WO2017167255A1 - 一种3-差向异构酶以及编码它的多核苷酸 - Google Patents
一种3-差向异构酶以及编码它的多核苷酸 Download PDFInfo
- Publication number
- WO2017167255A1 WO2017167255A1 PCT/CN2017/078923 CN2017078923W WO2017167255A1 WO 2017167255 A1 WO2017167255 A1 WO 2017167255A1 CN 2017078923 W CN2017078923 W CN 2017078923W WO 2017167255 A1 WO2017167255 A1 WO 2017167255A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- polypeptide
- protein
- epimerase
- tagatose
- psicose
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images


Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/24—Preparation of compounds containing saccharide radicals produced by the action of an isomerase, e.g. fructose
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L27/00—Spices; Flavouring agents or condiments; Artificial sweetening agents; Table salts; Dietetic salt substitutes; Preparation or treatment thereof
- A23L27/30—Artificial sweetening agents
- A23L27/33—Artificial sweetening agents containing sugars or derivatives
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/90—Isomerases (5.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/02—Monosaccharides
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y503/00—Intramolecular oxidoreductases (5.3)
- C12Y503/01—Intramolecular oxidoreductases (5.3) interconverting aldoses and ketoses (5.3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y501/00—Racemaces and epimerases (5.1)
- C12Y501/03—Racemaces and epimerases (5.1) acting on carbohydrates and derivatives (5.1.3)
- C12Y501/03001—Ribulose-phosphate 3-epimerase (5.1.3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y501/00—Racemaces and epimerases (5.1)
- C12Y501/03—Racemaces and epimerases (5.1) acting on carbohydrates and derivatives (5.1.3)
- C12Y501/03022—L-Ribulose-5-phosphate 3-epimerase (5.1.3.22)
Definitions
- the present invention relates to a polypeptide or protein having 3-epimerase activity, a polynucleotide sequence encoding a polypeptide or protein having 3-epimerase activity, a nucleic acid construct or expression comprising the polynucleotide sequence A carrier, a method of producing the enzyme, and the use of the enzyme in various industrial applications.
- the wood invention relates to a method for producing D-psicose by using a polypeptide or protein having 3-epimerase activity to catalyze a monosaccharide or a mixed sugar such as fructose, glucose, or starch sugar, and using the polypeptide or protein A method for producing D-tagatose by catalyzing a monosaccharide or a mixed sugar such as sorbose or starch sugar.
- D-psicose and D-tagatose have become natural hotspots for natural low-calorie functional sweeteners that are naturally occurring in nature but have little content.
- D-allulose also known as D-ribose-2-hexulose
- D-psicose is the epimer of the C-3 position of D-fructose.
- D-psicose is a naturally occurring but very low-calorie functional sweetener with a sweetness of 70% of sucrose but only 0.3% of sucrose, which can be used as a low-calorie diet food.
- the sweetener at the same time, D-psicose also has the function of inhibiting the activity of enzymes related to lipid synthesis in the liver, helping to reduce abdominal fat accumulation and controlling body weight to some extent, and can be used for health foods and the like.
- a variety of functional foods can also improve the flavor, appearance, etc. of food, and extend the shelf life of food. Therefore, D-psicose, a healthy and safe low-calorie functional sweetener, has attracted more and more scholar' attention and become one of the most competitive new sweeteners in the market.
- D-psicose Since D-psicose is a relatively rare natural monosaccharide, it has low yield and high cost from the natural environment, and it is difficult to meet the needs of people as low-calorie healthy sweeteners, and is not suitable for industrialized large-scale production. Therefore, in order to be applied to the food industry, there is a need for an efficient D-psicose production process.
- the traditional D-psicose production process is mainly chemical, which requires a lot of cost, many by-products and pollutants.
- the preparation of D-psicose by biological method has the characteristics of strong reaction specificity, simple and easy purification of product components and natural products, which has become a research hotspot.
- the most effective way to prepare D-psicose by biological methods is to find an enzyme that converts fructose into D-psicose.
- the current psicose-3-differential isomerase enzyme has problems such as reduced stability at high temperature or low activity and low reaction rate, which is not conducive to the cost control of industrial production of D-psicose. Therefore, there is a need to develop a psicose-3-epimerase having excellent high temperature stability and high activity to meet the demand for industrial production.
- D-Tagatose is the epimer of the C-3 position of D-sorbose.
- D-tagatose is a natural deposit
- the low-calorie functional sweetener with a very low content has a sweetness of 92% of sucrose and a low absorption rate of only 20% to 25% in the human body, which does not cause significant changes in the blood sugar level of the body. It is very suitable for diabetics, and most of the tagatose directly enters the colon, which is selectively fermented by the microbial flora, promotes the proliferation of beneficial bacteria, inhibits the growth of harmful bacteria, and plays a significant role in improving the intestinal flora. It is a good probiotic and can be used in a variety of functional foods such as health foods.
- D-tagatose can also improve food flavor, appearance and the like. Therefore, D-tagatose, a healthy and safe low-calorie functional sweetener, has attracted more and more scholars' attention and become one of the most promising new sweeteners.
- Isomerization of sorbose to D-tagatose is one of the most effective methods for the preparation of D-tagatose by biological methods.
- the key to this method is to find an enzyme that converts sorbose into D-tagatose.
- the current tagatose-3-differential isomerase has problems such as reduced stability at high temperature or low activity and low reaction rate, which is not conducive to cost control and scale-up of industrial production of D-tagatose. Therefore, it is necessary to develop a tagatose-3-diffusion isomerase having excellent high temperature stability and high activity to meet the demand for industrial production.
- the present inventors discovered a novel protein having 3-epimerase activity from the Thermomemmatispora carboxidivorans strain. Accordingly, the present invention provides novel proteins having 3-epimerase activity, and polynucleotides encoding the proteins or polypeptides.
- the protein or polypeptide has psicose-3-epimerase and tagatose-3-epimerase activity.
- the protein or polypeptide of the present invention has good activity and excellent thermal stability, both of which are excellent properties for producing D-psicose and D-tagatose at higher temperature conditions.
- the present invention relates to an isolated protein or polypeptide having psicose-3-epimerase and tagatose-3-epimerase activity selected from the group consisting of
- the invention also relates to nucleic acid constructs, expression vectors and recombinant host cells comprising the polynucleotides: A method of producing a polynucleotide.
