ES2713437T3 - Producción de mixopironina y de sus derivados - Google Patents
Producción de mixopironina y de sus derivados Download PDFInfo
- Publication number
- ES2713437T3 ES2713437T3 ES14725669T ES14725669T ES2713437T3 ES 2713437 T3 ES2713437 T3 ES 2713437T3 ES 14725669 T ES14725669 T ES 14725669T ES 14725669 T ES14725669 T ES 14725669T ES 2713437 T3 ES2713437 T3 ES 2713437T3
- Authority
- ES
- Spain
- Prior art keywords
- seq
- module
- amino acids
- mxnk
- enzyme
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P17/00—Preparation of heterocyclic carbon compounds with only O, N, S, Se or Te as ring hetero atoms
- C12P17/02—Oxygen as only ring hetero atoms
- C12P17/06—Oxygen as only ring hetero atoms containing a six-membered hetero ring, e.g. fluorescein
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Enzymes And Modification Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
Proceso para producir un compuesto que tiene una primera cadena (cadena occidental) y una segunda cadena (cadena oriental) que se condensan formando un anillo pirona al proporcionar a un microorganismo las enzimas de la ruta sintética que catalizan la síntesis del compuesto, en donde el microorganismo se manipula genéticamente para comprender genes que codifican las enzimas de la ruta sintética que comprenden las enzimas MxnA (SEQ ID NO: 1), MxnB (SEQ ID NO: 2), MxnC (SEQ ID NO: 3), MxnD (SEQ ID NO: 4), MxnE (SEQ ID NO: 5), MxnF (SEQ ID NO: 6), MxnG (SEQ ID NO: 7), MxnH (SEQ ID NO: 8), Mxnl (SEQ ID NO: 9), MxnJ (SEQ ID NO: 10) y MxnK (SEQ ID NO: 11), incubar el microorganismo y separar el compuesto del microorganismo, caracterizado por que al menos un dominio catalítico de una enzima está ausente o inactivado, dominio que se selecciona del grupo que consiste en a) la cetosintasa (KS) del módulo 1E de Mxnl que comprende los aminoácidos No. 694-885 de SEQ ID NO: 9, b) la cetosintasa (KS) del módulo 2E de Mxnl que comprende los aminoácidos No. 1945-2135 de SEQ ID NO: 9, c) la cetosintasa (KS) del módulo 3E de Mxnl que comprende los aminoácidos No. 3442-3633 de SEQ ID NO: 10, d) la cetosintasa (KS) del módulo 4E de MxnJ que comprende los aminoácidos No. 605-795 de SEQ ID NO: 10, e) la cetosintasa (KS) del módulo 5E de MxnJ que comprende los aminoácidos No. 2476-2660 de SEQ ID NO: 10, f) la cetosintasa (KS) del módulo 6E de MxnJ que comprende los aminoácidos No. 3208-3399 de SEQ ID NO: 10, g) la cetosintasa (KS) del módulo 1W de MxnK que comprende los aminoácidos No. 180-364 de SEQ ID NO: 11, h) la cetosintasa (KS) del módulo 2W de MxnK que comprende los aminoácidos No. 1926-2110 de SEQ ID NO: 11, i) la cetosintasa (KS) del módulo 3W de MxnK que comprende los aminoácidos No. 2684-2875 de SEQ ID NO: 11, j) la cetosintasa (KS) del módulo 4W de MxnK que comprende los aminoácidos No. 3504-3688 de SEQ ID NO: 11, k) la cetosintasa (KS) del módulo 5W de MxnK que comprende los aminoácidos No. 5409-5593 de SEQ ID NO: 11, I) la cetorreductasa (KR) del módulo 2E de Mxnl que comprende los aminoácidos No. 2903-3050 de SEQ ID NO: 9, m) la cetorreductasa (KR) del módulo 4E de MxnJ que comprende los aminoácidos No. 1568-1715 de SEQ ID NO: 10, n) la cetorreductasa (KR) del módulo 1W de MxnK que comprende los aminoácidos No. 1418-1560 de SEQ ID NO: 11, o) la cetorreductasa (KR) del módulo 4W de MxnK que comprende los aminoácidos No. 4470-4617 de SEQ ID NO: 11, p) la proteína transportadora (CP) del módulo L de Mxnl que comprende los aminoácidos No. 424-433 de SEQ ID NO:9, q) la proteína transportadora (CP) del módulo 1E de Mxnl que comprende los aminoácidos No. 1720-1726 de SEQ ID NO: 9, r) la proteína transportadora (CP) del módulo 2E de Mxnl que comprende los aminoácidos No. 3192-3198 de SEQ ID NO: 9, s) la proteína transportadora (CP) del módulo 3E de MxnJ que comprende los aminoácidos No. 366-372 de SEQ ID NO: 9, t) la proteína transportadora (CP) del módulo 4E de MxnJ que comprende los aminoácidos No. 2238-2244 de SEQ ID NO: 10, u) la proteína transportadora (CP) del módulo 5E de MxnJ que comprende los aminoácidos No. 2967-2973 de SEQ ID NO: 10, v) la proteína transportadora (CP) del módulo 6E de MxnJ que comprende los aminoácidos No. 3810-3816 de SEQ ID NO: 10, w) la proteína transportadora (CP*) del módulo 6E de MxnJ que comprende los aminoácidos No. 3566-3572 de SEQ ID NO: 12, x) la proteína transportadora (CP) del módulo 1W de MxnK, que comprende los aminoácidos No. 1683-1689 de SEQ ID NO: 11 y) la proteína transportadora (CP) del módulo 2W de MxnK que comprende los aminoácidos No. 2429-2435 de SEQ ID NO: 11, z) la proteína transportadora (CP) del módulo 3W de MxnK que comprende los aminoácidos No. 3234-3240 de SEQ ID NO: 11, aa) la proteína transportadora (CP) del módulo 4W de MxnK que comprende los aminoácidos No. 5165-5171 de SEQ ID NO: 11, bb) la proteína transportadora (CP*) del módulo 5W de MxnK que comprende los aminoácidos No. 5894-5900 de SEQ ID NO: 11, cc) la proteína transportadora (CP) del módulo 5W de MxnK que comprende los aminoácidos No. 5984-5990 de SEQ ID NO: 11, dd) la deshidratasa (DH) del módulo 2E de Mxnl que comprende los aminoácidos No. 2402-2414 de SEQ ID NO: 9, ee) la deshidratasa (DH) del módulo 3E de MxnJ que comprende los aminoácidos No. 53-65 de SEQ ID NO: 9, ff) la deshidratasa (DH) del módulo 4E de MxnJ que comprende los aminoácidos No. 1091-1103 de SEQ ID NO: 10, gg) la deshidratasa (DH) del módulo 1W de MxnK que comprende los aminoácidos No. 955-967 de SEQ ID NO: 11, hh) la deshidratasa (DH) del módulo 4W de MxnK que comprende los aminoácidos No. 3980-3992 de SEQ ID NO: 11, ii) la metiltransferasa (MT) del módulo 4E de MxnJ que comprende los aminoácidos No. 1982-1987 de SEQ ID NO: 10, jj) la metiltransferasa (MT) del módulo 4W de MxnK que comprende los aminoácidos No. 4914-4919 de SEQ ID NO: 11, en donde en las realizaciones en las que al menos un dominio catalítico está inactivado en las enzimas de la ruta sintética MxnK y/o MxnI-MxnJ, en donde el dominio catalítico inactivado es un dominio catalítico cuya inactivación en ausencia de precursor da lugar a una parada de la síntesis por la enzima cuando se expresa en un microorganismo, un precursor de la cadena occidental, y/o un precursor de la cadena oriental se añade a un microorganismo que expresa las enzimas de la ruta sintética, o en donde en las realizaciones en las que la inactivación es por intercambio de al menos un centro catalítico, dominio, módulo o enzima seleccionada de las enzimas CorJ (SEQ ID NO: 25), CorK (SEQ ID NO: 26), CorL (SEQ ID NO: 27), Orf1 (SEQ ID NO: 27), CorM (SEQ ID NO: 28), CorM (SEQ ID NO: 28), CorN (SEQ ID NO: 29) y CorO (SEQ ID NO: 30) y sus centros catalíticos, dominios y módulos, o la enzima MxnI (SEQ ID NO: 9) se reemplaza por la enzima CorI (SEQ ID NO: 24).
Description
DESCRIPCION
Produccion de mixopironina y de sus derivados
La presente invencion se refiere a un proceso para la produccion del compuesto mixobacteriano mixopironina y de derivados de mixopironina, p. ej., para la produccion de una composicion farmaceutica que contiene mixopironina y/o su derivado. Ademas, la invencion se refiere a los derivados que se pueden obtener por el proceso de produccion, especialmente para el uso del derivado en un tratamiento medico, p. ej., en el tratamiento medico de una infeccion bacteriana. El proceso de produccion usa enzimas de la ruta sintetica natural de la mixopironina para producir derivados de mixopironina y usa las enzimas que constituyen la ruta sintetica completa de la mixopironina para producir mixopironina. El proceso de produccion usa las enzimas expresadas en un microorganismo, que puede ser el organismo fuente de los genes que codifican las enzimas, o usa las enzimas expresadas en un microorganismo huesped heterologo, p. ej., una bacteria u hongo, incluyendo una levadura. Para la expresion de las enzimas en un microorganismo, los genes que codifican las enzimas estan contenidos en el microorganismo. Ademas de o alternativamente a la expresion de las enzimas en un microorganismo, el proceso de produccion puede usar las enzimas en un medio sin celulas, p. ej., despues de la disrupcion y/o aislamiento de las enzimas de un microorganismo que expresa las enzimas, opcionalmente con la inmovilizacion de las enzimas en un transportador, y/o preferiblemente con la adicion de al menos un compuesto precursor a mixopironina o su derivado.
Estado de la tecnica
Erol et al., ChemBioChem 2010, 11, 1253-1265 describen los genes de la agrupacion genica posible biosintetica de la coralopironina A de Corallococcus coralloides y experimentos de alimentacion usando acetato, bicarbonato de sodio, metil-metionina, y glicina marcados con 13C y 15N para deducir la ruta sintetica natural para la coralopironina A a partir de analisis de RMN.
Irschik et al., J. Antibiot. 1983 describen que Myxococcus fulvus produce mixopironina. La mixopironina puede describirse como dos mitades de molecula, tambien denominadas cadena occidental y cadena oriental, que estan fusionadas en un resto de anillo pirona:

Kohl et al., Liebigs Ann. Chem. 1983 describen el aislamiento y elucidacion de la estructura de la mixopironina A y B de Myxococcus fulvus Mx f50.
Kohl et al., Liebigs Ann. Chem. 1984 describen el origen biosintetico de la mixopironina A sobre la base de experimentos de alimentacion con precursores marcados con 13C y 15N y el analisis del patron de marcaje por espectroscopfa de RMN
Sucipto et al., ChemBioChem 2013, describe la produccion de mixopironina y los genes implicados.
Objeto de la invencion
Es un objeto de la invencion proporcionar un proceso alternativo para la produccion de mixopironina y de sus derivados, y preferiblemente proporcionar nuevos derivados de mixopironina y un proceso para su produccion.
Descripcion general de la invencion
La invencion consigue el objeto por las caractensticas de las reivindicaciones. Generalmente, la invencion proporciona un proceso para producir mixopironina y derivados de mixopironina proporcionando enzimas de la ruta sintetica en un microorganismo o in vitro, p. ej., extracelularmente, por ejemplo, despues de la expresion de las enzimas en un microorganismo y de la separacion de las enzimas del microorganismo. La mixopironina y los derivados de la mixopironina producidos segun la invencion son compuestos en los que una primera cadena, tambien denominada cadena occidental, y una segunda cadena, tambien denominada cadena oriental, se condensan formando un anillo
pirona. Generalmente, las realizaciones de la invencion pueden combinarse entre sft Las enzimas de la ruta sintetica pueden producirse por la expresion de al menos un casete de expresion que comprende las secuencias de acido nucleico que codifican las enzimas en un microorganismo, en donde una o mas secuencias codificadoras pueden estar contenidas en un casete de expresion, p. ej., cada secuencia codificadora puede estar contenida en un casete de expresion separado o al menos dos secuencias codificadoras pueden estar contenidas en un casete de expresion. En detalle, se describe un proceso para producir mixopironina por las enzimas que constituyen la ruta sintetica, en donde preferiblemente estas enzimas se expresan en un microorganismo que esta geneticamente manipulado para contener genes que codifican las enzimas, en donde el microorganismo preferiblemente es un microorganismo huesped heterologo, o proporcionando las enzimas en un medio sin celulas. Estas enzimas son MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnK-MxnL-MxnM, que en el proceso preferiblemente estan codificadas en 5' a 3' por los genes mxnA-mxnB-mxnC-mxnD-mxnE-mxnF-mxnG-mxnH-mxnl-mxnJ-mxnK-mxnL-mxnM. Las enzimas MxnA, Mxnl y MxnJ, preferiblemente adicionalmente MxnH, catalizan la smtesis de la parte de la molecula de la cadena oriental, y las enzimas MxnA y MxnK, preferiblemente adicionalmente MxnC-MxnD-MxnE-MxnF-MxnG, catalizan la ruta biosintetica para la smtesis de la parte de la molecula de la cadena occidental. Preferiblemente, la enzima MxnA se proporciona, p. ej., para catalizar la carga del modulo de la protema transportadora de Mxnl y/o del modulo de la protema transportadora de MxnK y/o del modulo de la protema transportadora de MxnJ.
Generalmente, las enzimas se describen en la presente memoria como que comprenden modulos como sus partes o que consisten en modulos como sus partes. Los modulos comprenden como sus partes dominios catalfticos que contienen centros catalfticos. Las secuencias de acidos nucleicos se proporcionan en 5'-3', las secuencias de aminoacidos se proporcionan del extremo N al extremo C.
Para la reaccion de condensacion, la parte de la molecula de la cadena oriental y la parte de la molecula de la cadena occidental se unen ambas por enzimas, p. ej., a un dominio de protema transportadora, a traves de un atomo de azufre (S), y se condensan en sus extremos unidos por S respectivos, dando lugar a un anillo pirona, reaccion de condensacion que esta catalizada por la enzima MxnB. En la alternativa, la parte de la molecula de la cadena oriental y/o la parte de la molecula de la cadena occidental puede activarse mediante su union al atomo de azufre de un grupo auxiliar, y la enzima MxnB puede estar presente en un medio sin celulas, p. ej., como una protema aislada, para catalizar la reaccion de condensacion al anillo pirona.
Las enzimas MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnK-MxnL-MxnM que constituyen la ruta sintetica natural pueden expresarse en un microorganismo huesped heterologo.
En la invencion, al menos una enzima de la misma puede inactivarse, p. ej., mutarse, para obtener disfuncionalidad, o delecionarse, para producir mixopironina o un derivado de la misma. Por ejemplo, la enzima MxnL y/o MxnM y/o MxnC-MxnG puede inactivarse y las enzimas activas pueden consistir en
MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-MxnI-MxnJ-MxnK-MxnM,
MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-MxnI-MxnJ-MxnK-MxnL,
MxnA-MxnB-MxnH-MxnI-MxnJ-MxnK-MxnL-MxnM,
MxnA-MxnB-MxnH-MxnI-MxnJ-MxnK-MxnM,
MxnA-MxnB-MxnH-MxnI-MxnJ-MxnK-MxnL o
MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-MxnI-MxnJ-MxnK.
Alternativamente o adicionalmente a la inactivacion de la enzima MxnL y/o MxnM y/o MxnC-MxnG, al menos un centro catalftico de al menos una de MxnK y Mxnl-MxnJ puede inactivarse, centro catalftico que es preferiblemente una cetosintasa (KS), cetorreductasa (KR), deshidrogenasa (DH), metiltransferasa (MT), hidroximetilglutaril-CoA sintasa (HMG-CoA), enoil-CoA hidratasa/isomerasa (ECH).
En el caso de la inactivacion de al menos una actividad catalftica de las enzimas de la ruta sintetica, las enzimas pueden expresarse en una cepa productora de mixopironina original que, p. ej., se ha manipulado geneticamente, p. ej., usando recombinacion homologa, para la inactivacion de al menos una enzima o centro catalftico, p. ej., cuando se proporcionan las enzimas por expresion homologa. En la primera realizacion, el proceso comprende cultivar un microorganismo que expresa las enzimas y aislar la mixopironina o su derivado del medio de cultivo y/o del microorganismo.
En una primera realizacion, la invencion proporciona un proceso para producir mixopironina o un proceso para producir al menos un derivado de mixopironina mediante el cultivo de un microorganismo que expresa al menos enzimas parcialmente modificadas, que se modifican, p. ej., por inactivacion y/o cambio de la actividad de al menos una enzima
de las enzimas MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnK-MxnL-MxnM y/o de al menos un dominio de una de estas enzimas y/o de al menos un centro catalftico de uno de estos dominios. La inactivacion puede ser por al menos la delecion parcial y/o intercambio, p. ej., de al menos un aminoacido o por la delecion completa de una enzima, dominio o centro catalftico.
La insercion o intercambio de al menos un centro catalftico, dominio, modulo o enzima puede ser por insercion o reemplazo de al menos un centro catalftico, dominio o enzima adicional en las enzimas de la ruta sintetica que comprende o consiste en MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnK-MxnL-MxnM o una parte de estas, centro catalftico, dominio, modulo o enzima que se selecciona de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnK-MxnL-MxnM, dando lugar, p. ej., al menos a un centro catalftico, dominio, modulo o enzima homologo adicional o intercambiado, y/o centro catalftico, dominio, modulo o enzima que se selecciona de las enzimas CorJ, CorK, CorL, Orf1, CorM, CorL, CorM, CorN y CorO y sus centros catalfticos, dominios y modulos, respectivamente.
Generalmente, los cambios en la secuencia de aminoacidos se hacen preferiblemente por cambios en la secuencia de acido nucleico que codifica la secuencia de aminoacidos de la enzima, dominio o centro catalftico, respectivamente. Las secuencias de aminoacidos de las enzimas se proporcionan en la Tabla 1.
Tabla 1: enzimas y funciones

Los dominios catalfticos son: PM, fosfomutasa; CP, protema transportadora; KS, cetosintasa; A, adenilacion; DH, deshidratasa; KR, cetorreductasa; AT, aciltransferasa; MT, metiltransferasa; ER, enoil reductasa; KS, cetosintasa. Un "*" indica la inactividad del dominio. En la alternativa a las enzimas de Myxococcus, puede usarse la enzima indicada de Corallococcus. La referencia a un gen de Myxococcus incluye, por lo tanto, la referencia a un gen de Corallococcus indicado en la misma fila de la tabla 1.
El modulo de elongacion C-terminal de MxnJ se incluye en el dominio 6E de MxnJ.
El conjunto completo de enzimas de la ruta sintetica de Myxococcus esta codificado por SEQ ID NO: 14. Las secuencias de aminoacidos de los dominios funcionales se proporcionan en la Tabla 2.
Tabla 2: modulos de enzimas

en donde aa es aminoacidos, seguido de la numeracion de los aminoacidos en las SEQ ID NO indicadas. Las secuencias de aminoacidos de los centros catalfticos de los dominios se proporcionan en la Tabla 3 mas adelante.
En una segunda realizacion, un compuesto precursor para la cadena occidental, tambien denominado precursor de la cadena occidental, y/o un compuesto precursor para la cadena oriental, tambien denominado precursor de la cadena oriental, se anade a un microorganismo que expresa las enzimas de la ruta sintetica de la primera y/o segunda realizacion, en la que al menos un dominio catalftico se inactiva en las enzimas de la ruta sintetica MxnK y/o MxnI-MxnJ, en donde el dominio catalftico inactivado es preferiblemente un dominio catalftico cuya inactivacion en ausencia de precursor da lugar a una parada de la smtesis por la enzima cuando se expresa en un microorganismo. Para la tercera realizacion, un dominio catalftico inactivado preferido se selecciona de una protema transportadora (CP) y una cetorreductasa (KR). La secuencia de aminoacidos respectiva de dominios catalfticos para MxnK y Mxnl-MxnJ se proporcionan en la Tabla 3 mas adelante.
En la segunda realizacion, una parte del conjunto completo de enzimas sinteticas se proporciona en un microorganismo, parte que cataliza la condensacion del anillo pirona, la parte, p. ej., que comprende o consiste en las enzimas MxnB, opcionalmente MxnA-MxnB, y se proporciona un precursor de la cadena occidental y al menos una parte de MxnK para la union del precursor de la cadena occidental y catalizar la conversion del precursor de la cadena occidental en un precursor de la cadena occidental convertido y MxnB, opcionalmente MxnH y/o MxnA, y ademas opcionalmente adicionalmente MxnC-MxnD-MxnE-MxnF-MxnG. Esta realizacion permite la produccion de compuestos en los que el precursor de la cadena occidental convertido se condensa en la parte de la molecula de la cadena oriental natural. Adicionalmente o alternativamente, se proporciona un precursor de la cadena oriental y se proporciona una parte de Mxnl-MxnJ para la union del precursor de la cadena oriental y catalizar la conversion del precursor de la
cadena oriental a un precursor de la cadena oriental convertido, que se condensa con el precursor de la cadena occidental convertido o con la parte de la molecula occidental. Cuando se anade un precursor de la cadena occidental, se proporcionan las siguientes enzimas por manipulacion genetica en un microorganismo con los genes que codifican las enzimas y la expresion de las enzimas, en combinacion con proporcionar un precursor de la cadena occidental. Las enzimas que se proporcionan pueden, p. ej., comprender o consistir en: MxnA-MxnB, Mxnl-MxnJ, preferiblemente MxnH, y MxnK en donde al menos un dominio catalftico se inactiva, dando lugar la inactivacion a la interrupcion de la ruta sintetica de MxnK, p. ej., MxnK que consiste opcionalmente en los modulos 3W-5W, modulos 4W-5W o modulo 5W de MxnK, y ademas opcionalmente adicionalmente MxnC-D-E-F-G.
En la segunda realizacion, en una primera variante, en la enzima MxnK de tipo salvaje (SEQ ID NO: 1) que consiste en los modulos 1W-5W, al menos un modulo puede estar funcionalmente ausente, p. ej., estar delecionado o inactivado, los modulos 1W-2W (los aminoacidos No. 1 a 2531, preferiblemente los aminoacidos No. 1 a 2467 en SEQ ID NO: 11) pueden estar ausentes o MxnK puede consistir en los modulos 3W a 5W, respectivamente. En una segunda variante, los modulos 1W a 3W (los aminoacidos No. 1 a 3348, preferiblemente los aminoacidos No. 1 a 3272 en SEQ ID NO: 11) pueden estar funcionalmente ausentes, p. ej., estar delecionados o inactivados, o la parte de enzima MxnK puede consistir en los modulos 4W a 5W, respectivamente. En una tercera variante, los modulos 1W a 4W (los aminoacidos No. 1 a 5251, preferiblemente los aminoacidos No. 1 a 5203 en SEQ ID NO: 11) pueden estar funcionalmente ausentes, p. ej., estar delecionados o inactivados, o la parte de enzima MxnK puede consistir en el modulo 5W (los aminoacidos No. 5252 a 6023, preferiblemente aminoacidos los No. 5204 a 6023 en SEQ ID NO: 11), respectivamente. En variantes, los modulos funcionalmente ausentes pueden, p. ej., ser modulos en los que los aminoacidos que forman al menos uno de sus dominios cataltticos, estan mutados o delecionados.
Cuando se proporciona un precursor de la cadena occidental, preferiblemente una parte de la enzima MxnK esta presente y el precursor de la cadena occidental puede tener una estructura que se une entonces a un dominio de MxnK, preferiblemente a la protema transportadora (CP), p. ej., el precursor de la cadena occidental preferiblemente tiene una estructura que comprende un grupo ceto unido a un atomo de azufre de un grupo auxiliar. Preferiblemente, la estructura del precursor de la cadena occidental mimetiza un intermedio biosintetico. El grupo auxiliar puede, p. ej., estar comprendido o consistir en un grupo p-ceto-A/-acetilcisteamina (SNAC) o un grupo tiofenol, cuyo atomo de azufre esta unido al C del grupo ceto.
Especialmente en la primera variante, especialmente para la incorporacion por el dominio 3W, el precursor de la cadena occidental puede tener la siguiente estructura: Rw-CH2-CH2-CH2-C(O)-X. Especialmente en la segunda variante, especialmente para la incorporacion por el dominio 4W, el precursor de la cadena occidental puede tener la siguiente estructura: Rw-CH2-CH2-C(CH3)=CH2-C(O)-X. Especialmente en la tercera variante, el precursor de la cadena occidental puede tener la siguiente estructura: Rw-CH2-CH2-C(CH3)=CH-CH=C(CH3)-C(O)-X. En cada variante, R puede, p. ej., ser cualquier alquilo, arilo o hidrocarburo aromatico, que contiene cada uno opcionalmente un heteroatomo, o R puede ser H. X es un grupo auxiliar que tiene un atomo de azufre que esta unido al atomo de carbono. Los ejemplos adicionales del precursor de la cadena occidental se proporcionan mas adelante, en donde cada uno de X1-X10 y Rw pueden ser independientemente cualquier heteroatomo, cualquier grupo metino (=CH o -CH-R con R, p. ej., que es un grupo alquilo, arilo o alcohol), un grupo metileno, un grupo carbonilo.
Ejemplos para el precursor de la cadena occidental:
Generalmente, en realizaciones que comprenden la smtesis de la parte de la molecula de la cadena oriental natural, el proceso puede ser opcionalmente en ausencia de MxnH e incluir el suministro de un compuesto precursor metilado. En esta realizacion, el proceso puede comprender el suministro de enzimas que comprenden o consisten en Mxnl y MxnJ, preferiblemente adicionalmente MxnA, para la smtesis de la parte de la molecula de la cadena oriental y el suministro de al menos una parte de las enzimas para la smtesis de la parte de la molecula de la cadena occidental, opcionalmente con el suministro de un precursor de la cadena occidental y de la enzima MxnB. El compuesto precursor metilado puede, p. ej., seleccionarse de los siguientes compuestos: A metilcarbonato (R1-O-C(O)-OCH3), especialmente un metilcarbonato de alquilo o arilo (R1 es un alquilo o arilo), p. ej., C1 a C12, especialmente C6, alternativamente R1-S-C(O)-OCH3.
En la segunda realizacion, una parte del conjunto completo de enzimas sinteticas se proporciona, p. ej., en un microorganismo, parte del conjunto completo de enzimas sinteticas que cataliza la condensacion del anillo pirona. La parte puede comprender o consistir en MxnB, preferiblemente MxnA-MxnB. En esta realizacion, se proporciona un precursor de la cadena oriental, y se proporciona al menos una parte de Mxnl-MxnJ, parte que cataliza la conversion de un precursor de la cadena oriental anadido con un precursor de la cadena oriental convertido y se proporcionan MxnA, opcionalmente MxnA-MxnB, preferiblemente MxnK, y opcionalmente MxnC-MxnD-MxnE-MxnF-MxnG. Esto permite la produccion de compuestos, en los que el precursor de la cadena oriental convertido se condensa con la parte de la molecula de la cadena occidental natural o con un precursor de la cadena occidental convertido.
Las enzimas que se proporcionan pueden, p. ej., comprender o consistir en MxnA-MxnB, y en una primera variante los modulos 1E-6E de Mxnl-MxnJ, en una segunda variante los modulos 2E-6E de Mxnl-MxnJ, en una tercera variante los modulos 4E a 6E o Mxnl-MxnJ, o en una cuarta variante el modulo 6E de MxnJ. En combinacion con el suministro de un precursor de la cadena occidental cuando al menos un dominio catalftico o un modulo de MxnK se inactiva como se ha descrito anteriormente, ademas opcionalmente en combinacion con MxnC-MxnD-MxnE-MxnF-MxnG.
En una primera variante, Mxnl-MxnJ que consiste en los modulos 1E-6E puede proporcionarse, p. ej., inactivando o delecionando al menos el dominio de carga (LM, los aminoacidos No. 1 a 530, preferiblemente aminoacidos los No. 1 a 466, en SEQ ID NO: 9 (Mxnl)), en una segunda variante, Mxnl-MxnJ que consiste en los modulos 2E-6E puede proporcionarse inactivando al menos el modulo 1E (los aminoacidos No. 1 a 1781, preferiblemente los aminoacidos No. 1 a 1759, en SEQ lD NO: 9 (Mxnl)), en una tercera variante, MxnJ que consiste en los modulos 4E-6E puede proporcionarse inactivando al menos el modulo 3E, p. ej., delecionando o inactivando Mxnl (SEQ lD NO: 9), y opcionalmente los aminoacidos No. 1 a 405, preferiblemente No. 1 a 450, de SEQ lD NO: 10, opcionalmente Mxnl (SEQ lD NO: 9), y ademas opcionalmente la parte de MxnJ comprendida por los aminoacidos No. 1 a 405, en SEQ lD NO: 10) puede estar funcionalmente ausente, p. ej., estar delecionada o inactivada, en una cuarta variante, Mxnl-MxnJ que consiste en los modulos 5E y 6E puede proporcionarse, p. ej., inactivando al menos el modulo 4E. En la tercera y cuarta variante, Mxnl (SEQ lD NO: 9) puede estar funcionalmente ausente, p. ej., estar mutada, especialmente delecionada, dando lugar a que el dominio de carga del modulo 4E este funcionalmente ausente, opcionalmente parte de MxnJ, p. ej., los aminoacidos No. 1 a 3052, preferiblemente Mxnl completa y la parte de MxnJ comprendida por los aminoacidos No. 1 a 3006 en SEQ lD NO: 10 puede estar funcionalmente ausente, p. ej., estar delecionada o inactivada. Preferiblemente, se proporciona una enzima que comprende o consiste en el modulo 6E (los aminoacidos No. 3007 a 3849, preferiblemente los aminoacidos No. 3053 a 3849 en SEQ lD NO: 10) de MxnJ para la union del precursor de la cadena oriental, opcionalmente en combinacion con el suministro de MxnA.
En la tercera realizacion, el precursor de la cadena oriental puede tener la estructura Re-O-C(O)-X. Especialmente en la segunda variante, para la incorporacion por el modulo 2E, el precursor de la cadena oriental puede tener la siguiente estructura: Re-O-C(O)-NH-CH2-C(O)-X. Especialmente en la tercera variante, para la incorporacion por el modulo 4E, el precursor de la cadena oriental puede tener la siguiente estructura: Re-O-C(O)-NH-CH=CH-CH2-C(O)-X. Especialmente en la cuarta variante, para la incorporacion por el modulo 6E, el precursor de la cadena oriental puede tener la siguiente estructura: Re-O-C(O)-NH-CH=CH-CH2-CH2-CH(CH3)-C(O)-X. En cada variante, Re puede, p. ej., ser cualquier alquilo, arilo o hidrocarburo aromatico, que contiene cada uno opcionalmente un heteroatomo, o Re puede ser H. X es un grupo auxiliar que tiene un atomo de azufre que esta unido al atomo de carbono.
En la primera y segunda realizaciones, el conjunto de enzimas sinteticas puede proporcionarse por manipulacion genetica en un microorganismo original productor de mixopironina mediante la inactivacion o delecion de los genes que codifican enzimas sinteticas, modulos o dominios de las mismas, que estan en exceso de los genes que codifican la parte del conjunto natural de las enzimas usadas en el proceso. De acuerdo con esto, la parte del conjunto natural de enzimas usadas en el proceso de la invencion forma las enzimas sinteticas.
De acuerdo con esto, en la primera realizacion del proceso para producir una mixopironina o un derivado, las etapas naturales para la smtesis de la parte de la molecula de la cadena occidental, p. ej., el suministro de las enzimas que catalizan la parte de la molecula de la cadena occidental, puede reemplazarse proporcionando un precursor de la cadena occidental y preferiblemente al menos una parte de las enzimas que catalizan la smtesis de la parte de la molecula de la cadena occidental, p. ej., una parte de los modulos o dominios de MxnK. En esta realizacion, el
precursor no esta necesariamente unido a la enzima sintetica, pero el precursor puede tener un grupo auxiliar, preferiblemente unido mediante un atomo de azufre del grupo auxiliar, p. ej., el precursor puede estar unido al atomo de azufre de un tioalquilo o un tioarilo que puede estar sustituido ademas con heteroatomos, p. ej., con un grupo amina, una alquilamina, una alquilamida, p. ej., al atomo de azufre de una amida del acido W-2-tioalquil carbonico, en donde la cadena de carbono del alquilo y del resto acido puede ser un grupo C1 a C6 lineal o ramificado, especialmente W-2-tioetilacetamida. Como un ejemplo para un tioarilo, el precursor puede estar unido al atomo de azufre de un grupo tiofenol.
En la segunda realizacion del proceso para producir una mixopironina o un derivado, las etapas naturales para la smtesis de la parte de la molecula de la cadena oriental, p. ej., el suministro de las enzimas que catalizan la parte de la molecula de la cadena oriental se reemplaza proporcionando un precursor de la cadena oriental y preferiblemente una enzima que comprende o consiste en el modulo 6E de Mxnl-MxnJ, los modulos 4E-6E de Mxnl-MxnJ, los modulos 2E-6E de Mxnl-MxnJ, o los modulos 1E-6E de Mxnl-MxnJ.
Para los propositos de esta invencion, la parte de la molecula de la cadena occidental y la parte de la molecula de la cadena oriental, respectivamente, se sintetizan por el microorganismo huesped, mientras el precursor de la cadena occidental y el precursor de la cadena oriental, respectivamente, son compuestos que se anaden al cultivo del microorganismo. El precursor de la cadena occidental y el precursor de la cadena oriental pueden producirse sinteticamente.
El precursor preferiblemente contiene o consiste en R1-(C=O)-CH2-(C=O)- que esta unido al S del grupo auxiliar X, en donde preferiblemente el R1 es R2(CH3)C- o R2(CH3CH2)C-, con R2 p. ej., seleccionado de un alquilo o arilo C1 a C12 ramificado o lineal o un compuesto aromatico, p. ej., que contiene uno dos enlaces dobles, preferiblemente dos enlaces dobles conjugados, y opcionalmente ademas sustituido con un arilo o un grupo que contiene heteroatomo, p. ej., un grupo fenilo, hidroxifenilo, hidroxilo o amina, preferiblemente en el extremo. R1 puede ser Rw para el precursor de la cadena occidental y Re para el precursor de la cadena oriental, respectivamente. Generalmente, el precursor puede tener la siguiente estructura

