WO2022224203A1 - Antibodies that bind sars-cov-2 spike protein - Google Patents
Antibodies that bind sars-cov-2 spike protein Download PDFInfo
- Publication number
- WO2022224203A1 WO2022224203A1 PCT/IB2022/053756 IB2022053756W WO2022224203A1 WO 2022224203 A1 WO2022224203 A1 WO 2022224203A1 IB 2022053756 W IB2022053756 W IB 2022053756W WO 2022224203 A1 WO2022224203 A1 WO 2022224203A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- seq
- antibody
- cov
- coronavirus
- sars
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—RNA viruses
- C07K16/102—Coronaviridae (F)
- C07K16/104—Severe acute respiratory syndrome coronavirus 2 [SARS‐CoV‐2]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/50—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates
- A61K47/51—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent
- A61K47/68—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent the modifying agent being an antibody, an immunoglobulin or a fragment thereof, e.g. an Fc-fragment
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/569—Immunoassay; Biospecific binding assay; Materials therefor for microorganisms, e.g. protozoa, bacteria, viruses
- G01N33/56983—Viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/54—Medicinal preparations containing antigens or antibodies characterised by the route of administration
- A61K2039/541—Mucosal route
- A61K2039/544—Mucosal route to the airways
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/22—Immunoglobulins specific features characterized by taxonomic origin from camelids, e.g. camel, llama or dromedary
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/569—Single domain, e.g. dAb, sdAb, VHH, VNAR or nanobody®
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/60—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments
- C07K2317/64—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments comprising a combination of variable region and constant region components
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/94—Stability, e.g. half-life, pH, temperature or enzyme-resistance
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/005—Assays involving biological materials from specific organisms or of a specific nature from viruses
- G01N2333/08—RNA viruses
- G01N2333/165—Coronaviridae, e.g. avian infectious bronchitis virus
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2469/00—Immunoassays for the detection of microorganisms
- G01N2469/10—Detection of antigens from microorganism in sample from host
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Definitions
- coronavirus spike polypeptide particularly the spike polypeptide of SARS-CoV-2 and variants thereof, and to the use of such antibodies for various applications including the detection of a coronavirus and/or treatment or prevention of a coronavirus infection.
- Coronavirus is a single-stranded enveloped RNA virus belonging to the subfamily Coronavirinae in the order Nidovirales.
- coronaviruses Based on genomic structure, coronaviruses have been classified into four genera; Alphacoronavirus, Betacoronavirus, Gammacoronavirus, and Deltacoronavirus; two of which (alphacoronaviruses and betacoronaviruses) infect mammals. Seven coronaviruses are known to cause human disease: HCoV 229E, HCov OC43, HCoVNL63, HCoVHKU1, SARS-CoV, MERS-CoV, and SARS-CoV-2. Three coronaviruses, SARS-CoV, MERS-CoV, and SARS-CoV-2, cause serious illness in humans, whereas the remaining four human coronaviruses are associated with mild illness.
- coronavirus spike protein which is a homotrimeric glycoprotein.
- the spike polypeptide includes three segments, an ectodomain, a single-pass transmembrane anchor, and an intracellular tail.
- the spike ectodomain is made up of a receptor-binding subunit (S1) and a membrane-fusion subunit (S2).
- S1 includes two major domains, an N-terminal domain (NTD) and a C-terminal domain (CTD), which is also known as the receptor binding domain (RBD).
- S1 contains two subdomains (SD1 and S1-SD2) as described in Lan et al., 2020.
- S1 binds to a host cell surface receptor and S2 fuses the host and viral membranes (Li, 2016).
- the host cell surface receptor bound by both SARS-CoV and SARS-CoV- 2 is a zinc peptidase angiotensin-converting enzyme 2 (ACE2), whereas MERS-CoV recognizes a serine peptidase (DPP4) (Li, 2016; Zhou et al, 2020).
- ACE2 zinc peptidase angiotensin-converting enzyme 2
- DPP4 serine peptidase
- the receptor binding domain (RBD) of SARS-CoV-2 has been characterized and the binding mode of the SARS-CoV-2 RBD to ACE2 has been found to be nearly identical to that observed for SARS-CoV (Lan et al., 2020).
- RBD receptor binding domain
- Antibodies that neutralize coronaviruses have significant potential as therapeutic agents. Further antibodies with high affinity for coronaviruses, such as SARS-CoV-2, may allow for detection, quantification, or capture of coronaviruses with high sensitivity and specificity.
- an isolated or purified antibody that specifically recognizes at least one coronavirus spike polypeptide, wherein the antibody comprises an antigen binding portion of an antibody heavy chain, wherein the antigen binding portion comprises a first complementarity determining region (CDR1), a second complementarity determining region (CDR2), and a third complementarity determining region (CDR3), and wherein CDR1, CDR2, and CDR3, respectively, comprise the amino acid sequence set forth in: SEQ ID NO: 1, SEQ ID NO: 45, and SEQ ID NO: 90; SEQ ID NO: 2, SEQ ID NO: 45, and SEQ ID NO: 91; SEQ ID NO: 1, SEQ ID NO: 46, and SEQ ID NO: 92; SEQ ID NO: 3, SEQ ID NO: 47, and SEQ ID NO: 93; SEQ ID NO: 4, SEQ ID NO: 48, and SEQ ID NO: 94; SEQ ID NO: 5, SEQ ID NO: 49, and SEQ ID NO: 95; SEQ ID NO: 1, SEQ ID NO:
- the antibody is a neutralizing antibody and CDR1, CDR2, and CDR3, respectively, comprise the amino acid sequence set forth in: SEQ ID NO: 1, SEQ ID NO: 45, and SEQ ID NO: 90; SEQ ID NO: 1, SEQ ID NO: 46, and SEQ ID NO: 92; SEQ ID NO: 2, SEQ ID NO: 45, and SEQ ID NO: 91; SEQ ID NO: 3, SEQ ID NO: 47, and SEQ ID NO: 93; SEQ ID NO: 4, SEQ ID NO: 48, and SEQ ID NO: 94; SEQ ID NO: 5, SEQ ID NO: 49, and SEQ ID NO: 95; SEQ ID NO: 6, SEQ ID NO: 50, and SEQ ID NO: 96; SEQ ID NO: 7, SEQ ID NO: 51, and SEQ ID NO: 97; SEQ ID NO: 8, SEQ ID NO: 52, and SEQ ID NO: 98; SEQ ID NO: 10, SEQ ID NO: 54, and SEQ ID NO: 100;
- the antibody specifically binds the S1-NTD domain of the coronavirus spike polypeptide and CDR1, CDR2, and CDR3, respectively, comprise the amino acid sequence set forth in: SEQ ID NO: 21, SEQ ID NO: 65, and SEQ ID NO: 111; SEQ ID NO: 22, SEQ ID NO: 66, and SEQ ID NO: 112; SEQ ID NO: 23, SEQ ID NO: 67, and SEQ ID NO: 113; SEQ ID NO: 24, SEQ ID NO: 68, and SEQ ID NO: 114; SEQ ID NO: 25, SEQ ID NO: 69, and SEQ ID NO: 115; SEQ ID NO: 26, SEQ ID NO: 70, and SEQ ID NO: 116; SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 126; SEQ ID NO: 37, SEQ ID NO: 82, and SEQ ID NO: 128; or SEQ ID NO: 38, SEQ ID NO: 83, and SEQ
- the antibody specifically binds the S2 subunit of the coronavirus spike polypeptide and CDR1, CDR2, and CDR3, respectively, comprise the amino acid sequence set forth in: SEQ ID NO: 27, SEQ ID NO: 71, and SEQ ID NO: 117; SEQ ID NO: 28, SEQ ID NO: 72, and SEQ ID NO: 118; SEQ ID NO: 29, SEQ ID NO: 73, and SEQ ID NO: 119; SEQ ID NO: 30, SEQ ID NO: 74, and SEQ ID NO: 120; SEQ ID NO: 31, SEQ ID NO: 75, and SEQ ID NO: 121; SEQ ID NO: 32, SEQ ID NO: 76, and SEQ ID NO: 122; SEQ ID NO: 33, SEQ ID NO: 77, and SEQ ID NO: 123; SEQ ID NO: 34, SEQ ID NO: 78, and SEQ ID NO: 124; SEQ ID NO: 39, SEQ ID NO: 84, and SEQ ID NO:
- the antibody specifically binds the S1-RBD domain of the coronavirus spike polypeptide and CDR1, CDR2, and CDR3, respectively, comprise the amino acid sequence set forth in: SEQ ID NO: 1, SEQ ID NO: 45, and SEQ ID NO: 90; SEQ ID NO: 2, SEQ ID NO: 45, and SEQ ID NO: 91; SEQ ID NO: 1, SEQ ID NO: 46, and SEQ ID NO: 92; SEQ ID NO: 3, SEQ ID NO: 47, and SEQ ID NO: 93; SEQ ID NO: 4, SEQ ID NO: 48, and SEQ ID NO: 94; SEQ ID NO: 5, SEQ ID NO: 49, and SEQ ID NO: 95; SEQ ID NO: 6, SEQ ID NO: 50, and SEQ ID NO: 96; SEQ ID NO: 7, SEQ ID NO: 51, and SEQ ID NO: 97; SEQ ID NO: 8, SEQ ID NO: 52, and SEQ ID NO: 98; SEQ ID NO: 9,
- the antibody is cross-reactive with the spike polypeptide of SARS- CoV-2 and SARS-CoV, and CDR1, CDR2, and CDR3, respectively, comprise the amino acid sequence set forth in: SEQ ID NO: 1, SEQ ID NO: 45, and SEQ ID NO: 92; SEQ ID NO: 9, SEQ ID NO: 53, and SEQ ID NO: 99; SEQ ID NO: 21, SEQ ID NO: 65, and SEQ ID NO: 111; SEQ ID NO: 22, SEQ ID NO: 66, and SEQ ID NO: 112; SEQ ID NO: 8, SEQ ID NO: 52, and SEQ ID NO: 98; SEQ ID NO: 11, SEQ ID NO: 55, and SEQ ID NO: 101; SEQ ID NO: 13, SEQ ID NO: 57, and SEQ ID NO: 103; SEQ ID NO: 18, SEQ ID NO: 62, and SEQ ID NO: 108; SEQ ID NO: 19, SEQ ID NO: 63, and SEQ ID NO:
- the antibody recognizes a linear epitope, and CDR1, CDR2, and CDR3, respectively, comprise the amino acid sequence set forth in: SEQ ID NO: 21, SEQ ID NO: 65, and SEQ ID NO: 111; SEQ ID NO: 24, SEQ ID NO: 68, and SEQ ID NO: 114; SEQ ID NO: 27, SEQ ID NO: 71, and SEQ ID NO: 117; SEQ ID NO: 28, SEQ ID NO: 72, and SEQ ID NO: 118; SEQ ID NO: 29, SEQ ID NO: 73, and SEQ ID NO: 119; SEQ ID NO: 31, SEQ ID NO: 75, and SEQ ID NO: 121; SEQ ID NO: 32, SEQ ID NO: 76, and SEQ ID NO: 122; SEQ ID NO: 30, SEQ ID NO: 74, and SEQ ID NO: 120; SEQ ID NO: 39, SEQ ID NO: 84, and SEQ ID NO: 130; SEQ ID NO: 40, SEQ ID NO: 27, SEQ ID NO
- the antibody comprises the amino acid sequence set forth in SEQ ID NO: 183.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 21, SEQ ID NO: 65, and SEQ ID NO: 111; SEQ ID NO: 23, SEQ ID NO: 67, and SEQ ID NO: 113; or SEQ ID NO: 24, SEQ ID NO: 68, and SEQ ID NO: 114.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 38, SEQ ID NO: 83, and SEQ ID NO: 129.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 126 or SEQ ID NO: 37, SEQ ID NO: 82, and SEQ ID NO: 128.
- the antibody comprises the amino acid sequence set forth in SEQ ID NO: 184.
- the antibody comprises the amino acid sequence set forth in SEQ ID NO: 158, SEQ ID NO: 157, SEQ ID NO: 172, SEQ ID NO: 175, SEQ ID NO: 176, SEQ ID NO: 160, SEQ ID NO: 161, SEQ ID NO: 159, or SEQ ID NO: 162, or an amino acid sequence having at least 75%, at least 80%, at least 85%, at least 90%, at least 95%, at least 96%, at least 97%, at least 98%, or at least 99% sequence identity to the full length of the amino acid sequence set forth in SEQ ID NO: 158, SEQ ID NO: 157, SEQ ID NO: 172, SEQ ID NO: 175, SEQ ID NO: 176, SEQ ID NO: 160, SEQ ID NO: 161, SEQ ID NO: 159, and/or SEQ ID NO: 162.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 31, SEQ ID NO: 75, and SEQ ID NO: 121; SEQ ID NO: 40, SEQ ID NO: 85, and SEQ ID NO: 131; SEQ ID NO: 41, SEQ ID NO: 86, and SEQ ID NO: 132; SEQ ID NO: 42, SEQ ID NO: 87, and SEQ ID NO: 133; SEQ ID NO: 43, SEQ ID NO: 88, and SEQ ID NO: 134; or SEQ ID NO: 44, SEQ ID NO: 89, and SEQ ID NO: 135.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in SEQ ID NO: 33, SEQ ID NO: 77, and SEQ ID NO: 123.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in SEQ ID NO: 32, SEQ ID NO: 76, and SEQ ID NO: 122.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in SEQ ID NO: 27, SEQ ID NO: 71, and SEQ ID NO: 117.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in SEQ ID NO: 28, SEQ ID NO: 72, and SEQ ID NO: 118.
- the antibody comprises the amino acid sequence set forth in SEQ ID NO: 185.
- the antibody comprises the amino acid sequence set forth in SEQ ID NO: 180, SEQ ID NO: 182, SEQ ID NO: 166, SEQ ID NO: 177, SEQ ID NO: 179, SEQ ID NO: 163, SEQ ID NO: 164, SEQ ID NO: 167, SEQ ID NO: 170, SEQ ID NO: 168, SEQ ID NO: 169, SEQ ID NO: 181, SEQ ID NO: 165, or SEQ ID NO: 178, or an amino acid sequence having at least 75%, at least 80%, at least 85%, at least 90%, at least 95%, at least 96%, at least 97%, at least 98%, or at least 99% sequence identity to the full length of the amino acid sequence set forth in SEQ ID NO: 180, SEQ ID NO: 182, SEQ ID NO: 166, SEQ ID NO: 177, SEQ ID NO: 179, SEQ ID NO: 163, SEQ ID NO: 164, SEQ ID NO: 167, SEQ ID NO: 180, SEQ ID
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 5, SEQ ID NO: 49, and SEQ ID NO: 95; SEQ ID NO: 14, SEQ ID NO: 58, and SEQ ID NO: 104; SEQ ID NO: 16, SEQ ID NO: 60, and SEQ ID NO: 106; SEQ ID NO: 10, SEQ ID NO: 54, and SEQ ID NO: 100; SEQ ID NO: 15, SEQ ID NO: 59, and SEQ ID NO: 105; or SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 127.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 3, SEQ ID NO: 47, and SEQ ID NO: 93; SEQ ID NO: 6, SEQ ID NO: 50, and SEQ ID NO: 96; SEQ ID NO: 14, SEQ ID NO: 58, and SEQ ID NO: 104; SEQ ID NO: 10, SEQ ID NO: 54, and SEQ ID NO: 100; SEQ ID NO: 15, SEQ ID NO: 59, and SEQ ID NO: 105; or SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 127.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 3, SEQ ID NO: 47, and SEQ ID NO: 93; SEQ ID NO: 6, SEQ ID NO: 50, and SEQ ID NO: 96; SEQ ID NO: 16, SEQ ID NO: 60, and SEQ ID NO: 106; SEQ ID NO: 10, SEQ ID NO: 54, and SEQ ID NO: 100; SEQ ID NO: 15, SEQ ID NO: 59, and SEQ ID NO: 105; or SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 127.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in SEQ ID NO: 7, SEQ ID NO: 51, and SEQ ID NO: 97.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 4, SEQ ID NO: 48, and SEQ ID NO: 94; SEQ ID NO: 9, SEQ ID NO: 53, and SEQ ID NO: 99; SEQ ID NO: 11, SEQ ID NO: 55, and SEQ ID NO: 101; or SEQ ID NO: 12, SEQ ID NO: 56, and SEQ ID NO: 102.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 3, SEQ ID NO: 47, and SEQ ID NO: 93; SEQ ID NO: 6, SEQ ID NO: 50, and SEQ ID NO: 96; SEQ ID NO: 10, SEQ ID NO: 54, and SEQ ID NO: 100; SEQ ID NO: 15, SEQ ID NO: 59, and SEQ ID NO: 105; or SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 127.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 10, SEQ ID NO: 54, and SEQ ID NO: 100; SEQ ID NO: 15, SEQ ID NO: 59, and SEQ ID NO: 105; or SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 127.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 14, SEQ ID NO: 58, and SEQ ID NO: 104; SEQ ID NO: 10, SEQ ID NO: 54, and SEQ ID NO: 100; SEQ ID NO: 15, SEQ ID NO: 59, and SEQ ID NO: 105; or SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 127.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 16, SEQ ID NO: 60, and SEQ ID NO: 106; SEQ ID NO: 10, SEQ ID NO: 54, and SEQ ID NO: 100; SEQ ID NO: 15, SEQ ID NO: 59, and SEQ ID NO: 105; or SEQ ID NO: 20, SEQ ID NO: 81, and SEQ ID NO: 127.
