WO2016013870A1 - 완전한 이뮤노글로불린 형태의 항체를 세포막을 투과하여 세포질에 위치시키는 방법 및 그의 이용 - Google Patents
완전한 이뮤노글로불린 형태의 항체를 세포막을 투과하여 세포질에 위치시키는 방법 및 그의 이용 Download PDFInfo
- Publication number
- WO2016013870A1 WO2016013870A1 PCT/KR2015/007626 KR2015007626W WO2016013870A1 WO 2016013870 A1 WO2016013870 A1 WO 2016013870A1 KR 2015007626 W KR2015007626 W KR 2015007626W WO 2016013870 A1 WO2016013870 A1 WO 2016013870A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- antibody
- chain variable
- variable region
- light chain
- cell
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images




































Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/50—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/32—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against translation products of oncogenes
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/46—Hybrid immunoglobulins
Definitions
- viruses HSV
- toxins choleratoxin, diphtheria toxin
- endocytosis an active intracellular transport mechanism.
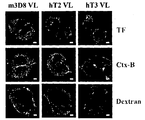
- These intracellularizations are largely classified into three pathways, including clathrin-induced endocytosis, which is involved in ligand-induced cellular internalization, or by caveolae, which is found in some toxins such as choleratoxin, dextran, and ebolavirus.
- caveolae which is found in some toxins such as choleratoxin, dextran, and ebolavirus.
- the endocytosis involving clathrin and caveolae begins primarily when membrane receptors bind to specific ligands.
- the present invention is to infiltrate the cell membrane of living animal cells through the process of endocytosis and actively immobilize the complete immunoglobulin-type antibody in the cytoplasm using the endosomal escape mechanism To provide a way.
- the cytoplasmic penetrating ability may be located in the cytoplasm by endosome escape after actively infiltrating living cells through endocytosis.
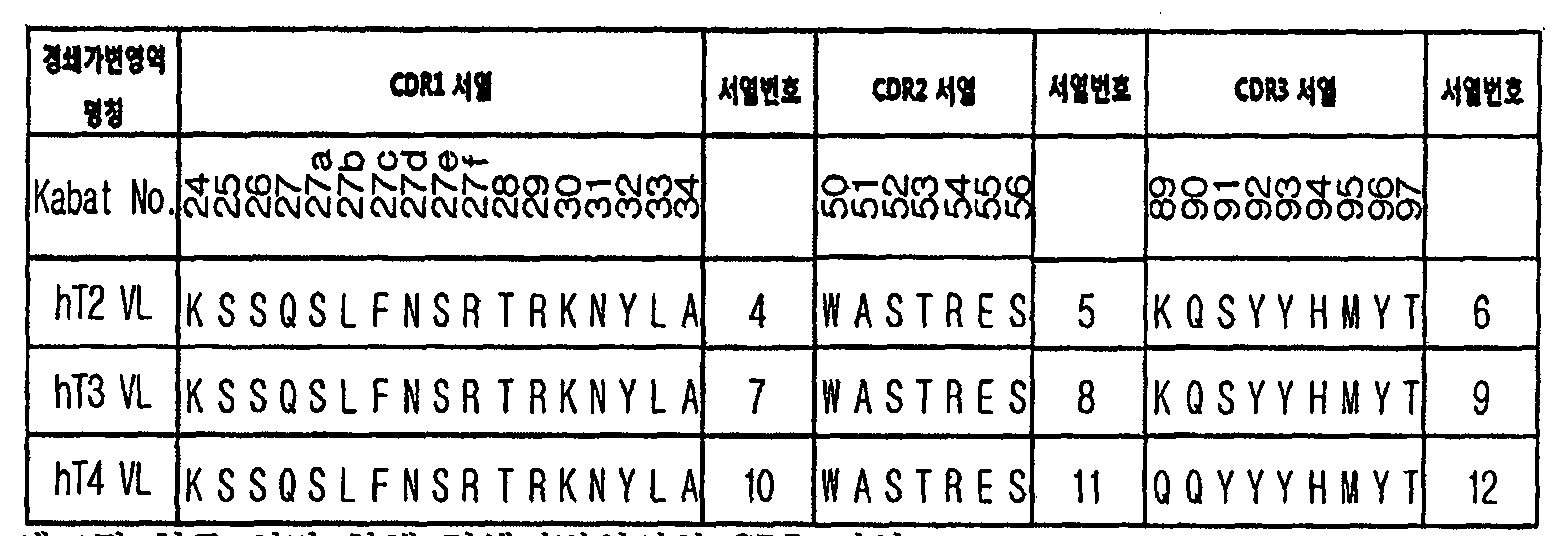
- the light chain variable region may include CDR1 of SEQ ID NO: 4, CDR2 of SEQ ID NO: 5, and CDR3 of SEQ ID NO: 6.
- the light chain variable region may include CDR1 of SEQ ID NO. 7, CDR2 of SEQ ID NO. 8, and CDR3 of SEQ ID NO.
- the heavy chain variable region that specifically binds to and inhibits the RAS was screened by the following method.
- KRas G12D which is an activated (GTP coupled) RAS mutant
- carcinogenesis-related RAS mutations occur mainly at residues 12, 13 and 61, and residues 12 and 13 are located in the P-loop of the RAS protein and are bound to the RAS protein.
- Hydrolysis of GTP affects the binding of GAP (GTPase-activating protein), which induces protein structural changes in an inactive form.
