JP2009284832A - 耐熱性菌類の検出方法 - Google Patents
耐熱性菌類の検出方法 Download PDFInfo
- Publication number
- JP2009284832A JP2009284832A JP2008141499A JP2008141499A JP2009284832A JP 2009284832 A JP2009284832 A JP 2009284832A JP 2008141499 A JP2008141499 A JP 2008141499A JP 2008141499 A JP2008141499 A JP 2008141499A JP 2009284832 A JP2009284832 A JP 2009284832A
- Authority
- JP
- Japan
- Prior art keywords
- primer
- base sequence
- seq
- set forth
- oligonucleotide represented
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images


















Landscapes
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
【解決手段】Loop mediated isothermal amplification(LAMP)法によって、耐熱性菌類のβ−チューブリン遺伝子、または28S rDNAのD1/D2領域及びITS領域の遺伝子中の標的領域を含むDNA断片を増幅させ、増幅産物の有無を確認する工程を含む耐熱性菌類の検出方法。対象菌種は、ビソクラミス(Byssochlamys)属の菌類、タラロマイセス(Talaromyces)属の菌類、ネオサルトリア(Neosartorya)属の菌類、ハミゲラ(Hamigera)属の菌類、およびアスペルギルス・フミガタス(Aspergillus fumigatus)から選ばれる。
【選択図】なし
Description
さらに、危害事故発生時における事故原因究明及び対策のためには、事故原因菌の同定が必要となる。従って、上記4属の耐熱性菌類を識別することができれば、より迅速な事故原因菌の検出、識別が可能となる。
DNAポリメラーゼと、
dATP、dCTP、dGTP及びdTTPを含むdNTPと、
を含む耐熱性菌類検出キットに関する。
本発明は、Loop mediated isothermal amplification(LAMP)法によって耐熱性菌類の遺伝子中の標的領域のDNA断片を増幅させ、増幅産物の有無を確認する工程を含む耐熱性菌類の検出方法である。本発明において、前記標的領域がβ−チューブリン遺伝子、または28S rDNAのD1/D2領域及びITS(内部転写スペーサー、internal transcribed spacer)領域の塩基配列から選択されることが好ましい。ここで、「β−チューブリン」とはa-チューブリンとともに真核細胞の微小管を形成しているタンパク質であり、「β−チューブリン遺伝子」とは、β−チューブリンをコードする遺伝子である。また、「28S rDNA」とはタンパク質への変換の場であるリボゾームの遺伝情報をコードするDNAである。
また、本発明の耐熱性菌類の検出方法において、遺伝子中の標的領域としてβ−チューブリン遺伝子または28S rDNAの特定領域を増幅する。これは、β−チューブリン遺伝子と28S rDNAは共に、コードする蛋白質自体は真菌に普遍的に存在するが、遺伝子中には塩基の変異が蓄積しやすく、属や種のレベルで保存されているため、遺伝子内の特定の領域に他属他種との識別に用いることができる塩基配列が存在する可能性が高いという共通点があるからである。
上記6つの塩基配列領域は、以下のように選定することができるが、本発明はこれに限定されるものではない。
検出対象となる菌種の塩基配列を用いてアライメントを行う。アライメントには、例えば、Clustal X等のソフトを用いることができる。次いで、得られたアライメント情報をもとに、Primer Explorer V4(栄研化学株式会社社製)等のソフトを用いて上記6つの塩基配列領域を選定し、LAMP法のプライマーを設計する。
一方、「アウタープライマー」とは、標的塩基配列上の『「ある特定のヌクレオチド配列領域」(例えば前記F2領域またはB2領域)の5'末端側に存在するある特定のヌクレオチド配列領域』を認識かつ合成起点を与える塩基配列を有するオリゴヌクレオチドであり、F3領域より選ばれた塩基配列を含むプライマーおよびB3領域より選ばれた塩基配列を含むプライマーが挙げられる。ここで、F3領域より選ばれた塩基配列を含むプライマーをアウタープライマーF(以下、F3プライマー)、B3領域より選ばれた塩基配列を含むプライマーをアウタープライマーB(以下、B3プライマー)と呼ぶ。
ここで、各プライマーにおけるFとは、標的塩基配列のアンチセンス鎖と相補的に結合し、合成起点を提供することを意味するプライマー表示であり、各プライマーにおけるBとは、標的塩基配列のセンス鎖と相補的に結合し、合成起点を提供することを意味するプライマー表示である。
ループプライマーの塩基配列は、上記ダンベル構造の5’側のループ構造の一本鎖部分の塩基配列に相補的であれば、標的領域の塩基配列又はその相補鎖から選択されてもよく、他の塩基配列でもよい。また、ループプライマーは1種類であっても、2種類であってもよい。
本明細書におけるビソクラミス属、タラロマイセス属、ネオサルトリア属およびハミゲラ属等の「耐熱性菌類」とは、マユハキタケ科(Trichocomaceae)に属する不整子嚢菌類であり、75℃、30分間の加熱処理後であっても生存可能な耐熱性菌類である。ビソクラミス属の菌類の例として、ビソクラミス ファルバ(Byssochlamys fulva)、ビソクラミス ニベア(Byssochlamys nivea)が挙げられる。タラロマイセス属の菌類の例として、タラロマイセス フラバス(Talaromyces flavus)、タラロマイセス ルテウス(Talaromyces luteus)、タラロマイセス トラキスペルムス(Talaromyces trachyspermus)、タラロマイセス ウォルトマニー(Talaromyces wortmannii)が挙げられる。ネオサルトリア属の菌類の例として、ネオサルトリア スピノサ(Neosartorya spinosa)、ネオサルトリア フィシェリ(Neosartorya ficheri)、ネオサルトリア グラブラ(Neosartorya glabra)、ネオサルトリア ヒラツカエ(Neosartorya hiratsukae)が挙げられる。ハミゲラ属の菌類の例として、ハミゲラ アベラネア(Hamigera avellanea)が挙げられる。
本発明における「アスペルギルス フミガタス」とは、テレオモルフ(有性世代)が発見されていない不完全菌類の一種であり、ネオサルトリア フィシェリとは、系統的に近縁である。
ビソクラミス属の菌類検出用プライマーセット(LB1プライマーセット)
LB1F3プライマー:CGGTCCTCGAGCGTATGG(配列番号1)
LB1B3プライマー:CCGTTACTGGGGCAATCC(配列番号2)
LB1FIPプライマー:AGTTAGGTGACCGTGAGGTCGTCTTTGTCACGCGCTCTGG(配列番号3)
LB1BIPプライマー:GGATCAGGTAGGGATACCCGCTGTTGGTTTCTTTTCCTCCGC(配列番号4)
図1に、ビソクラミス属の28S rDNAのITS領域及びD1/D2領域の塩基配列における、上記プライマーが認識する塩基配列の位置関係を示す。
ネオサルトリア属の菌類およびアスペルギルス フミガタス検出用プライマーセット(LN1プライマーセット)
LN1F3プライマー:GGCAACATCTCACGATCTGA(配列番号5)
LN1B3プライマー:CCCTCAGTGTAGTGACCCTT(配列番号6)
LN1FIPプライマー:ATGGTACCAGGCTCGAGATCGATACTAGGCCAACGGTGACA(配列番号7)
LN1BIPプライマー:GTCCCTTCGGCGAGCTCTTCGTTGTTACCAGCACCAGACT(配列番号8)
ネオサルトリア属の菌類およびアスペルギルス フミガタスを検出するために、上記プライマーに加えてループプライマーを用いるのが好ましい。ループプライマーとしては、以下のプライマーを用いるのが好ましい。
ネオサルトリア属の菌類およびアスペルギルス フミガタス検出用ループプライマー(LN1ループプライマー)
LN1LFループプライマー:ACGGCACGAGGAACATACT(配列番号9)
LN1LBループプライマー:CGATAACTTCGTCTTCGGCC(配列番号10)
図2に、ネオサルトリア グラブラと同じネオサルトリア属のネオサルトリア フィシェリのβ−チューブリン遺伝子の塩基配列における、上記プライマーが認識する塩基配列の位置関係を示す。
アスペルギルス フミガタス検出用プライマーセット(LAf2プライマーセット)
LAf2F3プライマー:GCCGCTTTCTGGTATGTCT(配列番号11)
LAf2B3プライマー:CGCTTCTTCCTTGTTTTCCG(配列番号12)
LAf2FIPプライマー:CCATGACAGTGAGGCTGAACCCCGGGTGATTGGGATCTCTCA(配列番号13)
LAf2BIPプライマー:ACCATCTCTGGTGAGCATGGCTTTCCGCCGCTTTCTCAA(配列番号14)
アスペルギルス フミガタスを検出するために、上記プライマーに加えてループプライマーを用いるのが好ましい。ループプライマーとしては、以下のプライマーを用いるのが好ましい。
アスペルギルス フミガタス検出用ループプライマー(LAf2ループプライマー)
LAf2LBループプライマー:AGTAAGTTCGACCTATATCCTCCC(配列番号15)
図3に、アスペルギルス フミガタスのβ−チューブリン遺伝子の塩基配列における、上記プライマーが認識する塩基配列の位置関係を示す。
なお、上記プライマーセットを用いた場合、アスペルギルス フミガタスを特異的に検出することはできるが、ネオサルトリア属の菌類を検出することはできない。従って、前記ネオサルトリア属の菌類およびアスペルギルス フミガタス検出用プライマーセット(LN1プライマーセット)と、上記アスペルギルス フミガタス検出用プライマーセット(LAf2プライマーセット)を組み合わせて用いることにより、ネオサルトリア属の菌類とアスペルギルス フミガタスとを識別することができる。
ハミゲラ属の菌類検出用プライマーセット(LH2プライマーセット)
LH2F3プライマー:GGATCCGAATACGACGTGTC(配列番号16)
LH2B3プライマー:CCCTCAGTGTAGTGACCCTT(配列番号17)
LH2FIPプライマー:CATGGTGCCAGGCTCGAGATCCAGGCCAGCGGTAACAAG(配列番号18)
LH2BIPプライマー:CCGGTCCTTTTGGCCAGCTCTGTTACCGGCACCAGACT(配列番号19)
ハミゲラ属の菌類を検出するために、上記プライマーに加えてループプライマーを用いるのが好ましい。ループプライマーとしては、以下のプライマーを用いるのが好ましい。
ハミゲラ属の菌類検出用ループプライマー(LH2ループプライマー)
LH2LFループプライマー:ACGGCACGGGGGACATA(配列番号20)
LH2LBループプライマー:TTCCGCCCAGACAACTTCG(配列番号21)
図4に、ハミゲラ属のβ−チューブリン遺伝子の塩基配列における、上記プライマーが認識する塩基配列の位置関係を示す。
タラロマイセス フラバス検出用プライマーセット(LTf2プライマーセット)
LTf2F3プライマー:CCAGTTGGAGCGTATGAACG(配列番号22)
LTf2B3プライマー:CCCAGTTGTTACCAGCACCG(配列番号23)
LTf2FIPプライマー:TTGTTGCCGGAGGCCTACACTTTACTTCAACGAGGTGCGT(配列番号24)