- CDR1, CDR2, and CDR3, respectively comprise the amino acid sequence set forth in: SEQ ID NO: 1, SEQ ID NO: 45, and SEQ ID NO: 90; SEQ ID NO: 1, SEQ ID NO: 46, and SEQ ID NO: 92; SEQ ID NO: 2, SEQ ID NO: 45, and SEQ ID NO: 91; SEQ ID NO: 8, SEQ ID NO: 52, and SEQ ID NO: 98; SEQ ID NO: 13, SEQ ID NO: 57, and SEQ ID NO: 103; SEQ ID NO: 17, SEQ ID NO: 61, and SEQ ID NO: 107; SEQ ID NO: 19, SEQ ID NO: 63, and SEQ ID NO: 109; SEQ ID NO: 35, SEQ ID NO: 79, and SEQ ID NO: 125; or SEQ ID NO: 36, SEQ ID NO: 80, and SEQ ID NO: 125.
- the antibody comprises the amino acid sequence set forth in SEQ ID NO: 186.
- the antibody comprises the amino acid sequence set forth in SEQ ID NO: 171, SEQ ID NO: 173, SEQ ID NO: 149, SEQ ID NO: 155, SEQ ID NO: 147, SEQ ID NO: 148, SEQ ID NO: 139, SEQ ID NO: 142, SEQ ID NO: 154, SEQ ID NO: 144, SEQ ID NO: 145, SEQ ID NO: 156, SEQ ID NO: 174, SEQ ID NO: 137, SEQ ID NO: 136, SEQ ID NO: 138, SEQ ID NO: 152, SEQ ID NO: 153, SEQ ID NO: 140, SEQ ID NO: 143, SEQ ID NO: 141, SEQ ID NO: 146, SEQ ID NO: 150, or SEQ ID NO: 151, or an amino acid sequence having at least 75%, at least 80%, at least 85%, at
- the antibody is a single domain antibody. In a further embodiment, the antibody is a V H H. [0040] In an embodiment the antibody is of camelid origin. [0041] In an embodiment, the antibody is in a multivalent display format. In a further embodiment, the antibody is linked to an Fc fragment. In a further embodiment, the Fc-linked antibody is in a bivalent display format. [0042] In an embodiment of the antibody, the at least one coronavirus spike polypeptide specifically binds an ACE2 receptor. [0043] In an embodiment of the antibody, the at least one coronavirus spike polypeptide comprises a SARS-CoV-2 spike polypeptide.
- the at least one coronavirus spike polypeptide is comprised within a homotrimer.
- an antibody cocktail composition comprising two or more of the antibodies as described herein.
- the composition may comprise two, three, four, five, or more different antibodies as described herein.
- the antibody cocktail composition may further comprise a pharmaceutically acceptable carrier and/or diluent.
- Another embodiment is a nucleic acid molecule encoding an antibody as described herein.
- a further embodiment is a vector comprising the nucleic acid molecule. In an embodiment of the vector, the nucleic acid molecule is operably linked to at least one promoter and/or regulatory element to enable expression in a host cell.
- An additional embodiment is a host cell comprising the vector.
- Another embodiment is a pharmaceutical composition comprising at least one antibody as defined herein and a pharmaceutically acceptable carrier and/or diluent. In an embodiment, the pharmaceutical composition is for delivery by inhalation or nebulization.
- Another embodiment is a composition comprising at least one antibody as defined herein, linked to another molecule. In an embodiment, the other molecule is a label or polypeptide. In an embodiment, the other molecule is an ACE2 polypeptide or a fragment thereof.
- Another embodiment is a composition or apparatus comprising at least one antibody as defined herein immobilized on a substrate.
- a further embodiment is a method for capturing a coronavirus or a coronavirus spike polypeptide or fragment thereof from a sample, the method comprising exposing the sample to the composition or apparatus.
- the coronavirus is a coronavirus that specifically binds an ACE2 receptor or the coronavirus spike polypeptide is a coronavirus spike polypeptide that specifically binds an ACE2 receptor.
- the coronavirus is SARS-CoV-2 or SARS-CoV.
- Another embodiment is use of an antibody as described herein to treat or detect a coronavirus infection.
- the coronavirus infection is caused by at least one coronavirus that specifically binds an ACE2 receptor.
- the coronavirus infection is caused by SARS-CoV-2 and/or SARS-CoV.
- Another embodiment is use of an antibody or composition as described herein to detect, quantify and/or capture a coronavirus; or to detect, quantify and/or capture a coronavirus spike polypeptide or fragment thereof.
- the coronavirus is a coronavirus that specifically binds an ACE2 receptor or the coronavirus spike polypeptide is a coronavirus spike polypeptide that specifically binds an ACE2 receptor.
- the coronavirus is SARS-CoV-2 or SARS-CoV.
- Another embodiment is a method for treating or preventing a coronavirus infection, the method comprising administering at least one antibody or composition as described herein to a subject in need thereof.
- the coronavirus infection is caused by at least one coronavirus that specifically binds an ACE2 receptor.
- the coronavirus infection is caused by SARS-CoV-2 and/or SARS-CoV.
- the administration is by inhalation or nebulization.
- Another embodiment is a method for detecting the presence of a coronavirus or a coronavirus spike polypeptide or fragment thereof in a sample, the method comprising exposing the sample to at least one antibody or composition as described herein and assaying for specific binding between the at least one antibody and the sample, wherein specific binding indicates a presence of the at least one coronavirus or coronavirus spike polypeptide or fragment thereof in the sample.
- the coronavirus is a coronavirus that specifically binds an ACE2 receptor or the coronavirus spike polypeptide is a coronavirus spike polypeptide that specifically binds an ACE2 receptor.
- coronavirus is SARS-CoV-2 or SARS-CoV
- the coronavirus spike polypeptide or fragment thereof is a SARS-CoV-2 or SARS-CoV coronavirus spike polypeptide or fragment thereof.
- Another embodiment is an antibody or composition as described herein for use to detect or treat a coronavirus infection.
- the coronavirus infection is caused by at least one coronavirus that specifically binds an ACE2 receptor.
- the at least one coronavirus is SARS-CoV-2 and/or SARS-CoV.
- Another embodiment is an antibody composition as described herein for use to detect, quantify and/or capture a coronavirus; or to detect, quantify and/or capture a coronavirus spike polypeptide or fragment thereof.
- the coronavirus is a coronavirus that specifically binds an ACE2 receptor, or the coronavirus spike polypeptide is a coronavirus spike polypeptide that specifically binds an ACE2 receptor.
- the coronavirus is SARS-CoV-2 or SARS-CoV or the coronavirus spike polypeptide or fragment thereof is a SARS- CoV-2 or SARS-CoV spike polypeptide or fragment thereof.
- Another embodiment is use of an antibody as described herein in the manufacture of a medicament for prevention or treatment of a coronavirus infection.
- the coronavirus infection is caused by at least one coronavirus that specifically binds an ACE2 receptor.
- the at least one coronavirus is SARS-CoV-2 and/or SARS-CoV.
- the medicament is for delivery by inhalation or nebulization.
- Figures 1A and 1B describe antigen validation by ELISA.
- Fig.1A shows the results of an ELISA assessing the binding of microtiter-well-adsorbed (S, S1, S2, S1-RBD) and microtiter- well-captured (AviTag-S1, AviTag-S1-RBD) SARS-CoV-2 spike protein fragments to cognate ACE2 receptor (ACE2-hFc).
- AviTag-S1 and AviTag-S1-RBD were captured on streptavidin- coated microtiter wells through their C-terminal biotins.
- FIG. 1B shows the results of an ELISA confirming the binding of microtiter-well-adsorbed SARS-CoV-2 spike protein fragments S, S1, S2 and S1-RBD to a commercial rabbit anti-SARS-CoV-2 S polyclonal antibody (pAb).
- Figures 2A and 2B show the results of llama serology.
- Fig. 2A shows the results of an ELISA performed with pre-immune (day 0) and immune (day 21 and 28) sera, demonstrating that spike protein-immunized Maple Red and Eva Green llamas generated a strong immune response against target antigens S, S1, S2 and S1-RBD.
- ELISA performed with day 0, 21 and 28 sera showed spike protein-immunized llamas did not react with non-target antigens (casein and dipeptidase 1 [DPEP1]), demonstrating specificity of the immune response.
- Fig.2B shows flow cytometry surrogate neutralization assays performed with pre-immune (day 0) and immune (day 21 and 28) sera demonstrating that the Eva Green llama mounted a polyclonal immune response that was more potent in inhibiting the binding of SARS-CoV-2 S to ACE2 than Maple Red’s. Due to a lack of complete curves, inhibitory serum titers for Maple Red sera were estimated assuming similar upper plateaus as those for Eva Green sera.
- Figure 3 provides a schematic representation of three different antibody formats: monomeric V H H, bivalent V H H-Fc and monovalent V H H-Fc.
- Figures 4A and 4B show size-exclusion chromatogram (SEC) profiles of anti-SARS- CoV-2 spike protein V H Hs.
- Fig.4A shows SEC profiles of Eva Green V H Hs.
- Fig.4B shows SEC profiles of Maple Red V H Hs. V e , elution volume: mAU, milliabsorbance unit.
- Figures 5A and 5B show data on the thermostability of anti-SARS-CoV-2 spike protein V H Hs.
- Fig.5A provides representative examples showing the thermal unfolding of NRCoV2-1d, NRCoV2-02, NRCoV2-07 and NRCoV2-11, as determined using CD spectroscopy.
- Fig. 5B provides a summary of V H H T m s. The dotted line across the graph in Fig.5B represents the median T m (70.4 ).
- Figures 6A, 6B, 6C, 6D and 6E show SPR/ELISA binding affinity, specificity and cross- reactivity data for anti-SARS-CoV-2 V H Hs and V H H-Fcs. Figs.
- FIGS. 6A and 6B show the results of ELISA assessing the cross-reactivity of anti-SARS-CoV-2 V H H-Fcs against to a collection of spike glycoproteins from various coronavirus genera and SARS-CoV-2 variants. Assays were performed at a fixed V H H-Fc concentration (13 nM). The V H H-72 (Wrapp et al., 2020) benchmark and human ACE-2 were included for comparison. The epitope bin numbers provided along the bottom of Fig. 6B correspond to the bins shown in Fig. 9G. Fig.
- 6C shows representative SPR sensorgrams showing single-cycle kinetic analysis of NRCoV2-02, NRCoV2-07, NRCoV2-SR03 and NRCoV2-S2A4 VHH binding to SARS-CoV S and SARS-CoV-2 S, S1, S2 and S1-RBD.
- Spike proteins were captured on CM5 sensorchip surfaces, followed by flowing V H Hs over the sensorchip surfaces at the concentration ranges shown in each panel.
- “NRCoV2-02/NRCoV2- 07”, “SR03” and “S2A4” represent SPR binding profiles for V H Hs specific to SARS-CoV-2 S1- RBD, S1-NTD and S2, respectively.
- NRCoV2-07 also represents binding profiles for V H Hs that cross-react with SARS-CoV.
- Figs. 6D and 6E show the results of ELISA assessing the domain specificity of a set of anti-SARS-CoV-2 V H H-Fcs. Assays were performed against SARS-CoV-2 S, S1, S1-NTD and S1-RBD at a fixed concentration (13 nM) (Fig.6D) or varying concentrations (Fig. 6E) of V H H-Fcs. In the graphs shown in Fig. 6E, NRCoV2-02 is included as an internal control (dashed line).
- Figures 7A and 7B show on-/off-rate maps summarizing V H H kinetic rate constants, k a s and k d s. Diagonal lines represent equilibrium dissociation constants, K D s. Maps were constructed using the V H H binding data against SARS-CoV-2 S (Fig.7A) and SARS-CoV S (Fig.7B). In Fig. 7A, V H Hs are clustered based on subunit/domain specificity determined in Example 5.
- FIGS. 8A and 8B show the results of flow cytometry assessing the binding of V H H-Fcs to SARS-CoV-2 S-expressing CHO-S cells.
- Fig. 8A shows representative examples.
- Fig. 8B summarizes affinity values, i.e., EC 50 s, determined from graphs in Fig.8A.
- V H H-72 (Wrapp et al., 2020; open circle) is included for comparison. The line through the data points is the median.
- Figures 9A, 9B, 9C, 9D, 9E, 9F, and 9G show epitope typing and binning data obtained by SDS-PAGE/WB, sandwich ELISA and SPR.
- Fig.9A and 9B show the results of epitope typing of anti-SARS-CoV-2 V H Hs by SDS-PAGE/WB. Binding of biotinylated V H Hs or V H H-Fcs to denatured SARS-CoV-2 S was detected using streptavidin-peroxidase conjugate (Fig.9A) or anti- human Ig Fc antibody-peroxidase conjugate (Fig. 9B), respectively. Presence of binding signals indicates V H H recognizing a linear epitope.
- FIG.9C shows representative sensorgrams showing SPR epitope binning on SARS-CoV-2 S-immobilized surfaces.
- Figs. 9D and 9E show epitope binning of S1-RBD-specific V H Hs by competitive sandwich ELISA. ELISA binding results for pair-wise combinations of V H Hs against S1 are presented as a heat map.
- Binding pairs giving binding signal were considered as recognizing non-overlapping epitopes hence belonging to different epitope bins or V H H clusters, while those giving no/week binding signals (colorless/pale shading) were considered to be recognizing overlapping epitopes belonging to the same epitope bins.
- ACE2-Fc and V H H-72 V H H/V H H-Fc benchmark were also included in assays.
- Fig.9F provides a schematic summary of the initial epitope binning results.
- NRCoV2- 1c and NRCoV2-MRed02 were assigned to bin 1 since their CDRs were essentially the same as to those of NRCoV2-1a/1d and NRCoV2-MRed04, respectively, with experimentally defined bins.
- Fig. 9G provides a schematic summary of binning results after further characterization. Unless specified otherwise, references to epitope bin numbers throughout the present disclosure refer to the bins identified in Fig.9F. The bin numbers provided in Fig.9E correspond to the bins shown in Fig.9G.
- FIG 10 shows the results of ELISA assessing the ability of monomeric V H Hs in blocking (“neutralizing”) the binding of human ACE2 receptor (ACE2-Fc) to its SARS-CoV-2 S1-RBD ligand (i.e., S).
- a 450 nm is a measure of blocking.
- V H H-72 V H H (Wrapp et al., 2020) and monomeric ACE2-H 6 served as positive antibody controls, while toxin A-specific A20.1 V H H (Hussack et al., 2011) was a negative antibody control.
- PBS represents assays in which VHH was substituted with PBS and, similar to the A20.1 control, provides a reference binding signal for lack of any blocking (“min inhibition”).
- FIG. 11 shows sensorgrams showing the ability of monomeric V H Hs in blocking (“neutralizing”) the binding of ACE2 receptor to its ligand SARS-CoV-2 S.
- a tandem SPR in two different orientation formats were performed where injection of V H H (orientation #1) or ACE2 (orientation #2) at 20 – 40 ⁇ K D concentration (V H H) or 1 ⁇ M (ACE2) over sensor chip- immobilized S was followed by injection of V H H + ACE2 mix at the same V H H and ACE2 concentrations.
- Solid and dashed profiles represent binding results with the two orientation formats.
- NRCoV2-02:ACE2 represents profiles for blocking (neutralizing) VHHs where the addition of the V H H or ACE2 results in no significant increase in binding over that achieved by the injection of the ACE2 or V H H over the antigen surface.
- “NRCoV2-11:ACE2” represents profiles for non-blocking (non-neutralizing V H Hs where the addition of the V H H or ACE2 results in significant increase in binding over that achieved by the injection of the ACE2 or V H H over the antigen surface.
- RUs, representing binding differences between the first and second injection were calculated from the sensorgrams and used to identify V H Hs that block (neutralize) the binding of ACE2 receptor to its ligand S1-RBD.
- FIG. 12A and 12B show the results of flow cytometry assessing the ability of monomeric V H Hs in blocking (“neutralizing”) the binding of SARS-CoV-2 S to ACE2-expressing Vero E6 cells at 100 nM (Fig. 12A) or increasing (Fig. 12B) V H H concentrations.
