WO2009142494A1 - Influenza cap-leader sequence - Google Patents
Influenza cap-leader sequence Download PDFInfo
- Publication number
- WO2009142494A1 WO2009142494A1 PCT/NL2009/050277 NL2009050277W WO2009142494A1 WO 2009142494 A1 WO2009142494 A1 WO 2009142494A1 NL 2009050277 W NL2009050277 W NL 2009050277W WO 2009142494 A1 WO2009142494 A1 WO 2009142494A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- leader
- cap
- viral
- amv3
- influenza
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
- C12N15/1131—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing against viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
- A61P31/16—Antivirals for RNA viruses for influenza or rhinoviruses
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/11—Antisense
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/31—Chemical structure of the backbone
- C12N2310/315—Phosphorothioates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/31—Chemical structure of the backbone
- C12N2310/317—Chemical structure of the backbone with an inverted bond, e.g. a cap structure
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/32—Chemical structure of the sugar
- C12N2310/323—Chemical structure of the sugar modified ring structure
- C12N2310/3233—Morpholino-type ring
Definitions
- This invention relates to the field of viral genome transcription, more particular to the field of cap-leader sequences involved in initiation of Influenza genome transcription, more especially to the field of strong consensus Influenza cap-leader sequences. Further the invention relates to novel therapeutic substances for inhibiting viral infections.
- Cap- snatching is a special mechanism of viral transcription initiation found among the segmented (-)ssRNA viruses and first discovered for Influenza A (Bouloy et al., 1978; Plotch et al., 1979; Krug et al., 1979; Caton and Robertson, 1980; Dhar et al., 1980; Beaton and Krug, 1981; Plotch et al., 1981).
- the viral transcriptase cleaves m 7 G-capped RNA leader sequences from host mRNAs to prime transcription of the viral genome. Nascent viral transcripts thereby are provided a 5' cap structure and a non- viral leader sequence, which allows them to be distinguished from (anti)genomic viral RNA molecules.
- Influenza A virus replicates in the nucleus of the infected cell where it uses nascently synthesized transcripts from the RNA polymerase II complex (Fodor et al., 2000; Engelhardt et al., 2005; Chan et al., 2006).
- the Influenza A polymerase complex consists of three viral proteins, PBl, PB2 and PA, which are all required for viral mRNA synthesis.
- PB2 binds to the 5' cap structure of cellular mRNAs (Licheng et al., 1995), which activates its endonuclease domain to cleave at a site 10-14 nucleotides downstream the 5' cap structure.
- PBl catalyses nucleotide addition (Braam et al, 1983).
- vRNA viral RNA template
- Endonuclease cleavage generally takes place around 15 nt from the 5' cap structure, though variation in leader length between 10 to 20 nucleotides is observed. Exceptions are reported for Dugbe nairovirus (Bunyaviridae) and Tacaribe arenavirus (Arenaviridae) which use relatively short capped RNA leaders (1-4 resp. 5-16 nts). For several viruses, sequence analyses of viral mRNAs have shown a nucleotide preference at the 3' end of the non- viral leader, which is assumed to reflect a sequence preference for endonuclease cleavage.
- the capped leader may as well have been cleaved further downstream the assumed 3' end, e.g. 1-2 nucleotides, to provide several residues which assist in the alignment of capped RNA leader molecules along the viral template by virtue of base pairing.
- This idea is not only plausible but also supported by several lines of evidence presented in literature.
- a "prime-and-realign" mechanism has been proposed for Hantaan virus genome transcription to explain the presence of repetitive sequences within the leader sequence of viral transcripts. During this process re-alignment of a capped leader sequence to the viral RNA template relies on base pairing interaction. Since its first description (Garcin et al, 1995), the presence of repeated sequences in viral leader sequences of viruses from the Bunyaviridae have been attributed, partly in retrospective, to the occurrence of "prime-and- realign" (Bishop et al.
- TSWV prefers capped leaders with multiple base complementarity to the viral template RNA, even when offered at relative low amounts compared to others with less base complementarity (van Knippenberg et al., 2005).
- Influenza viruses can obtain (via mutations) the capacity to spread between different host species as observed for the H5N1 virus (poultry and human). Accordingly it should be feared that H5N1 can mutate into a strain capable of efficient human-to-human transmission.
- Current options to limit the clinical impact of influenza virus infections include vaccination and the use of antiviral drugs. Since the current manufacturing capacity for vaccines is insufficient to cope with the demand during a pandemic, the availability of antiviral drugs may help to contain the emerging pandemic virus at its emergence.
- the efficacy of the two currently available classes of antiviral drugs, neuraminidase inhibitors (e.g. Tamiflu) and ion channel blockers, is limited and heavily dependent of a very early start of treatment. In addition, several cases of resistance have already been reported.
- the inventors have now further specified the requirements for the cap- leader sequence necessary for optimal initiation of the viral genome transcription of (-)ssRNA viruses.
- the current invention provides a cap-leader sequence comprising at least a single up to three base complementarity to the 3' residues of the viral RNA.
- This cap leader preferably has the consensus sequence 7m G (N) 6 -io-(A/U/G)-A/U-AGC.
- a modification of the cap-leader sequence to avoid elongation and/or endonuclease cleavage by the viral polymerase. Said modification preferably is a modification of the 3'-OH terminated cap-leader into a 3'-phosphate.
- said modification is a modification of the phosphodiester bonds within the cap-leader to avoid cleavage by the viral endonuclease (and other host nucleases) to render a cap- leader sequence capable of becoming elongated.
