WO2011034946A1 - Generation of high polyhydroxybutrate producing oilseeds - Google Patents
Generation of high polyhydroxybutrate producing oilseeds Download PDFInfo
- Publication number
- WO2011034946A1 WO2011034946A1 PCT/US2010/048963 US2010048963W WO2011034946A1 WO 2011034946 A1 WO2011034946 A1 WO 2011034946A1 US 2010048963 W US2010048963 W US 2010048963W WO 2011034946 A1 WO2011034946 A1 WO 2011034946A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- plant
- seeds
- transgenic
- expression
- transgenic plant
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8214—Plastid transformation
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
- C12N15/8257—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits for the production of primary gene products, e.g. pharmaceutical products, interferon
Definitions
- the invention is generally related to the field of polymer production in transgenic plants. Methods for generating industrial oilseeds producing high levels of polyhydroxybutyrate (PHB) and industrial oilseeds producing high levels of PHB are described.
- PHB polyhydroxybutyrate
- PHAs polyhydroxyalkanoates
- PHAs are a natural component of numerous organisms in multiple ecosystems and accumulate in a wide range of bacteria as a granular storage material when the microbes are faced with an unfavorable growth environment, such as a limitation in an essential nutrient (Madison et al., Microbiol. Mol. Biol Rev., 1999, 63, 21-53; Suriyamongkol et al.,
- transgenic oilseeds having commercially viable levels of polyhydroxyalkanoates in the seed, for example greater than 7%, 10%, 15%, or 19% polyhydroxyalkanoate or more of the total dry seed weight and capable of germinating.
- Transgenic oilseed plants are provided.
- the transgenic oilseed plants synthesize
- PHB polyhydroxybutyrate
- Host plants, plant tissue, and plant material have been engineered to express genes encoding enzymes in the biosynthetic pathway for PHB production such that polymer precursors in the plastid are polymerized to polymer.
- Genes utilized include phaA, phaB, phaC, all of which are known in the art.
- the genes can be introduced in the plant, plant tissue, or plant cell using conventional plant molecular biology techniques.
- the transgenes encoding PHA biosynthesis are expressed in a seed specific manner such that the PHA accumulates in the seed.
- the level of PHA accumulated is greater than %, 8%, 9%, 10%, 11%, 12%, 13%. 14%, 15%, 16%, 17%, 18% and 19% of the dry weight of the seed.
- these transgenic oilseeds encode one or more additional transgenes to improve the germination efficiency of high PHA producing oilseeds where the level of PHA in the oilseed is greater than 8% by weight and where the seeds germinate to at least 10%, 20%, 40%, 60%, 80%, 90%, 100% of the level of seeds from the unmodified parental line or seeds with low levels of PHA.
- These additional transgenes can encode siRNA for one or more of the genes encoding enzymes for producing PHA. These additional transgenes can encode one or more genes involved in the PHA degradation pathway. These additional transgenes can encode one or more enzymes involved in photosynthesis pathways. In a more preferred embodiment, these additional transgenes can be expressed under the control of an inducible regulatory element or promoter. In another embodiment, these additional transgenes can be placed under the control of a minimal promoter such that very low levels of expression are obtained.
- these additional transgenes can be placed under the control of a germination specific promoter, such as the promoter from Vigna mungo sulphydryl-endopeptidase gene (SH-EP promoter; Akasofu et al. , 1990 Nucleic Acids Research. 18, 1892).
- a germination specific promoter such as the promoter from Vigna mungo sulphydryl-endopeptidase gene (SH-EP promoter; Akasofu et al. , 1990 Nucleic Acids Research. 18, 1892).
- the transgenic oilseed may encode
- transgenes for example transgenes encoding siR A plus transgenes encoding one of more enzymes involved in photosynthesis pathways.
- additional transgenes or other transgenes and approaches to solving this previously unknown problem will be obvious to those skilled in the art.
- Transgenic plants useful for the invention include dicots or monocots.
- Preferred host plants are oilseed plants, but are not limited to members of the Brassica family including B. nap s, B. rapa, B. carinata and B.juncea;
- oilseeds such as Camelina sativa, Crambe, jatropha, castor;
- Arabidopsis thaliana Calendula, Cuph a; maize; soybean; cottonseed; sunflower; palm; coconut; safflower; peanut; mustards including Sinapis alba;; and tobacco.
- Other embodiments provide plant material and plant parts of the transgenic plants including seeds, flowers, stems, and leaves.
- the oilseeds can be used for the extraction of PHA biopolymer or as a source of PHA biopolymer based chemical intermediates.
- the residual parts of the seed can be used as meal for animal feed or steam and power generation and a source of vegetable oil for industrial oelochemicals or biofuel.
- Figure 1 is a schematic diagram describing an ecdysone inducible promoter system.
- Figure 2 is a bar graph showing percent PHB content in select T2 and
- T3 PHB producing seeds obtained from transformations of vectors containing the PHB pathway genes and a cassette for siRNA to either the thiolase or synthase gene.
- a lines were obtained from transformations with vector pPhaA-R Ai/35S.
- B lines were obtained from transformations with vector pPhaA-RNAi/glyP.
- C lines were obtained from transformations with vector pPhaC-RNAi/35S.
- D lines were obtained from transformations with vector pPhaC-RNAi/glyP.
- Figure 3 is a schematic diagram describing a strategy for using a polymer degradation pathway to enable seed germination.
- Figure 4 is a bar graph showing percent PHB content in select T2 and
- T3 PHB producing seeds obtained from transformations of vector pMBXVTl containing the PHB pathway genes expressed under the control of seed specific promoters and expression cassettes for a degradation pathway consisting of depolymerase and dehydrogenase expressed under the control of a germination specific promoter.
- Figure 5 is a schematic diagram describing a strategy for creating hybrid seeds using cytoplasmic male sterility.
- Figure 6 is a protein sequence alignment of FBPase/SBPase genes in transformation vectors pMBXS407 and pMBXS408.
- Vector pMBXS407 contains a gene encoding a FBPase/SBPase with 100% homology to the FBPase/SBPase protein from S nechococcus elongatw PCC 7942 listed in accession CP000100.
- Transformation vector pMBXS408 contains a gene encoding a FBPase/SBPase with 100% homology to the FBPase/SBPase protein from Synechococcus elongatus PCC 7942 listed in accession
- the disclosure encompasses all conventional techniques of plant breeding, microbiology, cell biology and recombinant DNA, which are within the skill of the art. See, e.g., Sambrook and Russell, Molecular Cloning: A Laboratory Manual, 3rd edition (2001); Current Protocols In Molecular Biology [(F. M. Ausubel, et al. eds., (1987)]; Plant Breeding: Principles and Prospects (Plant Breeding, Vol 1) M. D.
- PHB refers to polyhydroxybutyrate and is used
- PHA polyhydroxyalkanoate
- PHB also encompasses copolymers of hydroxybutyrate with other hydroxyacid monomers.
- PHA copolymer refers to a polymer composed of at least two different hydroxyalkanoic acid monomers.
- PHA homopolymer refers to a polymer that is composed of a single hydroxyalkanoic acid monomer.
- a "vector” is a replicon, such as a plasmid, phage, or cosmid, into which another DNA segment may be inserted so as to bring about the replication of the inserted segment
- the vectors can be expression vectors.
- an "expression vector” is a vector that includes one or more expression control sequences
- an "expression control sequence” is a DNA sequence that controls and regulates the transcription and/or translation of another DNA sequence.
- Control sequences that are suitable for prokaryotes include a promoter, optionally an operator sequence, a ribosome binding site, and the like.
- Eukaryotic cells are known to utilize promoters, polyadenylation signals, and enhancers.
- operably linked means incorporated into a genetic construct so that expression control sequences effectively control expression of a coding sequence of interest.
- transformed and transfected encompass the introduction of a nucleic acid into a cell by a number of techniques known in the art.
- heterologous means from another host.
- the other host can be the same or different species.
- cell refers to a membrane-bound biological unit capable of replication or division.
- construct refers to a recombinant genetic molecule including one or more isolated polynucleotide sequences.
- Genetic constructs used for transgene expression in a host organism comprise in the 5 -3' direction, a promoter sequence; a nucleic acid sequence encoding the desired transgene product; and a termination sequence.
- the open reading frame may be orientated in either a sense or anti-sense direction.
- the construct may also comprise selectable marker gene(s) and other regulatory elements for expression.
- plant is used in it broadest sense. It includes, but is not limited to, any species of woody, ornamental or decorative, crop or cereal, fruit or vegetable plant, and photosynthetic green algae (e.g.,
- Chlamydomonas reinhardtii It also refers to a plurality of plant cells that are largely differentiated into a structure that is present at any stage of a plant's development. Such structures include, but are not limited to, a fruit, shoot, stem, leaf, flower petal, etc.
- plant tissue includes differentiated and undifferentiated tissues of plants including those present in roots, shoots, leaves, pollen, seeds and tumors, as well as cells in culture (e.g., single cells, protoplasts, embryos, callus, etc.). Plant tissue may be in planta, in organ culture, tissue culture, or cell culture.
- plant part as used herein refers to a plant structure, a plant organ, or a plant tissue.
- a non-naturally occurring plant refers to a plant that does not occur in nature without human intervention.
- Non-naturally occurring plants include transgenic plants and plants produced by non-transgenic means such as plant breeding.
- plant cell refers to a structural and physiological unit of a plant, comprising a protoplast and a cell wall.
- the plant cell may be in form of an isolated single cell or a cultured cell, or as a part of higher organized unit such as, for example, a plant tissue, a plant organ, or a whole plant
- plant cell culture refers to cultures of plant units such as, for example, protoplasts, cell culture cells, cells in plant tissues, pollen, pollen tubes, ovules, embryo sacs, zygotes and embryos at various stages of development.
- plant material refers to leaves, stems, roots, flowers or flower parts, fruits, pollen, egg cells, zygotes, seeds, cuttings, cell or tissue cultures, or any other part or product of a plant.
- Plant organ refers to a distinct and visibly structured and differentiated part of a plant such as a root, stem, leaf, flower bud, or embryo.
- Plant tissue refers to a group of plant cells organized into a structural and functional unit. Any tissue of a plant, whether in a plant or in culture, is included. This term includes, but is not limited to, whole plants, plant organs, plant seeds, tissue culture and any groups of plant cells organized into structural and/or functional units. The use of this term in conjunction with, or in the absence of, any specific type of plant tissue as listed above or otherwise embraced by this definition is not intended to be exclusive of any other type of plant tissue.
- “Seed germination” refers to growth of an embryonic plant contained within a seed resulting in the formation and emergence of a seedling.
- Cotyledon refers to the embryonic first leaves of a seedling.
- Early plantlet development refers to growth of the cotyledon containing seedling to form a plantlet
- Transgenic plants have been developed that produce increased levels of biopolymers such as polyhydroxyalkanoates (PHAs) in seeds. Methods and constructs for engineering plants for seed specific production of PHA, in particular PHB, are described.
- PHA polyhydroxyalkanoates
- One embodiment provides transgenic plants for the direct, large scale production of PHAs in crop plants or in energy crops where a plant by-product, such as oil, can be used for production of energy.
- PHB polyhydroxybutyrate
- Transgenic oilseeds comprising at least about 8% dry weight PHA are provided.
- One embodiment provides transgenic oilseeds having at least 10% PHA dry weight and which are impaired in germination and plant survival.
- Suitable genetic constructs include expression cassettes for enzymes for production of polyhydroxyalkanoates, in particular from the
- the construct contains operatively linked in the 5' to 3' direction, a seed specific promoter that directs transcription of a nucleic acid sequence in the nucleus; a nucleic acid sequence encoding one of the PHB biosynthetic enzymes; and a 3' polyadenylation signal that increases levels of expression of transgenes.
- enzymes for formation of polymer precursors are targeted to the plastid using appropriate plastid-targeting signals.
- a cassette containing DNA sequences homologous to a portion of one of the transgenes and designed to promote R A interference (RNAi) is included.
- this cassette for RNAi contains an intron between an inverted repeat.
- a cassette with homology to one of the PHB pathway genes is designed to produce antisense RNA thus attenuating the level of translation into protein.
- the PHA pathway is expressed directly from the plastid genome using appropriate plastidial promoters and regulatory sequences.
- the construct contains operatively linked in the 5' to 3' direction, a promoter that directs transcription of a nucleic acid sequence in the nucleus; a nucleic acid sequence encoding genes for PHA degradation to enable seed germination; and a 3' polyadenylation signal that increases levels of expression of transgenes.
- enzymes for degradation of polymer are targeted to the plastid using appropriate plastid-targeting signals.
- enzymes for polymer degradation include a depolymerase and/or dehydrogenase.
- the construct contains operatively linked in the 5' to 3' direction, a promoter that directs transcription of a nucleic acid sequence in the nucleus; a nucleic acid sequence encoding a gene to capable of increasing photosynthesis in a plant; and a 3' polyadenylation signal that increases levels of expression of transgenes.
- genes to increase photosynthesis include enzymes capable of increasing carbon flow through the Calvin Cycle.
- enzymes for increasing photosynthesis are targeted to the plastid using appropriate plastid-targeting signals.
- DNA constructs useful in the methods described herein include transformation vectors capable of introducing transgenes into plants.
- transgenic refers to an organism in which a nucleic acid fragment containing a heterologous nucleotide sequence has been introduced.
- the transgenes in the transgenic organism are preferably stable and inheritable.
- the heterologous nucleic acid fragment may or may not be integrated into the host genome.
- Plant transformation vectors generally include one or more coding sequences of interest under the transcriptional control of 5' and 3' regulatory sequences, including a promoter, a transcription termination and/or polyadenylation signal, and a selectable or screenable marker gene.
- additional RNA processing signals and ribozyme sequences can be engineered into the construct (U.S. Pat. No. 5,519,164). This approach has the advantage of locating multiple transgenes in a single locus, which is advantageous in subsequent plant breeding efforts.
- Engineered minichromosomes can also be used to express one or more genes in plant cells.
- Cloned telomeric repeats introduced into cells may truncate the distal portion of a chromosome by the formation of a new telomere at the integration site.
- a vector for gene transfer can be prepared by Irimming off the arms of a natural plant chromosome and adding an insertion site for large inserts (Yu et al., Proc Natl Acad Sci U S A, 2006, 103, 17331-6; Yu et al, Proc Natl Acad Sci USA, 2007, 104, 8924- 9).
- the utility of engineered minichromosome platforms has been shown using Cre/lox and FRT FLP site-specific recombination systems on a maize minichromosome where the ability to undergo recombination was
- chromosome engineering in plants involves in vivo assembly of autonomous plant minichromosomes (Carlson et al, PLoS Genet, 2007, 3, 1965-74). Plant cells can be transformed with centromeric sequences and screened for plants that have assembled autonomous chromosomes de novo. Useful constructs combine a selectable marker gene with genomic DNA fragments containing centromeric satellite and retroelement sequences and/or other repeats.
- ETL Engineered Trait Loci
- US Patent 6,077,697 to Hadlaczky et al.; US Patent Application 2006/0143732 This system targets DNA to a heterochromatic region of plant chromosomes, such as the pericentric heterochromatin, in the short arm of acrocentric chromosomes.
- Targeting sequences may include ribosomal DNA (rDNA) or lambda phage DNA.
- rDNA ribosomal DNA
- the pericentric rDNA region supports stable insertion, low recombination, and high levels of gene expression.
- This technology is also useful for stacking of multiple traits in a plant (US Patent Application 2006/0246586, 2010/0186117 and PCT WO 2010/037209).
- Zinc-finger nucleases are also useful in that they allow double strand DNA cleavage at specific sites in plant chromosomes such that targeted gene insertion or deletion can be performed (Shukla et al., Nature, 2009; Townsend et al., Nature, 2009).
- a vector to transform the plant plastid chromosome by homologous recombination (as described in U.S. Pat. No. 5,545,818 to McBride et al.) is used in which case it is possible to take advantage of the prokaryotic nature of the plastid genome and insert a number of transgenes as an operon.
- WO 2010/061186 describes an alternative method for introducing genes into the plastid chromosome using an adapted endogenous cellular process for the transfer of RNAs from the cytoplasm to the plastid where they are incorporated by homologous recombination.
- a transgene may be constructed to encode a multifunctional enzyme through gene fusion techniques in which the coding sequences of different genes are fused with or without linker sequences to obtain a single gene encoding a single protein with the activities of the individual genes.
- a transgene may be constructed to encode a series of enzyme activities separated by intein sequences such that on expression, two or more enzyme activities are expressed from a single promoter as described by Snell in US patent No. 7,026,526 to Metabolix, Inc. 1. Genes involved in Polyhydroxyalkanoate Synthesis
- the products of the transgenes are enzymes and other factors required for production of a biopolymer, such as a polyhydroxyalkanoate (PHA).
- a biopolymer such as a polyhydroxyalkanoate (PHA).
- transgenes encode enzymes such as beta- ketothiolase, acetoacetyl-CoA reductase, PHB ("short chain”) synthase, PHA ("long chain”) synthase, threonine dehydratase, dehydratases such as 3-OH acyl ACP, isomerases such as ⁇ 3-cis, ⁇ 2-trans isomerase, propionyl-CoA synthetase, hydroxyacyl-CoA synthetase, hydroxyacyl-CoA transferase, R-3- hydroxyacyl-ACP:CoA transferase, thioesterase, fatty acid synthesis enzymes and fatty acid beta-oxidation enzymes.
- enzymes such as beta- ketothiolase, acetoacetyl-CoA reductase, PHB (“short chain”) synthase, PHA ("long chain”) synthase, threonine dehydratase, dehydratases such as 3-
- PHA synthases include a synthase with medium chain length substrate specificity, such as phaCl from Pseudomonas oleovorans (WO 91/000917; Huisman, et al. J. Biol Chem. 266, 2191-2198 (1991)) or Pseudomonas aeruginosa (Timm, A. & Steinbuchel, A. Eur. J. Biochem. 209: 15-30 (1992)), the synthase from Alcaligenes eutrophus with short chain length specificity (Peoples, O. P. & Sinskey, A. J. J. Biol. Chem.
