ES2741806T3 - Un método para la diferenciación directa de células madre pluripotentes en músculo cardíaco funcional - Google Patents
Un método para la diferenciación directa de células madre pluripotentes en músculo cardíaco funcional Download PDFInfo
- Publication number
- ES2741806T3 ES2741806T3 ES14767019T ES14767019T ES2741806T3 ES 2741806 T3 ES2741806 T3 ES 2741806T3 ES 14767019 T ES14767019 T ES 14767019T ES 14767019 T ES14767019 T ES 14767019T ES 2741806 T3 ES2741806 T3 ES 2741806T3
- Authority
- ES
- Spain
- Prior art keywords
- stem cells
- basal medium
- cells
- bhm
- pluripotent stem
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0652—Cells of skeletal and connective tissues; Mesenchyme
- C12N5/0657—Cardiomyocytes; Heart cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/05—Inorganic components
- C12N2500/10—Metals; Metal chelators
- C12N2500/20—Transition metals
- C12N2500/24—Iron; Fe chelators; Transferrin
- C12N2500/25—Insulin-transferrin; Insulin-transferrin-selenium
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/90—Serum-free medium, which may still contain naturally-sourced components
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/115—Basic fibroblast growth factor (bFGF, FGF-2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/155—Bone morphogenic proteins [BMP]; Osteogenins; Osteogenic factor; Bone inducing factor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/16—Activin; Inhibin; Mullerian inhibiting substance
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/30—Hormones
- C12N2501/38—Hormones with nuclear receptors
- C12N2501/39—Steroid hormones
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/40—Regulators of development
- C12N2501/415—Wnt; Frizzeled
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/70—Enzymes
- C12N2501/72—Transferases [EC 2.]
- C12N2501/727—Kinases (EC 2.7.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2506/00—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells
- C12N2506/02—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells from embryonic cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2506/00—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells
- C12N2506/45—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells from artificially induced pluripotent stem cells
Landscapes
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Life Sciences & Earth Sciences (AREA)
- Wood Science & Technology (AREA)
- Organic Chemistry (AREA)
- Chemical & Material Sciences (AREA)
- Biotechnology (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- Microbiology (AREA)
- Cell Biology (AREA)
- Rheumatology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Cardiology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Materials For Medical Uses (AREA)
Abstract
Un método para producir músculo cardíaco de bioingeniería a partir de células madre pluripotentes, que comprende las etapas de (i) cultivar células madre pluripotentes en un medio basal que comprende una cantidad efectiva de (a) BMP4, Activina A, FGF2, un inhibidor de GSK3, y (b) un suplemento libre de suero que tiene como resultado una concentración final de 0,5 - 50 mg/ml de albúmina, 1 - 100 μg/ml de transferrina, 0,1 - 10 μg/ml de etanolamina, 0,003 - 0,3 μg/ml de selenito sódico, 0,4 - 40 μg/ml de L-carnitina HCl, 0,1 - 10 μg/ml de hidrocortisona, 0,05 - 5 μl/ml de suplemento de ácido graso, 0,0001 - 0,1 μg/ml de triyodo-L-tironina (T3), induciendo así la diferenciación mesodérmica de dichas células madre pluripotentes; (ii) cultivar las células obtenidas en la etapa (i) en un medio basal que comprende una cantidad eficaz de un inhibidor de la ruta de señalización Wnt y un suplemento libre de suero como se define en (i), induciendo así la diferenciación cardíaca de las células; y (iii) cultivar las células obtenidas en la etapa (ii) en un medio basal que comprende una cantidad eficaz de un suplemento libre de suero como en (i), bajo estimulación mecánica, promoviendo así la maduración cardíaca.
Description
DESCRIPCIÓN
Un método para la diferenciación directa de células madre pluripotentes en músculo cardíaco funcional
Fundamento de la invención
Las células madre pluripotentes humanas (hPSCs) se usan ahora mucho para proporcionar un suministro teóricamente sin fin y también grande de cardiomiocitos humanos (Kehat et al. J Clin Invest 108, 407 - 414 (2001); Takahashi et al. Cell 131, 861 - 872 (2007); Zhang et al., Circ Res. 104, e30 - 41 (2009)). Los cardiomiocitos humanos se han derivado de células madre embrionarias humanas (hESCs) (Thomson et al. Science 282, 1145 - 1147 (1998)) y de células madre pluripotentes inducidas (hIPSC) (Takahashi et al., Cell 131, 861 - 872 (2007)) y tienen un uso demostrado para múltiples fines, incluyendo los modelos de desarrollo (Lian et al. Stem Cells 2012 (2012)), la eficacia de los medicamentos y/o los cribados de seguridad (Schaaf et al. PLoS ONE 6, 20 (2011)), modelación de hipertrofia y aplicaciones regenerativas. Además, con los recientes avances en la tecnología de las hIPSC, se pueden generar in vitro los cardiomiocitos que exhiben fenotipos hereditarios de enfermedades genéticas (Carvajal-Vergara, X. et al. Nature 465, 808 - 812 (2010); Itzhaki et al., Nature 471, 225 - 229 (2011); Malan y otros, Circ Res (2011); Moretti y otros, N Engl J Med 363, 1397- 1409 (2010); Yazawa y otros Nature 471,230-234 (2010)).
Ahora se acepta universalmente que el cultivo 2D de baja densidad de cardiomiocitos humanos derivados de una biopsia conduce a cambios rápidos en el fenotipo y la morfología de los cardiomiocitos (Bird et al. Cardiovasc Res. 58, 423 - 434 (2003)) y hace que sea difícil extrapolar los resultados a la situación in vivo. Para obtener un fenotipo de cardiomiocito más representativo de las condiciones in vivo, se ha utilizado la ingeniería de tejidos cardíacos (Eschenhagen et al. Fa SEB J 11, 683 - 694 (1997); Zimmermann et al. Biotechnol Bioeng 68, 106 - 114 (2000), Zimmermann et al. Circ Res 90, 223 - 230 (2002); Tulloch et al. Circ Res 109, 47 - 59 (2011); Tiburcy et al. Circ Res 109, 1105- 1114 (2011); Eschenhagen et al. Am J Physiol Heart Circ Physiol 303, 11 (2012)) para generar constructos con propiedades similares a las del tejido cardíaco nativo.
La ideología actual de la ingeniería de tejidos es generar/aislar los tipos de células requeridos y sembrarlos en un entorno diseñado para promover su diferenciación y generar tejidos similares a los del tipo in vivo. Por lo tanto, la ingeniería tisular puede considerarse como un proceso ineficaz por dos razones: 1) la desasociación de un tejido / cultivo de diferenciación destruye el entorno extracelular destruyendo así la información del desarrollo (por ejemplo, interconectividad célula-célula, posicionamiento geométrico de las células, conectividad célula-ECM), esto precisa aumentos muy grandes en la producción de la matriz extracelular (ECM) con el fin de reconstruir el entorno (Hudson et al. Tissue Eng Part A 17, 2279 - 2289 (2011)), y 2) el proceso de desasociación es variable entre las líneas de hPSC y puede conducir a una muerte celular considerable.
Otros protocolos reportados en la literatura pueden requerir la modificación del protocolo para permitir eficiencias de cardiomiocitos similares en múltiples líneas de hPSC. Sin embargo, los resultados del autor de la presente invención demuestran que los cambios en el protocolo de diferenciación pueden afectar en gran medida al fenotipo cardiomiocitario (por ejemplo, se demuestra que la dorsomorfina puede afectar en gran medida al músculo cardíaco de bioingeniería (BHM: Bioengineered Heart Muscle)). Esto puede conducir a cambios en las propiedades miocárdicas en el tejido construido que pueden enmascarar los efectos de diferentes condiciones experimentales o modelos genéticos de enfermedades, por lo tanto, se debe tener cuidado al usar diferentes protocolos en diferentes líneas.
Algunos protocolos publicados recientemente pueden permitir que se use el mismo protocolo para múltiples líneas, también producen cardiomiocitos con una pureza muy alta. Sin embargo, los cardiomiocitos puros no facilitan la formación de miocardio construido con ingeniería tisular funcional y se requieren tanto cardiomiocitos como células del estroma para la formación de miocardio con tejido funcional (Naito et al. Circulation 114, 172- 78 (2006), Hudson et al. Tissue Eng Part A 17, 2279 - 2289 (2011)).
Los documentos WO 2013/056072 A1, WO 2013/111875 A1 y US 2013/177535 describen métodos para producir cardiomiocitos pero no de un músculo cardíaco construido de bioingeniería.
Por lo tanto, en la técnica existe la necesidad de métodos para producir miocardio humano de bioingeniería, que sean capaces de superar los anteriores inconvenientes.
