BR102019017792A2 - Receptáculo proteico, método para produção do receptáculo, método de identificação de patógenos ou de diagnóstico de doenças, e, uso do receptáculo - Google Patents
Receptáculo proteico, método para produção do receptáculo, método de identificação de patógenos ou de diagnóstico de doenças, e, uso do receptáculo Download PDFInfo
- Publication number
- BR102019017792A2 BR102019017792A2 BR102019017792-6A BR102019017792A BR102019017792A2 BR 102019017792 A2 BR102019017792 A2 BR 102019017792A2 BR 102019017792 A BR102019017792 A BR 102019017792A BR 102019017792 A2 BR102019017792 A2 BR 102019017792A2
- Authority
- BR
- Brazil
- Prior art keywords
- seq
- protein
- receptacle
- receptacle according
- amino acid
- Prior art date
Links
Images











Classifications
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/58—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving labelled substances
- G01N33/582—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving labelled substances with fluorescent label
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/002—Protozoa antigens
- A61K39/005—Trypanosoma antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/099—Bordetella
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/35—Allergens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K1/00—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length
- C07K1/04—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length on carriers
- C07K1/042—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length on carriers characterised by the nature of the carrier
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/43504—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from invertebrates
- C07K14/43595—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from invertebrates from coelenteratae, e.g. medusae
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—RNA viruses
- C07K16/102—Coronaviridae (F)
- C07K16/104—Severe acute respiratory syndrome coronavirus 2 [SARS‐CoV‐2]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/70—Vectors or expression systems specially adapted for E. coli
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/569—Immunoassay; Biospecific binding assay; Materials therefor for microorganisms, e.g. protozoa, bacteria, viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55566—Emulsions, e.g. Freund's adjuvant, MF59
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/60—Fusion polypeptide containing spectroscopic/fluorescent detection, e.g. green fluorescent protein [GFP]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/20011—Rhabdoviridae
- C12N2760/20111—Lyssavirus, e.g. rabies virus
- C12N2760/20134—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/20011—Coronaviridae
- C12N2770/20034—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/005—Assays involving biological materials from specific organisms or of a specific nature from viruses
- G01N2333/08—RNA viruses
- G01N2333/165—Coronaviridae, e.g. avian infectious bronchitis virus
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/005—Assays involving biological materials from specific organisms or of a specific nature from viruses
- G01N2333/08—RNA viruses
- G01N2333/175—Bunyaviridae, e.g. California encephalitis virus, Rift valley fever virus, Hantaan virus
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/005—Assays involving biological materials from specific organisms or of a specific nature from viruses
- G01N2333/08—RNA viruses
- G01N2333/18—Togaviridae; Flaviviridae
- G01N2333/181—Alphaviruses or Group A arboviruses, e.g. sindbis, VEE, EEE, WEE or semliki forest virus
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/435—Assays involving biological materials from specific organisms or of a specific nature from animals; from humans
- G01N2333/44—Assays involving biological materials from specific organisms or of a specific nature from animals; from humans from protozoa
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Organic Chemistry (AREA)
- Biochemistry (AREA)
- Biomedical Technology (AREA)
- Virology (AREA)
- Genetics & Genomics (AREA)
- Microbiology (AREA)
- Urology & Nephrology (AREA)
- Hematology (AREA)
- Biophysics (AREA)
- Biotechnology (AREA)
- Analytical Chemistry (AREA)
- Physics & Mathematics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Tropical Medicine & Parasitology (AREA)
- Pharmacology & Pharmacy (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- General Physics & Mathematics (AREA)
- Pathology (AREA)
- Food Science & Technology (AREA)
- Cell Biology (AREA)
- Mycology (AREA)
- Epidemiology (AREA)
- Zoology (AREA)
- General Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Gastroenterology & Hepatology (AREA)
- Plant Pathology (AREA)
- Communicable Diseases (AREA)
- Oncology (AREA)
Abstract
A presente invenção refere-se a um receptáculo proteico capaz de receber diversas sequências de poli-aminoácidos exógenas, concomitantemente, para expressão em diversos sistemas e para diferentes usos. A presente invenção se insere no campo de aplicação da Química, Farmácia, Medicina, Biotecnologia e, mais especificamente, na área de preparações para finalidades biomédicas, uma vez que, se refere ao processo de produção de uma proteína receptáculo como um apresentador de diversas sequências de poli-aminoácidos, simultaneamente, em mais do que quatro diferentes sítios na proteína receptáculo, para uso em diagnósticos ou composições vacinais.
Description
[001] A presente invenção refere-se a um receptáculo proteico capaz de receber diversas sequências de poli-aminoácidos exógenas, concomitantemente, para expressão em diversos sistemas e para diferentes usos. A presente invenção se inserenocampo de aplicação da Química, Farmácia, Medicina, Biotecnologia e, mais especificamente, na área de preparações para finalidades biomédicas, uma vez que, se refere ao processo de produção de uma proteína receptáculo como um apresentador de diversas sequências de poli-aminoácidos, simultaneamente, em mais do que quatro diferentes sítios na proteína receptáculo, para uso em diagnósticos ou composições vacinais.
[002] Este documento apresenta várias referências ao longo do texto, as quais estão indicadas dentro de parênteses. As informações publicadas nestas referências estão aqui incluídas a fim de melhor descrever o estado da arte a qual a presente invenção pertence.
[003] A proteína verde fluorescente (GFP), produzida por um cnidário, a água viva Aequorea victoria, emite alta fluorescência na zona verde do espectro visível (Prasher et al., Gene 15, 111 (2): 229-33, 1992) e, classicamente, vem sendo utilizada como um marcador da expressão e de localização de genes, e, desta forma, conhecida como proteína repórter. Após a primeira observação de que a proteína GFP poderia emitir fluorescência, diversos usos foram descritos para a proteína.
[004] A utilização da GFP como proteína repórter tem sido patenteada para diferentes usos e em diferentes sistemas. O pedido de patente US2018298058 utiliza a GFP como sistema de produção e purificação de proteínas. O pedido de patente CN108303539 utiliza a GFP como insumo para teste de detecção de cânceres; o pedido CN108220313 apresenta um método de fusão e expressão de GFP de alta produção. A solicitação CN108192904 reivindica uma proteína de fusão GFP com capacidade de se inserir em membranas biológicas; o pedido CN107703219 já enfatiza o uso da GFP no estudo metabolômico de células mesenquimais. Ainda a solicitação de patente US2018016310 apresenta variações de GFP de fusão “superfolder”. Há diversas patentes, como esses exemplos: US6054321, US6096865, US6027881, US6025485 que apresentam GFPs mutantes para aumentar a expressão de fluorescência ou modificar os picos de comprimento de onda de fluorescência. Desse modo, é evidente que a proteína GFP, desde sua descrição, tem sido utilizada e patenteada para diversos usos como proteína repórter.
[005] Moléculas repórteres são frequentemente usadas em sistemas biológicos para monitorar a expressão de genes. A proteína GFP trouxe grande inovação neste cenário ao prescindir do uso de qualquer substrato ou cofator, ou seja, não requerendo a adição de qualquer outro reagente para que possa ser visualizada, como ocorre para a maioria de outras proteínas repórteres. Outra vantagem apresentada pela GFP é sua capacidade de apresentar autofluorescência, não necessitando de marcações fluorescentes que, por diversas causas, não apresentam a sensibilidade ou especificidade adequadas para o seu uso.
[006] Em face dessa característica, sua auto-produção de fluorescência verde detectável, a GFP tem sido largamente utilizada para estudar a expressão gênica e a localização de proteínas, sendo considerada como uma das mais promissoras proteínas repórteres na literatura.
[007] Em seu uso como proteína repórter, o gene que codifica a GFP pode ser empregado na produção de proteínas de fusão, ou seja, um determinado gene de interesse é fusionado ao gene que codifica a GFP. O cassete de genes fusionados pode ser inserido em um sistema vivo, permitindo a expressão dos genes fusionados e o monitoramento da localização intracelular dessa proteína de interesse (Santos-Beneit & Errington, Archives Microbiology, 199 (6): 875-880, 2017; Belardinelli & Jackson, Tuberculosis (Edinb), 105: 13-17, 2017; Wakabayashi et al, International Journal Food Microbiology, 19, 291: 144-150, 2018; Cai et al, Viruses, 20; 10 (11), 2018).
[008] A proteína GFP também tem sido útil como um arcabouço para a apresentação de peptídeos ou mesmo de bibliotecas de peptídeos, tanto em leveduras quanto em sistemas celulares de mamíferos (Kamb et al., Proc. Natl. Acad Sci. USA, 95: 7508-7513, 1998; WO 2004005322). A proteína GFP, como uma proteína arcabouço para a apresentação de peptídeos randômicos, pode ser utilizada para definir as características de uma biblioteca de peptídeos.
[009] Avanços no desenvolvimento denovas variações de GFP buscam alcançar melhorias nas propriedades da proteína, a fim de produzirno vos reagentes úteis para uma grande gama de propósitos de pesquisa. no vas versões de GFP têm sido desenvolvidas, via mutações, contendo sequências de DNA otimizadas para produção aumentada em sistemas celulares humanos, ou seja, proteínas GFP humanizadas (Cormack, et al., Gene 173, 33-38, 1996; Haas, et al., Current Biology 6, 315-324, 1996; Yang, et al., Nucleic Acids Research 24, 4592-4593, 1996).
[0010] Em uma destas versões, descreveu-se a proteína verde fluorescente melhorada, a “enhanced green fluorescent protein” (eGFP) (Heim & Cubitt, Nature, 373, pp. 663-664, 1995).
[0011] A GFP, codificada pelo gene gfp10, originário de um cnidário, a água-viva Aequorea victoria, é uma proteína de 238 aminoácidos. A proteína tem a capacidade de absorver a luz azul (com pico principal de excitação em 395 nm) e emitir luz verde, a partir de um cromóforo nocentro da proteína (pico principal de emissão em 509 nm) (Morin & Hastings, Journal Cell Physiology, 77(3): 313-8, 1971; Prasher et al., Gene 15, 111 (2): 229-33, 1992). O cromóforo é composto de seis peptídeos e se iniciano aminoácido 64, e é derivado da sequência primária dos aminoácidos, através da ciclização e oxidação da serina, tirosina e glicina (nas posições 65, 66 e 67) (Shimomura, 104 (2), 1979; Cody et al., Biochemistry, 32 (5): 1212-8, 1993). A luz emitida pela GFP é independente da espécie biológica celular onde é expressa e não requer qualquer tipo de substrato, cofatores ou produtos de genes adicionais da A. victoria (Chalfie et al., Science, 263 (5148): 802-5, 1994). Esta propriedade da GFP permite que sua fluorescência seja detectada em outras células vivas que não a A. victoria, desde que possa ser processada no sistema de expressão proteico da célula (Ormo et al, Science 273: 13921395, 1996;. Yang et al, Nature Biotech 14: 1246-1251, 1996).
[0012] A estrutura básica de uma GFP consiste de onze cadeias beta pregueadas antiparalelas, que se enovelam e formam uma estrutura terciária em forma de um beta barril. Cada cadeia é ligada à seguinte por um domínio de alça que se projeta para a superfície superior e inferior do barril, interagindo com o meio ambiente. Por convenção, cada cadeia e alça pode ser identificada por uma numeração a fim de melhor descrever a proteína.
[0013] Experimentos de mutação dirigida têm demonstrado que diversas propriedades bioquímicas da proteína GFP são decorrentes dessa estrutura de barril. E, por isso, mudanças de aminoácidos na estrutura primária são responsáveis por acelerar o enovelamento da proteína, reduzir a agregação dos produtos decorrentes da tradução e aumentar a estabilidade da proteína em solução.
[0014] Uma determinada alça se desloca para a cavidade do barril proteico, formando uma alfa hélice que é responsável pelas propriedades fluorescentes da proteína (Crone et al, GFP-Based Biosensors, InTech, 2013). Algumas intervenções na estrutura da proteína podem interferir na capacidade de emitir fluorescência. Mutações em determinados aminoácidos podem mudar desde a intensidade de emissão de fluorescência ao comprimento de onda, alterando a cor emitida. A mutação da Tyr66, resíduo interno participante do cromóforo fluorescente, pode gerar um grande número de variantes de proteínas fluorescentes com a estrutura do cromóforo alterado ou do ambiente circunvizinho. Essas alterações interferem na absorção e emissão de luz em diferentes comprimentos de onda, produzindo um grande leque de distintas cores emitidas (Heim & Tsien, Current Biology, 6 (2): 178-82, 1996).
[0015] Mudanças no pH também podem interferir na intensidade de fluorescência. Em pH fisiológico, a GFP exibe absorção máxima a 395nm, enquanto que em 475 nm absorve menor quantidade de luz. Entretanto, o aumento do pH para cerca de 12,0, faz com que a absorção máxima ocorra na faixa de 475 nm, e em 395 nm tenha absorção diminuída (Ward et al, Photochemistry and Photobiology, 35(6): 803-808, 1982).
[0016] A estrutura compacta do cerne proteico permite que a GFP seja altamente estável mesmo em condições adversas, tais como, tratamento por proteases, fazendo com que a proteína seja extremamente útil como uma proteína repórter de modo geral.
[0017] Há diferentes versões da GFP, sempre buscando aperfeiçoar a proteína ao agregar uma nova funcionalidade ou retirando alguma limitação. A eGFP se apresenta como um aprimoramento que permite uma maior flexibilidade da proteína em face das modificaçõesn os aminoácidos F64L e S65T (Heim et al, Nature 373: 663-664, 1995; Li et al, Journal Biology Chemistry, 272 (45): 28545-9, 1997). Este aprimoramento permite que a GFP alcance tanto sua forma tridimensional esperada quanto sua capacidade de expressar fluorescência, mesmo ao abrigar sequências heterólogas em sua sequência proteica (Pedelacq et al, Nat Biotechnology, 24 (1): 79-88, 2006).
[0018] A GFAb, é uma versão da proteína modificada que aceita as sequências exógenas em dois domínios de alça. Em seu desenvolvimento, foram necessárias várias rodadas de evolução direta para que se pudessem selecionar três clones proteicos mutados que suportassem a inserção de peptídeos exógenos nas duas regiões proximais, quais sejam, Glu-172-Asp-173 e Asp-102-Asp-103. Os autores ao utilizarem proteínas não mutadas comprovaram que a inserção de dois peptídeos exógenos impedia a produção de fluorescência da GFP e a expressão proteica na superfície celular da levedura. A inserção simultânea, em somente duas regiões, foi possível após uma série de mutações e seleções, mas ainda resultando em grande perda das atividades inerentes a GFP, por vezes, inviabilizando sua produção ou expressão do inserto (Pavoor et al., PNAS 106 (29): 11895-11900, 2009).
[0019] Mutantes circularmente permutados para os N- e C-terminais também têm demonstrado que eGFP é passível de manipulação na sequência codificadora sem comprometer os aspectos estruturais do cerne da proteína (Topell et al., FEBS Letters 457(2): 283-289, 1999). Contudo, a análise de 20 variantes proteicas permutadas circulares demonstrou a baixa tolerância das proteínas à inserção de um novo terminal e, em sua maioria, que perdem a capacidade de formar o cromóforo. Este fato indica que a manipulação da sequência proteica pode interferir drasticamente em suas características ou mesmo em sua expressão celular.
[0020] Várias tentativas têm sido feitas para inserir simultaneamente múltiplos epítopos nas regiões de alças da GFP com o objetivo de alcançar o uso da proteína para reações de ligação específicas a um alvo. Contudo, todos esses esforços têm apresentado sucesso limitado em face da sensibilidade estrutural da GFP e seu cromóforo.
[0021] Outras proteínas mutantes da proteína GFP apresentam versões melhoradas que emitem outros tipos de espectro de luz fluorescente. Por exemplo, Heim et al (Proc Natl Acad Sci USA, 91 (26): 12501-4, 1994) descreveram uma proteína mutante que emite fluorescência azul por conter uma histidina ao invés de uma tirosina no aminoácido 66. Heim et al (Nature, 373 (6516): 663-4, 1995) posteriormente também descreveram uma proteína GFP mutante, pela substituição de uma serina por uma treonina no aminoácido 65, que apresenta um espectro muito similar ao do obtido com a Renilla reniformis, que tem um coeficiente de extinção por monômero 10 vezes maior que o pico de comprimento de onda da GFP nativa proveniente da Aequorea. Outros documentos de patente descrevem proteínas GFP mutantes apresentando espectros de emissão de luz diferentes do verde, como azul, vermelho (US 5625048, WO 2004005322).
