WO2014065572A1 - ENOblock을 유효성분으로 포함하는 암 예방 또는 치료용 약제학적 조성물 - Google Patents
ENOblock을 유효성분으로 포함하는 암 예방 또는 치료용 약제학적 조성물 Download PDFInfo
- Publication number
- WO2014065572A1 WO2014065572A1 PCT/KR2013/009438 KR2013009438W WO2014065572A1 WO 2014065572 A1 WO2014065572 A1 WO 2014065572A1 KR 2013009438 W KR2013009438 W KR 2013009438W WO 2014065572 A1 WO2014065572 A1 WO 2014065572A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- cancer
- integer
- enoblock
- enolase
- composition
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/33—Heterocyclic compounds
- A61K31/395—Heterocyclic compounds having nitrogen as a ring hetero atom, e.g. guanethidine or rifamycins
- A61K31/53—Heterocyclic compounds having nitrogen as a ring hetero atom, e.g. guanethidine or rifamycins having six-membered rings with three nitrogens as the only ring hetero atoms, e.g. chlorazanil, melamine
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
Definitions
- Cancer prevention or therapeutic pharmaceutical composition comprising ENOblock as an active ingredient
- the present invention is made by the task number NRF-2012-003460 under the support of the Ministry of Education, Science and Technology, the research management specialized organization of the project is the Korea Research Foundation, the research project name is "general researcher support project / young researcher support project”, research The title of the project is "Exploration and Characterization of Insulin-Low-Operating Low Molecular Weight Compounds for the Development of Diabetic Therapeutics".
- the Organizing Agency is Gwangju Institute of Science and Technology.
- the present invention was made by the task number NRF-2012-000462 under the support of the Ministry of Education, Science and Technology, the research management professional organization of the task is the Korea Research Foundation, the research project name is "general researcher support project / women scientists support project” , The name of research project is “New anticancer mechanism and anticancer target discovery by control of cancer microenvironment”.
- the host institution is Gwangju Institute of Science and Technology and the research period is 2012.05.01 ⁇ 2015.04.30
- the present invention was made by the task number A120326 under the support of the Ministry of Health and Welfare, the research and management institution of the task is the Korea Health Industry criminal Institute, the research project name is "health medical research and development project”, the research project name is "small molecule differentiation Production of patient-derived iPSCs and their use in the treatment of myocardial infarction cells using regulators, the Gwangju Institute of Science and Technology, and the period of study are 2012.08.01 ⁇ 2014.07.31.
- This patent application claims priority to Korean Patent Application No. 10-2012-0117417 filed with the Korean Patent Office on October 22, 2012, the disclosure of which is hereby incorporated by reference.
- the present invention relates to an ENOblock, which is a non-substrate analogue having enolase inhibitory activity, and a pharmaceutical composition for preventing or treating cancer comprising the same.
- Enolase is an important component of the glycolysis pathway. Important for various intracellular processes not related to glycolytic function
- ENOblock is the first reported enolase inhibitor suitable for biological assays and an attractive molecule for drug development in cancer and diabetes.
- Glycolysis is an evolutionarily very well conserved metabolic pathway that converts one glucose molecule into two pyruvates. Free energy is released and used to form high energy-containing molecules, adenosine triphosphate (ATP) and reduced NADHCnicotinamide adenine dinucleotide (ATP).
- ATP adenosine triphosphate
- ATP NADHCnicotinamide adenine dinucleotide
- the glycolysis consists of 10 biochemical reactions, each stage being catalyzed by a different enzyme. Over the past two decades, there has been a surprising understanding of the complex functions that function in the various cellular processes of enzymes involved in glycolysis (1).
- Cancer cells with the Warburg effect shows an increased glycolysis to produce ATP, a known phenomenon (2).
- the metabolic modification is the fundamental difference between cancer cells and normal cells, which provides a therapeutic strategy for the selective killing of cancer cells using glycolysis inhibitors (3).
- glycolysis inhibitors more effectively induce cancer cell death in the hypoxic environment occurring in the tumors formed (4).
- the hypoxic environment described above makes cancer cells less susceptible to anticancer agents such as cytarabine and doxorubicin.
- ENOblock a triazine compound
- Another object of the present invention is to provide a method for preventing or treating cancer.
- the present invention is (a) a therapeutically effective amount of a triazine-based compound represented by the formula (I); And (b) a pharmaceutically acceptable carrier, the pharmaceutical composition for cancer prevention or treatment comprising:
- 3 ⁇ 4 is H or d-Cs straight or branched alkyl
- R 2 is H, dC 5 straight or branched alkyl, CC 5 straight or branched alkyl alcohol,-[(CH 2 ) ra — 0] n- (CH 2 ) p -N3 ⁇ 4 (m, n and p Are each an integer of 1-10,-[(C3 ⁇ 4) ra -0] n -CH 3 (where m and n are each an integer of 1 ⁇ 10),-[(CH 2 ) ra -0] n- (CH 2 ) p -C3 ⁇ 4 (where m, n and p are each an integer of 1-10), ⁇ (CH 2 ) q- (C0NH) -d- 5 linear or branched alkyl (where q is 0- Is an integer of 5),-(CH z ) q- (CONlD-ds linear or branched alkyl alcohol
- the present invention provides a pharmaceutical composition for preventing or treating enolase-associated disorders comprising the above-mentioned composition as an active ingredient.
- the present inventors have tried to isolate / identify small molecules that can inhibit cancer cell survival in the hypoxic state.
- the present inventors directly bind the triazine-based compound named ENOblock to enolase to inhibit its activity more effectively in the hypoxic state than in the normal oxygen state, and the compound inhibits the migration, metastasis and invasion of cancer cells.
- the cells induce glucose uptake, down-regulate PEPCK expression, and inhibit adipocyte differentiation and foam cell formation.
- the active ingredient AP-III-a4 (named 'ENOblock') is a triazine-based compound, and the inventors of the present invention from a tagged library containing 384 tagged triazine-based compounds. It is a compound identified as a small molecular compound that more effectively and specifically induces cancer cell death in the hypoxic state using the screening system established by Methods for synthesizing triazine compound libraries and their uses used in the present invention are described in International Patent Applications Publications WO 03/032903 and WO 03/050237, which are incorporated herein by reference.
- d-Cs straight or branched alkyl in the general formula (I) of the present invention refers to methylamine, ethylamine, n-propylamine, isopropylamine, n-butylamine, sec-butylamine, tert-butylamine, n- Amyl, tert-amyl, ' straight chain (branched chain) or branched (branched chain) alkyl group of the nuclear chamber.
- H ( 2 : alkyl, more preferably H.
- R 2 in formula (I) of the present invention is H, dC 5 linear or branched alkyl, dC 5 linear or branched alkylalcohol, one [(C3 ⁇ 4) m — 0] n- (CH 2 ) p -NH 2 (where m, n and are each an integer of 1 ⁇ 10),-[(C3 ⁇ 4) m -0] n — C3 ⁇ 4 (where m and n are each an integer of 1-10) ,-[(CH 2 ) m -0] n- (CH 2 ) p -CH 3 (where m, n and p are each an integer of 1-10),-(CH 2 ) q- (C0NH) -Ci -5 linear or branched alkyl (where q is an integer of 0-5),-(CH 2 ) q- (C0NH) -Ci- 5 linear or branched alkylalcohol (where q is an integer of 0-5),
- the triazine-based compound represented by Formula (I) includes a compound represented by Formula ( ⁇ ):
- the present invention is the first report for small molecules (ENOblock), which are non-substrate analogs that directly bind to and inhibit activity of enolase.
- the ENOblock of the present invention inhibited the dose-dependently specifically the activity of enolase in a normal or hypoxic state, thereby inhibiting cancer cell migration, metastasis and invasion (see FIG. 1 to 3).
- the above-described effects showed even better efficacy in a low oxygen state. Therefore, the ENOblock of the present invention can be effectively applied to the treatment of cancer or enolase-related diseases through inhibition of enolase activity.
- the composition of the present invention inhibits the activity of enolase and inhibits the migration, invasion and metastasis of cancer cells.
- the above inhibitory effect by the ENOblock of the present invention is normal oxygen It is promoted more in hypoxia than in normoxia.
- the composition of the present invention reduces the expression of apoptosis-inducible proteins and more preferably reduces the expression of AKT or Bcl-xL proteins.
- cancer that can be prevented or treated by the composition of the present invention is brain cancer, neuroendocrine cancer, gastric cancer lung cancer, breast cancer, ovarian cancer, liver cancer, bronchial cancer, nasopharyngeal cancer, laryngeal cancer, pancreatic cancer, bladder cancer Adrenal cancer, colon cancer, colon cancer cervical cancer, prostate cancer, bone cancer, skin cancer, thyroid cancer, parathyroid cancer and ureter cancers.
- Symptoms of the above-mentioned cancers include breast lumps, nipple changes, breast cysts, breast pain, death weight loss, weakness, excessive fatigue, difficulty eating and breathing, loss of appetite, chronic sneezing, blood loss, hematuria, bloody stools, motion sickness and vomiting.
- Lung metastasis Bone metastasis, colon metastasis, bladder metastasis, kidney metastasis, pancreatic metastasis, abdominal distension, bloating, intraperitoneal fluid, vaginal bleeding, constipation, colon perforation, acute peritonitis, pain, excessive sweating , Fever, high blood pressure, anemia, diarrhea, jaundice, dizziness, chills or muscle spasms.
- enolase-associated disorders refers to a condition in which overexpression of enolase is induced, and more preferably to a disease induced in a hypoxic.state rather than a normal oxygen state. Meaning, including but not limited to cancers, autoimmune disorders, ischemia and bacterial infections.
- composition of the present invention can also be used as a composition for preventing or treating diabetes.
- the composition of the present invention has an insulin-mimicking activity.
- the above-described insulin mimicking activity by the composition of the present invention includes promoting intracellular glucose uptake.
- the composition of the present invention is glucose It down-regulates the expression of PEPCK (hosphoeno 1 pyru uva te carboxykinase), an important protein for gluconeogenesis.
- PEPCK hosphoeno 1 pyru uva te carboxykinase
- the composition of the present invention inhibits adipogenesis and foam cell formation.
- pharmaceutically effective amount in this specification means an amount sufficient to achieve the efficacy or activity (eg, anti-cancer agent, anti-diabetic agent, etc.) of the triazine-based compound described above.
- the pharmaceutical composition of the present invention comprises a pharmaceutically acceptable carrier.
- Pharmaceutically acceptable carriers included in the pharmaceutical composition of the present invention are commonly used in the preparation, lactose, dextrose, sucrose, sorbbi, manny, starch, acacia rubber, calcium phosphate, alginate, Gelatin, calcium silicate, microcrystalline cellulose, polyvinylpyrrolidone, cellulose, water, syrup, methyl cellulose, methylhydroxybenzoate, propylhydroxybenzoate, talc, magnesium stearate and mineral oil Including, but not limited to.
- the pharmaceutical composition of the present invention may further include a lubricant, a humectant, a sweetener, a flavoring agent, an emulsifier, a suspension agent, a preservative, and the like, in addition to the above components.
- a lubricant e.g., talc, kaolin, kaolin, kaolin, kaolin, kaolin, kaolin, kaolin, kaolin, kaolin, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, sorbitol, mannitol, mannitol, mannitol, mannitol, mannitol, mannitol, mannitol, mann
- Appropriate dosages of the pharmaceutical compositions of the present invention may vary depending on factors such as formulation method, mode of administration, age, weight, sex, morbidity, food, time of administration, route of administration, rate of excretion and reaction response to the patient. Can be.
- the oral dosage of the pharmaceutical composition of the present invention is preferably 0.001-100 mg / kg (body weight) per day.
- the pharmaceutical composition of the present invention may be administered orally or parenterally, and when administered parenterally, may be administered by intravenous injection, subcutaneous injection, intramuscular injection, intraperitoneal injection, transdermal administration, or the like. It is preferable that the route of administration of the pharmaceutical composition of the present invention is determined according to the type of the disease to be applied.
- the concentration of the triazine-based compound of formula I, which is an active ingredient included in the composition of the present invention, may be determined in consideration of the purpose of treatment, the condition of the patient, the period of time required, and the like, and is not limited to a specific range of concentration.
- the pharmaceutical compositions of the present invention may be prepared in unit dosage form by formulating with a pharmaceutically acceptable carrier and / or excipient according to methods which can be easily carried out by those skilled in the art. Or may be prepared by incorporating into a multi-dose container.
- the formulation may be in the form of a solution, suspension or emulsion in an oil or aqueous medium, or may be in the form of axes, powders, granules, tablets or capsules, and may further include a dispersant or stabilizer.
- the present invention provides a pharmaceutical composition comprising (a) a therapeutically effective amount of a triazine-based compound represented by formula (I); And (b) administering to the subject a composition comprising a pharmaceutically acceptable carrier, wherein the method for preventing or treating cancer comprises:
- 3 ⁇ 4 is H or dC 5 straight or branched alkyl
- 3 ⁇ 4 is H, Ci-C 5 straight or branched alkyl, d-Cs straight or branched alkyl alcohol,-[(CH 2 ) m -0] n — (C3 ⁇ 4) p -N3 ⁇ 4 (m, n and p are each an integer of 1-10),-[(CH 2 ) m -0] n -CH 3 (where m and n are each an integer of 1-10),-[(CH 2 ) ra -0] n- (CH 2 ) p -CH 3 (wherein m, ⁇ and p are each an integer of 1-10),-(C3 ⁇ 4) q- (C0NH) -Ci- 5 linear or branched alkyl (where q is Is an integer of 0-5),-(C3 ⁇ 4) q — (C0NH) -Ci- 5 linear or branched alkyl alcohol (where Q is an integer of 5),-(CH 2 ) q- (C0
- the present invention is the surprising inhibition ratio having activity related disorder (enolase-associated) a medicament for the prevention or therapeutic It relates to a composition.
- the ENOblock of the present invention binds directly to enolase and inhibits its activity, which is more effective in the hypoxic state than normoxia.
- the ENOblock of the present invention inhibits the migration, metastasis and invasion of cancer cells.
- the ENOblock of the present invention induces glucose uptake in cells and down-regulates PEPCK expression.
- the ENOblock of the present invention inhibits adipogenesis and foam cell format ion.
- composition comprising the ENOblock of the present invention can be very effectively applied to the prevention or treatment of cancer or enolase-associated disease.
- ENOblock AP-III-a4 compound
- HCT116 cancer cells were dispensed into parallel 96-well culture plates. Hypoxic states were induced in one plate via 150 mM cobalt chloride (CoCl 2 ) treatment.
- Triazine compound library (5 mM) was added after 4 hours and cell death was Irradiation was performed 24 hours after hypoxia induction. 'Hit' compounds induced 25% or more cell death in the hypoxic state compared to normoxia (measured as MTT assay absorbance).
- Lb shows the chemical structure of AP-III-a4 (EN0block) of the present invention, the control compounds AP-IV-e3 and ⁇ -1-flO, GAPDS targeting GAPDH, and the well-known anti-diabetic drug rosiglitazone Figure showing.
- Lc shows that ENOblock significantly induces HCT116 colon cancer cell death in hypoxic state (Hypox) compared to normal oxygen state (Norm).
- error standard deviation (SD); *, Value indicating increased cell death compared to normal oxygen status ⁇ 0.05.
- FIG. ID shows the representative phase contrast microscopy images of HCT116 colon cancer cells treated in normal or hypoxic state.
- Figure 2 is a result that indicates that the 4-time processing of 150 ⁇ CoCl 2 in HCT116 cancer cells induced hypoxia, hypoxia-shows the increased expression of the surprising anti-male gene of claim.
- the other proteins contained in the process path (glyceraldehycle 3-phosphate dehydrogenase) GAPDH is a previous report (e. G., Said, et al, BMC Mol Biol 8:.. 55 (2007)) and "match enough low oxygen There was no increased expression even after the condition was induced.
- the red pair indicates the fold—change of expression calculated by densitometry analysis.
- 3 is a diagram schematically illustrating synthesis of an AP linker library.
- 3B shows a linker.
- Figure 3c shows the amines and amino alcohols used for the construction of building bltok I Showing results.
- Figure ID shows the
- 4 is a result showing that the AP-III-a4 compound (ENOblock) of the present invention is a direct inhibitor of enolase.
- ENOblock AP-III-a4 compound
- 4B shows the sequence of the enolase monomer subunit. Identified peptides are shown in red.
- 4C is a Western blot analysis confirming that enolase in HCT116 cancer cell lysate binds to the ENOblock affinity matrix.
- Free as a competitor Competitive analysis using EN0block (denoted abbreviated EN0) completely inhibited enolase binding to the ENOblock affinity matrix.
- 4D is a Western blot analysis showing that purified human enolase binds to the ENOblock affinity matrix.
- error standard deviation (SD); * , ⁇ Value ⁇ 0.05 compared to untreated group.
- FIG. 5 is a result showing that ⁇ -1-flO, a 'non-hit' compound obtained from the same triazine compound library used for ENOblock identification, does not significantly inhibit enolase activity.
- ENOblock was used as a positive control.
- Abbreviation: error standard deviation (SD); *, 7 ° value compared to non-treated group ⁇ 0.05.
- FIG. 6 is a result showing that siRNA-mediated knock-down of enolase expression in HCT116 cancer cells increases susceptibility to hypoxic states.
- Cells were treated with varying concentrations of enolase (EN01) siRNA or two siRNAs as negative controls: negative control siRNAs; (a) 80 pmols of scrambled siRNA or (b) 80 pmols of p57 (cyclin dependent kinase inhibitor) siRNA. After 24 hours of siRNA transfection, cells were transferred to 96-well culture plates at a density of 10 4 cells / well and incubated for 24 hours in normal oxygen or hypoxia induced by 150 mM CoCl 2 treatment. Cytotoxicity was assessed by MTT assay after 24 hours.
- FIG. 7 is a result showing that ENOblock can inhibit cancer cell invasion and migration (migrat ion).
- FIG. 7A shows that treatment of ENOblcok with HCT116 cancer cells in normal oxygen conditions inhibits invasion dependently.
- ENOblock treatment inhibited cell infiltration much more effectively than LY294002 (inhibition of phosphoinositide 3-kinase signaling), a small molecule inhibitor to previously known infiltration.
- error standard deviation (SD); * , P value ⁇ 0.05 compared to untreated group.
- FIG. 7A shows that treatment of ENOblcok with HCT116 cancer cells in normal oxygen conditions inhibits invasion dependently.
- ENOblock treatment inhibited cell infiltration much more effectively than LY294002 (inhibition of phosphoinositide 3-kin
- 3E shows the treatment of ENOblock against ENOblock treatment of HCT116 cancer cells.
- Results show increased sensitivity to anti-tubulin chemotherapeutic agents, taxol and vincristine, 10 nM Taxol and 10 nM bins in cells with or without 10 mM ENOblock.
- Christine was abbreviated: Error standard deviation (SD); *, values between groups indicated in the graph ⁇ 0.05.
- SD Error standard deviation
- Figure 7F shows that the treatment of ENOblock against ENOblock treatment of HCT116 cancer cells showed expression of AKT and Bcl-Xl, which are negative regulators of apoptosis.
- the results showed that the cells were treated with ENOblock for 24 hours for AKT Western Blot analysis; the cells were treated with ENOblock for 48 hours for Bcl-Xl Western Blot analysis.
- FIG. 8 is a result showing photographs of crystal vial-stained trans 3 ⁇ 4 inserts showing that treatment of AP-I—flO compounds (10 ⁇ ) did not reduce HCT116 cancer cell infiltration.
- Treatment of LY294002 (5 ⁇ ) was used as a positive control and showed a significant reduction in infiltrated cancer cells.
- 9 shows the in vivo assay results for toxicological study and anti-cancer activity of ENOblock.
- 9a is zebrafish pom Results of toxicity analysis of ENOblock treatment using the zebrafish larvae system. Measurement of various developmental parameters confirmed that the 10 ⁇ ENOblock dose was not harmful to fry, but the 20 ⁇ ENOblock dose inhibited zebrafish's swimming ability.
- FIG. 8 is a result showing photographs of crystal vial-stained trans 3 ⁇ 4 inserts showing that treatment of AP-I—flO compounds (10 ⁇ ) did not reduce HCT116 cancer cell infiltration.
- Treatment of LY294002 (5 ⁇ ) was used as
- FIG. 9B shows microscopic: observation results of Sebrafish fry at 72 days post fertilization (72 days post fertilization, 72 dfp) with increasing ENOblock dose. As a result, it was observed that the dose of 20 ⁇ ENOblock caused abnormally large swim (red arrow). Doses of 40 ⁇ ENOblock resulted in numerous abnormal phenotypes in the fry. FIG. 9C shows that the dose of 20 ⁇ or 40 ⁇ ENOblock also reduced zebrafish overall larvae viability while the dose of 10 ⁇ ENOblock did not affect viability.
- error standard deviation (SD); *, Value for change in survival compared to DMS0-treated group ⁇ 0.05.
- FIG. 10 is the result that means that ENOblock induces glucose uptake and inhibits PEPCK expression.
- 10A is a result showing that 24-hour treatment of 10 ⁇ ENOblock (abbreviated EN0) in Huh7 hepatocytes or HEK kidney cells induces glucose uptake as measured using the fluorescent glucose probe 2—NBDG. .
- 24-hour treatment of GAPDH, a small molecule regulator of the glycolytic enzyme GAPDH) could also induce glucose uptake in hepatocytes.
- well-known anti-diabetic drugs 24-hour treatment of rosiglitazone (abbreviated R0SI; 10 ⁇ ) could induce glucose uptake in hepatocytes, but not glucose intake in renal cells.