- residue 61 binds to the hydrolytic activity of GAP and prevents GTP hydrolysis. Therefore, various carcinogenic RAS mutations have the same region as the RAS G12D mutation and signal transduction (Switch I, Switch II). It is not limited to.
- liposomes are composed of one or more lipid bilayer membranes surrounding an aqueous internal compartment that can associate themselves.
- Liposomes can be specified by membrane type and size.
- Small unilamellar vesicles SUVs
- Large uni-lamellar vesicles LUV
- Oligolamella large vesicles and multilamellar large vesicles have multiple, generally concentric, membrane layers and may be 100 nm or more in diameter. Liposomes with multiple asymmetrical membranes, ie several small vesicles contained within larger vesicles, are called multivesicular vesicles.
- the present invention also provides a pharmaceutical composition for preventing or treating cancer, including a bioactive molecule selected from the group consisting of the antibody, or a peptide, a protein, a small molecule drug, a nanoparticle, and a liposome fused thereto.
- a bioactive molecule selected from the group consisting of the antibody, or a peptide, a protein, a small molecule drug, a nanoparticle, and a liposome fused thereto.
- Figure 2b is a diagram comparing the model structure using the WAM modeling of m3D8 VL and humanized light chain variable region monodomain hT0 VL, mutant hT2 VL, hT3 VL using a superimposing method.
- Figure 6d is a result of agarose gel nucleic acid hydrolysis verification experiment to confirm the nucleic acid hydrolytic ability in cytotransmab through human light chain variable region (hT4) substitution having a cell penetrating ability grafting the CDRs of autoimmune mouse-derived antibody.
- hT4 human light chain variable region
- Figure 8a is a result of observing the degree of cell penetration according to the concentration of TMab4 under a confocal microscope.
- FIG. 10 is a result of observing with a confocal microscope through a pulse-chase experiment to observe the transport process and stability of TMab4 introduced into the cell.
- FIG. 25 shows the results of in vitro evaluation of cell growth inhibition by treatment of RGD-TMab4 and RGD-RT4 in HCT116 and PANC-1 cell lines.
- Figure 1 is a schematic diagram showing the concept of a monoclonal antibody of the complete IgG form located in the cytoplasm through the cell infiltration named cytotransmab, in order to understand the cytoplasmic penetration of the humanized antibody light chain variable region in order to implement this, the light chain variable derived from the existing mouse Reference was made to the correlation of cytoplasmic permeability of the region single domain m3D8 VL with CDRs belonging to the light chain variable region fragment (Lee et al., 2013).
- Example 3 Cellular Infiltration Cellular infiltration capacity and cell infiltration mechanism validation of humanized light chain variable region (VL) single domain.
- VL variable region
- Figure 3a is a result of observing the cytoplasmic penetration capacity of the light chain variable region single domain by confocal microscopy (confocal microscopy).
- Figure 3b is a result of observation by confocal microscopy to verify the cytoplasmic penetration mechanism of the light chain variable region monodomain.
- the existing literature has approved hT3 VL and FDA based on data on the location of interface residues between human antibody variable regions, the frequency and binding of specific interface residues located on opposite variable regions, and the frequency of use of interface residues in human antibodies.
- the interface between the heavy and light chain variable regions of the therapeutic antibodies Bevacizumab (Avastin) and Adalimumab (Humira) was analyzed (Vargas-Madrazo and Paz-Garcia, 2003).
- residues 89 and 91 included in CDR3 involved in binding between variable regions in mouse-derived CDRs of hT3 VL are regions of high human antibody use and may affect the structure of CDR3 of heavy chain variable region (VH). It was confirmed that there is.
- the two residues were mutated to amino acids that are frequently used in human antibodies to develop hT4 VL that can be optimized for binding to human antibody heavy chain variable regions.
- the heavy chain variable regions (Bevacizumab VH, Adalimumab VH, fused with DNA encoding the secretion signal peptide at the 5 'end to construct a heavy chain expression vector for production in the form of monoclonal antibodies in the form of complete IgG)
- DNA encoding the heavy chain comprising the humanized hT0 VH) and the heavy chain constant region (CH1-hinge-CH2-CH3) was cloned into NotI / HindIII in the pcDNA3.4 (Invitrogen) vector, respectively.
- Proteins were purified from cell culture supernatants harvested with reference to standard protocols.
- the antibody was applied to a Protein A Sepharose column (GE healthcare) and washed with PBS (pH 7.4).
- the antibody was eluted at pH 3.0 with 0.1 M glycine buffer and then immediately neutralized with 1 M Tris buffer.
- the eluted antibody fraction was concentrated by exchanging buffer with PBS (pH7.4) through dialysis. Purified protein was quantified using absorbance and extinction coefficient at 280 nm wavelength.
- an elution buffer (12 mM phosphate, pH 7.4, 500 mM NaCl, 2.7 mM KCl) (SIGMA) having a high salt was used to suppress nonspecific binding with the resin due to electrical attraction due to basic residues.
- the flow rate is 0.5 ml / min.
- Proteins used as protein size markers were dehydrogenase (150 kDa), albumin (66 kDa), carbonic anhydrase (29 kDa). In all monoclonal antibodies and cytotransmabs, one pole was measured and confirmed to exist as a monolith.
- cytotransmab After combining the cytotransmab and monoclonal antibodies TMab4, Bevacizumab, Adalimumab, AvaT4, HuT4 and washed three times for 10 minutes with 0.1% PBST.