[0022] Ainda, outras proteínas mutantes de GFP apresentam espectros de excitação otimizados para serem utilizados especificamente em determinados equipamentos de citômetro de fluxo de laser por argônio (FACS) (US 5804387). Ainda há descrição de proteínas mutantes de GFP modificadas para serem melhor expressas em sistemas celulares de plantas (WO1996027675). O documento de patente US5968750 apresenta uma GFP humanizada que foi adaptada para ser expressa em células de mamíferos, inclusive humanas. A GFP humanizada incorpora códons preferenciais para leitura nos sistemas de expressão de genes de células humanas.
[0023] No estado da arte, é nítida a capacidade da GFP de abrigar genes em suas extremidades 5’ ou 3’ sem que haja interferência na sua expressão, enovelamento tridimensional e produção de fluorescência, como pode ser observado nos documentos de patente acima elencados. Adicionalmente, a GFP tem sido utilizada como suporte para a exibição de peptídeos ou mesmo bibliotecas de peptídeos in vivo. No caso de bibliotecas de peptídeos, a GFP pode auxiliar na apresentação de peptídeos randômicos, e assim, na definição das características da biblioteca de peptídeos (Kamb et al., Proc. Natl. Acad. Sci. USA, 95:7508-7513, 1998; WO 2004005322).
[0024] Por exemplo, Abedi et al (1998, Nucleic Acids Res. 26: 623300) inseriram peptídeos em proteínas GFP de Aequorea victoria, em regiões de alças expostas e demonstraram que as moléculas de GFP retêm a autofluorescência quando expressas em leveduras e Escherichia coli. Os autores ainda demonstraram que a fluorescência de um arcabouço de GFP pode ser utilizada para monitorar a diversidade de peptídeos, assim como a presença ou expressão de um dado peptídeo em uma determinada célula. Entretanto, a taxa de fluorescência das moléculas do arcabouço de GFP é relativamente baixa em comparação com a GFP natural. Kamb e Abedi (US 6025485) prepararam bibliotecas de arcabouços de GFP a partir de proteína verde fluorescente melhorada (eGFP) a fim de amplificar a intensidade de fluorescência.
[0025] Adicionalmente, Peele et al (Chem. & Bio. 8: 521-534, 2001) testaram bibliotecas de peptídeos utilizando eGFP como arcabouço com diferentes viéses estruturais em células de mamíferos. Anderson et al ainda amplificaram a intensidade de fluorescência ao inserirem peptídeos nas alças de GFP com ligantes de tetraglicina (US20010003650). Happe et al descreveram uma GFP humanizada que pode ser expressa em grandes quantidades em sistemas de células de mamíferos, tolera inserções de peptídeos e preserva a autofluorescência (WO 2004005322).
[0026] Entretanto, há ainda uma necessidade,no campo tecnológico, de arcabouços de moléculas de GFP que não somente apresentem fluorescência em intensidades apropriadas, mas também que possam ser expressas em altos níveis em sistemas celulares.
[0027] Há uma variabilidade na tolerância, entre as moléculas de GFP, para apresentação dos peptídeos enquanto retém a autofluorescência. Assim, há uma necessidade no estado da arte de desenvolver-se GFPs que possam ser expressas em altos níveis e tolerarem inserções, enquanto preservam a autofluorescência.
[0028] Ademais da habilidade de suportar a expressão de genes nas extremidades, a GFP pode permitir a inserção de epítopos nas alças superficiais da molécula expostas para o meio. Várias tentativas têm sido feitas para inserir simultaneamente múltiplos peptídeos nas regiões de alça da GFP, que pudessem permitir o uso da proteína para reações de ligação específica a um alvo. Contudo, todos esses esforços apresentaram sucesso limitado em face da sensibilidade estrutural da GFP e seu cromóforo.
[0029] Pavoor e colaboradores empreenderam esforços para desenvolver uma proteína modificada para aceitar sequências exógenas em dois domínios de alça. Em seu desenvolvimento, foram necessárias várias rodadas de evolução direta para que se pudessem selecionar três clones proteicos mutados que suportassem a inserção de peptídeos exógenos nas duas regiões proximais, quais sejam, Glu-172-Asp-173 e Asp-102-Asp-103. Os autores ao utilizarem proteínas não mutadas comprovaram que a inserção de dois peptídeos exógenos impedia a produção de fluorescência da GFP e a expressão proteica na superfície celular da levedura. A inserção simultânea, em somente duas regiões, foi possível após uma série de mutações e seleções, mas ainda resultava em grande perda das atividades inerentes a GFP, por vezes, inviabilizando sua produção ou expressão do inserto (Pavoor et al., PNAS 106 (29): 11895-11900, 2009).
[0030] Abedi et al (Nucleic Acids Research 26(2): 623-30, 1998) apresentaram 10 posições da proteína, em regiões de alças, das quais 8 posições entre folhas-β, para expressão de peptídeos. A proteína quimérica seria útil para experimentos que requeressem a expressão intracelular, e, portanto, a ininterrupção de fluorescência seria um fator limitante. Nesse estudo, somente três proteínas quiméricas (aquelas com sítios de inserção nos aminoácidos 157-158 172-173 e 194-195) apresentaram fluorescência (diminuídas a um quarto do original); e somente dois sítios de inserção (estudados separadamente) puderam abrigar peptídeos sem que houvesse perda da fluorescência. Os autores do trabalho ainda concluíram que “é curioso como a GFP é tão sensível a perturbações estruturais mesmo que seja nas folhas β”.
[0031] Li et al (Photochemistry and Photobiology, 84(1): 111-9, 2008) apresentam um estudo da proteína quimérica (red fluorescente protein -RFP), apontando nesta proteína seis sítios geneticamente distintos localizados em três diferentes alças onde sequências de cinco resíduos podem ser inseridos sem interferir na capacidade da proteína ser fluorescente. Entretanto, os autores não demonstraram o uso concomitante desses sítios para inserção de diferentes peptídeos.
[0032] O pedido de patente WO02090535 apresenta uma GFP fluorescente com inserções, não simultâneas, de peptídeos em 5 diferentes alças da proteína. O pedido de patente, em seu relatório descritivo, indica a possibilidade de inserção de peptídeos em mais de uma alça da proteína ao mesmo tempo, aumentando a complexidade da biblioteca e permitindo a apresentação em uma mesma face da proteína. Entretanto, o pedido de patente não atesta esta possibilidade, posto que somente apresenta ensaios de inserção de peptídeos, um de cada vez, em 5 diferentes alças proteicas. Vale destacar que o texto ainda enfatiza que as alças 1 e 5 não se apresentam como bons sítios de inserção, pois a inserção de peptídeos nestes sítios impediram a expressão da proteína. Outros documentos de patente ainda apresentam variações da GFP visando a expressão de peptídeos nas alças da proteína, entretanto, estes estudos não comprovam a viabilidade de expressão simultânea de mais de 4 peptídeos em diferentes sítios de inserção nas alças da proteína GFP, sem que houvesse perda de alguma de suas características essenciais (WO02090535, US2003224412, WO200134824).
[0033] A presente invenção refere-se a um receptáculo proteico capaz de apresentar diversas sequências de poli-aminoácidos exógenas, concomitantemente, em mais do que quatro diferentes sítios na proteína receptáculo.
[0034] Outra realização da presente invenção se refere à expressão eficiente do dito receptáculo proteico, baseado na sequência de uma GFP, carreando uma ou duas, ou mais do que duas, três ou quatro, ou mais do que dez sequências de poli-aminoácidos exógenas, concomitantemente, em dez diferentes sítios na proteína receptáculo, em um sistema celular. Especificamente, a presente invenção se refere à expressão eficiente do dito receptáculo proteico apresentando sequências de poli-aminoácidos exógenas, em dez diferentes sítios proteicos, simultaneamente. Mais especificamente, a presente invenção se refere à expressão eficiente do dito receptáculo carreando as ditas sequências de poli-aminoácidos exógenas, inseridos em diferentes sítios proteicos, sem contudo, perder suas características inerentes, tais como, autofluorescência.
[0035] Ainda, a presente invenção se refere a um sistema para expressão concomitante de múltiplas sequências de poli-aminoácidos, utilizando um mesmo receptáculo proteico, baseado na GFP, útil para diferentes usos, tais como reagente para pesquisa, para diagnóstico ou para composições vacinais. Especificamente, o dito sistema de expressão pode atuar como reagente de pesquisa útil para purificar anticorpos ao se ligar em epítopos. Adicionalmente, o dito sistema de expressão pode ainda atuar para uso em técnicas imunológicas e/ou moleculares para diagnósticos de doenças crônicas e infecciosas. Ainda, o dito sistema de expressão pode ser útil para uso em composições vacinais contendo múltiplos antígenos para imunização de animais e humanos.
[0036] Também é uma realização da presente invenção, o método para produção concomitante de múltiplas sequências de poli-aminoácidos, utilizando um mesmo receptáculo proteico, baseado na GFP, útil para diferentes usos, tais como reagente para pesquisa, para diagnóstico ou para composições vacinais.
[0037] Figura 1 - Purificação da proteína PlatCruzi por cromatografia de afinidade. (A) Perfil de eluição da proteína receptáculo numa coluna de níquel usando um sistema de cromatografia líquida Àktapurifier. (B) Análise por eletroforese em gel de poliacrilamida (SDS PAGE) dos eluatos 13-26 coletados. A eluição foi realizada com tampão B e em ordem crescente. PM - Marcador de peso molecular.
[0038] Figura 2 - Reatividade da PlatCruzi, por ELISA, com soros das Referências Biológicas de Padrão Internacional de Trypanosoma cruzi cedidos pela OMS. (A) Pool de soros de pacientes reconhecendo as cepas TcI, denominado de IS 09/188. (B) Pool de soros de pacientes reconhecendo as cepas TcII, denominado de IS 09/186.
[0039] Figura 3 - Determinação do título de anticorpos de soros de pacientes com doença de Chagas crônica usando como antígeno a proteína receptáculo PlatCruzi. Os soros foram cedidos pelos LACENS e uma concentração de 500 ng/poço e a diluição de soros 1: 50 - 1: 1000 foram usados nos ELISAS.
[0040] Figura 4 - Desempenho do antígeno de PlatCruzi por ELISA frente a soro de pacientes com diferentes doenças. O antígeno de PlatCruzi foi usado na concentração de 500 ng/poço e os soros diluídos 1:250.
[0041] Figura 5 - Detecção de epítopos específicos do vírus da raiva por soro de coelho anti-RxRabies2. Anticorpos de coelho imunizados com RxRabies2 foram purificados por cromatografia de afinidade com RxRabies2 e usados como anticorpo primário, por Western blot, para detectar RxRabies2 em um extrato bruto (*; coluna 2) ou semi-purificado (f) em duas diferentes concentrações de 1x e 0.5x (colunas 4 e 6, respectivamente). Controles negativos: proteína receptáculo Rx (coluna 3) e PlatCruzi (em duas concentrações: 1x e 0.5x nas colunas 5 e 7, respectivamente).
[0042] Figura 6 - Análise da proteína RxHoIgG3 por eletroforese em gel de poliacrilamida (SDS PAGE). A, extrato solúvel de E.coli não produtora de RxHoIgG3. B, Fração insolúvel em meio aquoso de bactéria produtora de RxHoIgG3. C, Fração solúvel da bactéria produtora de RxHoIgG3. As setas indicam a posição da proteína RxHoIgG3.
[0043] Figura 7 - Detecção de anticorpos IgM anti-RxOro por ELISA. C-: controle negativo (soro de paciente sem infecção pelo vírus Oropouche); C+: soro positivo padrão para infecção pelo vírus Oropouche. Pacientes: casos suspeitos de infecção por infecção pelo vírus Oropouche. Oro+: positivos para a infecção por Oropouche (detecção de anticorpos IgM anti-RxOro). Oro- (controle negativo): Ausência de reação IgM para RxOro. Concentração da proteína: 0,288μg/μL. Cutoff: 0,0613.
[0044] Figura 8 - Eletroforese em gel de poliacrilamida (SDS-PAGE) representando a produção das proteínas PlatCruzi, RxMayaro_IgG e RxMayaro_IgM. As colunas verticais 1 a 8, indicam: 1) peso molecular; 2) extrato total bacteriano sem indução de proteína recombinante; 3) extrato total bacteriano após indução da produção de PlatCruzi; 4) extrato total bacteriano após indução da produção de RxMayaro_IgG; 5) extrato total bacteriano após indução da produção de RxMayaro_IgM; 6) proteínas bacterianas insolúveis após indução da produção de PlatCruzi; 7) proteínas bacterianas insolúveis após indução da produção de RxMayaro_IgG; 8) proteínas bacterianas insolúveis após indução da produção de RxMayaro_IgM. A setas apontam para as bandas que representam PlatCruzi (colunas 3 e 6), RxMayaro_IgG (colunas 4 e 7) e RxMayaro_IgM (colunas 5 e 8).
[0045] Figura 9 - Reatividade de soros de pacientes positivos para o vírus Mayaro (S MAY) e indivíduos saudáveis (S N), por ELISA, empregando a proteína RxMayaro_IgG. A revelação foi realizada com imunoglobulina anti-IgG conjugado a enzima fosfatase alcalina (Cutoff = 0.0210).
[0046] Figura 10 - Reatividade de soros de indivíduos considerados saudáveis (S N) e positivos para vírus Mayaro (S MAY), por ELISA, empregando a proteína RxMayaro_IgM. A revelação foi realizada com imunoglobulina anti-IgM conjugada a enzima fosfatase alcalina. (Cut off = 0.0547).
[0047] Figura 11 - Eletroforese em gel de poliacrilamida (SDS-PAGE) apresentando a produção das proteínas insolúveis (I) e solúveis (S) de PlatCruzi, TxCruzi, RxPtx, TxNeuza e RxYFIgG. As colunas 1 a 10 contém: 1) proteínas insolúveis de bactéria induzida para produzir PlatCruzi; 2) proteínas solúveis de bactéria induzida para produzir PlatCruzi; 3) proteínas insolúveis de bactéria induzida para produzir TxCruzi; 4) proteínas solúveis de bactéria induzida para produzir TxCruzi; 5) proteínas insolúveis de bactéria induzida para produzir RxPtx; 6) proteínas solúveis de bactéria induzida para produzir RxPtx; 7) proteínas insolúveis de bactéria induzida para produzir TxNeuza; 8) proteínas solúveis de bactéria induzida para produzir TxNeuza; 9) proteínas insolúveis de bactéria induzida para produzir RxYFIgG; 10) proteínas solúveis de bactéria induzida para produzir RxYFIgG. A setas apontam para as bandas que representem PlatCruzi (colunas 1 e 2), TxCruzi (colunas 3 e 4), RxPtx (colunas 5 e 6), TxNeuza (colunas 7 e 8) e RxYFIgG (colunas 9 e 10).
[0048] Ao longo de todo este documento, algumas abreviações serão utilizadas. Segue abaixo um rol das abreviações:
Com relação às bases nitrogenadas:
C = citosina; A = adenosina; T = timidina; G = guanosina Com relação aos aminoácidos:
I = isoleucina; L = leucina; V = valina; F = fenilalanina; M = metionina; C = cisteína; A = alanina; G = glicina; P = prolina; T = treonina; S = serina; Y = tirosina; W = triptofano; Q = glutamina; N = asparagina; H = histidina; E = ácido glutâmico; D = ácido aspártico; K = lisina; R = arginina.
Com relação às bases nitrogenadas:
C = citosina; A = adenosina; T = timidina; G = guanosina Com relação aos aminoácidos:
I = isoleucina; L = leucina; V = valina; F = fenilalanina; M = metionina; C = cisteína; A = alanina; G = glicina; P = prolina; T = treonina; S = serina; Y = tirosina; W = triptofano; Q = glutamina; N = asparagina; H = histidina; E = ácido glutâmico; D = ácido aspártico; K = lisina; R = arginina.