en donde X es el grupo auxiliar, p. ej.,

En realizaciones en las que se proporcionan tanto un precursor de la cadena occidental como un precursor de la cadena oriental, el proceso comprende el suministro de la enzima MxnB o CorB, opcionalmente de MxnA-MxnB o CorA-CorB, ya que se encontro que MxnB es suficiente para catalizar la condensacion de ambos precursores en el resto del anillo pirona que porta los sustituyentes del grupo (C=O)-CH2-(CO) de cada uno del precursor de la cadena occidental y de la cadena oriental, preferiblemente del modulo 6E de MxnJ, especialmente para la union del precursor de la cadena oriental a la enzima, y preferiblemente para introducir un grupo intermedio en el precursor de la cadena oriental (etapa de elongacion introduciendo el grupo -CH2-CH2-), respectivamente.
De acuerdo con esto, MxnC-MxnM, o CorC-ORF1, respectivamente, incluyendo las enzimas intermedias proporcionadas en la Tabla 1 o los modulos o dominios de las mismas, pueden estar ausentes o estar inactivadas en esta realizacion. Preferiblemente, ademas del suministro de la enzima MxnB, preferiblemente de MxnA-MxnB, el proceso incluye el suministro de la enzima MxnL, MxnM o MxnM-MxnL. En esta realizacion, al menos una parte de las enzimas MxnK y MxnJ estan ausentes o inactivas, evitando la smtesis de la parte de la molecula de la cadena occidental y de la parte de la molecula de la cadena oriental, y las enzimas MxnC-MxnG y/o MxnH pueden estar opcionalmente presentes o ausentes. La adicion de un precursor de la cadena occidental y de un precursor de la cadena oriental da lugar a la produccion de derivados de mixopironina. Esquematicamente, esta realizacion en la que tanto MxnK como Mxnl estan inactivadas, puede representarse por

en donde la enzima preferiblemente es MxnB y X es un grupo auxiliar que esta unido por un S (atomo de azufre). El grupo auxiliar X es p. ej., SNAC o tiofenol. De forma similar, el producto se obtuvo cuando tanto MxnK como MxnJ se inactivaron, de nuevo con las demas enzimas opcionalmente activas.
En realizaciones para producir una mixopironina o un derivado de mixopironina que tiene, p. ej., la cadena oriental natural o una cadena oriental derivada de un precursor de la cadena oriental, el proceso puede comprender el suministro de un precursor de la cadena occidental y el suministro de la enzima MxnB, preferiblemente de MxnA-MxnB, y de las enzimas Mxnl-MxnJ, opcionalmente ademas MxnL y/o MxnM con el fin de proporcionar las enzimas de la ruta sintetica para la smtesis de la parte de la molecula de la cadena oriental en el microorganismo huesped. En la alternativa a MxnB, puede proporcionarse CorB, y en la alternativa a MxnA-MxnB, puede proporcionarse CorA-CorB, en la alternativa a Mxnl-MxnJ, puede proporcionarse Corl-CorJ, en la alternativa a MxnL y/o MxnM, puede proporcionarse ORF1. En esta realizacion, la enzima MxnK esta ausente o inactiva, evitando la smtesis de la parte de la molecula de la cadena occidental, y las enzimas MxnC-MxnG y/o MxnH pueden opcionalmente proporcionarse o estar ausentes. Opcionalmente, el precursor de la cadena oriental puede ser un metilcarbonato, preferiblemente activado por la union al S de un grupo auxiliar. En presencia de la enzima MxnH, el precursor de la cadena oriental se metila por MxnH.
Esquematicamente, esta realizacion puede representarse por

En realizaciones para producir una mixopironina o un derivado de mixopironina que tiene, p. ej., la cadena occidental natural o una cadena occidental derivada de un precursor de la cadena occidental, el proceso puede comprender el suministro de un precursor de la cadena oriental y el suministro de las enzimas MxnA-MxnB, alternativamente CorA-CorB, preferiblemente en combinacion con un modulo de elongacion C-terminal de MxnJ, p. ej., el modulo 6E de MxnJ, alternativamente un modulo de elongacion C-terminal de CorJ, y de la enzima MxnK, opcionalmente ademas MxnL y/o MxnM, alternativamente CorK-CorL, opcionalmente con ORF1, para la smtesis de la parte de la molecula de la cadena occidental en el microorganismo huesped. En esta realizacion, las enzimas Mxnl-MxnJ estan ausentes o inactivas, evitando la smtesis de la parte de la molecula de la cadena oriental, preferiblemente se proporciona una enzima que comprende o consiste en el modulo 6E de MxnJ, y las enzimas MxnC-MxnG y/o MxnH pueden opcionalmente proporcionarse o estar ausentes.
Esquematicamente, esta realizacion puede representarse por

Para el precursor de la cadena occidental, R2 puede, p. ej., ser una cadena alquilo ramificada o lineal, que tiene opcionalmente uno o dos enlaces dobles, y/o ademas opcionalmente esta unida a un arilo, p. ej., a un anillo fenilo, que esta sustituido opcionalmente con un grupo hidroxilo o amina.
Para el precursor de la cadena oriental, R2 preferiblemente es un alquilo o arilo C1 a C24, p. ej., C1 a C12 ramificado o lineal, que contiene opcionalmente uno o dos enlaces dobles, preferiblemente una cadena C4 lineal, que porta opcionalmente un grupo amina o amida terminal, p. ej., un grupo metoxi amida.
Preferiblemente, el precursor de la cadena occidental y/o el precursor de la cadena oriental puede tener la siguiente estructura:

en donde solo se muestra el atomo de azufre (S) del grupo auxiliar X.
El proceso puede proporcionar una parte de o todas las enzimas que catalizan la smtesis de la parte de la molecula de la cadena oriental, y proporcionar solo un precursor para la cadena occidental. En esta realizacion, la cadena oriental del producto corresponde a la cadena oriental natural de la mixopironina y la cadena occidental corresponde al precursor de la cadena occidental que esta unido a la molecula auxiliar.
Alternativamente o adicionalmente, el proceso puede proporcionar una parte de o todas las enzimas que catalizan la smtesis de la parte de la molecula de la cadena occidental, y proporcionar solo un precursor para la cadena oriental. En esta realizacion, la cadena occidental del producto se produce por el microorganismo y corresponde, p. ej., a la cadena occidental natural de la mixopironina, p. ej., cuando se proporcionan las enzimas MxnA-MxnK o a un derivado de la cadena occidental natural, p. ej., cuando MxnC-MxnG y/o MxnH estan funcionalmente ausentes del microorganismo, y la cadena oriental corresponde a la del precursor de la cadena oriental que esta unido a la molecula auxiliar con el anillo pirona producido por MxnB.
En una realizacion espedfica, el proceso proporciona todas las enzimas de la ruta sintetica que constituyen la ruta biosintetica natural de la smtesis de la mixopironina, de la que se inactivan genes opcionalmente, p. ej., genes que codifican MxnC-MxnG y/o MxnH y/o MxnL y/o MxnM se delecionan, dando lugar, p. ej., a una alteracion del producto sintetizado. En esta realizacion, las enzimas de la ruta se expresan en un microorganismo, y la mixopironina se sintetiza enteramente por el microorganismo. De acuerdo con esto, en esta realizacion, el proceso excluye el suministro de compuestos precursores espedficos.
A partir del analisis de la produccion de mixopironina o derivados de M. fulvus en el que se habfan inactivado genes espedficos, se encuentra que la produccion de mixopironina se produjo cuando todas las enzimas MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnK-MxnL-MxM estaban presentes.
Cuando se inactivaron las enzimas, se encontraron los siguientes efectos en la produccion de mixopironina:
- que la inactivacion de MxnC-MxnD-MxnE-MxnF-MxnG con MxnA-MxnB y MxnH-MxnI-MxnJ-MxnM-MxnL-MxnM activa dio lugar a la produccion del derivado de mixopironina 21-desmetil-mixopironina A,

- que la inactivacion de MxnH solo dio lugar a ausencia de produccion,
- que la inactivacion de MxnC-MxnG y la inactivacion de MxnH dieron lugar a ausencia de produccion,
- que la inactivacion de MxnA solo dio lugar a ausencia de produccion de mixopironina,
- que la inactivacion de MxnB solo dio lugar a ausencia de produccion de mixopironina,
- que la inactivacion de MxnL solo dio lugar a ausencia de produccion de mixopironina A y B,
- que la inactivacion de MxnM solo dio lugar a la produccion de mixopironina A y B,
- la inactivacion de Mxnl solo, es decir, MxnA-MxnH y MxnJ-MxnM activas con la adicion del precursor de la cadena oriental que contiene Re dio lugar a la produccion de

Derivado de mixopironina A: R = H
Derivado de mixopironina B: R = CH3,
- que la inactivacion tanto de MxnK como de Mxnl solo dio lugar a ausencia de produccion de mixopironina, y - que la inactivacion de MxnK y MxnJ dio lugar a ausencia de produccion de mixopironina.
Cuando se alimenta un precursor de la cadena occidental y/u oriental, se encontro la siguiente produccion para las enzimas inactivadas, en donde las enzimas se indican por sus secuencias codificadoras respectivas y los precursores se indican, proporcionandose los productos para cada fila en la columna de la derecha. En la siguiente tabla, las enzimas conectadas por un guion (-) indican las enzimas indicadas incluyendo las enzimas intermedias.






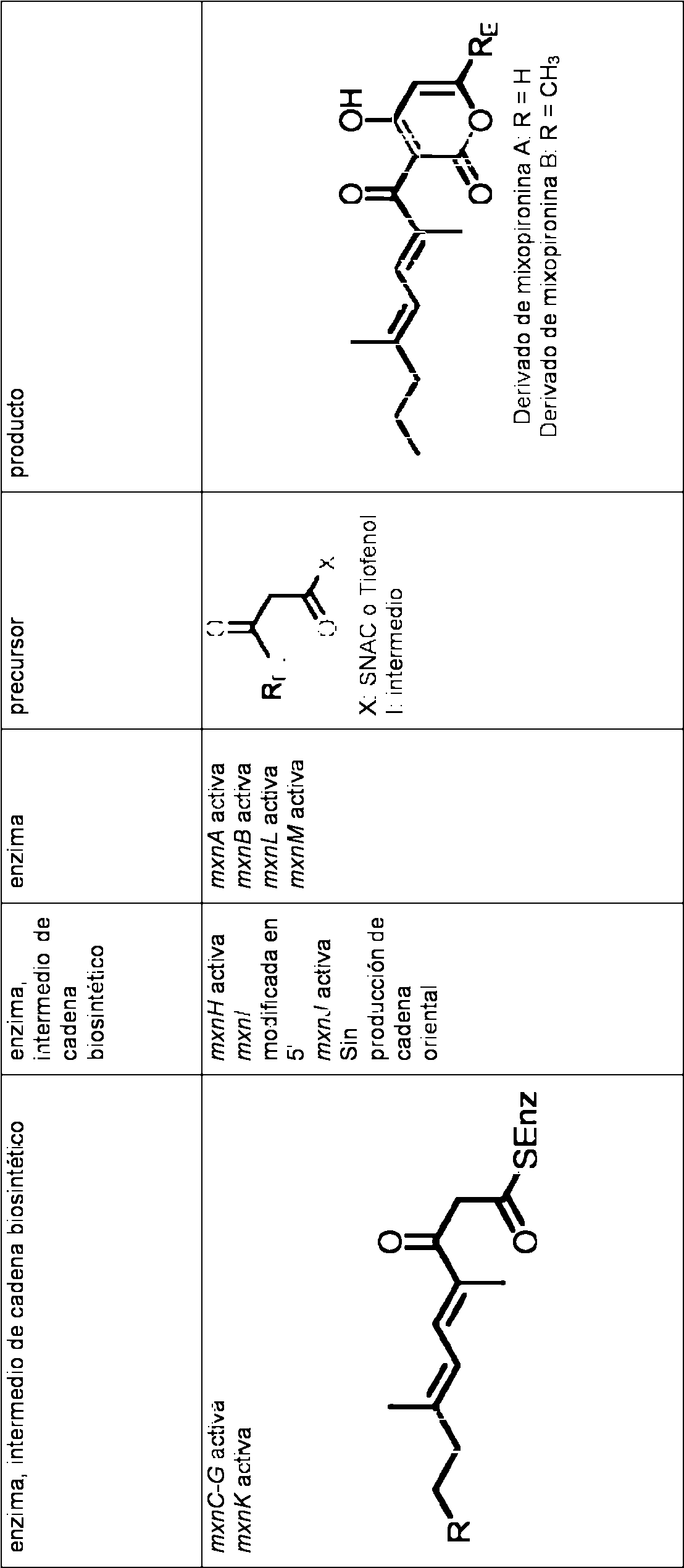
Estos resultados muestran que un proceso para la produccion de mixopironina puede comprender la expresion de MxnA-MxnK, con la expresion de MxnL-MxnM como una opcion al menos en Myxococcus (M. fulvus Mx f50), ya que la delecion de MxnL-MxnM no evito la smtesis de mixopironina en M. fulvus.
Un proceso para la produccion de mixopironina usando la smtesis de la parte de la molecula de la cadena occidental por la expresion de MxnK, pero sin la expresion de MxnC-MxnG, da lugar a derivados de mixopironina que no tienen grupo metilo en C21 de la cadena occidental. Opcionalmente, la produccion de mixopironina usando biosmtesis de la parte de la molecula de la cadena oriental por la expresion de Mxnl-MxnJ, y/o usando la biosmtesis de la parte de la molecula de la cadena occidental por la expresion de al menos una parte de MxnK, opcionalmente en presencia o ausencia de MxnC-MxnG, puede hacerse opcionalmente sin la expresion de MxnH, preferiblemente en presencia de MxnH con la adicion del precursor en la forma de un monoalquil ester de acido de carbono, en donde el acido de carbono contiene un alquilo C1 a C12 lineal o ramificado unido al grupo acido, y en donde el alquilo, p. ej., es un C1 a C12 lineal o ramificado, preferiblemente un monometil ester de acido de carbono. El monoalquil ester de acido de carbono en esta realizacion esta integrado en la cadena oriental.
Generalmente, la enzima o enzimas que catalizan la smtesis pueden proporcionarse por la expresion a partir de una secuencia de acido nucleico que codifica la enzima y enzimas, resp, en donde la secuencia de acido nucleico esta contenida en al menos un casete de expresion. La secuencia de acido nucleico puede codificar la enzima o enzimas por el uso de codones del microorganismo huesped. Como un microorganismo huesped, se prefieren las bacterias, mas preferiblemente las bacterias Gram-negativas, p. ej., Pseudomonas spp, Myxococcus spp. Generalmente en el proceso, las enzimas pueden proporcionarse in vivo por el microorganismo o pueden proporcionarse in vitro, p. ej., producirse y obtenerse por el microorganismo, p. ej., extraerse o aislarse del microorganismo.
Generalmente, la incubacion del microorganismo puede comprender cultivar el microorganismo en medio de cultivo bajo agitacion y/o aeracion.
La invencion se describe ahora con mayor detalle mediante los ejemplos con referencia a las figuras, que muestran en
- la Figura 1 un esquema de la organizacion natural de los genes sinteticos en Myxococcus fulvus Mx f50 (mxnA a mxnM),
- la Figura 2 en A) un esquema de la smtesis natural de la parte de la molecula oriental, en B) un esquema de la smtesis natural de la parte de la molecula occidental y en C) un esquema del mecanismo de la condensacion de las partes de la molecula al anillo pirona,
- las Figuras 3 y 4 esquemas para la subclonacion de enzimas de la ruta,
- la Figura 5 un esquema para la clonacion de una construccion de expresion para expresar las enzimas de la ruta sintetica a partir de un casete de expresion,
- la Figura 6 un esquema para clonar secuencias codificadoras de enzimas en casetes de expresion separados, - la Figura 7 resultados de HPLC-MS de extractos de Myxococcus xanthus DK 1622 AmchA::tet no productor, del transformante M. xanthus DK1622 AmchA-tet:: pHSU-mxn43 (M. xanthus DK1622 AmchA-tet::mixopironina A), y de Myxococcus fulvus Mx f50 de tipo salvaje (WT),
- la Figura 8 un esquema para la clonacion para inactivar la enzima Mxnl,
- la Figura 9 un esquema para la clonacion para inactivar la enzima MxnJ,
- la Figura 10 un esquema para la clonacion para inactivar la enzima MxnK,
- la Figura 11 un esquema para la clonacion para inactivar la enzima MxnK,
- la Figura 12 un esquema para la clonacion para inactivar la enzima MxnH,
- la Figura 13 un esquema para la clonacion para inactivar las enzimas MxnC-MxnD-MxnE-MxnG-MxnH,
- la Figura 14 un esquema para la clonacion para inactivar la enzima MxnC-MxnD-MxnE-MxnG,
- la Figura 15 resultados de HPLC-MS de extractos de cultivos de microorganismos en los que al menos una enzima de la ruta sintetica de la mixopironina A esta inactivada,
- la Figura 16 un esquema para la clonacion para inactivar la enzima MxnL,
- la Figura 17 un esquema para la clonacion para inactivar la enzima MxnM,
- la Figura 18 resultados analfticos para la produccion de la mixopironina A durante el cultivo de a) Myxococcus fulvus Mx f50, b) Myxococcus fulvus Mx f50 que tiene MxnL inactivada, c) Myxococcus fulvus Mx f50 que tiene MxnM inactivada (AmxnM),
- la Figura 19 un esquema del efecto de la inactivacion (X) de KR del modulo 2E (KR-2E) en la smtesis,
- la Figura 20 un esquema del efecto de la inactivacion (X) de DH del modulo 2E en la smtesis,
- la Figura 21 un esquema del efecto de la inactivacion (X) de MT del modulo 4E en la smtesis,
- la Figura 22 resultados analfticos de HPLC-MS para la smtesis in vitro usando precursores de la cadena occidental y de la cadena oriental,
- la Figura 23 espectros MS2 del producto en extractos de sobrenadantes de cultivo,
- la Figura 24 un esquema de clonacion para la inactivacion de la secuencia de acido nucleico que codifica el dominio catalftico CP del modulo 1W de la enzima MxnK,
- la Figura 25 un esquema de clonacion para la inactivacion de la secuencia de acido nucleico que codifica el dominio catalftico CP del modulo 4W de la enzima MxnK,
- la Figura 26 un esquema en a) de la smtesis de tipo salvaje de la parte de la molecula occidental por MxnK, en b) la conversion de un precursor anadido de la cadena occidental por MxnK que tiene el dominio catalftico CP del modulo 1W inactivado, en c) la conversion de un precursor anadido de la cadena occidental por MxnK que tiene el dominio catalftico CP del modulo 3W inactivado, en d) la conversion de un precursor anadido de la cadena occidental por MxnK que tiene el dominio catalftico CP del modulo 4W inactivado, indicando los cftculos oscuros dominios activos, mientras los cftculos blancos indican dominios inactivos,
- la Figura 27 un esquema en a) de la smtesis de tipo salvaje de la parte de la molecula oriental por Mxnl-MxnJ, en b) la conversion de un precursor anadido de la cadena oriental por Mxnl-MxnJ que tiene el dominio catalftico CP del modulo de carga (carga) inactivado, en c) la conversion de un precursor anadido de la cadena oriental por Mxnl-MxnJ que tiene el dominio catalftico CP del modulo 1E inactivado, en d) la conversion de un precursor anadido de la cadena oriental por Mxnl-MxnJ que tiene el dominio catalftico CP del modulo 3E inactivado, en e) la conversion de un precursor anadido de la cadena oriental por Mxnl-MxnJ que tiene el dominio catalftico CP del modulo 4E inactivado, indicando los cftculos oscuros dominios activos, mientras los cftculos blancos indican dominios inactivos,
- la Figura 28 muestra esquematicamente la conversion de un precursor anadido de la cadena occidental por MxnK que tiene el dominio catalftico CP del modulo 2W inactivado y la smtesis de la parte de la molecula oriental por Mxnl-MxnJ que tiene el dominio catalftico MT del modulo 4E inactivado, seguido de la condensacion de la parte de la molecula oriental alterada con el precursor convertido de la cadena occidental por MxnB, indicando los cftculos oscuros dominios activos, mientras los cftculos blancos indican dominios inactivos, y
- la Figura 29 muestra esquematicamente la smtesis de la parte de la molecula occidental por MxnK que tiene el dominio catalftico MT del modulo 4W inactivado y la conversion de un precursor anadido de la cadena oriental por Mxnl-MxnJ que tiene el dominio catalftico CP del modulo 1E inactivado, seguido de la condensacion de la parte de la molecula occidental alterada con el precursor convertido de la cadena oriental por MxnB, indicando los cftculos oscuros dominios activos, mientras los cftculos blancos indican dominios inactivos.
Generalmente, en las Figuras que representan genes que codifican enzimas o enzimas, en donde, p. ej., se indican los modulos, dominios o centros catalfticos, en combinacion con la smtesis de mixopironina o de un derivado, la secuencia de las etapas catalfticas es de izquierda a derecha, correspondiendo a la direccion desde el extremo N al extremo C de las protemas indicadas.
Como se muestra en la Figura 1, los genes que codifican las enzimas para la smtesis de mixopironina en Myxococcus estan organizados adyacentes uno de otro. En las figuras, * indica dominios catalfticos que pueden no tener enteramente la funcion catalftica indicada debido a alteraciones, p. ej., de dominios catalfticos conocidos. La Figura 2 A) muestra esquematicamente los dominios catalfticos de las enzimas codificadas por los genes respectivos que participan en la smtesis de la mixopironina. A partir del analisis, se asume que Mxnl proporciona un dominio fosfomutasa (PM) N-terminal que podna estar implicado en la activacion de biscarbonato de sodio como una unidad de partida por la transferencia del grupo fosforilo para permitir la union posterior al dominio CP del modulo de carga.
La smtesis de la parte de la molecula de la cadena oriental comprende la elongacion de la unidad de partida con glicina y tres unidades extensoras de malonil-CoA por una policetido sintetasa/peptido no ribosomal megasintetasa que consiste en dos subunidades (enzimas Mxnl y MxnJ) y que porta un total de seis modulos de elongacion (1E-6E) ademas del modulo de carga (PM-CP) indicado en el extremo N del modulo 1E de Mxnl. Mxnl-MxnJ son PKS/NPRS
(policetido sintetasa/peptido no ribosomal sintasa) que comprende un modulo de carga y seis modulos de extension. El modulo 1E (KS-A-CP) cataliza la incorporacion de glicina, que se elonga entonces por el modulo 2E (KS-DH-KR-CP) con malonil-CoA. El modulo 3E representa un modulo dividido (KS de Mxnl y DH*-CP de MxnJ), que no se asume que incorpore una unidad extensora, pero en donde el dominio de deshidrogenasa DH podna catalizar el desplazamiento de un enlace doble de la posicion a, p a la posicion p,Y. El modulo 4E (KS-DH-KR-MT-CP) incorpora otra unidad extensora malonil-CoA, que se metila por el dominio MT. La reduccion completa de la funcionalidad pcarbonilo se asume que esta catalizada por actividades DH-KR internas mas la actividad ER de MxnA, que se proporciona en trans. El dominio cetosintasa del modulo 5E (KS*-CP) posiblemente no cataliza la elongacion de la cadena, de manera que no puede incorporarse ninguna unidad extensora a partir de este modulo. Una tercera unidad extensora malonil-CoA se incorpora por el modulo 6E (KS-CP*-CP), en el que uno de los dominios CP en tandem se asume que es inactivo, para generar el intermedio final de la cadena oriental con una funcionalidad p-carbonilo, que es esencial para la condensacion de la cadena para la formacion del anillo pirona. Para las tres etapas de elongacion con malonil-CoA (por los modulos 2E, 4E y 6E), se requiere la actividad AT en trans de MxnA. MxnM podna tener una actividad de edicion/correccion para eliminar las unidades acilo aberrantes de los modulos Mxnl/MxnJ bloqueados, y opcionalmente tambien de los modulos MxnK bloqueados.
Como se muestra en la Fig. 2 B), la parte de la molecula de la cadena occidental esta catalizada por MxnK. Se asume que el inicio es con acetil-CoA para la mixopironina A o con propionil-CoA para la mixopironina B, que se carga directamente en el dominio KS N-terminal del modulo 1W; no se encontro un modulo de carga tfpico. Despues de la elongacion con la primera unidad de malonil-CoA por el modulo 1W (KS-DH-KR-CP), el compuesto p-ceto resultante se reduce completamente por las actividades DH-KR internas mas la actividad ER en trans de MxnA. A partir del analisis, se asume que el dominio KS* del modulo 2W, KS*-CP, es inactivo y de acuerdo con esto el modulo 2W no esta implicado en la elongacion de la cadena. Los modulos posteriores 3W, 4W, 5W se asume que catalizan la incorporacion de tres unidades extensoras malonil-CoA adicionales para rendir la parte de la molecula de la cadena occidental. La elongacion de la cadena por el modulo 3W incluye ramificacion en p para introducir un grupo metilo en la posicion p, catalizado por MxnC-MxnG. La elongacion de la cadena en el modulo 4W incluye una reduccion completa de la funcionalidad p-carbonilo por las actividades DH-KR internas mas la actividad ER en trans de MxnA. De manera similar a la biosmtesis de la cadena oriental, la actividad AT en trans de MxnA se requiere para las cuatro etapas de elongacion con malonil-CoA por los modulos 1W, 3W, 4W y 5W. MxnM podna tener una actividad de editado/correccion para eliminar las unidades acilo aberrantes de los modulos MxnK bloqueados, y tambien de los modulos Mxnl/MxnJ bloqueados.
La Fig. 2 C) muestra la condensacion de la parte de la molecula de la cadena occidental unida a enzima y la parte de la molecula de la cadena oriental unida a enzima al anillo pirona que esta sustituido con Rw y Re, como se proporciona por las enzimas biosinteticas naturales.
Las siguientes secuencias de acido nucleico se usaron como cebadores:





