WO2017122635A1 - 固定化ウイルス粒子を含むワクチン - Google Patents
固定化ウイルス粒子を含むワクチン Download PDFInfo
- Publication number
- WO2017122635A1 WO2017122635A1 PCT/JP2017/000485 JP2017000485W WO2017122635A1 WO 2017122635 A1 WO2017122635 A1 WO 2017122635A1 JP 2017000485 W JP2017000485 W JP 2017000485W WO 2017122635 A1 WO2017122635 A1 WO 2017122635A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- virus particles
- immobilized
- strain
- particles
- vaccine
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images




Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/145—Orthomyxoviridae, e.g. influenza virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/12—Ketones
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/16—Amides, e.g. hydroxamic acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/125—Picornaviridae, e.g. calicivirus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
- A61P31/16—Antivirals for RNA viruses for influenza or rhinoviruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5252—Virus inactivated (killed)
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/60—Medicinal preparations containing antigens or antibodies characteristics by the carrier linked to the antigen
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/62—Medicinal preparations containing antigens or antibodies characterised by the link between antigen and carrier
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16134—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16161—Methods of inactivation or attenuation
- C12N2760/16163—Methods of inactivation or attenuation by chemical treatment
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16211—Influenzavirus B, i.e. influenza B virus
- C12N2760/16234—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16211—Influenzavirus B, i.e. influenza B virus
- C12N2760/16261—Methods of inactivation or attenuation
- C12N2760/16263—Methods of inactivation or attenuation by chemical treatment
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Definitions
- the present invention relates to a vaccine containing immobilized virus particles. More specifically, the present invention relates to a vaccine comprising immobilized virus particles that suppress side reactions by immobilizing the virus particle structure with an immobilizing agent.
- Vaccination and the like are known as a method for protecting and preventing infection by such viruses.
- inactivated vaccines Two types of vaccines against viruses such as influenza virus and Japanese encephalitis virus are manufactured and marketed, inactivated vaccines and live vaccines.
- inactivated vaccines are prepared by treating purified virus particles with an inactivating agent such as formalin to prepare and destroying (split) the purified virus particles with an organic solvent or surfactant. It is roughly divided into split vaccines. All-particle vaccines are highly immunogenic and are excellent in preventing infection. However, all-particle vaccines tend to have strong side reactions such as local reactions and exothermic reactions. On the other hand, split vaccines are excellent in safety because local reactions are reduced and there is almost no fever reaction.
- split vaccines tends to be low in children who have not established basic immunity or in elderly people whose immune response has declined. Therefore, there is a demand for the development of a highly safe vaccine that exhibits superior efficacy (immunogenicity) than the split vaccine.
- the present invention has been made in view of the above circumstances, and an object thereof is to provide a vaccine with high immunogenicity and reduced side reactions.
- the present inventors surprisingly did not destroy (split) the particle structure of the virus particle, but immobilized it with an immobilizing agent, thereby realizing the original virus particle.
- Vaccine containing virus particles with the same components and structure (hereinafter sometimes referred to as “immobilized virus particles”) is equivalent in immunogenicity tests and excellent in side reaction results such as fever tests.
- immobilized virus particles is equivalent in immunogenicity tests and excellent in side reaction results such as fever tests.
- the present invention provides the following [1] to [15].
- a vaccine comprising immobilized virus particles, The sum of the exothermic reaction in the three rabbits in the exothermic test of the immobilized virus particles is based on the sum of the exothermic reaction in the three rabbits in the original virus particles or the corresponding inactivated virus particles. As a vaccine, less than 80%.
- the vaccine according to [1], wherein the sum of the exothermic reactions of the three rabbits in the exothermic test of the immobilized virus particles is 1.3 ° C. or lower.
- a vaccine comprising immobilized virus particles, The amount of inflammatory cytokines produced from human peripheral blood mononuclear cells stimulated by the immobilized virus particles was stimulated by the original virus particles of the immobilized virus particles or the corresponding inactivated virus particles A vaccine that is less than 80% based on the amount of inflammatory cytokines produced from human peripheral blood mononuclear cells.
- the vaccine according to any one of [1] to [3], wherein the virus particles that are the basis of the immobilized virus particles include orthomyxovirus particles, flavivirus particles, or picornavirus particles.
- influenza virus particles include influenza virus particles.
- influenza virus particles include influenza A virus particles or influenza B virus particles.
- the influenza virus particles are H1N1 subtype strain, H2N2 subtype strain, H3N2 subtype strain, H3N8 subtype strain, H5N1 subtype strain, H5N2 subtype strain, H5N6 subtype strain, H6N1 subtype strain, H7N3
- the vaccine according to [6] or [7] comprising influenza virus particles classified into subtype strains, H7N7 subtype strains, H7N9 subtype strains, H9N2 subtype strains, or H10N8 subtype strains.
- the relative value of the specific activity of the immobilized virus particles with respect to the specific activity (antigen content / protein content) of the virus particles as the basis of the immobilized virus particles is 0% to 95%.
- the immobilized virus particles have an average particle size of 80% to 150% of the particle size of the virus particles that are the basis of the immobilized virus particles or the corresponding inactivated virus particles.
- the vaccine in any one of. [14] The vaccine according to any one of [1] to [13], wherein a peak is detected at a sucrose concentration of 35% or higher when the immobilized virus particles are measured by sucrose density gradient centrifugation. [15] The vaccine according to any one of [1] to [14], wherein a single peak is observed when the immobilized virus particles are measured by high performance liquid chromatography. [16] The vaccine according to any one of [1] to [15], which induces IgG2a specific to the immobilized viral particle rather than IgG1 specific to the immobilized viral particle when immunized to a mouse .
- the present invention provides the following [17] to [31].
- [17] A method for producing immobilized virus particles, comprising a step of adding an immobilizing agent to a suspension containing the original virus particles or corresponding inactivated virus particles.
- the immobilizing agent contains aldehydes.
- the aldehyde is selected from the group consisting of formaldehyde, paraformaldehyde, glutaraldehyde, and combinations thereof.
- the production method according to [19] wherein the aldehyde contains formaldehyde.
- the carbodiimides are selected from the group consisting of dicyclohexylcarbodiimide, diisopropylcarbodiimide, 1-ethyl-3- (3-dimethylaminopropyl) carbodiimide hydrochloride, analogs thereof, and combinations thereof [24] ] The manufacturing method of description. [26] The production method according to [25], wherein the carbodiimide includes 1-ethyl-3- (3-dimethylaminopropyl) carbodiimide hydrochloride.
- the concentration of the 1-ethyl-3- (3-dimethylaminopropyl) carbodiimide hydrochloride is 0.05 to 1500 mM based on the total amount of the suspension and the immobilizing agent.
- the manufacturing method as described in. [28] The production method according to any one of [17] to [27], wherein the original virus particles are virus particles collected by infecting cultured cells, chicken eggs or mouse brain.
- a method for producing a vaccine comprising a step of adding immobilized virus particles obtained by the production method according to any one of [17] to [30].
- the vaccine according to the present embodiment is a vaccine containing immobilized virus particles, and the sum (° C.) of the exothermic reaction in three rabbits in the fever test of the immobilized virus particles is the origin of the immobilized virus particles. And the sum of the exothermic reactions of the three rabbits in the corresponding virus particles or the corresponding inactivated virus particles.
- the amount of inflammatory cytokines produced from human peripheral blood mononuclear cells stimulated by the immobilized virus particles is the virus particles from which the immobilized virus particles are based or the corresponding inactivated virus particles. The amount of inflammatory cytokines produced from human peripheral blood mononuclear cells stimulated by is reduced.
- the immunogenicity of the vaccine containing the immobilized virus particles is more than the immunogenicity of the split vaccine, and side reactions such as local reactions and fever reactions are suppressed to the same extent as the split vaccine.
- the immobilized viral particles are superior in stability compared to whole particle vaccines comprising conventional inactivated viral particles.
- Immobilized virus particle means a virus particle that has no ability to infect a host and whose particle structure is immobilized by cross-linking proteins on the surface of the virus particle.
- the immobilized virus particles are highly immunogenic because they maintain the same particle properties as the original virus particles or the corresponding inactivated virus particles.
- Immobilized virus particles can be obtained by treating the original virus particles or the corresponding inactivated virus particles with an immobilizing agent.
- the “fixing agent” means a drug that crosslinks the proteins of the virus particles by covalent bonds.
- the immobilizing agent is a drug that maintains the particle structure of virus particles by cross-linking surface antigens, surface antigens and matrix proteins or membrane proteins, or matrix proteins or membrane proteins.
- Inactivated virus particle means a virus particle that has no ability to infect the host and whose particle structure is not immobilized. Inactivated virus particles are obtained by treating the original virus particles with an inactivating agent. In the case of influenza virus particles, inactivated virus particles, for example, have a final concentration of 0.02 v / v% (0.0072 to 0.0076 w / v% in terms of formaldehyde) in a suspension containing influenza virus particles. Formalin (36-38 w / v% formaldehyde aqueous solution) may be added and reacted at 4 ° C. for 6 weeks.
- inactivated virus particles are, for example, commercially available Vero cell culture Japanese encephalitis vaccine drug substance (manufactured by Chemical and Serum Therapy Research Institute, trade name “Ensebak”, 0.08 v / v% formalin) (Including inactivated Japanese encephalitis virus particles).
- fever test is performed by a method according to the fever test method shown in the biological preparation standards (Japan, Ministry of Health, Labor and Welfare Notification No. 192).
- “Fever reaction” refers to the body temperature of a rabbit measured after injection of the sample into the ear vein (sometimes referred to as “measurement value”) and the body temperature of the rabbit measured before injection (“control body temperature”) Is the maximum value of the difference (sometimes referred to as “different body temperature”).
- measurement value the body temperature of the rabbit measured before injection
- control body temperature Is the maximum value of the difference
- the differential body temperature is a negative value
- the exothermic reaction is interpreted as zero.
- the heat generation test is performed according to the following procedure.
- the immobilized virus particles are diluted with physiological saline, and the protein content in 1 mL is set to 70 ⁇ g (in the case of Japanese encephalitis virus particles) or 240 ⁇ g (in the case of influenza virus particles) as a sample.
- a rabbit is inoculated with 1-3 mL of the above sample per kg of body weight, and an increase in fever is observed until 6 hours later.
- the difference between the body temperature of the rabbit before inoculating the sample (control body temperature) and the body temperature of the rabbit after inoculation is obtained, and the maximum value of the difference is defined as the exothermic reaction of the rabbit.
- the same test is performed on three rabbits, and the sum (° C.) of the exothermic reaction of the three rabbits is determined.
- the sum of the exothermic reaction in the three rabbits in the exothermic test of the immobilized virus particles is the sum of the exothermic reaction in the three rabbits in the virus particles that are the origin of the immobilized virus particles or the corresponding inactivated virus particles.
- it may be less than 80%, less than 60%, less than 40%, less than 20%, or less than 10%.
- the lower limit is not particularly limited, but may be 0% or more based on the sum of the exothermic reaction in the three rabbits in the virus particles as the basis of the immobilized virus particles or the corresponding inactivated virus particles, or 20% It may be the above.
- the sum of the exothermic reaction in three rabbits in the exothermic test of immobilized virus particles may be 1.3 ° C. or less, 0.9 ° C. or less, 0.5 It may be below °C.
- 0 degreeC or more may be sufficient and 0.6 degreeC or more may be sufficient.
- “Inflammatory cytokine” is a general term for cytokines produced with inflammation, and examples thereof include IL-1 ⁇ , IL-6, TNF- ⁇ , IFN- ⁇ , and IFN- ⁇ .
- Human peripheral blood mononuclear cells mean lymphocytes (including T cells, B cells, NK cells, etc.) and monocytes obtained from human peripheral blood.
- the amount of inflammatory cytokine is determined by quantifying the amount of inflammatory cytokine produced when human peripheral blood mononuclear cells (PBMC) are stimulated with virus particles by a method according to the European Pharmacopoeia MONOCYTE-ACTIVATION TEST. .
- the method may be a method in which the measurement conditions shown in Examples described later are changed in the European Pharmacopoeia MONOCYTE-ACTIVATION TEST.
- the amount of inflammatory cytokines produced from human peripheral blood mononuclear cells stimulated by the immobilized virus particles is stimulated by the virus particles that are the origin of the immobilized virus particles or the corresponding inactivated virus particles. Based on the amount of inflammatory cytokines produced from human peripheral blood mononuclear cells received, it may be less than 80%, less than 60%, less than 40%, 20% Or less than 10%.
- the lower limit is not particularly limited, but it is 0 on the basis of the amount of inflammatory cytokines produced from human peripheral blood mononuclear cells stimulated by the original virus particle of the immobilized virus particle or the corresponding inactivated virus particle. % Or more, or 40% or more.
- the concentration of the inflammatory cytokine produced from the human peripheral blood mononuclear cells stimulated by the immobilized virus particles is based on the culture medium containing the human peripheral blood mononuclear cells. Or 30 pg / ml or less, or 20 pg / ml or less.
- the lower limit is not particularly limited, but may be 0 pg / ml or more, or 5 pg / ml or more.
- the concentration of the inflammatory cytokine produced from human peripheral blood mononuclear cells stimulated by the immobilized virus particles is based on the culture medium containing the human peripheral blood mononuclear cells. Or 50 pg / ml or less, or 40 pg / ml or less.
- the lower limit is not particularly limited, but may be 0 pg / ml or more, or 5 pg / ml or more.
- orthomyxovirus particles include influenza virus particles.
- flavivirus particles include Japanese encephalitis virus particles.
- picornavirus particles include hepatitis A virus particles.
- influenza virus particles include influenza A virus particles and influenza B virus particles.
- influenza A virus particles for example, H1N1 subtype strain, H2N2 subtype strain, H3N2 subtype strain, H3N8 subtype strain, H5N1 subtype strain, H5N2 subtype strain, H5N6 subtype strain, H6N1 subtype strain Influenza particles classified into strains, H7N3 subtype strains, H7N7 subtype strains, H7N9 subtype strains, H9N2 subtype strains, or H10N8 subtype strains.
- Japanese encephalitis virus particles examples include Beijing-1 strain, Nakayama strain (Nakayama-Yoken strain), or SA14-14-2 strain and P3 strain of Japanese encephalitis virus particles.
- the immobilized virus particles contain genomic nucleic acids (DNA, RNA, etc.) derived from virus particles because the particle structure is not destroyed as in split vaccines.
- Genomic nucleic acid from viral particles can act as an adjuvant.
- inactivated poliovirus vaccines include D antigens that contain viral genomic RNA and C antigens that do not contain viral genomic RNA.
- C antigen is weakly immunogenic and has no effect as a vaccine antigen.
- the only molecular species that has an effect as a vaccine antigen is the D antigen. This suggests that the viral genomic RNA encapsulated in the vaccine is important for the expression of the effect. Therefore, the vaccine according to this embodiment can induce a Th1-type response.
- an IgG2a subclass antibody induced by a Th1-type response in mice has a better ability to protect against influenza virus than an IgG1 subclass antibody induced by a Th2-type response. From this, further improvement in the effectiveness of the vaccine can be expected. That is, the above vaccine may induce IgG2a specific to immobilized virus particles rather than IgG1 specific to immobilized virus particles when immunizing mice.
- Immobilized virus particles have substantially the same parameters as the original virus particles or corresponding inactivated virus particles (e.g., as measured by sucrose density gradient centrifugation, high performance liquid chromatography, and / or dynamic light scattering) , Molecular weight, average particle size, density, hemagglutinin (HA) content, etc.).
- the immobilized virus particle may be an immobilized virus particle having an average particle size of 80% to 150% of the particle size of the original virus particle or the corresponding inactivated virus particle, and 90% to 140% It may be an immobilized virus particle having an average particle size.
- the immobilized virus particles may be immobilized virus particles having an average particle size of 90% to 130% of the particle size of the original virus particles or the corresponding inactivated virus particles. It may be an immobilized virus particle having an average particle size of from% to 120%.
- the average particle diameter of the immobilized virus particles may be around 150 nm when measured by a dynamic light scattering method, or may be 120 nm to 180 nm. It may be 130 nm to 170 nm.
- the average particle diameter of the immobilized virus particles may be 100 nm or more when measured by a dynamic light scattering method, or 120 nm or more It may be 130 nm or more, 150 nm or more, or 170 nm or more.
- the average particle diameter may be 180 nm or less, 175 nm or less, or 170 nm or less.
- the average particle diameter of the immobilized virus particles may be around 90 nm when measured by a dynamic light scattering method, and is 80 nm to 110 nm. Also good.
- the average particle size of the immobilized virus particles may be 70 nm or more when measured by a dynamic light scattering method, and 80 nm The above may be sufficient and 90 nm or more may be sufficient.
- the average particle diameter may be 110 nm or less, or 100 nm or less.
- the content of the lipid component in the immobilized virus particle may be equivalent to the content of the lipid component in the virus particle.
- the immobilized virus particle may be an immobilized virus particle whose peak is detected at a sucrose concentration of 35% or more when measured by sucrose density gradient centrifugation, and the peak is at a sucrose concentration of 45% or more and 55% or less. May be an immobilized virus particle from which is detected.
- the immobilized virus particle may be an immobilized virus particle in which a unimodal peak is observed when measured by high performance liquid chromatography (size exclusion chromatography (SEC)).
- SEC size exclusion chromatography
- a unimodal peak when measured by high performance liquid chromatography (size exclusion chromatography (SEC) is performed using size exclusion chromatography (trade name: TSKgel G6000PWXL (manufactured by TOSOH) or Superose 6 10/300 GE (manufactured by GE Healthcare)) (eluent: PBS, flow rate: 0.00). 5 ml / min), a unimodal peak with an elution time of 14 to 15 minutes may be observed, and it may be an immobilized virus particle of Japanese encephalitis virus particles. This may be an immobilized virus particle of an influenza virus particle.
- 0% to 90% of the protein on the surface of the immobilized virus particle may not be immobilized, and 5% to 80% of the protein on the surface of the immobilized virus particle is immobilized. It does not have to be.
- proteins on the surface of the immobilized virus particle include surface antigens, matrix proteins, membrane proteins, and combinations thereof.
- examples of the surface antigen include hemagglutinin (HA) and neuraminidase (NA).
- HA hemagglutinin
- NA neuraminidase
- M1 protein is mentioned, for example.
- M2 protein is mentioned, for example.
- examples of the surface antigen include E protein and M protein.
- Examples of a method for calculating the ratio of proteins that are not immobilized include a method for obtaining a change rate (residual rate) of the amount of protein before and after immobilization in the original molecular weight of the target protein. It is done. For example, when the immobilized virus particles are based on influenza virus particles, after performing SDS-PAGE (polyacrylamide gel electrophoresis) under reducing conditions, the concentration of the band of the M1 protein, which is one of the constituent proteins of the virus, is increased. A method of calculating the residual ratio (%) of the M1 protein by obtaining the thickness by densitometry is mentioned. In this case, the residual ratio of the M1 protein tends to decrease as the concentration of the immobilizing agent increases. By applying this method, it is possible to analyze the degree of crosslinking with a fixing agent.
- a change rate residual rate
- the M1 protein residual rate when the immobilized virus particles are based on influenza virus particles, the M1 protein residual rate may be 5% to 90%, or the M1 protein residual rate may be 10%. It may be an immobilized virus particle that is 80% to 80%. In other words, 5% to 90% of the M1 protein may not be immobilized, and 10% to 80% may not be immobilized.
- the crosslinking degree analysis can also be performed by determining the specific activity (antigen content / protein content) of the immobilized virus particles.
- the monoclonal antibody used for measuring the antigen content of the virus particle recognizes the neutralizing epitope, and the specific activity decreases when the structural change of the neutralizing epitope occurs. Utilizing this property, the relative value (%) of the specific activity of the immobilized virus particles relative to the virus particles not immobilized is calculated, and the degree of crosslinking is evaluated. In the present embodiment, the relative value of specific activity is in the range of 0 to 95%.
- the relative value of the specific activity of the immobilized Japanese encephalitis virus particles may be 90 to 95%, 70 to 90%, 50 to It may be 70%.
- the amount of immobilized virus particles contained in the vaccine may be appropriately selected according to the type of virus or the administration target.
- the amount (concentration) of immobilized virus particles contained in the influenza vaccine may be 1 to 40 ⁇ g HA / mL per virus strain as the hemagglutinin concentration.
- the vaccine may be a simple vaccine containing antigens derived from the same type of virus or bacterium.
- the vaccine may be a mixed vaccine comprising antigens from multiple types of viruses or bacteria.
- the vaccine may be a multivalent vaccine that is a virus or bacterium of the same genus and contains antigens from multiple types.
- the vaccine when the vaccine is an influenza virus vaccine, it may contain either immobilized influenza A virus particles or immobilized influenza B virus particles, or both of them.
- the vaccine is a Japanese encephalitis virus vaccine, it may contain either cell culture-derived or mouse brain-derived virus.
- the influenza virus vaccine or Japanese encephalitis virus vaccine may contain an antigen derived from another virus or bacteria. For example, it may be mixed with diphtheria / pertussis / tetanus / inactivated poliovirus mixed vaccine (DPT-IPV vaccine).
- DPT-IPV vaccine diphtheria / pertussis / tetanus / inactiv
- the dosage form of the vaccine may be, for example, liquid, powder (freeze-dried powder, dry powder, etc.), capsule, tablet, or frozen state.
- the vaccine may contain a pharmaceutically acceptable carrier.
- a carrier usually used for vaccine production can be used without limitation.
- the carrier includes saline, buffered saline, dextrose, water, glycerol, isotonic aqueous buffer, and combinations thereof.
- an emulsifier, a preservative (for example, thimerosal), an isotonic agent, a pH adjusting agent, an inactivating agent (for example, formalin) and the like may be further appropriately blended.
- the route of administration of the vaccine is, for example, transdermal administration, sublingual administration, ophthalmic administration, intradermal administration, intramuscular administration, oral administration, enteral administration, nasal administration, intravenous administration, subcutaneous administration, intraperitoneal administration, mouth To the lung may be used.
- the method for administering the vaccine may be, for example, a syringe, a transdermal patch, a microneedle, an implantable sustained-release device, a syringe with a microneedle, a needleless device, or a method of spraying.
- the vaccine may contain an adjuvant.