[0049] A presente invenção é dirigida à produção e uso de um receptáculo proteico, baseado na sequência de uma proteína verde fluorescente, aqui denominada GFP, em uma variedade de métodos e composições que exploram a capacidade do dito receptáculo proteico em apresentar concomitantemente diversas sequências de poli-aminoácidos exógenas, diferentes ou idênticas em mais do que quatro diferentes sítios proteicos, e ainda, exibir intensidades adequadas de fluorescência, ser eficientemente expresso em sistemas celulares de produção de proteína e ser útil como reagente para pesquisa, diagnóstico ou em composições vacinais.
[0050] Esta invenção também provê uma proteína receptáculo isolada, produzida por qualquer sistema de expressão, a partir de uma molécula de DNA, composta por um elemento regulatório contendo a sequência nucleotídica codificadora da proteína receptáculo de escolha. A proteína receptáculo isolada pode ainda ser modificada in vitro para diferentes usos.
[0051] A proteína receptáculo pode ainda sofrer modificações necessárias para o desenvolvimento de seus usos, através da inserção de elementos acessórios. À proteína receptáculo podem ser adicionadas sequências que auxiliem no processo de purificação, tais como, mas não limitados a, cauda de poli-histidina, proteína ligadora de quitina, proteína ligadora de maltose, proteína ligadora de calmodulina, strep-tag e GST. Ainda podem ser incorporadas à proteína receptáculo sequências para estabilização tal como thiorodixino. Também podem ser adicionadas à proteína receptáculo, sequências que auxiliem em qualquer processo de detecção por anticorpos, tais como, sequências V5, Myc, HÁ, Spot, FLAG.
[0052] Ainda podem ser adicionados à proteína receptáculo sequências que atuem como sítios de clivagem alvo (sitio catalítico) para proteases, que permitam separar a proteína principal dos elementos acessórios acima, incluídos com o estrito intuito de otimizar a produção ou purificação da proteína, mas não contribuindo para o uso final aventado. Essas sequências, contendo sítios que servem como alvos de proteases, podem ser inseridos em qualquer lugar e incluem, mas não são limitados a, trombina, fator Xa, enteropeptidase, PreScission, TEV (Kosobokova et al., Biochemistry, 8: 187-200, 2015). Ainda outra sequência acessória marcadora de proteases pode ser a AviTag, que permite a biotinilação específica em um único ponto durante ou após a expressão proteica. Nesse sentido, é possível criar proteínas marcadas pela combinação de diferentes elementos (Wood, Current Opinion in Structural Biology, 26: 54-61, 2014).
[0053] A sequência de DNA codificadora para a proteína receptáculo difere da sequência de DNA de formas de GFP que ocorrem na natureza, em termos de identidade ou localização de um ou mais resíduos de aminoácidos, seja pela deleção, adição ou substituição de aminoácidos. Mas, ainda preservam algumas ou todas as características inerentes às formas que ocorrem na natureza, tais como, mas não limitadas a, produção de fluorescência, formato tridimensional característico, capacidade de ser expressa em diferentes sistemas e capacidade de receber peptídeos exógenos.
[0054] A sequência de DNA codificadora para a proteína receptáculo da presente invenção inclui: a incorporação de códons preferenciais para a expressão por determinados sistemas de expressão; a inserção de sítios de clivagem para enzimas de restrição; a inserção de sequências otimizadas para facilitação de construção de vetores de expressão; a inserção de sequências facilitadoras para abrigar as sequências de poli-aminoácidos de escolha a serem introduzidos na proteína receptáculo. Todas essas estratégias já bastante conhecidas no estado da arte.
[0055] A invenção também está direcionada para a produção e uso dos receptáculos proteicos, “Platform”, “Rx” e “Tx”, e de suas sequências de aminoácidos (descritas nas SEQ IDno. 1, SEQ IDno. 3 e SEQ IDn°. 77, respectivamente), de suas sequências de nucleotídeos (descritas nas SEQ IDn° . 2, SEQ IDno. 4 e SEQ IDno. 78, respectivamente) e de suas sequências de aminoácidos incluindo as sequências de poli-aminoácidos exógenos de escolha (descritas nas SEQ IDno.18, SEQ IDno. 20, SEQ IDno.31, SEQ IDno.33, SEQ IDno. 45, SEQIDn°. 51, SEQIDno.64, SEQIDno. 75, SEQIDno.88 e SEQIDno. 90).
[0056] Adicionalmente, a invenção ainda provê elementos genéticos adicionados das sequências de nucleotídeos codificadoras das proteínas receptáculo, tais como nas sequências descritas nas SEQ IDno. 2, SEQ IDno. 4 e SEQ IDno. 78. Ainda adicionalmente, provê tais elementos contendo sequências de nucleotídeos codificadoras das proteínas receptáculo adicionadas do DNA codificante para as sequências de poli-aminoácidos exógenos de escolha. Tais elementos genéticos contendo as sequências descritas nas SEQ IDno.. 17, SEQ IDno.. 19, SEQ IDno.. 32, SEQ IDno. 34, SEQ IDno.46, SEQIDno. 52, SEQIDno. 63, SEQIDno. 76, SEQIDno. 89 e SEQIDno. 91.
[0057] Elementos regulatórios requeridos para a expressão da proteína receptáculo incluem sequências de promotores para ligar a RNA polimerase e sequências de início de translação para a ligação ao ribossomo. Por exemplo, um vetor de expressão bacteriano deve incluir um codón de iniciação, um promotor adequado ao sistema celular e para início de translação uma sequência de Shine-Dalgarno. De igual forma, um vetor de expressão eucarioto inclui um promotor, um códon de início, um sinal de poliadenilação processo a jusante e um códon de término. Tais vetores podem ser obtidos comercialmente, ou construídos a partir de sequências já conhecidas do estado da arte.
[0058] A invenção ainda provê células contendo sequências de nucleotídeos codificadoras da proteína receptáculo, ou da proteína receptáculo adicionada do DNA codificante para as sequências de poli-amioácidos exógenos de escolha, para atuarem como sistemas de expressão para a proteína receptáculo. As células podem ser células bacterianas, fúngicas, leveduras, células de inseto, de planta ou mesmo animais. As sequências de DNA codificadoras das proteínas receptáculo adicionadas do DNA codificante para as sequências de poli-amioácidos exógenos de escolha também podem ser inseridos em vírus que podem ser utilizados para expressão e produção da proteína receptáculo, como sistemas de entrega, tais como baculovírus, adenovírus, vírus associados a adenovírus, alfavírus, herpes vírus, pox vírus, retrovírus, lentivirus, mas não limitados a esses.
[0059] Há uma grande variedade de métodos para introdução de material genético exógeno em células, todos já bastante conhecidos no estado da arte. Por exemplo, material de DNA exógeno pode ser introduzido em uma célula pela tecnologia de precipitação de fosfato de cálcio. Outras tecnologias, tais como tecnologias que utilizem eletroporação, lipofection, microinjeção, vetores retrovirais, e outros sistemas de vetores virais, como sistemas virais associados a adenovírus podem ser utilizados para desenvolvimento dessa invenção.
[0060] Esta invenção provê um organismo vivo compreendendo, ao menos, células que contenham uma molécula de DNA contendo um elemento regulatório para expressão da sequência codificadora da proteína receptáculo. A invenção é aplicável para produção de proteínas receptáculo em animais vertebrados, animais não vertebrados, plantas e microrganismos.
[0061] A expressão das proteínas receptáculos pode ser realizada em, mas não limitadas a, células de Escherichia coli, Bacillus subtilis, Saccharomyces cerevisiae, Pichia pastoris, Pichia methanolica, Candida boidinii, Pichia angusta, células de mamíferos, tais como, células CHO, HEK293 ou células de insetos, tais como, células Sf9. Todos os sistemas de expressão proteico de procariotos ou eucariotos, conhecidos no estado da arte, estão aptos a produzir as proteínas receptáculo.
[0062] Em um desenvolvimento da presente invenção, um vírus ou bacteriófago, carreando a sequência codificadora para a proteína receptáculo, pode infectar um tipo particular de bactéria ou célula eucariótica e propiciar a expressão da proteína receptáculo nesse sistema celular. A infecção pode ser facilmente observada através da detecção da expressão da proteína receptáculo. De forma similar, um vírus de célula eucariótica de planta ou animal, carreando a sequência codificadora da proteína receptáculo, pode infectar um tipo celular específico e propiciar a expressão da proteína receptáculo no sistema celular eucarioto.
[0063] A invenção ainda provê técnicas para produção das proteínas receptáculo contendo os poli-aminoácidos exógenos de escolha. Esta invenção apresenta um eficiente método para expressão de proteínas receptáculo contendo os poli-aminoácidos exógenos de escolha o qual promove a produção da proteína de interesse em grande quantidade. Métodos para a produção das proteínas receptáculos produção podem ser realizados em diferentes sistemas celulares, tais como, leveduras, plantas, células de plantas, células de insetos, células de mamíferos e animais transgênicos. Cada sistema pode ser utilizado pela incorporação de uma sequência de ácidos nucleicos códon-otimizada, a qual pode gerar a sequência de aminoácidos desejada, em um plasmídeo apropriado para o determinado sistema celular. Esse plasmídeo pode conter elementos, que conferem um número de atributos no sistema de expressão, que incluem, mas não estão limitados a, sequências que promovem a retenção e replicação, marcadores selecionáveis, sequências promotoras para transcrição, sequências estabilizadoras do RNA transcrito, sítio de ligação do ribossomo.
[0064] A transição de um plasmídeo para outro vetor pode ser alcançada pela alteração da sequência de nucleotídeos ao se adicionar ou eliminar sítios de restrição sem modificar a sequência de aminoácidos, que pode ser realizada pela reação de polimerização em cadeia.
[0065] A expressão otimizada em outros sistemas pode ser alcançada pela alteração da sequência de nucleotídeos para adicionar ou eliminar sítios de restrição e também para otimizar códons para alinhamento dos códons preferenciais donovo sistema de expressão, sem, contudo, alterar a sequência de aminoácidos final.
[0066] As proteínas receptáculo também podem ser produzidas por sistemas múltiplos de biologia sintética. A geração de genes totalmente sintéticos está ligada a basicamente três sistemas de síntese baseados em ligação, amplamente descritos no estado da arte.
[0067] Esta invenção provê métodos para produção da proteína receptáculo a partir da utilização de sistema de expressão de proteínas que compreendem a: introdução em células competentes de interesse de sequência de DNA codificadora para a proteína receptáculo adicionada do DNA codificante para as sequências de poli-amioácidos exógenos de escolha, a cultura dessas células em condições favoráveis à produção da proteína receptáculo contendo as sequências de poli-amioácidos de escolha e o isolamento da proteína receptáculo contendo os poli-amioácidos exógenos de escolha.
[0068] Métodos para isolar proteínas expressas são bastante conhecidas no estado da arte e, nesse sentido, as proteínas receptáculo pode ser isoladas facilmente por qualquer técnica. A presença da cauda de poli-histidina permite a purificação das proteínas recombinantes após sua expressão no sistema bacteriano (Hochuli et al Bio/Technology 6: 1321-25; Bornhorst and Falke, Methods Enzymology 326:245-54).
[0069] A presente invenção ainda contempla a escolha e seleção dos poli-amioácidos exógenos. As proteínas receptáculo podem abrigar diferentes sequências de poli-aminoácidos de diferentes origens, desde animais vertebrados, incluindo mamíferos, invertebrados, plantas, microrganismos ou vírus a fim de promover sua expressão, apresentação ou utilização em diferentes meios. Diferentes métodos de seleção de sequências de poliaminoácidos pode ser utilizados, tais como seleção específica por afinidade de ligação a anticorpos ou outras proteínas ligantes, por mapeamento de epítopos ou outras técnicas conhecidas no estado da arte.
[0070] As sequências de poli-aminoácidos são sequências de cinco a 30 aminoácidos que representam sensível ou especificamente um organismo, para qualquer fim descrito nesse pedido de patente. As sequências de poliaminoácidos podem representar, mas não limitados a esses exemplos: (i) epítopos lineares de células B; (ii) epítopos de células T; (iii) epítopos neutralizantes; (iv) regiões proteicas específicas de fontes patogênicas ou não patogênicas; (v) regiões proximais a sítios ativos de enzimas que não são no rmalmente alvos de uma resposta imunológica. Essas regiões de epítopos podem ser identificadas por uma grande variedade de métodos que incluem, mas não são limitados a: análise por síntese spot, bibliotecas de peptídeos randômicos, phage display, análises por software, uso de dados de cristalografia por raios X, bases de dados de epítopos ou outros métodos constantes no estado da arte.
[0071] A inserção de sequências de poli-aminoácidos exógenos nas proteínas receptáculo em sítios previamente identificados surpreendentemente não perturbou ou interferiu as características inerentes da proteína. Foram identificados oito sítios de introdução para sequências de poli-aminoácidos exógenas nas proteínas receptáculo (Kiss et al. Nucleic Acids Res 34: e132, 2006; Pavoor et al. Proc Natl Acad Sci U S A 106: 11895-900, 2009; Abedi et al, Nucleic Acids Res 26: 623-30, 1998; Zhong et al. Biomol Eng 21: 6772, 2004). Esses sítios de introdução podem abrigar uma, duas ou mais diferentes sequências de poli-aminoácidos exógenos, in tandem,no mesmo sítio de inserção, amplificando consideravelmente a expressão de diferentes sequências de poli-aminoácidos.
[0072] Assim, as proteínas receptáculo podem expressar um grande número de diferentes sequências de poli-aminoácidos, se caracterizando como proteínas receptáculo multivalentes, ampliando sua utilização para fins de composição vacinal, insumo para pesquisa e desenvolvimento tecnológico ou diagnóstico de doenças. Há uma real necessidade no estado da arte de se desenvolver proteínas receptáculo que não somente exibam intensidades adequadas de fluorescência, mas também possam ser expressas em grandes quantidades em sistemas celulares de produção e, ainda, tolerar a apresentação concomitante de múltiplas sequências de poli-aminoácidos exógenos sem deixar de apresentar a autofluorescência detectável.
[0073] A invenção ainda apresenta diversos usos para a proteína receptáculo. Esses usos incluem, mas não são limitados a, o uso do receptáculo proteico (i) como uma molécula repórter em ensaios celulares de triagem, incluindo ensaios intracelulares; (ii) como uma proteína para apresentação de bibliotecas randômicas ou selecionadas de peptídeos; (iii) como proteína apresentadora de antígenos como reagente para desenvolvimento de testes in vitro de diagnóstico imunológico, em geral, e para doenças infecciosas, parasitárias ou de outra origem imunológica; (iv) como proteína apresentadora de antígenos úteis para seleção, captura, triagem ou purificação de substâncias ligantes, tais como, anticorpos; (v) como proteína apresentadora de antígenos para composição vacinal; (vi) como proteína contendo sequências de anticorpos para ligação a antígenos; (vii) como proteína apresentadora de antígenos com atividade de imunização passiva.
[0074] As proteínas receptáculo podem ser úteis como composição vacinal ao, extraordinariamente, apresentarem: (a) um grande número, simultaneamente, de sequências de poli-aminoácidos indutores de respostas imunológicas, e (b) um cerne proteico não indutor de resposta imunológica.
[0075] A presente invenção é descrita detalhadamente através dos exemplos apresentados abaixo. É necessário frisar que a invenção não está limitada a esses exemplos, mas que também inclui variações e modificações dentro dos limites nos quais pode ser desenvolvida. Ainda vale informar que o acesso a todas as sequências biológicas do patrimônio genético brasileiro estão cadastradas no SISGEN, sob o número de cadastro AC53976.
[0076] As sequências de aminoácidos entre diferentes exemplos de proteína verde fluorescente, eGFP (GenBank: L29345.1; UniProtKB -P42212), Cycle-3 (GenBank: CAH64883.1), SuperFolder (GenBank: AOH95453.1), Split (Cabantous et al. Science Reports 3: 2854, 2013), Superfast (Fisher & DeLisa. PLoS One 3: e2351, 2008) foram utilizadas para a construção denovas proteínas para o presente pedido de invenção. Foram realizados alinhamentos e comparações das sequências pelo software Intaglio (Purgatory Design, V3.9.4). A partir desses dados, foram realizadas diversas alterações para que as proteínas receptáculo pudessem alcançar as características necessárias.