LTf2BIPプライマー:CGACTTGGAGCCCGGTACCAAAAGTTGTCGGGACGGAAGA(配列番号25)
タラロマイセス フラバスを検出するために、上記プライマーに加えてループプライマーを用いるのが好ましい。ループプライマーとしては、以下のプライマーを用いるのが好ましい。
タラロマイセス フラバス検出用ループプライマー(LTf2ループプライマー)
LTf2LBループプライマー:GCTGGTCCCTTTGGTCAGC(配列番号26)
図5に、タラロマイセス フラバスのβ−チューブリン遺伝子の塩基配列における、上記プライマーが認識する塩基配列の位置関係を示す。
タラロマイセス ウォルトマニー検出用プライマーセット(LTw4−3プライマーセット)
LTw4F3プライマー:TGGCTCCGGAATGTGAGTT(配列番号27)
LTw3B3プライマー:CAAATCGACGAGGACGGC(配列番号28)
LTw4FIPプライマー:CGCTCCAACTGGAGGTCGGAAAATTTCGACATCCCACCCT(配列番号29)
LTw3BIPプライマー:GGAATCTGCCCCGCGACATTCCGGGGGACGTACTTGTTG(配列番号30)
タラロマイセス ウォルトマニーを検出するために、上記プライマーに加えてループプライマーを用いるのが好ましい。ループプライマーとしては、以下のプライマーを用いるのが好ましい。
タラロマイセス ウォルトマニー検出用ループプライマー(LTw4−3ループプライマー)
LTw4LFループプライマー:GGTGCCATTGTAACTGGAAATGA(配列番号31)
LTw3LBループプライマー:ACTCATATCGTATAGGCTAGCGG(配列番号32)
図6に、タラロマイセス ウォルトマニーのβ−チューブリン遺伝子の塩基配列における、上記プライマーが認識する塩基配列の位置関係を示す。
タラロマイセス ルテウス検出用プライマーセット(LTl1プライマーセット)
LTl1F3プライマー:CGAATCACCACTGATGGGAA(配列番号33)
LTl1B3プライマー:GAAGAGCTGACCGAAAGGAC(配列番号34)
LTl1FIPプライマー:TTCGTGCTGTCGGTCGGTAATGTTCCGACCTCCAGTTAGAGC(配列番号35)
LTl1BIPプライマー:TAGGCTAGCGGCAACAAGTACGATAGTACCGGGCTCCAGATC(配列番号36)
タラロマイセス ルテウスを検出するために、上記プライマーに加えてループプライマーを用いるのが好ましい。ループプライマーとしては、以下のプライマーを用いるのが好ましい。
タラロマイセス ルテウス検出用ループプライマー(LTl1ループプライマー)
LTl1LFループプライマー:ACCTCGTTGAAATAGACGTTCA(配列番号37)
図7に、タラロマイセス ルテウスのβ−チューブリン遺伝子の塩基配列における、上記プライマーが認識する塩基配列の位置関係を示す。
タラロマイセス フラバスおよびタラロマイセス トラキスパーマム検出用プライマーセット(LT1プライマーセット)
LT1F3プライマー:GCGTCATTTCTGCCCTCAA(配列番号38)
LT1B3プライマー:AGTTCAGCGGGTAACTCCT(配列番号39)
LT1FIPプライマー:TACGCTCGAGGACCAGACGGCGGCTTGTGTGTTGGGTG(配列番号40)
LT1BIPプライマー:TCTGTCACTCGCTCGGGAAGGACCTGATCCGAGGTCAACC(配列番号41)
タラロマイセス フラバスおよびタラロマイセス トラキスパーマムを検出するために、上記プライマーに加えてループプライマーを用いるのが好ましい。ループプライマーとしては、以下のプライマーを用いるのが好ましい。
タラロマイセス フラバスおよびタラロマイセス トラキスパーマム検出用ループプライマー(LT1ループプライマー)
LT1LFループプライマー:GCTGCCTTTTGGGCAGGTC(配列番号42)
LT1LBループプライマー:TGGTCACACCACTATATTTTACCAC(配列番号43)
図8、図9及び図9−1に、タラロマイセス フラバスおよびタラロマイセス トラキスパーマムの28S rDNAのITS領域及びD1/D2領域の塩基配列における、上記プライマーが認識する塩基配列の位置関係を示す。
(a)前記B2領域を3’側に有し、前記B1c領域を5’側に有する塩基配列
(b)前記B3領域を有する塩基配列
(c)前記F2領域を3’側に有し、前記F1c領域を5’側に有する塩基配列
(d)前記F3領域を有する塩基配列
(e)前記B1領域と前記B2領域の間の部分と相補的な配列を有する塩基配列
(f)前記F1領域と前記F2領域の間の部分と相補的な配列を有する塩基配列
本発明のオリゴヌクレオチドは、LAMP法で用いられるプライマーとしてだけではなく、PCR法等のプライマー、核酸検出用プローブなどとしても用いることができる。
検体からDNAを調製する方法としては、耐熱性菌類の検出を行うのに十分な精製度および量のDNAが得られるのであれば特に制限されず、未精製の状態でも使用できるが、さらに分離、抽出、濃縮、精製等の前処理をして使用することもできる。例えば、フェノール及びクロロホルム抽出を行って精製したり、市販の抽出キットを用いて精製して、核酸の純度を高めて使用することができる。
また、LAMP法では核酸の合成により基質が大量に消費され、副産物であるピロリン酸イオンが、共存するマグネシウムイオンと反応してピロリン酸マグネシウムが算出される。ピロリン酸マグネシウムが算出されると、反応液が肉眼で確認できる程度にまで白濁する。この白濁を指標として、反応終了後あるいは反応中の濁度上昇を経時的に光学的に観察できる測定機器を用いて検出できる。例えば、分光光度計を用いて400nmにおける吸光度の変化を確認することによって、核酸の増幅反応を検出することができる(国際公開第01/83817号パンフレット)。
(1)プライマーの設計及び合成
シークエンシング法により決定した各種菌類(パエシロマイセス バリオッティー、ハミゲラ アベラネラ、タラロマイセス フラバス、タラロマイセス ルテウス、タラロマイセス トラキスパーマム、ビソクラミス ニベア、ビソクラミス フルバ、ネオサルトリア フィシェリ)の28SrDNAのITS領域及びD1/D2領域の塩基配列情報をもとに、DNA解析ソフトウエア(商品名:DNAsis pro、日立ソフトウエア社製)を用いてアライメント解析を行い、ビソクラミス属の菌類に特異的な塩基配列を特定した。特定した塩基配列をもとに、配列番号1〜4の塩基配列で表されるオリゴヌクレオチドからなるプライマーを設計し、E Genome order(株式会社富士通システムソリューションズ、配列番号1,2;5pmolスケール、配列番号3,4;40pmolスケール;全てカラム精製品)に合成依頼し、購入した。
ビソクラミス属の菌類としては、ビソクラミス ファルバおよびビソクラミス ニベアを使用した。ビソクラミス属の菌類の28SrDNAのITS領域及びD1/D2領域に対する配列番号1〜4の塩基配列で表されるオリゴヌクレオチドからなるプライマーの特異性を確かめるために、表1に示す菌類も使用した。これらの菌類は千葉大学医学部真菌医学研究センターが保管し、IFMナンバーなどにより管理されているものを入手し、使用した。
各菌体を至適条件下で培養した。培養条件についてはポテトデキストロース培地(商品名:パールコア ポテトデキストロース寒天培地、栄研化学株式会社製)を用いて25℃で7日間培養した。

ゲノムDNA調製用キット(商品名 PrepMan ultra、アプライドバイオ社製)を用いて、菌体からゲノムDNA溶液を調製した。具体的には、各培地から数個のコロニーを採取し、キットの付属試薬200μlに菌体を懸濁し、100℃、10分間の加熱処理で菌体を溶解させ、14800rpmで5分間遠心分離した後、上清を回収した。得られたゲノムDNA溶液の濃度を50ng/μlに調製した。このゲノムDNA溶液を鋳型DNAとして、以下のLAMP反応に用いた。
2x Reaction Mix(Tris−HCl(pH8.8) 40mM、KCl 20mM、MgSO4 16mM、(NH4)2SO4 20mM、0.2%Tween20、Betaine 1.6M、dNTPs 2.8mM:栄研化学株式会社製;Loopamp DNA増幅試薬キット)12.5μl、配列番号1で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LB1F3プライマー:5pmol/μl)1μl、配列番号2で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LB1B3プライマー:5poml/μl)1μl、配列番号3で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LB1FIPプライマー:40pmol/μl)1μl、配列番号4で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LB1BIPプライマー:40pmol/μl)1μl、Bst DNA Polymerase(8U/25μL、栄研化学株式会社製)1μl、および上記で調製した鋳型DNA 1μlを混合し、蒸留水を加えて全量25μlの反応液とした。
上記で調製した反応液を、リアルタイム濁度測定装置Loopamp RT−160C(栄研化学株式会社製)にて、63±2℃で60分間DNAの増幅反応を行った。同時に反応液の濁度を測定した(波長:400nm)。
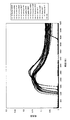
DNAの増幅の有無は、反応液の濁度が上昇しているかによって判断した。反応液の濁度の測定結果を、図10(a)及び図10(b)に示す。なお、図10(a)は表1の試料番号1〜8、図10(b)は表1の試料番号9〜16の結果を示す。
その結果、ビソクラミス属の菌類のゲノムDNAを鋳型とした系のみで、反応開始30分後から濁度の上昇、すなわちDNAの合成・増幅反応が認められた。濁度の上昇は反応開始後60〜70分後にピークとなりその後穏やかに下降した。
一方、ビソクラミス属以外の菌類のゲノムDNAを用いた系では、反応開始後90分までの間、反応液の濁度の上昇は認められなかった。なお、反応開始100分前後から、ビソクラミス属以外の菌類のゲノムDNAを用いた系においても反応液の濁度上昇が観察されたが、これはプライマー同士が反応して増幅してしまうこと、または、反応時間が長くなるにつれて標的配列以外の配列にも少数のプライマーがアニールしてしまうこと等が原因であると考えられる。
以上の結果から、本発明の方法によれば、簡便、迅速かつ特異的にビソクラミス属の菌類を検出することが可能である。
(1)プライマーの設計及び合成
シークエンシング法により決定した各種菌類(ネオサルトリア フィシェリ、ネオサルトリア・グラブラ、パエシロマイセス バリオッティー、ハミゲラ アベラネラ、タラロマイセス フラバス、タラロマイセス ルテウス、タラロマイセス トラキスパーマム、ビソクラミス ニベア、ビソクラミス フルバ)のβ−チューブリン遺伝子の塩基配列情報をもとに、DNA解析ソフトウエア(商品名:DNAsis pro、日立ソフトウエア社製)を用いてアライメント解析を行い、ネオサルトリア属の菌類およびアスペルギルス・フミガタスに特異的な塩基配列を特定した。特定した塩基配列をもとに、配列番号5〜10の塩基配列で表されるオリゴヌクレオチドからなるプライマーを設計し、E Genome order(株式会社富士通システムソリューションズ、配列番号5,6;5pmolスケール、配列番号7,8;40pmolスケール、配列番号9,10;20pmolスケール;全てカラム精製品)に合成依頼し、購入した。
ネオサルトリア属の菌類およびアスペルギルス フミガタスとしては、表2および表2−1に記載の菌類を使用した。これらの菌類のβ−チューブリン遺伝子に対する配列番号5〜10の塩基配列で表されるオリゴヌクレオチドからなるプライマーの特異性を確かめるために、表2および表2−1に示す菌類も使用した。これらの菌類は千葉大学医学部真菌医学研究センターが保管し、IFMナンバーなどにより管理されているものを入手し、使用した。