- an adjuvant for example, an oil-in-water emulsion adjuvant (AS03, MF59, etc.) containing aluminum adjuvant or squalene, CpG and 3-to-deacylated-4′-monophosphoryl lipid A (MPL) etc.
- Examples of the target mammal include mouse, rat, guinea pig, hamster, rabbit, cat, dog, sheep, pig, cow, horse, goat, monkey, human and the like.
- the vaccine which concerns on this embodiment may be used with respect to a human, may be used with respect to a child under 5 years old, and an elderly person 65 years or older.
- Vaccine varies depending on the purpose of administration, administration method, and administration target condition (sex, age, weight, medical condition, etc.), but when flu vaccine is administered to humans, 3.8 ⁇ g HA to 30 ⁇ g HA per administration
- the active ingredient immobilized virus particles may be used to be administered.
- the method for producing immobilized virus particles according to the present embodiment includes a step of immobilizing the particle structure on the original virus particles or the corresponding inactivated virus particles.
- a method of destroying (spliting) enveloped virus particles using a surfactant or an organic solvent has been conventionally known.
- these methods tend to reduce the effectiveness (immunogenicity) of the vaccine as the particles break down.
- the original virus particle or the corresponding inactivated virus particle is treated with an immobilizing agent that causes a covalent bond with the virus particle protein, so that the virus particle structure is maintained. It is possible to increase safety while maintaining effectiveness.
- the production method includes a step of culturing a host, a step of infecting a virus with a host, a step of growing a virus in the host, a step of recovering virus particles from the host, or a step of inactivating the recovered virus particles. Further, it may be included.
- the virus particles may be virus particles collected from the host after the virus particles are infected and propagated in the host. You may select a host suitably according to the kind of virus particle.
- a known method may be used to inactivate the virus particles, and examples thereof include a method of inactivating with an inactivating agent such as formalin.
- influenza virus particles examples of the host include cultured cells, chicken eggs and mouse brain.
- the cultured cells may be primary cells or established cells. Examples of cultured cells include Vero cells and MDCK cells.
- the step of immobilizing is exemplified by a method of treating the original virus particles or the corresponding inactivated virus particles with an immobilizing agent, for example, the original virus particles or the corresponding inactivated virus particles.
- a step of adding a fixing agent to the suspension containing You may change suitably the density
- the concentration of virus particles in the suspension may be 60 to 90 ⁇ g / mL, 300 to 3000 ⁇ g / mL, or 500 to 2500 ⁇ g / mL as the protein concentration of virus particles. It may be.
- the type of immobilizing agent can be changed as appropriate according to the type of virus.
- the fixing agent includes organic solvents, aldehydes, diimide esters, bis (3,5-dibromosalicyl) fumarate (DBBF), carbodiimides, and combinations thereof.
- the organic solvent include methanol, ethanol, acetone, and combinations thereof.
- aldehydes include formaldehyde (FA) (eg, formalin), paraformaldehyde, glutaraldehyde (GA), and combinations thereof.
- FA formaldehyde
- GA glutaraldehyde
- carbodiimides examples include dicyclohexylcarbodiimide (DCC), diisopropylcarbodiimide (DIC), 1-ethyl-3- (3-dimethylaminopropyl) carbodiimide hydrochloride (EDC), analogs thereof, and combinations thereof.
- DCC dicyclohexylcarbodiimide
- DIC diisopropylcarbodiimide
- EDC 1-ethyl-3- (3-dimethylaminopropyl) carbodiimide hydrochloride
- the concentration of the immobilizing agent may be appropriately changed according to the type of virus and the type of immobilizing agent.
- the concentration of formaldehyde may be 0.005 to 0.5 w / v% based on the total amount of the suspension containing virus particles and the immobilizing agent.
- the formaldehyde concentration is less than 0.005 w / v%, immobilization is weakened and the particle structure tends to be difficult to be maintained.
- the formaldehyde concentration exceeds 0.5 w / v%, immobilization is strong and chemical modification by crosslinking tends to proceed excessively.
- the concentration of formaldehyde may be 0.01 to 0.5 w / v% based on the total amount of the suspension and the fixing agent, and 0.018 to 0.005. It may be 152 w / v%, 0.029 to 0.152 w / v%, or 0.029 to 0.076 w / v%.
- the method using a fixing agent containing formaldehyde may be used when the virus particles are influenza virus particles or Japanese encephalitis virus particles.
- the immobilizing agent is formalin (36-38 w / v% aqueous formaldehyde solution)
- the formalin concentration is 0.014-0.4 v / v% based on the total amount of the suspension and the immobilizing agent. Or 0.05 to 0.4 v / v%, 0.08 to 0.4 v / v%, or 0.08 to 0.2 v / v%. .
- the concentration of glutaraldehyde may be 0.001 to 0.06 w / v% based on the total amount of the suspension and the immobilizing agent. It may be ⁇ 0.05 w / v%, 0.004 to 0.02 w / v%, or 0.005 to 0.01 w / v%.
- the concentration is less than 0.001 w / v%, when Japanese encephalitis virus particles are used as virus particles, the particles tend to aggregate.
- the concentration exceeds 0.06 w / v% when Japanese encephalitis virus particles are used as virus particles, the epitope of E protein, which is the main structural protein, tends to be inactivated.
- the method using glutaraldehyde as the immobilizing agent may be used when the virus particles are influenza virus particles or Japanese encephalitis virus particles.
- the concentration of EDC may be 0.05 to 1500 mM or 0.15 to 500 mM based on the total amount of the suspension and the immobilizing agent. It may be 5-50 mM.
- the method using an immobilizing agent containing EDC may be used when the virus particles are influenza virus particles or Japanese encephalitis virus particles.
- the temperature at the time of treatment with the immobilizing agent may be appropriately changed according to the type of virus, the type of immobilizing agent, the concentration of the immobilizing agent and the like.
- the temperature may be 0 ° C. (ice bath) to 37 ° C., 4 ° C. to 37 ° C., or 25 ° C. to 37 ° C.
- the period (treatment time) for treatment with the immobilizing agent may be appropriately changed according to the type of virus, the type of immobilizing agent, the concentration of the immobilizing agent, the temperature of treatment, and the like.
- the period may be 1 day to 4 weeks, 3 days to 4 weeks, or 1 week to 4 weeks.
- EDC is used as the immobilizing agent, the period may be 5 minutes to 24 hours, 0.5 hours to 24 hours, or 2 hours to 20 hours.
- a quenching treatment may be performed using an amino acid such as glycine.
- the quenching treatment may be performed for the purpose of improving vaccine stability, immunogenicity, and safety.
- a step of purifying the recovered immobilized virus particles may be further included.
- a method for purifying the immobilized virus particles can be appropriately performed by a known method, and examples thereof include a method of filtering using an ultrafiltration membrane.
- the vaccine production method includes a step of adding the immobilized virus particles obtained by the above-described method for producing immobilized virus particles.
- the method for producing a vaccine may further include a step of adding a pharmaceutically acceptable carrier, an emulsifier, a preservative, an isotonic agent, a pH adjusting agent, an inactivating agent and the like.
- the vaccine may contain all viral proteins as antigens for protection against infection.
- the best vaccine depends on the actual virus, given that the presence of virus-derived genomic nucleic acids to increase the immunogenicity of the viral protein, the size and shape of the virus particles, and so on, are each acting on the immune response.
- the present inventors consider that they have close components and structures.
- the immobilized virus particles according to the present embodiment are only immobilized with an immobilizing agent, and the other components and structures are the same as those of the original virus particles, so that the immunogenicity is high and side reactions are suppressed. It is possible to provide a new vaccine.
- Example 1 Preparation of antigen derived from influenza virus particles (1) Preparation of FA-immobilized influenza virus particles Formaldehyde (FA) treatment Influenza A virus H1N1 subtype strain (A / California / 07/2009 (X-179A) strain, (It may be described as “A / CA strain”) was inoculated into the chorioallantoic cavity of 11-day-old embryonated chicken eggs and cultured at 34 ° C. for 2 days. After clarifying the obtained chorioallantoic fluid, influenza virus particles were precipitated by ultracentrifugation. The influenza virus particles were resuspended in phosphate buffered saline (PBS) to obtain a suspension.
- PBS phosphate buffered saline
- the obtained fraction was diluted so that the final protein concentration of the purified influenza virus particles was 500 ⁇ g / mL to obtain a suspension.
- the formalin 36-38 w / v% formaldehyde aqueous solution
- the solution was added to the solution and reacted at 25 ° C. for 1 week.
- the reaction solution was dialyzed with PBS, and formaldehyde was removed to obtain immobilized influenza virus particles (hereinafter sometimes referred to as “FA-immobilized influenza virus particles”).
- split influenza antigen A split influenza virus antigen (hereinafter sometimes referred to as “split influenza antigen”) as a comparative control is an influenza HA vaccine (trade name “manufactured by Chemical and Serum Therapy Research Institute”. The stock solution of each strain contained in influenza HA vaccine “Kakekenken” ” was used.
- Fever test A fever test was performed in accordance with the biologics standard (Japan, Ministry of Health, Labor and Welfare Notification No. 192). Inactivated influenza virus particles, FA-immobilized influenza virus particles, and split influenza antigen were diluted with physiological saline, and the protein content in 1 mL was adjusted to 240 ⁇ g. A rabbit was inoculated with 1 to 3 mL of the above sample per kg body weight, and an increase in fever was observed until 6 hours later. The difference between the body temperature of the rabbit before inoculating the sample (control body temperature) and the body temperature of the rabbit after inoculation was determined, and the maximum value of the difference was defined as the exothermic reaction of the rabbit. A similar test was performed on three rabbits. Table 1 shows the sum of exothermic reactions (° C.) of three rabbits.
- Table 2 shows the results of cytokine production of FA-immobilized influenza virus particles and inactivated influenza virus particles. It was found that the production amount of IL-6 in FA-immobilized influenza virus particles was sufficiently low compared with inactivated influenza virus particles. From this, it was suggested that the FA-immobilized influenza virus particles have higher safety than the inactivated influenza virus particles.
- Example 2 Physical property evaluation Analysis by Sucrose Density Gradient Centrifugation The obtained fraction was diluted and suspended by the method according to Example 1 above so that the final protein concentration of influenza virus particles (A / CA strain) was 2500 ⁇ g / mL. A turbid liquid was obtained. Thereafter, formalin was added to the suspension so that the final concentration was 0.12 v / v%, and the mixture was reacted at 25 ° C. for 1 week. The reaction solution was dialyzed against PBS to obtain FA-immobilized influenza virus particles. The resulting FA-immobilized influenza virus particles were analyzed by sucrose density gradient centrifugation.
- the specimen was layered on a sucrose density gradient of 15 to 60%, and centrifuged at 18000 rpm (57500 ( ⁇ g)) at 4 ° C. for 16 hours. After centrifugation, 0.6 mL was fractionated per fraction, and the sucrose concentration, HA value and protein concentration of each fraction were measured.
- Table 3 shows the results of the A / CA strain (H1N1 subtype strain).
- the protein was widely distributed at a sucrose concentration of 25 to 50%, indicating that the virus particles were degraded.
- the FA-immobilized influenza virus particles were fractionated as a single peak (particulate) at a high sucrose concentration (44.3%).
- the HA activity was 10240 times.
- Fig. 1 shows a photograph of FA-immobilized influenza virus particles taken with an electron microscope.
- the FA-immobilized influenza virus particles had the same particle structure as the inactivated influenza virus particles.
- Dynamic light scattering Influenza A virus H3N2 subtype strain (A / New York / 39/2012 (X-233A) strain, hereinafter sometimes referred to as “A / NY strain”) and influenza B virus Victoria FA-immobilized influenza virus particles based on strains (B / Brisbane / 60/2008 strain, hereinafter sometimes referred to as “B / BR strain”) were prepared by a method according to Example 1.
- the average particle size of each was analyzed using Zetasizer Nano ZS (Malvern).
- Table 4 shows the average particle diameter in the liquid by the dynamic light scattering method.
- the FA-immobilized influenza virus particles were unimodal with an average particle size of around 140 to 150 nm. From this, it was found that the average particle size of the FA-immobilized influenza virus particles was equivalent to the inactivated influenza virus particles. In addition, the particle structure of the FA-immobilized influenza virus particles was maintained, and impurities such as aggregates were not observed.
- Molecular weight distribution measurement A method according to Example 1 above for Yamagata strain of influenza B virus (B / Massachetts / 02/022012 (BX-51B) strain (hereinafter sometimes referred to as “B / MA strain”)) The FA-immobilized influenza virus particles (B / MA strain) were prepared. For A / CA strain (H1N1 subtype strain), A / NY strain (H3N2 subtype strain), and B / MA strain (B type strain), molecular weight distribution measurement of split influenza antigen and FA-immobilized influenza virus particles was performed.
- the molecular weight was measured using size exclusion chromatography (trade name: TSKgel G6000PWXL (manufactured by TOSOH)) (PBS was used as the eluent, and the flow rate was 0.5 ml / min).
- the elution pattern is shown in Table 5.
- the FA-immobilized influenza virus particles had a unimodal peak around an elution time of 16 to 17 minutes.
- the split influenza antigen derived from the same strain had a trimodal peak around an elution time of 19 to 30 minutes.
- the FA-immobilized influenza virus particles (A / CA strain) prepared in Example 1 were analyzed for the degree of cross-linking with an immobilizing agent by the following procedure. First, SDS-Buffer (final concentration: Tris 0.76 w / v%, SDS 1 w / v%, Glycerol 10 v / v%, Bromophenol Blue (BPB) 0.01 w / v%) and 2-mercaptoethanol (final) (Concentration: 0.8 v / v%), boiled for 6 minutes, and then used PAGEL NPU-12.5L (trade name, manufactured by ATTO) or PAGEL NPU-R12.5L (trade name, manufactured by ATTO).
- SDS-Buffer final concentration: Tris 0.76 w / v%, SDS 1 w / v%, Glycerol 10 v / v%, Bromophenol Blue (BPB) 0.01 w / v%) and 2-mercaptoethanol (final) (Concentration
- M1 protein residual ratio (%) was calculated.
- Table 6 M1 protein residual ratios of FA-immobilized influenza virus particles (A / CA strain) immobilized at formalin concentrations of 0.05%, 0.08%, 0.11%, and 0.14% were 35.7% and 23, respectively. 8%, 11.0%, and 5.4%, suggesting that as the formalin concentration increases, cross-linking is promoted and the M1 protein residual rate decreases.
- Immunogenicity 1 Intramuscular inoculation of mice
- Immunogenicity in FA-immobilized influenza virus particles was evaluated using mice.
- ddY mice female, 8 weeks old
- three weeks after immunization all mice were bled and euthanized. Serum was obtained by centrifugation, and the HI antibody titer was measured according to the “Pathogen Detection Manual” (National Institute of Infectious Diseases).
- Table 7 shows the results of immunogenicity (HI antibody titer (GMT)) when mice were inoculated intramuscularly.
- FA-immobilized influenza virus particles A / CA strain
- Immunogenicity 2 (intradermal mouse inoculation) The immunogenicity in FA-immobilized influenza virus particles (A / CA strain) was evaluated by intradermal mouse inoculation in the same procedure as in “6. Immunogenicity 1 (intramuscular inoculation of mouse)”.
- ddY mice female, 8 weeks old were inoculated intracutaneously with split influenza antigen, inactivated influenza virus particles, or FA-immobilized influenza virus particles at a dose of 0.2 ⁇ g of protein (5 mice per group). .
- Table 8 shows the results of immunogenicity (HI antibody titer (GMT)) when mice were inoculated intradermally.
- the FA-immobilized influenza virus particles were significantly more immunogenic than the split influenza antigen and were as immunogenic as the inactivated influenza virus particles.
- Immunogenicity 3 (cynomolgus monkey subcutaneous inoculation) For A / CA strain (H1N1 subtype strain), A / NY strain (H3N2 subtype strain), B / BR strain and B / MA strain (B strain), split influenza antigen, inactivated influenza virus particles, and FA The immunogenicity of immobilized influenza virus particles was evaluated using cynomolgus monkeys (male or female, 17-25 months of age). Split influenza antigen, inactivated influenza virus particles, or FA-immobilized influenza virus particles at an inoculation amount of 7.5 ⁇ g HA equivalent (protein amount corresponding to the HA amount of split influenza antigen) per 8 or 9 mice per group, Inoculated subcutaneously twice daily.
- 7.5 ⁇ g HA equivalent protein amount corresponding to the HA amount of split influenza antigen
- Partial blood collection was performed before the second immunization and on the 21st day after the second immunization. Serum was obtained by centrifugation, and the HI antibody titer and neutralizing antibody titer were measured according to the “Pathogen Detection Manual” (National Institute of Infectious Diseases). The immunogenicity results are shown in Table 9 (HI antibody titer (GMT) 21 days after secondary immunization) and Table 10 (Neutralizing antibody titer (GMT) 21 days after secondary immunization). In the HI antibody titer, it was found that the FA-immobilized influenza virus particles have a higher immunogenicity than the split influenza antigen.
- the immunogenicity was significantly higher than that of the split influenza antigen.
- the FA-immobilized influenza virus particles were significantly more immunogenic in all of the A-type and B-type strains compared to the split influenza antigen.
- Antibody subclass analysis As antibody subclass analysis, A / CA strain (H1N1 subtype strain) and B / MA strain (B strain) obtained in “6. Immunogenicity 1 (intramuscular inoculation of mice)” above, IgG1 and IgG2a antibody titers specific to viral antigens contained in mouse serum were measured by enzyme immunoassay (ELISA). As a result, it was shown that FA-immobilized influenza virus particles induce antigen-specific IgG2a rather than antigen-specific IgG1 (Tables 11 and 12). This is the same result as inactivated influenza virus particles, and a stronger tendency to induce IgG2a was also observed in comparison with split influenza antigen. From this result, the FA-immobilized influenza virus particles can be expected to further improve the effectiveness of the vaccine, compared to the split influenza antigen that activates humoral immunity but hardly activates cellular immunity.
- RNA release amount For the FA-immobilized influenza virus particles (A / NY strain) prepared by the method according to Example 1, the RNA release amount over time during protease treatment was evaluated by the following procedure. First, FA-immobilized influenza virus particles were diluted with PBS, SDS and Proteinase K were added and reacted at 55 ° C., and RNA was extracted over time. For RNA extraction, TRIzol LS Reagent, PureLink RNA Mini Kit, and PureLink DNase (trade name, manufactured by Invitrogen) were used. The content of the extracted RNA was measured with Quant-iT RiboGreen RNA Reagent and Kit (trade name, manufactured by Invitrogen). The RNA content over time for each FA concentration is shown in Table 13. As a result, it was shown that RNA release was controlled gradually depending on the FA concentration. It was suggested that the sustained release of RNA release by FA fixation slows the production of inflammatory cytokines and produces high safety.
- Example 3 (1) Preparation of GA-immobilized influenza virus particles Glutaraldehyde (GA) treatment Influenza A virus (H3N2 subtype strain (A / NY strain)) and influenza B virus (type B Victoria strain (B / BR strain)) were cultured in the same manner as in Example 1. And purified. Each influenza virus particle was diluted so that the final protein concentration of each purified influenza virus particle was 1000 ⁇ g / mL to obtain a suspension. Next, using a 1 w / v% GA solution, the GA concentration was diluted to 0.016 w / v% or 0.008 w / v%.
- Example 2 An exothermic test was conducted in the same manner as in Example 1. Inactivated influenza virus particles and GA-immobilized influenza virus particles (A / NY strain and B / BR strain) were diluted with physiological saline, and the protein content in 1 mL was adjusted to 240 ⁇ g, which was used as a sample. A rabbit was inoculated with 1 mL of the above sample per 1 kg of body weight, and an increase in fever was observed until 6 hours later. Table 14 shows the sum of exothermic reactions (° C.) of the three rabbits of the A / NY strain, and Table 15 shows the sum of the exothermic reactions (° C.) of the three rabbits of the B / BR strain.
- Example 4 Evaluation of physical properties The physical properties of the GA-immobilized influenza virus particles obtained in Example 3 were evaluated by the following methods.
- the average particle diameter of GA-immobilized influenza virus particles was analyzed using Zetasizer Nano ZS (Malvern). Table 18 shows the average particle diameter in the liquid by the dynamic light scattering method.
- the GA-immobilized influenza virus particles were unimodal with an average particle size of around 130 to 160 nm. From this, it was confirmed that the average particle size of GA-immobilized influenza virus particles is equivalent to the original virus particles. That is, it was found that the average particle size of GA-immobilized influenza virus particles was single and did not change. Further, the particle structure of the GA-immobilized influenza virus particles was maintained, and impurities such as aggregates were not observed.
- the molecular weight distribution of GA-immobilized influenza virus particles was measured by the same method (SEC) as in Example 2. The elution pattern is shown in Table 19. The split influenza antigen had a four-peaked peak around an elution time of 16 to 30 minutes. On the other hand, the GA-immobilized influenza virus particles had a unimodal peak around the elution time of 16 to 17 minutes.
- Antibody subclass analysis As an antibody subclass analysis, IgG1 and IgG2a antibody titers specific to influenza virus antigens contained in the serum of the mouse obtained in “5. Immunogenicity (intramuscular inoculation of mouse)” were determined by ELISA. Measured by. As a result, in contrast to split influenza antigen, GA-immobilized influenza virus particles were shown to induce antigen-specific IgG2a rather than antigen-specific IgG1 (Table 22). This is the same result as inactivated influenza virus particles. From this result, GA-immobilized influenza virus particles can be expected to further improve the effectiveness of the vaccine, compared to split influenza antigen that activates humoral immunity but hardly activates cellular immunity.
- Example 5 (1) Preparation of EDC-immobilized influenza virus particles 1.1-ethyl-3- [3-dimethylaminopropyl] carbodiimide hydrochloride (EDC) treatment influenza A virus (H3N2 subtype strain (A / NY strain)) and Influenza B virus (Victoria strain B / BR strain) was cultured and purified in the same manner as in Example 1. The obtained fraction was diluted so that the final protein concentration of each purified influenza virus particle was 2500 ⁇ g / mL for the A / NY strain and 500 ⁇ g / mL for the B / BR strain to obtain a suspension.
- EDC EDC-immobilized influenza virus particles 1.1-ethyl-3- [3-dimethylaminopropyl] carbodiimide hydrochloride (EDC) treatment influenza A virus (H3N2 subtype strain (A / NY strain)) and Influenza B virus (Victoria strain B / BR strain) was cultured and
- the EDC solution is serially diluted with PBS to 0.1 to 4M, added to the suspension so that the final concentration is 50 to 500 mM, and allowed to react for 2 to 20 hours with ice cooling (0 ° C.). It was. After completion of the reaction, the reaction solution was dialyzed with PBS and EDC was removed to obtain immobilized influenza virus particles (hereinafter sometimes referred to as “EDC-immobilized influenza virus particles”). Fever test to evaluate the pyrogenic activity of the obtained EDC-immobilized influenza virus particles (A / NY strain, B / BR strain) based on the sum of pyrogenic reactions in 3 rabbits, and inflammatory cytokines when stimulated with human PBMC It was evaluated by quantifying the production amount of
- Example 2 An exothermic test was conducted in the same manner as in Example 1. A sample was prepared by diluting EDC-immobilized influenza virus particles with physiological saline and setting the protein content in 1 mL to 240 ⁇ g. The EDC-immobilized influenza virus particles were prepared by reacting the reaction solution with PBS at 50 mM or 500 mM EDC concentration for 2 hours, and then dialyzing the reaction solution with PBS (for A / NY strain, 5 mM EDC concentration was also used). Carried out.). For the A / NY strain, a sample obtained by reacting at 5 mM EDC concentration at 4 ° C. for 20 hours and then dialysis treatment of the reaction solution with PBS was also used. A rabbit was inoculated with 1 mL of the above sample per 1 kg of body weight, and an increase in fever was observed until 6 hours later. Tables 23 and 24 show the sum of exothermic reactions (° C.) of three rabbits.
- the EDC-immobilized influenza virus particles did not show a sum of exothermic reactions at 1.3 ° C. or higher under all EDC treatment conditions. This also suggests that the EDC-immobilized influenza virus particles have high safety like the split influenza antigen.
- Example 6 Evaluation of physical properties The physical properties of the EDC-immobilized influenza virus particles obtained in Example 5 were evaluated by the following methods.