- the labeling antibody binds to an alkaline phosphatase-conjugated anti-human mAb (SIGMA) conjugated with goat derived AP. 405 nm absorbance was quantified by reaction with p-nitrophenyl palmitate (pNPP) (SIGMA).
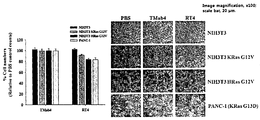
- ⁇ 10 4 cells HeLa, PANC-1) per well in a 96 well plate were diluted in 0.1 ml of medium containing 10% FBS, respectively, and cultured at 12 hours, 37 degrees, and 5% CO 2 conditions. Thereafter, 1 ⁇ M of TMab4, HuT4, Adalimumab, AvaT4, and Bevacizumab were treated for 20 hours or 44 hours, and then 20 ⁇ l of MTT solution (1 mg / ml PBS) was added and further incubated for 4 hours. Formed fomazan was dissolved in 200 ⁇ l of DMSO (Dimethyl Sulfoxide), and cell viability was determined by measuring absorbance at 595 nm with an absorbance meter.
- DMSO Dimethyl Sulfoxide
- pLJM1 a Lenti virus vector
- pLJM1 a Lenti virus vector
- 3 x 10 6 HEK293T cells were placed in 1 ml of medium containing 10% FBS and incubated at 5% CO 2 , 37 ° C for 12 hours.
- GFP11-SBP2 For expression of GFP11-SBP2 fused cytotransmab animal cells, GFP11-SBP2 was genetically fused to the heavy chain C-terminus using three GGGGS linkers. Afterwards, transient transfection of HEK293F protein-expressing cells simultaneously with the animal expression vector encoding GFP11-SBP2 and the animal expression vector encoding the cytoplasmic infiltration light chain and endosomal escape ability were performed. Was done. After the purification of GFP11-SBP2 fused cytoplasmic penetration monoclonal antibody was carried out in the same manner as in Example 5.
- VH variable domain monodomain
- FR (framework) of the used library uses IGHV3-23 * 04, J H 4, which are the most commonly used V genes in conventional antibodies, and a library having 9 residues in CDR3 length was used.
- IGHV3-23 * 04, J H 4 which are the most commonly used V genes in conventional antibodies
- a library having 9 residues in CDR3 length was used.
- the construction of the library and yeast surface expression methods are described in detail in a previously published paper (Baek and Kim, 2014).
- Selected Escherichia coli was cultured in LB medium in the presence of 100 ⁇ g / ml ampicillin antibiotic to absorbance of 0.6 at 37 to 600 nm, and then 0.1 mM IPTG was added for protein expression, followed by further incubation at 30 degrees for 5 hours. . Thereafter, E. coli was collected using a centrifuge, and then E. coli was pulverized using ultrasonic waves (SONICS). Only the supernatant from which E. coli pulverization was removed using a centrifuge was purified using Glutathione resin (Clontech), which specifically purified GST-tagged proteins.
- Glutathione resin Clontech
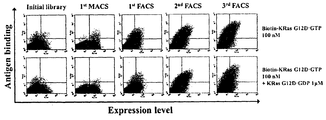
- the conjugated KRas G12D antigen was reacted with a library-expressed yeast at room temperature for 1 hour, followed by reaction with PE conjugated streptavidin (Streptavidin-R-phycoerythrin conjugate (SA-PE) (Invitrogen) to produce FACS (Fluorescence). Suspension was confirmed by activated cell sorting (FACS Caliber) (BD biosciences). After selection of the next screening condition by FACS analysis, antigens were bound to yeast expressing the suspended library under the same conditions as above, and then suspended by FACS ariaII instrument.
- Humanized heavy chain variable chain libraries suspended by primary MACS and primary FACS screening were expressed on the yeast surface in the form of Fab after yeast conjugation with yeast secreting cytoplasmic infiltration light chain single chain (hT4 VL), followed by secondary FACS, 3 The second FACS screening process was conducted.
- FIG. 16 is a flow cytometric analysis of binding ability of GTP-coupled KRas G12D alone and competitive conditions with GDP-coupled KRas G12D in the selection step to obtain specific high affinity to the above-described GTP-bound KRas G12D. Data. Through this, the heavy chain variable region (VH) -dependent library selection capable of specific binding to GTP-coupled KRas G12D was confirmed.
- VH heavy chain variable region
- RT4 clones were finally selected through individual clone analysis from a library having high affinity and specificity for GTP-coupled KRas G12D protein through high-speed screening.
- FIG. 17 is an analysis of 12% SDS-PAGE in reducing or non-reducing conditions after purification of anti-Ras.GTP iMab RT4.
- the cell penetrating ability of anti-Ras.GTP iMab RT4 was observed in cell lines with KRas mutations (PANC-1, HCT116) and cell lines with KRas wild type (HT29, HeLa).
- Figure 26a is an experimental result confirming the tumor growth inhibitory effect of RGD-fused anti-Ras.GTP iMab RT4 in mice transplanted with HCT116 cell line.
- Figure 26b is a graph measuring the weight of rats to identify non-specific side effects of RGD fused anti-Ras.GTP iMab RT4.