- Said modification preferably is a replacement of the phosphodiester bonds within the cap-leader into phosphorothioate bonds or phosphorodiamidate bonds.
- the complete molecule has phosphorothiate or phosphorodiamidate bonds.
- said modification is a replacement of one or more ribose ring(s) (or when a DNA consensus sequence is used of the dideoxyribose) within the cap-leader with one or more morpholino ring(s).
- the invention further provides for a method to inhibit genome transcription of a (-)ssRNA virus in a cell by providing said cell with a modified cap-leader which has at least three bases complementary to the 3'ultimate residues of the viral template wherein the modifications is chosen from the modifications listed above.
- a modified cap-leader which has at least three bases complementary to the 3'ultimate residues of the viral template wherein the modifications is chosen from the modifications listed above.
- said virus is Influenza (A, B or C) and the 3'-ultimate residues have the sequence UCG.
- the invention pertains to the use of the above describe sequence and method to inhibit or treat a viral infection wherein the viral infection preferably is an Influenza infection.
- the viral infection preferably is an Influenza infection.
- Figure 1 Analysis of in vitro synthesized Influenza niRNA
- Cap(ped)-leader sequence(s) or “cap(ped)-leader(s)” or “leader sequence(s)” or cap(ped) RNA primers are herein defined as m 7 G-capped RNA sequences that can be used to prime transcription of the viral genome.
- the 5' cap is found on the 5' end of an mRNA molecule and consists of a 7- methylguanosine residue connected to the RNA via an unusual 5' to 5' triphosphate linkage (7MeGpppN). It is referred to as a 7-methylguanosine cap, abbreviated m 7 G-cap.
- m 7 G-cap After completion of transcription the viral RNA is provided with a non- viral (not originating from the virus itself) 5' cap structure and non- viral leader sequences.
- Cap-leaders can be of any origin like viral, animal, human or plant, but are mostly of host origin.
- the process of initiation of viral transcription with a cap-leader sequence is also referred to as "cap- snatching". This cap-snatching is one of the key events in viral genome transcription.
- cap-leader sequences Although several reports addressed cap-leader sequences, the specific requirements for cap-leader sequences for Influenza genome transcription have still not been elucidated. The current inventors have now found the requirements for cap-leader sequences necessary for preferential usage during priming of transcription.
- This cap-leader comprises at least a three base complementarity to the 3' ultimate residues of the viral RNA.
- base complementarity refers to the complementarity between the 5'-cap-leader sequences and the 3' viral RNA.
- the viral RNA is herein also referred to as the viral template or virus template sequence.
- the complementary bases bind to the 3' ultimate end of the viral RNA wherein the A, G and C residues of the cap-leader subsequently bind to the 3' ultimate U, penultimate C and following G (also referred to as viral 3' terminal 1, 2, and 3 residues).
- Ultimate is herein defined as viral 3' terminal RNA base residue.
- Penultimate is herein defined as the RNA base subsequent to the ultimate base pair, the second most terminal viral RNA 3' base residue.
- the current invention further identifies that residues in immediate proximity of the three complementary bases are important for efficient initiation and priming of transcription.
- the positions of these residues are referred to as -1 for the base immediately 5' upstream of AGC and -2 for the base at the second position 5' upstream of the AGC in the cap-leader sequence.
- the examples show that an A or T at position -1 and an A, T, or G at position - 2 in the cap-leader results in a very strong privilege for use in priming and elongation.
- the cap leader preferably has the sequence 7m G (N)6-io-(AZUZG)-AZU- AGC which is herein referred to as the "consensus sequence".
- the final AGC in the consensus is predominant and is also referred to as three base complementarity or triple complementarity.
- the consensus sequence also identifies an optimal leader length.
- the experiments performed in the current invention allow for the identification of an optimal length of the leader sequence between 10 and 20 nucleotides, wherein the predominant AGC is preferably at position 10-12 nucleotides or 11-13 nucleotides from the cap 7m G ( 7m G).
- the leader can optionally be extended resulting in the following consensus sequence 7m G (N)6-io-(AZUZG)-AZU-AGC-(N) 6 -i7..
- the leader is cleaved (most likely after the three base complementary A, G, or C) whereafter elongation starts with the three base complementary ACG terminus.
- This elongation can be blocked by modification of the three base complementary AGC terminus.
- Said modification preferably is the addition of a 3'-phosphate group at the 3'-terminus of the cap leader.
- Another possible modification areis a replacement of one or more ribose ring(s) (or when a DNA consensus sequence is used of the dideoxyribose) within the cap-leader with one or more morpholino ring(s). Such a modification confers resistance to vial endonucleases.
- the base complementarity can be extended to 4 or 5 or more bases complementary to the virus 3' RNA.
- the corresponding complementary sequences after the three base AGC are different for respectively Influenza A, B and C.
- an extended cap- leader with a sequence specific for said virus.
- cap-leaders The alignment of cap-leaders to the viral RNA template by virtue of base pairing is the initial step during initiation/priming of transcription. This initiates viral endonuclease cleavage of the capped-leader 10-20 nucleotides from the m 7 G-cap 5' end. The resulting cap-leaders are used by the virus to prime transcription , i.e. elongation of the cap-leader.
- the chain elongation is herein also referred to as transcription, elongation or incorporation.
- “Viral genome transcription” can thus be divided in three steps: initiation (alignment/binding of the cap-leader also referred to as priming of transcription), endonuclease cleavage and elongation. It is obvious for a person skilled in the art that when cap-leaders of shorter length are used, the endonuclease cleavage in the first step of initiation can become unnecessary and direct priming can take place by binding of the cap-leader to the viral RNA after which elongation can take place. On the other hand, cap-leaders shorter than 9 nt are too short to prime transcription. In another aspect the invention relates to modified cap-leaders of the invention comprising a blocking group.