- EDC-immobilized influenza virus particles were analyzed by sucrose density gradient centrifugation.
- Table 27 shows the results of EDC-immobilized influenza virus particles (A / NY strain (H3N2 subtype strain)) after 2 hours of reaction with ice cooling at an EDC concentration of 50 mM.
- the protein was widely distributed at a sucrose concentration of 25 to 50%, indicating that the virus particles were degraded.
- EDC-immobilized influenza virus particles were fractionated as a single peak (particulate) at a high sucrose concentration (47.2%). The HA activity was 10240 times.
- EDC-immobilized influenza virus particles were analyzed using Zetasizer Nano ZS (Malvern). Table 28 shows the average particle diameter in the liquid by the dynamic light scattering method.
- EDC-immobilized influenza virus particles A / NY strain (H3N2 subtype strain), B / BR strain (type B strain)
- B / BR strain type B strain
- the average particle size of the EDC-immobilized influenza virus particles is equivalent to the original virus particles. That is, it was found that the average particle size of the immobilized influenza virus particles was single and did not change even when the EDC treatment was performed. Further, the particle structure of the EDC-immobilized influenza virus particles was maintained, and impurities such as aggregates were not observed.
- Molecular weight distribution measurement The molecular weight distribution of EDC-immobilized influenza virus particles based on the H3N2 subtype strain (A / NY strain) was measured. The measurement was performed using size exclusion chromatography (trade name: TSKgel G6000PWXL (manufactured by TOSOH)) (PBS was used as the eluent, and the flow rate was 0.5 ml / min). The elution pattern is shown in Table 29. The split influenza antigen had a four-peaked peak around an elution time of 16 to 30 minutes. On the other hand, EDC-immobilized influenza virus particles showed a unimodal peak at an elution time of 16 to 17 minutes.
- Example 5 Analysis by degree of crosslinking
- Table 30 shows the results of the A / NY strain (H3N2 subtype strain).
- Table 31 shows the results of the B / BR strain (B-type Victoria strain).
- the M1 protein residual ratio (%) of EDC-immobilized influenza virus particles fixed at EDC concentrations of 5 mM, 50 mM, and 500 mM was 85.1%, 56.1%, and 27.2%, respectively.
- a / NY strain H3N2 subtype strain
- Antibody subclass analysis As antibody subclass analysis, IgG1 and IgG2a antibody titers specific to viral antigens contained in the serum of mice obtained in “6. Immunogenicity (intramuscular inoculation of mice)” are measured by ELISA. did. As a result, it was shown that EDC-immobilized influenza virus particles induce antigen-specific IgG2a rather than antigen-specific IgG1 in contrast to split influenza antigen (Table 35, Table 36). From this result, EDC-immobilized influenza virus particles can be expected to further improve the effectiveness of the vaccine, compared to split influenza antigen that activates humoral immunity but hardly activates cellular immunity.
- Immunogenicity (cynomolgus monkey subcutaneous inoculation) The immunogenicity in EDC-immobilized influenza virus particles was evaluated by the following procedure using cynomolgus monkeys. First, cynomolgus monkeys (male or female, 29-35 months of age) were inoculated subcutaneously with split influenza antigen or EDC-immobilized influenza virus particles at an inoculum of 15 ⁇ g as HA content (8 per group). Subcutaneous inoculation was performed twice at intervals of 3 weeks, and blood was collected 4 weeks after the second immunization. Serum was obtained by the same method as in Example 2, and the HI antibody titer and neutralizing antibody titer were measured.
- the immunogenic HI antibody titer (GMT) results of EDC-immobilized influenza virus particles after reacting at 5 mM EDC concentration at 4 ° C. for 20 hours are shown in Table 37, and neutralizing antibody titer (GMT) Table 38 shows the results.
- the residual rate of M1 protein after the severe test of EDC-immobilized influenza virus particles fixed at an EDC concentration of 50 and 500 mM decreased by 18% in comparison with before the severe test, and the rate of change was smaller than that of inactivated influenza virus particles. , Suggested to be more stable.
- Stability test analysis by one-way radioimmunodiffusion test
- the stability of the EDC-immobilized influenza virus particles prepared according to Example 5 was evaluated using the change in HA content before and after the severe test as an index.
- the HA content was calculated by a single radioimmunodiffusion test (Biological Formulation Standard (Japan, Ministry of Health, Labor and Welfare Notification No. 192)), and the HA content before the severe test was 37%.
- the change rate of the HA content after a one-week severe test was calculated.
- Table 40 shows the results of A / CA strain (H1N1 subtype strain), A / NY strain (H3N2 subtype strain), B / MA strain (B type Yamagata strain), and B / BR strain (B type Victoria strain). Shown in It was suggested that the change rate of HA content after the severe test of EDC-immobilized influenza virus particles was small and stable.
- Table 41 shows the results of A / CA strain (H1N1 subtype strain), A / NY strain (H3N2 subtype strain), B / MA strain (B type Yamagata strain), and B / BR strain (B type Victoria strain). Shown in It was suggested that the change rate of the M1 protein residual rate of EDC-immobilized influenza virus particles under storage at 5 ° C. for 11 months was small and stable.
- Influenza virus particles were more immunogenic than split influenza antigen.
- Serum was obtained by the same method as in Example 2, and the HI antibody titer and neutralizing antibody titer were measured.
- the results of HI antibody titer (GMT) of EDC-immobilized influenza virus particles after reaction at 4 ° C. for 20 hours at 5 mM EDC concentration at 5 ° C. for 10 months are shown in Table 46.
- the antibody titer (GMT) results are shown in Table 47 (Table 46 and Table 47 are the reprints of Table 37 and Table 38, respectively).
- Influenza virus particles were significantly more immunogenic than split influenza antigen.
- RNA release amount The amount of RNA release over time during protease treatment of the EDC-immobilized influenza virus particles prepared according to Example 5 was evaluated. First, inactivated influenza virus particles and EDC-immobilized influenza virus particles were diluted with PBS, SDS and Proteinase K were added and reacted at 55 ° C., and RNA was extracted over time. For RNA extraction, TRIzol LS Reagent, PureLink RNA Mini Kit, and PureLink DNase (trade name, manufactured by Invitrogen) were used. The content of the extracted RNA was measured with Quant-iT RiboGreen RNA Reagent and Kit (trade name, manufactured by Invitrogen).
- the production amount of inflammatory cytokines over time of EDC-immobilized influenza virus particles prepared according to Example 5 was evaluated.
- the method was based on the European Pharmacopoeia MONOCYTE-ACTIVATION TEST. Specifically, the measurement was carried out by changing the use of human PBMC in the European Pharmacopoeia MONOCYTE-ACTIVATION TEST by pooling at least 4 donors to 1 donor.
- Table 49 shows changes over time in IL-1 ⁇ production of inactivated influenza virus particles and EDC-immobilized influenza virus particles in the A / NY strain (H3N2 subtype strain). Table 49 shows changes in IL-6 production over time. 50.
- Table 51 shows changes in IL-1 ⁇ production over time of inactivated influenza virus particles and EDC-immobilized influenza virus particles in the B / BR strain (type B Victoria strain). The changes are shown in Table 52.
- Table 52 shows changes in IL-1 ⁇ production over time of inactivated influenza virus particles and EDC-immobilized influenza virus particles in the B / BR strain (type B Victoria strain). The changes are shown in Table 52.
- the production of inflammatory cytokines by the EDC-immobilized influenza virus particles was gradually released as compared with the inactivated influenza virus particles. That is, it was suggested that the EDC-fixed influenza virus particles have high safety by slowing the release of RNA in the body by EDC fixation and delaying the production of inflammatory cytokines.
- Example 7 Preparation of GA-immobilized Japanese encephalitis virus particles Glutaraldehyde treatment (step of fixing the particle structure to the virus particles) Vero cell culture Japanese encephalitis vaccine drug substance (Chemical and Serological Therapy Research Institute, trade name “Ensebac”) containing Japanese encephalitis virus particles inactivated with 0.08 v / v% formalin in a protein concentration of 60 to 90 ⁇ g / ml.
- glutaraldehyde is added so that the final concentration is 0.005 to 0.02 w / v%, and may be described as “inactivated Japanese encephalitis virus particles”. Reacted.
- the resulting reaction solution was dialyzed against a PBS-like solution (PBS to which lactose as an activator (final concentration 5 w / v%) was added).
- PBS PBS-like solution
- lactose as an activator final concentration 5 w / v% was added
- Glutaraldehyde was removed by dialysis to obtain immobilized Japanese encephalitis virus particles (hereinafter sometimes referred to as “GA immobilized Japanese encephalitis virus particles”).
- the pyrogenic activity of the obtained GA-immobilized Japanese encephalitis virus particles was evaluated by a fever test in which the sum of the exothermic reaction in three rabbits was evaluated, and the amount of inflammatory cytokine produced when human PBMC was stimulated.
- Example 2 An exothermic test was conducted in the same manner as in Example 1. A GA-immobilized Japanese encephalitis virus particle or inactivated Japanese encephalitis virus particle was diluted with physiological saline, and the protein content in 1 mL was set to 70 ⁇ g. The rabbit was inoculated with 3 mL of the above sample per kg of body weight, and an increase in fever was observed until 6 hours later. Table 53 shows the sum of exothermic reactions (° C.) of three rabbits. As a representative, GA-immobilized Japanese encephalitis virus particles after a reaction for 3 days at 4 ° C. with a glutaraldehyde concentration of 0.01 w / v% were evaluated.
- the GA-immobilized Japanese encephalitis virus particles did not show a sum of the fever reaction above 1.3 ° C, and a decrease in the sum of the fever reactions above 1.6 ° C was observed compared to the inactivated Japanese encephalitis virus particles. . This also suggests that GA-immobilized Japanese encephalitis virus particles have high safety.
- Example 8 Evaluation of physical properties The physical properties of the GA-immobilized Japanese encephalitis virus particles obtained in Example 7 were evaluated by the following methods.
- FIG. 4 shows a photograph of GA-immobilized Japanese encephalitis virus particles after reaction at 0.01% w / v% glutaraldehyde concentration at 4 ° C. for 3 days.
- the GA-immobilized Japanese encephalitis virus particles had the same particle structure as the inactivated Japanese encephalitis virus particles by fixing with glutaraldehyde.
- the molecular weight distribution of GA-immobilized Japanese encephalitis virus particles was measured. The measurement was performed using size exclusion chromatography (trade name: Superose 6 10/300 GE (manufactured by GE Healthcare)) (PBS was used as the eluent, and the flow rate was 0.5 ml / min). ). The elution pattern is shown in Table 56. In the GA-immobilized Japanese encephalitis virus particles, a unimodal main peak was observed around an elution time of 14 to 15 minutes. Inactivated Japanese encephalitis virus particles also had a unimodal main peak around an elution time of 14 to 15 minutes.
- Antigen Content was measured by the following procedure by sandwich ELISA using an anti-Japanese encephalitis virus antibody.
- the E antigen contained in the specimen is captured on a microplate to which an anti-Japanese encephalitis virus rabbit IgG (primary antibody; polyclonal antibody) is bound. Thereafter, by reacting anti-Japanese encephalitis virus E protein monoclonal antibody (secondary antibody; monoclonal antibody) conjugated with horseradish peroxidase (HRP), anti-E antigen antibody / E antigen / secondary antibody bound to the plate A complex is formed.
- HRP horseradish peroxidase
- E antigen content (antigen content) was measured using the fact that the intensity of OPD color development was proportional to the amount of complex (reflecting the amount of E antigen).
- Table 57 shows the antigen contents of GA-immobilized Japanese encephalitis virus particles and inactivated Japanese encephalitis virus particles. As a representative, when the GA-immobilized Japanese encephalitis virus particles were reacted at a glutaraldehyde concentration of 0.01 w / v% at 4 ° C. for 3 days, an antigen equivalent to the inactivated Japanese encephalitis virus particles was contained.
- the GA-immobilized Japanese encephalitis virus particles prepared in Example 7 were evaluated for the degree of cross-linking with the immobilizing agent based on the specific activity (antigen content / protein content). Specifically, the monoclonal antibody (503) used for measuring the antigen content recognizes the neutralizing epitope, and the specific activity decreases when the structural change of the neutralizing epitope occurs. Relative value (%) of the specific activity of GA-immobilized Japanese encephalitis virus particles treated with each glutaraldehyde concentration relative to the specific activity of inactivated Japanese encephalitis virus particles not immobilized (hereinafter referred to as “503 antibody”). The reaction rate "(may be referred to as”% ”) was calculated.
- the results are shown in Table 58.
- the 503 antibody reaction rates of GA-immobilized Japanese encephalitis virus particles immobilized at glutaraldehyde concentrations of 0.005 w / v%, 0.01 w / v%, and 0.02 w / v% are 95.1% and 74.7%, respectively. 55.2%, and as the glutaraldehyde concentration increased, crosslinking was promoted, suggesting a decrease in reaction rate due to structural change of the 503 antibody epitope.
- Serum was obtained by centrifugation, and an equal volume was pooled for each group, and the neutralizing antibody titer was measured according to the “Pathogen Detection Manual” (edited by National Institute of Infectious Diseases). Table 59 shows the result calculated from the 50% plaque reduction rate. GA-immobilized Japanese encephalitis virus particles had a neutralizing antibody titer equivalent to or higher than inactivated Japanese encephalitis virus particles.
- GA-immobilized Japanese encephalitis virus particles were maintained for 1 month under 25 ° C storage.
- inactivated Japanese encephalitis virus particles showed an increase under 25 ° C storage.
- GA-immobilized Japanese encephalitis virus particles have improved stability compared to inactivated Japanese encephalitis virus particles.
- Example 9 Preparation of FA-immobilized Japanese encephalitis virus particles Formalin treatment (step of immobilizing particle structure to virus particles) With respect to inactivated Japanese encephalitis virus particles, formalin (36-38 w / v) so that the final concentration is 0.014-0.04 v / v% (formaldehyde conversion: 0.005-0.015 w / v%). % Formaldehyde aqueous solution) was added and reacted at 25 ° C. for 1 week.
- the reaction solution was dialyzed with a PBS-like solution, and formalin was removed to obtain immobilized Japanese encephalitis virus particles (hereinafter sometimes referred to as “FA-immobilized Japanese encephalitis virus particles”).
- the pyrogenic activity of the obtained FA-immobilized Japanese encephalitis virus particles was evaluated by quantifying the amount of inflammatory cytokine produced when human PBMC was stimulated.
- the FA-immobilized Japanese encephalitis virus particles were found to produce a sufficiently lower amount of inflammatory cytokines than the inactivated Japanese encephalitis virus particles. This also suggests that FA-immobilized Japanese encephalitis virus particles have higher safety than inactivated Japanese encephalitis virus particles.
- Example 10 Evaluation of physical properties The physical properties of the FA-immobilized Japanese encephalitis virus particles obtained in Example 9 were evaluated by the following methods.
- Antigen Content was measured in the same manner as in Example 8 by sandwich ELISA using an anti-Japanese encephalitis virus antibody.
- Table 66 shows the antigen contents of FA-immobilized Japanese encephalitis virus particles and inactivated Japanese encephalitis virus particles.
- evaluation of FA-immobilized Japanese encephalitis virus particles after reaction for 1 week at 25 ° C. at formalin concentrations of 0.014 v / v% and 0.02 v / v% revealed an antigen equivalent to inactivated Japanese encephalitis virus particles.
- Stability test (antigen content) FA-immobilized Japanese encephalitis virus particles and inactivated Japanese encephalitis virus particles were diluted with a PBS-like solution so that the final protein concentration was 8 ⁇ g / mL. The storage stability at 37 ° C. was evaluated using the antigen content as an index. As a representative, Table 68 shows the results after reaction at 25 ° C. for 1 week at a formalin concentration of 0.014 v / v%. FA-immobilized Japanese encephalitis virus particles were maintained for 1 week under storage at 37 ° C. On the other hand, inactivated Japanese encephalitis virus particles showed a decrease under storage at 37 ° C. FA-immobilized Japanese encephalitis virus particles were shown to have improved stability compared to inactivated Japanese encephalitis virus particles.
- Example 11 Preparation of EDC-immobilized Japanese encephalitis virus particles 1.1-ethyl-3- (3-dimethylaminopropyl) carbodiimide hydrochloride (EDC) treatment The final concentration of inactivated Japanese encephalitis virus particles was 0.15 to 15 mM. Then, EDC was added and reacted at 4 ° C. for 2 to 20 hours. When performing the quenching treatment, glycine was further added to the reaction solution in an amount 8 times (molar mass ratio) of EDC as a quenching agent.
- EDC EDC-immobilized Japanese encephalitis virus particles 1.1-ethyl-3- (3-dimethylaminopropyl) carbodiimide hydrochloride
- EDC-immobilized Japanese encephalitis virus particles immobilized Japanese encephalitis virus particles (hereinafter sometimes referred to as “EDC-immobilized Japanese encephalitis virus particles”). It was. The pyrogenic activity of the obtained EDC-immobilized Japanese encephalitis virus particles was evaluated by quantifying the amount of inflammatory cytokine produced when human PBMC was stimulated.
- EDC-immobilized Japanese encephalitis virus particles have a sufficiently lower inflammatory cytokine production than inactivated Japanese encephalitis virus particles. This also suggests that EDC-immobilized Japanese encephalitis virus particles have higher safety than inactivated Japanese encephalitis virus particles.
- Example 12 Evaluation of physical properties The physical properties of the EDC-immobilized Japanese encephalitis virus particles obtained in Example 11 were evaluated by the following methods.
- Example 2 Analysis by electron microscope
- FIG. 1 a photograph of EDC-immobilized Japanese encephalitis virus particles after reaction at 4 ° C. for 20 hours at an EDC concentration of 1.5 mM is shown in FIG.
- the particle structure of the EDC-immobilized Japanese encephalitis virus particles was maintained by fixing with EDC in the same manner as the inactivated Japanese encephalitis virus particles.
- EDC-immobilized Japanese encephalitis virus particles were analyzed using Zetasizer Nano ZS (Malvern). Table 70 shows the average particle diameter in the liquid by the dynamic light scattering method. EDC-immobilized Japanese encephalitis virus particles were unimodal with an average particle size of about 90 nm. On the other hand, inactivated Japanese encephalitis virus particles were unimodal at about 80 nm. In addition, the particle structure of EDC-immobilized Japanese encephalitis virus particles was maintained, and no impurities such as aggregates were observed.
- EDC-immobilized Japanese encephalitis virus particles The molecular weight distribution of EDC-immobilized Japanese encephalitis virus particles was measured in the same manner as in Example 8. The elution pattern is shown in Table 71. In the EDC-immobilized Japanese encephalitis virus particles, a unimodal main peak was observed around an elution time of 14 to 15 minutes. Inactivated Japanese encephalitis virus particles also had a unimodal main peak around an elution time of 14 to 15 minutes.
- Antigen Content was measured in the same manner as in Example 8 by sandwich ELISA using an anti-Japanese encephalitis virus antibody.
- Table 72 shows the antigen contents of EDC-immobilized Japanese encephalitis virus particles and inactivated Japanese encephalitis virus particles after reaction for 20 hours at 4 ° C. at an EDC concentration of 1.5 mM.
- EDC-immobilized Japanese encephalitis virus particles contained the same antigen as inactivated Japanese encephalitis virus particles.
- EDC-immobilized Japanese encephalitis virus particles and inactivated Japanese encephalitis virus particles were diluted with a PBS-like solution so that the final protein concentration was 8 ⁇ g / mL.
- the storage stability at 25 ° C. and 37 ° C. was evaluated using the antigen content as an index. The results are shown in Table 74 and Table 75.
- EDC-immobilized Japanese encephalitis virus particles were maintained for 1 month under 25 ° C storage and for 1 week under 37 ° C storage.
- the inactivated Japanese encephalitis virus particles showed a decrease in antigen content under both storage at 25 ° C. and storage at 37 ° C. It was shown that EDC-immobilized Japanese encephalitis virus particles have improved stability compared to inactivated Japanese encephalitis virus particles.
- the present invention is useful in the field of pharmaceuticals, particularly in the field of vaccines.
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Virology (AREA)
- General Health & Medical Sciences (AREA)
- Veterinary Medicine (AREA)
- Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Epidemiology (AREA)
- Mycology (AREA)
- Microbiology (AREA)
- Immunology (AREA)
- Pulmonology (AREA)
- Oncology (AREA)
- Communicable Diseases (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Organic Chemistry (AREA)
- Molecular Biology (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description
[1]固定化ウイルス粒子を含むワクチンであって、
上記固定化ウイルス粒子の発熱試験におけるウサギ3羽での発熱反応の和が、上記固定化ウイルス粒子の元となるウイルス粒子又は対応する不活化ウイルス粒子におけるウサギ3羽での発熱反応の和を基準として、80%未満である、ワクチン。
[2]上記固定化ウイルス粒子の発熱試験におけるウサギ3羽での発熱反応の和が、1.3℃以下である、[1]に記載のワクチン。
[3]固定化ウイルス粒子を含むワクチンであって、
上記固定化ウイルス粒子によって刺激を受けたヒト末梢血単核球から産生される炎症性サイトカインの量が、上記固定化ウイルス粒子の元となるウイルス粒子又は対応する不活化ウイルス粒子によって刺激を受けたヒト末梢血単核球から産生される炎症性サイトカインの量を基準として、80%未満である、ワクチン。
[4]上記固定化ウイルス粒子の元となるウイルス粒子が、オルソミクソウイルス粒子、フラビウイルス粒子、又はピコルナウイルス粒子を含む、[1]~[3]のいずれかに記載のワクチン。
[5]上記ウイルス粒子が、インフルエンザウイルス粒子、日本脳炎ウイルス粒子、又はA型肝炎ウイルス粒子を含む、[4]に記載のワクチン。
[6]上記ウイルス粒子が、インフルエンザウイルス粒子を含む、[5]に記載のワクチン。
[7]上記インフルエンザウイルス粒子が、A型インフルエンザウイルス粒子、又はB型インフルエンザウイルス粒子を含む、[6]に記載のワクチン。
[8]上記インフルエンザウイルス粒子が、H1N1亜型株、H2N2亜型株、H3N2亜型株、H3N8亜型株、H5N1亜型株、H5N2亜型株、H5N6亜型株、H6N1亜型株、H7N3亜型株、H7N7亜型株、H7N9亜型株、H9N2亜型株、又はH10N8亜型株に分類されるインフルエンザウイルス粒子を含む、[6]又は[7]に記載のワクチン。
[9]上記ウイルス粒子が、日本脳炎ウイルス粒子を含む、[5]に記載のワクチン。
[10]上記日本脳炎ウイルス粒子が、北京-1株、中山株、SA14-14-2株、又はP3株を含む、[9]に記載のワクチン。
[11]上記固定化ウイルス粒子の表面のタンパク質の0%~90%が、固定化されていない、[1]~[10]のいずれかに記載のワクチン。
[12]上記固定化ウイルス粒子の元となるウイルス粒子の比活性(抗原含量/タンパク質含量)に対する、上記固定化ウイルス粒子の比活性の相対値が、0%~95%である、[1]~[11]のいずれかに記載のワクチン。
[13]上記固定化ウイルス粒子が、上記固定化ウイルス粒子の元となるウイルス粒子又は対応する不活化ウイルス粒子の粒子径の80%~150%の平均粒子径を有する、[1]~[12]のいずれかに記載のワクチン。
[14]上記固定化ウイルス粒子をショ糖密度勾配遠心で測定した場合に、ショ糖濃度35%以上にピークが検出される、[1]~[13]のいずれかに記載のワクチン。
[15]上記固定化ウイルス粒子を高速液体クロマトグラフィーで測定した場合に、単峰性のピークが認められる、[1]~[14]のいずれかに記載のワクチン。
[16]マウスに免疫した場合、上記固定化ウイルス粒子に特異的なIgG1よりも、上記固定化ウイルス粒子に特異的なIgG2aを誘導する、[1]~[15]のいずれかに記載のワクチン。
[17]元となるウイルス粒子又は対応する不活化ウイルス粒子を含む懸濁液に固定化剤を加える工程を含む、固定化ウイルス粒子の製造方法。
[18]上記固定化剤が、アルデヒド類を含む、[17]に記載の製造方法。
[19]上記アルデヒド類が、ホルムアルデヒド、パラホルムアルデヒド、グルタルアルデヒド、及びそれらの組合せからなる群から選択される、[18]に記載の製造方法。
[20]上記アルデヒド類が、ホルムアルデヒドを含む、[19]に記載の製造方法。
[21]上記ホルムアルデヒドの濃度が、上記懸濁液及び上記固定化剤の全量を基準として、0.005~0.5w/v%である、[20]に記載の製造方法。
[22]上記アルデヒド類が、グルタルアルデヒドを含む、[19]に記載の製造方法。
[23]上記グルタルアルデヒドの濃度が、上記懸濁液及び上記固定化剤の全量を基準として、0.001~0.06w/v%である、[22]に記載の製造方法。
[24]上記固定化剤が、カルボジイミド類を含む、[17]に記載の製造方法。
[25]上記カルボジイミド類が、ジシクロヘキシルカルボジイミド、ジイソプロピルカルボジイミド、1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド塩酸塩、それらの類縁体及び、それらの組合せからなる群から選択される、[24]に記載の製造方法。
[26]上記カルボジイミド類が、1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド塩酸塩を含む、[25]に記載の製造方法。
[27]上記1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド塩酸塩の濃度が、上記懸濁液及び上記固定化剤の全量を基準として、0.05~1500mMである、[26]に記載の製造方法。
[28]元となる上記ウイルス粒子が、培養細胞、鶏卵又はマウス脳に感染させて、回収されたウイルス粒子である、[17]~[27]のいずれかに記載の製造方法。
[29]上記培養細胞が、初代細胞、又は株化細胞を含む、[28]に記載の製造方法。
[30]上記培養細胞が、Vero細胞、又はMDCK細胞を含む、[29]に記載の製造方法。
[31][17]~[30]のいずれかに記載の製造方法によって得られた固定化ウイルス粒子を加える工程を含む、ワクチンの製造方法。
本実施形態に係るワクチンは、固定化ウイルス粒子を含むワクチンであって、上記固定化ウイルス粒子の発熱試験におけるウサギ3羽での発熱反応の和(℃)が、上記固定化ウイルス粒子の元となるウイルス粒子又は対応する不活化ウイルス粒子におけるウサギ3羽での発熱反応の和を基準として、低下している。また、上記ワクチンは、固定化ウイルス粒子によって刺激を受けたヒト末梢血単核球から産生される炎症性サイトカインの量が、上記固定化ウイルス粒子の元となるウイルス粒子又は対応する不活化ウイルス粒子によって刺激を受けたヒト末梢血単核球から産生される炎症性サイトカインの量を基準として、低減している。すなわち、固定化ウイルス粒子を含むワクチンの免疫原性は、スプリットワクチンの免疫原性以上であり、局所反応及び発熱反応等の副反応がスプリットワクチンと同等に抑えられている。他の側面において、固定化ウイルス粒子は、従来の不活化ウイルス粒子を含む全粒子ワクチンと比較して安定性に優れている。
本実施形態に係る固定化ウイルス粒子の製造方法は、元となるウイルス粒子又は対応する不活化ウイルス粒子に対して粒子構造を固定化する工程を含む。
上記固定化する工程は、元となるウイルス粒子又は対応する不活化ウイルス粒子を固定化剤で処理する方法が例示され、例えば、元となるウイルス粒子又は対応する不活化ウイルス粒子を含む懸濁液に固定化剤を加える工程が挙げられる。懸濁液中のウイルス粒子の濃度は、ウイルスの種類、固定化剤の種類及びその濃度等に応じて適宜変更してもよい。例えば、懸濁液中のウイルス粒子の濃度は、ウイルス粒子のタンパク質の濃度として、60~90μg/mLであってもよいし、300~3000μg/mLであってもよいし、500~2500μg/mLであってもよい。
1.インフルエンザウイルス粒子に由来する抗原の調製
(1)FA固定化インフルエンザウイルス粒子の調製
ホルムアルデヒド(FA)処理
A型インフルエンザウイルスH1N1亜型株(A/California/07/2009(X-179A)株、以下、「A/CA株」と記載することがある。)を11日齢の発育鶏卵の漿尿膜腔内に接種し、34℃で2日間培養した。得られた漿尿液を清澄化した後、超遠心分離によってインフルエンザウイルス粒子を沈殿させた。上記インフルエンザウイルス粒子をリン酸緩衝生理食塩水(PBS)で再浮遊させて懸濁液を得た。得られた懸濁液をショ糖密度勾配遠心(RCF=57500(×g)、16時間)で遠心分離し、ショ糖濃度が33%~50%である画分を回収することによって、インフルエンザ粒子を精製した。精製したインフルエンザウイルス粒子のタンパク質の最終濃度が500μg/mLとなるように、得られた画分を希釈し、懸濁液を得た。その後、最終濃度が0.05~0.20v/v%(ホルムアルデヒド換算で、0.018~0.076w/v%)となるようにホルマリン(36~38w/v%ホルムアルデヒド水溶液)を上記懸濁液に添加し、25℃で1週間反応させた。反応終了後、PBSで反応液を透析し、ホルムアルデヒドを除くことで、固定化インフルエンザウイルス粒子(以下、「FA固定化インフルエンザウイルス粒子」と記載することがある。)を得た。
上記懸濁液に、最終濃度が0.02v/v%(ホルムアルデヒド換算で、0.0072~0.0076w/v%)となるようにホルマリン(36~38w/v%ホルムアルデヒド水溶液)を添加し、4℃で6週間~8週間反応させた。反応終了後、PBSで反応液を透析し、ホルムアルデヒドを除くことで不活化インフルエンザウイルス粒子を得た。同様の方法で、他のインフルエンザウイルス株(亜型株)についても不活化インフルエンザウイルス粒子を調製し、実施例1~6において比較対照として使用した。
比較対照とするスプリットインフルエンザウイルス抗原(以下、「スプリットインフル抗原」と記載することがある。)は、インフルエンザHAワクチン(化学及血清療法研究所製、商品名「インフルエンザHAワクチン“化血研”」)に含まれる各株の原液を使用した。
生物学的製剤基準(日本国、厚生労働省告示 第192号)に準じて、発熱試験を行った。不活化インフルエンザウイルス粒子、FA固定化インフルエンザウイルス粒子及びスプリットインフル抗原を生理食塩水で希釈し、1mL中のタンパク質含量を240μgとしたものを試料とした。上記試料を、体重1kg当たりにつき1~3mLをウサギに接種して、6時間後まで発熱の上昇を観察した。試料を接種する前のウサギの体温(対照体温)と接種した後のウサギの体温との差を求め、その差の最大値をそのウサギの発熱反応とした。同様の試験を3羽のウサギに対して行った。ウサギ3羽の発熱反応の和(℃)を表1に示す。