- Fig. 12B provides plots showing inhibition of SARS-CoV-2 S binding to Vero E6 cells as a function of V H H concentration.
- the NRCoV2-1d, NRCoV2-02, NRCoV2-05, and NRCoV2-11 V H Hs are S1-RBD, SR13, S1-NTD-specific.
- Monomeric ACE2 (ACE2-H 6 ) serves as positive “antibody” control and reference, and V H H-72 V H H (Wrapp et al., 2020) is included as benchmark.
- A20.1 and PBS represent negative control assays in which V H Hs were replaced with C. difficile toxin A-specific A20.1 V H H (Hussack et al., 2011) and PBS, respectively.
- Figures 13A and 13B show virus-neutralizing potential of V H H-Fcs in flow cytometry- based surrogate virus neutralization assays.
- Fig.13A shows flow cytometry assessing the ability of bivalent VHH-Fcs in blocking (“neutralizing”) the binding of SARS-CoV-2 S to ACE2- expressing Vero E6 cells at 250 nM V H H-Fc concentrations.
- Fig. 13B shows flow cytometry assessing the ability of bivalent V H H-Fcs in blocking (“neutralizing”) the binding of SARS-CoV- 2 S to ACE2-expressing Vero E6 cells at increasing V H H-Fc concentrations.
- V H H-Fcs are S1-RBD- specific, while NRCoV2-SR01 and NRCoV2-SR13 V H H-Fcs are S1-NTD-specific.
- V H H-72 V H H- Fc (Wrapp et al., 2020) is included as a benchmark.
- A20.1” and “PBS” represent negative control assays in which V H Hs were replaced with C. difficile toxin A-specific A20.1 V H H (Hussack et al., 2011) and PBS, respectively.
- Figures 14A and 14B show the results of a V H H-Fc in vitro live-virus micro- neutralization assay. Antibody concentrations that gave 100% neutralization, i.e., MN100s, were used to rank the neutralizing potency of V H H-Fcs. A lower MN 100 means a higher neutralization potency. V H H-72 (Wrapp et al., 2020) is included as benchmark.
- Fig.14A provides a plot showing the MN 100 s of bivalent V H H-Fcs. The inset shows MN 100 s of monomeric NRCoV2-02 and V H H- 72 V H Hs.
- Fig.14B provides a plot comparing the MN 100 s of bivalent V H H-Fcs to monovalent V H H-Fcs. Monovalent V H H-72-Fc did not show MN 100 at the highest concentration tested (350 nM). In monovalent V H H-Fc constructs, one heavy chain displays an S- specific V H H, while the other displays a C. difficile toxin A-specific, mock V H H (A26.8) (Hussack et al., 2011).
- FIG. 15A shows inhibition capability of S1-RBD-specific V H H-Fcs at high (312.5 nM) and low (2.5 nM) V H H-Fc concentrations.
- NRCoV2-08, NRCoV2-19 and NRCoV2-21 which showed no binding to spike protein-expressing CHO cells (CHO-S), do not neutralize either.
- V H H-72 Wrapp et al., 2020
- C. difficile toxin A-specific V H H A20.1 are included as benchmark and negative control, respectively.
- Figs.15B-D provide representative examples showing inhibition capability of V H H-Fcs as a function of V H H- Fc concentration, for select S-RBD specific antibodies (Fig. 15B), S1-NTD-specific antibodies (Fig. 15C), and S2-specific antibodies (Fig. 15D).
- Antibody concentrations that gave 50% neutralization, i.e., IC 50 s, were calculated from graphs and used to rank the neutralizing potency of VHH-Fcs.
- Fig. 15E shows a summary of IC50 categorized based on subunit/domain specificity and epitope bin. A lower IC 50 means a higher neutralization potency.
- V H H-72 is shown as open circle in bin 1.
- FIG. 16A shows SEC profiles of pre- vs post-aerosolized V H Hs, for representative V H Hs.
- NRCoV2-1d, NRCoV2-02 and NRCoV2-07 represent the vast majority of V H Hs which were resistant to aerosolization-induced aggregation, showing a homogenously monomeric peak.
- the V H H-72 benchmark forms a significant amount of soluble aggregates following aerosolization.
- NRCoV2-11 on the other hand represents the few V H Hs that formed visible, precipitating aggregates reflected in significant reduction of their monomeric peak areas (compare monomeric peak for pre- vs post-aeosolized NRCoV2-11). V e , elution volume.
- Fig. 16B summarizes the % recovery of all V H Hs and
- Fig. 16C summarizes the % recovery of a subset of V H Hs.
- % recovery represents the proportion of a V H H that remained monomerically soluble following aerosolization.
- the open circle in Fig.16B represents benchmark V H H-72. The line through the data points is the median.
- FIG.16D shows the results of ELISA assessing the effect of aerosolization on the functionality of V H Hs by comparing the binding activity of pre- vs post- aerosolized V H Hs against SARS-CoV-2 S.
- Essentially identical EC 50 s for pre- vs post-aerosolized V H Hs clearly indicate aerosolization had no effect on the functional activity of V H Hs.
- Figure 17 provides the results of sandwich ELISA demonstrating the potential utility of V H Hs in detecting/capturing SARS-CoV-2, SARS-CoV and related viruses, as well as their spike proteins.
- SARS-CoV-2 S, S1 and S1-RBD antigens were used as surrogates for viruses. Specific detection of S, S1 and S1-RBD was achieved using NRCoV2-02 V H H as the capture antibody and NRCoV2-1d, NRCoV2-02, NRCoV2-04, NRCoV2-07, or NRCoV2-11 V H H-Fcs as detecting antibodies. SC 50 is the concentration of antigen that gives 50% binding and were calculated from graphs. [0076] Figure 18 shows an alignment of amino acid sequences of S-specific V H H antibodies described herein. [0077] Figure 19 shows an alignment of amino acid sequences of S1-NTD-specific V H H antibodies described herein.
- Figure 20 shows an alignment of amino acid sequences of S2-specific V H H antibodies described herein.
- Figure 21 shows an alignment of amino acid sequences of S1-RBD-specific V H H antibodies described herein.
- Figures 22A, 22B, 22C, and 22D show the results of efficacy tests of V H H-Fcs in hamsters challenged with SARS-CoV-2.
- Fig.22A shows lung viral load in V H H-Fc-treated (V H H- 72 benchmark, 1d, 05, MRed05, SR01, S2A3, 1d/MRed05, 1d/SR01) and control groups treated with PBS or isotype A20.1 V H H-Fc at 5 dpi.
- Fig.22B shows the percent body weight change for antibody-treated and control groups.
- Fig. 22C shows the percent body weight change at 5 dpi.
- treatment effects assessed by one-way ANOVA with Dunnett’s multiple comparison post hoc test, were significant (*p ⁇ 0.05, **p ⁇ 0.01, ***p ⁇ 0.001 or ****p ⁇ 0.0001).
- Dunnett’s test was performed by comparing treatment groups against the isotype control. ns, not significant.
- FIG 23 shows immunohistochemical demonstration of SARS-CoV-2 nucleocapsid (N) protein in the lungs of V H H-Fcs-treated animals.
- Untreated (PBS) and A20.1 isotype-treated animals showed strong viral N protein immunoreactivity which was mainly found in large multifocal patches of consolidated areas.
- Black arrow indicates the presence of viral N protein in bronchiolar epithelial cells.
- Omission of anti-nucleocapsid antibody eliminated the staining (Negative). Shown also is the absence of staining in healthy animals (Na ⁇ ve).
- a marked reduction in viral N protein staining was seen in all lung tissues examined from V H H-Fc-treated animals (middle and bottom panels).
- FIG. 24 shows immunohistochemical detection of infiltrating macrophages in the lungs of V H H-Fc-treated animals. Untreated (PBS) and A20.1 isotype-treated animals showed an intense immune reaction to anti-Iba-1 antibody and an increased number of Iba-1-positive macrophagesin the consolidated areas.
- FIG. 25 shows immunohistochemical detection of T lymphocytes in the lungs of V H H- Fc-treated animals. Untreated (PBS) and A20.1 isotype-treated animals showed an increased number of T lymphocytes in the pulmonary interstitium. A dramatic decrease in the number of T lymphocytes was seen in the lungs of V H H-Fc-treated animals. Representative images are shown from a single experiment.
- Figure 26 shows immunohistochemical detection of apoptotic cells in the lungs of V H H- Fc-treated animals.
- Untreated (PBS) and A20.1 isotype-treated animals showed an increase in the number of TUNEL-positive cells with classical features of apoptotic cells in the pulmonary interstitium.
- a marked reduction in the TUNEL- positive cells was seen in the lungs of NRCoV2-05- and NRCoV2-MRed05-treated animals. Black arrows indicate occasional TUNEL-positive cells. Representative images are shown from a single experiment.
- Figure 27 shows on-/off-rate maps summarizing V H H kinetic rate constants, kas and kds determined by SPR for the binding of V H Hs to SARS-CoV S.
- Figures 28A and 28B show on-/off-rate maps summarizing V H H kinetic rate constants, kas and kds determined by SPR for the binding of V H Hs to SARS-CoV-2 Alpha S (Fig.28A) and SARS-CoV-2 Beta S (Fig.28B).
- Figure 29 shows representative SPR sensorgrams showing single-cycle kinetics analysis of NRCoV2-02, NRCoV2-15 and NRCoV2-MRed05 binding to Wuhan, Alpha and Beta S (NRCoV2-02, NRCoV2-15) and RBD (NRCoV2-MRed05).
- Figure 30 shows a summary of IC 50 s obtained by live virus neutralization assays (LVNAs) for V H H-Fcs against Wuhan, Alpha, and Beta SARS-CoV-2 variants.
- the epitope bin numbers provided in Fig.30 correspond to the bins shown in Fig.9G.
- Figures 31A, 31B, 31C, and 31D show results from live virus neutralization assays assessing the ability of SARS-CoV-2 V H H-Fcs in blocking the infection of ACE2-expressing Vero E6 cells by SARS-CoV-2 Alpha (Fig. 31A and Fig. 31C) and Beta (Fig. 31B and Fig. 31D) variants at fixed (Fig. 31A and Fig. 31B) or varying (Fig. 31C and Fig. 31D) V H H-Fc concentrations.
- Inhibition assays shown in Fig.31A and Fig.31B were performed at 312, 12.5 or 0.5 nM V H H-Fc concentrations.
- FIG. 31C and Fig. 31D are recorded in Table 19.
- V H H-72 and C. difficile toxin A-specific V H H A20.1 are included as a benchmark and negative antibody control, respectively.
- the epitope bin numbers provided in Figs. 31C and 31D correspond to the bins shown in Fig.9G.
- Figure 32 shows in vivo stability and persistence of V H Hs. Stability and persistence were determined by monitoring the concentration of a representative VHH-Fc (NRCoV2-1d) in hamster blood at various days post-injection by ELISA. V H H-72 V H H-Fc was used as the benchmark.
- DETAILED DESCRIPTION [0091] The following is a detailed description provided to aid those skilled in the art in practicing the present disclosure.
- coronavirus spike polypeptide or “coronavirus spike protein” (S) is the major coronavirus surface protein, and is a glycosylated homotrimer that binds to a host cell receptor and mediates coronavirus entry into a host cell.
- the coronavirus may be SARS-CoV-2, SARS-CoV, or another coronavirus.
- SARS-CoV-2 may be used herein to refer to any strain or variant of the SARS-CoV-2 virus.
- SARS-CoV may be used to refer to any strain or variant of the SARS-CoV virus.
- a SARS-CoV-2 variant is a strain of SARS-CoV-2 that comprises one or more mutations relative to the Wuhan strain of SARS-CoV-2.
- a variant may be, but need not be, a variant that has been designated as a variant of concern or a variant of interest by the World Health Organization.
- the term “polypeptide” refers to a molecule comprising two or more amino acid residues linked by peptide bonds.
- a polypeptide may have primary, secondary, and/or tertiary structure.
- a “protein” comprises at least one polypeptide and may have primary, secondary, tertiary, and/or quaternary structure.
- polypeptide and “protein” are often used interchangeably, and a polypeptide may be comprised by a protein.
- a protein may be a homo- or hetero-multimer that comprises two or more polypeptides, or a protein may comprise a single polypeptide.
- a polypeptide or protein may include one or more post- translational modifications, such as, but not limited to, glycosylation, phosphorylation, lipidation, S-nitrosylation, N-acetylation, or methylation.
- fragment in the context of a polypeptide, refers to a portion of a polypeptide comprising a series of consecutive amino acid residues from a parent polypeptide.
- fragment refers to an amino acid sequence of at least 5, at least 10, at least 15, at least 20, at least 25, at least 30, at least 35, at least 40, at least 45, or at least 50 consecutive amino acid residues from a parent polypeptide.
- a fragment may comprise an epitope or binding domain from a parent polypeptide.
- a fragment may be a biologically active fragment that retains one or more functional characteristics of a parent polypeptide.
- antibody refers to an antigen binding protein comprising at least a heavy chain variable region (V H ) that binds a target epitope.
- the term antibody includes monoclonal antibodies comprising immunoglobulin heavy and light chain molecules, single heavy chain variable domain antibodies, and variants and derivatives thereof, including chimeric variants of monoclonal and single heavy chain variable domain antibodies.
- the antibody may be a naturally-occurring antibody, it may be obtained by manipulation of a naturally-occurring antibody, or it may be produced using recombinant methods.
- an antibody may include, but is not limited to a Fv, single-chain Fv (scFv; a molecule consisting of VL and VH connected with a peptide linker), Fab, F(ab')2, single domain antibody (sdAb; an antibody composed of a single V L or V H ), or a multivalent presentation of any of these.
- Antibodies such as those just described may require linker sequences, disulfide bonds, or other types of covalent bond to link different portions of the antibody.
- the antibody may be a single domain antibody derived from a naturally- occurring source.
- Heavy chain antibodies of camelid origin Hamers-Casterman et al, 1993
- V H H sdAbs have also been observed in shark and are termed VNAR (Nuttall et al, 2003).
- single domain antibody includes single domain antibodies directly isolated from V H , V H H, V L , or VNAR reservoir of any origin through phage display or other technologies, single domain antibodies derived from the aforementioned single domain antibodies, recombinantly produced single domain antibodies, as well as single domain antibodies generated through further modification of such single domain antibodies by humanization, affinity maturation, stabilization, solubilization, camelization, or other methods of antibody engineering.
- Single domain antibodies possess desirable properties for antibody molecules, such as high thermostability, high detergent resistance, relatively high resistance to proteases (Dumoulin et al, 2002) and high production yield (Arbabi-Ghahroudi et al, 1997). They can also be engineered to have very high affinity by isolation from an immune library (Li et al, 2009) or by in vitro affinity maturation (Davies & Riechmann, 1996).
- a person of skill in the art would be well-acquainted with the structure of a single-domain antibody.
- a single domain antibody comprises a single immunoglobulin domain that retains the immunoglobulin fold; most notably, only three CDR/hypervariable loops form the antigen-binding site. However, and as would be understood by one of skill in the art, not all CDRs may be required for binding the antigen.
- one, two, or three of the CDRs may contribute to binding and recognition of the antigen by a single domain antibody.
- the CDRs of the single domain antibody or variable domain are referred to herein as CDR1, CDR2, and CDR3, and numbered as defined by Lefranc et al., 2003.
- the amino acid sequence and structure of a heavy chain variable domain can be considered—without however being limited thereto—to be comprised of four framework regions or ‘FR’, which are referred to in the art and herein as ‘Framework region 1’ or ‘FR1’; as ‘Framework region 2’ or ‘FR2’; as ‘Framework region 3’ or ‘FR3’; and as ‘Framework region 4’ or ‘FR4’, respectively; which framework regions are interrupted by three complementarity determining regions or ‘CDR s’, which are referred to in the art as ‘Complementarity Determining Region 1’ or ‘CDR1’; as ‘Complementarity Determining Region 2’ or ‘CDR2’; and as ‘Complementarity Determining Region 3’ or ‘CDR3’, respectively.
- FR framework regions or ‘FR’, which are referred to in the art and herein as ‘Framework region 1’ or ‘FR1’; as ‘Framework region 2’ or ‘FR2’
- binding refers to the process of a non-covalent interaction between molecules. Preferably, said binding is specific.
- the terms ‘specific’ or ‘specificity’ or grammatical variations thereof refer to the number of different types of antigens or their epitopes to which a particular antibody such as a V H H can bind.
- the specificity of an antibody also referred to as “specific binding”, can be determined based on affinity.
- a specific antibody preferably has a binding affinity (Kd) for its epitope of less than 10 7 M, preferably less than 10 8 M.