各菌体を至適条件下で培養した。培養条件についてはポテトデキストロース培地(商品名:パールコア ポテトデキストロース寒天培地、栄研化学株式会社製)を用いて25℃で7日間培養した。
ゲノムDNA調製用キット(商品名 PrepMan ultra、アプライドバイオ社製)を用いて、菌体からゲノムDNA溶液を調製した。具体的には、各培地から数個のコロニーを採取し、キットの付属試薬200μlに菌体を懸濁し、100℃、10分間の加熱処理で菌体を溶解させ、14800rpmで5分間遠心分離した後、上清を回収した。得られたゲノムDNA溶液の濃度を50ng/μlに調製した。このゲノムDNA溶液を鋳型DNAとして、以下のLAMP反応に用いた。
2x Reaction Mix(Tris−HCl(pH8.8) 40mM、KCl 20mM、MgSO4 16mM、(NH4)2SO4 20mM、0.2%Tween20、Betaine 1.6M、dNTPs 2.8mM:栄研化学株式会社製;Loopamp DNA増幅試薬キット)12.5μl、配列番号5で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LN1F3プライマー:5pmol/μl)1μl、配列番号6で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LN1B3プライマー:5pmol/μl)1μl、配列番号7で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LN1FIPプライマー:40pmol/μl)1μl、配列番号8で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LN1BIPプライマー:40pmol/μl)1μl、配列番号9で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LN1LFループプライマー:20pmol/μl)1μl、配列番号10で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LN1LBループプライマー:20pmol/μl)1μl、Bst DNA Polymerase(8U/25μL、栄研化学株式会社製)1μl、および上記で調製した鋳型DNA 1μlを混合し、蒸留水を加えて全量25μlの反応液とした。
上記で調製した反応液を、リアルタイム濁度測定装置Loopamp RT−160C(栄研化学株式会社製)にて、63±2℃で60分間DNAの増幅反応を行った。同時に反応液の濁度を測定した(波長:400nm)。
DNAの増幅の有無は、反応液の濁度が上昇しているかによって判断した。反応液の濁度の測定結果を、図11および図11−1に示す。
その結果、ネオサルトリア属の菌類(ネオサルトリア・スピノサ、ネオサルトリア・ヒラツカエ、ネオサルトリア・フィシェリ、ネオサルトリア・グラブラ)及びアスペルギルス・フミガタスのゲノムDNAを鋳型とした系のみで、反応開始20分前後から濁度の上昇、すなわちDNAの合成・増幅反応が認められた。
一方、その他の菌類のゲノムDNAを用いた系では、反応開始後50分までの間、反応液の濁度の上昇は認められなかった。なお、反応開始60分前後から、ネオサルトリア属の菌類及びアスペルギルス・フミガタス以外のゲノムDNAを用いた系においても反応液の濁度上昇が観察されたが、これはプライマー同士が反応して増幅してしまうこと、または、反応時間が長くなるにつれて標的配列以外の配列にも少数のプライマーがアニールしてしまうこと等が原因であると考えられる。
以上の結果から、本発明の方法によれば、簡便、迅速かつ特異的にネオサルトリア属の菌類及びアスペルギルス・フミガタスを検出することが可能である。
(1)プライマーの設計及び合成
シークエンシング法により決定した各種菌類(パエシロマイセス バリオッティー、ハミゲラ アベラネラ、タラロマイセス フラバス、タラロマイセス ルテウス、タラロマイセス トラキスパーマム、ビソクラミス ニベア、ビソクラミス フルバ、ネオサルトリア フィシェリ)のβ−チューブリン遺伝子の塩基配列情報をもとに、DNA解析ソフトウエア(商品名:DNAsis pro、日立ソフトウエア社製)を用いてアライメント解析を行い、ハミゲラ属の菌類に特異的な塩基配列を特定した。特定した塩基配列をもとに、配列番号16〜21の塩基配列で表されるオリゴヌクレオチドからなるプライマーを設計し、E Genome order(株式会社富士通システムソリューションズ、配列番号16,17;5pmolスケール、配列番号18,19;40pmolスケール、配列番号20,21;20pmolスケール;全てカラム精製品)に合成依頼し、購入した。
ハミゲラ属の菌類としては、表3に記載のハミゲラ アベラネア(Hamigera avellanea)を使用した。これらの菌類のβ−チューブリン遺伝子に対する配列番号16〜21の塩基配列で表されるオリゴヌクレオチドからなるプライマーのハミゲラ属の菌類特異性を示すために、表3に示す他の菌類も使用した。これらの菌類は千葉大学医学部真菌医学研究センターが保管し、IFMナンバーなどにより管理されているものを入手し、使用した。
各菌体を至適条件下で培養した。培養条件についてはポテトデキストロース培地(商品名:パールコア ポテトデキストロース寒天培地、栄研化学株式会社製)を用いて25℃で7日間培養した。
ゲノムDNA調製用キット(商品名 PrepMan ultra、アプライドバイオ社製)を用いて、菌体からゲノムDNA溶液を調製した。具体的には、各培地から数個のコロニーを採取し、キットの付属試薬200μlに菌体を懸濁し、100℃、10分間の加熱処理で菌体を溶解させ、14800rpmで5分間遠心分離した後、上清を回収した。得られたゲノムDNA溶液の濃度を50ng/μlに調製した。このゲノムDNA溶液を鋳型DNAとして、以下のLAMP反応に用いた。
2x Reaction Mix(Tris−HCl(pH8.8) 40mM、KCl 20mM、MgSO4 16mM、(NH4)2SO4 20mM、0.2%Tween20、Betaine 1.6M、dNTPs 2.8mM:栄研化学株式会社製;Loopamp DNA増幅試薬キット)12.5μl、配列番号16で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LH2F3プライマー:5pmol/μl)1μl、配列番号17で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LH2B3プライマー:5pmol/μl)1μl、配列番号18で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LH2FIPプライマー:40pmol/μl)1μl、配列番号19で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LH2BIPプライマー:40pmol/μl)1μl、配列番号20で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LH2LFループプライマー:20pmol/μl)1μl、配列番号21で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LH2LBループプライマー:20pmol/μl)1μl、Bst DNA Polymerase(8U/25μL、栄研化学株式会社製)1μl、および上記で調製した鋳型DNA 1μlを混合し、蒸留水を加えて全量25μlの反応液とした。
上記で調製した反応液を、リアルタイム濁度測定装置Loopamp RT−160C(栄研化学株式会社製)にて、63±2℃で60分間DNAの増幅反応を行った。同時に反応液の濁度を測定した(波長:400nm)。
DNAの増幅の有無は、反応液の濁度が上昇しているかによって判断した。反応液の濁度の測定結果を、図12に示す。
その結果、ハミゲラ属の菌類(ハミゲラ アベラネア)のゲノムDNAを鋳型とした系のみで、反応開始25分前後から濁度の上昇、すなわちDNAの合成・増幅反応が認められた。
一方、ハミゲラ属以外の菌類のゲノムDNAを用いた系では、反応開始後100分までの間、反応液の濁度の上昇は認められなかった。なお、反応開始110分前後から、ハミゲラ属以外の菌類のゲノムDNAを用いた系においても反応液の濁度上昇が観察されたが、これはプライマー同士が反応して増幅してしまうこと、または、反応時間が長くなるにつれて標的配列以外の配列にも少数のプライマーがアニールしてしまうこと等が原因であると考えられる。
以上の結果から、本発明の方法によれば、簡便、迅速かつ特異的にハミゲラ属の菌類を検出することが可能である。
(1)プライマーの設計及び調整
シークエンシング法により決定した各種菌類(アスペルギルス・フミガタス、ネオサルトリア フィシェリ、ネオサルトリア スピノサ)のβ−チューブリン遺伝子の塩基配列情報をもとに、DNA解析ソフトウエア(商品名:DNAsis pro、日立ソフトウエア社製)を用いてアライメント解析を行い、アスペルギルス・フミガタスに特異的な塩基配列を特定した。特定した塩基配列をもとに、配列番号11〜15の塩基配列で表されるオリゴヌクレオチドからなるプライマーを設計し、E Genome order(株式会社富士通システムソリューションズ、配列番号11,12;5pmolスケール、配列番号13,14;40pmolスケール、配列番号15;20pmolスケール;全てカラム精製品)に合成依頼し、購入した。
ネオサルトリア属に属する菌類およびアスペルギルス・フミガタスとしては、表4に記載の菌類を使用した。これらの菌類は千葉大学医学部真菌医学研究センターが保管し、IFMナンバーなどにより管理されているものを入手し、使用した。
各菌体を至適条件下で培養した。培養条件についてはポテトデキストロース培地(商品名:パールコア ポテトデキストロース寒天培地、栄研化学株式会社製)を用いて25℃で7日間培養した。
ゲノムDNA調製用キット(商品名 PrepMan ultra、アプライドバイオ社製)を用いて、菌体からゲノムDNA溶液を調製した。具体的には、各培地から数個のコロニーを採取し、キットの付属試薬200μlに菌体を懸濁し、100℃、10分間の加熱処理で菌体を溶解させ、14800rpmで5分間遠心分離した後、上清を回収した。得られたゲノムDNA溶液の濃度を50ng/μlに調製した。このゲノムDNA溶液を鋳型DNAとして、以下のLAMP反応に用いた。
2x Reaction Mix(Tris−HCl(pH8.8) 40mM、KCl 20mM、MgSO4 16mM、(NH4)2SO4 20mM、0.2%Tween20、Betaine 1.6M、dNTPs 2.8mM:栄研化学株式会社製;Loopamp DNA増幅試薬キット)12.5μl、配列番号11で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LAf2F3プライマー:5pmol/μl)1μl、配列番号12で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LAf2B3プライマー:5pmol/μl)1μl、配列番号13で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LAf2FIPプライマー:40pmol/μl)1μl、配列番号14で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LAf2BIPプライマー:40pmol/μl)1μl、配列番号15で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LAf2LBループプライマー:20pmol/μl)1μl、Bst DNA Polymerase((8U/25μL)栄研化学株式会社製)1μl、鋳型DNA 1μl、及び蒸留水で全量を25μlに調製した。