- a blocking group is herein defined as any group that will allow binding of the cap-leader to the viral RNA but inhibits subsequent steps during the process of transcription initiation. Two blocking groups are preferred,to convert cap-leaders into dysfunctional ones, i.e. phosphorothioate bonds and a 3'-phosphate terminus.
- RNA polymerases need a 3'-OH terminus for elongation
- the 3'-phosphate blocking preferably takes place at the 3' terminal C. This will give the following consensus 7m G (N)6-io-(A/U/G)-A/U-AGCp wherein p is a 3'-phosphate group which can be interchanged for any other blocking group with similar functionality.
- Phosphorodiamidate bounds also confer endonuclease resistance to DNA and RNA.
- Morpholino or phospohorodiamidate morpholino refer to chemicals containing a six-membered morpholino ring. When bound to the viral RNA the cap-leader containing the morpholino ring(s) will be more resistant to viral endonuclease cleavage and thus elongation is inhibited.
- the invention provides for a method to modify the cap-leaders or a method for preparing modified cap-leaders.
- a blocking group is added to the cap- leader or wherein DNA or RNA with one or more phosphorothioate bond(s) instead of phosphodiester bond(s) is produced.
- the blocking group preferably is a 3'-phosphate groups or one or more phosphorothioate bonds.
- a person skilled in the art will be able to produce such compounds with blocking groups.
- the 3' phosphate can be added via esterification of the cap-leader with phosphoric acid. It will be known to a skilled person in the art how to make RNA with phosphorothioate bonds.
- the invention relates to a method to modify the cap-leaders or a method for preparing modified cap-leaders, wherein the blocking group is a phosphorodiamidate bond instead of phosphorothioates or phosphodiester bonds, and/or morpholino rings instead of a ribose rings. It will be known to a skilled person in the art how to make RNA with phosphorodiamidate bonds and/or morpholino rings.
- RNA can be carried out by conventional phosphotriester, phosphite or phosphoramidite chemistry, using solid phase techniques such as those described in v Chemical and Enzymatic Synthesis of Gene Fragments--A Laboratory Manual" (ed. H. G. Gassen and A. Lang), Verlag Chemie, Weinheim (1982), For example chemically synthesis can be performed by using an DNA/RNA synthesizer and b-cyanoethyl chemistry.
- the invention provides for a method to inhibit viral genome transcription of a (-)ssRNA virus in a cell by providing said cell with a modified cap RNA leader which has at least three bases complementary to the 3'ultimate residues of the viral RNA wherein the modification consist of a modification on either of the three residues.
- Said virus is preferably Influenza and the 3'-ultimate residues of the virus have the sequence UCG.
- a "cell” is herein defined as any host cell that is or can be infected with the virus.
- the cap-leader can be applied to the cell using any known method used in the state of the art.
- the delivery of modified (dysfunctional) cap-leaders preferably takes place via delivery vehicles such as liposomes, polymers, viral particles (VP), virosomes, viral vectors, lipid particles or (degradable) nano- particles.
- delivery vehicles such as liposomes, polymers, viral particles (VP), virosomes, viral vectors, lipid particles or (degradable) nano- particles.
- virosomes, viral particles or liposomes containing Influenza H and N proteins are used, as these will specifically target the host cells (a.o. respiratory tract) that naturally become infected by Influenza virus.
- Liposomes are herein referred to as vesicles composed of a bilayer membrane, wherein said bilayer typically could be a phospholipid and cholesterol bilayer. Liposomes can be composed of several different kind of lipids for example naturally- derived phospholipids
- liposomes can vary depending on the lipid composition (cationic, anionic, neutral lipid species). Liposomes, have the ability to function as delivery vehicle for several different content like proteins, small molecules, drugs, compounds or nucleic acids like DNA or RNA. In the current invention said liposomes are used to deliver nucleic acids like DNA or RNA. Liposomal content can be delivered into the cell via fusion of the lipid bilayer with the cell membrane (but specific targeting or receptor mediated uptake can also be used). Liposomes can be produced via sonication of phospholipids in water.
- Liposomes of different sizes and different compositions can be made. Liposomes composed with opsonins and ligands to activate endocytosis in cell types, or liposomes with targeting ligands such as monoclonal antibodies (making an immunoliposome), vitamins, or specific antigens can be made. Targeted liposomes can target nearly any cell type in the body. In a preferred embodiment of the infection the liposomes comprise Influenza H and N proteins, as these will specifically target the host cells (a.o. cells in the respiratory tract) that naturally become infected by Influenza virus.
- Virosomes are lipid bilayer vesicles containing viral glycoproteins derived from enveloped viruses. Virosomes are generally produced by extraction of membrane proteins and lipids from enveloped viruses with a detergent, followed by removal of this detergent from the extracted lipids and viral membrane proteins.Virosomes thus represent reconstituted empty virus envelopes, devoid of nucleocapsid and genetic material. In this way virosomes are unable to replicate. In contrast to liposomes, virosomes contain functional viral envelope glycoproteins in the phospholipid bilayer membrane. Virosomes can be used to deliver for example proteins, small molecules, drugs, compounds or nucleic acids like DNA or RNA.
- said virosomes are used to deliver nucleic acids like DNA or RNA.
- the virosomes comprise Influenza H and N proteins, as these will specifically target the host cells (a.o. cells in the respiratory tract) that naturally become infected by Influenza virus.