Ejemplo comparativo: Produccion de mixopironina por la expresion heterologa de enzimas de la ruta sintetica
Para la expresion heterologa, los genes que codifican las enzimas de la ruta sintetica para la smtesis de mixopironina se aislaron del ADN genomico de Myxococcus fulvus Mxf50 y se subclonaron para generar construcciones de expresion que se transfirieron a microorganismos huesped, preferiblemente un huesped heterologo, p. ej., por electroporacion. Para la integracion de las construcciones de expresion en el genoma del microorganismo huesped, se construyeron plasmidos suicidas como se describe mas adelante.
En detalle, se construyeron tres plasmidos suicidas diferentes para la integracion en diferentes sitios de la region de la ruta de la mixopironina: (1) El promotor Tn5 se amplifico por PCR usando los cebadores mxn33/ mxn41 con el vector pCRII-TOPO como un molde para obtener el fragmento 1. Un fragmento de 1191 pb de mxnA se amplifico a partir del ADN genomico de M. fulvus Mxf50 por PCR usando los cebadores mxn42/mxn30 para obtener el fragmento 2. El fragmento 1 y el fragmento 2 se unieron entre sf usando PCR de superposicion con los cebadores mxn33/mxn30. Los fragmentos unidos se subclonaron entonces en un vector pJET1,2 para obtener el plasmido pTn5-mxnA/pJET. En paralelo, se obtuvo un vector de clonacion lineal mmimo p15A-Tet por PCR usando los cebadores mxn52/mxn53 con pACYC184 como un molde. El plasmido pTn5-mxnA/pJET se hidrolizo con PacI/EcoRV y se ligo en el vector de clonacion hidrolizado con PacI/PvuII p15A-Tet para generar pHSU-mxn16. (2) Un fragmento de 1491 pbjusto despues del extremo 3' de la agrupacion de genes de la mixopironina A se amplifico a partir del ADN genomico de M. fulvus Mxf50 usando los cebadores mxn64/mxn65. El fragmento amplificado se clono en el vector pCRII-TOPO para crear pHSU-mxn19. (3) Un fragmento de 1380 pb de mxnJ se amplifico a partir del ADN genomico de M. fulvus Mxf50 usando los cebadores mxn124 y mxn125. Un casete de resistencia a zeocina se amplifico a partir de pCDNA-Zeo como el molde usando los cebadores mxn128/mxn129 y se subclono en un vector de clonacion lineal mmimo p15A-Tet obtenido por PCR usando los cebadores mxn126/mxn127 con pACYC184 como un molde para obtener el plasmido p15A-Tet-Zeo. El fragmento de 1380 pb de mxnJ se hidrolizo entonces con Pvul y se clono posteriormente en p15A-Tet-Zeo hidrolizado con la misma enzima de restriccion para crear pHSU-mxn32.
En primer lugar, pHSU-mxn16 se transformo en M. fulvus Mx f50 para obtener Mx f50::pHSU-mxn16 mutante. Para el analisis genotfpico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn60/p15A-Tet1 y mxn61/p15A-Tet2 para verificar la integracion correcta del plasmido de inactivacion. Posteriormente, el siguiente unico entrecruzamiento se realizo transformando pHSU-mxn19 en Mx f50::pHSU-mxn16 para obtener Mx f50::pHSU-mxn16::pHSU-mxn19 como se representa en la Fig. 3. El mutante se confirmo por PCR usando las combinaciones de cebadores mxn94/pTOPO-out y mxn95/pTOPO-in. Ademas, la confirmacion de este mutante tambien se realizo por analisis de Transferencia Southern. En segundo lugar, pHSU-mxn32 se transformo en M. fulvus Mxf50 para obtener Mx f50::pHSU-mxn32 mutante como se representa en la Fig. 4. Para el analisis genotfpico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn136/p15A-Tet2 y p15A-Tet3/mxn137 para verificar la integracion correcta del plasmido de inactivacion.
La recuperacion de los fragmentos de la ruta se realizo como sigue: el ADN genomico de Mx f50::pHSU-mxn16::pHSU-mxn19 mutante se hidrolizo con EcoRV y posteriormente se volvio a ligar para generar pHSU-mxn26 (Fig. 3). Independientemente, tambien se hidrolizo con Pacl/Pvul y posteriormente se volvio a ligar para generar pHSU-mxn27 (Fig. 3). El ADN genomico de Mx f50::pHSU-mxn32 mutante se hidrolizo con Pvul y despues se volvio a ligar para generar pHSU-mxn35 (Fig. 4). Un casete de ampicilina se amplifico por PCR usando los cebadores mxn147/mxn148 con el vector pJET1,2 como el molde. pHSU-mxn35 se hidrolizo con Aflll/Pacl para reemplazar el casete de zeocina con el casete de ampicilina amplificado hidrolizado con el mismo conjunto de enzimas de restriccion para generar pHSU-mxn40 (Fig. 4).
El metodo de clonacion para la generacion de una construccion de expresion que porta la ruta completa se muestra en la Fig. 5. En primer lugar, la ruta de biosmtesis de la cadena occidental se completo por la integracion del fragmento Avrll de 18,5 kb de pHSU-mxn40 en pHSU-mxn27 mediante recombinacion Red/ET. Se realizo otra ronda de recombinacion Red/ET para intercambiar los 6,2 kb del extremo 3' de mxnl en pHSU-mxn26 con un casete de resistencia a cloranfenicol (CmR) para generar pHSU-mxn31. Ambos, pHSU-mxn31 y pHSU-mxn41, se hidrolizaron entonces con Spel/Pacl para ligar el fragmento mxnK-M en pHSU-mxn41 para generar pHSU-mxn42. Se amplifico un casete de resistencia a kanamicina a partir del vector pCRII-TOPO como un molde usando los cebadores mxn144/mxn145. El casete amplificado se subclono entonces en el vector pJET1,2 para obtener pHSU-mxn37. Ambos, pHSU-mxn37 y pHSU-mxn42 se hidrolizaron con PacI para ligar el casete de resistencia a kanamicina en pHSU-mxn42 para generar pHSU-mxn43.
Para la expresion heterologa, la construccion de expresion pHSU-mxn43 descrita anteriormente y mostrada en la Fig. 5 se transformo en el huesped de expresion M. xanthus DK1622 AmchA-tet por el siguiente procedimiento: M. xanthus DK1622 AmchA-tet se crecio en matraces con deflector con medio CTT suplementado con 6,25 pg/ml de oxitetraciclina a 30 °C hasta que se alcanzo una DO600 entre 0,6-0,9. Las celulas se recogieron entonces de 2-4 ml de cultivo (1-2 x 109 celulas/ml) por centrifugacion a 12,500 rpm durante 1 min a temperatura ambiente. Despues de dos etapas de lavado
con 1 ml de H2O a temperatura ambiente, las celulas se resuspendieron en 65 j l de H2O. Se mezclaron 1-2 |jg de ADN plasmndico con la suspension celular, y se llevo a cabo la electroporacion bajo las siguientes condiciones: 25 jF , 400 Q, 650 V usando cubetas de electroporacion de 0,1 cm y un dispositivo GenePulser XCell (Bio-Rad). Se anadio directamente 1 ml de medio CTT a la suspension celular inmediatamente despues de la electroporacion y las celulas se transfirieron a un tubo de centnfuga de 2 ml. Despues de 5 h de cultivo a 30 °C y 850 rpm en un thermomixer, las celulas se mezclaron con 2 ml de agar blando CY (medio CTT con 0,7 % de agar) y se sembraron en placas de agar CTT (medio CTT con 1,5 % de agar) suplementadas con 50 jg/m l de kanamicina. Las placas se incubaron a 30 °C durante 7-10 dfas hasta que las colonias fueron visibles. La seleccion se realizo en placas de agar CTT que conteman 50 jg/m l de kanamicina para aislar la cepa de expresion M. xanthus DK1622 AmchA-tet: pHSU-mxn43.
Alternativamente, para la construccion de construcciones de expresion que portan partes de las enzimas de la ruta sintetica, podna usarse una estrategia de multiples plasmidos. El metodo de clonacion para la generacion de dos construcciones de expresion compatibles que portan dos partes de la ruta se representa esquematicamente en la Fig. 6. En primer lugar, se ligo un marcador de seleccion adecuado, ejemplificado por el gen de resistencia a zeocina, zeoR, en pHSU-mxn41 usando los sitios de restriccion Aflll/PacI para generar la construccion de expresion pHSU-mxn44. En paralelo, la construccion pHSU-mxn31 se modifico por recombinacion Red/ET para insertar un promotor adecuado en 5' de mxnK junto con otro marcador de seleccion, ejemplificado aqu por el casete de resistencia a apramicina (ApraR), que estaba flanqueado por dos sitios de restriccion unicos (R1 y R2) para generar el plasmido pHSU-mxn45. El plasmido se modifico ademas por ligacion de una region de homologfa adecuada mediante los sitios de restriccion R1/R2 para rendir el segundo plasmido de expresion pHSU-mxn46.
Para el metodo con multiples plasmidos, las construcciones de expresion pHSU-mxn44 y pHSU-mxn46 descritas anteriormente se transformaron consecutivamente en M. xanthus DK1622 AmchA-tet segun el siguiente procedimiento. M. xanthus DK1622 AmchA-tet se crecio en matraces con deflector en medio CTT suplementado con 6,25 jg/m l de oxitetraciclina a 30 °C hasta que se alcanzo una DO600 entre 0,6-0,9. Las celulas se recogieron entonces de 2-4 ml de cultivo (1-2 x 109 celulas/ml) por centrifugacion a 12,500 rpm durante 1 min a temperatura ambiente. Despues de dos etapas de lavado con 1 ml de H2O a temperatura ambiente, las celulas se resuspendieron en 65 j l de H2O. Se mezclaron 1-2 jg de ADN plasirndico con la suspension celular, y se llevo a cabo la electroporacion bajo las siguientes condiciones: 25 jF , 400 Q, 650 V usando cubetas de electroporacion de 0,1 cm y un dispositivo GenePulser XCell (Bio-Rad). Se anadio directamente 1 ml de medio CTT a la suspension celular inmediatamente despues de la electroporacion y las celulas se transfirieron a un tubo de centnfuga de 2 ml. Despues de 5 h de cultivo a 30 °C y 850 rpm en un thermomixer, las celulas se mezclaron con 2 ml de agar blando CY (medio CTT con 0,7 % de agar) y se sembraron en placas de agar CTT (medio CTT con 1,5 % de agar) suplementadas con 50 jg/m l de kanamicina. Las placas se incubaron a 30 °C durante 7-10 dfas hasta que las colonias fueron visibles.
La seleccion se realizo en placas de agar CTT suplementadas con 12,5 jg/m l de zeocina para pHSU-mxn44 o 50 jg/m l de kanamicina para pHSU-mxn46. Despues, la segunda construccion de expresion respectiva se transformo en los mutantes para generar las cepas de expresion M. xanthus DK1622 AmchA-tet:: pHSU-mxn44/pHSU-mxn46.
Para la produccion heterologa de mixopironina, M. xanthus DK1622 AmchA-tet:: pHSU-mxn43 y Mxanthus DK1622 AmchA-tet:: pHSU-mxn44/pHSU-mxn46 se cultivaron en medio CTT a 30 °C, p. ej., en matraces con deflector de 300 ml con 50 ml de medio durante 2 dfas a 105 rpm. Despues de la centrifugacion, el sobrenadante se extrajo dos veces con un volumen igual de acetato de etilo, se evaporo el acetato de etilo del extracto, el residuo se disolvio en 1 ml de metanol para proporcionar el extracto. Una parte alfcuota de 5 j l del extracto se analizo por HPLC-MS (Thermo Ultimate 3000 RSLC, acoplado a un instrumento de trampa de iones Bruker Daltonics Amazon ESI-MS) operando en modo de ionizacion positivo. Los compuestos se separaron en Waters Acquity BEH C18 (50 x 2,1 mm; 1,7 jm de diametro de partícula; velocidad de flujo 600 jl/m in y 45 °C) con una fase movil de agua/acetonitrilo conteniendo cada uno 0,1 % de acido formico, usando un gradiente de 5 %-95 % de acetonitrilo durante 9 min. La deteccion se hizo tanto por matriz de diodos como ESI-MS. Se uso Cromatograffa Lfquida de Alta Resolucion-Espectrometna de Masa (HPLC-MS) (Thermo Ultimate 3000 RSLC, acoplado a un instrumento de ionizacion por electropulverizacion Bruker Daltonics Amazon (ESI)-MS trampa de iones) operando en modo de ionizacion positivo. Los compuestos se separaron en una columna Waters Acquity BEH C18 (50 x 2,1 mm; 1,7 jm de diametro de partícula) a una velocidad de flujo de 600 j l min'1 y 45 °C por un gradiente lineal con (A) H2O 0,1 % de acido formico (FA) a (B) acetonitrilo (ACN) 0,1 % de FA a una velocidad de flujo de 600 j l min-1 y 45 °C. El gradiente se inicio por una etapa isocratica de 0,33 min a 5 % de B, seguido de un incremento hasta 95 % de B en 9 min para finalizar con una etapa de lavado de 1 min a 95 % de B antes de volver a equilibrar con las condiciones iniciales. La deteccion se llevo a cabo tanto con una matriz de diodos (DAD) como por ESI-MS. Para el analisis por espectrometna de masa de alta resolucion, las mediciones se realizaron en un sistema Dionex Ultimate 3000 RSLC usando una columna Waters BEH C18 (50 x 2,1 mm, 1,7 jm dp) inyectando 5 j l del extracto metanolico. La separacion se consiguio por el mismo gradiente que anteriormente con una etapa isocratica de 0,33 min a 5 % de B. La deteccion por UV y MS se realizo simultaneamente. El acoplamiento de la HPLC a la MS se apoyo por un sistema Advion Triversa Nanomate nano-ESI unido a un Thermo Fisher Orbitrap. Los espectros de masas se adquirieron en modo centroide variando de 200-2000 m/z a una resolucion de R = 30000.
Para comparacion, la cepa productora natural Myxococcus fulvus Mx f50 (de tipo salvaje, wt) y la Myxococcus xanthus DK1622 AmchA-tet no transformada se cultivaron y extrajeron en paralelo. Los resultados mostrados en la Fig. 7 evidencian claramente que la M. xanthus DK1622 AmchA-tet no transformada no produce mixopironina y que la M. xanthus DK1622 AmchA-tet::pHSU-mxn43 transformante que porta los genes mxnA-mxnM (M. xanthus DK1622 AmchA-tet::mixopironina A) produce mixopironina A como la Myxococcus fulvus Mx f50 de tipo salvaje.
Ejemplo 1: Delecion de secuencias de acido nucleico en Myxococcus para la inactivacion de genes de la ruta sintetica
Para la introduccion de secuencias de acido nucleico en Myxococcus o para la delecion de secuencias de acido nucleico de Myxococcus, se uso preferiblemente el siguiente procedimiento sin marcador:
M. fulvus Mx f50, que se mostro en el Ejemplo 1 que contema los genes que codifican las enzimas de la ruta sintetica completa, se crecio en matraces con deflector con medio CY (0,3 % de casitona, 0,1 % de extracto de levadura, 0,1 % de CaCl2 x 2 H2O) suplementado con 5 x 10'4 pg/L de vitamina B12 a 30 °C hasta que se alcanzo una DO600 entre 0,6-0,9. Las celulas se recogieron entonces de 2-4 ml de cultivo (1-2 x 109 celulas/ml) por centrifugacion a 12,500 rpm durante 1 min a temperatura ambiente. Despues de dos etapas de lavado con 1 ml de H2O a temperatura ambiente, las celulas se resuspendieron en 65 pl de H2O. Se mezclo ADN plasirndico (1-2 pg) con la suspension celular, y se llevo a cabo la electroporacion bajo las siguientes condiciones: 25 pF, 400 Q, 650 V usando cubetas de electroporacion de 0,1 cm y un dispositivo GenePulser XCell (Bio-Rad). Se anadio directamente 1 ml de medio CY a la suspension celular inmediatamente despues de la electroporacion y las celulas se transfirieron a un tubo de centnfuga de 2 ml. Despues de 6-8 h de cultivo a 30 °C y 800 rpm en un thermomixer, las celulas se mezclaron con 2 ml de agar blando CY (medio CY con 0,7 % de agar) y se sembraron en placas de agar CY (medio CY con 1,7 % de agar) suplementadas con 50 pgml'1 de kanamicina o 6,25 pgml'1 de oxitetraciclina. Las placas se incubaron a 30 °C durante 7-10 dfas hasta que las colonias fueron visibles.
Para la construccion de mutantes sin marcador con doble entrecruzamiento se uso el siguiente procedimiento: despues de la verificacion de los mutantes con un unico entrecruzamiento (mutantes resistentes a kanamicina) por PCR, porque fue posible la integracion a traves de dos regiones de homologfa diferentes, un mutante con un unico entrecruzamiento seleccionado se crecio en medio CY en ausencia de antibiotico. Despues de 3-4 dfas, se transfirio 1 ml del cultivo crecido a 50 ml de medio fresco para empezar otro ciclo de cultivo a 30 °C durante 3-4 dfas. Este procedimiento se repitio tres veces para incrementar la posibilidad de un segundo evento de entrecruzamiento, que dana lugar a la perdida del plasmido de inactivacion. Dependiendo de la region de homologfa usada para el segundo entrecruzamiento, esto puede rendir bien el genotipo de tipo salvaje (revertante) o la cepa mutante esperada, en la que la region diana se esta delecionando (mutante con doble entrecruzamiento). Para seleccionar los clones, en los que tuvo lugar esta segunda reaccion de entrecruzamiento, se uso un sistema de contraseleccion basado en el gen sacB. Para esto, se sembraron en placas diferentes diluciones de la poblacion celular en agar CY suplementado con 6 % de sacarosa para la contraseleccion. Despues de 7-10 dfas, las primeras colonias fueron visibles, que se crecieron entonces en medio CY para aislar ADN genomico para la verificacion genotfpica y para extraer los cultivos para el analisis de la produccion de mixopironina.
Para la inactivacion de Mxnl, se empleo mutagenesis insercional de su gen codificador mxnl, usando el plasmido de inactivacion pHSU-mxn13, que se construyo por la amplificacion de un fragmento de 1474 pb para la recombinacion homologa del ADN genomico de M.fulvus Mx f50 por PCR usando los cebadores mxn50 (SEQ ID NO: 29) y mxn51 (SEQ ID NO: 30) que conteman sitios de restriccion BamHI y EcoRV. El fragmento resultante se clono en el vector pCRII-TOPO para generar pHSU-mxn13. El plasmido se transformo en M.fulvus Mx f50 por electroporacion. Para el analisis genotfpico, se llevo a cabo un conjunto de experimentos de PCR diferentes usando las combinaciones de cebadores mxn58 (SEQ ID NO: 31)/pTOPO-in (SEQ ID NO: 32) y mxn59 (SEQ ID NO: 33)/pTOPO-out (SEQ ID NO: 34) para verificar la integracion correcta del plasmido de inactivacion. El metodo de clonacion se representa esquematicamente en la Fig. 8. El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-MxnJ-MxnK-MxnL-MxnM.
Para la inactivacion de MxnJ, se empleo mutagenesis insercional de su gen codificador mxnJ, usando el plasmido de inactivacion pHSU-mxn30, que se construyo por la amplificacion de un fragmento de 1439 pb para la recombinacion homologa del ADN genomico de M.fulvus Mx f50 por PCR usando los cebadores mxn120 (SEQ ID NO: 35) y mxn121 (SEQ ID NO: 36). En otra PCR, el promotor Tn5 se amplifico a partir de la parte central del vector pCRII-TOPO usando los cebadores mxn33 (SEQ ID NO: 37) y mxn34 (SEQ ID NO: 38). Despues de la purificacion en gel, ambos productos de PCR se fusionaron por PCR de superposicion usando los cebadores mxn33 y mxn121. El producto resultante (fragmento homologo Tn5-mxnJ) se hidrolizo con EcoRV y BamHI y se ligo en el vector pSupKan hidrolizado con las mismas enzimas para generar pHSU-mxn30. El plasmido se transformo en M.fulvus Mx f50 por electroporacion. Para el analisis genotfpico, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn130 (SEQ ID NO: 39) /pSubB (SEQ ID NO: 40) y mxn131 (SEQ ID NO: 41) /pSupE (SEQ ID NO: 42) para verificar la integracion correcta del plasmido de inactivacion. El metodo de clonacion se representa esquematicamente en la Fig.
9. El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnK-MxnL-MxnM.
Para la inactivacion de MxnK, se empleo mutagenesis insercional de su gen codificador mxnK, usando el plasmido de inactivacion pHSU-mxn12, que se construyo por la amplificacion de un fragmento de 1448 pb para la recombinacion homologa del ADN genomico de M.fulvus Mx f50 por PCR usando los cebadores mxn35 (SEQ ID NO: 43) y mxn36 (SEQ ID NO: 44). En otra PCR, el promotor Tn5 se amplifico a partir de la parte central del vector pCRII-TOPO usando los cebadores mxn33 y mxn34. Despues de la purificacion en gel, ambos productos de PCR se fusionaron por PCR de superposicion usando los cebadores mxn33 y mxn36. El producto resultante (fragmento homologo Tn5-mxnK) se hidrolizo con EcoRV y BamHI y se ligo en el vector pSupKan hidrolizado con las mismas enzimas para generar pHSU-mxn12. Para el analisis genotfpico, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn48/pSubB y mxn49/pSupE para verificar la integracion correcta del plasmido de inactivacion. Este procedimiento se muestra esquematicamente en la Fig. 10, indicando la posicion de los cebadores en el genoma de tipo salvaje (WT) antes de la integracion por entrecruzamiento, y despues de la integracion en el genoma mutante (Mut). El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnL-MxnM.
En la alternativa, una enzima se inactivo inactivando el gen que la codifica por delecion en marco. Esto se muestra en el ejemplo de mxnK. Para disrumpir mxnK por delecion en marco, se construyo un plasmido de inactivacion genica que portaba dos fragmentos. Los fragmentos tienen un tamano de 1001 pb y 1262 pb y son homologos al area en 5' y en 3' de la region diana cromosomica. Estos fragmentos se amplificaron del ADN genomico de M.fulvus Mx f50 por PCR usando los cebadores mxn54 (SEQ ID NO: 45)/mxn55 (SEQ ID NO: 46) y mxn56 (SEQ ID NO: 47)/mxn57 (SEQ ID NO: 48). Despues de la hidrolisis del fragmento en 5' (producto mxn54/mxn55) con BamHI y Notl, y del fragmento en 3' (producto mxn56/57) con Notl y Sacl, los fragmentos se ligaron en pSWU41 previamente hidrolizado con BamHI y Sacl para generar pHSU-mxn18. Para el analisis genotfpico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn68 (SEQ ID NO: 49)/mxn69 (SEQ ID NO: 50), mxn70 (SEQ ID NO: 51)/mxn72 (SEQ ID NO: 52) y mxn73 (SEQ ID NO: 53)/mxn71 (SEQ ID NO: 54) para verificar la integracion correcta del plasmido de inactivacion. Para el analisis genotfpico de los posibles mutantes con doble entrecruzamiento, se llevaron a cabo PCR usando los cebadores mxn68/mxn71. Ademas, la confirmacion del mutante de delecion se obtuvo por analisis por Transferencia Southern. Este procedimiento de clonacion se muestra esquematicamente en la Fig. 11, indicando la posicion de los cebadores en el genoma de tipo salvaje (WT), en el genoma mutado por la integracion de pHSU-mxn18 (Mut.), y en el genoma despues del doble entrecruzamiento (Mut. Doble Entrecruz.). El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnL-MxnM.
La inactivacion de MxnH pudo obtenerse por delecion en marco de su gen codificador mxnH. En detalle, se construyo un plasmido de inactivacion genica que portaba dos fragmentos, que teman un tamano de 1121 pb y 1250 pb y que eran homologos al area en 5' y en 3' de la region diana cromosomica. Estos fragmentos se amplificaron del ADN genomico de M. fulvus Mx f50 por PCR usando los cebadores mxn117/mxn118 y mxn83/mxn84. Despues de la hidrolisis del fragmento en 5' (producto mxn117/mxn118) con Pvul y Notl, y del fragmento en 3' (producto mxn83/84) con Notl y Spel, los fragmentos se ligaron en pSWU41 hidrolizado con Pvul y Spel para generar el plasmido pHSU-mxn23. Para el analisis genotfpico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn110/mxn111 y mxn112/mxn107 para verificar la integracion correcta del plasmido de inactivacion. Para el analisis genotfpico de los posibles mutantes con doble entrecruzamiento, se llevaron a cabo PCR usando los cebadores mxn110/mxn111, mxn112/mxn107 y mxn110/mxn149 como se representa esquematicamente en la Fig. 12. El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-Mxnl-MxnJ-MxnK-MxnL-MxnM.
La inactivacion de MxnC-MxnH pudo obtenerse por delecion en marco de sus genes codificadores mxnC-mxnH. En detalle, para dicha disrupcion de los genes mxnCDEFGH por delecion en marco, se construyo un plasmido de inactivacion genica que portaba dos fragmentos, que teman un tamano de 1016 pb y 1237 pb y eran homologos al area en 5' y en 3' de la region diana cromosomica. Estos fragmentos se amplificaron del ADN genomico de M. fulvus Mx f50 por PCR usando los cebadores mxn85/mxn119 y mxn83/mxn84. Despues de la hidrolisis del fragmento en 5' (producto mxn85/mxn119) con Pvul y Notl, y del fragmento en 3' (producto mxn83/84) con Notl y Spel, los fragmentos se ligaron en pSWU41 hidrolizado con Pvul y Spel para generar pHSU-mxn22. Para el analisis genotfpico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn102/mxn104 y mxn106/mxn107 para verificar la integracion correcta del plasmido de inactivacion. Para el analisis genotfpico de los posibles mutantes con doble entrecruzamiento, se llevaron a cabo PCR usando los cebadores mxn102/mxn104, mxn106/mxn107 y mxn102/mxn149. El metodo de clonacion se representa esquematicamente en la Fig,13. El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnI-MxnJ-MxnK-MxnL-MxnM.
La inactivacion de MxnC-MxnG pudo obtenerse por delecion en marco de sus genes codificadores mxnC-mxnG. En detalle, para dicha disrupcion de mxnCDEFG por delecion en marco, se construyo un plasmido de inactivacion genica que portaba dos fragmentos, que teman un tamano de 1016 pb y 1250 pb y eran homologos al area en 5' y en 3' de la region diana cromosomica. Estos fragmentos se amplificaron del ADN genomico de M.fulvus Mx f50 por PCR usando los cebadores mxn85/mxn119 y mxn87/mxn88. Despues de la hidrolisis del fragmento en 5' (producto mxn85/mxn119) con Pvul y Notl, y del fragmento en 3' (producto mxn87/88) con Notl y SamHl, los fragmentos se ligaron en pSWU41 hidrolizado con Pvul y SamHl para generar pHSU-mxn21. Para el analisis genotipico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn102/mxn104 y mxn105/mxn103 para verificar la integracion correcta del plasmido de inactivacion. Para el analisis genotfpico de los posibles mutantes con doble entrecruzamiento, se llevaron a cabo PCR usando los cebadores mxn102/mxn104, mxn105/mxn103 y mxn102/mxn150. El metodo de clonacion se representa esquematicamente en la Fig,14. El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnH-MxnI-MxnJ-MxnK-MxnL-MxnM.
La produccion de mixopironina se analizo a partir de caldo de cultivo. Se usaron pre-cultivos crecidos (DO600 = 0,7-1,0) de M.fulvus Mx f50 o de mutantes para inocular cultivos de produccion en matraces con deflector de 300 ml que conteman 50 ml de medio CY (1 % de inoculo). Despues de cultivar durante 2 dfas a 30 °C y 105 rpm, los cultivos se recogieron por centrifugacion. La mixopironina esta casi exclusivamente presente en el sobrenadante, que se extrajo dos veces con un volumen igual de acetato de etilo. Despues de evaporar la fase organica, el residuo se disolvio en 1 ml de metanol. Se analizo una parte alfcuota de 5 pl del extracto por HPLC-MS (Thermo Ultimate 3000 RSLC, acoplado con un detector de trampa de iones Bruker Daltonics Amazon ESI-MS) operando en modo de ionizacion positivo. Los compuestos se separaron en una columna Waters Acquity BEH C18 (50 x 2,1 mm; 1,7 pm de diametro de partícula) a una velocidad de flujo de 600 pl/min y 45 °C con un gradiente lineal con (A) H2O 0,1 % de FA (acido formico) a (B) ACN (acetonitrilo) 0,1 % de FA a una velocidad de flujo de 600 pl/min y 45 °C. El gradiente se inicio por una etapa isocratica de 0,5 min a 5 % de B, seguido de un incremento hasta 95 % de B en 9 min para finalizar con una etapa de lavado de 1 min a 95 % de B antes de volver a equilibrar con las condiciones iniciales. La deteccion se hizo tanto con una matriz de diodos como con ESI-MS. Para la espectrometna de masa de alta resolucion, las mediciones se realizaron en un sistema Dionex Ultimate 3000 RSLC usando una columna Waters BEH C18, 50 x 2,1 mm, 1,7 pm dp por inyeccion de 2 pl de muestra metabolica. La separacion se consiguio con un gradiente lineal con (A) H2O 0,1 % de FA a (B) ACN 0,1 % de FA a una velocidad de flujo de 600 pl/min y 45 °C. El gradiente se inicio por una etapa isocratica de 0,33 min a 5 % de B, seguido de un incremento hasta 95 % B en 9 min para finalizar con una etapa de lavado de 1 min a 95 % de B antes de volver a equilibrar con las condiciones iniciales. La deteccion con UV y MS se realizo simultaneamente. El acoplamiento de HPLC con la MS se apoyo por un sistema Advion Triversa Nanomate nano-ESI unido a un Thermo Fisher Orbitrap. Los espectros de masas se adquirieron en modo centroide variando de 200-2000 m/z a una resolucion de R = 30000.
Los resultados de HPLC-MS se muestran en la Fig. 15. En la cepa productora de tipo salvaje (Mx f50 de Tipo Salvaje), la mixopironina pudo detectarse, pero en los extractos de las cepas mutantes con inactivacion genica obtenidas, p. ej., por inactivacion insercional de mxnl (Mx f50::pHSU-mxn13), inactivacion de mxnJ (Mx f50::pHSU-mxn30), inactivacion de mxnK (Mx f50::pHSU-mxn12), inactivacion por delecion en marco de mxnK (Mx f50::pHSU-mxn18), inactivacion de mxnH por delecion en marco (Mx f50::pHSU-mxn23), inactivacion de mxnC-mxnH por delecion en marco (Mx f50::pHSU-mxn22), no se encontro mixopironina. Para la inactivacion de mxnC-mxnG por delecion en marco (Mx f50::pHSU-mxn21), se encontro el derivado de mixopironina 21-desmetil-mixopironina A. Esto muestra que la biosmtesis de la mixopirina nativa se interrumpe por la inactivacion de las enzimas Mxnl, MxnK, MxnC-MxnH de la ruta biosintetica, mientras la inactivacion de MxnC-MxnG da lugar a la produccion del derivado 21-desmetilo de la mixopironina A.
Ejemplo 2: Delecion de secuencias de acido nucleico en Myxococcus para la inactivacion de enzimas de la ruta sintetica
Para la inactivacion de enzimas, se uso el productor Myxococcus M. fulvus Mx f50 del Ejemplo 2, alternativamente se uso la cepa de expresion M. xanthus DK1622 AmchA-tet:: pHSU-mxn43 del Ejemplo 1.
La inactivacion de mxnL pudo obtenerse por delecion en marco. En detalle, se construyo un plasmido de inactivacion genica que portaba dos fragmentos, que teman un tamano de 1232 pb y 1109 pb y que eran homologos al area en 5' y en 3' de la region diana cromosomica. Estos fragmentos se amplificaron del ADN genomico de M. fulvus Mx f50 por PCR usando los cebadores mxn113/mxn114 y mxn115/mxn116. Despues de la hidrolisis del fragmento en 5' (producto mxn113/mxn114) con Pvul y Notl, y del fragmento en 3' (producto mxn115/116) con Notl y BamHI, los fragmentos se ligaron en pSWU41 hidrolizado con Pvul y BamHI para generar pHSU-mxn20. Para el analisis genotfpico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn98/mxn100 y mxn101/mxn99 para verificar la integracion correcta del plasmido de inactivacion. El proceso de clonacion se representa esquematicamente en la Fig. 16. La confirmacion del mutante de delecion se obtuvo por
analisis por Transferencia Southern. El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-Mxnl-MxnJ-MxnK-MxnM.
La inactivacion de MxnM pudo obtenerse por delecion en marco. En detalle, se construyo un plasmido de inactivacion genica que portaba dos fragmentos, que teman un tamano de 1110 bp y 1199 pb y que eran homologos al area en 5' y en 3' de la region diana cromosomica. Estos fragmentos se amplificaron del ADN genomico de M. fulvus Mx f50 por PCR usando los cebadores mxn132/mxn133 y mxn134/mxn135. Despues de la hidrolisis del fragmento en 5' (producto mxn132/mxn133) con Pvul y Sacl, y del fragmento en 3' (producto mxn134/135) con Sacl y BamHI, los fragmentos se ligaron en pSWU41 hidrolizado con Pvul y BamHI para generar pHSU-mxn34. Para el analisis genotfpico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn140/mxn141 y mxn142/mxn143 para verificar la integracion correcta del plasmido de inactivacion. Para el analisis genotfpico de los posibles mutantes con doble entrecruzamiento, se llevaron a cabo PCR usando los cebadores mxn140/mxn141, mxn142/mxn143 y mxn140/mxn143. El proceso de clonacion se representa esquematicamente en la Figura 17. El microorganismo resultante contema las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-MxnI-MxnJ-MxnK-MxnL.
Se produjeron y analizaron extractos de los sobrenadantes de cultivos de estos microorganismos en muestras tomadas durante el cultivo como se describe en el Ejemplo 2. Los resultados medidos para mixopironina A durante el cultivo se muestran en la Fig. 18 a) para el M. fulvus Mx f50 de tipo salvaje (WT), en la Fig. 18 b) para el microorganismo que contiene las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-MxnI-MxnJ-MxnK-MxnM (AmxnL), y en la Fig. 18 c) para el microorganismo que contiene las enzimas de la ruta sintetica de MxnA-MxnB-MxnC-MxnD-MxnE-MxnF-MxnG-MxnH-MxnI-MxnJ-MxnK-MxnL, con el gen mxnM que codifica MxnM estando delecionado (AmxnM). Estos resultados muestran que estas enzimas de la ruta sintetica son suficientes para la smtesis de mixopironina o sus derivados.
Ejemplo 3: Inactivacion de centros catalfticos de dominios de genes de la ruta sintetica
Usando los metodos de clonaciones descritos en la presente memoria, preferiblemente el metodo del doble entrecruzamiento usando una construccion de acido nucleico de inactivacion, pueden inactivarse centros catalfticos unicos o multiples de dominios en las enzimas, preferiblemente mutando la secuencia codificadora de aminoacidos que definen el centro catalftico, p. ej., aminoacidos catalfticamente activos. Los aminoacidos preferidos para cambiarse para la inactivacion de un centro catalftico, p. ej., delecionarse o intercambiarse por un aminoacido catalfticamente inactivo, p. ej., por Leucina, Isoleucina, Alanina, o Valina, se proporcionan en la Tabla 3 siguiente en negrita, que aminoacidos participan en la actividad catalftica.
Tabla 3: aminoacidos de los centros catalfticos



