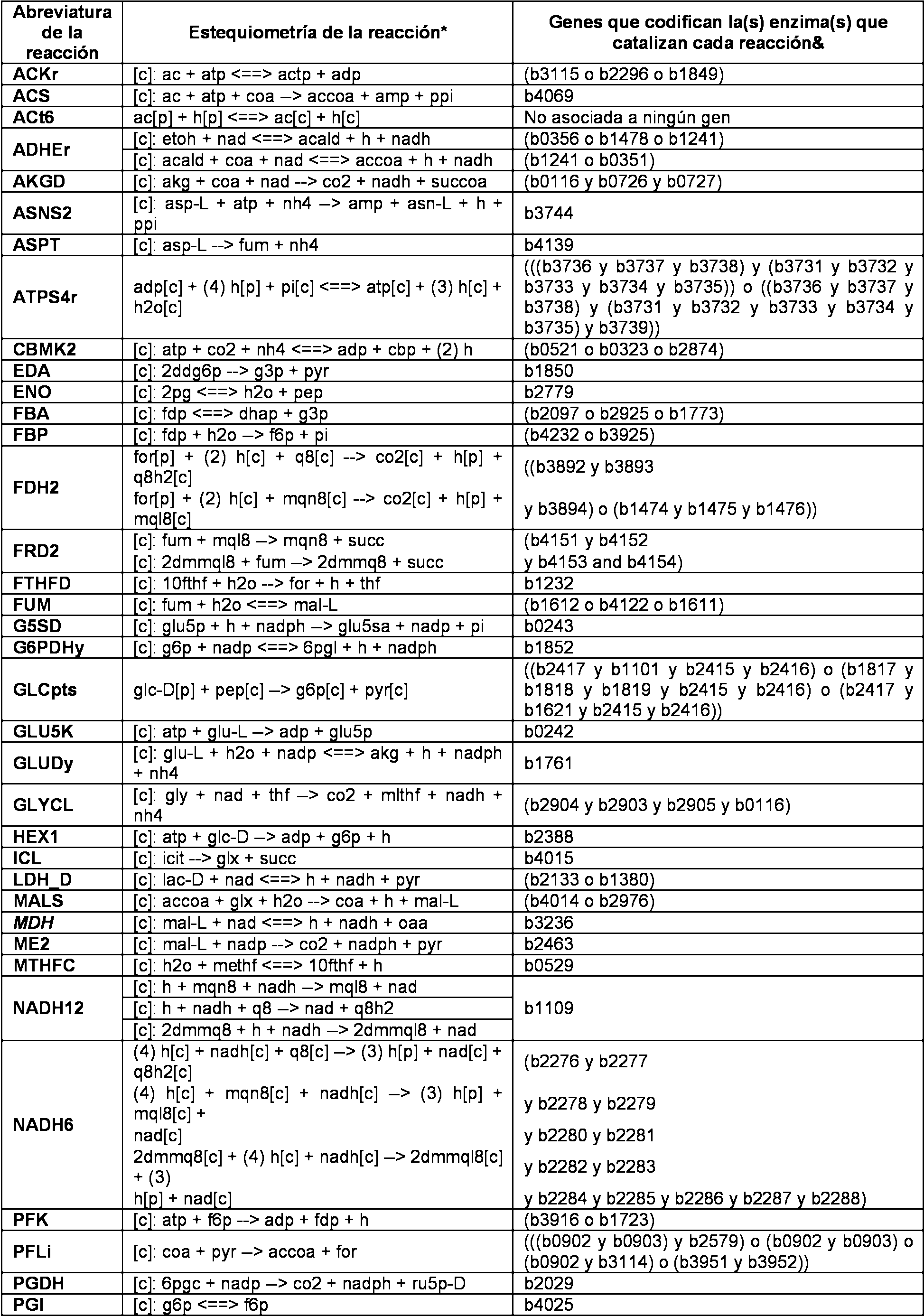

Tabla 9. Nombres de los metabolites correspondientes a las abreviaturas utilizadas en la Tabla 8.


Se aplicaron numerosos criterios para seleccionar los conjuntos de genes mas practicos como dianas de eliminacion. En primer lugar, los disenos se limitaron a incluir solamente desactivaciones que no reducen significativamente (esto es, >5%) el rendimiento teorico maximo de BDO en condiciones anaerobias, en presencia o ausencia de nitrato como aceptor de electrones. Dichas desactivaciones creanan un techo artificial para cualquier esfuerzo futuro de ingeniena metabolica, y esto, por tanto, es indeseable. Con este fin, se resolvieron una serie de problemas de programacion lineal (PL) que maximizaron el rendimiento en BDO para la red metabolica natural de E. coli suponiendo que cada reaccion se eliminara individualmente de la red. Tal como se usa en el presente documento, la referencia a una red natural de E. coli supone que la ruta de BDO esta disponible. El termino "natural” se utiliza por tanto como un nombre sustituto de red de E. coli. Las reacciones cuya delecion afectan negativamente el rendimiento maximo de BDO suponiendo que la PEP carboxiquinasa es reversible o irreversible se muestran en las Tablas 10 y 11, respectivamente. La Tabla 10 muestra reacciones que, cuando se suprimen, reducen el rendimiento teorico maximo de BDO en condiciones anaerobias en presencia o ausencia de nitrato, suponiendo que la PEP carboxiquinasa no puede utilizarse para producir oxaloacetato. La cepa AB3 contiene deleciones en ADHE, LDH_D, y PFLi. ‘ Inf indica que los requisitos de energfa no asociada con el crecimiento no se pueden satisfacer.
T ab la 10. Reacciones que, cuando se suprimen, reducen el rendimiento teorico maximo de BDO en condiciones anaerobias e suponiendo que la PEP carboxiquinasa no puede utilizarse para producir oxaloacetato.


Tab la 10 (cont.)


La Tabla 11 muestra reacciones que, cuando se suprimen, reducen el rendimiento teorico maximo de BDO en condiciones anaerobias en presencia o ausencia de nitrato, suponiendo que la PEP carboxiquinasa puede utilizarse para producir oxaloacetato. ‘ Inf indica que los requisitos de energfa no asociada con el crecimiento no se pueden satisfacer.
Tab la 11. Reacciones que, cuando se suprimen, reducen el rendimiento teorico maximo de BDO en condiciones anaerobias suponiendo que la PEP carboxiquinasa puede utilizarse para producir oxaloacetato.

Tab la 11 (cont.)

El analisis anteriormente descrito condujo a tres observaciones cnticas. Una observacion fundamental fue que la acetato quinasa y la fosfotransacetilasa son necesarias para conseguir el maximo rendimiento de BDO en todas las condiciones mediante la regeneracion de la acetil-CoA a partir del acetato producido por la 4-hidroxibutirato:acetil-CoA transferasa. Este hallazgo sugiere que la eliminacion de la formacion de acetato debido a la delecion de ackA-pta puede que no sea una opcion viable. As^ es probable que una cepa de exito haya tenido que proporcionar un entorno intracelular donde la conversion de acetato en acetil-CoA sea beneficiosa y termodinamicamente factible. Por otra parte, debena encontrarse un conjunto de enzimas capaces de realizar las reducciones de BDO necesarias sin pasar por un derivado de CoA o bien, debera aceptarse una produccion simultanea de 1 mol of acetato por mol de BDO.
Una segunda observacion fundamental fue que los enzimas del ciclo TCA citrato sintasa (CS), aconitasa (ACONT), e isocitrato deshidrogenasa (ICDHy) son necesarias para conseguir el maximo rendimiento de BDO en todas las condiciones. Esto indica que el flujo del ciclo TCA inverso desde oxaloacetato a succinato a succinil-CoA debe completarse en alguna medida mediante CS, ACONT, e ICDHy para una produccion maxima.
Una tercera observacion fundamental fue que sustituir la PEP carboxilasa por PEP carboxiquinasa en E. coli puede afectar positivamente el programa de BDO. El rendimiento maximo de BDO en condiciones anaerobias en presencia y ausencia de nitrato es del 3% y 12% inferior, respectivamente, si la PEP carboxilasa realiza la conversion de PEP en oxaloacetato en comparacion con la conversion realizada por PEP carboxiquinasa. Adicionalmente, en condiciones anaerobias sin adicion de nitrato, la PEP carboxiquinasa puede disminuir el requisito de actividad de la piruvato deshidrogenasa para una produccion de BDO maxima. Espedficamente, el rendimiento maximo de BDO disminuye un 11% si esta enzima tfpicamente aerobia no tiene actividad si se considera la PEP carboxiquinasa como irreversible en comparacion con una reduccion del 3% si la PEP carboxiquinasa puede catalizar la produccion de oxaloacetato. Una alternativa a la actividad piruvato deshidrogenasa se puede utilizar mediante el acoplamiento no natural de formiato deshidrogenasa, que puede catalizar la reduccion del formiato a dioxido de carbono, a la actividad de piruvato formiato liasa.
Los dos criterios siguientes aplicados para evaluar los disenos OptKnock fueron el numero de desactivaciones necesarias y el rendimiento de BDO previsto a crecimiento maximo. El analisis de las estrategias de seleccion uno, dos y tres de la Tabla 6 (esto es, PEP carboxiquinasa supuesta como irreversible) revelo que tan pocas desactivaciones eran insuficientes para evitar elevados rendimientos de acetato. Las proporciones de acetato/BDO previstas para los disenos de delecion uno, dos o tres, fue al menos 1,5. Incluso permitiendo cuatro deleciones, se consiguen solamente dos disenos, n° 99 y n° 100, con rendimientos de BDO previstos superiores a 0,35 g/g, y estos disenos sugieren la supresion del gen de la glicolisis, pgi, que codifica la fosfoglucoisomerasa. Dada la importancia prevista de la glicolisis en la fermentacion de E. coli, se decidio continuar con estos disenos de alto riesgo solamente como ultimo recurso. Adicionalmente, las deleciones sugeridas disminuyeron el rendimiento teorico maximo de los disenos n° 99 y n° 100 en un 8% y un 15%, respectivamente. La estrategia de cuatro deleciones de alto rendimiento que no afecto negativamente el rendimiento teorico maximo fue el diseno n° 129, que tema un rendimiento de BDO previsto a crecimiento maximo de solamente 0,26 g/g.
Solamente un diseno de cinco deleciones de la Tabla 6 satisface todos los criterios para un diseno de exito. Esta estrategia de desactivacion implica la eliminacion de ADHEr (alcohol deshidrogenasa), PFLi (piruvato formiato liasa), LDH_D (lactato deshidrogenasa), MDH (malato deshidrogenasa), y ASPT (aspartato transaminasa). Las desactivaciones sugeridas no reducen el rendimiento teorico maximo de BDO bajo ninguna de las condiciones examinadas. Se predice que una cepa disenada con estas desactivaciones conseguina un rendimiento de BDO para el crecimiento maximo de 0,37 g/g suponiendo condiciones anaerobias e irreversibilidad de PEP carboxiquinasa. Este diseno tiene varias propiedades deseables. De forma mas notable, evita que la red produzca altos rendimientos de los productos de fermentacion natural, etanol, formiato, lactato, y succinato. La prevencion de la produccion de homosuccinato mediante la delecion de MDH, en oposicion a la eliminacion de la PEP carboxilasa, fumarasa, o fumarato reductasa, es especialmente intrigante porque bloquea la ruta de fermentacion productora de energfa de oxaloacetato a succinato que podna aparecer si se supone que PEP carboxiquinasa es reversible sin afectar negativamente el rendimiento maximo de BDO. En este diseno, el succinato semialdeddo se puede fabricar mediante succinil-CoA o alfa-cetoglutarato. El succinil-CoA se forma a partir del succinato mediante la succinil-CoA sintetasa. El succinato se puede formar a partir tanto de las reacciones del ciclo TCA inverso (PEP carboxilasa, PEP carboxiquinasa, fumarasa, fumarato reductasa) y la derivacion del glioxilato (malato sintasa, isocitrato liasa).