欧州薬局方MONOCYTE-ACTIVATION TESTに準じた方法で、ホルマリン濃度0.05v/v%、0.08v/v%、0.11v/v%で固定したFA固定化インフルエンザウイルス粒子又は不活化インフルエンザウイルス粒子でヒト末梢血単核球(PBMC)を刺激した場合の炎症性サイトカイン(IL-6)の産生量を定量した。具体的には、欧州薬局方MONOCYTE-ACTIVATION TESTにおけるヒトPBMCを最低4ドナー分プールして使用するところを、1ドナー分に変更して測定した。FA固定化インフルエンザウイルス粒子及び不活化インフルエンザウイルス粒子のサイトカインの産生量の結果を表2に示す。FA固定化インフルエンザウイルス粒子におけるIL-6の産生量は、不活化インフルエンザウイルス粒子と比較して十分低いことがわかった。このことから、FA固定化インフルエンザウイルス粒子は、不活化インフルエンザウイルス粒子と比較して、高い安全性を有することが示唆された。

物性評価
1.ショ糖密度勾配遠心法による分析
上記実施例1に準じた方法で、インフルエンザウイルス粒子(A/CA株)のタンパク質の最終濃度が2500μg/mLとなるように、得られた画分を希釈し懸濁液を得た。その後、最終濃度が0.12v/v%となるようにホルマリンを上記懸濁液に添加し、25℃で1週間反応させた。反応液をPBSで透析することによって、FA固定化インフルエンザウイルス粒子を得た。得られたFA固定化インフルエンザウイルス粒子をショ糖密度勾配遠心法によって分析した。検体を15~60%のショ糖密度勾配に重層し、4℃、18000rpm(57500(×g))で16時間、遠心分離を行った。遠心後、1フラクションあたり0.6mLずつ分画し、各フラクションのショ糖濃度、HA価及びタンパク質濃度を測定した。A/CA株(H1N1亜型株)の結果を表3に示す。スプリットインフル抗原では、タンパク質がショ糖濃度25~50%に幅広く分布し、ウイルス粒子が分解していることが示された。これに対し、FA固定化インフルエンザウイルス粒子では高いショ糖濃度(44.3%)に単一ピーク(粒子性)として分画されていることが示された。HA活性については10240倍であった。

上記FA固定化インフルエンザウイルス粒子(A/CA株)の形状をさらに詳細に調べるために、電子顕微鏡による観察を実施した。検体を、グルタルアルデヒドを用いて室温で20分間固定した。その後、観察用のイオンコートしたシートメッシュ(日新EM社製)に、固定した検体を載せて約60秒間静置し、2%リンタングステン酸水溶液でネガティブ染色した。染色した検体を、透過型電子顕微鏡(FEIカンパニー社製テクナイG2:加速電圧120kV)を用いて観察、撮影した。
A型インフルエンザウイルスH3N2亜型株(A/New York/39/2012(X-233A)株、以下、「A/NY株」と記載することがある。)及びB型インフルエンザウイルスビクトリア系統株(B/Brisbane/60/2008株株、以下、「B/BR株」と記載することがある。)を元とするFA固定化インフルエンザウイルス粒子を実施例1に準じた方法で作製し、それぞれの平均粒子径をZetasizer Nano ZS(Malvern社製)を用いて解析した。動的光散乱法による液体中の平均粒子径を表4に示す。FA固定化インフルエンザウイルス粒子は平均粒子径が140~150nm前後で単峰性であった。このことから、FA固定化インフルエンザウイルス粒子の平均粒子径は、不活化インフルエンザウイルス粒子と同等であることがわかった。また、FA固定化インフルエンザウイルス粒子の粒子構造は保たれており、凝集体のような不純物は認められなかった。

B型インフルエンザウイルスの山形系統株(B/Massachusetts/02/2012(BX-51B)株(以下、「B/MA株」と記載することがある。))について、上記実施例1に準じた方法でFA固定化インフルエンザウイルス粒子(B/MA株)を作製した。A/CA株(H1N1亜型株)、A/NY株(H3N2亜型株)、及びB/MA株(B型株)について、スプリットインフル抗原、及びFA固定化インフルエンザウイルス粒子の分子量分布測定を、サイズ排除クロマトグラフィー(商品名:TSKgel G6000PWXL(TOSOH社製))を用いて分子量測定を行った(溶離液にはPBSを用いて、流速は0.5ml/minで行った。)。その溶出パターンを表5に示す。FA固定化インフルエンザウイルス粒子は溶出時間16~17分付近に単峰性のピークが認められた。一方、同じ株由来のスプリットインフル抗原は溶出時間19~30分付近に三峰性のピークが認められた。

実施例1で調製したFA固定化インフルエンザウイルス粒子(A/CA株)について、以下の手順で、固定化剤での架橋度を分析した。まず、検体にSDS-Buffer(終濃度:Tris 0.76w/v%、SDS 1w/v%、Glycerol 10v/v%、Bromophenol Blue(BPB) 0.01w/v%)と2-メルカプトエタノール(終濃度:0.8v/v%)を添加し、6分間煮沸後、PAGEL NPU-12.5L(ATTO社製、商品名)又はPAGEL NPU-R12.5L(ATTO社製、商品名)を用いて、SDS-PAGEを行った。泳動後、CBB(クマシーブリリアントブルー)染色を行い、LAS3000(富士フイルム株式会社製、商品名)にて画像を取り込んだ。固定化剤による架橋が進むと、ウイルスを構成するタンパク質の一つであるM1タンパク質は、本来のバンドの位置(25~37kDa)から高分子側へシフトする。そのため、本来の位置に検出されるM1タンパク質のバンド(M1バンド)は薄くなることが示唆されており、これを架橋度の指標とした。具体的には、固定化を行っていないインフルエンザウイルス粒子のM1バンドのデンシトメトリー値に対する、各ホルマリン濃度で処理したFA固定化インフルエンザウイルス粒子の、本来の位置に検出されるM1バンドのデンシトメトリー値の相対値(%)(以下、この相対値を「M1タンパク質残存率」(%)と記載することがある。)を算出した。結果を表6に示す。ホルマリン濃度0.05%、0.08%、0.11%、0.14%で固定したFA固定化インフルエンザウイルス粒子(A/CA株)のM1タンパク質残存率は、それぞれ35.7%、23.8%、11.0%、5.4%であり、ホルマリン濃度が高まるにつれて架橋が促進され、M1タンパク質残存率が低下することが示唆された。

FA固定化インフルエンザウイルス粒子(A/CA株)における免疫原性を、マウスを用いて評価した。ddYマウス(雌性、8週齢)に、スプリットインフル抗原、不活化インフルエンザウイルス粒子、又はFA固定化インフルエンザウイルス粒子(A/CA株)を、タンパク質量として0.8μgの接種量で筋肉内接種した(一群当たり13匹)。免疫3週後にマウスを全採血し、安楽死させた。遠心分離によって血清を得て、「病原体検出マニュアル」(国立感染症研究所編)に従って、HI抗体価を測定した。マウスに筋肉内接種を行った際の免疫原性(HI抗体価(GMT))の結果を表7に示す。FA固定化インフルエンザウイルス粒子(A/CA株)は、スプリットインフル抗原と比較して高い免疫原性であり、不活化インフルエンザウイルス粒子と同等の免疫原性であった。

FA固定化インフルエンザウイルス粒子(A/CA株)における免疫原性を、上記「6.免疫原性1(マウス筋肉内接種)」と同じ手順でマウス皮内接種によって評価した。ddYマウス(雌性、8週齢)に、スプリットインフル抗原、不活化インフルエンザウイルス粒子、又はFA固定化インフルエンザウイルス粒子を、タンパク質量として0.2μgの接種量で皮内接種した(一群当たり5匹)。免疫3週後にマウスを全採血し、安楽死させた。遠心分離によって血清を得て、HI抗体価を測定した。マウスに皮内接種を行った際の免疫原性(HI抗体価(GMT))の結果を表8に示す。FA固定化インフルエンザウイルス粒子は、スプリットインフル抗原と比較して有意に免疫原性が高く、不活化インフルエンザウイルス粒子と同等の免疫原性であった。

