JP2019508018A - SorCSペプチド及びそれらの用途 - Google Patents
SorCSペプチド及びそれらの用途 Download PDFInfo
- Publication number
- JP2019508018A JP2019508018A JP2018531443A JP2018531443A JP2019508018A JP 2019508018 A JP2019508018 A JP 2019508018A JP 2018531443 A JP2018531443 A JP 2018531443A JP 2018531443 A JP2018531443 A JP 2018531443A JP 2019508018 A JP2019508018 A JP 2019508018A
- Authority
- JP
- Japan
- Prior art keywords
- peptide
- amino acid
- seq
- acid residues
- sorcs2
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images
























Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/02—Drugs for disorders of the nervous system for peripheral neuropathies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/06—Antimigraine agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/08—Antiepileptics; Anticonvulsants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/14—Drugs for disorders of the nervous system for treating abnormal movements, e.g. chorea, dyskinesia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/14—Drugs for disorders of the nervous system for treating abnormal movements, e.g. chorea, dyskinesia
- A61P25/16—Anti-Parkinson drugs
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/18—Antipsychotics, i.e. neuroleptics; Drugs for mania or schizophrenia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/22—Anxiolytics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/24—Antidepressants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/28—Drugs for disorders of the nervous system for treating neurodegenerative disorders of the central nervous system, e.g. nootropic agents, cognition enhancers, drugs for treating Alzheimer's disease or other forms of dementia
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/06—Linear peptides containing only normal peptide links having 5 to 11 amino acids
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/08—Linear peptides containing only normal peptide links having 12 to 20 amino acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Neurosurgery (AREA)
- Neurology (AREA)
- Biomedical Technology (AREA)
- Animal Behavior & Ethology (AREA)
- Veterinary Medicine (AREA)
- Pharmacology & Pharmacy (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Public Health (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Gastroenterology & Hepatology (AREA)
- Zoology (AREA)
- Toxicology (AREA)
- Psychiatry (AREA)
- Pain & Pain Management (AREA)
- Cell Biology (AREA)
- Immunology (AREA)
- Psychology (AREA)
- Hospice & Palliative Care (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
【選択図】図1
Description
(i)X1はスレオニン(T)またはイソロイシン(I)であり、
(ii)X2はホスホセリン(J)、アスパラギン酸(D)、グルタミン酸(E)、アラニン(A)、及びセリン(S)からなる群より選択され、
(iii)X3はプロリン(P)またはセリン(S)であり、
(iv)X4はホスホセリン(J)、アスパラギン酸(D)、グルタミン酸(E)、アラニン(A)、及びセリン(S)からなる群より選択され、
(v)X5はヒスチジン(H)またはグルタミン(Q)であり、
(vi)X6はホスホセリン(J)、アスパラギン酸(D)、グルタミン酸(E)、アラニン(A)、及びセリン(S)からなる群より選択される。
(i)1つ以上の候補化合物、及びSorCS2、SorCS1、SorCS3を発現する細胞を準備するステップ、
(ii)該候補化合物の非存在下で、該細胞における(i)のタンパク質のリン酸化レベルを測定するステップ、
(iii)(ii)の細胞を該候補化合物と接触させるステップ、
(iv)(iii)のタンパク質のリン酸化レベルを測定するステップ、
(v)(ii)と(iv)のリン酸化レベルを比較するステップ、
(vi)SorCS2、SorCS1、またはSorCS3のC末端ドメインのリン酸化を調節することができる(v)の化合物を選択することにより化合物を特定するステップ
を含む。
(i)1つ以上の候補化合物、ならびにSorCS2、SorCS1、及び/またはSorCS3を発現する細胞を準備するステップ、
(ii)該候補化合物の非存在下で、該細胞における(i)のタンパク質の発現レベルを測定するステップ、
(iii)(ii)の細胞を該候補化合物と接触させるステップ、
(iv)(iii)のタンパク質の発現レベルを測定するステップ、
(v)(ii)と(iv)の発現レベルを比較するステップ、
(vi)SorCS2、SorCS1、及び/またはSorCS3の発現を調節することができる(v)の化合物を選択することにより、化合物を特定するステップ
を含む。
細胞透過性ペプチド(CPP):この用語は、哺乳類細胞の細胞膜を貫通できることを特徴とし、そのためペプチド、タンパク質、オリゴヌクレオチドなどのカーゴ分子に結合して該カーゴ分子の細胞内送達をもたらすことができるペプチドを指す。
シナプス可塑性とは、本明細書では、シナプスが強度を変える能力であり、記憶形成を含むほとんどの認知プロセスの細胞の基本と考えられている。シナプスの強化または弱化はそれぞれ長期増強(LTP)と長期抑制(LTD)により生じる。ニューロン活性は、脳からの神経栄養因子BDNFのシナプス前放出を誘発し、次いでBDNFがシナプス後チロシンキナーゼ受容体TrkBと相互作用することが、LTPの初期相(E−LTP)の誘発に必要である(23)。さらに、BDNFの前駆体であるproBDNFの放出と、それに続く細胞外プロテイナーゼによるproBDNFから成熟BDNFへの変換が、LTPの後期相及びシナプス強化に必要である(25)。一方、シナプスの弱化は異なるメカニズムにより促進され、これにはproBDNFとシナプス後パンニューロトロフィン受容体p75(p75NTR)の相互作用が含まれ、その結果としてLTDがもたらされる(24)。樹状突起の成長は、シナプス活性により、また近隣細胞からの分子シグナルによっても局所的に制御される。このような活性依存型の構造変化は、局所のシナプスの有効性と共作用して、神経系の特殊な接続性のパターンを形作る。
本発明は、各請求項で定められるとおりである。
(i)X1は、スレオニン(T)、イソロイシン(I)から選択され、
(ii)X3は、プロリン(P)、セリン(S)から選択され、
(iii)X5は、ヒスチジン(H)、グルタミン(Q)から選択され、
(iv)X2、X4、X6のうちの少なくとも1つは、ホスホセリン(J)、アスパラギン酸(D)、グルタミン酸(E)、アラニン(A)からなる群より選択される。