Los lfmites de la solucion de BDO comparado con la biomasa para las estrategias de desactivacion de ADHEr, PFLi, LDH_D, MDH, ASPT se muestran en las Figs. 7A y 7B, suponiendo la irreversibilidad o reversibilidad de la PEP carboxiquinasa, respectivamente. Se debe tener en cuenta que los lfmites de la solucion se obtienen usando un modelo a escala genomica del metabolismo de E. coli en oposicion al modelo reducido, porque dichos calculos requieren muchos recursos de CPU. Los lfmites de la solucion revelan que el acoplamiento del BDO con el crecimiento es solido con respecto a la asuncion de la reversibilidad de la PEP carboxiquinasa. Sin embargo, la delecion de la bomba de protones transhidrogenasa (THD2) o glutamato deshidrogenasa (GLUDy) puede ser necesaria para conseguir un acoplamiento obligatorio del crecimiento celular con la produccion de BDO. El unico aspecto negativo del diseno es que las deleciones de MDH y ASPT disminuyen el rendimiento maximo de ATP en la produccion de BDO ligeramente (~20%) si se supone la reversibilidad de la PEP carboxiquinasa. Esto hace que la
solucion de crecimiento optimo de la estrategia de diseno este por debajo de la lmea negra de BDO vs. biomasa de la red natural de la figura 7B. Sin embargo, este hallazgo sugiere la posibilidad de genomanipular un equilibrio optimo de la actividad de MDH cuando hay suficiente cantidad presente para garantizar una produccion eficaz de BDO, pero al mismo tiempo lo suficientemente limitada para evitar que el succinato se convierta en el producto principal de la fermentacion. A destacar que se preve que el succinato sea el producto de fermentacion principal de la red natural y se supone la reversibilidad de la PEP carboxiquinasa.
Las Tablas 10 y 11 listan reacciones cuya delecion afecta negativamente el rendimiento maximo de BDO en una cepa intermedia, denominada como AB3, que carece de ADHEr, LDH_D, y PFLi, asf como una cepa que carece de ASPT y MDH ademas de las deleciones de AB3. A destacar que las deleciones sugeridas otorgan gran importancia a la obtencion de piruvato deshidrogenasa, citrato sintasa, y actividad de aconitasa en condiciones completamente anaerobias. Si se puede conseguir suficiente actividad piruvato deshidrogenasa, una alternativa es dejar PFLi intacto y complementar su actividad con una formiato deshidrogenasa no natural que captura un equivalente reductor mientras convierte el formiato en dioxido de carbono.
La Figura 8 y la Tabla 12 representan graficamente los intervalos de flujo que la red de E. coli puede conseguir cuando alcanza bien el rendimiento maximo de BDO (casos 1-4) o el rendimiento maximo de biomasa (caso 5) en condiciones anaerobias. Los casos 1 y 2 suponen que no se han producido deleciones de genes. El caso 2 muestra intervalos de flujo mas restringidos que el caso 1 porque se ha impuesto una restriccion adicional que refuerza el rendimiento maximo de BDO. Los casos 3 y 4 son analogos a los casos 1 y 2, salvo que los flujos codifican las reacciones de ADHEr, ASPT, LDH_D, MDH, y PFLi se han ajustado a cero. En el caso 5 se muestran los intervalos de flujo, suponiendo una maximizacion del rendimiento de biomasa en presencia de desactivaciones de ADHE, ASPT, LDH_D, MDH, y PFLi.
La Tabla 12 muestra los intervalos de flujos metabolicos centrales que se pueden conseguir en condiciones anaerobias suponiendo que la PEP carboxiquinasa es reversible. Los valores de flujo en negrita se configuran como restricciones del sistema. Se consideran cinco casos: Caso 1, rendimiento maximo de BDO de la red natural; Caso 2, rendimiento maximo de ATP suponiendo rendimiento maximo de BDO de la red natural; Caso 3, rendimiento maximo de BDO de la red con flujos a traves de ADHEr, ASPT, LDH_D, MDH, y PFLi ajustados a cero; Caso 4, rendimiento maximo de ATP suponiendo el rendimiento maximo de BDO de la red con flujos a traves de ADHEr, ASPT, LDH_D, MDH, y PFLi ajustados a cero; y Caso 5, rendimiento maximo de biomasa de la red con flujos a traves de ADHEr, ASpT, LDH_D, MDH, y PFLi ajustados a cero.
Tabla 12. Intervalos de flujos metabolicos centrales que se pueden conseguir en condiciones anaerobias suponiendo que la PEP carboxiquinasa es reversible. Las reacciones que se suponen inactivas en los casos 3, 4, y 5 se indican en negrita.

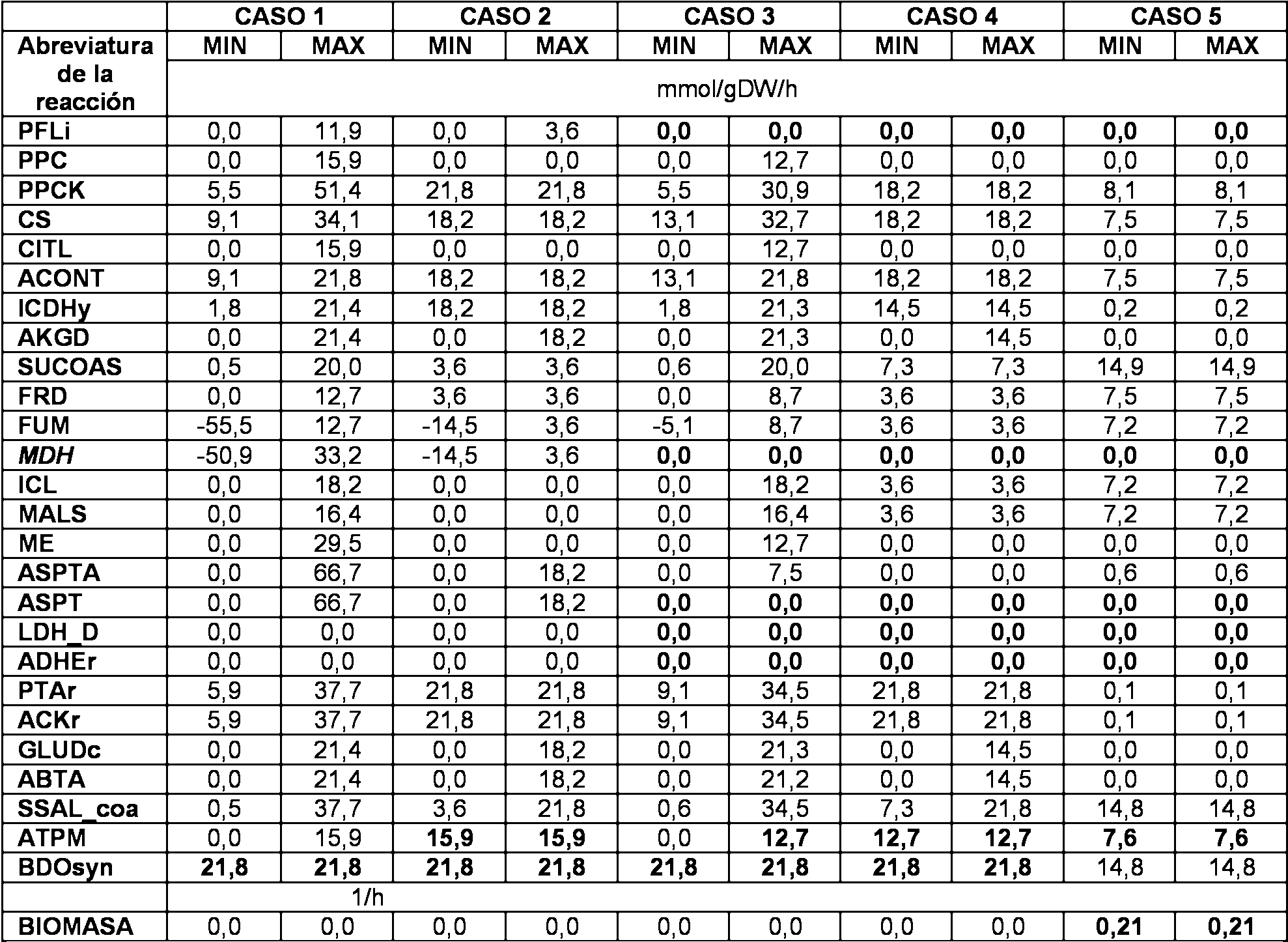
Los dos primeros disenos de cuatro deleciones (n° 92 y n° 93) relacionados en la Tabla 7 (esto es, PEP carboxiquinasa supuesta reversible) se consideran mas adelante debido a sus rendimientos de BDO previstos relativamente elevados. El diseno n° 92 (ADHEr, HEX1, PFLi, y PGI) necesitana desactivaciones adicionales que eliminaran la produccion de succinato y lactato y el rendimiento maximo de BDO del diseno n° 93 (ADHEr, e Da , NADH6, PGI) solamente fue el 90% del maximo teorico de la red natural. Ambos disenos requieren la eliminacion de PGI que, como se ha mencionado anteriormente, se descarto por indeseable debido a la importancia prevista de la glucolisis sobre la fermentacion. El diseno n° 98 (ADHEr, ATPS4r, FDH2, NADH6) fue el primer diseno de cuatro deleciones que requiere la eliminacion de la ATP sintasa sin requerir adicionalmente la desactivacion de PGI. Sin embargo, este diseno tambien requerina la delecion de genes para evitar la produccion de lactato y succinato para aumentar el rendimiento de BDO previsto a crecimiento maximo. Tras analisis posterior, se descubrio que casi todos los disenos prometedores de la Tabla 7 se podfan mejorar garantizando que las deleciones (es decir, ADHEr, LDH_D, MDH, ASPT, y PFLi) tambien se habfan implementado, si no se habfa especificado ya.
Los siguientes esfuerzos se centraron a continuacion en identificar las deleciones de reacciones que pudieran suministrar el conjunto fundamental (esto es, ADHEr, LDH_D, MDH, ASPT, PFLi). La Tabla 13 muestra genes de E. coli conocidos responsables de catalizar las reacciones diana para su eliminacion. La designacion "[c]” se refiere a citosolico. Los genes que codifican estas reacciones, junto las reacciones del conjunto fundamental, se proporcionan en la Tabla 13. El efecto de las deleciones adicionales sobre los lfmites de la solucion de BDO versus biomasa se muestra en la figura 9. De forma notable, el acoplamiento entre el BDO y la produccion de biomasa se vuelve cada vez mas pronunciado a medida que se acumulan deleciones adicionales. El rendimiento de BDO previsto a crecimiento maximo tras acumular todas las deleciones es de 0,46 g/g. Finalmente, ninguna de las deleciones afecto negativamente el rendimiento teorico maximo de BDO.
Tabla 13. Genes de E. coli conocidos responsables de catalizar las reacciones diana para su eliminacion.


En los resultados mostrados en la Tabla 13, OptKnock identifica reacciones a eliminar de un organismo para potenciar la produccion bioqmmica. Cualquier combinacion (esto es, al menos una y como maximo todas) de las deleciones de genes relacionadas podna tener, teoricamente, el efecto deseado de garantizar que la correspondiente reaccion no es funcional en E. coli. La estrategia experimental mas practica para eliminar las reacciones marcadas para su eliminacion se debe determinar caso por caso.
EJEMPLO VI
Generacion de cepas genomanipuladas
Para validar las predicciones informaticas del Ejemplo V, las cepas se construyeron, se hicieron evolucionar, y se sometieron a ensayo. Escherichia coli K-12 MG1655 sirve como la cepa natural en la que se introducen las deleciones. Las cepas se construyeron mediante la incorporacion de deleciones en marco usando recombinacion homologa usando el sistema X-Red recombinasa de (Datsenko y Wanner, Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)). El enfoque implica la sustitucion de una secuencia cromosomica (esto es, el gen marcado para eliminacion) por un gen de resistencia a antibioticos seleccionable, que posteriormente se elimina a su vez. Las desactivaciones se integran una por una en la cepa receptora. Despues de cada delecion no permanece ningun marcador de resistencia a farmacos ni cicatrices, lo que permite la acumulacion de multiples mutaciones en cada cepa diana. La tecnologfa de delecion elimina completamente el gen marcado para eliminacion de tal forma que se reduce sustancialmente la posibilidad de que los mutantes construidos reviertan al tipo natural. Durante las etapas iniciales del desarrollo de la cepa, los genes no nativos que permiten la produccion de BDO se expresan en un operon sintetico tras un promotor inducible en un plasmido con un numero de copias medio o alto; por ejemplo, el promotor PBAD que se induce por arabinosa, en un plasmido de la serie pBAD (Guzman et al., J. Bacteriol. 177:4121-4130 (1995)). Se sabe que este promotor es muy facil de titular, permitiendo afinar la expresion en un intervalo de 1000 veces el intervalo de concentraciones de arabinosa. Si la produccion de BDO tiene exito, estos genes posteriormente se integran en el cromosoma para promover la estabilidad.