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Medicinal Chemistry (AREA)
- Organic Chemistry (AREA)
- Immunology (AREA)
- General Health & Medical Sciences (AREA)
- Public Health (AREA)
- Biophysics (AREA)
- Epidemiology (AREA)
- Engineering & Computer Science (AREA)
- Veterinary Medicine (AREA)
- Pharmacology & Pharmacy (AREA)
- Biochemistry (AREA)
- Animal Behavior & Ethology (AREA)
- Genetics & Genomics (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Microbiology (AREA)
- Mycology (AREA)
- Oncology (AREA)
- Peptides Or Proteins (AREA)
Abstract
Description
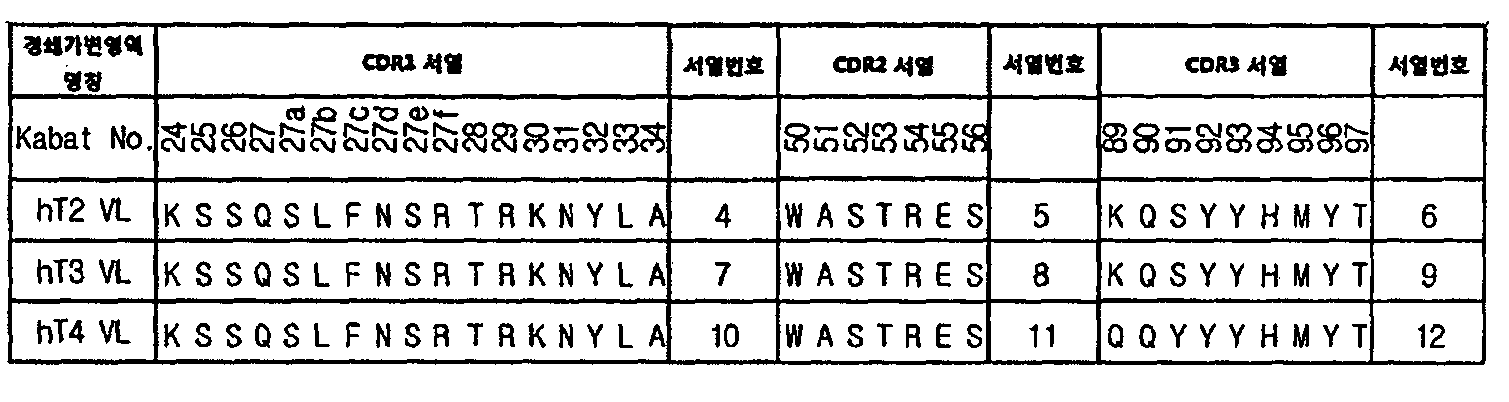



표 1

표 2



Claims (44)
- 완전한 이뮤노글로불린(immunoglobulin) 형태의 항체를 세포막을 투과하여 세포질에 위치시키는 방법으로서, 상기 항체는 세포질 침투능을 가지는 경쇄가변영역 (VL)을 포함하는 것인 방법.
- 청구항 1에 있어서, 상기 항체는 키메릭, 인간, 또는 인간화된 항체인 것인, 방법.
- 청구항 1에 있어서 상기 항체는 IgG, IgM, IgA, IgD 및 IgE로 이루어진 군으로부터 선택된 것인, 방법.
- 청구항 1에 있어서, 상기 세포질 침투능은 세포 내재화를 통해 침투한 후 엔도좀 탈출에 의한 것인, 방법.
- 청구항 1에 있어서, 상기 경쇄 가변영역은서열번호 4, 7 및 10으로 이루어진 군으로부터 선택된 아미노산 서열로 이루어진 CDR1 또는 이와 상동성이 90% 이상인 서열; 및서열번호 6, 9, 및 12로 이루어진 군으로부터 선택된 아미노산 서열로 이루어진 CDR3 또는 이와 상동성이 90% 이상인 서열을 포함하는 것인, 방법.
- 청구항 1에 있어서, 상기 경쇄가변영역 (VL)은 경쇄 가변 영역의 N 말단으로부터 2번째 및 4번째 아미노산이 각각 류신 (leucine, L) 및 메티오닌 (Methionine, M)으로 치환된 것인, 방법.(단, 상기 아미노산 위치는 카바트 (Kabat) 번호에 따름.)
- 청구항 1에 있어서, 상기 경쇄 가변 영역은 경쇄 가변 영역의 N 말단으로부터 9, 10, 13, 17, 19, 21, 22, 42, 45, 58, 60, 79, 및 85번째 아미노산이 각각세린 (serine, S), 세린 (serine, S), 알라닌 (Alanine, A), 발린 (Valine, V), 아스파트 산 (aspartic acid, D), 발린 (Valine, V), 이소류신 (Isoleucine, I), 트리오닌 (Threonine, T), 라이신 (Lysine, K), 라이신 (Lysine, K), 발린 (Valine, V), 세린 (serine, S), 글루타민 (Glutamine, Q) 및 트리오닌 (Threonine, T)으로 치환된 것인, 방법.(단, 상기 아미노산 위치는 카바트 (Kabat) 번호에 따름.)
- 청구항 1에 있어서, 상기 경쇄 가변 영역은 경쇄 가변 영역의 N 말단으로부터 89 번째 및 91번째 아미노산이 각각 글루타민 (Glutamine, Q) 및 타이로신 (Tyrosine, Y)으로 치환된 것인, 방법.(단, 상기 아미노산 위치는 카바트 (Kabat) 번호에 따름.)
- 청구항 1에 있어서, 상기 경쇄 가변영역은 서열번호 1, 2, 및 3으로 이루어진 군으로부터 선택된 아미노산으로 이루어진 것인, 방법.
- 청구항 1에 있어서, 상기 항체는 살아있는 세포에 능동적으로 침투하는 것인, 방법.