- affinity refers to the strength of a binding reaction between a binding domain of an antibody and an epitope. It is the sum of the attractive and repulsive forces operating between the binding domain and the epitope.
- affinity refers to the equilibrium dissociation constant, Kd.
- epitope refers to a part of an antigen that is recognized by an antibody.
- epitope includes linear epitopes and conformational epitopes.
- a linear epitope is an epitope that is recognized by an antibody based on its primary structure, and a stretch of contiguous amino acids is sufficient for binding.
- a conformational epitope is based on 3-D surface features and shape and/or tertiary structure of the antigen.
- neutralizing antibody refers to an antibody that, when bound to an epitope, interferes with at least one of the steps leading to the release of a virus genome, such as a coronavirus genome, into a host cell.
- subject refers to an animal that is susceptible to infection by a coronavirus.
- the subject may be an animal that is susceptible to infection by a coronavirus that binds an ACE2 receptor, such as SARS-CoV-2 or SARS-CoV.
- the subject may be a human or non-human animal.
- the subject is a human or non-human mammal.
- the ACE2 receptor may be a human ACE2 receptor or an animal ACE2 receptor.
- administration routes include, but are not limited to, parenteral (intravenous, intramuscular, and subcutaneous), oral, nasal, ocular, transmucosal (buccal, vaginal, and rectal), transdermal, and pulmonary administration.
- strong interaction and strong binding refer to the presence of salt bridges and cation-pi interactions between amino acid residues, as is known to the skilled person.
- weak interaction and weak binding refer to the presence of hydrogen bonds and non-bonded/hydrophobic interactions, as is known to the skilled person.
- purified refers to a molecule, e.g. a polypeptide or protein that has been identified and substantially separated and/or recovered from the components of its natural environment.
- isolated antibody refers to an antibody that is substantially freed from other antibody molecules having different antigenic specificities. Further, a purified or isolated antibody may be substantially free of one or more other cellular and/or chemical substances. Absolute purity is not required for a molecule to be considered purified or isolated.
- pharmaceutically acceptable means generally regarded as safe when administered to humans.
- the term “pharmaceutically acceptable” is approved by a federal or state government regulatory agency for use in animals, more preferably in humans.
- carrier means a diluent, adjuvant, excipient, or vehicle with which a compound is formulated and/or administered.
- Such pharmaceutical carriers can be water and sterile liquids, such as petroleum, animal, vegetable or synthetically derived oils such as peanut oil, soybean oil, mineral oil, sesame oil. Water or saline solutions and aqueous dextrose and glycerol solutions are preferably used as carriers for injectable solutions. Suitable pharmaceutical carriers are, for example, described in “Remington (23 rd edition), The Science and Practice of Pharmacy”.
- linker refers to a chemical group or molecule that can be used to join one molecule to another.
- An antibody may be linked to another molecule by a linker or an antibody may be directly linked (aka joined, fused, or bonded) to another molecule, without the use of a linker.
- Suitable linkers are known in the art and may be selected based on the chemical nature of the molecules being joined. Examples of linkers include peptide linkers and chemical cross-linkers. Peptide linkers may comprise a single amino acid residue or a plurality of amino acid residues.
- label refers to a molecule or compound that can be used to label a molecule, such as an antibody, to allow detection of the molecule. Suitable labels will be known to one skilled in the art and include, but are not limited to, radioisotopes; enzymes, such as horse radish peroxidase (HRP) or calf intestinal alkaline phosphate (AP); fluorophores; antigen binding fragments from cleaved antibodies (Fabs); and colloidal gold.
- HRP horse radish peroxidase
- AP calf intestinal alkaline phosphate
- fluorophores antigen binding fragments from cleaved antibodies (Fabs)
- colloidal gold colloidal gold.
- nucleic acid molecule refers to any nucleic acid-containing molecule including, but not limited to, DNA, RNA, and DNA/RNA hybrids, in any form and/or conformation.
- the term encompasses nucleic acids that include any of the known base analogs of DNA and RNA. For example, single-stranded, double-stranded, nuclear, extranuclear, extracellular, and isolated nucleic acids are all contemplated.
- vector refers to a synthetic nucleotide sequence used for manipulation of genetic material, including but not limited to cloning, subcloning, sequencing, or introduction of exogenous genetic material into cells, tissues or organisms. It is understood by one skilled in the art that vectors may contain synthetic DNA sequences, naturally occurring DNA sequences, or both. Examples of commonly used vectors include plasmids, viral vectors, cosmids, and artificial chromosomes.
- the term “regulatory sequence” includes promoters, enhancers and other expression control elements, such as polyadenylation sequences, matrix attachment sites, insulator regions for expression of multiple genes on a single construct, ribosome entry/attachment sites, introns that are able to enhance expression, and silencers. Promoters may be cell-specific or tissue- specific to facilitate expression in a desired target.
- promoters include promoters, enhancers and other expression control elements, such as polyadenylation sequences, matrix attachment sites, insulator regions for expression of multiple genes on a single construct, ribosome entry/attachment sites, introns that are able to enhance expression, and silencers. Promoters may be cell-specific or tissue- specific to facilitate expression in a desired target.
- the term “operably linked” is used herein to mean that the two sequences are associated in a manner that allows the regulatory sequence to affect expression of the other nucleotide sequence.
- the term “host cell” refers to a cell into which a nucleic acid molecule or vector may be introduced, for example to allow for replication of the nucleic acid molecule or vector by the host cell and/or to allow for expression of the nucleic acid molecule, or of a nucleic acid molecule comprised by the vector, by the host cell to produce a product of interest, such as an RNA or protein.
- the nucleic acid molecule may encode an antibody as described herein, and introduction of the nucleic acid molecule into the host cell may allow the antibody to be expressed by the host cell.
- a host cell may be any suitable cell, such as a bacterial cell or eukaryotic cell.
- host cells include E. coli, yeast, and mammalian cells, such as, but not limited to, Chinese hamster ovary (CHO) cells, mouse myeloma cells, and human embryonic kidney (HEK) cells.
- CHO Chinese hamster ovary
- HEK human embryonic kidney
- invention refers to the prophylactic administration of a therapeutic molecule or composition to a subject to prevent the occurrence of, or to reduce the severity of, an illness or disease in the subject.
- sample refers to a sample in which a coronavirus presence is suspected or expected.
- the sample may be a biological sample from a subject, such as, but not limited to, blood or a fraction thereof, saliva, cellular material, urine, or feces; a sample from a bioreactor; or an environmental sample.
- sequence identity refers to the percentage of sequence identity between two amino acid sequences or two nucleic acid sequences. To determine the percent identity of two amino acid sequences or of two nucleic acid sequences, the sequences are aligned for optimal comparison purposes (e.g. gaps can be introduced in the sequence of a first amino acid or nucleic acid sequence for optimal alignment with a second amino acid or nucleic acid sequence). The amino acid residues or nucleotides at corresponding amino acid positions or nucleotide positions are then compared. When a position in the first sequence is occupied by the same amino acid residue or nucleotide as the corresponding position in the second sequence, then the molecules are identical at that position.
- the determination of percent identity between two sequences can also be accomplished using a mathematical algorithm.
- One non-limiting example of a mathematical algorithm utilized for the comparison of two sequences is the algorithm of Karlin and Altschul, 1990, modified as in Karlin and Altschul, 1993. Such an algorithm is incorporated into the NBLAST and XBLAST programs of Altschul et al., 1990.
- BLAST nucleotide searches can be performed with the NBLAST nucleotide program parameters set, e.g.
- Gapped BLAST can be utilized as described in Altschul et al., 1997.
- PSI-BLAST can be used to perform an iterated search which detects distant relationships between molecules.
- XBLAST and NBLAST can be used (see, e.g. the NCBI website).
- Another non-limiting example of a mathematical algorithm utilized for the comparison of sequences is the algorithm of Myers and Miller, 1988. Such an algorithm is incorporated in the ALIGN program (version 2.0) which is part of the GCG sequence alignment software package.
- ALIGN program version 2.0
- a PAM120 weight residue table When utilizing the ALIGN program for comparing amino acid sequences, a PAM120 weight residue table, a gap length penalty of 12, and a gap penalty of 4 can be used.
- the percent identity between two sequences can be determined using techniques similar to those described above, with or without allowing gaps. In calculating percent identity, typically only exact matches are counted.
- the phrase "at least one,” in reference to a list of one or more elements, should be understood to mean at least one element selected from any one or more of the elements in the list of elements, but not necessarily including at least one of each and every element specifically listed within the list of elements and not excluding any combinations of elements in the list of elements.
- This definition also allows that elements may optionally be present other than the elements specifically identified within the list of elements to which the phrase "at least one" refers, whether related or unrelated to those elements specifically identified.
- Description [00127] The present disclosure relates to SARS-CoV-2 spike protein-specific antibodies and uses thereof. Provided are isolated or purified antibodies comprising complementarity determining region (CDR) 1, CDR2, and CDR3 sequences as outlined in Table 6.
- the antibodies described herein recognize a variety of spike protein epitopes in different subunit and domains of the coronavirus spike protein, specifically S2, the N-terminal domain of S1 (S1-NTD), and the receptor binding domain of S1 (S1-RBD). Within these subunits/domains, antibodies described herein recognize several different epitopes. Because of this epitopic diversity, antibodies described herein may be used in combination, for example for combination therapy, or as bi- specific or multi-specific antibodies.
- An antibody as described herein comprises an antigen binding portion of an antibody heavy chain, wherein the antigen binding portion comprises a first complementarity determining region (CDR1), a second complementarity determining region (CDR2), and a third complementarity determining region (CDR3), and wherein CDR1, CDR2, and CDR3, respectively, comprise the amino acid sequence set forth in: SEQ ID NO: 1, SEQ ID NO: 45, and SEQ ID NO: 90; SEQ ID NO: 2, SEQ ID NO: 45, and SEQ ID NO: 91; SEQ ID NO: 1, SEQ ID NO: 46, and SEQ ID NO: 92; SEQ ID NO: 3, SEQ ID NO: 47, and SEQ ID NO: 93; SEQ ID NO: 4, SEQ ID NO: 48, and SEQ ID NO: 94; SEQ ID NO: 5, SEQ ID NO: 49, and SEQ ID NO: 95; SEQ ID NO: 6, SEQ ID NO: 50, and SEQ ID NO: 96; SEQ ID NO: 7,
- the antibody comprises the amino acid sequence set forth in SEQ ID NO: 183, 184, 185, or 186.
- an antibody as described herein comprises the amino acid sequence set forth in SEQ ID NO: 136, SEQ ID NO: 137, SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144, SEQ ID NO: 145, SEQ ID NO: 146, SEQ ID NO: 147, SEQ ID NO: 148, SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151, SEQ ID NO: 152, SEQ ID NO: 153, SEQ ID NO: 154, SEQ ID NO: 155, SEQ ID NO: 156, SEQ ID NO: 157, SEQ ID NO: 158, SEQ ID NO: 159, SEQ ID NO: 160, SEQ ID NO:
- the antibody comprises an amino acid sequence having at least 75%, at least 80%, at least 85%, at least 90%, at least 95%, at least 96%, at least 97%, at least 98%, or at least 99% sequence identity to the full length of the amino acid sequence set forth in SEQ ID NO: 136, SEQ ID NO: 137, SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144, SEQ ID NO: 145, SEQ ID NO: 146, SEQ ID NO: 147, SEQ ID NO: 148, SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151, SEQ ID NO: 152, SEQ ID NO: 153, SEQ ID NO: 154, SEQ ID NO: 155, SEQ ID NO: 156, SEQ ID NO: 157, SEQ ID NO: 158, SEQ ID NO:
- nucleic acid molecule encoding an antibody as described herein.
- a further embodiment is a vector comprising the nucleic acid molecule.
- the nucleic acid molecule may be operably linked to at least one promoter and/or regulatory element to enable expression in a host cell.
- a further embodiment is a host cell comprising the nucleic acid or vector.
- An antibody as described herein may be comprised within a composition.
- the antibody may be comprised within a pharmaceutical composition that comprises a pharmaceutically acceptable carrier and/or diluent, the antibody may be linked to another molecule, or the antibody may be immobilized on a substrate.
- the pharmaceutical composition may be for delivery by inhalation or nebulization.
- Antibodies and compositions as described herein may be used, or for use, to treat or prevent a coronavirus infection, including an infection caused by at least one coronavirus that specifically binds an ACE2 receptor. Antibodies as described herein may also be used in the manufacture of a medicament for prevention or treatment of a coronavirus infection.
- the at least one coronavirus is SARS-CoV-2 and/or SARS-CoV.
- a method for prevention or treatment of a coronavirus infection comprising administering an antibody or composition as described herein to a subject in need thereof. In an embodiment, the administration is by inhalation or nebulization.
- Antibodies and compositions as described herein may also be used, or for use, to detect, quantify, and/or capture a coronavirus, a coronavirus spike polypeptide or a coronavirus spike polypeptide fragment. Further provided are methods for detecting, quantifying, and/or capturing a coronavirus, a coronavirus spike polypeptide or a coronavirus spike polypeptide fragment using an antibody or composition as described herein.
- the coronavirus or spike polypeptide is a coronavirus or spike polypeptide that specifically binds an ACE2 receptor.
- the ACE2 receptor may be a human ACE2 receptor or an animal ACE2 receptor.
- the coronavirus is SARS-CoV-2 or SARS-CoV
- the spike polypeptide or fragment thereof is from SARS-CoV-2 or SARS-CoV
- the antibodies described herein have the characteristics of neutralizing antibodies, and some have been demonstrated to be cross-reactive with the spike protein of other coronaviruses, such as SARS-CoV and related coronaviruses that infect bats, pangolin, and civet, suggesting that antibodies described herein may be useful for binding the spike protein of more than one coronavirus; including coronaviruses that bind an ACE2 receptor, such as SARS-CoV-2 and SARS-CoV.
- Antibodies described herein have also been demonstrated to bind various SARS- CoV-2 spike protein variants, such as the Wuhan-Hu-1 variant that was first identified in China; the B.1.1.7 variant that was first identified in the United Kingdom (also referred to herein as the UK variant, or the Alpha variant); the B.1.352 variant that was first identified in South Africa (also referred to herein as the South Africa variant, or the Beta variant), the B.1.617.1 variant that was first detected in India (also referred to herein as Kappa); the B.1.617.2 variant that was first detected in India (also referred to herein as Delta); and the B.1.1.529 variant that was first detected in South Africa (also referred to herein as Omicron).
- SARS- CoV-2 spike protein variants such as the Wuhan-Hu-1 variant that was first identified in China; the B.1.1.7 variant that was first identified in the United Kingdom (also referred to herein as the UK variant, or the Alpha variant); the B.1.352 variant that was first identified in South Africa (also referred
- Antibodies described herein may be linked to another molecule or substrate.
- they may be linked to a detectable label to allow detection, quantification, and/or visualization; they may be linked to a molecule that extends antibody half-life, such as polyethylene glycol (PEG), Ig Fc, serum albumin, serum-albumin-specific antibody, serum- albumin-specific peptide, or Fc-specific peptides, proteins or antibodies; they may be linked to a therapeutic molecule; they may be immobilized onto a substrate, such as a plastic surface, a magnetic bead or a protein sheet or bead; and/or they may be linked to a polypeptide.
- PEG polyethylene glycol
- Ig Fc Ig Fc
- serum albumin serum-albumin-specific antibody
- serum- albumin-specific peptide serum- albumin-specific peptide
- Fc-specific peptides proteins or antibodies
- antibodies described herein may be linked to an ACE2 polypeptide or a fragment thereof.
- Antibodies described herein may also be employed in various formats and combinations.
- antibodies described herein may be monoparatropic or multiparatropic (including biparatropic), or monospecific or multispecific (including bispecific).
- Antibodies described herein may be in a monovalent format or in a multivalent format (including a bivalent format).
- Antibodies described herein that are specific for the same or different epitopes, or for the same or different spike protein subunit or domains may be linked, for example to produce antibodies with different affinities and/or specificities.
- antibodies described herein may be linked to one or more other antibodies or antibody fragments.
- antibodies described herein may be used individually or in combination.
- a combination may comprise any two or more antibodies described herein, or it may comprise at least one antibody described herein and another antibody.
- the antibodies are V H H antibodies or V H H-Fc antibodies.
- Antibodies described herein may be useful for a variety of applications.
- they may be useful for detecting the presence of a coronavirus or a coronavirus spike polypeptide or fragment thereof; for capturing a coronavirus or a coronavirus spike polypeptide or fragment thereof; for quantifying the amount of a coronavirus or a coronavirus spike polypeptide or fragment thereof in a sample; for treatment or prevention of a coronavirus infection; for diagnosing a coronavirus infection; for monitoring the production of a coronavirus spike protein or fragment thereof; for purification of a coronavirus spike protein or fragment thereof; for detecting the level of expression of a coronavirus spike protein or fragment thereof, and/or for quantifying the amount of a coronavirus.