Las cepas genomanipuladas se caracterizan por la medicion de la tasa de crecimiento, tasa de captacion de sustrato, y tasa de secrecion de productos/subproductos. Se anticipo inicialmente que estas cepas mostranan tasas de crecimiento suboptimas hasta que sus redes metabolicas se hubieran ajustado a sus funcionalidades ausentes. Para permitir este ajuste, las cepas evolucionan de forma adaptativa. Al someter las cepas a evolucion adaptativa, la tasa de crecimiento celular se convierte en la presion de seleccion principal y las celulas mutantes se ven impulsadas a recolocar sus flujos metabolicos para aumentar sus tasas de crecimiento. Se ha demostrado recientemente esta reprogramacion del metabolismo para varios mutantes de E. coli que habfan evolucionado de forma adaptativa sobre distintos sustratos para alcanzar tasas de crecimiento previstas a priori mediante un modelo in silico (Fong y Palsson, Nat. Genet. 36:1056-1058 (2004)). Si las predicciones de OptKnock tuvieran exito, las mejoras en el crecimiento conseguidas por la evolucion adaptiva van acompanadas de las tasas de produccion de BDO mejoradas. La evolucion adaptativa se realiza por triplicado (realizado en paralelo) debido a las diferencias en los patrones evolutivos usados como testigos previamente en E. coli (Fong y Palsson, mas arriba, 2004; Fong et al., J. Bacteriol. 185:6400-6408 (2003); Ibarra et al., Nature 420:186-189 (2002)) que potencialmente podnan dar como resultado una cepa con cualidades de produccion superiores a las demas. Las evoluciones iw actuan durante un periodo de 2-6 semanas, dependiendo de la mejora en la tasa de crecimiento obtenida. En general, las evoluciones q43 se detienen una vez que se obtiene un fenotipo de crecimiento estable.
Tras el proceso de evolucion adaptativa, las cepas nuevas se vuelven a caracterizar por la medicion de la tasa de crecimiento, tasa de captacion de sustrato, y tasa de secrecion de productos/subproductos. Estos resultados se comparan con las predicciones de OptKnock representando graficamente los rendimientos de crecimiento y
produccion reales a lo largo de la produccion anteriormente descrita. Las combinaciones de diseno OptKnock/evolucion de mas exito se seleccionan para investigacion adicional, y se caracterizan en fermentaciones de laboratorio tanto continuas como discontinuas. El concepto de produccion bioqmmica acoplada al crecimiento que esta detras del enfoque OptKnock tambien da como resultado la generacion de productores en exceso geneticamente estables. Por tanto, los cultivos se mantienen en modo continuo durante un mes para evaluar su estabilidad a largo plazo. Se toman muestras periodicamente para garantizar que el rendimiento y la productividad se mantienen durante la totalidad del proceso.
EJEMPLO VII
Cepas OptKnock para la produccion del BDO
Como se describe en los Ejemplos V y VI, la aplicacion de la metodologfa OptKnock se ha aplicado a la generacion de dianas de delecion prometedoras que generan cepas productoras de BDO. OptKnock identifica reacciones a eliminar de un organismo para acoplar los rendimientos de produccion bioqmmica y de biomasa. El diseno proporciona una lista de las reacciones metabolicas que se deben considerar para su eliminacion mediante OptKnock. Tambien se proporcionaron los genes de E. coli conocidos que codifican los enzimas que catalizan cada reaccion para describir que modificaciones geneticas se deben implementar para conseguir los fenotipos de produccion acoplada al crecimiento previstos. Evidentemente, si nuevos descubrimientos revelan que otros genes del genoma de E. coli pueden transmitir una o mas de las funcionalidades de reaccion consideradas para eliminacion en un diseno dado, entonces, estos genes debenan eliminarse junto con los descritos en el presente documento. A destacar que evitar la actividad de solamente un subconjunto (esto es, al menos una y como maximo todas) de las reacciones en cada uno de los disenos puede ser a veces suficiente para transmitir un fenotipo de produccion acoplada al crecimiento. Por ejemplo, si un diseno requiere la eliminacion de una reaccion particular cuya actividad in vivo no es suficiente para desacoplar el crecimiento de la produccion de BDO, entonces, los genes que codifican los enzimas que catalizan esta reaccion pueden dejarse intactos. Ademas, cualquier combinacion (esto es, al menos una y como maximo todas) de las deleciones de genes relacionadas para una reaccion dada podna tener, teoricamente, el efecto deseado de garantizar que la reaccion no es funcional en E. coli.
En la Tabla 6 y 7 se relacionan multiples estrategias de delecion para potenciar el acoplamiento entre la produccion de 1,4-butanodiol y el crecimiento de E. coli suponiendo que PEP carboxiquinasa es irreversible y reversible, respectivamente. Un diseno (esto es, ADHEr, ASPT, MDH, LDH_D, PFLi) surgio como el mas prometedor que satisfada multiples criterios. Las deleciones sugeridas 1) condujeron a un elevado rendimiento de BDO previsto a crecimiento maximo, 2) requirio un numero de desactivaciones razonable, 3) no tuvo un efecto perjudicial sobre el rendimiento teorico maximo de BDO, 4) consiguio un acoplamiento fuerte entre la produccion de BDO y el crecimiento celular, y 5) era solido con respecto a la reversibilidad o irreversibilidad de la PEP carboxiquinasa. La siguiente lista especifica el conjunto mmimo de deleciones genicas necesarias previstas para convertir BDO en el principal producto de fermentacion de E. coli:
adhE (b1421), ldhA (b1380).
pflAB (b0902, b0903) no esta incluido en el conjunto mmimo porque su delecion fuerza una dependencia de la piruvato deshidrogenasa para proporcionar cantidad suficiente de acetil-CoA para el crecimiento celular y un equivalente de reduccion para el piruvato. Como la actividad de la piruvato deshidrogenasa es baja en condiciones anaerobias y esta inhibida por concentraciones elevadas de NADH, una alternativa plausible a la delecion pflAB es anadir una formiato deshidrogenasa no natural a E. coli que recoja el pode reductor que se pierde por otra parte mediante la secrecion del formiato. Sin embargo, la adicion de pflAB al conjunto de delecion mmimo proporciona: adhE (b1421), ldhA (b1380), pflAB (b0902, b0903).
mdh (b3236) no esta incluido en el conjunto mmimo porque hay multiples deleciones que pueden evitar que el succinato se convierta en el producto de fermentacion principal de E. coli en oposicion a BDO. Los ejemplos incluyen los genes que codifican la fumarasa y/o fumarato reductasa. Sin embargo, la eliminacion de la malato deshidrogenasa parece ser la eleccion mas logica para atenuar la produccion de succinato ya que deja intacta una ruta para la conversion del producto derivado de glioxolato, malato, en BDO. La adicion de la malato deshidrogenasa al conjunto mmimo proporciona:
adhE (b1421), ldhA (b1380), pflAB (b0902, b0903), mdh (b3236).
El gen, mqo, que codifica una malato:quinona-oxidorreductasa, se cree que cataliza la oxidacion de malato a oxalactato (van der Rest et al., J. Bacteriol. 182:6892-6899 (2000)). Sin embargo, si se demuestra que tambien cataliza la formacion de malato a partir de oxaloacetato, su eliminacion sera necesaria para garantizar que no evita la delecion de mdh. Esto conduce al conjunto de delecion:
adhE (b1421), ldhA (b1380), pflAB (b0902, b0903), mdh (b3236), mqo (b2210).
aspA queda fuera del conjunto mmimo porque existe la duda de si la aspartato desaminasa puede transportar flujo suficiente para evitar la delecion de la malato deshidrogenasa. Sin embargo, aunque este escenario es ciertamente posible, entonces la lista minima de deleciones necesarias se convierte en:
adhE (b1421), IdhA (b1380), pflAB (b0902, b0903), mdh (b3236), aspA (b4139).
Para los calculos anteriores, se supone que los enzimas malicas dependientes de NADH y NADPH de E. coli operan de forma irreversible, catalizando solamente la conversion del malato en dioxido de carbono y piruvato. Si estas enzimas pueden catalizar tambien la formacion de malato a partir de piruvato y dioxido de carbono, los genes que codifican una o ambas enzimas malicas deberan eliminarse para evitar que el succinato se convierta en el producto de fermentacion principal. Esto conduce al siguiente conjunto de deleciones:
adhE (b1421), mdh (b3236), IdhA (b1380), pflAB (b0902, b0903), sfcA (b1479)
adhE (b1421), mdh (b3236), IdhA (b1380), pflAB (b0902, b0903), maeB (b2463)
adhE (b1421), mdh (b3236), IdhA (b1380), pflAB (b0902, b0903), sfcA (b1479), maeB (b2463)
El conjunto mmimo de deleciones se puede complementar con deleciones adicionales destinadas a refinar el acoplamiento de la produccion de BDO con el crecimiento celular. Estos conjuntos de deleciones se relacionan a continuacion.
adhE (b1421), ldhA (b1380), pflAB (b0902, b0903), mdh (b3236), pntAB (b1602, b1603)
adhE (b1421), ldhA (b1380), pflAB (b0902, b0903), mdh (b3236), gdhA (b1761)
adhE (b1421), ldhA (b1380), pflAB (b0902, b0903), mdh (b3236), pykA (b1854), pykF (b1676), dhaKLM (b1198, b1199, b1200), deoC (b4381), edd (b1851), yiaE (b3553), ycdW (b1033)
adhE (b1421), ldhA (b1380), pflAB (b0902, b0903), mdh (b3236), pykA (b1854), pykF (b1676), dhaKLM (b1198, b1199, b1200), deoC (b4381), edd (b1851), yiaE (b3553), ycdW (b1033), prpC (b0333), gsk (b0477)
Las cepas que tienen las deleciones indicadas en este Ejemplo se pueden complementar con deleciones adicionales si se descubre que las deleciones sugeridas no reducen la actividad de sus correspondientes reacciones en la medida que exige alcanzar la produccion de BDO acoplada al crecimiento o si los genes de E. coli naturales, mediante evolucion adaptativa o mutagenesis, consiguen mutaciones que transmitan actividades capaces de evitar las estrategias de diseno propuestas.
Puntos divulgados:
1. Un microorganismo no natural que comprende una o mas perturbaciones genicas, produciendose dicha una o mas perturbaciones genicas en genes que codifican para una enzima obligatoria para acoplar la produccion de 1,4-butanodiol al crecimiento de dicho microorganismo cuando dicha perturbacion genica reduce una actividad de dicha enzima, mediante lo cual dicha una o mas perturbaciones genicas confieren produccion de 1,4-butanodiol acoplada al crecimiento estable a dicho microorganismo no natural.
2. El microorganismo no natural del punto 1, en el que dicha una o mas perturbaciones genicas comprenden una modificacion metabolica enumerada en la tabla 6 o 7.
3. El microorganismo no natural del punto 1, en el que dicha una o mas perturbaciones genicas comprenden una delecion de dicho uno o mas genes.
4. El microorganismo no natural del punto 1, en el que dicho microorganismo no natural se selecciona del grupo de microorganismos que tienen una modificacion metabolica enumerada en la tabla 6 o 7.
5. El microorganismo no natural del punto 1, en el que dicho microorganismo comprende una bacteria, levadura u hongo.
6. El microorganismo no natural del punto 5, en el que dicha bacteria comprende una especie seleccionada de Escherichia coli, Klebsiella oxytoca, Anaerobiospirillum succiniciproducens, Actinobacillus succinogenes, Mannheimia succiniciproducens, Rhizobium etli, Bacillus subtilis, Corynebacterium glutamicum, Gluconobacter oxydans, Zymomonas mobilis, Lactococcus lactis, Lactobacillus plantarum, Streptomyces coelicolor, Clostridium acetobutylicum, Pseudomonas fluorescens y Pseudomonas putida.