El desarrollo de un protocolo de diferenciación robusto es un paso muy importante que permite la producción consistente de BHM. En este estudio se produjeron n > 140 BHM en > 18 experimentos independientes y cada uno exhibió actividad de latido espontáneo. Adicionalmente, el protocolo permite producir BHM a partir de múltiples líneas de hPSC utilizando el mismo protocolo. Además, podían eliminarse todas las etapas de desasociación y las hPSCs se diferenciaron directamente en miocardio de bioingeniería, conservando así la memoria de desarrollo del tejido, previniendo cualquier respuesta de recreación de tejido y proporcionando un modelo in vitro más preciso del desarrollo del miocardio humano.
Sumario de la invención
La presente invención se refiere a un método para producir músculo cardíaco de bioingeniería a partir de células madre pluripotentes, que comprende las etapas de
(i) cultivar células madre pluripotentes en un medio basal que comprende una cantidad efectiva de (a) BMP4, Activina A, FGF2, un inhibidor de GSK3, y (b) un suplemento libre de suero que tiene como resultado una concentración final de 0,5 - 50 mg /ml de albúmina, 1 - 100 pg/ml de transferrina, 0,1 - 10 pg/ml de etanol amina, 0,003 - 0,3 pg/ml de selenito de sodio, 0,4 - 40 pg/ml de L-carnitina HCl, 0,1 - 10 pg/ml de hidrocortisona, 0,05 - 5 pl/ml de suplemento de ácido graso, 0,0001 - 0,1 pg/ml de triyodo-L-tironina (T3), induciendo así la diferenciación mesodérmica de dichas células madre pluripotentes;
(ii) cultivar las células obtenidas en la etapa (i) en un medio basal que comprende una cantidad eficaz de un inhibidor de la ruta de señalización Wnt y un suplemento libre de suero como se define en la etapa (i), induciendo así la diferenciación cardíaca de las células; y
(iii) cultivar las células obtenidas en la etapa (ii) en un medio basal que comprende una cantidad eficaz de un suplemento libre de suero como se define en la etapa (i), bajo estimulación mecánica, promoviendo así la maduración cardíaca.
Llevando a cabo el método descrito en el presente texto el músculo cardiaco de bioingeniería (BHM) derivado de células madre pluripotentes humanas (hPSC) es generado por la formación directa del tejido de hPSCs en hidrogeles de colágeno. Para formar BHM, se imitó el desarrollo in vivo utilizando un protocolo de inducción libre de suero dirigida que hace que el tejido progrese a través de distintas etapas de desarrollo conocidas, a través de la pluripotencia, el mesodermo temprano, el progenitor cardiaco, los cardiomiocitos inmaduros y, finalmente, el tejido cardiaco más maduro, compuesto de 50% de cardiomiocitos, siendo el resto predominantemente una fracción de células estromales. Los autores de la presente invención optimizaron su protocolo de BHM libre de suero y encontraron que las propiedades individuales del BHM son muy dependientes de estímulos particulares, indicando así que se requieren múltiples estímulos exógenos para propiedades de BHM óptimas. Al final, se produjo BHM rítmicamente contráctil con una fuerza contráctil medible, capacidad de estimulación e inotropía en respuesta al aumento de la longitud de reposo, la concentración de calcio y la estimulación p-adrenérgica. Este protocolo BHM, sin modificaciones, fue capaz de producir constantemente b Hm a partir de múltiples líneas de hPSC (realizado en cada BHM en cada experimento). Los datos presentes sugieren que el protocolo BHM descrito en el presente texto es una forma robusta, libre de suero y reproducible para producir miocardio humano para múltiples aplicaciones. Por ejemplo, se demuestra también que el BHM es un modelo potencial de desarrollo del miocardio humano y se demuestra que la inhibición de la señalización de BMP conduce a un fenotipo cardíaco más inmaduro con una fuerza contráctil reducida.
Por consiguiente, la presente invención también está dirigida a un BHM producido por el método de acuerdo con la invención, como se define en las reivindicaciones con más detalle.
También se contempla el uso de BHM de acuerdo con la invención en un modelo in vitro para el cribado de la toxicidad de fármacos. En otras palabras, la presente invención también se dirige a un método para el cribado de la toxicidad del fármaco, que comprende la etapa de poner en contacto un BHM de acuerdo con la invención, con un fármaco que se desea cribar.
Además, la presente invención está dirigida al uso de BHM de acuerdo con la invención en un método in vitro para ensayar la modulación de la función cardíaca por agentes farmacológicos candidatos. Así pues, también se describe un método para probar la modulación de la función cardíaca, que comprende la etapa de poner en contacto un BHM de acuerdo con la invención con un agente farmacológico candidato.
Finalmente, la presente invención también se dirige al uso de BHM según la invención como herramienta de investigación, así como a un BHM de acuerdo con la invención para uso en medicina.
Descripción detallada de las realizaciones preferidas
Un método para producir músculo cardíaco de bioingeniería a partir de células madre pluripotentes, que comprende los pasos de
(i) cultivar células madre pluripotentes en un medio basal que comprende una cantidad efectiva de (a) BMP4, Activina A, FGF2, un inhibidor de GSK3, y (b) un suplemento libre de suero que da como resultado una concentración final de 0,5 - 50 mg /ml de albúmina, 1 - 100 pg/ml de transferrina, 0,1 - 10 pg/ml de etanolamina, 0,003 - 0,3 pg/ml de selenito de sodio, 0,4 - 40 pg/ml de L-carnitina HCl, 0,1 - 10 pg/ml de hidrocortisona, 0,05 - 5 pl/ml de suplemento de ácido graso, 0,0001 - 0,1 pg/ml de triyodo-L-tironina (T3), induciendo así la diferenciación mesodérmica de dichas células madre pluripotentes;
(ii) cultivar las células obtenidas en la etapa (i) en un medio basal que comprende una cantidad eficaz de un inhibidor de la ruta de señalización Wnt y un suplemento libre de suero como se define en la etapa (i), induciendo así la diferenciación cardíaca de las células; y
(iii) cultivar las células obtenidas en la etapa (ii) en un medio basal que comprende una cantidad eficaz de un suplemento libre de suero como se define en la etapa (i), bajo estimulación mecánica, promoviendo así la maduración cardíaca.
En una realización preferida, las células madre pluripotentes son células madre pluripotentes de origen primate, más preferiblemente las células madre pluripotentes son células madre pluripotentes humanas. Las células madre pluripotentes pueden diferenciarse en cada tipo de célula del cuerpo. Como tales, las células madre pluripotentes humanas ofrecen la oportunidad única de obtener células cardiacas humanas genuinas. Actualmente, las células pluripotentes más utilizadas son las células madre embrionarias (ESC) o las células madre pluripotentes inducidas (iPSC). Las líneas de ESC humanas fueron establecidas por primera vez por Thomson y colaboradores (Thomson et al., Science 282: 1145 - 1147 (1998). La investigación de ESC humana permitió recientemente el desarrollo de una nueva tecnología para reprogramar las células del cuerpo en una célula similar a ES. Esta tecnología fue iniciada por Yamanaka y colaboradores en 2006 (Takahashi & Yamanaka Cell 126: 663 - 676 (2006). Las células pluripotentes inducidas resultantes (iPSC) muestran un comportamiento muy similar al de ESC y, lo que es más importante, también son capaces de diferenciarse en cada célula del cuerpo. Además, se publicó que también es probable que las células madre partenogenéticas sean adecuadas para la producción de BHM (Didie et al. J Clin Invest. 123, 1285 - 1298 (2013). Por consiguiente, las células madre pluripotentes pueden seleccionarse entre células madre embrionarias, células madre pluripotentes inducidas y células madre partenogenéticas. En el contexto de la presente invención, dichas células madre pluripotentes, sin embargo, no se producen utilizando un proceso que implica modificar la identidad genética de la línea germinal de los seres humanos o que implica el uso de un embrión humano para fines industriales o comerciales.
El medio basal utilizado en la etapa (i) se puede seleccionar entre DMEM/F12, StemPro, medio de Iscove, aMEM, DMEM y RPMI. Preferiblemente, el medio basal usado en la etapa (i) está suplementado en RPMI con piruvato. Sin embargo, se puede usar en el método cualquier medio basal adecuado. Los medios basales están disponibles comercialmente o pueden prepararse de acuerdo con recetas que están a disposición pública, por ejemplo, en catálogos de la ATCC. Si se considera apropiado, el medio basal puede suplementarse con aminoácidos no esenciales. Si se usa aMEM como medio basal, el medio basal no necesita ser suplementado adicionalmente con aminoácidos no esenciales. Los aminoácidos no esenciales están disponibles comercialmente como un suplemento combinado. Dicho suplemento, por ejemplo, comprende 750 mg/L de glicina, 890 mg/L de L-alanina, 1320 mg/L de L-asparagina, 1330 mg/L de ácido L-aspártico, 1470 mg/L de ácido L-glutámico, 1150 mg/L de L-prolina, y 1050 mg/L de L-serina.