(i)X7は、アスパラギン酸(D)、アスパラギン(N)、セリン(S)から選択され、
(ii)X8は、バリン(V)、アルギニン(R)、アラニン(A)から選択され、
(iii)X9は、プロリン(P)、グルタミン(Q)から選択され、
(iv)X10は、グリシン(G)、リジン(K)、アスパラギン(N)から選択され、
(v)X11は、アラニン(A)、イソロイシン(I)、バリン(V)から選択され、
(vi)X12は、バリン(V)、スレオニン(T)、プロリン(P)から選択され、
(vii)X13は、グルタミン(Q)、ロイシン(L)から選択され、
(viii)X14は、グリシン(G)、スレオニン(T)、セリン(S)から選択される。
(i)X2はアラニン(A)、グリシン(G)、ロイシン(L)、イソロイシン(I)、バリン(V)、プロリン(P)、メチオニン(M)からなる群より選択され、
(ii)X4はアラニン(A)、グリシン(G)、ロイシン(L)、イソロイシン(I)、バリン(V)、プロリン(P)、メチオニン(M)からなる群より選択され、または
(iii)X6はアラニン(A)、グリシン(G)、ロイシン(L)、イソロイシン(I)、バリン(V)、プロリン(P)、メチオニン(M)からなる群より選択される。そのような実施形態では、X4及びX6は好ましくはアラニン(A)であり、このようなP1は配列番号50〜74からなる群より選択される。
(i)X2はアスパラギン酸(D)、グルタミン酸(E)、ホスホセリン(J)からなる群より選択され、
(ii)X4はアスパラギン酸(D)、グルタミン酸(E)、ホスホセリン(J)からなる群より選択され、または
(iii)X6はアスパラギン酸(D)、グルタミン酸(E)、ホスホセリン(J)からなる群より選択される。
X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む29アミノ酸残基からなる。他の実施形態では、ペプチドまたはペプチドアナログは、8アミノ酸配列X1X2X3VX4X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む30アミノ酸残基からなる。他の実施形態では、ペプチドまたはペプチドアナログは、8アミノ酸配列X1X2X3VX4X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む31アミノ酸残基からなる。他の実施形態では、ペプチドまたはペプチドアナログは、8アミノ酸配列X1X2X3VX4X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む32アミノ酸残基からなる。他の実施形態では、ペプチドまたはペプチドアナログは、8アミノ酸配列X1X2X3VX4X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む33アミノ酸残基からなる。他の実施形態では、ペプチドまたはペプチドアナログは、8アミノ酸配列X1X2X3VX4X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む34アミノ酸残基からなる。他の実施形態では、ペプチドまたはペプチドアナログは、8アミノ酸配列X1X2X3VX4X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む35アミノ酸残基からなる。他の実施形態では、ペプチドまたはペプチドアナログは、8アミノ酸配列X1X2X3VX4X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む36アミノ酸残基からなる。他の実施形態では、ペプチドまたはペプチドアナログは、8アミノ酸配列X1X2X3VX4X5X6E(配列番号9)または13アミノ酸配列KEQEMX1X2X3VX4X5X6E(配列番号10)または16アミノ酸配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)または21アミノ酸配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含む76以下のアミノ酸残基からなる。
本発明で説明するペプチドまたはペプチドアナログはさらに、追加の部分(Y)に結合されていてもよい。該部分は、ペプチドまたはペプチドアナログの末端に結合されていてもよい。
本発明は、各請求項で定められるとおりである。
本発明の一態様では、本明細書で説明するペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物は、神経系疾患、たとえば主に中枢神経系を侵す全身性萎縮症、たとえばハンチントン病、及び中枢神経系の変性疾患、たとえばアルツハイマー病、錐体外路及び運動神経系障害、たとえばパーキンソン病のうちの1種以上の治療及び/または予防用途で有用である。
本発明の化合物または塩は、そのままペプチドまたはペプチドアナログとして投与することも可能であるが、好ましくは、医薬製剤として提供される。したがって、本発明はさらに、本明細書で定義される医薬製剤を提供し、該製剤は、本発明の化合物またはその製薬上許容される塩もしくはエステル、及びその製薬上許容される担体を含む。該医薬製剤は、たとえばRemington: The Science and Practice of Pharmacy 2005, Lippincott, Williams & Wilkinsで説明されているような一般的な製薬法により調製することができる。
本発明は、Vps10ドメインを含む受容体SorCS2、SorCS1、及び/またはSorCS3のC末端ドメインのリン酸化がシナプス可塑性にとって重要である、という知見に関する。SorCS2、SorCS1、及びSorCS3は、Vps10pと高い相同性を有するN末端Vps10pドメイン、及び細胞質C末端ドメインを含む。
一態様では、本発明は、SorCS2、SorCS1、及びSorCS3からなる群より選択されるVps10pドメイン受容体のC末端細胞質ドメインのリン酸化を調節することができる化合物を特定する方法に関し、該方法は、
(i)1つ以上の候補化合物、及びSorCS2、SorCS1、またはSorCS3を発現する細胞を準備するステップ、
(ii)該候補化合物の非存在下で、該細胞における(i)のタンパク質のリン酸化レベルを測定するステップ、
(iii)(ii)の細胞を該候補化合物と接触させるステップ、
(iv)(iii)のタンパク質のリン酸化レベルを測定するステップ、
(v)(ii)と(iv)のリン酸化レベルを比較するステップ、
(vi)SorCS2、SorCS1、またはSorCS3のC末端ドメインのリン酸化を調節することができる(v)の化合物を選択することにより化合物を特定するステップ
を含む。
本方法の第1のステップでは、1つ以上の候補化合物、及びSorCS2、SorCS1、またはSorCS3を発現する細胞を準備する。
Vps10ドメインを含む受容体SorCS2、SorCS1、またはSorCS3のこれらを発現する細胞におけるリン酸化レベルを候補化合物の非存在下で(ステップii)またはその存在下で(ステップiv)測定する。所与のタンパク質のリン酸化レベルを測定する方法は当業界では既知であり、限定ではないが、免疫組織化学、免疫細胞化学、ウェスターンブロット、SILAC、ELISA、及び任意選択によりオートラジオグラフィーと組み合わせた電気泳動が挙げられる。
a)配列番号1のアミノ酸残基1099〜1159、
b)配列番号1のアミノ酸残基1100〜1159、及び
c)配列番号1のアミノ酸残基1101〜1159
からなる群より選択される。
a)配列番号3のアミノ酸残基1099〜1173、
b)配列番号3のアミノ酸残基1100〜1173、及び
c)配列番号3のアミノ酸残基1101〜1173
からなる群より選択される。
a)配列番号5のアミノ酸残基1099〜1174、
b)配列番号5のアミノ酸残基1100〜1174、及び
c)配列番号5のアミノ酸残基1101〜1174
からなる群より選択される。
a)配列番号7のアミノ酸残基1099〜1160、
b)配列番号7のアミノ酸残基1100〜1160、及び
c)配列番号7のアミノ酸残基1101〜1160
からなる群より選択される。
本明細書で先に定義した化合物はいずれも、配列番号1、配列番号3、配列番号5、もしくは配列番号7に示すSorCS2のあらゆる型、SorCS1、またはSorCS3の発現を調節することができる場合がある。
(i)1つ以上の候補化合物、ならびにSorCS2、SorCS1、及び/またはSorCS3を発現する細胞を準備するステップ、
(ii)該候補化合物の非存在下で、該細胞における(i)のタンパク質の発現レベルを測定するステップ、
(iii)(ii)の細胞を該候補化合物と接触させるステップ、
(iv)(iii)のタンパク質の発現レベルを測定するステップ、
(v)(ii)と(iv)の発現レベルを比較するステップ、
(vi)SorCS2、SorCS1、及び/またはSorCS3の発現を調節することができる(v)の化合物を選択することにより、化合物を特定するステップ
を含む。