7. El microorganismo no natural del punto 5, en el que dicha levadura comprende una especie seleccionada de Saccharomyces cerevisiae, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces marxianus,
Aspergillus terreus, Aspergillus niger y Pichia pastoris.
8. Un microorganismo no natural que comprende un conjunto de modificaciones metabolicas obligatorias para acoplar la produccion de 1,4-butanodiol al crecimiento de dicho microorganismo, comprendiendo dicho conjunto de modificaciones metabolicas perturbacion de uno o mas genes, o un ortologo de los mismos, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de adhE y ldhA, en el que dicho microorganismo presenta produccion de 1,4-butanodiol acoplada al crecimiento estable.
9. El microorganismo no natural del punto 8, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de mdh.
10. El microorganismo no natural del punto 9, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de uno o mas genes seleccionados del conjunto de genes que comprende mqo, aspA, sfcA, maeB, pntAB y gdhA.
11. El microorganismo no natural del punto 10, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de sfcA y maeB.
12. El microorganismo no natural del punto 9, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de uno o mas genes seleccionados del conjunto de genes que comprende pykA, pykF, dhaKLM, deoC, edd, yiaE, ycdW, prpC y gsk.
13. El microorganismo no natural del punto 12, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de pykA, pykF, dhaKLM, deoC, edd, yiaE y ycdW.
14. El microorganismo no natural del punto 13, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de prpC y gsk.
15. El microorganismo no natural de cualquiera de los puntos 9-14, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de pflAB.
16. El microorganismo no natural del punto 8, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de uno o mas genes seleccionados del conjunto de genes que comprende adhE, ldhA, pflAB, mdh y aspA.
17. El metodo del punto 8, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de los genes adhE, ldhA, pflAB, mdh y aspA.
18. El microorganismo no natural del punto 8, en el que dicho microorganismo comprende ademas una ruta biosintetica de 1,4-butanodiol (BDO), comprendiendo dicha ruta al menos un acido nucleico exogeno que codifica para 4-hidroxibutanoato deshidrogenasa, semialdehndo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdehndo succmico deshidrogenasa dependiente de CoA, 4-hidroxibutirato:CoA transferasa, glutamato:semialdehndo succmico transaminasa, glutamato descarboxilasa, aldehndo deshidrogenasa independiente de CoA, aldehndo deshidrogenasa dependiente de CoA o alcohol deshidrogenasa, en el que dicho acido nucleico exogeno se expresa en cantidades suficientes para producir 1,4-butanodiol (BDO).
19. El microorganismo no natural del punto 8, en el que dicha perturbacion de uno o mas genes comprende una delecion.
20. El microorganismo no natural del punto 8, en el que dicho microorganismo comprende una bacteria, levadura u hongo.
21. El microorganismo no natural del punto 20, en el que dicha bacteria comprende una especie seleccionada de E. coli, K. oxytoca, A. succiniciproducens, A. succinogenes, M. succiniciproducens, R. etli, B. subtilis, C. glutamicum, G. oxydans, Z. mobilis, L. lactis, L. plantarum, S. coelicolor, C. acetobutylicum, P. fluorescens y P. putida.
22. El microorganismo no natural del punto 20, en el que dicha levadura comprende una especie seleccionada de S. cerevisiae, S. pombe, K. lactis, K. marxianus, A. terreus, A. niger y P. pastoris.
23. Un metodo de produccion de un microorganismo no natural que tiene produccion de 1,4-butanodiol acoplada al crecimiento estable, que comprende:
(a) identificar in silico un conjunto de modificaciones metabolicas que requieren produccion de 1,4-butanodiol durante el crecimiento exponencial, y
(b) modificar por ingeniena genetica un microorganismo para que contenga dicho conjunto de modificaciones
metabolicas que requieren produccion de 1,4-butanodiol.
24. El metodo del punto 23, en el que dicho conjunto de modificaciones metabolicas se identifican mediante un metodo in silico seleccionado de OptKnock o SimPheny.
25. El metodo del punto 23, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion funcional de una o mas reacciones metabolicas.
26. El metodo del punto 25, en el que dichas modificaciones metabolicas se perturban mediante delecion genica. 27. El metodo del punto 25, en el que dicho microorganismo no natural comprende un microorganismo que tiene una modificacion metabolica seleccionada del conjunto de modificaciones metabolicas enumeradas en la tabla 6 o 7.
28. El metodo del punto 23, que comprende ademas cultivar dicho microorganismo modificado por ingeniena genetica.
29. El metodo del punto 28, que comprende ademas evolucionar de manera adaptativa dicho microorganismo modificado por ingeniena genetica o en condiciones que requieren produccion de 1,4-butanodiol.
30. El metodo del punto 23, en el que dicho microorganismo no natural comprende una bacteria, levadura u hongo.
31. El metodo del punto 30, en el que dicha bacteria comprende una especie seleccionada de E. coli, K. oxytoca, A. succiniciproducens, A. succinogenes, M. succiniciproducens, R. etli, B. subtilis, C. glutamicum, G. oxydans, Z. mobilis, L. lactis, L. plantarum, S. coelicolor, C. acetobutylicum, P. fluorescens y P. putida.
32. El microorganismo no natural del punto 30, en el que dicha levadura comprende una especie seleccionada de S. cerevisiae, S. pombe, K. lactis, K. marxianus, A. terreus, A. niger y P. pastoris.
33. Un microorganismo producido mediante el metodo del punto 23 o 29.
34. Un metodo de produccion de 1,4-butanodiol acoplado al crecimiento de un microorganismo, que comprende:
(a) cultivar en fase de crecimiento exponencial en una cantidad suficiente de nutrientes y medios un microorganismo no natural que comprende un conjunto de modificaciones metabolicas obligatorias para el acoplamiento de la produccion de 1,4-butanodiol al crecimiento de dicho microorganismo, comprendiendo dicho conjunto de modificaciones metabolicas perturbacion de uno o mas genes, o un ortologo de los mismos, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de adhE y ldhA, en el que dicho microorganismo presenta produccion de 1,4-butanodiol acoplada al crecimiento estable; y
(b) aislar 1,4-butanodiol producido a partir de dicho microorganismo no natural.
35. El metodo del punto 34, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de mdh.
36. El metodo del punto 35, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de uno o mas genes seleccionados del conjunto de genes que comprende mqo, aspA, sfcA, maeB, pntAB y gdhA.
37. El metodo del punto 36, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de sfcA y maeB.
38. El metodo del punto 35, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de uno o mas genes seleccionados del conjunto de genes que comprende pykA, pykF, dhaKLM, deoC, edd, yiaE, ycdW, prpC y gsk.
39. El metodo del punto 38, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de pykA, pykF, dhaKLM, deoC, edd, yiaE y ycdW.
40. El metodo del punto 39, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de prpC y gsk.
41. El metodo de cualquiera de los puntos 34-40, en el que dicho conjunto de modificaciones metabolicas comprende ademas perturbacion de pflAB.
42. El metodo del punto 34, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de uno o mas genes seleccionados del conjunto de genes que comprende adhE, ldhA, pflAB, mdh y aspA.
43. El metodo del punto 34, en el que dicho conjunto de modificaciones metabolicas comprende perturbacion de los genes adhE, IdhA, pflAB, mdh y aspA.
44. El metodo del punto 34, en el que dicho microorganismo no natural comprende ademas una ruta biosintetica de 1,4-butanodiol (BDO), comprendiendo dicha ruta al menos un acido nucleico exogeno que codifica para 4-hidroxibutanoato deshidrogenasa, semialdehndo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdehndo succmico deshidrogenasa dependiente de CoA, 4-hidroxibutirato:CoA transferasa, glutamatê semialdelmdo succmico transaminasa, glutamato descarboxilasa, aldehfdo deshidrogenasa independiente de CoA, aldehfdo deshidrogenasa dependiente de CoA o alcohol deshidrogenasa, en el que dicho acido nucleico exogeno se expresa en cantidades suficientes para producir 1,4-butanodiol (BDO).
Claims (10)
1. Metodo para fabricar un producto posterior de 4-hidroxibutirato o 1,4-butanodiol, que comprende:
(a) cultivar en fase de crecimiento exponencial en una cantidad suficiente de nutrientes y medios un microorganismo no natural que comprende perturbacion de los genes adhE, IdhA, pflAB y mdh, en el que dicho microorganismo expresa al menos un acido nucleico exogeno que codifica para una enzima en una ruta biosintetica de 4-hidroxibutirato o 1,4-butanodiol en cantidades suficientes para producir 4-hidroxibutirato o 1,4-butanodiol, codificando dicho al menos un acido nucleico exogeno para 4-hidroxibutanoato deshidrogenasa, semialdetudo succmico deshidrogenasa independiente de CoA, succinil-CoA sintetasa, semialdetudo succmico deshidrogenasa dependiente de CoA, 4-hidroxibutirato:CoA transferasa, glutamato:semialdetndo succmico transaminasa, glutamato descarboxilasa, aldetudo deshidrogenasa independiente de CoA, aldetudo deshidrogenasa dependiente de CoA o alcohol deshidrogenasa, y en el que dicho microorganismo produce 4-hidroxibutirato o 1,4-butanodiol;
(b) aislar opcionalmente 4-hidroxibutirato o 1,4-butanodiol; y
(c) convertir qmmicamente 4-hidroxibutirato o 1,4-butanodiol en el producto posterior.
2. Metodo segun la reivindicacion 1, en el que el microorganismo presenta produccion de 1,4-butanodiol acoplada al crecimiento estable.
3. Metodo segun la reivindicacion 1 o 2, en el que el producto posterior es gamma-butirolactona.
4. Metodo segun la reivindicacion 3, que comprende ademas la etapa de aislar gamma-butirolactona.
5. Metodo segun la reivindicacion 3 o 4, en el que la gamma-butirolactona se convierte qmmicamente a partir de 4-hidroxibutirato.
6. Metodo segun la reivindicacion 3 o 4, en el que la gamma-butirolactona se convierte qmmicamente a partir de 1,4-butanodiol.
7. Metodo segun la reivindicacion 1 o 2, en el que el producto posterior es tetrahidrofurano.
8. Metodo segun la reivindicacion 1 o 2, en el que el producto posterior es pirrolidona.
9. Metodo segun una cualquiera de las reivindicaciones 1-8, en el que el 1,4-butanodiol, 4-hidroxibutirato o gamma-butirolactona se afsla a partir de un cultivo de fermentacion.
10. Metodo segun una cualquiera de las reivindicaciones 1-9, en el que el 1,4-butanodiol o gammabutirolactona se purifica mediante destilacion.