[0077] Alterações foram realizadas para criação de sítios de ação de enzimas de restrição. A inserção desses sítios foi projetada para que não houvesse alteração nas características físico-químicas da proteína GFP e, desta forma, não afetar as propriedades ou qualidades descritas nesse pedido de patente. Adicionalmente, a inserção desses sítios de restrição agrega outras propriedades às proteínas receptáculo ao permitir uma potencialidade de usos em métodos e processos de engenharia genética que permitiriam a manipulação genética dessas proteínas para incorporação de diversos peptídeos em diferentes regiões da proteína.
[0078] A sequência de nucleotídeos da proteína GFP foi manipulada para a introdução ou substituição de nucleotídeos a fim de se criarnovos sítios para enzimas de restrição. Desse modo, foram produzidas duasnovas proteínas receptáculo, a proteína “Platform” e a proteína “Rx”.
[0079] As alterações nas sequências nucleotídicas baseando-se na proteína eGFP, deram origem às seguintes alterações de aminoácidos nas proteínas receptáculo:
Proteína “Platform”
Posição 16, aminoácido I;
Posição 28, aminoácido F;
Posição 30, aminoácido R;
Posição 39, aminoácido I;
Posição 43, aminoácido S;
Posição 72, aminoácido S;
Posição 99, aminoácido Y;
Posição 105, aminoácido T;
Posição 111, aminoácido E;
Posição 124, aminoácido V;
Posição 128, aminoácido I;
Posição 145, aminoácido F;
Posição 153, aminoácido T;
Posição 163, aminoácido A;
Posição 166, aminoácido T;
Posição 167, aminoácido V;
Posição 171, aminoácido V;
Posição 205, aminoácido T;
Posição 206, aminoácido I;
Posição 208, aminoácido L.
Proteína “Platform”
Posição 16, aminoácido I;
Posição 28, aminoácido F;
Posição 30, aminoácido R;
Posição 39, aminoácido I;
Posição 43, aminoácido S;
Posição 72, aminoácido S;
Posição 99, aminoácido Y;
Posição 105, aminoácido T;
Posição 111, aminoácido E;
Posição 124, aminoácido V;
Posição 128, aminoácido I;
Posição 145, aminoácido F;
Posição 153, aminoácido T;
Posição 163, aminoácido A;
Posição 166, aminoácido T;
Posição 167, aminoácido V;
Posição 171, aminoácido V;
Posição 205, aminoácido T;
Posição 206, aminoácido I;
Posição 208, aminoácido L.
[0080] Proteína “Rx”
Posição 16, aminoácido V;
Posição 28, aminoácido S;
Posição 30, aminoácido R;
Posição 39, aminoácido I;
Posição 43, aminoácido T;
Posição 72, aminoácido A;
Posição 99, aminoácido S;
Posição 105, aminoácido K;
Posição 111, aminoácido V;
Posição 124, aminoácido V;
Posição 128, aminoácido T;
Posição 145, aminoácido F;
Posição 153, aminoácido T;
Posição 163, aminoácido A;
Posição 166, aminoácido T;
Posição 167, aminoácido V;
Posição 171, aminoácido V;
Posição 205, aminoácido T;
Posição 206, aminoácido V;
Posição 208, aminoácido S.
Posição 16, aminoácido V;
Posição 28, aminoácido S;
Posição 30, aminoácido R;
Posição 39, aminoácido I;
Posição 43, aminoácido T;
Posição 72, aminoácido A;
Posição 99, aminoácido S;
Posição 105, aminoácido K;
Posição 111, aminoácido V;
Posição 124, aminoácido V;
Posição 128, aminoácido T;
Posição 145, aminoácido F;
Posição 153, aminoácido T;
Posição 163, aminoácido A;
Posição 166, aminoácido T;
Posição 167, aminoácido V;
Posição 171, aminoácido V;
Posição 205, aminoácido T;
Posição 206, aminoácido V;
Posição 208, aminoácido S.
[0081] Substituições alternativas de aminoácidos ainda podem selecionadas, para todas as proteínas receptáculo, nas posições:
Posição 39, aminoácido N;
Posição 72, aminoácido S;
Posição 99, aminoácido S;
Posição 105, aminoácido Y ou K;
Posição 206, aminoácido I;
Posição 208, aminoácido L.
Posição 39, aminoácido N;
Posição 72, aminoácido S;
Posição 99, aminoácido S;
Posição 105, aminoácido Y ou K;
Posição 206, aminoácido I;
Posição 208, aminoácido L.
[0082] A presença de algumas mutações pode influenciar características bioquímicas da proteína. A mutação S30R influencia positivamente as características de enovelamento da proteína; as mutações Y145F e I171V previnem a tradução de intermediários indesejáveis; as mutações A206V ou I reduzem a possibilidade de agregação das proteínas nascentes.
[0083] Outras alterações foram realizadas nas proteínas receptáculo “Platform” e “Rx”, a partir da inclusão denovos códons de nucleotídeos para criação denovos sítios de restrição, que podem ser vistas na tabela 1 (abaixo):
Tabela 1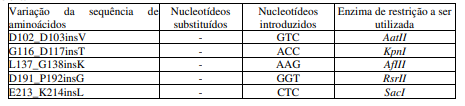
Tabela 1
[0084] Adicionalmente, a sequência de nucleotídeos das proteínas receptáculo “Platform” e “Rx” abriga mais dois outros sítios de restrição para NdeI and NheI,no terminal amino 5’ da proteína, a partir da inserção da sequência CATATGGTGGCTAGC (SEQ IDn°. 5) e outros dois sítios de restrição para EcoRI e XhoI,no terminal carboxi 3’, a partir da inserção da sequência GAATTCTAATGACTCGAG (SEQ IDn°6). Ainda foi incorporado às proteínas receptáculo dois códons de parada (stop codons) e uma cauda de polihistidina no amino-terminal.
[0085] A sequência de aminoácidos da proteína “Platform” pode ser observada na SEQ IDno.1 e sua sequência correspondente em nucleotídeos está descrita na SEQ IDno.2.
[0086] A sequência de aminoácidos da proteína “Rx” pode ser observada na SEQ IDno.3 e sua sequência correspondente em nucleotídeos está descrita na SEQ IDno.4.
[0087] Ao se criar sítios de restrição sem alterar a estrutura tridimensional da proteína original, permitiu-se o surgimento de 10novos sítios de inserção de sequências de poli-aminoácidos exógenas na proteína. Cadanovo sítio de inserção será aqui denominado de posição 1 a posição 10.
[0088] As localizações, nas sequências de nucleotídeos e de aminoácidos, das posições 1 a 10, das proteínas receptáculo “Platform” e “Rx” estão apontadas na Tabela 2 (abaixo):
Tabela 2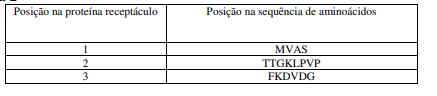

Tabela 2
[0089] A partir de alinhamentos das sequências de aminoácidos de diferentes proteínas GFP, foi concebida uma sequência dos aminoácidos de consenso, designada CGP (Dai et al. Protein Engineering, Design and Selection 20(2): 69-79 2007). Esta proteína fluorescente embora apresentasse alta estabilidade, foi melhorada por evolução dirigida para apresentar maior estabilidade em relação ao CGP (Kiss et al. Protein Engineering, Design & Selection 22(5): 313-23, 2009). Contudo, a proteína melhorada, devido à presença de três mutações, apresentava tendência a agregação. Outras mutações foram ainda incorporadas com base em uma análise de sua estrutura cristalina resultando na eliminação da agregação e produção da proteína denominada Proteína Térmica Verde (Thermal Green Protein; TGP) (Close et al. Proteins 83(7): 1225-37, 2015). Quando usada como receptáculo de proteína, a sequência é chamada de "Tx". A sequência de aminoácidos da proteína “Tx” pode ser observada na SEQ IDno. 77 e sua sequência correspondente em nucleotídeos está descrita na SEQ IDno. 78.
[0090] As localizações, nas sequências de nucleotídeos e de aminoácidos, das posições 1 a 12, das proteínas receptáculo “Tx” estão apontadas na Tabela 3 (abaixo):
Tabela 3

Tabela 3
[0091] A proteína receptáculo “Platform” foi geneticamente manipulada para abrigar epítopos de Trypanosoma cruzi, a qual aqui denominamos plataforma PlatCruzi. O gene correspondente à proteína PlatCruzi, aqui denominado gene PlatCruzi, está descrito na sequência de nucleotídeos SEQ IDno. 17.
[0092] Sequências de poli-aminoácidos originárias do T. cruzi foram selecionados a partir da literatura disponívelnoestado da arte, considerando dados experimentais de especificidade e sensibilidade para testes diagnósticos para a doença de Chagas (Peralta JM et al. J Clin Microbiol 32: 971-974, 1994; Houghton RL et al. J Infect Dis 179: 1226-1234, 1999; Thomas et al. Clin Exp Immunol 123: 465-471, 2001; Rabello et al, 1999; Gruber & Zingales, Exp Parasitology, 76(1): 1-12, 1993; Lafaille et al, Moleular Biochemistry Parasitology, 35(2): 127-36, 1989). Foram selecionados dez sequências de poli-aminoácidos para inserçãonos dez pontos de inserção na proteína “Platform”, aqui denominados TcEp1 a TcEp10, conforme a tabela 4 (abaixo).
[0093] Após a seleção das sequências de poli-aminoácidos de T. cruzi, o gene sintético correspondente a proteína PlatCruzi foi produzido por síntese química, pela metodologia de gene síntese por ligação e inserido em plasmídeos para experimentação. A sequência de aminoácidos correspondente ao gene PlatCruzi, contendo os epítopos TcEp1 a TcEp10, está descrito na SEQ IDn°. 18.
Tabela 4
Tabela 4
[0094] O gene sintético foi introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a PlatCruzi, uma cepa DH5a de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento.
[0095] O método de sequenciamento utlizado foi o enzimático, dideoxi ou de terminação em cadeia, que se baseia na síntese enzimática de uma fita complementar, cujo crescimento é interrompido pela adição de um dideoxinucleotídeo (Sanger et al., Proceeding National Academy of Science, 74(12): 5463-5467, 1977). Esta metodologia consistenos seguintes passos: Reação de sequenciamento (replicação do DNA em 25 ciclos no termociclador), precipitação do DNA por isopropanol/etanol, desnaturação da dupla fita (95°C por 2 min) e a leitura da sequência nucleotídica foi realizada no sequenciador automático ABI 3730XL (ThermoFischer SCIENTIFIC) (Otto et al,. Genetics and Molecular Research 7: 861-871, 2008). As análises das sequências obtidas foram feitas com o auxílio do programa 4Peaks (Nucleobytes; Mac OS X, 2004). Na reação foram utilzados os inciadores do vetor pET-28a (T7 Promoter e T7 Terminator).
[0096] O clone de plasmídeo analisado abrigando a sequência correta para a PlatCruzi foi transferido para E. coli, cepa BL21, a fim de se produzir a proteína PlatCruzi. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa foi cultivada durante anoite em meio LB e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h.
[0097] A cultura foi submetida à centrifugação e o pellet ressuspenso em tampão de uréia (100 mM NaH2PO4, 10 mM Tris-base, 8 M ureia, pH 8,0). A solução foi submetida à cromatografia por coluna de afinidade HisTrap™, 1 mL, (GE Healthcare Life Sciences) que permite a purificação de alta resolução de proteínas marcadas com histidina (histidine-tagged). O sobrenadante foi aplicado em coluna de afinidade a níquel (HisTrapTM, 1 mL, GE Healthcare Life Sciences) com o fluxo de 0,5 mL por minuto, previamente equlibrado em tampão A (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 5 mM imidazol). Após ligação, a resina foi lavada com 10 mL do tampão A. A proteína foi eluída em gradiente de 100% do tampão B (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 500 mM imidazol) com fluxo de 0,7 mL/min por 45 minutos. A purificação da PlatCruzi foi acompanhada a 280 nm (linha preta) e está apresentada na Figura 1A. A porcentagem de imidazol está marcado em vermelho. A proteína foi eluída em um volume, aproximadamente, entre 19 e 25 ml.
[0098] Alíquotas das proteínas recombinantes (1 μg/poço) foram submetidas a eletroforese em gel de poliacrilamida contendo SDS (SDS-PAGE) (Laemmli, Nature 227: 680-685, 1970). Os géis de concentração (stacking gel) e de separação (running gel) foram preparados a uma concentração de acrilamida de 4% e 11%, respectivamente (tabela 5, abaixo). As amostras foram preparadas em condições desnaturantes em tampão 62,5 mM Tris-HCl, pH 6,8, 2% SDS, 5% β-mercaptoetanol, 10% glicerol e fervidas a 95 °C durante 5 min (Hames BD, Gel electrophoresis of proteins: a practical approach. 3. ed. Oxford. 1998). Após a eletroforese, as proteínas foram detectadas por coloração com azul de comassie Blue R250 (Bio-Rad, EUA). O marcador Kaleidoscope™ Prestained Standards foi utilizado como referência de peso molecular (Bio-Rad, EUA). A Figura 1B, mostra a purificação da proteína PlatCruzi por cromatografia de afinidade usando uma coluna de níquel-agarose usando um sistema de cromatografia líquida Äktapurifier.
Tabela 5: Volume e concentração dos reagentes usados no preparo do gel de concentração de amostras a 4% e para o gel de separação a 11%.
Tabela 5: Volume e concentração dos reagentes usados no preparo do gel de concentração de amostras a 4% e para o gel de separação a 11%.
[0099] A performance da proteína PlatCruzi foi avaliada contra um painel de amostras biológicas de referência e de indivíduos acometidos por infecções por T. cruzi. A proteína PlatCruzi, em solução tampão carbonato/bicarbonato (50 mM pH 9,6), foi adicionada a uma placa de ELISA de 96 orifícios, nas quantidades de 0,1, 0,25, 0,5 e 1,0 μg/orifício a 4 °C por 12-18 h. Os orifícios foram lavados com solução de tampão salina-fosfato (PBS) adicionado de Tween 20 (PBS-T, fosfato de sódio 10 mM - Na3PO4, cloreto de sódio 150 mM - NaCl e Tween-20 0,05%, pH 7,4) e, em seguida, incubados com tampão PBS 1x contendo leite desnatado desidradato 5% (peso/volume) por 2 h a 37°C.
[00100] Em seguida, os orifícios foram lavados 3 vezes com tampão PBS-T e incubadas com as amostras biológicas de referência TCl (IS 09/188) ou TCII (IS 09/186) (Organização Mundial da Saúde) diluídas 2, 4, 8, 16, 32 e 64 vezes por 1 h a 370C. Após o período de incubação, os orifícios foram lavados três vezes com PBS-T e, em seguida, incubados com anticorpo IgG humano marcado com fosfatase alcalina, na diluição de 1:5000, por 1 h a 370C. Os orifícios foram lavados novamente três vezes com tampão PBS-T e o substrato para-nitrofenilfosfato (PNPP, 1mg/mL, ThermoFischer SCIENTIFIC) foi adicionado. Após 30 minutos e sob abrigo de luz, a absorbância foi medida em leitor de placa de ELISA a 405 nm.
[00101] Os resultados mostraram respostas satisfatórias em todas as diluições de amostras biológicas de referência TCI e TCII utilizadas independentemente da quantidade utilizada da PlatCruzi (Figura 2A e 2B). Os resultados fortemente suportam o uso da PlatCruzi para detecção de infecções por T. cruzi causadas por qualquer das seis DTUs (discrete typing units/unidades de tipagem distintas), abrangendo toda a área geográfica das cepas de T. cruzi circulantes. A mesma performance pode ser observada ao se utilizar soros de pacientes com doença de Chagas, com baixos e altos títulos de detecção de anticorpos (Figura 3).
[00102] Placas de Elisa contendo 500ng (em 0,3M Urea, pH 8,0) de PlatCruzi foram elaboradas conforme anteriormente descrito e, após três lavagens com PBS-T, foram incubadas com soros de quatro pacientes com altos índices de anticorpos anti-T.cruzi (6C-CE, 9C-CE, 15C-CE and 12-SE) e de quatro pacientes com baixos índices de anticorpos anti-T.cruzi (3C-PB, 6C-PB, 16C-PB and 17C-PB) em diferentes diluições: 1:50, 1:100, 1:250, 1:500, 1:1000 por 1 h a 37°C. Após esse período, os orifícios foram lavados três vezes com PBS-T e incubados com anticorpo IgG humano marcado com fosfatase alcalina, na diluição de 1:5000, por 1 h. Os orifícios foram lavados novamente três vezes com tampão PBS-T e o substrato para-nitrofenilfosfato (PNPP, 1mg/mL, ThermoFischer SCIENTIFIC) foi adicionado. Após 30 minutos, a absorbância foi medida em leitor de placa de ELISA a 405 nm.