A/CA株(H1N1亜型株)、A/NY株(H3N2亜型株)、B/BR株及びB/MA株(B型株)について、スプリットインフル抗原、不活化インフルエンザウイルス粒子、及びFA固定化インフルエンザウイルス粒子の免疫原性を、カニクイザル(雄性又は雌性、17~25ヶ月齢)を用いて評価した。スプリットインフル抗原、不活化インフルエンザウイルス粒子、又はFA固定化インフルエンザウイルス粒子を、1群当たり8又は9匹に7.5μgHA相当(スプリットインフル抗原のHA量に相当するタンパク質量)の接種量で、21日間隔で2回皮下接種した。2次免疫前、及び2次免疫後21日目に部分採血した。遠心分離によって血清を得て、「病原体検出マニュアル」(国立感染症研究所編)に従って、HI抗体価及び中和抗体価を測定した。免疫原性の結果を、表9(2次免疫後21日目のHI抗体価(GMT))、表10(2次免疫後21日目の中和抗体価(GMT))に示す。HI抗体価では、FA固定化インフルエンザウイルス粒子はスプリットインフル抗原と比較して高い免疫原性を有することがわかった。特に、B/MA株以外では、スプリットインフル抗原と比較して有意に免疫原性が高かった。中和抗体価においても、FA固定化インフルエンザウイルス粒子はスプリットインフル抗原と比較してA型株及びB型株全てにおいて有意に免疫原性が高かった。


抗体サブクラス解析として、上記「6.免疫原性1(マウス筋肉内接種)」で得られたA/CA株(H1N1亜型株)、及びB/MA株(B型株)について、マウスの血清中に含まれる、ウイルス抗原に特異的なIgG1及びIgG2a抗体価を酵素免疫測定(ELISA)法によって測定した。その結果、FA固定化インフルエンザウイルス粒子は、抗原特異的IgG1よりも抗原特異的IgG2aを誘導することが示された(表11、表12)。これは、不活化インフルエンザウイルス粒子と同様の結果であり、スプリットインフル抗原との比較においても、より強いIgG2a誘導傾向が見られた。この結果から、FA固定化インフルエンザウイルス粒子は、液性免疫を活性化するが細胞性免疫をほとんど活性化することができないスプリットインフル抗原に比べて、ワクチンの有効性のさらなる向上が期待できる。


実施例1に準じた方法で調製したFA固定化インフルエンザウイルス粒子(A/NY株)について、以下の手順で、プロテアーゼ処理中の経時的なRNA放出量を評価した。まず、FA固定化インフルエンザウイルス粒子をPBSで希釈し、SDSとProteinase Kを加えて55℃で反応させ、経時的にRNAを抽出した。RNA抽出には、TRIzol LS ReagentとPureLink RNA Mini Kit、PureLink DNase (invitrogen社製、商品名)を用いた。抽出したRNAの含有量は、Quant-iT RiboGreen RNA Reagent and Kit (invitrogen社製、商品名)で測定した。各FA濃度の経時的なRNA含量を表13に示す。その結果、FA濃度依存的にRNA放出が徐放化されていることが示された。FA固定によるRNA放出の徐放化が、炎症性サイトカイン産生を徐放化させ、高い安全性を生み出していることが示唆された。

(1)GA固定化インフルエンザウイルス粒子の調製
1.グルタルアルデヒド(GA)処理
A型インフルエンザウイルス(H3N2亜型株(A/NY株))及びB型インフルエンザウイルス(B型ビクトリア系統株(B/BR株))を実施例1と同様の方法で培養、精製した。精製した各インフルエンザウイルス粒子のタンパク質の最終濃度が1000μg/mLとなるように、各インフルエンザウイルス粒子を希釈し、懸濁液を得た。次に、1w/v%GA溶液を用いて、GA濃度が0.016w/v%又は0.008w/v%となるように希釈した。上記懸濁液と、希釈したGA溶液(0.016w/v%又は0.008w/v%)とを等量混合し、4℃で3日間反応させた。反応終了後、PBSで反応液を透析し、GAを除くことで、固定化インフルエンザウイルス粒子(以下、「GA固定化インフルエンザウイルス粒子」と記載することがある。)を得た。得られたGA固定化インフルエンザウイルス粒子(A/NY株及びB/BR株)の発熱活性を、ウサギ3羽における発熱反応の和で評価する発熱試験、及びヒトPBMCを刺激した場合の炎症性サイトカインの産生量の定量によって評価した。
実施例1と同様の方法によって、発熱試験を行った。不活化インフルエンザウイルス粒子及びGA固定化インフルエンザウイルス粒子(A/NY株及びB/BR株)を生理食塩水で希釈し、1mL中のタンパク質含量を240μgとしたものを試料とした。上記試料を、体重1kg当たりにつき1mLをウサギに接種して、6時間後まで発熱の上昇を観察した。A/NY株のウサギ3羽の発熱反応の和(℃)を表14、B/BR株のウサギ3羽の発熱反応の和(℃)を表15に示す。


欧州薬局方MONOCYTE-ACTIVATION TESTに準じた方法で、GA固定化インフルエンザウイルス粒子又は不活化インフルエンザウイルス粒子でヒトPBMCを刺激した場合のサイトカイン(IL-1β)の産生量を定量した。具体的には、欧州薬局方MONOCYTE-ACTIVATION TESTにおけるヒトPBMCを最低4ドナー分プールして使用するところを、1ドナー分に変更して測定した。GA固定化インフルエンザウイルス粒子(A/NY株及びB/BR株)のサイトカインの産生量の結果を表16、表17に示す。GA固定化インフルエンザウイルス粒子は、不活化インフルエンザウイルス粒子と比較して、炎症性サイトカインの産生量が十分低いことがわかった。このことから、GA固定化インフルエンザウイルス粒子は、スプリットインフル抗原と同様に、高い安全性を有することが示唆された。

物性評価
上記実施例3で得られたGA固定化インフルエンザウイルス粒子の物性を以下の方法で評価した。
GA固定化インフルエンザウイルス粒子(A/NY株)の形状をさらに詳細に調べるために、実施例2と同様の方法で、電子顕微鏡による観察を実施した。代表として、0.008w/v%のGA濃度で4℃、3日間反応させたGA固定化インフルエンザウイルス粒子(A/NY株)を撮影した写真を図2に示す。GA固定化インフルエンザウイルス粒子は粒子構造が保たれており、粒子同士が結合しあった凝集体は認められなかった。
GA固定化インフルエンザウイルス粒子の平均粒子径をZetasizer Nano ZS(Malvern社製)を用いて解析した。動的光散乱法による液体中の平均粒子径を表18に示す。GA固定化インフルエンザウイルス粒子は、平均粒子径が130~160nm前後で単峰性であった。このことから、GA固定化インフルエンザウイルス粒子の平均粒子径は、元となるウイルス粒子と同等であることが確認された。すなわち、GA固定化インフルエンザウイルス粒子の平均粒子径は単一で変わらないことがわかった。また、GA固定化インフルエンザウイルス粒子の粒子構造は保たれており、凝集体のような不純物は認められなかった。

GA固定化インフルエンザウイルス粒子の分子量分布を実施例2と同様の方法(SEC)で測定した。その溶出パターンを表19に示す。スプリットインフル抗原は、溶出時間16~30分付近に四峰性のピークが認められた。一方、GA固定化インフルエンザウイルス粒子は、溶出時間16~17分付近に単峰性のピークが認められた。

GA固定化インフルエンザウイルス粒子の固定化剤での架橋度を、実施例2と同様の方法で分析した。B/BR株(B型ビクトリア系統株)の結果を表20に示す。グルタルアルデヒド濃度0.004w/v%、0.008w/v%で固定したGA固定化インフルエンザウイルス粒子のM1タンパク質残存率は、それぞれ23.8%、10.8%であり、グルタルアルデヒド濃度が高まるにつれて架橋が促進され、M1タンパク質残存率が低下することがわかった。

GA固定化インフルエンザウイルス粒子(B/BR株)における免疫原性を、以下の手順で評価した。まず、ddYマウス(雌性、8週齢)に、スプリットインフル抗原、又はGA固定化インフルエンザウイルス粒子をタンパク質量として0.8μgの接種量で筋肉内接種した(一群当たり16匹)。免疫3週後にマウスを全採血し、安楽死させた。遠心分離によって血清を得て、中和抗体価を測定した。代表としてB/BR株(B型ビクトリア系統株)の免疫原性(中和抗体価(GMT))の結果を表21に示す。B型株の場合、GA固定化インフルエンザウイルス粒子は、スプリットインフル抗原と比較して高い免疫原性であった。

抗体サブクラス解析として、上記「5.免疫原性(マウス筋肉内接種)」で得られたマウスの血清中に含まれる、インフルエンザウイルスの抗原に特異的なIgG1及びIgG2a抗体価をELISA法によって測定した。その結果、スプリットインフル抗原とは対照的に、GA固定化インフルエンザウイルス粒子は、抗原特異的IgG1よりも抗原特異的IgG2aを誘導することが示された(表22)。これは、不活化インフルエンザウイルス粒子と同様の結果である。この結果から、GA固定化インフルエンザウイルス粒子は、液性免疫を活性化するが細胞性免疫をほとんど活性化することができないスプリットインフル抗原に比べて、ワクチンの有効性のさらなる向上が期待できる。

(1)EDC固定化インフルエンザウイルス粒子の調製
1.1-エチル-3-[3-ジメチルアミノプロピル]カルボジイミド塩酸塩(EDC)処理
A型インフルエンザウイルス(H3N2亜型株(A/NY株))及びB型インフルエンザウイルス(ビクトリア系統株B/BR株)を実施例1と同様に培養、精製した。精製した各インフルエンザウイルス粒子のタンパク質の最終濃度がA/NY株は2500μg/mL、B/BR株は500μg/mLとなるように、得られた画分を希釈し、懸濁液を得た。EDC溶液を、0.1~4MとなるようにPBSで段階希釈し、最終濃度が50~500mMとなるように懸濁液に添加し、氷冷(0℃)で、2~20時間反応させた。反応終了後、PBSで反応液を透析し、EDCを除くことで、固定化インフルエンザウイルス粒子(以下、「EDC固定化インフルエンザウイルス粒子」と記載することがある。)を得た。得られたEDC固定化インフルエンザウイルス粒子(A/NY株、B/BR株)の発熱活性を、ウサギ3羽における発熱反応の和で評価する発熱試験、及びヒトPBMCを刺激した場合の炎症性サイトカインの産生量の定量によって評価した。
実施例1と同様の方法によって、発熱試験を行った。EDC固定化インフルエンザウイルス粒子を生理食塩水で希釈し、1mL中のタンパク質含量を240μgとしたものを試料とした。EDC固定化インフルエンザウイルス粒子は、50mM又は500mMのEDC濃度で、氷冷で2時間反応した後、PBSで反応液を透析処理した検体を用いた(A/NY株については、5mMのEDC濃度も実施した。)。また、A/NY株については、5mMのEDC濃度で、4℃で20時間反応した後、PBSで反応液を透析処理した検体も用いた。上記試料を、体重1kg当たりにつき1mLをウサギに接種して、6時間後まで発熱の上昇を観察した。ウサギ3羽の発熱反応の和(℃)を表23、表24に示す。


欧州薬局方MONOCYTE-ACTIVATION TESTに準じた方法で、EDC固定化インフルエンザウイルス粒子又は不活化インフルエンザウイルス粒子でヒトPBMCを刺激した場合のサイトカイン(IL-1β及びIL-6)の産生量を定量した。具体的には、欧州薬局方MONOCYTE-ACTIVATION TESTにおけるヒトPBMCを最低4ドナー分プールして使用するところを、1ドナー分に変更して測定した。A/NY株及びB/BR株のEDC固定化インフルエンザウイルス粒子及び不活化インフルエンザウイルス粒子のサイトカインの産生量の結果を表25、表26に示す。EDC固定化インフルエンザウイルス粒子は、不活化インフルエンザウイルス粒子と比較して、炎症性サイトカイン産生量が十分低いことがわかった。このことから、EDC固定化インフルエンザウイルス粒子は、不活化インフルエンザウイルス粒子と比較して、副反応を抑制しやすいことが示唆された。

物性評価
上記実施例5で得られたEDC固定化インフルエンザウイルス粒子の物性を以下の方法で評価した。
実施例2と同様の方法によって、EDC固定化インフルエンザウイルス粒子をショ糖密度勾配遠心法によって分析した。代表として、50mMのEDC濃度にて、氷冷で2時間反応した後のEDC固定化インフルエンザウイルス粒子(A/NY株(H3N2亜型株))の結果を表27に示す。スプリットインフル抗原では、タンパク質がショ糖濃度25~50%に幅広く分布し、ウイルス粒子が分解していることが示された。これに対し、EDC固定化インフルエンザウイルス粒子では高いショ糖濃度(47.2%)に単一ピーク(粒子性)として分画されていることが示された。HA活性については10240倍であった。

EDC固定化インフルエンザウイルス粒子の形状を詳細に調べるために、実施例2と同様の方法で、電子顕微鏡による観察を実施した。代表として、500mMのEDC濃度にて、氷浴で2時間反応した後のEDC固定化インフルエンザウイルス粒子を撮影した写真を図3に示す。EDC固定化インフルエンザウイルス粒子は粒子構造が保たれており、抗原同士が結合しあった凝集体は認められなかった。
EDC固定化インフルエンザウイルス粒子の平均粒子径をZetasizer Nano ZS(Malvern社製)を用いて解析した。動的光散乱法による液体中の平均粒子径を表28に示す。EDC固定化インフルエンザウイルス粒子(A/NY株(H3N2亜型株)、B/BR株(B型株))は、平均粒子径が130~160nm前後で単峰性であった。このことから、EDC固定化インフルエンザウイルス粒子の平均粒子径は、元となるウイルス粒子と同等であることが確認された。すなわち、EDC処理されても固定化インフルエンザウイルス粒子の平均粒子径は単一で変わらないことがわかった。また、EDC固定化インフルエンザウイルス粒子の粒子構造は保たれており、凝集体のような不純物は認められなかった。

H3N2亜型株(A/NY株)を元とするEDC固定化インフルエンザウイルス粒子の分子量分布を測定した。測定は、サイズ排除クロマトグラフィー(商品名:TSKgel G6000PWXL(TOSOH社製))を用いて行った(溶離液にはPBSを用いて、流速は0.5ml/minで行った)。その溶出パターンを表29に示す。スプリットインフル抗原は溶出時間16~30分付近に四峰性のピークが認められた。一方、EDC固定化インフルエンザウイルス粒子は溶出時間16~17分付近に単峰性のピークが認められた。
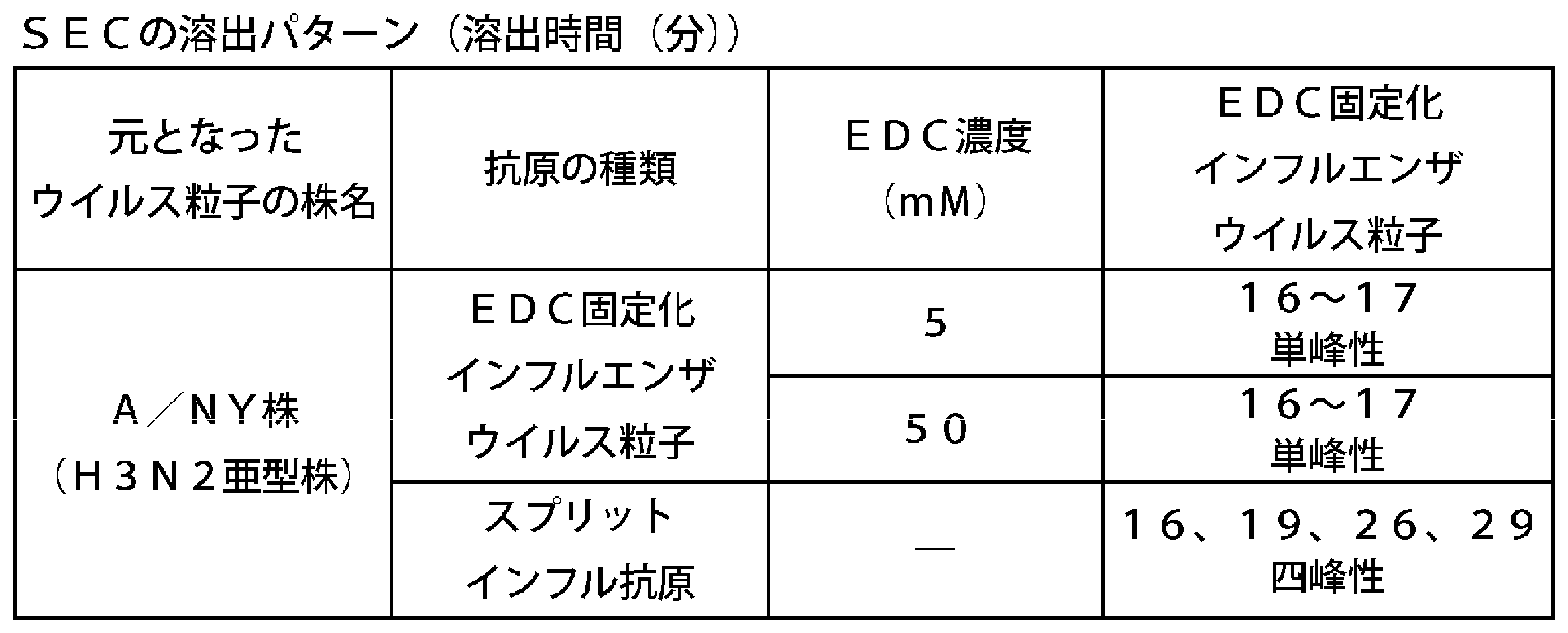
実施例5によって調製したEDC固定化インフルエンザウイルス粒子の固定化剤での架橋度を実施例2と同様の方法で分析した。A/NY株(H3N2亜型株)の結果を表30に示す。EDC濃度50mM、500mMで固定したEDC固定化インフルエンザウイルス粒子のM1タンパク質残存率(%)は、それぞれ85.7%、53.4%であり、EDC濃度が高まるにつれて架橋が促進され、M1タンパク質残存率が低下することがわかった。B/BR株(B型ビクトリア系統株)の結果を表31に示す。EDC濃度5mM、50mM、500mMで固定したEDC固定化インフルエンザウイルス粒子のM1タンパク質残存率(%)は、それぞれ85.1%、56.1%、27.2%であった。A/NY株(H3N2亜型株)と同様に、EDC濃度が高まるにつれて架橋が促進され、M1タンパク質残存率が低下することがわかった。

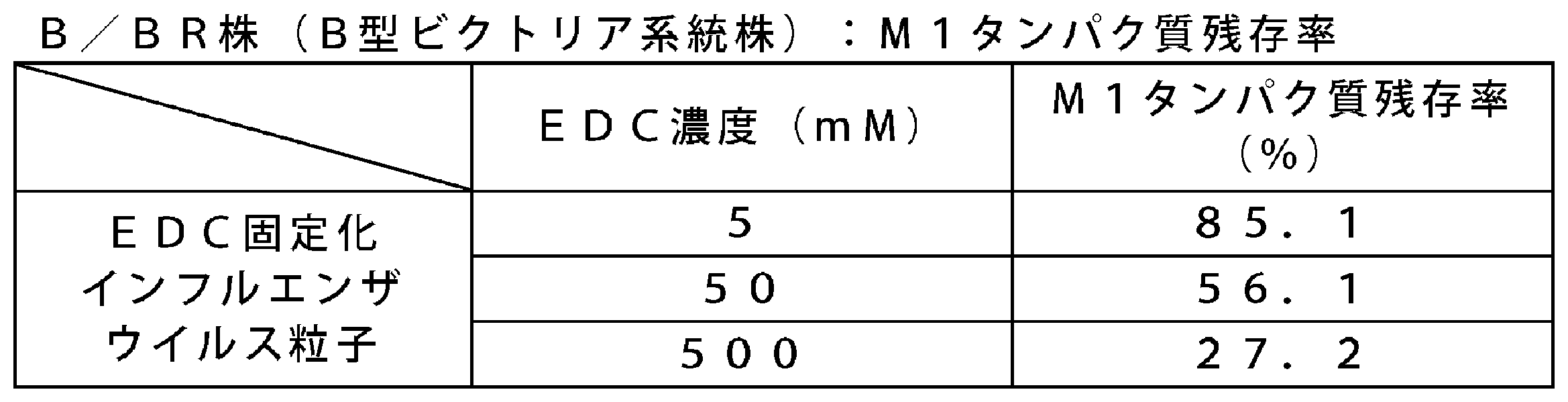
EDC固定化インフルエンザウイルス粒子における免疫原性を、マウスを用いて評価した。実施例4と同様の方法によって血清を得て、HI抗体価及び中和抗体価を測定した。代表として50mM及び500mMのEDC濃度にて、氷浴で2時間反応した後のEDC固定化インフルエンザウイルス粒子の免疫原性の結果を、表32(A/NY株のHI抗体価(GMT))、表33(A/NY株の中和抗体価(GMT))及び表34(B/BR株の中和抗体価(GMT))に示す。A/NY株(H3N2亜型株)、及びB/BR株(B型ビクトリア系統株)の場合、EDC固定化インフルエンザウイルス粒子はスプリットインフル抗原と比較し有意に高い免疫原性であった。