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US11/891,602 US7947483B2 (en) | 2007-08-10 | 2007-08-10 | Methods and organisms for the growth-coupled production of 1,4-butanediol |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2695125T3 true ES2695125T3 (es) | 2019-01-02 |
Family
ID=39768484
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES08782639.2T Active ES2583179T3 (es) | 2007-08-10 | 2008-08-06 | Métodos y organismos para la producción de 1,4-butanodiol acoplada al crecimiento |
| ES13001197T Active ES2695125T3 (es) | 2007-08-10 | 2008-08-06 | Métodos y organismos para la producción de 1,4-butanodiol acoplada al crecimiento |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES08782639.2T Active ES2583179T3 (es) | 2007-08-10 | 2008-08-06 | Métodos y organismos para la producción de 1,4-butanodiol acoplada al crecimiento |
Country Status (13)
| Country | Link |
|---|---|
| US (5) | US7947483B2 (es) |
| EP (5) | EP3447133B1 (es) |
| AU (1) | AU2008287072B2 (es) |
| BR (1) | BRPI0815138A2 (es) |
| CA (2) | CA3175485A1 (es) |
| DK (1) | DK2679676T3 (es) |
| ES (2) | ES2583179T3 (es) |
| HU (1) | HUE040654T2 (es) |
| LT (1) | LT2679676T (es) |
| PL (1) | PL2679676T3 (es) |
| SI (1) | SI2679676T1 (es) |
| TR (1) | TR201814888T4 (es) |
| WO (1) | WO2009023493A1 (es) |
Families Citing this family (108)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US8673601B2 (en) * | 2007-01-22 | 2014-03-18 | Genomatica, Inc. | Methods and organisms for growth-coupled production of 3-hydroxypropionic acid |
| LT2821494T (lt) * | 2007-03-16 | 2017-04-25 | Genomatica, Inc. | 1,4-butandiolio ir jo pirmtakų biosintezės būdai ir kompozicijos |
| US7947483B2 (en) | 2007-08-10 | 2011-05-24 | Genomatica, Inc. | Methods and organisms for the growth-coupled production of 1,4-butanediol |
| WO2009045637A2 (en) | 2007-08-10 | 2009-04-09 | Genomatica, Inc. | Methods for the synthesis of acrylic acid and derivatives from fumaric acid |
| KR101042242B1 (ko) * | 2007-09-07 | 2011-06-17 | 한국과학기술원 | 1,4-부탄디올 생성능을 가지는 변이체 및 이를 이용한1,4-부탄디올의 제조방법 |
| CN101978042B (zh) * | 2008-01-22 | 2017-03-08 | 基因组股份公司 | 利用合成气或其它气态碳源和甲醇的方法和有机体 |
| EP2262901B1 (en) * | 2008-03-05 | 2018-11-21 | Genomatica, Inc. | Primary alcohol producing organisms |
| CN102066551B (zh) | 2008-03-27 | 2013-10-09 | 基因组股份公司 | 用于产生己二酸和其他化合物的微生物 |
| WO2009135074A2 (en) | 2008-05-01 | 2009-11-05 | Genomatica, Inc. | Microorganisms for the production of methacrylic acid |
| US8129154B2 (en) | 2008-06-17 | 2012-03-06 | Genomatica, Inc. | Microorganisms and methods for the biosynthesis of fumarate, malate, and acrylate |
| BRPI0915749A2 (pt) * | 2008-07-08 | 2018-07-10 | Opx Biotechnologies Inc | métodos, composições e sistemas para produção biossintética de 1,4-butanodiol |
| US20100021978A1 (en) * | 2008-07-23 | 2010-01-28 | Genomatica, Inc. | Methods and organisms for production of 3-hydroxypropionic acid |
| US7858350B2 (en) * | 2008-09-10 | 2010-12-28 | Genomatica, Inc. | Microorganisms for the production of 1,4-butanediol |
| US20100184173A1 (en) * | 2008-11-14 | 2010-07-22 | Genomatica, Inc. | Microorganisms for the production of methyl ethyl ketone and 2-butanol |
| EP2373781A4 (en) * | 2008-12-16 | 2012-10-10 | Genomatica Inc | MICRO-ORGANISMS AND METHODS OF CONVERTING SYNGAS AND OTHER CARBON RESOURCES IN USEFUL PRODUCTS |
| BRPI1013505A2 (pt) | 2009-04-30 | 2018-02-14 | Genomatica Inc | organismos para a produção de isopropanol, n-butanol, e isobutanol |
| EP4321615A3 (en) | 2009-04-30 | 2024-02-21 | Genomatica, Inc. | Organisms for the production of 1,3-butanediol |
| DK2427544T3 (da) | 2009-05-07 | 2019-10-07 | Genomatica Inc | Mikroorganismer og fremgangsmåder til biosyntese af adipat, hexamethylendiamin og 6-aminokapronsyre |
| BRPI1012877A2 (pt) * | 2009-05-15 | 2016-04-05 | Genomatica Inc | organismo para produção de ciclohexanona |
| SI3199511T1 (sl) * | 2009-06-04 | 2020-07-31 | Genomatica, Inc. | Proces separacije komponent fermentacijske brozge |
| WO2010141920A2 (en) | 2009-06-04 | 2010-12-09 | Genomatica, Inc. | Microorganisms for the production of 1,4-butanediol and related methods |
| EP2440669A4 (en) * | 2009-06-10 | 2013-08-28 | Genomatica Inc | MICROOGANISMS AND PROCEDURES FOR THE CARBON EFFECTIVE BIOSYNTHESIS MEK AND 2-BUTANOL |
| US8669397B2 (en) | 2009-06-13 | 2014-03-11 | Rennovia, Inc. | Production of adipic acid and derivatives from carbohydrate-containing materials |
| CA2763177C (en) | 2009-06-13 | 2017-08-01 | Thomas R. Boussie | Production of glutaric acid and derivatives from carbohydrate-containing materials |
| EP2440515B1 (en) | 2009-06-13 | 2018-08-15 | Archer-Daniels-Midland Company | Production of adipic acid and derivatives from carbohydrate-containing materials |
| WO2011017560A1 (en) | 2009-08-05 | 2011-02-10 | Genomatica, Inc. | Semi-synthetic terephthalic acid via microorganisms that produce muconic acid |
| EP2933338A3 (en) | 2009-09-09 | 2016-01-06 | Genomatica, Inc. | Microorganisms and methods for the co-production of isopropanol with primary alcohols, diols and acids |
| US8377666B2 (en) * | 2009-10-13 | 2013-02-19 | Genomatica, Inc. | Microorganisms for the production of 1,4-butanediol, 4-hydroxybutanal, 4-hydroxybutyryl-coa, putrescine and related compounds, and methods related thereto |
| KR20180014240A (ko) | 2009-10-23 | 2018-02-07 | 게노마티카 인코포레이티드 | 아닐린의 제조를 위한 미생물 |
| CN102933715B (zh) * | 2009-10-29 | 2015-04-29 | 韩国生命工学研究院 | 从甘油生产3-羟基丙酸的方法 |
| EP2501797A4 (en) | 2009-11-18 | 2013-04-17 | Myriant Corp | TECHNIQUES OF MICROBES FOR THE EFFECTIVE MANUFACTURE OF CHEMICALS |
| US8530210B2 (en) * | 2009-11-25 | 2013-09-10 | Genomatica, Inc. | Microorganisms and methods for the coproduction 1,4-butanediol and gamma-butyrolactone |
| US8268607B2 (en) | 2009-12-10 | 2012-09-18 | Genomatica, Inc. | Methods and organisms for converting synthesis gas or other gaseous carbon sources and methanol to 1,3-butanediol |
| TWI537386B (zh) * | 2009-12-22 | 2016-06-11 | Dsm智慧財產有限公司 | 由發酵法獲得的6-胺己酸製備己內醯胺 |
| EP2529011A4 (en) * | 2010-01-29 | 2015-07-15 | Genomatica Inc | MICROORGANISMS AND METHODS FOR BIOSYNTHESIS OF P-TOLUATE AND TEREPHTHALATE |
| US8637286B2 (en) * | 2010-02-23 | 2014-01-28 | Genomatica, Inc. | Methods for increasing product yields |
| US8048661B2 (en) * | 2010-02-23 | 2011-11-01 | Genomatica, Inc. | Microbial organisms comprising exogenous nucleic acids encoding reductive TCA pathway enzymes |
| US8669393B2 (en) | 2010-03-05 | 2014-03-11 | Rennovia, Inc. | Adipic acid compositions |
| CA2792546C (en) | 2010-03-09 | 2015-11-24 | Mitsui Chemicals, Inc. | Highly productive isopropyl alcohol-producing bacterium |
| US9200288B2 (en) | 2010-04-27 | 2015-12-01 | The Regents Of The University Of California | Production of 1,4-butanediol by recombinant microorganisms |
| US9023636B2 (en) | 2010-04-30 | 2015-05-05 | Genomatica, Inc. | Microorganisms and methods for the biosynthesis of propylene |
| KR101814648B1 (ko) | 2010-05-05 | 2018-01-04 | 게노마티카 인코포레이티드 | 부타디엔을 생합성하기 위한 미생물 및 방법 |
| US9770705B2 (en) | 2010-06-11 | 2017-09-26 | Rennovia Inc. | Oxidation catalysts |
| BR112013001635A2 (pt) | 2010-07-26 | 2016-05-24 | Genomatica Inc | micro-organismo e métodos para a biossíntese de aromáticos, 2, 4-pentadienoato e 1,3-butadieno |
| EP2652122B1 (en) | 2010-12-13 | 2018-08-22 | Myriant Corporation | Method of producing succinic acid and other chemicals using sucrose-containing feedstock |
| WO2012095244A2 (en) * | 2010-12-20 | 2012-07-19 | Lek Pharmaceuticals D.D. | Enzymatic synthesis of active pharmaceutical ingredient and intermediates thereof |
| WO2012106516A1 (en) | 2011-02-02 | 2012-08-09 | Genomatica, Inc. | Microorganisms and methods for the biosynthesis of butadiene |
| BR112013027555A2 (pt) * | 2011-04-29 | 2021-08-10 | Danisco Us Inc. | células recombinantes capazes de maior produção de isopreno, mevalonato e isoprenoides e método para produzir isopreno, mevalonato e isoprenoides |
| WO2012177943A1 (en) | 2011-06-22 | 2012-12-27 | Genomatica, Inc. | Microorganisms for producing 1,4-butanediol and methods related thereto |
| US8617862B2 (en) | 2011-06-22 | 2013-12-31 | Genomatica, Inc. | Microorganisms for producing propylene and methods related thereto |
| US9169486B2 (en) | 2011-06-22 | 2015-10-27 | Genomatica, Inc. | Microorganisms for producing butadiene and methods related thereto |
| EP2744906B1 (en) | 2011-08-19 | 2019-05-22 | Genomatica, Inc. | Microorganisms and methods for producing 2,4-pentadienoate, butadiene, propylene, 1,3-butanediol and related alcohols |
| KR20140070605A (ko) | 2011-09-16 | 2014-06-10 | 게노마티카 인코포레이티드 | 알켄을 생산하기 위한 유기체 및 방법 |
| BR112014010448A2 (pt) | 2011-11-02 | 2017-04-18 | Genomatica Inc | microorganismos e métodos para a produção de caprolactona |
| AU2013202630B2 (en) | 2012-01-20 | 2015-10-01 | Genomatica, Inc. | Microorganisms and processes for producing terephthalic acid and its salts |
| KR101351989B1 (ko) | 2012-02-16 | 2014-01-27 | 서강대학교산학협력단 | 2,3-부탄다이올 제조용 효모의 제조방법 |
| WO2013184602A2 (en) | 2012-06-04 | 2013-12-12 | Genomatica, Inc. | Microorganisms and methods for production of 4-hydroxybutyrate, 1,4-butanediol and related compounds |
| WO2013192237A1 (en) * | 2012-06-18 | 2013-12-27 | The Regents Of The University Of California | Escherichia coli engineered for isobutyraldehyde production |
| KR20140014648A (ko) * | 2012-07-25 | 2014-02-06 | 삼성전자주식회사 | 1,4-부탄디올의 고효율 생산을 위한 변형 미생물 |
| KR102023618B1 (ko) | 2012-07-27 | 2019-09-20 | 삼성전자주식회사 | 1,4-bdo 생성능이 개선된 변이 미생물 및 이를 이용한 1,4-bdo의 제조방법 |