SorCS2ノックアウトマウスは、10代戻し交配してC57/BL6Jとされている。行動試験は、戻し交配に用いられた同じC57/BL6J亜株と比較してホモ接合である戻し交配マウスで行った(Glerup, Olsen et al., 2014)。ここに記載する各実験では、すべてのマウス株を10代戻し交配してC57/BL6Jbom(Taconic)とし、同腹のコントロールを実験に用いた。すべての実験は、デンマーク国法務省管轄のDanish Animal Experiments Inspectorateの承認を受け(許可番号2011/561−119)、施設及び国のガイドラインにしたがい実施した。動物はすべて、Aarhus大学の実験動物施設で繁殖し収容した。動物の収容は、プラスチック製ケージ(42x25x15cm)1つにつきマウス5匹までのグループとし、病原体フリー条件で、12時間/12時間の明暗サイクルとし、標準餌(Altromin#1324)と水を自由に摂らせた。毎週ケージを掃除し、床敷及び巣材料、木の棒、及び金属のトンネルを入れた。行動実験は、午前9時から午後5時の間の明サイクル中に12〜16週齢の雄マウスで実施した。後述する各行動試験では無処置動物を用い、マウス遺伝子型を盲検として担当者が無作為の順で試験した。後続の分析から除外した動物はいなかった。実験終了時、頚部脱臼により動物を屠殺した。
野生型マウスとSorCS2−/−マウスに、食塩水のみ、または20mg/ml(20mg/kg)のフルオキセチン(商品名Prozac(商標))を含む食塩水を注射した。先行研究では、フルオキセチン注射によるTrkBのリン酸化は1時間後にピークに達することがわかっている(Lieto et al, Plos one 2012)。そこで、マウスをもとのケージ内に1時間放置してから、床敷上に12個のガラス玉を置いた別のケージに移した。そしてマウスをこのケージ内に20分放置し、その後各ケージで50%よりも深く埋められたガラス玉の数を数えた。
BDNFが誘発する抑制性シナプスの形成についての試験では、p0の野生型(wt)の子とSorCS2−/−の子のニューロンを、カバーガラス当たり100,000ニューロンの密度で播種した。インビトロで7日後、培地を1nMのBDNFまたは同量の滅菌D−PBSのいずれかを含む培地に変えた。ニューロンを72時間、37℃、5%CO2でインキュベートしてから、氷冷の4%PFA中で20分固定した。D−PBSで5分ずつ3回洗った後、ニューロンを、抑制性マーカーであるゲフィリンに対し染色し、また樹状突起樹のマーカーであるMap2に対し同時染色した。共焦点顕微法で撮像し、Imarisソフトウェアを用いて分析した。
インビトロのニューロンの樹状突起の分枝を決定するために、P0マウスから海馬ニューロンを単離し、カバーガラス当たり5000ニューロンの密度で培養し増殖させた。24時間後、培地を1nMのBDNFまたは同量の滅菌PBSのいずれかを含む培地に変えた。その後細胞を37℃のインキュベーター内に72時間置いてから、4%PFA中で20分、室温で固定した。細胞の形態を決定するため、ニューロンをβ−チューブリンに対し染色した(β−チューブリンマウスmAb chemicon MAB3408)。共焦点顕微鏡で撮像し、分枝パターンをZen 2011 Image Processing(Carl Zeiss)により次のように分析した。ニューロンの第1次、第2次、第3次、及び第4次の分枝パターンを計数し、ここで第1次の枝は細胞体から突出している神経突起と定義し、細胞体の直径よりも枝が長い場合のみ第1次の枝とした。第2次の枝は、第1次の枝から伸びる突起と定義し、以下同じとした。
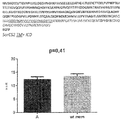
SorCS2のリン酸化についての試験では(図4A)、wtニューロンを6ウェル当たり250万の密度で播種した。培養して5日後、培地をホスファートを含まない培地+0.4mCi/mlの32Pラベル化ホスファートに変え、4時間、37℃、5%CO2で放置した。その後、ホスファートを含まない培地でニューロンを1度洗ってから、1nMのBDNFまたは同量の滅菌PBSのいずれかを含む培地を加えた。これを10分放置してから、培地を回収し、プロテアーゼ阻害剤及びホスファターゼ阻害剤を含む溶解バッファーを加えた。対SorCS2抗体を用いて試料からの免疫沈降を実施した。次いで試料をSDSゲル、続いてオートラジオグラフィーで分析して、SorCS2のリン酸化を評価した。
3匹の野生型マウスに、普通の水道水または0.8mg/mlのフルオキセチンを含む水道水を、1週間に1度water−chanceで3週間与えた。その後、マウスを頚部脱臼により安楽死させ、海馬及び皮質を切除した。試料をプロテアーゼ阻害剤及びホスファターゼ阻害剤を含む溶解バッファー中で溶解した。次いで試料をウェスターンブロット分析して、フルオキセチン処置によりSorCS2レベルが上昇するかどうかを調べた。
BDNF媒介性シグナリングにおけるSorCS2の細胞外部分の役割を調べるため、SorCS2−/−ニューロンをカバーガラス当たり100,000ニューロンの密度で播種した。ニューロンを、膜内への挿入を容易にするようにSorCS2の細胞外部分がGFPで置換されている構築物(wt memと表記する)でトランスフェクトした。ここで、全長SorCS2でトランスフェクトしたニューロンとwt memでトランスフェクトしたニューロンの差異は観察されなかった(図8)。しかし、wt memでトランスフェクトしBDNFで刺激したニューロンは、全長SorCS2でトランスフェクトしたニューロンよりも形態が大幅に増加し(図8)、細胞外ドメインは十分なBDNF応答にとって必要ではないことが示された。
SorCS2尾部の可溶性バリアントが形態的応答の誘発に十分であるかどうかを調べるため、SorCS2−/−ニューロンをカバーガラス当たり100,000ニューロンの密度で播種した。これらのニューロンをSorCS2の細胞内部分を含む構築物でトランスフェクトした(図10.1)(「wt可溶性尾部」と表記する)。wt可溶性尾部でトランスフェクトしたニューロンは、全長SorCS2でトランスフェクトしたニューロンと比べて形態の増加を示した。この応答はさらに、BDNFを付加すると増加した。次に発明者らは、リン酸化セリンを模倣するため1125、1128、及び1130の位置のセリンをアスパラギン酸に変えたSorCS2の細胞内部分を含む構築物を作成した(図10.2)(「活性化された可溶性尾部」と表記する)。活性可溶性尾部でトランスフェクトしたニューロンを全長SorCS2でトランスフェクトしたニューロンと比較すると、有意な応答が観察された。この応答は、BDNFを活性可溶性尾部に加えてもさらに増加はしなかった。このことは、活性可溶性尾部がニューロンの形態的応答の誘発に十分であることを示す。要約すると、SorCS2は、ニューロンにおけるBDNFが媒介する応答にとって重要であるだけでなく、十分でもある。
発明者らは次に、SorCS2の細胞内尾部と同様のペプチドを作成した。第1のペプチドは、アスパラギン酸に変えた1125、1128、及び1130の位置のセリンを除いては、野生型SorCS2尾部と同じ配列であった。さらに、ニューロンへの侵入を容易にするためTAT配列が付加されていた(図11)(「Ppep」と表記する)。このペプチドが神経栄養性応答を誘発できるかどうかを調べるため、内在性レベルのSorCS2を発現する野生型ニューロンをカバーガラス当たり20,000ニューロンの密度で、0.1μM、1μM、または10μMを1nMのBDNFまたは同量の滅菌食塩水と組み合わせて含む培地中に播種した。0.1μMも10μMも応答を誘発しなかったが、1μMのPpepは、コントロール(無刺激)と比べて有意な応答を誘発した(図11.4)。また、BDNF処理してもさらなる増加は観察されず(図11.2)、Ppepはニューロンにおける神経栄養性応答の誘発に十分であることが示された。
pPepの付加は、BDNFの非存在下でも神経栄養性応答の誘発に十分であったため、発明者らは、不活性ペプチドの付加がBDNFの存在下でもBDNFシグナリングの阻害に十分であるかどうか、検証したいと考えた。そこで発明者らは、不活性SorCS2尾部を模倣するためSorCS2尾部の1125、1128、及び1130の位置のセリンをアラニンに変えたペプチドを作成した(図12ではApepと表記する)。さらに、このペプチドは、細胞への侵入を容易にするためTAT配列(HIVタンパク質形質導入ドメイン)を備えている。0.1μMのApepでは、BDNFはまだ、有意な応答を誘発できた(図12.1)。しかし、1μMのApep(図12.2)または10μMのApep(図12.3)の添加ではそうではなかった。コントロールと比較すると、コントロール(BDNFを加えていない)と1μM Apep+BDNF(図12.4)では差異が観察されなかった。このことは、Apepは、1μMで、BDNFの存在下でも、BDNF誘発性形態的応答を阻害することができることを示している。
より小さいペプチドを作成し、これらが神経栄養性応答の誘発に十分であるかどうかを調べた。1nMのBDNFの付加では(図13.2)、wtニューロンは、BDNFなし(図13.1)のWTニューロンと比べて複雑性が増加した。実施例9及び10からわかるように、25、28、及び30の位置のセリンがアラニンに変異しているペプチドは、BDNFの存在下でも、形態的応答を阻害し(図13.3)、25、28、及び30の位置のセリンがアスパラギン酸に変えられているペプチドは、BDNFの非存在下でも、神経栄養性応答を誘発できた(図13.4)。そこで発明者らは、セリンをアラニンに変え、より小さいペプチドを作成した。このペプチドは、BDNFが誘発する神経栄養性応答を阻害することができた(図13.5)。さらに、セリンがアスパラギン酸に変えられている短ペプチドは、BDNFの非存在下でも、神経栄養性応答を誘発できた(図13.6)。
一般的な高度に保存されている配列のほかに、図14に示すように、SorCS1では1125、1128、及び1130の位置のセリンが保存されている。SorCS3では、1128及び1130の位置のセリンが保存されている(これらのセリンは、BDNF応答の媒介に重要な2つのセリンであることがわかっている。図5参照)。したがって、SorCS1及びSorCS3の尾部も、SorCS2と同様に神経栄養シグナリングを媒介し得る。
