WO2022181515A1 - Pd-1シグナル阻害剤の併用療法 - Google Patents
Pd-1シグナル阻害剤の併用療法 Download PDFInfo
- Publication number
- WO2022181515A1 WO2022181515A1 PCT/JP2022/006843 JP2022006843W WO2022181515A1 WO 2022181515 A1 WO2022181515 A1 WO 2022181515A1 JP 2022006843 W JP2022006843 W JP 2022006843W WO 2022181515 A1 WO2022181515 A1 WO 2022181515A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- cells
- mice
- cell
- tcr
- signaling
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images













Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K45/00—Medicinal preparations containing active ingredients not provided for in groups A61K31/00 - A61K41/00
- A61K45/06—Mixtures of active ingredients without chemical characterisation, e.g. antiphlogistics and cardiaca
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/13—Amines
- A61K31/135—Amines having aromatic rings, e.g. ketamine, nortriptyline
- A61K31/136—Amines having aromatic rings, e.g. ketamine, nortriptyline having the amino group directly attached to the aromatic ring, e.g. benzeneamine
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/16—Amides, e.g. hydroxamic acids
- A61K31/165—Amides, e.g. hydroxamic acids having aromatic rings, e.g. colchicine, atenolol, progabide
- A61K31/167—Amides, e.g. hydroxamic acids having aromatic rings, e.g. colchicine, atenolol, progabide having the nitrogen of a carboxamide group directly attached to the aromatic ring, e.g. lidocaine, paracetamol
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K35/00—Medicinal preparations containing materials or reaction products thereof with undetermined constitution
- A61K35/12—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
- A61K39/39533—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum against materials from animals
- A61K39/3955—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum against materials from animals against proteinaceous materials, e.g. enzymes, hormones, lymphokines
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2827—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against B7 molecules, e.g. CD80, CD86
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
Definitions
- Non-Patent Document 1- 4 The results of recent clinical trials have revealed that PD-1 signal inhibition therapy is more effective than conventional therapies in many cancer types (1-4) (Non-Patent Document 1- 4).
- the response rate of anti-PD-1 antibody treatment was 20-30%, which was dramatically improved compared to conventional anticancer agents.
- the purpose of the present invention is to provide a new combination therapy with PD-1 signal inhibition therapy.
- T cell activation P. S. Chowdhury, K. Chamoto, T. Honjo, Combination therapy strategies for improving PD-1 blockade efficiency: a new era in cancer immunotherapy. J Intern Med 283, 110-120 (2016).; K. Chamoto et al., Mitochondrial activation chemicals synergize with surface receptor PD-1 blockade for T cell-dependent antitumor activity. A 114, E761-E770 (2017).).
- T cell activation is facilitated by the T cell receptor (TCR) signaling cascade. Phosphorylation and dephosphorylation of TCR signaling molecules affect signal complex formation and TCR signal transduction.
- CD45 is a transmembrane phosphatase that plays a major role in regulating TCR signaling by regulating the level of tyrosine phosphorylation of lymphocyte protein kinase (Lck) and its downstream factors (J. Rossy, D J. Williamson, K. Gauss, How does the kinase Lck phosphorylate the T cell receptor? Spatial organization as a regulatory mechanism. Front Immunol 3, 167 (2012).) (R. J. Brownlie, R. Zamoyska, T cell receptor signaling networks: branched, diversified and bounded. Nat Rev Immunol 13, 257-269 (2013).).
- Lck lymphocyte protein kinase
- CD45 inhibitors support anti-tumor immunity when used in combination with PD-1 signal inhibitors, maintain synergistic effects of suppressing cancer growth, and can be used as monotherapy for PD-1 signal inhibition. It is thought to be effective even against cancers that exhibit unresponsiveness. A similar effect was seen with cell transplantation (xenogeneic cells, allogeneic cells). Enhanced TCR signaling was also confirmed by cell transplantation.
- the gist of the present invention is as follows.
- a pharmaceutical composition comprising a substance capable of enhancing T-cell receptor (TCR) signaling and administered either before, after or simultaneously with administration of a PD-1 signaling inhibitor.
- TCR T-cell receptor
- the CD45 inhibitor is 2-(4-Acetylanilino)-3-chloronaphthoquinone, N-(9,10-Dioxo-9,10-dihydrophenanthren-2-yl)-2,2-dimethylpropionamide and analogues thereof
- the pharmaceutical composition according to (2) which is at least one compound selected from the group consisting of: (4) The pharmaceutical composition according to (2), wherein the cells are xenocells, allocells or a combination thereof.
- a PD-1 signal inhibitory activity enhancer comprising a CD45 inhibitor and/or cells.
- a TCR signal enhancer comprising a CD45 inhibitor and/or cells.
- a method of preventing and/or treating cancer, an infectious disease, or a combination thereof comprising: (11) administration of an effective amount of a substance capable of enhancing T cell receptor (TCR) signaling to the subject either before, after, or concurrently with the administration of the PD-1 signaling inhibitor; A method of enhancing PD-1 signaling inhibitory activity, comprising: (12) A method of enhancing TCR signaling, comprising administering an effective amount of a CD45 inhibitor and/or cells to a subject. (13) A substance capable of enhancing T-cell receptor (TCR) signaling for use in preventing and/or treating cancer, infectious disease, or a combination thereof, which is a PD-1 signal inhibitor The substance administered either before, after, or concurrently with the administration of
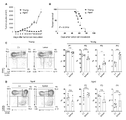
- MC38 cells were inoculated into the skin of young and aged PD-1 KO mice.
- A MC38 tumor size in young (3-4 months old) and aged (15 months old) PD-1 KO mice.
- B Kaplan-Meier plot of survival of MC38 tumor-bearing PD-1 KO mice.
- C and D Analysis of CD8-positive T cell subsets in young (C, 2-3 months old) or aged (D, 15-21 months old) PD-1 KO mice with or without inoculation of MC38 cells. . Peripheral and regional lymph node cells stained on Day 9.
- CD44 and CD62L expression in CD3+CD8+ cells and percentages of CD8+ cell subsets CD44 low /CD62L high (naive; P1), CD44 high /CD62L high (central memory; P2), CD44 high /CD62L low (effects/memory; P3), CD44 low /CD62L low (P4).
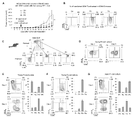
- MC38-OVA cells were injected into young OT-1 mice via the tail vein and the respective subsets were isolated from splenocytes. Five days after intradermal implantation of MC38-OVA cells, P3 or P4 subset cells were adoptively transferred into CD8 KO mice.
- A Tumor volume in MC38-OVA tumor-bearing CD8 KO mice with or without transfer of P3 or P4 cells.
- B FACS analysis of transplanted CD8-positive T cells in day 11 peripheral blood.
- C Scheme of CD8-positive T-cell subset isolation from splenocytes of young PD-1 KO mice.
- a to E Microarray analysis of P1, P2, P3, and P4 cells isolated from young PD-1 KO mice (1 to 3 months old; mix of 9 mice).
- A Hierarchical clustering heatmap of all genes.
- B Scatter plot showing the normalized log intensity of each probe. Dotted lines indicate log 2-fold differences. Genes that have been shown to be involved in the activation and differentiation of CD8-positive cells are listed.
- C Top 10 gene ontology (GO) involving genes upregulated in P4 cells. GO terms involved in 1C metabolism are shown in red.
- D Schematic representation of the 1C metabolic pathway. THF, tetrahydrofolate.
- E Heatmap showing the expression of 1C metabolic-related genes in CD8-positive T-cell subsets.
- OCR Oxygen consumption rate
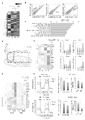
- FIG. 1 Representative diagrams showing the expression of CD44 and CD62L on CD8-positive T cells and the percentage of each CD8-positive T-cell subset in the mice shown 5 days after MC38-OVA transplantation.
- C Percentage of pZAP70-positive CD8-positive T cells in peripheral lymph nodes from young or aged OT-1 mice with or without MC38-OVA transplantation.
- D Representative plots of CD45RB expression levels and fluorescence intensity in total CD8+ T cells or respective subsets from young (2-3 months old) or aged (14-17 months old) OT-1 mice. (MFI).
- A Schematic diagram of the experimental schedule.
- mice Ten days after the transplantation (day 0), the mice were implanted with MC38 (E to G) or MC38-OVA (H and I) cells in the skin and used for the experiments described below.
- G Percentage of pZAP70-positive cells in CD8-positive T cells from day 6 regional lymph nodes.
- a and B MC38 cells were inoculated into the skin of young (2 months old) and aged (14 months old) C57BL/6 wild-type mice. The mice then received anti-PD-L1 antibody on days 5, 11, and 17.
- A MC38 tumor size in young and aged wild-type mice.
- B Kaplan-Meier plot of survival of MC38 tumor-bearing wild-type mice. P values were calculated by log-rank test.
- C and D Analysis of CD8-positive T cell subsets in young (C, 2 months old) or aged (D, 15 months old) wild-type mice with or without inoculation of MC38 cells.
- mice (B and C) MC38 tumor size in young (B) and aged (C) wild-type mice.
- (D and E) MC38 tumor-bearing young (D) and aged (E) wild-type mouse survival. Data represent the mean ⁇ s.e.m. of mice (n 5-6). *p ⁇ 0.05; **p ⁇ 0.01; n.s., no significant difference (evaluated by ANOVA followed by Tukey's test or log-rank test). Restoration of age-related mitochondrial damage by xenotransplantation.
- MC38-OVA cells were implanted into the skin of young (3-4 months old) and aged (17-18 months old) PD-1 KO mice that had been transplanted with PBS (Ctrl) or Daudi cells 10 days earlier. .
- OCR trace (A), basal respiration (B) and reserve respiratory capacity were calculated from OCR values (C).
- MMC-treated splenocytes from C57BL/6 (Ctrl) or Balb/c (Allo) mice were injected into the tail vein of young (1 month old) and aged (14 to 18 months old) C57BL/6 wild-type mice. ported by Subsequent experimental conditions are the same as in FIG. 9A.
- C and D MC38 tumor-bearing young (C) and aged (D) survival of C57BL/6 wild-type mice.
- Experimental method and flow Enhanced PD-1 signaling inhibitory effect by CD45 inhibition. Pre-administration: PD-1 knockout (KO) mice were administered vehicle (control) or 211 and 3 days later were transplanted with MC38 cells (1 ⁇ 10 6 ).
- 211 was administered every 2 days from Day 5, 5 days after MC38 cell transplantation to PD-1 KO mice, for a total of 3 doses.
- the present invention provides a pharmaceutical composition comprising a substance capable of enhancing T cell receptor (TCR) signaling and administered either before, after or simultaneously with administration of a PD-1 signaling inhibitor. do.
- TCR T cell receptor
- T cell receptor is an antigen receptor molecule expressed on the cell membrane of T cells.
- TCR T cell receptor
- intracellular signals are transduced through the phosphorylation of downstream factors such as Lck and ZAP-70, resulting in activation of T cells.
- the enhancement of TCR signals can be confirmed by evaluating the phosphorylation of ZAP-70 in CD8-positive T cells under antigen stimulation. A higher level of ZAP-70 phosphorylation indicates an enhanced TCR signal. Phosphorylation of ZAP-70 can be detected by flow cytometric analysis using an anti-p-ZAP-70 antibody.
- TCR signal By enhancing the TCR signal, it is possible to enhance the anti-tumor immune effect of CD8-positive T cells. Therefore, by enhancing the TCR signal, it is possible to enhance the antitumor effect of PD-1 inhibition therapy, and to treat aging animals and humans that are resistant to PD-1 signal inhibition, as well as resistant cancers. can have an effect.
- CD45 inhibitors and/or cells can be exemplified as substances capable of enhancing TCR signals. These substances may also have the effect of inducing P4 cells.
- P4 cells are a CD44 low CD62L low CD8 positive T cell subset.
- the surface markers CD44 and CD62L L-selectin are used to distinguish between three major CD8+ T cells: naive (also called P1; CD44 low CD62L high ) and central memory (P2; CD44 high CD62L high ). ), effector/memory (P3; CD44 high CD62L low ).
- the remaining CD44 low CD62L low (P4) CD8 positive T cell subset is a very minor population.
- CD45 inhibitors examples include 2-(4-Acetylanilino)-3-chloronaphthoquinone, N-(9,10-Dioxo-9,10-dihydrophenanthren-2-yl)-2,2-dimethylpropionamide and their analogues. can do.
- analog is a concept including salts of compounds, derivatives of compounds, prodrugs for active metabolites, active metabolites for prodrugs, solvates thereof, and the like.
- Compound salts include salts formed with acids such as hydrochloric acid, mesylic acid (methanesulfonic acid), fumaric acid, and phosphoric acid when the compound has an amino group (including substituted amino groups) or an amide group. can do.
- a derivative of a compound is a compound that has undergone modifications such as introduction or substitution of a functional group, oxidation, reduction, or replacement of an atom, to the extent that the structure or properties of the parent substance are not significantly changed.
- Derivatives of -chloronaphthoquinone include those in which the chloro (-Cl) group is replaced with other halogen groups (e.g., fluoro (-F) group, bromo (-Br) group, etc.), and those in which the acetyl (CH 3 CO-) group is substituted.
- Derivatives of ,10-dihydrophenanthren-2-yl)-2,2-dimethylpropionamide include those in which the dimethylpropionyl group is substituted with other alkyl groups (e.g., methyl, ethyl, propyl, benzyl, etc.). can be exemplified.
- prodrugs include compounds in which the amino group of the active compound is acylated, alkylated, or phosphorylated (for example, the amino group of the active compound is eicosanoylated, alanylated, pentylaminocarbonylated, (5-methyl-2 -oxo-1,3-dioxolen-4-yl)methoxycarbonylated, tetrahydrofuranylated, pyrrolidylmethylated, pivaloyloxymethylated, t-butylated compounds, etc.), where the amide group of the active compound is Alkylated (for example, methylated, ethylated, provylated, etc.) and the like can be exemplified.