- 청구항 1에 있어서, 상기 항체는 세포질, 핵, 미토콘드리아, 소포체, 세포내 고분자 (organelle macromolecule)를 표적하는 것인, 방법.
- 청구항 11에 있어서, 상기 세포내 고분자는 단백질, 지질, DNA 또는 RNA인 것인, 방법.
- 청구항 12에 있어서 상기 단백질은 세포 성장, 세포증식, 세포주기, DNA 수리, DNA 보전, 전사, 복제, 번역 또는 세포내 이동의 제어와 관련된 것인, 방법.
- 청구항 12에 있어서, 상기 단백질은 인산기, 카르복실산기, 메틸기, 설페이트기, 지질, 수산기, 또는 아미드기로 변형 또는 활성화되거나, 돌연변이된 것인, 방법.
- 청구항 1에 있어서, 상기 항체는 세포질 내 활성화된 RAS에 특이적으로 결합하는 것인 방법.
- 청구항 15에 있어서, 상기 결합은 세포질 내의 활성화된 RAS에 특이적으로 결합하는 중쇄 가변영역 (VH)에 의한 것인, 방법.
- 청구항 15에 있어서, 상기 활성화된 RAS는 돌연변이된 형태인 것인, 방법.
- 청구항 16에 있어서, 상기 중쇄가변영역은서열번호 14의 CDR1 또는 이와 상동성이 90% 이상인 아미노산 서열;서열번호 15의 CDR2 또는 이와 상동성이 90% 이상인 아미노산서열; 및서열번호 16의 CDR3 또는 이와 상동성이 90% 이상인 아미노산 서열;을 포함하는 것인, 방법.
- 청구항 16에 있어서, 상기 중쇄가변영역은 서열번호 13의 아미노산으로 이루어진 것인, 방법.
- 완전한 이뮤노글로불린(immunoglobulin) 형태의 항체가 세포막을 투과하여 세포질에 위치하는 것을 유도하는 경쇄가변영역 (VL).
- 청구항 20에 있어서, 상기 경쇄가변영역은서열번호 4, 7 및 10으로 이루어진 군으로부터 선택된 아미노산 서열로 이루어진 CDR1 또는 이와 상동성이 90% 이상인 서열; 및서열번호 6, 9, 및 12로 이루어진 군으로부터 선택된 아미노산 서열로 이루어진 CDR3 또는 이와 상동성이 90% 이상인 서열;을 포함하는 것인, 경쇄가변영역 (VL).
- 청구항 20에 있어서, 상기 경쇄가변영역 (VL)은 경쇄 가변 영역의 N 말단으로부터 2번째 및 4번째 아미노산이 각각 류신 (leucine, L) 및 메티오닌 (Methionine, M)으로 치환된 것인, 경쇄가변영역 (VL).(단, 상기 아미노산 위치는 카바트 (Kabat) 번호에 따름.)
- 청구항 20에 있어서, 상기 경쇄가변영역은 경쇄 가변 영역의 N 말단으로부터 9, 10, 13, 17, 19, 21, 22, 42, 45, 58, 60, 79, 및 85번째 아미노산이 각각세린 (serine, S), 세린 (serine, S), 알라닌 (Alanine, A), 발린 (Valine, V), 아스파트 산 (aspartic acid, D), 발린 (Valine, V), 이소류신 (Isoleucine, I), 트리오닌 (Threonine, T), 라이신 (Lysine, K), 라이신 (Lysine, K), 발린 (Valine, V), 세린 (serine, S), 글루타민 (Glutamine, Q) 및 트리오닌 (Threonine, T)으로 치환된 것인, 경쇄가변영역 (VL).(단, 상기 아미노산 위치는 카바트 (Kabat) 번호에 따름.)
- 청구항 20에 있어서, 상기 경쇄가변영역은 경쇄 가변 영역의 N 말단으로부터 89 번째 및 91번째 아미노산이 각각 글루타민 (Glutamine, Q) 및 타이로신 (Tyrosine, Y)으로 치환된 것인, 경쇄가변영역 (VL).(단, 상기 아미노산 위치는 카바트 (Kabat) 번호에 따름.)
- 청구항 20에 있어서, 상기 경쇄 가변영역은 서열번호 1, 2, 및 3으로 이루어진 군으로부터 선택된 아미노산 서열로 이루어진 것인, 경쇄가변영역 (VL).
- 청구항 20에 있어서, 상기 세포질 침투능은 세포 내재화를 통해 침투한 후 엔도좀 탈출에 의한 것인, 경쇄가변영역 (VL).
- 청구항 20 내지 26 중 어느 한 항의 경쇄가변영역을 포함하는 항체.
- 청구항 27에 있어서, 상기 항체는 세포막을 투과하여 세포질에 위치하는 것인, 항체
- 청구항 27에 있어서, 상기 항체는 키메릭, 인간, 또는 인간화된 항체인 것인, 항체.
- 청구항 27에 있어서, 항체는 IgG, IgM, IgA, IgD 및 IgE로 이루어진 군으로부터 선택된 것인, 항체.
- 청구항 27에 있어서, 상기 항체는 세포질, 핵, 미토콘드리아, 소포체, 세포내 고분자 (organelle macromolecule)를 표적하는 것인, 항체.
- 청구항 31에 있어서, 상기 세포내 고분자는 단백질, 지질, DNA 또는 RNA인 것인, 항체.
- 청구항 32에 있어서, 상기 단백질은 세포 성장, 세포증식, 세포주기, DNA 수리, DNA 보전, 전사, 복제, 번역 또는 세포내 이동의 제어와 관련된 것인, 항체.