Tabla 3 continuacion:

Tabla 3 continuacion:


Tabla 3 continuacion:

Tabla 3 continuacion:


En la Tabla 3, — indica huecos insertados en las secuencias de aminoacidos para un mejor alineamiento de aminoacidos similares durante el analisis asistido por ordenador. Para los alineamientos de KS y KR, los huecos insertados comprenden mas de 3 aminoacidos. En la Tabla 3, las secciones de aminoacidos proporcionadas como aminoacidos Nos. de la secuencia de aminoacidos de una enzima comprenden o definen la seccion de la enzima respectiva que contiene el dominio. El alineamiento de aminoacidos para los centros catalfticos muestra la importancia de los aminoacidos conservados, que se escriben en negrita. De acuerdo con esto, la mutacion de la secuencia codificadora a cambio, p. ej., eliminacion de al menos uno de estos aminoacidos, preferiblemente dos o tres de estos aminoacidos conservados o todos los aminoacidos proporcionados para un centro catalftico, puede usarse para la inactivacion del dominio respectivo, preferiblemente del modulo indicado.
Los ejemplos para los metodos de clonacion para inactivar un centro catalftico espedfico se proporcionan en el Ejemplo 7 mas adelante para el dominio CP de los modulos 1W y 4W, respectivamente.
Preferiblemente, para la produccion de un derivado de mixopironina, se inactiva al menos un centro catalftico de cetorreductasa (KR). Como un ejemplo, la Fig. 19 muestra esquematicamente el efecto de la inactivacion del dominio KR del modulo 2E que da lugar a la produccion de un derivado de mixopironina. La inactivacion del dominio KR del modulo 2E da lugar a que el intermedio p-cetoester no se reduzca al intermedio hidroxi ester, y de acuerdo con esto el grupo p-cetoester aparece en la cadena lateral del anillo pirona.
Preferiblemente, para la produccion de un derivado de mixopironina, se inactiva al menos un centro catalftico de un dominio catalftico deshidrogenasa (DH). Como un ejemplo, la Fig. 20 muestra esquematicamente el efecto de la inactivacion del dominio DH del modulo 2E que da lugar a la produccion de un derivado de mixopironina. La inactivacion del dominio DH del modulo 2E da lugar a que el intermedio hidroxiester no se deshidrate, y de acuerdo con esto el grupo p-hidroxilo aparece en la cadena lateral del anillo pirona.
Preferiblemente, para la produccion de un derivado de mixopironina, se inactiva el centro catalftico de al menos un dominio metiltransferasa (MT). Como un ejemplo, la Fig. 21 muestra esquematicamente el efecto de la inactivacion del dominio MT del modulo 4E que da lugar a la produccion del efecto de la inactivacion del dominio MT del modulo 4E que da lugar a la produccion de un derivado de mixopironina. La inactivacion de un dominio MT, p. ej., del modulo 4W da lugar en ausencia de un grupo metilo en la cadena lateral del anillo pirona, en este caso en ausencia del grupo metilo unido al carbono a.
Ejemplo 4: Produccion de derivados de mixopironina usando enzimas en un proceso in vitro
La produccion de mixopironina o un derivado se catalizo in vitro por MxnB en combinacion con el centro catalftico del dominio CP del modulo 6E de MxnJ (CP-6E) o con el centro catalftico CP del dominio 5W de MxnK (CP-5W) a partir de los precursores JHS486 y JHS274. Esto muestra que MxnB cataliza la condensacion de precursores para formar el resto de anillo pirona que tiene las cadenas de los precursores. En presencia del centro catalftico del dominio CP del modulo 6E de MxnJ y en presencia del centro catalftico del dominio CP del modulo 5W de MxnK, respectivamente, se produce la transferencia del resto diceto de cada precursor a MxnB formando el anillo pirona, indicando que los derivados de mixopironina pueden producirse por estos peptidos a partir de precursores que estan activados, p. ej., activados por portar un grupo auxiliar, p. ej., como el grupo SNAC, unido al grupo diceto.
Los peptidos se expresaron a partir de casetes de expresion separados: para la expresion de MxnB, la secuencia codificadora de acido nucleico mxnB se amplifico del ADN genomico de M. fulvus Mx f50 por PCR usando los cebadores mxn9 y mxn10. Los productos de PCR se clonaron en el vector romo pJET1,2 (Thermo Fisher Scientific) dando lugar a pJET-MxnB y se secuencio. El pJET-HSU-mxn5 resultante se digirio con Ndel y HindIII y el fragmento de ADN que portaba la region codificadora de MxnB se clono en el sitio Ndel y HindIII de pColdl dando lugar a pColdl-HSU-mxn5, que codifica MxnB con una etiqueta N-terminal de hexahistidina.
La secuencia codificadora para CP-6E (dominio CP del modulo 6E) se amplifico de ADN genomico de M. fulvus Mx f50 por PCR usando los cebadores mxn214 y mxn215. Los productos de PCR se clonaron en el vector romo pJET1,2 (Thermo Fisher Scientific) dando lugar a pJET-CP-6E y se secuencio. El pJET-HSU-mxn59 se digirio con Ndel y Hindlll, y el fragmento de ADN que portaba la region codificadora de CP-6E se clono en los sitios Ndel y Hindlll de pColdl, dando lugar a pColdI-HSU-mxn59 con una etiqueta N-terminal de hexahistidina. Para obtener CP-6E soluble, se
coexpreso una chaperona usando pGro7 para obtener pColdI-pGro7-mxn59. Independientemente, se coexpreso pColdI-HSU-mxn59 con pSUMtaA para obtener pColdl-pSUMtaHSU-mxn64.
La secuencia codificadora para CP-5W (dominio CP del modulo 5W) se amplifico del ADN genomico de M.fulvus Mx f50 por PCR usando los cebadores mxn230 y mxn231. Los productos de PCR se clonaron en el vector romo pJET1,2 (Thermo Fisher Scientific) dando lugar a pJET-MxnB y se secuencio usando los cebadores pJET1,2For/pJET1,2Rev, respectivamente. pJET-HSU-mxn63 se digirio con Ndel y Hindlll y el fragmento de ADN que portaba la secuencia codificadora de CP-5W se clono en los sitios Ndel y Hindlll de pColdl dando lugar a pColdl-HSU-mxn63 con una etiqueta N-terminal de hexahistidina. Ademas, se coexpreso pColdI-HSU-mxn63 con pSUMtaA para obtener pColdlpSUMtaHSU-mxn65.
MxnB, el centro catalttico del dominio CP del modulo 6E de MxnJ (CP-6E) y el centro catalftico del dominio CP del modulo 5W de MxnK (CP-5W) se produjeron cada uno separadamente por expresion heterologa. Los plasmidos de expresion respectivos, pHSU-mxn5, pHSU-mxn57, pHSU-mxn59, y pHSU-mxn63, se introdujeron en Escherichia coli BL21(DE3) en transformaciones separadas. Las E. coli BL21(DE3) que portaban los plasmidos de expresion se cultivaron en medio LB que contema 100 pg/mL de ampicilina (para la expresion de pColdl) o medio LB modificado que contema 100 pg/mL de ampicilina, 25 pg/mL de cloranfenicol y 2 mg/mL de L-arabinosa (para la expresion de pColdI-pGro7) a 37 °C hasta una DO600 = 0,5 a 0,6. La expresion se indujo por la adicion de isopropil-p-dtiogalactopiranosido (IPTG, 0,15 mM) y el cultivo se continuo a 16 °C durante 20 h. Las celulas se recogieron por centrifugacion (8000 rpm; 20 min) y se resuspendieron en tampon de lisis (tampon Tris-HCl 40 mM pH 7,5 que contema glicerol al 10%, NaCl 300 mM e imidazol 10 mM). Las celulas se lisaron por sonicacion y se preparo un extracto sin celulas eliminando los restos celulares por centrifugacion a 8000 rpm durante 10 min a 4 °C. El extracto sin celulas se purifico usando resina Ni-NTA Superflow (Qiagen), lavada con tampon que contema imidazol (20 mm), y eluida con imidazol (200 mm) en el mismo tampon. El tampon de elucion se intercambio con tampon (tampon Tris-HCl 40 mM pH 7,5 que contema glicerol al 10%, NaCl 300 mM) usando ultrafiltracion (Amicon, Milipore). Las concentraciones de protemas se estimaron por el ensayo de Bradford (Bio-Rad) con albumina de suero bovino como estandar. Las protemas purificadas se congelaron rapidamente en nitrogeno lfquido y se almacenaron a -80 °C.
Para la reaccion, se preparo una mezcla (total de 100 pL) que contema CP-6E o CP-5W100 pM, MxnB 30 pM, JHS274 1 mM y JHS486 1 mM y se incubo a 37 °C durante 8 h. Las mezclas de reaccion se extrajeron con acetato de etilo y la capa organica se evaporo. El residuo se disolvio en 100 de metanol y una parte almuota de 5 pl del extracto se analizo por el residuo se disolvio en 100 pl de metanol y una almuota 5 pl del extracto se analizo por HPLC/MS como se describe en el Ejemplo 2. Los resultados de HPLC-MS se muestran en la Fig. 22, indicando que los precursores en presencia de MxnB solo se condensan en un anillo pirona, y que en presencia de MxnB y el dominio CP del modulo 6E y/o dominio CP del modulo 5W, los precursores se condensan en un anillo pirona (formula estructural anadida) que porta la cadena lateral unida a SNAC en JHS486 como la cadena occidental y que porta la cadena lateral unida a SNAC en JHS274 como la cadena oriental.
Ejemplo 5: Produccion de derivados de mixopironina usando enzimas con MxnK inactivada y alimentacion de precursor de la cadena occidental
Se ha encontrado que el mutante de bloqueo que se produce a partir de la inactivacion de MxnK por mutagenesis sin marcador de mxnK(M.fulvus Mx f50 ApHSU-mxn18, que contiene pHSU-mxn18 descrito en el Ejemplo 2), no produce mixopironina A. Para los experimentos de mutasmtesis, se crecio un precultivo de M.fulvus Mx f50ApHSU-mxn18 en medio CY a 30 °C hasta una DO600 entre 0,3 a 0,6. El precultivo se uso para inocular 20 mL de medio CY (7,2 x 106 celulas/mL). El cultivo se llevo a cabo durante 12-24 horas mas antes de alimentar el precursor que actua como un mimetico de sustrato. Se alimentaron esteres de N-acetil-cisteamina (SNAC) que se usaron como un precursor de la cadena occidental cada uno en los diferentes cultivos hasta una concentracion final de 100 pM. Se cultivaron ademas durante otras 48 h antes de recoger las celulas.
Los precursores y los derivados de mixopironina respectivos encontrados se proporcionan en la Tabla 4 siguiente. Los espectros de MS2 del producto en los extractos de los sobrenadantes de los cultivos se muestran en la Fig. 23. El analisis del extracto crudo se realizo por HPLC/MS como se ha descrito anteriormente. La medicion por espectrometna de masa de alta resolucion confirmo la incorporacion de las cadenas de los precursores mostrando la misma manera de patron de fragmentacion.




Estos resultados muestran que los derivados de mixopironina pueden producirse por la expresion de las enzimas sinteticas, en donde MxnK esta inactivada, p. ej., las enzimas proporcionadas consisten en MxnA, Mxnl, MxnJ, MxnK, y opcionalmente comprenden MxnL y MxnM, p. ej., por la expresion en un microorganismo, en combinacion con la adicion de un precursor de la cadena occidental.
Para la produccion de derivados de mixopironina, MxnK podna inactivarse en uno de sus dominios catalfticos, p. ej., en un dominio CP, en combinacion con la alimentacion de un precursor de la cadena occidental. En esta realizacion, la parte remanente de MxnK convierte el precursor de la cadena occidental en un precursor de la cadena occidental convertido que se condensa entonces con la cadena oriental por MxnB, dando lugar a un derivado de mixopironina que tiene una cadena occidental que se origina del precursor de la cadena occidental convertido.
Ejemplo 6: Produccion de derivados de mixopironina usando enzimas con un centro catalftico de MxnK inactivado y alimentacion con el precursor de la cadena occidental
Como un ejemplo para la inactivacion de un centro catalftico, un dominio catalftico de CP (protema transportadora) se inactivo en diferentes modulos de MxnK. MxnK podna inactivarse parcialmente, p. ej., mediante la inactivacion del dominio catalftico de CP del modulo 1W de MxnK. Para inactivar este dominio catalftico de MxnK, se indujo una mutacion puntual en su secuencia codificadora mxnK usando un plasmido de inactivacion genica que portaba un fragmento de 2445 pb. El fragmento se amplifico a partir el plasmido pHSU-mxn43 del Ejemplo 1 por PCR de superposicion usando los cebadores mxn169/mxn170 y mxn171/mxn172. Despues de la hidrolisis del fragmento con Pvul y Notl, se ligo en pSWU41 hidrolizado con Pvul y Notl para generar pHSU-mxn48. Para el analisis genotipico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn185/mxn70 y pSWU41-F/mxn187 para verificar la integracion correcta del plasmido de inactivacion. Para el analisis genotfpico del doble entrecruzamiento, se realizo una PCR usando la combinacion de cebadores mxn189/mxn205 y el producto amplificado se secuencio. El metodo de clonacion se muestra esquematicamente en la Fig. 24.
MxnK podna inactivarse parcialmente en el dominio catalftico CP de su modulo 4W por mutacion puntual. Para inactivar mxnK en el dominio CP del modulo 4W, se construyo un plasmido de inactivacion genica que portaba un fragmento de 2486 pb. El fragmento se amplifico a partir del plasmido pHSU-mxn43 por PCR de superposicion usando los cebadores mxn174/mxn175 y mxn176/mxn177. Despues de la hidrolisis del fragmento con Pvul y Notl, se ligo en pSWU41 hidrolizado con Pvul y Notl para generar pHSU-mxn49. Para el analisis genotfpico del unico entrecruzamiento, se llevo a cabo un conjunto de diferentes PCR usando las combinaciones de cebadores mxn191/mxn70 y pSWU41-F/mxn193 para verificar la integracion correcta del plasmido de inactivacion. Para el analisis genotfpico del doble entrecruzamiento, se realizo una PCR usando la combinacion de cebadores mxn195/mxn206 y el producto amplificado se secuencio. El metodo de clonacion se muestra esquematicamente en la Fig. 25.
Para la produccion de derivados de mixopironina, un mutante de bloqueo de al menos un dominio catalftico de protema transportadora (CP) de MxnK inactivado se cultivo con alimentacion de un precursor de la cadena occidental. El precursor de la cadena occidental puede convertirse por centros catalfticos organizados en C-terminal respecto al centro catalftico inactivado. Como ejemplos, la Fig. 26 en a) muestra esquematicamente la smtesis de tipo salvaje de la parte de la molecula de la cadena occidental, en b) MxnK con el dominio CP del modulo 1W inactivado con alimentacion de un precursor de la cadena occidental que esta unido al dominio CP del modulo 3W y se convierte posteriormente por los modulos 3W a 5W antes de condensarse en el anillo pirona, en c) MxnK con el dominio CP del modulo 3W inactivado con alimentacion de un precursor de la cadena occidental que esta unido al dominio CP del modulo 4W y se convierte posteriormente por los modulos 4W a 5W antes de condensarse en el anillo pirona, y en d) MxnK con el dominio CP del modulo 4W inactivado con alimentacion de un precursor de la cadena occidental que esta unido al dominio CP del modulo 5W y se convierte posteriormente por el modulo 5W antes de condensarse en el anillo pirona. Generalmente, especialmente en las Figuras 26-28, las enzimas en las que al menos un dominio catalftico esta inactivado puede denominarse Mutasinton.
Ejemplo 7: Produccion de derivados de mixopironina usando enzimas con un centro catalftico de Mxnl-MxnJ inactivado y alimentacion con precursor de la cadena oriental
Como se ha mostrado anteriormente, la inactivacion de MxnJ en M.fulvus Mx f50 ::pHSU-mxn30 suprime la produccion de mixopironina A. Como un ejemplo para la inactivacion de un dominio catalftico, un dominio catalftico CP (protema transportadora) se inactivo en diferentes modulos de Mxnl-MxnJ.
La Fig. 27 muestra esquematicamente en a) la smtesis de tipo salvaje (wt) de la parte de la molecula de la cadena oriental, en b) Mxnl-MxnJ que tiene un dominio CP inactivado en el modulo de carga N-terminal y la union del precursor de la cadena oriental alimentado por el dominio CP del modulo 1E y su conversion posterior por los siguientes modulos de Mxnl-MxnJ, en c) Mxnl-MxnJ que tiene un dominio CP inactivado en el modulo 1E y la union del precursor de la
cadena oriental alimentado por el dominio CP del modulo 2E y su conversion posterior por los siguientes centros catalfticos de Mxnl-MxnJ, en d) Mxnl-MxnJ que tiene un dominio CP inactivado en el modulo 3E y la union del precursor de la cadena oriental alimentado por el dominio CP del modulo 4E y su conversion posterior por los siguientes modulos de Mxnl-MxnJ, y en e) Mxnl-MxnJ que tiene un dominio CP inactivado en el modulo 4E y la union del precursor de la cadena oriental alimentado por el dominio CP del modulo 6E y su condensacion posterior al anillo pirona, es decir, su condensacion sin conversion adicional. En la alternativa a la inactivacion del dominio CP en el modulo 4E, el dominio CP en el modulo 5E puede inactivarse.
Para la produccion, se crecio un precultivo de M.fulvus Mx f50::pHSU-mxn30 en medio CY a 30 °C hasta una DO600 entre 0,3-0,6. El precultivo se uso para inocular 20 mL de medio CY (7,2 x 106 celulas/mL). El cultivo se llevo a cabo durante 12-24 horas adicionales antes de alimentar los mimeticos de sustrato. Se anadieron esteres de W-acetilcisteamina (SNAC) que mimetizan la cadena oriental a los cultivos a la final de 100 pM. Se crecieron ademas durante otras 48 horas antes de recoger las celulas. El analisis del extracto crudo se realizo por HPLC/MS.
Ejemplo 8: Produccion de derivados de mixopironina usando enzimas con un centro catalftico CP de MxnK inactivado y alimentacion del precursor de la cadena occidental y un centro catalftico de Mxnl-MxnJ inactivado
Como un ejemplo para la combinacion de la segunda y tercera realizaciones, M.fulvus Mx f50 ::pHSU-mxn48, en donde el dominio catalftico CP del modulo 1W de MxnK esta inactivado, se usa en combinacion con la inactivacion de al menos un dominio catalftico de Mxnl-MxnJ. La inactivacion de al menos un dominio catalftico de Mxnl-MxnJ se ejemplifica por la inactivacion del dominio catalftico MT en el modulo 4E de MxnJ. La Fig. 28 muestra esquematicamente la smtesis de la cadena occidental empezando a partir del precursor de la cadena occidental que esta unido por el dominio catalftico CP del modulo 3W y se convierte posteriormente por los siguientes dominios catalfticos de los modulos 4W-5W, mientras la smtesis de la parte de la molecula de la cadena oriental esta alterada por la ausencia de la actividad del dominio MT en el modulo 4E. La condensacion del precursor de la cadena occidental convertido con la parte de la molecula de la cadena oriental alterada esta catalizada por MxnB.
Ejemplo 9: Produccion de derivados de mixopironina usando enzimas con un centro catalftico de MxnK inactivado y un centro catalftico CP de Mxnl-MxnJ inactivado con alimentacion de precursor de la cadena oriental
Como un ejemplo para la combinacion de la segunda y tercera realizaciones, M.fulvus Mx f50, en donde el dominio catalftico MT del modulo 4W de MxnK esta inactivado se describe en combinacion con la inactivacion del domino catalftico CP del modulo 1E de Mxnl-MxnJ, con alimentacion de un precursor de la cadena oriental durante el cultivo. La ruta sintetica del derivado de mixopironina resultante se representa esquematicamente en la Fig. 29, en donde la parte de la molecula de la cadena occidental esta alterada debido a la ausencia de la actividad de la metil transferasa y en donde el precursor de la cadena oriental esta unido por el dominio catalftico CP del modulo 2E y se convierte posteriormente por los siguientes modulos de Mxnl-MxnJ, seguido de la condensacion al anillo pirona como se cataliza por MxnB.
Ejemplo 10: Produccion de derivados de mixopironina usando enzimas, en donde al menos un centro catalftico o dominio o enzima de Mxnl-MxnJ-MxnK se intercambia por al menos un centro catalftico o dominio o enzima de Corl-CorJ-CorK-CorL
Produccion heterologa de coralopironinas en Myxococcus xanthus DK1622
El metodo de clonacion para la generacion de una construccion de expresion que porta la ruta "hftbrida" se ilustra en la Figura 8. La ruta de biosmtesis de la cadena oriental es pHSU-mxn41 (que consiste en mxnA-mxnJ) de la ruta de la mixopironina y la parte occidental incluyendo genes adaptados (corK-corO) generada de una biblioteca de cosmidos de coralopironina AM24 y BA5.
En primer lugar, se amplifico un casete de resistencia a apramicina por PCR usando los cebadores mxn200/mxn201 con el vector pKC1132 como el molde. El fragmento Z (~13 kb) en el cosmido BA5 se intercambio con el casete de resistencia a apramicina (ApraR) mediante recombinacion Red/ET para generar pHSU-mxn51. En paralelo, otro casete de resistencia a apramicina se amplifico por PCR usando los cebadores mxn202/mxn203 con el vector pKC1132 como el molde. El fragmento Y en el cosmido AM24 se intercambio entonces con el casete de resistencia a apramicina (ApraR) mediante recombinacion Red/ET para generar pHSU-mxn52. Se realizo otra ronda de recombinacion Red/ET para intercambiar el fragmento X en el cosmido AM24 con un casete de resistencia a cloranfenicol (CmR) para generar pHSU-mxn53. La siguiente ronda de recombinacion Red/ET se realizo entonces para integrar el fragmento corK* de pHSU-mxn53 en pHSU-mxn51 para generar pHSU-mxn54. Tanto pHSU-mxn41 como pHSU-mxn54 se hidrolizaron entonces con Spel/Pacl para ligar el fragmento corK-O en pHSU-mxn41 para generar pHSU-mxn55.
Posteriormente, tanto pHSU-mxn37 como pHSU-mxn42 se hidrolizaron con Pacl para ligar el casete de resistencia a kanamicina en pHSU-mxn55 para generar pHSU-mxn56. La construccion de expresion pHSU-mxn56 descrita
anteriormente mostrada en la Figura 8 puede transformarse entonces en un huesped de expresion, p. ej., M. xanthus DK1622 AmchA-tet por electroporacion como se ha descrito anteriormente.
Los precursores podnan obtenerse como sigue:
Esquema 1: smtesis general de esteres de p-ceto-A/-acetilcisteamina (SNAC) insaturados

Esquema 4: smtesis de esteres de p-ceto-tiofenol insaturados

Se prefieren los siguientes precursores:


Los compuestos precursores podnan sintetizarse por los siguientes metodos:
Procedimiento general para la smtesis de acil acidos de meldrum a partir de cloruros de acilo: Procedimiento general A:
A una disolucion de acido de meldrum (10,8 g, 75,0 mmoles) y piridina (12,0 g, 0,15 moles) en diclorometano anhidro (50 mL) a 0 °C se anadio gota a gota una disolucion del cloruro de acilo apropiado (80,0 mmoles) en diclorometano anhidro (20 mL). La mezcla naranja resultante se agito durante 1 h a 0 °C y despues durante 1 h a temperatura ambiente. La mezcla de reaccion se diluyo con diclorometano (50 mL) y se lavo con HCl 2M acuoso (3 x 50 mL), salmuera (2 x 50 mL), se seco (MgSO4) y se concentro bajo presion reducida. El material crudo se filtro sobre una almohadilla corta de SiO2 eluyendo con n-hexano/EtOAc 2:1.
Procedimiento general para la smtesis de oxinonas a partir de acil acidos de meldrum: Procedimiento general B El acil acido de meldrum apropiado (20,0 mmoles) se disolvio en tolueno (25 mL) y acetona (2,0 mL). La disolucion se calento a reflujo durante 2 h. Los disolventes se eliminaron bajo presion reducida y el material crudo se purifico por cromatograffa flash (SO 2, n-hexano/EtOAc) o destilacion en vacm.
Procedimiento general para la smtesis de fosfonatos de dioxinona: Procedimiento general C: A una disolucion de diisopropilamina (7,18 mL, 51,1 mmoles) en THF (30 mL) se anadio gota a gota una disolucion de n-BuLi (2,5M en n-hexano, 20,5 mL, 51,1 mmoles) a 0 °C. Despues de agitar durante 30 minutos, la mezcla de reaccion se enfrio hasta -78 °C y se anadio la 2,2-dimetil-4H-1,3-dioxin-4-ona apropiada (36,5 mmoles) durante 5 min. Despues de 40 min se anadio clorodietilfosfito (7,62 mL, 53,0 mmoles) y el bano de enfriamiento se retiro. Cuando la mezcla alcanzo la temperatura ambiente, se diluyo con Et2O (200 mL), se lavo con salmuera semisaturada (100 mL), se seco (MgSO4) y se concentro en vacm. El aceite amarillo resultante se disolvio en CH2Cl2 (60 mL) y se anadio cuidadosamente H2O2 acuoso (al 30 %, 13 mL) a 0 °C. La disolucion se agito durante 1 h y se diluyo con EtOAc (200 mL). Despues de la separacion, la capa organica se lavo con salmuera (2 x 100 mL), se seco (MgSO4) y se concentro en vacm. El material crudo se purifico por cromatograffa flash (SO 2, EtOAc).
Procedimiento general para la olefinacion de Horner-Wadsworth-Emmons para la preparacion de dioxinonas insaturadas: Procedimiento general D
A una disolucion de diisopropilamina (1,04 mL, 7,13 mmoles) en THF (20 mL) se anadio una disolucion de n-BuLi (2,5M en n-hexano, 2,85 mL, 7,13 mmoles) a 0 °C. Despues de agitar durante 30 min, la mezcla de reaccion se enfrio hasta -78 °C. Se anadio una disolucion del fosfonato apropiado (6,84 mmoles) en THF (10 mL) a la mezcla de reaccion, y la mezcla resultante se calento hasta 0 °C y se agito durante 15 min. Despues de enfriar hasta -78 °C, se anadio HMPA (2,10 mL, 12,1 mmoles) y la agitacion se continuo durante 30 min. Se anadio lentamente una disolucion del aldetffdo apropiado (6,84 mmoles) en THF (10 mL). La mezcla de reaccion de dejo calentar lentamente hasta 0 °C y se agito durante 1 h. Despues de diluir con EtOAc (80 mL), la mezcla se lavo con NH4O acuoso saturado (3 x 40 mL) y salmuera (50 mL). La capa organica se seco (MgSO4) y se concentro. El material crudo se purifico por cromatograffa flash (SiO2, n-hexano/EtOAc).
Procedimiento general para la smtesis de esteres de p-ceto-W-acetilcisteamina (SNAC) a partir de dioxinonas: Procedimiento general E:
Una disolucion desgasificada de la dioxinona apropiada (0,95 mmoles), W-acetilcisteamina (SNAC-H, 0,15 g, 1,25 mmoles) en tolueno ( 6 mL) se puso a reflujo durante 16 h bajo una atmosfera de nitrogeno. Despues de enfriar hasta temperatura ambiente, el disolvente se evaporo bajo presion reducida y el producto crudo se purifico por cromatograffa en columna flash (SiO2, n-hexano/EtOAc = 1 :1 ^ EtOAc) para rendir el ester de SNAC deseado.
Procedimiento general para la smtesis de acil acidos de meldrum a partir de acidos carboxflicos mediante acoplamiento con DCC: Procedimiento general F:
Una disolucion del acido apropiado (10,6 mmoles) y DCC (2,19 g, 10,6 mmoles) en CH2Cl2 anhidro (20 mL) se agito durante 30 min a 0 °C. Un solido blanco precipito. Se anadieron posteriormente DMAP (1,94 g, 15,9 mmoles) y acido de meldrum (1,52 g, 10,6 mmoles). El bano de enfriamiento se retiro y la mezcla de reaccion se agito durante 2 h a temperatura ambiente. La urea insoluble se retiro por filtracion y la torta del filtro se lavo con CH2CI2 enfriado en hielo (10 mL). Los filtrados combinados se lavaron con HCl 1M (2 x 50 mL), H2O (2 x 50 mL) y salmuera (75 mL), se secaron (MgSO4) y se concentraron. Para eliminar los ultimos residuos de urea, el material crudo se filtro sobre algodon.
Procedimiento general para la smtesis de esteres de p-ceto-W-acetilcisteamina (SNAC) a partir de acil acidos de meldrum: Procedimiento general G:
Una disolucion desgasificada del acil acido de meldrum apropiado (0,80 mmoles), W-acetilcisteamina (SNAC-H, 0,12 g, 1,00 mmol) en tolueno (6 mL) se puso a reflujo durante 16 h bajo una atmosfera de nitrogeno. Despues de enfriar hasta temperatura ambiente, el disolvente se evaporo bajo presion reducida y el producto crudo se purifico por cromatograffa en columna flash (SiO2, n-hexano/EtOAc = 1 :1 ^ EtOAc) para rendir el ester de p-ceto-SNAC deseado.
Procedimiento general para la smtesis de esteres de W-acetilcisteamina (SNAC) a partir de acidos carboxflicos mediante acoplamiento con DCC: Procedimiento general H:
Una disolucion del acido apropiado (1,06 mmoles) y DCC (0,41 g, 1,06 mmoles) en CH2CI2 anhidro (5 mL) se agito durante 30 min a 0 °C. Un solido blanco precipito. Se anadieron posteriormente DMAP (0,37 g, 1,59 mmoles) y SNAC-H (0,24 g, 1,06 mmoles). El bano de enfriamiento se retiro y la mezcla de reaccion se agito durante 2 h a temperatura ambiente. La urea insoluble se retiro por filtracion y la torta del filtro se lavo con CH2CI2 enfriado en hielo (10 mL). Los filtrados combinados se evaporaron en vacm y el producto crudo se purifico por cromatograffa en columna flash (SiO2, n-hexano/EtOAc = 1 :1 ^ EtOAc) para rendir el ester de SNAC deseado.
Procedimiento general para la smtesis de esteres de tiofenol a partir de acil acidos de meldrum: Procedimiento general I: Una disolucion desgasificada de la dioxinona (0,92 mmoles), tiofenol (0,13 g, 1,16 mmoles) en tolueno (6 mL) se puso a reflujo durante 16 h bajo una atmosfera de nitrogeno. Despues de enfriar hasta temperatura ambiente, el disolvente se evaporo bajo presion reducida y el producto crudo se purifico por cromatograffa en columna flash (SiO2, nhexano/EtOAc = 12:1 —^ 8:1) para rendir el tioester deseado.
Smtesis y datos espectroscopicos de los intermedios.
(E)-3-(3-Hidroxifenil)acrilaldehndo (IM-1) (JHS216)

El compuesto del tftulo se preparo a partir de 3-hidroxibenzaldehndo segun el siguiente procedimiento:
Una mezcla de 3-hidroxibenzaldehndo (5,00 g, 41,0 mmoles) y acetato de vinilo (4,15 mL, 45,1 mmoles) en acetonitrilo (15 mL) se anadio a temperatura ambiente a una suspension de carbonato de potasio (6,80 g, 49,2 mmoles) en acetonitrilo (50 mL) y agua (0,2 mL). La mezcla de reaccion se puso a reflujo durante 40 h y se enfrio hasta temperatura ambiente. La mezcla de reaccion se diluyo con agua (50 mL) y EtOAc (50 mL). La capa acuosa se separo y se extrajo con EtOAc (2 x 50 mL). Las capas organicas combinadas se secaron (MgSO4) y se concentraron en vado a sequedad. El residuo se purifico por cromatograffa en columna flash (SO 2, n-hexano/EtOAc 8:1— 2:1) para rendir el compuesto del tftulo.
Rendimiento: 25%, solido naranja. 1H RMN (DMSO-d6): 8 = 9,68 (br. s, 1H), 9,65 (d, J = 7,7 Hz, 1H), 7,65 (d, J =15,8 Hz, 1H), 7,27 (t, J = 8,0 Hz, 1H), 7,17 (d, J = 8,0 Hz, 1H), 7,08 (t, J = 2,4 Hz, 1H), 6,89 (ddd, J = 8,0, 2,4, 1,0 Hz, 1H), 6,73 (dd, J =15,8, 7,7 Hz, 1H) ppm. 13C RMN (DMSO-d6): 8 = 194,4, 157,8, 153,5, 135,3, 130,1, 128,3, 119,6, 118,4, 115,0 ppm.
(E)-3-(3-((terc-Butildimetilsilil)oxi)fenil)acrilaldemdo (IM-2) (JHS221)

El compuesto del tftulo se preparo a partir de IM-1 segun el siguiente procedimiento:
Se anadio imidazol (0,86 g, 12,7 mmoles) a una disolucion de IM-1 (0,76 g, 5,13 mmoles) y ferc-butildimetilclorosilano (0,92 g, 6,10 mmoles) en W,A/-dimetilformamida anhidra (10 mL). La mezcla se agito durante 3 h, se diluyo con NaHCO3 acuoso al 5 % (50 mL), y se extrajo con EtOAc (4 x 50 mL). Las capas organicas combinadas se lavaron con salmuera (100 mL), se secaron (MgSO4), y se evaporaron en vado para rendir el compuesto del tftulo.
Rendimiento: 82 %, aceite amarillo. 1H RMN (DMSO-d6): 8 = 9,66 (d, J =7,7 Hz, 1H), 7,71 (d, J =15,9 Hz, 1H), 7,34 7,38 (m, 2H), 7,19-7,22 (m, 1H), 6,94-6,99 (m, 1H), 6,84 (dd, J = 15,9, 7,7 Hz, 1H), 0,96 (s, 9H), 0,21 (s, 6H) ppm. 13C RMN (DMSO-d6): 8 = 194,3, 155,5, 152,9, 135,6, 130,2, 128,7, 122,7, 121,9, 119,8, 25,5, 17,9, -4,6 ppm.
(E)-3-(3-Bromofenil)acrilaldelddo (IM-3) (JHS487)

El compuesto del tftulo se preparo a partir de 3-bromobenzaldehndo segun el siguiente procedimiento:
Una disolucion de 3-bromobenzaldehndo (5,00 g, 27,0 mmoles) en H2O (6 mL) se utilizo para llenar un matraz de fondo redondo de tres bocas de 100 mL equipado con un termometro. Se anadio NaOH solido (54,0 mg, 1,35 mmoles) y se disolvio. Se anadio gota a gota una disolucion de acetato de vinilo (0,41 g, 10,4 mmoles) en H2O (1 mL). La temperatura se mantuvo a 25 °C y se anadio con una jeringa NaOH (270 mg, 0,68 mmoles) en H2O (1 mL). Posteriormente, se anadio mas acetato de vinilo (0,41 g, 10,4 mmoles) en H2O (1 mL) y la mezcla de reaccion se agito toda la noche a
temperatura ambiente. La reaccion se neutralizo con acido acetico, se diluyo con 50 mL de H2O y se extrajo con Et2O (2 x 100 mL). Las capas organicas se combinaron, se lavaron con salmuera (100 mL), se secaron (MgSO4), y se evaporaron en vado. El residuo se purifico por cromatograffa flash (SO 2, n-hexano/EtOAc = 20: 1^- 15: 1).
Rendimiento: 12 %, aceite amarillo. 1H RMN (CDCh): δ = 9,72 (d, J = 7,5 Hz, 1H), 7,71 (s, 1H), 7,57 (d, J = 7,7 Hz, 1H), 7,50 (d, J = 7,7 Hz, 1H), 7,40 (d, J = 15,9 Hz, 1H), 7,32 (t, J = 7,7 Hz, 1H), 6,70 (dd, J = 15,9, 7,5 Hz, 1H) ppm.
13C RMN (CDCla): δ = 193,2, 150,6, 136,1, 134,0, 131,2, 130,6, 129,7, 126,9, 123,2 ppm.
Pent-4-in-1-ol (IM-4) (JHS490)

El compuesto del tftulo se preparo segun el siguiente procedimiento:
Se disolvio acido 4-pentinoico (3,00 g, 30,6 mmoles) en THF anhidro (90 mL) y se enfrio hasta 0 °C. Se anadieron posteriormente trietilamina (4,69 mL, 33,6 mL) y metilcloroformato (2,60 mL, 33,6 mmoles) y la reaccion se agito a 0 °C durante 20 min. El precipitado blanco que aparecio se retiro por filtracion, posteriormente bajo una atmosfera de nitrogeno y el filtrado se anadio a una disolucion de NaBH4 (1,85 g, 49,9 mmoles) en agua (60 mL) a 0 °C. La mezcla resultante se agito durante 1 h a 0 °C y posteriormente durante 2 h a temperatura ambiente. La reaccion se paro cuidadosamente con HCl 1M (20 mL) hasta que ceso el burbujeo. La mezcla se extrajo con EtOAc (3 x 100 mL). Las capas organicas combinadas se lavaron con disolucion saturada de NaHCO3 (100 mL) y salmuera (100 mL), se secaron (MgSO4) y se concentraron en vado para rendir el compuesto del tftulo.
Rendimiento: 39 %, ftquido incoloro. 1H RMN (CDCl3): δ = 3,75 (t, J = 6,4 Hz, 2H), 2,31 (dt, J = 7,0, 2,7 Hz, 2H), 1,97 (t, J = 2,7 Hz, 1H), 1,85 (s, 1H), 1,77 (quin, J = 6,4 Hz, 2H) ppm,13C RMN (CDCl3): 8 = 83,8, 68,7, 61,4, 31,0, 14,9 ppm.
Pent-4-inal (IM-5) (JHS491)

El compuesto del tftulo se preparo a partir de IM-4 segun el procedimiento general mostrado para IM-7 y se uso en la siguiente etapa sin mas purificacion.
4-(terc-Butildimetilsililoxi)butan-1-ol (IM-6 ) (JHS225)

El compuesto del tftulo se preparo segun el siguiente procedimiento:
A una suspension de hidruro de potasio (1,32 g, 33,0 mmoles) en THF (40 mL) se anadio 1,4-butanodiol (3,00 g, 33,3 mmoles) a temperatura ambiente y se agito durante 45 min. Se formo un precipitado blanco voluminoso. A la mezcla de reaccion se anadio cloruro de ferc-butildimetilsililo (4,97 g, 33,0 mmoles) y la agitacion se continuo durante 45 min. La mezcla se diluyo con Et2O (200 mL), se lavo con K2CO3 acuoso (al 10 %, 50 mL) y salmuera (2 x 100 mL), se seco (MgSO4) y se concentro en vado.
Rendimiento: 99 %, aceite incoloro. 1H RMN (CDCla): δ = 3,63-3,69 (m, 4H), 2,51 (t, J = 5,6 Hz, 1H), 1,62-1,69 (m, 4H), 0,91 (s, 9H), 0,08 (s, 6 H) ppm,13C RMN (CDCh): δ = 63,3, 62,7, 30,2, 29,8, 25,9, 18,3,-5,4 ppm.
4-(terc-Butildimetilsililoxi)butanal (IM-7) (JHS227)

El compuesto del tftulo se preparo a partir de IM- 6 segun el siguiente procedimiento:
A una suspension agitada de PCC (10,6 g, 49,2 mmoles) y NaOAc (0,49 g, 9,84 mmoles) en CH2O 2 (100 mL) se anadio una disolucion del alcohol IM- 6 (6,70 g, 32,8 mmoles) en CH2O 2 (100 mL). Despues de 7 h, la mezcla de reaccion se diluyo con Et20 (100 mL) y la mezcla se filtro sobre una almohadilla corta de celite. Los disolventes se evaporaron en vado y el residuo se purifico por cromatograffa flash (SO 2, n-hexano/EtOAc = 20:1^-10:1).
Rendimiento: 30%, aceite incoloro. 1H RMN (CDCI3): 5 = 9,79 (t, J =1,7 Hz, 1H), 3,65 (t, J = 6,0 Hz, 2H), 2,51 (dt, J = 7,1, 1,7 Hz, 2H), 1,86 (quin, J = 6,5 Hz, 2H), 0,89 (s, 9H), 0,05 (s, 6 H) ppm. 13C RMN (CDCls): 5 = 202,6, 62,1, 40,8, 25,9, 25,5, 18,3, -5,4.
(E)-6-(terc-Butildimetilsililoxi)hex-2-enal (IM-8 ) (JHS228)

El compuesto del tftulo se preparo a partir de IM-7 segun el siguiente procedimiento:
A una disolucion de formilmetilentrifenilfosforano (3,01 g, 9,90 mmoles) en benceno (80 mL), se anadio una disolucion del aldehudo IM-7 (2,00 g, 9,90 mmoles) en benceno (15 mL) y la mezcla se calento a reflujo durante 24 h. Despues de eliminar el disolvente, el material crudo se purifico por cromatograffa flash (SiO2, n-hexano/EtOAc = 12:1^-10:1). Rendimiento: 58%, aceite amarillo claro. 1H RMN (CDCl3): 5 = 9,46 (d, J = 7,9 Hz, 1H), 6,84 (dd, J =15,7, 6 ,8 Hz, 1H), 6,08 (ddt, J = 15,7, 7,9, 1,5 Hz, 1H), 3,61 (t, J = 6,1 Hz, 2H), 2,34-2,41 (m, 2H), 1,63-1,72 (m, 2H), 0,84 (s, 9H), 0,00 (s, 6 H) ppm. 13C RMN (CDCla): 5 = 194,0, 158,6, 133,0, 62,0, 30,9, 29,3, 25,9, 18,3, 15,4 ppm.
Acido 3-((terc-butildimetilsilil)oxi)benzoico (IM-9) (JHS423)

El compuesto del tftulo se preparo a partir de acido 3-hidroxibenzoico segun el siguiente procedimiento:
Se anadio imidazol (1,36 g, 20,0 mmoles) a una disolucion de acido 3-hidroxibenzoico (1,00 g, 7,24 mmoles) y terc-butildimetilclorosilano (1,31 g, 8,69 mmoles) en W,W-dimetilformamida anhidra (15 mL). La mezcla se agito durante 3 h a temperatura ambiente y despues se diluyo con acetona (50 mL). Los disolventes se evaporaron en vado y el residuo se disolvio en EtOAc (50 mL). La capa organica se lavo con H2O (2 x 40 mL) y salmuera (30 mL), se seco (MgSO4), y se evaporo en vado para rendir el compuesto del tftulo.
Rendimiento: 38 %, cristales incoloros. 1H RMN (CDCla) 57,73 (td, J = 8,0, 1,1 Hz, 1H), 7,58 (t, J = 2,5 Hz, 1H), 7,34 (t, J = 8,0 Hz, 1H), 7,10 (ddd, J = 8,0, 2,5, 1,1 Hz, 1H), 1,01 (s, 9H), 0,24 (s, 6 H) ppm. 13C RMN (CDCI3) 171,7, 155,8, 130,7, 129,5, 125,7, 123,2, 121,5, 25,6, 18,2, -4,5 ppm.
(S)-3-((terc-butildimetilsilil)oxi)benzotioato de (2-acetamidoetilo) (IM-10) (JHS425)
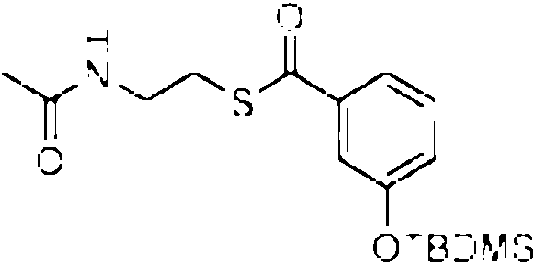
El compuesto del tftulo se preparo a partir de IM-9 segun el procedimiento general H:
Rendimiento 51 %, solido blanco. 1H RMN (CDCls) 5 = 7,57 (d, J =7,7 Hz, 1H), 7,39-7,44 (m, 1H), 7,32 (t, J = 7,70 Hz, 1H), 7,03-7,12 (m, 1H), 5,92 (br. s., 1H), 3,50-3,60 (m, 1H), 3,23 (t, J = 6,38 Hz, 1H), 1,98 (s, 3H), 1,01 (s, 9H), 0,23 (s, 6 H) ppm. 13C RMN (CDCI3) 5 = 191,6, 169,9, 155,6, 137,6, 129,3, 125,1, 119,9, 118,1, 76,8, 76,6, 76,2, 39,3, 28,2, 25,2, 22,8, -4,9 ppm.
5-(1-Hidroxietiliden)-2,2-dimetil-1,3-dioxano-4,6-diona (IM-11) (JHS197)

El compuesto del tftulo se prepare a partir de cloruro de acetilo segun el procedimiento general A: Rendimiento: 83 %, solido amarillo. 1H RMN (CDCls): δ = 15,12 (s, 1H), 2,68 (s, 3H), 1,74 (s, 6 H) ppm. 13C RMN (CDCla): δ = 194,6, 170,2, 160,4, 104,9, 91,8, 26,8, 23,5 ppm.
6-Metil-2,2-dimetil-4H-1,3-dioxin-4-ona (IM-12) (JHS198)

El compuesto del trtulo se preparo a partir de IM-11 segun el procedimiento general B:
Rendimiento: 85%, aceite amarillo. 1H RMN (CDCI3): δ = 5,19 (s, 1H), 1,94 (s, 3H), 1,64 (s, 6 H) ppm. 13C RMN (CDCI3): 8 = 168,6, 161,1, 106,2, 93,7, 24,9, 19,8 ppm.
1-(2,2-Dimetil-4-oxo-4H-1,3-dioxin-6-il)metilfosfonato de dietilo (IM-13) (JHS208)

El compuesto del tftulo se preparo a partir de IM-12 segun el procedimiento general C:
Rendimiento: 62 %, resina amarilla. 1H RMN (CDCla) 8 = 5,39 (d, J = 3,6 Hz, 1H), 4,08-4,24 (m, 4H), 2,80 (d, J = 22,1 Hz, 2H), 1,71 (s, 6 H), 1,35 (t, J =7,1 Hz, 6 H) ppm,13C RMN (75 MHz, CDCI3): δ = 163,0 (d, J = 9,7 Hz), 160,5, 107,1, 96,2 (d, J = 8,2 Hz), 62,6 (d, J = 6,7 Hz), 32,4 (d, J =137,1 Hz), 24,9, 16,3 (d, J = 6,0 Hz) ppm.
6-((1E,3E)-Hepta-1,3-dien-1-il)-2,2-dimetil-4H-1,3-dioxin-4-ona (IM-14) (JHS209)

El compuesto del tftulo se preparo a partir de IM-13 segun el procedimiento general D:
Rendimiento: 42%. 1H RMN (CDCls): δ = 6,92 (dd, J =15,2, 9,9 Hz, 1H), 5,98-6,14 (m, 2H), 5,90 (d, J =15,2 Hz, 1H), 5,21 (s, 1H), 2,16 (q, J = 7,2 Hz, 2H), 1,72 (s, 6 H), 1,46 (sxt, J = 7,4 Hz, 2H), 0,95 (t, J = 7,4 Hz, 3H) ppm,13C RMN (CDCls): δ = 163,5, 161,7, 143,4, 138,5, 128,9, 120,6, 105,9, 93,6, 34,8, 24,8, 21,8, 13,4 ppm.
5-(1-Hidroxipropiliden)-2,2-dimetil-1,3-dioxano-4,6-diona (IM-15) (JHS190D)

El compuesto del tftulo se preparo a partir de cloruro de propionilo segun el procedimiento general A:
Rendimiento: 93 %, cristales naranjas. 1H RMN (CDCI3): δ = 15,35 (br. s, 1H), 3,10 (q, J = 7,4 Hz, 2H), 1,72 (s, 6 H), 1,25 (t, J = 7,4 Hz, 3H) ppm,13C RMN (CDCI3): δ = 198,9, 170,2, 160,2, 104,8, 90,9, 29,4, 26,7, 9,6 ppm.
6-Etil-2,2-dimetil-4H-1,3-dioxin-4-ona IM-16 (MAGR35)

El compuesto del tftulo se preparo a partir de IM-15 segun el procedimiento general B:
Rendimiento: 80 %, aceite amarillo. 1H RMN (CDCl3): 8 = 5,22 (t, J =1,0 Hz, 1H), 2,24 (dq, J = 7,6, 1,0 Hz, 2H), 1,67 (s, 6H), 1,12 (t, J = 7,6 Hz, 3H) ppm. 13C RMN (CDCI3): 8 = 173,1, 161,5, 106,2, 92,2, 26,7, 24,9, 9,9 ppm.
1-(2,2-Dimetil-4-oxo-4H-1,3-dioxin-6-il)etilfosfonato de dietilo (IM-17) (JHS 195)

El compuesto del tftulo se preparo a partir de IM-16 segun el procedimiento general C:
Rendimiento: 75 %. Aceite amarillo claro. 1H RMN (CDCh): 8 = 5,37 (d, J = 3,5 Hz, 1H), 4,08-4,18 (m, 4H), 2,77 (dq, J = 23,9, 7,3 Hz, 1H), 1,69 (s, 6H), 1,30-1,45 (m, 9H) ppm,13C RMN (CDCh): 8 = 168,1 (d, J =8,2 Hz), 160,8 (d, J = 2,2 Hz), 106,8, 94,9 (d, J = 6,7 Hz), 62,1 (dd, J = 6,7, 4,5 Hz), 36,9 (d, J =137,1 Hz), 24,8 (d, J = 70,8 Hz), 16,4 (d, J = 6,0 Hz), 12,1 (d, J = 6,0 Hz) ppm.
(E)-5-(1-Hidroxi-2-metilhept-2-en-6-in-1-iliden)-2,2-dimetil-1,3-dioxano-4,6-diona (IM-18) (JHS492)

El compuesto del tftulo se preparo a partir de IM-17 segun el procedimiento general D:
Rendimiento: 53 %, aceite incoloro. 1H RMN (CDCI3) 8 = 6,40 (dt, J = 7,2, 1,1 Hz, 1H), 5,43 (s, 1H), 2,39-2,50 (m, 2H), 2,29-2,37 (m, 2H), 1,95-2,00 (m, 1H), 1,81 (s, 3H), 1,70 (s, 6H) ppm. 13C RMN (CDCI3) 8 = 165,3, 162,2, 134,9, 128,5, 106,0, 91,8, 83,0, 69,1, 27,6, 24,9, 17,9, 12,3 ppm.
2,2-Dimetil-6-((2E,4E)-octa-2,4-dien-2-il)-4H-1,3-dioxin-4-ona (IM-19) (MAGR38)
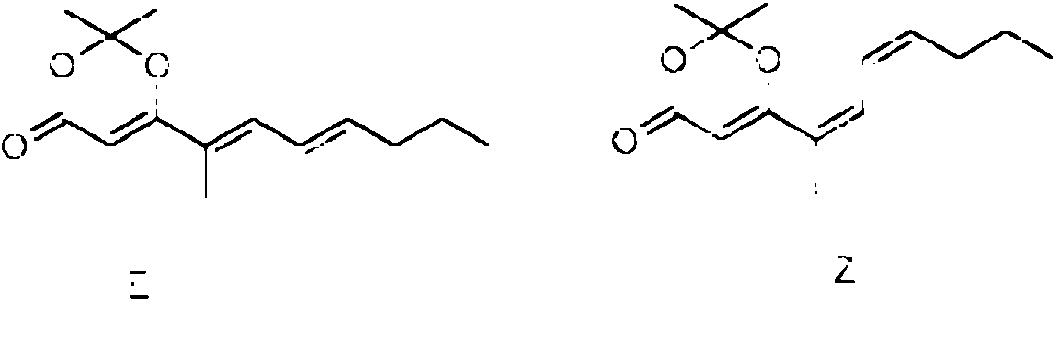
El compuesto del tftulo se preparo a partir de IM-17 segun el procedimiento general D:
Rendimiento: 42 %. 1H RMN (CDCh): 8 = 6,85 (d, J= 11,3 Hz, 1H), 6,36 (dd, J = 14,8, 11,3 Hz, 1H), 6,07 (dt, J= 14,8, 7,5 Hz, 1H), 5,42 (s, 1H), 2,15-2,20 (m, 2H), 1,86 (s, 3H) 1,68 (s, 6H) 1,39-1,48 (m, 2H), 0,90 (t, J = 7,4 Hz, 1H) ppm.
13C RMN (CDCI3): 8 = 165,7, 162,2, 143,3, 134,3, 125,9, 124,4, 105,8, 91,6, 35,3, 24,9, 22,1, 13,6, 12,1 ppm.
5-((2E,4E)-1-Hidroxi-2,5-dimetilocta-2,4-dien-1-iliden)-2,2-dimetil-1,3-dioxano-4,6-diona (IM-20) (JHS485)

El compuesto del tftulo se preparo a partir de IM-17 segun el procedimiento general D:
Rendimiento: 60 %, aceite amarillo. 1H RMN (CDCh) 8 = 7,09 (d, J= 11,8 Hz, 1H), 6,14 (dd, J= 11,8, 1,2 Hz, 1H), 5,41 (s, 1H), 2,12 (t, J= 7,5 Hz, 2H), 1,85 (s, 3H), 1,83 (s, 3H), 1,68 (s, 6H), 1,47 (sxt, J= 7,4 Hz, 2H), 0,87 (t, J = 7,4 Hz,
3H) ppm. 13C RMN (CDCI3) 5 = 166,0, 162,3, 148,5, 130,0, 124,0, 120,6, 105,7, 91,2, 42,7, 24,8, 20,9, 17,0, 13,6, 11,9 ppm.
2,2-Dimetil-6-((2E,4E)-nona-2,4-dien-2-il)-4H-1,3-dioxin-4-ona (IM-21) (MAGR40)

El compuesto del tftulo se preparo a partir de IM-17 segun el procedimiento general D:
Rendimiento: 19 %, resina amarilla. 2E,4E/2Z,4E = 14:1, 1H RMN (CDCI3) 2E,4E solo: 5 = 6,83 (d, J = 11,1 Hz, 1H), 6,36 (dd, J= 14,8, 11,3 Hz, 1H), 6,10 (dt, J = 14,8, 7,4 Hz, 1H), 5,46 (s, 1H), 2,16-2,27 (m, 2H), 1,89 (s, 3H), 1,72 (s, 6 H) 1,39-1,46 (m, 2H), 1,32-1,39 (m, 2H), 0,92 (t, J= 7,3 Hz, 1H) ppm. 13C RMN (CDCls): 5 = 165,8, 162,4, 143,7, 134,3, 125,9, 124,6, 105,9, 91,8, 33,1, 31,1, 25,0, 22,2, 13,9, 12,2 ppm.
(E)-2,2-Dimetil-6-(5-fenilpent-2-en-2-il)-4H-1,3-dioxin-4-ona (IM-22) (MAGR165)