てんかん重積状態を原因とするTrkBの過剰な活性化は、側頭葉(temporal lope)てんかん(TLE)の発症を促進する(3)。TrkBの全阻害は、ニューロン生存率の低下をもたらすので望ましくない(3)。リード化合物が、ニューロン生存率を低下させることなく、てんかん重積状態を原因とするTrkBの過剰なシグナリングの悪影響を阻害することができるかどうかを試験するため、発明者らは、(3)で説明されているのと同様に、カイニン酸を注射する前に、リード化合物を注射する。ここで発明者らは、リード化合物が、てんかん発作の発症を阻害することができるかどうか、及び/またはTrkBの過剰な活性化を原因とする悪影響を阻害し、そうすることで側頭葉(temporal lope)てんかんの発症を阻害することができるかどうかを試験する。
ハンチントン病は、BDNFシグナリングの損失によりニューロンへの栄養補助が不足することで起きると考えられている、常染色体優性神経変性障害である(4)。リード化合物が、ハンチントン病において下流シグナリングカスケードを活性化できるかどうかを調べるため、発明者らは、全長ヒトミュータントハンチンチンが挿入されている、BACHDと称されるハンチントン病のトランスジェニックマウスモデルを用いる。これらのマウスでは、BDNFは、シナプス強化またはLTPを誘発することができない(4)。発明者らは、リード化合物の付加が、BACHDマウスの脳においてLTPを誘発することができるかどうか、電気生理学的に調査する。
BDNFは、中枢神経系のニューロンの栄養補助に重要であると考えられており、アルツハイマー病患者で観察されるBDNFレベルの低下は、BDNFが該疾患において果たし得る役割を示している(5)。ここでBDNFは、アルツハイマー病の病理特徴と考えられているアミロイド斑の形成を阻害する役割を果たしている可能性がある。リード化合物が該疾患の発症を減衰させることができるかどうかを調べるため、海馬ニューロンをカバーガラス当たり100,00ニューロンの密度で播種する。リード化合物は、(6)で説明されているように、アミロイド斑Aβ1−42の毒性成分を付加する4時間前に付加する。Aβ1−42の付加は、ニューロン死をもたらすと記載されているので、発明者らは、Aβ1−42を発明者らのリード化合物またはビヒクルと一緒に付加した場合のニューロン生存率を調査する。
論文(7)では、著者らは、WAGR症候群の子どもの一群を調査している。この疾患は、染色体11p13の一部が欠失している疾患である。一サブグループの子どもは、bdnf遺伝子も欠失しているためBDNFハプロ不全を有する。bdnfが欠失している子どもの100%が、10歳までに肥満症になっていた。ここではBDNFは、視床下部内で作用してエネルギー吸収を制御すると考えられている。BDNFハプロ不全のマウスも肥満であり(8)、脳室内BDNF注射により救済することが可能である(9)。エネルギー恒常性にとって重要な受容体であるレプチン受容体に変異のあるマウス(db/dbマウス)と、BDNFハプロ不全マウスは、どちらも野生型の動物よりも肥満である。db/dbマウスでは、BDNFの中枢または末梢投与により食物摂取量が減少し、エネルギー消費量が増加し、糖尿病が予防された(10、11)。リード化合物が糖尿病の発症を減衰することができるかどうかを調べるため、発明者らはdb/dbマウス及びBDNFハプロ不全マウスを調査する。ここで、リード化合物またはビヒクルを4週齢のdb/dbマウス、BDNFハプロ不全マウス、または野生型マウスに、12週間注射する。この間、血液試料を週2回採取して、グルコースレベル及びインスリンレベルを測定する。さらに、動物の体重、エネルギー消費量、食物摂取量を、実験の間中測定する。
野生型の出生後0日目の子を断頭により安楽死させ、脳を回収し、海馬を切除して氷冷のLeibovitz’s L15培地に入れる。切除後、組織を、予め活性化させた20U/mlのパパイン中30分かけて分離させる。その後、組織を、0.01mg/mlのDNase及び10%ウシ胎仔血清(FBS)を含むDMEMで洗ってから、0.01mg/mlのDNase及び10%FBSを含むDMEM中ですり潰す。
Wtマウスに、注射で、または水に混ぜて、薬物を3週間与える。その後マウスを頚部脱臼により安楽死させ、目標部位をマウスから切除する。これらの部位を、次いでウェスターンブロットまたはELISAで分析して、SorCS2などいくつかの遺伝子の制御について調査する。
初代海馬ニューロンにおいてSorCS2のリン酸化を誘発する、またはSorCS2の発現を増加させる能力について、CNS化合物ライブラリーのスクリーニングを実施する。候補化合物はさらに、WT及びKOの海馬ニューロンを用いて、神経突起のSorCS2依存性の成長を誘発する能力について試験する。リード化合物を1種類特定し、これが動物モデル、たとえばBDNFヘテロ接合マウスの精神神経性行動を調節する能力を試験する。
配列番号1:配列尾部Aを有するSorCS2タンパク質;ホモサピエンス

X1X2X3VX4X5X6E
配列番号10:切断型Vps10pドメイン受容体尾部
KEQEMX1X2X3VX4X5X6E
配列番号11:切断型Vps10pドメイン受容体尾部
X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14
配列番号12:切断型Vps10pドメイン受容体尾部
KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14
配列番号13:切断型SorCS2受容体尾部
TX2PVX4HX6E
配列番号14:切断型SorCS1受容体尾部
IX2PVX4HX6E
配列番号15:切断型SorCS3受容体尾部
IX2SVX4QX6E
配列番号16:切断型SorCS2受容体尾部
KEQEMTX2PVX4HX6E
配列番号17:切断型SorCS1受容体尾部
KEQEMIX2PVX4HX6E
配列番号18:切断型SorCS3受容体尾部
KEQEMIX2SVX4QX6E
配列番号19:切断型SorCS2受容体尾部
TX2PVX4HX6EDVQGAVQG
配列番号20:切断型SorCS1受容体尾部
IX2PVX4HX6ESRPNVPQT
配列番号21:切断型SorCS3受容体尾部
IX2SVX4QX6ENAPKITLS
配列番号22:切断型SorCS2受容体尾部
KEQEMTX2PVX4HX6EDVQGAVQG
配列番号23:切断型SorCS1受容体尾部
KEQEMIX2PVX4HX6ESRPNVPQT
配列番号24:切断型SorCS3受容体尾部
KEQEMIX2SVX4QX6ENAPKITLS
配列番号25:切断型SorCS2受容体尾部(活性化)
TX2PVX4HX6E
配列番号26:切断型SorCS1受容体尾部(活性化)
IX2PVX4HX6E
配列番号27:切断型SorCS3受容体尾部(活性化)
IX2SVX4QX6E
配列番号28:切断型SorCS2受容体尾部(活性化)
KEQEMTX2PVX4HX6E
配列番号29:切断型SorCS1受容体尾部(活性化)
KEQEMIX2PVX4HX6E
配列番号30:切断型SorCS3受容体尾部(活性化)
KEQEMIX2SVX4QX6E
配列番号31:切断型SorCS2受容体尾部(活性化)
TX2PVX4HX6EDVQGAVQG
配列番号32:切断型SorCS1受容体尾部(活性化)
IX2PVX4HX6ESRPNVPQT
配列番号33:切断型SorCS3受容体尾部(活性化)
IX2SVX4QX6ENAPKITLS
配列番号34:切断型SorCS2受容体尾部(活性化)
KEQEMTX2PVX4HX6EDVQGAVQG
配列番号35:切断型SorCS1受容体尾部(活性化)
KEQEMIX2PVX4HX6ESRPNVPQT
配列番号36:切断型SorCS3受容体尾部(活性化)
KEQEMIX2SVX4QX6ENAPKITLS
配列番号37:切断型SorCS2受容体尾部(活性化)
TDPVDHDE
配列番号38:切断型SorCS2受容体尾部(活性化)
KEQEMTDPVDHDE
配列番号39:切断型SorCS2受容体尾部(活性化)
TDPVDHDEDVQGAVQG
配列番号40:切断型SorCS2受容体尾部(活性化)
KEQEMTDPVDHDEDVQGAVQG
配列番号41:切断型SorCS2受容体尾部(活性化)
YARAAARNARAEKEQEMTDPVDHDEDVQGAVQ
配列番号42:切断型SorCS1受容体尾部(活性化)
IDPVDHDE
配列番号43:切断型SorCS1受容体尾部(活性化)
KEQEMIDPVDHDE
配列番号44:切断型SorCS1受容体尾部(活性化)
IDPVDHDESRPNVPQT
配列番号45:切断型SorCS1受容体尾部(活性化)
KEQEMIDPVDHDESRPNVPQT
配列番号46:切断型SorCS3受容体尾部(活性化)
IDSVDQDE
配列番号47:切断型SorCS3受容体尾部(活性化)
KEQEMIDSVDQDE
配列番号48:切断型SorCS3受容体尾部(活性化)
IDSVDQDENAPKITLS
配列番号49:切断型SorCS3受容体尾部(活性化)
KEQEMIDSVDQDENAPKITLS
配列番号50:切断型SorCS2受容体尾部(不活性化)
TX2PVX4HX6E
配列番号51:切断型SorCS1受容体尾部(不活性化)
IX2PVX4HX6E
配列番号52:切断型SorCS3受容体尾部(不活性化)
IX2SVX4QX6E
配列番号53:切断型SorCS2受容体尾部(不活性化)
KEQEMTX2PVX4HX6E
配列番号54:切断型SorCS1受容体尾部(不活性化)
KEQEMIX2PVX4HX6E
配列番号55:切断型SorCS3受容体尾部(不活性化)
KEQEMIX2SVX4QX6E
配列番号56:切断型SorCS2受容体尾部(不活性化)