抗体サブクラス解析として、上記「6.免疫原性(マウス筋肉内接種)」で得られたマウスの血清中に含まれる、ウイルス抗原に特異的なIgG1及びIgG2a抗体価をELISA法によって測定した。その結果、スプリットインフル抗原とは対照的に、EDC固定化インフルエンザウイルス粒子は抗原特異的IgG1よりも抗原特異的IgG2aを誘導することが示された(表35、表36)。この結果から、EDC固定化インフルエンザウイルス粒子は、液性免疫を活性化するが細胞性免疫をほとんど活性化することができないスプリットインフル抗原に比べて、ワクチンの有効性のさらなる向上が期待できる。


EDC固定化インフルエンザウイルス粒子における免疫原性を、カニクイザルを用いて以下の手順で評価した。まず、カニクイザル(雄性又は雌性、29~35ヶ月齢)に、スプリットインフル抗原、又はEDC固定化インフルエンザウイルス粒子をHA含量として15μgの接種量で皮下接種した(一群当たり8匹)。3週間隔で2回皮下接種し、2次免疫後4週間目に採血した。実施例2と同様の方法によって血清を得て、HI抗体価及び中和抗体価を測定した。代表として5mMのEDC濃度にて、4℃で20時間反応した後のEDC固定化インフルエンザウイルス粒子の免疫原性のHI抗体価(GMT)の結果を表37に示し、中和抗体価(GMT)の結果を表38に示す。A/CA株(H1N1亜型株)、及びA/NY株(H3N2亜型株)、B/MA株(B型山形系統)、B/BR株(B型ビクトリア系統株)において、EDC固定化インフルエンザウイルス粒子はスプリットインフル抗原と比較して有意に免疫原性が高かった。


実施例5によって調製したEDC固定化インフルエンザウイルス粒子の安定性を、苛酷試験の前後における架橋度の変化を指標として評価した。まず、実施例2と同様の方法によってM1タンパク質残存率(%)を算出し、さらに、苛酷試験前のM1タンパク質残存率を100%とした場合の、37℃、1週間の苛酷試験後のM1タンパク質残存率の割合を算出した。A/NY株(H3N2亜型株)の結果を表39に示す。不活化インフルエンザウイルス粒子の苛酷試験後のM1タンパク質残存率は、苛酷試験前に比べて24%上昇した。一方、EDC濃度50、500mMで固定したEDC固定化インフルエンザウイルス粒子の苛酷試験後のM1タンパク質残存率は、苛酷試験前に比べてそれぞれ18%減少し、不活化インフルエンザウイルス粒子よりも変化率が小さく、より安定であることが示唆された。

実施例5によって調製したEDC固定化インフルエンザウイルス粒子の安定性を、苛酷試験の前後におけるHA含量の変化を指標として評価した。まず、一元放射免疫拡散試験(生物学的製剤基準(日本国、厚生労働省告示 第192号))によってHA含量を算出し、さらに、苛酷試験前のHA含量を100%とした場合の、37℃、1週間の苛酷試験後のHA含量の変化率を算出した。A/CA株(H1N1亜型株)、及びA/NY株(H3N2亜型株)、B/MA株(B型山形系統)、B/BR株(B型ビクトリア系統株)の結果を表40に示す。EDC固定化インフルエンザウイルス粒子の苛酷試験後のHA含量の変化率は小さく、安定であることが示唆された。

実施例5によって調製したEDC固定化インフルエンザウイルス粒子の安定性を、5℃、11ヶ月保存の前後における架橋度の変化を指標として評価した。まず、実施例2と同様の方法によってM1タンパク質残存率(%)を算出し、さらに、5℃、11ヶ月保存前のM1タンパク質残存率を100%とした場合の、5℃、11ヶ月保存下でのM1タンパク質残存率の割合を算出した。A/CA株(H1N1亜型株)、及びA/NY株(H3N2亜型株)、B/MA株(B型山形系統)、B/BR株(B型ビクトリア系統株)の結果を表41に示す。EDC固定化インフルエンザウイルス粒子の5℃、11ヶ月保存下でのM1タンパク質残存率の変化率は小さく、安定であることが示唆された。

実施例5によって調製したEDC固定化インフルエンザウイルス粒子の安定性を、5℃、11ヶ月保存の前後におけるHA含量の変化を指標として評価した。まず、一元放射免疫拡散試験(生物学的製剤基準(日本国、厚生労働省告示 第192号))によってHA含量を算出し、さらに、5℃、11ヶ月保存前のHA含量を100%とした場合の、5℃、11ヶ月保存下でのHA含量の変化率を算出した。A/CA株(H1N1亜型株)、及びA/NY株(H3N2亜型株)、B/MA株(B型山形系統)、B/BR株(B型ビクトリア系統株)の結果を表42に示す。EDC固定化インフルエンザウイルス粒子の5℃、11ヶ月保存下でのHA含量の変化率は小さく、安定であることが示唆された。

実施例5によって調製したEDC固定化インフルエンザウイルス粒子の5℃、9ヶ月保存下での安定性を、マウス免疫原性(筋肉内接種)を用いて評価した。実施例4と同様の方法によって血清を得て、HI抗体価及び中和抗体価を測定した。代表として5mMのEDC濃度にて、4℃で20時間反応した後のEDC固定化インフルエンザウイルス粒子の免疫原性のHI抗体価(GMT)の結果を表43に示し、中和抗体価(GMT)の結果を表44に示す。A/CA株(H1N1亜型株)、及びA/NY株(H3N2亜型株)、B/MA株(B型山形系統)、B/BR株(B型ビクトリア系統株)において、EDC固定化インフルエンザウイルス粒子はスプリットインフル抗原と比較して免疫原性が高かった。
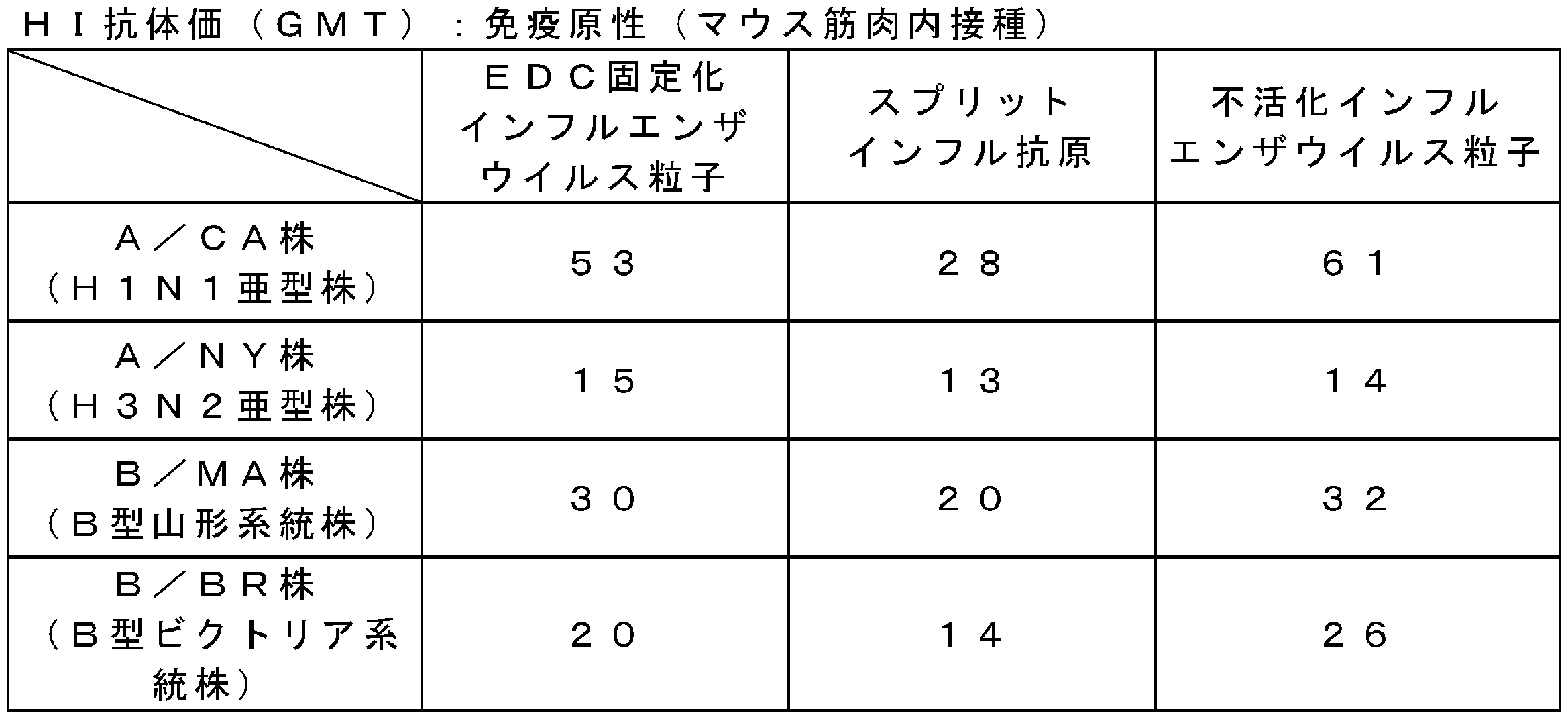

抗体サブクラス解析として、上記「13.5℃、9ヶ月保存下での安定性(マウス免疫原性(筋肉内接種))」で得られたマウスの血清中に含まれる、ウイルス抗原に特異的なIgG1及びIgG2a抗体価をELISA法によって測定した。その結果、5℃、9ヶ月保存後においても、スプリットインフル抗原とは対照的に、EDC固定化インフルエンザウイルス粒子は抗原特異的IgG1よりも抗原特異的IgG2aを誘導することが示された(表45)。この結果から、EDC固定化インフルエンザウイルス粒子が活性化させる細胞性免疫は5℃、9ヶ月保存後も維持されていることが期待できる。
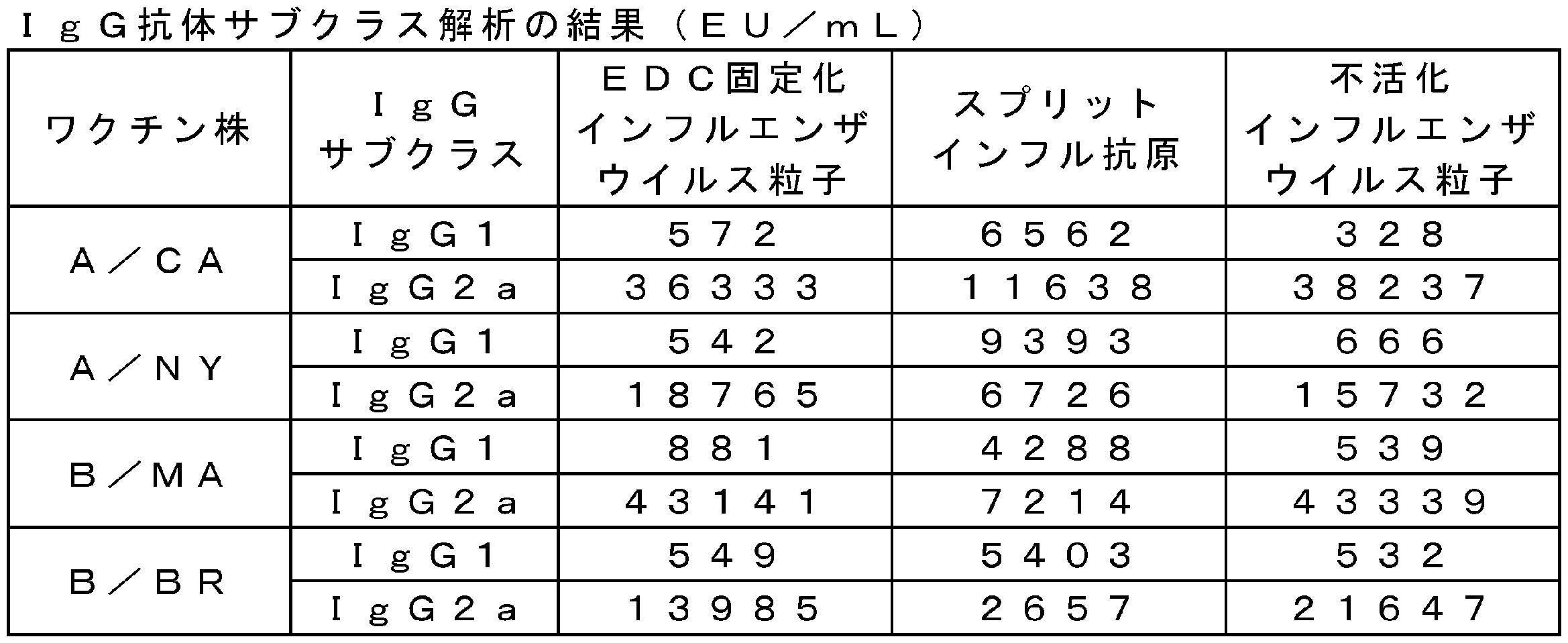
実施例5によって調製したEDC固定化インフルエンザウイルス粒子の5℃、10ヶ月保存下での安定性を、カニクイザル免疫原性(皮下接種)を用いて評価した。まず、カニクイザル(雄性又は雌性、29~35ヶ月齢)に、スプリットインフル抗原、EDC固定化インフルエンザウイルス粒子、及び不活化インフルエンザウイルス粒子をHA含量として15μgの接種量で皮下接種した(一群当たり8匹)。3週間隔で2回皮下接種し、2次免疫後4週間目に採血した。実施例2と同様の方法によって血清を得て、HI抗体価及び中和抗体価を測定した。代表として5mMのEDC濃度にて、4℃で20時間反応した後のEDC固定化インフルエンザウイルス粒子の5℃、10ヶ月保存下でのHI抗体価(GMT)の結果を表46に示し、中和抗体価(GMT)の結果を表47に示す(表46、表47は、それぞれ表37、表38の再掲)。A/CA株(H1N1亜型株)、及びA/NY株(H3N2亜型株)、B/MA株(B型山形系統)、B/BR株(B型ビクトリア系統株)において、EDC固定化インフルエンザウイルス粒子はスプリットインフル抗原と比較して有意に免疫原性が高かった。


実施例5によって調製したEDC固定化インフルエンザウイルス粒子のプロテアーゼ処理中の経時的なRNA放出量を評価した。まず、不活化インフルエンザウイルス粒子及びEDC固定化インフルエンザウイルス粒子をPBSで希釈し、SDSとProteinase Kを加えて55℃で反応させ、経時的にRNAを抽出した。RNA抽出には、TRIzol LS ReagentとPureLink RNA Mini Kit、PureLink DNase (invitrogen社製、商品名)を用いた。抽出したRNAの含有量は、Quant-iT RiboGreen RNA Reagent and Kit (invitrogen社製、商品名)で測定した。代表として、A/NY株(H3N2亜型株)の結果を表48に示す。その結果、不活化インフルエンザウイルス粒子に比べて、EDC固定化インフルエンザウイルス粒子のRNA放出が徐放化されていることが示された。
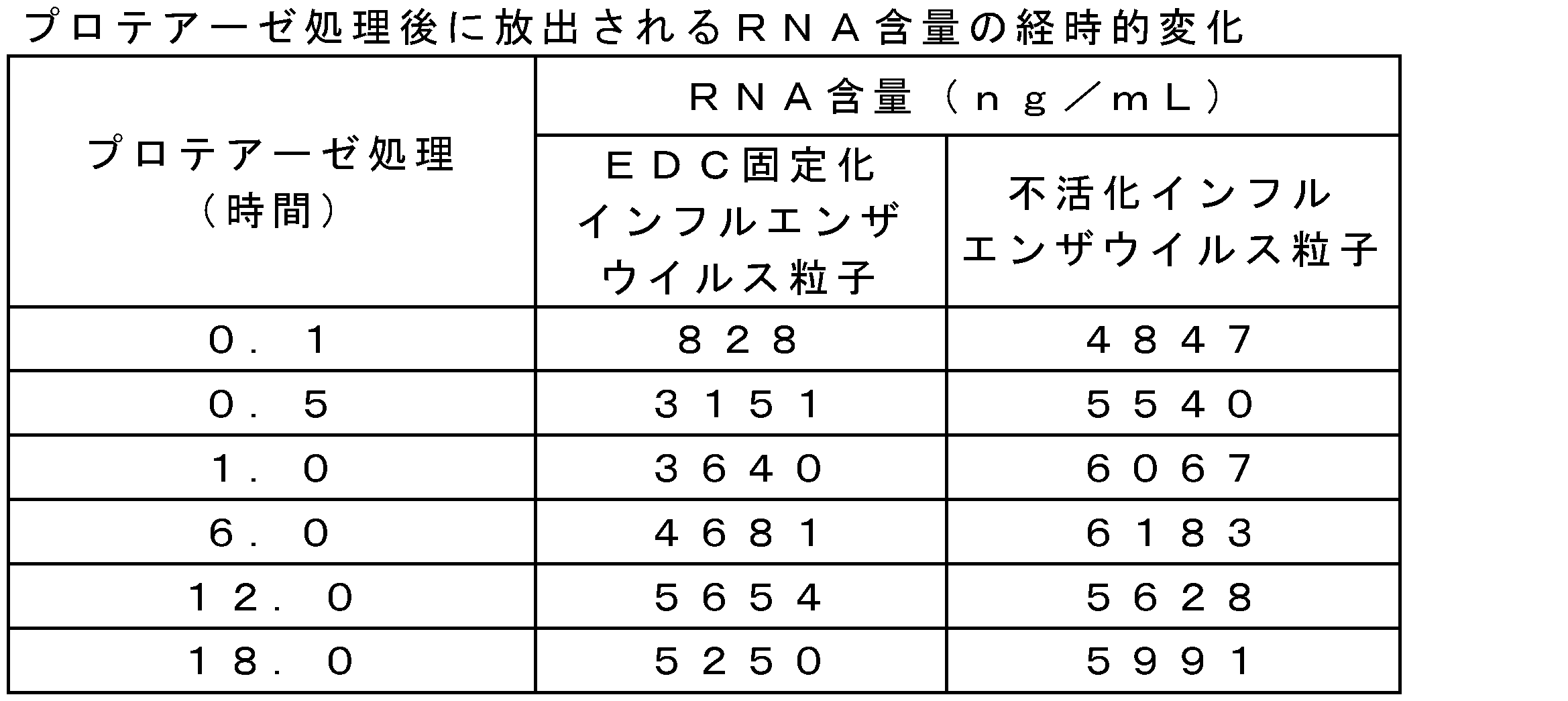
実施例5によって調製したEDC固定化インフルエンザウイルス粒子の炎症性サイトカインの経時的な産生量を評価した。方法は、欧州薬局方MONOCYTE-ACTIVATION TESTに準じた。具体的には、欧州薬局方MONOCYTE-ACTIVATION TESTにおけるヒトPBMCを最低4ドナー分プールして使用するところを、1ドナーに変更して測定した。A/NY株(H3N2亜型株)における、不活化インフルエンザウイルス粒子及びEDC固定化インフルエンザウイルス粒子のIL-1β産生量の経時的変化を表49に、IL-6産生量の経時的変化を表50に示す。また、B/BR株(B型ビクトリア系統株)における、不活化インフルエンザウイルス粒子及びEDC固定化インフルエンザウイルス粒子のIL-1β産生量の経時的変化を表51に、IL-6産生量の経時的変化を表52に示す。その結果、不活化インフルエンザウイルス粒子に比べて、EDC固定化インフルエンザウイルス粒子の炎症性サイトカイン産生が徐放化されていることが示された。つまり、EDC固定によって体内でのRNA放出が徐放化され、炎症性サイトカイン産生を遅延させることで、EDC固定化インフルエンザウイルス粒子が高い安全性を有していることが示唆された。
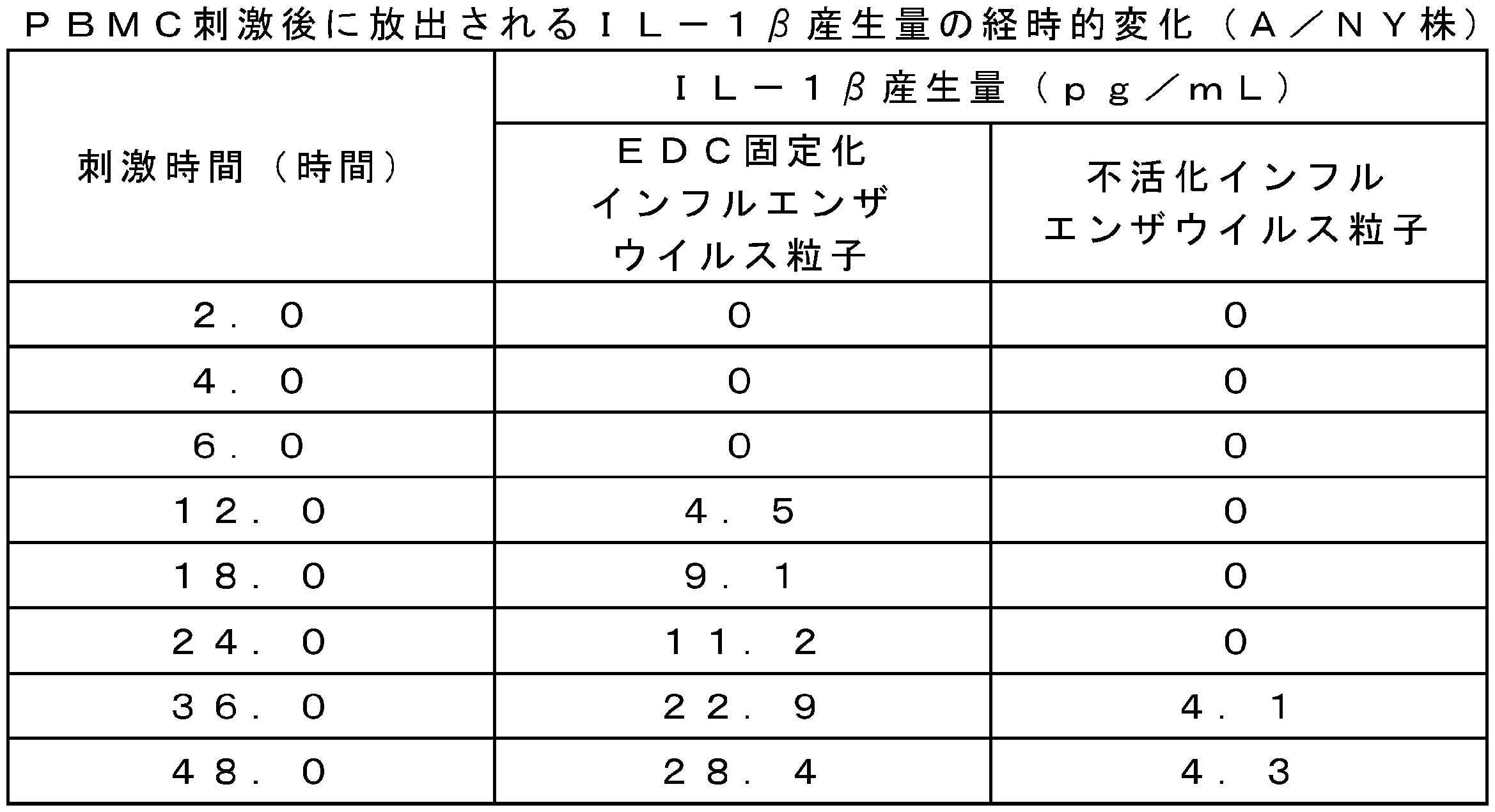