[00103] Os resultados demonstraram que para os soros com baixos títulos de anticorpos, sinais de leitura nas diluições 1:50, 1:100 and 1:250 ficaram inequivocamente acima do limiar alcançados pelos controle negativo, evidenciando o potencial da plataforma PlatCruzi para detecção de anticorpos anti-T.cruzi tanto em soros de pacientes com altos quanto com baixos índices de anticorpos.
[00104] O uso de amostras de soros de pacientes para propósitos experimentais foi aprovado pelo Comitê de Ética da Fiocruz, conforme autorização CEP/IOC - CAAE: 52892216.8.0000.5248.
[00105] As placas de Elisa desenvolvidas no exemplo anterior contendo 500 ng (0,3 M Urea, pH 8,0) de PlatCruzi foram incubadas com 71 soros de pacientes diagnosticados para T. cruzi, e mais 18 soros de pacientes diagnosticados para leishmaniose (negativos para T. cruzi), 20 soros diagnosticados para dengue (negativos para T. cruzi) e 39 soros negativos (outras infecções e indivíduos sem presença de alguma infecção) , à diluição de 1:250, por 1 h a 37°C. Após esse período, as placas foram submetidas a processos de lavagem e marcação com anticorpo, revelação e leitura tal como previamente realizado.
[00106] Os resultados apontaram para uma excelente sensibilidade e especificidade utilizando a plataforma PlatCruzi (Figura 4) a partir da análise da correlação da curva de característica de operação do receptor (ROC). Não foram observados resultados falso negativos para os soros previamente identificados como positivos para T. cruzi; assim como não foi observado falsos positivos para os demais soros reconhecidamente negativos para T. cruzi, inclusive para aqueles positivos para outras doenças infecciosas. Ambos os índices de sensibilidade e especificidade foram de 100%.
[00107] A proteína “Rx” foi testada para verificação de sua performance e habilidade para expressar epítopos provenientes de outros microrganismos, incluindo vírus. A literatura aponta para um expressivo número de sequências de poli-aminoácidos específicos que podem ser utilizados como alvos para anticorpos neutralizantes. Entretanto, pequenas variações nas sequências observadas entre as cepas virais podem interferir na neutralização. Dessa forma, um estudo aprofundado sobre as melhores sequências de poli-aminoácidos a serem utilizados requer um grande conhecimento da biologia do vírus e da epidemiologia de sua interação com o seu hospedeiro.
[00108] Sequências de poli-aminoácidos do vírus da Raiva foram selecionados a partir da literatura disponível no estado da arte, considerando dados experimentais de especificidade e sensibilidade para o diagnóstico da doença causada pelo vírus da raiva (Kuzmina et al., J Antivir Antiretrovir 5:2: 37-43, 2013; Cai et al., Microbes Infect 12: 948-955, 2010).
[00109] Foram selecionados dez sequências de poli-aminoácidos para inserçãonos dez pontos de inserção na proteína Rx, aqui denominados RaEp1 a RaEp10, conforme abaixo. A combinação da sequência dessas sequências de poli-aminoácidos com a proteína Rx deu origem a proteína RxRabies2. O gene correspondente à proteína RxRabies2, aqui denominado gene RxRabies2, está descrito na sequência de nucleotídeos SEQ IDn°. 19. A sequência de aminoácidos correspondente ao gene RxRabies2 o qual contem as sequências de poli-aminoácidos RaEp1 a RaEp10, está descrito na SEQ IDno. 20.
Tabela 6
Tabela 6
[00110] Um gene sintético contendo uma sequência codificante para a proteína Rx e sequências codificantes para as sequências de poli-aminoácidos descritas na tabela 6 acima foi sintetizado.
[00111] O gene sintétido foi introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a RxRabies2, uma cepa DH5a de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, realizados da mesma forma descrita em PlatCruzi.
[00112] O plasmídeo analisado abrigando a sequência correta para a RxRabies2 foi transferido para E. coli, cepa BL21, a fim de se produzir a proteína RxRabies2. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h a 37°C.
[00113] A cultura foi submetida à centrifugação e o pellet ressuspenso em tampão de uréia (100 mM NaH2PO4, 10 mM Tris-base, 8 M ureia, pH 8,0). A solução foi submetida à cromatografia por coluna de afinidade HisTrap™, 1 mL, GE Healthcare Life Sciences, que permite a purificação de alta resolução de proteínas marcadas com histidina (histidine-tagged), com o fluxo de 0,5 mL por minuto, previamente equlibrado em tampão A (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 5 mM imidazol). Após ligação, a resina foi lavada com 10 mL do tampão A. A proteína foi eluída em gradiente de 100% do tampão B (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 500 mM imidazol) com fluxo de 0,7 mL/min por 45 minutos.
[00114] A produção de RxRabies2 foi analisada em três diferentes volumes de cultura; 3, 25 e 50 ml. Como visto na Tabela 7 (abaixo), a expressão de RxRabies2 foi de 123 μg / ml em média.
Tabela 7
Tabela 7
[00115] A proteina RxRabies2 foi produzida tal como descrito no exemplo anterior. 100 pg de proteína foi suspensa em adjuvante incompleto de Freud’s (0,5 mL) e inoculados, intramuscular, na quadriceps de dois coelhos machos denova Zelândia, com 6 meses de vida. Os coelhos foram novamente inoculados sete e quatorze dias depois da inoculação inicial, com a proteina RxRabies suspensa em PBS (100 μg/0,5 mL).
[00116] Após 21 dias da inoculação da primeira dose da composição vacinal, foi realizada a coleta de sangue dos animais. O plasma foi coletado por centrifugação e submetido a purificação por afinidade através da ligação a proteína Rx-Rabies2 adsorvida a superfície de uma membrana de nitrocelulose.
[00117] A membrana de nitrocelulose contendo a proteína Rx-Rabies2 foi preparada conforme a descrição abaixo, a fim de isolá-la de potenciais contaminantes. Após a eletroforese, as proteínas foram transferidas para uma membrana de nitrocelulose usando técnicas de Western blot conhecidas no estado da arte.
preparação do gel de SDS-PAGE a 11%, conforme descrito em exemplo anterior e na Tabela 5;
aplicação de 10 μg da proteína Rx-Rabies2 em gel de SDS-PAGE 11% (Tabela 5) e submissão a corrente eletroforética de 100 volts, por apoximandante 2 horas;
transferência das proteínas para membrana de nitrocelulose: as proteínas foram transferidas para membranas de nitrocelulose através do Trans-Blot Cell (Bio-Rad, EUA), com o tampão de transferência (Tris base 25 mM, glicina 192 mM e metanol 20%) por uma hora a 100V;
coloração com vermelho de Ponceau S (Ponceau S 0,1%, ácido acético 5%), para a confirmação da presença da proteína recombinante. A membrana foi cortada de forma a especificamente obter um pedaço somente com a proteína RxRabies2;
descoloração das membranas em água destilada e permanência em TBS (0,1%) entre 12 a 18 horas (overnight);
incubação com solução de bloqueio (25 mM Tris-HCl, 125 mM NaCl pH 7,4 (TBS) contendo 0,05% (v/v) de Tween 20 (TBS-T) e 5% (p/v) de leite desnatado desidratado) por toda a noite; em seguida, as membranas foram incubadas novamente em solução de bloqueio, por uma hora e, posteriormente, feitas três lavagens com TBS-T por 5 minutos cada e mais três lavagens de 5 minutos com TBS.
preparação do gel de SDS-PAGE a 11%, conforme descrito em exemplo anterior e na Tabela 5;
aplicação de 10 μg da proteína Rx-Rabies2 em gel de SDS-PAGE 11% (Tabela 5) e submissão a corrente eletroforética de 100 volts, por apoximandante 2 horas;
transferência das proteínas para membrana de nitrocelulose: as proteínas foram transferidas para membranas de nitrocelulose através do Trans-Blot Cell (Bio-Rad, EUA), com o tampão de transferência (Tris base 25 mM, glicina 192 mM e metanol 20%) por uma hora a 100V;
coloração com vermelho de Ponceau S (Ponceau S 0,1%, ácido acético 5%), para a confirmação da presença da proteína recombinante. A membrana foi cortada de forma a especificamente obter um pedaço somente com a proteína RxRabies2;
descoloração das membranas em água destilada e permanência em TBS (0,1%) entre 12 a 18 horas (overnight);
incubação com solução de bloqueio (25 mM Tris-HCl, 125 mM NaCl pH 7,4 (TBS) contendo 0,05% (v/v) de Tween 20 (TBS-T) e 5% (p/v) de leite desnatado desidratado) por toda a noite; em seguida, as membranas foram incubadas novamente em solução de bloqueio, por uma hora e, posteriormente, feitas três lavagens com TBS-T por 5 minutos cada e mais três lavagens de 5 minutos com TBS.
[00118] Os soros dos coelhos imunizados foram diluídos 1:500 em TBS e 10ml foram colocados em contato com os segmentos de membrana de nitrocelulose contendo a proteína RxRabies2 por 1 h, sob agitação e, posteriormente, lavados três vezes por 5 minutos com TBS-T e novamente lavados 3 vezes por 5 minutos com TBS. Em seguida, os anticorpos ligados especificamente foram liberados pela adição de 1 ml de 100 mM de glicina (pH 3,0). O pH da solução foi elevado para 7 pela adição de 100 μl de 1M Tris (pH 9,0) para purificação dos anticorpos de coelho. A purificação de anticorpos que se ligam especificamente aos antígenos é uma importante etapa para uso desses anticorpos para terapias, pois permite a drástica redução do quantitativo a ser administrado minimizando os potenciais efeitos adversos.
[00119] Diferentes extratos foram utilizados para demonstrar a habilidade específica dos anticorpos de coelho produzidos em se ligar a RxRabies2. Foram utilizados:
extrato bacteriano bruto com expressão da proteína Rx, obtido através das mesmas condições citadas em PlatCruzi;
proteína RxRabies2n°extrato bruto bacteriano e purificada nas concentrações 1x e 0,5x;
proteína Platcruzi purificada, nas concentrações 1x e 0,5x.
extrato bacteriano bruto com expressão da proteína Rx, obtido através das mesmas condições citadas em PlatCruzi;
proteína RxRabies2n°extrato bruto bacteriano e purificada nas concentrações 1x e 0,5x;
proteína Platcruzi purificada, nas concentrações 1x e 0,5x.
[00120] Os potenciais ligantes foram submetidos uma eletroforese em gel de poliacrilamida (SDS-PAGE a 11%, Tabela 5), conforme descrito anteriormente e, em seguida, tranferidos para membrana de nitrocelulose e submetidos a Western blot, cujo o detalhamento do método já fora descrito para RxRabies2.
[00121] As membranas foram incubadas por uma hora com o soro anti-RxRabies2 purificados como descrito acima, sob agitação. Em seguida, foram feitas novamente três lavagens de 5 minutos com TBS-T e três lavagens de 5 minutos com TBS. Posteriormente, as membranas foram incubadas, por uma hora, com os anticorpos secundários anti-coelho IgG com peroxidase diluído a 1:10.000. Após a incubação com o anticorpo secundário, foram feitas três lavagens com TBS-T por 5 minutos e três lavagens com TBS por 5 minutos. A revelação foi realizada com auxílio do SigmaFast™ DAB Peroxidase Substrate Tablet.
[00122] Os resultados apresentaram a ligação específica dos anticorpos de coelho produzidos pela inoculação de RxRabies2 aos ligantes que continham proteínas do vírus da raiva 2. A Figura 5 apresenta bandas de leitura somente nas faixas 2, 4 e 6 as quais contém o extrato bruto bacteriano contendo a proteína RxRabies2, e a proteína RxRabies2 purificada e diluída nas concentrações 1x e 0,5x, respectivamente.
[00123] A especificidade da resposta imunológica também pode ser observada pela inexistência de banda nas linhas 3, 5 e 7, contendo (i) a proteína receptáculo Rx sem a introdução de epítopos, (ii) a plataforma Platcruzi, diluída 1x e 0,5x, respectivamente. Esses resultados confirmam que a resposta imune foi restrita aos epítopos do vírus da raiva, indicando que a proteína Rx per si não se apresenta imunogênica.
[00124] A partir de estudos de mapeamento de sequências de poliaminoácidos de imunoglobulinas de cavalo (De-Simone et al., Toxicon 78: 83-93, 2014; Wagner et al., Journal of Immunology 173: 3230-3242, 2004), foi identificado uma sequência de poli-aminoácidos de IgG3 de cavalos reconhecidos por IgG e IgE humanas, hábil para ser utilizado em ensaios laboratoriais de diagnóstico da alergia ao soro de cavalo.
[00125] A sequência de poli-aminoácidos DVLFTWYVDGTEV (SEQ IDno. 30) foi incorporado a proteína Rx nas posições 1, 5, 6, 8, 9 e 10, dando origem a proteína RxHolgG3. A sequência de aminoácidos da proteína RxHolgG3 está descrita na SEQ IDno. 31. A sequência de nucleotídeos da proteína RxHolgG3 está descrita na SEQ IDno.32. Um gene sintético contendo uma sequência codificante para a proteína Rx e a sequência codificante para a sequência de poli-aminoácidos descrita acima (SEQ IDno. 30) foi sintetizado.
[00126] O gene sintético foi introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a proteína RxHoIgG3, uma cepa DH5α de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, como já especificado em PlatCruzi.
[00127] O plasmídeo analisado abrigando a sequência correta para a RxHoIgG3 foi transferido para E. coli, cepa BL21, a fim de se produzir a proteína RxHoIgG3. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h.
[00128] A cultura foi submetida à centrifugação e o pellet ressuspenso em tampão de uréia (100 mM NaH2PO4, 10 mM Tris-base, 8 M uréia, pH 8,0). A solução foi submetida à cromatografia por uma coluna de afinidade a níquel (HisTrap™, 1 mL, GE Healthcare Life Sciences) e com o fluxo de 0,5 mL por minuto, previamente equlibrado em tampão A (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 5 mM imidazol). Após ligação, a resina foi lavada com 10 mL do tampão A. A proteína foi eluída em gradiente de 100% do tampão B (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 500 mM imidazol) com fluxo de 0,7 mL/min por 45 minutos. A produção da proteína RxHolgG3 pode ser evidenciada no eluato após a eletroforese por SDS-PAGE a 11% (Tabela 5). Os resultados estão apresentados na Figura 6, e mostram a expressão de RxHoIgG3 como uma proteína recombinante. Nenhuma banda foi detectada no extracto solúvel bacteriano não induzido (Figura 6, coluna 1), e uma nas frações insolúveis (coluna 2) e solúveis (coluna 3).
[00129] A partir de estudo de mapeamento de sequências de poliaminoácidos do vírus oropouche (cepa Q71MJ4 e Q9J945, Uniprot) (Acrani et al., Journal of General Virology 96: 513-523, 2014; Tilston-Lunel et al., Journal of General Virology 96 (Pt 7): 1636-1650, 2015), foram selecionadas sequências de poli-aminoácidos a partir de técnicas spot síntese ou microarranjo de peptídeo disponível no estado da arte, considerando seu potencial diagnóstico. Foram selecionados 6 sequências de poli-aminoácidos para inserção em nove pontos de inserção na proteína Rx, aqui denominados OrEp 1 a OrEp 7, conforme tabela 8 abaixo:
Tabela 8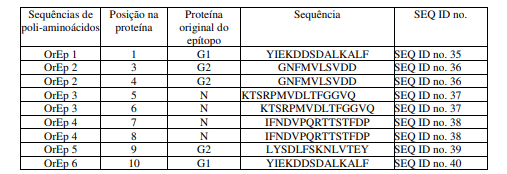
Tabela 8
[00130] A combinação da sequência dessas sequências de poliaminoácidos descritas acima com a proteína Rx deu origem a proteína RxOro. A sequência de aminoácidos correspondente ao gene RxOro, contendo as sequências de poli-aminoácidos OrEp1 a OrEp6, está descrita na SEQ IDno.33. O gene correspondente à proteína RxOro, aqui denominado gene RxOro, está descrito na sequência de nucleotídeos SEQ IDno. 34.