- cross-reactive antibodies described herein have been shown to be stable against aerosolization, indicating that they may be suitable for delivery to the lungs by inhalation or nebulization.
- cross-reactive antibodies may have general applicability for the treatment, prevention, detection, quantification or capture of coronaviruses, in addition to SARS-CoV-2, or coronavirus spike polypeptides or fragments thereof from coronaviruses in addition to SARS-CoV-2.
- cross-reactive antibodies may be used to bind coronaviruses or coronavirus spike polypeptides that bind an ACE2 receptor, including fragments of such coronavirus spike polypeptides.
- Antibodies described herein may be classified based on the spike protein subunit or domain to which they bind. Nine antibodies were generated that bind to the S1-NTD domain, 24 antibodies were generated that bind to the S1-RBD domain, and 14 antibodies were generated that bind to the S2 subunit (see Tables 5 and 6). Neutralization assays, as described in the Examples, identified antibodies with neutralizing properties within each of these three groups. To the inventors’ knowledge, this is the first known observation of single domain antibodies neutralizing the SARS-CoV-2 virus by targeting a non-S1-RBD region of S, i.e., S1-NTD or S2.
- Antibodies described herein may also be classified based on their pattern of cross- reactivity with different coronavirus spike proteins and/or spike protein variants, as shown in Figures 6A and 6B. Antibodies that recognize the same set of spike proteins and/or spike protein variants may be viewed as a single group.
- V H H spike protein antibody V H H-72 (Wrapp et al., 2020). Further, many of the antibodies described herein are demonstrated to outperform V H H-72 in neutralization assays, and some are demonstrated to be more broadly neutralizing than V H H-72. Additionally, some antibodies described herein are demonstrated to be more broadly cross-reactive than V H H-72. [00142]
- the antibodies described in the following examples may be modified, while still retaining antigen specificity. For example, changes may be introduced into the amino acid sequence of the framework regions, or the antibodies may be humanized. The antibodies may also be linked to other molecule(s).
- Coronavirus spike protein fragments used for library selection, binding and epitope study experiments Reference Accession describing RBD/SD1 (aa319-591) b Wrapp et al., (Wuhan) QHD43416.1 NRC 6xHis 2020 t SARS-CoV S AAU04646.1 NRC F Galipeau et al., Cive 1 a b LAG-Dual Strep-6xHis 2021 na, not applicable
- spike protein fragments used in the following Examples were produced as described in Stuible et al., 2021.
- Materials and Methods Binding to cognate human angiotensin converting enzyme (ACE2) receptor
- ELISA was performed to determine if spike proteins were able to bind to human ACE2 when passively adsorbed (S, S1, S1-RBD and S2) or captured (S1, S1-RBD) on microtiter wells.
- Table 2 Binding affinity (EC 50 ) of passively absorbed spike fragments and streptavidin- captured spike fragments to ACE2 Antigen S S1 S2 S1-RBD AviTag-S1-RBD AviTag-S1 “nb”
- Table 3 Binding affinity (EC 50 ) of passively absorbed spike fragments to a polyclonal antibody known to be specific for SARS-CoV-2 spike protein Antigen S S1 S2 S1-RBD
- Example 2 Llama Immunization and Serum Analyses Introduction
- two llamas were immunized with SARS-CoV-2 S or S/S1-RBD to trigger the generation of a diverse pool of antibodies targeting manifold sites over the surface of S, and targeting the S1-RBD sub-domain of S which is used by the virus to start the process of host cell infection through interaction with the ACE2 receptor.
- Llama sera were assessed by ELISAs for generation of immune responses against SARS-CoV-2 spike proteins, and by flow cytometry surrogate neutralization assays for generation of neutralizing antibodies.
- Materials and Methods [00155] Llama immunization [00156] Immunizations were performed at Cedarlane Laboratories (Burlington, ON, Canada) and essentially as described (Hussack et al., 2011).
- a second llama (Maple Red) was immunized with 100 g of S in 500 ⁇ L PBS combined with 500 ⁇ L of Freund’s complete adjuvant on day 0, followed by immunization with 100 g of S mixed with Freund’s incomplete adjuvant on day 7, and immunization with 50 g of S mixed with Freund’s incomplete adjuvant on each of days 14 and 21.
- Serum ELISA [00158] Llama sera were tested for antigen-specific immune response by ELISA essentially as described (Hussack et al., 2011; Henry et al., 2016). Briefly, dilutions of sera in PBST were added to wells pre-coated with S, S1, S2 or S1-RBD.
- Negative antigen control wells were pre-coated with casein (100 ⁇ L of 1% v/w) or recombinant human dipeptidase 1 ectodomain, DPEP1 (50 ng/well; Sino Biological, Cat#13543-H08H). Following 1 h incubation at room temperature, wells were washed 10 times with PBST and incubated with HRP-conjugated polyclonal goat anti-llama IgG heavy and light chain antibody (Bethyl, Cat#A160-100P) for 1 h at room temperature. After 10 washes, the peroxidase activity was determined as described above.
- Trimeric SARS-CoV-2 S was chemically biotinylated using EZ-LinkTM NHS-LC-LC- Biotin following manufacturer’s instructions (Thermo Fisher, Cat#21343). Vero E6 cells (ATCC, Cat#CRL-1586) were maintained according to ATCC protocols.
- DMEM medium Thermo Fisher, Cat#11965084
- FBS Thermo Fisher, Cat#10438034
- 2 mM Glutamax TM Thermofisher, Cat#35050061
- Percent inhibition 100 x [1 - (F n - F min ) / (F max - F min )], where, F n is the measured fluorescence at any given competitor serum dilution, F min is the baseline fluorescence measured in the presence of cells and SAPE only, and F max is the maximum fluorescence, measured in the absence of competitor serum.
- Phage display library construction [00166] On day 28, 100 mL of blood from each of the two llamas was drawn and peripheral blood mononuclear cells (PBMCs) were purified by Ficoll® gradient at Cedarlane Laboratories (Burlington, ON, Canada). Two independent phage-displayed V H /V H H libraries were constructed from 5 ⁇ 10 7 PBMCs as described previously (Henry et al., 2016; Rossotti et al., 2015; Henry et al., 2015).
- PBMCs peripheral blood mononuclear cells
- V H /V H H genes were amplified using semi-nested PCR and cloned into the phagemid vector pMED1, followed by transformation of E. coli TG1 to construct two libraries with sizes of 1 ⁇ 10 7 and 2 ⁇ 10 7 independent transformants for Eva Green and Maple Red, respectively.
- library phages were diluted at 1 x 10 11 colony-forming units (cfu)/mL in PBSBT [PBS supplemented with 1% [w/v] BSA and 0.05% Tween® 20] and incubated in antigen-coated microtiter wells for 2 h at 4°C.
- phages were added to wells with passively-adsorbed S (10 pg/well; PI), passively-adsorbed S2 (10 pg/well; P2), streptavidin-captured biotinylated SI (0.5 pg/well; P3) and streptavidin-captured biotinylated Sl-RBD (0.5 pg/well; P4).
- phages were pre-absorbed on passively-adsorbed Sl-RBD wells (10 pg/well) for 1 h at 4°C and then the unbound phage in the solution was transferred to wells with streptavidin-captured biotinylated SI (0.5 pg/well) in the presence of non-biotinylated Sl-RBD competitor in solution (10 pg/well).
- wells were washed 10 times with PBST and bound phages were eluted by treatment with 100 mM glycine pH 2.2 for 10 min at room temperature, followed by immediate neutralization of phages with 2 M Tris.
- phages were bound on streptavidin-captured biotinylated Sl-RBD but elution of bound phages were carried out competitively with 50 nM ACE2-Fc following the washing step.
- a small aliquot of eluted phage was used to determine their titer on LB-agar/ampicillin plates and the remaining were used for their subsequent amplification in E.coli TGI strain (Hussack et al., 2011).
- the amplified phages were used as input for the next round of selection as described above.
- Example 4 V H H Cloning/Expression in E. coli, Stability/Affinity Validation and Cross- Reactivity Studies Introduction [00171] Hits identified by monoclonal phage ELISA and DNA sequencing were cloned into the expression vector pMRo.BAP.H 6 (Rossotti et al., 2019), produced as His 6 -tagged V H Hs in the periplasmic space of E. coli BL21(DE3) and purified by immobilized metal-ion affinity chromatography (IMAC). V H Hs were subsequently validated for binding and further explored for cross-reactivity soluble ELISA against SARS-CoV-2, SARS-CoV and MERS-CoV spike proteins.
- IMAC immobilized metal-ion affinity chromatography
- V H Hs were validated for aggregation resistance by size exclusion chromatography (SEC) and thermostability by circular dichroism T m measurement assays.
- Lead V H Hs were produced in mammalian cells in fusion with human IgG1 Fc and were subsequently tested in a comprehensive cross-reactivity ELISA against a collection of various coronavirus spike proteins (S).
- SEC size exclusion chromatography
- S coronavirus spike proteins
- V H Hs were subsequently cloned into pET expression vector (Novagen, Madison, WI) for their production in BL21(DE3) E.coli as monomeric soluble protein (Rosotti et al., 2019). Briefly, individual colonies were cultured overnight in 10 mL of LB supplemented with 50 ⁇ g/mL of kanamycin (LB/Kan) at 37°C and 250 rpm. After 16 h, cultures were added to 250 mL LB/Kan and grown to an OD 600 of 0.6. Expression of V H Hs was induced with 10 ⁇ M of IPTG (isopropyl -D-1-thiogalactopyranoside) overnight at 28°C and 250 rpm.
- IPTG isopropyl -D-1-thiogalactopyranoside
- V H /V H Hs were extracted by sonication and purified by IMAC as described previously (Rosotti et al., 2019).
- ELISA see below
- a small fraction was biotinylated by incubating 1 mg of purified V H Hs with 10 ⁇ M of ATP (Alfa Aesar, Cat#CAAAJ61125-09), 100 ⁇ M of D-(+)-biotin (VWR, Cat#97061-446) and a bacterial cell extract overexpressing E.coli BirA as described previously (Rossotti et al., 2015b).
- V H H binding validation and preliminary cross-reactivity studies by ELISA Binding validation studies were performed with S1-RBD-specific clones. Briefly, microtiter well plates were coated with 50 ng/well SARS-CoV-2 S1-RBD in 100 ⁇ L PBS overnight at 4 .
- V H H T m s were measured by circular dichroism as previously described (Henry et al., 2017). Ellipticity of V H Hs were determined at 200 ⁇ g/mL V H H concentrations and 205 nm wavelength in 100 mM sodium phosphate buffer, pH 7.4.
- V H Hs Production of V H Hs in mammalian cells in fusion with human IgG1 Fc
- Codon-optimized genes for bivalent V H H-Fcs were synthesized (GenScript).
- VHH genes were PCR amplified as described previously and cloned into pTT5-hIgG1Fc between the genes for human V H leader sequence and the human IgG1 hinge/Fc sequences, using NarI/HindIII restriction sites.
- Bivalent V H H-Fcs were produced by transient transfection of HEK293-6E cells followed by protein A affinity chromatography as previously described (Rosotti et al., 2019).
- Heterodimeric monovalent V H H-Fcs were produced by co-transfection of HEK293-6E cells with two pTT5 vectors, one encoding for a 6xHis-tagged heavy chain (V H H1-hinge-C H 2-C H 3-His 6 ), the other for a non-tagged heavy chain of a different V H H (V H H2-hinge-C H 2-C H 3).
- the heterodimeric antibodies were purified by sequential protein A affinity chromatography and IMAC.
- V H H The sequence of the V H H was ordered as GeneBlock (IDT DNA) flanked by SfiI sites for cloning into pMRo.BAP.H6, and NarI/HindIII for cloning into pTT5-hIgG1Fc. Protein purity was evaluated by SDS-PAGE using 4–20% Mini-PROTEAN® TGX Stain-FreeTM Gels (Bio-Rad, Cat#17000435).
- V H H-Fcs were washed five times with PBSTC and binding of V H H-Fcs was detected using 1 ⁇ g/mL HRP-conjugated goat anti-human IgG. Finally plates were washed five times and peroxidase (HRP) activity was measured as described above.
- HRP-conjugated goat anti-human IgG HRP-conjugated goat anti-human IgG.
- HRP-conjugated goat anti-human IgG HRP-conjugated goat anti-human IgG.
- HRP-conjugated goat anti-human IgG HRP-conjugated goat anti-human IgG
- V H H hits were cloned in E. coli, confirmed by DNA sequencing, and expressed and purified by IMAC. Following expression of V H Hs, the binding of a sample set of V H Hs was validated by ELISA. Affinities, expressed as EC 50 s, were high, ranging from 0.4 to 7.2 nM (data not shown). V H Hs were also tested for aggregation resistance and stability, and cross-reactivity.
- Aggregation resistance and stability are desirable attributes of biotherapeutics, as they affect both efficacy and manufacturability.
- V H Hs tested were found to be aggregation resistant (Figs.4A and 4B), except for NRCoV2-08, which showed some degree of aggregation
- the V H Hs were also tested for thermal stability and found to be highly thermostable. With the exception of NRCoV2-11, which had a relatively lower T m of 60.4°C, the remaining 25 V H Hs tested had T m s higher than 65°C, with a T m range and median of 65.5 – 79.8°C and 70.4°C, respectively (Figs.5A and 5B).
- V H Hs had T m s that were higher than that of the V H H-72 benchmark (73.0°C).
- V H Hs with antigen binding activity were produced as monomeric and dimeric V H H-Fcs for subsequent binding and neutralization assays.
- the schematic formats of these fusion molecules are depicted in Fig.3.
- Fig.6A and 6B The results of cross-reactivity studies using SARS-CoV-2 variants and various coronaviruses are shown in Fig.6A and 6B. Initial experiments showed that for the UK (Alpha) and South Africa (Beta) variants of SARS-CoV-2, eight out of nine S1-NTD-specific V H Hs tested were cross-reactive to both variants (Fig.6A).
- S1-RBD-specific V H Hs 15/20 cross- reacted to both variants and an additional four cross-reacted with the UK variant. Only one (NRCoV2-08) V H H was not cross-reactive at all. Additionally, one S1-NTD-specific V H H, six S1- RBD-specfic and eight S2-specific V H Hs cross-reacted with SARS-CoV. Many antibodies also cross-reacted with pangolin CoV, with fewer, but still significant, numbers cross-reacting to SARS-like CoV W1V1, bat SARS-like CoV and civet SARS-CoV with similar cross-reactivity patterns.
- V H H-Fcs cross-reacted with the S protein from variants Alpha, Beta, Gamma, Delta and Kappa (B.1.617.1; Variant Being Monitored [VBM]).
- VBM Variant Being Monitored
- the exceptions were: 1) RBD-specific V H Hs NRCoV2-02/ NRCoV2-05 did not cross-react with Beta and Gamma and NRCoV2-04/ NRCoV2-14/ NRCoV2-15, did not cross- react with Kappa and 2) S2-specific V H Hs NRCoV2-MRed18 and NRCoV2-MRed19 did not cross-react with Kappa. All nine NTD-specific V H Hs cross-reacted with all variants tested.
- VHHs cross-reacted with pangolin CoV with fewer cross-reacting to SARS- CoV, SARS-like CoV WIV1, bat SARS-like CoV and civet SARS CoV.
- These viruses, including variants, are all of the Betacoronavirus Sarbecovirus subgenus. None of the antibodies tested cross- reacted with the remaining 11 non-Sarbecovirus Betacoronavirus, or with Alphacoronavirus, Deltacoronavirus or Gammacoronavirus. 29 V H Hs cross-reacted with the Omicron variant (Fig. 6B).
- V H Hs recognizing 10 – 12 viruses, including SARS-CoV-2 variants, were two NTD binders (NRCoV2-SR01, NRCoV2-SR02), six RBD binders (NRCoV2-1d, NRCoV2-07, NRCoV2-11, NRCoV2-12, NRCoV2-20, NRCoV2- MRed04) and six S2 binders (NRCoV2-S2F3, NRCoV2-S2G3, NRCoV2-S2G4, NRCoV2- MRed18, NRCoV2-MRed19, NRCoV2-MRed20).
- the VHH-72 benchmark was also broadly cross-reactive.
- V H Hs had similar cross-reactivity profiles to human ACE2, except that ACE2 did not bind civet SARS-CoV S and, unsurprisingly, bound HCoV-NL63 S.
- ACE2 did not bind civet SARS-CoV S and, unsurprisingly, bound HCoV-NL63 S.
- 12 out of 14 ELISA-positive V H Hs cross- reacted with SARS-CoV S, most with comparably high affinities (Table 11. Seven of these V H Hs were S2-specific, four were RBD-specific and one was NTD-specific.