- medium chain length substrate specificity such as phaCl from Pseudomonas oleovorans (WO 91/000917; Huisman, et al. J. Biol Chem. 266, 2191-2198 (1991)) or Pseudomonas aeruginosa (Ti
- PHA synthase genes have been isolated from, for example, Alcaligenes latus (Accession ALU47026), Burkholderia sp.
- strain RA3849 accesion L37761
- Rhodospirillum rubrum U.S. Patent No. 5,849,894
- Rhodococcus ruber Rhodococcus ruber
- PHA synthases with broad substrate specificity useful for producing copolymers of 3-hydroxybutyrate and longer chain length (from 6 to 14 carbon atoms) hydroxyacids have also been isolated from Pseudomonas sp. A33 (Appl. Microbiol. Biotechnol. 42: 901-909 (1995)) and Pseudomonas sp. 61-3 (Accession AB014757; ato, et ah Appl. Microbiol. Biotechnol. 45: 363-370 (1996)) .
- An alpha subunit of beta-oxidation multienzyme complex pertains to a multifunctional enzyme that minimally possesses hydratase and
- a ⁇ subunit of the ⁇ - oxidation complex refers to a polypeptide capable of forming a
- ⁇ subunits possesses thiolase activity.
- FadA from E, coli (DiRusso, C. C. J. Bacteriol. 172: 6459-6468 (1990)
- FaoB from E, coli (DiRusso, C. C. J. Bacteriol. 172: 6459-6468 (1990)
- FaoB from E, coli (DiRusso, C. C. J. Bacteriol. 172: 6459-6468 (1990)
- FaoB from E, coli (DiRusso, C. C. J. Bacteriol. 172: 6459-6468 (1990)
- FaoB from E, coli (DiRusso, C. C. J. Bacteriol. 172: 6459-6468 (1990)
- FaoB from E, coli (DiRusso, C. C. J. Bacteriol. 172: 6459-6468 (1990)
- Pseudomonas fragi (Sato, S., Hayashi, M., Imamura, S., Ozeki, Y.,
- the transgene can encode a reductase.
- a reductase refers to an enzyme that can reduce ⁇ -ketoacyl Co As to R-3-OH-acyl Co As, such as the NADH dependent reductase from Chromatium vinosum (Liebergesell, M., & Steinbuchel, A. Eur. J. Biochem. 209: 135-150 (1992)), the NADPH dependent reductase from Alcaligenes eutrophus (Accession J04987, Peoples, O. P. & Sinskey, A. J. J. Biol. Chem. 264: 15293-15297 (1989))), the NADPH reductase from Zoogloea ramigera (Accession P23238;
- the transgene can encode a thiolase.
- a beta-ketothiolase refers to an enzyme that can catalyze the conversion of acetyl CoA and an acyl CoA to a p -ketoacyl CoA, a reaction that is reversible.
- An example of such thioiases are PhaA from Alcaligenes eutropus (Accession J04987, Peoples, O. P. & Sinskey, A. J. J. Biol. Chem. 264: 15293-15297 (1989)), BktB from
- Alcaligenes eutrophus (Slater et al. J Bacteriol 180(8): 1979-87 (1998)), and thioiases from the following Rhizobium meliloti (Accession RMU 17226), Z ramigera (Accession P07097), Paracoccus denitrificans (Accession D49362), Burkholderia sp. (Accession AFl 53086), Alcaligenes latus (Accession ALU47026), Allochromatium vinosum (Accession P45369), Thiocystis violacea (Accession P45363); Pseudomonas sp.
- strain 61-3 accesion ABO 14757
- acyl CoA oxidase refers to an enzyme capable of converting saturated acyl CoAs to ⁇ 2 unsaturated acyl CoAs.
- Examples of acyl CoA oxidases are POX1 from Saccharomyces cerevisiae (Dmochowska, et al. Gene. 1990, 88, 247-252) and ACX1 from Arabidopsis thaliana (Genbank Accession # AF057044).
- the transgene can also encode a catalase.
- a catalase refers to an enzyme capable of converting hydrogen peroxide to hydrogen and oxygen. Examples of catalases are KatB from Pseudomonas aeruginosa (Brown, et al J. Bacterid. 177: 6536-6544 (1995)) and KatG from E. coli (Triggs- Raine, B. L. & Loewen, P. C. Gene 52: 121-128 (1987)).
- the disclosed constructs and transgenic plants may also produce small inhibitory RNA molecules (siRNA) that can be single stranded or double stranded RNA molecules generally less than 200 nucleotides in length. Such molecules are generally less than 100 nucleotides and usually vary from 10 to 100 nucleotides in length.
- siRNA molecules have 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 or 30 nucleotides and bind to and inhibit translation of mRNA encoding one or more of the genes involved in production of
- siRNA means a small interfering RNA that is a short-length, preferably double-stranded RNA that is not toxic. Generally, there is no particular limitation in the length of siRNA as long as it does not show toxicity. "siRNAs” can be, for example, 15 to 49 bp, preferably 15 to 35 bp, and more preferably 21 to 30 bp long. Alternatively, the double-stranded RNA portion of a final transcription product of siRNA to be expressed can be, for example, 15 to 49 bp, preferably 15 to 35 bp, and more preferably 21 to 30 bp long.
- the double- stranded RNA portions of siRNAs in which two RNA strands pair up are not limited to the completely paired ones, and may contain nonpairing portions due to mismatch (the corresponding nucleotides are not complementary), bulge (lacking in the corresponding complementary nucleotide on one strand), and the like. Nonpairing portions can be contained to the extent that they do not interfere with siRNA formation.
- the "bulge” used herein preferably comprise 1 to 2 nonpairing nucleotides, and the double-stranded RNA region of siRNAs in which two RNA strands pair up contains preferably 1 to 7, more preferably 1 to 5 bulges.
- the "mismatch" used herein is contained in the double-stranded RNA region of siRNAs in which two RNA strands pair up, preferably 1 to 7, more preferably 1 to 5, in number.
- one of the nucleotides is guanine, and the other is uracil.
- Such a mismatch is due to a mutation from C to T, G to A, or mixtures thereof in DNA coding for sense RNA, but not particularly limited to them.
- the double-stranded RNA region of siRNAs in which two RNA strands pair up may contain both bulge and mismatched, which sum up to, preferably 1 to 7, more preferably 1 to 5 in number.
- the terminal structure of siRNA may be either blunt or cohesive (overhanging) as long as siRNA can silence, reduce, or inhibit the target gene expression due to its RNAi effect.
- the cohesive (overhanging) end structure is not limited only to the 3' overhang, and the 5' overhanging structure may be included as long as it is capable of inducing the RNAi effect.
- the number of overhanging nucleotide is not limited to the already reported 2 or 3, but can be any numbers as long as the overhang is capable of inducing the RNAi effect.
- the overhang consists of 1 to 8, preferably 2 to 4 nucleotides.
- the total length of siRNA having cohesive end structure is expressed as the sum of the length of the paired double-stranded portion and that of a pair comprising overhanging single-strands at both ends.
- the total length is expressed as 23 bp.
- this overhanging sequence since this overhanging sequence has low specificity to a target gene, it is not necessarily complementary (antisense) or identical (sense) to the target gene sequence.
- siRNA may contain a low molecular weight RNA (which may be a natural RNA molecule such as tRNA, rRNA or viral RNA, or an artificial RNA molecule), for example, in the overhanging portion at its one end.
- RNA which may be a natural RNA molecule such as tRNA, rRNA or viral RNA, or an artificial RNA molecule
- the terminal structure of the "siRNA” is not necessarily the cut off structure at both ends as described above, and may have a stem- loop structure in which ends of one side of double-stranded RNA are connected by a linker RNA.
- the length of the double-stranded RNA region can be, for example, 15 to 49 bp, preferably 15 to 35 bp, and more preferably 21 to 30 bp long.
- the length of the double- stranded RNA region that is a final transcription product of siRNAs to be expressed is, for example, 15 to 49 bp, preferably 15 to 35 bp, and more preferably 21 to 30 bp long.
- the length of the linker there is no particular limitation in the length of the linker as long as it has a length so as not to hinder the pairing of the stem portion.
- the linker portion may have a clover-leaf tRN A structure. Even though the linker has a length that hinders paking of the stem portion, it is possible, for example, to construct the linker portion to include introns so that the introns are excised during processing of precursor RNA into mature RNA, thereby allowing pairing of the stem portion.
- either end (head or tail) of RNA with no loop structure may have a low molecular weight RNA.
- this low molecular weight RNA may be a natural RNA molecule such as tRNA, rRNA or viral RNA, or an artificial RNA molecule.
- siRNA molecules can be achieved using conventional software for example at http://www.sirnawi2ard.com/. Because the nucleotide sequences of all of the genes involved in PHA production are known, one of skill in the art could input this sequence data into the siRNA software to design specific siRNA molecules that can be expressed by the transgenic plant to inhibit expression of one or more transgenes involved in PHA production.
- the disclosed constructs may contain a transgene expressing a PHA depolymerase.
- depolymerases There are two kinds of depolymerases, one that is used by micro-organisms to degrade polymer intraceilularly (intracellular depolymerases, and another that is secreted from the micro-organism to degrade extracellular polymer (extracellular depolymerases).
- extracellular depolymerases There are also depolymerases with specificity for short chain length polymers such as PHB (EC 3.1.1.75) and depolymerases with specificity for medium chain length polymers ( EC 3.1.1.76).
- Depolymerases suitable for this invention include but are not limited to the intracellular depolymerase PhaZ3 from Cupriavidus necator (formerly known as Ralstonia eutropha) (Accession AAP74581), the intracellular depolymerase PhaZ2 from Cupriavidus necator (Accession AAP74580), the intracellular depolymerase PhaZl from Ralstonia eutropha (Accession ABO 17612) (Saegusa, H., M. Shiraki, et at, 2001, J. Bacteriol. 183: 94-100; York, G. M. et al., 2003, J. Bacteriol. 185: 3788-3794), the extracellular depolymerase from Rhodospirillum rubrum (Accession AAP74581), the intracellular depolymerase PhaZ2 from Cupriavidus necator (Accession AAP74580), the intracellular depolymerase Pha
- the disclosed constructs may also contain a transgene encoding a 3- hydroxybutyrate dehydrogenase (EC 1.1.1.30). This enzyme catalyzes the conversion of 3-hydroxybutrate to acetoacetate ( Figure 3).
- Suitable 3- hydroxybutrate dehydrogenases include but are not limited to theD(-)-3- hydroxybutyrate dehydrogenase (hbdh) from Pseudomonas fragi (Accession AB183516), Bordetella pertussis (Accession BX640418), Ralstonia eutropha (Accession AF145230), Pseudomonas aeruginosa (Accession AE004626), Azospirillum brasilense (Accession AF355575), Caulobacter crescentus (Accession AE005999), Brucella melitensis (Accession AE009469), and Rhodobacter (Accession AF03
- the disclosed constructs may also contain expression cassettes for one or more transgenes encoding enzymes capable of increasing
- Candidate enzymes include but are not limited to sedoheptulose 1,7- bisphosphatase (SBPase, EC 3.1.3.37), fructose 1 ,6-bisphosphatase (FBPase, EC 3.1.3.11), a bi-functional enzyme encoding both SBPase and FBPase activities, transketolase (EC 2.2.1.1), and aldolase (EC 4.1.2.13). SBPase, transketolase, and aldolase activities have been shown to have an impact on the control of carbon fixed by the Calvin cycle (Raines, 2003, Photosynthesis Research, 75, 1-10) which could be attributed to an increase in ribulose 1 ,5- bisphosphate regenerative capacity.
- Bifunctional enzymes that contain both FBPase and SBPase activities have been reported from for example Ralstonia eutropha H16 (Accession number AAA69974), Synechococcus elongates PCC 7942 (Accession numbers D83512 and CP000100), Synechococcus sp.
- WH 7805 (Accession number ZP 01124026), B tyrivibrio crossotus DSM 2876 (Accession number EFF67670), Rothia m cilaginosa DY-18 (Accession number YP_003363264), Thiobacillus denitriflcans ATCC 25259 (Accession number AAZ98530), Methylacidiphilwn infernorum V4 (Accession number ACD83413), Nitrosomonas europaea ATCC 19718 (Accession number
- CAD84432 Vibrio vulnificus CMCP6 (Accession number AAO09802), and Methanohalophilus mahii DSM 5219 (Accession number YP_003542799).
- the FBPase/SBPase gene from Synechococcus elongatus PCC 7942 has previously been expressed in tobacco and enhanced both photosynthesis and plant growth (Miyagawa, 2001 f Nat. BiotechnoL, 19, 965-969).
- Enzymes possessing SBPase activity that could be used to increase the flow of carbon within the Calvin cycle include for example the sedoheptulose- 1,7-bisphosphatase from Zea mays (Accession
- NP_001148402 the sedoheptulose- 1,7-bisphosphatase from Arabidopsis th liana (Accession AAB33001), or the sedoheptulose-l,7-bisphosphatase from Triticum aestivum (Accession P46285).
- Enzymes possessing FBPase that could be used to increase the flow of carbon within the Calvin cycle include for example the protein encoded by the fopl gene from Synechococcus elongates PCC 6301 (Accession number AP008231.1), a D-fructose 1 ,6-bisphosphatase from Synechococcus elongatus PCC 7942 (Accession number CP000100), the gene encoding fructose- 1,6-bisphosphatase from Zea mays (Accession NP_001147459), the gene encoding fructose-1, 6-bisphosphatase from Saccharum hybrid cultivar H65-7052 (Accession CAA61409) and the fructose- 1,6-bisphosphatase from Pisum sativum (Accession AAD10213).
- Enzymes possessing transketolase activity that could be used to increase the flow of carbon within the Calvin cycle include for example the transketolase from Cyanobacterium UCYN-A (Accession YP_003421778), the transketolase from Spinacia oleracea (Accession AAD 10219), the transketolase from Rhodbacter capsulatus SB 1003 (Accession AAC32307), and the transketolase from Esherichia coli -12 MG1655 (Accession AAA69102).
- Enzymes possessing adolase activity that could be used to increase the flow of carbon within the Calvin cycle include for example the aldolase from Synechococcus sp. CC9902 (ACCESSION YP J78043) the ketose- bisphosphate aldolase from Crocosphaera watsonii WH 8501 (ACCESSION EAM501 8), the fructose-bisphosphate aldolase 1 from Rhodobacter sphaeroides (Accession number P27995), and the fructose-1 ,6- /sedoheptulose- 1 ,7-bisphosphate aldolase from Nitrobacter vulgaris
- Co-expression of RUBISCO with one or more of the above enzymes could further increase the rate of photosynthesis.
- Plant promoters can be selected to control the expression of the transgene in different plant tissues or organelles for all of which methods are known to those skilled in the art (Gasser & Fraley, Science 244:1293-99 (1989)).
- promoters are selected from those of eukaryotic or synthetic origin that are known to yield high levels of expression in plant and algae cytosol.
- promoters are selected from those of plant or prokaryotic origin that are known to yield high expression in plastids.
- the promoters are inducible. Inducible plant promoters are known in the art.
- Suitable constitutive promoters for nuclear-encoded expression include, for example, the core promoter of the Rsyn7 promoter and other constitutive promoters disclosed in U.S. Pat. No. 6,072,050; the core CAMV 35S promoter, (Odell et al. (1985) Nature 313:810-812); rice actin (McElroy et al. (1990) Plant Cell 2:163-171); ubiquitin (Christensen et al. (1989) Plant Mot. Biol 12:619-632 and Christensen et al. (1992) Plant Mot Biol 18:675- 689); pEMU (Last et al. (1991) Theor. Appl Genet.
- Tissue-preferred promoters can be used to target a gene expression within a particular tissue such as seed, leaf or root tissue.
- Tissue-preferred promoters include Yamamoto et al. (1997) Plant J. 12(2)255-265; Kawamata et al. (1997) Plant Cell Physiol 38(7):792-803; Hansen et al (1997) Mol Gen. Genet. 254(3):337-343; Russell et al. (1997) Transgenic Res. 6(2):157- 168; Rinehart et al. (1996) Plant Physiol. 112(3):1331-1341; Van Camp et al (1996) Plant Physiol. 112(2):525-535; Canevascini et al. (1996) Plant Physiol. 112(2):513-524; Yamamoto et al. (1994) Plant Cell Physiol.
- seed-preferred promoters include both “seed-specific” promoters (those promoters active during seed development such as promoters of seed storage proteins) as well as “seed-germinating” promoters (those promoters active during seed germination). See Thompson et al. (1989) BioEssays 10:108.
- seed-preferred promoters include, but are not limited to, Ciml (cytokinin-induced message); cZ19Bl (maize 19 kDa zein); milps (myoinositol-! -phosphate synthase); and eel A (cellulose synthase).
- Gama-zein is a preferred endosperm-specific promoter.
- Glob-1 is a preferred embryo- specific promoter.
- seed-specific promoters include, but are not limited to, bean ⁇ -phaseolin, napin ⁇ -conglycinin, soybean lectin, cruciferin, oleosin, the Lesquerella hydroxylase promoter, and the like.
- seed-specific promoters include, but are not limited to, maize 15 kDa zein, 22 kDa zein, 27 kDa zein, g-zein, waxy, shrunken 1, shrunken 2, globulin 1, etc. Additional seed specific promoters useful for practicing this invention are described in the Examples disclosed herein.
- Leaf-specific promoters are known in the art. See, for example, Yamamoto et al. (1997) Plant J. 12(2):255-265; Kwon et al. (1994) Plant Physiol. 105:357-67; Yamamoto et al. (1994) Plant Cell Physiol 35(5):773- 778; Gotor et al. (1993) Plant J. 3:509-18; Orozco et al. (1993) Plant MoL Biol 23(6): 1129-1138; and Matsuoka et al. (1993) Proc. Natl Acad. Set USA 90(20):9586-9590.