- the amino group of the active compound is acylated, alkylated, or phosphorylated
- the amino group of the active compound is eicosanoylated, alanylated, pentylaminocarbonylated, (5
- solvates include solvates with solvents such as water, methanol, ethanol, and acetonitrile.
- the solvent may be a single solvent or a mixture of multiple solvents.
- Cells capable of enhancing TCR signals may be xenocells (heterologous cells) or allocells (allogeneic cells) for humans or animals to which the pharmaceutical composition is administered. or a combination thereof.
- xenogeneic Daudi cells and allogeneic splenocytes were transplanted into mice.
- xenocells are heterogeneous cells, and allocells are homogenous cells with different HLA (human leukocyte antigen) or MHC (major histocompatibility complex).
- Various cells xenocells, allocells
- tissues to be administered to living bodies for the prevention or treatment of diseases are known, and these cells may be used in the present invention.
- Allogeneic e.g., breast cancer cell lines, chronic myelogenous leukemia cell lines, lung cancer cells, non-small cell lung cancer cell lines, melanoma cell lines, monocytes, pancreatic cancer cell lines, prostate cancer cell lines, renal cancer cell lines, etc.
- Cells are used to treat various cancers (Human Vaccines & Immunotherapeutics 10:1, 52-63; January 2014). These cells may be irradiated to prevent proliferation and may be genetically modified to secrete immune stimulators such as GM-CSF.
- Heterologous cells such as murine melanoma B16 and Lewis lung carcinoma (LLC) cells have also been used in clinical trials to administer to melanoma patients (Eur J Dermatol 2016 Apr 1;26(2):138- 43).
- cells used for blood products for transfusion erythrocyte products, platelet products, whole blood products
- hematopoietic stem cells used for bone marrow transplantation can also be used. These cells may be either allo cells or xeno cells.
- PD-1 signal refers to the information transduction mechanism carried by PD-1, and as one of them, PD-1 is its ligand PD-L1, in cooperation with PD-L2, Signal transduction mechanisms that suppress activation of T cells can be exemplified.
- PD-1 programmed cell death-1
- PD-L1 and PD-L2 are antigens of monocytes and dendritic cells. It is expressed in various cells such as presenting cells and cancer cells.
- PD-1, PD-L1 and PD-L2 act as suppressors that suppress T cell activation.
- Certain cancer cells and virus-infected cells express ligands for PD-1 to suppress T-cell activation and evade host immune surveillance.
- PD-1 signal inhibitors include substances that specifically bind to PD-1, PD-L1 or PD-L2, and such substances include proteins, polypeptides, oligopeptides, nucleic acids (natural (including nucleic acids and artificial nucleic acids), low-molecular-weight organic compounds, inorganic compounds, cell extracts, extracts from animals, plants, soil, and the like. Substances may be natural or synthetic.
- Preferred PD-1 signal inhibitors are antibodies, more preferably antibodies such as anti-PD-1 antibodies, anti-PD-L1 antibodies and anti-PD-L2 antibodies.
- Antibodies may be any of polyclonal antibodies, monoclonal antibodies, chimeric antibodies, single-chain antibodies, humanized antibodies, and humanized antibodies as long as they can inhibit PD-1 signaling. Methods for producing those antibodies are known. Antibodies can be derived from any organism, such as human, mouse, rat, rabbit, goat, guinea pig. Also, as used herein, the antibody refers to Fab, F (ab) '2, ScFv, Diabody, VH, VL, Sc (Fv) 2, Bispecific sc (Fv) 2, Minibody, scFv-Fc monomer, scFv- This concept also includes low-molecular-weight compounds such as Fc dimer.
- PD-1 signaling inhibitors that target PD-1 include nivolumab (Bristol-Myers Squibb), pembrolizumab (Merck), cemiplimab (Regeneron/Sanofi), camrelizumab (Jiangsu Hengrui), sintilimab (Innovent Biologics/Lilly), toripalimab (Shanghai Junshi Bioscience), PD-L1-targeting atezolizumab (Genetech/Roche), durvalumab (AstraZeneca), avelumab (Merck/Pfizer) are approved globally and also target PD-1 tislelizumab (BeiGene), dostarlimab (AnaptysBio/Tesaro), HLX-04 (Shanghai Henlius Biotech), AK-105 (Akeso Biopharma), spartalizumab (Novartis), BCD-100 (Biocad), MGA-012 (
- the pharmaceutical composition of the present invention can be used as an anticancer agent, a therapeutic agent for infectious diseases, or a combination thereof.
- target cancers or tumors include leukemia, lymphoma (Hodgkin's disease, non-Hodgkin's lymphoma, etc.), multiple myeloma, brain tumor, breast cancer, and uterine body.
- lymphoma Hodgkin's disease, non-Hodgkin's lymphoma, etc.
- multiple myeloma brain tumor, breast cancer, and uterine body.
- cancer cervical cancer, ovarian cancer, esophageal cancer, stomach cancer, appendix cancer, colon cancer, liver cancer, gallbladder cancer, bile duct cancer, pancreatic cancer, adrenal cancer, gastrointestinal stromal tumor, mesothelioma, head and neck cancer (Laryngeal cancer, etc.), Oral cancer (Floor of the mouth cancer, etc.), Gingival cancer, Tongue cancer, Buccal mucosa cancer, Salivary gland cancer, Paranasal sinus cancer (Maxillary sinus cancer, Frontal sinus cancer, Ethmoid sinus cancer, Sphenoid sinus cancer) ), thyroid cancer, kidney cancer, lung cancer, osteosarcoma, prostate cancer, testicular tumor (testicular cancer), renal cell carcinoma, bladder cancer, rhabdomyosarcoma, skin cancer (basal cell carcinoma, spinous cell cancer) cancer, malignant melanoma, actinic keratosis, Bowen's disease, Paget's disease, etc.), anal cancer,
- target infectious diseases include bacterial infections (streptococci (group A ⁇ -hemolytic streptococci, pneumococci, etc.), Staphylococcus aureus (MSSA, MRSA), Staphylococcus epidermidis, Enterococcus, Listeria, Meningococcus, Neisseria gonorrhoeae, Pathogenic Escherichia coli (0157:H7, etc.), Klebsiella (Klebsiella pneumoniae), Proteus, Pertussis, Pseudomonas aeruginosa, Serratia, Citrobacter, Acinetobacter, Various infectious diseases caused by Enterobacter, Mycoplasma, Clostridium, etc., tuberculosis, cholera, plague, diphtheria, dysentery, scarlet fever, anthrax, syphilis, tetanus, lepros
- bacterial infections streptococci (group A ⁇ -
- the pharmaceutical composition of the present invention contains a substance capable of enhancing TCR signaling, and is administered either before, after, or at the same time as administration of a PD-1 signaling inhibitor.
- the PD-1 signal inhibitor and the substance capable of enhancing the TCR signal may be administered separately at different times or at the same time, or may be administered together at the same time.
- good. comprising a PD-1 signal inhibitor and a substance capable of enhancing TCR signal when the PD-1 signal inhibitor and the substance capable of enhancing TCR signal are administered together at the same time It may be used as a compounding agent.
- composition of the present invention is administered to a subject systemically or locally, orally or parenterally.
- Subjects include humans and non-human animals.
- PD-1 signal inhibitors e.g., anti-PD-1 antibody, anti-PD-L1 antibody, anti-PD-L2 antibody
- a buffer solution such as PBS, physiological saline, sterile water, etc.
- this solution contains additives (for example, colorants, emulsifiers, suspending agents, surfactants, solubilizers, stabilizers, preservatives, antioxidants, buffers, tonicity agents, etc.).
- additives for example, colorants, emulsifiers, suspending agents, surfactants, solubilizers, stabilizers, preservatives, antioxidants, buffers, tonicity agents, etc.
- the route of administration includes intravenous, intramuscular, intraperitoneal, subcutaneous, and intradermal administration.
- PD-1 signal inhibitors e.g., anti-PD-1 antibody, anti-PD-L1 antibody, anti-PD-L2 antibody
- content in the formulation varies depending on the type of formulation, usually 1 to 100% by weight, preferably 50-100% by weight.
- the formulations may be formulated in unit dosage formulations.
- Dosage, number and frequency of administration of PD-1 signal inhibitors are subject to symptoms, age, body weight, administration method, administration Although it varies depending on the form and the like, for example, in terms of the amount of the active ingredient, an adult usually administers 0.1 to 100 mg/kg body weight, preferably 1 to 10 mg/kg body weight, at least once to achieve the desired effect. It should be administered at a frequency that can be confirmed.
- Substances capable of enhancing TCR signaling may be included in formulations containing PD-1 signaling inhibitors, alone or mixed with excipients or carriers, in tablets, capsules, powders, granules. , liquids, syrups, aerosols, suppositories, injections and the like.
- Excipients or carriers are conventionally used in the art and may be pharmaceutically acceptable, and their type and composition may be changed as appropriate. For example, water, vegetable oil and the like are used as the liquid carrier.
- solid carriers saccharides such as lactose, white sugar and glucose, starches such as potato starch and corn starch, and cellulose derivatives such as crystalline cellulose are used.
- a lubricant such as magnesium stearate, a binder such as gelatin and hydroxypropylcellulose, a disintegrant such as carboxymethylcellulose, and the like may be added.
- antioxidants, coloring agents, corrigents, preservatives and the like may be added.
- Substances capable of enhancing TCR signals can be administered by various routes such as oral, nasal, rectal, transdermal, subcutaneous, intravenous and intramuscular.
- the content of a substance that can enhance the TCR signal in the formulation varies depending on the type of formulation, but is usually 1-100% by weight, preferably 50-100% by weight.
- the content of the substance capable of enhancing the TCR signal in the formulation is preferably 1-100% by weight.
- the content of the substance capable of enhancing the TCR signal in the formulation is usually about 10-100% by weight, preferably 50-100% by weight, and the balance is the carrier. is.
- the formulations may be formulated in unit dosage formulations.
- the dose, frequency and frequency of administration of the substance capable of enhancing the TCR signal vary depending on the type of the substance capable of enhancing the TCR signal, subject's symptoms, age, body weight, administration method, administration form, etc.
- the substance capable of enhancing the TCR signal is a CD45 inhibitor
- it is usually 0.005 ⁇ g (or ⁇ l) to 25000 mg (or ml) per adult in terms of the amount of the active ingredient.
- the substance capable of enhancing the TCR signal is a cell, it is usually about 1 x 10 6 cells/person to 1 x 10 11 cells/person, preferably 5 x 10 6 cells/person to an adult. About 2 x 10 10 cells/person should be administered at least once at a frequency that allows confirmation of the desired effect.
- the ratio of the PD-1 signal inhibitor (e.g., anti-PD-1 antibody, anti-PD-L1 antibody, anti-PD-L2 antibody) and the substance capable of enhancing the TCR signal is the substance capable of enhancing the TCR signal is a CD45 inhibitor, 1:0.1 to 1:100 (mass ratio) is suitable, preferably 1:1 to 1:50, and the substance capable of enhancing the TCR signal is the cell , 1 x 10 6 to 1 x 10 10 cells, preferably 5 x 10 6 to 2 x 10 9 cells, are suitable for 240 mg of the PD-1 signal inhibitor.
- the present invention also provides administration of a pharmaceutically effective amount of a substance capable of enhancing TCR signaling to a subject either before, after, or at the same time as administration of a PD-1 signaling inhibitor. Also provided are methods of preventing and/or treating cancer, infectious diseases, or combinations thereof, including: Furthermore, the present invention relates to the use of a substance capable of enhancing TCR signaling for the treatment of cancer, infectious disease or a combination thereof, wherein the agent is administered before, after or simultaneously with a PD-1 signaling inhibitor. A substance capable of enhancing the TCR signal is administered during any of the above uses.
- the present invention relates to the use of a substance capable of enhancing TCR signaling for use in a method of treating cancer, infectious disease or a combination thereof, comprising administering a PD-1 signaling inhibitor Also provided is said use wherein a substance capable of enhancing the TCR signal is administered either before, after or simultaneously.
- the present invention provides a substance capable of enhancing T cell receptor (TCR) signaling for use in preventing and/or treating cancer, infectious diseases, or a combination thereof, which inhibits PD-1 signaling Also provided are said substances administered either before, after or concurrently with the administration of the agent.
- TCR T cell receptor
- the present invention also provides agents that enhance PD-1 signaling inhibitory activity, including substances capable of enhancing TCR signaling.
- the present invention is to administer to a subject an effective amount of a substance capable of enhancing T cell receptor (TCR) signals either before, after or at the same time as administration of a PD-1 signal inhibitor.
- TCR T cell receptor
- methods of enhancing PD-1 signaling inhibitory activity comprising:
- the drug of the present invention can be used as a combination drug or combination drug with a PD-1 signal inhibitor. Combinations and formulations of PD-1 signal inhibitors and substances capable of enhancing TCR signals are described above.
- the agent of the present invention can be used not only as a medicine but also as an experimental reagent.
- the present invention provides a TCR signal enhancer comprising a CD45 inhibitor and/or cells.
- the invention also provides a method of enhancing TCR signaling comprising administering to a subject an effective amount of a CD45 inhibitor and/or cells.
- CD45 inhibitors and cells are described above.
- the agent of the present invention can be used not only as a medicine but also as an experimental reagent.
- CD8-positive cells play a major role in anti-tumor responses and directly kill cancer cells. Strong suppression of CD8-positive cell immunity in the elderly is associated with cancer and other age-related diseases.
- This study investigated the role of T-cell function in age-related resistance to PD-1 inhibitory therapy. We found that formation of CD44 low CD62L low CD8 positive T cells was reduced in regional lymph nodes of aged tumor-bearing mice. First, P4 cells were induced from naive CD8+ T cells by in vitro stimulation and then differentiated into effector/memory cells.
- Immunosenescence is caused by quantitative and qualitative changes in immune cells.
- One of the major quantitative changes in T cells is a reduction in TCR repertoire diversity due to thymic regression, leading to a reduction in naive cells in the periphery (13-15).