TX2PVX4HX6EDVQGAVQG
配列番号57:切断型SorCS1受容体尾部(不活性化)
IX2PVX4HX6ESRPNVPQT
配列番号58:切断型SorCS3受容体尾部(不活性化)
IX2SVX4QX6ENAPKITLS
配列番号59:切断型SorCS2受容体尾部(不活性化)
KEQEMTX2PVX4HX6EDVQGAVQG
配列番号60:切断型SorCS1受容体尾部(不活性化)
KEQEMIX2PVX4HX6ESRPNVPQT
配列番号61:切断型SorCS3受容体尾部(不活性化)
KEQEMIX2SVX4QX6ENAPKITLS
配列番号62:切断型SorCS2受容体尾部(不活性化)
TAPVAHAE
配列番号63:切断型SorCS2受容体尾部(不活性化)
KEQEMTAPVAHAE
配列番号64:切断型SorCS2受容体尾部(不活性化)
TAPVAHAEDVQGAVQG
配列番号65:切断型SorCS2受容体尾部(不活性化)
KEQEMTAPVAHAEDVQGAVQG
配列番号66:切断型SorCS2受容体尾部(不活性化)
YARAAARNARAEKEQEMTAPVAHAEDVQGAVQ
配列番号67:切断型SorCS1受容体尾部(不活性化)
IAPVAHAE
配列番号68:切断型SorCS1受容体尾部(不活性化)
KEQEMIAPVAHAE
配列番号69:切断型SorCS1受容体尾部(不活性化)
IAPVAHAESRPNVPQT
配列番号70:切断型SorCS1受容体尾部(不活性化)
KEQEMIAPVAHAESRPNVPQT
配列番号71:切断型SorCS3受容体尾部(不活性化)
IASVAQAE
配列番号72:切断型SorCS3受容体尾部(不活性化)
KEQEMIASVAQAE
配列番号73:切断型SorCS3受容体尾部(不活性化)
IASVAQAENAPKITLS
配列番号74:切断型SorCS3受容体尾部(不活性化)
KEQEMIASVAQAENAPKITLS
配列番号75:切断型SorCS3受容体尾部(不活性化)
KEQEMIASVAQAENAPKITLS
Claims (81)
- ペプチドまたはペプチドアナログ(P1)であって、配列X1X2X3VX4X5X6E(配列番号9)を含むかまたはそれからなり、
(i)X1はスレオニン(T)またはイソロイシン(I)であり、
(ii)X2はホスホセリン(J)、アスパラギン酸(D)、グルタミン酸(E)、アラニン(A)、及びセリン(S)からなる群より選択され、
(iii)X3はプロリン(P)またはセリン(S)であり、
(iv)X4はホスホセリン(J)、アスパラギン酸(D)、グルタミン酸(E)、アラニン(A)、及びセリン(S)からなる群より選択され、
(v)X5はヒスチジン(H)またはグルタミン(Q)であり、
(vi)X6はホスホセリン(J)、アスパラギン酸(D)、グルタミン酸(E)、アラニン(A)、及びセリン(S)からなる群より選択される、
前記ペプチドまたはペプチドアナログ。 - P1は、配列KEQEMX1X2X3VX4X5X6E(配列番号10)を含むかまたはそれからなる、請求項1に記載のペプチドまたはペプチドアナログ。
- P1は、配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)を含むかまたはそれからなり、
(i)X7はアスパラギン酸(D)、アスパラギン(N)、セリン(S)から選択され、
(ii)X8はバリン(V)、アルギニン(R)、アラニン(A)から選択され、
(iii)X9はプロリン(P)またはグルタミン(Q)であり、
(iv)X10はグリシン(G)、リジン(K)、アスパラギン(N)から選択され、
(v)X11はアラニン(A)、イソロイシン(I)、バリン(V)から選択され、
(vi)X12はバリン(V)、スレオニン(T)、プロリン(P)から選択され、
(vii)X13はグルタミン(Q)またはロイシン(L)であり、
(viii)X14はグリシン(G)、スレオニン(T)、セリン(S)から選択される、
請求項1に記載のペプチドまたはペプチドアナログ。 - P1は、配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)を含むかまたはそれからなる、請求項1に記載のペプチドまたはペプチドアナログ。
- P1は、配列X1X2X3VX4X5X6E(配列番号9)からなり、X1はスレオニン(T)であり、X3はプロリン(P)であり、X5はヒスチジン(H)であり、このようなP1は配列番号13である、請求項1に記載のペプチドまたはペプチドアナログ。
- P1は、配列KEQEMX1X2X3VX4X5X6E(配列番号10)からなり、X1はスレオニン(T)であり、X3はプロリン(P)であり、X5はヒスチジン(H)であり、このようなP1は配列番号16である、請求項2に記載のペプチドまたはペプチドアナログ。
- P1は、配列X1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号11)からなり、X1はスレオニン(T)であり、X3はプロリン(P)であり、X5はヒスチジン(H)であり、X7はアスパラギン酸(D)であり、X8はバリン(V)であり、X9はグルタミン(Q)であり、X10はグリシン(G)であり、X11はアラニン(A)であり、X12はバリン(V)であり、X13はグルタミン(Q)であり、X14はグリシン(G)であり、このようなP1は配列番号19である、請求項3に記載のペプチドまたはペプチドアナログ。
- P1は、配列KEQEMX1X2X3VX4X5X6EX7X8X9X10X11X12X13X14(配列番号12)からなり、X1はスレオニン(T)であり、X3はプロリン(P)であり、X5はヒスチジン(H)であり、X7はアスパラギン酸(D)であり、X8はバリン(V)であり、X9はグルタミン(Q)であり、X10はグリシン(G)であり、X11はアラニン(A)であり、X12はバリン(V)であり、X13はグルタミン(Q)であり、X14はグリシン(G)であり、このようなP1は配列番号22である、請求項4に記載のペプチドまたはペプチドアナログ。
- P1は、X2、X4、X6のうちの少なくとも1つを含み、
(i)X2はアスパラギン酸(D)、グルタミン酸(E)、ホスホセリン(J)からなる群より選択され、
(ii)X4はアスパラギン酸(D)、グルタミン酸(E)、ホスホセリン(J)からなる群より選択され、または
(iii)X6はアスパラギン酸(D)、グルタミン酸(E)、ホスホセリン(J)からなる群より選択される、
請求項1〜8のいずれか1項に記載のペプチドまたはペプチドアナログ。 - P1は、配列番号25〜49のいずれかから選択される配列を含むかまたはそれからなる、請求項9に記載のペプチドまたはペプチドアナログ。
- P1は、配列番号37を含むかまたはそれからなる、請求項9に記載のペプチドまたはペプチドアナログ。
- P1は、X2、X4、X6のうちの少なくとも1つを含み、X2、X4、及び/またはX6はアラニン(A)である、請求項1〜8のいずれか1項に記載のペプチドまたはペプチドアナログ。
- P1は、配列番号50〜74のいずれかから選択される配列を含むかまたはそれからなる、請求項0に記載のペプチドまたはペプチドアナログ。
- P1は、配列番号62を含むかまたはそれからなる、請求項0に記載のペプチドまたはペプチドアナログ。
- P1は、キナーゼ活性を増加させることができる、請求項1〜11のいずれか1項に記載のペプチドまたはペプチドアナログ。
- P1は、ホスファターゼ活性を阻害することができる、請求項1〜11のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 外来性BDNFの貢献がなくても、キナーゼ活性が増加し、またはホスファターゼ活性が阻害される、請求項15〜16のいずれか1項に記載のペプチドまたはペプチドアナログ。
- P1は、キナーゼ活性を阻害することができる、請求項1〜8及び請求項0〜14のいずれか1項に記載のペプチドまたはペプチドアナログ。
- P1は、ホスファターゼ活性を増加させることができる、請求項1〜8及び請求項0〜14のいずれか1項に記載のペプチドまたはペプチドアナログ。
- P1は、8〜76アミノ酸残基を含む、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- P1は、少なくとも9アミノ酸残基、たとえば少なくとも10アミノ酸残基、たとえば少なくとも11アミノ酸残基、たとえば少なくとも12アミノ酸残基、たとえば少なくとも13アミノ酸残基、たとえば少なくとも14アミノ酸残基、たとえば少なくとも15アミノ酸残基、たとえば少なくとも16アミノ酸残基、たとえば少なくとも17アミノ酸残基、たとえば少なくとも18アミノ酸残基、たとえば少なくとも19アミノ酸残基、たとえば少なくとも20アミノ酸残基、たとえば少なくとも21アミノ酸残基、たとえば少なくとも22アミノ酸残基、たとえば少なくとも23アミノ酸残基、たとえば少なくとも24アミノ酸残基、たとえば少なくとも25アミノ酸残基、たとえば少なくとも26アミノ酸残基、たとえば少なくとも27アミノ酸残基、たとえば少なくとも28アミノ酸残基、たとえば少なくとも29アミノ酸残基、たとえば少なくとも30アミノ酸残基、たとえば少なくとも31アミノ酸残基、たとえば少なくとも32アミノ酸残基、たとえば少なくとも33アミノ酸残基、たとえば少なくとも34アミノ酸残基、たとえば少なくとも35アミノ酸残基、たとえば少なくとも36アミノ酸残基、たとえば少なくとも40アミノ酸残基、たとえば少なくとも45アミノ酸残基、たとえば少なくとも50アミノ酸残基、たとえば少なくとも55アミノ酸残基、たとえば少なくとも60アミノ酸残基、たとえば少なくとも65アミノ酸残基、たとえば少なくとも70アミノ酸残基、たとえば少なくとも75アミノ酸残基、たとえば76アミノ酸残基を含む、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- P1は、少なくとも1つの部分、たとえば2つの結合部分(Y1)及び(Y2)に結合されている、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 