[00131] Um gene sintético RxOro foi sintetizado e introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a RxOro, uma cepa DH5α de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, conforme descrito em PlatCruzi.
[00132] O plasmídeo analisado abrigando a sequência correta para a proteína RxOro foi transferido para E. coli, cepa BL21. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h.
[00133] A cultura foi submetida à centrifugação e o pellet ressuspenso em tampão de uréia (100 mM NaH2PO4, 10 mM Tris-base, 8 M uréia, pH 8,0). A solução foi submetida à cromatografia por uma coluna de afinidade a níquel (HisTrap™, 1 mL, GE Healthcare Life Sciences) e com o fluxo de 0,5 mL por minuto, previamente equlibrado em tampão A (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 5 mM imidazol). Após ligação, a resina foi lavada com 10 mL do tampão A. A proteína foi eluída em gradiente de 100% do tampão B (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 500 mM imidazol) em um fluxo de 0,7 mL/min por 45 minutos. A produção de RxOro foi examinada em três diferentes volumes de crescimentto bacteriano; 3, 25 e 50 ml. O nível de expressão de RxOro foi de 203 μg / ml em média e os resultaddos estão apresentados na Tabela 9.
Tabela 9
Tabela 9
[00134] A performance da proteína RxOro foi avaliada contra um painel de soros de indivíduos acometidos por infecções por Oropouche. A proteína RxOro, em solução (0,3 M Urea, pH 8,0), foi adicionada a uma placa de ELISA de 96 orifícios, na quantidade de 500 ng/orifício a 4 °C por 12-18 h. Os orifícios foram lavados com solução de tampão salina-fosfato (PBS) adicionado de Tween 20 (PBS-T, fosfato de sódio 10 mM - Na3PO4, cloreto de sódio 150 mM - NaCl e Tween-20 0,05%, pH 7,4) e, em seguida, incubados com tampão PBS 1x contendo leite desnatado desidradato 5% (peso/volume) por 2 h a 37°C.
[00135] Em seguida, os orifícios foram lavados 3 vezes com tampão PBS-T e incubadas com 98 amostras de soros de pacientes suspeitos de infecção pelo vírus Oropouche e 51 amostras de soros de pacientes saudáveis diluídas 1:100, por 1 h a 37°C. Após o período de incubação, os orifícios foram lavados três vezes com PBS-T e, em seguida, incubados com anticorpo IgG humano marcado com fosfatase alcalina, na diluição de 1:5000, por 1 h a 370C. Os orifícios foram lavados novamente três vezes com tampão PBS-T e o substrato para-nitrofenilfosfato (PNPP, 1mg/mL, ThermoFischer SCIENTIFIC) foi adicionado. Após 30 minutos e sob abrigo de luz, a absorbância foi medida em leitor de placa de ELISA a 405 nm.
[00136] Os resultados apontaram para uma excelente sensibilidade e especificidade utilizando a RxOro (Figura 7). Os resultados fortemente suportam o uso da RxOro para detecção de infecções pelo vírus Oropouche.
[00137] O uso de amostras de soros de pacientes para propósitos experimentais foi aprovado pelo Comitê de Ética da Fiocruz, conforme autorização CEP/IOC - CAAE: 52892216.8.0000.5248.
[00138] A partir de estudo de mapeamento de sequências de poliaminoácidos do vírus Mayaro (cepa Q8QZ73 e Q8QZ72, Uniprot) (Espósito et al., Genome Announcement 3: e01372-15, 2015), foram selecionadas sequências de poli-aminoácidos a partir da literatura disponível no estado da arte, considerando seu potencial diagnóstico. Foram selecionadas 4 sequências de poli-aminoácidos para inserção em nove pontos de inserção na proteína Rx, aqui denominados MGEp1 a MGEp 4, conforme Tabela 10 abaixo:
Tabela 10
Tabela 10
[00139] A combinação da sequência dessas sequências de poliaminoácidos descritas acima com a proteína Rx deu origem a proteína RxMayaro_IgG. A sequência de aminoácidos correspondente ao gene RxMayaro_IgG, contendo as sequências de poli-aminoácidos MGEp 1 a MGEp 4, está descrito na SEQ IDno. 45. O gene correspondente à proteína RxMayaro_IgG, aqui denominado gene RxMayaro_IgG, está descrito na sequência de nucleotídeos SEQ IDno. 46.
[00140] Um gene sintético RxMayaro_IgG foi sintetizado e introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a RxMayaro_IgG, uma cepa DH5α de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, conforme citado em PlatCruzi.
[00141] O plasmídeo analisado abrigando a sequência correta para a proteína RxMayaro_IgG foi transferido para E. coli, cepa BL21. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h.
[00142] A cultura foi submetida à centrifugação e o pellet ressuspenso em tampão de uréia (100 mM NaH2PO4, 10 mM Tris-base, 8 M uréia, pH 8,0). A solução foi submetida à cromatografia por uma coluna de afinidade a níquel (HisTrap™, 1 mL, GE Healthcare Life Sciences) e com o fluxo de 0,5 mL por minuto, previamente equlibrado em tampão A (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 5 mM imidazol). Após ligação, a resina foi lavada com 10 mL do tampão A. A proteína foi eluída em gradiente de 100% do tampão B (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 500 mM imidazol) com fluxo de 0,7 mL/min por 45 minutos. A produção da proteína RxMayaro_IgG pode ser evidenciada no eluído a partir da realização de eletroforese SDS-PAGE a 11% (Tabela 5). A proteína RxMayaro_IgG também foi examinada por SDS-PAGE para confirmar sua produção e determinar sua distribuição entre as frações solúvel e insolúvel. Como visto na Figura 8, as colunas 4 e 7 (setas) mostram que RxMayaro_IgG é produzido como solúvel e insolúvel.
[00143] A produção de RxMayaro_IgG foi examinada em três diferentes volumes de crescimento: 3, 25 e 50 ml. Como visto na Tabela 11 (abaixo), o nivel de expressão de RxMayaro_IgG foi de 130 μg / ml em média.
Tabela 11
Tabela 11
[00144] A performance da proteína RxMayaro_IgG foi avaliada contra um painel de soros de indivíduos acometidos por infecções por vírus Mayaro. A proteína RxMayaro_IgG, em solução (0,3 M Urea, pH 8,0), foi adicionada a uma placa de ELISA de 96 orifícios, na quantidade de 500 ng/orifício a 4 °C por 12-18 h. Os orifícios foram lavados com solução de tampão salina-fosfato (PBS) adicionado de Tween 20 (PBS-T, fosfato de sódio 10 mM - Na3PO4, cloreto de sódio 150 mM - NaCl e Tween-20 0,05%, pH 7,4) e, em seguida, incubados com tampão PBS 1x contendo leite desnatado desidradato 5% (peso/volume) por 2 h a 37°C.
[00145] Em seguida, os orifícios foram lavados 3 vezes com tampão PBS-T e incubadas com 6 amostras de soros de pacientes suspeitos de infecção pelo vírus Mayaro e 29 amostras de soros de pacientes saudáveis diluídas 1:100, por 1 h a 37°C. Após o período de incubação, os orifícios foram lavados três vezes com PBS-T e, em seguida, incubados com anticorpo IgG humano marcado com fosfatase alcalina, na diluição de 1:5000, por 1 h a 37°C. Os orifícios foram lavados novamente três vezes com tampão PBS-T e o substrato para-nitrofenilfosfato (PNPP, 1mg/mL, ThermoFischer SCIENTIFIC) foi adicionado. Após 30 minutos e sob abrigo de luz, a absorbância foi medida em leitor de placa de ELISA a 405 nm.
[00146] Os resultados apontaram para uma excelente sensibilidade e especificidade utilizando a RxMayaro_IgG (Figura 9). Os resultados fortemente suportam o uso da RxMayaro_IgG para detecção de infecções pelo vírus Mayaro.
[00147] O uso de amostras de soros de pacientes para propósitos experimentais foi aprovado pelo Comitê de Ética da Fiocruz, conforme autorização CEP/IOC - CAAE: 52892216.8.0000.5248.
[00148] A partir de estudo de mapeamento de sequências de poliaminoácidos do vírus Mayaro (cepa Q8QZ73 e Q8QZ72, Uniprot) (Espósito et al., Genome Announcement 3: e01372-15, 2015), foram selecionadas sequências de poli-aminoácidos a partir da literatura disponível no estado da arte, considerando seu potencial diagnóstico. Foram selecionadas 4 sequências de poli-aminoácidos para inserção em nove pontos de inserção na proteína Rx, aqui denominados MMEpl a MMEp 4, conforme tabela 12 (abaixo):
Tabela 12
Tabela 12
[00149] A combinação da sequência dessas sequências de poliaminoácidos descritas acima com a proteína Rx deu origem a proteína RxMayaro_IgM. A sequência de aminoácidos correspondente ao gene RxMayaro_IgM, contendo as sequências de poli-aminoácidos MMEp 1 a MMEp 4, está descrito na SEQ IDno. 51. O gene correspondente à proteína RxMayaro_IgM, aqui denominado gene RxMayaro_IgM, está descrito na sequência de nucleotídeos SEQ IDno. 52.
[00150] Um gene sintético RxMayaro_IgM foi sintetizado e introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a RxMayaro_IgM, uma cepa DH5a de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, como previamente descrito em PlatCruzi.
[00151] O plasmídeo analisado abrigando a sequência correta para a proteína RxMayaro_IgM foi transferido para E. coli, cepa BL21. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200 rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h.
[00152] A cultura foi submetida à centrifugação e o pellet ressuspenso em tampão de uréia (100 mM NaH2PO4, 10 mM Tris-base, 8 M uréia, pH 8,0). A solução foi submetida à cromatografia por uma coluna de afinidade a níquel (HisTrap™, 1 mL, GE Healthcare Life Sciences) e com o fluxo de 0,5 mL por minuto, previamente equlibrado em tampão A (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 5 mM imidazol). Após ligação, a resina foi lavada com 10 mL do tampão A. A proteína foi eluída em gradiente de 100% do tampão B (50 mM Tris-HCl, pH 8,0, 100 mM NaCl e 500 mM imidazol) com fluxo de 0,7 mL/min por 45 minutos.
[00153] A produção da proteína RxMayaro_IgM pode ser evidenciada no eluído a partir da realização de eletroforese SDS-PAGE e sua distribuição ser observada entre as frações solúvel e insolúvel. Como pode ser visto na Figura 8, as colunas 5 e 8 (setas) mostram que RxMayaro_IgM é produzido como solúvel e insolúvel.
[00154] A produção de RxMayaro_IgM também foi examinada em três diferentes volumes de crescimento: 3, 25 e 50 ml. Como visto na Tabela 13 (abaixo), o nível de expressão de RxMayaro_IgM foi de 205 μg / ml em média.
Tabela 13
Tabela 13
[00155] A performance da proteína RxMayaro_IgM foi avaliada contra um painel de soros de indivíduos acometidos por infecções por vírus Mayaro. A proteína RxMayaro_IgM, em solução (0,3 M Urea, pH 8,0), foi adicionada a uma placa de ELISA de 96 orifícios, na quantidade de 500 ng/oriffcio a 4 °C por 12-18 h. Os orifícios foram lavados com solução de tampão salina-fosfato (PBS) adicionado de Tween 20 (PBS-T, fosfato de sódio 10 mM - Na3PO4, cloreto de sódio 150 mM - NaCl e Tween-20 0,05%, pH 7,4) e, em seguida, incubados com tampão PBS 1x contendo leite desnatado desidradato 5% (peso/volume) por 2 h a 37°C.
[00156] Em seguida, os orifícios foram lavados 3 vezes com tampão PBS-T e incubadas com 6 amostras de soros de pacientes suspeitos de infecção pelo vírus Mayaro e 29 amostras de soros de pacientes saudáveis diluídas 1:100, por 1 h a 37°C. Após o período de incubação, os orifícios foram lavados três vezes com PBS-T e, em seguida, incubados com anticorpo IgM humano marcado com fosfatase alcalina, na diluição de 1:5000, por 1 h a 37°C. Os orifícios foram lavados novamente três vezes com tampão PBS-T e o substrato para-nitrofenilfosfato (PNPP, 1mg/mL, ThermoFischer SCIENTIFIC) foi adicionado. Após 30 minutos e sob abrigo de luz, a absorbância foi medida em leitor de placa de ELISA a 405 nm.
[00157] Os resultados apontaram para uma excelente sensibilidade e especificidade utilizando a RxMayaro_IgM (Figura 10). Os resultados fortemente suportam o uso da RxMayaro_IgM para detecção de infecções pelo vírus Mayaro.
[00158] O uso de amostras de soros de pacientes para propósitos experimentais foi aprovado pelo Comitê de Ética da Fiocruz, conforme autorização CEP/IOC - CAAE: 52892216.8.0000.5248.
[00159] A partir de estudo de mapeamento de sequências de poliaminoácidos da toxina proteica da bactéria Bordetella pertussis (P04977; P04978; P04979; P0A3R5 e P04981: Uniprot) foram selecionadas sequências de poli-aminoácidos a partir da literatura disponível no estado da arte, considerando seu potencial diagnóstico. Foram selecionadas 10 sequências de poli-aminoácidos para inserção em nove pontos de inserção na proteína Rx, aqui denominados PtxEp1 a PtxEp 10, conforme Tabela 14 abaixo. Neste exemplo, dois epítopos foram localizados na posição 1 usando um espaçador (SEQ IDno. 95: SYWKGS) entre eles. Dois epítopos também foram inseridos na posição 10 usando outro espaçador (SEQ IDno. 96: EAAKEAAK). O intuito da introdução destes espaçadores é gerar um espaço físico inerte entre os poli-aminoáciodos consecutivos, ajudando assim a impedir a competição de ligação entre anticorpos adjacentes.
Tabela 14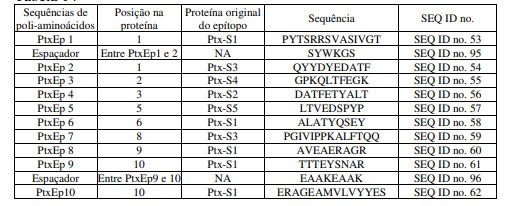
Tabela 14
[00160] A combinação da sequência dessas sequências de poliaminoácidos descritas acima com a proteína Rx deu origem a proteína RxPtx. A sequência de aminoácidos correspondente ao gene RxPtx, contendo os epítopos PtxEp 1 a PtxEp 10, está descrito na SEQ IDno.64. O gene correspondente à proteína RxPtx, aqui denominado gene RxPtx, está descrito na sequência de nucleotídeos SEQ IDno. 63.
[00161] Um gene sintético RxPtx foi sintetizado e introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a RxPtx, uma cepa DH5α de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, como previamente descrito em PlatCruzi.
[00162] O plasmídeo analisado abrigando a sequência correta para a proteína RxPtx foi transferido para E. coli, cepa BL21. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200 rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h.
[00163] A cultura foi submetida à centrifugação e o pellet ressuspenso em 2 mL de PBS com CelLytic (0,5x) durante uma hora a 4 °C. Após outra centrifugação, o sobrenadante foi recolhido e o sedimento foi ressuspenso em solução de ureia 8 M (pH 8,0)no mesmo volume que o sobrenadante. Volumes iguais foram carregados no gel de SDS-PAGE (11%, Tabela 5).
[00164] A proteína RxPtx foi examinada por eletroforese de SDS-PAGE para confirmar sua produção e determinar sua distribuição entre as frações solúvel e insolúvel. Como visto na Figura 11, as colunas 5 e 6 (setas) mostram que RxPtx é produzido como solúvel e insolúvel com maior proporção na fração insolúvel.
[00165] A partir de estudo de mapeamento de sequências de poliaminoácidos do vírus da febre amarela (cepa 17DD e as sequências no arquivo p03314-Uniprot) foram selecionadas sequências de poli-aminoácidos a partir da literatura disponível no estado da arte, considerando seu potencial diagnóstico. Foram selecionadas 10 sequências de poli-aminoácidos para inserção em nove pontos de inserção na proteína Rx, aqui denominados YFIgGEp1 a YFIgGEp10, conforme tabela 15 abaixo. Dois epítopos foram localizados na posição 10 usando um espaçador (SEQ IDno. 97: TSYWKGS) entre eles. O espaçador tem a função de criar um espaço físico entre os epítopos consecutivos, auxiliando a preservação da interação com os anticorpos.