- functional deficits such as impaired TCR signaling, reduced ability to differentiate into effector and memory cells, and reduced cytokine production accumulate in peripheral T cells in the elderly (16, 17).
- age-related changes greatly affect the proportion of well-defined CD8-positive T-cell subsets (18-20).
- CD44 and CD62L are used to distinguish between three major CD8+ T cells: naive (also called P1; CD44 low CD62L high ) and central memory (P2; CD44 high CD62L high ). ), effector/memory (P3; CD44 high CD62L low ).
- the CD44 low CD62L low (P4) CD8 positive T cell subset that remains in naive mice is a very minor population.
- Naive T cells differentiate into effector T cells upon antigen stimulation (14, 21). Some effector cells differentiate into memory cells and are capable of rapidly responding to the same antigenic stimulus and effector cell differentiation.
- TCR T cell receptor
- Phosphorylation and dephosphorylation of TCR signaling molecules affect signal complex formation and TCR signal propagation.
- CD45 is a transmembrane phosphatase that plays a major role in regulating TCR signaling by regulating the level of tyrosine phosphorylation of lymphocyte protein kinase (Lck). This Lck acts as an activator of TCRz, z-chain-associated protein (ZAP)-70, and the linker of activated T cells (LAT), which are downstream factors of TCR signaling (24, 25).
- Lck lymphocyte protein kinase
- 1C metabolism In activated and proliferating cells, 1C metabolism is generally enhanced and supports survival and differentiation through purine and thymidine biosynthesis, amino acid production, epigenetic regulation, and redox regulation (29, 29). 30). Recent reports indicate that 1C metabolism is one of the most frequently induced metabolic pathways during early CD4+ or CD8+ T cell activation (31, 32). After T cell activation, increased 1C metabolism promotes serine processing required for the biosynthesis of new nucleic acids derived from the 1C unit, which is responsible for antigen-specific T cell proliferation and differentiation and effector function. is necessary. This 1C metabolic pathway is also involved in energy production through regulation of mitochondrial protein synthesis by mitochondrial tRNA methylation (33, 34). Since mitochondrial function and 1C metabolism are lower in aged CD4+ T cells than in young CD4+ T cells (35), aging of T cells may be involved in the inactivation of 1C metabolism. .
- P4 cells are intermediate between P1 and P3 cells and retain potential for anti-tumor activity.
- the P3 and P4 subsets of CD8-positive T cells were induced by intravenous injection of OVA-expressing MC38 tumor cells (MC38-OVA) into the tail vein of OT-1 mice and isolated from the spleens 5 days later.
- the same number of P3 or P4 cells were adoptively transferred to young CD8 KO mice engrafted with MC38-OVA.
- MC38-OVA proliferation was strongly suppressed in mice transfected with P3 or P4 cells (Fig. 2A).
- Fig. 2B all transplanted P4 cells in peripheral blood 6 days after transplantation differentiated into P3 cells.
- P1 and P2 cells are known to differentiate into P3 cells during activation, but the characteristics of P4 cells are scarce due to their low numbers.
- P1, P2, and P4 cells were isolated from young PD-1 KO mice and treated with anti-CD3/CD28 antibodies in vitro. stimulated by IL-2 (Fig. 2C). Consistent with the in vivo results, isolated P4 cells differentiated predominantly into P3 cells under these conditions (Fig. 2D).
- FIG. 2E and 2F P4 cells were formed from P1 cells, but not from P2 cells, two days after in vitro stimulation.
- Tumor inoculation increased the expression of 1C metabolic-related genes in total CD8+ T cells from young wild-type and PD-1 KO mice (Fig. 3G), which could be explained by the formation of P4 cells. ( Figures 1C and 6C). Consistent with the high efficiency of P4 cell induction in PD-1 KO mice (Fig. 1C and Fig. 6C), CD8-positive T cells in young PD-1 KO mice were more likely to express most 1C metabolism-related genes than in wild-type mice. was expressed at high levels in response to MC38 tumor cell inoculation (Fig. 3G). In contrast, the expression of these genes in total CD8-positive T cells from aged mice showed no appreciable variation with tumor inoculation, regardless of mouse genotype.
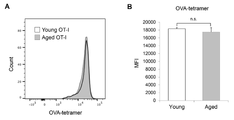
- MC38-OVA injection increased the percentage of CD8-positive T cells expressing phosphorylated ZAP-70 in young mice (Fig. 4C). This suggests that the activation of TCR signaling by antigen stimulation is inhibited in aged OT-1 mice. Remarkably, no significant difference was found in the intensity of OVA-specific MHC tetramers (Fig. 7A&B), indicating no difference in TCR expression levels and avidity of OT-I T cells from young and aged mice. matter was shown. Therefore, the inhibition of antigen-stimulated TCR activation seen in aged mice may involve mechanisms other than TCR-MHC/antigen interactions.
- CD45RB an isoform of CD45 that is predominantly expressed on naive and memory T cells in mice.
- the effect of aging on CD45RB expression was greater in P1 and P4 cells than in P2 and P3 cells (Fig. 4D).
- the highest increase in CD45RB expression was observed in P1 cells of aged wild-type and PD-1 KO mice (Fig.
- Heterologous cell therapy enhances P4 cell induction in aged mice and restores the antitumor potential of CD8+ T cells.
- strong in vitro stimulation restores suppression of the P1 to P4 transition in aged mice, suggesting that strong stimulation using heterologous cells (human Burkitt's lymphoma, Daudi cells) is associated with aging.
- Heterologous cells human Burkitt's lymphoma, Daudi cells
- Fig. 5A Ten days after tail vein injection of Daudi cells (day 0: before MC38 tumor cell transplantation), the proportion of P4 cells was strongly increased in the peripheral lymph nodes of young and aged PD-1 KO mice, but the proportion of P1 to P3 cells decreased. The percentage did not increase significantly (Fig.
- Daudi cell injection strongly suppressed MC38 tumor growth in aged mice and prolonged survival (Fig. 5E&F). Age-related unresponsiveness to anti-PD-L1 antibodies in wild-type mice was also restored by Daudi cell injection (FIGS. 9A-E).
- Fig. 5G the percentage of p-ZAP-70-positive cells was increased in PD-1 KO mice injected with Daudi cells (Fig. 5G), indicating that xenostimulation enhances TCR signals to tumor antigens. It was suggested.
- Daudi cell therapy enhanced the proliferation of tumor antigen-reactive CD8+ T cells.
- MC38-OVA cells were transplanted into PD-1 KO mice 10 days after Daudi cell injection, and the appearance of OVA tetramer-positive CD8-positive T cells was monitored.
- the percentage of OVA tetramer-positive CD8-positive T cells was increased by infusion of Daudi cells in regional lymph nodes and tumor tissues of aged PD-1 KO mice (Fig. 5H&I), indicating that this treatment resulted in antigen-specific CD8 It was shown to increase the induction of positive T cells. Given that prestimulation with Daudi cells enhances P4 cell induction and 1C metabolism in CD8+ T cells from aged mice, it is plausible that Daudi cell infusion increased both basal respiration and respiratory reserve. result (Fig. 10A-C).
- T cells are directly (eg, donor peptide presented by donor MHC) or indirectly (eg, donor peptide presented by recipient MHC) activated by allo- or heterologous MHC molecules (54-56).
- donor MHC donor peptide presented by donor MHC
- recipient MHC donor peptide presented by recipient MHC
- mice and Cells All mice were bred at Kyoto University graduate School of Medicine Animal Research Facility or RIKEN Life Science Research Center under an SPF environment free of specific pathogens that would interfere with experiments, and in accordance with appropriate experimental guidelines. and was used in PD-1-/- (PD-1KO) mice were generated in our laboratory by homologous recombination (59). C57BL/6 wild-type mice were obtained from Charles River Laboratories Japan. OT-1 TCR transgenic and CD8 ⁇ / ⁇ (CD8 KO) mice were purchased from The Jackson Laboratory (Bar Harbor, Maine USA) (deposited by M. B. Bevan, Washington Medical Center or T. Mak, University of Toronto). A mouse colon carcinoma (MC38) cell line was kindly provided by J. P.
- OVA-expressing MC38 cells were isolated from MC38 by a pMXs-based OVA-F2A-EGFO retrovirus. generated by transformation. Cells were cultured in the absence of mycobacterial infection in RPMI 1640 (Gibco; 11875-093) containing 10% inactivated fetal bovine serum and 1% penicillin-streptomycin (Nacalai Tesque; 26253-84). did
- mice Mouse Treatment Model MC38 cells (5 ⁇ 10 5 or 2 ⁇ 10 6 ) were implanted intradermally into the right abdomen of mice (day 0). Five days after MC38 cell transplantation (day 5), the mice were treated with an anti-PD-L1 antibody (clone 1-111A.4, produced and stored at Kyoto University graduate School of Medicine Immunology and Genomic Medicine Laboratory (Immunology Letters 84 (2002) ) 57-62) (6 mg/kg) was administered intraperitoneally, and this anti-PD-L1 antibody administration was repeated three times every 6 days (days 5, 11, and 17).
- an anti-PD-L1 antibody clone 1-111A.4, produced and stored at Kyoto University graduate School of Medicine Immunology and Genomic Medicine Laboratory (Immunology Letters 84 (2002) ) 57-62
- this anti-PD-L1 antibody administration was repeated three times every 6 days (days 5, 11, and 17).
- Antibody isotype (Rat IgG2a) was used as a control.Tumor growth was monitored by measuring tumor size using a caliper, and volume was calculated by the formula ⁇ (length ⁇ width ⁇ height)/6.
- the transfer of non-autologous cells was Burkitt's lymphoma Daudi cells or BALB/c treated with mitomycin C (MMC) 10 days before tumor cell transplantation, or C57BL/6N-derived splenocytes (2 ⁇ 10 6 cells/mouse) as a control. was performed by injection through the tail vein of
- Peripheral and regional lymph node cells and tumor-infiltrating lymphocytes were isolated as previously described (39).
- splenocyte analysis spleens were briefly disaggregated and suspended in potassium ammonium chloride solution to lyse red blood cells. Splenocytes were washed and treated with RPMI containing 10% fetal bovine serum, L-glutamine, 55 ⁇ M 2-mercap and ethanol, penicillin-streptomycin, 1 mM sodium pyruvate (Gibco), 1% MEM non-essential amino acids solution (Gibco). Resuspended in 1640 internal solution (T cell culture medium).
- CD8a Ly-2 microbeads
- FACSAria BD Biosciences
- peripheral blood cells collected peripheral blood cells were treated with potassium ammonium chloride solution and washed with T cell culture medium. Purified cells were stimulated with T cell culture medium supplemented with anti-CD3/28 Dynabeads (Gibco) and recombinant human IL-2 (20 U/mL; PeptoTech).
- CD8-positive T cells using MC38-OVA cells
- CD8-positive T cells isolated from lymph nodes of OT-1 mice were combined with MMC-treated MC38-OVA cells in T-cell culture medium. cultured. One day after stimulation, the cells were treated with 0.05 ⁇ M CD45 inhibitor VI (EDM Millipore) for 1 hour.
- CD3 (clone 145-2C11) and CD8 (clone 53-6.7) (Invitrogen); phospho-ZAP70/Syk (clone n3kobu5) (Invitrogen), CD45.1 (clone A20), CD45.2 (clone 104), CD44 ( Clone IM7) and CD62L (clone MEL-14) (TONBO biosciences or BioLegend); CD8 (clone KT15, MBL Life Science); CD45RB (clone 16A, BD Pharmingen).
- H2Kb Negative (SIY) Tetramer-SIRYYGL-APC and H-2Kb OVA Tetramer-SINFEKL-APC (TS-M008-2) were analyzed by MBL Life Science flow cytometric analysis using FACSCant II (BD Biosciences) and FlowJo software (FlowJo LLC). was used.
- CD8 KO and OT-1 Mouse Models To induce P3 and P4 cells, MC38-OVA cells (1-2 ⁇ 10 6 ) were transferred into OT-1 mice by tail vein injection. Five days later, P3 and P4 cells were isolated from those mice. Each isolated CD8-positive T cell was injected via the tail vein of CD8 KO mice that had been intradermally implanted with 2 ⁇ 10 5 MC38-OVA cells 5 days earlier. Peripheral blood was then obtained 6 days later to analyze the differentiation of the transplanted cells.
- Real-time PCR was performed to amplify the indicated mRNAs using the Applied Biosystems 7500 Fast Real-Time PCT system (ABI) and PowerUp SYBR Green Master Mix (Applied Biosystems). The expression level of each gene was normalized by the ⁇ -actin mRNA level.
- the primer sequences are as follows: Shmt1 forward (Fw), 5'-CCAGAGTGCTGTGGCAACTC-3' (SEQ ID NO: 1); Shmt1 reverse (Rv), 5'-GCAAAACACAGGCTGTTCCTG-3' (SEQ ID NO: 2); Shmt2 Fw, 5'-GACAGTTGAGGACACCTGGC-3' (SEQ ID NO: 3); Shmt2 Rv, 5'-CCAGAGAGGAGTGACATCTC-3' (SEQ ID NO: 4); Phgdh Fw, 5'-TGGCCTCGGCAGAATTGGAAG-3' (SEQ ID NO: 5); Phgdh Rv, 5'-TGTCATTCAGCAAGCCTGTGGT-3' (SEQ ID NO: 6); Psat1 Fw, 5'-GATGAACATCCCATTTCGCATTGG-3' (SEQ ID NO: 7); Psat1 Rv, 5'-GCGTTATACAGAGAGGCACGAATG-3' (SEQ ID
- Oxygen Consumption Rate (OCR) Analysis The OCR assay was performed as previously described with some modifications (39). Oxygen consumption rates of CD8+ T cells isolated from regional lymph nodes were analyzed with the XFe96 Extracellular Flux analyzer (Agilent Technologies) using the Seahorse XFe96 Extracellular Flux assay kit and Seahorse XF Cell Mito Stress test kit (Agilent Technologies). rice field. CD8-positive T cells were seeded (3 ⁇ 10 5 /well) on XFe96 plates coated with Cell-Tak (Corning). Reserve respiratory capacity was calculated from OCR graphs as previously described (60). Basal respiration was calculated by subtraction with non-mitochondrial respiration (values before oligomycin addition to values after rotenone/antimycin A addition) (61).