- Root-preferred promoters are known and may be selected from the many available from the literature or isolated de novo from various compatible species. See, for example, Hire et al. (1992) Plant Mol. Biol. 20(2): 207-218 (soybean root-specific glutamine synthetase gene); Keller and Baumgartner (1991) Plant Cell 3(10): 1051-1061 (root-specific control element in the GRP 1.8 gene of French bean); Sanger et al. (1990) Plant Mol. Biol. 14(3):433-443 (root-specific promoter of the mannopine synthase (MAS) gene of Agrobacterium tumefaciens); and Miao et al.
- MAS mannopine synthase
- Plastid specific promoters include the PrbcL promoter [Allison L. A. et al., EMBO 15: 2802-2809 (1996); Shiina T. et al., Plant Cell 10: 1713- 1722 (1998)3; the PpsbA promoter [Agrawal GK, et al., Nucleic Acids Research 29: 1835-1843 (2001)]; the Prrn 16 promoter [Svab Z & Maliga P., Proc. Natl. Acad. Sci.
- Chemical-regulated promoters can be used to modulate the rate of the enzyme
- the promoter may be a chemical-inducible promoter, where application of the chemical induces gene expression, or a chemical-repressible promoter, where application of the chemical represses gene expression.
- Chemical-inducible promoters are known in the art and include, but are not limited to, the maize ln2-2 promoter, which is activated by benzenesulfonamide herbicide safeners, the maize GST promoter, which is activated by hydrophobic electrophilic compounds that are used as pre-emergent herbicides, and the tobacco PR-1 a promoter, which is activated by salicylic acid.
- promoters of interest include steroid-responsive promoters (see, for example, the glucocorticoid-inducible promoter in Schena et al. Proc. Natl Acad Sci. USA 88:10421-10425 (1991) and cNellis etal Plant J. 14(2):247- 257(1998)) and tetracycline-inducible and tetracycline-repressible promoters (see, for example, Gatz etal Mol Gen. Genet 227:229-237 (1991), and U.S. Pat. Nos. 5,814,618 and 5,789,156), herein incorporated by reference in their entirety.
- coordinated expression of the three transgenes, phaA,phaB t and phaC, necessary for conversion of acetyl-CoA to PHB is controlled by a seed specific promoter, such as the soybean oleosin promoter (Rowley et al, Biochim Biophys Acta, 1997, 1345, 1-4) or the promoter from the lesquerlla hydroxylase gene (US Patent No. 6,437,220 Bl).
- a seed specific promoter such as the soybean oleosin promoter (Rowley et al, Biochim Biophys Acta, 1997, 1345, 1-4) or the promoter from the lesquerlla hydroxylase gene (US Patent No. 6,437,220 Bl).
- coordinated expression of the three transgenes, phaA,phaB, and phaC, necessary for conversion of acetyl-CoA to PHB is controlled by a promoter active primarily in the biomass plant, such as the maize chlorophyll A/B binding protein promoter (Sullivan et al, Mol Gen. Genet, 1989, 215, 431-40). It has been previously shown that plants transformed with multi- gene constructs produced higher levels of polymer than plants obtained from crossing single transgene lines (Valentin et al, Int. J. Biol Macromol, 1999, 25, 303-306; Bohmert et al, Planta, 2000, 211, 841-845).
- the final molecular weight of the polymer produced is controlled by the choice of promoter for expression of the PHA synthase gene. As described in US Patent No. 5,811,272, high PHA synthase activity will lower polymer molecular weight and low PHA synthase activity will increase polymer molecular weight.
- a strong promoter is used for expression of the genes encoding plastid-targeted monomer producing enzymes while a weaker promoter is used to control expression of synthase.
- polyadenylation signal can be engineered.
- a polyadenylation signal refers to any sequence that can result in polyadenylation of the mR A in the nucleus prior to export of the mRNA to the cytosol, such as the 3' region of nopaline synthase (Bevan, M, Barnes, W. M., Chilton, M. D. Nucleic Acids Res. 1983, 11, 369-385).
- Genetic constructs may encode a selectable marker to enable selection of plastid transformation events. There are many methods that have been described for the selection of transformed plants [for review see (Miki et ai, Journal of Biotechnology, 2004, 107, 193-232) and references incorporated within]. Selectable marker genes that have been used extensively in plants include the neomycin phosphotransferase gene nptll (U.S. Patent Nos. 5,034,322, U.S. 5,530,196), hygromycin resistance gene (U.S. Patent No. 5,668,298), the bar gene encoding resistance to
- aminoglycoside 3"-adenyltransferase (aadA) to confer spectinomycin resistance (U.S. Patent No. 5,073,675), the use of inhibition resistant 5- enolpyruvyl-3-phosphoshikimate synthetase (U.S. Patent No. 4,535,060) and methods for producing glyphosate tolerant plants (U.S. Patent No. 5,463,175; U.S. Patent No. 7,045,684).
- Methods of plant selection that do not use antibiotics or herbicides as a selective agent have been previously described and include expression of glucosamine-6-phosphate deaminase to inactive glucosamine in plant selection medium (U.S. Pat No.
- EP 0 530 129 A 1 describes a positive selection system which enables the transformed plants to outgrow the non-transformed lines by expressing a transgene encoding an enzyme that activates an inactive compound added to the growth media.
- U.S. Patent No. 5,767,378 describes the use of mannose or xylose for the positive selection of transgenic plants.
- Screenable marker genes include the beta-glucuronidase gene (Jefferson et al., 1987, EMBO J. 6: 3901-3907; U.S. Patent No. 5,268,463) and native or modified green fluorescent protein gene (Cubitt et al., 1995, Trends Biochem. Set 20: 448- 455; Pan et al., 1996, Plant Physiol. 112: 893-900).
- Transformation events can also be selected through visualization of fluorescent proteins such as the fluorescent proteins from the
- a preferred selectable marker is the spectinomycin-resistant allele of the plastid 16S ribosomal R A gene (Staub JM, Maliga P, Plant Cell 4: 39-45 (1992); Svab Z, Hajdukiewicz P, Maliga P, Proc. Natl. Acad Sci. USA 87: 8526-8530 (1990)).
- Selectable markers that have since been successfully used in plastid transformation include the bacterial aadA gene that encodes aminoglycoside 3'- adenyltransferase (AadA) conferring spectinomycin and streptomycin resistance (Svab et al, Proc. Natl. Acad. Sci.
- Plastid targeting sequences include the chloroplast small subunit of ribulose-1,5-bisphosphate carboxylase (Rubisco) (de Castro Silva Filho et al Plant Mol. Biol. 30:769-780 (1996); Schnell et al. J. Biol. Chem. 266(5):3335-3342 (1991)); 5-(enolpyruvyI)shikimate-3- phosphate synthase (EPSPS) (Archer et al J. Bioenerg. Biomemb.
- Rubisco chloroplast small subunit of ribulose-1,5-bisphosphate carboxylase
- EPSPS 5-(enolpyruvyI)shikimate-3- phosphate synthase
- Plants transformed in accordance with the present disclosure may be monocots or dicots.
- the transformation of suitable agronomic plant hosts using vectors for nuclear transformation or direct plastid transformation can be accomplished with a variet of methods and plant tissues.
- Representative plants useful in the methods disclosed herein include the Brassica family including B. napus, B. rapa, B. carinata and B. juncea; industrial oilseeds such as Camelina sativa, Crambe, jatropha, castor; Calendula, Cuphea, Arabidopsis thaliana; maize; soybean; cottonseed; sunflower; palm;
- coconut coconut; safflower; peanut; mustards including Sinapis alba; sugarcane flax and tobacco, also are useful with the methods disclosed herein.
- Representative tissues for transformation using these vectors include protoplasts, cells, callus tissue, leaf discs, pollen, and meristems.
- Transformation protocols as well as protocols for introducing nucleotide sequences into plants may vary depending on the type of plant or plant cell targeted for transformation. Suitable methods of introducing nucleotide sequences into plant cells and subsequent insertion into the plant genome include microinjection (Crossway et al (1986) Biotechniques 4:320- 334), electroporation (Riggs et al (1986) Proc. Natl Acad. Sci USA
- plastid transformation may be accomplished by transactivation of a silent plastid-borne transgene by tissue-preferred expression of a nuclear-encoded and plastid-directed RNA
- Plastid transformation vectors can be designed such that the transgenes are expressed from a promoter sequence that has been inserted with the transgene during the plastid transformation process or, alternatively, from an endogenous plastidial promoter such that an extension of an existing plastidial operon is achieved (Herz et al.
- Recombinase technologies which are useful for producing the disclosed transgenic plants include the cre-lox, FLP/FRT and Gin systems. Methods by which these technologies can be used for the purpose described herein are described for example in (U.S. Pat, No. 5,527,695; Dale And Ow, 1991, Proc. Natl. Acad. Sci. USA 88: 10558-10562; Medberry etal., 1995, Nucleic Acids Res. 23: 485-490).
- the following procedures can be used to obtain a transformed plant expressing the transgenes: select the plant cells that have been transformed on a selective medium; regenerate the plant cells that have been transformed to produce differentiated plants; select transformed plants expressing the transgene producing the desired level of desired polypeptide(s) in the desired tissue and cellular location.
- further rounds of regeneration of plants from explants of a transformed plant or tissue can be performed to increase the number of transgenic plastids such that the transformed plant reaches a state of homoplasmy (all plastids contain uniform plastomes containing transgene insert).
- the cells that have been transformed may be grown into plants in accordance with conventional techniques. See, for example, McCormick et at Plant Cell Reports 5:81-84(1986). These plants may then be grown, and either pollinated with the same transformed variety or different varieties, and the resulting hybrid having constitutive expression of the desired phenotypic characteristic identified. Two or more generations may be grown to ensure that constitutive expression of the desired phenotypic characteristic is stably maintained and inherited and then seeds harvested to ensure constitutive expression of the desired phenotypic characteristic has been achieved.
- Hybrid lines can be created by crossing a line containing one or more PHB genes with a line containing the other gene(s) needed to complete the PHB biosynthetic pathway.
- Use of lines that possess cytoplasmic male sterility (Esser, K. et al., 2006, Progress in Botany, Springer Berlin Heidelberg. 67, 31-52) with the appropriate maintainer and restorer lines allows these hybrid lines to be produced efficiently.
- Cytoplasmic male sterility systems are already available for some Brassicaceae species (Esser, K. et al., 2006, Progress in Botany, Springer Berlin Heidelberg. 67, 31-52). These Brassicaceae species can be used as gene sources to produce cytoplasmic male sterility systems for other oilseeds of interest such as Camelina.
- the expression of the PHB genes during germination could divert stored carbon to PHB instead of plant development
- Possible solutions to this include inhibiting expression of the PHB genes during germination and early plant development using additional transgene(s) encoding siRNA genes to inhibit expression of one or more of the PHB genes during germination and early development.
- An alternative solution is to use different seed specific promoters whose expression profile is high enough during seed development to achieve PHB levels of greater 8% but whose expression is low enough during germination and early seed development that the plant is not affected. These alternative promoters can be used to control the expression of one or more of the PHA biosynthetic genes. In some of our Examples described herein we have identified a series of promoters for this approach.
- Another possible scenario is that both the presence of PHB and /or expression of PHB genes during germination impairs photosynthesis during the critical stages of germination and early plantlet development resulting in failure of the seedlings to survive.
- the first two cotyledons of high PHB producers do become chlorotic or bleached .
- a possible solution to this would be to express additional transgenes encoding enzymes involved in the photosynthetic pathway to enhance photosynthetic flux of carbon.
- One example of such an enzyme is the cyanobacterial FBPase/SBPase.
- the disclosed genetic constructs can be used to produce industrial oilseed plants for high levels of PHA production. Specifically, PHA is produced in the seed.
- the transgenic plants can be grown and harvested.
- polyhydroxyalkanoate can be isolated from the oilseeds and the remaining plant material can be used as a feedstock for industrial use, preferably for the production of oleochemicals, energy or for use as feed for animals.
- the polyhydroxyalkanoate harvested from the plants can then be used to produce plastics, rubber material, coating material, and binders for paints, or as a feedstock for producing chemical derivatives such as hydroxyacids, esters, alkenoic acids or amines.
- PHA also has several medical applications.
- Vector pMBXS490 a pCAMBIA based plasmid (Centre for Application of Molecular Biology to International Agriculture, Canberra, Australia), contains the following gene expression cassettes: (1) an expression cassette for PHA synthase containing the promoter from the soybean oleosin isoform A gene, a DNA fragment encoding the signal peptide of the small subunit of rubisco from pea (P.
- DsRed a protein that can be visualized in seeds by placing them in light of the appropriate wavelength, containing the promoter from the cassava mosaic virus (CMV), a DNA fragment encoding a modified red fluorescent protein from Discosoma sp, (DsRed) in which eleven amino acids have been added to the C-terminus to increase solubility and/or prevent aggregation of the protein, and a termination sequence from the Agrobacterium tumefaciens nopaline synthase gene.
- CMV cassava mosaic virus
- Promoters are as follows: LH, promoter from the Lesqueretta fendleri Afunctional oleate 12-hydroxylase:saturate gene (US Patent No. 6,437,220 Bl); Oleosin, promoter from the soybean oleosin isoform A gene (Rowley and Herman, 1997, Biochim. Biophys. Acta 1345, 1-4); Napin, promoter from the Brassica napus napin gene (Ellenstrom, M. et al., 1996, Plant Molecular Biology, 32: 1019-1027); Glycinin, promoter from the soybean glycinin (gyl) gene (Iida, A. et al., 1995, Plant Cell Reports, 14,:539-544).
- Vectors pMBXS364, pMBXS355, pMBXS491, and pMBXS492 contain the same PHB pathway genes as pMBXS490 with the exception that the expression of these genes is under the control of different promoters as outlined in Table 1.
- Vector pMBXS355 contains an expression cassette for the bar gene, encoding phosphinothricin acetyltransferase whose expression is under the control of the 35S promoter. Expression of the bar gene allows selection of transformants based on their resistance to bialaphos. All other vectors in Table 1 contain expression cassettes for DsRed allowing the identification of transgenic seeds under the appropriate wavelength of light.
- Agrobacterium strain GV3101 was transformed with the construct of interest using electroporation
- a single colony of GV3101 containing the construct of interest was obtained from a freshly streaked plate and was inoculated into 5 mL LB medium. After overnight growth at 28°C, 2 mL of culture was transferred to a 500-mL flask containing 300 mL of LB and incubated overnight at 28°C.
- Agrobacterium inoculum contained in a 500-ml beaker. A vacuum (85 kPa) was applied and held for 5 min. Plants were removed from the desiccator and were covered with plastic bags in the dark for 24 h at room temperature. Plants were removed from the bags and returned to normal growth conditions within the greenhouse for seed formation.
- DsRed Camelina seeds expressing DsRed
- fully mature seeds were harvested from transformed plants and placed in a desiccator with anhydrous calcium sulfate as desiccant for at least 2 days prior to screening.
- DsRed expressing seeds were visualized in a darkroom with a green
- LumaMax LED flashlight (Lab Safety Supply, Lac, Janesville, WI) and a pair of KD's Dark Red glasses ( Pacific Coast Sunglasses Inc., Santa Maria, CA).
- bialaphos resistant seeds To identify bialaphos resistant seeds, seeds from floral dip transformations were sterilized in 70% ethanol and 10% bleach, and washed in water. Sterilized seeds were placed on germination and selection medium in square Petri dishes. The germination and selection medium contained 10 mg/L bialaphos (Gold BioTechnology, B0178-500) in 1/2X MS medium, which was made with Murashige & Skoog medium mixture (Caisson Labs, MSP09) at half concentration.
- T2 seeds were isolated containing 8% PHB in DsRed seeds picked from the segregating population. These seeds possessed a 75% germination rate and a 60% survival rate under greenhouse conditions in soil. The cotyledons after 11 days were chlorotic and the growth of this line was significantly delayed compared to wild-type.
- T2 seeds were isolated containing 6.9% PHB in DsRed seeds picked from the segregating population. These seeds possessed a 75% germination rate and a 70% survival rate under greenhouse conditions in soil. After 11 days, the cotyledons and first true leaves of this transformant were green. The growth of this line was somewhat delayed compared to wild-type but faster than the pMBXS491 line.
- DsRed as a visual marker in Camelina enabled the identification of high PHB producing seeds that would not have germinated in a typical seed screening procedure where an antibiotic or herbicide selectable marker, such as glyphosate resistance, is employed to provide resistance to the selection agent during seed germination and seedling development in tissue culture medium.
- an antibiotic or herbicide selectable marker such as glyphosate resistance
- Brassica carinata can be transformed using a previously described floral dip method (Shiv et al., 2008, Journal of Plant Biochemistry and Biotechnology 17, 1-4). Briefly constructs of interest are transformed into Agrobacterium strain GV-3101 and cells are grown in liquid medium. Cells are harvested and resuspended in a transformation medium consisting of 1 ⁇ 2 MS salts, 5% sucrose, and 0.05% Silwet L-77. Brassica carinata plants are grown in a greenhouse until inflorescences develop and approximately 25% of their flowers are opened. Plants are submerged in the prepared
- Agrobacterium solution for approximately 1 minute, and covered for 24 hours. Plants are returned to the greenhouse and allowed to set seed.
- Transformed seeds are screened by picking DsRed seeds under the appropriate wavelength of light as described above.
- Brassica seeds are surface sterilized in 10% commercial bleach (Javex, Colgate-Palmolive) for 30 min with gentle shaking. The seeds are washed three times in sterile distilled water and placed in germination medium comprising Murashige-Skoog (MS) salts and vitamins, 3% (w/v) sucrose and 0.7% (w/v) phytagar, pH 5.8 at a density of 20 per plate and maintained at 24°C an a 16 h light 8h dark photoperiod at a light intensity of 60-80 ⁇ -2 s "1 for 4-5 days.
- MS Murashige-Skoog
- Constructs of interest are introduced into Agrobacterium tumefacians strain EHA101 (Hood et. al., 1986, J. Bacterid. 168: 1291-1301) by electroporation. Prior to transformation of cotyledonary petioles, single colonies of strain EHA101 harboring each construct are grown in 5 ml of minimal medium supplemented with appropriate antibiotics for 48 hr at 28°C. One ml of bacterial suspension was pelleted by centrifugation for 1 min in a microfuge. The pellet was resuspended in 1 ml minimal medium.