- FIG. 10B shows that 24-hour treatment of EN0block (10 ⁇ ) in Huh7 hepatocytes inhibits expression of PEPCK, a key positive regulator of glucose gluconeogenesis. 10 24 hours of treatment ⁇ rosiglitazone could also, to suppress the expression of PEPCK. However, 24-hour treatment of 10 ⁇ GAPDS did not inhibit PEPCK expression.
- FIG. 10D shows that G6Pase (glucose 6-phosphatase), another enzyme that regulates glucose synthetase, or 24 'treatment of BN0block (10 ⁇ ) in hepatocytes or AMPK (5' AMP-activated protein kinase) that regulates energy homeostasis The results show that they do not affect expression. Similarly, 24-hour treatment of 10 ⁇ GAPDS or 10 ⁇ rosiglitazone did not affect the expression of the enzymes described above.
- error standard deviation (SD).
- FIG. 11 shows that siRNA-mediated knock-down of enolase expression in hepatocytes or kidney cells induces glucose uptake, as evidenced by increasing labeling of the fluorescent glucose probe 2-NBDG.
- Cells were treated with varying concentrations of enolase (EN01) siRNA or two siRNAs as negative controls: negative control siRNAs; (a) 80 pmols of scrambled siRNA or (b) 80 pmols cyclin dependent kinase inhibitor (p57) siRNA. After 24 hours of siRNA transfection, cells were transferred to 96-well culture plates at a density of 10 4 cells / well and treated with 100 uM 2-NBDG for 30 minutes. Thereafter, 2-NBDG fluorescence measurement was performed as described in the experimental method herein.
- FIG. 12 shows that ENOblock can inhibit PEPCK expression and induce glucose uptake in phosphorus.
- 12A is a schematic representation of our approach to measuring the effect of ENOblock treatment on PEPCK expression in zebrafish.
- 12B is a result showing that 3-hour treatment of EN0block (10 ⁇ ) inhibits PEPCK expression in liver tissues of zebrafish.
- error standard deviation (SD); *, Value for PEPCK expression results compared to non-treated group ⁇ 0.05.
- FIG. 12C shows our approach to measure the effect of ENOblock treatment on ingestion of fluorescence-tagged glucose bioprobe (2-NBDG) in zebrafish larvae (easily observable due to transparency of the pom) Diagrammatically). 12D is the result showing that 4-hour treatment of ENOblock (10 ⁇ ) increases glucose uptake in zebrafish fry. Increased glucose uptake can be observed through developing embryos, particularly the eye (white arrow), small intestine and egg yolk sac.
- zebrafish treated with known anti-diabetic natural product, emodine (10 yg / mL), which promotes glucose intake increased glucose uptake in a similar but stronger pattern to that of ENOblock-treated zebrafish.
- emodine 10 yg / mL
- FIG. 13 shows that 1 hour treatment of ENOblockCLO ⁇ ) in 72 hpf zebrafish induces glucose uptake, and 2-NBDG signals were measured using a fluorescent plate reader in the dissolved fry.
- error standard deviation (SD); *, ⁇ Values for increased fluorescence detection results compared to NBDG alone-treated group.
- FIG. 14 shows that ENOblock inhibits adipocyte differentiation (3 ⁇ 1> (61 ⁇ 3) and foam cell formation.
- FIG. 14A (i) shows progenitor cells undergoing adipocyte differentiation.
- error standard deviation (SD); *, ⁇ value for lipid accumulation results compared to non-treated adipocytes ⁇ 0.05.
- FIG. 14B shows microscopic analysis of macrophages treated with oleic acid. Treatment with 10 ⁇ ENOblock inhibited foam cell formation as indicated by the reduction of Oil Red 0 staining. 14C is a quantitative graph showing that ENOblock treatment (10 ⁇ ) inhibits foam cell formation, as shown by counting cells showing lipid accumulation .. In contrast, macrophages developing into foam cells. Anti-diabetic drug rosiglitazone (10 ⁇ ) did not affect the number of cells showing lipid accumulation.
- FIG. 14D is a result showing that treatment of EN0block (10 ⁇ ) to monocytes through counting the cells and numbers attached to the culture dish inhibits differentiation into macrophages. In contrast, treatment of rosiglitazone (10 ⁇ ) with monocytes did not affect differentiation into macrophages.
- error standard deviation (SD); *, ⁇ value ⁇ 0.05 for counting the reduced number of attached cells compared to ⁇ -treated groups.
- FIG. 14E is a graph showing that ENOblock treatment effects on macrophages are not due to cytotoxicity, as can be seen in the ⁇ assay assay.
- 48 h treatment of 3 ⁇ 40 2 (3 mM) was used as a positive control.
- error standard deviation (SD); *, Value for reduced absorbance (570 nm) result compared to untreated group ⁇ 0.05.
- FIG. 15 is an immunoblot analysis showing enolase expression knock-down through siRNA treatment in HEK kidney cells and Huh7 hepatocytes. Cells were transfected with 80 pmols of siRNAs for 48 hours.
- the tagged triazine compound library is based on previously reported procedures. Was prepared (FIG. 1; 5). The purity of the synthesized compounds was determined by analytical HPUX Agilent Technologies, Snata Clara, USA. Screening for Apoptosis Inducers Maintaining Efficiency Under Hypoxic Conditions
- HCT116 colon carcinoma cells were dispensed into two sets of 96-well plates at a density of 5 ⁇ 10 3 cells per well.
- the hypoxic state was induced in a set of 96 microwell plates via treatment with 0.22 ⁇ -filtered 150 ⁇ CoCl 2 (Sigma) and the volume of the culture medium (49) was 50% (per well) as described previously. Reduced to 100 L).
- test compounds from the tagged triazine compound library (5) were screened at a concentration of 5 ⁇ in two independent wells. Cytotoxicity was examined via ⁇ assay 24 hours after drug treatment. For further analysis, 'hit' was classified as compounds exhibiting 25% or more cytotoxicity (measured as MTT assay absorbance) in the hypoxic state as compared to normoxia. 2-DG (50 ⁇ M; 50), a glycolysis inhibitor that selectively kills cancer cells in the hypoxic state, was used as a positive control.
- Enolase expression was investigated to verify CoCl 2 treatment-induced hypoxic conditions. Enolase expression is known to increase in response to hypoxia (51). Four hour back treatment of 150 yM CoCl 2 on HCT116 carcinoma cells induced enolase expression (FIG. 2). Enolase Activity Assays
- Enolase purified from rabbit muscle was purchased from Sigma-Aldrich.
- One unit of enolase is an enzyme that produces 1 umol of phosphoroenol pyruvate from phospho-D-glycerate (Sigma) per minute in a standard assay. Defined as the quantity (52).
- Enolase activity assays include 50 mM imidazole-HCl (pH 6.8) (JUNSEI), 2.0 mM MgS0 4 (JU SEI) and 400 mM KCKJUNSEI) in the absence or presence of ENOblock or NaF (Sigma). Purified enolase (3-9 U) in 37T was performed by reaction at 37T.
- reaction was initiated by the addition of 1 urn of 2-phospho-D-glycerate (2-phospho- glycerate), the 0D value being the spectrophotometer after 10 minutes of reaction time (VERSA max, Molecular Devices, USA) at 240 nm.
- Zebrafish tumor cell xenograft model 2-phospho-D-glycerate (2-phospho- glycerate), the 0D value being the spectrophotometer after 10 minutes of reaction time (VERSA max, Molecular Devices, USA) at 240 nm.
- Zebrafish tumor cell xenograft model 2-phospho- glycerate
- Zebrafish (Lotte Mart, Gwangju, Republic of Korea) was maintained according to standard protocols (53). The management and treatment of zebrafish was carried out in accordance with the guidelines established by Animal Care and Ethics Committees of Gwangju Institute of Science and Technology (Korea). Zebrafish embryos were obtained using standard mating conditions and differentiated up to 48 hours post fertilization for cell xenografts (ceil ⁇ 131 3 011). After staining of cancer cells, embryos were de-chor ionized using micro-pliers and anesthetized with 0.0016% tr icaine (Sigma), followed by 1.0% wet agar. The right side was placed on an Osvitrogen.
- Tumor cells were separated from the culture dish with 0.05% trypsin-EDTA (Gibco) and washed twice with PBS (Sigma) at room temperature. Cells were stained with Dil (2 ug / ml; Invitrogen) diluted in PBS and washed four times: once with FBS (Invitrogen); Twice with PBS; And once with FBS (103 ⁇ 4) diluted in PBS. The cells were then placed on ice before injection. Cancer cells were counted by microscope (OLYMPUS CKX41) and resuspended in 10% FBS, of which 100 cells were infused with borosilicate glass capillaries (PV820 pneumatic icopump, World Precision Instruments). Infused into the center of yolk sac.
- the injected embryos were transferred to 96-well plates (1 embryo / well) containing the desired drug diluted in 200 mL of methylene blue free E3 medium and maintained at pre-selected incubation temperature. 4 days after injection, The number of embryos showing the dissemination of cancer cells from the injection site was counted and photographed with an upright microscope (Leica DM2500 microscope, Germany). Embryos that counted the number of migrated cells and showed at least 5 fluorescent microfoci distants from the yolk sac were scored for cell dissemination. Measurement of Glucose Intake in Zebrafish
- ENOblock binds to and inhibits enolase
- ENOblock The binding of ENOblock to enolase in cancer cell lysates was again confirmed by Western blot analysis of the proteins eluted from the ENOblock affinity matrix.
- Competitive analysis using free ENOblock has shown to inhibit enolase binding to the ENOblock affinity matrix (FIG. 4C).
- ENOblock could bind to purified human enolase, which means a direct interaction between ENOblock and enolase (FIG. 4D).
- FIG. 4E Subsequent analysis revealed that ENOblock Dependently, it was confirmed that enolase: activity could be inhibited (FIG. 4E).
- AP-I—? 0 (3) a Daron non-heat compound obtained from a tagged triazine compound library as an additional control.
- Enolase is a metabolic protein with a 'moonlighting' function that performs many functions that are not related to intrinsic glycolytic function (6).
- ENOblock means a powerful chemical means that characterizes the collateral functions of enolase.
- FIG. 7 we investigated the role of enolase in cancer cell progression (FIG. 7).
- enolase inhibition by ENOblock can reduce cancer cell invasion, which was the first confirmation to our knowledge that enolase activity is linked to metastasis (FIG. 7A-). 7b).
- the inventors tested ⁇ - ⁇ - ⁇ 0 (3), another compound obtained from a library of tagged triazine compounds as an additional control. The compound was found not to reduce cell invasion (FIG.
- ENOblock treatment also inhibited cancer cell migration (FIGS. 7C-7D). ENOblock treatment reduced cancer cell infiltration / migration in normal oxygen at concentrations that did not cause cytotoxicity (see, FIGS. 7A-7B and lc).
- Previous studies have reported that knock-down of enolase expression can increase cytotoxicity caused by the anticancer drugs taxol and vincristine (7). In line with this, we have found that ENOblock treatment can also increase cancer cytotoxicity induced by Taxol and vincristine (FIG. 7E). To investigate how ENOblock induces cancer cytotoxicol, we measured two protein expressions (AKT (8) Bcl-xL (9)) linked to induction of apoptosis.
- ENOblock treatment reduced the expression of AKT and Bcl-xL proteins (FIG. 7F).
- Zebrafish G3 ⁇ 4 / 7 / i? rerio) Cancer cell xenograft models are an effective and convenient means used to gain increasing research results in the testing of anticancer drug candidates in phosphorus (10, 11).
- zebrafish is a suitable vertebrate platform for predicting toxicological effects in mammals (12).
- treatment of 10 ⁇ M ENOblock with developing zebrafish fry was non-toxic (FIGS. 9A-9C).
- the use of a recently published zebrafish tumor xenograft model was effective for anti-cancer drug testing (10).
- ENOblock treatment reduces cancer cell seeding, which means that ENOblock inhibits cancer cell migration and invasion processes (FIG. 9 (1)).
- ENOblock induces intracellular glucose uptake and down-regulates PEPCK expression
- ENOblock (Compound AP-III-a4) was one of the triazine compounds previously identified through screening to find new regulators of glucose uptake, although the mechanism of action was not identified (13 ).
- 2-NBDG (2- (N- (7—nitrobenz-2-oxa—1,3-diaz 1-4-), a fluorescent probe that can be used to monitor intracellular glucose flux.
- y 1) am i no) -2-deoxyg 1 ucose; 14) to confirm the activity of ENOblock to increase glucose uptake in cells (FIG. 10A).
- the identification of the present invention that regulation of enolase function is linked to increased glucose uptake is the first proof.
- the role of enolase in promoting cellular glucose uptake was reaffirmed by siRNA-mediated enolase expression knock-down (FIG. 11).
- ENOblock downregulates the expression of phosphoeno 1 pyru uvat e carboxykinase (PEPC) in hepatocytes, wherein the PEPCK enzyme is a rate-limiting step of liver tissue glucose neosynthesis. It is an enzyme catalyzing 15 (FIG. 9B).
- PEPC phosphoeno 1 pyru uvat e carboxykinase
- GAPDS (4) may be synthesized by GAPDH (glyceraldehyde 3-) to promote glucose intake.
- AMPKC5 'AMP-activated protein kinase enzymes play a key role in cellular energy homeostasis (20). Similar to G6Pase, we observed that liver cell treatment of ENOblock, GAPDS or rosiglitazone did not affect AMPK expression (FIG. 10D).
- ENOblock down-regulates PEPCK expression and induces glucose uptake in vivo
- zebrafish To investigate the in vivo effect of ENOblock on glucose homeostasis, we chose zebrafish because the animal model provides a convenient and rapid experimental format that requires a small amount of test compound. Moreover, zebrafish and mammals have been known to share similar glucose regulatory reactions (17, 21). Zebrafish tuna treated with ENOblock or rosiglitazone induced down-regulated liver ⁇ expression 12A-12B, which reconfirmed our cell-based findings. The fluorescent glucose probe, 2-NBDG, has been used to evaluate the glucose uptake in zebrafish cheer (22), which is transparent and allows visualization of 2-NBDG fluorescence. We observed that ENOblock treatment induces glucose uptake in zebrafish fry (FIGS. 12C-12D).
- ENOblock treatment inhibits adipogenesis and foam cell format ion
- ENOblock treatment inhibited both foam cell differentiation from macrophages and macrophage differentiation from monocytes (FIGS. 14C-14D).
- the inhibitory effect of ENOblock described above was observed at treatment concentrations that did not induce cytotoxicity (FIG. 14E). Additional discussion
- ENOblock the first non-substrate derivative that directly binds to enolase, which can be used as a probe to characterize enolase activity in biological systems.
- Enolase is the second phase of glycolysis (second from end) to 2-phospho-D-glycerate.
- Enolase is part of the assimilation pathway of glucose neosynthesis and catalyzes counter reactions depending on the environmental concentration of the substrates (6). Enolase also performs many functions that are not related to intrinsic glycolytic function (1, 6). To the best of our knowledge, there are few small molecular means that clearly reveal the various functions of enolase. The most widely reported enolase inhibitor is phosphonoacetohydroxamate (PhAH). PhAH is thought to mimic the aci-carboxylate form of carbanion, an intermediate of reaction, and is only applied for crystallographic studies (29, 30).
- ENOblock In discussing the activity of the ENOblock, the inventors believe that it is important to distinguish the effects of the compound in the hypoxic or normal oxygen state.
- the inventors have discovered ENOblock by screening for compounds that can induce cancer cell death more in the hypoxic state as compared to the normal oxygen state.
- We have adopted the above approach because drugs for general cancer chemotherapy are less effective in hypoxic conditions.
- ENOMock which kills cancer cells in the hypoxic state (FIG. 2) and has inhibitory activity against enolase.
- cancer cells can be characterized by the Warburg effect, which shows an increasing series of metabolic modifications of anabolic glycolysis for energy generation (2).
- glycolysis inhibitors such as 3-bromopyruvate and 6-aminonicotinamide can kill cancer cells (3).
- ENOblock can selectively kill cancer cells under hypoxia (Fig. Lc-ld), which is due to the function of enolase involved in glycolysis (Fig. 6).
- the various functional roles of enolase are found under normal oxygen conditions.
- enolase is non-classical Discharging path (non-classical export pathway) for: moving the eukaryotic cell surface through and have been proposed as being able to cause a transition to mediate cancer cell infiltration (36,37).
- ENOblock treatment on cancer cells in the normal oxygen state inhibits cancer cell infiltration and migration at non-apoptotic concentrations (FIGS. 7A-7D).
- the above findings mean that ENOblock can target cell surface-bound enolase to regulate cancer cell metastasis.
- our in vivo analysis of ENOblock treatment was conducted even in normal oxygen conditions.
- ENOblock can inhibit cancer cell metastasis without significantly affecting cancer cell viability (FIGS. 9D-9E; cancer cells appear to be maintained at the injection site without decreasing cell numbers. ). Furthermore, enolase was known to be associated with cellular microtubule systems that could negatively affect the effects of anticancer agents that destroy microtubules (7, 38). Our results show that ENOblock treatment can increase the effect of microtubule-stabilizing drugs that kill cancer cells (FIG. 7E). Thus, the present inventors have found that ENOblock has a potent activity to inhibit cancer cell metastasis and enhance the effect of microtubule-stabilizing drugs in normal oxygen, as well as to selectively kill cancer cells in the niche of hypoxia in the tumor. Is deduced as a potent new drug candidate for cancer therapy (cancer stem cells are known to express hypoxic state-inducing factors that promote survival under hypoxia; 39).
- ENOblock treatment may inhibit PEPCK expression in ⁇ and induce glucose uptake means that ENOblock may be an attractive candidate for further development as an anti-diabetic drug. This may be supported by the fact that enolase expression is increased in diabetic patients compared to normal people (42). Moreover, our results suggest that it inhibits some complications resulting from the use of diabetic drugs such as increased adipocyte differentiation and foam cell formation (FIG. 14). Insulin signaling is associated with accelerated foam cell formation (43), and commonly prescribed diabetes drugs can induce adipocyte differentiation or weight gain (27).
- ENOblock which inhibits foam cell formation or adipocyte differentiation
- metformin the most commonly prescribed anti-diabetic drug, metformin, also inhibits hepatic glucose neosynthesis and has recently been applied to various clinical trials as an anti-cancer drug (44).
- zebrafish In vivo analyzes of the ENOblock treatment presented herein used a zebrafish vertebrate model. Zebrafish have many advantages as primary animals for testing new therapeutics, including: (a) glucose homeostasis mechanisms preserved in mammals; (b) the usefulness of a system for testing effective anticancer drugs; (c) toxic reactions associated with mammalian testing; And (d) the need for relatively small amounts of test compounds (10, 12, 17, 21, 46). Also, Highly detailed studies of mammalian cell behavior can be conducted in zebrafish (47). Thus, our zebrafish-based series of assays is a suitable format for the initial reporting of ENOblock activity.
- ENOblock the first non-substrate analog inhibitor that directly binds to enolase, and a small molecule that can be used to probe the various non- glycolytic functions of enolase.
- ENOblock we have used ENOblock to assess the effect of enolase inhibition on cancer growth, and it is the first time that enolase inhibition can reduce cancer cell metastasis in phosphorus.
- the inventors show for the first time that enolase inhibition is able to inhibit PEPCK, a regulator of glucose neosynthesis and is a novel target for developing anti-diabetic drugs.
- ENOblock may be the testament of forward chemical genetics, providing novel chemical probes, drug targets and candidate therapeutics for unidentified intracellular mechanisms that regulate human disease. .
- Bacterial o infections eg, Yersinia pest is, Borrelia spp., Streptococcus pneumonia, etc.
- etiology of parasites eg, trypanosomat id parasites; 48
Landscapes
- Health & Medical Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Pharmacology & Pharmacy (AREA)
- Life Sciences & Earth Sciences (AREA)
- Animal Behavior & Ethology (AREA)
- Epidemiology (AREA)
- General Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
Abstract
본 발명은 에놀라제 억제 활성을 가지는 비-기질성 유사체(non-substrate analogue)인 ENOblock 및 이를 포함하는 암 또는 에놀라제-관련된 질환(enolase-associated) 예방 또는 치료용 약제학적 조성물에 관한 것이다. 본 발명의 ENOblock은 에놀라제에 직접적으로 결합하여 이의 활성을 억제하고 이는 정상산소 상태(normoxia)보다 저산소 상태에서 더욱 효과적이다. 또한, 본 발명의 ENOblock은 암 세포의 이동(migration), 전이(metastasis) 및 침윤(invasion)을 억제한다. 한편, 본 발명의 ENOblock은 세포 내 글루코오스 섭취(glucose uptake)를 유도하고 PEPCK 발현을 하향-조절(down-regulation)할 뿐 아니라, 지방세포분화(adipogenesis) 및 거품세포 형성(foam cell formation)을 억제한다. 따라서, 본 발명의 ENOblock을 포함하는 조성물은 암 또는 에놀라제-관련된 질환(enolase-associated) 예방 또는 치료에 매우 효과적으로 적용될 수 있다.
Description
【명세서] : .