- the invention also relates to a process for the enzymatic production of D-psicose and D-tagatose.
- Ipsicose-3-epimerase activity means catalyzing the conversion of the C-3 position of fructose to D-psicose active.
- Tagatose-3-epimerase activity means the activity of catalyzing the conversion of the C-3 position of sorbose to D-tagatose.
- Isolated protein or polypeptide means a polypeptide that has been human modified with respect to a protein or polypeptide as found in nature. In one aspect, the polypeptide is at least 1% pure, as determined by SDS-PAGE,
- Non-limiting examples of isolated proteins or polypeptides include: (1) any non-naturally occurring protein or polypeptide, (2) any protein or polypeptide including, but not limited to, any enzyme, variant, nucleic acid, protein, peptide or accessory. Factors that are at least partially removed from one or more or all of the naturally occurring components that are associated with them in nature; (3) any protein or polypeptide that has been artificially modified relative to a protein or polypeptide found in nature: or (4) Any protein or polypeptide that is modified by increasing the amount of the protein or polypeptide relative to other components with which it is naturally associated.
- An isolated protein or polypeptide can be present in the fermentation broth sample.
- substantially pure protein or polypeptide means a protein or polypeptide preparation containing up to 10% by weight of its natural or recombinant association. Other protein or polypeptide materials. Preferably, the protein or polypeptide is at least 90% pure by weight of all protein or polypeptide material present in the preparation.
- the protein or polypeptide of the invention is preferably in a substantially pure form, for example, this can be achieved by preparing the polypeptide by well known recombinant methods or by classical purification methods.
- Sequence identity The parameter "sequence identity" describes the correlation between two amino acid sequences or between two nucleotide sequences.
- the degree of sequence identity between two amino acid sequences is achieved by using, for example, the EMBOSS software package (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al, 2000, Trends in Genetics 16: 276-277) (preferred)
- EMBOSS software package EMBOSS: The European Molecular Biology Open Software Suite, Rice et al, 2000, Trends in Genetics 16: 276-277
- the Needleman-Wunsch algorithm (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453) performed in the Needle program of version 3.0.0 or higher) was determined.
- the optional parameters used were a gap open penalty of 10, a gap extension penalty of 0.5 and an EBLOSUM62 (EMBOSS version of BLOSUM62) substitution matrix.
- Use Needle to mark "highest identity” "The output (obtained with the -nobrief option) as a percentage of identity and calculated as follows:
- the degree of sequence identity between two nucleotide sequences is achieved by using, for example, the EMBOSS software package (EMBOSS: The European Molecular Bi010 Open Software Suite, Rice et al, supra) (preferably version 3.0.0 or more).
- EMBOSS European Molecular Bi010 Open Software Suite, Rice et al, supra
- the Needleman-Wunsch algorithm (Needleman and Wunsch, 1970, supra), performed in the higher version of the Needle program, was determined.
- the optional parameters used were gap open penalty of 10, gap extension penalty of 0.5 and EDNAFULL (EMBOSS version of NCBI NUC4.4) substitution matrix.
- fragment means a polypeptide having one or more (several) amino acids deleted from the amino and/or carboxyl terminus of a mature polypeptide; wherein the fragment has 3-epimerase activity.
- allelic variant means any two or more alternative forms of a gene occupying the same chromosomal locus. Allelic variation occurs naturally by mutation and can result in polymorphism within the population. A genetic mutation can be silent (no change in the encoded polypeptide) or can encode a polypeptide having an altered amino acid sequence.
- An allelic variant of a polypeptide is a polypeptide encoded by an allelic variant of the gene.
- Isolated polynucleotide means a human modified polynucleotide relative to a polynucleotide found in nature. In one aspect, the isolated polynucleotide is 1% to 95% pure as determined by agarose electrophoresis.
- the polynucleotide can be of genomic, cDNA, RNA, semi-synthetic, synthetic origin, or any combination thereof.
- a pure polynucleotide means a polynucleotide preparation free of other foreign or undesired nucleotides, and the polynucleotide preparation is suitable for A form used in genetically engineered protein production systems.
- a substantially pure polynucleotide contains from 0.5% to 10% by weight of other polynucleotide material with which it is naturally or recombinantly bound.
- substantially pure polynucleotides can include naturally occurring 5' and 3' untranslated regions, such as promoters and terminators.
- the substantially pure polynucleotide is 90% to 99.5% pure by weight.
- the polynucleotide of the present invention is preferably in a pure form on the basis of wood.
- Coding sequence means a polynucleotide that directly specifies the amino acid sequence of a polypeptide.
- the boundaries of the coding sequence are usually determined by an open reading frame, usually starting with the ATG start codon or alternative start codons such as GTG and TTG, and ending with stop codons such as TAA, TAG and TGA.
- the coding sequence can be a DNA, cDNA, synthetic or recombinant polynucleotide.
- cDNA means a DNA molecule capable of being prepared by reverse transcription from a mature, spliced mRNA molecule derived from a eukaryotic cell.
- the cDNA lacks an intron sequence that is normally present in the corresponding genomic DNA.
- the beginning (initial), a primary RNA transcript is a precursor to mRNA that is processed through a series of steps, including splicing, and then appears as mature, spliced mRNA.
- nucleic acid construct means a single-stranded or double-stranded nucleic acid molecule which is isolated from a naturally occurring gene or which is modified to contain a nucleic acid in a manner not inherent in nature. Section, or it is synthetic.
- nucleic acid construct is synonymous with the term “expression cassette” when the nucleic acid construct contains the regulatory sequences required for expression of a coding sequence of the invention.
- Control sequence means all components necessary for expression of a polynucleotide encoding a polypeptide of the present invention.
- Each control sequence may be native or foreign to the nucleotide sequence encoding the polypeptide, or the individual regulatory sequences may be native or foreign to each other.
- These regulatory sequences include, but are not limited to, a leader sequence, a polyadenylation sequence, a propeptide sequence, a promoter, a signal peptide sequence, and a transcription terminator.
- regulatory sequences include promoters as well as termination signals for transcription and translation.
- the control sequences can be provided with a linker for the purpose of introducing a specific restriction site that facilitates ligation of the regulatory sequence to the coding region of the polynucleotide encoding the polypeptide.
- operably linked means a configuration in which a regulatory sequence is placed at an appropriate position relative to the coding sequence of a polynucleotide such that the regulatory sequence directs expression of the coding sequence.