- Virus like particles or Viral particles (VP) are composed of viral protein(s) of a virus.
- said proteins are embedded within a lipid bi- layer.
- VPs can be used to deliver for example proteins, small molecules, drugs, compounds or nucleic acids like DNA or RNA.
- said VPs are used to deliver nucleic acids like DNA or RNA.
- the VPs comprise Influenza H and N proteins, as these will specifically target the host cells (a.o. cells in the respiratory tract) that naturally become infected by Influenza virus.
- the modified cap-leader sequence of the invention is used to treat viral infections.
- the viral infection can be any (-)ssRNA virus, but preferably is a Influenza infection.
- This aspect of the invention comprises administration of a pharmaceutical vehicle or pharmaceutical composition comprising an active amount of the modified cap- leader.
- the pharmaceutical composition is comprised of the modified cap-leader in or on a delivery vehicle and a pharmaceutical excipient.
- Administration can be in any suitable form known in the state of the art and is not limited to pharmaceutical formulations like, pills, tablets, capsules, powders, lotions, ointments, soft-gels, gels, creams, transdermal patches, solutions, suspensions or inhalers (powders and nebulized solutions or suspensions).
- compositions can be applied orally (pills, tablets and capsules), nasal or pneumonial (via inhalation), topically, intramuscular or intravenous or subcutaneous (via injection). Preferred are injections and inhalations.
- Preparation and administration of the pharmaceutical compositions can be via any known method in the art (See also Martindale the complete drug reference, S Sweetman, 35 the edition 2007). The invention will be explained in more detail in the following, non- limiting examples.
- Virus Influenza virus strain A/Puerto Rico/8/34 was propagated and purified from embryonated chicken eggs.
- allantoic fluid was collected and cleared by low speed centrifugation for 10 min at lOOOg and filtrated through a 0,45 ⁇ m-pore-size filter.
- the filtrate was layered onto 0,5 ml 60% (wt/vol) sucrose (in PBS) - 2,0 ml 20% (wt/vol) sucrose (in PBS) step gradients and centrifuged at 150,00Og for 2 h at 4°C, using a SW41 rotor (Beckmann).
- Virus particles were recovered from the interface, adjusted to 20% (wt/vol) sucrose (in PBS), and loaded onto the top of a 12 ml 20 to 60% (wt/vol) continuous sucrose gradient (in PBS). After centrifugation at 150,00Og for 16 h at 4°C in a SW41 rotor (Beckmann), the virus fraction was collected. Purified virus (10-15 ⁇ g/ ⁇ l) was stocked in small aliquots at -80 0 C prior to use in transcription assays.
- AMV RNA3 (AMV3) constructs and wild-type (wt-AMV3) were generated from plasmid pXO32NcoP3, which contains a full length cDNA clone of AMV3
- Wt-AMV3, Mut-2 and Mut-3 were PCR amplified by using primer AMV3-Rv (complementary to nt 339-313 of the AMV3 sequence) and primers wt-AMV3, mut2AMV3 or mut3-AMV3 respectively.
- MiIt-N 14 an AMV3-Influenza NP mutant which could base pair over a stretch of 14 nt to the 3' end of the viral NSl template, was amplified with primers MutN 14 - AMV3 and NPUP2.
- Point mutants of AMV3 at nt 10 or 11 with the additional three base pairing residues are referred to as G10C11A12G13C14 (in which nt 10 was changed from the wild-type C residue into an G, and nts at positions 13 and 14 into a G and C respectively), A10C11A12G13C14 and U10C11A12G13C14 or C10G11A12G13C14, C10A11A12G13C14 and C10U11A12G13C14 respectively.
- Mutant C10C11A12G13C14 was made by changing only the nucleotides at positions 13 and 14 from the wild- type TT residues into a G and C respectively.
- PCR fragments were purified using the GFX PCR purification kit (Roche), restriction enzyme digested with BamHI/EcoRI and ligated into pUCl9 using T4 DNA ligase (Promega). Individual clones were verified by sequence analysis. Constructs to determine the leader length requirement were made by first amplifying pXO32NcoP3 with primer AMV3-Rv and primers C 7 As, CsAg, C9A10, CioAii, C11A12, C12A13, C13A14, C14A15, C15A16, Ci6Ai 7 or Ci 7 AiS. Primer T7prom3-AMV3 was used in the second PCR amplification. For these constructs, capped transcripts were made directly from the purified PCR fragments without additional cloning. Primer sequences are listed Table 6 (Supplemental Data).
- capped RNA leaders were performed by using the Ambion T7 mMESSAGE mMACHINE kit according to the manufacturer's instruction. Transcription was performed in the presence of cap-analog m 7 G(5')ppp(5')G. Prior to transcription each DNA template was linearized with ⁇ JcoRI. T7 RNA polymerase promoter sequences were introduced upstream of the AMV3 leader sequence by PCR amplification as described above.
- Influenza transcription assays were performed using approximately 10 ⁇ g of purified Influenza in a final volume of 25 ⁇ l, in analogy as previously described for TSWV (van Knippenberg et al., 2002). These assays contained 4 mM Mg acetate, 1 mM of each NTP, 0.1% NP-40, 0.8 U/ ⁇ l RNasin, 2.5 ⁇ l translation mix (amino acid mixture of 1:1:1 — cys; -lys; -met), and were supplemented by the AP-Biotech Rabbit Reticulocyte Lysate system according to the manufacturer's procedures. The amount of AMV3 capped leaders added to the reactions is indicated in the Tables. After incubation for 2 hr at 37 0 C, the reaction mixture was extracted with phenol- chloroform and the RNA ethanol precipitated.