- cotyledons are excised from 4 or in some cases 5 day old seedlings so that they included -2 mm of petiole at the base.
- Individual cotyledons with the cut surface of their petioles are immersed in diluted bacterial suspension for 1 s and immediately embedded to a depth of ⁇ 2mm in co-cultivation medium, MS medium with 3% (w/v) sucrose and 0.7% phytagar and enriched with 20 ⁇ benzyladenine.
- the inoculated cotyledons are plated at a density of 10 per plate and incubated under the same growth conditions for 48 h.
- the cotyledons are transferred to regeneration medium comprising MS medium supplemented with 3% sucrose, 20 ⁇ benzyladenine, 0.7% (w/v) phytagar, pH 5.8, 300 mg/L timentinin and 20 mg/L kanamycin sulfate.
- “shoot elongation” medium (MS medium containing, 3% sucrose, 300mg/L timentin, 0.7% (w/v) phytagar, 300 mg/L timentinin and 20 mg/L kanamycin sulfate, pH 5.8) in Magenta jars.
- the elongated shoots are transferred to "rooting" medium comprising MS medium, 3% sucrose, 2mg/L indole butyric acid, 0.7% phytagar and 500mg/L carbenicillin.
- plantlets are transferred to potting mix (Redi Earth, W.R. Grace and Co.). The plants are maintained in a misting chamber (75% relative humidity) under the same growth conditions. Plants are allowed to self pollinate to produce seeds. Seeds are screened by visualization of DsRed as described above.
- Brassica napus can also be transformed using the floral dip procedure described by Shiv et al. (Shiv et al., 2008, Journal of Plant Biochemistry and Biotechnology 17, 1-4) as described above for Brassica carinata.
- Brassica juncea can be transformed using hypocotyl explants according to the methods described by Barfield and Pua (Barfield and Pua, Plant Cell Reports, 10, 308-314) or Pandian et al. (Pandian, et al., 2006, Plant Molecular Biology Reporter 24: 103a-103i) as follows.
- B. juncea seeds are sterilized 2 min in 70% (v/v) ethanol and washed for 20 min in 25% commercial bleach (10 g/L hypochlorite). Seeds are rinsed 3X in sterile water. Surface-sterilized seeds are plated on germination medium (lx MS salts, lx MS vitamins, 30 g L sucrose, 500 mg L MES. pH 5.5) and kept in the cold room for 2 days. Seeds are incubated for 4-6 days at 24°C under low light (20 ⁇ m ' V 1 ).
- hypocotyl segments are excised and rinsed in 50 mL of callus induction medium (lx MS salts, Ix B5 vitamins, 30 g L sucrose, 500 mg/L MES, 1.0 mg/L 2,4-D, 1.0 mg/L kinetin pH 5.8) for 30 min without agitation. This procedure is repeated but with agitation on orbital shaker ( ⁇ 140 g) for 48 h at 24°C in low light (10 ⁇ rn V 1 ).
- Agrobacterium can be prepared as follows: Cells of Agrobacterium strain AGL1 (Lazo, G. et al. (1991),Biotechnology, 9: 963-967) containing the construct of interest are grown in 5 mL of LB medium with appropriate antibiotic at 28°C for 2 days. The 5 mL culture is transferred to 250 mL flask with 45 mL of LB and cultured for 4 h at 28°C. Cells is pelleted and resuspended in BM medium (l MS salts, lx B5 vitamins, 30 g L sucrose, 500 mg/L MES, pH 5.8). The optical density at 600 nm is adjusted to 0.2 with BM medium and used for inoculation.
- Explants are cocultivated with Agrobacterium for 20 min after which time the Agrobacterium suspension is removed. Hypocotyl explants are washed once in callus induction medium after which cocultivation proceeds for 48 h with gentle shaking on orbital shaker. After several washes in C1M, explants are transferred to selective shoot-inducing medium (500 mg/L AgN02, 0.4 mg/L zeatin riboside, 2.0 mg/L benzylamino purine, 0.01 mg/L GA, 200 mg/L Timentin appropriate selection agent and 8 gL agar added to basal medium) plates for regeneration at 24°C.
- selective shoot-inducing medium 500 mg/L AgN02, 0.4 mg/L zeatin riboside, 2.0 mg/L benzylamino purine, 0.01 mg/L GA, 200 mg/L Timentin appropriate selection agent and 8 gL agar added to basal medium
- Root formation is induced on root-inducing medium (0.5x MS salts, 0.5x B5 vitamins, 10 g/L sucrose, 500 mg L MES, 0.1 mg/L indole-3 -butyric acid, 200 mg/L Timentin, appropriate selection agent and 8 g/L agar, pH 5.8).
- root-inducing medium 0.5x MS salts, 0.5x B5 vitamins, 10 g/L sucrose, 500 mg L MES, 0.1 mg/L indole-3 -butyric acid, 200 mg/L Timentin, appropriate selection agent and 8 g/L agar, pH 5.8.
- Plantlets are transferred to or removed from agar, gently washed, and transferred to potting soil in pots. Plants are grown in a humid environment for a week and then transferred to the greenhouse.
- RNA interference R Ai constructs
- the RNAi element was targeted to either synthase, thiolase, or reductase.
- the RNAi element was designed with an intron between an inverted repeat of the stretch of the gene targeted for RNAi interference. Expression of the RNAi element was controlled by a chemically inducible promoter.
- Table 3 All constructs for RNAi interference contain the PHB expression cassettes and DsRed expression cassette of pMBXS490.
- Plasmid phaA-RNAi/35S contains the following expression cassettes for inducible expression of the RNAi element with homology to a stretch of the phaA gene: (1) an expression cassette for a chimeric ecdysone receptor consisting of the double enhanced version of the 35S promoter from cauliflower mosaic virus , the grvH gene encoding a chimeric ecdysone receptor that contains a DNA-binding domain derived from the human glucocorticoid receptor, the transcriptional activation domain from the Herpes simplex viral protein VP16, and the ligand-binding domain from the ecdysone receptor of Heliothis virescens, and the 3' termination sequence of the nopaline synthase gene from Agrobacterium tumefaciens; (2) an expression cassette for the RNAi element consisting of a DN A fragment encoding six copies of glucocorticoid response element (GRE) derived from the promoter region of mouse mammary
- RNAi Ribonucleic acid
- rbcs rib-1,5-bisphospate carboxylase
- Seeds were germinated and plants were grown in a greenhouse and treated with methoxyfenozide inducing agent during flowering and seed formation. A portion of the seed was used for analysis of PHB. Seeds containing 10% PHB were isolated ( Figure 2). T2 seeds were placed on a piece of filter paper and soaked in inducing agent prior to transfer to soil.
- T2 seeds from the above transformations were germinated and grown in soil in a greenhouse producing T2 seedlings. Untreated T2 plants were allowed to set seed and T3 seeds from select lines were collected and the polymer content was measured using the previously described gas chromatography butanolysis procedures. Several lines producing greater than 7% dwt PHB in both the T2 and T3 generations were obtained ( Figure 3). No difference between plants treated with inducing agent or treated with water was observed. This suggests that the inducible promoter element is not controllable under the conditions used for the experiments but that there is some basal level of expression from the minimal promoter i front of the RNAi element
- the lighting program used in the HID chamber was as follows: 6 am to 7 am, 300 microMoles m'V 1 ; 7 am to 8 am, 750 microMoles m "2 s -1 ; 8 am to 3 pm, 1250 microMoles m "2 s -1 ; 3 pm to 5 pm, ramp down from 1250 to
- RNAi lines tested possessed greater than 50% survival, with some as high as 85-95%.
- High PHB containing seeds can be screened for germination ability prior to planting in soil by plating the seeds on wet filter paper to determine if they germinate. If seeds are impaired in germination or possess chlorotic seedlings, this filter paper can be transferred to tissue culture medium containing 1/2X MS agar medium (prepared from Murashige & Skoog salts with vitamins, Caisson Labs, MSP09) supplemented with 2% sucrose,
- Example 6 Managing gene expression during germination, controlled polymer degradation during germination.
- constructs were designed containing genes encoding a pathway for controlled polymer degradation during seed germination. PHB production would proceed during seed formation and polymer degradation would occur during seed germination ( Figure 3). Genes encoding PHA depolymerase and 3- hydroxybutyrate dehydrogenase were chosen for degradation of polymer. These genes are expected to convert PHB to 3 -hydroxybutyrate and 3- hydroxybutryate to acetoacetate, compounds that could be further metabolized by the germinating seedling. Since construct pMBXS490 enabled high PHB production, albeit with poor germination/seedling survival, it was used as a starting plasmid to build future transformation constructs.
- Plant transformation construct pMBXVTl is a pCAMBIA based vector containing seed specific expression of PHA genes and cassettes for expression of the depolymerase and 3 -hydroxybutyrate dehydrogenase under the control of germination specific promoters. Expression cassettes for the PHB biosynthetic genes and DsRed are as described for pMBXS490.
- Additional expression cassettes in pMBXVTl are as follows: 1) an expression cassette for depolymerase containing the promoter from Vig a mtmgo sulphydryl-endopeptidase gene (SH-EP promoter; Akasofu et al. t 1990 Nucleic Acids Research. 18, 1892), a DNA fragment encoding the signal peptide and the first 24 amino acids of the mature protein of the small subunit of rubisco from pea, a DNA fragment encoding an intracellular polyhydroxybutyrate depolymerase (PhaZal) from Ralstonia eutropha (Saegusa et al., 2001, J. Bacterid.
- PVZal polyhydroxybutyrate depolymerase
- Construct pMBXVTl was transformed into Camelina as previously described and Ti seeds were selected by visualization of DsRed. Ti seeds were either planted directly into soil or germinated on filter paper and transplanted into soil. The resulting T2 seeds were tested for PHB using the previously described gas chromatography butanolysis techniques. T2 seeds containing up to 11.3 % PHB were isolated (Table 5) however these seeds produced seedlings that did not survive in soil conditions. Germination of T2 seeds on filter paper was measured and the percent survival was calculated. One line containing 5.75% PHB with 100% survival in soil was isolated. Lines that possessed severely impaired germination in soil or on filter paper (i.e. line containing 11.3% PHB) were rescued by germination on tissue culture medium as follows.
- Seeds were surface sterilized with 70% alcohol for 2 minutes and with 10% commercial bleach for 10 minutes. The seeds were washed thoroughly at least 3 times with sterile water before transferring them on to agar plates. Seeds were cold treated at 4°C by plating them on agar media containing 1 ⁇ 2 strength Murasbige and Skoog basal salts and Gamborg's vitamins (Sigma Chemical Company, St. Louis, MO) supplemented with 2% sucrose. Plates were incubated at 4°C for 72 hours and then transferred to a tissue culture chamber set at 20°C. Seedlings were transferred to soil once they had obtained true leaves and were then transferred to the greenhouse. T3 seeds were generated from the T 2 lines and evaluated for PHB content. A graph comparing T2 and T3 seeds from select lines is shown in Figure 4.
- pMBXS490 for the PHB biosynthetic pathway and DsRed genes as well as inducible expression cassettes for PHB depolymerase and 3-hydroxybutyrate dehydrogenase.
- the inducible expression cassettes rely on the binding of a chimeric receptor (VP16:GAL4:CJEcR gene), whose expression is under the control of a constitutive promoter, to the inducing agent and response element ( Figure 1 ).
- the chimeric receptor contains a transcriptional activation domain from Herpes simplex viral protein (VP1 AD), a binding domain from yeast GAL4 transcription activator (GAL4 DBD), and a ligand binding domain from the Choristone ra fumiferana ecdysone receptor (CfEcR). This binding initiates transcription of the PHB depolymerase and 3-hydroxybutyrate dehydrogenase genes placed behind a DNA sequence containing a minimal 35S promoter with five copies of the 19 bp yeast GAL4 response elements upstream of the minimal promoter for chemical induction.
- VP1 AD Herpes simplex viral protein
- GAL4 DBD yeast GAL4 transcription activator
- CfEcR Choristone ra fumiferana ecdysone receptor
- the chimeric receptor protein Upon addition of a chemical inducing agent, the chimeric receptor protein transactivates expression of the target gene(s) cloned under the control of the GAL4 response elements and the minimal promoter.
- a chemical inducing agent Upon addition of a chemical inducing agent, the chimeric receptor protein transactivates expression of the target gene(s) cloned under the control of the GAL4 response elements and the minimal promoter.
- Four separate constructs were constructed that differ in the length of their minimal promoter sequence and/or the promoter that drives the expression of the chimeric receptor (Table 6).
- inducing agent was expected to yield good expression of the PHB depolymerase and 3-hydroxybutyrate dehydrogenase at the growth stage in which the inducing agent was applied. In the absence of inducing agent, a basal level of expression due to the leakiness of the promoter was expected.
- Constructs were transformed into Camelina, using the transformation methods described above, and the chemical inducing agent was applied from flowering to harvest of the Ti seeds.
- the chemical inducing agent used for this purpose was methoxyfenozide applied to the plants in the form of the commercial pesticide Intrepid (Dow AgroSciences, Indianapolis, IN).
- Concentrations for application ranged from 66 to 100 uM.
- Intrepid was also applied during germination of Tj seeds, and again from flowering to harvest of the T2 seeds. The T 2 seeds were then split into two groups. The first received no inducing agent (allowing the accumulation of PHB in the seeds). The other was treated with the inducing agent to limit PHB accumulation in the seeds, possibly improving seed germination. No significant difference in the levels of PHB in seeds that had been treated with Intrepid during flowering and seed development were observed compared to controls.
- T2 seedlings The survival of T2 seedlings was determined by germinating seeds on filter paper and then transferring seedlings to soil (Table 7) T seeds with poor germination were rescued by germinating on 1 ⁇ 2 strength Murashige and Skoog basal salts with Gamborg's vitamins supplemented with 2% sucrose as described above. Lines were grown in the greenhouse to produce T 3 seeds.
- T2 seeds from these lines had in general better germination and seedling viability than seeds obtained from transformations with plasmid pMBXS490, leaky expression from the inducible promoter controlling the expression of depolymerase and 3-hydroxybutyrate dehydrogenase may have occurred such that sufficient amounts of these enzymes are produced to increase germination and seedling viability of high PHB producing seeds without significantly compromising PHB yield.
- T 2 seeds that were unable to germinate and survive on filter paper were rescued by germinating on 1 ⁇ 2 strength Murashige and Skoog basal salts with Gamborg's vitamins supplemented with 2% sucrose and 15 ⁇ methoxyfenozide as described above. All lines were grown in the greenhouse to produce T 3 seeds.
- High PHB containing seeds can be screened for germination ability by plating the seeds on wet filter paper to determine if they germ inate. If seeds are impaired in germination or possess chlorotic seedlings, this filter paper can be transferred to tissue culture medium containing 1/2X MS agar medium (prepared from Murashige & Skoog salts with vitamins, Caisson Labs, MSP09) supplemented with 2% sucrose,
- Example 7 Expression of Depolymerase and 3-Hydroxybutyrate Dehydrogenase using a Heat Shock Promoter.
- Plasm d pMBXS430 was prepared to test the use of a heat shock inducible promoter to control expression of depolymerase and 3- hydroxybutyrate dehydrogenase genes.
- This plasmid is the same as pMBXVTl with the exception that the germination specific promoter controlling the expression of depolymerase and 3-hydroxybutyrate dehydrogenase genes has been replaced by a heat shock inducible promoter from the soybean small heat shock (Gmhspl7.5E) gene (Czarnecka, E. et al., 1989, MoL Cell Biol. 9, 3457-3463).
- Plasmid pMBXS430 was transformed into Camelina according to the methods described above and seeds were screened for DsRed expression. Isolated T 1 seeds were germinated on 1/2X MS agar medium (Murashige & Skoog salts with vitamins, Caisson Labs, MSP09) supplemented with 2% sucrose, transferred to soil in the
- Example 8 Production of hybrid lines that are not capable of germinating.
- lower levels of PHB were obtained when lines expressing individual PHB genes were crossed to produce a plant containing the entire PHB biosynthetic pathway (Nawrath, C, Y. Poirier, et ai.,1994, Proc. Natl. Acad. Sci. USA 91, 12760-12764) than when multi-gene constructs containing the entire PHB biosynthetic pathway were constructed and transformed (Bohmert, K., I. et al., 2000, Planta 211, 841-845;US Patent 6,448,473). This observation led to the subsequent predominant use of multi-gene constructs for PHB production in plants.
- Hybrid lines can be created by crossing a line containing one or more PHB genes with a line containing the other gene(s) needed to complete the PHB biosynthethic pathway.
- Cytoplasmic male sterility systems are already available for some Brassicaceae species (Esser, . et al., 2006, Progress in Botany, Springer Berlin Heidelberg. 67, 31 -52). These Brassicaceae species can be used as gene sources to produce cytoplasmic male sterility systems for other oilseeds of interest such as Camelina. Cytoplasmic male sterility has also been reported upon expression of a ⁇ -ketothiolase from the chloroplast genome in tobacco (Ruiz, O. N. and H. Daniell, 2005, Plant Physiol. 138, 1232-1246). Male sterility has also been reported upon expression of the faoA gene encoding the a-subunit of the fatty acid ⁇ - oxidation complex from Pseudomonas putida (US Patent 6586658).
- High PHB producing lines that are not capable of germination can be produced using oilseed lines that possess cytoplasmic male sterility (CMS) controlled by an extranuclear genome (i.e. mitochondria or chloroplast).
- CMS cytoplasmic male sterility
- the male sterile line is typically maintained by crossing with a maintainer line that is genetically identical except that it possesses normal fertile cytoplasm and is therefore male fertile. Transformation of the maintainer line with one or more genes for the PHB biosynthetic pathway and crossing this modified maintainer line [Figure 5, M line (phaA and phaC ] with the original male sterile line [Figure 5, S line (CMS)] will produce a male sterile line possessing a portion of the PHB biosynthetic pathway.