GA固定化日本脳炎ウイルス粒子の調製
1.グルタルアルデヒド処理(ウイルス粒子に対して粒子構造を固定化する工程)
Vero細胞培養日本脳炎ワクチン原薬(化学及血清療法研究所製、商品名「エンセバック」、0.08v/v%ホルマリンで不活化済の日本脳炎ウイルス粒子をタンパク質濃度で60~90μg/ml含む。以下、「不活化日本脳炎ウイルス粒子」と記載することがある。)に対して、最終濃度が0.005~0.02w/v%となるようにグルタルアルデヒドを添加し、4℃で3日間反応させた。反応終了後、PBS様溶液(賦活剤である乳糖(終濃度5w/v%)を添加したPBS)で、得られた反応液を透析した。透析によって、グルタルアルデヒドを除くことで、固定化日本脳炎ウイルス粒子(以下、「GA固定化日本脳炎ウイルス粒子」と記載することがある。)を得た。得られたGA固定化日本脳炎ウイルス粒子の発熱活性を、ウサギ3羽における発熱反応の和で評価する発熱試験、ヒトPBMCを刺激した場合の炎症性サイトカインの産生量の定量によって評価した。
実施例1と同様の方法によって、発熱試験を行った。GA固定化日本脳炎ウイルス粒子又は不活化日本脳炎ウイルス粒子を生理食塩水で希釈し、1mL中のタンパク質含量を70μgとしたものを試料とした。上記試料を、体重1kg当たりにつき3mLをウサギに接種して、6時間後まで発熱の上昇を観察した。ウサギ3羽の発熱反応の和(℃)を表53に示す。代表として、0.01w/v%のグルタルアルデヒド濃度で4℃、3日間反応後のGA固定化日本脳炎ウイルス粒子を評価した。

欧州薬局方MONOCYTE-ACTIVATION TESTに準じた方法で、GA固定化日本脳炎ウイルス粒子又は不活化日本脳炎ウイルス粒子でヒトPBMCを刺激した場合のサイトカイン(IL-1β及びIL-6)の産生量を定量した。具体的には、欧州薬局方MONOCYTE-ACTIVATION TESTにおけるヒトPBMCを最低4ドナー分プールして使用するところを、1ドナー分に変更して測定した。その結果を表54に示す。GA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子よりも炎症性サイトカインの産生量が十分低いことがわかった。このことからも、GA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子よりも高い安全性を有することが示唆された。

物性評価
上記実施例7で得られたGA固定化日本脳炎ウイルス粒子の物性を以下の方法で評価した。
GA固定化日本脳炎ウイルス粒子の形状を詳細に調べるために、実施例2と同様の方法で、電子顕微鏡による観察を実施した。代表として、0.01w/v%のグルタルアルデヒド濃度で4℃、3日間反応後のGA固定化日本脳炎ウイルス粒子を撮影した写真を図4示す。GA固定化日本脳炎ウイルス粒子は、グルタルアルデヒドで固定することで不活化日本脳炎ウイルス粒子と同様に粒子構造が保たれていた。
GA固定化日本脳炎ウイルス粒子の平均粒子径を、Zetasizer Nano ZS(Malvern社製)を用いて解析した。動的光散乱法による液体中の平均粒子径を表55に示す。GA固定化日本脳炎ウイルス粒子は、平均粒子径が約90nmで単峰性であった。一方、不活化日本脳炎ウイルス粒子は、約80nmで単峰性であった。また、GA固定化日本脳炎ウイルス粒子の粒子構造は保たれており、凝集体のような不純物は認められなかった。

GA固定化日本脳炎ウイルス粒子の分子量分布を測定した。測定は、サイズ排除クロマトグラフィー(商品名:Superose 6 10/300 GE(GEヘルスケア社製))を用いて行った(溶離液にはPBSを用いて、流速は0.5ml/minで行った)。その溶出パターンを表56に示す。GA固定化日本脳炎ウイルス粒子は、溶出時間14~15分付近に単峰性の主ピークが認められた。また、不活化日本脳炎ウイルス粒子も溶出時間14~15分付近に単峰性の主ピークが認められた。

抗日本脳炎ウイルス抗体を用いたサンドイッチELISA法によって、抗原の含有量(抗原含量)を以下の手順で測定した。検体中に含まれるE抗原は、抗日本脳炎ウイルスウサギIgG(一次抗体;ポリクローナル抗体)が結合したマイクロプレートに捕捉される。その後、西洋わさび由来ペルオキシダーゼ(HRP)を結合した抗日本脳炎ウイルスEタンパク質モノクローナル抗体(二次抗体;モノクローナル抗体)を反応させることで、プレートに結合した抗E抗原抗体/E抗原/二次抗体の複合体が形成される。反応せず残った試薬及び検体を洗浄によって除去し、酵素基質液(o-フェニレンジアミン溶液:OPD溶液)と反応させるとE抗原複合体上のHRPが反応し、発色が起きる。OPDの発色の強度は、複合体の量(E抗原量を反映)に比例することを利用し、E抗原含量(抗原含量)を測定した。

実施例7で調製したGA固定化日本脳炎ウイルス粒子について、比活性(抗原含量/タンパク質含量)により、固定化剤での架橋度を評価した。具体的には、抗原含量の測定に使用するモノクローナル抗体(503)は、中和エピトープを認識しており、中和エピトープの構造変化が起こると比活性が低下する。固定化を行っていない不活化日本脳炎ウイルス粒子の比活性に対する、各グルタルアルデヒド濃度で処理したGA固定化日本脳炎ウイルス粒子の比活性の相対値(%)(以下、この相対値を「503抗体反応率」(%)と記載することがある。)を算出した。結果を表58に示す。グルタルアルデヒド濃度0.005w/v%、0.01w/v%、0.02w/v%で固定したGA固定化日本脳炎ウイルス粒子の503抗体反応率は、それぞれ95.1%、74.7%、55.2%であり、グルタルアルデヒド濃度が高まるにつれて架橋が促進され、503抗体エピトープの構造変化による反応率の低下が示唆された。
ddYマウス(雌性、4週齢)に、代表として、0.005若しくは0.01w/v%のグルタルアルデヒド濃度で4℃、3日間反応後のGA固定化日本脳炎ウイルス粒子又は不活化日本脳炎ウイルス粒子を1μg又は0.25μgの接種量で腹腔内接種した(一群当たり10匹)。免疫1週間後に再度免疫し、その1週間後にマウスを全採血し、安楽死させた。遠心分離によって血清を得て、群毎に等量ずつプールし、中和抗体価を「病原体検出マニュアル」(国立感染症研究所編)に従って、測定した。50%プラック減少率から算出した結果を表59に示す。GA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子と比較して同等又はそれ以上の中和抗体価であった。

最終タンパク質濃度が8μg/mLとなるように、PBS様溶液でGA固定化日本脳炎ウイルス粒子及び不活化日本脳炎ウイルス粒子を希釈した。抗原含量を指標に、25℃での保存安定性を評価した。代表として、0.01w/v%のグルタルアルデヒド濃度で4℃、3日間反応後のGA固定化日本脳炎ウイルス粒子又は不活化日本脳炎ウイルス粒子の結果を表60に示す。GA固定化日本脳炎ウイルス粒子は、25℃保存下では1ヶ月間維持された。一方、不活化日本脳炎ウイルス粒子は、25℃保存下では低下を示した。GA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子と比較し安定性が向上することが示された。
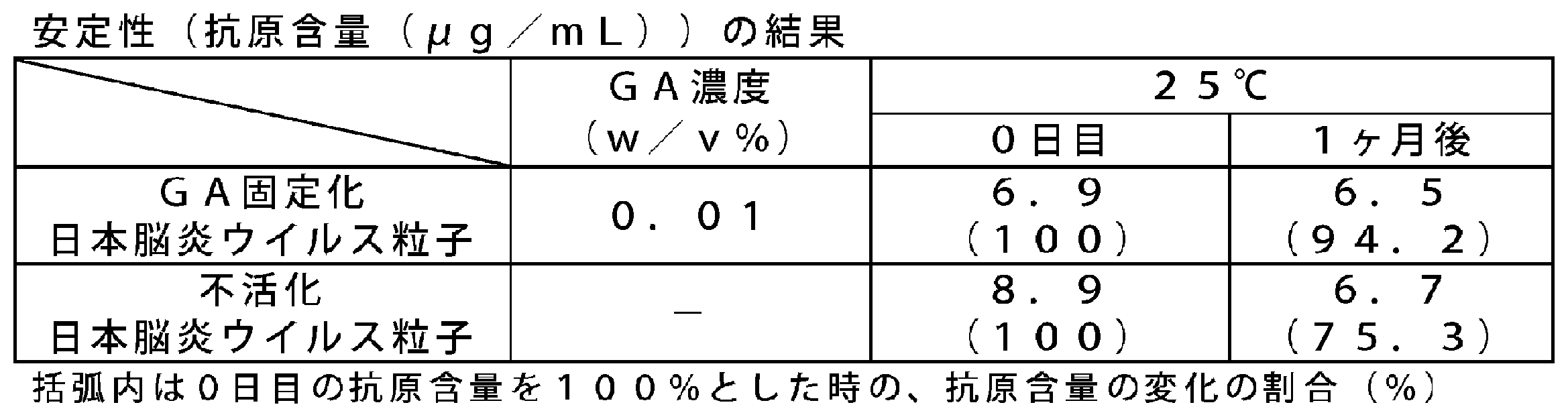
最終タンパク質濃度が8μg/mLとなるように、PBS様溶液でGA固定化日本脳炎ウイルス粒子及び不活化日本脳炎ウイルス粒子を希釈した。Zetasizer Nano ZSを用いた動的光散乱法による液体中の平均粒子径を指標に、25℃での保存安定性を評価した。代表として、0.01w/v%のグルタルアルデヒド濃度で4℃、3日間反応後のGA固定化日本脳炎ウイルス粒子又は不活化日本脳炎ウイルス粒子の結果を表61に示す。GA固定化日本脳炎ウイルス粒子は、25℃保存下では1ヶ月間維持された。一方、不活化日本脳炎ウイルス粒子は、25℃保存下では増加を示した。これにより、GA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子と比較し安定性が向上することが示された。

最終タンパク質濃度が8μg/mLとなるように、PBS様溶液でGA固定化日本脳炎ウイルス粒子及び不活化日本脳炎ウイルス粒子を希釈した。マウスでの免疫原性(中和抗体価)を指標に、4℃での保存安定性を評価した。代表として、0.01w/v%のグルタルアルデヒド濃度で4℃、15ヶ月間反応後のGA固定化日本脳炎ウイルス粒子又は不活化日本脳炎ウイルス粒子の結果を表62に示す。0ヶ月と15ヶ月後で投与量が異なるものの、GA固定化日本脳炎ウイルス粒子は、4℃保存下では15ヶ月間維持されることが推察された。一方、不活化日本脳炎ウイルス粒子は、4℃保存下では低下することが推察された。GA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子と比較して、安定性が向上することが推察された。

FA固定化日本脳炎ウイルス粒子の調製
1.ホルマリン処理(ウイルス粒子に対して粒子構造を固定化する工程)
不活化日本脳炎ウイルス粒子に対して、最終濃度が0.014~0.04v/v%(ホルムアルデヒド換算で、0.005~0.015w/v%)となるようにホルマリン(36~38w/v%ホルムアルデヒド水溶液)を添加し、25℃で1週間反応させた。反応終了後、PBS様溶液で反応液を透析し、ホルマリンを除くことで、固定化日本脳炎ウイルス粒子(以下、「FA固定化日本脳炎ウイルス粒子」と記載することがある。)を得た。得られたFA固定化日本脳炎ウイルス粒子の発熱活性を、ヒトPBMCを刺激した場合の炎症性サイトカインの産生量の定量によって評価した。
欧州薬局方MONOCYTE-ACTIVATION TESTに準じた方法で、FA固定化日本脳炎ウイルス粒子又は不活化日本脳炎ウイルス粒子でヒトPBMCを刺激した場合のサイトカイン(IL-1β及びIL-6)の産生量を定量した。具体的には、欧州薬局方MONOCYTE-ACTIVATION TESTにおけるヒトPBMCを最低4ドナー分プールして使用するところを、1ドナー分に変更して測定した。その結果を表63に示す。FA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子よりも炎症性サイトカインの産生量が十分低いことがわかった。このことからも、FA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子よりも高い安全性を有することが示唆された。
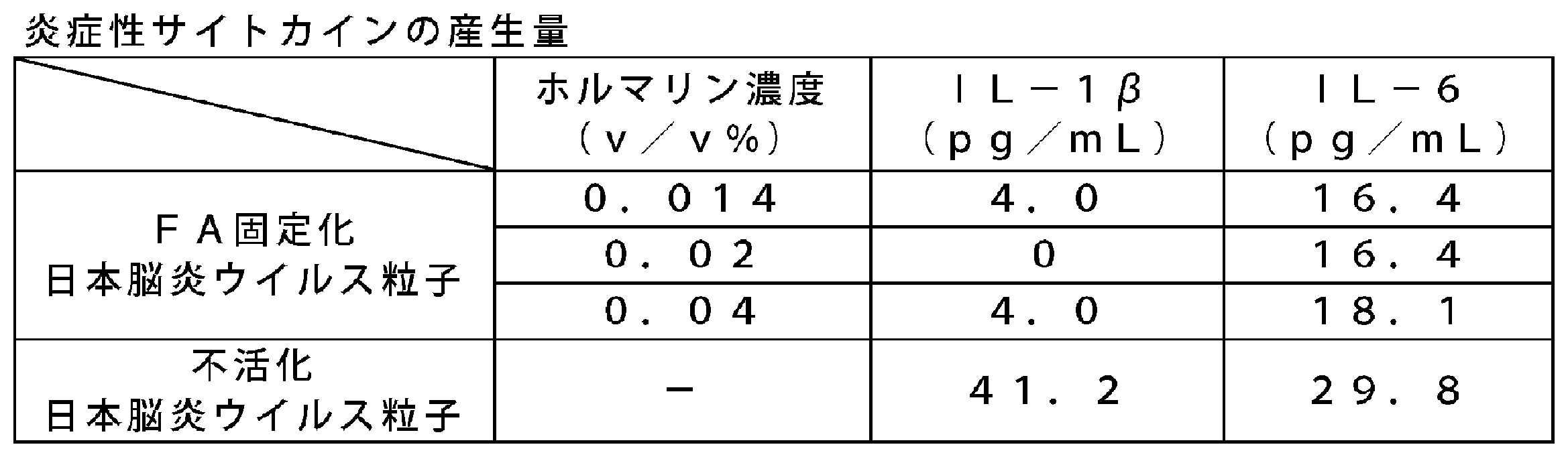
物性評価
上記実施例9で得られたFA固定化日本脳炎ウイルス粒子の物性を以下の方法で評価した。
FA固定化日本脳炎ウイルス粒子の形状を詳細に調べるために、実施例2と同様の方法で、電子顕微鏡による観察を実施した。代表として、0.014v/v%のホルマリン濃度で25℃、1週間反応後のFA固定化日本脳炎ウイルス粒子を撮影した写真を示す(図5)。FA固定化日本脳炎ウイルス粒子は、ホルマリンで固定することで不活化日本脳炎ウイルス粒子と同様に粒子構造が保たれていた。
FA固定化日本脳炎ウイルス粒子の平均粒子径を、Zetasizer Nano ZS(Malvern社製)を用いて解析した。動的光散乱法による液体中の平均粒子径を表64に示す。FA固定化日本脳炎ウイルス粒子は、平均粒子径が約90nmで単峰性であった。一方、不活化日本脳炎ウイルス粒子は、約80nmで単峰性であった。また、FA固定化日本脳炎ウイルス粒子の粒子構造は保たれており、凝集体のような不純物は認められなかった。

FA固定化日本脳炎ウイルス粒子の分子量分布を実施例8と同様の方法(SEC)で測定した。溶出パターンを表65に示す。FA固定化日本脳炎ウイルス粒子は、溶出時間14~15分付近に単峰性の主ピークが認められた。また、不活化日本脳炎ウイルス粒子も溶出時間14~15分付近に単峰性の主ピークが認められた。

抗日本脳炎ウイルス抗体を用いたサンドイッチELISA法によって、抗原の含有量(抗原含量)を実施例8と同様の方法で測定した。

実施例8と同様の方法で、代表として、0.02v/v%のホルマリン濃度で25℃、1週間反応後のFA固定化日本脳炎ウイルス粒子の中和抗体価を測定した。50%プラック減少率から算出した結果を表67に示す。FA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子と比較して同等又はそれ以上の中和抗体価であった。
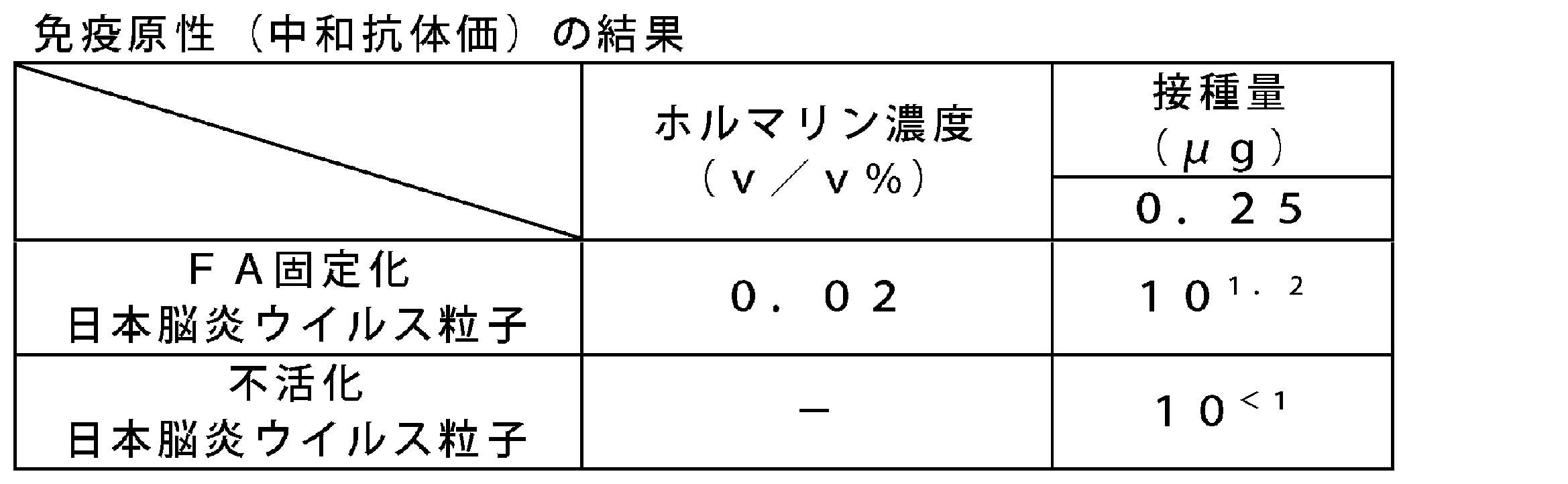
最終タンパク質濃度が8μg/mLとなるようにPBS様溶液で、FA固定化日本脳炎ウイルス粒子及び不活化日本脳炎ウイルス粒子を希釈した。抗原含量を指標に、37℃での保存安定性を評価した。代表として、0.014v/v%のホルマリン濃度で25℃、1週間反応後の結果を表68に示す。FA固定化日本脳炎ウイルス粒子は、37℃保存下では1週間維持された。一方、不活化日本脳炎ウイルス粒子は、37℃保存下では低下を示した。FA固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子と比較し安定性が向上することが示された。