[발명의 명칭】 : ;
ENOblock을 유효성분으로 포함하는 암 예방:또는 치료용 약제학적 조성물
'
【기술분야】
본 발명은 대한민국 교육과학기술부의 지원 하에서 과제번호 NRF- 2012-003460에 의해 이루어진 것으로서, 상기 과제의 연구관리전문기관은 한국연구재단, 연구사업명은 "일반연구자지원사업 /신진연구자지원사업" , 연구과제명은 "당뇨병 치료제 개발을 위한 인슐린양 (樣) 작용 저분자 화합물의 탐색과 특성 규명" , 주관기관은 광주과학기술원, 연구기간은 2012.05.01 ~ 2015.04.30이다. 또한, 본 발명은 대한민국 교육과학기술부의 지원 하에서 과제번호 NRF-2012-000462에 의해 이루어진 것으로서, 상기 과제의 연구관리전문기관은 한국연구재단, 연구사업명은 "일반연구자지원사업 /여성과학자지원사업" , 연구과제명은 "암 미세환경 제어에 의한 새로운 항암 기전 및 항암 표적발굴" , 주관기관은 광주과학기술원, 연구기간은 2012.05.01 ~ 2015.04.30이다. 또한, 본 발명은 대한민국 보건복지부의 지원 하에서 과제번호 A120326에 의해 이루어진 것으로서, 상기 과제의 연구관리전문기관은 한국보건산업진홍원, 연구사업명은 "보건의료연구개발사업" , 연구과제명은 "저분자 분화조절제를 이용한 환자 자가유래 iPSCs의 생산과 심근 경색 세포 치료에의 웅용" , 주관기관은 광주과학기술원, 연구기간은 2012.08.01~2014.07.31이다. 본 특허출원은 2012년 10월 22일에 대한민국 특허청에 제출된 대한민국 특허출원 제 10-2012-0117417 호에 대하여 우선권을 주장하며, 상기 특허출원의 개시 사항은 본 명세서에 참조로서 삽입된다.
본 발명은 에놀라제 억제 활성을 가지는 비-기질성 유사체 (non- substrate analogue)인 ENOblock 및 이를 포함하는 암 예방 또는 치료용 약제학적 조성물에 관한 것이다.
【배경기술】
에놀라제는 해당과정 경로 (glycolysis pathway)의
해당과정에서의 기능과 연관되지 않은 다양한 세포 내 과정들에서 중요한
'부가적 기능 (moonlighting)' 을 하는 다양한 기능의 단백질이다. 하지만, 에놀라제 기능을 탐침하는 :작은 분자 도구들은 결정학 (crystallography) 또는 효소학 (enzymology)에 국한되어 왔다. 본 연구에서, 본 발명자들은 에놀라제에 직접 결합하여 이의 활성을 억제하는 최초의 비-기질성 (non- substrate) 유사체로 작은 분자인 'ENOblock' 의 발견을 보고한다. ENOblock은 이전에 약물 저항성 (drug resistance)을 유도하는 것으로 알려진 저산소 상태 (hypoxia)에서 기능하는 세포독성 (cytotoxicity) 제제들을 검출하기 위한 암 세포 어세이를 이용한 작은 분자 스크리닝을 통해 동정되었다. 추가적인 분석을 통해, ENOblock은 인 ^^에서 암 세포 전이를 억제할 수 있다는 것을 확인하였다. 더 나아가, 글루코오스 항상성에서 에놀라제의 예상치 못한 기능이 인 비보 분석에 의해 확인되었다. 따라서, ENOblock은 생물학적 어세이에 적합한 최초로 보고된 에놀라제 억제제이며 암 및 당뇨병의 약물 개발을 위한 매력적인 분자이다. 해당과정은 하나의 글루코오스 분자를 2개의 피루베이트로 전환시키는 진화적으로 매우 잘 보존된 대사 경로이다. 자유 에너지가 방출되어 고-에너지를 함유하는 분자인 ATP(adenosine triphosphate) 및 환원된 NADHCnicotinamide adenine dinucleot ide)를 형성하는 데 이용된다. 해당과정은 10개의 생화학적 반웅들로 구성되고 각 단계는 다른 효소에 의해 촉매도ᅵ어 조절된다. 지난 20년 동안, 해당과정에 관여하는 효소들의 다양한 세포 과정들에서 기능하는 복합 기능들에 대한 놀라운 이해가 있었다 (1).
암 세포는 와버그 효과 (Warburg effect )로; 알려진 현상인 ATP를 생산하기 위해 증가된 해당과정을 나타낸다 (2). 상기 대사적 변형은 암 세포와 정상 세포 간의 기본적인 차이로, 이는 해당과정 억제제들을 이용한 암 세포의 선택적 사멸에 대한 치료 전략올 제공한다 (3). 또한, 해당과정 억제제들은 형성된 종양에서 발생하는 저산소 환경에서 암 세포 사멸을 보다 효과적으로 유도한다는 사실이 알려져 있다 (4). 더욱이, 상술한 저산소 환경은 암 세포가 시타라빈 (cytarabine) 및 독소루비신 (doxorubicin) 같은 항암제에 덜 민감하도록 만든다 (4).
본 명세서 전체에 걸쳐 다수의 논문 및 특허문헌이 참조되고 그 인용이 표시되어 있다. 인용된 논문 및 특허문헌꾀 개시 내용은 그 전체로서 본 명세서에 참조로 삽입되어 본 발명이 속하는 기술 분야의 수준 및 본 발명의 내용이 보다 명확하게 설명된다.
【발명의 내용】
【해결하려는 과제】
본 발명자들은 저산소 상태 (hypoxia)에서 암 세포 생존을 억제할 수 있는 작은 분자를 분리 /동정하고자 노력하였다. 그 결과, 본 발명자들은 ENOblock으로 명명한 트리아진계 화합물 (triazine compound)이 에놀라제에 직접적으로 결합하여 이의 활성을 정상산소 상태 (normoxia)보다 저산소 상태에서 더 효과적으로 억제하몌 상기: 화합물은 암 세포의 이동 (migration), 전이 (metastasis) 및 침윤 (invasion)을 억제할 뿐 아니라; : 세포 내 글루코오스 섭취 (glucose uptake)를 유도하고 PEPCK 발현을 하향- 조절 (down-regulation)하며 지방세포분화 (adipogenesis) 및 거품세포 형성 (foam cell format ion)을 억제한다는 사실을 확인함으로써, 본 발명을 완성하게 되었다.
본 발명의 목적은 암 예방 또는 치료용 약제학적 조성물을 제공한다. 본 발명의 다른 목적은 에놀라제ᅳ-관계된 질환 (enolase-associated disorders) 예방 또는 치료용 약제학적 조성물을 제공한다.
본 발명의 또 목적은 암 예방 또는 치료 방법을 제공한다. 본 발명의 다른 목적 및 이점은 하기의 발명의 상세한 설명, 청구범위 및 도면에 의해 보다 명확하게 된다.
【과제의 해결 수단】
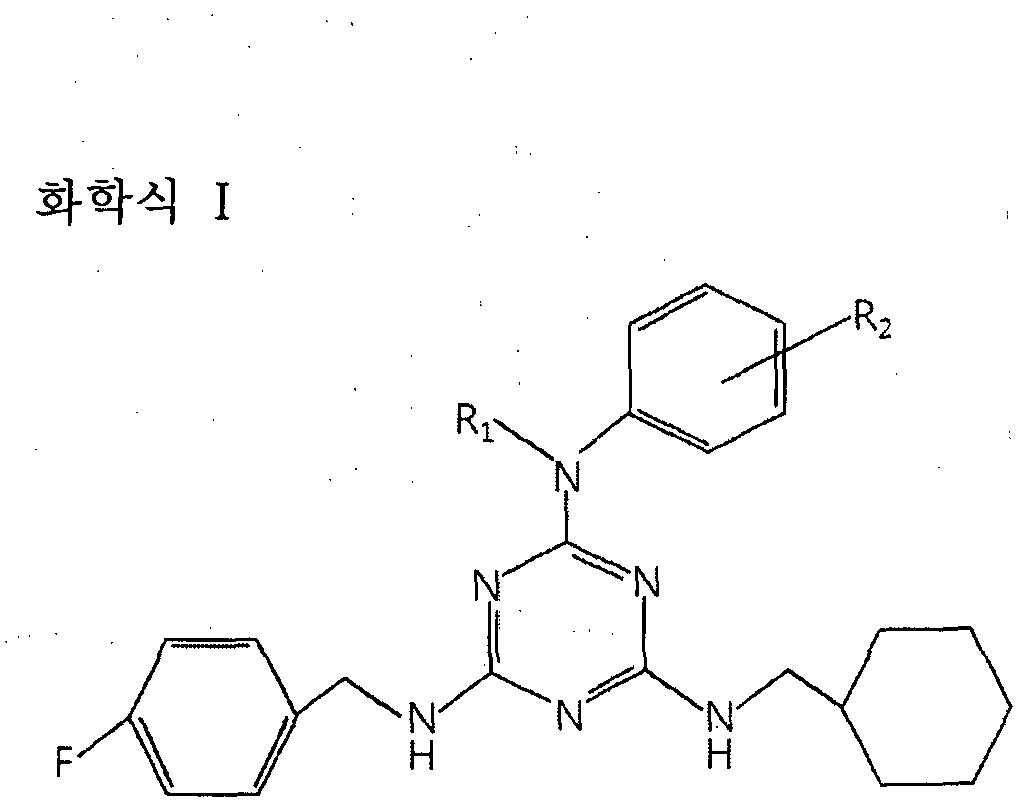
본 발명의 일 양태에 따르면, 본 발명은 (a) 하기 화학식 I로 표시되는 트리아진 (triazine)계 화합물의 치료학적 유효량; 및 (b) 약제학적으로 허용되는 담체를 포함하는 암 예방 또는 치료용 약제학적 조성물을 제공한다:
본 발명의 다른 양태에 따르면, 본 발명은 상술한 조성물을 유효성분으로 포함하는 에놀라제-관계된 질환 (enolase-associated disorders) 예방 또는 치료용 약제학적 조성물을 제공한다. 본 발명자들은 저산소 상태에서 암 세포 생존을 억제할 수 있는 작은 분자를 분리 /동정하고자 노력하였다. 그 결과, 본 발명자들은 ENOblock으로 명명한 트리아진계 화합물이 에놀라제에 직접적으로 결합하여 이의 활성을 정상산소 상태보다 저산소 상태에서 더 효과적으로 억제하며, 상기 화합물은 암 세포의 이동, 전이 및 침윤을 억제할 뿐 아니라, 세포 내 글루코오스 섭취를 유도하고 PEPCK 발현을 하향-조절하며 지방세포분화 및 거품세포 형성을 억제한다는 사실을 확인하였다.
본 발명와 :유효성분인 AP-III-a4( 'ENOblock' 으로 명명됨)는 트리아진 (triazine)계 화합물로, 384종의 태깅된 트리아진계 화합물을 포함하는 화합물 라이브러리 (tagged library)로부터 본 발명자들에 의해 확립된 스크리닝 시스템을 이용하여 저산소 상태에서 암 세포 사멸을 보다 효과적이고 특이적으로 유도하는 작은 분자 화합물로서 동정된 화합물이다. 본 발명에서 사용된 트리아진 화합물 라이브러라의 합성방법 및 이의 용도에 대해서는 국제특허출원 공개공보 W0 03/032903 및 W0 03/050237에 설명되어 있으며, 이들 문헌은 본 명세서에 참조로써 삽입된다.
본 발명의 화학식 I에서 용어 "d-Cs 직쇄 또는 가지쇄 알킬" 은 메틸아민, 에틸아민, n-프로필아민, 이소프로필아민, n-부틸아민, sec- 부틸아민, tert-부틸아민, n-아밀 (amyl), tert-아밀, ' 핵실 등의 직쇄 (straight chain) 또는 가지쇄 (branched chain)의 알킬기를 의미한다. 본 명세서에서 화학식 I에서 알킬 알코올은 메탄을, 에탄올, 프로판을/ 부탄을, 벤질 알코을, 페네틸 알코을 및 이의 유사체를 포함하지만, 이에' 한정되는 것은 아니다.
본 발명의 바람직한 구현예에 따르면, 본 발명의 화학식 I에서 ¾은
H또는 (:广(:2 알킬을 포함하며, 보다 바람직하게는 H를 포함한다.
본 발명의 바람직한 구현예에 따르면, 본 발명의 화학식 I에서 R2는 H, d-C5직쇄 또는 가지쇄의 알킬, d-C5 직쇄 또는 가지쇄의 알킬알코을, 一 [(C¾)m— 0]n-(CH2)p-NH2(상기 m, n 및 는 각각 1ᅳ10의 정수이다), -[(C¾)m- 0]n— C¾(상기 m 및 n은 각각 1-10의 정수이다), -[ (CH2)m-0]n-(CH2)p-CH3(상기 m, n 및 p는 각각 1-10의 정수이다), -(CH2)q-(C0NH)-Ci-5 직쇄 또는 가지쇄의 알킬 (상기 q는 0-5의 정수이다), -(CH2)q-(C0NH)-Ci-5 직쇄 또는 가지쇄의 알킬알코을 (상기 q는 0-5의 정수이다), -(CH2)q-(C0NH)-[(CH2)m- 0]n-(CH2)p-NH2(상기 m, n 및 p는 각각 1ᅳ 10의 정수이고, 상기 q는 0—5의 정수이다), -(C¾)q-(C0NH)-[(CH2)m-0]n-C¾(상기 m 및 n은 각각 1-10의 정수이고, 상기 q는 0-5의 정수이다) 또는 -(CH2)q-(C0NH)-[(CH2)m— 0]n- (C¾)p— CH3(상기 m, n 및 p는 각각 1-10의 정수이고, 상기 q는 0-5의 정수이다)를 포함하고 보다 바람직하게는, H, Ci-C5 직쇄 또는 가지쇄의 알킬, d-C5 직쇄 또는 가지쇄의 알킬알코올, -[(C¾)m-0]n-(CH2)p-N¾(상기 m, n 및 p는 각각 1-5의 정수이다), -[(CH2)m-0]n-CH3(상기 m 및 n은 각각 1-5의
정수이다), -[(C¾)m-0]n-(CH2)pᅳ C¾(상기 m, n 및 p는 각각 1-5의 정수이다), -(CH2)q-(C0NH)-C1-5 직쇄 또는 가지쇄의 알킬 (상기 q는 0-2의 정수이다), - (CH2)q-(C0NH)-C1-5 직쇄 또는 가지쇄의 알킬알코올 (상기 Q는 0-2의 정수이다), -(CH^q- MO-^Cifc -OL- H p-NH^상기 m, n 및 p는 각각 1- 5의 정수이고, 상기 q는 0-2의 정수이다), -(CH2)q-(C0NH)-[(CH2)ra-0]n- C (상기 m 및 n은 각각 1-5의 정수이고, 상기 q는 0-2의 정수이다) 또는 - (C¾)q-(C0NH)-[(CH2V0]n-(CH2)p-C¾(상기 m, n 및 p는 각각 1-5의 정수이고, 상기 q는 0-2의 정수이다)를 포함한다.
본 발명의 보다 바람직한 구현예에 의하면, 상기 화학식 I로 표시되는 트리아진계 화합물은 하기 화학식 Π로 표시되는 화합물을 포함한다:

본 발명은 에놀라제에 직접 결합하여 이의: 활성을 억제하는 비- 기질성 유사체인 작은 분자 (ENOblock)에 대한 최초 보고이다.
본 발명에 따르면, 본 발명의 ENOblock은 정상산소 상태 또는 저산소 상태에서 에놀라제의 활성을 투여량 -의존적으로 특이적으로 억제하였으며, 이를 통해 암 세포 이동, 전이 및 침윤을 억제하였다 (참고: 도 1 내지 도 3). 상술한 효과는 저산소 상태에서 더욱 우수한 효능을 발휘하였다. 따라서, 본 발명의 ENOblock은 에놀라제 활성 억제를 통해 암 또는 에놀라제ᅳ관련된 질환 치료제로 효과적으로 적용될 수 있다.
본 발명의 바람직한 구현예에 따르면, 본 발명의 조성물은 에놀라제의 활성을 억제하고 암 세포의 이동 (migration), 침윤 (invasion) 및 전이 (metastasis)를 억제한다. 본 발명의 보다 바람직한 구현예에 따르면, 본 발명의 ENOblock에 의한 상술한 억제 효과는 정상산소
상태 (normoxia)보다 저산소 상태 (hypoxia)에서 더욱 촉진된다. 본 발명의 바람직한 구현예에 따르면, 본 발명의 조성물은 아팝토시스-유도성 단백질 (apoptosis-inducible proteins)의 발현을 감소시키고, 보다 바람작하게는 AKT 또는 Bcl-xL 단백질의 발현을 감소시킨다.
본 발명의 바람직한 구현예에 따르면, 본 발명의 조성물에 의해 예방 또는 치료될 수 있는 암은 뇌암, 신경 내분비 암, 위암 폐암, 유방암, 난소암, 간암, 기관지암, 비인두암, 후두암, 췌장암, 방광암, 부신암, 대장암, 결장암 자궁경부암, 전립선암, 골암, 피부암, 갑상선암, 부갑상선암 및 요관암으로 구성된 군으로부터 선택되는 암을 포함한다.
상술한 암의 증후는 유방 소괴, 유두 변화, 유방 낭종, 유방 고통, 죽음 몸무게 감소, 허약, 과도한 피로, 섭취 및 숨쉬기의 어려움, 식욕 감퇴, 만성 재채기, 각혈, 혈뇨, 혈변, 멀미, 구토 간 전이, 폐 전이, 골 전이, 대장 전이, 방광 전이, 신장 전이, 췌장 전이, 복부팽만 (abdominal distension), 더부룩함, 복강 내 액체, 질출혈, 변비, 대장천공, 급성 복막염, 고통, 과도한 땀홀림, 열, 고혈압, 빈혈, 설사, 황달, 어지러움, 오한 또는 근육 경련을 포함한다.
본 명세서의 용어 "에놀라제-관련된 질환 (enolase-associated disorders)" 은 에놀라제의 과다발현 (overexpression)이 유도되는 질환으로, 보다 바람직하게는 정상산소 상태보다는 저산소.상태에서 유도되는 질환을 의미하며, 예를 들어 암 (cancers), 자가면역질환 (autoimmune disorders), 허혈 (ischemia) 및 박테리아 감염을 포함하지만, 이에 한정되는 것은 아니다.
한편, 본 발명의 조성물은 당뇨병 예방 또는 치료용 조성물로서도 이용될 수 있다.
본 발명의 바람직한 구현예에 따르면, 본 발명의 조성물은 인슬린 모방 활성 (insulin-mimicking activity)을 가진다.
본 발명의 바람직한 구현예에 따르면, 본 발명의 조성물에 의한 상술한.인슐린 모방 활성은 세포 내 글루코오스 섭취 (glucose uptake)를 촉진시키는 것을 포함한다.
본 발명의 바람직한 구현예에 따르면, 본 발명의 조성물은 포도당
신생합성 (gluconeogenesis)에 중요 단백질인 PEPCK ( hosphoeno 1 pyr uva t e carboxykinase)의 발현을 하향一조절 (down— regulat ion)시킨다.
본 발명의 바람직한 구현예에 따르면, 본 발명의 조성물은 지방세포분화 (adipogenesis) 및 거품세포 형성 (foam cell formation)을 억제한다.
본 명쎄서에서 용어 "약제학적 유효량" 은 상술한 트리아진계 화합물의 효능 또는 활성 (예컨대, 항-암제, 항 -당뇨병제, 등)을 달성하는 데 층분한 양을 의미한다 .
본 발명의 약제학적 조성물은 약제학적으로 허용되는 담체를 포함한다. 본 발명의 약제학적 조성물에 포함되는 약제학적으로 허용되는 담체는 제제시에 통상적으로 이용되는 것으로서, 락토스, 덱스트로스, 수크로스, 솔비를, 만니를, 전분, 아카시아 고무, 인산 칼슘, 알기네이트, 젤라틴, 규산 칼슘, 미세결정성 셀를로스, 폴리비닐피롤리돈, 셀를로스, 물, 시럽, 메틸 셀를로스, 메틸히드록시벤조에이트, 프로필히드록시벤조에이트, 활석, 스테아르산 마그네슘 및 미네랄 오일 등을 포함하나, 이에 한정되는 것은 아니다. 본 발명의 약제학적 조성물은 상기 성분들 이외에 윤활제, 습윤제, 감미제, 향미제, 유화제, 현탁제 , 보존제 등을 추가로 포함할 수 있다. 적합한 약제학적으로 허용되는 담체 및 제제는 Remington's Pharmaceutical Sciences (19th ed. , 1995)에 상세히 기재되어 있다.
본 발명의 약제학적 조성물의 적합한 투여량은 제제화 방법, 투여 방식, 환자의 연령, 체중, 성, 병적 상태, 음식, 투여 시간, 투여 경로, 배설 속도 및 반웅 감응성과 같은 요인들에 의해 다양하게 처방될 수 있다. 한편, 본 발명의 약제학적 조성물의 경구 투여량은 바람직하게는 1일 당 0.001-100 mg/kg (체중)이다.
본 발명의 약제학적 조성물은 경구 또는 비경구로 투여할 수 있고, 비경구로 투여되는 경우, 정맥내 주입, 피하 주입, 근육 주입, 복강 주입, 경피 투여 등으로 투여할 수 있다. 본 발명의 약제학적 조성물은 적용되는 질환의 종류에 따라, 투여 경로가 결정되는 것이 바람직하다.
본 발명의 조성물에 포함되는 유효성분인 화학식 I의 트리아진 계열 화합물의 농도는 치료 목적, 환자의 상태, 필요기간 등을 고려하여 결정할 수 있으며 특정 범위의 농도로 한정되지 않는다.