- Fertility can be restored to this line using a "restorer line” that carries the appropriate nuclear restorer genes.
- the restorer line can be transformed with the remaining genes required to complete the PHB biosynthetic pathway [Figure 5, R line (phaB)] and crossed with the previously created male sterile line containing phaA and phaC [Figure 5, S line (phaA and phaC)] to produce a hybrid line containing the entire PHB biosynthetic pathway [ Figure 5, Hybrid seeds (phaA, phaB, and phaC) ⁇ .
- Crosses can be performed in the field by planting multiple rows of the male sterile line, the line that will produce the seed, next to a few rows of the male fertile line.
- Harvested seed can be used for subsequent plantings or as the PHB containing seed for crushing and extraction.
- expression cassettes for the PHB genes in tins example are controlled by strong promoters, such as the soybean oleosin promoter, high PHB producing seeds generated in this manner will possess weak seedlings upon germination and will not be able to survive field conditions under normal growth
- Cytoplasmic male sterility systems are already available for some Brassicacea species (Esser, ., 2006, Progress in Botany, Springer Berlin Heidelberg. 67, 31-52). These Brassicaceae species can be used as gene sources to produce cytoplasmic male sterility systems for other oilseeds of interest such as Camelina. Cytoplasmic male sterility has also been reported upon expression of a p-ketothiolase from the chloroplast genome in tobacco (Ruiz, O. N. and H. Daniell, 2005, Plant Physiol. 138, 1232-1246). Overexpression of ⁇ -ketothiolase in Camelinato generate a male sterile line and subsequent crossing with a line expressing phaB and phaC could also be used for hybrid seed production.
- Double haploid technology can be used to speed up the breeding process.
- immature pollen grains haploids
- haploids immature pollen grains
- Example 9 Improved germination efficiency of high PHB producing seeds using promoters that are not active or minimally active daring seed germination and seedling development
- each promoter was put in an expression cassette with the reporter gene ⁇ -glucuronidase (GUS). Seedlings were germinated and seedlings were stained with X-Gluc (5-bromo-4-chloro-3-indolyl beta-D- glucuronide). GUS expression was observed with all seed specific promoters tested in germinating seedlings (Table 8).
- promoters from the lesquerella hydroxylase gene, the napin gene, and the glycinin gene yielded GUS staining in their first true leaves.
- Table 8 GUS expression patterns of seed specific promoters during seed formation and germination.
- Promoters are as follows: 35S, promoter from the cauliflower mosaic virus 35S gene; LH, promoter from the Lesquerella fendleri bifunctional oleate 12- hydroxylase:saturate gene ; Oleosin, promoter from the soybean oleosin isoform A gene; P3, promoter from a seed specific gene in Arabidopsis 10 thaliana (US patent 7405345); Napin, promoter from the Brassica napus
- Glycinin promoter from the soybean glycinin (gyl) gene.
- Reference profile 1 20 was set to be highly expressed at the 24-h post-pollination seed
- Reference profiles 2 and 3 were set to be highly expressed in both the globular-stage and cotyledon-stage seed, since these stages are developmental ⁇ close and were identified to exhibit similar expression patterns.
- Reference profiles 4 and 5 were also set to be highly expressed in both the mature-green-stage and postenature-green-stage seed. All non-seed stages, including the unfertilized ovule, seedling, leaf, root, stem, and floral buds were set to zero.
- Table 9 Predefined search profiles to identify genes with similar expression patterns.
- OV unfertilized ovule
- 24H 24-h postpollination seed
- GLOB globular-stage seed
- COT cotyledon-stage seed
- MG mature-green-stage seed
- PMG postmature-green-stage seed
- SDLG seedling
- L leaf
- R root
- S stem
- F floral buds.
- Hierarchical clustering analysis identified several genes which showed similar expression patterns as the five reference profiles. Genes with expression values in non-seed stages were removed from the set of identified genes. 81 genes whose promoter region may be suitable for PHB production in seeds with little to no PHB gene expression in seedlings were identified (Table 10).
- genes were selected that exhibited different temporal profiles, le. were highest expressed in a particular seed development stage; (2) genes with medium and high expression levels were chosen and genes with low expression levels were omitted; and (3) preference was given to genes whose function was established. These criteria resulted in the selection of 17 genes, three of which appear to encode isoenzymes due to their high sequence homology (Table 11). Use of the promoters from these genes may lead to seeds with high PHB content and high germination/survival.
- suitable promoters may be identified by modifying the predefined search profiles described in Table 9.
- Example 10 Increasing Flux through the Calvin cycle: Design and construction of transformation vectors expressing a gene encoding FBPaseSBPase with genes encoding the PHB biosynthetic enzymes in oilseeds.
- Transformation vectors pMBXS407 and pMBXS408 were prepared that contain title expression cassettes for plastid targeted PHB enzymes from plasmid pMBXS490 and an additional cassette for expression of a
- FBPase/SBPase gene under the control of the 35S promoter from the cauliflower mosaic virus.
- Two different sequences for FBPase/SBPase gene from Synechococcus elongatus PCC 7942 are listed in the NCBI database, accession numbers D83512 and CP000100. These two sequences differ at amino acidsl45 to 148 and at their C-terminus ( Figure 6). Transformation vectors pMBXS407 and pMBXS408 were constructed in which the
- FBPase/SBPase genes were fused at the 5' end to a DNA sequence encoding a signal peptide of the small subunit of pea and the first 24 amino acids of the mature protein [Cashmore, A. R. (1983). Nuclear Genes Encoding the Small Subunit of Ribulose-1,5-Bisphosphate Carboxylase. Genetic
- Transformation vector pMBXS407 contains a gene encoding a
- FBPase/SBPase with 100% homology to the FBPase SBPase protein from Synechococcus elongatus PCC 7942 listed in accession CP000100.
- Transformation vector pMBXS408 contains a gene encoding a
- FBPase/SBPase with 100% homology to the FBPase/SBPase protein from Synechococcus elongatus PCC 7942 listed in accession D83512. Even though this gene is listed in accession D83512 as a fructose- 1,6- bisphosphatase-I gene, the presence of both FBPase and SBPase activities in the encoded protein has been verified enzymatically (Tamoi, M, et al., 1996, Archives of Biochemistry and Biophysics, 334, 27-36).
- Transformation vectors pMBXS407 and pMBXS408 were transformed into Camelina and Tl seeds were isolated based on DsRed expression. Tl lines were further propagated and second generation (T2) transgenic seeds were produced. The highest PHB producing lines (i.e. greater than 10% PHB) were generated by germination of seeds in tissue culture medium containing 2 % sucrose. The base tissue culture medium was 1/2 x MS agar medium made with Murashige and Skoog medium mixture [Caisson Labs]. Further propagation yielded T3 transgenic seeds that produced PHB at levels up to 13% of the seed weight. Select lines were used in germination trials under controlled greenhouse conditions (Table 12).
- seedlings generated from the pMBXS407 transformations possessed healthier seedlings and with greater survival rates than seedlings generated from pMBXS408 or pMBXS490 transformations.
- transgenic seedlings from the pMBXS407 transformation showed significant increases in growth and biomass production when compared to transgenic seedlings transformed with pMBXS408 and pMBXS490 transformed plants. This increased growth and biomass production persisted through growth of the plants to maturity.
- the change in shoot biomass in the transgenic plants that may be due to overexpression of the FBPase/SBPase gene in pMBXS407 was correlated to both an increase in stem diameter and leaf surface area.
- transformation vector pMBXSSl 1 was prepared. This vector contains the PHB gene and DsRed expression cassettes in pMBXS490 and an additional cassette for expression of the Synechococcus elongatus PCC 7942 FBPase SBPase gene listed in accession gb
- pMBXS511 the plastid targeting sequence from pea including the first 24 amino acids of the mature protein is attached to the 5' end of the
- FBPase/SBPase to direct the import of the protein into the plastids.
Landscapes
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Cell Biology (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Plant Pathology (AREA)
- Medicinal Chemistry (AREA)
- Pharmacology & Pharmacy (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
Description



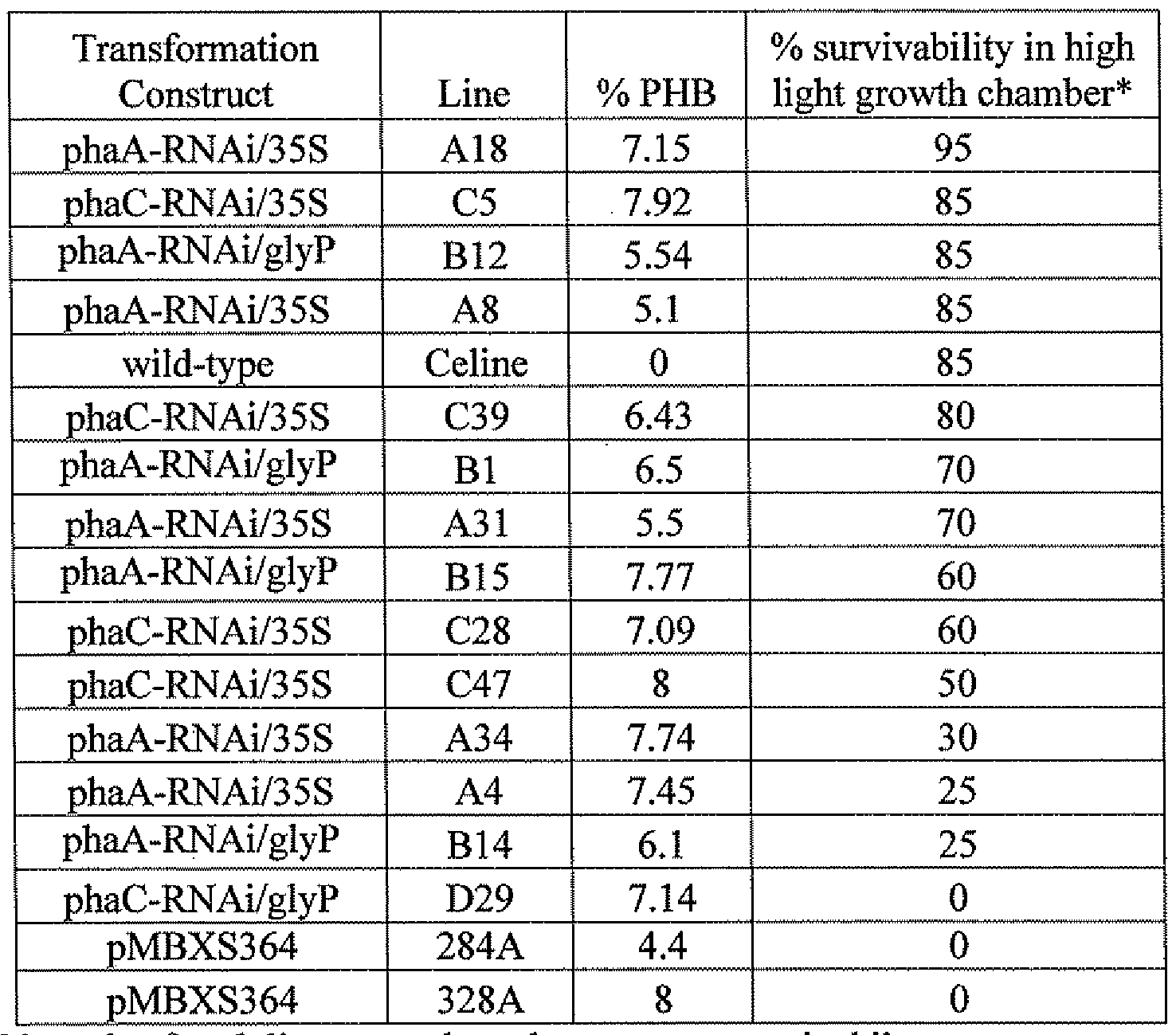



































































Claims
Priority Applications (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP10757363A EP2478105A1 (en) | 2009-09-15 | 2010-09-15 | Generation of high polyhydroxybutrate producing oilseeds |
| CA2773707A CA2773707A1 (en) | 2009-09-15 | 2010-09-15 | Generation of high polyhydroxybutrate producing oilseeds with improved germination and seedling establishment |
| AU2010295638A AU2010295638B2 (en) | 2009-09-15 | 2010-09-15 | Generation of high polyhydroxybutrate producing oilseeds |
| BRBR112012005591-7A BR112012005591A2 (en) | 2009-09-15 | 2010-09-15 | Generation of oilseeds producing high polyhydroxybutyrate. |
| US13/395,702 US9181559B2 (en) | 2009-09-15 | 2010-09-15 | Generation of high polyhydroxybutyrate producing oilseeds |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US24252209P | 2009-09-15 | 2009-09-15 | |
| US61/242,522 | 2009-09-15 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| WO2011034946A1 true WO2011034946A1 (en) | 2011-03-24 |
| WO2011034946A9 WO2011034946A9 (en) | 2011-05-19 |
Family
ID=43063232
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US2010/048962 Ceased WO2011034945A1 (en) | 2009-09-15 | 2010-09-15 | Generation of high polyhydroxybutrate producing oilseeds |
| PCT/US2010/048963 Ceased WO2011034946A1 (en) | 2009-09-15 | 2010-09-15 | Generation of high polyhydroxybutrate producing oilseeds |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US2010/048962 Ceased WO2011034945A1 (en) | 2009-09-15 | 2010-09-15 | Generation of high polyhydroxybutrate producing oilseeds |
Country Status (6)
| Country | Link |
|---|---|
| US (2) | US9181559B2 (en) |
| EP (2) | EP2477477A1 (en) |
| AU (1) | AU2010295637B2 (en) |
| BR (2) | BR112012005592A2 (en) |
| CA (2) | CA2773707A1 (en) |
| WO (2) | WO2011034945A1 (en) |
Cited By (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2012037324A3 (en) * | 2010-09-15 | 2012-05-18 | Metabolix, Inc. | Increasing carbon flow for polyhydroxybutyrate production in biomass crops |
| CN102864167A (en) * | 2012-09-26 | 2013-01-09 | 浙江大学 | Plant expression carrier and method for cultivating low-phytic-acid rice |
| WO2014100289A1 (en) | 2012-12-18 | 2014-06-26 | Metabolix, Inc. | Transcriptional regulation for improved plant productivity |
| WO2014127053A2 (en) | 2013-02-13 | 2014-08-21 | Metabolix, Inc. | Process for ultra pure chemical production from biobased raw starting materials |
| WO2014133159A1 (en) * | 2013-02-28 | 2014-09-04 | 株式会社ユーグレナ | Method for introducing gene to euglena, and transformant therefrom |
| JP2014193153A (en) * | 2013-02-28 | 2014-10-09 | Euglena Co Ltd | Method for introducing gene into euglena |
| JP2014193154A (en) * | 2013-02-28 | 2014-10-09 | Euglena Co Ltd | Transformant of euglena |
| US10316330B2 (en) | 2012-05-08 | 2019-06-11 | Monsanto Technology Llc | Corn event MON 87411 |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| MY185155A (en) | 2012-05-31 | 2021-04-30 | Micromidas Inc | Polyhydroxyalkanoate derivatives, preparation and uses thereof |
| AU2014278519B2 (en) * | 2013-06-11 | 2020-09-10 | Syngenta Crop Protection Ag | Methods for generating transgenic plants |
| CN103329793A (en) * | 2013-06-25 | 2013-10-02 | 华中农业大学 | Breeding, multiplying and seed production methods for non-heading cabbage cytoplasmic male sterility line |
| CN103329792A (en) * | 2013-06-25 | 2013-10-02 | 华中农业大学 | Breeding, propagating and seed-producing method of purple pakchoi cytoplasmic male sterile line |
| EP2868748A1 (en) * | 2013-10-31 | 2015-05-06 | Locusia Oy | A method for protein production in doubled haploid plants |
| CN104429917B (en) * | 2014-03-06 | 2016-08-31 | 贵州省油菜研究所 | The pure sterile line of seed selection cabbage type rape homogeneity and the method for temporary maintainer line while of a kind of |
| AR109598A1 (en) * | 2016-09-09 | 2018-12-26 | Koch Biological Solutions Llc | IMPROVING PHOTOSYNTHETIC AND STRESS HEAT RISK THROUGH CONTACT OF A PLANT WITH A BiosTIMULANT THAT INCLUDES PSYCHROTOLERANS PSEUDOMONES |
| WO2020251932A1 (en) * | 2019-06-10 | 2020-12-17 | Yield10 Bioscience, Inc. | Transgenic land plants that express a polyhydroxyalkanoate synthase seed specifically with cytosolic localization |
Citations (59)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US704684A (en) | 1900-02-01 | 1902-07-15 | Herbert E Irwin | Alarm device for binders. |
| US4535060A (en) | 1983-01-05 | 1985-08-13 | Calgene, Inc. | Inhibition resistant 5-enolpyruvyl-3-phosphoshikimate synthetase, production and use |