上記で調製した反応液を、リアルタイム濁度測定装置Loopamp RT−160C(栄研化学株式会社製)にて、63±2℃で60分間DNAの増幅反応を行った。同時に反応液の濁度を測定した(波長:400nm)。
DNAの増幅の有無は、反応液の濁度が上昇しているかによって判断した。反応液の濁度の測定結果を、図13に示す。
その結果、アスペルギルス・フミガタスのゲノムDNAを鋳型とした系のみで、反応開始45分前後から濁度の上昇、すなわちDNAの合成・増幅反応が認められた。
一方、ネオサルトリア属の菌類のゲノムDNAを用いた系では、反応開始後90分までの間、反応液の濁度の上昇は認められなかった。なお、反応開始100分前後から、ネオサルトリア属の菌類のゲノムDNAを用いた系においても反応液の濁度上昇が観察されたが、これはプライマー同士が反応して増幅してしまうこと、または、反応時間が長くなるにつれて標的配列以外の配列にも少数のプライマーがアニールしてしまうこと等が原因であると考えられる。
以上の結果から、本発明の方法によれば、簡便、迅速かつ特異的にアスペルギルス・フミガタスを同定することが可能である。
また、本実施例のアスペルギルス・フミガタスの検出方法と、実施例2に示した配列番号5〜10で示される塩基配列で表されるオリゴヌクレオチドからなるプライマーを用いた方法とを組合わせることで、ネオサルトリア属の菌類とアスペルギルス・フミガタスとを識別することも可能となる。
(1)プライマーの設計及び合成
シークエンシング法により決定した各種菌類(パエシロマイセス バリオッティー、ハミゲラ アベラネラ、タラロマイセス フラバス、タラロマイセス ルテウス、タラロマイセス トラキスパーマム、ビソクラミス ニベア、ビソクラミス フルバ、ネオサルトリア フィシェリ)のβ−チューブリン遺伝子の塩基配列情報をもとに、DNA解析ソフトウエア(商品名:DNAsis pro、日立ソフトウエア社製)を用いてアライメント解析を行い、タラロマイセス・フラバスに特異的な塩基配列を特定した。特定した塩基配列をもとに、配列番号22〜26の塩基配列で表されるオリゴヌクレオチドからなるプライマーを設計し、E Genome order(株式会社富士通システムソリューションズ、配列番号22,23;5pmolスケール、配列番号24,25;40pmolスケール、配列番号26;20pmolスケール;全てカラム精製品)に合成依頼し、購入した。
表5に記載のタラロマイセス・フラバスを使用した。タラロマイセス・フラバスのβ−チューブリン遺伝子に対する配列番号22〜26の塩基配列で表されるオリゴヌクレオチドからなるプライマーの特異性を示すために、表5に示す他の菌類も使用した。これらの菌類は千葉大学医学部真菌医学研究センターが保管し、IFMナンバーなどにより管理されているものを入手し、使用した。
各菌体を至適条件下で培養した。培養条件についてはポテトデキストロース培地(商品名:パールコア ポテトデキストロース寒天培地、栄研化学株式会社製)を用いて25℃で7日間培養した。
ゲノムDNA調製用キット(商品名 PrepMan ultra、アプライドバイオ社製)を用いて、菌体からゲノムDNA溶液を調製した。具体的には、各培地から数個のコロニーを採取し、キットの付属試薬200μlに菌体を懸濁し、100℃、10分間の加熱処理で菌体を溶解させ、14800rpmで5分間遠心分離した後、上清を回収した。得られたゲノムDNA溶液の濃度を50ng/μlに調製した。このゲノムDNA溶液を鋳型DNAとして、以下のLAMP反応に用いた。
2x Reaction Mix(Tris−HCl(pH8.8) 40mM、KCl 20mM、MgSO4 16mM、(NH4)2SO4 20mM、0.2%Tween20、Betaine 1.6M、dNTPs 2.8mM:栄研化学株式会社製;Loopamp DNA増幅試薬キット)12.5μl、配列番号22で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTf2F3プライマー:5pmol/μl)1μl、配列番号23で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTf2B3プライマー:5pmol/μl)1μl、配列番号24で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTf2FIPプライマー:40pmol/μl)1μl、配列番号25で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTf2BIPプライマー:40pmol/μl)1μl、配列番号26で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTf2LBループプライマー:20pmol/μl)1μl、Bst DNA Polymerase(8U/25μL、栄研化学株式会社製)1μl、および上記で調製した鋳型DNA 1μlを混合し、蒸留水を加えて全量25μlの反応液とした。
上記で調製した反応液を、リアルタイム濁度測定装置Loopamp RT−160C(栄研化学株式会社製)にて、63±2℃で60分間DNAの増幅反応を行った。同時に反応液の濁度を測定した(波長:400nm)。
DNAの増幅の有無は、反応液の濁度が上昇しているかによって判断した。反応液の濁度の測定結果を、図14(a)及び図14(b)に示す。なお、図14(a)は表5の試料番号1〜8、図14(b)は表5の試料番号9〜16の結果を示す。
その結果、タラロマイセス・フラバスのゲノムDNAを鋳型とした系のみで、反応開始30分前後から濁度の上昇、すなわちDNAの合成・増幅反応が認められた。
一方、タラロマイセス・フラバス以外の菌類のゲノムDNAを用いた系では、反応開始後70分までの間、反応液の濁度の上昇は認められなかった。なお、反応開始80分前後から、タラロマイセス・フラバス以外の菌類のゲノムDNAを用いた系においても反応液の濁度上昇が観察されたが、これはプライマー同士が反応して増幅してしまうこと、または、反応時間が長くなるにつれて標的配列以外の配列にも少数のプライマーがアニールしてしまうこと等が原因であると考えられる。
以上の結果から、本発明の方法によれば、タラロマイセス・フラバスのDNAのみが増幅され、かつ増幅による濁度の上昇が顕著な反応開始60分前後の時点までの反応液の濁度を計測することで、簡便かつ迅速にタラロマイセス・フラバスを検出することが可能である。
(1)プライマーの設計及び合成
シークエンシング法により決定した各種菌類(タラロマイセス ウォルトマンニー、パエシロマイセス バリオッティー、ハミゲラ アベラネラ、タラロマイセス フラバス、タラロマイセス ルテウス、タラロマイセス トラキスパーマム、ビソクラミス ニベア、ビソクラミス フルバ、ネオサルトリア フィシェリ)のβ−チューブリン遺伝子の塩基配列情報をもとに、DNA解析ソフトウエア(商品名:DNAsis pro、日立ソフトウエア社製)を用いてアライメント解析を行い、タラロマイセス・ウォルトマニーに特異的な塩基配列を特定した。特定した塩基配列をもとに、配列番号27〜32の塩基配列で表されるオリゴヌクレオチドからなるプライマーを設計し、E Genome order(株式会社富士通システムソリューションズ、配列番号27,28;5pmolスケール、配列番号29,30;40pmolスケール、配列番号31,32;20pmolスケール;全てカラム精製品)に合成依頼し、購入した。
表6に記載のタラロマイセス・ウォルトマニーを使用した。タラロマイセス・ウォルトマニーのβ−チューブリン遺伝子に対する配列番号27〜32の塩基配列で表されるオリゴヌクレオチドからなるプライマーの特異性を示すために、表6に示す他の菌類も使用した。これらの菌類は千葉大学医学部真菌医学研究センターが保管し、IFMナンバーなどにより管理されているものを入手し、使用した。
各菌体を至適条件下で培養した。培養条件についてはポテトデキストロース培地(商品名:パールコア ポテトデキストロース寒天培地、栄研化学株式会社製)を用いて25℃で7日間培養した。
ゲノムDNA調製用キット(商品名 PrepMan ultra、アプライドバイオ社製)を用いて、菌体からゲノムDNA溶液を調製した。具体的には、各培地から数個のコロニーを採取し、キットの付属試薬200μlに菌体を懸濁し、100℃、10分間の加熱処理で菌体を溶解させ、14800rpmで5分間遠心分離した後、上清を回収した。得られたゲノムDNA溶液の濃度を50ng/μlに調製した。このゲノムDNA溶液を鋳型DNAとして、以下のLAMP反応に用いた。
2x Reaction Mix(Tris−HCl(pH8.8) 40mM、KCl 20mM、MgSO4 16mM、(NH4)2SO4 20mM、0.2%Tween20、Betaine 1.6M、dNTPs 2.8mM:栄研化学株式会社製;Loopamp DNA増幅試薬キット)12.5μl、配列番号27で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTw4F3プライマー:5pmol/μl)1μl、配列番号28で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTw3B3プライマー:5pmol/μl)1μl、配列番号29で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTw4FIPプライマー:40pmol/μl)1μl、配列番号30で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTw3BIPプライマー:40pmol/μl)1μl、配列番号31で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTw4LFループプライマー:20pmol/μl)1μl、配列番号32で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTw3LBループプライマー:20pmol/μl)1μl、Bst DNA Polymerase(8U/25μL、栄研化学株式会社製)1μl、および上記で調製した鋳型DNA 1μlを混合し、蒸留水を加えて全量25μlの反応液とした。
上記で調製した反応液を、リアルタイム濁度測定装置Loopamp RT−160C(栄研化学株式会社製)にて、63±2℃で60分間DNAの増幅反応を行った。同時に反応液の濁度を測定した(波長:400nm)。
DNAの増幅の有無は、反応液の濁度が上昇しているかによって判断した。反応液の濁度の測定結果を、図15に示す。
その結果、タラロマイセス・ウォルトマニーのゲノムDNAを鋳型とした系のみで、反応開始20分前後から濁度の上昇、すなわちDNAの合成・増幅反応が認められた。
一方、タラロマイセス・ウォルトマニー以外の菌類のゲノムDNAを用いた系では、反応開始後40分までの間、反応液の濁度の上昇は認められなかった。なお、反応開始50分前後から、タラロマイセス・ウォルトマニー以外の菌類のゲノムDNAを用いた系においても反応液の濁度上昇が観察されたが、これはプライマー同士が反応して増幅してしまうこと、または、反応時間が長くなるにつれて標的配列以外の配列にも少数のプライマーがアニールしてしまうこと等が原因であると考えられる。
以上の結果から、簡便、迅速かつ特異的にタラロマイセス・ウォルトマニーを検出することが可能である。
(1)プライマーの設計及び合成
シークエンシング法により決定した各種菌類(タラロマイセス ルテウス、パエシロマイセス バリオッティー、ハミゲラ アベラネラ、タラロマイセス フラバス、タラロマイセス ウォルトマンニー、タラロマイセス トラキスパーマム、ビソクラミス ニベア、ビソクラミス フルバ、ネオサルトリア フィシェリ)のβ−チューブリン遺伝子の塩基配列情報をもとに、DNA解析ソフトウエア(商品名:DNAsis pro、日立ソフトウエア社製)を用いてアライメント解析を行い、タラロマイセス・ルテウスに特異的な塩基配列を特定した。特定した塩基配列をもとに、配列番号33〜37の塩基配列で表されるオリゴヌクレオチドからなるプライマーを設計し、E Genome order(株式会社富士通システムソリューションズ、配列番号33,34;5pmolスケール、配列番号35,36;40pmolスケール、配列番号37;20pmolスケール;全てカラム精製品)に合成依頼し、購入した。