- expression includes any step involved in the production of a polypeptide including, but not limited to, transcription, post-transcriptional modification, translation, post-translational modification, and secretion.
- Expression vector means a linear or circular DNA molecule comprising a polynucleotide encoding a polypeptide, and the polynucleotide is operably linked to additional nucleotides provided for its expression.
- host cell means any cell type that is susceptible to transformation, transfection, transduction, etc., using a nucleic acid construct or expression vector comprising a polynucleotide of the present invention.
- host cell encompasses the progeny of any parental cell that differs from the parental cell due to mutations that occur during replication.
- variant means a protein or polypeptide having 3-epimerase activity that comprises an alteration at one or more (several) positions, ie one or more (several) amino acid residues Substitutions, insertions and/or deletions. Substitution means replacing an amino acid occupying a position with a different amino acid: deletion means removal of an amino acid occupying a position; and insertion means addition of 1-3 amino acids to an amino acid adjacent to a position.
- the present inventors studied the characteristics of an epimerase polypeptide derived from Thermomemmatispora carboxidivorans, and found that when converting fructose into D-psicose and converting sorbose into D-tagatose, The above polypeptide has a function of epimerization reaction of fructose and sorbose at the third carbon position to convert it into D-psicose or D-tagatose, which is carried out by the following steps: synthesis source Epimerase group from Thermomemmatispora carboxidivorans Therefore, the microorganism containing the gene expression vector is cultured and the polypeptide is overexpressed.
- the polypeptide was found to have good psicose-3-epimerase and tagatose-3-epimerase activity and excellent thermal stability. Accordingly, the present invention provides a polypeptide having psicose-3-epimerase and tagatose-3-epimerase activity, and using the aldose--3-difference A method of producing D-psicose or D-tagatose to a polypeptide of isomerase and tagatose-3-epimerase activity.
- the gene labeled as a polypeptide of the epimerase is obtained from the Thermomemmatispora carboxidivorans strain by gene synthesis, and the labeled epimerase gene is It is based only on the DNA base sequence, not on the results of its functional characterization.
- the obtained epimerase gene is then inserted into a suitable expression vector to produce a recombinant vector containing the epimerase gene, and the recombinant vector is transformed into a suitable microorganism.
- the transformed microorganism is cultured in a fermentation medium, and the polypeptide product of the epimerase gene is overexpressed in the microorganism, and then the polypeptide product of the epimerase gene is isolated and purified for use.
- the polypeptide has psicose-3-epimerase and tagatose-3-epimerase activity, which can transform fructose to produce D-psicose, and can also transform sorbose. D-tagatose.
- novel 3-epimerase-active polypeptide produced by the method of the present invention may have an amino acid sequence which is not limited to the amino acid sequence of SEQ ID No: 2 but includes the amino acid sequence of SEQ ID No: 2.
- the expression vector which can be used to produce a recombinant expression vector can be any expression vector conventionally used in genetic recombination techniques, and can be, for example, pET-22b(+).
- the microorganism capable of being transformed by the recombinant expression vector may be Escherichia coli BL21 (DE3).
- the microorganism is not limited as long as it is any microorganism capable of overexpressing the gene after being transformed with a recombinant expression vector containing the desired gene, and capable of producing an active protein as a result of overexpression. Or a polypeptide.
- the following process of culturing the transformed microorganism and inducing overexpression of the protein or polypeptide of the present invention can be carried out according to an exemplary experimental protocol of the present invention as described below.
- the cryopreserved recombinant Escherichia coli was inoculated into a 250 mL flask containing 50 mL of LB medium, and the strain was cultured in a shaker maintained at 37 ° C until the absorbance at 600 nm reached 2.0.
- the culture solution was added to a 7 L fermentor containing 5 L of fermentation broth, 25 g/L peptone, 25 g/L yeast extract, 10 g/L sodium chloride, 2 g/L glucose, 3 g.
- the stirring rate is 500 rpm
- the aeration rate is 1.0 vvm
- the culture temperature is 37 ° C, and the above culture conditions are favorable for mass production. 3-epimerase.
- the recombinant E. coli culture solution was centrifuged at 6,000 x g for 30 minutes at 4 ° C and then washed twice with 0.85% NaCl. Subsequently, the cells were resuspended in 50 mM sodium phosphate buffer solution (containing 300 mM NaCl) of pH 8.0, and the buffer solution containing the cells was placed in an ice bath for 30 minutes.
- the cells in the buffer solution were disrupted by a high-pressure homogenizer, and the disrupted cells were centrifuged at 13,000 ⁇ g, 4 ° C for 20 minutes, and removed, and the supernatant was filtered through a filter having a pore size of 0.45 ⁇ m and passed through It is purified by rapid protein chromatography under low temperature conditions.
- the filtrate containing the protein of the present invention was added to a HisTrap HP column equilibrated with 50 mM sodium phosphate buffer solution of pH 8.0 containing 300 mM NaCl and 10 mM imidazole.
- the HisTrap HP column was washed with the same sodium phosphate buffer solution, and the protein attached to the column was eluted with the same sodium phosphate buffer solution containing imidazole having a concentration gradient of from 10 mM to 200 mM at a flow rate of 1 mL/min.
- the eluate containing the protein of the present invention was added to a HiPrep 16/60 resin column which was equilibrated with a 50 mM sodium phosphate buffer solution of pH 7.5 and further at 6 mL/min. The rate of elution of the protein.
- the protein solution thus collected was added to a Sephacryl S-100 HR column which was equilibrated with a pH 7.5 50 mM sodium phosphate buffer solution containing 0.15 M NaCl at a flow rate of At 6 mL/min, the eluted white matter was finally dialyzed against a 50 mM sodium phosphate buffer solution.
- the protein of the present invention obtained as described above is a 3-epimerase, and the monomeric molecular weight of the 3-epimerase is 31,770 Da.
- the 3-epimerase is a metalloenzyme, and the metal ion significantly promotes its activity.
- the 3-epimerase is reacted in the presence of a metal ion to achieve the purpose of increasing the production of D-psicose from fructose, and also to improve the production of D- from sorbose.
- the metal ion is selected from the group consisting of manganese, magnesium and cobalt in a concentration ranging from 0.5 to 5 mM, for example, 1 mM. When the concentration of the metal ions is less than 0.5 mM, the effect of increasing the conversion is not remarkable, and when the concentration of the metal ions is higher than 5 mM, there is no significant difference in the conversion rate.