- the SPR cross-reactivity data performed with 37 V H Hs, were consistent with ELISA, except for NRCoV2-04 and NRCoV2-14, which were negative or very weak for binding to the Beta variant by SPR (Tables 11 and 12). All 37 V H Hs tested bound the Alpha variant S protein, and 34 were also cross-reactive to the Beta variant S protein (Figs.28A (Alpha) and 28B (Beta); Fig.29; Table 11; Table 12).
- VHHs bound all three variants with similar affinities, except for V H Hs NRCoV2-10, NRCoV2-15 and NRCoV2-17 which bound to the Beta variant with 40 – 50-fold weaker affinity; the remaining four that did not bind the Beta variant showed cross-reactivity with the Alpha variant with similar (NRCoV2-04, NRCoV2-14) or reduced ( ⁇ 5-fold [NRCoV2-05] and ⁇ 20-fold [NRCoV2-02]) affinity relative to the Wuhan variant. All NTD-specific and S2-specific V H Hs cross-reacted with the three variants with essentially the same or similar affinities.
- V H Hs and V H H-Fcs The cross-reactivity of the V H Hs and V H H-Fcs is significant, as it is believed that the progenitor of SARS-CoV was generated by recombination among bat SARS-like coronaviruses that spread to humans via civet cat as an intermediate host (Zheng et al, 2020). Further, most new emerging viruses are derived from strains circulating in zoonotic reservoirs. Antibodies that can cross-react against a variety of animal and human coronaviruses have potential to be used for detection and/or treatment of emerging coronavirus outbreaks.
- V H Hs and V H H-Fcs Surface Plasmon Resonance (SPR) and ELISA Binding Studies Introduction
- SPR Surface Plasmon Resonance
- ELISA ELISA Binding Studies Introduction
- Binding of anti-SARS-CoV-2 V H Hs against various SARS-CoV-2 spike protein fragments (Wuhan) was assayed using SPR and ELISA to determine their affinity and domain/sub- domain specificity.
- Binding of V H Hs against SARS-CoV, SARS-CoV-2 UK (Alpha) variant and SARS-CoV-2 South African (Beta) variant spike protein S was also carried out to determine their virus cross-reactivity patterns.
- V H Hs, ACE2 receptor Prior to SPR analyses all analytes in flow (V H Hs, ACE2 receptor) were SEC-purified on a Superdex TM 75 Increase 10/300 GL column (Cytiva) in HBS-EP buffer at a flow rate of 0.8 mL/min to obtain monomeric proteins.
- SARS-CoV spike (S), SARS-CoV-2 spike trimer (S) and various SARS-CoV-2 spike fragments were immobilized on CM5 sensor chips through standard amine coupling (10 mM acetate buffer, pH 4.0; Cytiva).
- V H Hs at various concentration ranges were flowed over all surfaces at a flow rate of 40 ⁇ L/min with 180 s of contact time and 600 s of dissociation time.
- Surfaces were regenerated with a 12 s pulse of 10 mM glycine, pH 1.5, at a flow rate of 100 ⁇ L/min.
- V H H EK2 Injection of EGFR-specific V H H EK2 served as a negative control for the SARS-CoV and SARS-CoV-2 surfaces and as a positive control for the EGFR surface.
- the ACE2 affinity was determined using similar conditions by flowing a range of monomeric ACE2 concentrations (31.53 – 500 nM). All affinities were calculated by fitting reference flow cell-subtracted data to a 1:1 interaction model using BIAevaluation Software v3.0 (Cytiva). [00193] For V H H 12 and MRed05, V H H-Fc formats were used in SPR experiments.
- V H H-Fcs Approximately 200 RUs of V H H-Fcs (2 ⁇ g/mL) were captured on goat anti-human IgG surfaces (4000 RUs, Jackson ImmunoResearch, Cat#109-005-098) at a flow rate of 10 ⁇ L/min for 30 s.
- a range of SEC-purified RBD fragments (Table 1; SARS-CoV, Wuhan, Alpha and Beta) at 0.62 – 10 nM were flowed over the captured V H H-Fc at a flow rate of 40 ⁇ L/min with 180 s of contact time and 300 s of dissociation. Surfaces were regenerated with a 120 s pulse of 10 mM glycine, pH 1.5, at a flow rate of 50 ⁇ L/min.
- V H Hs that bound to the S1 subunit but not its S1-RBD domain in SPR assays, were further examined by ELISA to determine if they were binding to the S1-NTD domain of S1. Briefly, S, S1, S1-NTD and S1-RBD were coated onto NUNC® MaxiSorpTM 4BX plates (Thermo Fisher) at 100 ng/well in 100 ⁇ L PBS, pH 7.4.
- V H Hs were tested by SPR against SARS-CoV-2 S, S1, S1-RBD and S2 to determine their affinity and domain/sub-domain specificity. Binding data are presented in Fig. 6C, Figs. 7A-B, Table 8 and Table 9.
- SPR binding assays NRCoV2-SR01, NRCoV2-SR02, NRCoV2-SR03, NRCoV2-SR04, NRCoV2-SR13, NRCoV2-SR16, NRCoV2-MRed03, NRCoV2-MRed06 and NRCoV2-MRed07 bound to the S1 subunit but not to its S1-RBD domain.
- V H Hs were S1-NTD-specific (Figs. 6D and 6E and Table 10).
- V H Hs displayed high affinity towards their target (i.e., S) with the vast majority having K D s in the range of single-digit-nM to pM.
- Three clusters of V H Hs based on domain/subdomain specificity were identified: (i) S1-RBD-specific V H Hs; (ii) S1-NTD- spepcfic V H Hs; and (iii) S2-specific V H Hs (Fig.7A).
- V H Hs As for the S1-RBD-specific V H Hs, with the exception of NRCoV2-06, which had an affinity of 223 nM (Table 11), the remaining 16 cluster members displayed high affinities ranging from 0.02 - 10 nM, all vastly outperforming the benchmark V H H-72 V H H, which had a K D of 86.2 nM.
- V H Hs were S1-NTD-specific and, similar to S1-RBD-specific V H Hs, displayed high affinities (K D s) in the range of 0.1 – 5.2 nM.
- 11 V H Hs were S2 subunit-specific, with similarly high affinities (K D s) ranging from 0.09 – 12.8 nM.
- V H Hs were tested against SARS-CoV (S) in SPR assays for quantitative determination of cross-reactivity.
- V H Hs were first screened for cross-reactivity at fixed concentrations. Twelve out of 37 V H Hs screened showed cross-reactivity to SARS-CoV S. These 12 V H Hs were subsequently subjected to comprehensive binding analysis against both SARS-CoV S and SARS- CoV-2 S at multiple V H H concentrations.
- the SPR cross-reactivity results which agreed with those from ELISAs, are presented in Fig.27 and Table 11. Seven out of the 12 VHHs tested were S2-specific, four were S1-RBD-specific and one was S1-NTD-specific.
- NRCoV2-MRed04 showed weak binding to SARS-CoV S compared to SARS-CoV-2 S (300 nM for SARS-CoV S vs 1 nM for SARS-CoV-2 S), but the remaining V H Hs cross-reacted with high/comparable affinities to both SARS-CoV-2 S and SARS-CoV S.
- NRCoV2-07, NRCoV2-12, NRCoV2-MRed18, NRCoV2-MRed19 and NRCoV2-MRed20 cross-reacted with SARS-CoV S with relatively lower affinities in comparison to SARS-CoV-2 S, but nonetheless with high absolute affinities in the low nanomolar KD range.
- V H Hs bound all three variants with similar affinities, except for V H Hs NRCoV2-10, NRCoV2-15 and NRCoV2-17 which bound to the Beta variant with 40 – 50-fold weaker affinity; the remaining four that did not bind the Beta variant showed cross- reactivity with the Alpha variant with similar (NRCoV2-04, NRCoV2-14) or reduced ( ⁇ 5-fold [NRCoV2-05] and ⁇ 20-fold [NRCoV2-02]) affinity relative to the Wuhan variant. All NTD- specific and S2-specific V H Hs cross-reacted with the three variants with essentially the same or similar affinities.
- Example 6 Cell Binding Assays by Flow Cytometry Introduction
- a stable Chinese hamster ovary (CHO) cell line CHO BRI TM /55E1 (Stuible et al., 2021) overexpressing SARS-CoV-2 S (CHO-S) was grown in BalanCDTM CHO Growth A medium (Irvine Scientific) supplemented with 50 ⁇ M of methionine sulfoximine (MSX) at 120 rpm and 37°C in a humidified 5% CO 2 atmosphere. When the cell count reached 2 x 10 6 /mL, the expression of the membrane anchored SARS-CoV-2 trimeric spike protein (SmT1, described in Stuible et al, 2021) was induced by adding cumate at 2 ⁇ g/mL.
- SmT1 membrane anchored SARS-CoV-2 trimeric spike protein
- V H H-Fcs V-Bottom 96-well microtest plates (Globe Scientific, Cat# 120130) and mixed with 50 ⁇ L of CHO-S cells.
- VHH-Fcs (NRCoV2-08, NRCoV2-19, NRCoV2-21, NRCoV2-S202) which bound to SARS-CoV-2 S in purified form did not bind to SARS-CoV-2 S-displaying target cells.
- the remaining 41 V H H-Fcs bound to cells in a dose dependent manner (Fig.8A- B; Table 13).
- affinities for S2-specific V H H-Fcs were also high (EC 50 range: 0.1 – 6.5; EC 50 median: 1 nM).
- Table 13 Summary of V H H-Fc bindings to SARS-CoV-2 S expressing CHO-S cells S1-RBD-specific S1-NTD-specific S2-specific x NRCoV2- 1.3 280 NRCoV2- 10 44 MRed18 0.9 10205
- Example 7 Epitope Studies Introduction [00211] Western blotting experiments were performed to determine if V H Hs bind to conformational or linear epitopes.
- V H H epitope binning was performed by SPR dual injection experiments on the SARS-CoV-2 S at a flow rate of 40 ⁇ L/min in HBS-EP buffer. Dual injections consisted of injection of V H H1 (at 50 – 100 ⁇ K D concentration) for 150 s, followed by immediate injection of a mixture of V H H1 + V H H2 (both at 50 – 100 ⁇ K D concentration) for 150 s. The opposite orientation was also performed (VHH2 followed by VHH2 + VHH1) (Fig.9C). Surfaces were regenerated using a 12 s pulse of 10 mM glycine, pH 1.5, at a flow rate of 100 ⁇ L/min.
- V H Hs All pairwise combinations of V H Hs were analyzed and distinct or overlapping epitope bins determined.
- Epitope binning by ELISA [00217] The pairwise ability of V H Hs to bind to their antigen in a sandwich ELISA format was evaluated as described previously (Rosotti et al., 2015a; Delfin-Riela et al., 2020), (Fig. 9D).
- a matrix of 14 wells (row) 23 wells (column) was generated using six NUNC® MaxiSorp TM 4BX plates (Thermo Fisher) and coated overnight at 4°C with 4 ⁇ g/mL streptavidin (Jackson ImmunoResearch, Cat#016-000-113) in 100 ⁇ L PBS, pH 7.4.
- Wells were blocked with 200 ⁇ L PBSC for 1 h at room temperature and then biotinylated V H Hs (10 ⁇ g/mL in 100 ⁇ L PBSCT) were captured in each row (same V H H in each row; 14 rows for a total of 14 V H Hs) for 1 h at room temperature.
- V H H-Fcs/ACE2-Fc at 1 ⁇ g/mL used as detector antibodies (same V H H-Fc in each column; 23 column for a total of 22 V H H-Fcs and ACE2-Fc).
- the binding of V H H-Fcs/ACE2-Fc to S1 was detected using 100 ⁇ L 1 ⁇ g/mL HRP-conjugated goat anti-human IgG (SIGMA, Cat#A0170).
- VHHs monomeric VHHs as probe
- three out of 26 VHHs tested bound to denatured S indicating they were recognizing linear epitopes, while the remaining V H Hs appeared to be conformational epitope-specific based on their lack of significant binding to denatures S.
- V H H-Fc was used instead of V H H
- V H Hs linear epitope-specific V H Hs give the option of virus detection against denatured S by robust diagnostic techniques such as SDS-PAGE/Western blot, where the additional molecular weight information provided by the SDS-PAGE would serve as a second, confirmatory piece of information to eliminate/reduce false positives obtained by binding data alone.
- V H Hs were subjected to epitope binning experiments by SPR and sandwich ELISA.
- V H H1 V H H1
- V H H2 V H H2 + VHH2
- Fig.9C (left panel) exemplifies a V H H pair (NRCoV2-02/NRCoV2-05) binding to an overlapping epitope, hence belonging to the same epitope bin, as the addition of the second V H H does not result in any increased binding (i.e., increase in RU) over that obtained for the addition of the first V H H.
- Fig. 9C (right panel), on the other hand, exemplifies a V H H pair (NRCoV2-02/NRCoV2-07) binding to non-overlapping epitopes, hence belonging to different epitope bins, as the addition of the second V H H results in significant increase in binding over that already achieved by the addition of the first V H H.
- the ELISA experiments confirmed the results of epitope binning by SPR, expanded the number of binders within each epitope bin, and identified new epitope bins.
- the epitope binning results obtained by SPR and ELISA are summarized in Figs. 9F (initial results), 9G (further results) and Table 14.
- Initial binning results identified 14 non-overlapping/partially overlapping bins: six for S1-RBD-specific V H Hs, three for S1-NTD-specific V H Hs and five for S2-specific V H Hs.
- Example 8 Surrogate Virus Neutralization Assays Introduction
- Surrogate neutralization assays were performed to identify potential neutralizing V H Hs/V H H-Fcs, i.e., V H Hs/V H H-Fcs inhibiting SARS-CoV-2 viruses from entering host cells.
- Three different surrogate assays were performed: ELISA, SPR and flow cytometry.
- ELISA and SPR ACE2 and SARS-CoV-2 S acted as surrogates for an ACE2-containing host cell and an S- containing invading virus, respectively.
- flow cytometry assays which were performed directly against the host cell (Vero E6), S1-RBD or S served as surrogate virus.
- Antibodies that interfered with the binding of spike fragment proteins to ACE2 in the surrogate assays were considered to be neutralizing antibodies.
- Materials and Methods [00222] ACE2 competition assay by ELISA [00223] Wells of NUNC® MaxiSorp TM microtiter plates (Thermo Fisher) were coated overnight at 4°C with 50 ng/well of S in 100 ⁇ L PBS, pH 7.4. Next day, plates were blocked with 250 ⁇ L PBSC for 1 h at room temperature.
- ACE2-Fc (ACROBiosystems, Cat#AC2-H5257) at 400 ng/mL was mixed with 50 ⁇ L of V H H at 1 ⁇ M, and then transferred to SARS-CoV-2 S coated microtiter plate wells. After 1 h incubation at room temperature, plates were washed 10 times with PBST and the ACE2-Fc binding was detected using 1 ⁇ g/mL goat anti-human IgG (Fc specific) peroxidase antibody (SIGMA, Cat# A0170) in 100 ⁇ L PBSCT. After 10 washes with PBST, the peroxidase activity was determined as described above.
- ACE2 competition assay by SPR [00225] Standard SPR techniques were used for binding studies. All SPR assays were performed on a Biacore TM T200 instrument (Cytiva) at 25°C with HBS-EP running buffer (10 mM HEPES, 150 mM NaCl, 3 mM EDTA, 0.005 % [v/v] Tween® 20, pH 7.4) and CM5 sensor chips (Cytiva).
- V H Hs Prior to SPR analyses, all analytes in flow (V H Hs, ACE2 receptor) were SEC-purified on a Superdex TM 75 Increase 10/300 GL column (Cytiva) in HBS-EP buffer at a flow rate of 0.8 mL/min to obtain monomeric proteins.
- V H Hs were analyzed for their ability to block the SARS- CoV-2 spike trimer (S) interaction with ACE2 using SPR dual injection experiments.
- S SARS- CoV-2 spike trimer
- V H Hs and ACE2 were flowed over the SARS-CoV-2 S surface at 40 ⁇ L/min in HBS-EP buffer.
- Dual injections consisted of injection of ACE2 (1 ⁇ M) for 150 s, followed by immediate injection of a mixture of ACE2 (1 ⁇ M) + V H H (at 20 – 40 ⁇ K D concentration) for 150 s. The opposite orientation was also performed (V H H followed by V H H + ACE2). Surfaces were regenerated using a 12 s pulse of 10 mM glycine, pH 1.5, at a flow rate of 100 ⁇ L/min. All pairwise combinations of VHHs and ACE2 were analyzed. V H Hs that competed with ACE2 for SARS-CoV-2 spike trimer binding showed no increase in binding response during the second injection. Conversely, a binding response was seen during the second injection for V H Hs that did not compete with ACE2.