El compuesto del tftulo se preparo a partir de IM-17 segun el procedimiento general D:
Rendimiento: 13 %, resina incolora, isomero E solo. 1H RMN (CDCla): 5 = 7,12-7,32 (m, 5H), 6,44 (t, J= 7,5 Hz, 1H), 5,40 (s, 1H), 2,76 (t, J = 7,6 Hz, 2H), 2,48-2,56 (m, 2H), 1,73 (s, 3H), 1,70 (s, 6 H) ppm. 13C RMN (CDCls): 5 = 165,6, 162,4, 141,0, 136,5, 128,4, 128,3, 127,8, 126,2, 106,0, 91,5, 35,0, 30,6, 24,9, 12,1 ppm.
2,2-Dimetil-6-((2E,4E)-5-fenilpenta-2,4-dien-2-il)-4H-1,3-dioxin-4-ona (IM-23) (JHS203)

El compuesto del tftulo se preparo a partir de IM-17 segun el procedimiento general D:
Rendimiento: 50%, aceite amarillo. 1H RMN (CDCls): 5 = 7,48-7,51 (m, 2H), 7,30-7,40 (m, 4H), 7,14 (dd, J =14,1, 11,3 Hz, 1H), 7,06 (d, J =11,3 Hz, 1H), 6,90 (d, J = 14,1 Hz, 1H), 5,54 (s, 1H) 2,02 (s, 3H), 1,75 (s, 6 H) ppm,13C RMN (CDCls): 5 = 165,3, 162,2, 139,2, 136,6, 134,0, 128,8, 127,0, 127,0, 123,7, 106,0, 92,3, 25,0, 12,5 ppm.
6-((2E,4E)-5-(3-Bromofenil)penta-2,4-dien-2-il)-2,2-dimetil-4H-1,3-dioxin-4-ona (IM-24) (JHS488)
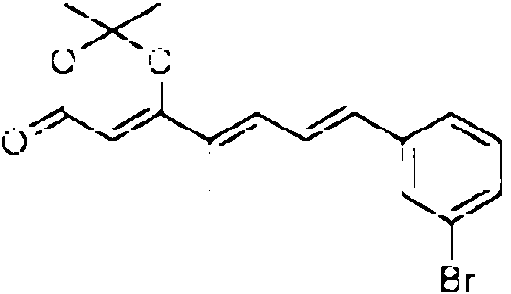
El compuesto del tftulo se preparo a partir de IM-17 segun el procedimiento general D:
Rendimiento: 50%, solido amarillo. 1H RMN (CDCls) 5 = 7,61 (t, J =1,6 Hz, 1H), 7,34-7,44 (m, 2H), 7,22 (t, J = 7,8 Hz, 1H), 7,10 (dd, J = 14,3, 11,4 Hz, 1H), 7,02 (d, J =11,4 Hz, 1H), 6,78 (d, J = 14,3 Hz, 1H), 5,54 (s, 1H), 2,02 (s, 3H), 1,74 (s, 6 H) ppm,13C RMN (CDCls): 5 = 165,0, 162,0, 138,7, 137,2, 133,3, 131,4, 130,2, 129,5, 128,0, 125,7, 125,0, 122,9, 106,0, 92,6, 25,0, 12,5 ppm.
6-((2E,4E)-5-(3-((terc-Butildimetilsilil)oxi)fenil)penta-2,4-dien-2-il)-2,2-dimetil-4H-1,3-dioxin-4-ona (IM-25) (JHS223)

El compuesto del tftulo se preparo a partir de IM-17 usando IM-2 segun el procedimiento general D:
Rendimiento: 16 %, aceite amarillo. 1H RMN (CDCls): δ = 7,22 (dd, J = 7,8, 7,8 Hz, 1H), 7,02-7,12 (m, 3 H), 6,93 (dd, J =1,9, 1,8 Hz, 1H), 6,75-6,88 (m, 2H), 5,53 (s, 1H), 2,02 (s, 3H), 1,75 (s, 6 H), 1,01 (s, 9H), 0,23 (s, 6 H) ppm,13C RMN (CDCla): δ = 165,3, 162,3, 156,1, 139,2, 138,1, 134,0, 129,7, 127,0, 123,9, 120,6, 120,2, 118,6, 106,0, 92,3, 25,7, 25,1, 18,2, 12,6, 4,4 ppm.
(4E,6E)-(S)-2-Acetamidoetil-7-(3-((terc-butildimetilsilil)oxi)fenil)-4-metil-3-oxohepta-4,6-dienotioato (IM-26) (JHS224)

El compuesto del tftulo se preparo a partir de IM-25 segun el procedimiento general E:
Rendimiento: 53%, aceite amarillo. 1H RMN (CDCls): δ = 7,11-7,30 (m, 2H), 6,96-7,11 (m, 2H), 6,70-6,96 (m, 3H), 6,11 (br. s, 1H), 4,01 (s, 2H), 3,35-3,51 (m, 2H), 3,08 (q, J = 6,1 Hz, 2H), 1,97 (s, 3H), 1,93 (s, 3H), 0,96 (s, 9H), 0,18 (s, 6 H) ppm. 13C RMN (CDCls): δ = 193,1, 193,0, 170,3, 156,0, 141,3, 137,5, 135,5, 129,8, 128,1, 124,0, 121,1, 120,5, 118,8, 52,8, 39,2, 29,2, 25,6, 23,1, 18,2, 11,8, -4,4 ppm.
6-((2E,4E)-8-((terc-Butildimetilsilil)oxi)octa-2,4-dien-2-il)-2,2-dimetil-4H-1,3-dioxin-4-ona (IM-27) (JHS240)

El compuesto del tftulo se preparo a partir de IM-17 usando IM- 8 segun el procedimiento general D:
Rendimiento: 29 %, aceite amarillo. 1H RMN (CDCls): δ = 6,83 (d, J =11,2 Hz, 1H), 6,39 (ddd, J= 14,9, 11,2, 1,0 Hz, 1H), 6,09 (td, J =14,9, 7,2 Hz, 1H), 5,44 (s, 1H), 3,61 (t, J= 6,2 Hz, 2H), 2,26 (q, J = 7,2 Hz, 2H), 1,87 (s, 3H), 1,71 (s, 6 H), 1,58-1,67 (m, 2H), 0,89 (s, 9H), 0,04 (s, 6 H). 13C RMN (CDCI3): δ = 165,4, 162,3, 143,0, 137,8, 134,2, 126,1, 124,7, 105,9, 91,8, 62,4, 32,0, 29,3, 25,9, 25,0, 18,3, 12,2, -5,3ppm.
(E)-2,2-Dimetil-6-(oct-2-en-2-il)-4H-1,3-dioxin-4-ona (IM-28) (JHS288)

El compuesto del tftulo se preparo a partir de IM-17 segun el procedimiento general D:
Rendimiento: 37%, aceite incoloro. 1H RMN (CDCls): δ = 6,41 (t, J = 7,4 Hz, 1H), 5,40 (s, 1H), 2,20 (q, J = 7,4 Hz, 2H), 1,79 (s, 3H), 1,70 (s, 6 H), 1,40-1,49 (m, 2H), 1,27-1,36 (m, 4H), 0,90 (t, J = 6,9 Hz. 3H) ppm. 13C RMN (CDCls): 8 = 165,8, 162,5, 138,1, 126,9, 105,9, 91,2, 31,6, 28,7, 28,5, 24,9, 22,4, 13,9, 12,1 ppm.
5-(1-Hidroxibutiliden)-2,2-dimetil-1,3-dioxano-4,6-diona (IM-29) (JHS192)

El compuesto del titulo se preparo a partir de cloruro de butirilo segun el procedimiento general A:
Rendimiento: 92%, aceite amarillo. 1H RMN (CDCls): δ = 15,30 (s, 1H), 3,05 (t, J = 7,5 Hz, 2H), 1,73 (s, 6 H), 1,72 (sxt, J = 7,5 Hz, 2H), 1,03 (t, J = 7,5 Hz, 3H) ppm. 13C RMN (CDCla): δ = 198,0, 170,5, 160,2, 104,7, 91,3, 37,4, 26,8, 19,6, 13,8 ppm.
6-Propil-2,2-dimetil-4H-1,3-dioxin-4-ona (IM-30) (JHS194)

El compuesto del titulo se preparO a partir de IM-29 segUn el procedimiento general B:
Rendimiento: 82 %, aceite amarillo. 1H RMN (CDCI3): δ = 5,22 (s, 1H), 2,18 (t, J = 7,5 Hz, 2H), 1,66 (s, 6 H), 1,57 (sxt, J = 7,5 Hz, 2H), 0,95 (t, J = 7,5 Hz, 3H) ppm. 13C RMN (CDCls): δ = 171,9, 161,4, 106,2, 93,1, 35,4, 24,9, 19,1, 13,4 ppm.
1-(2,2-Dimetil-4-oxo-4H-1,3-dioxin-6-il)propilfosfonato de dietilo (IM-31) (JHS 199)

El compuesto del titulo se preparO a partir de IM-30 segUn el procedimiento general C:
Rendimiento: 45 %, aceite amarillo claro. 1H RMN (acetona-d6): δ = 5,46 (d, J = 3,4 Hz, 1H), 4,08-4,23 (m, 4H), 2,83 (ddd, J = 22,9, 10,8, 4,3 Hz, 1H), 1,77-2,01 (m, 2H), 1,69 (s, 6 H), 1,26-1,34 (m, 6 H), 1,00 (t, J = 7,4 Hz, 3H) ppm,13C RMN (acetona-d6): δ = 167,5 (d, J = 8,2 Hz), 160,6 (d, J = 2,2 Hz), 107,6, 97,1 (d, J = 8,2 Hz), 63,7 (d, J = 6,7 Hz), 45,7 (d, J = 136,4 Hz), 25,3 (d, J = 7,5 Hz), 21,0 (d, J = 3,7 Hz), 16,8 (d, J = 6,0 Hz), 12,7 (d, J =15,7 Hz) ppm. 2,2-Dimetil-6-((3E,5E)-6-fenilhexa-3,5-dien-3-il)-4H-1,3-dioxin-4-ona (IM-32) (JHS241)

El compuesto del titulo se preparO a partir de IM-31 segUn el procedimiento general D:
Rendimiento: 25 %, sOlido amarillo. 1H RMN (CDCls) isOmero 3E,5E solo: δ = 7,41 (m, 2H), 7,19-7,32 (m, 3H), 7,03 (dd, J =14,8, 11,3 Hz, 1H), 6,92 (d, J =11,3 Hz, 1H), 6,82 (d, J =14,8 Hz, 1H), 5,50 (s, 1H), 2,41 (q, J = 7,6 Hz, 2H), 1,67 (s, 6 H), 1,06 (t, J = 7,6 Hz, 3H) ppm. 13C RMN (CDCls): 8=164,6, 162,4, 139,5, 136,6, 133,7, 133,5, 128,8, 127,0, 123,4, 106,0, 92,0, 25,0, 20,1, 14,4 ppm.
Acido 6 -(metoxicarbonilamino)hexanoico (IM-33) (JHS271)

El compuesto del t^tulo se preparo segun el siguiente procedimiento:
A una disolucion agitada de acido Y-aminobutftico (5,79 g, 56,1 mmoles) y KOH (3,21 g, 57,2 mmoles) en H2O (20 mL) se anadieron por partes, alternativamente metilclorofomato (4,82 mL, 62,3 mmoles) y una disolucion de KOH (57,6 mmoles) en H2O (5 mL). Despues de 2 h, la mezcla de reaccion se basifico anadiendo una disolucion de KOH (4,50 g, 80,2 mmoles) en H2O (10 mL). La disolucion se lavo con CH2CI2 (2 x 75 mL). La capa acuosa se acidifico con H2SO4 al 30 % causando la precipitacion de un solido blanco. La mezcla se extrajo con CH2CI2 (4 x 200 mL) y se seco (MgSO4). El disolvente se evaporo en vado.
Rendimiento: 88 %, aceite incoloro. 1H RMN (CDCls): δ = 10,33 (br. s, 1H), 5,88 (br. s, 0,30H, NH), 4,81 (br. s, 0,70H, NH), 3,66 (s, 3H), 3,08-3,20 (m, 2H), 2,35 (t, J =7,4 Hz, 2H), 1,60-1,70 (m, 2H), 1,47-1,56 (m, 2H), 1,31-1,41 (m, 2H) ppm. 13C RMN (CDCI3): δ = 178,9, 157,1, 52,0, 40,8, 33,8, 29,6, 26,0, 24,2 ppm.
6-(2,2-Dimetil-4,6-dioxo-1,3-dioxan-5-iliden)-6-hidroxihexilcarbamato de metilo (IM-34) (JHS273)

El compuesto del tftulo se preparo a partir de IM-33 segun el procedimiento general F:
Rendimiento: 99 %, aceite naranja. 1H RMN (CDCla): δ = 15,31 (br. s, 1H), 4,70 (br. s, 1H), 3,66 (s, 3H), 3,12-3,25 (m, 2H), 3,07 (t, J = 7,6 Hz, 2H), 1,30-1,75 (m, 6 H), 1,73 (s, 6 H) ppm. 13C RMN (CDCls): 8 = 197,8, 171,1, 157,1, 104,8, 91,3, 60,3, 52,0, 40,7, 29,6, 26,8, 26,3, 14,2 ppm.
5-(1-Hidroxi-2-metilpent-4-eniliden)-2,2-dimetil-1,3-dioxano-4,6-diona (IM-35) (JHS286)
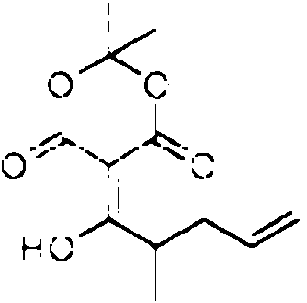
El compuesto del tftulo se preparo a partir de acido 2-metil-4-pentenoico segun el procedimiento general F:
Rendimiento: 48 %, aceite naranja. Ceto/enol = 1:61H RMN (CDCls): δ = 15,49 (br. s, 1H), 5,77 (m, 1H), 5,01-5,14 (m, 2H), 4,11-4,23 (m, 1H), 2,43-2,53 (m, 1H), 2,22-2,31 (m, 1H), 1,74 (s, 6 H), 1,24 (d, J = 6 ,8 Hz, 3H) ppm. 13C RMN (CDCls): δ = 200,9, 170,7, 160,1, 135,0, 117,3, 104,7, 91,2, 38,1, 37,6, 26,6, 17,0 ppm.
2-Metiloctanoato de etilo (IM-36) (JHS278)

El compuesto del tftulo se preparo segun el siguiente procedimiento:
El ester insaturado IM-39 (1,20 g, 6,60 mmoles) se disolvio en EtOH (50 mL). Despues de la adicion de Pd/C (al 5 %, 6 mg), la mezcla se hidrogeno en un aparato Parr a 3 bares de H2 durante 24 h. El catalizador se retiro por filtracion y la torta del filtro se lavo con EtOAc (30 mL). Los filtrados combinados se concentraron en vado. El residuo se disolvio
en CH2CI2 (50 mL) y se filtro sobre una almohadilla corta de SiO2 eluyendo con CH2CI2. El disolvente se elimino en vado.
Rendimiento: 73%, l^quido incoloro. 1H RMN (CDCl3): δ = 4,13 (q, J = 7,1 Hz, 2H), 2,42 (tq, J = 6,9, 6,5 Hz, 1H), 1,59 1,71 (m, 1H), 1,33-1,46 (m, 1H), 1,22-1,33 (m, 8 H), 1,26 (t, J = 7,1 Hz, 3H), 1,14 (d, J = 6,9 Hz, 3H), 0,88 (t, J = 6,7 Hz, 3H) ppm. 13C RMN (CDCI3): δ = 177,0, 60,0, 39,6, 33,8, 31,7, 29,2, 27,2, 22,6, 17,1, 14,3, 14,0 ppm.
Acido 2-metiloctanoico (IM-37) (JHS281)

El compuesto del tttulo se preparo segun el siguiente procedimiento:
A una disolucion del ester IM-36 (0,90 g, 4,80 mmoles) en MeOH (25 mL) se anadio una disolucion de NaOH (0,97 g, 24,2 mmoles) en H2O (8 mL). La mezcla resultante se calento a reflujo durante 16 h. Despues de enfriar hasta temperatura ambiente, el MeOH se evaporo en vado. La capa acuosa se enfrio hasta 0 °C y se acidifico hasta pH 2 por la adicion de HCl (2M). La mezcla se extrajo con EtOAc (3 x 40 mL). Las capas organicas combinadas se secaron (MgSO4) y se concentraron bajo presion reducida.
Rendimiento: 92 %, solido blanco. 1H RMN (CDCl3): δ = 10,97 (br. s, 1H), 2,45 (tq, J = 7,1, 6,9 Hz, 1H), 1,64-1,75 (m, 1H), 1,69 (quin, J = 7,1, 1H), 1,25-1,37 (m, 8 H), 1,18 (d, J = 6,9 Hz, 3H), 0,89 (t, J = 6,7 Hz, 3H) ppm. 13C RMN (CDCl3):
8 = 183,5, 39,4, 33,5, 31,7, 29,2, 27,1, 22,6, 16,8, 14,0 ppm.
5-(1-Hidroxi-2-metiloctiliden)-2,2-dimetil-1,3-dioxano-4,6-diona (IM-38) (JHS284)

El compuesto del tftulo se preparo a partir de IM-37 segun el procedimiento general F:
Rendimiento: 64 %, aceite naranja. Ceto/enol = 1:3. 1H RMN (CDCI3): δ = 15,46 (br. s, 1H), 4,03 (sxt, J = 6 ,8 Hz, 1H), 1,72 (s, 6 H), 1,17-1,50 (m, 10H), 1,20 (d, J = 6 ,8 Hz, 3H), 0,85 (t, J = 6 ,8 Hz, 3H) ppm,13C RMN (CDCI3): 8 = 202,1, 170,8, 160,1, 104,6, 91,0, 37,9, 34,0, 31,6, 29,2, 27,2, 26,8, 22,5, 17,4, 14,0 ppm.
(2E,4E)-2-metilocta-2,4-dienoato de etilo (IM-39) (JHS276)

El compuesto del tftulo se preparo segun el siguiente procedimiento:
Se disolvieron bromuro de (1-etoxicarbonilmetil)trifenilfosfonio (8 ,86 g, 20,0 mmoles) y K2CO3 (4,00 g, 28,9 mmoles) en una mezcla de CH2Cl2 (60 mL) y H2O (50 mL). Despues de la adicion gota a gota de E-2-hexen-1-al (1,50 g, 15,0 mmoles), la mezcla de reaccion se calento a reflujo toda la noche. La capa organica se lavo con H2O (2 x 50 mL), se seco (MgSO4) y se concentro en vado. El residuo se purifico por cromatograffa flash (SiO2, n-hexano/EtOAc = 10:1).
Rendimiento: 87 %, aceite amarillo claro. 2E,4E/2Z,4E = 20:1. 1H RMN (CDCI3) isomero 2E,4E solo: 8 = 7,17 (d, J = 11,2 Hz, 1H), 6,35 (tq, J =15,0, 11,2, 1,4 Hz, 1H), 6,07 (dt, J = 15,0, 7,5 Hz, 1H), 4,21 (q, J = 7,1 Hz, 2H), 2,17 (q, J = 7,3 Hz, 2H), 1,93 (s, 3H), 1,47 (sxt, J = 7,3 Hz, 2H), 1,31 (t, J = 7,1 Hz, 3H), 0,93 (t, J= 7,3 Hz, 3H) ppm. 13C RMN (CDCl3): δ = 168,7, 142,9, 138,6, 126,1, 125,1, 60,4, 35,3, 22,2, 14,3, 13,7, 12,5 ppm.
Acido (2E,4E)-2-metilocta-2,4-dienoico (IM-40) (JHS279)

El compuesto del tftulo se preparo a partir de IM-39 segun el siguiente procedimiento:
A una disolucion del ester IM-39 (1,30 g, 7,10 mmoles) en MeOH (30 mL) se anadio una disolucion de NaOH (1,32 g, 33,1 mmoles) en H2O (10 mL). La mezcla resultante se calento a reflujo durante 16 h. Despues de enfriar hasta temperatura ambiente, el MeOH se evaporo en vado. La capa acuosa se enfrio hasta 0 °C y se acidifico hasta pH 2 por la adicion de HCI (2M). La mezcla se extrajo con EtOAc (3 x 50 mL). Las capas organicas combinadas se secaron (MgSO4) y se concentraron bajo presion reducida.
Rendimiento. 91 %; solido amarillo. 2E,4E/2Z,4E = 20:1. 1H RMN (CDCI3) isomero 2E,4E solo: δ = 11,36 (br. s, 1H), 7,30 (d, J =11,3 Hz, 1H), 6,38 (dd, J = 15,1, 11,3 Hz, 1H), 6,15 (dt, J = 15,1, 7,5 Hz, 1H), 2,12-2,28 (m, 2H), 1,94 (s, 3H), 1,41-1,52 (m, 2H), 0,94 (t, J = 7,4 Hz, 3H) ppm. 13C RMN (CDCI3): δ = 174,3, 144,5, 141,1, 126,1, 124,1, 35,4, 22,1, 13,7, 12,1 ppm.
(2E,4E)-2-metil-5-fenilpenta-2,4-dienoato de etilo (IM-41) (JHS277)

El compuesto del título se preparó según el siguiente procedimiento:
Se disolvieron bromuro de (1-etoxicarbonilmetil)trifenilfosfonio (8 ,86 g, 20,0 mmoles) y K2CO3 (4,00 g, 28,9 mmoles) en una mezcla de CH2Cl2 (60 mL) y H2O (50 mL). Despues de la adicion gota a gota de E-cinamaldetudo (1,98 g, 15,0 mmoles), la mezcla de reaccion se calento a reflujo toda la noche. La capa organica se lavo con H2O (2 x 50 mL), se seco (MgSO4) y se concentro en vado. El residuo se purifico por cromatograffa flash (SiO2, n-hexano/EtOAc = 10 :1 ).
Rendimiento: 93 %, aceite amarillo claro. 2E,4E/2Z,4E = 11:1. 1H RMN (CDCh) isomero 2E,4Esolo: δ = 7,41 (m, 2H), 7,20 7,32 (m, 3H), 7,20 (d, J =11,3 Hz, 1H),m m (dd, J =15,5, 11,3 Hz, 1H), 6,80 (d, J =15,5 Hz, 1H), 4,15 (q, J= 7,1 Hz, 2H), 1,98 (s, 3H), 1,26 (t, J = 7,1 Hz, 3H) ppm. 13C RMN (CDCla): δ = 168,4, 138,9, 138,1, 136,6, 128,7, 128,6, 127,5, 127,0, 124,0, 60,6, 14,3, 12,9 ppm.
Acido (2E,4E)-2-metil-5-fenilpenta-2,4-dienoico (IM-42) (JHS280)

El compuesto del tftulo se preparo a partir de IM-41 segun el siguiente procedimiento:
A una disolucion del ester IM-41 (3,00 g, 14,8 mmoles) en MeOH (80 mL) se anadio una disolucion de NaOH (2,77 g, 69,3 mmoles) en H2O (20 mL). La mezcla resultante se calento a reflujo durante 16 h. Despues de enfriar hasta temperatura ambiente, el MeOH se evaporo en vado. La capa acuosa se enfrio hasta 0 °C y se acidifico hasta pH 2 por la adicion de HCI (2M). La mezcla se extrajo con EtOAc (3 x 100 mL). Las capas organicas combinadas se secaron (MgSO4) y se concentraron bajo presion reducida.
Rendimiento: 97 %, solido blanco. 2E,4E/2Z,AE = 11:1. 1H RMN (CDCls) isomero 2E,4E solo: δ = 11,27 (br. s, 1H), 7,40-7,44 (m, 3H), 7,23-7,32 (m, 2H), 7,22 (d, J = 11,2 Hz, 1H ),rnm (dd, J = 15,4, 11,2 Hz, 1H), 6,85 (d, J= 15,4 Hz, 1H), 1,99 (s, 3H) ppm. 13C RMN (CDCh): δ = 173,9, 140,6, 140,2, 136,4, 128,9, 128,8, 127,2, 126,4, 123,7, 12,5 ppm. (E)-3-Metilhex-2-enoato de etilo (IM-43) (JHS406)

El compuesto del tftulo se prepare segun el siguiente procedimiento:
A una disolucion de NaH (1,46 g, 60,9 mmoles) en THF (250 mL) se anadio gota a gota una disolucion de fosfonoacetato de trietilo (13,0 g, 58,1 mmoles) en THF (50 mL) a 0 °C. La mezcla resultante se dejo calentar hasta temperatura ambiente y se agito durante 30 minutos. Se anadio lentamente una disolucion de 2-pentanona (5,00 g, 58,1 mmoles) en THF (50 mL) y la reaccion se calento hasta 80 °C toda la noche. La reaccion se paro por la adicion de una disolucion saturada de NH4CI (200 mL). La mezcla resultante se extrajo con Et2O (3 x 100 mL). Las capas organicas combinadas se lavaron con H2O (100 mL) y salmuera (100 mL) y se secaron sobre MgSO4. Los disolventes se evaporaron en vado para rendir un ftquido amarillo. El material crudo se purifico por cromatograffa en columna flash (SO 2, n-hexano/EtOAc = 20:1) para rendir el compuesto del tftulo como un ftquido incoloro.
Rendimiento: 88 %, ftquido incoloro. E/Z = 3,2:1, 1H RMN (CDCI3) isomero E solo: 8 = 5,66 (q, J = 1,2 Hz, 1H), 4,14 (q, J =7,1 Hz, 2H), 2,15 (d, J = 1,3 Hz, 3H), 2,08-2,14 (m, 2H), 1,51 (sxt, J =7,4, 2H), 1,28 (t, J = 7,1 Hz, 3H), 0,91 (t, J = 7,4 Hz, 3H) ppm. 13C RMN (CDCla): δ = 166,9, 159,9, 115,6, 59,4, 42,9, 20,5, 18,6, 14,3, 13,6 ppm.
Acido (E)-3-metilhex-2-enoico (IM-44) (JHS408)

El compuesto del tftulo se prepare a partir de IM-43 segun el siguiente procedimiento:
A una disolucion del ester IM-43 (2,60 g, 16,6 mmoles) en MeOH (80 mL) se anadio una disolucion de NaOH (3,30 g, 83,2 mmoles) en H2O (20 mL). La mezcla resultante se calento a reflujo durante 2 h. Despues de enfriar hasta temperatura ambiente, el MeOH se evaporo en vado. La capa acuosa se enfrio hasta 0 °C y se acidifico hasta pH 2 por la adicion de HCI (2M). La mezcla se extrajo con EtOAc (3 x 100 mL). Las capas organicas combinadas se secaron (MgSO4) y se concentraron bajo presion reducida.
Rendimiento: 97 %, solido blanco. 1H RMN (CDCla): δ = 11,75 (br. s, 1H), 5,70 (s, 1H), 2,17 (d, J = 1,3 Hz, 3H), 2,12 2,20 (m, 2H), 1,53 (sxt, J = 7,4 Hz, 2H), 0,89-0,96 (m, 3H), 0,92 (t, J = 7,4 Hz, 3H) ppm. 13C RMN (CDCls): 8 = 172,1, 163,5, 115,0, 43,2, 20,5, 19,0, 13,6 ppm.
(E)-3-Metilhex-2-en-1 -ol (IM-45) (JHS411)

El compuesto del tftulo se prepare a partir de IM-43 segun el siguiente procedimiento:
Una suspension de LiAlH4 en Et2O anhidro se enfrio hasta 0 °C. Se anadio AICI3 y la mezcla resultante se agito durante 15 minutos. Se anadio una disolucion de IM-43 (5,00 g, 32,0 mmoles) en Et2O (15 mL) y la agitacion se continuo durante 1,5 h a 0 °C. La parada con una disolucion acuosa de NaOH (al 5 % m/m, 20 mL) dio lugar a la precipitacion de hidroxidos de aluminio. La mezcla de reaccion se filtro sobre un lecho corto de MgSO4 que se lavo con Et2O (50 mL). La eliminacion del disolvente en vado rindio el compuesto del tftulo como un ftquido incoloro.
Rendimiento: 44 %, aceite incoloro. E/Z = 4:1, 1H RMN (CDCI3) isomero E solo: 8 = 5,41 (tq, J = 6,9, 1,3 Hz, 1H), 4,07 4,20 (m, 2H), 2,00 (t, J = 7,5 Hz, 2H), 1,66 (d, J =1,3 Hz, 3H) 1,39-1,51 (m, 2H) 0,89 (t, J = 7,4 Hz, 3H) ppm. 13C RMN (CDCls): δ = 139,9, 123,3, 59,4, 41,6, 20,7, 16,1, 13,7 ppm.
(E)-3-Metilhex-2-enal (IM-46) (JHS412)