- the reaction between the 3-epimerase and fructose or sorbose can utilize a substrate having a concentration of 10-75% (w/w), pH 6-8 and a temperature of 50-90 ° C (ie fructose or sorbose solution). )get on.
- concentration of the substrate, fructose or sorbose is in the range of 10-75% (w/w)
- the yield of D-psicose or D-tagatose is good
- the conversion rate is high
- the pH and temperature conditions are the pH and temperature ranges that are optimal for 3-epimerase activity.
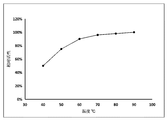
- the 3-epimerase has excellent thermal stability. After the incubation at 60 ° C for 12 hours, the activity is still not detected. After incubation at 80 ° C for 12 hours, the activity remains above 80%, and the temperature is maintained at 90 ° C. There is still 50% residual activity after 8 hours.
- the excellent thermal stability of the 3-phase isomerase is excellent for producing D-psicose and D-tagatose under higher temperature conditions.
- the 3-epimerase conversion of fructose to produce D-psicose and the conversion of sorbose to produce D-tagatose can be achieved by 3-epimer isomerization during the reaction.
- the enzyme immobilization is carried out on a carrier because the 3-epimerase immobilized on the carrier is capable of maintaining enzyme activity for a long period of time and is easy to reuse.
- the carrier used in the present embodiment of the present invention may be any one known for its use in enzyme immobilization, and may be, for example, sodium alginate.
- sodium alginate is a natural colloidal polysaccharide which is abundant in algal cell walls and contains ⁇ -D-mannuronic acid and ⁇ -L-guluronic acid residues, said ⁇ -D-mannuronic acid And the ⁇ -L-guluronic acid residue is randomly linked by a ⁇ -1,4 bond. Therefore, sodium alginate allows for stable immobilization of the 3-epimerase and is advantageous for obtaining higher D-psicose or D-tagatose production.
- sodium alginate can be used for the immobilization of 3-epimerase at a concentration of 1.5-4% (w/v), for example at a concentration of 2.5% (w/v).
- the solution of the 3-epimerase is added to an aqueous solution of sodium alginate, and the volume of the aqueous solution of sodium alginate is 3 - one or two times the volume of the epimerase solution, and then the mixture is added dropwise to the 0.2 M calcium ion solution by means of a syringe pump and a vacuum pump to form a 3-epimerase-sodium alginate complex.
- a syringe pump and a vacuum pump to form a 3-epimerase-sodium alginate complex.
- These 3-epimerase-alginic acid complex spheres can be used in the conversion of fructose to produce D-psicose and the conversion of sorbose to produce D-tagatose.
- the 3-epimerase of the present invention has good activity and excellent thermal stability for fructose and sorbose, both of which produce D-psicose and D-tagatose under higher temperature conditions. Words are excellent performance.
- the method for producing D-psicose and D-tagatose according to an embodiment of the present invention is environmentally friendly because an enzyme derived from microorganisms is used, and the method requires a simple enzyme immobilization process, and Significantly increased production yield and production efficiency of D-psicose and D-tagatose, thereby reducing production costs and maximizing production.
- the D-psicose and D-tagatose thus produced can be effectively used as an additive for foods or medicines.
- the 3-epimerase of the present invention has good activity and excellent thermal stability for fructose and sorbose, both of which are for producing D-psicose and D-tagatose under high temperature conditions. It is excellent performance.
- the method for producing D-psicose and D-tagatose according to an embodiment of the present invention is environmentally friendly because an enzyme derived from microorganisms is used, and the method requires a simple enzyme immobilization process, and Significantly increased production yield and production efficiency of D-psicose and D-tagatose, thereby reducing production costs and maximizing production.
- the D-psicose and D-tagatose thus produced can be effectively used as an additive for foods or medicines.
- Figure 1 is a flow chart showing the production of 3-epimerase in Example 1 of the present invention.
- Figure 2-1 is a graph showing the effect of pH on the activity of 3-epimerase in Example 5 of the present invention.
- Example 5 of the present invention is a graph showing the effect of temperature on the activity of 3-epimerase in Example 5 of the present invention.
- Figure 3 is a graph showing the relationship of temperature activity in Example 6 of the present invention.
- Figure 4 is a schematic diagram showing the conversion of D-psicose produced by the conversion of fructose by 3-epimerase in Example 7 of the present invention
- Figure 5 is a graph showing the conversion of D-tagatose produced by the conversion of sorbose by 3-epimerase in Example 11 of the present invention.
- the enzymatic activity was measured using fructose and sorbose as substrates.
- 3-epimerase was mixed with 50 mM pH 7.5 sodium phosphate buffer solution containing 10% fructose or sorbose, reacted at 60 ° C for 20 minutes, and then heated at 100 ° C. 15 minutes to terminate the reaction.
- the sodium phosphate buffer solution containing fructose or sorbose is prepared by dissolving fructose or sorbose in a sodium phosphate buffer solution of pH 7-8 to achieve a concentration of 60-70% (w/v), and The sodium phosphate buffer solution containing fructose or sorbose was continuously added to the bioreactor maintained at 60 °C.
- one unit of psicose-3-epimerase is defined as required to produce 1 mole of D-psicose per minute at pH 7.5 and 60 °C.
- Amount of psicose-3-epimerase; one unit of tagatose-3-epimerase is defined as producing 1 mole of D-tower per minute at pH 7.5 and 60 °C
- the amount of tagatose-3-epimerase required for the sugar was measured by high performance liquid chromatography using a BP-100 calcium ion hydrocarbon column and an RI detector. The column temperature was 80 ° C, the mobile phase was ultrapure water, and the flow rate was 0.5 mL/min.
- the 3-epimerase gene is obtained by synthesizing a gene of a polypeptide labeled as a epimerase in Thermogemmatispora carboxidivorans, which is based only on the sequence, not on the functional aspects thereof. Defined.
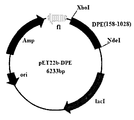
- the obtained epimerase gene was inserted into the expression vector pET-22b(+) by using restriction enzymes NdeI and XhoI, thereby producing a recombinant expression vector pET-22b(+)/epimerase ( see picture 1).
- the recombinant vector was transformed into E. coli BL21 (DE3) by a conventional transformation method.
- the transformed recombinant Escherichia coli BL21 (DE3) was stored in an ultra-low temperature freezer at -80 °C.