Como se estableció anteriormente, el medio basal de la etapa (i) comprende una cantidad efectiva de BMP4, Activina A, FGF2 y un inhibidor de GSK3. Por ejemplo, dicho medio basal comprende 1 - 20 ng/ml de BMP4, preferiblemente 2 - 15 ng/ml, más preferiblemente 2,5 -10 ng/ml, más preferiblemente 3 -8 ng/ml, lo más preferiblemente 4 -6 ng/ml, e incluso lo más preferiblemente aproximadamente 5 ng/ml;
0,1 -10 ng/ml de FGF2, preferiblemente 1 - 9 ng/ml, más preferiblemente 2 - 8 ng/ml, aún más preferiblemente 3 -7 ng/ml, lo más preferiblemente 4 -6 ng/ml, e incluso lo más preferiblemente aproximadamente 5 ng/ml;
1 - 20 ng/ml de Activina A, preferiblemente 2 ,5-18 ng/ml, más preferiblemente 5 - 16 ng/ml, aún más preferiblemente 7,5 - 14 ng/ml, aún más preferiblemente 8 - 12 ng/ml, lo más preferiblemente 8,5 - 10 ng/ml, e incluso lo más preferiblemente aproximadamente 9 ng/ml.
El inhibidor de GSK3 en el medio basal de la etapa (i) se puede seleccionar, por ejemplo, entre el grupo que consiste en CHIR99021, CHIR98014, SB216763, TWS119, Tide-glusib, SB415286 y LY2090314. Sin embargo, se puede aplicar cualquier inhibidor de GSK3 adecuado en el método de la invención. En una realización preferida, el inhibidor de GSK3 en el medio basal de la etapa (i) es CHIR99021.
Un experto en la materia entenderá que la concentración de una cantidad eficaz de un inhibidor de GSK3 varía con la disponibilidad y la constante de inhibición del inhibidor en cuestión. En el contexto de la presente invención, la expresión "cantidad eficaz" como se usa en el presente documento en el contexto de un inhibidor de GSK3 pretende significar una concentración inactivadora de enzima. Por ejemplo, en el caso de CHIR99021, el medio basal en la etapa (i) comprende 0,1 -10 pM de CHIR99021, preferiblemente 0,2 - 9 pM, más preferiblemente 0,3 - 8 pM, aún más preferiblemente 0,4 - 7 pM, aún más preferiblemente 0,5 - 6 pM, más preferiblemente 0,6 - 5 pM, más preferiblemente 0,7 - 4 pM, más preferiblemente 0,8 - 3 pM, lo más preferiblemente 0,9 - 2 pM, e incluso lo más preferiblemente aproximadamente 1 pM CHIR99021. Se entenderá que una concentración eficaz de cualquier receptor/agonista o inhibidor enzimático varía con la disponibilidad y la actividad biológica del compuesto respectivo. El suplemento libre de suero aplicado en las etapas (i), (ii) y (iii) del método está formulado para que resulte una concentración final de 0,5 - 50 mg/ml de albúmina (preferiblemente 1 - 40 mg/ml, más preferiblemente 2 - 30 mg/ml, aún más preferiblemente
3 - 20 mg/ml, lo más preferiblemente 4 - 10 mg/ml, e incluso lo más preferiblemente 4,5 - 7,5 mg/ml (aproximadamente 5 mg/ml),
1 -100 |jg/ml transferrina (preferiblemente 2 - 90 jg/ml, más preferiblemente 3 - 80 jg/ml, aún más preferiblemente 4 - 70 jg/ml, aún más preferiblemente 5 - 60 jg/ml, más preferiblemente 6 - 50 jg/m l, más preferiblemente 7 - 40 jg/ml, más preferiblemente 8 - 30 jg/ml, más preferiblemente 9 - 20 mg/ml, tal como aproximadamente 10 jg/ml),
0,1 - 10 jg/m l etanolamina (preferiblemente 0,2 - 9 jg/ml, más preferiblemente 0,3 - 8 jg/ml, incluso más preferiblemente 0,4 - 7 jg/ml, aún más preferiblemente 0,5 - 6 jg/ml, más preferiblemente 0,6 - 5 jg/ml, más preferiblemente 0,7 -  jg /ml, más preferiblemente 0,8 - 3 jg/ml, más preferiblemente 1 - 2,5 jg/ml, tal como aproximadamente 2 jg/ml),
jg /ml, más preferiblemente 0,8 - 3 jg/ml, más preferiblemente 1 - 2,5 jg/ml, tal como aproximadamente 2 jg/ml),
0,003 - 0,3 jg/m l de selenito sódico (preferiblemente 0,005 - 0,2 jg/ml, más preferiblemente 0,01 - 0,1 jg/ml, aún más preferiblemente 0,02 - 0,05 jg/ml, y lo más preferiblemente aproximadamente 0,03 jg/ml, tal como aproximadamente 0,032 jg/ml),
0,4 - 40 jg/m l de L-carnitina HCl (preferiblemente 0,5 - 30 jg/ml, más preferiblemente 1 - 20 jg/ml, aún más preferiblemente 2 - 10 jg/ml, más preferiblemente 3 -5 jg/ml, e incluso lo más preferiblemente alrededor de 4 jg/ml),
0,1 - 10 jg /l de hidrocortisona (preferiblemente 0,2 - 9 jg/ml, más preferiblemente 0,3 - 8 jg/ml, aún más preferiblemente 0,4 - 7 jg/ml, aún más preferiblemente 0,5 - 6 jg/ml, más preferiblemente 0,6 - 5 jg/ml, más preferiblemente 0,7 - 4 jg/ml, más preferiblemente 0,8 - 3 jg/ml, más preferiblemente 0,9 - 2 jg/ml, tal como aproximadamente 1 jg/ml),
0,05 - 5 jl/m l de suplemento de ácido graso (preferiblemente 0 , 1 - 4 jl/m l, más preferiblemente 0,2 - 3 jl/m l, aún más preferiblemente 0,3 - 3 jl/m l, lo más preferiblemente 0,4 - 2 jl/m l, e incluso lo más preferiblemente 0,45 - 1 jl/m l, tal como alrededor de 0,5 jl/m l), y
0,0001 - 0,1 jg/m l de triyodo-L-tironina (T3) (preferiblemente 0,001 - 0,01 jg/ml, más preferiblemente 0,002 - 0,0075 jg/ml, aún más preferiblemente 0,003 - 0,005 jg/ml, y lo más preferiblemente aproximadamente 0,004 jg/ml).
Además, el suplemento libre de suero puede comprender además uno o más componentes seleccionados entre el grupo que consiste en vitamina A, D-galactosa, L-carnitina, ácido linoleico, ácido linolénico, progesterona y putrescina.
Estos componentes son favorables para la viabilidad de las células. Las concentraciones adecuadas de los componentes respectivos son conocidas por el experto en la materia o pueden determinarse fácilmente usando medidas de rutina.
El suplemento libre de suero mencionado en la etapa (i) está también disponible comercialmente. Por ejemplo, se puede usar suplemento B27® o suplemento B27® menos insulina. En una realización preferida, el suplemento B27® o el suplemento B27® menos insulina usado en la etapa (i) del método anterior se aplica en una cantidad de 0,1 -10% de B27® o B27® menos insulina, preferiblemente 0,5 - 8%, más preferiblemente 1 - 6%, incluso más preferiblemente 1,5-4 %, y lo más preferiblemente aproximadamente 2% de B27® o B27® menos insulina.
Como se demuestra en los ejemplos que siguen, se ha probado que es ventajoso incluir una cantidad eficaz de ácido ascórbico o una sal del mismo en el medio basal de la etapa (i). En una realización preferida, el medio basal de la etapa (i) comprende 10 - 1000 jM , preferiblemente 50 - 400 jM , más preferiblemente 100 - 300 jM , aún más preferiblemente 150 - 250 jM , y lo más preferiblemente aproximadamente 200 jM de ácido ascórbico o una sal del mismo. El ácido ascórbico se puede administrar en forma libre o como una sal. Dado que el ascorbato es el ingrediente activo, se puede usar cualquier sal o derivado del ácido ascórbico, que proporcione el ascorbato a las células, siempre y cuando el ion contrario no tenga un efecto perjudicial sobre las células. Como se muestra en los ejemplos, una sal o derivado adecuado del ácido ascórbico es el ascorbato-2-fosfato.
La longitud de la etapa (i) y la concentración de factores como BMP4, Activina A, FGF2 y el inhibidor de GSK3 pueden optimizarse monitorizando la eficiencia de la inducción de la diferenciación del mesodermo. Esto se puede lograr controlando la expresión de la superficie celular o los marcadores de pluripotencia, es decir, mediante (a) una disminución de las células positivas para TRA-1-60 y OCT4 (células madre pluripotentes) y (b) un aumento de las células positivas MIXL1 y Mesp1 (mesodermo) (véase también la Fig. 4f en el presente texto).