| US9493746B2 (en) | 2012-07-30 | 2016-11-15 | Samsung Electronics Co., Ltd. | Enzyme used in biosynthesis of 1, 4-BDO and screening method of the same |
| KR20140016654A (ko) | 2012-07-30 | 2014-02-10 | 삼성전자주식회사 | 대장균 내에서 1,4-부탄디올의 생합성에 사용되는 효소의 개량과 개선된 유전자를 스크리닝 하는 방법 |
| US9657316B2 (en) | 2012-08-27 | 2017-05-23 | Genomatica, Inc. | Microorganisms and methods for enhancing the availability of reducing equivalents in the presence of methanol, and for producing 1,4-butanediol related thereto |
| US10501714B2 (en) | 2012-10-08 | 2019-12-10 | Calysta Energy, Inc. | Gas-fed fermentation systems |
| KR20210118216A (ko) | 2012-10-15 | 2021-09-29 | 게노마티카 인코포레이티드 | 특정 길이의 지방 알콜 및 관련 화합물의 생산을 위한 미생물 및 방법 |
| EP2909325A4 (en) | 2012-10-22 | 2016-05-25 | Genomatica Inc | MICROORGANISMS AND METHOD FOR IMPROVING THE AVAILABILITY OF REDUCING EQUIVALENTS IN THE PRESENCE OF METHANOL AND FOR PRODUCING SUCCINATE THEREOF |
| US9909150B2 (en) | 2012-11-05 | 2018-03-06 | Genomatica, Inc. | Microorganisms and methods for enhancing the availability of reducing equivalents in the presence of methanol, and for producing 1,2-propanediol, n-propanol, 1,3-propanediol, or glycerol related thereto |
| US9346902B2 (en) | 2012-11-05 | 2016-05-24 | Genomatica, Inc. | Microorganisms and methods for enhancing the availability of reducing equivalents in the presence of methanol, and for producing 3-hydroxyisobutyrate or methacrylic acid related thereto |
| TWI716752B (zh) | 2012-12-17 | 2021-01-21 | 美商奇諾麥提卡公司 | 用於增加甲醇存在下之還原當量可利用性及用於製造與其相關己二酸、6-胺基己酸、己二胺或己內醯胺之微生物及方法 |
| KR20150108367A (ko) * | 2013-01-21 | 2015-09-25 | 세키스이가가쿠 고교가부시키가이샤 | 재조합 세포, 및 1,4-부탄디올의 생산 방법 |
| BR112015020904A2 (pt) | 2013-03-15 | 2017-10-10 | Genomatica Inc | microrganismos e métodos para produção de butadieno e compostos relacionados por assimilação de formiato |
| WO2014152665A1 (en) | 2013-03-15 | 2014-09-25 | Genomatica, Inc. | Process and systems for obtaining 1,4-butanediol from fermentation broths |
| BR112015024633B1 (pt) | 2013-04-12 | 2022-03-15 | Toray Industries, Inc | Processos de produção de 1,4-butanodiol e de um poliéster |
| WO2014176514A2 (en) | 2013-04-26 | 2014-10-30 | Genomatica, Inc. | Microorganisms and methods for production of 4-hydroxybutyrate, 1,4-butanediol and related compounds |
| US11814664B2 (en) | 2013-05-24 | 2023-11-14 | Genomatica, Inc. | Microorganisms and methods for producing (3R)-hydroxybutyl (3R)-hydroxybutyrate |
| EP2818551B1 (en) * | 2013-06-28 | 2017-04-05 | Samsung Electronics Co., Ltd | Corynebacterium, in which a gene encoding CoA independent succinate semialdehyde dehydrogenase and a gene encoding L-lactate dehydrogenase are attenatued and method of producing C4 compounds, e.g. 4-hydroxybutyrate |
| EP3041942B1 (en) | 2013-09-03 | 2020-07-01 | PTT Global Chemical Public Company Limited | A process for manufacturing acrylic acid, acrylonitrile and 1,4-butanediol from 1,3-propanediol |
| KR20150035654A (ko) * | 2013-09-27 | 2015-04-07 | 삼성전자주식회사 | 1,4-bdo 생산능을 가진 미생물 및 그를 이용한 1,4-bdo를 생산하는 방법 |
| US20160376600A1 (en) | 2013-11-25 | 2016-12-29 | Genomatica, Inc. | Methods for enhancing microbial production of specific length fatty alcohols in the presence of methanol |
| EP4296364B1 (en) | 2013-12-03 | 2025-12-03 | Genomatica, Inc. | Microorganisms and methods for improving product yields on methanol using acetyl-coa synthesis |
| US10597684B2 (en) | 2013-12-27 | 2020-03-24 | Genomatica, Inc. | Methods and organisms with increased carbon flux efficiencies |
| WO2015158716A1 (en) | 2014-04-16 | 2015-10-22 | Novamont S.P.A. | Process for the production of 1,4-butanediol |
| WO2016004334A1 (en) | 2014-07-03 | 2016-01-07 | Genomatica, Inc. | Microorganisms for producing 4c-5c compounds with unsaturation and methods related thereto |
| US10487342B2 (en) | 2014-07-11 | 2019-11-26 | Genomatica, Inc. | Microorganisms and methods for the production of butadiene using acetyl-CoA |
| EP3741865B1 (en) | 2014-09-18 | 2024-03-13 | Genomatica, Inc. | Non-natural microbial organisms with improved energetic efficiency |
| WO2016092517A1 (en) | 2014-12-12 | 2016-06-16 | Versalis S.P.A. | Process for the production of 1, 3-butadiene from 1, 4 -butanediol via tetrahydrofuran |
| JP7127987B2 (ja) | 2014-12-23 | 2022-08-30 | ジェノマティカ, インコーポレイテッド | ジアミンの製造および処理方法 |
| CN108026214B (zh) | 2015-05-30 | 2021-03-02 | 基因组股份公司 | 乙烯基异构酶-脱水酶、烯醇脱水酶、芳樟醇脱水酶和/或巴豆醇脱水酶及其制备和使用 |
| US11441128B2 (en) | 2015-10-30 | 2022-09-13 | Genomatica, Inc. | Methanol dehydrogenase fusion proteins |
| WO2017165397A1 (en) | 2016-03-22 | 2017-09-28 | University Of Georgia Research Foundation, Inc. | Genetically engineered microbes and methods for producing citramalate |
| US10829789B2 (en) | 2016-12-21 | 2020-11-10 | Creatus Biosciences Inc. | Methods and organism with increased xylose uptake |
| CA3047841A1 (en) | 2016-12-21 | 2018-06-28 | Creatus Biosciences Inc. | Xylitol producing metschnikowia species |
| CN110691847A (zh) | 2017-03-31 | 2020-01-14 | 基因组股份公司 | 3-羟基丁酰基-CoA脱氢酶变体以及使用方法 |
| CA3058219A1 (en) | 2017-03-31 | 2018-10-04 | Genomatica, Inc. | Aldehyde dehydrogenase variants and methods of use |
| CN111601648B (zh) | 2017-11-27 | 2022-08-02 | 诺瓦蒙特股份公司 | 用于生产来自可再生资源的1,4-丁二醇的方法和由其获得的聚酯 |
| US20210079334A1 (en) | 2018-01-30 | 2021-03-18 | Genomatica, Inc. | Fermentation systems and methods with substantially uniform volumetric uptake rate of a reactive gaseous component |
| WO2020006058A2 (en) | 2018-06-26 | 2020-01-02 | Genomatica, Inc. | Engineered microorganisms with g3p---> 3pg enzyme and/or fructose-1,6-bisphosphatase including those having synthetic or enhanced methylotrophy |
| DE102018006128B8 (de) | 2018-08-03 | 2019-10-10 | Pepperl + Fuchs Gmbh | Landwirtschaftliche Erfassungsvorrichtung und Erfassungsverfahren zur Erfassung von landwirtschaftlichen Objekten |
| EP3856896A1 (en) | 2018-09-26 | 2021-08-04 | Genomatica, Inc. | Aldehyde dehydrogenase variants and methods of using same |
| US12540315B2 (en) | 2019-04-12 | 2026-02-03 | Renew Biopharma, Inc. | Compositions and methods for using genetically modified enzymes |
| KR20220023757A (ko) | 2019-04-24 | 2022-03-02 | 게노마티카 인코포레이티드 | 알데하이드 데하이드로게나아제 활성을 개선하기 위한 조작된 미생물 및 방법 |
| BR112022007541A2 (pt) | 2019-10-23 | 2022-07-12 | Genomatica Inc | Organismo microbiano de ocorrência não natural e métodos para aumentar a disponibilidade de nadph |
| US20230174454A1 (en) | 2020-05-12 | 2023-06-08 | Genomatica, Inc. | Process of synthesizing and purifying (3r)-hydroxybutyl (3r)-hydroxybutanoate |
| IT202000013243A1 (it) | 2020-06-04 | 2021-12-04 | Novamont Spa | Processo per la purificazione di una miscela di dioli |
| US11788053B2 (en) | 2020-06-15 | 2023-10-17 | Melio Peptide Systems Inc. | Microorganisms and methods for reducing bacterial contamination |
| JP2024503868A (ja) | 2021-01-17 | 2024-01-29 | ジェノマティカ, インコーポレイテッド | アミド化合物を生成する方法および組成物 |
| IT202100030572A1 (it) | 2021-12-02 | 2023-06-02 | Novamont Spa | 1,3-butandiolo purificato da una miscela di dioli |
| IT202300019440A1 (it) | 2023-09-21 | 2025-03-21 | Novamont Spa | Idrolizzato proteico di cellule di escherichia coli residue da un processo di fermentazione |
Family Cites Families (135)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB1230276A (es) | 1968-12-09 | 1971-04-28 | ||
| JPS4831084B1 (es) | 1970-09-04 | 1973-09-26 | ||
| GB1344557A (en) | 1972-06-23 | 1974-01-23 | Mitsubishi Petrochemical Co | Process for preparing 1,4-butanediol |
| DE2455617C3 (de) | 1974-11-23 | 1982-03-18 | Basf Ag, 6700 Ludwigshafen | Verfahren zur Herstellung von Butandiol und/oder Tetrahydrofuran über die Zwischenstufe des γ-Butyrolactons |
| DE2501499A1 (de) | 1975-01-16 | 1976-07-22 | Hoechst Ag | Verfahren zur herstellung von butandiol-(1.4) |
| GB1583517A (en) * | 1977-05-04 | 1981-01-28 | Jackson J F | Solid bowl decanter centrifuges of the scroll discharge type |
| DE2842575A1 (de) * | 1977-10-04 | 1979-04-12 | Broadbent & Sons Ltd Thomas | Vollmantel-abklaerzentrifuge |
| US4344002A (en) | 1978-02-22 | 1982-08-10 | Supertex, Inc. | Detection circuit and structure therefor |
| JPS575693A (en) | 1980-06-13 | 1982-01-12 | Ajinomoto Co Inc | Production of l-arginine through fermentation process |
| US4301077A (en) | 1980-12-22 | 1981-11-17 | Standard Oil Company | Process for the manufacture of 1-4-butanediol and tetrahydrofuran |
| IT1190783B (it) | 1981-04-29 | 1988-02-24 | Davy Mckee Oil & Chem | Processo per l'idrogenolisi di esteri di acidi carbossilici |
| US4652685A (en) | 1985-11-15 | 1987-03-24 | General Electric Company | Hydrogenation of lactones to glycols |
| JPS62285779A (ja) * | 1986-06-04 | 1987-12-11 | Chiyoda Chem Eng & Constr Co Ltd | 1,4−ブタンジオ−ル産生バチルス属細菌及びそれを用いる1,4−ブタンジオ−ルの製造方法 |
| US5250430A (en) | 1987-06-29 | 1993-10-05 | Massachusetts Institute Of Technology | Polyhydroxyalkanoate polymerase |
| US5245023A (en) | 1987-06-29 | 1993-09-14 | Massachusetts Institute Of Technology | Method for producing novel polyester biopolymers |
| US4876331A (en) | 1987-08-18 | 1989-10-24 | Mitsubishi Kasei Corporation | Copolyester and process for producing the same |
| EP0556387B1 (en) * | 1988-04-27 | 1995-08-09 | Daicel Chemical Industries, Ltd. | Process for preparing optically active 1,3-butanediol |
| EP0373230B1 (en) | 1988-12-12 | 1993-02-17 | Unilever N.V. | Process for the microbiological preparation of 1,3-propane-diol from glycerol |
| IE893328A1 (en) | 1989-10-16 | 1991-04-24 | Mcdaid Denis | Telephone lock |
| GB9011777D0 (en) | 1990-05-25 | 1990-07-18 | Ici Plc | Hv/hb copolymer production |