表7に記載のタラロマイセス・ルテウスを使用した。タラロマイセス・ルテウスのβ−チューブリン遺伝子に対する配列番号33〜37の塩基配列で表されるオリゴヌクレオチドからなるプライマーの特異性を示すために、表7に示す他の菌類も使用した。これらの菌類は千葉大学医学部真菌医学研究センターが保管し、IFMナンバーなどにより管理されているものを入手し、使用した。
各菌体を至適条件下で培養した。培養条件についてはポテトデキストロース培地(商品名:パールコア ポテトデキストロース寒天培地、栄研化学株式会社製)を用いて25℃で7日間培養した。
ゲノムDNA調製用キット(商品名 PrepMan ultra、アプライドバイオ社製)を用いて、菌体からゲノムDNA溶液を調製した。具体的には、各培地から数個のコロニーを採取し、キットの付属試薬200μlに菌体を懸濁し、100℃、10分間の加熱処理で菌体を溶解させ、14800rpmで5分間遠心分離した後、上清を回収した。得られたゲノムDNA溶液の濃度を50ng/μlに調製した。このゲノムDNA溶液を鋳型DNAとして、以下のLAMP反応に用いた。
2x Reaction Mix(Tris−HCl(pH8.8) 40mM、KCl 20mM、MgSO4 16mM、(NH4)2SO4 20mM、0.2%Tween20、Betaine 1.6M、dNTPs 2.8mM:栄研化学株式会社製;Loopamp DNA増幅試薬キット)12.5μl、配列番号33で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTl1F3プライマー:5pmol)1μl、配列番号34で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTl1B3プライマー:5pmol)1μl、配列番号35で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTl1FIPプライマー:40pmol)1μl、配列番号36で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTl1BIPプライマー:40pmol)1μl、配列番号37で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LTl1LFループプライマー:20pmol)1μl、Bst DNA Polymerase(8U/25μL、栄研化学株式会社製)1μl、および上記で調製した鋳型DNA 1μlを混合し、蒸留水を加えて全量25μlの反応液とした。
上記で調製した反応液を、リアルタイム濁度測定装置Loopamp RT−160C(栄研化学株式会社製)にて、63±2℃で60分間DNAの増幅反応を行った。同時に反応液の濁度を測定した(波長:400nm)。
DNAの増幅の有無は、反応液の濁度が上昇しているかによって判断した。反応液の濁度の測定結果を、図16(a)及び図16(b)に示す。なお、図16(a)は表7の試料番号1〜8、図16(b)は表7の試料番号9〜16の結果を示す。
その結果、タラロマイセス・ルテウスのゲノムDNAを鋳型とした系のみで、反応開始25分前後から濁度の急激な上昇、すなわちDNAの合成・増幅反応が認められた。
一方、タラロマイセス・ルテウス以外の菌類のゲノムDNAを用いた系では、反応開始後80分までの間、反応液の濁度の上昇は認められなかった。なお、反応開始90分前後から、タラロマイセス・ルテウス以外の菌類のゲノムDNAを用いた系においても反応液の濁度上昇が観察されたが、これはプライマー同士が反応して増幅してしまうこと、または、反応時間が長くなるにつれて標的配列以外の配列にも少数のプライマーがアニールしてしまうこと等が原因であると考えられる。また、試料番号8では、反応開始10分後から徐々に濁度の上昇が見られたが、これはゲノムDNAの特異的な塩基配列に対応した遺伝子の増幅ではなく、プライマー同士が反応した緩慢な遺伝子増幅であることが理由であると考えられる。この緩慢な遺伝子増幅は濁度の測定結果に上昇ピークが見られない等、プライマーが鋳型DNAにアニールして増幅反応が起こる本来のLAMP反応とは測定結果等において明らかに相違し、明確に区別することができる。
以上の結果から、本発明の方法によれば、簡便、迅速かつ特異的にタラロマイセス・ルテウスを同定することが可能である。
(1)プライマーの設計及び合成
シークエンシング法により決定した各種菌類(タラロマイセス フラバス、タラロマイセス トラキスパーマム、パエシロマイセス バリオッティー、ハミゲラ アベラネラ、タラロマイセス ウォルトマンニー、ビソクラミス ニベア、ビソクラミス フルバ、ネオサルトリア フィシェリ)の28SrDNA遺伝子のITS領域及びD1/D2領域の塩基配列情報をもとに、DNA解析ソフトウエア(商品名:DNAsis pro、日立ソフトウエア社製)を用いてアライメント解析を行い、タラロマイセス・フラバスおよびタラロマイセス・トラキスパーマムに特異的な塩基配列を特定した。特定した塩基配列をもとに、配列番号38〜43の塩基配列で表されるオリゴヌクレオチドからなるプライマーを設計し、E Genome order(株式会社富士通システムソリューションズ、配列番号38,39;5pmolスケール、配列番号40,41;40pmolスケール、配列番号42,43;20pmolスケール;全てカラム精製品)に合成依頼し、購入した。
表8に記載のタラロマイセス・フラバスおよびタラロマイセス・トラキスパーマムを使用した。これらの菌類の28SrDNAのITS領域及びD1/D2領域に対する配列番号38〜43の塩基配列で表されるオリゴヌクレオチドからなるプライマーの特異性を示すために、表8に示す他の菌類も使用した。これらの菌類は千葉大学医学部真菌医学研究センターが保管し、IFMナンバーなどにより管理されているものを入手し、使用した。
各菌体を至適条件下で培養した。培養条件についてはポテトデキストロース培地(商品名:パールコア ポテトデキストロース寒天培地、栄研化学株式会社製)を用いて25℃で7日間培養した。
ゲノムDNA調製用キット(商品名 PrepMan ultra、アプライドバイオ社製)を用いて、菌体からゲノムDNA溶液を調製した。具体的には、各培地から数個のコロニーを採取し、キットの付属試薬200μlに菌体を懸濁し、100℃、10分間の加熱処理で菌体を溶解させ、14800rpmで5分間遠心分離した後、上清を回収した。得られたゲノムDNA溶液の濃度を50ng/μlに調製した。このゲノムDNA溶液を鋳型DNAとして、以下のLAMP反応に用いた。
2x Reaction Mix(Tris−HCl(pH8.8) 40mM、KCl 20mM、MgSO4 16mM、(NH4)2SO4 20mM、0.2%Tween20、Betaine 1.6M、dNTPs 2.8mM:栄研化学株式会社製;Loopamp DNA増幅試薬キット)12.5μl、配列番号38で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LT1F3プライマー:5pmol/μl)1μl、配列番号39で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LT1B3プライマー:5poml/μl)1μl、配列番号40で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LT1FIPプライマー:40pmol/μl)1μl、配列番号41で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LT1BIPプライマー:40pmol/μl)1μl、配列番号42で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LT1LFプライマー:20pmol/μl)1μl、配列番号43で示される塩基配列で表されるオリゴヌクレオチドからなるプライマー(LT1LBループプライマー:20pmol/μl)1μl、Bst DNA Polymerase(8U/25μL、栄研化学株式会社製)1μl、および上記で調製した鋳型DNA 1μlを混合し、蒸留水を加えて全量25μlの反応液とした。
上記で調製した反応液を、リアルタイム濁度測定装置Loopamp RT−160C(栄研化学株式会社製)にて、63±2℃で60分間DNAの増幅反応を行った。同時に反応液の濁度を測定した(波長:400nm)。
DNAの増幅の有無は、反応液の濁度が上昇しているかによって判断した。反応液の濁度の測定結果を、図17に示す。
その結果、タラロマイセス・フラバスおよびタラロマイセス・トラキスパーマムのゲノムDNAを鋳型とした系のみで、反応開始40分前後から濁度の上昇、すなわちDNAの合成・増幅反応が認められた。
一方、タラロマイセス・フラバスおよびタラロマイセス・トラキスパーマム以外の菌類のゲノムDNAを用いた系では、反応開始後45分までの間、反応液の濁度の上昇は認められなかった。なお、反応開始50分後から、タラロマイセス・フラバスおよびタラロマイセス・トラキスパーマム以外の菌類のゲノムDNAを用いた系においても反応液の濁度上昇が観察されたが、これはプライマー同士が反応して増幅してしまうこと、または、反応時間が長くなるにつれて標的配列以外の配列にも少数のプライマーがアニールしてしまうこと等が原因であると考えられる。
以上の結果から、本発明の方法によれば、簡便、迅速かつ特異的にタラロマイセス・フラバスおよびタラロマイセス・トラキスパーマムを検出することが可能である。
Claims (44)
- Loop mediated isothermal amplification(LAMP)法によって耐熱性菌類の遺伝子中の標的領域を含むDNA断片を増幅させ、増幅産物の有無を確認する工程を含む耐熱性菌類の検出方法。
- 前記標的領域がβ−チューブリン遺伝子、または28S rDNAのD1/D2領域及びITS領域の塩基配列から選択される請求項1記載の検出方法。
- 前記耐熱性菌類がビソクラミス(Byssochlamys)属の菌類、タラロマイセス(Talaromyces)属の菌類、ネオサルトリア(Neosartorya)属の菌類、ハミゲラ(Hamigera)属の菌類、およびアスペルギルス フミガタス(Aspergillus fumigatus)からなる群より選ばれる少なくとも1種である請求項1または2記載の検出方法。
- 前記標的領域を含むDNA断片が、
配列番号44に記載の塩基配列の一部であって当該塩基配列の400〜600位の塩基配列の全部又は一部を含む塩基配列、
配列番号45に記載の塩基配列の一部であって当該塩基配列の10〜250位及び/又は350〜559位の塩基配列の全部又は一部を含む塩基配列、
配列番号46に記載の塩基配列の一部であって当該塩基配列の10〜250位及び/又は350〜559位の塩基配列の全部又は一部を含む塩基配列、
配列番号47に記載の塩基配列の一部であって当該塩基配列の300〜550位の塩基配列の全部又は一部を含む塩基配列、
配列番号48に記載の塩基配列の一部であって当該塩基配列の200〜450位の塩基配列の全部又は一部を含む塩基配列、
配列番号49に記載の塩基配列の一部であって当該塩基配列の150〜420位の塩基配列の全部又は一部を含む塩基配列、
配列番号50に記載の塩基配列の一部であって当該塩基配列の150〜450位の塩基配列の全部又は一部を含む塩基配列、
配列番号51に記載の塩基配列の一部であって当該塩基配列の250〜550位の塩基配列の全部又は一部を含む塩基配列、
配列番号52に記載の塩基配列の一部であって当該塩基配列の250〜550位の塩基配列の全部又は一部を含む塩基配列、および
これらの塩基配列において1若しくは数個の塩基が欠失、置換若しくは付加された塩基配列、
からなる群より選ばれる塩基配列を含むことを特徴とする請求項1〜3のいずれか記載の検出方法。 - 請求項1〜3のいずれか記載の検出方法であって、ビソクラミス属の菌類検出用プライマーセットとして配列番号1〜4に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーセットを用いてLAMP法により28S rDNAのD1/D2領域及びITS領域中の標的領域を含むDNA断片を増幅する請求項1〜3のいずれか記載の検出方法。