Brevemente, las células se fijan usando etanol, se bloquean usando protocolos estándar y luego se tiñen con anticuerpos primarios dirigidos contra TRA-1-60, OCT4, MlXL1 y/o Mesp1 (véase la Tabla 2 a continuación) en un tampón de bloqueo durante 45 minutos, seguido opcionalmente por anticuerpos secundarios (si el anticuerpo primario no está marcado con fluorescencia) en tampón de bloqueo y Hoechst durante 30 minutos a 4 °C (véase la Tabla 2 a continuación). Se utiliza un LSRII BD para el análisis de citometría de flujo (BD Biosystems). Para las células vivas, las poblaciones se clasifican basándose en los perfiles de dispersión frontal y lateral. El software BD FACSDiva (BD Bioscience) o Cyflologic v1.2.1 (Cyflo Ltd) se utilizan para el análisis. La inducción de la diferenciación del mesodermo está indicada si
(a) menos del 50%, preferiblemente menos del 40%, más preferiblemente menos del 30%, aún más preferiblemente menos del 20%, lo más preferiblemente menos del 10%, e incluso más preferiblemente menos del 5% de las células de la población de células vivas son positivas para TRA-1-60; y/o menos del 50%, preferiblemente menos del 40%, más preferiblemente menos del 30%, incluso más preferiblemente menos del 20%, lo más preferiblemente menos del 10%, y aún más preferiblemente menos del 5% de las células de la población de células vivas es positiva para OCT4; y
(b) más del 20%, preferiblemente más del 30%, más preferiblemente más del 40%, incluso más preferiblemente más del 50%, y más preferiblemente más del 60%, de las células de la población de células vivas son positivas para MIXL1; y/o más del 20%, preferiblemente más del 30%, más preferiblemente más del 40%, incluso más preferiblemente más del 50%, y más preferiblemente más del 60% de las células de la población viva son positivas para Mesp1.
Normalmente, la etapa (i) se realiza durante 48 - 96 h. Preferiblemente, la etapa (i) se lleva a cabo durante 60 - 84 h, y más preferiblemente la etapa (i) se lleva a cabo durante 66 - 78 h.
El medio basal utilizado en la etapa (ii) se puede seleccionar entre DMEM/F12, StemPro, medio de Iscove, aMEM, DME y RPMI. Preferiblemente, el medio basal usado en la etapa (ii) es RPMI suplementado con piruvato. Sin embargo, se puede usar en el método cualquier medio basal adecuado.
Si se considera apropiado, el medio basal de la etapa (ii) puede suplementarse con aminoácidos no esenciales. Si se usa aMEM como medio basal en la etapa (ii), el medio basal no necesita suplementarse adicionalmente con aminoácidos no esenciales. Los aminoácidos no esenciales son disponibles comercialmente como un suplemento combinado. Dicho suplemento, por ejemplo, comprende 750 mg/L de glicina, 890 mg/L de L-alanina, 1320 mg/L de L-asparagina, 1330 mg/L de ácido L-aspártico, 1470 mg/L de ácido L-glutámico, 1150 mg/L de L-prolina, y 1050 mg/L de L-serina.
El medio basal en la etapa (ii) puede seleccionarse independientemente del medio basal aplicado en la etapa (i). Sin embargo, en una realización preferida, el medio basal en los pasos (i) y (ii) es el mismo.
El inhibidor de la ruta de señalización Wnt en el medio basal de la etapa (ii) puede ser cualquier inhibidor de la ruta de señalización Wnt, que puede aplicarse adecuadamente en el método de la invención. Preferiblemente, dicho inhibidor de la ruta de señalización Wnt se selecciona entre el grupo que consiste en IWP4, IWP2, IWR-1, IWP1, IWP3, IWR-2, IWR-3, IWR-4, IWR-5, XAV939, DKK1, quercetina, ICG-001, pirvinio, CCT031374, iCRT-3,5,14, CPG049090 y NC043. Más preferiblemente, dicho inhibidor de la ruta de señalización Wnt se selecciona entre el grupo que consiste en IWP4, IWP2, IWR-1, IWP1, IWP3, IWR-2, IWR-3, IWR-4, IWR-5, XAV939, DKK1. Como se demuestra en los ejemplos IWR-2, IWR-3, IWR-4, IWR-5, XAV939, DKK1. Como se demuestra en los ejemplos que siguen, un inhibidor particularmente útil de la vía de señalización Wnt en el medio basal de la etapa (ii) es IWP4.
El suplemento libre de suero mencionado en la etapa (ii) es como se define para la etapa (i) anterior. Los suplementos libres de suero aplicados en las etapas (i) y (ii) pueden ser iguales o no. Del mismo modo, el suplemento B27® o el suplemento B27® menos insulina se pueden usar en la etapa (ii). En una realización preferida, el suplemento B27® o el suplemento B27® menos insulina usado en la etapa (ii) del método anterior se aplica en una cantidad de 0,1 -10% de b27® o B27® menos insulina, preferiblemente 0,5 - 8%, más preferiblemente 1 - 6%, aún más preferiblemente 1,5 - 4 %, y lo más preferiblemente aproximadamente 2% de B27® o B27® menos insulina.
El profesional experto en la técnica entenderá que la concentración de una cantidad eficaz de un inhibidor de la ruta de señalización Wnt varía con la disponibilidad y la constante de inhibición del inhibidor en cuestión. Por ejemplo, en el caso de IWP4, el medio basal de la etapa (ii) puede comprender 0,1 -10 pM IWP4, preferiblemente 1 - 9 pM, más preferiblemente 2 - 8 pM, aún más preferiblemente 3 - 7 pM, aún más preferiblemente 4 - 6 pM, y lo más preferiblemente aproximadamente 5 pM de IWP4. Se entenderá que una concentración eficaz de cualquier receptor/enzima agonista o inhibidor varía con la disponibilidad y la actividad biológica del compuesto respectivo.
Como se demuestra en los ejemplos más adelante, se ha probado que es ventajoso incluir una cantidad eficaz de ácido ascórbico o una sal del mismo en el medio basal de la etapa (ii). En una realización preferida, el medio basal de la etapa (ii) comprende 10 - 1000 pM, preferiblemente 50 - 400 pM, más preferiblemente 100 - 300 pM, aún más preferiblemente 150 - 250 pM, y lo más preferiblemente aproximadamente 200 pM de ácido ascórbico o una sal del mismo. El ácido ascórbico se puede suministrar en forma libre o como una sal. Dado que el ascorbato es el ingrediente activo, se puede usar cualquier sal del ácido ascórbico, que proporciona el ascorbato a las células siempre que el ión contrario no tenga un efecto perjudicial sobre las células. Como se muestra en los ejemplos, una sal o derivado adecuado de ácido ascórbico para uso en el medio basal en la etapa (ii) es ascorbato-2-fosfato.
La longitud de la etapa (ii) y la concentración de los restantes constituyentes, tal como el inhibidor de la ruta de señalización Wnt, pueden optimizarse controlando la eficiencia de la inducción de la diferenciación cardíaca de las células. Esto puede conseguirse controlando la expresión de los marcadores de diferenciación, es decir, mediante un aumento de Nkx2.5 y actinina.
Brevemente, las células se fijan con etanol, se bloquean y luego se tiñen con anticuerpos primarios dirigidos contra Nkx2.5 y/o actinina (véase la Tabla 2 a continuación) en tampón de bloqueo durante 45 minutos, seguido opcionalmente por anticuerpos secundarios (si el anticuerpo primario no está marcado con fluorescencia) en tampón de bloqueo y Hoechst durante 30 min a 4 °C (véase la Tabla 2 a continuación). Se utiliza un BD LSRII para el análisis de citometría de flujo (BD Biosystems). Para las células vivas, las poblaciones se clasifican según los perfiles de dispersión frontal y lateral. El software BD FACSDiva (BD Bioscience) o Cyflologic v1.2.1 (Cyflo Ltd) se utilizan para el análisis. La inducción de la diferenciación cardíaca está indicada si más del 20%, preferiblemente más del 30%, más preferiblemente más del 40%, incluso más preferiblemente más del 50%, y lo más preferiblemente más del 60% de las células de la población de células vivas son positivas para Nkx2.5; y/o más del 20%, preferiblemente más del 30%, más preferiblemente más del 40%, incluso más preferiblemente más del 50%, y lo más preferiblemente más del 60% de las células de la población de células vivas son positivas para la actinina (véanse también las figuras 4d y 4f del presente documento).
Por lo general, la etapa (ii) se lleva a cabo durante 8 - 12 días. Preferiblemente, la etapa (ii) se lleva a cabo durante 9 -11 días, y lo más preferiblemente la etapa (ii) se lleva a cabo durante 10 días.