- 청구항 32에 있어서, 상기 단백질은 인산기, 카르복실산기, 메틸기, 설페이트기, 지질, 수산기, 또는 아미드기로 변형 또는 활성화되거나, 돌연변이된 것인, 항체.
- 청구항 27에 있어서, 상기 항체는 세포질 내 활성화된 RAS에 특이적으로 결합하는 것인 항체.
- 청구항 35에 있어서, 상기 결합은 세포질 내의 활성화된 RAS에 특이적으로 결합하는 중쇄 가변영역 (VH)에 의한 것인, 항체.
- 청구항 35에 있어서, 상기 활성화된 RAS는 돌연변이된 형태인 것인, 항체.
- 청구항 36에 있어서, 상기 중쇄가변영역은서열번호 14의 CDR1 또는 이와 상동성이 90% 이상인 아미노산 서열;서열번호 15의 CDR2 또는 이와 상동성이 90% 이상인 아미노산서열; 및서열번호 16의 CDR3 또는 이와 상동성이 90% 이상인 아미노산 서열;을 포함하는 것인, 항체.
- 청구항 36에 있어서, 상기 중쇄가변영역은 서열번호 13의 아미노산으로 이루어진 것인, 항체.
- 청구항 27 내지 39 중 어느 한 항의 항체에 융합된, 펩타이드, 단백질, 소분자 약물, 나노입자 및 리포좀으로 이루어진 군으로부터 선택된 생체활성분자.
- 청구항 27 내지 39 중 어느 한 항의 항체, 또는 이에 융합된 펩타이드, 단백질, 소분자 약물, 나노입자 및 리포좀으로 이루어진 군으로부터 선택된 생체활성분자를 유효성분으로 포함하는, 암의 예방 또는 치료용 약학적 조성물.
- 청구항 27 내지 39 중 어느 한 항의 항체, 또는 이에 융합된 펩타이드, 단백질, 소분자 약물, 나노입자 및 리포좀으로 이루어진 군으로부터 선택된 생체활성분자를 포함하는, 암의 진단용 조성물.
- 청구항 20 내지 26 중 어느 한 항의 경쇄가변영역 또는, 청구항 27 내지 39 중 어느 한 항의 항체를 코딩하는 폴리뉴클레오티드.
- 항체의 경쇄가변영역을, 살아있는 세포 내부로 침투하여 세포질에 위치하는 특성을 가진 경쇄가변영역으로 치환하는 단계를 포함하는,세포내부로 침투하여 세포질에 위치하는 항체의 제조하는 방법.
Priority Applications (11)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| RU2017103675A RU2017103675A (ru) | 2014-07-22 | 2015-07-22 | Способ размещения в цитоплазме антител в виде полного иммуноглобулина путем проникновения антител через клеточную мембрану и его применение |
| EP15825418.5A EP3173428B1 (en) | 2014-07-22 | 2015-07-22 | Method for positioning, in cytoplasm, antibody having complete immunoglobulin form by penetrating antibody through cell membrane, and use for same |
| CA2955272A CA2955272A1 (en) | 2014-07-22 | 2015-07-22 | Method for positioning, in cytoplasm, antibody having complete immunoglobulin form by penetrating antibody through cell membrane, and use for same |
| JP2017525495A JP6595592B2 (ja) | 2014-07-22 | 2015-07-22 | 完全型免疫グロブリン形態の抗体を細胞膜を透過して細胞質に位置させる方法およびその利用 |
| AU2015292955A AU2015292955B2 (en) | 2014-07-22 | 2015-07-22 | Method for positioning, in cytoplasm, antibody having complete immunoglobulin form by penetrating antibody through cell membrane, and use for same |
| US15/327,369 US10844136B2 (en) | 2014-07-22 | 2015-07-22 | Method for positioning, in cytoplasm, antibody having complete immunoglobulin form by penetrating antibody through cell membrane, and use for same |
| BR112017001304-5A BR112017001304A2 (ko) | 2014-07-22 | 2015-07-22 | The complete method for placing the antibody of the immunoglobulin type is transmitted through the cell membrane into the cytoplasm, and use his |
| CN201580045139.0A CN107001481A (zh) | 2014-07-22 | 2015-07-22 | 通过使抗体穿透细胞膜将具有完全免疫球蛋白形式的抗体定位在细胞质中的方法及其用途 |
| MX2017001010A MX2017001010A (es) | 2014-07-22 | 2015-07-22 | Metodo para posicionar en citoplasma, anticuerpo que tiene forma de inmunoglobulina completa penetrando el anticuerpo a traves de la membrana celular y uso para el mismo. |
| IL250203A IL250203B (en) | 2014-07-22 | 2017-01-19 | Method for positioning, in cytoplasm, antibody having complete immunoglobulin form by penetrating antibody through cell membrane, and use for same |
| ZA2017/00488A ZA201700488B (en) | 2014-07-22 | 2017-01-20 | Method for positioning, in cytoplasm, antibody having complete immunoglobulin form by penetrating antibody through cell membrane, and use for same |
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR20140092673 | 2014-07-22 | ||
| KR10-2014-0092673 | 2014-07-22 | ||
| KR1020150103163A KR101602870B1 (ko) | 2014-07-22 | 2015-07-21 | 완전한 이뮤노글로불린 형태의 항체를 세포막을 투과하여 세포질에 위치시키는 방법 및 그의 이용 |
| KR10-2015-0103163 | 2015-07-21 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2016013870A1 true WO2016013870A1 (ko) | 2016-01-28 |
Family
ID=55163334
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/KR2015/007626 Ceased WO2016013870A1 (ko) | 2014-07-22 | 2015-07-22 | 완전한 이뮤노글로불린 형태의 항체를 세포막을 투과하여 세포질에 위치시키는 방법 및 그의 이용 |
Country Status (1)
| Country | Link |
|---|---|
| WO (1) | WO2016013870A1 (ko) |