El compuesto del tftulo se prepare a partir de IM-45 segun el siguiente procedimiento:
A una suspension agitada de PCC (4,28 g, 19,9 mmoles) y NaOAc (200 mg, 3,98 mmoles) en CH2Cl2 (100 mL) se anadio una disolucion del alcohol IM-45 (1,60 g, 14,0 mmoles) en CH2Cl2 (100 mL). Despues de 7 h, la mezcla de reaccion se diluyo con Et2O (100 mL) y la mezcla se filtro sobre una almohadilla corta de celite. Los disolventes se evaporaron en vado y el residuo se purifico por cromatograffa flash (SiO2, n-hexano/EtOAc = 20 :1 ^ 10:1).
Rendimiento: 46 %, aceite incoloro. E/Z = 2:1, 1H RMN (CDCI3) isomero E solo: 8 = 9,99 (d, J = 8,0 Hz, 1H), 5,87 (dq, J = 8,0, 1,2 Hz, 1H), 2,19 (t, J =7,4 Hz, 2H), 2,15 (d, J = 1,2 Hz, 3H), 1,54 (q, J = 7,4 Hz, 2H), 0,93 (t, J =7,4 Hz, 3H) ppm. 13C RMN (CDCI3): δ = 191,3, 164,1, 127,4, 42,6, 20,3, 17,4, 13,6 ppm.
(2E,4E)-2,5-Dimetilocta-2,4-dienoato de etilo (IM-47) (JHS413)
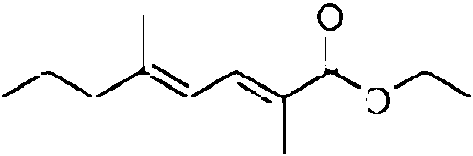
El compuesto del tftulo se preparo a partir de IM-46 segun el siguiente procedimiento:
A una suspension de NaH (0,16 g, 6,55 mmoles) en THF (30 mL) se anadio gota a gota una disolucion fosfonoacetato de trietilo (1,49 g, 6,24 mmoles) en THF (5 mL) a 0 °C. La mezcla resultante se dejo calentar hasta temperatura ambiente y se agito durante 30 minutos. Se anadio lentamente IM-46 (0,70 g, 6,24 mmoles), disuelto en THF (5 mL), y la reaccion se agito a temperatura ambiente toda la noche. La reaccion se paro por la adicion de una disolucion saturada de NH4CI (50 mL). La mezcla resultante se extrajo con Et2O (3 x 30 mL). Las capas organicas combinadas se lavaron una vez con H2O (30 mL) y salmuera (30 mL) y se secaron sobre MgSO4. Los disolventes se evaporaron en vacfo para rendir un ftquido amarillo. El residuo se purifico por cromatograffa en columna flash (SO 2, nhexano/EtOAc = 20:1) para rendir el compuesto del tftulo como un aceite incoloro.
Rendimiento: 57 %, aceite incoloro. Mezcla de regioisomeros, aprox: 2E,4E = 53 %, 2E,4Z = 26 %, 2Z,4E = 13 %, 2Z,4Z = 7%; 1H RMN (CDCls): isomeros 2E,4E y 2E,4Z solo: δ = m 58-m 67 (m, 1H), 6,17-6,21 (m, 0,35H), 6,16 (dq, J =11,8, 1,3 Hz, 0,65H), 4,22 (q, J = 7,1 Hz, 2H), 2,28 (t, J = 7,5 Hz, 0,70H), 2,14 (t, J = 7,5 Hz, 1,30H), 1,85-1,95 (m, 6 H), 1,43-1,57 (m, 2H), 1,31 (t, J = 7,1 Hz, 3H), 0,86-0,98 (m, 3H) ppm. 13C RMN (CDCla) mezcla de regioisomeros: 8 = 169,1, 169,0, 148,5, 148,3, 148,1, 134,4, 134,1, 124,6, 124,4, 121,5, 120,7, 60,4, 42,8, 39,6, 34,6, 33,1, 31,9, 30,3, 29,7, 24,7, 21,5, 21,0, 20,8, 20,6, 20,5, 17,1, 14,3, 14,2, 14,1, 14,1, 13,9, 13,7, 12,4, 12,3 ppm.
Acido (2E,4E)-2,5-dimetilocta-2,4-dienoico (IM-48) (JHS414)

El compuesto del tftulo se preparo a partir de IM-47 segun el siguiente procedimiento:
A una disolucion del ester IM-47 (0,70 g, 3,57 mmoles) en MeOH (15 mL) se anadio una disolucion de NaOH (0,70 g, 17,5 mmoles) en H2O (5 mL). La mezcla resultante se calento a reflujo durante 2 h. Despues de enfriar hasta temperatura ambiente, el MeOH se evaporo en vacfo. La capa acuosa se enfrio hasta 0 °C y se acidifico hasta pH 2 por la adicion de HCl (2M). La mezcla se extrajo con EtOAc (3 x 100 mL). Las capas organicas combinadas se secaron (MgSO4) y se concentraron bajo presion reducida.
Rendimiento: 83 %, aceite amarillo. Mezcla de regioisomeros, aprox: 2E,4E = 53 %, 2E,4Z = 26 %, 2Z,4E = 13 %, 2Z,4Z = 7 %; 1H RMN (CDCls) isomeros 2E,4Ey 2E,4Z solo: δ = ^ 44 - ^ 51 (m, 1H), 6,12-6,18 (m, 0,35H), 6,12 (dq, J =11,9, 1,2 Hz, 0,65H), 4,22 (q, J =7,1 Hz, 2H), 2,30 (t, J = 7,5 Hz, 0,70H), 2,17 (t, J = 7,5 Hz, 1,30H), 1,85-1,95 (m, 6 H), 1,44-1,59 (m, 2H), 0,86-1,03 (m, 3H) ppm. 13C RMN (CDCls) mezcla de regioisomeros: 8 = 174,6, 174,5, 173,8, 152,3, 150,2, 149,9, 136,8, 136,5, 124,7, 123,5, 123,4, 121,6, 120,8, 42,8, 39,6, 34,7, 33,2, 24,8, 21,6, 21,0, 20,6, 20,5, 20,5, 17,2, 13,9, 13,7, 12,0, 11,9 ppm.
Smtesis y datos espectroscopicos de los compuestos precursores
(S)-2-Acetamidoetil-3-oxobutanotioato (1) (MAGR169)

El compuesto del tftulo se preparo a partir de IM-11 segun el procedimiento general G:
Rendimiento 43 %. Ceto/enol = 3:1 1H RMN (CDCls): δ = 12,56 (s, 0,25H, enol C=COH), 6,12 (s, 1H, NH), 5,45 (s, 0,25H, enol CO=CH), 3,70 (s, 1,50H, ceto COCH2CO), 3,41-3,47 (m, 2H, CH2N), 3,04-3,10 (m, 2H, CH2S), 2,25 (s,
2,25H, ceto COCH3), 1,93-1,96 (m, 3,75H, enol COHCH3 + NHCOCH3) ppm,13C RMN (CDCI3): 5 = 199,8, 194,2, 192,2, 174,0, 170,5, 170,3, 99,8, 58,0, 39,8, 39,1, 30,4, 29,1, 27,7, 23,1, 23,1, 21,0 ppm.
(S)-2-Acetamidoetil-3-oxohexanotioato (2) (MAGR170 / JHS 201)

0
El compuesto del tftulo se preparo a partir de IM-29 segun el procedimiento general G:
Rendimiento 41 %. Ceto/enol = 3:1 1H RMN (CDCI3): 5 = 12,60 (s, 0,25H, enol C=COH), 5,94 (s, 1H, NH), 5,47 (s, 0,25H, enol CO=CH), 3,70 (s, 1,50H, ceto COCH2CO), 3,44-3,52 (m, 2H, CH2N), 3,07-3,13 (m, 2H, CH2S), 2,53 (t, J = 7,2 Hz, 1,50H, ceto COCH2), 2,17 (t, J = 7,5 Hz, 0,50H, enol COHCH2), 1,99 (s, 3H, COCH3), 1,58-1,70 (m, 2H, CH2CH3), 0,92-0,99 (m, 3H, CH2CH3) ppm. 13C RMN (CDCl3): 5 = 202,1, 194,3, 192,4, 177,4, 170,4, 170,3, 99,2, 57,2, 45,3, 39,9, 39,2, 36,8, 35,8, 29,2, 27,8, 23,2, 23,1, 19,6, 19,5, 16,9, 13,6, 13,5 ppm.
(S)-2-Acetamidoetil-(£)-4-metil-3-oxonon-4-en-8-intioato (3) (JHS493)

El compuesto del titulo se preparo a partir de IM-18 segun el procedimiento general G:
Rendimiento 50 %. Ceto/enol = 3:1. 1H RMN (CDCI3) ceto solo: 5 = 6,62-6,72 (m, 1H), 6,08 (br. s, 1H), 3,98 (s, 2H), 3,47 (t, J = 6,2 Hz, 2H), 3,10 (q, J = 6,2 Hz, 2H), 2,28-2,57 (m, 4H), 2,01 (t, J = 2,6 Hz, 1H), 1,97 (s, 3H), 1,82 (s, 3H) ppm. 13C RMN (CDCI3): 5 = 193,3, 193,1, 170,5, 143,0, 138,1, 82,6, 69,5, 52,6, 39,2, 29,2, 28,1,23,2, 17,6, 11,5 ppm. (4E,6E)-(S)-2-Acetamidoetil-3-oxodeca-4,6-dienotioato (4) (JHS 210)

El compuesto del tftulo se preparo a partir de IM-14 segun el procedimiento general E:
Rendimiento 50 %. Polvo naranja. Ceto/enol = 1:21H RMN (acetona-d6) enol 4E,6E solo: 5 = 12,46 (s, 1H), 7,36 (br. s, 1H), 7,13 (dd, J =15,2, 10,7, 1H), 6,27-6,35 (m, 1H), 6,11-6,20 (m, 1H), 5,94 (d, J =15,2 Hz, 1H), 5,65 (s, 1H), 3,37 (q, J = 6,6 Hz, 2H), 3,08 (t, J = 6,6 Hz, 2H), 2,12-2,22 (m, 2H), 1,87 (s, 3H), 1,41-1,51 (m, 2H), 0,92 (t, J = 7,5 Hz, 3H) ppm. 13C RMN (acetona-d6): 5 = 195,1, 192,3, 168,5, 143,8, 140,5, 130,5, 123,6, 101,3, 39,9, 35,8, 28,8, 22,9, 22,8, 14,0 ppm.
(4E,6E)-(S)-2-Acetamidoetil-4-metil-3-oxodeca-4,6-dienotioato (5) (MAGR41 / MAGR168)

El compuesto del titulo se preparo a partir de IM-19 segun el procedimiento general E:
Rendimiento 25 %. Ceto/enol = 4:1 4E,6E/4Z,6E = 8:1. 1H RMN (DMSO-d6) isomero ceto 4E,6E solo: 5 = 8,03 (br. s, 1H), 7,21 (d, J = 11,0 Hz, 1H), 6,48-6,54 (m, 1H), 6,30 (dt, J = 15,1, 7,5 Hz, 1H), 4,15 (s, 2H), 3,17 (m, 2H), 2,92 (t, J = 6,9 Hz, 2H), 2,14-2,22 (m, 2H), 1,77-1,79 (m, 6H), 1,40-1,48 (m, 2H), 0,90 (t, J = 7,5 Hz, 3H) ppm. 13C RMN (DMSO-d6): 5 = 193,8, 193,0, 169,4, 145,6, 141,9, 132,9, 126,8, 52,4, 38,1, 34,8, 28,5, 22,5, 21,6, 13,6, 11,3 ppm.
(S)-2-Acetamidoetil-(4E,6E)-4,7-dimetil-3-oxodeca-4,6-dienotioato (6) (JHS486)
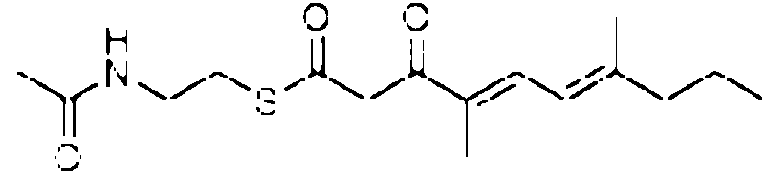
El compuesto del tftulo se prepare a partir de IM-20 segun el procedimiento general E:
Rendimiento 65 %. Ceto/enol = 4:1, 4£, 6E/4Z, 6E = 2:1. 1H RMN (acetona-d6) isomero ceto 4E,6E solo: 8 = 7,49 (dd, J =11,5, 1,2 Hz, 1H), 7,29 (br. s, 1H), 6,27-6,37 (m, 1H), 4,12 (s, 2H), 3,32 (q, J = 6,8 Hz, 2H), 3,02 (t, J = 6,8 Hz, 2H), 2,15-2,25 (m, 2H), 1,95 (s, 3H), 1,85 (s, 3H), 1,84 (s, 3H), 1,53 (sxt, J = 7,4 Hz, 2H), 0,91 (t, J =7,4 Hz, 3H) ppm. 13C RMN (acetona-d6) 8194,0, 193,5, 170,2, 151,7, 138,2, 133,9, 122,1, 53,9, 43,5, 39,6, 29,8, 22,9, 21,7, 17,5, 14,1, 11,4 ppm.
(4E,6E)-(S)-2-Acetamidoetil-4-metil-3-oxoundeca-4,6-dienotioato (7) (MAGR42)

El compuesto del tftulo se prepare a partir de IM-21 segun el procedimiento general E:
Rendimiento 27 %. Ceto/enol = 5,6:1, 4E,6E/4Z,6E = 15:1, 1H RMN (DMSO-d6) isomero 4E,6E solo: 8 = 12,80 (s, 0,15H, enol C=COH), 8,07 (br. s, 0,15H enol NH), 8,03 (br. s, 0,85H ceto NH), 7,22(d, J =11,0 Hz, 0,85H, ceto CH=CMe), 7,05 (d, J= 11,9 Hz, 0,15H, enol CH=CMe), 6,48-6,54 (m, 1H, CH=CH), 6,30 (dt, J = 15,1, 7,6 Hz, 0,85H, ceto CHCH2), 6,16 (dt, J = 14,8, 7,5 Hz, 0,15H, enol CHCH2), 5,87 (s, 0,15H, enol CO=CH), 4,15 (s, 1,70H, ceto COCH2), 3,19-3,25 (m, 0,30H, enol CH2N), 3,11-3,19 (m, 1,70H, ceto CH2N), 3,00 (t, J = 7,0 Hz, 0,30H, enol CH2S), 2,92 (t, J = 6,9 Hz, 1,70H, ceto CH2S), 2,18-2,25 (m, 2H, C=CCH2), 1,78-1,79 (m, 6H, C=CCH y COCH3), 1,28-1,44 (m, 4H, CH2CH2), 0,90 (t, J = 7,6 Hz, 3H, CH2CH3) ppm. 13C RMN (DMSO-d6): isomero ceto 4E,6E solo: 8 = 193,7, 192,9, 169,2, 145,7, 141,8, 132,8, 126,6, 52,4, 38,1, 32,4, 30,4, 28,4, 22,5, 21,7, 13,7, 11,2 ppm.
(E)-(S)-2-Acetamidoetil-4-metil-3-oxo-7-fenilhept-4-enotioato (8) (MAGR171)

El compuesto del tftulo se preparo a partir de IM-22 segun el procedimiento general E:
Rendimiento 62 %. Ceto/enol = 3:1, 1H RMN (acetona-d6): 8 = 12,84 (s, 0,25H, enol C=COH), 7,47 (br. s, 1H, NH), 7,21-7,32 (m, 4H, Ar-H), 7,16-7,21 (m, 1H, Ar-H), 6,85 (t, J = 7,2 Hz, 0,75H, ceto CMe=CH), 6,67 (t, J= 7,4 Hz, 0,25H, enol CMe=CH), 5,73 (s, 0,25H, COCH), 4,04 (s, 1,50H, ceto COCH2), 3,31-3,42 (m, 2H, CH2N), 3,08 (t, J = 6,8 Hz, 0,50H, ceto CH2S), 3,02 (t, J = 6,9 Hz, 1,50H, enol CH2S), 2,75-2,84 (m, 2 H, CH2Ph), 2,53-2,65 (m, 2H, CH2CH=C), 1,90 (s, 0,75H, enol COCH3), 1,90 (s, 2,25H, ceto COCH3), 1,73 (s, 0,75H, enol C=CCHa), 1,67 (s, 2,25H, ceto C=CCHa) ppm. 13C RMN (acetona-d6): ceto solo: 8 = 194,1, 193,4, 171,2, 145,7, 145,6, 142,2, 138,0, 129,4, 129,3, 127,0, 53,4, 39,7, 35,1, 32,0, 22,9, 11,3 ppm.
(4E,6E)-(S)-2-Acetamidoetil-4-metil-3-oxo-7-fenilhepta-4,6-dienotioato (9) (JHS206)

El compuesto del tftulo se preparo a partir de IM-23 segun el procedimiento general E:
Rendimiento 60 %, solido amarillo. Ceto/enol = 2:1, 1H RMN (acetona-d6): 8 = 12,82 (s, 0,35H, enol C=COH), 7,99 (br. s, 1H, NH), 7,60-7,66 (m, 2H), 7,26-7,52 (m, 5H), 7,13 (d, J = 14,7 Hz, 0,65H, ceto CMe=CH), 6,99 (d, J = 13,9 Hz, 0,35H, enol CMe=CH), 5,88 (s, 0,35H, enol CO=CH), 4,19 (s, 1,30H, ceto COCH2), 3,37-3,47 (m, 2H, CH2N), 3,13 (t, J = 6,8 Hz, 0,70H, ceto CH2S), 3,06 (t, J = 6,7 Hz, 1,30H, enol CH2S), 1,98-1,99 (m, 6H, COCH3, C =C C ^) ppm,13C
RMN (acetona-da): δ = 194,4, 194,2, 173,3, 173,2, 142,6, 142,1, 139,8, 137,9, 137,5, 136,3, 135,8, 130,0, 129,8, 129,7, 129,5, 128,4, 128,1, 125,3, 125,3, 98,4, 53,3, 40,4, 39,9, 29,3, 28,3, 22,7, 20,9, 12,6, 11,9 ppm.
(S)-2-Acetamidoetil-(4£,6£)-7-(3-bromofenil)-4-metil-3-oxohepta-4,6-dienotioato (10) (JHS489)

El compuesto del titulo se preparo a partir de IM-24 segun el procedimiento general E:
Rendimiento 40 %, solido amarillo. Ceto/enol = 2:1, 1H RMN (CDCl3) ceto solo: 8 = 7,22 (t, J = 7,9 Hz, 1H), 7,07-7,15 (m, 1H), 6,89 (d, J = 14,8 Hz, 1H), 6,07 (br. s, 1H), 4,05 (s, 1H), 3,48 (quin, J = 6 , 6 Hz, 2H), 3,11 (t, J = 6 , 6 Hz, 2H), 2,02 (d, J =1,0 Hz, 3H), 1,97 (s, 3H) ppm,13C RMN (CDCl3): δ = 193,1, 193,0, 170,4, 140,5, 139,4, 138,9, 136,4, 130,2, 129,9, 126,0, 125,7, 125,2, 123,0, 52,9, 39,2, 29,2, 23,1, 11,9 ppm.
(4£,6£)-(S)-2-Acetamidoetil-7-(3-hidroxifenil)-4-metil-3-oxohepta-4,6-dienotioato (11) (JHS226)

El compuesto del tftulo se preparo a partir de IM-26 segun el siguiente procedimiento:
A una disolucion de IM-26 (0,08 g, 0,17 mmoles) en THF (1,50 mL) se anadio una disolucion de nBu4NF (disolucion de THF 1,0 M, 0,21 mL, 0,21 mmoles) a 0 °C. Despues de agitar durante 6 h a temperatura ambiente, la reaccion se paro por la adicion de NH4CI acuoso saturado (10 mL). Los productos crudos se extrajeron con EtOAc (3 x 10 mL), y las capas organicas combinadas se lavaron con salmuera, se secaron (MgSO4), y se concentraron en vado. El residuo se purifico por cromatograffa en columna flash (SO 2, n-hexano/EtOAc = 1:1^- EtOAc) para rendir el compuesto del tftulo.
Rendimiento 92%, solido amarillo. 1H RMN (DMSO-d6) ceto solo: δ = 9,51 (s, 1H), 8,03 (t, J = 5,30 Hz, 1H), 7,13-7,44 (m, 3H), 6,86-7,13 (m, 3H), 6,68-6,80 (m, 1H), 4,20 (s, 2H), 3,18 (q, J = 6,70 Hz, 2H), 2,94 (t, J = 6,70 Hz, 2H), 1,92 (s, 3H), 1,79 (s, 3H) ppm,13C RMN (DMSO-d6): δ = 193,5, 192,9, 169,3, 157,7, 141,6, 141,2, 137,4, 134,8, 129,8, 124,4, 118,5, 116,4, 113,9, 52,5, 38,1, 28,5, 22,5, 11,5 ppm.
(4E,6E)-(S)-2-Acetamidoetil-10-((terc-butildimetilsilil)oxi)-4-metil-3-oxodeca-4,6-dienotioato (12) (JHS247B)

El compuesto del tftulo se preparo a partir de IM-27 segun el procedimiento general E:
Rendimiento 99 %. Ceto/enol = 3:1 1H RMN (acetona-d6): δ = 12,86 (s, 0,25H, enol C=COH), 7,40-7,60 (m, 1H, NH), 7,22 (d, J = 10,7 Hz, 0,75H, ceto CH=CMe), 7,11 (d, J =11,3 Hz, 0,25H, enol CH=CMe), 6,84 (m, 0,25H), 6,48-6,61 (m, 0,75H, CH=CH), 6,32 (dt, J = 15,0, 7,3 Hz, 0,75H, ceto CHCH2), 6,13 (dt, J = 14,8, 7,2 Hz, 0,25H, enol CHCH2), 5,81 (s, 0,25H, enol CO=CH), 4,09 (s, 1,50H, ceto COCH2), 3,66-3,71 (m, 2H, CH2O), 3,32-3,43 (m, 2H, CH2N), 3,00 3,12 (m, 2H, CH2S), 1,91-1,60 (m, 8 H), 0,90 (s, 9H), 0,06 (s, 6 H) ppm. 13C RMN (acetona-d6): 8 = 194,2, 193,6, 171,3, 171,0, 146,0, 142,5, 134,4, 127,7, 62,9, 60,6, 53,4, 39,7, 32,9, 32,8, 32,4, 26,4, 22,8, 20,9, 18,9, 14,6, 11,6, -5,1 ppm.
(4E,6E)-(S)-2-Acetamidoetil-10-hidroxi-4-metil-3-oxodeca-4,6-dienotioato (13) (JHS266)

El compuesto del tftulo se prepare a partir de 12 segun el siguiente procedimiento:
A una disolucion de 12 (0,17 g, 0,40 mmoles) en THF (4,0 mL) se anadio una disolucion de nBu4NF (disolucion de THF 1,0M, 1,0 mL, 1,00 mmol) a 0 °C. Despues de agitar durante 6 h a temperatura ambiente, la reaccion se paro por la adicion de NH4Cl acuoso saturado (30 mL). Los productos crudos se extrajeron con EtOAc (3 x 30 mL), y las capas organicas combinadas se lavaron con salmuera, se secaron (MgSO4), y se concentraron en vado. El residuo se purifico por cromatograffa en columna flash (SiO2, n-hexano/EtOAc = 1:1^- EtOAc) para rendir el compuesto del tftulo.
Rendimiento 96 %, aceite amarillo. Ceto/enol = 3:1. 1H RMN (acetona-d6): 8 = 12,75 (s, 0,25H, enol C=COH), 7,92 8,01 (m, 1H, NH), 7,22 (d, J =10,7 Hz, 0,75H, ceto CH=CMe), 7,05 (d, J =11,3 Hz, 0,25H, enol CH=CMe), 6,46-6,60 (m, 1H, CH=CH), 6,32 (dt, J = 15,0, 7,3 Hz, 0,75H, ceto CHCH2), 6,13 (dt, J =14,8, 7,2 Hz, 0,25H, enol CHCH2), 5,77 (s, 0,25H, enol CO=CH), 4,09 (s, 1,50H, ceto COCH2), 3,57 (t, J = 6,5 Hz, 2H, CH2OH), 3,30-3,42 (m, 2H, CH2N), 3,08 (t, J = 6 , 8 Hz, 0,50H, enol CH2S), 3,01 (t, J = 6,7 Hz, 1,50H, cetoCH2S), 2,21-2,31 (m, 2H, C=CCH2), 1,95 (s, 0,75H, enol COCH3), 1,94 (s, 2,25H, ceto COCH3), 1,84 (s, 0,75H, enol C=CCH3), 1,79 (s, 2,25H, ceto C=CCH3), 1,60-1,69 (m, 2H, CH2CH2OH) ppm. 13C RMN (acetona-d6): δ = 194,7, 194,4, 173,5, 173,4, 146,6, 143,7, 143,1, 136,1, 134,2, 127,5, 127,3, 62,1, 53,1, 40,4, 40,3, 39,9, 38,6, 32,8, 32,5, 30,7, 28,1, 22,7, 12,3, 11,6 ppm.
(E)-(S)-2-Acetamidoetil-4-metil-3-oxodec-4-enotioato (14) (JHS 291)

El compuesto del titulo se preparo a partir de IM-28 segun el procedimiento general E:
Rendimiento 66 %, aceite amarillo. Ceto/enol = 3:1 1H RMN (acetona-d6) isomero ceto 4E,6E solo: 8 = 7,35 (br. s, 1H, NH), 6,81 (t, J = 7,3 Hz, 1H), 4,06 (s, 2H), 3,30-3,41 (m, 2H), 3,01 (t, J = 6,7 Hz, 2H), 2,21-2,34 (m, 2H), 2,53-2,65 (m, 2H), 1,87 (s, 3H), 1,75 (s, 3H), 1,42-1,55 (m, 2H), 1,30-1,37 (m, 4H), 0,89 (t, J = 7,0 Hz. 3H) ppm. 1H RMN (acetonad6): δ = 194,1, 193,4, 170,6, 146,9, 137,5, 53,5, 39,6, 32,3, 29,8, 29,6, 28,9, 23,2, 22,9, 14,3, 11,4 ppm.
(4E,6E)-(S)-2-Acetamidoetil-4-etil-3-oxo-7-fenilhepta-4,6-dienotioato (15) (JHS246)

El compuesto del tftulo se preparo a partir de IM-32 segun el procedimiento general E:
Rendimiento 88 %, resina amarilla. Ceto/enol = 2:1, 4E,6E/4Z,6E = 10:1. 1H RMN (acetona-d6) isomero 4E,6E solo: 8 = 12,99 (br. s, 0,35H, enol C=COH), 7,58-7,69 (m, 2H), 7,24-7,57 (m, 6 H), 7,08-7,18 (m, 0,65H). 7,00 (d, J = 14,5 Hz, 0,35H), 5,95 (s, 0,35H, enol CO=CH), 4,19 (s, 1,30H, ceto COCH2), 3,33-3,45 (m, 2H, CH2N), 3,12 (t, J = 6 , 8 Hz, 0,70H, enol CH2S), 3,05 (t, J = 6,7 Hz, 1,30H, ceto CH2S), 2,56 (q, J = 7,5 Hz, 0,70H, enol CH2CH3), 2,55 (q, J = 7,5 Hz, 1,30H, ceto CH2CH3), 1,92 (s, 1,05H, enol COCH3), 1,90 (s, 1,95H, ceto COCH3), 1,11 (s, 1,05H, enol CH2CH3), 1,00 (s, 1,95H, ceto CH2CH3) ppm,13C RMN (acetona-d6): δ = 193,6, 193,5, 171,0, 142,7, 142,1, 142,0, 140,1, 137,9, 135,4, 130,0, 129,8, 129,7, 129,6, 128,4, 128,1, 125,0, 124,9, 98,2, 53,6, 40,0, 39,7, 29,6, 28,8, 22,9, 20,3, 19,8, 15,2, 14,7 ppm.
(S)-2-Acetamidoetil-8-(metoxicarbonilamino)-3-oxooctanotioato (16) (JHS 274)

El compuesto del tftulo se preparo a partir de IM-34 segun el procedimiento general G:
Rendimiento 56 %, solido blanco. Ceto/enol = 5:1 1H RMN (CDCh) ceto solo: 8 = 6,25 (br. s, 1H), 4,90 (br. s, 1H, NHCOOCH3), 3,62 (s, 2H), 3,58 (s, 3H), 3,37 (q, J = 6,2 Hz, 2H), 3,09 (q, J = 6,4 Hz, 2H), 3,02 (t, J = 6,4 Hz, 2H), 2,48 (t, J =7,2 Hz, 2H), 1,91 (s, 3H), 1,38-1,59 (m, 4H), 1,19-1,30 (m, 2H) ppm. 13C RMN (CDCh): 8 = 202,0, 192,2, 170,6, 157,2, 99,3, 57,3, 52,0, 43,1, 39,1, 29,7, 29,2, 25,9, 23,1, 22,8 ppm.
(S)-2-Acetamidoetil-4-metil-3-oxohept-6-enotioato (17) (JHS287)

El compuesto del titulo se prepare a partir de IM-35 segun el procedimiento general G:
Rendimiento 49 %, aceite amarillo. Ceto/enol = 2,3:1, 1H RMN (acetona-d6) ceto solo: 8 = 7,63 (br. s, 1H), 5,69-5,83 (m, 1H), 5,00-5,10 (m, 2H), 3,89 (s, 2H), 3,32-3,41 (m, 2H), 3,04 (t, J = 6,7 Hz, 2H), 2,78 (sxt, J = 7,0, 1H), 2,31-2,42 (m, 1H), 2,05-2,22 (m, 1H), 1,91 (s, 3H), 1,08 (d, J = 7,0 Hz, 3H) ppm. 13C RMN (acetona-da): 8 = 205,7, 192,8, 171,3, 136,5, 117,3, 56,4, 46,8, 39,7, 37,3, 29,5, 22,9, 15,8 ppm.
(S)-2-Acetamidoetil-4-metil-3-oxodecanotioato (18) (JHS 285)
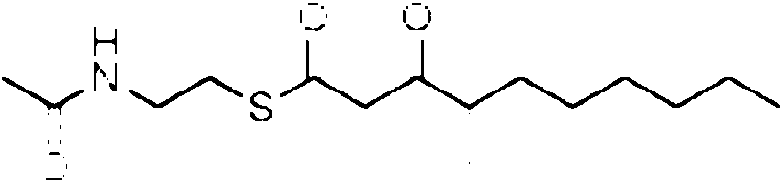
El compuesto del titulo se prepare a partir de IM-38 segUn el procedimiento general G:
Rendimiento 75 %, resina amarilla. Ceto/enol = 3:1 1H RMN (acetona-d6) ceto solo: 8 = 7,60 (br. s, 1H), 3,87 (s, 2H), 3,35 (quin, J = 6,8 Hz, 2H), 3,05 (q, J = 6,8 Hz, 2H), 2,70 (quin, J = 6,8 Hz, 1H), 1,91 (s, 3H), 1,25-1,39 (m, 10H), 1,07 (d, J = 6,8 Hz, 3H), 0,87 (t, J = 6,8 Hz, 3H) ppm. 13C RMN (acetona-d6): 8 = 206,0, 193,0, 172,3, 56,2, 47,3, 39,8, 33,2, 32,4, 30,0, 29,3, 27,7, 23,3, 22,8, 16,1, 14,4 ppm.
(2E,4E)-(S)-2-Acetamidoetil-2-metilocta-2,4-dienotioato (19) (JHS282)
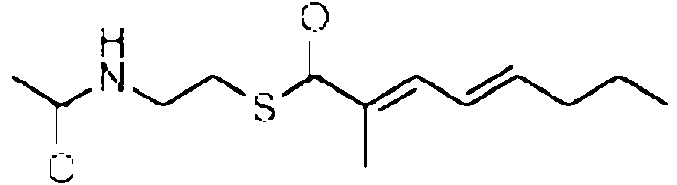
El compuesto del titulo se prepare a partir de IM-40 segUn el procedimiento general H:
Rendimiento 30 %, aceite amarillo. 1H RMN (acetona-d6): 8 = 7,69 (br. s, 1H), 7,16 (d, J =10,9 Hz, 1H), 6,46 (d, J = 15,1, 10,9 Hz, 1H), 6,24 (d, J = 15,1, 7,2 Hz, 1H), 3,31-3,39 (m, 2H), 3,05 (t, J = 6,8 Hz, 2H), 2,18-2,24 (m, 2H), 1,94 (s, 3H), 1,93 (s, 3H), 1,44-1,52 (m, 2H), 0,92 (t, J = 7,4 Hz, 3H) ppm,13C RMN (acetona-d6): 8 = 193,0, 171,3, 145,6, 138,4, 133,5, 126,7, 40,0, 36,0, 28,9, 22,9, 22,8, 14,0, 12,7 ppm.
(2E,4E)-(S)-2-Acetamidoetil-2-metil-5-fenilpenta-2,4-dienotioato (20) (JHS283)