- Influenza NP or NSl mRNAs containing capped 5' leader sequences derived from AMV3 were amplified by RT-PCR (Schmidt and Mueller, 1999; Schramm et al., 2000).
- First- strand cDNA was synthesized using an internal primer for the Influenza NP or NSl gene (NPUPl or NSlUPl respectively) and in the presence of a template- switch oligonucleotide (TS-oligo).
- Superscript II did add a few C residues at the 3' end of the cDNA, which allowed the template- switch oligonucleotide, containing a few G-residues at its 3' end, to subsequently base pair to the first- strand cDNA.
- Superscript II recognized this oligonucleotide as a new template and switched from the viral RNA transcript to the oligonucleotide as template.
- the first-strand cDNA became extended with residues complementary to the template- switch oligonucleotide.
- primer AMV3-leader2 was used instead of primer AMV31eaderl during the PCR amplification.
- PCR products of expected size were gel-purified using the GFX PCR purification kit (Roche), cloned into pGEM-T Easy
- NSl transcripts were reverse transcribed into single- stranded (ss) cDNA as described earlier, using an internal primer for the NSl gene (NSlUPl) and the TS-oligo primer.
- the first-strand cDNA was applied in a SYBR Green 1-based quantitative real-time PCR according to Zwart et al. (2007), with minor modifications: an annealing temperature of 50° C was used and template concentrations were determined using the standard curve method (RotorGene 6.0 software, Corbett Research). Template cDNA was diluted 500-fold.
- a PCR reaction was performed with the AMV3-leaderl and NS1UP2 primers to determine the AMV3-NS1 cDNA concentration in the sample.
- Influenza A virus particles support genome transcription in the presence of RRL
- an in vitro transcription assay was established (Materials and Methods), in analogy to TSWV (van Knippenberg et al., 2002), in which mature virus particles support transcription.
- In vitro reaction products from assays performed in the presence of radionucleotides were resolved by RNA electrophoresis and clearly showed that Influenza virus purified from embryonated eggs, supported the synthesis of RNA molecules in the presence of RRL, but hardly or not when absent from the reaction (Figure IA), like what earlier was observed for TSWV.
- NSl mRNA molecules from the in vitro reaction were cloned by reverse transcription (RT)-PCR and sequence analyzed for the presence of 5' non- viral leader sequences, either corresponding to the leader sequence of ⁇ -globin mRNAs present from the RRL, or of those from exogenously added AMV3 cap donors. Since the 5' non- viral leader sequence of Influenza transcripts are only 9-13 nts, primers to these sequences would not be specific enough due to their short length. The selective RT-PCR strategy as earlier applied to amplify de novo synthesized TSWV transcripts could thus not be used (Duijsings et al,
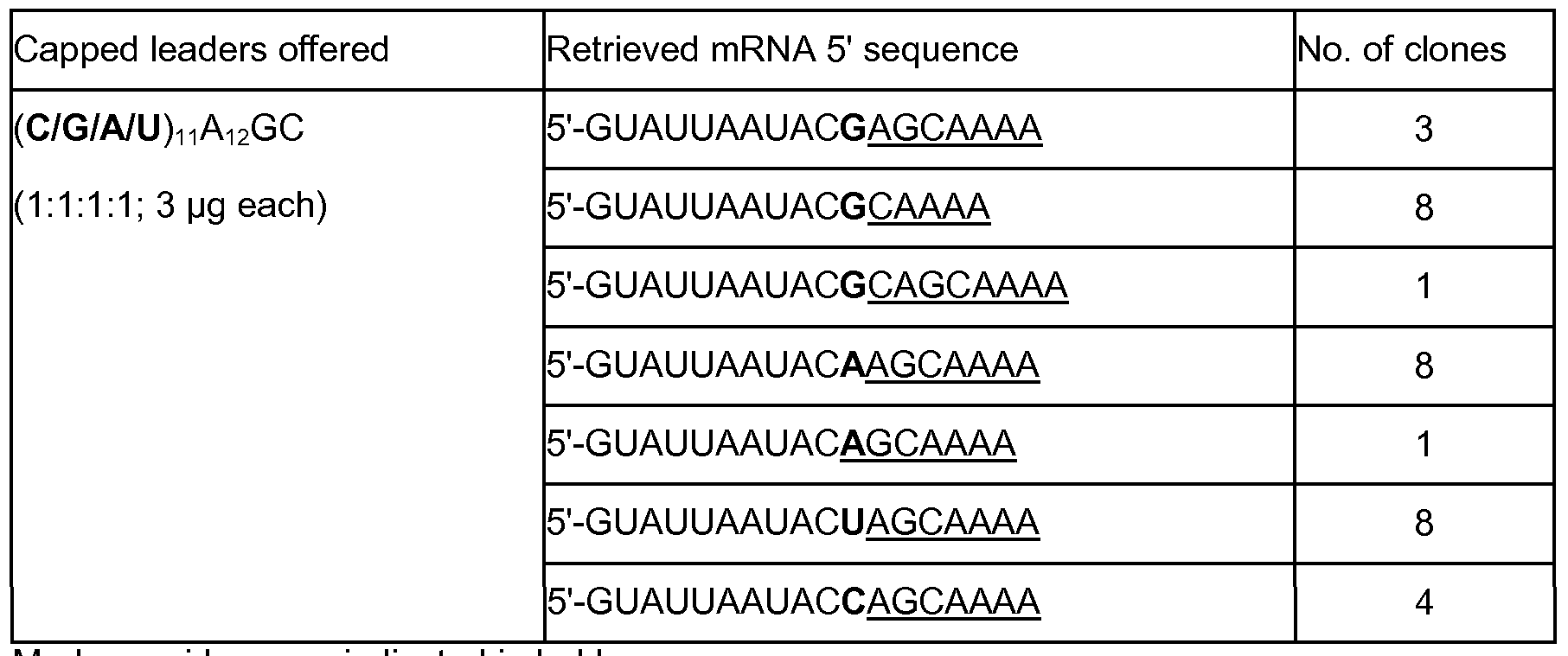
- Example 2 Influenza A virus prefers capped RNA leaders with more base complementarity to the viral RNA template
- Influenza A prefers capped RNA leader molecules with more base complementarity to the viral RNA template, in analogy to what has earlier been demonstrated for TSWV (van Knippenberg et al., 2005), four AMV3 leader constructs (Table 1) were made that only differed in their base complementarity to the 3' end of the viral RNA template, and provided to the in vitro Influenza A virus transcription assay to be tested as cap donor in pair- wise competition.