Cited By (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2017204606A1 (ko) * | 2016-05-27 | 2017-11-30 | 오름테라퓨틱 주식회사 | 세포질 침투 항체 및 이의 용도 |
| CN109790212A (zh) * | 2016-05-27 | 2019-05-21 | 奥隆制药 | 细胞溶质穿透抗体及其用途 |
| KR20190132338A (ko) * | 2019-11-21 | 2019-11-27 | 오름테라퓨틱 주식회사 | 세포질 침투 항체 및 이의 용도 |
| WO2019235426A1 (ja) | 2018-06-04 | 2019-12-12 | 中外製薬株式会社 | 細胞質内での半減期が変化した抗原結合分子 |
| WO2019235581A1 (ja) | 2018-06-06 | 2019-12-12 | 国立大学法人大阪大学 | Regnase-1が関与する疾患の治療および/または予防方法 |
| WO2020047345A1 (en) | 2018-08-31 | 2020-03-05 | Yale University | Compositions and methods of using cell-penetrating antibodies in combination with immune checkpoint modulators |
| US10787487B2 (en) | 2018-06-21 | 2020-09-29 | Orum Therapeutics Inc. | Cell/tissue-specific cell-penetrating antibodies |
| US10851177B2 (en) | 2014-07-22 | 2020-12-01 | Orum Therapeutics Inc. | Method for inhibiting intracellular activated RAS using intact immunoglobulin-type antibody having cytosol-penetrating ability and use thereof |
| US10961301B2 (en) | 2011-04-01 | 2021-03-30 | Yale University | Cell-penetrating anti-DNA antibodies and uses thereof inhibit DNA repair |
| US11590242B2 (en) | 2016-06-15 | 2023-02-28 | Yale University | Antibody-mediated autocatalytic, targeted delivery of nanocarriers to tumors |
| WO2024081736A2 (en) | 2022-10-11 | 2024-04-18 | Yale University | Compositions and methods of using cell-penetrating antibodies |
| EP4267745A4 (en) * | 2020-12-23 | 2025-06-18 | Chugai Seiyaku Kabushiki Kaisha | Antigen-binding molecules with improved cytosol-penetrating activity |
| US12486318B2 (en) | 2016-06-15 | 2025-12-02 | Yale University | Anti-guanosine antibody as a molecular delivery vehicle |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20130266570A1 (en) * | 2012-03-30 | 2013-10-10 | Richard H. Weisbart | Targeting Intracellular Target-binding Determinants with Intracellular Antibodies |
-
2015
- 2015-07-22 WO PCT/KR2015/007626 patent/WO2016013870A1/ko not_active Ceased
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20130266570A1 (en) * | 2012-03-30 | 2013-10-10 | Richard H. Weisbart | Targeting Intracellular Target-binding Determinants with Intracellular Antibodies |
Non-Patent Citations (5)
Cited By (18)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US10961301B2 (en) | 2011-04-01 | 2021-03-30 | Yale University | Cell-penetrating anti-DNA antibodies and uses thereof inhibit DNA repair |
| US10851177B2 (en) | 2014-07-22 | 2020-12-01 | Orum Therapeutics Inc. | Method for inhibiting intracellular activated RAS using intact immunoglobulin-type antibody having cytosol-penetrating ability and use thereof |
| CN109790212A (zh) * | 2016-05-27 | 2019-05-21 | 奥隆制药 | 细胞溶质穿透抗体及其用途 |
| JP2019520086A (ja) * | 2016-05-27 | 2019-07-18 | オラム・セラピューティクス・インコーポレイテッド | 細胞質浸透抗体およびその用途 |
| WO2017204606A1 (ko) * | 2016-05-27 | 2017-11-30 | 오름테라퓨틱 주식회사 | 세포질 침투 항체 및 이의 용도 |
| EP3466970A4 (en) * | 2016-05-27 | 2020-01-15 | Orum Therapeutics Inc. | ANTIBODIES PENETRATING IN CYTOSOL AND USES THEREOF |
| AU2017271189B2 (en) * | 2016-05-27 | 2020-02-20 | Orum Therapeutics Inc. | Cytosol-penetrating antibody and use thereof |
| US11155641B2 (en) | 2016-05-27 | 2021-10-26 | Orum Therapeutics Inc. | Cytosol-penetrating antibody and use thereof |
| US12486318B2 (en) | 2016-06-15 | 2025-12-02 | Yale University | Anti-guanosine antibody as a molecular delivery vehicle |
| US11590242B2 (en) | 2016-06-15 | 2023-02-28 | Yale University | Antibody-mediated autocatalytic, targeted delivery of nanocarriers to tumors |
| WO2019235426A1 (ja) | 2018-06-04 | 2019-12-12 | 中外製薬株式会社 | 細胞質内での半減期が変化した抗原結合分子 |
| WO2019235581A1 (ja) | 2018-06-06 | 2019-12-12 | 国立大学法人大阪大学 | Regnase-1が関与する疾患の治療および/または予防方法 |
| US10787487B2 (en) | 2018-06-21 | 2020-09-29 | Orum Therapeutics Inc. | Cell/tissue-specific cell-penetrating antibodies |