El medio basal utilizado en la etapa (iii) se puede seleccionar entre DMEM/F12, StemPro, medio de Iscove, aMEM, DMEM y RPMI. Preferiblemente, el medio basal usado en la etapa (iii) está suplementado en RPMI con piruvato. Sin embargo, se puede usar en el método cualquier medio basal adecuado.
Si se considera apropiado, el medio basal de la etapa (iii) puede suplementarse con aminoácidos no esenciales. Si se utiliza aMEM como medio basal en la etapa (iii), el medio basal no necesita suplementarse adicionalmente con aminoácidos no esenciales. Los aminoácidos no esenciales están disponibles comercialmente como un suplemento combinado. Dicho suplemento, por ejemplo, comprende 750 mg/L de glicina, 890 mg/L de L-alanina, 1320 mg/L de L-asparagina, 1330 mg/L de ácido L-aspártico, 1470 mg/L de ácido L-glutámico, 1150 mg/L de L-prolina, y 1050 mg/L de L-serina.
El medio basal en la etapa (iii) puede seleccionarse independientemente del medio basal aplicado en las etapas (i) y/o (ii). Sin embargo, en una realización preferida, el medio basal en los pasos (ii) y (iii) es el mismo. Más preferiblemente, el medio basal en las etapas (i), (ii) y (iii) es el mismo.
Como se demuestra en los ejemplos que siguen, se ha probado que es ventajoso incluir una cantidad eficaz de ácido ascórbico o una sal del mismo en el medio basal de la etapa (iii). En una realización preferida, el medio basal de la etapa (iii) comprende 10 - 1000 pM, preferiblemente 50 - 400 pM, más preferiblemente 100 - 300 pM, aún más preferiblemente 150 - 250 pM, y lo más preferiblemente aproximadamente 200 pM de ácido ascórbico o una sal del mismo. El ácido ascórbico se puede administrar en forma libre o como una sal. Dado que el ascorbato es el ingrediente activo, se puede usar cualquier sal o derivado del ácido ascórbico, que proporciona el ascorbato a las células, siempre que el ion contrario no tenga un efecto perjudicial sobre las células. Como se muestra en los ejemplos, una sal adecuada de ácido ascórbico para uso en el medio basal en la etapa (iii) es ascorbato-2-fosfato.
El suplemento libre de suero al que se hace referencia en la etapa (iii) es un suplemento sin suero como se define para la etapa (i) anterior. Los suplementos libres de suero aplicados en los pasos (i), (ii) y (iii) pueden ser iguales o no. Del mismo modo, el suplemento B27® o el suplemento B27® menos insulina se pueden usar en la etapa (iii). En una realización preferida, el suplemento B27® o el suplemento B27® menos insulina usado en la etapa (iii) del método anterior se aplica en una cantidad de 0,1 - 10% de B27® o B27® menos insulina, preferiblemente 0,5 - 8%, más preferiblemente 1 - 6%, aún más preferiblemente 1,5-4 %, y lo más preferiblemente aproximadamente 2% de B27® o B27® menos insulina.
El medio basal de la etapa (iii) comprende además una cantidad eficaz de TGFp1. Por ejemplo, el medio basal de la etapa (iii) puede comprender 0,1 - 10 ng/ml de TGFp1, preferiblemente 0,2 - 9 ng/ml, más preferiblemente 0,3 - 8 ng/ml, incluso más preferiblemente 0,4 - 7 ng/ml, aún más preferiblemente 0,5 - 6 ng/ml, más preferiblemente 0,6 - 5 ng/ml, más preferiblemente 0,7 - 4 ng/ml, más preferiblemente 0,8 - 3 ng/ml, lo más preferiblemente 0,9 - 2 ng/ml, e incluso lo más preferiblemente aproximadamente 1 ng/ml de TGFp1.
Como se muestra en los ejemplos, para la maduración cardíaca es ventajoso que el medio basal de la etapa (iii) no comprenda una cantidad efectiva de FGF2. Al contrario que esto, se ha demostrado que el calcio mejora la maduración cardíaca. Por consiguiente, en una realización preferida, el medio basal de la etapa (iii) comprende 0,5 - 3 mM de Ca2+, preferiblemente 0,5 - 2,75 mM de Ca2+, más preferiblemente 1 - 2,25 mM de Ca2+, incluso más preferiblemente 1-1,5 mM mM de Ca2+, y lo más preferiblemente aproximadamente 1,2 mM de Ca2+.
Normalmente, la etapa (iii) del método de la invención se lleva a cabo bajo estimulación mecánica, por ejemplo, en un dispositivo de estiramiento, como se conoce generalmente en la técnica. Preferiblemente, el dispositivo de estiramiento aplica un estiramiento estático, fásico o dinámico al BHM. Más específicamente, el estiramiento mecánico puede ser (a) estático, (b) dinámico o (c) flexible contra una carga resiliente. Preferiblemente, la estimulación mecánica en la etapa (iii) es una estimulación mecánica dinámica o un estiramiento estático. En una realización más preferida, la
estimulación mecánica en la etapa (iii) es una estimulación mecánica dinámica contra una carga resiliente para facilitar las contracciones auxotónicas.
Se puede probar si se promueve la maduración cardíaca mediante inspección óptica para observar contracciones espontáneas o estimuladas eléctricamente. Preferiblemente, la maduración cardíaca se controla mediante un experimento de contracción isométrica, en el que un desarrollo de fuerza de contracción de > 0,01 mN es indicativo de la maduración cardíaca.
Brevemente, los experimentos de contracción se realizan en baños de órganos a 37 °C bajo burbujeo constante con 5% de CO2 y 95% de O2 para mantener un pH fisiológico en la solución de Tyrode que contiene (todo en mM): 120 NaCl, 1 MgCl2, 0,2 CaCl2, 5,4 KCl, 22,6 NaHCO3, 4,2 NaH2PO4, 5,6 glucosa y 0,56 ascorbato. El calcio se ajusta utilizando una solución de cloruro de calcio 0,2 M. Todos los BHM se analizan a 3 Hz con pulsos cuadrados de 5 ms de 200 mA de corriente eléctrica para estimar aproximadamente la frecuencia cardíaca embrionaria. La frecuencia de estimulación se altera para confirmar la respuesta correcta de fuerza-frecuencia (mecanismo de Bowditch). Los BHM se estiran mecánicamente a intervalos de 125 pm hasta que se observa la fuerza máxima de contracción (respuesta de fuerza-longitud; mecanismo de Frank-Starling).
Normalmente, la etapa (iii) se realiza durante al menos 72 h. Aunque no hay un límite superior particular para la longitud de la etapa (iii), dicha etapa se lleva a cabo generalmente durante menos de 100 días. En realizaciones específicas, la etapa (iii) puede llevarse a cabo durante 4 - 50 días, tal como durante aproximadamente 15 días.
La etapa (i) del método de la invención puede estar precedida por una etapa de siembra, en donde dichas células madre pluripotentes se siembran en una relación de (2,5 - 6 x 106 células/1 mg de colágeno)/1 ml de medio en un molde adecuado. Preferiblemente, la etapa de siembra se lleva a cabo 18-30 h antes de la etapa (i).
El medio utilizado en la etapa de siembra generalmente comprende 0,2 - 2 mg/ml de colágeno (preferiblemente 0,3 -1,9 mg/ml, más preferiblemente 0,4 - 1,8 mg/ml, aún más preferiblemente 0,4 - 1,7 mg/ml, aún más preferiblemente 0,5 -1,6 mg/ml, más preferiblemente 0,6 -1,5 mg/ml, más preferiblemente 0,7 -1,4 mg/ml, más preferiblemente 0,8 -1,3 mg/ml, más preferiblemente 0,9 -1,2 mg/ml, tal como aproximadamente 1 mg/ml). Preferiblemente el colágeno es de grado médico y se selecciona entre el grupo que consiste en colágeno de tipo I, colágeno de tipo III, colágeno de tipo V y una mezcla de los mismos. En una realización más preferida, al menos el 90% de dicho colágeno es colágeno de tipo I. Sin embargo, dicho colágeno también puede comprender además uno o más componentes de la matriz extracelular seleccionados entre el grupo que consiste en elastina, laminina, entactina, nidógeno, proteoglicano y fibronectina. Normalmente, la composición exacta del colágeno dependerá del origen del que se deriva. El colágeno es preferiblemente de origen humano, pero también se contempla el origen bovino o porcino, o el origen marino, tal como a partir de algas o peces. Alternativamente, también se puede usar colágeno recombinante.