본 발명의 약제학적 조성물은 당해 발명이 속하는 기술분야에서 통상의 지식을 가진 자가 용이하게 실시할 수 있는 방법에 따라, 약제학적으로 허용되는 담체 및 /또는 부형제를 이용하여 제제화 함으로써 단위 용량 형태로 제조되거나 또는 다용량 용기내에 내입시켜 제조될 수 있다. 이때 제형은 오일 또는 수성 매질중의 용액, 현탁액 또는 유화액 형태이거나 액스제, 분말제, 과립제, 정제 또는 캅셀제 형태일 수도 있으며 분산제 또는 안정화제를 추가적으로 포함할 수 있다 . 본 발명의 또 다른 양태에 따르면, 본 발명은 (a) 하기 화학식 I로 표시되는 트리아진 (triazine)계 화합물의 치료학적 유효량; 및 (b) 약제학적으로 허용되는 담체를 포함하는 조성물을 대상 (subject)에게 투여하는 단계를 포함하는 암 예방또는 치료 방법을 제공한다:
화학식 I

¾는 H, Ci-C5 직쇄 또는 가지쇄의 알킬, d-Cs 직쇄 또는 가지쇄의 알킬알코올, -[(CH2)m-0]n— (C¾)p-N¾ (상기 m, n 및 p는 각각 1-10의 정수이다), -[(CH2)m-0]n-CH3(상기 m 및 n은 각각 1-10의 정수이다), - [(CH2)ra-0]n-(CH2)p-CH3(상기 m, η 및 p는 각각 1-10의 정수이다), -(C¾)q - (C0NH)-Ci-5 직쇄 또는 가지쇄의 알킬 (상기 q는 0-5의 정수이다), -(C¾)q— (C0NH)-Ci-5 직쇄 또는 가지쇄의 알킬알코올 (상기 Q는 으 5의 정수이다), - (CH2)q-(C0NHH(CH2)m-0]n-(C¾)p— NH2(상기 m, n 및 p는 각각 1-10의 정수이고, 상기 q는 0-5의 정수이다), -(CH2)q-(CONH)-[ ((¾) -()]„-(¾(상기 m 및 n은 각각 1-10의 정수이고, 상기 q는 0—5의 정수이다) 또는 -(C¾)q- (C0NH)-[(CH2)m-0]n-(CH2)p-C¾(상기 m, n 및 p는 각각 1—10의 정수이고,
상기 q는 0-5의 정수이다)이다.
본 발명의 방법은 상술한 조성물을 이용하기 때문에, 이 둘 사이의 공통된 내용은 본 명세서의 과도한 복잡성을 회피하기 위하여 그 기재를 생략한다ᅵ.
【발명의 효과】
본 발명의 특징 및 이점을 요약하면 다음과 같다:
(a) 본 발명은 에놀라제 억제 활성을 가지는 비-기질성 유사체 (ήοη- substrate analogue)인 ENOblock 및 이를 포함하는 암 또는 에놀라제— 관련된 질환 (enolase-associated) 예방 또는 치료용 약제학적 조성물에 관한 것이다.
(b) 본 발명의 ENOblock은 에놀라제에 직접적으로 결합하여 이의 활성을 억제하고 이는 정상산소 상태 (normoxia)보다 저산소 상태에서 더욱 효과적이다.
(c) 또한, 본 발명의 ENOblock은 암 세포의 이동 (migration), 전이 (metastasis) 및 침윤 (invasion)을 억제한다.
(d) 한편, 본 발명의 ENOblock은 세포 내 글루코오스 섭취 (glucose uptake)를 유도하고 PEPCK발현을 하향 -조절 (down-regulation)한다.
(e) 또한, 본 발명의 ENOblock은 지방세포분화 (adipogenesis) 및 거품세포 형성 (foam cell format ion)을 억제한다.
(f) 따라세 본 발명의 ENOblock을 포함하는 조성물은 암 또는 에놀라제ᅳ관련된 질환 (enolase-associated) 예방 또는 치료에 매우 효과적으로 적용될 수 있다. 【도면의 간단한 설명】
도 1은 AP-III-a4화합물 (ENOblock)의 발견올 보여주는 도면이다. 도 la는 저산소 상태 (hypoxia) 하에서 효과적인 아팝토시스 유도인자들을 검출하기 위해 이용된 스크리닝 시스템을 도식적으로 보여준다. HCT116 암 세포들이 평행 96-웰 배양 플레이들에 분주되었다. 저산소 상태는 150 mM 코발트 클로라이드 (CoCl2) 처리를 통해 하나의 플레이트에서 유도하였다. 트리아진 화합물 라이브러리 (5 mM)이 4시간 후에 첨가되고 세포 사멸은
저산소 상태 유도 후 24시간 째에 조사하였다. '히트 (Hit)' 화합물들은 정상산소 상태 (normoxia)와 비교하여 저산소 상태에서 25% 또는 그 이상의 세포 사멸을 유도하였다 (MTT 어세이 흡광도로서 측정되었음). 도 lb는 본원발명의 AP-III-a4(EN0block), 대조군 화합물인 AP-IV-e3 및 ΑΡ-1-flO, GAPDH를 타겟팅하는 GAPDS, 그리고 잘-알려진 항—당뇨병 약물인 로지글리타존의 화학적 구조를 보여주는 도면이다. 도 lc는 정상산소 상태 (Norm)와 비교하여 저산소 상태 (Hypox)에서 ENOblock가 HCT116 대장암 세포 사멸을 현저하게 유도한다는 것올 보여주는 결과이다. 약어: 오차 (error) = 표준편차 (SD); *, 정상산소 상태와 비교하여 증가된 세포 사멸을 나타내는 값 < 0.05. 각각 유의한 변수값 (variable)에 대한 정확한 P 값은 다음과 같다: hypox 10 = 1.23X10"6; hypox 5 = 2.38X10"6; hypox 2.5 = 1.16X10"5; hypox 1.25 = 0.000334. 도 Id는 정상산소 상태 또는 저산소 상태로 처리된 HCT116 대장암 세포들의 대표적인 위상차 현미경 이미지를 보여주는 결과이다.
도 2는 HCT116 암 세포에 150 μΜ CoCl2의 4시간 처리가 저산소 상태를 유발한다는 것을 의미하는 결과로, 저산소 상태-반웅성 유전자인 에놀라제의 증가된 발현을 보여준다. 이와 대조적으로, 해당과정 경로에 포함된 다른 단백질인 GAPDH(glyceraldehycle 3-phosphate dehydrogenase)는 이전 보고들 (예컨대, Said, et al., BMC Mol Biol. 8: 55(2007))과' 일치하게도 저산소 상태가 유도된 후에도 증가된 발현을 나타내지 않았다. 빨간색 슷자는 밀도계측 분석 (densitometry analysis)을 통해 계산된 발현의 배수ᅳ변화 (fold— change)를 지시한다.
도 3은 AP 링커 라이브러리 (AP linker library)의 합성을 개략적으로 나타내는 도면이다. 도 3a는 직교 합성 시약 및 조건에 대한 통상적인 도식을 보여준다. 반웅조건: (a) RiNH2 또는 ΝΗ20Η(5당량), THF에 녹여진 2% 아세트산, 상은, 1시간; NaB(0Ac)3H(7당량), 상온, 12시간; (b) THF에 녹여진 빌딩 블록 (Building block) 11(4당량), 60 °C, 1시간, DIEA(n,n- diisopropyl ethyl amine); (c) ' NH, DIEA, NMP:n— Bu0H=l: 1, 120 °C, 3시간; 및 (d) 디클로로메탄 (dichloromethane)에 녹여진 10% TFACtrifluoroacetic acid), 30분. 도 3b는 링커 (Linker)를 나타낸다. 도 3c는 빌딩 블톡 I의 구축을 위해 이용된 아민류 및 아미노 알코올류를
보여주는 결과이다. 도 Id는 빌딩 블록 III의 구축을 위해 이용된 아민류를 보여주는 결과이다.
도 4는 본원발명의 AP-III-a4 화합물 (ENOblock)이 에놀라제의 직접적인 억제제라는 것을 나타내는 결과이다. 도 4a는 HCT116 암 세포에서 ENOblock에 대한 친화성 크로마토그래피 연구 결과를 보여준다. 'a' 및 'b' 로 지시된 단백질 밴드들은 질량분석올 통해 에놀라제의 헤테로다이머의 서브유니트들로 동정되었다. 이와 대조적으로, ENOblock 친화성 매트릭스로부터 얻어진 용출액 내 다른 뚜렷한 단백질 밴드들은 질량분석 (mass spectrometry)을 통해 동정할 수 없었다. 대조군 매트릭스 = HCT116 용해물 단백질들에 결합하지 않았던 AP-IV-e3(2) 친화성 매트릭스. 도 4b는 에놀라제 모노머 서브유니트의 서열을 보여준다. 동정된 펩타이드는 빨간색으로 제시된다. 100 이상의 마스코트 스코어 (Mascot scores)는 유의한 것으로 간주된다. 도 4c는 HCT116 암 세포 용해물 내 에놀라제가 ENOblock 친화성 매트릭스에 결합한다는 것을 확인시켜 주는 웨스턴 블랏 분석이다. 경쟁인자로 프리 (free): EN0block(EN0 약자로 표기됨)을 이용한 경쟁 분석은 ENOblock 친화성 매트릭스에 대한 에놀라제 결합올 완벽하게 억제하였다. YD-10B 구강 암 세포 또는 Huh7 간세포 (hepatocytes)로부터 유래한 세포 용해물 (20 μ§)은 양성 대조군으로서 이용하였다. 도 4d는 정제된 인간 에놀라제가 ENOblock 친화성 매트릭스에 결합한다는 것을 보여주는 웨스턴 블랏 분석이다. 경쟁인자로 프리 ENOblock을 이용한 경쟁 분석은 ENOblock 친화성 매트릭스에 대한 정제된 에놀라제 결합을 완벽하게 억제하였다. 이와 대조적으로, 대조군인 AP-IV-e3 화합물 친화성 매트릭스는 정제된 에놀라제에 결합할 수 없었다. 양성 대조군으로서, ENOblock 친화성 매트릭스는 HCT116 세포 용해물 (200 μ§)과 반응시켰다. HCT116 암 세포로부터 얻어진 세포 용해물 (50 g)은 에놀라제 항체에 대한 양성 대조군으로서 이용하였다. 도 4e는 ENOblock이 정제된 에놀라제의 활성을 투여량 -의존적으로 억제한다는 것을 보여주는 결과이다. ENOblock은 잘- 알려진 에놀라제 억제제인 NAF보다 현저하게 더 낮은 농도에서 에놀라제 활성을 억제하였고, 2.5 μΜ의 ENOblock 처리는 1 mM의 NaF 처리와 유사한 레벨로 에놀라제 활성을 감소시켰다. 약어: 오차 = 표준편차 (SD); *,
비처리된 그룹과 비교한 ^값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: 플루오라이드 0.5 mM = 0.007708; 플투오라이드 1 mM = 0.000543; 플루오라이드 2 mM = 0.000376; EnoBlock 2.5 μΜ = 0.000527; EnoBlock 5 μΜ = 0.000474; EnoBlock 10 μΜ = 0.000335.
도 5는 ENOblock 동정에 이용된 동일한 트리아진 화합물 라이브러리로부터 얻어진 '비—히트 (non-hit)' 화합물인 ΑΡ-1-flO가 에놀라제 활성을 ,현저하게 억제하지 않는다는 것을 보여주는 결과이다. ENOblock은 양성 대조군으로서 이용하였다. 약어: 오차 = 표준편차 (SD); *, 비-처리된 그룹과 비교하여 7°값 < 0.05.
도 6은 HCT116 암 세포에서 에놀라제 발현의 siRNA-매개된 넉-다운이 저산소 상태에 대한 민감성 (susceptibility)을 증가시킨다는 것올 보여주는 결과이다. 세포에 에놀라제 (EN01) siRNA 또는 음성 대조군인 2개의 siRNA를 다양한 농도로 처리하였다: 음성대조군 siRNAs; (a) 80 pmols의 스크램블 (scrambled) siRNA 또는 (b) 80 pmols의 p57(cyclin dependent kinase inhibitor) siRNA. 24시간의 siRNA 트랜스펙션 후, 세포들은 104 세포 /웰의 밀도로 96-웰 배양 플레이트로 옮겨 정상산소 상태 또는 150 mM CoCl2 처리에 의해 유도된 저산소 상태에서 24시간 동안 배양시켰다. 세포독성은 24시간 후에 MTT 어세이로 평가하였다. 증가하는 농도의 EN01 siRNA는 저산소 상태 하에서 투여량ᅳ의존적으로 암 세포 사멸을 유도하였다. 약어: 오차 = 표준편차 (SD); *, 정상산소 상태와 비교하여 저산소 상태 하의 증가된 세포독성 결과에 대한 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: 저산소 상태 하의 에놀라제 (40 pmols) = 0.00990; 저산소 상태 하의 에놀라제 (60 pmols) = 3.32X10"6; 저산소 상태 하의 에놀라제 (80 pmols) = 1.43X10"8.
도 7은 ENOblock가 암 세포 침윤 (invasion) 및 이동 (migrat ion)을 억제할 수 있다는 것을 보여주는 결과이다. 도 7a은 정상산소 상태에서 HCT116 암 세포들에 ENOblcok의 처리는 투여량ᅳ의존적으로 침윤을 억제한다는 결과를 나타낸다. ENOblock 처리는 이전에 알려진 침윤에 대한 작은 분자 억제제인 LY294002(포스포이노시티드 3-키나제 시그널링의 억제)보다 훨씬 더 효과적으로 세포 침윤을 억제하였다. 약어: 오차 = 표준편차 (SD); *, 비처리된 그룹과 비교한 P 값 < 0.05. 각각 유의한
변수값에 대한 정확한 P 값은 다음과 같다: EnoBlock 2.5 μΜ = 0.0191; EnoBlock 1.25 μΜ = 0.0286; EnoBlock 0.625 μΜ = 0.0481. 도 7b는 트랜스웰 인서트 (transwell inserts)에 침투된 크리스탈 바이을렛 (crystal violet)-염색된 HCT116 세포들의 현미경 이미지들을 나타낸다 [축척 막대 (scale bar) = 100 μπι]. 도 7c은 정상산소 상태에서 HCT116 암 세포들에 대한 ENOblock의 처리가 투여량 -의존적으로 세포 이동을 억제한다는 것을 보여주는 결과이다. 세포 침윤과 유사하게도, 세포 이동을 억제함에 있어 ENOblock은 LY294002보다 더욱 효과적이었다. 약어: 오차 = 표준편차 (SD); *, 비처리된 그룹과 비교한 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: EnoBlock 2.5 μΜ = 0.00939. 도 7(1는 트랜스웰 인서트 (transwell inserts)로 이등된 크리스탈 바이올렛— 염색된 HCT116 세포들의 현미경 이미지들을 나타낸다 (축척 막대 = 100 ym). 도 3e는 ENOblock treatment of HCT116 암 세포에 대한 ENOblock의 처리는 항-튜블린 (ant i tubulin) 화학치료제인 탁솔 (taxol) 및 빈크리스틴 (vincristine)에 대한 민감성을 증가시켰다는 것을 보여주는 결과이다. 10 mM ENOblock의 존재 또는 부존재 하에서 세포에 10 nM 탁솔 및 10 nM 빈크리스틴을 처리하였다. 약어: 오차 표준편차 (SD); *, 그래프에 지시된 그룹들 간의 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: EN0block+탁솔과 비교된 탁솔 2.55X10'6; EnoBIock+빈크리스틴과 비교된 빈크리스틴 = 0.000304. 도 7f는 ENOblock treatment of HCT116 암 세포에 대한 ENOblock의 처리가 아팝토시스의 음성 조절자들인 AKT 및 Bcl-Xl의 발현을 감소시켰다는 것을 보여주는 결과이다. AKT 웨스턴 블랏 분석을 위해, 세포에 ENOblock을 24시간 동안 처리하였고; Bcl-Xl 웨스턴 블랏 분석을 위해, 세포에 ENOblock을 48시간 동안 처리하였다.
도 8은 AP-I— flO 화합물 (10 μΜ)의 처라가 HCT116 암 세포 침윤올 감소시키지 않는다는 것을 나타내는 크리스탈 바이을렛-염색된 트랜스 ¾ 인서트의 사진을 보여주는 결과이다. LY294002(5 μΜ)의 처리는 양성 대조군으로 이용되었으며, 침윤된 암 세포들에서 현저한 감소를 나타냈다. 도 9는 ENOblock의 독성 연구 (toxicological study) 및 항-암 활성에 대한 인 비보 분석 결과를 보여준다. 도 9a는 제브라피쉬 치어
시스템 (zebrafish larvae system)을 이용한 ENOblock 처리의 독성 분석 결과이다. 다양한 발생학적 변수 (developmental parameters)의 측정을 통해 10 μΜ ENOblock 투여량은 치어에 해롭지 않으나, 20 μΜ ENOblock 투여량은 제브라피쉬의 수영 능력올 억제한다는 것을 확인하였다. 도 9b는 증가하는 ENOblock 투여량에 따른 수정 후 72일 째 (72 days post fertilization, 72 dfp) 쎄브라피쉬 치어의 현미경 :관찰 결과를 보여준다. 그 결과, 20 μΜ ENOblock의 투여량은 비정상적으로 큰 부레 (swim bladder; 빨간색 화살표)를 발생시킨다는 것을 관찰할 수 있었다. 40 μΜ ENOblock의 투여량은 치어에서 수많은 비정상적 표현형을 발생시켰다. 도 9c는 20 μΜ 또는 40 μΜ ENOblock의 투여량도 전체적으로 제브라피쉬의 치어 생존율 (larvae viability)을 감소시킨 반면에 10 μΜ ENOblock의 투여량은 생존율에 영향을 미치지 않는다는 것을 보여주는 결과이다. 약어: 오차 = 표준편차 (SD); *, DMS0-처리된 그룹과 비교하여 생존율의 변화에 대한 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: 20 μΜ ENOblock = 0.0121; 40 μΜ ENOblock = 0.0019. 도 9d는 HCT116-이종이식된 제브라피쉬에 xenotranspl anted zebrafish with a dose of 10 μΜ ENOblock의 96시간 처리가 난황 주머니 주입 위치로부터 이동 및 전이 (광범위한 암 세포들)을 나타내는 배아들의 수를 감소시킨다는 것을 보여주는 결과이다. DMS0 처리는 대조군으로 이용하였다. 3개의 대표적인 배아들이 각 실험 그룹으로부터 보여지고 광범위한 암 세포 병소들 (foci)이 파란색 화살표들로 제시된다. 이종이식된 암 세포 미세병소들의 정량을 통해, 본 발명자들은 ENOblock 처리가 암 세포 이동 및 전이를 현저하게 감소시켰다는 것을 확인하였다. 약어: 오차 = 표준편차 (SD); *, DMS0- 처리된 그룹과 비교하여 항-전이의 효과에 대한 / ^값 < 0.05.
도 10은 ENOblock은 글루코오스 섭취를 유도하고 PEPCK 발현을 억제한다는 것을 의미하는 결과이다. 도 10a는 Huh7 간세포 또는 HEK 신장 세포에 10 μΜ ENOblock (약어로 EN0)의 24시간 처리가 형광성 글루코오스 프로브인 2— NBDG를 이용하여 측정된 바와 같이 글루코오스 섭취 (glucose uptake)를 유도한다는 것을 보여주는 결과이다. 해당과정 효소인 GAPDH의 작은 분자 조절자인 GAPDSC10 μΜ)의 24시간 처리도 간세포에서 글루코오스 섭취를 유도할 수 있었다. 이와 대조적으로, 잘-알려진 항-당뇨병 약물인
로지글리타존 (약어로 R0SI; 10 μΜ)의 24시간 처리는 간세포에서 글루코오스 섭취를 유도할 수 있었지만, 신장 세포에서는 글루코오스 섭취를 유도하지 않았다. 약어: 오차 = 표준편차 (SD); *, 비처리된 그룹과 비교하여 글루코오스 섭취 결과에 대한 7값 < 0.05. 각각 유의한 변수값에 대한 정확한 Ρ 값은 다음과 같다: 간세포의 경우, R0SI = 0.0227; GAPDS = 0.0302; EN0 = 0.0213; 신장 세포의 경우, GAPDS = 0.0231; EN0=0.0271. 도 10b는 Huh7 간세포에 EN0block(10 μΜ)의 24시간 처리는 포도당 신생합성 (gluconeogenesis)의 핵심 양성 조절자인 PEPCK의 발현을 억제한다는 것을 보여주는 결과이다. 10 μΜ 로지글리타존의 24시간 처리도 PEPCK 발현을 억제할 ' 수 있었다. 하지만, 10 μΜ GAPDS의 24시간 처리는 PEPCK 발현을 억제하지 않았다. 약어: 오차 = 표준편차 (SD); *, 약물- 처리되지 않은 그룹 (DMS0-처리된 그룹)과 비교하여 PEPCK 발현 결과에 대한 ^ 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: GAPDS = 0.00213; EN0 = 0.00466. 도 10c는 HEK 세포에 BN0block(10 μΜ)의 24시간 처리가 PEPCK의 발현을 억제한다는 것을 보여주는 결과이다. 이와 대조적으로, 10 μΜ GAPDS, 1 pg/mL 인슐린 (약어로 Ins) 또는 10 μΜ 로지글리타존의 24시간 처리는 PEPCK 발현을 감소시키지 않았다. 약어: 오차 = 표준편차 (SD); *, 약물-처리되지 않은 그룹 (DMS0-처리된 그룹)과 비교하여 PEPCK의 증가된 발현 결과에 대한 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: EN0 = 0.000393. 도 10d는 간세포에 BN0block(10 μΜ)의 24시간 처리가 포도당 신생합성올 조절하는 다른 효소인 G6Pase(glucose 6-phosphatase) 또는 세포 내 에너지 항상성을 조절하는 AMPK(5' AMP-activated protein kinase)의 발현에 영향을 미치지 않는다는 것올 보여주는 결과이다. 유사하게도, 10 μΜ GAPDS 또는 10 μΜ rosiglitazone의 24시간 처리도 상술한 효소들의 발현에 영향을 미치지 않았다. 약어: 오차 = 표준편차 (SD).