前記結合部分Y1及びY2は、細胞透過性ペプチド(CPP)、アルブミン結合部分(ABM)、及び検出可能部分(Z)からなる群より選択される、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 前記結合部分はCPPである、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 前記CPPはアミド結合によりP1に連結され、前記CPPはアルギニン及び/またはリジンから選択される少なくとも4アミノ酸残基を含む、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 前記CPPは、レトロインベルソペプチドを含む、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 前記CPPは、アミノ酸配列GRKKRRQRRRを有するTatペプチド、またはアミノ酸配列rrrqrrkkrgを有するレトロインベルソ−d−Tatペプチドである、請求項1〜26のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 前記検出可能部分(Z)は、フルオロフォアである、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 前記(Z)はTMRである、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ
- アミノ酸残基のいずれか1つは、別のアミノ酸に変えられているが、ただしそのように変えられているアミノ酸は4つ以下である、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- アミノ酸残基のいずれか1つは、別のアミノ酸に変えられているが、ただしそのように変えられているアミノ酸は3つ以下である、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- アミノ酸残基のいずれか1つは、別のアミノ酸に変えられているが、ただしそのように変えられているアミノ酸は2つ以下である、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 前記ペプチドは、配列番号1〜8のいずれか1つのアミノ酸1100〜1173の配列バリアントである、先行請求項のいずれか1項に記載のペプチドまたはペプチドアナログ。
- 先行請求項のいずれか1項で定めたペプチドをコードする、ポリヌクレオチド。
- 請求項34で定めたポリヌクレオチドを含む、ベクター。
- 請求項34に記載のポリヌクレオチドまたは請求項35に記載のベクターを含む、宿主細胞。
- 前記宿主細胞はバクテリア細胞である、請求項36に記載の宿主細胞。
- 前記宿主細胞は哺乳類細胞である、請求項36に記載の宿主細胞。
- 前記宿主細胞はヒト細胞である、請求項36に記載の宿主細胞。
- 請求項1〜33のいずれか1項で定めたペプチドもしくはペプチドアナログ、請求項34に記載のポリヌクレオチド、請求項35に記載のベクター、または請求項36〜39のいずれか1項に記載の宿主細胞を含む、組成物。
- 前記組成物は医薬組成物である、請求項40に記載の使用される組成物。
- 医薬として使用される、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、請求項34に記載のポリヌクレオチド、請求項35に記載のベクター、請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜41のいずれか1項に記載の組成物。
- 神経系疾患の治療及び/または予防に使用される、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、請求項34に記載のポリヌクレオチド、請求項35に記載のベクター、請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜41のいずれか1項に記載の組成物。
- 前記神経系疾患は突発性及び発作性疾患である、請求項43に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記突発性及び発作性疾患は、てんかん、てんかん重積状態、及び片頭痛からなるリストから選択される、請求項44に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記神経系疾患は、主に中枢神経系を侵す全身性萎縮症である、請求項43に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記主に中枢神経系を侵す全身性萎縮症はハンチントン病である、請求項46に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記神経系疾患は中枢神経系の変性疾患である、請求項43に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記中枢神経系の変性疾患はアルツハイマー病である、請求項48に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記神経系疾患は神経系の錐体外路及び運動障害である、請求項43に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記神経系の錐体外路及び運動障害はパーキンソン病である、請求項50に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記神経系疾患は神経障害性疼痛である、請求項43に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記神経障害性疼痛疾患は、痛覚過敏症の一因となる、請求項52に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 精神及び行動障害の治療及び/または予防に使用される、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、請求項34に記載のポリヌクレオチド、請求項35に記載のベクター、請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜41のいずれか1項に記載の組成物。
- 前記精神または行動障害は、鬱病、不安症、強迫性障害(OCD)、双極性障害(BD)、統合失調症(SZ)、及び広汎性発達障害からなる群より選択される、請求項54に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記広汎性発達障害はRett症候群である、請求項55に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 先天性神経発達異常及び染色体異常の治療及び/または予防に使用される、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、請求項34に記載のポリヌクレオチド、請求項35に記載のベクター、請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜41のいずれか1項に記載の組成物。
- 前記先天性神経発達異常及び染色体異常はルビンシュタイン・テイビ症候群である、請求項57に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 脳血管疾患の脳血管症候群を含む心血管疾患の治療及び/または予防に使用される、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、請求項34に記載のポリヌクレオチド、請求項35に記載のベクター、請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜41のいずれか1項に記載の組成物。
- 前記脳血管疾患の脳血管症候群は卒中である、請求項59に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 代謝及び摂食障害の治療及び/または予防に使用される、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、請求項34に記載のポリヌクレオチド、請求項35に記載のベクター、請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜41のいずれか1項に記載の組成物。
- 前記代謝及び摂食障害は、糖尿病、肥満、インスリン抵抗症、及び神経性食欲不振症からなる群より選択される、請求項61に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 腫瘍性疾患の治療及び/または予防に使用される、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、請求項34に記載のポリヌクレオチド、請求項35に記載のベクター、請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜41のいずれか1項に記載の組成物。