- 請求項1〜3のいずれか記載の検出方法であって、ネオサルトリア属の菌類およびアスペルギルス フミガタス検出用プライマーセットとして配列番号5〜8に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーセットを用いてLAMP法によりβ−チューブリン遺伝子中の標的領域を含むDNA断片を増幅する請求項1〜3のいずれか記載の検出方法。
- 請求項6記載の検出方法であって、さらに配列番号9に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーと、配列番号10に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーのうちの少なくとも一方を用いる請求項6記載の検出方法。
- 請求項1〜3のいずれか記載の検出方法であって、アスペルギルス フミガタス検出用プライマーセットとして配列番号11〜14に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーセットを用いてLAMP法によりβ−チューブリン遺伝子中の標的領域を含むDNA断片を増幅する請求項1〜3のいずれか記載の検出方法。
- 請求項8記載の検出方法であって、さらに配列番号15に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーを用いる請求項8記載の検出方法。
- 請求項1〜3のいずれか記載の検出方法であって、ハミゲラ属の菌類検出用プライマーセットとして配列番号16〜19に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーセットを用いてLAMP法によりβ−チューブリン遺伝子中の標的領域を含むDNA断片を増幅する請求項1〜3のいずれか記載の検出方法。
- 請求項10記載の検出方法であって、さらに配列番号20に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーと、配列番号21に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーのうちの少なくとも一方を用いる請求項10記載の検出方法。
- 請求項1〜3のいずれか記載の検出方法であって、タラロマイセス属の菌類のうちタラロマイセス フラバス(Talaromyces flavus)を検出するためのプライマーセットとして配列番号22〜25に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーセットを用いてLAMP法によりβ−チューブリン遺伝子中の標的領域を含むDNA断片を増幅する請求項1〜3のいずれか記載の検出方法。
- 請求項12記載の検出方法であって、さらに配列番号26に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーを用いる請求項12記載の検出方法。
- 請求項1〜3のいずれか記載の検出方法であって、タラロマイセス属の菌類のうちタラロマイセス ウォルトマニー(Talaromyces wortmannii)を検出するためのプライマーセットとして配列番号27〜30に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーセットを用いてLAMP法によりβ−チューブリン遺伝子中の標的領域を含むDNA断片を増幅する請求項1〜3のいずれか記載の検出方法。
- 請求項14記載の検出方法であって、さらに配列番号31に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーと、配列番号32に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーのうちの少なくとも一方を用いる請求項14記載の検出方法。
- 請求項1〜3のいずれか記載の検出方法であって、タラロマイセス属の菌類のうちタラロマイセス ルテウス(Talaromyces luteus)を検出するためのプライマーセットとして配列番号33〜36に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーセットを用いてLAMP法によりβ−チューブリン遺伝子中の標的領域を含むDNA断片を増幅する請求項1〜3のいずれか記載の検出方法。
- 請求項16記載の検出方法であって、さらに配列番号37に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーを用いる請求項16記載の検出方法。
- 請求項1〜3のいずれか記載の検出方法であって、タラロマイセス属の菌類のうちタラロマイセス フラバス(Talaromyces flavus)及び/またはタラロマイセス トラキスパーマム(Talaromyces trachyspermus)を検出するためのプライマーセットとして配列番号38〜41に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーセットを用いてLAMP法により28S rDNAのD1/D2領域及びITS領域中の標的領域を含むDNA断片を増幅する請求項1〜3のいずれか記載の検出方法。
- 請求項18記載の検出方法であって、さらに配列番号42に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーと、配列番号43に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマーのうちの少なくとも一方を用いる請求項18記載の検出方法。
- ビソクラミス(Byssochlamys)属の菌類をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号1に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号2に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号3に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号4に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - ネオサルトリア(Neosartorya)属の菌類およびアスペルギルス フミガタス(Aspergillus fumigatus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号5に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号6に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号7に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号8に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - ネオサルトリア(Neosartorya)属の菌類およびアスペルギルス フミガタス(Aspergillus fumigatus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号5に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号6に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号7に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号8に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号9に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号10に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - アスペルギルス フミガタス(Aspergillus fumigatus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号11に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号12に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号13に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号14に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - アスペルギルス フミガタス(Aspergillus fumigatus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号11に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号12に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号13に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号14に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号15に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - ハミゲラ(Hamigera)属の菌類をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号16に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号17に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号18に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号19に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - ハミゲラ(Hamigera)属の菌類をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号16に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号17に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号18に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号19に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号20に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号21に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - タラロマイセス フラバス(Talaromyces flavus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号22に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号23に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号24に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号25に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - タラロマイセス