- the first, single base pairing construct was represented by wild type (wt) AMV3, showing single base complementarity to the viral RNA template with the A residue at nucleotide position 12 downstream the 5' capped end.
- the second, double base pairing construct (denoted Mut-2) was derived from wt-AMV3 and harbored two base pairing (AG) residues at position 12 and 13 downstream the 5' capped end.
- a third, triple base pairing construct (denoted Mut-3) derived from wt- AMV3, harbored three base pairing (AGC) residues at position 12 to 14 downstream the 5' capped end.
- the fourth construct denoted MUt-N 14 , reflected a genuine Influenza A virus transcript, and consisted of the AMV3 leader sequence 5' of the Influenza A virus NP gene which harbored 14 nts complementary to the first 14 nts of the NSl genomic RNA template (Table 1).
- a marker nucleotide was additionally introduced in each of these constructs by changing the Cio residue within the leader sequence into A, G or T respectively (Table 1).
- AMV3 constructs were made that harbored either a C (construct wt-AMV3, also referred to as C10C11A12), A (C10A11A12) , G (C10G11A12) or U (C10U11A12) at the -1 position of the assumed (single) base pairing residue A12 (Table 1).
- the nucleotide changes at position 11 were simultaneously used as a marker nucleotide to discriminate between leaders derived from wt-AMV3 and the 3 derived mutants.
- Synthetic capped transcripts of all four AMV3 constructs were offered in equimolar amounts to the in vitro assay and de novo synthesized NSl gene transcripts were RT-PCR amplified, cloned and sequenced.
- 18 out of the 32 clones analyzed contained the G nucleotide at position 11 (Table 3), indicating a preference for the C10G11A12 sequence.
- the resulting constructs were referred to as C10C11A12G13C14, C10A11A12G13C14, C10G11A12G13C14 and C10U11A12G13C14, and C10C11A12G13C14, A10C11A12G13C14, G10C11A12G13C14 and U10C11A12G13C14 respectively.
- the changed nucleotide at position 11 (-1 relative to the first base pairing residue) or 10 (-2 relative to the first base pairing residue) were simultaneously used as marker nucleotide.
- RNA leader with reciprocal 5' upstream sequences Based on the data described above, AMV3-derived single base pairing capped leader molecules harboring a 3' CAA were relatively well used in competitions involving other AMV3 single base pairing capped leader molecules but those harboring a 3' CCA not. To analyze whether this influence was also found in pair- wise competitions of these molecules with triple base pairing cap donor molecules, thereby underscore the importance of nucleotide sequences just upstream the putative base pairing residues, two additional pair- wise competition assays were performed.
- C10A11A12 and C10C11A12G13C14 (Table 1), that differed in their base complementarity to the viral RNA template and one residue at position -1 just upstream the putative base pairing residues of the RNA leader sequence, were tested and in the second one their reciprocal leader variants, referred to as C10C11A12 (identical to wt-AMV3) and C10A11A12G13C14 (Table 1).
- C10C11A12 identical to wt-AMV3
- C10A11A12G13C14 Table 1
- 14 out of 18 clones analyzed contained a leader from mutant C10C11A12G13C14 and 4 from C10A11A12.
- All cap donor molecules offered contained a CAGC sequence, embedded within an oligo U stretch, to guarantee a proper alignment of the capped RNA leader along the viral RNA template by base pairing of the trinucleotide AGC.
- Transcripts of all leader mutants were simultaneously offered in equimolar amounts to an in vitro Influenza transcription assay.
- RT- PCR cloning and sequence analysis of de novo synthesized viral transcripts revealed that 9 out 19 transcripts contained capped leaders originating from AMV3 mutant C9A10G11C12, and 5 out of 19 were originated from mutant C10A11G12C13 (Table 4), suggesting an optimal leader length of 10-11 nucleotides.
- Several clones revealed the presence of repetitive sequences, again pointing towards the occurrence of "prime-and-realign".
- the relative amount of ⁇ -globin leader initiated NSl transcription was analyzed by RT-PCR and revealed that increasing amounts of C10C11A12 reduced, by competition, globin leader initiated NSl transcription to a lesser extent than C10C11A12G13C14 and C10A11A12G13C14 did (data not shown).
- the ratio of AMV3-NS1 to ⁇ -globin-NSl was determined by means of a two step quantitative real-time PCR. The de novo synthesized NSl transcripts were first reverse transcribed into single- stranded cDNA and then PCR analyzed (see Materials and Methods).
- capped-RNA leaders of wt AMV, Mut-2, Mut-3 and Mut-N14 (Table 7) where transfected to Influenza virus-infected cells to be tested as cap-donor in pair-wise competitions.