| US4945050A (en) | 1984-11-13 | 1990-07-31 | Cornell Research Foundation, Inc. | Method for transporting substances into living cells and tissues and apparatus therefor |
| WO1991000917A1 (en) | 1989-07-10 | 1991-01-24 | Massachusetts Institute Of Technology | Method for producing novel polyester biopolymers |
| US5023179A (en) | 1988-11-14 | 1991-06-11 | Eric Lam | Promoter enhancer element for gene expression in plant roots |
| US5034322A (en) | 1983-01-17 | 1991-07-23 | Monsanto Company | Chimeric genes suitable for expression in plant cells |
| US5073675A (en) | 1989-05-26 | 1991-12-17 | Dna Plant Technology Corporation | Method of introducing spectinomycin resistance into plants |
| US5110732A (en) | 1989-03-14 | 1992-05-05 | The Rockefeller University | Selective gene expression in plants |
| EP0530129A1 (en) | 1991-08-28 | 1993-03-03 | Sandoz Ltd. | Method for the selection of genetically transformed cells and compounds for use in the method |
| US5240855A (en) | 1989-05-12 | 1993-08-31 | Pioneer Hi-Bred International, Inc. | Particle gun |
| US5268463A (en) | 1986-11-11 | 1993-12-07 | Jefferson Richard A | Plant promoter α-glucuronidase gene construct |
| US5276268A (en) | 1986-08-23 | 1994-01-04 | Hoechst Aktiengesellschaft | Phosphinothricin-resistance gene, and its use |
| US5322783A (en) | 1989-10-17 | 1994-06-21 | Pioneer Hi-Bred International, Inc. | Soybean transformation by microparticle bombardment |
| US5324646A (en) | 1992-01-06 | 1994-06-28 | Pioneer Hi-Bred International, Inc. | Methods of regeneration of Medicago sativa and expressing foreign DNA in same |
| US5399680A (en) | 1991-05-22 | 1995-03-21 | The Salk Institute For Biological Studies | Rice chitinase promoter |
| US5401836A (en) | 1992-07-16 | 1995-03-28 | Pioneer Hi-Bre International, Inc. | Brassica regulatory sequence for root-specific or root-abundant gene expression |
| US5459252A (en) | 1991-01-31 | 1995-10-17 | North Carolina State University | Root specific gene promoter |
| US5463175A (en) | 1990-06-25 | 1995-10-31 | Monsanto Company | Glyphosate tolerant plants |
| US5466785A (en) | 1990-04-12 | 1995-11-14 | Ciba-Geigy Corporation | Tissue-preferential promoters |
| US5519164A (en) | 1990-02-01 | 1996-05-21 | Hoechst Aktiengesellschaft | Expression of a multigene RNA having self-splicing activity |
| US5527695A (en) | 1993-01-29 | 1996-06-18 | Purdue Research Foundation | Controlled modification of eukaryotic genomes |
| US5530196A (en) | 1983-01-17 | 1996-06-25 | Monsanto Company | Chimeric genes for transforming plant cells using viral promoters |
| US5545818A (en) | 1994-03-11 | 1996-08-13 | Calgene Inc. | Expression of Bacillus thuringiensis cry proteins in plant plastids |
| US5563055A (en) | 1992-07-27 | 1996-10-08 | Pioneer Hi-Bred International, Inc. | Method of Agrobacterium-mediated transformation of cultured soybean cells |
| US5569597A (en) | 1985-05-13 | 1996-10-29 | Ciba Geigy Corp. | Methods of inserting viral DNA into plant material |
| US5604121A (en) | 1991-08-27 | 1997-02-18 | Agricultural Genetics Company Limited | Proteins with insecticidal properties against homopteran insects and their use in plant protection |
| WO1997006250A1 (en) | 1995-08-10 | 1997-02-20 | Rutgers University | Nuclear-encoded transcription system in plastids of higher plants |
| US5608144A (en) | 1994-08-12 | 1997-03-04 | Dna Plant Technology Corp. | Plant group 2 promoters and uses thereof |
| US5608149A (en) | 1990-06-18 | 1997-03-04 | Monsanto Company | Enhanced starch biosynthesis in tomatoes |
| US5608142A (en) | 1986-12-03 | 1997-03-04 | Agracetus, Inc. | Insecticidal cotton plants |
| US5633363A (en) | 1994-06-03 | 1997-05-27 | Iowa State University, Research Foundation In | Root preferential promoter |
| US5659026A (en) | 1995-03-24 | 1997-08-19 | Pioneer Hi-Bred International | ALS3 promoter |
| US5668298A (en) | 1984-12-24 | 1997-09-16 | Eli Lilly And Company | Selectable marker for development of vectors and transformation systems in plants |
| US5736369A (en) | 1994-07-29 | 1998-04-07 | Pioneer Hi-Bred International, Inc. | Method for producing transgenic cereal plants |
| US5750386A (en) | 1991-10-04 | 1998-05-12 | North Carolina State University | Pathogen-resistant transgenic plants |
| US5767378A (en) | 1993-03-02 | 1998-06-16 | Novartis Ag | Mannose or xylose based positive selection |
| US5789156A (en) | 1993-06-14 | 1998-08-04 | Basf Ag | Tetracycline-regulated transcriptional inhibitors |
| US5811272A (en) | 1996-07-26 | 1998-09-22 | Massachusetts Institute Of Technology | Method for controlling molecular weight of polyhydroxyalkanoates |
| US5814618A (en) | 1993-06-14 | 1998-09-29 | Basf Aktiengesellschaft | Methods for regulating gene expression |
| US5837876A (en) | 1995-07-28 | 1998-11-17 | North Carolina State University | Root cortex specific gene promoter |
| US5849894A (en) | 1995-11-29 | 1998-12-15 | Monsanto Company | Rhodospirillum rubrum poly-β-hydroxyalkanoate synthase |
| US6011144A (en) | 1994-09-16 | 2000-01-04 | Monsanto Company | PHA E and PHA C components of poly(hydroxy fatty acid) synthase from thiocapsa pfennigii |
| US6072050A (en) | 1996-06-11 | 2000-06-06 | Pioneer Hi-Bred International, Inc. | Synthetic promoters |
| US6077697A (en) | 1996-04-10 | 2000-06-20 | Chromos Molecular Systems, Inc. | Artificial chromosomes, uses thereof and methods for preparing artificial chromosomes |
| US6316262B1 (en) | 1997-09-19 | 2001-11-13 | Metabolix, Inc. | Biological systems for manufacture of polyhydroxyalkanoate polymers containing 4-hydroxyacids |
| US6437220B1 (en) | 1995-09-20 | 2002-08-20 | Monsanto Company, Inc. | Strong early seed-specific gene regulatory region |
| US6444878B1 (en) | 1997-02-07 | 2002-09-03 | Danisco A/S | Method of plant selection using glucosamine-6-phosphate deaminase |
| US6448473B1 (en) | 1999-03-05 | 2002-09-10 | Monsanto Technology Llc | Multigene expression vectors for the biosynthesis of products via multienzyme biological pathways |
| US20020182690A1 (en) * | 1999-01-07 | 2002-12-05 | Cannon Maura C. | Polyhydroxyalkanoate biosynthesis associated proteins and coding region in bacillus megaterium |
| US6586658B1 (en) | 1998-03-06 | 2003-07-01 | Metabolix, Inc. | Modification of fatty acid metabolism in plants |
| US7026526B2 (en) | 2000-02-11 | 2006-04-11 | Metabolix, Inc. | Multi-gene expression constructs containing modified inteins |
| US20060143732A1 (en) | 2001-05-30 | 2006-06-29 | Carl Perez | Plant artificial chromosomes, uses thereof and methods of preparing plant artificial chromosomes |
| WO2006101983A2 (en) * | 2005-03-16 | 2006-09-28 | Metabolix, Inc. | Chemically inducible expression of biosynthetic pathways |
| US20060246586A1 (en) | 2001-05-30 | 2006-11-02 | Edward Perkins | Chromosome-based platforms |
| US20080263728A1 (en) | 2002-06-21 | 2008-10-23 | Genoplante-Valor | Plastidial targeting peptide |
| WO2010037209A1 (en) | 2008-10-03 | 2010-04-08 | Agrisoma Biosciences Inc. | Production of modified fatty acids in plants |
| WO2010061186A2 (en) | 2008-11-25 | 2010-06-03 | Algentech Sas | Plant plastid transformation method |
| WO2010102293A1 (en) | 2009-03-06 | 2010-09-10 | Metabolix, Inc. | Method of positive plant selection using sorbitol dehydrogenase |
| US9801268B2 (en) | 2012-07-10 | 2017-10-24 | Endress + Hauser Gmbh + Co. Kg | Circuit board equipped with a high-frequency component emitting interference waves |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6103956A (en) * | 1998-03-31 | 2000-08-15 | Regents Of The University Of Minnesota | Polyhydroxyalkanoate synthesis in plants |
| ATE483023T1 (en) | 1998-05-22 | 2010-10-15 | Metabolix Inc | POLYHYDROXYALKANOOATBIOPOLYMERS |
| US7045684B1 (en) | 2002-08-19 | 2006-05-16 | Mertec, Llc | Glyphosate-resistant plants |
| US20090007294A1 (en) * | 2004-09-01 | 2009-01-01 | Henry Daniell | Genetic Engineering of Male Sterility in Plants |
-
2010
- 2010-09-15 WO PCT/US2010/048962 patent/WO2011034945A1/en not_active Ceased
- 2010-09-15 CA CA2773707A patent/CA2773707A1/en not_active Abandoned
- 2010-09-15 EP EP10757680A patent/EP2477477A1/en not_active Withdrawn
- 2010-09-15 CA CA2773703A patent/CA2773703A1/en not_active Abandoned
- 2010-09-15 US US13/395,702 patent/US9181559B2/en active Active
- 2010-09-15 EP EP10757363A patent/EP2478105A1/en not_active Withdrawn
- 2010-09-15 BR BR112012005592A patent/BR112012005592A2/en not_active IP Right Cessation
- 2010-09-15 AU AU2010295637A patent/AU2010295637B2/en not_active Ceased
- 2010-09-15 WO PCT/US2010/048963 patent/WO2011034946A1/en not_active Ceased
- 2010-09-15 BR BRBR112012005591-7A patent/BR112012005591A2/en not_active IP Right Cessation
- 2010-09-15 US US13/395,616 patent/US20120174253A1/en not_active Abandoned
Patent Citations (61)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US704684A (en) | 1900-02-01 | 1902-07-15 | Herbert E Irwin | Alarm device for binders. |
| US4535060A (en) | 1983-01-05 | 1985-08-13 | Calgene, Inc. | Inhibition resistant 5-enolpyruvyl-3-phosphoshikimate synthetase, production and use |
| US5034322A (en) | 1983-01-17 | 1991-07-23 | Monsanto Company | Chimeric genes suitable for expression in plant cells |
| US5530196A (en) | 1983-01-17 | 1996-06-25 | Monsanto Company | Chimeric genes for transforming plant cells using viral promoters |
| US4945050A (en) | 1984-11-13 | 1990-07-31 | Cornell Research Foundation, Inc. | Method for transporting substances into living cells and tissues and apparatus therefor |
| US5668298A (en) | 1984-12-24 | 1997-09-16 | Eli Lilly And Company | Selectable marker for development of vectors and transformation systems in plants |
| US5569597A (en) | 1985-05-13 | 1996-10-29 | Ciba Geigy Corp. | Methods of inserting viral DNA into plant material |
| US5276268A (en) | 1986-08-23 | 1994-01-04 | Hoechst Aktiengesellschaft | Phosphinothricin-resistance gene, and its use |
| US5268463A (en) | 1986-11-11 | 1993-12-07 | Jefferson Richard A | Plant promoter α-glucuronidase gene construct |
| US5608142A (en) | 1986-12-03 | 1997-03-04 | Agracetus, Inc. | Insecticidal cotton plants |
| US5023179A (en) | 1988-11-14 | 1991-06-11 | Eric Lam | Promoter enhancer element for gene expression in plant roots |
| US5110732A (en) | 1989-03-14 | 1992-05-05 | The Rockefeller University | Selective gene expression in plants |
| US5240855A (en) | 1989-05-12 | 1993-08-31 | Pioneer Hi-Bred International, Inc. | Particle gun |
| US5073675A (en) | 1989-05-26 | 1991-12-17 | Dna Plant Technology Corporation | Method of introducing spectinomycin resistance into plants |
| WO1991000917A1 (en) | 1989-07-10 | 1991-01-24 | Massachusetts Institute Of Technology | Method for producing novel polyester biopolymers |
| US5322783A (en) | 1989-10-17 | 1994-06-21 | Pioneer Hi-Bred International, Inc. | Soybean transformation by microparticle bombardment |
| US5519164A (en) | 1990-02-01 | 1996-05-21 | Hoechst Aktiengesellschaft | Expression of a multigene RNA having self-splicing activity |
| US5466785A (en) | 1990-04-12 | 1995-11-14 | Ciba-Geigy Corporation | Tissue-preferential promoters |
| US5608149A (en) | 1990-06-18 | 1997-03-04 | Monsanto Company | Enhanced starch biosynthesis in tomatoes |
| US5463175A (en) | 1990-06-25 | 1995-10-31 | Monsanto Company | Glyphosate tolerant plants |
| US5459252A (en) | 1991-01-31 | 1995-10-17 | North Carolina State University | Root specific gene promoter |
| US5399680A (en) | 1991-05-22 | 1995-03-21 | The Salk Institute For Biological Studies | Rice chitinase promoter |
| US5604121A (en) | 1991-08-27 | 1997-02-18 | Agricultural Genetics Company Limited | Proteins with insecticidal properties against homopteran insects and their use in plant protection |
| EP0530129A1 (en) | 1991-08-28 | 1993-03-03 | Sandoz Ltd. | Method for the selection of genetically transformed cells and compounds for use in the method |
| US5750386A (en) | 1991-10-04 | 1998-05-12 | North Carolina State University | Pathogen-resistant transgenic plants |
| US5324646A (en) | 1992-01-06 | 1994-06-28 | Pioneer Hi-Bred International, Inc. | Methods of regeneration of Medicago sativa and expressing foreign DNA in same |
| US5401836A (en) | 1992-07-16 | 1995-03-28 | Pioneer Hi-Bre International, Inc. | Brassica regulatory sequence for root-specific or root-abundant gene expression |
| US5563055A (en) | 1992-07-27 | 1996-10-08 | Pioneer Hi-Bred International, Inc. | Method of Agrobacterium-mediated transformation of cultured soybean cells |
| US5527695A (en) | 1993-01-29 | 1996-06-18 | Purdue Research Foundation | Controlled modification of eukaryotic genomes |
| US5767378A (en) | 1993-03-02 | 1998-06-16 | Novartis Ag | Mannose or xylose based positive selection |
| US5814618A (en) | 1993-06-14 | 1998-09-29 | Basf Aktiengesellschaft | Methods for regulating gene expression |
| US5789156A (en) | 1993-06-14 | 1998-08-04 | Basf Ag | Tetracycline-regulated transcriptional inhibitors |
| US5545818A (en) | 1994-03-11 | 1996-08-13 | Calgene Inc. | Expression of Bacillus thuringiensis cry proteins in plant plastids |
| US5633363A (en) | 1994-06-03 | 1997-05-27 | Iowa State University, Research Foundation In | Root preferential promoter |
| US5736369A (en) | 1994-07-29 | 1998-04-07 | Pioneer Hi-Bred International, Inc. | Method for producing transgenic cereal plants |
| US5608144A (en) | 1994-08-12 | 1997-03-04 | Dna Plant Technology Corp. | Plant group 2 promoters and uses thereof |
| US6011144A (en) | 1994-09-16 | 2000-01-04 | Monsanto Company | PHA E and PHA C components of poly(hydroxy fatty acid) synthase from thiocapsa pfennigii |