- ACE2 competition assay by flow cytometry [00226] Experiments were performed essentially as described in Example 2. Briefly, 400 ng of chemically biotinylated trimeric SARS-CoV-2 S was mixed with 5 10 4 Vero E6 cells in the presence of decreasing concentrations of V H Hs or V H H-Fcs in a final volume of 150 ⁇ L.
- a competition assay with recombinant human ACE2-His 6 in lieu of V H H was also included.
- A20.1 a C. difficile toxin A-specific V H H (Hussack et al., 2011) was used as negative control VHH.
- V H Hs 14 S1-RBD-specific, 6 S1-NTD-specific and 6 S2-specific
- a total of 26 V H Hs 14 S1-RBD-specific, 6 S1-NTD-specific and 6 S2-specific
- a total of 26 V H Hs 14 S1-RBD-specific, 6 S1-NTD-specific and 6 S2-specific
- a total of 26 V H Hs 14 S1-RBD-specific, 6 S1-NTD-specific and 6 S2-specific
- Fig.10 the majority of S1-RBD binders were significantly neutralizing, with NRCoV2-1d, NRCoV2-02, NRCoV2-05 and NRCoV2-07 displaying essentially 100% inhibition and outperforming the V H H-72 benchmark.
- NRCoV2-SR01 Two of the S1-NTD- specific V H Hs (NRCoV2-SR01, NRCoV2-SR02) showed significant neutralization, with NRCoV2-SR02 essentially neutralizing at 100%. None of the S2 binders showed significant neutralizing activity.
- a conceptually similar assay to ELISA was performed by a competitive SPR. The results are shown in Fig. 11 and Table 16. The four lead neutralizers identified by ELISA, i.e., NRCoV2-01d, NRCoV2-02, NRCoV2-05 and NRCoV2-07, were confirmed by SPR to be complete neutralizers (‘blockers”).
- NRCoV2-14, NRCoV2-15, NRCoV2-18 and NRCoV2-20 showed partial neutralization (“+/-“; Table 16). The remaining V H Hs tested were judged to be non-neutralizing. Although the ELISA and SPR results agreed in the case of the majority of V H Hs, there was some disagreement. For example, while NRCoV2-SR02, NRCoV2-06, NRCoV2-10, and NRCoV2-11 were neutralizing by ELISA, they were not found to be neutralizing by SPR. Conversely, NRCoV2-20 was judged to be somewhat neutralizing by SPR, but non-neutralizing by ELISA.
- NRCoV2-1d, NRCoV2-02, NRCoV2-05 and NRCoV2-07 led others with IC 50 / I max % values of 8.6 nM/72%, 5.1 nM/100%, 9.5 nM/97%, and 7.5 nM/86%, respectively.
- a second group of V H Hs, including NRCoV2-10, NRCoV2-14, NRCoV2-15, NRCoV2-18, NRCoV2-20 and NRCoV2-MRed04 were also potent/efficacious neutralizers (Table 17). All of these antibodies outperformed the benchmark V H H-72, which had a far higher IC 50 (59 nM).
- NRCoV2-11 and NRCoV2-17 although showing high potencies (IC 50 s of 16.8 nM and 9.4 nM, respectively), had weak efficacies (I max % values of 20% and 18%, respectively). None of the S1-NTD or S2 binders was neutralizing. The results obtained by flow cytometry correlated well with those obtained by ELISA and SPR. [00230] To increase the neutralization potency and efficacy of the V H Hs, they were reformatted as bivalent V H H-Fcs.
- V H H-Fcs were generated and tested in flow cytometry surrogate neutralization assays as described above.
- the majority of V H H-Fcs demonstrated high potencies and efficacies (Figs.13A- B; Table 17). Reformatting had a significant effect on the neutralization potencies/efficacies of V H Hs.
- NRCoV2-11 (anti- RBD) and SR01 (anti-NTD) were also efficient, achieving neutralization as potent as was observed against Wuhan spike protein. From the list of antibodies tested NRCoV2-12, -20, -11 and -SR01 are the leads, showing efficient pan-neutralization against the SARS-CoV-2 variants generated so far, and outperforming the benchmark VHH-72.
- Table 15 Flow cytometry SVNAs against SARS-CoV-2 variants and SARS-CoV SVNA IC 50 (nM) SARS-CoV-2 S V H H-Fc SARS-CoV S VHH-72 5.6 10.6 5.1 3.3 10.5 8.5 - 7.8 3,4 17 8.6 10.6 26.4 214 - - 3.4 - 10 8.8 11.3 10.8 21.8 - - 4.3 - 7,9,10 SR13 7.7 22.4 - 16.5 - 12.2 - 15
- Table 17 Neutralization capabilities of SARS-CoV-2-specific V H Hs/V H H-Fcs obtained by surrogate virus neutralization flow cytometry assays against SARS-CoV-2 S (Wuhan) V H H/ACE2-H 6 2 V H-Fc/ACE2-Fc 2 Domain/ H V H H/ACE2 bd i 3 3 3 3 3 3 NRCoV2-S2A4 S2 - - - - - - - NRCoV2-S2F3 S2 - - - - - app eric ACE2.
- IC 50 concentration of V H H/V H H/Fc giving 50% neutralization
- IC 99 concentration of V H H/V H H/Fc giving 99% neutralization
- I max % maximal inhibitory effect
- IC 50 , IC 99 and I max % values were extracted from graphs exemplified in Figs. 12B and Figs. 13B. Dash indicate V H H/V H H-Fc does not neutralize the interaction between Vero E6 cell-displayed ACE2 and soluble S. 4 ICs cannot be determined with certainty due to low I max % values. nd, not determined, due to lack of sufficient quantities and/or neutralization as V H H-Fc.
- Example 9 Live-Virus Neutralization Assays Introduction [00235] V H H-Fcs were subjected to authentic-virus neutralizations assays, i.e., micro- neutralization assays, to identify those that neutralized infection of host cells by the invading SARS-CoV-2 virus. Materials and Methods Authentic-virus neutralizations assays [00236] Neutralization activity of antibodies to SARS-CoV-2 was determined with the microneutralization assay.
- V H H-Fc and V H H were prepared at 1 mg/mL in PBS and sterilized by passing through 0.22 ⁇ M filters.1:5 serial dilutions of 50 ⁇ g/mL of each antibody was carried out in DMEM, high glucose media supplemented with 1 mM sodium pyruvate, 1mM non-essential amino acids, 100 U/ml penicillin-streptomycin, and 1% heat- inactivated fetal bovine serum.
- SARS-CoV-2 strain SARS-CoV-2/Canada/VIDO-01/2020 was incubated at 250 pfu with antibody dilution in 1:1 ratio at 37 o C for 1 h.
- Vero E6 cells seeded in 96-well plates were infected with virus/antibody mix and incubated at 37 o C in humidified/5% CO 2 incubator for 72 hours post-infection (hpi). Cells were then fixed in 10% formaldehyde overnight and virus infection was detected with mouse anti-SARS-CoV-2 nucleocapsid antibody (R&D Systems, clone #1035111) and counterstained with rabbit anti-mouse IgG-HRP (Rockland Inc.). Colorimetric development was obtained with o-phenylenediamine dihydrochloride peroxidate substrate (Sigma-Aldrich) and detected on Biotek Synergy H1 plate reader at 490 nm.
- IC 50 was determined from non-linear regression on GraphPad Prism 9. For determining neutralization potencies by measuring cytopathic effect (CPE), infected Vero E6 cells were incubated at 37 for 96 h until the virus-only control wells had nearly 100% CPE (cell-only controls were also included). Neutralization was scored by MN 100 , lowest antibody concentration that gave no CPE, i.e., 100% neutralization. Assays were performed in technical duplicates. [00237] Results and Discussion [00238] A select set of lead V H H-Fcs were subjected to preliminary authentic-virus micro- neutralization assays to assess their SARS-CoV-2 virus-neutralizing activity.
- CPE cytopathic effect
- S1-RBD binders The most potent neutralizers were amongst the S1-RBD binders: NRCoV2-02 (MN 100 0.01 nM); NRCoV2-1d (MN 100 0.25 nM); NRCoV2-04 and NRCoV2-07 (MN 100 1.25 nM); NRCoV2-03 (MN 100 6.25 nM). NRCoV2-02 and NRCoV2-1d were far more potent neutralizers than the benchmark (V H H-72), by five- and 125-fold, respectively.
- S1-NTD binders had MN 100 s of 6.25 nM (NRCoV2-SR01, NRCoV2-SR02).
- NRCoV2-02 also outperformed the benchmark in V H H format by 125-fold (Fig.14A inset).
- monovalent V H H-Fc versions of select V H H-Fcs were generated. Based on MN 100 values, neutralization potencies were decreased by five-fold for NRCoV2-SR01, 25-fold for NRCoV2-1d and NRCoV2- 07 and more than 125-fold for NRCoV2-02, with their conversion from bivalent to monovalent V H H-Fcs, demonstrating the sizable contribution of bivalency to their neutralization potency.
- V H H-Fcs recognized epitopes 2/3/4 and had IC 50 s of 0.0008- 3.1 nM (Fig.15E and Fig.30; Table 19).
- the leads were NRCoV-05 (IC 50 0.0008 nM) followed closely by NRCoV-02 (IC 50 0.12 nM) and NRCoV2-MRed 05 (IC 50 0.17 nM).
- V H H-Fcs recognizing epitope 1 showed intermediate potencies with IC 50 s of 1.94 – 9.6 nM, with V H H-72 (belonging to the same bin 1) having similar IC 50 (8.46 nM).
- V H H- Fcs recognizing epitope 5 and 6 showed IC 50 s of 9.96 – 76 nM.
- S1-NTD-specific V H H six out of nine V H H-Fcs tested were neutralizing, with the lead V H H-Fcs having IC 50 s of 9.42, 14.31 and 54.2 nM. The remaining two had IC 50 s in the high nM - micromolar range.
- V H H-Fcs were neutralizing with IC 50 s from 12.2 nM for S2A3 to high nM - micromolar range for S2G3 and S2G4. These belonged to three different epitope bins.
- V H H-Fcs outperformed the V H H-72 benchmark by 2.5 – 10,000-fold.
- the NRCoV2-05, NRCoV2-02 and NRCoV2- MRed05 leads showed 10,000-fold, 70-fold and 50-fold higher potency than V H H-72, respectively.
- Beta mutations in the RBD were completely abrogated presumably by the Beta mutations in the RBD (K417N, E484K, N501Y), several others including NRCoV2-MRed05, NRCoV2-10 and NRCov2-15 did retain their high neutralizing potencies against both Alpha and Beta variants.
- MN 100 Neutralization capabilities (MN 100 ) of SARS-CoV-2-specific V H H-Fcs obtained by authentic-virus (aka live virus) neutralization assays
- MN 100 H H (nM) NRCoV2-07 x S1-RBD x null 31.25 A26.8 1 V H H-72 ben SARS-CoV-2 S (Wrapp et al. tive control V H H (Hussack et al., 2011).
- 2 MN 100 is the lowest antibody concentration that gave no cytopathic effect (100% neutralization). Dash indicate V H H-Fc does not neutralize SARS-CoV-2 virus at the highest V H H-Fc concentration used.
- MN 100 values were used to construct Fig.14A-B graphs. 3
- the MN 100 of monovalent V H H-72 and NRCoV2-02 V H Hs were 156.25 and 1.25 nM, respectively.
- Table 19 Neutralization capabilities (IC 50 ) of SARS-CoV-2-specific V H H-Fcs obtained by authentic-virus (aka live virus) neutralization assays
- IC 50 Neutralization capabilities
- nM NTD specific V H 1 V H H-72 benchmark is a SARS-CoV S-specific V H H that cross-reacts with SARS-CoV-2 S (Wrapp et al., 2020);
- A20.1 is C.
- V H H H Hussack et al., 2011. Epitope bin numbers correspond to the bins shown in Fig.9G.
- Example 10 Stability of V H Hs against Aerosolization Introduction
- One effective therapeutic approach against COVID-19 might be the direct delivery of aerosolized antibodies to the nasal and lung epithelia by inhalation.
- V H Hs in particular are advantageously fit for such administration approach due to their high stability and robustness. Since aerosolization could compromise the structural integrity and function of antibodies that lack sufficient stability, such as mAbs (Detalle et al., 2016; Respaud et al., 2015), the effect of aerosolization on the stability of V H Hs was tested.
- Aerosolized V H Hs were collected into 15 mL Round-Bottom Polypropylene test tubes (Falcon, Cat#C352059) for 5 min to allow condensation and were subsequently quantified and kept at 4 C until use. Then 200 ⁇ L aliquots of pre- and post- aerosolized V H Hs were subjected to SEC to obtain chromatogram profiles. Additionally, condensed V H Hs were closely monitored for the formation of any visible aggregates, and in cases where aggregate formation was observed, aggregates were removed by centrifugation prior to concentration determination, SEC analysis and ELISA. % soluble aggregate was determined as the proportion of a V H H that gave elution volumes (V e s) smaller than that of the monomeric V H H fraction.
- % recovery was determined as the proportion of a V H H that remained monomerically soluble following aerosolization.
- V H Hs including the benchmark V H H-72 were examined for their aggregation resistance/stability against aerosolization.
- V H Hs For a few V H Hs, e.g., NRCoV2-MRed20, NRCoV2- S2A4, as well as the V H H-72 benchmark, aerosolization induced some soluble aggregation formation as determined by SEC (Fig. 16A; Table 20).
- V H Hs e.g., NRCoV2-11, NRCoV2-SR03
- Fig. 16A-C the majority of VHHs (20 out of 30 VHHs tested) were highly stable against aerosolization, that is, they did not form any soluble or visible aggregates and demonstrated high % recovery upon aerosolization treatment.
- V H Hs examples include NRCoV2-1c/1d, NRCoV2-02, NRCoV2-07, NRCoV2-17, NRCoV2-18 and NRCoV2-20.
- High % recovery indicates these V H Hs advantageously lack non-specific binding to nebulizer surfaces. For therapeutic V H Hs, this is expected to translate to a more effective drug delivery to the site of viral infection.
- V H Hs i.e., NRCoV2-04, NRCoV2-14, NRCoV2-15, NRCoV2-SR04 and NRCoV2-MRed04, while forming some visible aggregates, still showed a good % recovery upon aerosolization (52 – 69 %).
- V H Hs the activities (EC 50 s) of post- aerosolized V H Hs were determined by ELISA and compared to those for pre-aerosolized V H Hs. ELISAs were performed on a sample of four V H Hs: NRCoV2-1d, NRCoV2-02, NRCoV2-07 and NRCoV2-11 (Fig. 16D). Comparison of EC 50 s for post-aerosolized V H Hs vs pre-aerosolized V H Hs demonstrated that aerosolization did not compromise the functionality of V H Hs (Fig.16D; Table 21).
- V H Hs for Diagnosis and Capture of SARS-CoV-2 Introduction
- Pre pre-aerosolized
- Post post-aerosolized
- V H Hs EC (nM) V H 50 H t
- Example 11 V H Hs for Diagnosis and Capture of SARS-CoV-2 Introduction
- V H Hs described herein are promising diagnostic/capture agents against SARS-CoV-2, SARS-CoV and related viruses as well as their spike proteins.
- four of the V H Hs were tested in sandwich ELISA for their diagnostic/capturing capability against SARS-CoV-2.
- V H H-Fc that binds to a non- overlapping epitope in relation to NRCoV2-02 was added as the detecting antibody followed by the addition of a HRP-conjugated probing antibody binding to the detecting antibody.
- the different V H H-Fcs tested as detecting antibodies were: NRCoV2-1d, NRCoV2-04, NRCoV2-07, and NRCoV2-11. Very low SC 50 values were obtained in ELISA assays (Fig. 17, Table 22).
- V H H-Fc was chosen as a representative V H H and VHH-72 V H H-Fc, whose modified/enhanced version is currently in a phase 1 clinical trial, was included as a reference.
- Hamsters were injected intraperitoneally (IP) with 1 mg of each antibody and serum antibody concentration was monitored for up to four days by ELISA.
- Significant and comparable VHH-Fc concentrations were present in the hamster sera for both 1d and VHH-72 V H H-Fcs on days 1 and 4 post injection (Fig. 32), indicating that V H H-Fcs would have the required serum stability and persistence in vivo for the duration of the animal studies.
- V H H-Fcs which were neutralizing by live virus neutralization assay was then assessed in a hamster model of SARS-CoV-2 infection.
- Five V H H- Fcs were selected to cover a wide range of important attributes including in vitro neutralization potencies and breadth, epitope bin, subunit/domain specificity and cross-reactivity pattern. These included three RBD-specific (1d, 05, MRed05), one NTD-specific (SR01) and one S2-specific (S2A3) V H H-Fcs.
- V H H-Fcs Cocktails of two V H H-Fcs were also included to explore synergy between the antibody pairs recognizing distinct epitopes within the RBD (1d/MRed05) or RBD and NTD (1d/SR01).