- the presence of the marker nucleotide in de novo synthesized viral transcripts enabled the identification of RNA leader sequences.
- Mut-3 Like in the in vitro analysis, the competition between Mut-2 and Mut-3 did not show a strong preference for Mut-3 (Table 8). This is possibly explained by the sequence context of the Mut-3 leader in which a trinucleotide repetition (AGC) may offer various possibilities for base-pairing to the viral RNA template, and thereby possibly weakening the competitor strength with Mut-2.
- AGC trinucleotide repetition
- Influenza virus in uiuo/during infection of cell cultures prefers capped RNA leaders with more base-complementarity (around position 10-12 from the cap- structure) to the 3'- end of the viral RNA template.
- nucleotide residues potentially base pairing to the viral template (3'- UCG7) are underlined; the marker nucleotide used to discriminate between the different AMV3 leaders is shaded.
- Table 2 NSl mRNA 5' end sequences resulting from cap-snatching of the indicated capped leaders
- Influenza viral sequence is underlined; the marker nucleotide of each mutant is shaded.
- Table 3 NSl mRNA 5' end sequences resulting from cap-snatching of the indicated capped leaders
- Influenza viral sequence is underlined; the marker nucleotide of each mutant is shaded. Table 4. NP mRNA 5' end sequences resulting from cap-snatching of the indicated capped leaders
- nucleotide residues potentially base pairing to the viral template (3'- UCG7) are underlined; the marker nucleotide used to discriminate between the different AMV3 capped leaders is shaded.
- Influenza viral sequence is underlined; the 3'-terminal nucleotide of a capped leader which could potentially influence the use of the leader in the cap- snatching process is shaded.
- the proteins encoded by the cloned segments are indicated on the right.
- NPUP2 ⁇ '-CCCGAATTCGCTTGGCGCCAGATTCGC C wt-AMV3 5'-CCCGGATCCTAATACGACTCACTATAGTATTAATACCATTTTCAAAATATTCC Mut2-AlVr 5'-CCCGGATCCTAATACGACTCACTATAGTATTAATAACAGTTTCAAAATATTCC Mut3-AlVr 5'-CCCGGATCCTAATACGACTCACTATAGTATTAATAGCAGCTTCAAAATATTCC
- Ci 4 Ai 5 ⁇ '-CGACTCACTATAGTATTTTTTTTTTCAGCTTTTTTTCAAAATATTCCAAT
- Cianci C. et al. 1995, J. Virol. 69:3995-3999.
- Van Knippenberg I. Et al., 2002, Virology 303:278-286. Van Knippenberg, I. et al., 2005, Virology 335:122-130.
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Virology (AREA)
- Biomedical Technology (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Molecular Biology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Biophysics (AREA)
- Animal Behavior & Ethology (AREA)
- Biochemistry (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Veterinary Medicine (AREA)
- Communicable Diseases (AREA)
- Oncology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Pharmacology & Pharmacy (AREA)
- Microbiology (AREA)
- Public Health (AREA)
- Pulmonology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description



Claims
Priority Applications (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CA2725289A CA2725289A1 (en) | 2008-05-20 | 2009-05-20 | Influenza cap-leader sequence |
| US12/993,829 US20110165226A1 (en) | 2008-05-20 | 2009-05-20 | Influenza cap-leader sequence |
| JP2011510444A JP2011520464A (en) | 2008-05-20 | 2009-05-20 | Influenza cap-leader sequence |
| AU2009249883A AU2009249883A1 (en) | 2008-05-20 | 2009-05-20 | Influenza cap-leader sequence |
| EP09750814A EP2297324A1 (en) | 2008-05-20 | 2009-05-20 | Influenza cap-leader sequence |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP08156582A EP2123758A1 (en) | 2008-05-20 | 2008-05-20 | Influenza cap-leader sequence |
| EP08156582.2 | 2008-05-20 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2009142494A1 true WO2009142494A1 (en) | 2009-11-26 |
Family
ID=39590398
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/NL2009/050277 Ceased WO2009142494A1 (en) | 2008-05-20 | 2009-05-20 | Influenza cap-leader sequence |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20110165226A1 (en) |
| EP (2) | EP2123758A1 (en) |
| JP (1) | JP2011520464A (en) |
| AU (1) | AU2009249883A1 (en) |
| CA (1) | CA2725289A1 (en) |
| WO (1) | WO2009142494A1 (en) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2023018560A1 (en) * | 2021-08-13 | 2023-02-16 | Georgia State University Research Foundation, Inc. | Bicyclic fused pyrazole derivatives for the treatment of respiratory infections including rsv |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US10731161B2 (en) | 2013-03-11 | 2020-08-04 | The Johns Hopkins University | Influenza-activated constructs and methods of use thereof |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5837852A (en) * | 1993-10-14 | 1998-11-17 | Bristol-Myers Squibb Company | Capped nucleic acid oligomers that inhibit cap-dependent transcription of the influenza virus endonuclease |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20030026782A1 (en) * | 1995-02-07 | 2003-02-06 | Arthur M. Krieg | Immunomodulatory oligonucleotides |
| EP1144632A3 (en) * | 1998-12-23 | 2001-11-07 | Corixa Corporation | Compounds for immunotherapy and diagnosis of colon cancer and methods for their use |
-
2008