Para lograr alcanzar celulares adecuadas, para algunas líneas celulares pluripotentes puede ser útil suplementar el medio utilizado en la etapa de siembra con un inhibidor de ROCK. Por lo tanto, en una realización preferida, el medio usado en la etapa de siembra comprende además un inhibidor de ROCK. El inhibidor de ROCK puede ser cualquier inhibidor de ROCK que pueda aplicarse adecuadamente en el método de la invención. Preferiblemente, dicho inhibidor de ROCK se selecciona entre Y27632, H-1152P, Tiazovivina, Fasudil, Hidroxifasudil, GSK429286A, y RKI-1447, preferiblemente seleccionado entre Y27632, H-1152P, Tiazovivina, Fasudil, Hidroxifasudil, y más preferiblemente el inhibidor de ROCK se selecciona entre Y27632 o H-1152P. Como se demuestra en los ejemplos a continuación, un inhibidor de ROCK particularmente útil es Y27632.
El experto en la técnica entenderá que la concentración de una cantidad eficaz de un inhibidor de ROCK varía con la disponibilidad y la constante de inhibición del inhibidor en cuestión. Por ejemplo, en el caso de Y27632, el medio utilizado en la etapa de siembra puede comprender 1 - 50 pM, preferiblemente 2,5 - 40 pM, más preferiblemente 5 -30 pM, incluso más preferiblemente 7,5 - 20 pM, lo más preferiblemente 8 - 12 pM, y lo más preferiblemente aproximadamente 10 pM Y27632.
Se entenderá que una concentración eficaz de cualquier agonista o inhibidor de receptor/enzima varía con la disponibilidad y la actividad biológica del compuesto respectivo.
Aparte del método descrito anteriormente, la invención se refiere además a un BHM producido por dicho método, como se define en las reivindicaciones. A pesar del aumento de madurez observada en el protocolo BHM de los presentes autores, también se debe tener en cuenta que el BHM es todavía un tejido relativamente inmaduro. En comparación con el tejido cardíaco adulto, el BHM todavía tiene una relación ¡3-MHC/a-MHC inferior, y una expresión baja pero todavía retenida de genes progenitores (por ejemplo, ISL1). Sin embargo, el cultivo prolongado bajo condiciones de cultivo apropiadas con estimulación biofísica puede incrementar aún más la madurez. Ya hay evidencia morfológica que sugiere que esto también puede ser el caso en el sistema BHM.
El BHM obtenido por el método descrito en el presente texto presenta las siguientes características: Puede ser estimulado a múltiples frecuencias hasta al menos 3 Hz, exhibe un valor de EC50 de calcio superior a 0,2 mM, preferiblemente en el rango fisiológico de 4 - 8 mM, y una tensión decontracción de más de 200 pN. La tensión de
contracción aumenta en respuesta al aumento de la longitud de reposo y la tensión de reposo. En respuesta a isoprenalina 1 j M, el BHM exhibe una respuesta inotrópica de más de 40 j N en condiciones estimuladas a 0,6 mM de calcio, preferiblemente más de 45 j N, más preferiblemente más de 50 j N. En resumen, todos los experimentos de contracción se realizan en baños de órganos a 37 °C y pH fisiológico en solución de Tyrode que contiene (todo en mM): 120 NaCl, 1 MgCh/0,2 CaCh, 5,4 KCl, 22,6 NaHCOa, 4,2 NaH2PO4, 5,6 glucosa y 0,56 ascorbato. El calcio se ajusta utilizando una solución de cloruro de calcio 0,2 M. Todos los BHM se analizan a 3 Hz con pulsos cuadrados de 5 ms de corriente eléctrica de 200 mA para poder estimar aproximadamente la frecuencia cardíaca embrionaria. La frecuencia de estimulación se altera para confirmar la respuesta correcta frecuencia - fuerza (mecanismo de Bowditch). Los BHM se estiran mecánicamente a intervalos de 125 jm hasta que se observa la fuerza de contracción máxima a 2 mM de calcio (mecanismo de Frank-Starling). Posteriormente, los BHM se someten a diferentes concentraciones de calcio (0,2, 0,4, 0,8, 1,2, 1,6, 2,0, 2,4 mM) y se registra la fuerza de contracción. Para los experimentos con isoprenalina, la concentración de calcio se ajusta a 0,6 mM, y posteriormente la concentración de isoprenalina se ajusta a 1 j M.
Otra característica de la BHM obtenida por el método descrito en el presente texto es que comprende células estromales CD90+. La expresión de CD90 se puede determinar utilizando citometría de flujo. Brevemente, las células se fijan usando etanol, se bloquean y luego se tiñen con anticuerpos primarios dirigidos contra CD90 (véase la Tabla 2 a continuación) en tampón de bloqueo durante 45 minutos, seguido opcionalmente por anticuerpos secundarios (si el anticuerpo primario no está marcado con fluorescencia) en tampón de bloqueo y Hoechst durante 30 min a 4 °C (véase la Tabla 2 a continuación). Se utiliza un BD LSRII para el análisis de citometría de flujo (BD Biosystems). Para las células vivas, las poblaciones se clasifican basándose en los perfiles de dispersión frontal y lateral. El software BD FACSDIva (BD Bioscience) o Cyflologic v1.2.1 (Cyfio Ltd) se utilizan para el análisis.
El BHM puede proporcionar un buen sistema modelo para investigar mecanismos impulsores de la maduración en un entorno libre de suero, y los autores de la presente invención ya han demostrado que con el aumento de los periodos de cultivo la madurez puede aumentar (mostraron una mayor sensibilidad a la isoprenalina y mejoraron la morfología del tejido). La capacidad del cultivo BHM a largo plazo sin pérdida de función (al menos 63 días) también sugiere que es posible realizar experimentos de eficacia y seguridad farmacológica a largo plazo. Por lo tanto, en una realización preferida, el BHM obtenido por el método descrito aquí se puede mantener durante al menos 63 días.
Usando el planteamiento tradicional de diferenciación seguido de la ingeniería de tejidos, los cultivos de diferenciación requieren extensos protocolos de digestión para producir células individuales o pequeños grupos necesarios para las aplicaciones de ingeniería de tejido cardíaco. Estos protocolos de digestión destruyen el entorno extracelular y la distribución espacial formada durante el desarrollo y, por tanto, pueden tener un efecto inhibitorio difícil de controlar en el protocolo de diferenciación cardíaca.
Utilizando el BHM como modelo, los presentes autores demostraron que los factores que afectan el desarrollo temprano (ASC-2-P, dorsomorfina) y el desarrollo tardío (estimulación mecánica, FGF2, TGFp1 y concentración de calcio) tuvieron un profundo impacto sobre la función y las propiedades de BHM. Por lo tanto, el protocolo BHM de los presentes autores puede ser una herramienta útil en el estudio de los procesos de desarrollo que no solo rigen la cardiogénesis sino también la formación de tejidos y sus propiedades.
Por consiguiente, el BHM obtenido por el método descrito en el presente documento puede usarse adecuadamente como herramienta de investigación. Por ejemplo, se contempla el uso del BHM obtenido por el método descrito en el presente documento en un modelo in vitro para el cribado de la toxicidad de medicamentos. En otras palabras, se contempla un método para el cribado de la toxicidad del fármaco, que comprende la etapa de poner en contacto un BHM obtenido mediante el método descrito en el presente documento con el fármaco a examinar. Alternativamente, el BHM obtenido por el método descrito en el presente documento puede usarse en un método in vitro para probar la modulación de la función cardíaca por agentes candidato farmacológicos. Por lo tanto, también se describe un método para probar la modulación de la función cardíaca, que comprende la etapa de poner en contacto un BHM de acuerdo con la invención con un agente candidato farmacológico.
Finalmente, el BHM obtenido por el método descrito aquí se puede usar en medicina. Simplemente a modo de ejemplo, se contempla que el BHM obtenido por el método descrito en el presente documento se puede usar ventajosamente en la reparación del corazón.
La invención se describe adicionalmente mediante las siguientes realizaciones:
1. Un método para producir músculo cardíaco de bioingeniería a partir de células madre pluripotentes, que comprende las etapas de
(i) cultivar células madre pluripotentes en un medio basal que comprende una cantidad efectiva de (a) BMP4, Activina A, FGF2, un inhibidor de GSK3, y (b) un suplemento libre de suero que tenga como resultado una concentración final de 0,5 - 50 mg/ml de albúmina, 1 -100 jg/m l de transferrina, 0,1 -10 jg/m l de etanolamina, 0,003 - 0,3 jg/m l de selenito de sodio, 0,4 - 40 jg/m l de L-carnitina HCl, 0,1 - 10 jg/m l de hidrocortisona, 0,05 - 5 jl/m l suplemento de ácido graso, 0,0001 - 0,1 jg/m l de
triyodo-L-tironina (T3), induciendo así la diferenciación mesodérmica de dichas células madre pluripotentes;
(ii) cultivar las células obtenidas en la etapa (i) en un medio basal que comprende una cantidad eficaz de un inhibidor de la ruta de señalización Wnt y un suplemento libre de suero como en (i), induciendo así la diferenciación cardíaca de las células; y
(iii) cultivar las células obtenidas en la etapa (ii) en un medio basal que comprende una cantidad efectiva de un suplemento libre de suero como en (i), bajo estimulación mecánica, promoviendo así la maduración cardíaca.