| WO2020047345A1 (en) | 2018-08-31 | 2020-03-05 | Yale University | Compositions and methods of using cell-penetrating antibodies in combination with immune checkpoint modulators |
| KR102091195B1 (ko) | 2019-11-21 | 2020-03-19 | 오름테라퓨틱 주식회사 | 세포질 침투 항체 및 이의 용도 |
| KR20190132338A (ko) * | 2019-11-21 | 2019-11-27 | 오름테라퓨틱 주식회사 | 세포질 침투 항체 및 이의 용도 |
| EP4267745A4 (en) * | 2020-12-23 | 2025-06-18 | Chugai Seiyaku Kabushiki Kaisha | Antigen-binding molecules with improved cytosol-penetrating activity |
| WO2024081736A2 (en) | 2022-10-11 | 2024-04-18 | Yale University | Compositions and methods of using cell-penetrating antibodies |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| WO2016013870A1 (ko) | 완전한 이뮤노글로불린 형태의 항체를 세포막을 투과하여 세포질에 위치시키는 방법 및 그의 이용 | |
| WO2019098682A1 (ko) | 항-her2 항체 또는 그의 항원 결합 단편, 및 이를 포함하는 키메라 항원 수용체 | |
| WO2014189303A1 (ko) | 뉴로필린에 특이적인 종양 침투성 펩타이드 및 이 펩타이드가 융합된 융합 단백질 | |
| WO2016153276A1 (ko) | 뉴로필린1 특이적 결합 펩타이드 및 이 펩타이드가 융합된 융합 단백질, 및 이의 용도 | |
| WO2015005632A1 (ko) | Dll4와 vegf에 특이적으로 결합하는 신규 이중표적 단백질 및 이의 용도 | |
| WO2021235894A1 (en) | Anti-HER2 Antibody or Antigen-binding Fragment thereof, and Chimeric Antigen Receptor Comprising Same | |
| WO2019107812A1 (ko) | 세포의 세포질에 침투하여 세포내 활성화된 ras를 억제하는 항체 및 이의 용도 | |
| WO2019225787A1 (ko) | 항-b7-h3 항체 및 그 용도 | |
| WO2019225777A1 (ko) | 항-ror1 항체 및 그 용도 | |
| WO2017204606A1 (ko) | 세포질 침투 항체 및 이의 용도 | |
| WO2019190206A1 (ko) | 클라우딘 3의 ecl-2에 특이적으로 결합하는 항체, 이의 단편 및 이들의 용도 | |
| WO2014077648A1 (ko) | 인간 및 마우스 l1cam 단백질에 특이적으로 결합하는 항체 및 이의 용도 | |
| WO2010056043A2 (en) | Cell-penetrating, sequence-specific and nucleic acid-hydrolyzing antibody, method for preparing the same and pharmaceutical composition comprising the same | |
| WO2016013871A1 (ko) | 완전한 이뮤노글로불린 형태의 세포질 침투능을 갖는 항체를 이용하여 세포내 활성화된 ras를 억제하는 방법 및 그의 이용 | |
| WO2018174544A2 (ko) | Muc1에 특이적으로 결합하는 항체 및 그의 용도 | |
| WO2022039490A1 (en) | Anti-b7-h4/anti-4-1bb bispecific antibodies and use thereof | |
| WO2021101346A1 (en) | Anti-ror1/anti-4-1bb bispecific antibodies and uses thereof | |
| WO2019132533A1 (ko) | 항-pd-l1 항체 및 이의 용도 | |
| WO2020251316A1 (ko) | α-SYN/IGF1R에 대한 이중 특이 항체 및 그 용도 | |
| WO2022149837A1 (ko) | 항-fgfr3 항체 및 이의 용도 | |
| WO2014021693A2 (ko) | Tm4sf5 단백질에 특이적으로 결합하는 신규한 단일클론항체 및 이의 용도 | |
| WO2019132579A2 (ko) | 세포질 침투 항체에 융합된 rna 분해효소를 포함하는 면역독소 | |
| WO2017171373A2 (ko) | Egfr 표적 제제에 대한 저항성을 억제하기 위한 조성물 | |
| WO2022085905A1 (ko) | SARS-CoV-2 스파이크 단백질에 특이적으로 결합하는 항체 및 이의 용도 | |
| WO2018026249A1 (ko) | 프로그램화된 세포 사멸 단백질 리간드-1 (pd-l1)에 대한 항체 및 이의 용도 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 15825418 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2955272 Country of ref document: CA |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 15327369 Country of ref document: US |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 250203 Country of ref document: IL |
|
| WWE | Wipo information: entry into national phase |
Ref document number: MX/A/2017/001010 Country of ref document: MX |
|
| ENP | Entry into the national phase |
Ref document number: 2017525495 Country of ref document: JP Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| REG | Reference to national code |
Ref country code: BR Ref legal event code: B01A Ref document number: 112017001304 Country of ref document: BR |
|
| ENP | Entry into the national phase |
Ref document number: 2017103675 Country of ref document: RU Kind code of ref document: A |
|
| REEP | Request for entry into the european phase |
Ref document number: 2015825418 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2015825418 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: 2015292955 Country of ref document: AU Date of ref document: 20150722 Kind code of ref document: A |
|
| ENP | Entry into the national phase |
Ref document number: 112017001304 Country of ref document: BR Kind code of ref document: A2 Effective date: 20170120 |