El compuesto del titulo se prepare a partir de IM-42 segUn el procedimiento general H:
Rendimiento 43 %, aceite amarillo. 2E,4E/2Z,4E = 14:1, 1H RMN (acetona-d6) isomero 4E,6E solo: 8 = 7,74 (t, J =5,1 Hz, 1H), 7,60-7,63 (m, 2H), 7,28-7,40 (m, 4H), 7,28 (dd, J = 14,6, 11,0 Hz, 1H), 7,05 (d, J = 14,6 Hz, 1H), 3,38-3,44 (m, 2H), 3,11 (t, J = 6,7 Hz, 2H), 2,08 (s, 3H) ppm,13C RMN (acetona-d6): 8 =192,9, 170,9, 141,5, 138,2, 137,6, 135,7, 129,9, 129,7, 128,3, 124,6, 39,9, 29,2, 22,9, 13,0 ppm.
(S)-2-Acetamidoetil butanotioato (21) (JHS401)

El compuesto del tftulo se prepare a partir de cloruro de butirilo segun el procedimiento general H:
Rendimiento 81 %, aceite amarillo. 1H RMN (acetona-da) 8 = 7,36 (br. s, 1H), 3,28-3,34 (m, 2H), 2,98 (t, J = 6,9 Hz, 2H), 2,55 (t, J =7,4 Hz, 2H), 1,87 (s, 3H), 1,59-1,70 (m, 2 H), 0,92 (t, J = 7,4 Hz, 3H) ppm. 13C RMN (acetona-da) 8 = 199,1, 170,6, 46,4, 39,8, 29,0, 22,9, 19,8, 13,7 ppm.
(S)-2-Acetamidoetil-(£)-3-metilhex-2-enotioato (22) (JHS410)

El compuesto del tftulo se prepare a partir de IM-44 segun el procedimiento general H:
Rendimiento 66 %, aceite amarillo claro. E/Z = 3:1, 1H RMN (acetona-d6): isomero E solo: 8 = 7,29 (br. s, 1H), 6,01 (q, J =1,2 Hz, 1H), 3,30-3,35 (m, 2H), 3,01 (t, J = 6,9 Hz, 2 H), 2,13-2,17 (m, 2H), 2,13 (d, J =1,2 Hz, 3H), 1,87 (s, 3H), 1,48-1,57 (m, 2H), 0,91 (t, J= 7,6 Hz, 3H) ppm. 13C RMN (acetona-d6) 8 = 189,0, 170,3, 158,5, 123,4, 43,2, 40,0, 29,1, 22,9, 21,4, 19,7, 14,0 ppm.
(S)-2-Acetamidoetil-(2E,4E)-2,5-dimetilocta-2,4-dienotioato (23) (JHS415)

El compuesto del tftulo se prepare a partir de IM-48 segun el procedimiento general H:
Rendimiento 43 %, aceite amarillo. Mezcla de regioisomeros, aprox: 2E,4E = 53 %, 2E,4Z = 26 %, 2Z,4E = 13 %, 2Z,4Z= 7 %; 1H RMN (acetona-d6) isomeros 2E,4Ey 2E,4Z solo: 8 = ^ 58 - ^ 67 (m, 2H, C=CH NH), 6,25-6,29 (m, 0,35H, C=CH), 6,22 (dq, J =11,5, 1,3 Hz, 0,65H, C=CH), 3,29-3,37 (m, 2H, CH2), 3,01-3,06 (m, 2H), 2,30 (t, J= 7,5 Hz, 0,70H, CH2C=C), 2,18 (t, J = 7,5 Hz, 1,30H, CH2C=C), 1,85-1,95 (m, 9H, CHaC=C CH3CO), 1,45-1,57 (m, 2H, CH2CH3), 0,84-0,94 (m, 3H, CH2CH3) ppm. 13C RMN (acetona-d6) mezcla de regioisomeros: 8 = 192,9, 192,9, 170,4, 151,1, 150,9, 147,5, 144,5, 134,0, 133,8, 133,2, 133,0, 122,1, 121,3, 43,3, 40,2, 39,9, 39,7, 35,3, 33,9, 29,1, 24,9, 22,9, 22,3, 21,7, 21,3, 21,2, 21,0, 20,9, 17,4, 14,5, 14,2, 14,0, 12,6, 12,5 ppm.
(S)-2-Acetamidoetil benzotioato (24) (JHS417)

El compuesto del tftulo se prepare a partir de acido benzoico segun el procedimiento general H:
Rendimiento 75 %, resina blanca. 1H RMN (acetona-d6) 8 = 7,92-8,00 (m, 2H), 7,67 (tt, J= 7,4, 1,3 Hz, 1H), 7,54 (tt, J = 7,4, 1,3, 2H), 7,38 (br. s, 1H), 3,36-3,50 (m, 2H), 3,20 (t, J = 6,9 Hz, 2H), 1,88 (s, 3H) ppm. 13C RMN (acetonad6) 8 = 191,8, 170,3, 138,0, 134,6, 129,8, 127,9, 39,7, 22,9 ppm.
(S)-2-Acetamidoetil-3-hidroxibenzotioato (25) (JHS427)

El compuesto del tftulo se prepare a partir de acido 3-hidroxibezoico segun el procedimiento general H:
Rendimiento 62 %, solido blanco. 1H RMN (acetona-d6): 8 = 9,15 (br. s, 1H), 7,54 (br. s, 1H), 7,40-7,46 (m, 2H), 7,29-7,39 (m, 1H), 7,07-7,17 (m, 1H), 3,38-3,50 (m, 2H), 3,19 (t, J = 6,7 Hz, 2H), 1,92 (s, 3H) ppm. 13C RMN (acetona-d6) 8 = 191,6, 171,0, 158,9, 139,3, 130,9, 121,7, 119,1, 114,4, 39,8, 29,4, 22,9 ppm.
Claims (16)
1. Proceso para producir un compuesto que tiene una primera cadena (cadena occidental) y una segunda cadena (cadena oriental) que se condensan formando un anillo pirona al proporcionar a un microorganismo las enzimas de la ruta sintetica que catalizan la smtesis del compuesto, en donde el microorganismo se manipula geneticamente para comprender genes que codifican las enzimas de la ruta sintetica que comprenden las enzimas MxnA (SEQ ID NO: 1), MxnB (SEQ ID NO: 2), MxnC (SEQ ID NO: 3), MxnD (SEQ ID NO: 4), MxnE (SEQ ID NO: 5), MxnF (SEQ ID NO: 6), MxnG (SEQ ID NO: 7), MxnH (SEQ ID NO: 8), Mxnl (SEQ ID NO: 9), MxnJ (SEQ ID NO: 10) y MxnK (SEQ ID NO: 11), incubar el microorganismo y separar el compuesto del microorganismo,
caracterizado por que
al menos un dominio catalttico de una enzima esta ausente o inactivado, dominio que se selecciona del grupo que consiste en
a) la cetosintasa (KS) del modulo 1E de Mxnl que comprende los aminoacidos No. 694-885 de SEQ ID NO: 9, b) la cetosintasa (KS) del modulo 2E de Mxnl que comprende los aminoacidos No. 1945-2135 de SEQ ID NO: 9, c) la cetosintasa (KS) del modulo 3E de Mxnl que comprende los aminoacidos No. 3442-3633 de SEQ ID NO: 10, d) la cetosintasa (KS) del modulo 4E de MxnJ que comprende los aminoacidos No. 605-795 de SEQ ID NO: 10, e) la cetosintasa (KS) del modulo 5E de MxnJ que comprende los aminoacidos No. 2476-2660 de SEQ ID NO: 10, f) la cetosintasa (KS) del modulo 6E de MxnJ que comprende los aminoacidos No. 3208-3399 de SEQ ID NO: 10, g) la cetosintasa (KS) del modulo 1W de MxnK que comprende los aminoacidos No. 180-364 de SEQ ID NO: 11, h) la cetosintasa (KS) del modulo 2W de MxnK que comprende los aminoacidos No. 1926-2110 de SEQ ID NO: 11, i) la cetosintasa (KS) del modulo 3W de MxnK que comprende los aminoacidos No. 2684-2875 de SEQ ID NO: 11, j) la cetosintasa (KS) del modulo 4W de MxnK que comprende los aminoacidos No. 3504-3688 de SEQ ID NO: 11, k) la cetosintasa (KS) del modulo 5W de MxnK que comprende los aminoacidos No. 5409-5593 de SEQ ID NO: 11,
l) la cetorreductasa (KR) del modulo 2E de Mxnl que comprende los aminoacidos No. 2903-3050 de SEQ ID NO: 9, m) la cetorreductasa (KR) del modulo 4E de MxnJ que comprende los aminoacidos No. 1568-1715 de SEQ ID NO: 10,
n) la cetorreductasa (KR) del modulo 1W de MxnK que comprende los aminoacidos No. 1418-1560 de SEQ ID NO: 11,
o) la cetorreductasa (KR) del modulo 4W de MxnK que comprende los aminoacidos No. 4470-4617 de SEQ ID NO: 11,
p) la protema transportadora (CP) del modulo L de Mxnl que comprende los aminoacidos No. 424-433 de SEQ ID NO:9,
q) la protema transportadora (CP) del modulo 1E de Mxnl que comprende los aminoacidos No. 1720-1726 de SEQ ID NO: 9,
r) la protema transportadora (CP) del modulo 2E de Mxnl que comprende los aminoacidos No. 3192-3198 de SEQ ID NO: 9,
s) la protema transportadora (CP) del modulo 3E de MxnJ que comprende los aminoacidos No. 366-372 de SEQ ID NO: 9,
t) la protema transportadora (CP) del modulo 4E de MxnJ que comprende los aminoacidos No. 2238-2244 de SEQ ID NO: 10,
u) la protema transportadora (CP) del modulo 5E de MxnJ que comprende los aminoacidos No. 2967-2973 de SEQ ID NO: 10,
v) la protema transportadora (CP) del modulo 6E de MxnJ que comprende los aminoacidos No. 3810-3816 de SEQ ID NO: 10,
w) la protema transportadora (CP*) del modulo 6E de MxnJ que comprende los aminoacidos No. 3566-3572 de SEQ ID NO: 12,
x) la protema transportadora (CP) del modulo 1W de MxnK, que comprende los aminoacidos No. 1683-1689 de SEQ ID NO: 11
y) la protema transportadora (CP) del modulo 2W de MxnK que comprende los aminoacidos No. 2429-2435 de SEQ ID NO: 11,
z) la protema transportadora (CP) del modulo 3W de MxnK que comprende los aminoacidos No. 3234-3240 de SEQ ID NO: 11,
aa) la protema transportadora (CP) del modulo 4W de MxnK que comprende los aminoacidos No. 5165-5171 de SEQ ID NO: 11,
bb) la protema transportadora (CP*) del modulo 5W de MxnK que comprende los aminoacidos No. 5894-5900 de SEQ ID NO: 11,
cc) la protema transportadora (CP) del modulo 5W de MxnK que comprende los aminoacidos No. 5984-5990 de SEQ ID NO: 11,
dd) la deshidratasa (DH) del modulo 2E de Mxnl que comprende los aminoacidos No. 2402-2414 de SEQ ID NO: 9, ee) la deshidratasa (DH) del modulo 3E de MxnJ que comprende los aminoacidos No. 53-65 de SEQ ID NO: 9, ff) la deshidratasa (DH) del modulo 4E de MxnJ que comprende los aminoacidos No. 1091-1103 de SEQ ID NO: 10, gg) la deshidratasa (DH) del modulo 1W de MxnK que comprende los aminoacidos No. 955-967 de SEQ ID NO: 11, hh) la deshidratasa (DH) del modulo 4W de MxnK que comprende los aminoacidos No. 3980-3992 de SEQ ID NO: 11,
ii) la metiltransferasa (MT) del modulo 4E de MxnJ que comprende los aminoacidos No. 1982-1987 de SEQ ID NO: 10,
jj) la metiltransferasa (MT) del modulo 4W de MxnK que comprende los aminoacidos No. 4914-4919 de SEQ ID NO: 11,
en donde en las realizaciones en las que al menos un dominio catalftico esta inactivado en las enzimas de la ruta sintetica MxnK y/o MxnI-MxnJ, en donde el dominio catalftico inactivado es un dominio catalftico cuya inactivacion en ausencia de precursor da lugar a una parada de la smtesis por la enzima cuando se expresa en un microorganismo, un precursor de la cadena occidental, y/o un precursor de la cadena oriental se anade a un microorganismo que expresa las enzimas de la ruta sintetica, o en donde en las realizaciones en las que la inactivacion es por intercambio de al menos un centro catalftico, dominio, modulo o enzima seleccionada de las enzimas CorJ (SEQ ID NO: 25), CorK (SEQ ID NO: 26), CorL (SEQ ID NO: 27), Orf1 (SEQ ID NO: 27), CorM (SEQ ID NO: 28), CorM (SEQ ID NO: 28), CorN (SEQ ID NO: 29) y CorO (SEQ ID NO: 30) y sus centros catalfticos, dominios y modulos, o la enzima MxnI (SEQ ID NO: 9) se reemplaza por la enzima CorI (SEQ ID NO: 24).
2. El proceso segun la reivindicacion 1, caracterizado por que al menos una de la enzima MxnA (SEQ ID NO: 1) se reemplaza por la enzima CorA (SEQ ID NO: 16), la enzima MxnB (SEQ ID NO: 2) se reemplaza por la enzima CorB (SEQ ID NO: 17), la enzima MxnC (SEQ ID NO: 3) se reemplaza por la enzima CorC (SEQ ID NO: 18), la enzima MxnD (SEQ ID NO: 4) se reemplaza por la enzima CorD (SEQ ID NO: 19), la enzima MxnE (SEQ ID NO: 5) se reemplaza por la enzima CorE (SEQ ID NO: 20), la enzima MxnF (SEQ ID NO: 6) se reemplaza por la enzima CorF (SEQ ID NO: 21), la enzima MxnG (SEQ ID NO: 7) se reemplaza por la enzima CorG (SEQ ID NO: 22), la enzima MxnH (SEQ ID NO: 8) se reemplaza por la enzima CorH (SEQ ID NO: 23), la enzima MxnJ (SEQ ID NO: 10) se reemplaza por la enzima Cor J (SEQ ID NO: 25), la enzima MxnK (SEQ ID NO: 11) se reemplaza por las enzimas CorK (SEQ ID NO: 26) y CorL (SEQ ID NO: 27).
3. El proceso segun la reivindicacion 2, caracterizado por que al menos un dominio catalftico de una enzima esta ausente o inactivado, dominio que se selecciona del grupo que consiste en
a) la cetosintasa (KS) del modulo 1E de Corl que comprende los aminoacidos No. 688-880 de SEQ ID NO: 23, b) la cetosintasa (KS) del modulo 2E de Corl que comprende los aminoacidos No. 1925-2115 de SEQ ID NO: 23, c) la cetosintasa (KS) del modulo 3E de Corl que comprende los aminoacidos No. 3416-3607 de SEQ ID NO: 23,
d) la cetosintasa (KS) del modulo 4E de CorJ que comprende los aminoacidos No. 586-776 de SEQ ID NO: 24, e) la cetosintasa (KS) del modulo 5E de CorJ que comprende los aminoacidos No. 2448-2632 de SEQ ID NO: 24, f) la cetosintasa (KS) del modulo 6E de CorJ que comprende los aminoacidos No. 3172-3363 de SEQ ID NO: 24, g) la cetosintasa (KS) del modulo 1W de CorK que comprende los aminoacidos No. 171-355 de SEQ ID NO: 25, h) la cetosintasa (KS) del modulo 2W de CorK que comprende los aminoacidos No. 1898-2082 de SEQ ID NO: 25, i) la cetosintasa (KS) del modulo 3W de CorK que comprende los aminoacidos No. 3362-3550 de SEQ ID NO: 25, j) la cetosintasa (KS) del modulo 4W de CorL que comprende los aminoacidos No. 449-633 de SEQ ID NO: 26, k) la cetosintasa (KS) del modulo 5W de CorL que comprende los aminoacidos No. 1678-1869 de SEQ ID NO: 26, l) la cetosintasa (KS) del modulo 6W de CorL que comprende los aminoacidos No. 2484-2668 de SEQ ID NO: 26, m) la cetosintasa (KS) del modulo 7W de CorL que comprende los aminoacidos No. 4355-4539 de SEQ ID NO: 26, n) la cetorreductasa (KR) del modulo 2E de Corl que comprende los aminoacidos No. 2874-3021 de SEQ ID NO: 23, o) la cetorreductasa (KR) del modulo 4E de CorJ que comprende los aminoacidos No. 1548-1695 de SEQ ID NO: 24,
p) la cetorreductasa (KR) del modulo 1W de CorK que comprende los aminoacidos No. 1401-1543 de SEQ ID NO: 25,
q) la cetorreductasa (KR) del modulo 2W de CorK, que comprende los aminoacidos No. 2826-2972 de SEQ ID NO: 25,
r) la cetorreductasa (KR) del modulo 4W de CorL que comprende los aminoacidos No. 1139-1285 de SEQ ID NO: 26,
s) la cetorreductasa (KR) del modulo 6W de CorL que comprende los aminoacidos No. 3427-3574 de SEQ ID NO: 26,
t) la protema transportadora (CP) del modulo L de Corl que comprende los aminoacidos No. 416-425 de SEQ ID NO: 23,
u) la protema transportadora (CP) del modulo 1E de Corl que comprende los aminoacidos No. 1706-1712 de SEQ ID NO: 23,
v) la protema transportadora (CP) del modulo 2E de Corl que comprende los aminoacidos No. 3166-3172 de SEQ ID NO: 23,
w) la protema transportadora (CP) del modulo 3E de CorJ que comprende los aminoacidos No. 350-356 de SEQ ID NO: 24,
x) la protema transportadora (CP) del modulo 4E de CorJ que comprende los aminoacidos No. 2206-2212 de SEQ ID NO: 24,
y) la protema transportadora (CP) del modulo 5E de CorJ que comprende los aminoacidos No. 2935-2941 de SEQ ID NO: 24,
z) la protema transportadora (CP) del modulo 6E* de CorJ que comprende los aminoacidos No. 3530-3536 de SEQ ID NO: 24,
aa) la protema transportadora (CP) del modulo 6E de CorJ que comprende los aminoacidos No. 3765-3771 de SEQ ID NO: 24,
bb) la protema transportadora (CP) del modulo 1W de CorK que comprende los aminoacidos No. 1663-1669 de SEQ ID NO: 25,
cc) la protema transportadora (CP) del modulo 2W de CorK que comprende los aminoacidos No. 3109-3115 de SEQ ID NO: 25,
dd) la protema transportadora (CP) del modulo 3W* de CorK que comprende los aminoacidos No. 3900-3906 de SEQ ID NO: 25,
ee) la protema transportadora (CP) del modulo 3W de CorK que comprende los aminoacidos No. 3994-4090 de SEQ ID NO: 25,
ff) la protema transportadora (CP) del modulo 4W de CorL que comprende los aminoacidos No. 1443-1449 de SEQ ID NO: 25,
gg) la protema transportadora (CP) del modulo 5W de CorL que comprende los aminoacidos No. 2208-2214 de SEQ ID NO: 26,
hh) la protema transportadora (CP) del modulo 6W de CorL que comprende los aminoacidos No. 4116-4122 de SEQ ID NO: 26,
ii) la protema transportadora (CP) del modulo 7W* de CorL que comprende los aminoacidos No. 4826-4832 de SEQ ID NO: 26,
jj) la protema transportadora (CP) del modulo 7W de CorL que comprende los aminoacidos No. 4920-4926 de SEQ ID NO:26,
kk) la deshidratasa (DH) del modulo 2E de Corl que comprende los aminoacidos No. 2380-2392 de SEQ ID NO: 23, ll) la deshidratasa (DH) del modulo 3E de CorJ que comprende los aminoacidos No. 44-56 de SEQ ID NO: 24, mm) la deshidratasa (DH) del modulo 4E de CorJ que comprende los aminoacidos No. 1073-1085 de SEQ ID NO: 24,
nn) la deshidratasa (DH) del modulo 1W de CorK que comprende los aminoacidos No. 942-954 de SEQ ID NO: 25, oo) la deshidratasa (DH) del modulo 2W de CorK que comprende los aminoacidos No. 2319-2331 de SEQ ID NO: 25,
pp) la deshidratasa (DH) del modulo 4W de CorL que comprende los aminoacidos No. 19-31 de SEQ ID NO: 26, qq) la deshidratasa (DH) del modulo 6W de CorL que comprende los aminoacidos No. 2949-2961 de SEQ ID NO: 26,
rr) la metiltransferasa (MT) del modulo 4E de CorJ que comprende los aminoacidos No. 1962-1967 de SEQ ID NO: 24,
ss) la metiltransferasa (MT) del modulo 6W de CorL que comprende los aminoacidos No. 3865-3870 de SEQ ID NO: 26.
4. Proceso segun una de las reivindicaciones precedentes, caracterizado porque en la enzima MxnK (SEQ ID NO: 11) el modulo 1W esta inactivado y por la adicion de un precursor de la cadena occidental que tiene la estructura o

en donde Rw es un alquilo C1-C12, arilo o grupo aromatico,
y en donde X es un grupo auxiliar que esta unido por un atomo de azufre.
5. Proceso segun una de las reivindicaciones precedentes, caracterizado por que en la enzima MxnK (SEQ ID NO: 11) el modulo 2W esta inactivado y por la adicion de un precursor de la cadena occidental que tiene la estructura o

en donde Rw es un alquilo C1-C12, arilo o grupo aromatico,
y en donde X es un grupo auxiliar que esta unido por un atomo de azufre.
6. Proceso segun una de las reivindicaciones precedentes, caracterizado por que en la enzima MxnK (SEQ ID NO: 11) el modulo 3W esta inactivado y por la adicion de un precursor de la cadena occidental que tiene la estructura o

en donde Rw es un alquilo C1-C12, arilo o grupo aromatico,
y en donde X es un grupo auxiliar que esta unido por un atomo de azufre.
7. Proceso segun una de las reivindicaciones precedentes, caracterizado por que en la enzima MxnK (SEQ ID NO: 11) el modulo 4W esta inactivado y por la adicion de un precursor de la cadena occidental que tiene la estructura o

en donde Rw es un alquilo C1-C12, arilo o grupo aromatico,
y en donde X es un grupo auxiliar que esta unido por un atomo de azufre.
8. Proceso segun una de las reivindicaciones 5 a 7, caracterizado por que el precursor de la cadena occidental tiene una estructura seleccionada de Rw-CH2-CH2-CH2-C(O)-X, Rw-CH2-CH2-C(CH3)=CH2-C(O)-X, RW-CH2-CH2-C(CHa)=CH-CH=C(CHa)-C(O)-X.
9. Proceso segun una de las reivindicaciones precedentes, caracterizado por que en la enzima Mxnl (SEQ ID NO: 9) el modulo de carga (LM) que comprende los aminoacidos 1 a 466 de SEQ ID NO: 9 esta inactivado y por la adicion de un precursor de la cadena oriental que tiene la estructura

en donde X es un grupo auxiliar unido por un atomo de azufre.
10. Proceso segun una de las reivindicaciones precedentes, caracterizado por que en la enzima Mxnl (SEQ ID NO: 9) el modulo 1E esta inactivado y por la adicion de un precursor de la cadena oriental que tiene la estructura

en donde X es un grupo auxiliar unido por un atomo de azufre.
11. Proceso segun una de las reivindicaciones precedentes, caracterizado porque en la enzima Mxnl (SEQ ID NO: 9) el modulo de carga y el modulo 3E que comprende los aminoacidos 1 a 1759 de SEQ ID NO: 9 estan inactivados y por la adicion de un precursor de la cadena oriental que tiene la estructura

en donde X es un grupo auxiliar unido por un atomo de azufre
12. Proceso segun una de las reivindicaciones precedentes, caracterizado por que la enzima Mxnl (SEQ ID NO: 9) y el modulo 3E, que comprende los aminoacidos No. 1 a 405 de SEQ ID NO: 10 estan funcionalmente ausentes y por la adicion de un precursor de la cadena oriental que tiene la estructura
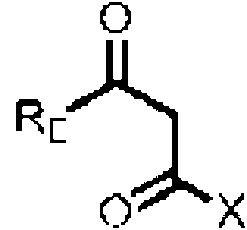
en donde X es un grupo auxiliar unido por un atomo de azufre.
13. Proceso segun una de las reivindicaciones 5 a 12, en donde el precursor se selecciona de


14. Proceso segun una de las reivindicaciones precedentes, caracterizado por que en la enzima Mxnl el modulo 4E que comprende los aminoacidos No. 1 a 3052 de SEQ ID NO: 9 esta inactivado y por la adicion de un precursor de la cadena oriental que tiene la estructura

en donde X es un grupo auxiliar unido por un atomo de azufre.
15. Proceso segun una de las reivindicaciones precedentes, caracterizado por que la enzima Mxnl (SEQ ID NO: 9) esta funcionalmente ausente y porque la enzima MxnJ consiste en el modulo 6E que consiste en los aminoacidos No.
3007 a 3849 de SEQ ID NO: 10.
16. Proceso segun una de las reivindicaciones precedentes, caracterizado por que las enzimas, que al menos comprenden MxnB se separan del microorganismo y el compuesto se produce por las enzimas en una reaccion sin celulas.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP13167392 | 2013-05-10 | ||
| PCT/EP2014/059677 WO2014181000A1 (en) | 2013-05-10 | 2014-05-12 | Production of myxopyronin and of its derivatives |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2713437T3 true ES2713437T3 (es) | 2019-05-21 |
Family
ID=48325491
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES14725669T Active ES2713437T3 (es) | 2013-05-10 | 2014-05-12 | Producción de mixopironina y de sus derivados |
Country Status (4)
| Country | Link |
|---|---|
| EP (1) | EP2994535B1 (es) |
| ES (1) | ES2713437T3 (es) |
| HU (1) | HUE041663T2 (es) |
| WO (1) | WO2014181000A1 (es) |
-
2014
- 2014-05-12 WO PCT/EP2014/059677 patent/WO2014181000A1/en not_active Ceased
- 2014-05-12 EP EP14725669.7A patent/EP2994535B1/en active Active
- 2014-05-12 ES ES14725669T patent/ES2713437T3/es active Active
- 2014-05-12 HU HUE14725669A patent/HUE041663T2/hu unknown
Also Published As
| Publication number | Publication date |
|---|---|
| HUE041663T2 (hu) | 2019-05-28 |
| EP2994535A1 (en) | 2016-03-16 |
| EP2994535B1 (en) | 2018-12-05 |
| WO2014181000A1 (en) | 2014-11-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2240259T3 (es) | Produccion recombinante de poliquetidos aromaticos. | |
| US8604004B2 (en) | Moenomycin analogs, methods of synthesis, and uses thereof | |
| AU2018301674B2 (en) | Generation of water-soluble cannabinoid compounds in yeast and plant cell suspension cultures and compositions of matter | |
| ES2287960T3 (es) | Produccion de policetidas en basct4rias y levadura. | |
| CN111373045A (zh) | 水溶性大麻素在植物系统中高水平体内生物合成和分离 | |
| ES2856887T3 (es) | Proceso para aislar y purificar Ambrox | |
| US20100016250A1 (en) | Toll-like receptor 9 agonists | |
| JP4599357B2 (ja) | プラジエノライドの生合成に関与するポリペプチドをコードするdna | |
| ES2899121T3 (es) | Producción de manool | |
| Oleksak et al. | The oxazolomycin family: a review of current knowledge | |
| Dickschat et al. | Biosynthesis of iso-fatty acids in myxobacteria | |
| CN105255921B (zh) | 异源合成广霍香醇的方法和微生物 | |
| ES2713437T3 (es) | Producción de mixopironina y de sus derivados | |
| CN115368439B (zh) | 一组fk228衍生物及其制备方法与应用 | |
| ES2933968T3 (es) | Producción de manool | |
| CN104427870B (zh) | Uk‑2生物合成基因和使用其提高uk‑2生产率的方法 | |
| US9394553B2 (en) | Biosynthetic gene cluster for chejuenolide of marine microorganism Hahella chejuensis | |
| CN102791871A (zh) | 通过酶法生产啶南平衍生物的方法 | |
| JP2009072152A (ja) | 天然型のホスファチジルイノシトールを製造する方法およびそのための改変型ホスホリパーゼd | |
| CN121319146B (zh) | 用于合成昆虫性信息素前体化合物的蛋白及方法 | |
| WO2025194409A1 (en) | Ionizable cationic lipid compounds for delivery system and uses thereof | |
| Nakou | Discovery and biosynthesis of gladiostatin: a novel glutarimide antibiotic from Burkholderia gladioli | |
| US20230159961A1 (en) | Biosynthesis of chemically diversified non-natural terpene products | |
| JP4590625B2 (ja) | 新規イソクマリン系蛍光物質 | |
| CN120536392A (zh) | 莨菪碱4β-羟化酶及其应用 |