도 11는 간세포 또는 신장 세포에서 에놀라제 발현의 siRNA-매개된 넉-다운은 글루코오스 섭취를 유도한다는 결과로, 형광성 글루코오스 프로브인 2-NBDG의 증가하는 표지로 확인할 수 있다. 세포에 에놀라제 (EN01) siRNA 또는 음성 대조군인 2개의 siRNA를 다양한 농도로 처리하였다: 음성대조군 siRNAs; (a) 80 pmols의 스크램블 (scrambled) siRNA 또는 (b) 80
pmols의 p57(cyclin dependent kinase inhibitor) siRNA. 24시간의 siRNA 트랜스펙션 후, 세포들은 104 세포 /웰의 밀도로 96-웰 배양 플레이트로 옮겨 100 uM 2-NBDG를 30분 동안 처리하였다. 이후, 2-NBDG 형광 측정은 본 명세서의 실험방법에 기재된 바와 같이 실시하였다. 약어: 오차 = 표준편차 (SD); *, 스크램블 siRNA-처리된 세포와 비교하여 증가된 글루코오스 섭취 결과에 대한 5값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: (a) 신장 세포의 경우, i) 에놀라제 (40 pmols) = 1.83X10-5, ii) 에놀라제 (60 pmols) = 1.76X10— 6' iii) 에놀라제 (80 pmols) = 5.1X10"6; (b) 간세포의 경우ᅳ I) 에놀라제 (40 pmols) = 2.42X10— 5, ii) 에놀라게 (60 pmols) = 0.00264, iii) 에놀라제 (80 pmols) = 0.00239.
도 12는 ENOblock이 인 에서 PEPCK 발현을 억제하고 글루코오스 섭취를 유도할 수 있다는 것을 보여주는 결과이다. 도 12a는 제브라피쉬에서 PEPCK 발현 상에 ENOblock 처리의 효과를 측정하기 위한 본 발명자들의 접근방법을 도식적으로 보여주는 도면이다. 도 12b는 EN0block(10 μΜ)의 3시간 처리가 제브라피쉬 성어의 간 조직에서 PEPCK 발현을 억제한다는 것을 보여주는 결과이다. 간세포에서 PEPCK 발현을 억제하였던 로지글리타존 (10 uM)의 3시간 처리도 제브라피쉬 간 조직에서 PEPCK 발현올 억제한다는 것을 나타내는 결과이다. 약어: 오차 = 표준편차 (SD); *, 비-처리된 그룹과 비교하여 PEPCK 발현 결과에 대한 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: R0SI = 0.00310; EN0 = 5.02X10—5. 도 12c는 제브라피쉬 치어 (larvae)에서 형광- 태깅된 글루코오스 바이오프로브 (2-NBDG)의 섭취 상에 ENOblock 처리의 효과를 측정하기 위한 본 발명자들의 접근방법 (치어의 투명성으로 인해 용이하게 관찰할 수 있음)을 도식적으로 보여주는 도면이다. 도 12d는 ENOblock (10 μΜ)의 4시간 처리가 제브라피쉬 치어에서 글루코오스 섭취를 증대시킨다는 것을 보여주는 결과이다. 증가된 글루코오스 섭취는 발달하는 배아, 특히 눈 (하얀색 화살표), 소장 (intestine) 및 난황 주머니를 통해 관찰될 수 있다. 비교군으로서, 글루코오스 섭취를 촉진하는 알려진 항- 당뇨병 천연산물인 에모딘 (10 yg/mL)이 처리된 제브라피쉬도 ENOblock이 처리된 제브라피쉬의 패턴과 유사하지만 더욱 강한 패턴으로 글루코오스 섭취를 증가시켰다. 도 12e는 치어 단계에서 수많은 글루코오스
트랜스포터들을 발현하는 72 hpf 째의 제브라피쉬 치어의 눈에서 형광성 글루코오스 프로브인 2-NBDG로부터 발생한 형광 시그널 강도의 정량 결과를 보여주는 그래프이다. 10 μΜ ENOblock 또는 10 yg/mL 에모딘이 4시간 동안 투여된 제브라피쉬 치어는 제브라피쉬 눈에서 현저하게 큰 형광- 태깅된 글루코오스 섭취를 유도하였다. 약어: 오차 = 표준편차 (SD); *, 2- NBDG 단독-처리된 제브라피쉬와 비교하여 측정된 형광 강도 결과에 대한 p 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: NBDG+ENOblock = 0.0252; NBDG+에모딘 = 0.0203.
도 13은 72 hpf 제브라피쉬에 ENOblockCLO μΜ)의 1시간 처리가 글루코오스 섭취를 유도한다는 것을 보여주는 결과로, 용해된 치어에서 2- NBDG 시그널을 형광 판독기 (fluorescent plate reader)를 이용하여 측정하였다. 약어: 오차 = 표준편차 (SD); *, NBDG 단독-처리된 그룹과 비교하여 증가된 형광 검출 결과에 대한 ^값 < 0.05.
도 14는 ENOblock이 지방세포분화(3^1>( 61 ^3) 및 거품세포 형성 (foam cell formation)을 억제한다는 것을 의미하는 결과이다. 도 14a(i)는 지방세포분화를 거치는 지방전구세포 (pnradipocytes)의 현미경 분석 결과로, 10 μΜ ENOblock의 처리는 Oil Red 0 염색의 부재에서 알 수 있듯이 지질 축적 (lipid accumulation)을 억제하였다. 이와 대조적으로, 지방세포분화-유도성 인자의 부재 하에서 지방전구세포에 대한 인슐린의 처리는 여전히 지질 축적을 유도하였다. 도 14a( )는 ENOblock 처리가 지방세포분화 과정동안 지질 축적을 차단한다는 것을 보여주는 정량 그래프 결과이다. 약어: 오차 = 표준편차 (SD); *, 비-처리된 지방세포 (adipocytes)와 비교하여 지질 축적 결과에 대한 尸 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: 인술린 단독 = 0.004793; 지방세포분화성 칵테일 (adipogenic cocktail) 단독 = 0.0252. 도 14b는 을레산 (oleic acid)이 처리된 대식세포의 현미경 분석 결과로, 10 μΜ ENOblock의 처리는 Oil Red 0 염색의 감소에서 알 수 있듯이 거품세포 형성을 억제하였다. 도 14c는 지질 축적을 보여주는 세포들의 카운팅에서 알 수 있듯이 ENOblock 처리 (10 μΜ)가 거품세포 형성올 억제한다는 것을 보여주는 정량 그래프 결과이다. 이와 대조적으로, 거품세포로 발생 (development)하는 대식세포에 항-당뇨병 약물인 로지글리타존 (10
μΜ)의 처리는 지질 축적을 보이는 세포들의 수에 영향을 미치지 않았다. 약어: 오차 = 표준편차 (SD); *, 올레산-처리된 지방세포 그룹과 비교하여 감소된 지질을 포함하는 세포 카운팅 결과에 대한 尸 값 < 0.05. 각각 유의한 변수값에 대한 정확한 Ρ 값은 다음과 같다: 을레산 +EN0block = 0.0335. 도 14d는 배양 디쉬에 부착된 세포들와 수를 카운팅을 통해 단핵구 (monocytes)에 대한 EN0block(10 μΜ)의 처리가 대식세포로의 분화 (differentiation)를 억제한다는 것을 보여주는 결과이다. 이와 대조적으로, 단핵구에 로지글리타존 (10 μΜ)의 처리는 대식세포로의 분화에 영향을 미치지 않았다. 약어: 오차 = 표준편차 (SD); *, ΡΜΑ-처리된 그룹과 비교하여 부착된 세포의 감소된 수를 카운팅한 결과에 대한 Ρ 값 < 0.05. 각각 유의한 변수값에 대한 정확한 Ρ 값은 다음과 같다: PMA+ENOblock = 0.0235. 도 14e는 ΜΊΤ 어세이 분석에서 알 수 있듯이 대식세포에 대한 ENOblock 처리 효과들이 세포독성에서 기인하지 않는다는 것을 보여주는 그래프 결과이다. ¾02(3 mM)의 48시간 처리가 양성 대조군으로서 이용되었다 . 약어 : 오차 = 표준편차 (SD); *, 비-처리된 그룹과 비교하여 감소된 흡광도 (570 nm) 결과에 대한 값 < 0.05. 각각 유의한 변수값에 대한 정확한 P 값은 다음과 같다: 3 mM H202 = 8.84X10"7.
도 15는 HEK 신장 세포 및 Huh7 간세포에서 siRNA 처리를 통한 에놀라제 발현 넉-다운을 보여주는 면역블롯 분석 결과이다. 세포에 80 pmols의 siRNA들을 48시간 동안 트랜스펙션시켰다.
【발명을 실시하기 위한 구체적인 내용】
이하, 실시예를 통하여 본 발명을 더욱 상세히 설명하고자 한다. 이들 실시예는 오로지 본 발명을 보다 구체적으로 설명하기 위한 것으로, 본 발명의 요지에 따라 본 발명의 범위가 이들 실시예에 의해 제한되지 않는다는 것은 당업계에서 통상의 지식을 가진 자에 있어서 자명할 것이다. 실시예
실험방법
태깅된 트리아진 (tagged triazine) 화합물 라이브러리의 구축
태깅된 트리아진 화합물 라이브러리는 이전에 보고된 과정에 따라
제조되었다 (도 1; 5). 합성된 화합물들의 순도는 분석 HPUXAgilent Technologies, Snata Clara, USA)로 결정하였다. 저산소 상태 하에서 효율성을 유지하는 아팝토시스 (apoptosis) 유발인자들에 대한 스크리닝
HCT116 대장 암종 세포들 (ATCC)이 웰 당 5X103 세포의 밀도로 2세트의 96-웰 플레이트에 분주되었다. 5 mM 글루코오스 (Sigma)를 포함하는 '저농도 글루코오스 (Low glucose)' DMEMdnvitrogen, OR, USA) 배지가 스크리닝에 이용되었는데, 이는 2-DG(2-Deoxy-D-glucose; Sigma) 같은 해당과정 억제제들이 글루코오스와의 경쟁에서 효과적이기 때문이다. 24시간 후, 저산소 상태는 0.22 μΐΏ-여과된 150 μΜ CoCl2(Sigma)의 처리를 통해 한 세트의 96ᅳ웰 플레이트에서 유도되었고 이전에 기재된 바와 같이 (49) 배양 배지의 용량이 50% (웰 당 100 L)까지 감소되었다. 네 시간 후에 태깅된 트리아진 화합물 라이브러리 (5)로부터 테스트 화합물들이 2개의 독립적인 웰 (duplicate wells)에서 5 μΜ의 농도로 스크리닝되었다. 세포독성은 약물 처리 후 24시간 째에 ΜΤΤ 어세이를 통해 조사하였다. 추가적인 분석을 위한, '히트 (hit)' 는 정상산소 상태 (normoxia)와 비교하여 저산소 상태에서 25% 또는 그 이상의 세포독성 (MTT 어세이 흡광도로서 측정되었음)을 나타내는 화합물들로 분류되었다. 저산소 상태에서 선택적으로 암 세포들을 죽이는 해당과정 억제제인 2-DG(50 μΜ; 50)는 양성 대조군으로서 이용하였다.
CoCl2 처리-유도된 저산소 상태를 검증하기 위해, 에놀라제 발현이 조사되었다. 에놀라제 발현은 저산소 상태에 대한 반웅에서 증가하는 것으로 알려져 있다 (51). HCT116 암종 세포에 150 yM CoCl2의 4시간 등안 처리는 에놀라제 발현을 유도하였다 (도 2). 에놀라제 활성 어세이
래빗 근육에서 정제된 에놀라제는 Sigma-Aldrich로부터 구매하였다. 일 유니트의 에놀라제는 표준 어세이에서 분 당 포스포 -D- 글라이세레이트 (phospho-D— glycerate; Sigma)로부터 1 umol의 포스포에놀 피루베이트 (phosphoenol pyruvate)를 생산하는 효소의 양으로 정의된다 (52).
에놀라제 활성 어세이는 ENOblock 또는 NaF(Sigma)의 부존재 또는 존재 하에서 50 mM 이미다졸 (iinidazole)-HCl(pH 6.8)(JUNSEI) , 2.0 mM MgS04(JU SEI) 및 400 mM KCKJUNSEI)을 포함하는 완층액에서 정제된 에놀라제 (3-9 U)를 37T에서 반웅시킴으로써 실시하였다. 반웅은 1 urn 의 2-포스포 -D-글라이세레이트 (2-phospho-으 glycerate)를 첨가하여 개시하였으며, 0D 값은 10분의 반웅 시간 후 분광광도계 (spectrophotometer; VERSA max, Molecular Devices, USA)를 이용하여 240 nm에서 측정하였다. 제브라피쉬 종양 세포 이종이식 모델 (xenograft model)
제브라피쉬 (Lotte Mart, Gwangju, Republic of Korea)는 표준 프로토콜에 따라 유지되었다 (53). 제브라피쉬의 관리 및 처리는 광주과학기술원 (한국)의 동물 관리 및 윤리위원회 (Animal Care and Ethics Committees)에 의해 확립된 가이드라인에 따라 실시하였다. 제브라피쉬 배아들은 표준 교배 조건을 이용하여 얻어졌으며 세포 이종이식 (ceil ^01131 3 011)을 위해 수정 후 48시간 째까지 분화되었다. 하기와 같이 암 세포의 염색 후, 배아들은 미세-집게를 이용하여 탈융모화 (de- chor ionization)시키고 0.0016% 트리케인 (tr icaine; Sigma)으로 마취시킨 후, 1.0% 습식 (wet) 아가로오스 패드 (Invitrogen) 위에 오른쪽 사이드를 위치시켰다. 종양 세포들은 0.05% 트립신 (trypsin)-EDTA(Gibco)로 배양 접시로부터 분리되어 상온에서 두 번에 걸쳐서 PBS(Sigma)로 세척되었다. 세포들은 PBS에 희석된 Dil(2 ug/ml; Invitrogen)로 염색되고 네 번에 걸쳐서 세척되었다: FBS( Invitrogen)로 한 번; PBS로 두 번; 및 PBS에 희석된 FBS(10¾)로 한 번. 이후, 세포들은 주입 전에 얼음에 놓았다. 암 세포들은 현미경 (OLYMPUS CKX41)에 의해 카운팅되었고, 10% FBS에 재현탁하여 이중 100개의 세포들은 보로실리케이트 글래스 모세관 (borosi licate glass capillaries; PV820 pneumatic icopump, World Precision Instruments)이 장착된 주입기를 이용하여 난황주머니 (yolk sac)의 중앙으로 주입되었다. 주입된 배아들은 200 mL의 메틸렌 블루가 없는 E3 배지에 희석된 목적 약물을 포함하는 96-웰 플레이트 (1개의 배아 /웰)에 옮겨 전-선택된 배양 온도에서 유지하였다. 주입 후 4일 째에,
주입 위치로부터 암 세포의 파종 (dissemination)을 나타내는 배아들의 수가 카운팅되고 업라이트 현미경 (Leica DM2500 microscope, Germany)으로 사진을 찍었다. 이동된 세포들의 수가 카운팅되고 난황주머니로부터 5개 이상의 형광 미세초점 거리 (microfoci distant)를 보여준 배아들이 세포 파종 (cell dissemination)으로 스코어링되었다. 제브라피쉬에서 글루코오스 섭취의 측정
수정 후 72시간 째 (72 hp f (hour post fertilization)), 치어 (larvae)를 96-웰 플레이트 (0.2 mM 2ᅳ페닐티오우레아 (phenylthiourea; Sigma)가 첨가된 200 의 E3 물을 포함하는 웰 당 6개의 난자)에 위치시켰다. 목적 약물을 1시간 동안 처리하였다. 이후ᅳ상기 용액은 600 μΜ 2-NBDG(Invitroge)가 첨가된 E3 물로 대체되어 3시간 동안 반웅시켰다. 상기 치어를 E3 물로 세척하고 0.02%트리케인-보충된 E3 물로 마취시켰다. 한 마리의 치어가 형광현미경 (DFC425C 디지털 카메라로 장착된 Leica DM2500 현미경) 관찰을 위해 3¾ 메틸셀를로오스 (methylceiluiose)를 포함하는 E3 물올 포함하는 챔버 슬라이드 (chamber slide)에 위치되었다. 이미지들은 50배율로 캡쳐되었다. 나머지 5마리의 치어들은 120 iiL의 CelLytic M 용액 (Sigma-Aldr ich) 및 초음파 (4°C, 10' ' /5' ' 필스, 10분)로 용해되었다. 용해된 치어들은 10분 동안 10,000 rpm으로 원심분리하였다. 상층액 (100 L)이 96-웰 플레이트로 옮겨진 후, 형광 판독기 (fluorescent microplate reader; SpectraMAX Gemini XS, Molecular Devices; Xex=466 nm, λεχ=540纖)를 이용하여 형광 측정되었다. 통계 분석
실험 그룹 간의 비교를 위핸 student' s t-검정이 이용되었다 (Microsoft Excel). 실시된 모든 테스트에서 0.05 이하의 广값이 통계적으로 유의성을 가지는 것으로 간주하였다. 특별히 언급되지 않는 한, 제시된 모든 결과들은 3개의 독립적인 실험들의 대표적인 예이다. 실험결과 ..
AP-in-a4(EN0block)의 동정
본 발명자들은 저산소 환경에서 암 세포들을 우선적으로 죽이는 분자들을 동정하기 위한 신규한 이중 스크리닝 시스템을 개발하였다 (도 la). 고형 지지체 (solid support) 상에 제조된 384개의 트리아진 화합물을 포함하는 하나의 작은 분자 라이브러리가 스크리닝에 이용되었다 (5; 도 3). '히트 (hit)' 분자는 정상상태정상산소 상태와 비교하여 저산소 상태에서 적어도 25% 이상의 암 세포독성 (ΜΊΤ 어세이 흡광도로서 측정되었음)올 나타내는 화합물들 (inducer)로 정의되었다. 384개의 트리아진계 화합물들 중 5개의 화합물이 저산소 상태 하에서 우수한 암 세포독성을 나타냈다. 가장 우수한 히트 분자는 AP-III-a4이었다 (1; 도 lb). 저산소 상태에서 배양된 암 세포들에 대한 AP-III-a4 처리는 투여량 -의존적으로 세포 생존율을 감소시켰다 (도 lc-d).
ENOblock은 에놀라제에 결합하여 이의 활성을 억제한다
본 발명자들은 AP-III-a4의 세포 내 타겟을 동정하기 위해 친화성 크로마토그래피 (Affinity chromatography)를 이용하였다. 본 연구에서 이용된 트리아진 화합물 라이브러리에 대한 타겟 동정 전략은 매우 간편한 (straight forward) 데, 이는 상기 분자들이 내장성 링커 모이어티 (built-in linker moiety)를 포함하기 때문이다. 상기 링커 모이어티는 친화성 매트릭스 (affinity matrix)으로의 컨쥬게이션을 가능하게 하는 데, 이를 통해 생물학적 활성의 감소될 수 있는 위험을 초래할 수 있다. AP-III-a4 친화성 매트릭스로부터 용출된 단백질들의 실버 염색 결과는 도 4a에 제시된다. 질량분석 (Mass spectrometry analysis)을 통해, 약 45 kD의 2개의 단백질 밴드들이 해당과정 효소인 에놀라제의 서브유니트라는 것을 확인하였다 (도 4b). 이에, 본 발명자들은 AP-III-a4 분자를 'ENOblock' 로 재명명하였다. 암 세포 용해물에서 에놀라제에 대한 ENOblock의 결합은 ENOblock 친화성 매트릭스로부터 용출된 단백질들에 대한 웨스턴 블랏 분석으로 다시 확인하였다. 프리 (free) ENOblock를 이용한 경쟁 분석은 ENOblock 친화성 매트릭스에 대한 에놀라제 결합을 억제하는 것을 보여주었다 (도 4c). 더욱이, ENOblock은 정제된 인간 에놀라제에 결합할 수 있었는데, 이는 ENOblock과 에놀라제 간의 직접적인 상호작용을 의미한다 (도 4d). 이후의 분석을 통해, ENOblock이 투여량-
의존적으로 에놀라제: 활성을 억제할 수 있다는 것을 확인하였다 (도 4e). 또한, 본 발명자들은 추가적인 대조군으로서 태깅된 트리아진 화합물 라이브러리로부터 얻어진 다론 비 -히트 화합물인 AP-I— Π0(3)을 테스트하였다. 상기 비 -히트 화합물은 에놀라제 활성을 감소시키지 않는 것으로 확인되었다 (도 5). 저산소 상태에서 암 세포 생존을 증대시키는 에놀라제의 기능은 siRNA-매개된 에놀라제 발현의 넉 -다운 (knock-down) HCT116 암 세포에서 재확인하였다 (도 6).