- the recombinant Escherichia coli was inoculated into a 250 mL Erlenmeyer flask containing 50 mL of liquid LB medium, and culture-activated in a shaker at 37 ° C until the absorbance of the culture solution at a wavelength of 600 nm reached 2.0.
- This culture solution was added to a 7 L fermentor containing 5 L of fermentation medium, and subjected to fermentation culture to mass-produce 3-epimerase.
- the stirring rate was maintained at 500 rpm, the ventilation rate was 1.0 vvm, and the culture temperature was 37 °C.
- the molecular weight of the purified 3-epimerase was measured, and it was found that the 3-epimerase monomer had a molecular weight of 31,770 Da.
- the amino acid sequence of the 3-epimerase was confirmed to be identical to the amino acid sequence of NCBI Accession No. WP_052889376.
- Example 3 in order to investigate the effect of metal ions on the 3-epimerase, the effect of fructose as a substrate on the activity of the substrate was determined in the presence of different metal ions, and the measurement was performed by EDTA. After the epimerization enzyme, 1 mM of various metal ions shown in Table 1 below was added to the 3-epimerase solution. The 3-epimerase reaction was carried out in a 50 mM Tris buffer solution of pH 7.5 at 60 ° C for 20 minutes, wherein the Tris buffer solution contained 0.04 U/mL of 3-epimerase and 10% ( The fructose of w/v) was further heated at 100 ° C for 15 minutes to terminate the reaction, and then the activity of the 3-epimerase was measured.
- the 3-epimerase reaction was carried out in a 50 mM sodium phosphate buffer solution at pH 7.5 and 60 ° C for 20 minutes, wherein the sodium phosphate buffer solution contained 0.04 U/ml of the 3-epimer
- Example 5 in order to investigate the effects of different pH and temperature on the activity of the 3-epimerase, the effect of fructose as a substrate on its activity was measured under different conditions of temperature and pH, and the differences were compared. Enzyme activity at temperature and pH.
- the 3-epimerase reaction was carried out in a sodium phosphate buffer solution having a pH range of 6.0-8.5 50 mM, wherein the sodium phosphate buffer solution contained 0.04 U/mL of the 3-difference Enzyme and 10% (w/v) fructose.
- the respective reactions were carried out at 60 ° C for 20 minutes without metal ion addition, followed by heating at 100 ° C for 15 minutes to terminate the reaction, and the enzyme activity was measured.
- the result is illustrated in Figure 2-1.
- the reaction was carried out in a 50 mM sodium phosphate buffer solution at a temperature ranging from 40 to 90 ° C for 20 minutes, wherein the sodium phosphate buffer solution contained 0.04 U/mL of the 3-epimerase. And 10% (w/v) fructose was reacted at pH 7.5. The reaction was terminated by heating at 100 ° C for 15 minutes, and its enzymatic activity was measured. The result is illustrated in Figure 2-2.
- Figure 2-1 is a graph showing the effect of pH on 3-epimerase activity under the conditions of an embodiment of the invention.
- Figure 2-2 is a graph showing the effect of temperature on 3-epimerase activity under the conditions of an embodiment of the invention.
- Example 6 in order to study the thermal stability of the 3-epimerase, the 3-epimerase was separately incubated at different temperature conditions, and samples were taken at different times to measure fructose as a substrate. Residual activity. The measurement was carried out every 1 hour after the 3-epimerase was incubated at 50 ° C, 60 ° C, 70 ° C, 80 ° C and 90 ° C water bath conditions, respectively.
- the 3-epimerase reaction was carried out in a sodium phosphate buffer solution of 50 mM pH 7.5 at 60 ° C for 20 minutes, wherein the sodium phosphate buffer solution contained 0.04 U/mL of 3-epimerase and 10
- the reaction solution was heated at 100 ° C for 15 minutes to terminate the reaction, and then the activity of the 3-epimerase was measured. The result is illustrated in FIG.
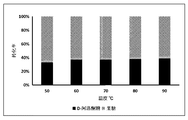
- Example 7 the 3-epimerase reaction was carried out in a pH 7.5 50 mM sodium phosphate buffer solution at a temperature range of 40-90 ° C. The reaction was allowed to proceed sufficiently for 12 hours, wherein the sodium phosphate buffer solution contained the 3-epimerase 0.04 U/mL, 1 mM cobalt ion, and 10% (w/v) fructose. . Then, the reaction was terminated by heating at 100 ° C for 15 minutes, and the contents of fructose and D-psicose in the sample were measured. The result is illustrated in FIG.
- the reaction is carried out in a pH 7.5 50 mM sodium phosphate buffer solution at 60 ° C, wherein the sodium phosphate buffer solution contains 10 U/mL of the 3-differential Isomerase, 1 mM cobalt ion and 700 g/L fructose. Samples were then taken at different reaction time points, the reaction was stopped by heating at 100 ° C for 15 minutes, and the concentration of D-psicose in the sample was measured. The D-psicose yields for different reaction times are shown in Table 3 below.
- the 3-epimerase was immobilized.
- the yield of immobilized 3-epimerase production was measured and compared to the production yield of unfixed (free) 3-epimerase.
- a 3-epimerase-alginate complex sphere was used, which was prepared by the following method: 3-epimerase solution Adding to 2.5% (w/v) sodium alginate solution, the volume of the sodium alginate solution is 1.5 times the volume of the 3-epimerase solution, and the mixture is added by a syringe pump and a vacuum pump. Into a 0.2M calcium ion solution.
- This reaction was carried out in the same manner as in Example 7 except that a fixed 3-epimerase was used.
- the amount of the 3-epimerase used in the reaction was 10 U/mL, and the D-psicose production rate was measured. The results are shown in Table 4 below.
- Example 10 Production yield of D-psicose in a bioreactor
- the immobilized 3-epimerase and fructose were prepared in the same manner as in Example 9, and fructose was added to the immobilized 3-epimerase, and the mixture was adjusted to a volume. 100 mL Subsequently, a bioreactor of 100 cm in height and 2.6 cm in diameter was filled with the mixture of the immobilized 3-epimerase and fructose, and the reaction was carried out at flow rates of 10 mL/h and 60 °C. The amount of 3-epimerase used was 500 U, and the concentration of fructose used was limited to 600 g/L due to precipitation of excess fructose during prolonged operation. The results are shown in Table 5 below.
- the present invention can provide a D-psicose production system using a bioreactor capable of mass production on an industrial scale.