EDC固定化日本脳炎ウイルス粒子の調製
1.1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド塩酸塩(EDC)処理
不活化日本脳炎ウイルス粒子に対して、最終濃度が0.15~15mMとなるようにEDCを添加し、4℃で2~20時間反応させた。クエンチング処理を行う場合には、さらにクエンチング剤としてグリシンを、EDCの8倍量(モル質量比)で反応液に添加した。反応終了後、PBS様溶液で反応液を透析し、EDC及びグリシンを除くことで、固定化日本脳炎ウイルス粒子(以下、「EDC固定化日本脳炎ウイルス粒子」と記載することがある。)を得た。得られたEDC固定化日本脳炎ウイルス粒子の発熱活性を、ヒトPBMCを刺激した場合の炎症性サイトカインの産生量の定量によって評価した。
欧州薬局方MONOCYTE-ACTIVATION TESTに準じた方法で、代表として、0.15mM又は1.5mMのEDC濃度で4℃、20時間反応後のEDC固定化日本脳炎ウイルス粒子又は不活化日本脳炎ウイルス粒子でヒトPBMCを刺激した場合のサイトカイン(IL-1β及びIL-6)の産生量を定量した。具体的には、欧州薬局方MONOCYTE-ACTIVATION TESTにおけるヒトPBMCを最低4ドナー分プールして使用するところを、1ドナー分に変更して測定した。その結果を表69に示す。EDC固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子よりも炎症性サイトカイン産生量が十分低いことがわかった。このことからも、EDC固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子よりも高い安全性を有することが示唆された。

物性評価
上記実施例11で得られたEDC固定化日本脳炎ウイルス粒子の物性を以下の方法で評価した。
EDC固定化日本脳炎ウイルス粒子の形状を詳細に調べるために、実施例2と同様の方法で、電子顕微鏡による観察を実施した。代表として、1.5mMのEDC濃度で4℃、20時間反応後のEDC固定化日本脳炎ウイルス粒子を撮影した写真を図6に示す。EDC固定化日本脳炎ウイルス粒子は、EDCで固定することで不活化日本脳炎ウイルス粒子と同様に粒子構造が保たれていた。
EDC固定化日本脳炎ウイルス粒子の平均粒子径を、Zetasizer Nano ZS(Malvern社製)を用いて解析した。動的光散乱法による液体中の平均粒子径を表70に示す。EDC固定化日本脳炎ウイルス粒子は、平均粒子径が約90nmで単峰性であった。一方、不活化日本脳炎ウイルス粒子は、約80nmで単峰性であった。また、EDC固定化日本脳炎ウイルス粒子の粒子構造は保たれており、凝集体のような不純物は認められなかった。

EDC固定化日本脳炎ウイルス粒子の分子量分布を実施例8と同様の方法で測定した。溶出パターンを表71に示す。EDC固定化日本脳炎ウイルス粒子は、溶出時間14~15分付近に単峰性の主ピークが認められた。また、不活化日本脳炎ウイルス粒子も溶出時間14~15分付近に単峰性の主ピークが認められた。

抗日本脳炎ウイルス抗体を用いたサンドイッチELISA法によって、抗原の含有量(抗原含量)を実施例8と同様の方法で測定した。代表として、1.5mMのEDC濃度で4℃、20時間反応後のEDC固定化日本脳炎ウイルス粒子、及び、不活化日本脳炎ウイルス粒子それぞれの抗原含有量を表72に示す。EDC固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子と同等の抗原が含まれていた。

実施例8と同様の方法で、EDC固定化日本脳炎ウイルス粒子の中和抗体価を測定した。50%プラック減少率から算出した結果を表73に示す。代表として、1.5mMのEDC濃度で4℃で2時間反応後にクエンチング剤としてグリシンを、EDCの8倍量(モル質量比)で反応液に添加したEDC固定化日本脳炎ウイルス粒子を評価したところ、不活化日本脳炎ウイルス粒子と比較して同等又はそれ以上の中和抗体価であった。
最終タンパク質濃度が8μg/mLとなるようにPBS様溶液で、EDC固定化日本脳炎ウイルス粒子及び不活化日本脳炎ウイルス粒子を希釈した。抗原含量を指標に、25℃、37℃での保存安定性を評価した。結果を表74、表75に示す。EDC固定化日本脳炎ウイルス粒子は、25℃保存下では1ヶ月間及び37℃保存下では1週間維持された。一方、不活化日本脳炎ウイルス粒子は、25℃保存下、37℃保存下のいずれにおいても、抗原含量の低下を示した。EDC固定化日本脳炎ウイルス粒子は、不活化日本脳炎ウイルス粒子と比較して安定性が向上することが示された。


Claims (31)
- 固定化ウイルス粒子を含むワクチンであって、
前記固定化ウイルス粒子の発熱試験におけるウサギ3羽での発熱反応の和が、前記固定化ウイルス粒子の元となるウイルス粒子又は対応する不活化ウイルス粒子におけるウサギ3羽での発熱反応の和を基準として、80%未満である、ワクチン。 - 前記固定化ウイルス粒子の発熱試験におけるウサギ3羽での発熱反応の和が、1.3℃以下である、請求項1に記載のワクチン。
- 固定化ウイルス粒子を含むワクチンであって、
前記固定化ウイルス粒子によって刺激を受けたヒト末梢血単核球から産生される炎症性サイトカインの量が、前記固定化ウイルス粒子の元となるウイルス粒子又は対応する不活化ウイルス粒子によって刺激を受けたヒト末梢血単核球から産生される炎症性サイトカインの量を基準として、80%未満である、ワクチン。 - 前記固定化ウイルス粒子の元となるウイルス粒子が、オルソミクソウイルス粒子、フラビウイルス粒子、又はピコルナウイルス粒子を含む、請求項1~3のいずれか一項に記載のワクチン。
- 前記ウイルス粒子が、インフルエンザウイルス粒子、日本脳炎ウイルス粒子、又はA型肝炎ウイルス粒子を含む、請求項4に記載のワクチン。
- 前記ウイルス粒子が、インフルエンザウイルス粒子を含む、請求項5に記載のワクチン。
- 前記インフルエンザウイルス粒子が、A型インフルエンザウイルス粒子、又はB型インフルエンザウイルス粒子を含む、請求項6に記載のワクチン。
- 前記インフルエンザウイルス粒子が、H1N1亜型株、H2N2亜型株、H3N2亜型株、H3N8亜型株、H5N1亜型株、H5N2亜型株、H5N6亜型株、H6N1亜型株、H7N3亜型株、H7N7亜型株、H7N9亜型株、H9N2亜型株、又はH10N8亜型株に分類されるインフルエンザウイルス粒子を含む、請求項6又は7に記載のワクチン。
- 前記ウイルス粒子が、日本脳炎ウイルス粒子を含む、請求項5に記載のワクチン。
- 前記日本脳炎ウイルス粒子が、北京-1株、中山株、SA14-14-2株、又はP3株を含む、請求項9に記載のワクチン。
- 前記固定化ウイルス粒子の表面のタンパク質の0%~90%が、固定化されていない、請求項1~10のいずれか一項に記載のワクチン。
- 前記固定化ウイルス粒子の元となるウイルス粒子の比活性(抗原含量/タンパク質含量)に対する、前記固定化ウイルス粒子の比活性の相対値が、0~95%である、請求項1~11のいずれか一項に記載のワクチン。
- 前記固定化ウイルス粒子が、前記固定化ウイルス粒子の元となるウイルス粒子又は対応する不活化ウイルス粒子の粒子径の80%~150%の平均粒子径を有する、請求項1~12のいずれか一項に記載のワクチン。
- 前記固定化ウイルス粒子をショ糖密度勾配遠心で測定した場合に、ショ糖濃度35%以上にピークが検出される、請求項1~13のいずれか一項に記載のワクチン。
- 前記固定化ウイルス粒子を高速液体クロマトグラフィーで測定した場合に、単峰性のピークが認められる、請求項1~14のいずれか一項に記載のワクチン。
- マウスに免疫した場合、前記固定化ウイルス粒子に特異的なIgG1よりも、前記固定化ウイルス粒子に特異的なIgG2aを誘導する、請求項1~15のいずれか一項に記載のワクチン。
- 元となるウイルス粒子又は対応する不活化ウイルス粒子を含む懸濁液に固定化剤を加える工程を含む、固定化ウイルス粒子の製造方法。
- 前記固定化剤が、アルデヒド類を含む、請求項17に記載の製造方法。
- 前記アルデヒド類が、ホルムアルデヒド、パラホルムアルデヒド、グルタルアルデヒド、及びそれらの組合せからなる群から選択される、請求項18に記載の製造方法。
- 前記アルデヒド類が、ホルムアルデヒドを含む、請求項19に記載の製造方法。
- 前記ホルムアルデヒドの濃度が、前記懸濁液及び前記固定化剤の全量を基準として、0.005~0.5w/v%である、請求項20に記載の製造方法。
- 前記アルデヒド類が、グルタルアルデヒドを含む、請求項19に記載の製造方法。
- 前記グルタルアルデヒドの濃度が、前記懸濁液及び前記固定化剤の全量を基準として、0.001~0.06w/v%である、請求項22に記載の製造方法。
- 前記固定化剤が、カルボジイミド類を含む、請求項19に記載の製造方法。
- 前記カルボジイミド類が、ジシクロヘキシルカルボジイミド、ジイソプロピルカルボジイミド、1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド塩酸塩、それらの類縁体及び、それらの組合せからなる群から選択される、請求項24に記載の製造方法。
- 前記カルボジイミド類が、1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド塩酸塩を含む、請求項25に記載の製造方法。
- 前記1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド塩酸塩の濃度が、前記懸濁液及び前記固定化剤の全量を基準として、0.05~1500mMである、請求項26に記載の製造方法。
- 元となる前記ウイルス粒子が、培養細胞、鶏卵又はマウス脳に感染させて、回収されたウイルス粒子である、請求項17~27のいずれか一項に記載の製造方法。
- 前記培養細胞が、初代細胞、又は株化細胞を含む、請求項28に記載の製造方法。
- 前記培養細胞が、Vero細胞、又はMDCK細胞を含む、請求項29に記載の製造方法。
- 請求項17~30のいずれか一項に記載の製造方法によって得られた固定化ウイルス粒子を加える工程を含む、ワクチンの製造方法。
Priority Applications (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CA3010049A CA3010049C (en) | 2016-01-15 | 2017-01-10 | Vaccine containing immobilized virus particles |
| CN201780005292.XA CN108697786B (zh) | 2016-01-15 | 2017-01-10 | 含有固定化病毒粒子的疫苗 |
| KR1020187018457A KR102405736B1 (ko) | 2016-01-15 | 2017-01-10 | 고정화 바이러스 입자를 포함하는 백신 |
| AU2017207076A AU2017207076B2 (en) | 2016-01-15 | 2017-01-10 | Vaccine containing immobilized virus particles |
| US16/066,664 US10881723B2 (en) | 2016-01-15 | 2017-01-10 | Vaccine containing immobilized virus particles |
| JP2017561109A JP6941566B2 (ja) | 2016-01-15 | 2017-01-10 | 固定化ウイルス粒子を含むワクチン |
| EP17738389.0A EP3403671B1 (en) | 2016-01-15 | 2017-01-10 | Vaccine containing immobilized virus particles |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2016006133 | 2016-01-15 | ||
| JP2016-006133 | 2016-01-15 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2017122635A1 true WO2017122635A1 (ja) | 2017-07-20 |
Family
ID=59311333
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2017/000485 Ceased WO2017122635A1 (ja) | 2016-01-15 | 2017-01-10 | 固定化ウイルス粒子を含むワクチン |
Country Status (9)
| Country | Link |
|---|---|
| US (1) | US10881723B2 (ja) |
| EP (1) | EP3403671B1 (ja) |
| JP (1) | JP6941566B2 (ja) |
| KR (1) | KR102405736B1 (ja) |
| CN (1) | CN108697786B (ja) |
| AU (1) | AU2017207076B2 (ja) |
| CA (1) | CA3010049C (ja) |
| TW (2) | TWI781088B (ja) |
| WO (1) | WO2017122635A1 (ja) |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2019050772A (ja) * | 2017-09-15 | 2019-04-04 | 国立大学法人 熊本大学 | ワクチン接種後副反応を予測する検査方法 |
| JP2020065459A (ja) * | 2018-10-22 | 2020-04-30 | キヤノンメディカルシステムズ株式会社 | ウイルスrna単離方法及びウイルス定量方法 |
| WO2024019113A1 (ja) | 2022-07-21 | 2024-01-25 | デンカ株式会社 | 不活化全粒子インフルエンザワクチンの発熱活性低減方法 |
| JP2024097058A (ja) * | 2020-07-17 | 2024-07-17 | 富士フイルム株式会社 | 日本脳炎予防剤及び日本脳炎ワクチン剤 |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN111018970B (zh) * | 2019-12-27 | 2021-09-14 | 中牧实业股份有限公司 | 一种猪乙型脑炎病毒特异性阳性血清及其制备方法 |
| CN113755480B (zh) * | 2021-03-19 | 2023-12-05 | 威海纽普生物技术有限公司 | 病毒固相感染体外培养细胞的方法 |
| CN113025653B (zh) | 2021-05-24 | 2021-09-24 | 华南农业大学 | 全禽源遗传系统及其在制备h7n9禽流感疫苗中的应用 |
Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2001076624A1 (en) * | 2000-04-07 | 2001-10-18 | Juridical Foundation The Chemo-Sero-Therapeutic Research Institute | Inactivated japanese b encephalitis vaccine and process for producing the same |
| JP2002528422A (ja) * | 1998-10-28 | 2002-09-03 | ゾンターク ハンス−ギュンター | 抗ウイルス剤の製造方法 |
| JP2003517834A (ja) * | 1999-12-20 | 2003-06-03 | アグリカルチュラル・リサーチ・カウンセル | 微小有機体の不活化方法 |
| JP2010514817A (ja) * | 2007-01-04 | 2010-05-06 | グラクソスミスクライン バイオロジカルズ ソシエテ アノニム | ワクチンの製造方法 |
| CN104498446A (zh) * | 2014-12-15 | 2015-04-08 | 北京科兴生物制品有限公司 | 一种在疫苗生产中灭活病毒的方法 |
Family Cites Families (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN1122531C (zh) * | 1995-08-01 | 2003-10-01 | 巴斯德梅诺血清和疫苗公司 | 抗日本脑炎疫苗的工业生产方法和所获得的疫苗 |
| DK1025209T3 (da) | 1997-08-28 | 2007-06-25 | Cheil Jedang Corp | Et attenueret japansk encephalitis-virus tilpasset til Vero-celle og en japansk encephalitis-vaccine |
| WO2008073490A1 (en) * | 2006-12-12 | 2008-06-19 | Carrington Laboratories Inc., | Purification of influenza viral antigens |
| WO2009106629A1 (en) * | 2008-02-29 | 2009-09-03 | Universiteit Gent | Viral inactivation process |
| US9956281B2 (en) * | 2011-05-04 | 2018-05-01 | Streck, Inc. | Inactivated virus compositions and methods of preparing such compositions |
| EP2906239A1 (en) * | 2012-10-12 | 2015-08-19 | GlaxoSmithKline Biologicals SA | Non-cross-linked acellular pertussis antigens for use in combination vaccines |
| WO2016010081A1 (ja) * | 2014-07-18 | 2016-01-21 | 一般財団法人化学及血清療法研究所 | ウイルス様粒子を含むワクチン |
| CN105056225B (zh) * | 2015-07-30 | 2018-01-16 | 成都天邦生物制品有限公司 | 一种灭活疫苗的制备方法 |
-
2017
- 2017-01-10 CA CA3010049A patent/CA3010049C/en active Active
- 2017-01-10 AU AU2017207076A patent/AU2017207076B2/en active Active
- 2017-01-10 WO PCT/JP2017/000485 patent/WO2017122635A1/ja not_active Ceased
- 2017-01-10 US US16/066,664 patent/US10881723B2/en active Active
- 2017-01-10 JP JP2017561109A patent/JP6941566B2/ja active Active
- 2017-01-10 CN CN201780005292.XA patent/CN108697786B/zh active Active
- 2017-01-10 EP EP17738389.0A patent/EP3403671B1/en active Active
- 2017-01-10 KR KR1020187018457A patent/KR102405736B1/ko active Active
- 2017-01-13 TW TW106101131A patent/TWI781088B/zh active
- 2017-01-13 TW TW110127455A patent/TWI781700B/zh active
Patent Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2002528422A (ja) * | 1998-10-28 | 2002-09-03 | ゾンターク ハンス−ギュンター | 抗ウイルス剤の製造方法 |
| JP2003517834A (ja) * | 1999-12-20 | 2003-06-03 | アグリカルチュラル・リサーチ・カウンセル | 微小有機体の不活化方法 |
| WO2001076624A1 (en) * | 2000-04-07 | 2001-10-18 | Juridical Foundation The Chemo-Sero-Therapeutic Research Institute | Inactivated japanese b encephalitis vaccine and process for producing the same |
| JP2010514817A (ja) * | 2007-01-04 | 2010-05-06 | グラクソスミスクライン バイオロジカルズ ソシエテ アノニム | ワクチンの製造方法 |
| CN104498446A (zh) * | 2014-12-15 | 2015-04-08 | 北京科兴生物制品有限公司 | 一种在疫苗生产中灭活病毒的方法 |
Non-Patent Citations (2)
| Title |
|---|
| DELRUE, I. ET AL.: "Inactivated virus vaccines from chemistry to prophylaxis: Merits, risks and challenges", EXPERT REVIEW OF VACCINES, vol. 11, no. 6, 2012, pages 695 - 719, XP055399927 * |
| See also references of EP3403671A4 * |
Cited By (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2019050772A (ja) * | 2017-09-15 | 2019-04-04 | 国立大学法人 熊本大学 | ワクチン接種後副反応を予測する検査方法 |
| JP2020065459A (ja) * | 2018-10-22 | 2020-04-30 | キヤノンメディカルシステムズ株式会社 | ウイルスrna単離方法及びウイルス定量方法 |
| JP7233880B2 (ja) | 2018-10-22 | 2023-03-07 | キヤノンメディカルシステムズ株式会社 | ウイルスrna単離方法及びウイルス定量方法 |
| JP2024097058A (ja) * | 2020-07-17 | 2024-07-17 | 富士フイルム株式会社 | 日本脳炎予防剤及び日本脳炎ワクチン剤 |
| WO2024019113A1 (ja) | 2022-07-21 | 2024-01-25 | デンカ株式会社 | 不活化全粒子インフルエンザワクチンの発熱活性低減方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| US10881723B2 (en) | 2021-01-05 |
| JPWO2017122635A1 (ja) | 2018-11-08 |
| JP6941566B2 (ja) | 2021-09-29 |
| EP3403671B1 (en) | 2023-06-07 |
| EP3403671C0 (en) | 2023-06-07 |
| CN108697786A (zh) | 2018-10-23 |
| AU2017207076B2 (en) | 2021-04-29 |
| TWI781088B (zh) | 2022-10-21 |
| US20190000960A1 (en) | 2019-01-03 |
| TW201733616A (zh) | 2017-10-01 |
| EP3403671A4 (en) | 2020-04-22 |
| CA3010049C (en) | 2021-10-12 |
| TWI781700B (zh) | 2022-10-21 |
| TW202144003A (zh) | 2021-12-01 |
| KR102405736B1 (ko) | 2022-06-07 |
| CA3010049A1 (en) | 2017-07-20 |
| EP3403671A1 (en) | 2018-11-21 |
| KR20180101354A (ko) | 2018-09-12 |
| CN108697786B (zh) | 2022-05-27 |
| AU2017207076A1 (en) | 2018-07-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6941566B2 (ja) | 固定化ウイルス粒子を含むワクチン | |
| KR20070116650A (ko) | 조성물 | |
| US10980871B2 (en) | Vaccine compositions | |
| US20200093915A1 (en) | Vaccine Containing Virus-Like Particles | |
| JP5918870B2 (ja) | インフルエンザに対する改善されたワクチン接種 | |
| JP2008519042A5 (ja) | ||
| JP7770301B2 (ja) | 不活化インフルエンザワクチンの製造方法及びそのワクチン組成物 | |
| JP2022536120A (ja) | インフルエンザウイルスバックボーン | |
| JPWO2008041710A1 (ja) | 沈降不活化インフルエンザワクチンおよびその製造方法 | |
| Geeraedts et al. | Influenza vaccines: what do we want and how can we get it? | |
| JP2016094359A (ja) | ワクチン及び感染防御キット |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 17738389 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2017561109 Country of ref document: JP Kind code of ref document: A |
|
| ENP | Entry into the national phase |
Ref document number: 3010049 Country of ref document: CA |
|
| ENP | Entry into the national phase |
Ref document number: 20187018457 Country of ref document: KR Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2017207076 Country of ref document: AU Date of ref document: 20170110 Kind code of ref document: A |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2017738389 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: 2017738389 Country of ref document: EP Effective date: 20180816 |