| US5659026A (en) | 1995-03-24 | 1997-08-19 | Pioneer Hi-Bred International | ALS3 promoter |
| US5837876A (en) | 1995-07-28 | 1998-11-17 | North Carolina State University | Root cortex specific gene promoter |
| WO1997006250A1 (en) | 1995-08-10 | 1997-02-20 | Rutgers University | Nuclear-encoded transcription system in plastids of higher plants |
| US6437220B1 (en) | 1995-09-20 | 2002-08-20 | Monsanto Company, Inc. | Strong early seed-specific gene regulatory region |
| US5849894A (en) | 1995-11-29 | 1998-12-15 | Monsanto Company | Rhodospirillum rubrum poly-β-hydroxyalkanoate synthase |
| US6077697A (en) | 1996-04-10 | 2000-06-20 | Chromos Molecular Systems, Inc. | Artificial chromosomes, uses thereof and methods for preparing artificial chromosomes |
| US6072050A (en) | 1996-06-11 | 2000-06-06 | Pioneer Hi-Bred International, Inc. | Synthetic promoters |
| US5811272A (en) | 1996-07-26 | 1998-09-22 | Massachusetts Institute Of Technology | Method for controlling molecular weight of polyhydroxyalkanoates |
| US6444878B1 (en) | 1997-02-07 | 2002-09-03 | Danisco A/S | Method of plant selection using glucosamine-6-phosphate deaminase |
| US6316262B1 (en) | 1997-09-19 | 2001-11-13 | Metabolix, Inc. | Biological systems for manufacture of polyhydroxyalkanoate polymers containing 4-hydroxyacids |
| US6586658B1 (en) | 1998-03-06 | 2003-07-01 | Metabolix, Inc. | Modification of fatty acid metabolism in plants |
| US20020182690A1 (en) * | 1999-01-07 | 2002-12-05 | Cannon Maura C. | Polyhydroxyalkanoate biosynthesis associated proteins and coding region in bacillus megaterium |
| US6835820B2 (en) | 1999-01-07 | 2004-12-28 | The University Of Massachusetts | Polyhydroxyalkanoate biosynthesis associated proteins and coding region in bacillus megaterium |
| US6448473B1 (en) | 1999-03-05 | 2002-09-10 | Monsanto Technology Llc | Multigene expression vectors for the biosynthesis of products via multienzyme biological pathways |
| US7026526B2 (en) | 2000-02-11 | 2006-04-11 | Metabolix, Inc. | Multi-gene expression constructs containing modified inteins |
| US20060143732A1 (en) | 2001-05-30 | 2006-06-29 | Carl Perez | Plant artificial chromosomes, uses thereof and methods of preparing plant artificial chromosomes |
| US20060246586A1 (en) | 2001-05-30 | 2006-11-02 | Edward Perkins | Chromosome-based platforms |
| US20080263728A1 (en) | 2002-06-21 | 2008-10-23 | Genoplante-Valor | Plastidial targeting peptide |
| WO2006101983A2 (en) * | 2005-03-16 | 2006-09-28 | Metabolix, Inc. | Chemically inducible expression of biosynthetic pathways |
| WO2010037209A1 (en) | 2008-10-03 | 2010-04-08 | Agrisoma Biosciences Inc. | Production of modified fatty acids in plants |
| US20100186117A1 (en) | 2008-10-03 | 2010-07-22 | Agrisoma Biosciences Inc. | Production of modified fatty acids in plants |
| WO2010061186A2 (en) | 2008-11-25 | 2010-06-03 | Algentech Sas | Plant plastid transformation method |
| WO2010102293A1 (en) | 2009-03-06 | 2010-09-10 | Metabolix, Inc. | Method of positive plant selection using sorbitol dehydrogenase |
| US9801268B2 (en) | 2012-07-10 | 2017-10-24 | Endress + Hauser Gmbh + Co. Kg | Circuit board equipped with a high-frequency component emitting interference waves |
Non-Patent Citations (234)
| Title |
|---|
| AGRAWAL GK ET AL., NUCLEIC ACIDS RESEARCH, vol. 29, 2001, pages 1835 - 1843 |
| AKASOFU ET AL., NUCLEIC ACIDS RESEARCH, vol. 18, 1990, pages 1892 |
| ALLISON L.A. ET AL., EMBO, vol. 15, 1996, pages 2802 - 2809 |
| ALLISON LA ET AL., EMBO, vol. 15, 1996, pages 2802 - 2809 |
| APPL. MICROBIOL. BIOTECHNOL., vol. 42, 1995, pages 901 - 909 |
| ARAI ET AL., PLANT BIOTECHNOL., vol. 18, 2001, pages 289 - 293 |
| ARCHER ET AL., J BIOENERG. BIOMEMB., vol. 22, no. 6, 1990, pages 789 - 810 |
| AUSUBEL ET AL.: "Current Protocols in Molecular Biology", 1987, GREEN PUBLISHING |
| BARFIELD; PUA, PLANT CELL REPORTS, vol. 10, pages 308 - 314 |
| BEVAN, M.; BARNES, W. M.; CHILTON, M. D., NUCLEIC ACIDS RES., vol. 11, 1983, pages 369 - 385 |
| BEVIS; GLICK, NAT BIOTECH, vol. 20, 2002, pages 83 - 87 |
| BOHMERT ET AL., MOLECULAR BIOLOGY AND BIOTECHNOLOGY OF PLANT ORGANELLES, 2004, pages 559 - 585 |
| BOHMERT ET AL., PLANT PHYSIOL., vol. 128, 2002, pages 1282 - 1290 |
| BOHMERT ET AL., PLANTA, vol. 211, 2000, pages 841 - 845 |
| BOHMERT, K., I. ET AL., PLANTA, vol. 211, 2000, pages 841 - 845 |
| BOHMERT: "Molecular Biology and Biotechnology of Plant Organelles", 2004, KLUWER ACADEMIC PUBLISHERS, pages: 559 - 585 |
| BROWN ET AL., J. BACTERIOL., vol. 177, 1995, pages 6536 - 6544 |
| BYTEBIER ET AL., PROC. NATL. ACAD SCI. USA, vol. 84, 1987, pages 5345 - 5349 |
| CANEVASCINI ET AL., PLANT PHYSIOL., vol. 112, no. 2, 1996, pages 513 - 524 |
| CARLSON ET AL., PLOS GENET, vol. 3, 2007, pages 1965 - 74 |
| CASHMORE, A. R.: "Genetic Engineering of Plants", 1983, PLENUM, article "Nuclear Genes Encoding the Small Subunit of Ribulose-1,5-Bisphosphate Carboxylase", pages: 29 - 38 |
| CASHMORE, A.R., GENETIC ENGINEERING OF PLANTS, 1983, pages 29 - 38 |
| CHAWLA, R. ET AL., PLANT BIOTECHNOL J, vol. 4, 2006, pages 209 - 218 |
| CHOI, S. ET AL., NUCLEIC ACIDS RES, vol. 28, 2000, pages E19 |
| CHRISTENSEN ET AL., PLANT MOL. BIOL., vol. 12, 1989, pages 619 - 632 |
| CHRISTENSEN ET AL., PLANT MOL. BIOL., vol. 18, 1992, pages 675 - 689 |
| CHRISTOU AND FORD ANNALS OF BOTANY, vol. 75, 1995, pages 407 - 413 |
| CHRISTOU ET AL., PLANT PHYSIOL, vol. 87, 1988, pages 671 - 674 |
| CLARK ET AL., J. BIOL. CHEM., vol. 264, 1989, pages 17544 - 17550 |
| CROSSWAY ET AL., BIOTECHNIQUES, vol. 4, 1986, pages 320 - 334 |
| CUBITT ET AL., TRENDS BIOCHEM. SCI., vol. 20, 1995, pages 448 - 455 |
| CZARNECKA, E. ET AL., MOL. CELL BIOL., vol. 9, 1989, pages 3457 - 3463 |
| DAFTA ET AL., BIOTECHNOLOGY, vol. 8, 1990, pages 736 - 740 |
| DALE; OW, PROC. NATL. ACAD. SCI. USA, vol. 88, 1991, pages 10558 - 10562 |
| DANIELL H ET AL., CURR. GENET., vol. 39, 2001, pages 109 - 116 |
| DAVIS; VIERSTRA, PLANT MOLECULAR BIOLOGY, vol. 36, 1998, pages 521 - 528 |
| DE CASTRO SILVA FILHO ET AL., PLANT MOL. BIOL., vol. 30, 1996, pages 769 - 780 |
| DE WET ET AL.: "The Experimental Manipulation of Ovule Tissues", 1985, pages: 197 - 209 |
| DELLA-CIOPPA ET AL., PLANT PHYSIOL., vol. 84, 1987, pages 965 - 968 |
| D'HALLUIN ET AL., PLANT CELL, vol. 4, 1992, pages 1495 - 1505 |
| DIRUSSO, C. C., J. BACTERIOL., vol. 172, 1990, pages 6459 - 6468 |
| DJUKANOVIC, V ET AL., PLANT BIOTECHNOL J, vol. 4, 2006, pages 345 - 357 |
| DMOCHOWSKA ET AL., GENE, vol. 88, 1990, pages 247 - 252 |
| EISEN ET AL., PROC. NATL. ACAD. SCI. USA, vol. 95, 1998, pages 14863 - 14868 |
| ELLENSTROM, M. ET AL., PLANT MOLECULAR BIOLOGY, vol. 32, 1996, pages 1019 - 1027 |
| ERIKSON ET AL., NAT BIOTECHNOL, vol. 22, 2004, pages 455 - 8 |
| ESSER, K. ET AL.: "Progress in Botany", vol. 67, 2006, SPRINGER, pages: 31 - 52 |
| ESSER, K.: "Progress in Botany", vol. 67, 2006, SPRINGER, pages: 31 - 52 |
| EVANS ET AL.: "Handbook of Plant Cell Culture", vol. 1, 1983, MACMILLAN PUBLISHING CO. |
| F. M. AUSUBEL, ET AL.: "Current Protocols In Molecular Biology", 1987 |
| FINER; MCMULLEN, IN VITRO CELL DEV. BIOL., vol. 27P, 1991, pages 175 - 182 |
| FROMM ET AL., BIOTECHNOLOGY, vol. 8, 1990, pages 833 - 839 |
| FUKUI; DOI, J. BACTERIOL., vol. 179, 1997, pages 4821 - 30 |
| GASSER; FRALEY, SCIENCE, vol. 244, 1989, pages 1293 - 99 |
| GATZ ET AL., MOL. GEN. GENET., vol. 227, 1991, pages 229 - 237 |
| GOTOR ET AL., PLANT J., vol. 3, 1993, pages 509 - 18 |
| GUEVARA-GARCIA ET AL., PLANT J., vol. 4, no. 3, 1993, pages 495 - 505 |
| HAJDUKIEWICZ PTJ ET AL., EMBO J., vol. 16, 1997, pages 4041 - 4048 |
| HALL, CAN. J. MICROBIOL., vol. 44, 1998, pages 687 - 91 |
| HANSEN ET AL., MOL. GEN. GENET., vol. 254, no. 3, 1997, pages 337 - 343 |
| HEM ET AL., ARCHIVES OF MICROBIOLOGY, vol. 170, 1998, pages 162 - 170 |
| HERZ ET AL., TRANSGENIC RESEARCH, vol. 14, 2005, pages 969 - 982 |
| HIRAI, SAYAKA ET AL: "RNAi Vectors for manipulation of gene expression in higher plants", OPEN PLANT SCIENCE JOURNAL , 2, 21-30 CODEN: OPSJA4; ISSN: 1874-2947 URL: HTTP://WWW.BENTHAM.ORG/OPEN/TOPSJ/OPENACCESS2.HTM, 2008, XP009142252 * |
| HIRE ET AL., PLANT MOL. BIOL., vol. 20, no. 2, 1992, pages 207 - 218 |
| HOOD, J. BACTERIOL., vol. 168, 1986, pages 1291 - 1301 |
| HOOYKAAS-VAN SLOGTEREN ET AL., NATURE, vol. 311, 1984, pages 763 - 764 |
| HOUMIEL ET AL., PLANTA, vol. 209, 1999, pages 547 - 550 |
| HOUMIEL KATHRYN L ET AL: "Poly(beta-hydroxybutyrate) production in oilseed leukoplasts of Brassica napus", PLANTA, SPRINGER VERLAG, DE, vol. 209, no. 4, 1 October 1999 (1999-10-01), pages 547 - 550, XP002195214, ISSN: 0032-0935, DOI: DOI:10.1007/S004250050760 * |
| HUANG F-C ET AL., MOL. GENET. GENOMICS, vol. 268, 2002, pages 19 - 27 |
| HUISMAN ET AL., J. BIOL. CHEM., vol. 266, 1991, pages 2191 - 2198 |
| IIDA ET AL., PLANT CELL REPORTS, vol. 14, 1995, pages 539 - 544 |
| ITO ET AL., J. MOL. BIOL., vol. 355, 2006, pages 722 - 733 |
| ITO ET AL: "d-3-Hydroxybutyrate Dehydrogenase from Pseudomonas fragi: Molecular Cloning of the Enzyme Gene and Crystal Structure of the Enzyme", JOURNAL OF MOLECULAR BIOLOGY, LONDON, GB, vol. 355, no. 4, 27 January 2006 (2006-01-27), pages 722 - 733, XP005216917, ISSN: 0022-2836, DOI: DOI:10.1016/J.JMB.2005.10.072 * |
| JEFFERSON ET AL., EMBO J., vol. 6, 1987, pages 3901 - 3907 |
| JENDROSSEK, D., J. BACTERIOL., vol. 191, no. 10, 2009, pages 3195 - 3202 |
| JENDROSSEK; HANDRICK, ANNU REV MICROBIOL, vol. 56, 2002, pages 403 - 432 |
| JOHN ET AL., PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 93, 1996, pages 12768 - 12773 |
| KAEPPLER ET AL., PLANT CELL REPORTS, vol. 9, 1990, pages 415 - 418 |
| KAEPPLER ET AL., THEOR. APPL. GENET., vol. 84, 1992, pages 560 - 566 |
| KATO ET AL., APPL. MICROBIOL. BIOTECHNOL., vol. 45, 1996, pages 363 - 370 |
| KAWAMATA ET AL., PLANT CELL PHYSIOL, vol. 38, no. 7, 1997, pages 792 - 803 |
| KELLER; BAUMGARTNER, PLANT CELL, vol. 3, no. 10, 1991, pages 1051 - 1061 |
| KENDREW ET AL.: "The Encyclopedia of Molecular Biology", 1999, WILEY-INTERSCIENCE |
| KERBACH, S. ET AL., THEOR APPL GENET, vol. 111, 2005, pages 1608 - 1616 |
| KLEIN ET AL., BIOTECHNOLOGY, vol. 6, 1988, pages 559 - 563 |
| KLEIN ET AL., PLANT PHYSIOL., vol. 91, 1988, pages 440 - 444 |
| KLEIN ET AL., PROC. NATL. ACAD. SCI. USA, vol. 85, 1988, pages 4305 - 4309 |
| KOURTZ ET AL., TRANSGENIC RES, vol. 16, 2007, pages 759 - 769 |
| KOURTZ, L., KL ET AL., PLANT BIOTECBNOL., vol. 3, 2005, pages 435 - 447 |
| KWON ET AL., PLANT PHYSIOL., vol. 105, 1994, pages 357 - 67 |
| LAM, RESULTS PROBL. CELL DIFFER., vol. 20, 1994, pages 181 - 196 |
| LAMPPA ET AL., J BIOL. CHEM., vol. 263, 1988, pages 14996 - 14999 |
| LAST ET AL., THEOR. APPL. GENET., vol. 81, 1991, pages 581 - 588 |
| LAWRENCE ET AL., J. BIOL. CHEM., vol. 272, no. 33, 1997, pages 20357 - 20363 |
| LAZO, G. ET AL., BIOTECHNOLOGY, vol. 9, 1991, pages 963 - 967 |
| LE ET AL., PROC. NATL. ACAD. SCI. USA, vol. 107, 2010, pages 8063 - 8070 |
| LEFEBVRE ET AL., PLANT PHYSIOL., vol. 138, 2005, pages 451 - 460 |
| LEWIN: "Genes VII", 2000, OXFORD UNIVERSITY PRESS |
| LI ET AL., PLANT CELL REPORTS, vol. 12, 1993, pages 250 - 255 |
| LI, W. ET AL., PLANT MOL BIOL, 2010 |
| LIDA, A. ET AL., PLANT CELL REPORTS, vol. 14, 1995, pages 539 - 544 |
| LIEBERGESELL, M.; STEINBUCHEL, A., EUR. J. BIOCHEM., vol. 209, 1992, pages 135 - 150 |
| LOSSL ET AL., PLANT CELL PHYSIOL, vol. 46, 2005, pages 1462 - 1471 |
| LÖSSL ET AL., PLANT CELL PHYSIOL, vol. 46, 2005, pages 1462 - 1471 |
| LOSSL ET AL., PLANT CELL REPORTS, vol. 21, 2003, pages 891 - 899 |
| LUTZ ET AL., PLANT J, vol. 37, 2004, pages 906 - 13 |
| LUTZ ET AL., PLANT PHYSIOL, vol. 145, 2007, pages 1201 - 10 |
| LUTZ KA ET AL., PLANT J, vol. 37, 2004, pages 906 - 913 |
| LUTZ KA ET AL., PLANT PHYSIOL., vol. 145, 2007, pages 1201 - 1210 |
| LYZNIK, LA ET AL., NUCLEIC ACIDS RES, vol. 21, 1993, pages 969 - 975 |
| M. D. HAYWARD; N. O. BOSEMARK; I. ROMAGOSA: "Plant Breeding: Principles and Prospects", vol. 1, 1993, CHAPMAN & HALL |
| M. J. MACPHERSON, B. D. HAMES AND G. R. TAYLOR: "the series Methods in Enzymology", 1995, ACADEMIC PRESS, INC. |
| MADISON ET AL., MICROBIAL. MOL. BIOL. REV., vol. 63, 1999, pages 21 - 53 |
| MADISON; HUISMAN, MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, vol. 63, 1999, pages 21 - 53 |
| MALIGA, ET AL.: "Methods in Plant Molecular Biology: A Laboratory Course Manual", 1995, COLD SPRING LABORATORY PRESS |
| MATSUMOTO, JOURNAL OF POLYMERS AND THE ENVIRONMENT, vol. 14, 2006, pages 369 - 374 |
| MATSUOKA ET AL., PROC NATL. ACAD SCI. USA, vol. 90, no. 20, 1993, pages 9586 - 9590 |
| MATSUOKA ET AL., PROC. NATL. ACAD SCI. USA, vol. 90, no. 20, 1993, pages 9586 - 9590 |
| MATZ ET AL., NAT BIOTECHNOL, vol. 17, 1999, pages 969 - 73 |
| MCB.RIDE ET AL., PROC. NATL. ACAD SCI. USA, vol. 91, 1994, pages 7301 - 7305 |
| MCCABE ET AL., BIOTECHNOLOGY, vol. 6, 1988, pages 923 - 926 |
| MCCORMICK ET AL., PLANT CELL REPORTS, vol. 5, 1986, pages 81 - 84 |
| MCELROY ET AL., PLANT CELL, vol. 2, 1990, pages 163 - 171 |
| MCNELLIS ET AL., PLANT J., vol. 14, no. 2, 1998, pages 247 - 257 |
| MEDBERRY ET AL., NUCLEIC ACIDS RES., vol. 23, 1995, pages 485 - 490 |
| MIAO ET AL., PLANT CELL, vol. 3, no. 1, 1991, pages 11 - 22 |
| MIKI ET AL., JOURNAL OF BIOTECHNOLOGY, vol. 107, 2004, pages 193 - 232 |
| MIRAS, S. ET AL., J BIOL CHEM, vol. 277, no. 49, 2002, pages 47770 - 8 |
| MIRAS, S. ET AL., J BIOL CHEM, vol. 282, 2007, pages 29482 - 29492 |
| MITTENDORF ET AL., PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 95, 1998, pages 13397 - 13402 |