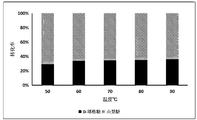
- Example 11 the 3-epimerase reaction was carried out in a pH 7.5 50 mM sodium phosphate buffer solution at a temperature ranging from 40 to 90 ° C for 12 hours, thereby allowing the reaction to proceed sufficiently, wherein
- the sodium phosphate buffer solution contained the 3-epimerase 0.04 U/mL, 1 mM cobalt ion, and 10% sorbose.
- the reaction was terminated by heating at 100 ° C for 15 minutes, and the contents of sorbose and D-tagatose in the samples were measured. The result is illustrated in FIG. 5.
- the reaction is carried out in a pH 7.5 50 mM sodium phosphate buffer solution at 60 ° C, wherein the sodium phosphate buffer solution contains 20 U/mL of the 3-differential Constructed enzyme, 1 mM cobalt ion and 500 g/L sorbose. Samples were then taken at different reaction time points, terminated by heating at 100 °C for 15 minutes, and the concentration of D-tagatose in the samples was measured. The D-tagatose yields for different reaction times are shown in Table 6 below.
- the 3-epimerase was immobilized.
- the production yield of the solid 3-densitized 3-epimerase was measured and compared with the production yield of the unfixed (free) 3-epimerase.
- a 3-epimerase-alginate complex sphere was used, which was prepared by the following method: 3-epimerase solution Adding to 2.5% (w/v) sodium alginate solution, the volume of the sodium alginate solution is 1.5 times the volume of the 3-epimerase solution, and the mixture is added by a syringe pump and a vacuum pump. Into a 0.2M calcium ion solution.
- This reaction was carried out in the same manner as in Example 12 except that a fixed 3-epimerase was used.
- the amount of 3-epimerase used in the reaction was 20 U/mL, and D-tagatose productivity was measured. The results are shown in Table 7 below.
- Example 14 Production yield of D-tagatose in a bioreactor
- the immobilized 3-epimerase and sorbose were prepared in the same manner as in Example 13, sorbose was added to the immobilized 3-epimerase, and the mixture was adjusted.
- a bioreactor 100 cm high and 2.6 cm in diameter was filled with the mixture of immobilized 3-epimerase and sorbose, and the reaction was carried out at flow rates of 10 mL/h and 60 °C. .
- the amount of 3-epimerase used was 400 U, and the concentration of sorbose used was 400 g/L. The results are shown in Table 8 below.
- the present invention can provide a D-tagatose production system using a bioreactor capable of mass production on an industrial scale.
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Nutrition Science (AREA)
- Food Science & Technology (AREA)
- Polymers & Plastics (AREA)
- Enzymes And Modification Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
Description
| 金属离子 | 相对活性(%) |
| 无 | 100 |
| Co2+ | 170 |
| Ca2+ | 65 |
| Mn2+ | 168 |
| Mg2+ | 130 |
| Zn2+ | 0 |
| Cu2+ | 0 |
| Na+ | 120 |
| K+ | 113 |
| 相对活性(%) | |
| 果糖 | 63.9 |
| D-阿洛酮糖 | 100 |
| 山梨糖 | 21 |
| D-塔格糖 | 40.7 |
| D-阿洛酮糖(g/L) | |
| 1 | 98 |
| 2 | 154 |
| 3 | 182 |
| 4 | 210 |
| 6 | 259 |
| 8 | 257 |
| 反应时间(小时) | D-阿洛酮糖(g/L) |
| 1 | 89 |
| 2 | 141 |
| 3 | 165 |
| 4 | 190 |
| 6 | 231 |
| 8 | 258 |
| 10 | 260 |

| 反应时间(小时) | D-塔格糖(g/L) |
| 1 | 65 |
| 2 | 101 |
| 3 | 120 |
| 4 | 139 |
| 6 | 171 |
| 8 | 170 |
| 反应时间(小时) | D-塔格糖(g/L) |
| 1 | 59 |
| 2 | 95 |
| 3 | 108 |
| 4 | 125 |
| 6 | 153 |
| 8 | 170 |
| 10 | 171 |

Claims (13)
- 一种新型3-差向异构酶,其选自下组:(a)多肽或蛋白质,其氨基酸序列与SEQ ID No:2具有至少70%,例如至少75%,至少80%,至少85%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%,或甚至100%序列同一性;(b)多肽或蛋白质,其由多核苷酸编码,所述多核苷酸在中等-高严格条件下与以下杂交:(i)SEQ ID No:1的多肽编码序列,(ii)包含SEQ ID No:1的多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;(c)多肽或蛋白质,其由多核苷酸编码,所述多核苷酸与SEQ ID NO:1的多肽编码序列具有至少70%,例如至少75%,至少80%,至少85%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%,或甚至100%序列同一性;(d)SEQ ID NO:2的多肽或蛋白质包含一个或多个(几个)氨基酸的取代、缺失和/或插入的变体;(e)任何(a)、(b)或(c)的多肽或蛋白质,其氨基酸序列包含SEQ ID No:2或由SEQ ID No:2组成,和(f)(a),(b),(c),(d),或(e)的多肽或蛋白质的片段,其具有3-差向异构酶活性。
- 权利要求1的分离的多肽或蛋白质,其与SEQ ID No:2的多肽或蛋白质的氨基酸序列具有至少70%,例如至少75%,至少80%,至少85%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%,或甚至100%序列同一性。