- mice and Cells All mice were bred at the Kyoto University graduate School of Medicine Animal Experimental Facility under an SPF environment free of specific pathogens that would interfere with experiments, and were used under appropriate experimental guidelines.
- C57BL/6 was obtained from Charles River Laboratories Japan.
- PD-1-/- (PD-1KO) mice were generated in our laboratory by homologous recombination (see Nishimura et al.) (11).
- Mouse colon cancer-derived MC38 cells were a gift from J. P. Allison, Memorial Sloan-Kettering Cancer (New York, NY). LLC cells derived from Lewis lung carcinoma were purchased from the American Type Culture Collection (ATCC).
- ATCC American Type Culture Collection
- Mouse therapeutic model CD45 inhibition ( Figure 12): 2-(4-Acetylanilino)-3-chloronaphthoquinone (211) (530197, EDM Millipore) (12) (3 mg/kg) or N-(9,10-Dioxo-9) ,10-dihydrophenanthren-2-yl)-2,2-dimethylpropionamide (PTP) (540215, Sigma-Aldrich) (13) (3 mg/kg) was administered to mice before or after (5 or 7 days after) tumor cell implantation. was administered intraperitoneally. MC38 cells (5 ⁇ 10 5 or 1 ⁇ 10 6 ) or LLC cells (2 ⁇ 10 6 ) were implanted intradermally into the right flank of mice (day 0).
- mice Seven days after LLC cell transplantation (day 7), the mice were intraperitoneally administered anti-PD-L1 antibody (clone 1-111A.4) (2 mg/kg). This anti-PD-L1 antibody administration was repeated three times every 6 days (days 7, 13, and 19). The tumor volume was measured with a caliper and calculated by the formula ⁇ (length ⁇ width ⁇ height)/6.
- PD-1 KO mice (2 months old) were intraperitoneally injected with MC38 mouse colon cancer cells once 3 days before transplantation (pre-administration) or 3 times 5 days after transplantation (post-administration) with CD45 inhibitor 211 at 3 mg/day. kg was dosed (FIGS. 13A & B). MC38 cells were implanted intradermally into the right flank of mice at a rate of 1 ⁇ 10 6 /tumor. As a result, the tumor volume was significantly reduced in the group administered with 211 before or after MC38 transplantation, compared with the control group administered with only the vehicle (FIGS. 14A & B). These results suggest that CD45 inhibition enhances the antitumor effect of PD-1 signal inhibition.
- Balb/c (allogeneic) spleen treated with anti-PD-L1 antibody (clone 1-111A.4) (1.5 mg/kg) intraperitoneally plus mitomycin C after 9, 16 and 23 days
- Cells (5x105/mouse or 2x106/mouse) were injected (A) only once after 6 days, (B) only once after 9 days, or (C) after 9, 16 and 23 days for a total of 3 It was intravenously injected twice, and changes in tumor size over time were measured (Fig. 18). Points in each line graph are means of 5-6 mice, error bars are standard errors. * means p-value less than 0.05, ns means no significant difference (comparison by Tukey test after one-way ANOVA). As a result, it was found that three administrations of allogeneic cells had a higher effect in combination with an immune checkpoint inhibitor than one administration.
- the pharmaceutical composition of the present invention can be used as an anticancer agent, a therapeutic agent for infectious diseases, or a combination thereof.
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Veterinary Medicine (AREA)
- Pharmacology & Pharmacy (AREA)
- Epidemiology (AREA)
- Immunology (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Endocrinology (AREA)
- Pain & Pain Management (AREA)
- Mycology (AREA)
- Microbiology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Biotechnology (AREA)
- Cell Biology (AREA)
- Genetics & Genomics (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Biomedical Technology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biophysics (AREA)
- Developmental Biology & Embryology (AREA)
- Virology (AREA)
- Zoology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Abstract
Description
(1)T細胞受容体(TCR)シグナルを増強させることができる物質を含み、PD-1シグナル阻害剤を投与する前、後又は同時のいずれかの時期に投与する、医薬組成物。
(2)TCRシグナルを増強させることができる物質が、CD45阻害剤及び/又は細胞である(1)記載の医薬組成物。
(3)CD45阻害剤が、2-(4-Acetylanilino)-3-chloronaphthoquinone、N-(9,10-Dioxo-9,10-dihydrophenanthren-2-yl)-2,2-dimethylpropionamide及びそれらの類似物からなる群より選択される少なくとも1つの化合物である(2)記載の医薬組成物。
(4)細胞が、ゼノ細胞、アロ細胞又はそれらの組み合わせである(2)記載の医薬組成物。
(5)PD-1シグナル阻害剤が抗体である(1)~(4)のいずれかに記載の医薬組成物。
(6)抗体が、抗PD-1抗体、抗PD-L1抗体及び抗PD-L2抗体からなる群より選択される少なくとも1つの抗体である(5)記載の医薬組成物。
(7)抗がん剤、感染症治療剤又はそれらの組み合わせとして使用される(1)~(6)のいずれかに記載の医薬組成物。
(8)CD45阻害剤及び/又は細胞を含む、PD-1シグナル阻害活性増強剤。
(9)CD45阻害剤及び/又は細胞を含む、TCRシグナル増強剤。
(10)PD-1シグナル阻害剤を投与する前、後又は同時のいずれかの時期に、T細胞受容体(TCR)シグナルを増強させることができる物質を有効な量で被験者に投与することを含む、がん、感染症又はそれらの組み合わせを予防及び/又は治療する方法。
(11)PD-1シグナル阻害剤を投与する前、後又は同時のいずれかの時期に、T細胞受容体(TCR)シグナルを増強させることができる物質を有効な量で被験者に投与することを含む、PD-1シグナル阻害活性を増強する方法。
(12)CD45阻害剤及び/又は細胞を有効な量で被験者に投与することを含む、TCRシグナルを増強する方法。
(13)がん、感染症又はそれらの組み合わせを予防及び/又は治療することに使用するためのT細胞受容体(TCR)シグナルを増強させることができる物質であって、PD-1シグナル阻害剤を投与する前、後又は同時のいずれかの時期に投与される前記物質。
本明細書は、本願の優先権の基礎である日本国特許出願、特願2021‐31041の明細書および/または図面に記載される内容を包含する。
〔実施例1〕
[要旨]
CD8陽性細胞は、抗腫瘍反応において主要な役割を果たし、がん細胞を直接殺傷する。高齢者におけるCD8陽性細胞免疫の強い抑制は、がんや他の加齢性疾患に関係する。本研究は、加齢に伴うPD-1阻害治療耐性におけるT細胞機能の役割を検討した。我々は、高齢担癌マウスの所属リンパ節において、CD44lowCD62Llow CD8陽性T細胞の形成が低下することを見出した。はじめに、P4細胞はin vitro刺激によりナイーブCD8陽性T細胞から誘導され、次に、エフェクター/メモリー細胞へと分化した。P4細胞では、抗原特異的なT細胞の活性化やミトコンドリアの機能に重要な、1炭素(1C)代謝に関連する遺伝子の発現が特異的に高かった。この知見と一致して、1C代謝関連遺伝子の発現やミトコンドリア呼吸が若齢CD8陽性T細胞に比べて、高齢CD8陽性T細胞では低下していた。オブアルブミン(OVA)特異的なT細胞受容体(TCR)遺伝子を導入した高齢マウスでは、OVAを発現する腫瘍細胞の移植によるZAP-70の活性化は誘導されなかった。このTCRシグナル抑制は加齢CD8陽性細胞におけるCD45RB脱リン酸化酵素の発現上昇が原因であることが示唆された。驚くべき事に、非自己細胞の加齢PD-1欠損マウスへの移植による強い刺激は、CD45RB発現を正常レベルへと減少させ、P4細胞の誘導やCD8陽性T細胞における1炭素代謝酵素の発現、抗腫瘍活性を回復させた。これらの知見は、P4サブセットの誘導不全が加齢に伴うPD-1阻害耐性に関与することを示唆し、それらが、強いTCR刺激により回復可能なことを示す。
加齢は抗腫瘍免疫を抑制することが知られているにも関わらず、詳細なメカニズムはほとんど分かっていなかった。ここで我々は、前エフェクター様のT細胞であるCD44lowCD62Llow CD8陽性T細胞サブセット(P4)の形成抑制により加齢マウスにおけるPD-1阻害抗腫瘍治療に対する耐性が説明出来ることを見出した。加齢ナイーブCD8陽性T細胞におけるCD45RB発現の増加はTCRシグナルを抑制し、その結果、1C代謝遺伝子を高発現するサブセットであるP4細胞が減少する。このP4細胞の減少や抗腫瘍活性の抑制は、非自己細胞による強い免疫刺激により回復した。これらの知見は加齢に伴う抗腫瘍免疫の抑制メカニズムについての有用な洞察を提供し、高齢がん患者のための治療戦略開発のための基礎となるであろう。
加齢は多くの生理機能に影響し、その結果、多様な疾病の発症に関与する。ゲノムにおける変異の蓄積やがん細胞に対する免疫監視の低下などにより、がんの危険性は加齢に伴い増加する(1, 2)。抑制された免疫機能を再活性化する免疫治療は、がん治療のために効果的な方法として登場した。様々ながん免疫治療のうちで、PD-1/PD-L1経路を遮断する抗体は様々ながん種の患者において非常に高い有効性を示した(3-6)。しかしながら、臨床試験により多くのがん患者がPD-1阻害治療に対して反応しないことが示された(7, 8)。そのような治療は、加齢マウスモデルにおいて効果が示されず、また、同様の傾向がいくつかの臨床試験で示されている(9-12)。
加齢マウスにけるCD44lowCD62Llow(P4)CD8陽性T細胞誘導の抑制
若齢PD-1 KOマウスは強く腫瘍増殖を抑制する一方で、加齢PD-1 KOマウスでは腫瘍増殖の拒絶は見られなかった(図1A&B)。我々はまた高齢の野生型マウスがPD-1阻害腫瘍治療に対して耐性を示すことを観察した(図6A&B)。担がん有り無しのPD-1 KOマウス由来の末梢リンパ節と所属リンパ節の細胞におけるCD8陽性T細胞サブセットの比較解析によりマイナーCD8陽性T細胞サブセットであるP4のパーセンテージが若齢マウスにおいて腫瘍移植により増加し、高齢マウスにおいては増加しないことが示された(図1C&D)。他のCD8陽性T細胞における腫瘍移植による変化は若齢と高齢PD-1KOの両マウスにおいてP4サブセットの変化よりも小さかった(図1C&D)。同様の結果が野生型のマウスにおいても観察された(図6C&D)。これらの結果は、P4サブセットは腫瘍拒絶の間に誘導され、PD-1阻害又は欠損で見られる抗腫瘍活性に必要である可能性が示唆された。
P4細胞誘導の抑制が高齢マウスにおけるPD-1阻害治療耐性に関与するかどうかを調べるために、CD8陽性P4サブセットの機能的な特徴を調べた。我々ははじめに、P4細胞の抗腫瘍活性を若齢の担がんCD8 KOマウスに単離したP4細胞を移植することで検討した。TCRレパトアの偏りを防ぐために、我々はOVAペプチドに対して特異的に反応するTCRを発現するCD8陽性T細胞を保持するTCR遺伝子導入OT-1マウスを使用した。CD8陽性T細胞のP3とP4サブセットはOVAを発現するMC38腫瘍細胞(MC38-OVA)をOT-1マウスの尾静脈内に投与することにより誘導し、その5日後に脾臓から単離した。同じ数のP3又はP4細胞はMC38-OVAを移植した若齢CD8 KOマウスに養子導入された。MC38-OVA増殖はP3又はP4細胞を導入したマウスにおいて強く抑制された(図2A)。注目すべきことに、移植から6日後の末梢血において移植された全てのP4細胞はP3細胞へと分化していた(図2B)。これらの結果は、P4細胞はP3細胞へと分化することで腫瘍を拒絶する能力を保持することを示唆する。
加齢マウスにおけるP4細胞の抗腫瘍活性における役割を理解するために、我々は若齢PD-1KOマウスにおけるCD8陽性T細胞サブセット間(P1, P2, P3とP4)の遺伝子発現プロファイルを比較した。グローバルな遺伝子発現の階層的なクラスタリングはCD8陽性T細胞のそれぞれのサブセットが異なる遺伝子発現特性を持つことを示した(図3A)。活性化T細胞で発現する遺伝子は(Ctl14, Prdm1やIl2rbのような)、P1細胞よりもP4細胞でより高い発現を示した(図3B)。一方、分化したT細胞で発現する遺伝子は(Il7r, IfngやTbx21のような)、P2やP3細胞よりもP4細胞において低い発現を示した(図3B)。これらの結果は、P4細胞は、活性化しているが成熟前の状態でありP2やP3細胞のようなより分化したCD8陽性T細胞のサブセットとは異なることを示唆し、P4細胞はP1細胞由来の前エフェクター様の細胞であるという我々の仮説を支持している。