- Hamsters were administered IP with 1 mg of V H H-Fcs 24 h prior to intranasal challenge with SARS-CoV-2 Wuhan isolate. Daily weight change and clinical symptoms were monitored. At 5 dpi, lungs were collected to determine viral titers. Viral titer decrease and reversal of weight loss in antibody treated versus control animals were taken as measures of antibody efficacy.
- Animals treated with RBD binders 1d, 05, and MRed05 showed reduced lung viral burden by three, five and six orders of magnitude, respectively, relative to PBS or VHH-Fc isotype controls, with 05 and MRed05 reducing viral burden to below detectable levels (Fig. 22A).
- the RBD-specific VHH-72 benchmark caused a mean viral decrease of four orders of magnitude.
- the NTD binder SR01, and interestingly, the S2 binder S2A3, were also effective neutralizers, decreasing mean viral titers by four and three orders of magnitude, respectively.
- Both 1d/SR01 and 1d/MRed05 cocktails decreased viral titers by 6 orders of magnitude to undetectable levels of virus infection.
- SARS-CoV-2 infection is characterized by an overt inflammatory response in the respiratory tract accompanied by an increased infiltration of inflammatory immune cells, e.g., macrophages and T lymphocytes, in the lung parenchyma 70. As expected, this was the case for the non-treated PBS and isotype control groups.
- a neutralizing human antibody binds to the N-terminal domain of the Spike protein of SARS-CoV-2. Science 369, 650-655. Colwill K, et al.
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Virology (AREA)
- Immunology (AREA)
- Molecular Biology (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Organic Chemistry (AREA)
- Urology & Nephrology (AREA)
- Biomedical Technology (AREA)
- Hematology (AREA)
- Biochemistry (AREA)
- Pathology (AREA)
- Public Health (AREA)
- Biotechnology (AREA)
- Cell Biology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Microbiology (AREA)
- Veterinary Medicine (AREA)
- Food Science & Technology (AREA)
- Physics & Mathematics (AREA)
- Analytical Chemistry (AREA)
- General Physics & Mathematics (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Communicable Diseases (AREA)
- Oncology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Epidemiology (AREA)
- Peptides Or Proteins (AREA)
- Pulmonology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description

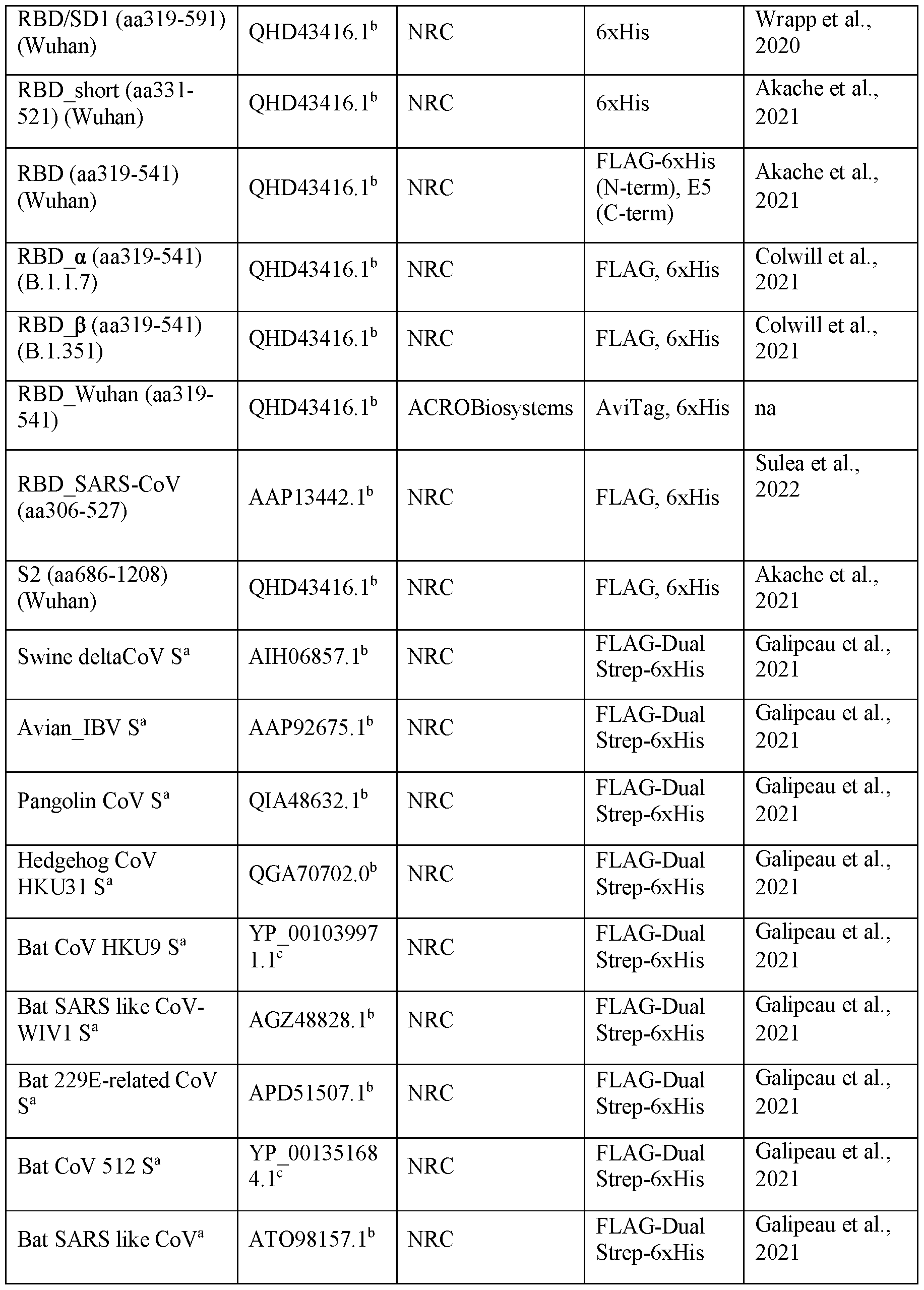





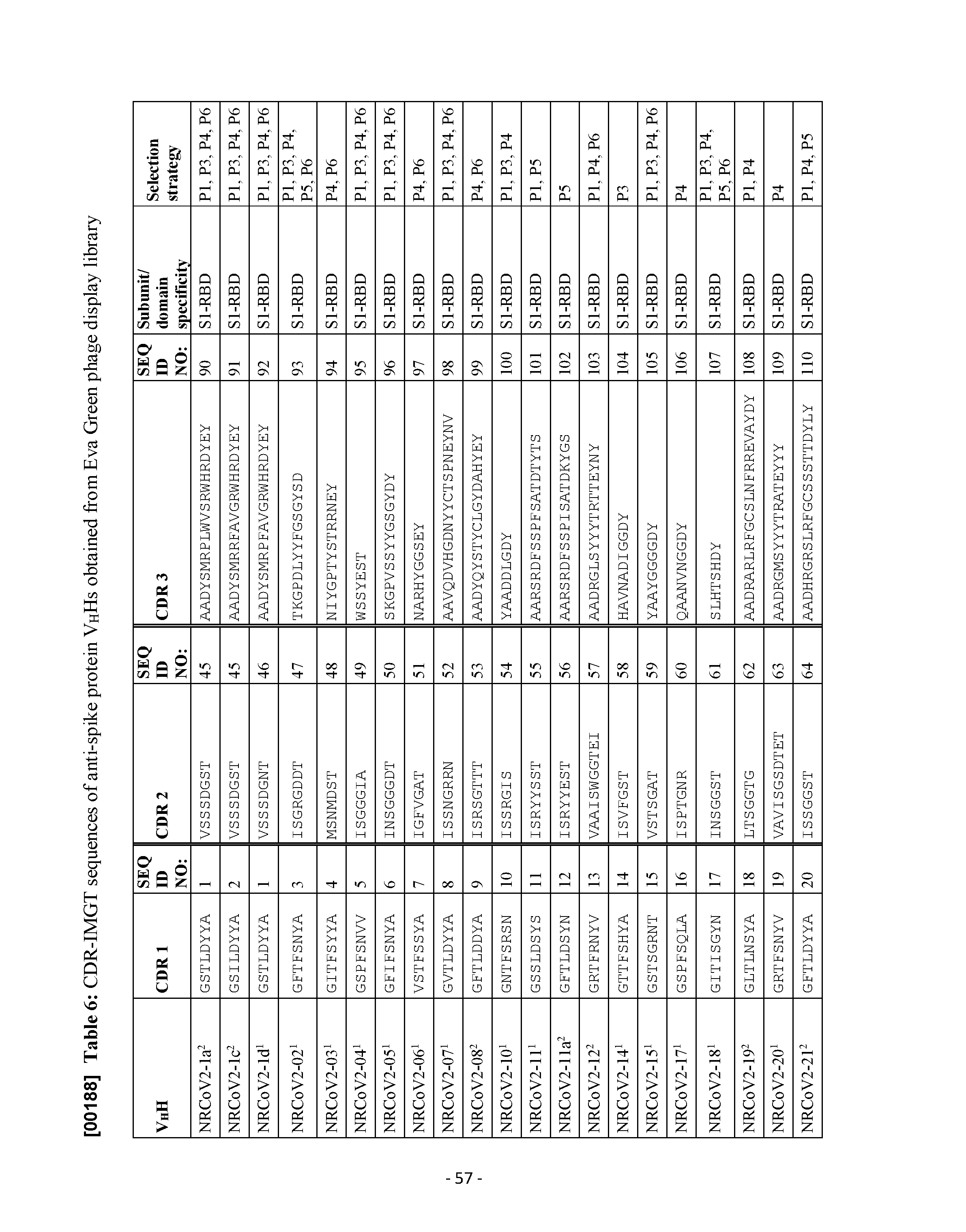
















































Claims
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2023560880A JP2024515525A (en) | 2021-04-22 | 2022-04-22 | Antibodies that bind to SARS-COV-2 spike protein |
| EP22791236.7A EP4326763A4 (en) | 2021-04-22 | 2022-04-22 | SARS-CoV-2 spike protein binding antibodies |
| CA3216274A CA3216274A1 (en) | 2021-04-22 | 2022-04-22 | Antibodies that bind sars-cov-2 spike protein |
| US18/556,415 US20240270826A1 (en) | 2021-04-22 | 2022-04-22 | Antibodies that bind sars-cov-2 spike protein |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CA3115877A CA3115877A1 (en) | 2021-04-22 | 2021-04-22 | Antibodies that bind sars-cov-2 spike protein |
| CA3,115,877 | 2021-04-22 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2022224203A1 true WO2022224203A1 (en) | 2022-10-27 |
Family
ID=83688473
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/IB2022/053756 Ceased WO2022224203A1 (en) | 2021-04-22 | 2022-04-22 | Antibodies that bind sars-cov-2 spike protein |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US20240270826A1 (en) |
| EP (1) | EP4326763A4 (en) |
| JP (1) | JP2024515525A (en) |
| CA (2) | CA3115877A1 (en) |
| WO (1) | WO2022224203A1 (en) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN117866903A (en) * | 2024-03-12 | 2024-04-12 | 北京贝来药业有限公司 | Single domain antibody modified stem cells and their use in disease treatment |
| WO2026062389A1 (en) * | 2024-09-20 | 2026-03-26 | The Rosalind Franklin Institute | Single domain antibodies against beta coronavirus spike protein s2 domain |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN116063474B (en) * | 2023-02-09 | 2024-07-23 | 中国海洋大学 | Shark single domain antibody targeting SARS-CoV-2-S1-RBD, application and kit thereof |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN112500480B (en) * | 2020-07-17 | 2022-04-01 | 上海洛启生物医药技术有限公司 | Nanobodies against novel coronaviruses and uses thereof |
-
2021
- 2021-04-22 CA CA3115877A patent/CA3115877A1/en active Pending
-
2022
- 2022-04-22 EP EP22791236.7A patent/EP4326763A4/en active Pending
- 2022-04-22 WO PCT/IB2022/053756 patent/WO2022224203A1/en not_active Ceased
- 2022-04-22 CA CA3216274A patent/CA3216274A1/en active Pending
- 2022-04-22 US US18/556,415 patent/US20240270826A1/en active Pending
- 2022-04-22 JP JP2023560880A patent/JP2024515525A/en active Pending
Non-Patent Citations (1)
| Title |
|---|
| DANIEL WRAPP, DE VLIEGER DORIEN, CORBETT KIZZMEKIA S., TORRES GRETEL M., WANG NIANSHUANG, VAN BREEDAM WANDER, ROOSE KENNY, VAN SCH: "Structural Basis for Potent Neutralization of Betacoronaviruses by Single-Domain Camelid Antibodies", CELL, ELSEVIER, AMSTERDAM NL, vol. 181, no. 5, 28 May 2020 (2020-05-28), Amsterdam NL , pages 1004 - 1015.e15, XP055764639, ISSN: 0092-8674, DOI: 10.1016/j.cell.2020.04.031 * |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN117866903A (en) * | 2024-03-12 | 2024-04-12 | 北京贝来药业有限公司 | Single domain antibody modified stem cells and their use in disease treatment |
| CN117866903B (en) * | 2024-03-12 | 2024-06-04 | 北京贝来药业有限公司 | Single domain antibody modified stem cells and their use in disease treatment |
| WO2026062389A1 (en) * | 2024-09-20 | 2026-03-26 | The Rosalind Franklin Institute | Single domain antibodies against beta coronavirus spike protein s2 domain |
Also Published As
| Publication number | Publication date |
|---|---|
| EP4326763A4 (en) | 2025-09-24 |
| JP2024515525A (en) | 2024-04-10 |
| CA3115877A1 (en) | 2022-10-22 |
| US20240270826A1 (en) | 2024-08-15 |
| EP4326763A1 (en) | 2024-02-28 |
| CA3216274A1 (en) | 2022-10-27 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Silva et al. | Identification of a conserved S2 epitope present on spike proteins from all highly pathogenic coronaviruses | |
| Rossotti et al. | Arsenal of nanobodies shows broad-spectrum neutralization against SARS-CoV-2 variants of concern in vitro and in vivo in hamster models | |
| US10196438B2 (en) | Human antibodies binding to RSV G protein | |
| CA2908921C (en) | Human antibodies binding to rsv g protein | |
| US20240270826A1 (en) | Antibodies that bind sars-cov-2 spike protein | |
| US20230374116A1 (en) | Single domain antibodies that target sars-cov-2 | |
| KR20230058434A (en) | Antigen binding molecules targeting SARS-CoV-2 | |
| WO2011050001A2 (en) | Anti-botulinum neurotoxin antibodies | |
| KR20230035350A (en) | Antigen binding molecules targeting SARS-CoV-2 | |
| CN113583115A (en) | Antibodies against SARS-CoV-2 and uses thereof | |
| US20230227538A1 (en) | Sars-cov2 neutralizing single domain antibody constructs | |
| JP2023534922A (en) | Antigen-binding molecule targeting SARS-CoV-2 | |
| RS66591B1 (en) | ANTIBODIES CAPABLE OF BINDING TO THE SPIKE PROTEIN OF CORONAVIRUS SARS-COV-2 | |
| US20230203134A1 (en) | Potent neutralizing antibodies against sars-cov-2, generation and uses thereof | |
| KR102899181B1 (en) | An antibody specific to C-terminal region of coronavirus nucleocapsid protein and uses thereof | |
| JP2025542128A (en) | Antibodies to metapneumovirus fusion (F) protein and uses thereof | |
| Rossotti et al. | Arsenal of Nanobodies for Broad-Spectrum Countermeasures against Current and Future SARS-CoV-2 Variants of Concerns | |
| KR102899180B1 (en) | An antibody specific to N-terminal region of coronavirus nucleocapsid protein and uses thereof | |
| WO2025083216A1 (en) | Antibodies specific for the spike protein of sars cov-2 coronavirus | |
| EP4594348A1 (en) | Antivirals against human parainfluenza virus | |
| WO2025003500A1 (en) | Antibodies against sars-cov2 spike protein | |
| CA3267635A1 (en) | Antivirals against human parainfluenza virus | |
| CN120441690A (en) | Antibody against RSV and hMPV or its antigen-binding fragment and application thereof | |
| CN117343168A (en) | Coronavirus binding molecules and medicinal uses | |
| CN116472284A (en) | SARS-COV2 neutralizing single domain antibody construct |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 22791236 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2023560880 Country of ref document: JP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 3216274 Country of ref document: CA |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 202317078934 Country of ref document: IN |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2022791236 Country of ref document: EP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2022791236 Country of ref document: EP Effective date: 20231122 |