ENOblock은 암 세포 이동 (migration) 및 침윤 (invasion)을 억제한다
에놀라제는 고유의 해당과정 기능 (6)과 연관되지 않은 많은 기능들을 실행하는 '부가적 (moonlighting)' 기능을 하는 대사 단백질이다. 따라서, 본 발명자들은 ENOblock이 에놀라제의 부수적 기능들을 특징화하는 강력한 화학 수단올 의미한다고 추론하였다. 첫 번째 테스트로서, 본 발명자들은 암 세포 성장 (progression)에서의 에놀라제의 역할을 조사하였다 (도 7). 본 발명자들은 ENOblock에 의한 에놀라제 억제가 암 세포 침윤을 감소시킬 수 있다는 것을 발견하였는 데, 이는 우리의 지식 하에서 에놀라제 활성이 전이 (metastasis)와 연결되어 있다는 최초의 확인이었다 (도 7a-7b). 또한, 본 발명자들은 추가적인 대조군으로서 태깅된 트리아진 화합물 라이브러리로부터 얻어진 다른 화합물인 ΑΡ-Ι-Π0(3)을 테스트하였다. 상기 화합물은 세포 침윤 (cell invasion)을 감소시키지 않는 것으로 확인되었다 (도 8). 또한, ENOblock 처리는 암 세포 이동을 억제하였다 (도 7c-7d) . ENOblock 처리는 세포독성을 유발하지 않는 농도에서 정상산소상태에서 암 세포 침윤 /이동을 감소시켰다 (참조, 도 7a-7b와 도 lc의 비교). 이전의 연구들은 에놀라제 발현의 넉—다운은 항암제인 탁솔 (taxol) 및 빈크리스틴 (vincristine)에 의해 유발된 세포독성을 증가시킬 수 있다는 것을 보고하였다 (7). 이와 일치하게도, 본 발명자들은 ENOblock 처리도 탁솔 및 빈크리스틴에 의해 유도된 암 세포독성을 증가시 ¾ 수 있다는 것올 발견하였다 (도 7e). 어떻게 ENOblock가 암 세포독성올 유도하는 지를 조사하기 위해, 본 발명자들은 아팝토시스의 유도와 연결된 2개의 단백질 발현 (AKT(8) Bcl-xL(9))을 측정하였다. 그 결과, ENOblock처리는 AKT 및 Bcl-xL단백질의 발현을 감소시켰다 (도 7f).
제브라피쉬 G¾/7/i? rerio) 암 세포 이종이식 모델은 인 /^에서 항암제 후보물질의 테스팅에 있어서 증가하는 탁월한 연구 성과들을 얻기 위해 이용되는 유효하고 편리한 수단이다 (10, 11). 또한, 제브라피쉬는 포유동물에서 독성 효과 (toxicological effects)를 예측하기 위한 적절한 척추동물 플랫품이다 (12). 본 발명자들은 발생중인 제브라피쉬 치어에 10 μΜ ENOblock를 처라하는 것이 독성이 없음을 확인하였다 (도 9a-9c). 최근 공개된 제브라피쉬 종양 이종이식 모델을 이용하는 것이 항 -암제 테스팅에 유효하였다 (10). 본 발명자들은 ENOblock 처리가 암 세포 파종을 감소시킨다는 것을 관찰하였는데, 이는 ENOblock이 암 세포 이동 및 침윤 프로세스를 억제한다는 것을 의미한다 (도 9(1).
ENOblock은 세포 내 글루코오스 섭취 (uptake)를 유도하고 PEPCK 발현을 하향—조절 (down— regul at ion)한다
홍미롭게도, ENOblock (화합물 AP-III-a4)은, 그 활성 기작에 대해 규명하지 않았을 지라도, 글루코오스 섭취의 신규한 조절자를 발견하기 위한 스크리닝을 통해 이전에 동정된 트리아진 화합물들 중 하나였다 (13). 따라서, 본 발명자들은 세포 내 글루코오스 흐름 (flux)올 모니터링하는 데 이용할 수 있는 형광 프로브인 2-NBDG(2-(N-(7— nitrobenz-2-oxa— 1,3- d i az 1 -4-y 1 ) am i no ) -2-deoxyg 1 ucose; 14)를 이용하여 세포에서 글루코오스 섭취를 증가시키는 ENOblock의 활성을 확인하였다 (도 10a). 현재까지, 에놀라제 기능의 조절이 증가된 글루코오스 섭취와 연결되어 있다는 본 발명의 규명은 첫 번째 증명이다. 세포 내 글루코오스 섭취를 촉진하는 에놀라제의 역할은 siRNA-매개된 에놀라제 발현 넉-다운에 의해 재확인되었다 (도 11).
ENOblock이 글루코오스 섭취를 촉진시킬 수 있는 기작을 규명하기 위해, 본 발명자들은 글루코오스 항상성과 결합된 핵심 효소의 발현올 조사하였다. 본 발명자들은 ENOblock이 간세포 (hepatocytes)에서 PEPC (phosphoeno 1 pyr uvat e carboxykinase)의 발현을 하향ᅳ조절한다는 것을 확인하였는 데, 상기 PEPCK 효소는 간 조직 포도당 신생합성의 속도- 제한 (rate-limiting) 단계 (15)를 촉매하는 효소이다 (도 9b). 주지하다시피, GAPDS(4)가 글루코오스 섭취를 촉진하기 위해 GAPDH(glyceraldehyde 3-
phosphate dehydrogenase)를 타겟팅한다 (GAPDH는 에놀라제의 업스트림인 해당과정의 6번째 단계를 촉매한다)는 보고 (16)와 함께, 해당과정 효소에 대한 작은 분자 조절이 글루코오스 섭취를 조절한다는 선행 발견이 있다. 따라서 , 본 발명자들도 GAPDS 또는 PEPCK 발현을 하향-조절할 수 있는 것으로 잘 알려진 당뇨병 제제인 로지글리타존 (rosiglitazone; 5)을 처리한 후 간세포에서 PEPCK 발현을 측정하였다 (17). 본 발명자들은 로지글리타존이 PEPCK 발현을 하향-조절할 수 있는 반면에 GAPDS는 효과가 없다는 것을 발견하였는데, 이는 GAPDS 및 ENOblock이 다른 세포 내 기작들에 의해 글루코오스 섭취를 촉진한다는 것을 의미한다 (도 10b). 홍미롭게도, 본 발명자들은 포도당 신생합성의 위치 (18)인 신장에서 ENOblock 처리가 PEPCK 발현을 하향-조절할 수 있다는 것을 확인하였다 (도 5c). 이와 대조적으로, 신장 세포에서 GAPDS, 로지글리타존 또는 인술린의 처리는 PEPCK 발현에 영향을 미치지 않았다 (도 10c). G6Pase(glucose 6- phosphatase) 효소는 포도당 신생합성에서 마지막 단계를 촉매하고 간에 의한 글루코오스 섭취의 항상성 조절에서 핵심 기능을 한다 (19). : 본 발명자들은 간 세포에 대한 ENOblock 처리가 G6Pase 발현에 영향을 미치지 않는다는 것을 확인하였으며, 상기 G6Pase 발현은 GAPDS 또는 로지글리타존의 처리 후에도 영향을 받지 않았다 (도 10d). AMPKC5' AMP- activated protein kinase) 효소는 세포 내 에너지 항상성에서 핵심 기능을 한다 (20). G6Pase와 유사하게도, 본 발명자들은 ENOblock, GAPDS 또는 로지글리타존의 간 세포 처리가 AMPK 발현에 영향을 미치지 않는다는 것을 관찰하였다 (도 10d).
ENOblock은 PEPCK 발현을 하향-조절하고 인 비보에서 글루코오스 섭취를 유도한다
글루코오스 항상성에서 ENOblock의 인 비보 효과를 조사하기 위해, 본 발명자들은 제브라피쉬를 선택하였는데, 이는 상기 동물 모델이 적은 양의 테스트 화합물을 필요로 하는 편리하고 신속한 실험 포멧을 제공하기 때문이다. 더욱이, 제브라피시와 포유동물은 유사한 글루코오스 조절성 반웅들을 공유한다고 알려져 왔다 (17, 21). ENOblock 또는 로지글리타존으로 처리된 제브라피쉬 성어는 하향-조절된 간 ΡΕΚΣ 발현을
나타냈으며 (도 12a— 12b), 이는 본 발명자들의 세포-기반된 발견들을 재확인시켜 주었다. 형광성 글루코오스 프로브인 2-NBDG는 투명하여 2-NBDG 형광의 시각화가 가능한 제브라피쉬 치어 (22)에서 글투코오스 섭취를 평가하는 데 이용되어 왔다. 본 발명자들은 ENOblock 처리가 제브라피쉬 치어에서 글루코오스 섭취를 유도한다는 것을 관찰하였다 (도 12c-12d). 비교군으로서, 본 발명자들은 세포 내 글루코오스 섭취를 촉진하는 것으로 알려진 식물의 생물학적 활성 성분인 에모딘(6ᅳ111(^1^1-1,3,8- trihydroxyanthraquinone; 23)의 효과를 테스트하였다. 2-NBDG—처리된 치어에 대한 형광현미경 분석 결과 에모딘 처리는 글투코오스 섭취를 증가시켰다 (도 12e). 2-NBDG 섭취는 72 hfp에 제브라피쉬 치어의 눈에서 2- NBDG 형광 강도를 측정함으로써 정량화되었는데, 상기 조직이 상기 발달 단계에서 상대적으로 많은 수의 글투코오스 트랜스포터 이소형들 (glucose transporter isoforms)을 발현하는 것으로 알려져 있기 때문이다 (24). 이미지 J 분석 (National Institutes of Health, ,USA)은 ENOblock 또는 에모딘 처리가 제브라피쉬에서 글루코오스 섭취를 촉진할 수 있다는 것을 확증하였다. 제브라피쉬에서 글루코오스 섭취를 정량화하기 위한 택일적인 접근방법으로서, 용해된 치어에서 2-NBDG 형광 시그널을 형광 판독기를 이용하여 측정하였다 (도 13). 상기 접근방법을 통해 조사한 결과들도 ENOblock 처리가 인 에서 글루코오스 섭취를 유도한다는 것을 확인시켜 주었다.
ENOblock 처리는 지방세포분화 (adipogenesis) 및 거품세포 형성 (foam cell format ion)을 억제한다
당뇨병 환자에 대한 일반적으로 처방되는 약물들은 체중증가 (weight gain) 또는 심혈관계 질환들 (cardiovascular events) 같은 부작용들을 초래한다 (25). 이에, 본 발명자들은 지방세포 전구세포들의 분화 상에 지질 축적에 대한 ENOblock의 효과를 테스트하였으며, 이는 신규한 항ᅳ비만 제제들에 대한 편리한 테스트 방법을 제공한다 (26). 지방세포분화 상에 로지글리타존의 양성적 효과는 이미 보고되어 있다 (27). 이와 대조적으로, 본 발명자들은 ENOblock 처리가 지방세포형성 인자들 (adipogenic factors)에 노출된 지방세포 전구세포들에서 지질 축적을 억제한다는 것을
발견하였다 (도 14a). 혈관벽에 부착된 대식세포로부터의 거품세포 분화는 죽상동맥경화증 (atherosclerosis)의 진행에 필수적인 단계이다 (28). ENOblock 처리는 거품세포 분화를 거치도록 유도된 대식세포에서 지질 축적을 억제하였다 (도 14b). 세포 카운팅 결과, ENOblock 처리는 대식세포로부터의 거품세포 분화 및 단핵구로부터의 대식세포 분화 모두를 억제하였다 (도 14c-14d). 상술한 ENOblock의 억제 효과는 세포독성을 유발하지 않는 처리 농도에서 관찰되었다 (도 14e). 추가논의사항
본 연구에서, 본 발명자들은 에놀라제에 직접적으로 결합하는 최초의 비-기질성 유도체인 신규한 작은 분자인 ENOblock을 보고하며, 이는 생물학적 시스템에서 에놀라제 활성을 특징지을 수 있는 프로브로서 이용될 수 잇다. 에놀라제는 해당과정의 9번째 단계 (끝에서 두 번째 단계)인 2- 포스포 -D-글라이세레이트의
포스포에놀피루베이트 (phosphoenolpyruvate)로의 탈수화 (dehydration )를 촉매하는 금속이온 효소 (metalloenzyme)이다 (6). 에놀라제는 포도당 신생합성의 동화 경로의 한 부분으로서 기질들의 환경적 농도에 따라 역반웅을 촉매한다 (6). 또한, 에놀라제는 고유의 해당과정 상 기능과 관련없는 많은 기능들을 수행한다 (1, 6). 우리가 아는 한, 에놀라제의 다양한 기능들을 명확하게 알려주는 작은 분자 수단들은 거의 없다 . 가장 널리 보고된 에놀라제 억제제는 PhAH(phosphonoacetohydroxamate)이다. PhAH는 반웅의 중간산물인 카바니온 (carbanion)의 아시- 카르복실레이트 (aci-carboxylate) 형태를 모방하는 것으로 생각되고 있고, 결정학적 연구들을 위해서만 적용되고 있다 (29, 30). 다른 2개의 기질 유사체들 (D—tartronate semi aldehyde phosphate 및 3—aminoenol pyruvate phosphate)이 개발되었지만, 에놀라제 활성 위치의 직접적인 분광학적 적정 및 효소 동력학의 stopped-flow 연구들에만 적용되어 왔다 (31, 32). 더욱이 상술한 기질 유사체들은 상업적으로 유용하지 않다. 또한, 무기 소듬 플루오라이드 (sodium fluoride)도 에놀라제에 대한 기질 경쟁자이다. 하지만, 플루오라이드는 포스파타제 억제 및 증가된 산화적 스트레스 또는 흔란된 항—산화 방어 기작들의 유도와 같은 다양한 비-특이적 독성
W
효과들 (33, 34)로 인해 생물학적 시스템에서 에놀라제를 연구하는 데 적합하지 않다. 최근의 한 보고는 말라리아 약물인 메플로퀸 (mefloquine)을 Schistosoma z?a7soi?/)에서 에놀라제 억제제로 기술하였다 (34). 하지만ᅳ 메플로퀸은 상기 기생생물로부터 유래된 재조합 에놀라제에 직접적으로 결합할 수 없고 정제된 에놀라제 활성을 억제하지 못 했다. 이는, 본 발명의 ENOblock과는 달리, 메플로퀸이 에놀라제에 직접 결합하는 것이 아니라 세포 추출물 내 규명되지 않은 에놀라제-조절성 분자와의 상호작용을 통해 상기 효과를 나타낸다는 것을 의미한다. 예를 들에 에놀라제는 액포에 결합하거나 또는 마이토콘드리아와 연계된 거대 고분자 복합체 (macromolecular complexes)를 형성할 수 있는 것으로 알려져 왔다 (34). 따라서, 본 발명자들은 ENOblock이 에놀라제의 다양하고 비- 해당과정성 '부가적 기능들 (35)을 규명하기 위한 강력한 화학적 수단이라는 것을 제안한다.
ENOblock의 활성을 논의함에 있어, 본 발명자들은 저산소 상태 또는 정상산소 상태에서 상기 화합물의 효과들을 구별하는 것이 증요하다고 생각한다. 본 발명자들은 정상산소 상태와 비교하여 저산소 상태에서와 암 세포 사멸을 보다 크게 유도할 수 있는 화합물에 대해 스크리닝함으로써 ENOblock을 발견하였다. 본 발명자들은 일반적인 암 화학치료법을 위한 약물들이 저산소 상태에서 덜 효과적이기 때문에 (4) 상기 접근방법을 채택하였다. 저산소 상태의 활성화 이후에 에놀라제 발현이 빠르게 상향ᅭ 조절된다는 본 발명자들의 규명은 저산소 상태에서 암 세포들을 죽이고 (도 2) 에놀라제에 대한 억제 활성을 가지는 ENOMock와 연관되어 있다. 더 나아가, 암 세포들은 에너지 발생을 위한 동화성 해당과정의 증대시키는 일련의 대사 변형들을 보여주는 와버그 효과 (Warburg effect)에 의해 특징지을 수 있다 (2). 따라서, 3-브로모피루베이트 (bromopyruvate) 및 6- 아미노니코틴아마이드 (aminonicotinamide) 같은 해당과정 억제제들은 암 세포들을 죽일 수 있다 (3). 본 연구에서, 본 발명자들은 ENOblock이 저산소 상태 하에서 암 세포들을 선택적으로 죽일 수 있으며 (도 lc-ld), 이는 해당과정ᅳ관련된 에놀라제의 기능 (도 6)으로 인한 것미라는 것을 보였다. 또한, 에놀라제의 다양한 기능적 역할들은 정상산소 상태 하에서
ENOblock을 이용하여 탐침될 수 있다. 예를 들어, 에놀라제는 비-고전적
배출 경로 (non-classical export pathway)를 : 통해 진핵세포 표면으로 이동되고 암 세포 침윤을 매개하여 전이를 유발할 수 있는 것으로 제안되어져 왔다 (36, 37). 본 발명자들은 정상산소 상태에서 암 세포에 대한 ENOblock 처리가 비ᅳ세포득성 농도에서 암 세포 침윤 및 이동을 억제한다는 것을 관찰하였다 (도 7a-7d). 상술한 발견은 ENOblock이 암 세포 전이를 조절하기 위해 세포 표면-결합된 에놀라제를 타겟팅할 수 있다는 것을 의미한다. 암 전이와 관련하여, ENOblock 처리에 대한 본 발명자들의 인 비보 분석은 정상산소 상태에서도 실시되었다. 그 결과, 본 발명자들은 ENOblock이 암 세포 생존율에 현저한 영향을 미치지 않고 암 세포 전이를 억제할 수 있다는 것을 확인하였다 (도 9d-9e; 암 세포들은 세포 수의 감소 없이 주입 위치에서 유지되고 있는 것으로 보인다). 더 나아가, 에놀라제는 미세소관 (microtubule)을 파괴하는 항암제들의 효과에 음성적으로 영향을 미칠 수 있는 세포 미세소관 시스템과 연관되어 있는 것으로 알려져 있었다 (7, 38). 본 발명자들의 결과들은 ENOblock 처리가 암 세포들을 죽이는 미세소관-불안정화 약물들의 효과를 증가시킬 수 있다는 것을 보여준다 (도 7e). 따라서, 본 발명자들은 암 세포 전이를 억제하고 정상산소 상태에서 미세소관-불안정화 약물들의 효과를 증대시킬 뿐 아니라 종양 내 저산소 상태의 환경 (niches)에서 암 세포들을 선택적으로 죽일 수 있는 강력한 활성을 가지는 ENOblock을 암 치료법을 위한 강력한 신규 약물 후보라고 추론한다 (암 줄기세포들은 저산소 상태 하에서 생존율을 촉진하는 저산소 상태-유도성 인자들올 발현하는 것으로 알려져 있다; 39).
한편, 본 발명자들의 연구는 ENOblock에 의한 에놀라제 억제가 세포 내 글루코오스 섭취를 유도할 수 있다는 것을 보여주었다 (도 10a). 본 발명자들의 결과들은 글루코오스 섭취를 증가시키는 ENOblock의 활성이 인슬린에 의해서도 억제 (40)되는 PEPCK의 억제로 인한 것이라는 것을 보여준다. 간에서의 PEPCK 억제는 포도당 신생합성을 감소시킬 뿐 아니라 동시에 글루코오스 섭취를 촉진한다 (15, 41). 흥미롭게도, 신장에서의 포도당 신생합성은 혈액 내 글루코오스 레벨을 조절하는 데 중요한 기능을 하며 (18), 본 발명자들의 결과들은 ENOblock이 신장 세포들에서 PEPCK 발현을 억제할 수 있고 인슐린 또는 당뇨병 약물인 로지글리타존은 억제하지 않는다는 것을 보여준다 (도 10c). 또한, 본 발명자들은 siR A를
이용한 에놀라제 넉-다운도 글루코오스 섭취를 유도할 수 있다는 것을 확인하였다 (도 15). ENOblock 처리가 PEPCK 발현을 인 ^^에서 억제하고 글루코오스 섭취를 유도할 수 있다는 본 발명자들의 발견은 ENOblock이 항- 당뇨병 약물로서 추가적인 개발을 위한 매력적인 후보물질일 수 있다는 것을 의미한다. 이것은 에놀라제 발현이 정상인과 비교하여 당뇨병 환자들에서 증가된다는 사실을 통해 뒷받침될 수 있다 (42). 더욱이, 본 발명자들의 결과들은 증가된 지방세포분화 및 거품세포 형성 (도 14) 같은 당뇨병 약물들의 사용에 따른 몇몇 합병증들을 억제한다는 것을 제시한다. 인슬린 시그널링은 촉진된 거품세포 형성과 연관되어 있으며 (43), 일반적으로 처방된 당뇨병 약물들은 지방세포분화 또는 체중증가를 유도할 수 있다 (27). 본 발명자들은 거품세포 형성 또는 지방세포분화를 억제하는 ENOblock의 활성이 항-당뇨병 약물 후보물질로서 개발될 수 있는 가능성을 더욱 더 지지해준다고 생각한다. 놀람게도, 가장 일반적으로 처방되고 있는 항-당뇨병 약물인 메트포르민 (metformin)도 간세포의 포도당 신생합성올 억제하고 최근에는 항-암 약물로서 다양한 임상실험들에 적용되고 있다 (44).
본 발명자들이 아는 한, 본 연구는 에놀라제 억제와 PEPCK 발현의 하향 -조절 (포도당 신생합성의 억제) 간의 최초의 연결점을 제공한다. 하지만, 상기 관계의 선례가 자연계에 존재한다. ^^o Aspergillus nidulans)^] 대한 연구들은 에놀라제를 인코딩하는 acuN 유전자의 돌연변이가 해당과정 탄소 소스 (glycolytic carbon sources)가 아니라 포도당 신생합성 탄소 소스 (gluconeogenic carbon sources)에 따른 성장 억제를 유도한다는 것을 보고하였다 (45). 따라서, 본 발명자들은 다른 해당과정 효소들이 포유동물에서 포도당 신생합성에 관여하는 조절성 기능을 가지는 지 여부를 평가하기 위한 이후의 연구들이 필요하다고 생각한다.