フラバス(Talaromyces flavus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号22に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号23に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号24に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号25に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号26に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - タラロマイセス ウォルトマニー(Talaromyces wortmannii)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号27に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号28に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号29に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号30に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - タラロマイセス ウォルトマニー(Talaromyces wortmannii)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号27に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号28に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号29に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号30に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号31に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号32に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - タラロマイセス ルテウス(Talaromyces luteus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号33に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号34に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号35に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号36に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - タラロマイセス ルテウス(Talaromyces luteus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号33に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号34に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号35に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号36に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号37に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - タラロマイセス フラバス(Byssochlamys fulva)及び/またはタラロマイセス トラキスパーマム(Talaromyces trachyspermus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号38に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号39に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号40に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号41に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - タラロマイセス フラバス(Byssochlamys fulva)及び/またはタラロマイセス トラキスパーマム(Talaromyces trachyspermus)をLoop mediated isothermal amplification(LAMP)法で検出するのに用いるプライマーセットであって、
配列番号38に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号39に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号40に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号41に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、
配列番号42に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー、および
配列番号43に記載の塩基配列で表されるオリゴヌクレオチドからなるプライマー
を含むプライマーセット。 - ビソクラミス(Byssochlamys)属の菌類検出用オリゴヌクレオチドであって、配列番号1〜4のいずれかの配列番号に記載の塩基配列で表されるオリゴヌクレオチド、または当該塩基配列に対して70%以上の相同性を有しかつプライマーとして使用できる塩基配列で表されるオリゴヌクレオチド。
- ネオサルトリア(Neosartorya)属の菌類およびアスペルギルス フミガタス(Aspergillus fumigatus)検出用オリゴヌクレオチドであって、配列番号5〜10のいずれかの配列番号に記載の塩基配列で表されるオリゴヌクレオチド、または当該塩基配列に対して70%以上の相同性を有しかつプライマーとして使用できる塩基配列で表されるオリゴヌクレオチド。
- アスペルギルス フミガタス(Aspergillus fumigatus)検出用オリゴヌクレオチドであって、配列番号11〜15のいずれかの配列番号に記載の塩基配列で表されるオリゴヌクレオチド、または当該塩基配列に対して70%以上の相同性を有しかつプライマーとして使用できる塩基配列で表されるオリゴヌクレオチド。
- ハミゲラ(Hamigera)属の菌類検出用オリゴヌクレオチドであって、配列番号16〜21のいずれかの配列番号に記載の塩基配列で表されるオリゴヌクレオチド、または当該塩基配列に対して70%以上の相同性を有しかつプライマーとして使用できる塩基配列で表されるオリゴヌクレオチド。
- タラロマイセス フラバス(Talaromyces flavus)検出用オリゴヌクレオチドであって、配列番号22〜26のいずれかの配列番号に記載の塩基配列で表されるオリゴヌクレオチド、または当該塩基配列に対して70%以上の相同性を有しかつプライマーとして使用できる塩基配列で表されるオリゴヌクレオチド。
- タラロマイセス ウォルトマニー(Talaromyces wortmannii)検出用オリゴヌクレオチドであって、配列番号27〜32のいずれかの配列番号に記載の塩基配列で表されるオリゴヌクレオチド、または当該塩基配列に対して70%以上の相同性を有しかつプライマーとして使用できる塩基配列で表されるオリゴヌクレオチド。
- タラロマイセス ルテウス(Talaromyces luteus)検出用オリゴヌクレオチドであって、配列番号33〜37のいずれかの配列番号に記載の塩基配列で表されるオリゴヌクレオチド、または当該塩基配列に対して70%以上の相同性を有しかつプライマーとして使用できる塩基配列で表されるオリゴヌクレオチド。
- タラロマイセス フラバス(Talaromyces flavus)及び/またはタラロマイセス トラキスパーマム(Talaromyces trachyspermus)検出用オリゴヌクレオチドであって、配列番号38〜43のいずれかの配列番号に記載の塩基配列で表されるオリゴヌクレオチド、または当該塩基配列に対して70%以上の相同性を有しかつプライマーとして使用できる塩基配列で表されるオリゴヌクレオチド。
- β−チューブリン遺伝子、または28S rDNAのD1/D2領域及びITS領域の塩基配列から選択される標的領域において、配列の5’側から順に、塩基配列領域としてF3、F2及びF1を選択し、
前記標的領域の3’側から順に、塩基配列領域としてB3c、B2c及びB1cを選択し、
前記F3、F2及びF1の相補的塩基配列を、それぞれF3c、F2c及びF1cとし、
前記B3c、B2c及びB1cに相補的な塩基配列を、それぞれB3、B2及びB1としたとき、
以下の(a)〜(f)のいずれかに該当する塩基配列からなる、耐熱性菌類検出用のオリゴヌクレオチド。
(a)前記B2領域を3’側に有し、前記B1c領域を5’側に有する塩基配列
(b)前記B3領域を有する塩基配列
(c)前記F2領域を3’側に有し、前記F1c領域を5’側に有する塩基配列
(d)前記F3領域を有する塩基配列
(e)前記B1領域と前記B2領域の間の部分と相補的な配列を有する塩基配列
(f)前記F1領域と前記F2領域の間の部分と相補的な配列を有する塩基配列 - 請求項20〜34のいずれか記載のプライマーセットまたは請求項35〜43のいずれか記載のオリゴヌクレオチドと、
DNAポリメラーゼと、
dATP、dCTP、dGTP及びdTTPを含むdNTPと
を含む耐熱性菌類検出キット。
Priority Applications (16)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2008141499A JP5548345B2 (ja) | 2008-05-29 | 2008-05-29 | 耐熱性菌類の検出方法 |
| EP13198462.7A EP2712922B1 (en) | 2008-05-28 | 2009-05-28 | Method of detecting heat-resistant fungi |
| CN200980126323.2A CN102105584B (zh) | 2008-05-28 | 2009-05-28 | 耐热菌类的检测方法 |
| EP13198494.0A EP2712923B1 (en) | 2008-05-28 | 2009-05-28 | Method of detecting heat-resistant fungi |
| EP09754787.1A EP2298884B1 (en) | 2008-05-28 | 2009-05-28 | Method for detection of thermotolerant bacterium |
| CN201410140749.6A CN103923990B (zh) | 2008-05-28 | 2009-05-28 | 耐热菌类的检测方法 |
| EP13198538.4A EP2712924B1 (en) | 2008-05-28 | 2009-05-28 | Method of detecting heat-resistant fungus of the genus neosartorya and aspergillus fumigates |