Tabela 15
Tabela 15
[00166] A combinação da sequência desses poli-aminoácidos descritos acima com a proteína Rx deu origem a proteína RxYFIgG. A sequência de aminoácidos correspondente ao gene RxYFIgG, contendo os epítopos YFIgGEp 1 a YFIgGEp 10, está descrito na SEQ IDno. 75. O gene correspondente à proteína RxYFIgG, aqui denominado gene RxYFIgG, está descrito na sequência de nucleotídeos SEQ IDno. 76.
[00167] Um gene sintético RxYFIgG foi sintetizado e introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a RxYFIgG, uma cepa DH5α de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, como previamente descrito em PlatCruzi.
[00168] O plasmídeo analisado abrigando a sequência correta para a RxYFIgG foi transferido para E. coli, cepa BL21, a fim de se produzir a proteína RxYFIgG. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-l-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB a 37 °C e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200 rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h a 37 °C.
[00169] A cultura foi submetida à centrifugação e o pellet ressuspenso em 2 mL de PBS com CelLytic (0,5x) durante uma hora a 4 °C. Após outra centrifugação, o sobrenadante foi recolhido e o sedimento foi ressuspenso em solução de ureia 8 M (pH 8,0) no mesmo volume que o sobrenadante. Volumes iguais foram carregados no gel de SDS-PAGE (11%, Tabela 5). A proteína RxYFIgG foi examinada por eletroforese por SDS-PAGE para confirmar sua produção e determinar sua distribuição entre as frações solúvel e insolúvel. Como visto na Figura 11, as colunas 9 e 10 (setas) mostram que RxYFIgG é produzido como solúvel e insolúvel, com maior proporção na fração insolúvel.
[00170] A proteína receptáculo “Tx” foi geneticamente manipulada para abrigar epítopos de células T da Dermatophogoides pteronyssinus, a qual aqui denominamos plataforma TxNeuza. O gene correspondente à proteína TxNeuza, aqui denominado gene TxNeuza, está descrito na sequência de nucleotídeos SEQ IDn°. 89.
[00171] A partir de estudo de mapeamento de sequências de poliaminoácidos de células T da D. pteronyssinus, foram selecionadas sequências de poli-aminoácidos a partir da literatura disponível no estado da arte, considerando seu potencial diagnóstico das alergias causada pelo D. pteronyssinus (Hinz et al., Clin Exp Allergy 45: 1601-1612, 2015; Oseroff et al., Clin Exp Allergy 47:577-592, 2017). Foram selecionados nove sequências de poli-aminoácidos para inserção em nove pontos de inserção na proteína Tx, aqui denominados NeuzaEpl a NeuzaEp9, conforme tabela 16 abaixo. Neste exemplo, dois epítopos foram localizados na posição 12 usando um espaçador (SEQ IDno. 98: GGSG) entre eles.
Tabela 16
Tabela 16
[00172] A combinação da sequência desses epítopos descritos acima com a proteína Tx deu origem a proteína TxNeuza. A sequência de aminoácidos correspondente ao gene TxNeuza, contendo os epítopos NeuzaEp 1 a NeuzaEp9, está descrito na SEQ IDno. 88.
[00173] Um gene sintético TxNeuza foi sintetizado e introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Ndel e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a TxNeuza, uma cepa DH5a de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, como previamente descrito em PlatCruzi.
[00174] O plasmídeo analisado abrigando a sequência correta para a TxNeuza foi transferido para E. coli, cepa BL21, a fim de se produzir a proteína TxNeuza. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB a 37 °C e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200 rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h a 37 °C.
[00175] A cultura foi submetida à centrifugação e o pellet ressuspenso em 2 mL de PBS com CelLytic (0,5x) durante uma hora a 4 °C. Após outra centrifugação, o sobrenadante foi recolhido e o sedimento foi ressuspenso em solução de ureia 8 M (pH 8,0)no mesmo volume que o sobrenadante. Volumes iguais foram carregados no gel de SDS-PAGE (11%, Tabela 5). A proteína TxNeuza foi examinada por SDS-PAGE para confirmar sua produção e determinar sua distribuição entre as frações solúvel e insolúvel. Como visto na Figura 11, as colunas 7 e 8 (setas) mostram que a proteína TxNeuza é produzida como insolúvel.
[00176] Sequências de poli-aminoácidos de T. cruzi foram selecionados a partir da literatura disponível no estado da arte, considerando dados experimentais de especificidade e sensibilidade para testes diagnósticos para a doença de Chagas (Balouz, et al., Clin Vaccine Immunol 22, 304-312, 2015; Alvarez, et al., Infect Immun 69, 7946-7949, 2001; Fernandez-Villegas, et al., J Antimicrob Chemother 71, 2005-2009, 2016; Thomas, et al., Clin Vaccine Immunol 19, 167-173, 2012). Foram selecionados dez sequências de poli-aminoácidos para inserçãonos dez pontos de inserção na proteína “Tx”, aqui denominados TcEp 1, TcEp 3, TcEp 4, TcEp 6, TcEp 8, TcEp 9, TcEp 10, TcEp 11, TcEp 12 e TcEp 13, conforme a tabela 17 abaixo. Dois epítopos foram localizados na posição 12 usando um espaçador (SEQ IDn°. 99: GGASG) entre eles.
Tabela 17
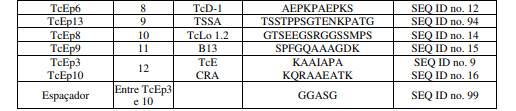
Tabela 17
[00177] Após a seleção das sequências de poli-aminoácidos de T. cruzi, o gene sintético correspondente a proteína TxCruzi foi produzido por síntese química, pela metodologia de gene síntese por ligação e inserido em plasmídeos para experimentação. A sequência de nucleotídeos correspondente ao gene TxCruzi, contendo os epítopos TcEp 1, TcEp 3, TcEp 4, TcEp 6, TcEp 8, TcEp 9, TcEp 10, TcEp 11, TcEp 12 e TcEp 13, está descrito na SEQ IDn°. 91.
[00178] A combinação da sequência desses epítopos descritos acima com a proteína Tx deu origem a proteína TxCruzi. A sequência de aminoácidos correspondente ao gene TxCruzi, contendo os epítopos TcEp 1, TcEp 3, TcEp 4, TcEp 6, TcEp 8, TcEp 9, TcEp 10, TcEp 11, TcEp 12 e TcEp 13, está descrito na SEQ IDno. 90.
[00179] O gene sintétido foi introduzido em plasmídeos pET28a, utilizando os sítios de restrição para as enzimas Xbal e Xhol através de métodos de biologia molecular conhecidos no estado da arte. A fim de se identificar se o gene sintético correspondia a sequência desenhada para a proteína TxCruzi, uma cepa DH5α de Escherichia coli foi transformada e o material plasmidial analisado por técnicas de digestão por enzimas de restrição e, posteriormente, sequenciamento, como já especificado em PlatCruzi.
[00180] O plasmídeo analisado abrigando a sequência correta para a TxCruzi foi transferido para E. coli, cepa BL21, a fim de se produzir a proteína TxCruzi. A cepa BL21 pode expressar RNA polimerase T7 quando induzida por Isopropil β-D-1-thiogalactopiranoside (IPTG). A cepa de BL21 foi cultivada durante anoite em meio LB a 37 °C e, posteriormente ressemeada no mesmo meio, adicionado de canamicina (30 μg/ml), em agitador a 200 rpm, até atingir a densidade óptica de turbidez de 0,6-0,8 (600 nm). Em seguida, IPTG (q.s.p. 1 mM) é adicionado à cultura e as mesmas condições de cultivo são mantidas por mais 3 h a 37 °C.
[00181] A cultura foi submetida à centrifugação e o pellet ressuspenso em 2 mL de PBS com CelLytic (0,5x) durante uma hora a 4 °C. Após outra centrifugação, o sobrenadante foi recolhido e o sedimento foi ressuspenso em solução de ureia 8 M (pH 8,0)no mesmo volume que o sobrenadante. Volumes iguais foram carregados no gel de SDS-PAGE (11%, Tabela 5).
[00182] A proteína TxCruzi foi examinada por SDS-PAGE para confirmar sua produção e determinar sua distribuição entre as frações solúvel e insolúvel. Como visto na Figura 11, as colunas 3 e 4 (setas) mostram que TxCruzi é produzido como solúvel e insolúvel. Em comparação com PlatCruzi (colunas 1 e 2), TxCruzi apresentou uma melhora na proporção de proteína solúvel produzida, o que pode ser atribuído ao uso de Tx como proteína receptáculo.
Claims (48)
- Receptáculo proteico, caracterizado pelo fato de que compreende uma estrutura proteica estável que suporte, em diferentes sítios, mais do que sequências de poli-aminoácidos exógenas simultaneamente.
- Receptáculo proteico de acordo com a reivindicação 1, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ ID n. 1.
- Receptáculo proteico de acordo com a reivindicação 2, caracterizado pelo fato de que compreende a sequência de nucleotídeos apresentada na SEQ ID n. 2.
- Receptáculo proteico de acordo com a reivindicação 1, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ ID n. 3.
- Receptáculo proteico de acordo com a reivindicação 4, caracterizado pelo fato de que compreende a sequência de nucleotídeos apresentada na SEQ ID n. 4.
- Receptáculo proteico de acordo com a reivindicação 1, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ ID n. 77.
- Receptáculo proteico de acordo com a reivindicação 4, caracterizado pelo fato de que compreende a sequência de nucleotídeos apresentada na SEQ ID n. 78.
- Receptáculo proteico de acordo com a reivindicação 1, caracterizado apresentar um cerne proteico estável a inserção de quatro ou mais sequências de poli-aminoácidos exógenos simultaneamente.
- Receptáculo proteico de acordo com a reivindicação 1, caracterizado pelo fato de que apresenta sítios de inserção das sequências de poli-aminoácidos exógenas em alças proteicas voltadas para o meio externo.
- Receptáculo proteico de acordo com a reivindicação 1, caracterizado pelo fato de que a inserção das sequências de poli-aminoácidos exógenas simultaneamente não interfere nas condições de produção da proteína receptáculo.
- Receptáculo proteico de acordo com a reivindicação 2, 4 ou 6, caracterizado pelo fato de que contém sequências de poli-aminoácidos exógenas, simultaneamente, para uso em composições vacinais, em diagnósticos ou no desenvolvimento de reagentes laboratoriais.
- Receptáculo proteico de acordo com a reivindicação 2, 4 ou 6, caracterizado pelo fato de que as sequências de poli-aminoácidos exógenas não perderam suas características imunogênicas ao serem inseridas simultaneamente nas alças proteicas da proteína receptáculo.
- Receptáculo proteico de acordo com a reivindicação 2 ou 3, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ IDn°. 18.
- Receptáculo proteico de acordo com a reivindicação 2 ou 3, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ IDn°. 17.
- Receptáculo proteico de acordo com a reivindicação 13 ou 14, caracterizado pelo fato de que abriga as sequências de poli-aminoácidos exógenas, simultaneamente, SEQ IDn°5, SEQ IDn°6, SEQ IDn°7, SEQ IDn° 8, SEQ IDn°9, SEQ IDn°10, SEQ IDn°11, SEQ IDn°12, SEQ IDn° 13, SEQ IDn°14, SEQ IDn°15 e SEQ IDn°16.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ IDn°. 20.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ IDn°. 19.
- Receptáculo proteico de acordo com a reivindicação 16 ou 17, caracterizado pelo fato de que abriga as sequências de poli-aminoácidos exógenas, simultaneamente, SEQ IDn°21, SEQ IDn°22, SEQ IDn°23, SEQ IDn°24, SEQ IDn°25, SEQ IDn°26, SEQ IDn°27, SEQ IDn°28 e SEQ IDn°29.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ IDn°. 31.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ IDn°. 32.
- Receptáculo proteico de acordo com a reivindicação 19 ou 20, caracterizado pelo fato de que abriga diversas cópias da sequência de poliaminoácido exógena, simultaneamente, SEQ IDn°30.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ IDn°. 33.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ IDn°. 34.
- Receptáculo proteico de acordo com a reivindicação 22 ou 23, caracterizado pelo fato de que abriga as sequências de poli-aminoácido exógenas, simultaneamente, SEQ IDn°35, SEQ IDn°36, SEQ IDn°37, SEQ IDn°38, SEQ IDn°39 e SEQ IDn°40.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ IDn°. 45.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ IDn°. 46.
- Receptáculo proteico de acordo com a reivindicação 25 ou 26, caracterizado pelo fato de que abriga as sequências de poli-aminoácido exógenas, simultaneamente, SEQ IDn°41, SEQ IDn°42, SEQ IDn°43 e SEQ IDn°44.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ IDn°. 51.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ IDn°. 52.
- Receptáculo proteico de acordo com a reivindicação 28 ou 29, caracterizado pelo fato de que abriga as sequências de poli-aminoácido exógenas, simultaneamente, SEQ IDn°47, SEQ IDn°48, SEQ IDn°49 e SEQ IDn°50.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ IDn°. 64.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ IDn°. 63.
- Receptáculo proteico de acordo com a reivindicação 31 ou 32, caracterizado pelo fato de que abriga as sequências de poli-aminoácido exógenas, simultaneamente, SEQ IDn°53, SEQ IDn°54, SEQ IDn°55, SEQ IDn°56, SEQ IDn°57, SEQ IDn°58, SEQ IDn°59, SEQ IDn°60, SEQ IDn°61, SEQ IDn°62, SEQ IDn°95 e SEQ IDn°96.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ IDn°. 75.
- Receptáculo proteico de acordo com a reivindicação 4 ou 5, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ ID n°. 76.
- Receptáculo proteico de acordo com a reivindicação 34 ou 35, caracterizado pelo fato de que abriga as sequências de poli-aminoácido exógenas, simultaneamente, SEQ ID n° 65, SEQ ID n° 66, SEQ ID n° 67, SEQ ID n° 68, SEQ ID n° 69, SEQ ID n° 70, SEQ ID n° 71, SEQ ID n° 72, SEQ ID n° 73, SEQ IDn° 74 e SEQ ID n° 97.
- Receptáculo proteico de acordo com a reivindicação 6 ou 7, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ ID n°. 88.
- Receptáculo proteico de acordo com a reivindicação 6 ou 7, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ ID n°. 89.
- Receptáculo proteico de acordo com a reivindicação 37 ou 38, caracterizado pelo fato de que abriga as sequências de poli-aminoácido exógenas, simultaneamente, SEQ ID n° 79, SEQ ID n° 80, SEQ ID n° 81, SEQ ID n° 82, SEQ ID n° 83, SEQ ID n° 84, SEQ IDn° 85, SEQ ID n° 86, SEQ ID n° 87 e SEQ ID n°98.
- Receptáculo proteico de acordo com a reivindicação 6 ou 7, caracterizado pelo fato de que compreende a sequência de aminoácidos apresentada na SEQ ID n°. 90.
- Receptáculo proteico de acordo com a reivindicação 6 ou 7, caracterizado pelo fato de ser expressa através da sequência de nucleotídeos apresentada na SEQ IDn°. 91.
- Receptáculo proteico de acordo com a reivindicação 40 ou 41, caracterizado pelo fato de que abriga as sequências de poli-aminoácido exógenas, simultaneamente, SEQ ID n° 7, SEQ ID n° 9, SEQ ID n° 10, SEQ ID n° 12, SEQ ID n° 14, SEQ ID n° 15, SEQ ID n° 16, SEQ ID n° 92, SEQ ID n° 93, SEQ ID n° 94 e SEQ ID n° 99.
- Método para produção do receptáculo proteico, caracterizado pelo fato de que não sofre interferência pela inserção de diversas sequências de poli-aminoácidos exógenas.
- Método para produção do receptáculo proteico de acordo com a reivindicação 43, caracterizado pelo fato de que introduz em células competentes de interesse as sequências de DNA codificadora para a proteína receptáculo adicionada do DNA codificante para as sequências de poliaminoácidos exógenos de escolha; realiza a cultura das células competentes da etapa anterior; e realiza o isolamento da proteína receptáculo contendo os poli-aminoácidos exógenos de escolha.