P4細胞はin vitro 刺激によりP1細胞から形成されることが示されたため、我々は高齢マウスで見られた腫瘍移植に伴うP4細胞誘導の低下は高齢P1細胞におけるTCRシグナルの非効率的な伝達により引き起こされうると考えた。TCRシグナルとP4細胞形成との間の関係を検討するために、我々はOT-1マウスにMC38-OVAを注射した。末梢リンパ節におけるP4細胞のパーセンテージは若齢マウスにおいて著しく増加したが、加齢マウスでは増加しなかった(図4A&B)。TCRシグナル伝達を評価するために、我々は若齢と加齢OT-1マウス由来のCD8陽性T細胞全体におけるTCRシグナリングに重要なZAP-70のリン酸化(40)を調べた。MC38-OVA注入は若齢マウスにおけるリン酸化ZAP-70を発現するCD8陽性T細胞の割合を増加させた(図4C)。このことは抗原刺激によるTCRシグナルの活性化は高齢OT-1マウスにおいて阻害されていることを示唆している。注目すべきことに、顕著な違いがOVA特異的なMHCテトラマーの強度において見られなかったため(図7A&B)、若齢や高齢マウスのOT-I T細胞のTCR発現レベル・結合力に違いが無い事が示された。そのため、加齢マウスで見られた抗原刺激によるTCRシグナルの活性化の阻害はTCRとMHC/抗原との相互作用以外の機構が関与するだろうと考えられる。
上記に示すように、加齢マウスにおけるP1からP4への移行の抑制が強いin vitro刺激により回復することから、我々は異種細胞(ヒトバーキットリンパ腫、Daudi細胞)を用いた強い刺激が加齢PD-1 KOマウスにおけるP4 CD8陽性T細胞の誘導を回復させることができるかどうかを調べた(図5A)。Daudi細胞の尾静脈注射から10日後(day 0: MC38腫瘍細胞移植前)、P4細胞の割合は若齢と加齢PD-1 KOマウスの末梢リンパ節において強く増加したが、P1からP3細胞の割合は有意には増加しなかった(図5B)。予想通りに、加齢PD-1 KOマウス由来のCD8陽性T細胞において、1C代謝関連遺伝子の発現レベルはDaudi細胞の注入により増加した(図5C)。これらの知見と一致して、Daudi細胞注入は加齢PD-1 KOマウス由来のP1細胞におけるCD45RBの発現を減少させた(図5D)。
高齢者や加齢動物は加齢に伴うCD8陽性T細胞反応の抑制により免疫治療に対して不応答性を示すことが知られている(9-11)。CD8陽性T細胞の腫瘍抗原に対する反応抑制のメカニズムはほとんど分かっていない。我々は、高齢担がんマウスがCD45RB発現の増加によりTCRシグナルが減弱させられているため、P4細胞が少ないことを見出した。このP4細胞の特徴はこれまでに分かっていなかった。P4サブセットはナイーブP1サブセットから分化し、そして、抗腫瘍活性に重要なエフェクター/メモリーサブセット(P3)へと成熟する(42, 43)。P4サブセットは発達や増殖細胞の生存と関係する1C代謝関連遺伝子の発現が高いという特徴を示す(29, 32, 44, 45)。これらの加齢に伴うCD8陽性T細胞の分化やPD-1阻害治療効果の阻害は非自己の細胞を用いた強い免疫刺激により回復する。
マウスと細胞
全てのマウスは京都大学大学院医学部動物実験施設又は理化学研究所生命科学研究センターにおいて、実験の障害になるような特定の病原体が存在しないSPF環境下で飼育され、適切な実験ガイドラインのもとで使用された。PD-1-/-(PD-1KO)マウスは相同組換えにより当研究室で作製したものを使用した(59)。C57BL/6野生型マウスはCharles River Laboratories Japanより入手した。OT-1 TCR遺伝子導入やCD8-/-(CD8 KO)マウスはThe Jackson Laboratory (Bar Harbor, Maine USA)から購入した(ワシントンメディカルセンターのM. B. Bevan又はトロント大学のT. Makにより寄託された)。マウス大腸がん(MC38)細胞系はJ. P. Allison、Memorial Sloan-Kettering Cancer(New York, NY)により譲渡され、オブアルブミン(OVA)発現MC38細胞はpMXs-based OVA-F2A-EGFOレトロウイルスによるMC38の形質転換により作製された。細胞は10%の非動化したウシ胎児血清および1%ペニシリン-ストレプトマイシン(Nacalai Tesque; 26253-84) を含むRPMI 1640 (Gibco; 11875-093) を用いて、マイコバクテリア感染の無い状況下で培養を行った。
MC38細胞は(5 × 105 又は 2 × 106)、マウスの右腹部の皮内に移植された(day 0)。MC38細胞移植から5日後(day 5)、それらのマウスは抗PD-L1抗体(clone 1-111A.4, 京都大学大学院医学研究科免疫ゲノム医学研究室にて作製・保管(Immunology Letters 84(2002) 57-62) (6 mg/kg)を腹腔内に投与された。この抗PD-L1抗体投与は、6日毎に3回繰り返された(day 5, 11, and 17)。抗PD-L1抗体のアイソタイプ(Rat IgG2a)はコントロールとして使用された。腫瘍増殖は測径器を使用した腫瘍サイズの計測によりモニターされ、体積は、π (縦 × 横 × 高さ)/6の式により算出した。非自己細胞の移入は腫瘍細胞移植10日前にバーキットリンパ腫Daudi細胞又はマイトマイシンC(MMC)処理をしたBALB/c又はコントロールとしてのC57BL/6N由来の脾細胞(2 × 106細胞/マウス)の尾静脈からの注射により行った
末梢や所属リンパ節細胞や腫瘍浸潤リンパ球は以前記述したように単離した(39)。脾細胞解析のために、脾臓は手早くほぐされ、赤血球を溶解させるために塩化アンモニウムカリウム溶液に懸濁された。脾細胞は洗浄され、10% ウシ胎児血清、L-グルタミン、55 μM 2-メルカプとエタノール、ペニシリン-ストレプトマイシン、1 mM ピルビン酸ナトリウム(Gibco)、1% MEM非必須アミノ酸溶液(Gibco)を含むPRMI 1640内溶液(T細胞培養液)に再懸濁された。我々はさらに、CD8陽性T細胞のサブセットをCD8a(Ly-2)マイクロビーズ(Miltenyi Biotec; 130-117-044)とFACSAria(BD Biosciences)により単離した。末梢血細胞の解析のために、集めた末梢血細胞は塩化アンモニウムカリウム溶液で処理され、T細胞培養液により洗浄された。精製された細胞は、抗CD3/28 Dynabeads(Gibco)とリコンビナントヒトIL-2(20 U/mL; PeptoTech)を加えたT細胞培養液で刺激された。MC38-OVA細胞を用いてCD8陽性T細胞を刺激するために、OT-1マウスのリンパ節から単離されたCD8陽性T細胞はT細胞培養液中でMMC処理をしたMC38-OVA細胞と共培養された。刺激から1日後に、その細胞は0.05 μM CD45阻害剤VI(EDM Millipore)で1時間、処理された。
解析のために以下の抗体を使用した。CD3 (クローン145-2C11)とCD8 (クローン53-6.7)(Invitrogen); phospho-ZAP70/Syk(クローンn3kobu5)(Invitrogen)、CD45.1(クローンA20)、CD45.2(クローン104)、CD44 (クローンIM7)と CD62L (クローンMEL-14) (TONBO biosciences又はBioLegend); CD8 (クローンKT15、MBL Life Science); CD45RB (クローン16A、BD Pharmingen)。H2Kb Negative(SIY)Tetramer-SIRYYGL-APCとH-2Kb OVA Tetramer-SINFEKL-APC(TS-M008-2)はMBL Life Scienceフローサイトメトリー解析は、FACSCant II (BD Biosciences) と FlowJo software (FlowJo LLC)を用いて行った。
P3とP4細胞を誘導するために、MC38-OVA細胞(1-2 × 106)はOT-1マウスに尾静脈注射により移入された。その5日後にP3とP4細胞はそれらのマウスから単離された。それぞれの単離されたCD8陽性T細胞は5日前に2 × 105のMC38-OVA細胞を皮内移植されたCD8 KOマウスの尾静脈より注入された。末梢血はそれから6日後に移植した細胞の分化を解析するために取得された。
トータルRNAは3から6匹を統合したトータルCD8陽性T細胞又は9匹を統合したCD8陽性T細胞サブセットからNucleospin RNA(Macherey-Nagel)を使用して生産者のプロトコールに従い抽出した。マイクロアレイ解析はマクロジェン株式会社によりマウス 8×60K v2マイクロアレイを使用して行われた。データはthe GEO repositoryに関連番号GSE161659とGSE161660として登録された(http://www.ncbi.nlm.nih.gov/geo/)。Gene ontology enrichment解析は、Annotation, Visualization, and Integrated Discovery(DAVID; http://david.abcc.ncifcrf.gov/)のためのデータベースを用いて行われた。遺伝子発現変動はR3.1.2又はRStudio version 1.1.383を用いて可視化された。
トータルRNAは細胞からNucleospin RNA(Macherey-Nagel)を使用して抽出し、製造者のマニュアルに沿ってRevatraAce逆転写酵素(Toyobo)とランダムプライマーを用いたcDNA合成のために使用された。リアルタイムPCRはthe Applied Biosystems 7500 Fast Real-Time PCT system(ABI)とPowerUp SYBR Green Master Mix(Applied Biosystems)を用いて示したmRNAを増幅するために行った。それぞれの遺伝子の発現レベルはβ-actinのmRNAレベルでノーマライズした。プライマー配列は以下の通りである:Shmt1 forward (Fw), 5’-CCAGAGTGCTGTGGCAACTC-3’(配列番号1); Shmt1 reverse (Rv), 5’-GCAAACACAGGCTGTTCCTG-3’ (配列番号2); Shmt2 Fw, 5’-GACAGTTGAGGACACCTGGC-3’ (配列番号3); Shmt2 Rv, 5’-CCAGAGAGGAGTGACATCTC-3’ (配列番号4); Phgdh Fw, 5’-TGGCCTCGGCAGAATTGGAAG-3’ (配列番号5); Phgdh Rv, 5’-TGTCATTCAGCAAGCCTGTGGT-3’ (配列番号6); Psat1 Fw, 5’-GATGAACATCCCATTTCGCATTGG-3’ (配列番号7); Psat1 Rv, 5’-GCGTTATACAGAGAGGCACGAATG-3’ (配列番号8); β-actin Fw, 5’-TAAGGCCAACCGTGAAAG-3’ (配列番号9); and β-actin Rv, 5’-GAGGCATACAGGGACAGCAC-3’ (配列番号10)。
OCRアッセイは若干の改良をし、以前の記述に従い行った(39)。所属リンパ節から単離したCD8陽性T細胞の酸素消費速度はthe Seahorse XFe96 Extracellular Flux assay kit and Seahorse XF Cell Mito Stress test kit(Agilent Technologies)を使用してXFe96 Extracellular Flux analyzer(Agilent Technologies)で解析された。CD8陽性T細胞は(3 × 105/ウェル)Cell-Tak(Corning)でコートしたXFe96プレートに播種された。予備呼吸能はOCRグラフから以前に記述したように算出された(60)。基礎呼吸は非ミトコンドリア呼吸との引き算によって(オリゴマイシン添加前の値からロテノン/アンチマイシンA添加後の値)算出された(61)。
データはPrism 7 (Graph pad)を用いて解析し、結果は平均値±標準誤差として表される。二つの群を比較する場合は、スチューデントの検定(独立2群の両側検定)を行った。二つ以上の群を比較する場合、統計的有意性は一元配置分散分析(ANOVA)を用いて複数群について評価し、次に多重比較(Tukey’s post hoc test)により差を評価した。生存率はカプラン・マイヤー法により評価し、統計的有意性はログ・ランク検定により決定された。
全てのシークエンスデータはNCBI GEOにアクセッションID GSE161659 and GSE161660として登録されている。
1. T. Finkel, M. Serrano, M. A. Blasco, The common biology of cancer and ageing. Nature 448, 767-774 (2007).
2. K. Tomihara, T. J. Curiel, B. Zhang, Optimization of immunotherapy in elderly cancer patients. Crit Rev Oncog 18, 573-583 (2013).
3. S. L. Topalian et al., Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med 366, 2443-2454 (2012).
4. A. Ribas et al., Association of Pembrolizumab With Tumor Response and Survival Among Patients With Advanced Melanoma. JAMA 315, 1600-1609 (2016).
5. M. Reck et al., Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer. N Engl J Med 375, 1823-1833 (2016).
6. P. S. Chowdhury, K. Chamoto, T. Honjo, Combination therapy strategies for improving PD-1 blockade efficacy: a new era in cancer immunotherapy. J Intern Med 283, 110-120 (2018).
7. W. Zou, J. D. Wolchok, L. Chen, PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: Mechanisms, response biomarkers, and combinations. Sci Transl Med 8, 328rv324 (2016).
8. A. L. Shergold, R. Millar, R. J. B. Nibbs, Understanding and overcoming the resistance of cancer to PD-1/PD-L1 blockade. Pharmacol Res 145, 104258 (2019).
9. J. Brahmer et al., Nivolumab versus Docetaxel in Advanced Squamous-Cell Non-Small-Cell Lung Cancer. N Engl J Med 373, 123-135 (2015).
10. C. Helissey, C. Vicier, S. Champiat, The development of immunotherapy in older adults: New treatments, new toxicities? J Geriatr Oncol 7, 325-333 (2016).
11. A. Padron et al., Age effects of distinct immune checkpoint blockade treatments in a mouse melanoma model. Exp Gerontol 105, 146-154 (2018).
12. J. Sceneay et al., Interferon Signaling Is Diminished with Age and Is Associated with Immune Checkpoint Blockade Efficacy in Triple-Negative Breast Cancer. Cancer Discov 9, 1208-1227 (2019).
13. K. Naylor et al., The influence of age on T cell generation and TCR diversity. J Immunol 174, 7446-7452 (2005).
14. M. Ahmed et al., Clonal expansions and loss of receptor diversity in the naive CD8 T cell repertoire of aged mice. J Immunol 182, 784-792 (2009).
15. Q. Qi et al., Diversity and clonal selection in the human T-cell repertoire. Proc Natl Acad Sci U S A 111, 13139-13144 (2014).
16. J. Nikolich-Zugich, G. Li, J. L. Uhrlaub, K. R. Renkema, M. J. Smithey, Age-related changes in CD8 T cell homeostasis and immunity to infection. Semin Immunol 24, 356-364 (2012).
17. M. T. Ventura, M. Casciaro, S. Gangemi, R. Buquicchio, Immunosenescence in aging: between immune cells depletion and cytokines up-regulation. Clin Mol Allergy 15, 21 (2017).