| EP13198558.2A EP2712935B1 (en) | 2008-05-28 | 2009-05-28 | Method of detecting heat-resistant fungus of the genus Hamigera |
| US12/994,566 US9074261B2 (en) | 2008-05-28 | 2009-05-28 | Method of detecting heat-resistant fungus |
| CN201410141182.4A CN103923992B (zh) | 2008-05-28 | 2009-05-28 | 耐热菌类的检测方法 |
| CN201410141157.6A CN103923991B (zh) | 2008-05-28 | 2009-05-28 | 耐热菌类的检测方法 |
| CN201410140150.2A CN103923989B (zh) | 2008-05-28 | 2009-05-28 | 耐热菌类的检测方法 |
| PCT/JP2009/059818 WO2009145279A1 (ja) | 2008-05-28 | 2009-05-28 | 耐熱性菌類の検出方法 |
| US14/747,571 US10006095B2 (en) | 2008-05-28 | 2015-06-23 | Method of detecting heat-resistant fungus |
| US15/987,401 US10093993B1 (en) | 2008-05-28 | 2018-05-23 | Method of detecting heat-resistant fungus |
| US15/987,370 US10093992B1 (en) | 2008-05-28 | 2018-05-23 | Method of detecting heat-resistant fungus |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2008141499A JP5548345B2 (ja) | 2008-05-29 | 2008-05-29 | 耐熱性菌類の検出方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2009284832A true JP2009284832A (ja) | 2009-12-10 |
| JP5548345B2 JP5548345B2 (ja) | 2014-07-16 |
Family
ID=41454653
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2008141499A Expired - Fee Related JP5548345B2 (ja) | 2008-05-28 | 2008-05-29 | 耐熱性菌類の検出方法 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP5548345B2 (ja) |
Cited By (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2010115122A (ja) * | 2008-11-11 | 2010-05-27 | Kao Corp | アスペルギルスフミガタス(Aspergillusfumigatus)類縁菌の検出方法 |
| JP2011250746A (ja) * | 2010-06-03 | 2011-12-15 | Kao Corp | ビソクラミス属に属する菌類の検出方法 |
| WO2012169099A1 (ja) | 2011-06-09 | 2012-12-13 | 東洋製罐株式会社 | カビの検出方法、pcr用反応液、及びカビ検出用担体 |
| JP2014018091A (ja) * | 2012-07-12 | 2014-02-03 | Kao Corp | ケトミウム・フニコラの検出方法 |
| CN108315469A (zh) * | 2018-04-09 | 2018-07-24 | 中国农业科学院农产品加工研究所 | 环介导等温扩增法检测致病镰刀菌的引物组合物和试剂盒及其用途 |
| CN113322192A (zh) * | 2021-07-15 | 2021-08-31 | 云南省农业科学院药用植物研究所 | 一种黄精内生真菌抑制大肠杆菌剂及其制备方法和应用 |
-
2008
- 2008-05-29 JP JP2008141499A patent/JP5548345B2/ja not_active Expired - Fee Related
Non-Patent Citations (2)
| Title |
|---|
| JPN6009031761; 果実協会報, (2006), [569], p.4-15 * |
| JPN6013024710; 第26回日本食品微生物学会学術総会講演要旨集, (2005), p.42 * |
Cited By (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2010115122A (ja) * | 2008-11-11 | 2010-05-27 | Kao Corp | アスペルギルスフミガタス(Aspergillusfumigatus)類縁菌の検出方法 |
| JP2011250746A (ja) * | 2010-06-03 | 2011-12-15 | Kao Corp | ビソクラミス属に属する菌類の検出方法 |
| WO2012169099A1 (ja) | 2011-06-09 | 2012-12-13 | 東洋製罐株式会社 | カビの検出方法、pcr用反応液、及びカビ検出用担体 |
| EP2719772A4 (en) * | 2011-06-09 | 2014-11-26 | Toyo Seikan Group Holdings Ltd | METHOD FOR DETECTION OF FUNGI, REACTION SOLUTION FOR PCR AND VECTOR FOR DETECTION OF FUNGI |
| US10370729B2 (en) | 2011-06-09 | 2019-08-06 | Toyo Seikan Group Holdings, Ltd. | Method for detecting fungi, reaction solution for PCR, and carrier for detecting fungi |
| JP2014018091A (ja) * | 2012-07-12 | 2014-02-03 | Kao Corp | ケトミウム・フニコラの検出方法 |
| CN108315469A (zh) * | 2018-04-09 | 2018-07-24 | 中国农业科学院农产品加工研究所 | 环介导等温扩增法检测致病镰刀菌的引物组合物和试剂盒及其用途 |
| CN108315469B (zh) * | 2018-04-09 | 2021-10-22 | 中国农业科学院农产品加工研究所 | 环介导等温扩增法检测致病镰刀菌的引物组合物和试剂盒及其用途 |
| CN113322192A (zh) * | 2021-07-15 | 2021-08-31 | 云南省农业科学院药用植物研究所 | 一种黄精内生真菌抑制大肠杆菌剂及其制备方法和应用 |
| CN113322192B (zh) * | 2021-07-15 | 2023-05-16 | 云南省农业科学院药用植物研究所 | 一种黄精内生真菌抑制大肠杆菌剂及其制备方法和应用 |
Also Published As
| Publication number | Publication date |
|---|---|
| JP5548345B2 (ja) | 2014-07-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN102105584B (zh) | 耐热菌类的检测方法 | |
| JP5548345B2 (ja) | 耐熱性菌類の検出方法 | |
| JP5670623B2 (ja) | パエシロマイセスバリオッティの検出方法 | |
| JP5934038B2 (ja) | サーモアスカス属真菌の検出方法 | |
| CN102272305B (zh) | 属于Geosmithia属的菌类的检测方法 | |
| JP5654265B2 (ja) | ビソクラミス属に属する菌類の検出方法 | |
| JP5548357B2 (ja) | アスペルギルスフミガタス(Aspergillusfumigatus)類縁菌の検出方法 | |
| JP5548388B2 (ja) | ネオサルトリア属に属する菌類及びアスペルギルスフミガタスの検出方法 | |
| JP5548387B2 (ja) | タラロマイセス属に属する菌類の検出方法 | |
| JP5548389B2 (ja) | ビソクラミス属に属する菌類の検出方法 | |
| JP5697881B2 (ja) | パエシロマイセスサトゥラタス及びパエシロマイセスディバリカタスの検出方法 | |
| JP5548386B2 (ja) | ハミゲラ属に属する菌類及びクラドスポリウム属に属する菌類の検出方法 | |
| JP5799139B2 (ja) | パエシロマイセスサトゥラタス及びパエシロマイセスディバリカタスの検出方法 | |
| JP5764619B2 (ja) | アスペルギルス・フミガタス(Aspergillusfumigatus)類縁菌の検出方法 | |
| JP2015195774A (ja) | アスペルギルス・オクラセウス及びアスペルギルス・ウェスタジキアエの検出方法 | |
| JP2015195773A (ja) | アスペルギルス・カーボナリウスの検出方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20110310 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20110311 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20110524 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20130528 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20130729 Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20130729 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20140304 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20140411 Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20140411 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20140513 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20140519 |
|
| R151 | Written notification of patent or utility model registration |
Ref document number: 5548345 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R151 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| LAPS | Cancellation because of no payment of annual fees |