본 발명에서 제시된 ENOblock 처리의 인 비보 분석들은 제브라피쉬 척추동물 모델을 이용하였다. 제브라피쉬는 새로운 치료제를 테스트하기 위한 일차 동물로서 다음과 같은 많은 장점들을 가진다: (a) 포유동물에서 보존된 글루코오스 항상성 기작들; (b) 유효한 항암 약물을 테스팅하는 시스템의 유용성; (c) 포유동물 테스트와 관련된 독성 반웅들; 및 (d) 상대적으로 적은 양의 테스트 화합물의 필요성 (10, 12, 17, 21, 46). 또한,
포유동물 세포 반웅 (cell behavior)에 대한 매우 상세한 연구들이 제브라피쉬에서 실시될 수 있다 (47). 따라서, 본 발명자들의 제브라피쉬- 기반된 일련의 분석들은 ENOblock 활성의 최초 보고를 위해 적절한 포맷이다.
요약하면, 본 발명자들의 연구는 에놀라제에 직접적으로 결합하는 최초의 비 -기질 유사체 억제제이며 에놀라제의 다양한 비-해당과정 기능들을 탐침하는 데 이용될 수 있는 작은 분자인 ENOblock을 보고한다. 본 발명자들은 암 성장 상에 에놀라제 억제의 효과를 평가하는 데 ENOblock을 아용하였고, 에놀라제 억제가 암 세포 전이를 인 ^^에서 감소시킬 수 있다는 것을 최초로 보인다. 또한, 본 발명자들은 에놀라제 억제가 포도당 신생합성의 조절자인 PEPCK를 억제할 수 있으며 항-당뇨병 약물들을 개발하기 위한 신규한 타겟이라는 것을 처음으로 보인다. ENOblock의 발견은 인간 질환을 조절하는 규명되지 않은 세포 내 기작들에 대한 신규한 화학 프로브들, 약물 타켓들 및 후보물질 치료학을 제공하는 순화학유전학 (forward chemical genetics)의 종언 (testament)일 수 있다. 암 치료법을 위한 신규한 해당과정 억제제들의 발견의 필요성 이외에도, 박터)리 o 감염 (예컨대, Yersinia pest is, Borrelia spp. , Streptococcus pneumonia, 등) 및 기생층 (예컨대, trypanosomat id parasites; 48)의 병인학에서 에놀라제의 잠재적인 기능에 비추어 볼 때, 본 발명자들은 ENOblock이 상술한 질환들에 대한 이해를 현저하게 높일 수 있을 것으로 판단한다. 이상으로 본 발명의 특정한 부분을 상세히 기술하였는 바, 당업계의 통상의 지식을 가진 자에게 있어서 이러한 구체적인 기술은 단지 바람직한 구현 예일 뿐이며, 이에 본 발명의 범위가 제한되는 것이 아닌 점은 명백하다. 따라서, 본 발명의 실질적인 범위는 첨부된 청구항과 그의 등가물에 의하여 정의된다고 할 것이다. 참조문헌
1. Kim, J. W. and Dang , C. V. , Multi faceted roles
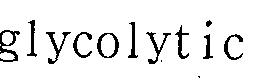 enzymes. Trends Biochem Sci 30 (3), 142 (2005) .
2. Warburg, 0. , Wind, F., and Negelein, E. , The Metabolism of Tumors in the Body. J Gen Physiol 8 (6), 519 (1927).
enzymes. Trends Biochem Sci 30 (3), 142 (2005) .
2. Warburg, 0. , Wind, F., and Negelein, E. , The Metabolism of Tumors in the Body. J Gen Physiol 8 (6), 519 (1927).
3. Pelicano, H. , Martin, D. S. , Xu, R. H. , and Huang, P., Glycolysis inhibit ion for anticancer treatment . Oncogene 25 (34), 4633 (2006) . 4. Xu, R. H. et al . , Inhibit ion of glycolysis in cancer cells: a novel strategy to overcome drug resistance associated with mitochondrial respiratory defect and hypoxia. Cancer Res 65 (2);, 613 (2005) .
5. Khersonsky, S. M. et al . , Facilitated forward chemical genetics using a tagged tr iazine library and zebraf ish embryo screening. J Am Chem Soc 125 (39), 11804 (2003) .
6. Pancho 1 i , V. , Multifunctional alphaᅳ enolase: : its role in diseases . Cell Mol Life Sci 58 (7), 902 (2001).
7. Georges , E., Bonneau , A. M. , and Prinos, P., RNAi -mediated knockdown of alphaᅳ enolase increases the sensitivity of tumor eel Is to ant i tubulin chemotherapeut ics . Int J Biochem Mol Biol 2 (4), 303
(2011) .
8. Kauf fmann-Zeh, A. et al., Suppression of c-Myc- induced apoptosis by Ras signalling through PI(3)K and PKB. Nature 385; (6616), 544 (1997).
9. Boise, L. H. et al . , bcl-x, a bcl-2-related gene that functions as a dominant regulator of apoptotic cell death. Cell 74 (4), 597 (1993).
10. Jung, D. . et al . , A novel zebraf ish human tumor xenograft model validated for ant i -cancer drug screening. Mol Biosyst 8 (7), 1930
(2012) .
11. Corkery , D. P., Dell aire, G. , and Berman , J. N. , Leukaemia xenotransplantation in zebraf ish― chemotherapy response assay in vivo.
Br J Haematol 153 (6), 786 (2011).
12. Sipes , N. S. , Padilla, S. , and Knudsen , T. B. , Zebraf ish: as an integrative model for twenty-first century toxicity testing. Birth Defects Res C Embryo Today 93 (3), 256 (2011).
13. Jung , D. W. et al . , Novel use of fluorescent glucose analogues to identify a new class of tr iazine一 based insulin mimetics possessing
useful secondary effects. Mol Biosyst 7 (2), 346 (2011).
14. Kim, W. H. , Lee, J., Jung, D. W. , and Williams, D. . , Visualizing sweetness- ' increasingly diverse applications for f luorescent-tagged glucose bioprobes and their recent structural modifications. Sensors (Basel) 12 (4), 5005 (2012).
15. Rosel la, G. et al., Impaired suppression of gluconeogenesis induced by overexpression of a noninsul in-responsive phosphoen이 pyruvate carboxykinase gene. Mol Endocrinol 7 (11), 1456 (1993).
16. Min, J. et al . , Forward chemical genetic approach identifies new role for GAPDH in insulin signaling. Nat Chem Biol 3 CO, 55 (2007) .
17. Elo, B. , Vi llano, C M. , Govorko, D. , and White, L. A., Larval zebraf ish as a model for glucose metabolism: expression of phosphoenol pyruvate carboxykinase as a marker for exposure to anti- diabetic compounds . J Mol Endocrinol 38 (4), 433 (2007).
18. Mitrakou, A., Kidney: its impact on glucose homeostasis and hormonal regulation. Diabetes Res Clin Pract 93 Suppl 1, S66 (2011) .
19. Hers , H. G. , Mechanisms of blood glucose homeostasis . J Inherit Me tab Dis 13 (4) , 395 (1990).
20. Hardie, D. G. and Carling, D. , The AMP-act ivated protein kinase— fuel gauge of the ma隱 alian cell? Eur J Biochem 246 (2), 259 (1997). 21. Jurczyk, A. et al . , Dynamic glucoregulat ion and mammal i an- 1 ike responses to metabolic and developmental disruption in zebraf ish. Gen Comp Endocrinol 170 (2), 334 (2011).
22. Jensen, P. J. , Gunter , L. B. , and Carayannopoulos , M. 0., Akt2 modulates glucose availability and downstream apoptot ic pathways during development. J Biol Chem 285 (23), 17673 (2010).
23. Yang, Y. et al . , Emodin with PPARgamma ligandᅳ binding activity promotes adipocyte differentiation and increases glucose uptake in 3T3-L1 cells. Biochem Biophys Res Commun 353 (2), 225 (2007) .
24. Tseng , Y, C. et al ., Specific expression and regul tion of glucose
transporters in zebraf ish ionocytes. Am J Physiol Regul Integr Comp Physiol 297 (2), R275 (2009) .
25. Agabegi , ED and Steven, S, Step-Up to Medicine (Step-Up Series) . (Lippincott Williams & Wilkins, 2008).
26. Zeng, X. Y. et al . , Screening for the efficacy on lipid accumulat ion in 3T3-L1 eel Is is an effective tool for the identification of new ant i -diabetic compounds . Biochem Pharmacol (2012).
27. Mads en, L. et al . , Adipocyte differentiation of 3T3-L1 preadipocytes is dependent on lipoxygenase activity during t e initial stages of the differentiation process. Biochem J 375 (Pt 3), 539 (2003).
28. Chait , A. , Progression of atherosclerosis: the cell biology. Eur Heart J 8 Suppl E, 15 (1987).
29. Zhang , E. , Hatada„ M., Brewer , J. M. , and Lebioda, L. , Catalytic metal ion binding in enolase: the crystal structure of an enolase— Mn2+-phosphonoacetohydroxamate com lex at 2.4— A resolution. Biochemistry 33 (20), 6295 (1994).
30. Carmiel i , R. et al., The catalytic Mn2+ sites in the enolaseᅳ inhibitor complex'- crystal lography , single-crystal EPR, and DFT calculations'. J Am Chem Soc 129 (14), 4240 (2007).
31. Brewer , J. M. , McKinnon, J. S. , and Phillips, R. S. , Stopped- flow studies of the reaction of D-tartronate semi aldehyde一 2ᅳ phosphate with human neuronal enolase and yeast enolase 1. FEBS Lett 584 (5), 979 (2010).
32. Spring, T. G. and Wold, F. , Studies on two high-affinity enolase inhibitors. Reaction with enolases. Biochemistry 10 (25) , 4655 (1971) .
33. Mansour , H. H. and Tawfik, S. S. , Efficacy of lycopene against fluoride toxicity in rats. Pharm Biol (2011) .
34. Manneck, T. , Keiser , J., and Muller, J., Mefloquine interferes with glycolysis in schistosomula of Schistosoma mansoni via inhibition
,pg s ().907 Getcs6320us.eni 171usidlanserill n A ^ ^^ :resss,cssori ino cot이 olualnIftins ret tfl-s Μ·0 Hr 3-

46. Selderslaghs, I. W. , Blust, R. , and Witters, H. E. , Feasibility study of the zebraf ish assay as an alternat ive method to screen for developmental toxicity and embryotoxicity using a training set of 27 compounds . Reprod Toxicol 33 (2), 142 (2012).
47. Zhu, S. et al . , Activated ALK collaborates with MYCN in neuroblastoma pathogenesis. Cancer Cell 21 (3) , 362 (2012) .
48. Ghosh , A. R. and Jacobs Lorena, M. , Surf ace expressed enolases of Plasmodium and other pathogens . Mem Inst Osvvaldo Cruz 106 Sup l 1, 85 (2011).
49. Piret, J. P., Mottet, D. , Raes, M. , and Michiels, C. , CoC12, a chemical inducer of hypoxia-inducible factor— 1 and hypoxia reduce apoptotic cell death in hepatoma cell line HepG2. Ann N Y Acad Sci 973 443 (2002). ,
50. Maher , J. C. , Kr i shan , A., and Lampidis , T. J. , Greater cell cycle inhibition and cytotoxicity induced by 2 deo y D— glucose in tumor eel Is treated under hypoxic vs aerobic conditions. Cancer Chemother Pharmacol 53 (2), 116 (2004) .
51. Sedoris, K. C. , Thomas , S. D. , and Miller, D. M. Hypoxia induces differential translation of enolase/MBP-1. BMC Cancer 10 157 (2010). 52. Gorsich, S. W. Barrows, V. , Halbert , J., and Farrar , W. W. , Purification and properties of ga麵 aga讓 a enolase from pig brain. J Protein Chem 18 (1), 103 (1999).
53. Nusslein-Volhard, C and Dahm, R, Zebraf ish: A Practical Approach (Practical Approach Series). (0UP, Oxford, 2002) .
Claims
【특허청구범위】
【청구항 II
(a) 하기 화학식 I로 표시되는 트리아진 (triazine)계 화합물의 치료학적 유효량; 및 (b) 약제학적으로 허용되는 담체를 포함하는 암 예방 또는 치료용 약제학적 조성물:

상기 화학식에서 , ^은 H 또는 d-C5 직쇄 또는 가지쇄의 알킬이고; ¾는 H, Ci-C5 직쇄 또는 가지쇄의 알킬, C厂 C5 직쇄 또는 가지쇄의 알킬알코올, -[(CH2)m-0]n-(CH2)p— NH2(상기 m, n 및 p는 각각 1-10의 정수이다), — [(C¾)m-0]n-C¾(상기 m 및 n은 각각 1-10의 정수이다), - [(CH2)m-0]n-(C¾)p— C¾(상기 m, n 및 p는 각각 1-10의 정수이다), -(CH2)q - (C0NH)-d-5 직쇄 또는 가지쇄의 알킬 (상기 q는 0-5의 정수이다), -(C¾)q- (CONH)-Ci-s 직쇄 또는 가지쇄의 알킬알코올 (상기 q는 으5의 정수이다), - (CH2)q-(C0NH)-[(CH2)m-0]n-(C¾)p-NH2(상기 m, n 및 p는 각각 1-10의 정수이고, 상기 q는 0-5의 정수이다), — (C¾)q-(C0NH)ᅳ [(C¾)ra-0]n-C¾(상기 m 및 n은 각각 1-10의 정수이고, 상기 Q는 으 5의 정수이다) 또 -(CH2)Q - (CONH)— [(C¾)m-0]n-(CH2)p-C¾(상기 m, n 및 p는 각각 1-10의 "정수이고, 상기 q는 0-5의 정수이다)이다.
【청구항 2】
제 1 항에 있어서, 상기 화학식 I로 표시되는 트리아진계 화합물은 하기 화학식 II로 표시되는 화합물인 것을 특징으로 하는 약제학적 조성물:

【청구항 3】
제 1 항에 있어서, 상기 조성물은 에놀라제의 활성을 억제하는 것을 특징으로 하는 약제학적 조성물ᅳ
【청구항 4】
제 1 항에 있어서, 상기 조성물은 암 세포의 이동 (migration), 침윤 (invasion) 및 전이 (metastasis)를 억제하는 것을 특징으로 하는 약제학적 조성물.
【청구항 5]
제 3 항 또는 제 4 항에 있어서, 상기 억제는 정상산소 상태 (normoxia)보다 저산소 상태 (hypoxia)에서 더욱 촉진되는 것을 특징으로 하는 약제학적 조성물.
【청구항 6】
제 1 항에 있어서, 상기 조성물은 아팝토시스-유도성 단백질 (apoptosis-inducible proteins)의 발현을 감소시키는 것을 특징으로 하는 약제학적 조성물.
【청구항 7】
제 6 항에 있어서, 상기 아팝토시스ᅳ유도성 단백질은 AKT 또는 Bcl- xL 단백질인 것을 특징으로 하는 약제학적 조성물.
【청구항 8】
제 1 항에 있어서, 상기 조성물은 인슐린 모방 활성 (insulin- mimicking activity)을 가지는 것올 특징으로 하는 약제학적 조성물.
【청구항 9】
제 8 항에 있어서, 상기 인슐린 모방 활성은 세포 내 글루코오스 섭취 (glucose uptake)를 촉진시키는 것을 특징으로 하는 약제학적 조성물.
【청구항 10】
제 1 항에 있어서, 상기 조성물은 PEPCK(phosphoenolpyruvate carboxykinase)의 발현을 하향 -조절 (down— regulat ion)하는 것을 특징으로 하는 약제학적 조성물.
【청구항 11]
제 1 항에 있어서, 상기 조성물은 지방세포분화 (adipogenesis) 및 거품세포 형성 (foam cell format ion)을 억제하는 것을 특징으로 하는 약제학적 조성물.
【청구항 12]
제 1 항에 있어서, 상기 암은 뇌암, 신경 내분비 암, 위암, 폐암, 유방암, 난소암, 간암, 기관지암, 비인두암, 후두암/ 췌장암, 방광암, 부신암, 대장암, 결장암, 자궁경부암, 전립선암, 골암, 피부암, 갑상선암, 부갑상선암 및 요관암으로 구성된 군으로부터 선택되는 암인 것을 특징으로 하는 약제학적 조성물.
【청구항 13]
제 1 항 내지 제 12 항 증 어느 한 항의 조성물올 유효성분으로 포함하는 에놀라제ᅳ관계된 질환 (enolase-associated disorders) 예방 또는 치료용 약제학적 조성물.
【청구항 14】
제 13 항에 있어서, 상기 에놀라제-관계된 질환은 자가면역질환 (auto immune disorders), 허혈 (ischemia) 및 박테리아 감염인 것을 특징으로 하는 약제학적 조성물.
【청구항 15】
(a) 하기 화학식 I로 표시되는 트리아진 (triazine)계 화합물의 치료학적 유효량; 및 (b) 약제학적으로 허용되는 담체를 포함하는 조성물을 대상 (subject)에게 투여하는 단계를 포함하는 암 예방 또는 치료 방법 :
화학식 I

【청구항 16】
제 15 항에 있어서, 상기 화학식 I로 표시되는 트리아진계 화합물은 하기 화학식 Π로 표시되는 화합물인 것을 특징으로 하는 방법:
화학식 II

【청구항 17】
제 15 항에 있어서, 상기 조성물은 에놀라제의 활성을 억제하는 것을 특징으로 하는 방법 .
【청구항 18]
제 15 항에 있어서, 상기 조성물은 암 세포의 이동 (migration), 침윤 (invasion) 및 전이 (metastasis)를 억제하는 것을 특징으로 하는 방법 .
【청구항 19】
제 17 항 또는 제 18 항에 있어서, 상기 억제는 정상산소 상태 (normoxia)보다 저산소 상태 (hypoxia)에서 더욱 촉진되는 것을 특징으로 하는 방법 . I
【청구항 20】
제 15 항에 있어서, 상기 조성물은 아팝토시스-유도성 단백질 (apoptosis-inducible proteins)의 발현을 감소시키는 것을 특징으로 하는 방법 .
【청구항 21】
제 20 항에 있어서, 상기 아팝토시스-유도성 단백질은 AKT 또는
Bcl-xL단백질인 것을 특징으로 하는 방법 .
【청구항 22】
제 15 항에 있어서, 상기 조성물은 인술린 모방 활성 (insulin- mimicking activity)을 가지는 것을 특징으로 하는 방법.
【청구항 23]
제 22 항에 있어서, 상기 인술린 모방 활성은 세포 내 글루코오스 섭취 (glucose uptake)를 촉진시키는 것을 특징으로 하는 방법 .
【청구항 24】
제 15 항에 있어서, 상기 조성물은 PEPCK(phosphoenolpyruvate carboxykinase)의 발현을 하향—조절 (down-regulation)하는 것을 특징으로 하는 방법 .
[청구항 25】
제 15 항에 있어서, 상기 조성물은 지방세포분화 (adipogenesis) 및 거품세포 형성 (foam cell formation)을 억제하는 것올 특징으로 하는 방법.
【청구항 26】
제 15 항에 있어서, 상기 암은 뇌암, 신경 내분비 암, 위암, 폐암 유방암 난소암, 간암, 기관지암, 비인두암, 후두암, 췌장암, 방광암, 부신암, 대장암, 결장암, 자궁경부암, 전립선암, 골암, 피부암, 갑상선암, 부갑상선암 및 요관암으로 구성된 군으로부터 선택되는 암인 것을 특징으로 하는 방법 .
【청구항 27】
제 15 항 내지 제 26 항 중 어느 한 항의 조성물을 유효성분으로 포함하는 에놀라제-관계된'질환 (enolase-associated disorders) 예방 또는 치료 방법 .
【청구항 28】
제 27 항에 있어서, 상기 에놀라제-관계된 질환은 자가면역질환 (autoi醒 une disorders), 허혈 (ischemia) 및 박테리아 감염인 것을 특징으로 하는 방법.