18. E. J. Wherry, R. Ahmed, Memory CD8 T-cell differentiation during viral infection. J Virol 78, 5535-5545 (2004).
19. J. C. Nolz, G. R. Starbeck-Miller, J. T. Harty, Naive, effector and memory CD8 T-cell trafficking: parallels and distinctions. Immunotherapy 3, 1223-1233 (2011).
20. J. Nikolich-Zugich, The twilight of immunity: emerging concepts in aging of the immune system. Nat Immunol 19, 10-19 (2018).
21. B. Youngblood, J. S. Hale, R. Ahmed, T-cell memory differentiation: insights from transcriptional signatures and epigenetics. Immunology 139, 277-284 (2013).
22. D. Sauce et al., Lymphopenia-driven homeostatic regulation of naive T cells in elderly and thymectomized young adults. J Immunol 189, 5541-5548 (2012).
23. N. Minato, M. Hattori, Y. Hamazaki, Physiology and pathology of T-cell aging. Int Immunol 32, 223-231 (2020).
24. J. Rossy, D. J. Williamson, K. Gaus, How does the kinase Lck phosphorylate the T cell receptor? Spatial organization as a regulatory mechanism. Front Immunol 3, 167 (2012).
25. R. J. Brownlie, R. Zamoyska, T cell receptor signalling networks: branched, diversified and bounded. Nat Rev Immunol 13, 257-269 (2013).
26. J. H. Cho et al., CD45-mediated control of TCR tuning in naive and memory CD8(+) T cells. Nat Commun 7, 13373 (2016).
27. A. C. Maue et al., T-cell immunosenescence: lessons learned from mouse models of aging. Trends Immunol 30, 301-305 (2009).
28. J. Nikolich-Zugich, Aging of the T cell compartment in mice and humans: from no naive expectations to foggy memories. J Immunol 193, 2622-2629 (2014).
29. M. Yang, K. H. Vousden, Serine and one-carbon metabolism in cancer. Nat Rev Cancer 16, 650-662 (2016).
30. G. S. Ducker, J. D. Rabinowitz, One-Carbon Metabolism in Health and Disease. Cell Metab 25, 27-42 (2017).
31. N. Ron-Harel et al., Mitochondrial Biogenesis and Proteome Remodeling Promote One-Carbon Metabolism for T Cell Activation. Cell Metab 24, 104-117 (2016).
32. E. H. Ma et al., Serine Is an Essential Metabolite for Effector T Cell Expansion. Cell Metab 25, 345-357 (2017).
33. R. J. Morscher et al., Mitochondrial translation requires folate-dependent tRNA methylation. Nature 554, 128-132 (2018).
34. D. R. Minton et al., Serine Catabolism by SHMT2 Is Required for Proper Mitochondrial Translation Initiation and Maintenance of Formylmethionyl-tRNAs. Mol Cell 69, 610-621 e615 (2018).
35. N. Ron-Harel et al., Defective respiration and one-carbon metabolism contribute to impaired naive T cell activation in aged mice. Proc Natl Acad Sci U S A 115, 13347-13352 (2018).
36. C. H. June, Principles of adoptive T cell cancer therapy. J Clin Invest 117, 1204-1212 (2007).
37. N. S. Joshi, S. M. Kaech, Effector CD8 T cell development: a balancing act between memory cell potential and terminal differentiation. J Immunol 180, 1309-1315 (2008).
38. N. E. Scharping et al., The Tumor Microenvironment Represses T Cell Mitochondrial Biogenesis to Drive Intratumoral T Cell Metabolic Insufficiency and Dysfunction. Immunity 45, 701-703 (2016).
39. K. Chamoto et al., Mitochondrial activation chemicals synergize with surface receptor PD-1 blockade for T cell-dependent antitumor activity. Proc Natl Acad Sci U S A 114, E761-E770 (2017).
40. H. Wang et al., ZAP-70: an essential kinase in T-cell signaling. Cold Spring Harb Perspect Biol 2, a002279 (2010).
41. M. Perron, H. U. Saragovi, Inhibition of CD45 Phosphatase Activity Induces Cell Cycle Arrest and Apoptosis of CD45(+) Lymphoid Tumors Ex Vivo and In Vivo. Mol Pharmacol 93, 575-580 (2018).
42. M. Sharpe, N. Mount, Genetically modified T cells in cancer therapy: opportunities and challenges. Dis Model Mech 8, 337-350 (2015).
43. R. W. Jenkins, D. A. Barbie, K. T. Flaherty, Mechanisms of resistance to immune checkpoint inhibitors. Br J Cancer 118, 9-16 (2018).
44. R. J. DeBerardinis, N. S. Chandel, Fundamentals of cancer metabolism. Sci Adv 2, e1600200 (2016).
45. N. M. Chapman, M. R. Boothby, H. Chi, Metabolic coordination of T cell quiescence and activation. Nat Rev Immunol 20, 55-70 (2020).
46. E. S. Egorov et al., The Changing Landscape of Naive T Cell Receptor Repertoire With Human Aging. Front Immunol 9, 1618 (2018).
47. C. S. Palmer et al., Emerging Role and Characterization of Immunometabolism: Relevance to HIV Pathogenesis, Serious Non-AIDS Events, and a Cure. J Immunol 196, 4437-4444 (2016).
48. H. Tsukamoto, S. Senju, K. Matsumura, S. L. Swain, Y. Nishimura, IL-6-mediated environmental conditioning of defective Th1 differentiation dampens antitumour immune responses in old age. Nat Commun 6, 6702 (2015).
49. R. A. Miller, Effect of aging on T lymphocyte activation. Vaccine 18, 1654-1660 (2000).
50. G. Pawelec, K. Hirokawa, T. Fulop, Altered T cell signalling in ageing. Mech Ageing Dev 122, 1613-1637 (2001).
51. R. L. Whisler, M. Chen, B. Liu, Y. G. Newhouse, Age-related impairments in TCR/CD3 activation of ZAP-70 are associated with reduced tyrosine phosphorylations of zeta-chains and p59fyn/p56lck in human T cells. Mech Ageing Dev 111, 49-66 (1999).
52. T. Fulop, Jr., A. Larbi, G. Dupuis, G. Pawelec, Ageing, autoimmunity and arthritis: Perturbations of TCR signal transduction pathways with ageing - a biochemical paradigm for the ageing immune system. Arthritis Res Ther 5, 290-302 (2003).
53. T. Furukawa, M. Itoh, N. X. Krueger, M. Streuli, H. Saito, Specific interaction of the CD45 protein-tyrosine phosphatase with tyrosine-phosphorylated CD3 zeta chain. Proc Natl Acad Sci U S A 91, 10928-10932 (1994).
54. K. F. Lindahl, D. B. Wilson, Histocompatibility antigen-activated cytotoxic T lymphocytes. II. Estimates of the frequency and specificity of precursors. J Exp Med 145, 508-522 (1977).
55. M. Cascalho, J. L. Platt, Xenotransplantation and other means of organ replacement. Nat Rev Immunol 1, 154-160 (2001).
56. N. Degauque, S. Brouard, J. P. Soulillou, Cross-Reactivity of TCR Repertoire: Current Concepts, Challenges, and Implication for Allotransplantation. Front Immunol 7, 89 (2016).
57. L. J. D'Orsogna, D. L. Roelen, Doxiadis, II, F. H. Claas, TCR cross-reactivity and allorecognition: new insights into the immunogenetics of allorecognition. Immunogenetics 64, 77-85 (2012).
58. M. Hebeisen et al., Molecular insights for optimizing T cell receptor specificity against cancer. Front Immunol 4, 154 (2013).
59. H. Nishimura, N. Minato, T. Nakano, T. Honjo, Immunological studies on PD-1 deficient mice: implication of PD-1 as a negative regulator for B cell responses. Int Immunol 10, 1563-1572 (1998).
60. P. S. Chowdhury, K. Chamoto, A. Kumar, T. Honjo, PPAR-Induced Fatty Acid Oxidation in T Cells Increases the Number of Tumor-Reactive CD8(+) T Cells and Facilitates Anti-PD-1 Therapy. Cancer Immunol Res 6, 1375-1387 (2018).
61. M. D. Brand, D. G. Nicholls, Assessing mitochondrial dysfunction in cells. Biochem J 435, 297-312 (2011).
[材料及び方法]
マウスと細胞
全てのマウスは京都大学大学院医学部動物実験施設において、実験の障害になるような特定の病原体が存在しないSPF環境下で飼育され、適切な実験ガイドラインのもとで使用された。C57BL/6はCharles River Laboratories Japanより入手した。PD-1-/-(PD-1KO)マウスは相同組換えにより当研究室で作製したものを使用した(西村らの文献参照)(11)。マウス大腸がん由来のMC38細胞はJ. P. Allison, Memorial Sloan-Kettering Cancer (New York, NY)から譲り受けた。ルイス肺癌由来のLLC細胞は、American Type Culture Collection(ATCC)より購入した。細胞は10%の非動化したウシ胎児血清および1%ペニシリン-ストレプトマイシン(Nacalai Tesque; 26253-84) を含むRPMI 1640 (Gibco; 11875-093) を用いて、マイコバクテリア感染の無い状況下で培養を行った。
CD45阻害(図12):2-(4-Acetylanilino)-3-chloronaphthoquinone(211) (530197, EDM Millipore) (12) (3 mg/kg)又はN-(9,10-Dioxo-9,10-dihydrophenanthren-2-yl)-2,2-dimethylpropionamide(PTP) (540215, Sigma-Aldrich) (13) (3 mg/kg)は腫瘍細胞移植の前又は後(5又は7日後)にマウスの腹腔内に投与された。MC38細胞(5 × 105 又は 1 × 106)又はLLC細胞(2 × 106)は、マウスの右腹部の皮内に移植された(day 0)。LLC細胞移植から7日後(day 7)、それらのマウスは抗PD-L1抗体(clone 1-111A.4) (2 mg/kg)を腹腔内に投与された。この抗PD-L1抗体投与は、6日毎に3回繰り返された(day 7, 13, and 19)。腫瘍体積は測径器により計測し、π (縦 × 横 × 高さ)/6の式により算出した。
データはPrism 7 (Graph pad)を用いて解析した。二つ以上の群を比較する場合、統計的有意性は一元配置分散分析(ANOVA)を用いて複数群について評価し、次に多重比較(Tukey’s post hoc test)により差を評価した。二つの群を比較する場合は、Student-T検定を行った。
本実施例では、CD45阻害剤として2-(4-Acetylanilino)-3-chloronaphthoquinone(211)又はN-(9,10-Dioxo-9,10-dihydrophenanthren-2-yl)-2,2-dimethylpropionamide(PTP)を用い、PD-1シグナル阻害との併用による腫瘍増殖抑制効果を検討した(図12)。
若齢(1~2ヶ月齢)又は高齢(14~16ヶ月齢)のPD-1 KOマウスの腹腔内にCD45阻害剤211を3 mg/kgの割合で投与した。211の投与から3日後に、MC38細胞を5×105/腫瘍の割合でマウスの右腹側に皮内移植し、腫瘍増殖を検討した。その結果、若齢PD-1 KOマウスで見られた腫瘍増殖の抑制は、高齢PD-1 KOマウスでは全く見られず、PD-1シグナル阻害耐性を示すことが明らかになった(図15A & B)。一方で、加齢マウスに211を投与した場合は、強く腫瘍増殖が抑えられた(図15A & B)。これらの結果から、CD45阻害は加齢に伴うPD-1シグナル阻害耐性を示す腫瘍の増殖を抑制させることが示された。
若齢(1~2ヶ月齢)の野生型マウスの皮内にLLC細胞を2×106/腫瘍の割合でマウスの右腹側に移植した(図16A)。その7日後から、抗PD-L1抗体単独又はCD45阻害剤(211又はPTP)との併用治療を行い、腫瘍増殖を検討した(図16A)。その結果、抗PD-L1単独治療ではほとんどみられなかった腫瘍抑制効果が、CD45阻害剤との併用治用により腫瘍増殖が有意に抑制された(図16B & C)。これら2種類のCD45阻害剤のどちらも、同様に抗PD-L1抗体との併用治療により腫瘍増殖に対する抑制効果を示した(図16B & C)。さらに、CD45阻害剤の単独治療効果を同様の条件下で検討した結果、LLC腫瘍増殖に対して有為な抑制効果を示さなかった(図16D & E)。これらの結果から、CD45阻害はPD-1シグナル阻害治療の効果を亢進させることで不応答性を示すがんの進行を抑制させる可能性が示された。
〔実施例3〕
若齢(6週齢)のC57BL/6マウスにMC38がん細胞を皮内注射した(5x10^5個)。9、16、23日後に抗PD-L1抗体(clone 1-111A.4)(1.5 mg/kg)を腹腔内投与し、それに加え、マイトマイシンCで処理したBalb/cの(同種異系)脾臓細胞(5x10^5個/マウス or 2x10^6個/マウス)を(A) 6日後に1回のみ、(B) 9日後に1回のみ、または(C) 9、16、23日後に計3回、静注し腫瘍サイズの経時的変化を測定した(図18)。各折れ線グラフにおける点は5ないし6マウスの平均値であり、エラーバーは標準誤差である。*はp値が0.05未満、n.s.は有意差が無いことを意味する(one-way ANOVA後のTukey検定による比較)。この結果、同種異系細胞を3回投与した方が1回よりも免疫チェックポイント阻害剤との併用効果が高いことがわかった。
1. S. L. Topalian et al., Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med 366, 2443-2454 (2012).
2. A. Ribas et al., Association of Pembrolizumab With Tumor Response and Survival Among Patients With Advanced Melanoma. JAMA 315, 1600-1609 (2016).
3. M. Reck et al., Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer. N Engl J Med 375, 1823-1833 (2016).
4. P. S. Chowdhury, K. Chamoto, T. Honjo, Combination therapy strategies for improving PD-1 blockade efficacy: a new era in cancer immunotherapy. J Intern Med 283, 110-120 (2018).