| US5998366A (en) | 1990-09-21 | 1999-12-07 | The Regents Of The University Of California | Method for ameliorating glutamic acid decarboxylase associated autoimmune disorders |
| US5475086A (en) | 1990-09-21 | 1995-12-12 | The Regents Of The University Of California | Cloned glutamic acid decarboxylase peptides |
| US5674978A (en) | 1990-09-21 | 1997-10-07 | The Regents Of The University Of California | Peptides derived from glutamic acid decarboxylase |
| US6682906B1 (en) | 1990-09-21 | 2004-01-27 | The Regents Of The University Of California | Cloned glutamic acid decarboxylase |
| GB9108756D0 (en) | 1991-04-24 | 1991-06-12 | Ici Plc | Production of polyalkanoate in plants |
| US5286842A (en) | 1991-07-01 | 1994-02-15 | Mitsubishi Kasei Corporation | Process for producing a biodegradable polymer |
| GB9115245D0 (en) | 1991-07-16 | 1991-08-28 | Ici Plc | Production of polyalkanoate |
| DE69233512T2 (de) | 1991-07-19 | 2005-10-13 | Michigan State University, East Lansing | Transgene pflanzen die polyhydroxyalkanoate produzieren |
| US5610041A (en) | 1991-07-19 | 1997-03-11 | Board Of Trustees Operating Michigan State University | Processes for producing polyhydroxybutyrate and related polyhydroxyalkanoates in the plastids of higher plants |
| BR9203053A (pt) | 1991-08-07 | 1993-03-30 | Ajinomoto Kk | Processo para produzir acido l-glutamico pro fermentacao |
| JP2777757B2 (ja) | 1991-09-17 | 1998-07-23 | 鐘淵化学工業株式会社 | 共重合体およびその製造方法 |
| DE4220759A1 (de) * | 1992-06-24 | 1994-01-05 | Inst Genbiologische Forschung | DNA-Sequenzen für Oligosaccharid-Transporter, Plasmide, Bakterien und Pflanzen enthaltend einen Transporter sowie Verfahren zur Herstellung und Transformation von Hefestämmen zur Identifikation des Transporteers |
| EP0625006A4 (en) | 1992-11-20 | 1996-04-24 | Agracetus | Transgenic cotton plants producing heterologous bioplastic. |
| JP3263710B2 (ja) | 1992-12-11 | 2002-03-11 | 高砂香料工業株式会社 | 生分解性光学活性ポリマー及びその製造方法 |
| JP3241505B2 (ja) | 1993-08-11 | 2001-12-25 | 高砂香料工業株式会社 | 生分解性光学活性コポリマー及びその製造方法 |
| SK283058B6 (sk) | 1993-10-28 | 2003-02-04 | Ajinomoto Co., Inc. | Spôsob produkcie cieľovej látky použitím mikroorganizmov |
| GB9414506D0 (en) | 1994-07-18 | 1994-09-07 | Zeneca Ltd | Improved production of polyhydroxyalkanoate |
| DE4433134A1 (de) | 1994-09-16 | 1996-03-21 | Buck Chem Tech Werke | Verfahren zur Herstellung von Polyhydroxyfettsäuren sowie rekombinanter Bakterienstämme zur Durchführung des Verfahrens |
| US5563239A (en) | 1994-11-09 | 1996-10-08 | Eastman Chemical Company | Composition and process for the production of poly(3-hydroxyalkanoates) |
| US5478952A (en) | 1995-03-03 | 1995-12-26 | E. I. Du Pont De Nemours And Company | Ru,Re/carbon catalyst for hydrogenation in aqueous solution |
| US5747311A (en) | 1995-08-22 | 1998-05-05 | Microgen Corporation | Process for chemical modification of reactants by microbes |
| US5869301A (en) | 1995-11-02 | 1999-02-09 | Lockhead Martin Energy Research Corporation | Method for the production of dicarboxylic acids |
| US5770435A (en) | 1995-11-02 | 1998-06-23 | University Of Chicago | Mutant E. coli strain with increased succinic acid production |
| DE19543635A1 (de) | 1995-11-23 | 1997-05-28 | Hp Chemie Pelzer Res & Dev | Verbundwerkstoffe aus Polyhydroxyfettsäuren und Fasermaterialien |
| US5849894A (en) | 1995-11-29 | 1998-12-15 | Monsanto Company | Rhodospirillum rubrum poly-β-hydroxyalkanoate synthase |
| US5908924A (en) * | 1996-02-27 | 1999-06-01 | Michigan State University | Cloning and expression of the gene encoding thermoanaerobacter ethanolicus 39E secondary-alcohol dehydrogenase and enzyme biochemical characterization |
| US6091002A (en) | 1996-03-13 | 2000-07-18 | Monsanto Company | Polyhydroxyalkanoates of narrow molecular weight distribution prepared in transgenic plants |
| US5959179A (en) | 1996-03-13 | 1999-09-28 | Monsanto Company | Method for transforming soybeans |
| US5958745A (en) | 1996-03-13 | 1999-09-28 | Monsanto Company | Methods of optimizing substrate pools and biosynthesis of poly-β-hydroxybutyrate-co-poly-β-hydroxyvalerate in bacteria and plants |
| US5750848A (en) | 1996-08-13 | 1998-05-12 | Monsanto Company | DNA sequence useful for the production of polyhydroxyalkanoates |
| US6730503B1 (en) | 1996-09-19 | 2004-05-04 | Roche Vitamins Inc. | Alcohol/aldehyde dehydrogenase |
| JPH10166664A (ja) * | 1996-12-12 | 1998-06-23 | Casio Electron Mfg Co Ltd | カラー印刷装置 |
| EP0977865A1 (en) | 1997-02-13 | 2000-02-09 | James Madison University | Methods of making polyhydroxyalkanoates comprising 4-hydroxybutyrate monomer units |
| US6759219B2 (en) | 1997-03-03 | 2004-07-06 | Metabolix, Inc. | Methods for the biosynthesis of polyesters |
| US6156852A (en) | 1997-04-21 | 2000-12-05 | Monsanto Company | Hydroxy-terminated polyhydroxyalkanoates |
| US5994478A (en) | 1997-04-21 | 1999-11-30 | Monsanto Company | Hydroxy-terminated polyhydroxyalkanoates |
| KR19990013007A (ko) | 1997-07-31 | 1999-02-25 | 박원훈 | 형질전환된 대장균 ss373(kctc 8818p)과 이를 이용한숙신산의 생산방법 |
| EP1015565B1 (en) | 1997-09-19 | 2006-04-12 | Metabolix, Inc. | Biological systems for manufacture of polyhydroxyalkanoate polymers containing 4-hydroxyacids |
| US6228579B1 (en) * | 1997-11-14 | 2001-05-08 | San Diego State University Foundation | Method for identifying microbial proliferation genes |
| US6280986B1 (en) | 1997-12-01 | 2001-08-28 | The United States Of America As Represented By The Secretary Of Agriculture | Stabilization of pet operon plasmids and ethanol production in bacterial strains lacking lactate dehydrogenase and pyruvate formate lyase activities |
| US6159738A (en) | 1998-04-28 | 2000-12-12 | University Of Chicago | Method for construction of bacterial strains with increased succinic acid production |
| EP1975236B1 (en) | 1998-08-04 | 2015-05-06 | Metabolix, Inc. | Polyhydroxyalkanoate production from polyols |
| DE69930276T2 (de) | 1998-08-18 | 2006-10-05 | Metabolix, Inc., Cambridge | Transgene polyhydroxyalkanoat produzierende mikroben |
| US6835820B2 (en) | 1999-01-07 | 2004-12-28 | The University Of Massachusetts | Polyhydroxyalkanoate biosynthesis associated proteins and coding region in bacillus megaterium |
| WO2000046405A2 (en) | 1999-02-02 | 2000-08-10 | Bernhard Palsson | Methods for identifying drug targets based on genomic sequence data |
| US6686310B1 (en) | 1999-02-09 | 2004-02-03 | E. I. Du Pont De Nemours And Company | High surface area sol-gel route prepared hydrogenation catalysts |
| US6448473B1 (en) | 1999-03-05 | 2002-09-10 | Monsanto Technology Llc | Multigene expression vectors for the biosynthesis of products via multienzyme biological pathways |
| KR100329019B1 (ko) * | 1999-04-13 | 2002-03-18 | 윤덕용 | 유기산의 고효율 생산방법 |
| WO2001012833A2 (en) | 1999-08-18 | 2001-02-22 | E.I. Du Pont De Nemours And Company | Process for the biological production of 1,3-propanediol |
| US6361983B1 (en) | 1999-09-30 | 2002-03-26 | E. I. Du Pont De Nemours And Company | Process for the isolation of 1,3-propanediol from fermentation broth |
| DE19956686A1 (de) | 1999-11-25 | 2001-05-31 | Degussa | Neue für die Gene sucC und sucD codierende Nukleotidsequenzen |
| US6897055B2 (en) | 1999-11-25 | 2005-05-24 | Degussa Ag | Nucleotide sequences coding for the genes sucC and sucD |
| KR20040014389A (ko) * | 2000-07-21 | 2004-02-14 | 메타볼릭스 인코포레이티드 | 폴리올로부터 폴리하이드록시알칸노에이트를 제조하는 방법 |
| US6495152B2 (en) | 2000-08-18 | 2002-12-17 | Tepha, Inc. | Sulfur containing polyhydroxyalkanoate compositions and method of production |
| US6916637B2 (en) | 2000-09-30 | 2005-07-12 | Degussa Ag | Fermentation process for the preparation of L-amino acids using strains of the family Enterobacteriaceae |
| CA2429039A1 (en) | 2000-11-20 | 2002-05-30 | Cargill, Incorporated | 3-hydroxypropionic acid and other organic compounds |
| US7501268B2 (en) * | 2000-12-28 | 2009-03-10 | Toyota Jidosha Kabushiki Kaisha | Methods of producing prenyl alcohols |
| JP4776146B2 (ja) | 2001-01-10 | 2011-09-21 | ザ・ペン・ステート・リサーチ・ファンデーション | 細胞代謝をモデル化する方法及びシステム |
| US7127379B2 (en) * | 2001-01-31 | 2006-10-24 | The Regents Of The University Of California | Method for the evolutionary design of biochemical reaction networks |
| CA2439260C (en) | 2001-03-01 | 2012-10-23 | The Regents Of The University Of California | Models and methods for determining systemic properties of regulated reaction networks |
| US7052883B2 (en) | 2001-04-03 | 2006-05-30 | Degussa Ag | Process for the production of L-amino acids using strains of the family Enterobacteriaceae that contain an attenuated fruR gene |
| WO2003008616A2 (en) | 2001-07-18 | 2003-01-30 | Degussa Ag | PROCESS FOR THE PREPARATION OF L-AMINO ACIDS USING STRAINS OF THE ENTEROBACTERIACEAE FAMILY WHICH CONTAIN AN ATTENUATED aceK GENE |
| US20090100536A1 (en) | 2001-12-04 | 2009-04-16 | Monsanto Company | Transgenic plants with enhanced agronomic traits |
| US20090158452A1 (en) | 2001-12-04 | 2009-06-18 | Johnson Richard G | Transgenic plants with enhanced agronomic traits |
| CN1217001C (zh) * | 2002-01-04 | 2005-08-31 | 清华大学 | 一种生产d-(-)-3-羟基丁酸的方法 |
| ES2329778T3 (es) * | 2002-01-18 | 2009-12-01 | Novozymes A/S | 2,3-aminomutasa de alanina. |
| US7314974B2 (en) | 2002-02-21 | 2008-01-01 | Monsanto Technology, Llc | Expression of microbial proteins in plants for production of plants with improved properties |
| US20030224363A1 (en) | 2002-03-19 | 2003-12-04 | Park Sung M. | Compositions and methods for modeling bacillus subtilis metabolism |