- 权利要求1或2任一项的分离的多肽或蛋白质,其包含SEQ ID No:2或由SEQID No:2组成。
- 一种分离的多核苷酸,其编码权利要求1至3任一项的多肽或蛋白质。
- 一种核酸构建体或表达载体,其包含权利要求4的多核苷酸,所述多核苷酸可操作地连接于一个或多个(几个)调控序列,所述调控序列指导所述多肽或蛋白质在表达载体中的产生。
- 一种重组宿主细胞,其包含权利要求4的多核苷酸,所述多核苷酸可操作地连接于一个或多个调控序列,所述调控序列指导所述多肽或蛋白质的产生。
- 一种产生权利要求1至3任一项的多肽或蛋白质的方法,其包括:(a)在有助于所述多肽或蛋白质的产生的条件下培养细胞,其以其野生型形式产生所述多肽或蛋白质;和(b)回收所述多肽或蛋白质。
- 一种产生权利要求1至3任一项的多肽或蛋白质的方法,其包括:(a)在有助于所述多肽或蛋白质的产生的条件下培养权利要求6的宿主细胞;和(b)回收所述多肽或蛋白质。
- 一种组合物,其包含权利要求1至3任一项的多肽或蛋白质和其它酶。
- 权利要求1至3任一项的多肽或蛋白质或权利要求9的组合物在用于生产D-阿洛酮糖的工艺中的用途。
- 权利要求1至3任一项的多肽或蛋白质或权利要求9的组合物在用于生产D-塔格糖的工艺中的用途。
- 一种用于生产D-阿洛酮糖的方法,其包括将果糖、葡萄糖及淀粉糖等混合糖与权利要求1至3任一项的多肽或蛋白质的水性溶液相接触生产D-阿洛酮糖。
- 一种用于生产D-塔格糖的方法,其包括将山梨糖及淀粉糖等混合糖与权利要求1至3任一项的多肽或蛋白质的水性溶液相接触生产D-塔格糖。
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020187028940A KR102068113B1 (ko) | 2016-04-01 | 2017-03-31 | 3-에피머화 효소 및 이를 코딩하는 폴리뉴클레오티드 |
| US16/088,821 US10781467B2 (en) | 2016-04-01 | 2017-03-31 | 3-epimerase and polynucleotide encoding same |
| EP17773285.6A EP3438256B1 (en) | 2016-04-01 | 2017-03-31 | 3-epimerase and polynucleotide encoding same |
| MX2018011980A MX2018011980A (es) | 2016-04-01 | 2017-03-31 | 3-epimerasa y polinucleotido que la codifica. |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN201610047300.4A CN105821027B (zh) | 2016-04-01 | 2016-04-01 | 一种3-差向异构酶的用途 |
| CN201610047300.4 | 2016-04-01 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2017167255A1 true WO2017167255A1 (zh) | 2017-10-05 |
Family
ID=56986929
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/CN2017/078923 Ceased WO2017167255A1 (zh) | 2016-04-01 | 2017-03-31 | 一种3-差向异构酶以及编码它的多核苷酸 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US10781467B2 (zh) |
| EP (1) | EP3438256B1 (zh) |
| KR (1) | KR102068113B1 (zh) |
| CN (1) | CN105821027B (zh) |
| MX (1) | MX2018011980A (zh) |
| WO (1) | WO2017167255A1 (zh) |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2019132558A1 (ko) * | 2017-12-27 | 2019-07-04 | 씨제이제일제당 주식회사 | 신규 내열성 과당-6-인산 3-에피머화 효소 및 이를 이용한 알룰로스 제조방법 |
| WO2019132556A1 (ko) * | 2017-12-27 | 2019-07-04 | 씨제이제일제당 주식회사 | 신규 내열성 과당-6-인산 3-에피머화 효소 및 이를 이용한 알룰로스 제조방법 |
| WO2020067781A1 (ko) * | 2018-09-28 | 2020-04-02 | 씨제이제일제당 (주) | 신규 과당-4-에피머화 효소 및 이를 이용한 타가토스의 제조 방법 |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN105821027B (zh) | 2016-04-01 | 2023-11-21 | 南京朗奈生物技术有限公司 | 一种3-差向异构酶的用途 |
| CA3088598C (en) * | 2018-01-24 | 2021-03-30 | Matsutani Chemical Industry Co., Ltd. | Ketose 3-epimerase with improved thermal stability |
| CN108251468A (zh) * | 2018-02-06 | 2018-07-06 | 南京朗奈生物技术有限公司 | 生物法生产d-阿洛酮糖的工艺 |
| CN111793616B (zh) * | 2020-08-07 | 2022-04-12 | 天津科技大学 | 一种差向异构酶的突变体及其应用 |
| KR102691596B1 (ko) | 2022-12-20 | 2024-08-06 | 대상 주식회사 | 항시발현용 신규 프로모터 변이체 및 이의 용도 |
Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN101189332A (zh) * | 2005-06-01 | 2008-05-28 | Cj第一制糖株式会社 | 利用d-阿洛酮糖差向异构酶的d-阿洛酮糖生产方法 |
| CN103131721A (zh) * | 2011-11-25 | 2013-06-05 | 天津工业生物技术研究所 | 瘤胃菌d-塔格糖3-差向异构酶的核苷酸序列及其应用 |
| CN103710330A (zh) * | 2014-01-03 | 2014-04-09 | 江南大学 | 一种高催化活性的d-阿洛酮糖 3-差向异构酶的突变体酶及其应用 |
| CN104160023A (zh) * | 2011-08-24 | 2014-11-19 | Cj第一制糖株式会社 | 热稳定性得到改善的d-阿洛酮糖3-差向异构酶突变体和使用其的d-阿洛酮糖的连续制备 |
| CN105821027A (zh) * | 2016-04-01 | 2016-08-03 | 南京朗奈生物技术有限公司 | 一种新型3-差向异构酶以及对其进行编码的多核苷酸 |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US9932617B2 (en) | 2013-01-08 | 2018-04-03 | Matsutani Chemical Industry Co., Ltd. | Ketose 3-epimerase produced by arthrobacter globiformis |
| DK3298127T3 (da) * | 2015-05-22 | 2022-04-25 | Archer Daniels Midland Co | Anvendelse af epimerase-enzymer til omdannelse af fructose til allulose |
-
2016
- 2016-04-01 CN CN201610047300.4A patent/CN105821027B/zh active Active
-
2017
- 2017-03-31 KR KR1020187028940A patent/KR102068113B1/ko active Active
- 2017-03-31 EP EP17773285.6A patent/EP3438256B1/en active Active
- 2017-03-31 US US16/088,821 patent/US10781467B2/en active Active
- 2017-03-31 MX MX2018011980A patent/MX2018011980A/es unknown
- 2017-03-31 WO PCT/CN2017/078923 patent/WO2017167255A1/zh not_active Ceased