- Método de identificação de patógenos ou de diagnóstico de doenças, caracterizado pelo fato de que utiliza a proteína receptáculo como definida na reivindicação 1.
- Método de identificação de patógenos ou de diagnóstico de doenças de acordo a reivindicação 45, caracterizado pelo fato de que promove o diagnóstico da doença de Chagas, da raiva, da coqueluche, da febre amarela, das infecções pelos vírus Oropouche, Mayaro, da hipersensibilidade a IgE, da alergia por D. pteronyssinus.
- Uso do receptáculo proteico como definido em qualquer uma das reivindicações 1 a 42, caracterizado pelo fato de ser como um reagente laboratorial.
- Uso do receptáculo proteico como definido em qualquer uma das reivindicações 1 a 42, caracterizado pelo fato de ser em uma composição vacinal.
Priority Applications (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| BR102019017792-6A BR102019017792A2 (pt) | 2019-08-27 | 2019-08-27 | Receptáculo proteico, método para produção do receptáculo, método de identificação de patógenos ou de diagnóstico de doenças, e, uso do receptáculo |
| US17/638,108 US20230365622A1 (en) | 2019-08-27 | 2020-08-27 | Protein receptacle, polynucleotide, vector, expression cassette, cell, method for producing the receptacle, method of identifying pathogens or diagnosing diseases, use of the receptacle and diagnostic kit |
| EP20859528.0A EP4039696A4 (en) | 2019-08-27 | 2020-08-27 | PROTEIN CONTAINER, POLYNUCLEOTIDE, VECTOR, EXPRESSION CASSETTE, CELL, METHOD FOR PRODUCING THE CONTAINER, METHOD FOR IDENTIFYING PATHOGENS OR DIAGNOSING DISEASES, USE OF THE CONTAINER AND DIAGNOSTIC KIT |
| CN202510495376.2A CN120441713A (zh) | 2019-08-27 | 2020-08-27 | 蛋白质容器、多核苷酸、载体、表达盒、细胞、容器的生产方法、病原体识别或疾病诊断方法、容器的用途、和诊断试剂盒 |
| CN202080058664.7A CN114258399B (zh) | 2019-08-27 | 2020-08-27 | 蛋白质容器、多核苷酸、载体、表达盒、细胞、容器的生产方法、病原体识别或疾病诊断方法、容器的用途、和诊断试剂盒 |
| PCT/BR2020/050341 WO2021035325A1 (pt) | 2019-08-27 | 2020-08-27 | Receptáculo proteico, polinucleotideo, vetor, cassete de expressão, célula, método para produção do receptáculo, método de identificação de patógenos ou de diagnóstico de doenças, uso do receptáculo, e, kit diagnóstico |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| BR102019017792-6A BR102019017792A2 (pt) | 2019-08-27 | 2019-08-27 | Receptáculo proteico, método para produção do receptáculo, método de identificação de patógenos ou de diagnóstico de doenças, e, uso do receptáculo |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| BR102019017792A2 true BR102019017792A2 (pt) | 2021-11-16 |
Family
ID=74683213
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| BR102019017792-6A BR102019017792A2 (pt) | 2019-08-27 | 2019-08-27 | Receptáculo proteico, método para produção do receptáculo, método de identificação de patógenos ou de diagnóstico de doenças, e, uso do receptáculo |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US20230365622A1 (pt) |
| EP (1) | EP4039696A4 (pt) |
| CN (2) | CN114258399B (pt) |
| BR (1) | BR102019017792A2 (pt) |
| WO (1) | WO2021035325A1 (pt) |
Families Citing this family (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20180128545A1 (en) | 2016-11-08 | 2018-05-10 | Berry Metal Company | Modular furnace cooling wall |
| US12384811B2 (en) | 2019-04-05 | 2025-08-12 | Academia Sinica | Sialidase-resistant saccharide and method of making and using the same |
| WO2021198999A1 (en) * | 2020-04-03 | 2021-10-07 | Axon Neuroscience Se | Epitope-based vaccines for treatment of coronavirus associated diseases |
| AU2021258419A1 (en) * | 2020-04-20 | 2022-11-17 | NEC Laboratories Europe GmbH | SARS-CoV-2 vaccines |
| WO2021226520A1 (en) * | 2020-05-08 | 2021-11-11 | Kiromic BioPharma, Inc. | Peptide compositions for the treatment of pathogenic infections |
| WO2022025264A1 (ja) * | 2020-07-31 | 2022-02-03 | 東亞合成株式会社 | SARS-CoV-2由来のアミノ酸配列およびその利用 |
| US20240192211A1 (en) * | 2021-02-24 | 2024-06-13 | Biotome Pty Ltd | Peptides and their use in diagnosis of sars-cov-2 infection |
| BR112023000666A2 (pt) | 2021-04-12 | 2023-10-24 | Academia Sinica | Imunógeno, composição imunogênica, método para induzir uma resposta imune contra um coronavírus 2 da síndrome respiratória aguda grave e uso da composição imunogênica |
| EP4337955A4 (en) * | 2021-05-13 | 2025-07-23 | La Jolla Inst For Immunology | CORONAVIRUS T-CELL EPITOPES, MEGAPOOLS, AND RELATED USES |
| WO2022270624A1 (ja) * | 2021-06-24 | 2022-12-29 | 良広 渡部 | 抗体誘導性ポリペプチド及びワクチン |
| JP2023004912A (ja) * | 2021-06-24 | 2023-01-17 | 国立大学法人金沢大学 | 抗体誘導性ポリペプチド及びワクチン |
| CN114832099B (zh) * | 2022-04-08 | 2023-11-28 | 国科宁波生命与健康产业研究院 | 一种用于治疗SARS-CoV-2变异毒株感染的多肽制剂 |
| US12318485B2 (en) | 2023-04-08 | 2025-06-03 | Rock Biomedical, Inc. | Methods and compositions for targeted delivery by polymersomes |
| AU2024252371B2 (en) | 2023-04-08 | 2026-02-12 | Rock Biomedical Inc. | Methods and compositions for dendritic cell targeting nano-delivery |
| WO2024238734A2 (en) | 2023-05-17 | 2024-11-21 | Rock Biomedical Inc. | Mgat1 deficient cells and uses thereof |
Family Cites Families (32)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5625048A (en) | 1994-11-10 | 1997-04-29 | The Regents Of The University Of California | Modified green fluorescent proteins |
| GB9504446D0 (en) | 1995-03-06 | 1995-04-26 | Medical Res Council | Improvements in or relating to gene expression |
| US5874304A (en) | 1996-01-18 | 1999-02-23 | University Of Florida Research Foundation, Inc. | Humanized green fluorescent protein genes and methods |
| US5804387A (en) | 1996-02-01 | 1998-09-08 | The Board Of Trustees Of The Leland Stanford Junior University | FACS-optimized mutants of the green fluorescent protein (GFP) |
| US6096865A (en) | 1996-05-06 | 2000-08-01 | Amgen Inc. | Mutants of the green fluorescent protein having improved fluorescent properties at 37° |
| US6027881A (en) | 1996-05-08 | 2000-02-22 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Mutant Aequorea victoria fluorescent proteins having increased cellular fluorescence |
| US6124128A (en) | 1996-08-16 | 2000-09-26 | The Regents Of The University Of California | Long wavelength engineered fluorescent proteins |
| US6025485A (en) | 1997-02-14 | 2000-02-15 | Arcaris, Inc. | Methods and compositions for peptide libraries displayed on light-emitting scaffolds |
| US7090976B2 (en) | 1999-11-10 | 2006-08-15 | Rigel Pharmaceuticals, Inc. | Methods and compositions comprising Renilla GFP |
| US6180343B1 (en) | 1998-10-08 | 2001-01-30 | Rigel Pharmaceuticals, Inc. | Green fluorescent protein fusions with random peptides |
| US6936421B2 (en) * | 1998-10-08 | 2005-08-30 | Rigel Pharmaceuticals, Inc. | Structurally biased random peptide libraries based on different scaffolds |
| US7297482B2 (en) | 1998-10-08 | 2007-11-20 | Rigel Pharmaceuticals, Inc. | Structurally biased random peptide libraries based on different scaffolds |
| DE60032927T2 (de) | 1999-11-10 | 2007-11-08 | Rigel Pharmaceuticals, Inc., South San Francisco | Verfahren and zusammenstellungen, die renilla gfp enthalten |
| AU2003274897A1 (en) | 2002-07-10 | 2004-01-23 | Stratagene California | Humanized renilla reniformis green fluorescent protein as a scaffold |
| GB0423974D0 (en) * | 2004-10-28 | 2004-12-01 | Ares Trading Sa | Proteins |
| EP2072527A1 (en) * | 2007-12-21 | 2009-06-24 | Altonabiotec AG | Fusion polypeptides comprising a SHBG dimerization component and uses thereof |
| WO2009088950A2 (en) * | 2007-12-31 | 2009-07-16 | Boehringer Ingelheim Vetmedica, Inc. | Pcv2 orf2 virus like particle with foreign amino acid insertion |
| US20100137158A1 (en) * | 2008-10-31 | 2010-06-03 | Wisconsin Alumni Research Foundation | GFABS: GFP-based biosensors possessing the binding properties of antibodies |
| US20120077960A1 (en) * | 2009-03-13 | 2012-03-29 | Los Alamos National Security, Llc | Fluorobodies: intrinsically fluorescent binding ligands |
| EP2630502B1 (en) * | 2010-10-19 | 2017-08-02 | Georgia State University Research Foundation, Inc. | Analyte sensors, using such sensors, and methods of detecting analyte activity |
| JP5856821B2 (ja) * | 2010-11-26 | 2016-02-10 | 旭化成メディカル株式会社 | 腹水濾過濃縮装置 |
| US9714274B2 (en) * | 2012-03-23 | 2017-07-25 | Suzhou Kunpeng Biotech Co., Ltd. | Fusion proteins of superfolder green fluorescent protein and use thereof |
| WO2014144583A2 (en) * | 2013-03-15 | 2014-09-18 | Northwestern University | Methods for cell- free protein synthesis |
| US10155025B2 (en) * | 2014-07-17 | 2018-12-18 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Method for inhibiting or reducing transplant rejection by adminstration of interleukin-34 |
| WO2018057441A1 (en) * | 2016-09-20 | 2018-03-29 | Boehringer Ingelheim Vetmedica Gmbh | Canine adenovirus vectors |
| CN106591343B (zh) * | 2016-11-29 | 2020-02-18 | 湖北大学 | 一种超折叠绿色荧光蛋白介导异源蛋白在大肠杆菌中分泌表达方法 |
| CN106986922B (zh) | 2017-04-14 | 2020-03-06 | 江南大学 | 一种自组装双亲短肽及其应用 |
| CN107703219B (zh) | 2017-07-28 | 2019-09-27 | 浙江大学 | 基于CILLC-MS的评价GFP基因转染对hPMSCs代谢组学影响的方法 |
| CN108220313A (zh) | 2018-01-12 | 2018-06-29 | 辽宁科技大学 | 一种绿色荧光蛋白的融合表达方法 |
| CN108192904B (zh) | 2018-01-29 | 2019-10-08 | 怀化学院 | 一种穿膜荧光蛋白及其编码基因与应用 |
| CN108303539B (zh) | 2018-01-31 | 2020-08-11 | 刘双萍 | 一种乳腺癌细胞检测生物试剂及应用 |
| CN109652433A (zh) * | 2018-12-19 | 2019-04-19 | 南京理工大学 | 可视化的离子通道蛋白bR可溶性融合表达载体及其构建方法 |
-
2019
- 2019-08-27 BR BR102019017792-6A patent/BR102019017792A2/pt not_active Application Discontinuation
-
2020
- 2020-08-27 CN CN202080058664.7A patent/CN114258399B/zh active Active
- 2020-08-27 EP EP20859528.0A patent/EP4039696A4/en active Pending
- 2020-08-27 US US17/638,108 patent/US20230365622A1/en active Pending
- 2020-08-27 WO PCT/BR2020/050341 patent/WO2021035325A1/pt not_active Ceased
- 2020-08-27 CN CN202510495376.2A patent/CN120441713A/zh active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| CN120441713A (zh) | 2025-08-08 |
| EP4039696A1 (en) | 2022-08-10 |
| EP4039696A4 (en) | 2023-12-06 |
| US20230365622A1 (en) | 2023-11-16 |
| CN114258399A (zh) | 2022-03-29 |
| WO2021035325A1 (pt) | 2021-03-04 |
| CN114258399B (zh) | 2025-06-24 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| BR102019017792A2 (pt) | Receptáculo proteico, método para produção do receptáculo, método de identificação de patógenos ou de diagnóstico de doenças, e, uso do receptáculo | |
| CN113416259B (zh) | 一种β冠状病毒抗原、其制备方法和应用 | |
| Dalvie et al. | Engineered SARS-CoV-2 receptor binding domain improves manufacturability in yeast and immunogenicity in mice | |
| Ogun et al. | The oligomerization domain of C4-binding protein (C4bp) acts as an adjuvant, and the fusion protein comprised of the 19-kilodalton merozoite surface protein 1 fused with the murine C4bp domain protects mice against malaria | |
| Derivery et al. | The Wave complex is intrinsically inactive | |
| CN115246874B (zh) | 一种重组新型冠状病毒s-rbd三聚体蛋白、其制备方法和应用 | |
| US20130129726A1 (en) | Peptide having cell membrane penetrating activity | |
| US10253072B2 (en) | Prophylactic vaccine against egg drop syndrome (EDS) | |
| CN113943373A (zh) | 一种β冠状病毒多聚体抗原、其制备方法和应用 | |
| ES2340154T3 (es) | Proteinas fluorescentes de aequorea coerulescens y uso de las mismas. | |
| WO2021165538A1 (en) | Improved anti-senescence compounds and uses thereof | |
| CN113637695A (zh) | 靶向刺激体液免疫和细胞免疫的新冠病毒mRNA疫苗 | |
| Yan et al. | Preparation of a new monoclonal antibody against subgroup A of avian leukosis virus and identifying its antigenic epitope | |
| CN114989266A (zh) | 一种非洲猪瘟病毒pA104R蛋白免疫抑制相关氨基酸位点及其应用 | |
| RU2458129C1 (ru) | ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ БЕЛОК KillerRed (ВАРИАНТЫ), ВЫДЕЛЕННЫЙ БЕЛОК (ВАРИАНТЫ), КАССЕТА ЭКСПРЕССИИ, КЛЕТКА, СОДЕРЖАЩАЯ КАССЕТУ ЭКСПРЕССИИ | |
| CN105968211B (zh) | 一种重组抗病毒蛋白及其制备方法和应用 | |
| Kalyoncu et al. | Development of a Recombinant Omicron BA. 1 Subunit Vaccine Candidate in Pichia pastoris | |
| RU2491342C1 (ru) | ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ОПЕРАТИВНО СЛИТЫЙ ИНТРАМОЛЕКУЛЯРНЫЙ ДИМЕР БЕЛКА KillerRed, КАССЕТА ЭКСПРЕССИИ, КЛЕТКА, ПРОДУЦИРУЮЩАЯ ХИМЕРНЫЙ БЕЛОК И СОДЕРЖАЩАЯ КАССЕТУ ЭКСПРЕССИИ, ВЫДЕЛЕННЫЙ ХИМЕРНЫЙ БЕЛОК | |
| Islam | Structure and functional analysis of the African Swine Fever virus encoded CD2v/EP402R protein and investigation of its role in virus persistence in pigs | |
| Gillespie | Structure-guided SCHEMA recombination of VRC01-class antibodies for reduced polyreactivity | |
| Qin et al. | Development of monoclonal antibodies for GoAstV-2 VP27 protein and precise mapping of linear antigenic epitopes | |
| Ren et al. | Improved method to raise polyclonal antibody using enhanced green fluorescent protein transgenic mice | |
| Wyatt | Intein mediated engineering of cancer-selective toxic proteins | |
| HK40101648A (en) | Improved anti-senescence compounds and uses for the treatment of cancer | |
| CN121779571A (zh) | 牛结节性皮肤病病毒融合抗原及其产品和应用 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| B03A | Publication of a patent application or of a certificate of addition of invention [chapter 3.1 patent gazette] | ||
| B10A | Cessation: cessation confirmed |