| CA2480216A1 (en) | 2002-03-29 | 2003-10-09 | Genomatica, Inc. | Human metabolic models and methods |
| JPWO2003095651A1 (ja) * | 2002-05-10 | 2005-09-15 | 協和醗酵工業株式会社 | メバロン酸の製造方法 |
| US7856317B2 (en) | 2002-06-14 | 2010-12-21 | Genomatica, Inc. | Systems and methods for constructing genomic-based phenotypic models |
| AU2003256480B2 (en) | 2002-07-10 | 2008-03-06 | The Penn State Research Foundation | Method for determining gene knockout strategies |
| US7413878B2 (en) * | 2002-07-15 | 2008-08-19 | Kosan Biosciences, Inc. | Recombinant host cells expressing atoAD and capable of making a polyketide using a starter unit |
| JP4578240B2 (ja) | 2002-08-09 | 2010-11-10 | コデクシス, インコーポレイテッド | 4−置換3−ヒドロキシ酪酸誘導体の生成のための酵素的プロセス |
| WO2004024876A2 (en) | 2002-09-12 | 2004-03-25 | Metabolix, Inc. | Polyhydroxyalkanoate production by coenzyme a-dependent aldehyde dehydrogenase pathways |
| KR101070110B1 (ko) | 2002-09-27 | 2011-10-05 | 디에스엠 아이피 어셋츠 비.브이. | 알데히드 데하이드로게나제 유전자 |
| US7734420B2 (en) | 2002-10-15 | 2010-06-08 | The Regents Of The University Of California | Methods and systems to identify operational reaction pathways |
| AU2003287625A1 (en) * | 2002-11-06 | 2004-06-03 | University Of Florida | Materials and methods for the efficient production of acetate and other products |
| JP2007502111A (ja) | 2003-08-11 | 2007-02-08 | コデクシス, インコーポレイテッド | 4−置換3−ヒドロキシ酪酸誘導体およびビシナルなシアノ,ヒドロキシ置換カルボン酸エステルの生成のための酵素的プロセス |
| US7927859B2 (en) | 2003-08-22 | 2011-04-19 | Rice University | High molar succinate yield bacteria by increasing the intracellular NADH availability |
| FR2864967B1 (fr) * | 2004-01-12 | 2006-05-19 | Metabolic Explorer Sa | Microorganisme evolue pour la production de 1,2-propanediol |
| DE102004031177A1 (de) * | 2004-06-29 | 2006-01-19 | Henkel Kgaa | Neue Geruchsstoffe bildende Genprodukte von Bacillus licheniformis und darauf aufbauende verbesserte biotechnologische Produktionsverfahren |
| US7601858B2 (en) | 2004-08-17 | 2009-10-13 | Gs Cleantech Corporation | Method of processing ethanol byproducts and related subsystems |
| KR101245428B1 (ko) | 2004-08-27 | 2013-03-19 | 라이스 유니버시티 | 숙신산 생산이 증가된 돌연변이 대장균 균주 |
| GB0419424D0 (en) | 2004-09-02 | 2004-10-06 | Viragen Scotland Ltd | Transgene optimisation |
| JP4712716B2 (ja) * | 2004-09-09 | 2011-06-29 | 財団法人地球環境産業技術研究機構 | プロモーター機能を有するdna断片 |
| JP2008513023A (ja) * | 2004-09-17 | 2008-05-01 | ライス ユニバーシティー | 高コハク酸生産細菌 |
| WO2006069174A2 (en) | 2004-12-22 | 2006-06-29 | Rice University | Simultaneous anaerobic production of isoamyl acetate and succinic acid |
| KR100679638B1 (ko) | 2005-08-19 | 2007-02-06 | 한국과학기술원 | 포메이트 디하이드로게나제 d 또는 e를 코딩하는 유전자로 형질전환된 미생물 및 이를 이용한 숙신산의 제조방법 |
| KR100676160B1 (ko) | 2005-08-19 | 2007-02-01 | 한국과학기술원 | 말릭효소를 코딩하는 유전자로 형질전환된 재조합 미생물 및 이를 이용한 숙신산의 제조방법 |
| JP2009507493A (ja) * | 2005-09-09 | 2009-02-26 | ジェノマティカ・インコーポレイテッド | 増殖連動型のコハク酸塩生成のための方法と生物 |
| US9297028B2 (en) * | 2005-09-29 | 2016-03-29 | Butamax Advanced Biofuels Llc | Fermentive production of four carbon alcohols |
| US20080274526A1 (en) | 2007-05-02 | 2008-11-06 | Bramucci Michael G | Method for the production of isobutanol |
| US8206970B2 (en) | 2006-05-02 | 2012-06-26 | Butamax(Tm) Advanced Biofuels Llc | Production of 2-butanol and 2-butanone employing aminobutanol phosphate phospholyase |
| US8017364B2 (en) * | 2006-12-12 | 2011-09-13 | Butamax(Tm) Advanced Biofuels Llc | Solvent tolerant microorganisms |
| US8673601B2 (en) * | 2007-01-22 | 2014-03-18 | Genomatica, Inc. | Methods and organisms for growth-coupled production of 3-hydroxypropionic acid |
| LT2821494T (lt) * | 2007-03-16 | 2017-04-25 | Genomatica, Inc. | 1,4-butandiolio ir jo pirmtakų biosintezės būdai ir kompozicijos |
| CA2688938C (en) * | 2007-03-20 | 2018-01-16 | University Of Florida Research Foundation, Inc. | Materials and methods for efficient succinate and malate production |
| CN101680007A (zh) | 2007-04-18 | 2010-03-24 | 纳幕尔杜邦公司 | 使用高活性酮醇酸还原异构酶来发酵生产异丁醇 |
| WO2008131286A1 (en) | 2007-04-18 | 2008-10-30 | Gevo, Inc. | Engineered microorganisms for producing isopropanol |
| US8426173B2 (en) | 2007-05-02 | 2013-04-23 | Butamax (Tm) Advanced Biofuels Llc | Method for the production of 1-butanol |
| US8426174B2 (en) | 2007-05-02 | 2013-04-23 | Butamax(Tm) Advanced Biofuels Llc | Method for the production of 2-butanol |
| US20090023182A1 (en) * | 2007-07-18 | 2009-01-22 | Schilling Christophe H | Complementary metabolizing organisms and methods of making same |
| US7947483B2 (en) | 2007-08-10 | 2011-05-24 | Genomatica, Inc. | Methods and organisms for the growth-coupled production of 1,4-butanediol |
| KR101042242B1 (ko) | 2007-09-07 | 2011-06-17 | 한국과학기술원 | 1,4-부탄디올 생성능을 가지는 변이체 및 이를 이용한1,4-부탄디올의 제조방법 |
| EP2198039A4 (en) | 2007-10-12 | 2012-05-09 | Univ California | MODIFIED MICROORGANISMS FOR THE MANUFACTURE OF ISOPROPANOL |
| CN101978042B (zh) | 2008-01-22 | 2017-03-08 | 基因组股份公司 | 利用合成气或其它气态碳源和甲醇的方法和有机体 |
| WO2009103026A1 (en) | 2008-02-15 | 2009-08-20 | Gevo, Inc. | Engineered microorganisms for producing isopropanol |
| BRPI0915749A2 (pt) | 2008-07-08 | 2018-07-10 | Opx Biotechnologies Inc | métodos, composições e sistemas para produção biossintética de 1,4-butanodiol |
| US7858350B2 (en) | 2008-09-10 | 2010-12-28 | Genomatica, Inc. | Microorganisms for the production of 1,4-butanediol |
| US8354563B2 (en) * | 2008-10-16 | 2013-01-15 | Maverick Biofuels, Inc. | Methods and apparatus for synthesis of alcohols from syngas |
| EP2373781A4 (en) | 2008-12-16 | 2012-10-10 | Genomatica Inc | MICRO-ORGANISMS AND METHODS OF CONVERTING SYNGAS AND OTHER CARBON RESOURCES IN USEFUL PRODUCTS |
| US20110014669A1 (en) | 2009-01-23 | 2011-01-20 | Microbia, Inc. | Production of 1,4 Butanediol in a Microorganism |
| WO2010141920A2 (en) * | 2009-06-04 | 2010-12-09 | Genomatica, Inc. | Microorganisms for the production of 1,4-butanediol and related methods |
| US8268607B2 (en) | 2009-12-10 | 2012-09-18 | Genomatica, Inc. | Methods and organisms for converting synthesis gas or other gaseous carbon sources and methanol to 1,3-butanediol |
-
2007
- 2007-08-10 US US11/891,602 patent/US7947483B2/en active Active
-
2008
- 2008-08-06 EP EP18183168.6A patent/EP3447133B1/en active Active
- 2008-08-06 EP EP13001196.8A patent/EP2679685B1/en not_active Not-in-force
- 2008-08-06 EP EP13001195.0A patent/EP2679684A1/en not_active Withdrawn
- 2008-08-06 PL PL13001197T patent/PL2679676T3/pl unknown
- 2008-08-06 CA CA3175485A patent/CA3175485A1/en active Pending
- 2008-08-06 HU HUE13001197A patent/HUE040654T2/hu unknown
- 2008-08-06 BR BRPI0815138-5A2A patent/BRPI0815138A2/pt not_active Application Discontinuation
- 2008-08-06 ES ES08782639.2T patent/ES2583179T3/es active Active
- 2008-08-06 EP EP08782639.2A patent/EP2185708B1/en active Active
- 2008-08-06 EP EP13001197.6A patent/EP2679676B1/en active Active
- 2008-08-06 TR TR2018/14888T patent/TR201814888T4/en unknown
- 2008-08-06 LT LTEP13001197.6T patent/LT2679676T/lt unknown
- 2008-08-06 ES ES13001197T patent/ES2695125T3/es active Active
- 2008-08-06 DK DK13001197.6T patent/DK2679676T3/en active
- 2008-08-06 CA CA2695706A patent/CA2695706C/en active Active
- 2008-08-06 WO PCT/US2008/072362 patent/WO2009023493A1/en not_active Ceased
- 2008-08-06 AU AU2008287072A patent/AU2008287072B2/en active Active
- 2008-08-06 SI SI200832003T patent/SI2679676T1/sl unknown
-
2011
- 2011-03-18 US US13/065,303 patent/US8470582B2/en active Active
-
2013
- 2013-06-10 US US13/914,422 patent/US20140030780A1/en not_active Abandoned
-
2017
- 2017-02-03 US US15/424,723 patent/US20170240931A1/en not_active Abandoned
-
2019
- 2019-05-31 US US16/428,627 patent/US20200095616A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| EP3447133B1 (en) | 2023-01-04 |
| EP2185708A1 (en) | 2010-05-19 |
| AU2008287072B2 (en) | 2015-05-14 |
| EP2679685A1 (en) | 2014-01-01 |
| ES2583179T3 (es) | 2016-09-19 |
| US20110201071A1 (en) | 2011-08-18 |
| EP2679676B1 (en) | 2018-07-18 |
| US20200095616A1 (en) | 2020-03-26 |
| AU2008287072A1 (en) | 2009-02-19 |
| BRPI0815138A2 (pt) | 2014-10-21 |
| DK2679676T3 (en) | 2018-10-29 |
| PL2679676T3 (pl) | 2019-03-29 |
| US20170240931A1 (en) | 2017-08-24 |
| EP2679685B1 (en) | 2018-07-18 |
| US8470582B2 (en) | 2013-06-25 |
| HUE040654T2 (hu) | 2019-03-28 |
| US20090047719A1 (en) | 2009-02-19 |
| CA2695706A1 (en) | 2009-02-19 |
| CA2695706C (en) | 2022-09-27 |
| US20140030780A1 (en) | 2014-01-30 |
| US7947483B2 (en) | 2011-05-24 |
| EP3447133A1 (en) | 2019-02-27 |
| WO2009023493A1 (en) | 2009-02-19 |
| EP2679676A1 (en) | 2014-01-01 |
| CA3175485A1 (en) | 2009-02-19 |
| SI2679676T1 (sl) | 2018-12-31 |
| TR201814888T4 (en) | 2018-11-21 |
| LT2679676T (lt) | 2018-11-12 |
| EP2185708B1 (en) | 2016-05-04 |
| EP2679684A1 (en) | 2014-01-01 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2695125T3 (es) | Métodos y organismos para la producción de 1,4-butanodiol acoplada al crecimiento | |
| US20230134936A1 (en) | Compositions and methods for the biosynthesis of 1,4-butanediol and its precursors | |
| AU2013203440A1 (en) | Methods and organisms for the growth-coupled production of 1,4-butanediol | |
| AU2013204038B2 (en) | Compositions and methods for the biosynthesis of 1,4-butanediol and its precursors | |
| BR122018011330B1 (pt) | Micro-organismo de ocorrência não-natural e método para a produção de 1,4-butanodiol | |
| BR122018011328B1 (pt) | Microrganismo de ocorrência não natural e método para a produção de 1,4-butanodiol | |
| BR122018011304B1 (pt) | Métodos e microrganismos para a produção de 1,4-butanodiol |