| MIYAGAWA, NAT. BIOTECHNOL., vol. 19, 2001, pages 965 - 969 |
| MIYAGAWA, Y., NAT BIOTECHNOL, vol. 19, 2001, pages 965 - 9 |
| MULLANEY; REHM, JOURNAL OF BIOTECHNOLOGY, vol. 147, 2010, pages 31 - 36 |
| NAGAI, T. ET AL., NAT BIOTECH, vol. 20, 2002, pages 87 - 90 |
| NAWRATH, C.; Y. POIRIER ET AL., PROC. NATL. ACAD. SCI. USA, vol. 91, 1994, pages 12760 - 12764 |
| ODELL ET AL., NATURE, vol. 313, 1985, pages 810 - 812 |
| OROZCO ET AL., PLANT MOL BIOL., vol. 23, no. 6, 1993, pages 1129 - 1138 |
| OROZCO ET AL., PLANT MOL. BIOL., vol. 23, no. 6, 1993, pages 1129 - 1138 |
| OSJODA ET AL., NATURE BIOTECHNOLOGY, vol. 14, 1996, pages 745 - 750 |
| OWEN, ET AL.,: "Transgenic Plants: A Production System for Industrial and Pharmaceutical Prot", 1996, JOHN WILEY & SONS LTD. |
| PAN ET AL., PLANT PHYSIOL., vol. 112, 1996, pages 893 - 900 |
| PANDIAN ET AL., PLANT MOLECULAR BIOLOGY REPORTER, vol. 24, 2006, pages 103A - 103I |
| PASZKOWSKI ET AL., EMBO J., vol. 3, 1984, pages 2717 - 2722 |
| PEOPLES, O. P.; SINSKEY, A. J., J BIOL. CHEM., vol. 264, 1989, pages 15298 - 15303 |
| PEOPLES, O. P.; SINSKEY, A. J., J. BIOL. CHEM, vol. 264, 1989, pages 15293 - 15297 |
| PEOPLES, O. P.; SINSKEY, A. J., J. BIOL. CHEM., vol. 264, 1989, pages 15293 - 15297 |
| PEOPLES, O. P.; SINSKEY, A. J., MOLECULAR MICROBIOLOGY, vol. 3, 1989, pages 349 - 357 |
| PEOPLES, O.; A. SINSKEY, J. BIOL. CHEM., vol. 264, 1989, pages 15293 - 15297 |
| PIEPER; STEINBUECHEL, FEMS MICROBIOL.LETT., vol. 96, no. 1, 1992, pages 73 - 80 |
| POIRIER ET AL., PLANT PHYSIOL., vol. 121, 1999, pages 1359 - 1366 |
| POIRIER ET AL.: "Biopolymers", vol. 3A, 2002, WILEY-VHC VERLAG GMBH, article "Polyhydroxyalkanoate production in transgenic plants", pages: 401 - 435 |
| POTRYKUS ET AL., MOL. GEN. GENET., vol. 199, 1985, pages 183 - 188 |
| POTRYKUS ET AL., PLANT MOLECULAR BIOLOGY REPORTER, vol. 3, 1985, pages 117 - 128 |
| POTRYKUS, ET AL.,: "Gene Transfer to Plants", 1995, SPRINGER-VERLAG |
| PURNELL ET AL., PLANT BIOTECHNOL. J., vol. 5, 2007, pages 173 - 184 |
| RAINES, PHOTOSYNTHESIS RESEARCH, vol. 75, 2003, pages 1 - 10 |
| RIGGS ET AL., PROC. NATL. ACAD SCI. USA, vol. 83, 1986, pages 5602 - 5606 |
| RINEHART ET AL., PLANT PHYSIOL., vol. 112, no. 3, 1996, pages 1331 - 1341 |
| ROBERT A. MEYERS: "Molecular Biology and Biotechnology, a Comprehensive Desk Reference", 1995, VCH PUBLISHERS, INC. |
| ROMANO ET AL., PLANTA, vol. 220, 2005, pages 455 - 464 |
| ROMER ET AL., BIOCHEM. BIOPHYS. RES. COMMUN., vol. 196, 1993, pages 1414 - 1421 |
| ROWLEY ET AL., BIOCHIM BIOPHYS ACTA, vol. 1345, 1997, pages 1 - 4 |
| ROWLEY; HERMAN, BIOCHIM. BIOPHYS. ACTA, vol. 1345, 1997, pages 1 - 4 |
| RUIZ, O. N.; H. DANIELL, PLANT PHYSIOL., vol. 138, 2005, pages 1232 - 1246 |
| RUSSELL ET AL., TRANSGENIC RES, vol. 6, no. 2, 1997, pages 157 - 168 |
| SAEGUSA ET AL., J. BACTERIOL., vol. 183, 2001, pages 94 - 100 |
| SAEGUSA H ET AL: "Cloning of an intracellular poly(D(-)-3-hydroxybutyrate) depolymerase gene from Ralstonia eutropha H16 and characterization of the gene product", JOURNAL OF BACTERIOLOGY, AMERICAN SOCIETY FOR MICROBIOLOGY, WASHINGTON, DC; US, vol. 183, no. 1, 1 January 2001 (2001-01-01), pages 94 - 100, XP002282418, ISSN: 0021-9193, DOI: DOI:10.1128/JB.183.1.94-100.2001 * |
| SAEGUSA, H.; M. SHIRAKI ET AL., J. BACTERIOL., vol. 183, 2001, pages 94 - 100 |
| SAMBROOK; RUSSELL: "Molecular Cloning: A Laboratory Manual, 3rd edition", 2001 |
| SANFORD ET AL., PARTICULATE SCIENCE AND TECHNOLOGY, vol. 5, 1987, pages 27 - 37 |
| SANGER ET AL., PLANT MOL. BIOL., vol. 14, no. 3, 1990, pages 433 - 443 |
| SARUUL ET AL., CROP SCI., vol. 42, 2002, pages 919 - 927 |
| SATO, S., HAYASHI ET AL., J. BIOCHEM., vol. 111, 1992, pages 8 - 15 |
| SATO, S.; HAYASHI, M.; IMAMURA, S.; OZEKI, Y.; KAWAGUCHI, A., J BIOCHEM., vol. 111, 1992, pages 8 - 15 |
| SCHENA ET AL., PROC. NATL. ACAD SCI. USA, vol. 88, 1991, pages 10421 - 10425 |
| SCHMIDT ET AL., J. BIOL. CHEM., vol. 268, no. 36, 1993, pages 27447 - 27457 |
| SCHNELL ET AL., J. BIOL. CHEM., vol. 266, no. 5, 1991, pages 3335 - 3342 |
| SHAH ET AL., SCIENCE, vol. 233, 1986, pages 478 - 481 |
| SHEEN ET AL., PLANT J, vol. 8, 1995, pages 777 - 84 |
| SHIINA T. ET AL., PLANT CELL, vol. 10, 1998, pages 1713 - 1722 |
| SHIV ET AL., JOURNAL OF PLANT BIOCHEMISTRY AND BIOTECHNOLOGY, vol. 17, 2008, pages 1 - 4 |
| SHUKLA ET AL., NATURE, 2009 |
| SINGH ET AL., THEOR. APPL. GENET., vol. 96, 1998, pages 319 - 324 |
| SLATER ET AL., JBACTERIOL., vol. 180, no. 8, 1998, pages 1979 - 87 |
| SLATER ET AL., NAT. BIOTECHNOL, vol. 17, 1999, pages 1011 - 1016 |
| SLATER ET AL., NAT. BIOTECHNOL., vol. 17, 1999, pages 1011 - 1016 |
| SNELL; PEOPLES, METAB. ENG., vol. 4, 2002, pages 29 - 40 |
| SOMLEVA ET AL., PLANT BIOTECHNOL. J, vol. 6, 2008, pages 663 - 678 |
| SOMLEVA ET AL., PLANT BIOTECHNOL. J., 2008, pages 663 - 678 |
| SPEICHER AND WINGFELD,: "Current Protocols in Protein Science", 1995, JOHN WILEY & SONS, INC. |
| SRIVASTAVA, V; OW, DW, PLANT MOL BIOL, vol. 46, 2001, pages 561 - 566 |
| STAUB JM; MALIGA P, PLANT CELL, vol. 4, 1992, pages 39 - 45 |
| SUDESH ET AL., CLEAN - SOIL, AIR, WATER, vol. 36, 2008, pages 433 - 442 |
| SUDESH ET AL., PROG. POLYM. SCI., vol. 25, 2000, pages 1503 - 1555 |
| SULLIVAN ET AL., MOL. GEM GENET., vol. 215, 1989, pages 431 - 40 |
| SURIYAMONGKOL ET AL., BIOTECHNOL ADV, vol. 25, 2007, pages 148 - 175 |
| SURIYAMONGKOL ET AL: "Biotechnological approaches for the production of polyhydroxyalkanoates in microorganisms and plants - A review", BIOTECHNOLOGY ADVANCES, ELSEVIER PUBLISHING, BARKING, GB, vol. 25, no. 2, 26 January 2007 (2007-01-26), pages 148 - 175, XP005862313, ISSN: 0734-9750, DOI: DOI:10.1016/J.BIOTECHADV.2006.11.007 * |
| SVAB ET AL., PROC. NATL. ACAD. SCI. USA, vol. 87, 1990, pages 8526 - 8530 |
| SVAB ET AL., PROC. NATL. ACAD. SCI. USA, vol. 90, 1993, pages 913 - 917 |
| SVAB Z; HAJDUKIEWICZ P; MALIGA P, PROC. NATL. ACAD. SCI. USA, vol. 87, 1990, pages 8526 - 8530 |
| SVAB Z; MALIGA P., MOL. GEN. GENET., vol. 241, 1993, pages 49 - 56 |
| SVAB Z; MALIGA P., PROC. NATL. ACAD. SCI. USA, vol. 90, 1993, pages 913 - 917 |
| SVAB; MALIGA, EMBO J., vol. 12, 1993, pages 601 - 606 |
| SVAB; MALIGA, PROC. NATL. ACAD SCI. USA, vol. 90, 1993, pages 913 - 917 |
| TAMOI, M. ET AL., ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS, vol. 334, 1996, pages 27 - 36 |
| TARONCHER-OLDENBURG ET AL., APPL. ENVIRON. MICROBIOL., vol. 66, 2000, pages 4440 - 4448 |
| THOMPSON ET AL., BIOESSAYS, vol. 10, 1989, pages 108 |
| TIMM, A.; STEINBUCHEL, A., EUR. J. BIOCHEM., vol. 209, 1992, pages 15 - 30 |
| TOKIWA; CALABIA, BIOTECHNOLOGY LETTERS, vol. 26, 2004, pages 1181 - 1189 |
| TOMES ET AL.: "Plant Cell, Tissue, and Organ Culture: Fundamental Methods", 1995, SPRINGER-VERLAG |
| TOWNSEND ET AL., NATURE, 2009 |
| TRIGGS-RAINE, B. L.; LOEWEN, P. C., GENE, vol. 52, 1987, pages 121 - 128 |
| TZFIRA ET AL., PLANT MOLECULAR BIOLOGY, vol. 57, 2005, pages 503 - 516 |
| VALENTIN ET AL., INT. J. BIOL. MACROMOL., vol. 25, 1999, pages 303 - 306 |
| VALENTIN H E ET AL: "PHA PRODUCTION, FROM BACTERIA TO PLANTS", INTERNATIONAL JOURNAL OF BIOLOGICAL MACROMOLECULES, ELSEVIER BV, NL, vol. 25, no. 1-03, 1 June 1999 (1999-06-01), pages 303 - 306, XP000892982, ISSN: 0141-8130, DOI: DOI:10.1016/S0141-8130(99)00045-8 * |
| VAN BEILEN ET AL., THE PLANT JOURNAL, vol. 54, 2008, pages 684 - 701 |
| VAN CAMP, PLANT PHYSIOL, vol. 112, no. 2, 1996, pages 525 - 535 |
| VASIL, IK: "Cell Culture and Somatic Cell Genetics", 1984, ACADEMIC |
| VELTEN ET AL., EMBO J., vol. 3, 1984, pages 2723 - 2730 |
| VERHOUNIG ET AL., PROC NATL ACAD SCI U S A, vol. 107, 2010, pages 6204 - 6209 |
| VERKHUSHA, V. V.; K. A. LUKYANOV, NAT BIOTECH, vol. 22, 2004, pages 289 - 296 |
| VON HEIJNE ET AL., PLANT MOL. BIOL. REP., vol. 9, 1991, pages 104 - 126 |
| WANG ET AL., CHINESE SCIENCE BULLETIN, vol. 50, 2005, pages 1113 - 1120 |
| WEISSINGER ET AL., ANN. REV. GENET., vol. 22, 1988, pages 421 - 477 |
| WROBEL ET AL., J. BIOTECHNOL., vol. 107, 2004, pages 41 - 54 |
| WROBEL-KWIATKOWSK ET AL., BIOTECHNOL PROG, vol. 23, 2007, pages 269 - 277 |
| YAMAMOTO ET AL., PLANT CELL PHYSIOL, vol. 35, no. 5, 1994, pages 773 - 778 |
| YAMAMOTO ET AL., PLANT J, vol. 12, no. 2, 1997, pages 255 - 265 |
| YAMAMOTO ET AL., PLANT J., vol. 12, no. 2, 1997, pages 255 - 265 |
| YORK, G. M. ET AL., J. BACTERIOL., vol. 185, 2003, pages 3788 - 3794 |
| YU ET AL., PROC NATL ACAD SCI U S A, vol. 103, 2006, pages 17331 - 6 |
| YU ET AL., PROC NATL ACAD SCI USA, vol. 103, 2006, pages 17331 - 6 |
| YU ET AL., PROC NATL ACAD SCI USA, vol. 104, 2007, pages 8924 - 9 |
| ZHAO ET AL., J. BIOL. CHEM., vol. 270, no. 11, 1995, pages 6081 - 6087 |
Cited By (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2012037324A3 (en) * | 2010-09-15 | 2012-05-18 | Metabolix, Inc. | Increasing carbon flow for polyhydroxybutyrate production in biomass crops |
| US10316330B2 (en) | 2012-05-08 | 2019-06-11 | Monsanto Technology Llc | Corn event MON 87411 |
| US11859198B2 (en) | 2012-05-08 | 2024-01-02 | Monsanto Technology Llc | Corn event MON 87411 |
| US11788099B2 (en) | 2012-05-08 | 2023-10-17 | Monsanto Technology Llc | Corn event MON 87411 |
| US11414672B2 (en) | 2012-05-08 | 2022-08-16 | Monsanto Technology Llc | Corn event MON 87411 |
| CN102864167A (en) * | 2012-09-26 | 2013-01-09 | 浙江大学 | Plant expression carrier and method for cultivating low-phytic-acid rice |
| CN102864167B (en) * | 2012-09-26 | 2014-04-09 | 浙江大学 | Plant expression carrier and method for cultivating low-phytic-acid rice |
| EP3702364A1 (en) | 2012-12-18 | 2020-09-02 | Yield10 Bioscience, Inc. | Transcriptional regulation for improved plant productivity |
| WO2014100289A1 (en) | 2012-12-18 | 2014-06-26 | Metabolix, Inc. | Transcriptional regulation for improved plant productivity |
| WO2014127053A2 (en) | 2013-02-13 | 2014-08-21 | Metabolix, Inc. | Process for ultra pure chemical production from biobased raw starting materials |
| JP2014193154A (en) * | 2013-02-28 | 2014-10-09 | Euglena Co Ltd | Transformant of euglena |
| JP2014193153A (en) * | 2013-02-28 | 2014-10-09 | Euglena Co Ltd | Method for introducing gene into euglena |
| WO2014133159A1 (en) * | 2013-02-28 | 2014-09-04 | 株式会社ユーグレナ | Method for introducing gene to euglena, and transformant therefrom |
Also Published As
| Publication number | Publication date |
|---|---|
| US9181559B2 (en) | 2015-11-10 |
| CA2773707A1 (en) | 2011-03-24 |
| US20120174253A1 (en) | 2012-07-05 |
| AU2010295637B2 (en) | 2013-12-12 |
| AU2010295638A1 (en) | 2012-04-26 |
| BR112012005591A2 (en) | 2015-09-01 |
| EP2477477A1 (en) | 2012-07-25 |
| WO2011034946A9 (en) | 2011-05-19 |
| WO2011034945A1 (en) | 2011-03-24 |
| CA2773703A1 (en) | 2011-03-24 |
| EP2478105A1 (en) | 2012-07-25 |
| BR112012005592A2 (en) | 2017-05-02 |
| US20120180162A1 (en) | 2012-07-12 |
| AU2010295637A1 (en) | 2012-04-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| WO2011034946A1 (en) | Generation of high polyhydroxybutrate producing oilseeds | |
| AU770120B2 (en) | Plant multi-gene expression constructs | |
| US11965182B2 (en) | Plants with enhanced yield and methods of construction | |
| WO2005063988A1 (en) | Alteration of oil traits in plants | |
| US8487159B2 (en) | Production of polyhydroxybutyrate in switchgrass | |
| JP2009291204A (en) | Modification of fatty acid metabolism in plant | |
| US12305179B2 (en) | Transgenic land plants that express a polyhydroxyalkanoate synthase seed specifically with cytosolic localization | |
| US7176349B1 (en) | Production of polyhydroxyalkanoate in plants | |
| CN102295691B (en) | BCCP2 gene and application on enhancing lipid content of plant and algae thereof | |
| US20210180078A1 (en) | Transgenic rice plants overexpressing acyl-coa-binding protein2 show enhanced grain size | |
| CA2754261A1 (en) | Stable, fertile, high polyhydroxyalkanoate producing plants and methods of producing them | |
| AU2010295638B2 (en) | Generation of high polyhydroxybutrate producing oilseeds | |
| US7341856B2 (en) | Production of polyhydroxyalkanoate in plants | |
| AU2020293986B2 (en) | Transgenic land plants that express a polyhydroxyalkanoate synthase seed specifically with cytosolic localization | |
| US7129395B2 (en) | OAR polynucleotides, polypeptides and their use in PHA production in plants | |
| CA3141340C (en) | Transgenic land plants that express a polyhydroxyalkanoate synthase seed specifically with cytosolic localization | |
| CA3170510A1 (en) | Genetically engineered plants that express 6-phosphogluconate dehydratase and/or 2-keto-3-deoxy-6-phosphogluconate aldolase |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 10757363 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2773707 Country of ref document: CA |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 13395702 Country of ref document: US |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2010295638 Country of ref document: AU |
|
| REEP | Request for entry into the european phase |
Ref document number: 2010757363 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2010757363 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: 2010295638 Country of ref document: AU Date of ref document: 20100915 Kind code of ref document: A |
|
| REG | Reference to national code |
Ref country code: BR Ref legal event code: B01A Ref document number: 112012005591 Country of ref document: BR |
|
| ENP | Entry into the national phase |
Ref document number: 112012005591 Country of ref document: BR Kind code of ref document: A2 Effective date: 20120313 |