- 前記腫瘍性疾患は固形腫瘍である、請求項63に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記固形腫瘍は尿生殖器異常または性腺芽腫である、請求項64に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記固形腫瘍は、尿路の悪性新生物である、請求項64に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 前記尿路の悪性新生物は腎臓の新生物である、請求項66に記載の使用されるペプチドもしくはペプチドアナログ、ポリヌクレオチド、ベクター、細胞、または組成物。
- 先行請求項のいずれか1項に記載の使用されるペプチドをコードする、ポリヌクレオチド。
- 請求項68で定めるポリヌクレオチドを含む、ベクター。
- 請求項68に記載のポリヌクレオチドまたは請求項69に記載のベクターを含む、宿主細胞。
- 前記宿主細胞はバクテリア細胞である、請求項70に記載の宿主細胞。
- 前記宿主細胞は哺乳類細胞である、請求項70に記載の宿主細胞。
- 前記宿主細胞はヒト細胞である、請求項70に記載の宿主細胞。
- 請求項1〜33のいずれか1項に記載のポリペプチドもしくはペプチドアナログ、または請求項34に記載のポリヌクレオチド、または請求項35に記載のベクター、または請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜42のいずれか1項に記載の組成物の、医薬製造への、請求項43〜67のいずれか1項に記載の疾患の治療への、及び/またはその予防への使用。
- 請求項43〜67のいずれか1つ以上に記載の疾患の治療または予防の方法であって、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、または請求項34に記載のポリヌクレオチド、または請求項35に記載のベクター、または請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜42のいずれか1項に記載の組成物を、それを必要とする対象に投与することを含む、前記方法。
- SorCS2、SorCS1、及びSorCS3からなる群より選択されるVps10pドメイン受容体のC末端細胞質ドメインのリン酸化を調節することができる化合物を特定する方法であって、
(i)1つ以上の候補化合物、及びSorCS2、SorCS1、またはSorCS3を発現する細胞を準備するステップ、
(ii)前記候補化合物の非存在下で、前記細胞における(i)のタンパク質のリン酸化レベルを測定するステップ、
(iii)(ii)の細胞を前記候補化合物と接触させるステップ、
(iv)(iii)のタンパク質のリン酸化レベルを測定するステップ、
(v)(ii)と(iv)のリン酸化レベルを比較するステップ、
(vi)SorCS2、SorCS1、またはSorCS3のC末端ドメインのリン酸化を調節することができる(v)の化合物を選択することにより化合物を特定するステップ
を含む、前記方法。 - SorCS2、SorCS1、及び/またはSorCS3の発現を調節することができる化合物を特定する方法であって、
(i)1つ以上の候補化合物、ならびにSorCS2、SorCS1、及び/またはSorCS3を発現する細胞を準備するステップ、
(ii)前記候補化合物の非存在下で、前記細胞における(i)のタンパク質の発現レベルを測定するステップ、
(iii)(ii)の細胞を前記候補化合物と接触させるステップ、
(iv)(iii)のタンパク質の発現レベルを測定するステップ、
(v)(ii)と(iv)の発現レベルを比較するステップ、
(vi)SorCS2、SorCS1、及び/またはSorCS3の発現を調節することができる(v)の化合物を選択することにより、化合物を特定するステップ
を含む、前記方法。 - シナプスの数を増やす方法であって、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、または請求項34に記載のポリヌクレオチド、または請求項35に記載のベクター、または請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜42のいずれか1項に記載の組成物を、それを必要とする対象に投与することを含む、前記方法。
- ニューロン形態の変化を促進する方法であって、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、または請求項34に記載のポリヌクレオチド、または請求項35に記載のベクター、または請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜42のいずれか1項に記載の組成物を、それを必要とする対象に投与することを含む、前記方法。
- ニューロン形態の変化は、樹状突起の及び/または軸索の分枝の増加である、請求項79に記載の方法。
- 初代海馬ニューロンにおけるSorCS2、SorCS1、及び/またはSorCS3のC末端ドメインのリン酸化を誘発する方法であって、請求項1〜33のいずれか1項に記載のペプチドもしくはペプチドアナログ、または請求項34に記載のポリヌクレオチド、または請求項35に記載のベクター、または請求項36〜39のいずれか1項に記載の宿主細胞、または請求項40〜42のいずれか1項に記載の組成物を、それを必要とする対象に投与することを含む、前記方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| DKPA201570835 | 2015-12-18 | ||
| DKPA201570835 | 2015-12-18 | ||
| PCT/DK2016/050445 WO2017101956A1 (en) | 2015-12-18 | 2016-12-19 | SorCS PEPTIDES AND USES THEREOF |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2019508018A true JP2019508018A (ja) | 2019-03-28 |
| JP2019508018A5 JP2019508018A5 (ja) | 2020-01-30 |
| JP6915810B2 JP6915810B2 (ja) | 2021-08-04 |
Family
ID=59055867
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2018531443A Expired - Fee Related JP6915810B2 (ja) | 2015-12-18 | 2016-12-19 | SorCSペプチド及びそれらの用途 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US20190345216A1 (ja) |
| EP (1) | EP3390435A1 (ja) |
| JP (1) | JP6915810B2 (ja) |
| WO (1) | WO2017101956A1 (ja) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP4192844A1 (en) | 2020-08-06 | 2023-06-14 | Aarhus Universitet | Cyclic peptides and uses thereof |
| JP2025506481A (ja) | 2022-02-09 | 2025-03-11 | テイトゥル トロフィックス エーピーエス | 新規ペプチド |
| CN121925267A (zh) | 2023-08-16 | 2026-04-24 | 泰图尔特罗菲克斯有限公司 | 用于治疗内耳或乳突疾病的肽类药物 |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2003287930A1 (en) * | 2002-12-20 | 2004-07-14 | Neuronicon Aps | Modulation of activity of neurotrophins |
| US8460657B2 (en) * | 2008-06-25 | 2013-06-11 | H. Lundbeck A/S | Modulation of the TrpV: Vps10p receptor system for the treatment of pain |
| EP3165537A1 (en) * | 2008-12-19 | 2017-05-10 | H. Lundbeck A/S | Modulation of the vps 10-domain receptor family for the treatment of mental and behavioural disorders |
| US20130336988A1 (en) * | 2010-11-17 | 2013-12-19 | New York University | Methods for treating early stage or mild neurological disorders |
-
2016
- 2016-12-19 EP EP16828710.0A patent/EP3390435A1/en not_active Withdrawn
- 2016-12-19 WO PCT/DK2016/050445 patent/WO2017101956A1/en not_active Ceased
- 2016-12-19 JP JP2018531443A patent/JP6915810B2/ja not_active Expired - Fee Related