Priority Applications (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US14/437,277 US9364480B2 (en) | 2012-10-22 | 2013-10-22 | Pharmaceutical composition for preventing or treating cancer, containing enoblock as active ingredient |
| EP13849288.9A EP2910246A4 (en) | 2012-10-22 | 2013-10-22 | PHARMACEUTICAL COMPOSITION FOR THE PREVENTION OR TREATMENT OF CANCER WITH ENOBLOCK AS AN ACTIVE SUBSTANCE |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020120117417A KR101472083B1 (ko) | 2012-10-22 | 2012-10-22 | ENOblock을 유효성분으로 포함하는 암 예방 또는 치료용 약제학적 조성물 |
| KR10-2012-0117417 | 2012-10-22 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2014065572A1 true WO2014065572A1 (ko) | 2014-05-01 |
Family
ID=50544895
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/KR2013/009438 Ceased WO2014065572A1 (ko) | 2012-10-22 | 2013-10-22 | ENOblock을 유효성분으로 포함하는 암 예방 또는 치료용 약제학적 조성물 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US9364480B2 (ko) |
| EP (1) | EP2910246A4 (ko) |
| KR (1) | KR101472083B1 (ko) |
| WO (1) | WO2014065572A1 (ko) |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR102059649B1 (ko) * | 2017-02-07 | 2019-12-27 | 광주과학기술원 | ENOblock을 유효성분으로 포함하는 비만 예방 또는 치료용 약학 조성물 |
| EP4099996A4 (en) * | 2020-02-03 | 2024-02-21 | TDL Innovations, LLC | FOAM COMPOSITIONS FOR CANCER TREATMENT |
| CN113288899B (zh) * | 2021-05-17 | 2022-10-11 | 浙江工业大学 | 一种杂环硫醇类化合物在制备抗肿瘤药物中的应用 |
| CN113995841A (zh) * | 2021-11-04 | 2022-02-01 | 上海市胸科医院 | Mettl3/alkbh5/eno1调控轴作为靶位点在制备治疗肺腺癌药物中的应用 |
| WO2025096754A1 (en) | 2023-10-31 | 2025-05-08 | Musc Foundation For Research Development | The use of enolase inhibitor pomhex for treatment of fibrosis |
Citations (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5536722A (en) * | 1991-11-12 | 1996-07-16 | Pfizer Inc. | Triazine derivatives for enhancing antitumor activity |
| WO2003032903A2 (en) | 2001-10-12 | 2003-04-24 | New York University | Trisubstituted triazines compounds with antitubulin activity |
| WO2003050237A2 (en) | 2001-12-12 | 2003-06-19 | New York University | Triazine library with linkers |
| EP1479397A1 (en) * | 2002-02-05 | 2004-11-24 | Yamanouchi Pharmaceutical Co. Ltd. | 2,4,6-triamino-1,3,5-triazine derivative |
| WO2012015249A2 (ko) * | 2010-07-29 | 2012-02-02 | 광주과학기술원 | 당뇨병 치료제의 스크리닝 방법 |
| KR20120011664A (ko) * | 2010-07-29 | 2012-02-08 | 광주과학기술원 | 트리아진계 화합물을 유효성분으로 포함하는 당뇨병 예방 또는 치료용 약제학적 조성물 |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6262053B1 (en) | 1999-06-23 | 2001-07-17 | Parker Hughes Institute | Melamine derivatives as potent anti-cancer agents |
-
2012
- 2012-10-22 KR KR1020120117417A patent/KR101472083B1/ko not_active Expired - Fee Related
-
2013
- 2013-10-22 WO PCT/KR2013/009438 patent/WO2014065572A1/ko not_active Ceased
- 2013-10-22 EP EP13849288.9A patent/EP2910246A4/en not_active Withdrawn
- 2013-10-22 US US14/437,277 patent/US9364480B2/en not_active Expired - Fee Related
Patent Citations (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5536722A (en) * | 1991-11-12 | 1996-07-16 | Pfizer Inc. | Triazine derivatives for enhancing antitumor activity |
| WO2003032903A2 (en) | 2001-10-12 | 2003-04-24 | New York University | Trisubstituted triazines compounds with antitubulin activity |
| WO2003050237A2 (en) | 2001-12-12 | 2003-06-19 | New York University | Triazine library with linkers |
| EP1479397A1 (en) * | 2002-02-05 | 2004-11-24 | Yamanouchi Pharmaceutical Co. Ltd. | 2,4,6-triamino-1,3,5-triazine derivative |
| WO2012015249A2 (ko) * | 2010-07-29 | 2012-02-02 | 광주과학기술원 | 당뇨병 치료제의 스크리닝 방법 |
| KR20120011664A (ko) * | 2010-07-29 | 2012-02-08 | 광주과학기술원 | 트리아진계 화합물을 유효성분으로 포함하는 당뇨병 예방 또는 치료용 약제학적 조성물 |
Non-Patent Citations (57)
| Title |
|---|
| "Remington's Pharmaceutical Sciences", 1995 |
| AGABEGI, ED; STEVEN, S: "Step-Up to Medicine (Step-Up Series", 2008, LIPPINCOTT WILLIAMS & WILKINS |
| BOISE, L. H. ET AL.: "bcl-x, a bcl-2-related gene that functions as a dominant regulator of apoptotic cell death", CELL, vol. 74, no. 4, 1993, pages 597, XP024246088, DOI: doi:10.1016/0092-8674(93)90508-N |
| BREWER, J. M.; MCKINNON, J. S.; PHILLIPS, R. S.: "Stopped-flow studies of the reaction of D-tartronate semialdehyde-2-phosphate with human neuronal enolase and yeast enolase 1", FEBS LETT, vol. 584, no. 5, 2010, pages 979, XP026940314, DOI: doi:10.1016/j.febslet.2010.01.042 |
| CARMIELI, R. ET AL.: "The catalytic Mn2+ sites in the enolase-inhibitor complex: crystallography, single-crystal EPR, and DFT calculations", J AM CHEM SOC, vol. 129, no. 14, 2007, pages 4240 |
| CHAIT, A.: "Progression of atherosclerosis: the cell biology", EUR HEART J, vol. 8, no. E, 1987, pages 15 |
| CORKERY, D. P.; DELLAIRE, G.; BERMAN, J. N.: "Leukaemia xenotransplantation in zebrafish--chemotherapy response assay in vivo", BR J HAEMATOL, vol. 153, no. 6, 2011, pages 786 |
| ELO, B.; VILLANO, C. M.; GOVORKO, D.; WHITE, L. A.: "Larval zebrafish as a model for glucose metabolism: expression of phosphoenolpyruvate carboxykinase as a marker for exposure to anti-diabetic compounds", J MOL ENDOCRINOL, vol. 38, no. 4, 2007, pages 433, XP002751354, DOI: doi:10.1677/JME-06-0037 |
| GALLAGHER, E. J.; LEROITH, D.: "Diabetes, cancer, and metformin: connections of metabolism and cell proliferation", ANN N Y ACAD SCI, vol. 1243, 2011, pages 54 |
| GEORGES, E.; BONNEAU, A. M.; PRINOS, P.: "RNAi-mediated knockdown of alpha-enolase increases the sensitivity of tumor cells to antitubulin chemotherapeutics", INT J BIOCHEM MOL BIOL, vol. 2, no. 4, 2011, pages 303 |
| GHOSH, A. K.; JACOBS-LORENA, M.: "Surface-expressed enolases of Plasmodium and other pathogens", MEM INST OSWALDO CRUZ, vol. 106, no. 1, 2011, pages 85 |
| GORSICH, S. W.; BARROWS, V.; HALBERT, J.; FARRAR, W. W.: "Purification and properties of gammagamma-enolase from pig brain", J PROTEIN CHEM, vol. 18, no. 1, 1999, pages 103, XP019283972 |
| HARDIE, D. G.; CARLING, D.: "The AMPactivated protein kinase--fuel gauge of the mammalian cell?", EUR J BIOCHEM, vol. 246, no. 2, 1997, pages 259 |
| HERS, H. G.: "Mechanisms of blood glucose homeostasis", J INHERIT METAB DIS, vol. 13, no. 4, 1990, pages 395 |
| HYNES, M. J. ET AL.: "Transcriptional control of gluconeogenesis in Aspergillus nidulans", GENETICS, vol. 176, no. 1, 2007, pages 139 |
| IORI, E. ET AL.: "Glycolytic enzyme expression and pyruvate kinase activity in cultured fibroblasts from type 1 diabetic patients with and without nephropathy", BIOCHIM BIOPHYS ACTA, vol. 1782, no. 11, 2008, pages 627, XP025625145, DOI: doi:10.1016/j.bbadis.2008.08.012 |
| JEFFERY, C. J.: "Moonlighting proteins", TRENDS BIOCHEM SCI, vol. 24, no. 1, 1999, pages 8, XP027392942 |
| JENSEN, P. J.; GUNTER, L. B.; CARAYANNOPOULOS, M. 0.: "Akt2 modulates glucose availability and downstream apoptotic pathways during development", J BIOL CHEM, vol. 285, no. 23, 2010, pages 17673 |
| JUNG, D. W. ET AL.: "A novel zebrafish human tumor xenograft model validated for anti-cancer drug screening", MOL BIOSYST, vol. 8, no. 7, 2012, pages 1930 |
| JUNG, D. W. ET AL.: "Novel use of fluorescent glucose analogues to identify a new class of triazine-based insulin mimetics possessing useful secondary effects", MOL BIOSYST, vol. 7, no. 2, 2011, pages 346, XP055193380, DOI: doi:10.1039/C0MB00089B |
| JUNG, DA-WOON ET AL.: "Novel use of fluorescent glucose analogues to identify a new class of triazine-based insulin mimetics possessing useful secondary effects.", MOLECULAR BIOSYSTEMS, vol. 7, no. 2, 2011, pages 346 - 358, XP055193380 * |
| JURCZYK, A. ET AL.: "Dynamic glucoregulation and mammalian-like responses to metabolic and developmental disruption in zebrafish", GEN COMP ENDOCRINOL, vol. 170, no. 2, 2011, pages 334, XP027578784 |
| KAUFFMANN-ZEH, A. ET AL.: "Suppression of c-Myc-induced apoptosis by Ras signalling through PI(3)K and PKB", NATURE, vol. 385, no. 6616, 1997, pages 544, XP000942138, DOI: doi:10.1038/385544a0 |
| KHERSONSKY, S. M ET AL.: "Facilitated forward chemical genetics using a tagged triazine library and zebrafish embryo screening", J AM CHEM SOC, vol. 125, no. 39, 2003, pages 11804, XP002301137, DOI: doi:10.1021/ja035334d |
| KIM, J. W; DANG, C. V.: "Multifaceted roles of glycolytic enzymes", TRENDS BIOCHEM SCI, vol. 30, no. 3, 2005, pages 142, XP004777031, DOI: doi:10.1016/j.tibs.2005.01.005 |
| KIM, W. H.; LEE, J.; JUNG, D. W.; WILLIAMS, D. R.: "Visualizing sweetness: increasingly diverse applications for fluorescent-tagged glucose bioprobes and their recent structural modifications", SENSORS, vol. 12, no. 4, 2012, pages 5005 |
| LI, Z.; RICH, J. N.: "Hypoxia and hypoxia inducible factors in cancer stem cell maintenance", CURR TOP MICROBIOL IMMUNOL, vol. 345, 2010, pages 21 |
| LIU, KO-JIUNN; SHIH, NENG-YAO: "The Role of Enolase in Tissue Invasion and Metastasis of Pathogens and Tumor Cells", JOURNAL OF CANCER MOLECULES, vol. 3, no. 2, 2007, pages 45, XP055203817 |
| MADSEN, L. ET AL.: "Adipocyte differentiation of 3T3-L1 preadipocytes is dependent on lipoxygenase activity during the initial stages of the differentiation process", BIOCHEM J, vol. 375, 2003, pages 539, XP002269730, DOI: doi:10.1042/BJ20030503 |
| MAHER, J. C.; KRISHAN, A.; LAMPIDIS, T. J.: "Greater cell cycle inhibition and cytotoxicity induced by 2-deoxy-D-glucose in tumor cells treated under hypoxic vs aerobic conditions", CANCER CHEMOTHER PHARMACOL, vol. 53, no. 2, 2004, pages 116, XP002538171, DOI: doi:10.1007/S00280-003-0724-7 |
| MANNECK, T.; KEISER, J.; MULLER, J.: "Mefloquine interferes with glycolysis in schistosomula of Schistosoma mansoni via inhibition of enolase", PARASITOLOGY, vol. 139, no. 4, 2012, pages 497 |
| MANSOUR, H. H.; TAWFIK, S. S.: "Efficacy of lycopene against fluoride toxicity in rats", PHARM BIOL, 2011 |
| MILES, L. A. ET AL.: "Role of cell-surface lysines in plasminogen binding to cells: identification of alpha-enolase as a candidate plasminogen receptor", BIOCHEMISTRY, vol. 30, no. 6, 1991, pages 1682 |
| MIN, J. ET AL.: "Forward chemical genetic approach identifies new role for GAPDH in insulin signaling", NAT CHEM BIOL, vol. 3, no. 1, 2007, pages 55 |
| MITRAKOU, A.: "Kidney: its impact on glucose homeostasis and hormonal regulation", DIABETES RES CLIN PRACT, vol. 93, no. 1, 2011, pages 66 |
| NUSSLEIN-VOLHARD, C; DAHM, R: "Zebrafish: A Practical Approach (Practical Approach Series", 2002, OUP |
| O'BRIEN, R. M. ET AL.: "Identification of a sequence in the PEPCK gene that mediates a negative effect of insulin on transcription", SCIENCE, vol. 249, no. 4968, 1990, pages 533, XP000606183, DOI: doi:10.1126/science.2166335 |
| PANCHOLI, V.: "Multifunctional alpha-enolase: its role in diseases", CELL MOL LIFE SCI, vol. 58, no. 7, 2001, pages 902, XP002288673 |
| PELICANO, H.; MARTIN, D. S.; XU, R. H.; HUANG, P.: "Glycolysis inhibition for anticancer treatment", ONCOGENE, vol. 25, no. 34, 2006, pages 4633, XP055009657, DOI: doi:10.1038/sj.onc.1209597 |
| PIRET, J. P; MOTTET, D.; RAES, M.; MICHIELS, C.: "CoCl2, a chemical inducer of hypoxia-inducible factor-1, and hypoxia reduce apoptotic cell death in hepatoma cell line HepG2", ANN N Y ACAD SCI, vol. 973, 2002, pages 443 |
| QUINN, P. G.; YEAGLEY, D.: "Insulin regulation of PEPCK gene expression: a model for rapid and reversible modulation", CURR DRUG TARGETS IMMUNE ENDOCR METABOL DISORD, vol. 5, no. 4, 2005, pages 423 |
| ROSELLA, G. ET AL.: "Impaired suppression of gluconeogenesis induced by overexpression of a noninsulin-responsive phosphoenolpyruvate carboxykinase gene", MOL ENDOCRINOL, vol. 7, no. 11, 1993, pages 1456 |
| SAID ET AL., BMC MOL BIOL., vol. 8, 2007, pages 55 |
| SEDORIS, K. C.; THOMAS, S. D.; MILLER, D. M.: "Hypoxia induces differential translation of enolase/MBP-1", BMC CANCER, vol. 10, 2010, pages 157, XP021074996, DOI: doi:10.1186/1471-2407-10-157 |
| See also references of EP2910246A4 * |
| SEKIYA, M. ET AL.: "Ablation of neutral cholesterol ester hydrolase 1 accelerates atherosclerosis", CELL METAB, vol. 10, no. 3, 2009, pages 219 |
| SELDERSLAGHS, I. W.; BLUST, R.; WITTERS, H. E.: "Feasibility study of the zebrafish assay as an alternative method to screen for developmental toxicity and embryotoxicity using a training set of 27 compounds", REPROD TOXICOL, vol. 33, no. 2, 2012, pages 142, XP028907667, DOI: doi:10.1016/j.reprotox.2011.08.003 |
| SIPES, N. S.; PADILLA, S.; KNUDSEN, T. B.: "Zebrafish: as an integrative model for twenty-first century toxicity testing", BIRTH DEFECTS RES C EMBRYO TODAY, vol. 93, no. 3, 2011, pages 256 |
| SPRING, T. G.; WOLD, F.: "Studies on two high-affinity enolase inhibitors", REACTION WITH ENOLASES. BIOCHEMISTRY, vol. 10, no. 25, 1971, pages 4655 |
| TSENG, Y. C. ET AL.: "Specific expression and regulation of glucose transporters in zebrafish ionocytes", AM J PHYSIOL REGUL INTEGR COMP PHYSIOL, vol. 297, no. 2, 2009, pages R275 |
| WALSH, J. L.; KEITH, T. J.; KNULL, H. R.: "Glycolytic enzyme interactions with tubulin and microtubules", BIOCHIM BIOPHYS ACTA, vol. 999, no. 1, 1989, pages 64, XP023579167, DOI: doi:10.1016/0167-4838(89)90031-9 |
| WARBURG, 0.; WIND, F.; NEGELEIN, E.: "The Metabolism of Tumors in the Body", J GEN PHYSIOL, vol. 8, no. 6, 1927, pages 519 |
| XU, R. H. ET AL.: "Inhibition of glycolysis in cancer cells: a novel strategy to overcome drug resistance associated with mitochondrial respiratory defect and hypoxia", CANCER RES, vol. 65, no. 2, 2005, pages 613, XP002365256 |
| YANG, Y. ET AL.: "Emodin with PPARgamma ligand-binding activity promotes adipocyte differentiation and increases glucose uptake in 3T3-Ll cells", BIOCHEM BIOPHYS RES COMMUN, vol. 353, no. 2, 2007, pages 225 |
| ZENG, X. Y. ET AL.: "Screening for the efficacy on lipid accumulation in 3T3-L1 cells is an effective tool for the identification of new anti-diabetic compounds", BIOCHEM PHARMACOL, 2012 |
| ZHANG, E.; HATADA, M.; BREWER, J. M.; LEBIODA, L.: "Catalytic metal ion binding in enolase: the crystal structure of an enolase-Mn2+-phosphonoacetohydroxamate complex at 2.4-A resolution", BIOCHEMISTRY, vol. 33, no. 20, 1994, pages 6295 |
| ZHU, S. ET AL.: "Activated ALK collaborates with MYCN in neuroblastoma pathogenesis", CANCER CELL, vol. 21, no. 3, 2012, pages 362, XP028473638, DOI: doi:10.1016/j.ccr.2012.02.010 |
Also Published As
| Publication number | Publication date |
|---|---|
| US20150246050A1 (en) | 2015-09-03 |
| KR20140052184A (ko) | 2014-05-07 |
| EP2910246A1 (en) | 2015-08-26 |
| EP2910246A4 (en) | 2015-09-30 |
| US9364480B2 (en) | 2016-06-14 |
| KR101472083B1 (ko) | 2014-12-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Huang et al. | Bruceine D inhibits HIF-1α-mediated glucose metabolism in hepatocellular carcinoma by blocking ICAT/β-catenin interaction | |
| Amengual-Cladera et al. | Metformin: from diabetes to cancer—unveiling molecular mechanisms and therapeutic strategies | |
| Rana et al. | Small molecule adenosine 5′-monophosphate activated protein kinase (AMPK) modulators and human diseases | |
| Park et al. | Berberine inhibits human colon cancer cell migration via AMP-activated protein kinase-mediated downregulation of integrin β1 signaling | |
| Zuo et al. | Targeting PP2A with lomitapide suppresses colorectal tumorigenesis through the activation of AMPK/Beclin1-mediated autophagy | |
| Fan et al. | Ameliorative effects of mangiferin derivative TPX on insulin resistance via PI3K/AKT and AMPK signaling pathways in human HepG2 and HL-7702 hepatocytes | |
| Zhu et al. | Extracellular α-crystallin protects astrocytes from cell death through activation of MAPK, PI3K/Akt signaling pathway and blockade of ROS release from mitochondria | |
| Qiao et al. | Curcumin prevents neuroinflammation by inducing microglia to transform into the M2-phenotype via CaMKKβ-dependent activation of the AMP-activated protein kinase signal pathway | |
| WO2014065572A1 (ko) | ENOblock을 유효성분으로 포함하는 암 예방 또는 치료용 약제학적 조성물 | |
| Huang et al. | Signaling pathways involved in HSP32 induction by hyperbaric oxygen in rat spinal neurons | |
| Chen et al. | Novel direct AMPK activator suppresses non-small cell lung cancer through inhibition of lipid metabolism | |
| Guerra et al. | Protein kinase CK2 inhibition is associated with the destabilization of HIF-1α in human cancer cells | |
| Ren et al. | Fraxetin inhibits the growth of colon adenocarcinoma cells via the Janus kinase 2/signal transducer and activator of transcription 3 signalling pathway | |
| Li et al. | Cardioprotective effects of Amentoflavone by suppression of apoptosis and inflammation on an in vitro and vivo model of myocardial ischemia-reperfusion injury | |
| Meng et al. | Bazedoxifene-induced ROS promote mitochondrial dysfunction and enhance osimertinib sensitivity by inhibiting the p-STAT3/SOCS3 and KEAP1/NRF2 pathways in non-small cell lung cancer | |
| Ye et al. | Secondary metabolites of mulberry leaves exert anti-lung cancer activity through regulating the PD-L1/PD-1 signaling pathway | |
| CN113813387A (zh) | Ppar激动剂在制备治疗急性髓细胞白血病的药物中的应用 | |
| Zhou et al. | Negative regulation of Sirtuin 1 by AMP-activated protein kinase promotes metformin-induced senescence in hepatocellular carcinoma xenografts | |
| Yang et al. | The flavonoid baicalin improves glucose metabolism by targeting the PH domain of AKT and activating AKT/GSK 3β phosphorylation | |
| Zong et al. | Macelignan inhibits the inflammatory response of microglia and regulates neuronal survival | |
| Yang et al. | Chlorquinaldol alleviates lung fibrosis in mice by inhibiting fibroblast activation through targeting methionine synthase reductase | |
| Bai et al. | The small molecule P7C3-A20 exerts neuroprotective effects in a hypoxic–ischemic encephalopathy model via activation of PI3K/AKT/GSK3β signaling | |
| Feng et al. | ATPR-induced differentiation and G0/G1 phase arrest in acute promyelocytic leukemia by repressing EBP50/NCF1 complex to promote the production of ROS | |
| Che et al. | The therapeutic effects of Longikaurin A, a natural ent-kauranoid, in esophageal squamous cell carcinoma depend on ROS accumulation and JNK/p38 MAPK activation | |
| He et al. | Identification of a molecular activator for insulin receptor with potent anti-diabetic effects |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 13849288 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 14437277 Country of ref document: US |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2013849288 Country of ref document: EP |