Patent Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN101189332A (zh) * | 2005-06-01 | 2008-05-28 | Cj第一制糖株式会社 | 利用d-阿洛酮糖差向异构酶的d-阿洛酮糖生产方法 |
| CN104160023A (zh) * | 2011-08-24 | 2014-11-19 | Cj第一制糖株式会社 | 热稳定性得到改善的d-阿洛酮糖3-差向异构酶突变体和使用其的d-阿洛酮糖的连续制备 |
| CN103131721A (zh) * | 2011-11-25 | 2013-06-05 | 天津工业生物技术研究所 | 瘤胃菌d-塔格糖3-差向异构酶的核苷酸序列及其应用 |
| CN103710330A (zh) * | 2014-01-03 | 2014-04-09 | 江南大学 | 一种高催化活性的d-阿洛酮糖 3-差向异构酶的突变体酶及其应用 |
| CN105821027A (zh) * | 2016-04-01 | 2016-08-03 | 南京朗奈生物技术有限公司 | 一种新型3-差向异构酶以及对其进行编码的多核苷酸 |
Non-Patent Citations (3)
| Title |
|---|
| DATABASE GenBanK [O] 20 August 2015 (2015-08-20), XP055427217, Database accession no. WP_052889376 * |
| DATABASE GenBank [O] 20 August 2015 (2015-08-20), XP055427219, Database accession no. WP_052889376 * |
| See also references of EP3438256A4 * |
Cited By (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2019132558A1 (ko) * | 2017-12-27 | 2019-07-04 | 씨제이제일제당 주식회사 | 신규 내열성 과당-6-인산 3-에피머화 효소 및 이를 이용한 알룰로스 제조방법 |
| WO2019132556A1 (ko) * | 2017-12-27 | 2019-07-04 | 씨제이제일제당 주식회사 | 신규 내열성 과당-6-인산 3-에피머화 효소 및 이를 이용한 알룰로스 제조방법 |
| KR20190079351A (ko) * | 2017-12-27 | 2019-07-05 | 씨제이제일제당 (주) | 신규 내열성 과당-6-인산 3-에피머화 효소 및 이를 이용한 알룰로스 제조방법 |
| KR20190079352A (ko) * | 2017-12-27 | 2019-07-05 | 씨제이제일제당 (주) | 신규 내열성 과당-6-인산 3-에피머화 효소 및 이를 이용한 알룰로스 제조방법 |
| KR102055875B1 (ko) | 2017-12-27 | 2019-12-13 | 씨제이제일제당 주식회사 | 신규 내열성 과당-6-인산 3-에피머화 효소 및 이를 이용한 알룰로스 제조방법 |
| KR102063908B1 (ko) | 2017-12-27 | 2020-01-08 | 씨제이제일제당 주식회사 | 신규 내열성 과당-6-인산 3-에피머화 효소 및 이를 이용한 알룰로스 제조방법 |
| WO2020067781A1 (ko) * | 2018-09-28 | 2020-04-02 | 씨제이제일제당 (주) | 신규 과당-4-에피머화 효소 및 이를 이용한 타가토스의 제조 방법 |
| US11459594B2 (en) | 2018-09-28 | 2022-10-04 | Cj Cheiljedang Corporation | Fructose-4-epimerase and method of preparing tagatose using the same |
Also Published As
| Publication number | Publication date |
|---|---|
| KR20180117190A (ko) | 2018-10-26 |
| CN105821027A (zh) | 2016-08-03 |
| MX2018011980A (es) | 2019-02-13 |
| EP3438256B1 (en) | 2020-07-01 |
| CN105821027B (zh) | 2023-11-21 |
| EP3438256A4 (en) | 2019-08-21 |
| US20190136282A1 (en) | 2019-05-09 |
| US10781467B2 (en) | 2020-09-22 |
| EP3438256A1 (en) | 2019-02-06 |
| KR102068113B1 (ko) | 2020-01-20 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| WO2017167255A1 (zh) | 一种3-差向异构酶以及编码它的多核苷酸 | |
| US11859224B2 (en) | Methods for manufacturing a product using a 3-epimerase | |
| CN101189332B (zh) | 利用d-阿洛酮糖差向异构酶的d-阿洛酮糖生产方法 | |
| CN111836889B (zh) | 热稳定性提高的酮糖3-差向异构酶 | |
| CN107723307A (zh) | 一种高效制备d‑阿洛酮糖3‑差向异构酶的方法及其应用 | |
| CN108018278B (zh) | 一种催化效率提高的d-阿洛酮糖3-差向异构酶突变体 | |
| TW201823456A (zh) | D-阿洛酮糖表異構酶、使用其製備d-阿洛酮糖的方法、編碼其的聚何苷酸、重組載體、微生物以及組成物 | |
| WO2021244005A1 (zh) | 阿洛酮糖3-差向异构酶突变体、表达其的工程菌及其固定化酶和固定化方法 | |
| CN112831489A (zh) | 一种阿洛酮糖3-差向异构酶固定化酶、其固定化方法与应用 | |
| CN105713883B (zh) | 一种l-脯氨酸-4-羟基化酶及其应用 | |
| CN104561194A (zh) | 一种n-乙酰神经氨酸醛缩酶在催化合成n-乙酰神经氨酸中的应用 | |
| CN117247916B (zh) | 一种提高糖原分支酶热稳定性的方法及其应用 | |
| KR20190001934A (ko) | 기능성 감미료의 제조방법 | |
| CN119859622B (zh) | 葡萄糖基转移酶突变体及其应用 | |
| KR102312806B1 (ko) | 비피도박테리움 속 유래인 저항전분 분해 활성을 나타내는 아밀라아제들 및 이의 용도 | |
| JPWO2009119538A1 (ja) | エピメラーゼ活性を有する新規ポリペプチド、その製造方法ならびにその利用方法 | |
| KR20240136938A (ko) | 효율적인 트랜스프럭토실화 활성을 갖는 돌연변이체 FTase | |
| CN117683750A (zh) | 重组α-葡萄糖苷酶及其制备方法 | |
| CN121472205A (zh) | 一种催化特性提高的d-阿洛酮糖3-差向异构酶突变体 | |
| KR101237857B1 (ko) | 신규한 알파-엘-아라비노퓨라노시데이즈 및 그 효소를 이용한 반섬유소 아라비난으로부터 아라비노스의 제조 방법 | |
| JP2001321177A (ja) | 新規耐熱性α−ガラクトシダーゼ遺伝子 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| DPE2 | Request for preliminary examination filed before expiration of 19th month from priority date (pct application filed from 20040101) | ||
| WWE | Wipo information: entry into national phase |
Ref document number: MX/A/2018/011980 Country of ref document: MX |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 20187028940 Country of ref document: KR Kind code of ref document: A |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2017773285 Country of ref document: EP |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 17773285 Country of ref document: EP Kind code of ref document: A1 |