5. W. Zou, J. D. Wolchok, L. Chen, PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: Mechanisms, response biomarkers, and combinations. Sci Transl Med 8, 328rv324 (2016).
6. A. L. Shergold, R. Millar, R. J. B. Nibbs, Understanding and overcoming the resistance of cancer to PD-1/PD-L1 blockade. Pharmacol Res 145, 104258 (2019).
7. K. Chamoto et al., Mitochondrial activation chemicals synergize with surface receptor PD-1 blockade for T cell-dependent antitumor activity. Proc Natl Acad Sci U S A 114, E761-E770 (2017).
8. J. Rossy, D. J. Williamson, K. Gaus, How does the kinase Lck phosphorylate the T cell receptor? Spatial organization as a regulatory mechanism. Front Immunol 3, 167 (2012).
9. R. J. Brownlie, R. Zamoyska, T cell receptor signalling networks: branched, diversified and bounded. Nat Rev Immunol 13, 257-269 (2013).
10. J. H. Cho et al., CD45-mediated control of TCR tuning in naive and memory CD8(+) T cells. Nat Commun 7, 13373 (2016).
11. H. Nishimura, N. Minato, T. Nakano, T. Honjo, Immunological studies on PD-1 deficient mice: implication of PD-1 as a negative regulator for B cell responses. Int Immunol 10, 1563-1572 (1998).
12. M. D. Perron et al., Allosteric noncompetitive small molecule selective inhibitors of CD45 tyrosine phosphatase suppress T-cell receptor signals and inflammation in vivo. Mol Pharmacol 85, 553-563 (2014).
13. B. Y. Ma et al., The lectin Jacalin induces human B-lymphocyte apoptosis through glycosylation-dependent interaction with CD45. Immunology 127, 477-488 (2009).
14. S. M. Krummey et al., CD45RB Status of CD8(+) T Cell Memory Defines T Cell Receptor Affinity and Persistence. Cell Rep 30, 1282-1291 e1285 (2020).
本明細書で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書にとり入れるものとする。
プライマーの塩基配列を示す。
Claims (13)
- T細胞受容体(TCR)シグナルを増強させることができる物質を含み、PD-1シグナル阻害剤を投与する前、後又は同時のいずれかの時期に投与する、医薬組成物。
- TCRシグナルを増強させることができる物質が、CD45阻害剤及び/又は細胞である請求項1記載の医薬組成物。
- CD45阻害剤が、2-(4-Acetylanilino)-3-chloronaphthoquinone、N-(9,10-Dioxo-9,10-dihydrophenanthren-2-yl)-2,2-dimethylpropionamide及びそれらの類似物からなる群より選択される少なくとも1つの化合物である請求項2記載の医薬組成物。
- 細胞が、ゼノ細胞、アロ細胞又はそれらの組み合わせである請求項2記載の医薬組成物。
- PD-1シグナル阻害剤が抗体である請求項1~4のいずれかに記載の医薬組成物。
- 抗体が、抗PD-1抗体、抗PD-L1抗体及び抗PD-L2抗体からなる群より選択される少なくとも1つの抗体である請求項5記載の医薬組成物。
- 抗がん剤、感染症治療剤又はそれらの組み合わせとして使用される請求項1~6のいずれかに記載の医薬組成物。
- CD45阻害剤及び/又は細胞を含む、PD-1シグナル阻害活性増強剤。
- CD45阻害剤及び/又は細胞を含む、TCRシグナル増強剤。
- PD-1シグナル阻害剤を投与する前、後又は同時のいずれかの時期に、T細胞受容体(TCR)シグナルを増強させることができる物質を有効な量で被験者に投与することを含む、がん、感染症又はそれらの組み合わせを予防及び/又は治療する方法。
- PD-1シグナル阻害剤を投与する前、後又は同時のいずれかの時期に、T細胞受容体(TCR)シグナルを増強させることができる物質を有効な量で被験者に投与することを含む、PD-1シグナル阻害活性を増強する方法。
- CD45阻害剤及び/又は細胞を有効な量で被験者に投与することを含む、TCRシグナルを増強する方法。
- がん、感染症又はそれらの組み合わせを予防及び/又は治療することに使用するためのT細胞受容体(TCR)シグナルを増強させることができる物質であって、PD-1シグナル阻害剤を投与する前、後又は同時のいずれかの時期に投与される前記物質。
Priority Applications (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US18/276,583 US20240148676A1 (en) | 2021-02-26 | 2022-02-21 | Combination therapy with pd-1 signal inhibitor |
| EP22759551.9A EP4299075A4 (en) | 2021-02-26 | 2022-02-21 | COMBINATION THERAPY WITH PD-1 SIGNAL INHIBITORS |
| JP2023502370A JP7844035B2 (ja) | 2021-02-26 | 2022-02-21 | Pd-1シグナル阻害剤の併用療法 |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2021031041 | 2021-02-26 | ||
| JP2021-031041 | 2021-02-26 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2022181515A1 true WO2022181515A1 (ja) | 2022-09-01 |
Family
ID=83047998
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2022/006843 Ceased WO2022181515A1 (ja) | 2021-02-26 | 2022-02-21 | Pd-1シグナル阻害剤の併用療法 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US20240148676A1 (ja) |
| EP (1) | EP4299075A4 (ja) |
| JP (1) | JP7844035B2 (ja) |
| WO (1) | WO2022181515A1 (ja) |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2004072286A1 (ja) * | 2003-01-23 | 2004-08-26 | Ono Pharmaceutical Co., Ltd. | ヒトpd−1に対し特異性を有する物質 |
| WO2017004167A1 (en) * | 2015-07-01 | 2017-01-05 | Augusta University Research Institute, Inc. | Methods and compositions for treating cancer |
| JP2021031041A (ja) | 2019-08-29 | 2021-03-01 | ジー・オー・ピー株式会社 | キャスターおよび運搬台車 |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ITCO20120056A1 (it) | 2012-11-07 | 2014-05-08 | Nuovo Pignone Srl | Metodo per operare un compressore in caso di malfunzionamento di uno o piu' segnali di misura |
| US20190282624A1 (en) | 2015-11-30 | 2019-09-19 | Beihao Stem Cell And Regenerative Medicine Research Institute Co., Ltd. | Improved methods for reprograming non-pluripotent cells into pluripotent stem cells |
-
2022
- 2022-02-21 EP EP22759551.9A patent/EP4299075A4/en active Pending
- 2022-02-21 JP JP2023502370A patent/JP7844035B2/ja active Active
- 2022-02-21 WO PCT/JP2022/006843 patent/WO2022181515A1/ja not_active Ceased
- 2022-02-21 US US18/276,583 patent/US20240148676A1/en active Pending
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2004072286A1 (ja) * | 2003-01-23 | 2004-08-26 | Ono Pharmaceutical Co., Ltd. | ヒトpd−1に対し特異性を有する物質 |
| WO2017004167A1 (en) * | 2015-07-01 | 2017-01-05 | Augusta University Research Institute, Inc. | Methods and compositions for treating cancer |
| JP2021031041A (ja) | 2019-08-29 | 2021-03-01 | ジー・オー・ピー株式会社 | キャスターおよび運搬台車 |
Non-Patent Citations (72)
| Title |
|---|
| A. C. MAUE ET AL.: "T-cell immunosenescence: lessons learned from mouse models of aging", TRENDS IMMUNOL, vol. 30, 2009, pages 301 - 305, XP026251275, DOI: 10.1016/j.it.2009.04.007 |
| A. L. SHERGOLDR. MILLARR. J. B. NIBBS: "Understanding and overcoming the resistance of cancer to PD-1/PD-L1 blockade", PHARMACOL RES, vol. 145, 2019, pages 104258 |
| A. PADRON ET AL.: "Age effects of distinct immune checkpoint blockade treatments in a mouse melanoma model", EXP GERONTOL, vol. 105, 2018, pages 146 - 154, XP085365724, DOI: 10.1016/j.exger.2017.12.025 |
| A. RIBAS ET AL.: "Association of Pembrolizumab With Tumor Response and Survival Among Patients With Advanced Melanoma", JAMA, vol. 315, 2016, pages 1600 - 1609 |
| B. Y MA ET AL.: "The lectin Jacalin induces human B-lymphocyte apoptosis through glycosylation-dependent interaction with CD45", IMMUNOLOGY, vol. 127, 2009, pages 477 - 488 |
| B. YOUNGBLOODJ. S. HALER. AHMED: "T-cell memory differentiation: insights from transcriptional signatures and epigenetics", IMMUNOLOGY, vol. 139, 2013, pages 277 - 284, XP071276396, DOI: 10.1111/imm.12074 |
| C. H. JUNE: "Principles of adoptive T cell cancer therapy", J CLIN INVEST, vol. 117, 2007, pages 1204 - 1212, XP055761950, DOI: 10.1172/JCI31446 |
| C. HELISSEYC. VICIERS. CHAMPIAT: "The development of immunotherapy in older adults: New treatments, new toxicities?", J GERIATR ONCOL, vol. 7, 2016, pages 325 - 333, XP029748707, DOI: 10.1016/j.jgo.2016.05.007 |
| C. S. PALMER: "Emerging Role and Characterization of Immunometabolism:Relevance to HIV Pathogenesis, Serious Non-AIDS Events, and a Cure", J IMMUNOL, vol. 196, 2016, pages 4437 - 4444, XP055624585, DOI: 10.4049/jimmunol.1600120 |
| D. R. MINTON ET AL.: "Serine Catabolism by SHMT2 Is Required for Proper Mitochondrial Translation Initiation and Maintenance of Formylmethionyl-tRNAs", MOL CELL, vol. 69, 2018, pages 610 - 621 |
| D. SAUCE ET AL.: "Lymphopenia-driven homeostatic regulation of naive T cells in elderly and thymectomized young adults", J IMMUNOL, vol. 189, 2012, pages 5541 - 5548 |
| E. H. MA ET AL.: "Serine Is an Essential Metabolite for Effector T Cell Expansion", CELL METAB, vol. 25, 2017, pages 345 - 357, XP029914179, DOI: 10.1016/j.cmet.2016.12.011 |
| E. J. WHERRYR. AHMED: "Memory CD8 T-cell differentiation during viral infection", J VIROL, vol. 78, 2004, pages 5535 - 5545 |
| E. S. EGOROV ET AL.: "The Changing Landscape of Naive T Cell Receptor Repertoire With Human Aging", FRONT IMMUNOL, vol. 9, 2018, pages 1618 |
| EUR J DERMATOL, vol. 26, no. 2, 1 April 2016 (2016-04-01), pages 138 - 43 |
| FERNANDES RICARDO A.; SU LEON; NISHIGA YOKO; REN JUNMING; BHUIYAN ALADDIN M.; CHENG NING; KUO CALVIN J.; PICTON LORA K.; OHTSUKI S: "Immune receptor inhibition through enforced phosphatase recruitment", NATURE, vol. 586, no. 7831, 21 October 2020 (2020-10-21), London, pages 779 - 784, XP037282854, ISSN: 0028-0836, DOI: 10.1038/s41586-020-2851-2 * |
| G. PAWELECK. HIROKAWAT. FULOP: "Altered T cell signalling in ageing", MECH AGEING DEV, vol. 122, 2001, pages 1613 - 1637 |
| G. S. DUCKERJ. D. RABINOWITZ: "One-Carbon Metabolism in Health and Disease", CELL METAB, vol. 25, 2017, pages 27 - 42, XP029879738, DOI: 10.1016/j.cmet.2016.08.009 |
| H. NISHIMURAN. MINATOT. NAKANOT. HONJO: "Immunological studies on PD-1 deficient mice: implication of PD-1 as a negative regulator for B cell responses", INT IMMUNOL, vol. 10, 1998, pages 1563 - 1572, XP000971789, DOI: 10.1093/intimm/10.10.1563 |
| H. TSUKAMOTOS. SENJUK. MATSUMURAS. L. SWAINY. NISHIMURA: "IL-6-mediated environmental conditioning of defective Thl differentiation dampens antitumour immune responses in old age", NAT COMMUN, vol. 6, 2015, pages 6702 |
| H. WANG ET AL.: "ZAP-70: an essential kinase in T-cell signaling", COLD SPRING HARB PERSPECT BIOL, vol. 2, 2010, pages a002279, XP055392786, DOI: 10.1101/cshperspect.a002279 |
| HIROAKI IKEDA , NAOZUMI HARADA: "Possibility of cancer treatment vaccine", PHARMACOLOGY, vol. 76, no. 1, 1 January 2016 (2016-01-01), JP , pages 32 - 38, XP009539283, ISSN: 0372-7629, DOI: 10.14843/jpstj.76.32 * |
| IMMUNOLOGY LETTERS, vol. 84, 2002, pages 57 - 62 |
| J. BRAHMER ET AL.: "Nivolumab versus Docetaxel in Advanced Squamous-Cell Non-Small-Cell Lung Cancer", N ENGL J MED, vol. 373, 2015, pages 123 - 135, XP055391742, DOI: 10.1056/NEJMoa1504627 |
| J. C. NOLZG. R. STARBECK-MILLERJ. T. HARTY: "Naive, effector and memory CD8 T-cell trafficking: parallels and distinctions", IMMUNOTHERAPY, vol. 3, 2011, pages 1223 - 1233 |
| J. H. CHO ET AL.: "CD45-mediated control of TCR tuning in naive and memory CD8(+) T cells", NAT COMMUN, vol. 7, 2016, pages 13373 |
| J. NIKOLICH-ZUGICH: "Aging of the T cell compartment in mice and humans: from no naive expectations to foggy memories", J IMMUNOL, vol. 193, 2014, pages 2622 - 2629 |
| J. NIKOLICH-ZUGICH: "The twilight of immunity: emerging concepts in aging of the immune system", NAT IMMUNOL, vol. 19, 2018, pages 10 - 19, XP036582832, DOI: 10.1038/s41590-017-0006-x |
| J. NIKOLICH-ZUGICHG. LIJ. L. UHRLAUBK. R. RENKEMAM. J. SMITHEY: "Age-related changes in CD8 T cell homeostasis and immunity to infection", SEMIN IMMUNOL, vol. 24, 2012, pages 356 - 364 |
| J. ROSSYD. J. WILLIAMSONK. GAUS: "How does the kinase Lck phosphorylate the T cell receptor? Spatial organization as a regulatory mechanism", FRONT IMMUNOL, vol. 3, 2012, pages 167 |
| J. SCENEAY ET AL.: "Interferon Signaling Is Diminished with Age and Is Associated with Immune Checkpoint Blockade Efficacy in Triple-Negative Breast Cancer", CANCER DISCOV, vol. 9, 2019, pages 1208 - 1227, XP055763897, DOI: 10.1158/2159-8290.CD-18-1454 |