El método de la realización 1, en el que las células madre pluripotentes son células madre pluripotentes de origen primate, preferiblemente células madre pluripotentes humanas.
El método de la realización 1 o 2, en el que las células madre pluripotentes se seleccionan entre células madre embrionarias, células madre pluripotentes inducidas y células madre partenogenéticas.
El método de una cualquiera de las realizaciones 1 - 3 , en el que el medio basal de la etapa (i) comprende 10 - 1000 |jM, preferiblemente 50 - 400 j M, más preferiblemente 100 - 300 j M, incluso más preferiblemente 150 - 250 j M, y lo más preferiblemente aproximadamente 200 j M de ácido ascórbico o una sal del mismo. El método de la realización 4, en el que la sal de ácido ascórbico es ascorbato-2-fosfato.
El método de una cualquiera de las realizaciones 1-5 , en el que la etapa (i) se lleva a cabo durante 48 - 96 h, preferiblemente en la que la etapa (i) se lleva a cabo durante 60 - 84 h, más preferiblemente en el que la etapa (i) se lleva a cabo durante 66 a 78 h.
El método de una cualquiera de las realizaciones 1-6 , en el que el medio basal en la etapa (i) comprende 1 - 20 ng/ml de BMP4, preferiblemente 2 - 15 ng/ml, más preferiblemente 2,5 - 10 ng/ml, más preferiblemente 3 -8 ng/ml, más preferiblemente 4 -6 ng/ml, e incluso lo más preferiblemente aproximadamente 5 ng/ml. El método de una cualquiera de las realizaciones 1 - 7 , en el que el medio basal en la etapa (i) comprende 0,1 - 10 ng/ml de FGF2, preferiblemente 1 - 9 ng/ml, más preferiblemente 2 - 8 ng/ml, incluso más preferiblemente 3 - 7 ng/ml, más preferiblemente 4 - 6 ng/ml, e incluso lo más preferiblemente aproximadamente 5 ng/ml.
El método de una cualquiera de las realizaciones 1 - 8, en el que el medio basal en la etapa (i) comprende 1 - 20 ng/ml de Activina A, preferiblemente 2,5 - 18 ng/ml, más preferiblemente 5 - 16 ng/ml, incluso más preferiblemente 7 ,5-14 ng/ml, aún más preferiblemente 8 - 12 ng/ml, lo más preferiblemente 8 ,5-10 ng/ml, e incluso lo más preferiblemente aproximadamente 9 ng/ml.
El método de una cualquiera de las realizaciones 1-9 , en el que el inhibidor de GSK3 en el medio basal de la etapa (i) se selecciona entre el grupo que consiste en CHIR99021, CHIR98014, SB216763, TWS119, Tideglusib, SB415286 y LY2090314.
El método de la realización 10, en el que el inhibidor de GSK3 en el medio basal de la etapa (i) es CHIR99021. El método de la realización 11, en el que el medio basal en la etapa (i) comprende 0,1 -10 j M CHIR99021, preferiblemente 0,2 - 9 j M, más preferiblemente 0,3 - 8 j M, incluso más preferiblemente 0,4 - 7 j M, aún más preferiblemente 0,5 - 6 j M, más preferiblemente 0,6 - 5 j M, más preferiblemente 0,7 - 4 j M, más preferiblemente 0,8 - 3 j M, más preferiblemente 0,9 - 2 j M, e incluso lo más preferiblemente aproximadamente 1 j l CHR99021.
El método de una cualquiera de las realizaciones 1 - 12, en el que el suplemento libre de suero en la etapa (i) comprende 0,1 - 10% B27 o B27 menos insulina, preferiblemente 0,5 - 8%, más preferiblemente 1 - 6%, incluso más preferiblemente 1,5-4 %, y lo más preferiblemente aproximadamente 2% de B27 o B27 menos insulina.
El método de una cualquiera de las realizaciones 1-13, en el que el medio basal usado en la etapa (i), es DMEM/F12, StemPro, medio de Iscove, aMEM, DMEM y RPMI.
El método de la reivindicación 14, en el que el medio basal usado en la etapa (i) es RPMI suplementado con piruvato.
El método de una cualquiera de las realizaciones 1-15, en el que el inhibidor de la ruta de señalización Wnt en el medio basal de la etapa (ii) se selecciona entre el grupo que consiste en IWP4, IWP2, IWR-1, IWP1,
IWP3, IWR-2, IWR-3, IWR-4, IWR-5, XAV939, DKK1, quercetina, ICG-001, pirvinio, CCT031374, iCRT-3,5,14, CPG049090, NC043; preferiblemente seleccionado entre el grupo que consiste en IWP4, IWP2, IWR-1, IWP1, IWP3, IWR-2, IWR-3, IWR-4, IWR-5, XAV939, DKK1.
El método de la realización 16, en el que el inhibidor de la ruta de señalización Wnt en el medio basal de la etapa (ii) es IWP4.
El método de la realización 17, en el que el medio basal de la etapa (ii) comprende 0 , 1 - 10 pM de IWP4, preferiblemente 1 - 9 pM, más preferiblemente 2 - 8 pM, incluso más preferiblemente 3 - 7 pM, aún más preferiblemente 4 -6 pM , y lo más preferiblemente aproximadamente 5 pM de IWP4.
El método de una cualquiera de las realizaciones 1-18, en el que el medio basal de la etapa (ii) comprende 10 - 1000 pM, preferiblemente 50 - 400 pM, más preferiblemente 100 -300 pM, aún más preferiblemente 150 - 250 pM, y lo más preferiblemente aproximadamente 200 pM de ácido ascórbico o una sal del mismo.
El método de la realización 19, en el que la sal o derivado del ácido ascórbico es ascorbato-2-fosfato.
El método de una cualquiera de las realizaciones 1 - 20, en el que la etapa (ii) se lleva a cabo durante 8 - 12 días, preferiblemente en la que la etapa (ii) se lleva a cabo durante 9 - 11 días, lo más preferiblemente en la que se realiza la etapa (ii) durante 10 días.
El método de una cualquiera de las realizaciones 1-21, en el que el suplemento libre de suero en la etapa (ii) comprende 0,1 - 10% de B27 o B27 menos insulina, preferiblemente 0,5 - 8%, más preferiblemente 1 -6%, incluso más preferiblemente 1,5 - 4%, y lo más preferiblemente alrededor de 2% de B27 o B27 menos insulina.
El método de una cualquiera de las realizaciones 1 - 22, en el que el medio basal usado en la etapa (ii), es DMEM/F12, StemPro, medio de Iscove, aMEM, DMEM y RPMI.
El método de la realización 23, en el que el medio basal usado en la etapa (ii) es RPMI suplementado con piruvato.
El método de una cualquiera de las realizaciones 1 - 24, en el que el medio basal de la etapa (iii) comprende además 10 - 1000 pM, preferiblemente 50 - 400 pM, más preferiblemente 100 - 300 pM, aún más preferiblemente 150 - 250 pM, y lo más preferiblemente aproximadamente 200 pM de ácido ascórbico o una sal del mismo.
El método de la realización 25, en el que la sal del ácido ascórbico es ascorbato-2-fosfato.
El método de una cualquiera de las realizaciones 1 - 26, en el que la etapa (iii) se lleva a cabo durante al menos 72 h, preferiblemente en el que la etapa (iii) se lleva a cabo durante menos de 100 días, más preferiblemente en el que se realiza la etapa (iii) durante 4 - 50 días, y más preferiblemente en el que la etapa (iii) se lleva a cabo durante aproximadamente 15 días.
El método de una cualquiera de las realizaciones 1 - 27, en el que el suplemento libre de suero en la etapa (iii) comprende 0,1 - 10% de B27 o B27 menos insulina, preferiblemente 0,5 - 8%, más preferiblemente 1 -6% 7, incluso más preferiblemente 1,5 - 4%, y lo más preferiblemente aproximadamente 2% de B27 o B27 menos insulina.
El método de una cualquiera de las realizaciones 1 - 28, en el que el medio basal usado en la etapa (iii), es DMEM/F12, StemPro, medio de Iscove, aMEM, DMEM y RPMI.
El método de la realización 29, en el que el medio basal usado en la etapa (i) es RPMI suplementado con piruvato.
El método de una cualquiera de las realizaciones 1 - 30, en el que el medio basal de la etapa (iii) comprende además 0,1 - 10 ng/ml de TGFp1, preferiblemente 0,2 - 9 ng/ml, más preferiblemente 0,3 - 8 ng/ml, incluso más preferiblemente 0,4 - 7 ng/ml, aún más preferiblemente 0,5 - 6 ng/ml, más preferiblemente 0,6 - 5 ng/ml, más preferiblemente 0,7 - 4 ng/ml, más preferiblemente 0,8 - 3 ng/ml, más preferiblemente 0,9 - 2 ng/ml, e incluso lo más preferiblemente aproximadamente 1 ng/ml de TGFp1.