- 2008-05-20 EP EP08156582A patent/EP2123758A1/en not_active Ceased
-
2009
- 2009-05-20 AU AU2009249883A patent/AU2009249883A1/en not_active Abandoned
- 2009-05-20 CA CA2725289A patent/CA2725289A1/en not_active Abandoned
- 2009-05-20 JP JP2011510444A patent/JP2011520464A/en active Pending
- 2009-05-20 WO PCT/NL2009/050277 patent/WO2009142494A1/en not_active Ceased
- 2009-05-20 EP EP09750814A patent/EP2297324A1/en not_active Withdrawn
- 2009-05-20 US US12/993,829 patent/US20110165226A1/en not_active Abandoned
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5837852A (en) * | 1993-10-14 | 1998-11-17 | Bristol-Myers Squibb Company | Capped nucleic acid oligomers that inhibit cap-dependent transcription of the influenza virus endonuclease |
Non-Patent Citations (4)
| Title |
|---|
| CHUNG T D Y ET AL: "BIOCHEMICAL STUDIES ON CAPPED RNA PRIMERS IDENTIFY A CLASS OF OLIGONUCLEOTIDE INHIBITORS OF THE INFLUENZA VIRUS RNA POLYMERASE", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE USA, NEW YORK, NY, US, vol. 91, no. 6, 15 March 1994 (1994-03-15), pages 2372 - 2376, XP000600722 * |
| DE CLERCQ ERIK: "Antiviral agents active against influenza A viruses.", NATURE REVIEWS. DRUG DISCOVERY DEC 2006, vol. 5, no. 12, December 2006 (2006-12-01), pages 1015 - 1025, XP002488967, ISSN: 1474-1776 * |
| GABRIEL GUELSAH ET AL: "Morpholino oligomers targeting the PB1 and NP genes enhance the survival of mice infected with highly pathogenic influenza A H7N7 virus", JOURNAL OF GENERAL VIROLOGY, vol. 89, no. Part 4, April 2008 (2008-04-01), pages 939 - 948, XP002488966, ISSN: 0022-1317 * |
| TADO M ET AL: "Inhibitory effect of modified 5'-capped short RNA fragments on influenza virus RNA polymerase gene expression", ANTIVIRAL CHEMISTRY & CHEMOTHERAPY, BLACKWELL SCIENTIFIC PUBL., LONDON, GB, vol. 12, no. 6, 1 November 2001 (2001-11-01), pages 353 - 358, XP009103188, ISSN: 0956-3202 * |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2023018560A1 (en) * | 2021-08-13 | 2023-02-16 | Georgia State University Research Foundation, Inc. | Bicyclic fused pyrazole derivatives for the treatment of respiratory infections including rsv |
Also Published As
| Publication number | Publication date |
|---|---|
| AU2009249883A1 (en) | 2009-11-26 |
| JP2011520464A (en) | 2011-07-21 |
| EP2123758A1 (en) | 2009-11-25 |
| EP2297324A1 (en) | 2011-03-23 |
| US20110165226A1 (en) | 2011-07-07 |
| CA2725289A1 (en) | 2009-11-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7177127B2 (en) | Conjugated antisense compounds and their uses | |
| EP3004347B1 (en) | Double-stranded agents for delivering therapeutic oligonucleotides | |
| Lin et al. | Intronic microrna (mirna) | |
| CN107236735B (en) | Oligonucleotide-based inhibitors comprising locked nucleic acid motifs | |
| JP2021519071A (en) | Nucleic acid molecule for pseudouridine formation | |
| CA2308852A1 (en) | Rnase l activators and antisense oligonucleotides effective to treat rsv infections | |
| AU2010258875A1 (en) | Chemical modification motifs for miRNA inhibitors and mimetics | |
| BRPI0618214A2 (en) | inhibition of replication of influenza virus replication | |
| EP3353301A1 (en) | Antisense oligonucleotides and uses thereof | |
| JPH11137260A (en) | Anti-influenza viral cyclic dumbbell type rna-dna chimera compound and anti-influenza viral agent | |
| JP7634542B2 (en) | Use of SEPT9 inhibitors for treating hepatitis B virus infection - Patents.com | |
| Geerts-Dimitriadou et al. | Base-pairing promotes leader selection to prime in vitro influenza genome transcription | |
| WO2023009396A2 (en) | Structure-based design of antisense oligonucleotide drugs | |
| US20100292299A1 (en) | Nucleotide Motifs Providing Localization Elements and Methods of Use | |
| EP2123758A1 (en) | Influenza cap-leader sequence | |
| Abe et al. | Antisense therapy of influenza | |
| US20090117179A1 (en) | siDNA against Influenza Virus | |
| Abe et al. | Specific inhibition of influenza virus RNA polymerase and nucleoprotein gene expression by liposomally encapsulated antisense phosphorothioate oligonucleotides in MDCK cells | |
| US20170183653A1 (en) | Pri-mirna libraries and methods for making and using pri-mirna libraries | |
| WO1999033970A1 (en) | Medicinal compositions for treating or preventing influenza and novel capped oligoribonucleotides | |
| WO2016190899A1 (en) | Pri-mirna libraries and methods for making and using pri-mirna libraries | |
| US9546364B2 (en) | Synthetic lariat RNA for RNA interference | |
| JP2004283024A (en) | New antisense oligonucleotide and anti-hiv agent | |
| CN115087451A (en) | Methods and compositions for inhibiting hepatitis B and hepatitis D virus infection | |
| Lewin | Regulatory RNA in gene therapy |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 09750814 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2725289 Country of ref document: CA Ref document number: 2011510444 Country of ref document: JP Ref document number: 2009249883 Country of ref document: AU |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2009750814 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: 2009249883 Country of ref document: AU Date of ref document: 20090520 Kind code of ref document: A |