| K. CHAMOTO ET AL.: "Mitochondrial activation chemicals synergize with surface receptor PD-1 blockade for T cell-dependent antitumor activity", PROC NATL ACAD SCI U S A, vol. 114, 2017, pages E761 - E770, XP055616829, DOI: 10.1073/pnas.1620433114 |
| K. F. LINDAHLD. B. WILSON: "Histocompatibility antigen-activated cytotoxic T lymphocytes. II. Estimates of the frequency and specificity of precursors", J EXP MED, vol. 145, 1977, pages 508 - 522 |
| K. NAYLOR ET AL.: "The influence of age on T cell generation and TCR diversity", J IMMUNOL, vol. 174, 2005, pages 7446 - 7452 |
| K. TOMIHARAT. J. CURIELB. ZHANG: "Optimization of immunotherapy in elderly cancer patients", CRIT REV ONCOG, vol. 18, 2013, pages 573 - 583 |
| L. J. D'ORSOGNAD. L. ROELENDOXIADIS, IIF. H. CLAAS: "TCR cross-reactivity and allorecognition: new insights into the immunogenetics of allorecognition", IMMUNOGENETICS, vol. 64, 2012, pages 77 - 85 |
| M. AHMED ET AL.: "Clonal expansions and loss of receptor diversity in the naive CD8 T cell repertoire of aged mice", J IMMUNOL, vol. 182, 2009, pages 784 - 792, XP055344575, DOI: 10.4049/jimmunol.182.2.784 |
| M. CASCALHOJ. L. PLATT: "Xenotransplantation and other means of organ replacement", NAT REV IMMUNOL, vol. 1, 2001, pages 154 - 160 |
| M. D. BRANDD. G. NICHOLLS: "Assessing mitochondrial dysfunction in cells", BIOCHEM J, vol. 435, 2011, pages 297 - 312 |
| M. D. PERRON ET AL.: "Allosteric noncompetitive small molecule selective inhibitors of CD45 tyrosine phosphatase suppress T-cell receptor signals and inflammation in vivo", MOL PHARMACOL, vol. 85, 2014, pages 553 - 563 |
| M. HEBEISEN ET AL.: "Molecular insights for optimizing T cell receptor specificity against cancer", FRONT IMMUNOL, vol. 4, 2013, pages 154 |
| M. PERRONH. U. SARAGOVI: "Inhibition of CD45 Phosphatase Activity Induces Cell Cycle Arrest and Apoptosis of CD45(+) Lymphoid Tumors Ex Vivo and In Vivo", MOL PHARMACOL, vol. 93, 2018, pages 575 - 580, XP055883450, DOI: 10.1124/mol.117.110908 |
| M. RECK ET AL.: "Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer", N ENGL J MED, vol. 375, 2016, pages 1823 - 1833, XP093013864, DOI: 10.1056/NEJMoa1606774 |
| M. SHARPEN. MOUNT: "Genetically modified T cells in cancer therapy: opportunities and challenges", DIS MODEL MECH, vol. 8, 2015, pages 337 - 350 |
| M. T. VENTURA, M. CASCIARO, S. GANGEMI, R. BUQUICCHIO: "Immunosenescence in aging:between immune cells depletion and cytokines up-regulation", CLIN MOL ALLERGY, vol. 15, 2017, pages 21 |
| M. YANGK. H. VOUSDEN: "Serine and one-carbon metabolism in cancer", NAT REV CANCER, vol. 16, 2016, pages 650 - 662 |
| N. DEGAUQUES. BROUARDJ. P. SOULILLOU: "Cross-Reactivity of TCR Repertoire: Current Concepts, Challenges, and Implication for Allotransplantation", FRONT IMMUNOL, vol. 7, 2016, pages 89 |
| N. E. SCHARPING ET AL.: "The Tumor Microenvironment Represses T Cell Mitochondrial Biogenesis to Drive Intratumoral T Cell Metabolic Insufficiency and Dysfunction", IMMUNITY, vol. 45, 2016, pages 701 - 703 |
| N. M. CHAPMANM. R. BOOTHBYH. CHI: "Metabolic coordination of T cell quiescence and activation", NAT REV IMMUNOL, vol. 20, 2020, pages 55 - 70, XP036976385, DOI: 10.1038/s41577-019-0203-y |
| N. MINATOM. HATTORIY. HAMAZAKI: "Physiology and pathology of T-cell aging", INT IMMUNOL, vol. 32, 2020, pages 223 - 231 |
| N. RON-HAREL ET AL.: "Defective respiration and one-carbon metabolism contribute to impaired naive T cell activation in aged mice", PROC NATL ACAD SCI U S A, vol. 115, 2018, pages 13347 - 13352 |
| N. RON-HAREL: "Mitochondrial Biogenesis and Proteome Remodeling Promote One-Carbon Metabolism for T Cell Activation", CELL METAB, vol. 24, 2016, pages 104 - 117, XP029638760, DOI: 10.1016/j.cmet.2016.06.007 |
| N. S. JOSHIS. M. KAECH: "Effector CD8 T cell development: a balancing act between memory cell potential and terminal differentiation", J IMMUNOL, vol. 180, 2008, pages 1309 - 1315 |
| NAKAJIMA YUKA, CHAMOTO KENJI, OURA TAKUMA, HONJO TASUKU: "Critical role of the CD44 low CD62L low CD8 + T cell subset in restoring antitumor immunity in aged mice", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES, vol. 118, no. 23, 8 June 2021 (2021-06-08), pages 1 - 10, XP055961621, ISSN: 0027-8424, DOI: 10.1073/pnas.2103730118 * |
| P. S. CHOWDHURYK. CHAMOTOA. KUMART. HONJO: "PPAR-Induced Fatty Acid Oxidation in T Cells Increases the Number of Tumor-Reactive CD8(+) T Cells and Facilitates Anti-PD-1 Therapy", CANCER IMMUNOL RES, vol. 6, 2018, pages 1375 - 1387, XP055956806, DOI: 10.1158/2326-6066.CIR-18-0095 |
| P. S. CHOWDHURYK. CHAMOTOT. HONJO: "Combination therapy strategies for improving PD-1 blockade efficacy: a new era in cancer immunotherapy", J INTERN MED, vol. 283, 2018, pages 110 - 120, XP055703710, DOI: 10.1111/joim.12708 |
| Q. QI ET AL.: "Diversity and clonal selection in the human T-cell repertoire", PROC NATL ACAD SCI U S A, vol. 111, 2014, pages 13139 - 13144, XP055568721, DOI: 10.1073/pnas.1409155111 |
| R. A. MILLER: "Effect of aging on T lymphocyte activation", VACCINE, vol. 18, 2000, pages 1654 - 1660, XP004189055, DOI: 10.1016/S0264-410X(99)00502-2 |
| R. J. BROWNLIER. ZAMOYSKA: "T cell receptor signaling networks: branched, diversified and bounded", NAT REV IMMUNOL, vol. 13, 2013, pages 257 - 269, XP093005882, DOI: 10.1038/nri3403 |
| R. J. BROWNLIER. ZAMOYSKA: "T cell receptor signalling networks: branched, diversified and bounded", NAT REV IMMUNOL, vol. 13, 2013, pages 257 - 269, XP093005882, DOI: 10.1038/nri3403 |
| R. J. DEBERARDINISN. S. CHANDEL: "Fundamentals of cancer metabolism", SCI ADV, vol. 2, 2016, pages e 1600200 |
| R. J. MORSCHER ET AL.: "Mitochondrial translation requires folate-dependent tRNA methylation", NATURE, vol. 554, 2018, pages 128 - 132, XP037473713, DOI: 10.1038/nature25460 |
| R. L. WHISLERM. CHENB. LIUY. G. NEWHOUSE: "Age-related impairments in TCR/CD3 activation of ZAP-70 are associated with reduced tyrosine phosphorylations of zeta-chains and p59fyn/p561ck in human T cells", MECH AGEING DEV, vol. 111, 1999, pages 49 - 66 |
| R. W. JENKINSD. A. BARBIEK. T. FLAHERTY: "Mechanisms of resistance to immune checkpoint inhibitors", BR J CANCER, vol. 118, 2018, pages 9 - 16 |
| S. L. TOPALIAN ET AL.: "Safety, activity, and immune correlates of anti-PD-1 antibody in cancer", N ENGL J MED, vol. 366, 2012, pages 2443 - 2454, XP055098235, DOI: 10.1056/NEJMoa1200690 |
| S. M. KRUMMEY ET AL.: "CD45RB Status of CD8(+) T Cell Memory Defines T Cell Receptor Affinity and Persistence", CELL REP, vol. 30, 2020, pages 1282 - 1291 |
| See also references of EP4299075A4 |
| T. FINKELM. SERRANOM. A. BLASCO: "The common biology of cancer and ageing", NATURE, vol. 448, 2007, pages 767 - 774, XP037798393, DOI: 10.1038/nature05985 |
| T. FULOP, JR., A. LARBI, G. DUPUIS, G. PAWELEC: "Ageing, autoimmunity and arthritis:Perturbations of TCR signal transduction pathways with ageing - a biochemical paradigm for the ageing immune system", ARTHRITIS RES THER, vol. 5, 2003, pages 290 - 302 |
| T. FURUKAWAM. ITOHN. X. KRUEGERM. STREULIH. SAITO: "Specific interaction of the CD45 protein-tyrosine phosphatase with tyrosine-phosphorylated CD3 zeta chain", PROC NATL ACAD SCI U S A, vol. 91, 1994, pages 10928 - 10932, XP055171132, DOI: 10.1073/pnas.91.23.10928 |
| W. ZOU, J. D. WOLCHOK, L. CHEN: "PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: Mechanisms, response biomarkers, and combinations", SCI TRANSL MED, vol. 8, 2016, pages 328rv324 |
| YUKA NAKAJIMA: "PD-1 inhibitory treatment resistance mechanism with aging mediated by a decrease in specific CD8 positive T cell subpopulation", ABSTRACTS OF THE 78TH ANNUAL MEETING OF THE JAPANESE CANCER ASSOCIATION; SEPTEMBER 26 TO 28, 2019, 1 January 2019 (2019-01-01), JP, pages 806, XP009539392 * |
Also Published As
| Publication number | Publication date |
|---|---|
| JP7844035B2 (ja) | 2026-04-13 |
| EP4299075A1 (en) | 2024-01-03 |
| JPWO2022181515A1 (ja) | 2022-09-01 |
| EP4299075A4 (en) | 2025-02-19 |
| US20240148676A1 (en) | 2024-05-09 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Hossain et al. | Reinvigorating exhausted CD8+ cytotoxic T lymphocytes in the tumor microenvironment and current strategies in cancer immunotherapy | |
| ES2981569T3 (es) | Métodos de tratamiento usando terapia celular adoptiva | |
| Riccaboni et al. | Spleen tyrosine kinases: biology, therapeutic targets and drugs | |
| EP1897548B2 (en) | T cell regulation | |
| ES2643887T3 (es) | Combinación de anticuerpos anti-KIR y anticuerpos anti-PD-1 para tratar el cáncer | |
| Ashino et al. | Janus kinase 1/3 signaling pathways are key initiators of TH2 differentiation and lung allergic responses | |
| US20210301020A1 (en) | Combination of lilrb1/2 pathway inhibitors and pd-1 pathway inhibitors | |
| EP3184117B1 (en) | Compositions and their use in the treatment of cancer | |
| CA3205637A1 (en) | Treatment of cancer with nk cells and a cd20 targeted antibody | |
| ES2777227T3 (es) | CD33 soluble para tratar síndromes mielodisplásicos (MDS) | |
| CN104853776A (zh) | 免疫应答的增强 | |
| AU2016270996A1 (en) | Methods to induce conversion of regulatory T cells into effector T cells for cancer immunotherapy | |
| US20240262909A1 (en) | Tim-3 modulates anti-tumor immunity by regulating inflammasome activation | |
| Benson et al. | Modeling the clinical phenotype of BTK inhibition in the mature murine immune system | |
| KR20210035805A (ko) | 세포후 신호전달 인자의 조절을 통한 면역 활성의 증가 | |
| JP2023500334A (ja) | がん治療のためのマイクロバイオータ及び代謝物の力の活用 | |
| JP2025527534A (ja) | 抗-ヒト表皮成長因子受容体2(her2)キメラ抗原受容体(car)を含むナチュラルキラー細胞を投与する方法 | |
| US20230173095A1 (en) | Living cells engineered with polyphenol-functionalized biologically active nanocomplexes | |
| JP7844035B2 (ja) | Pd-1シグナル阻害剤の併用療法 | |
| EP4492059A2 (en) | Methods of modulating antigenicity to enhance recognition by t-cells | |
| US20250381272A1 (en) | Treatment methods for second line therapy of cd19-targeted car t cells | |
| Rhee | Regulation of T follicular helper cells in the germinal center reaction of chronic graft-versus-host disease | |
| Ghobakhloo | Investigating the physiological role of leupaxin in the function of CD8+ T cells and their contribution to antitumor immunity | |
| Freitas | Regulation of Immune Cell Activation and Function by the nBMPp2 Protein and the CD5 Co-Receptor | |
| Han | The Role of the Tumor Microenvironment in Waldenström Macroglobulinemia |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 22759551 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2023502370 Country of ref document: JP Kind code of ref document: A |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2022759551 Country of ref document: EP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2022759551 Country of ref document: EP Effective date: 20230926 |