- 2016-12-19 US US16/061,085 patent/US20190345216A1/en not_active Abandoned
Non-Patent Citations (1)
| Title |
|---|
| JBC, 2003, VOL.278, PP.7390-7396, JPN6020050973, ISSN: 0004518635 * |
Also Published As
| Publication number | Publication date |
|---|---|
| US20190345216A1 (en) | 2019-11-14 |
| EP3390435A1 (en) | 2018-10-24 |
| JP6915810B2 (ja) | 2021-08-04 |
| WO2017101956A1 (en) | 2017-06-22 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Duan et al. | Semaphorin 5A inhibits synaptogenesis in early postnatal-and adult-born hippocampal dentate granule cells | |
| Lechauve et al. | Neuroglobin involvement in respiratory chain function and retinal ganglion cell integrity | |
| Hudry et al. | Inhibition of the NFAT pathway alleviates amyloid beta neurotoxicity in a mouse model of Alzheimer's disease | |
| Shioda et al. | Expression of a truncated form of the endoplasmic reticulum chaperone protein, σ1 receptor, promotes mitochondrial energy depletion and apoptosis | |
| Neymeyer et al. | Activation of annexin A1 signalling in renal fibroblasts exerts antifibrotic effects | |
| Wyszynski et al. | Biochemical and immunocytochemical characterization of GRIP, a putative AMPA receptor anchoring protein, in rat brain | |
| US9605042B2 (en) | Compositions and methods related to tauopathy | |
| Ehrlich et al. | Mapping GPR88-Venus illuminates a novel role for GPR88 in sensory processing | |
| US7314864B1 (en) | Humanin, a polypeptide suppressing neuronal death | |
| Masana et al. | The stress-inducible actin-interacting protein DRR1 shapes social behavior | |
| Elbaz et al. | Reduced synaptic density and deficient locomotor response in neuronal activity‐regulated pentraxin 2a mutant zebrafish | |
| Takagishi et al. | Localization of myosin II and V isoforms in cultured rat sympathetic neurones and their potential involvement in presynaptic function | |
| JP6915810B2 (ja) | SorCSペプチド及びそれらの用途 | |
| Yang et al. | Drosophila Vap-33 is required for axonal localization of Dscam isoforms | |
| Vervenne et al. | Lpp is involved in Wnt/PCP signaling and acts together with Scrib to mediate convergence and extension movements during zebrafish gastrulation | |
| AU2003288434B2 (en) | Peptides, antibodies thereto, and their use in the treatment of central nervous system damage | |
| Reuss et al. | Serine/threonine-kinase 33 (Stk33)–Component of the neuroendocrine network? | |
| CA2421834A1 (en) | Use of carp inhibitors for the treatment of heart diseases | |
| Alexander et al. | Creation of a myosin Va‐TAP‐tagged mouse and identification of potential myosin Va‐interacting proteins in the cerebellum | |
| Cresto et al. | The C-terminal fragment of LRRK2 with the G2019S substitution increases the neurotoxicity of mutant A53T α-synuclein in dopaminergic neurons in vivo | |
| Fallon | Pathological implications of the interaction between neurexins and alpha-synuclein in synucleinopathies | |
| Palata | Investigating the role of IRE1 in brain regeneration using a larval zebrafish model | |
| Kotova | Molecular Mechanisms Underlying Regulation and Function of Neuronal Gap Junction Proteins: Connexin 36 and Connexin 27.5. | |
| Nicolas | Understanding Molecular Mechanisms of Striated Muscle Laminopathies Using Cellular and Zebrafish Models | |
| EP1785728A1 (en) | Methods and devices for selecting compounds that interact with importin alpha7 and related uses thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20181116 |
|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20181220 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20181220 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20181226 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20190919 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20191210 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20191210 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20210105 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210401 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20210608 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20210706 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6915810 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| LAPS | Cancellation because of no payment of annual fees |