El método de una cualquiera de las realizaciones 1 - 31, en el que el medio basal de la etapa (iii) no comprende una cantidad efectiva de FGF2.
33. El método de una cualquiera de las realizaciones 1 - 32, en donde el medio basal de la etapa (Ni) comprende 0,5 - 3 mM Ca2+, preferiblemente 0,5 - 2,75 mM Ca2+, más preferiblemente 1 - 2,25 mM Ca2+, incluso más preferiblemente 1-1,5 mM Ca2+, y lo más preferiblemente aproximadamente 1,2 mM Ca2+.
34. El método de una cualquiera de las realizaciones 1 - 33, en el que la estimulación mecánica en la etapa (iii) es estimulación mecánica dinámica o estiramiento estático.
35. El método de la realización 34, en el que la estimulación mecánica en la etapa (iii) es estimulación mecánica dinámica.
36. El método de una cualquiera de las realizaciones 1 - 35, que comprende antes de la etapa (i) una etapa de siembra, en donde dichas células madre pluripotentes se siembran en una relación de (2,5 - 6 x 106 células/1 mg de colágeno)/ml de medio en un molde adecuado.
37. El método de la realización 36, en el que el colágeno es colágeno I.
38. El método de una cualquiera de las realizaciones 36 - 37, en el que dicho colágeno es de origen humano, de origen bovino o de origen marino.
39. El método de una cualquiera de las realizaciones 36 - 38, en el que el medio usado en la etapa de siembra comprende además un inhibidor de ROCK.
40. El método de la realización 39, en el que el inhibidor de ROCK se selecciona entre Y27632, H-1152P, tiazovivina, fasudil, hidroxifasudil, GSK429286A y RKI-1447, preferiblemente seleccionado entre Y27632, H-1152P, tiazovivina, fasudil, hidroxifasudil, y más preferiblemente el inhibidor de ROCK es Y27632 o H-1152P, y lo más preferiblemente el inhibidor de ROCK es Y27632.
41. El método de la realización 40, en el que el medio utilizado en la etapa de siembra comprende 1 - 50 pM, preferiblemente 2,5 - 40 pM, más preferiblemente 5 - 30 pM, incluso más preferiblemente 7,5 - 20 pM, lo más preferiblemente 8-12 pM, y lo más preferiblemente aproximadamente 10 pM Y27632.
42. El método de una cualquiera de las realizaciones 36 - 41, en el que la etapa de siembra se lleva a cabo 18 -30 h antes de la etapa (i).
Descripción de las figuras
Figura 1: Optimización de la diferenciación cardíaca temprana para una diferenciación cardíaca robusta y eficiente. (A) Esquema del protocolo de diferenciación cardíaca desarrollado. (B, C) Efecto de la adición de FGF-2 sobre la diferenciación cardíaca en el cultivo 2D. (D, E) Efecto de la variación de la concentración de BMP4 sobre la diferenciación cardíaca en el cultivo 2D mientras CHIR está presente. (F, G) Efecto de la eliminación de cada factor individualmente del protocolo de diferenciación cardíaca 2D, todos los factores se añadieron diariamente desde los días 0 - 3 , excepto IWP4, que fue añadido cada 2 - 3 días desde los días 3 -13. (H, I, J) Análisis de la presencia de tipos de células contaminantes utilizando qPCR. (K) Inmuno-tinción para marcadores de cardiomiocitos. (L) Citometría de flujo para cardiomiocitos (n = 6 experimentos). (M) Inmuno-tinción para marcadores de células estromales. (N) Citometría de flujo para células estromales (n = 6 experimentos). Todos los datos son n = 3 experimentos a menos que se indique otra cosa. Los datos de qPCR (MESP-1, OCT4, s Ox 17 y NEUROD1) se normalizan a GAPDH. * Indica una diferencia estadísticamente significativa (P < 0,05) al usar ANOVA con la prueba post-Hoc de comparación múltiple de Tukey. ** Indica una diferencia estadísticamente significativa entre las muestras suplementadas sin factores. *** Indica una diferencia estadísticamente significativa de las muestras suplementadas sin factores, todos los factores menos BMP4, todos los factores menos ACT-A y todos los factores menos IWP-4.
Figura 2: BHM puede formarse directamente partir de hPSC. (A) BHM a los 22 días de diferenciación. (B) inmunotinción whole-mount. (C) Tensión de contracción isométrica (fuerza de contracción) en respuesta a la variación de la concentración de calcio, n = 7 a partir de 4 experimentos. (D) Perfiles de citometría de flujo de pluripotencia (TRA-1-60/OCT4) y marcadores cardíacos (a-actinina), n = 3 -4 experimentos. (E) Citometría de flujo de marcadores de células estromales en el día 22, n = 3 experimentos. (F) perfiles de expresión qPCR de marcadores para pluripotencia, diferenciación mesodérmica y diferenciación cardíaca; los datos se normalizan a la expresión de GAPDH, n = 3 experimentos. * Indica una diferencia estadísticamente significativa (P < 0,05) en comparación con -1 días usando ANOVA con la prueba post-hoc de comparación múltiple de Tukey.
Figura 3: La optimización de la condición de cultivo BHM revela diferentes parámetros que responden específicamente a diferentes estímulos. (A) La suplementación con ASC-2-P mejora el BHM, la tensión de contracción isométrica (fuerza de contracción) en respuesta a la concentración variable de calcio, n = 8 - 9 de 3 experimentos, * indica una diferencia estadísticamente significativa (P < 0,05) en comparación con el control usando ANOVA de dos vías con pruebas post hoc de Bonferroni. (B) Análisis de citometría de flujo de experimentos de ASC-2-P para marcadores de cardiomiocitos y células estromales, n = 7 - 8 de 3 experimentos. (C) La estimulación mecánica mejora la función de
BHM, la tensión de contracción isométrica (fuerza de contracción) en respuesta a la concentración variable de calcio, n = 9-11 en 4 experimentos, * indica una diferencia estadísticamente significativa (P < 0,05) para ambos regímenes de estimulación mecánica en comparación con el control utilizando pruebas post hoc ANOVA de dos vías de Bonferroni. (D) Dispositivos de estimulación mecánica. (E) Inmuno-tinción whole-mount de BHM bajo control y regímenes de estimulación mecánica. (F) Factores de crecimiento (FGF2: 10 ng/ml y TGFbl: 1 ng/ml) añadidos durante la maduración cardíaca regulan la función de BHM, tensión de contracción isométrica (fuerza de contracción) en respuesta a la concentración variable de calcio, n = 9 - 11 desde 4 experimentos. (G) Análisis de tamaño de células de cardiomiocitos para los experimentos del factor de crecimiento utilizando citometría de flujo, n = 6 de 3 experimentos, * indica una diferencia estadísticamente significativa (P < 0,05) en comparación con el control usando la prueba t de Student. (H) La expresión qPCR de la relación p-MHC/a-MHC para los experimentos de factor de crecimiento, n = 3 - 6 experimentos, * indica una diferencia estadísticamente significativa (P < 0,05) en comparación con el control que utiliza ANOVA con la prueba post hoc de comparación múltiple de Tukey. (I) Expresión de qPCR de ANP y Sk Act para los experimentos del factor de crecimiento, n = 3 experimentos. (J) El ajuste del calcio a 1,2 mmol/L durante la maduración cardíaca mejora la función de BHM, la tensión de contracción isométrica (fuerza de contracción) en respuesta a la concentración variable de calcio, n = 10 - 11 en 4 experimentos, * indica una diferencia estadísticamente significativa (P < 0,05) en comparación con el control utilizando las pruebas post hoc de ANOVA de dos vías de Bonferroni. (K) tensión de reposo de BHM para los experimentos de calcio, n = 10-11 de 4 experimentos. (L) Módulo elástico de BHM para los experimentos de calcio, n = 10-11 de 4 experimentos, * indica una diferencia estadísticamente significativa (P < 0,05) en comparación con el control que utiliza ANOVA con la prueba post hoc de comparación múltiple de Tukey.
Figura 4: El BHM producido utilizando el protocolo optimizado exhibe propiedades de tipo in vivo. (A) BHM puede ser estimulado eléctricamente a varias velocidades. (B) BHM responde con mayor tensión de contracción (fuerza de contracción) al aumento de longitud (mecanismo de Frank-Starling). (C) Comparación de la tensión de contracción (fuerza de contracción) de BHM de 22 días de edad (de datos anteriores) y BHM de 29 - 30 días, n = 11 de 4 experimentos durante 22 días y n = 7 de 2 experimentos de 29 - 30 días. (D) Respuesta inotrópica de BHM de 22 días a la isoprenalina (1 pmol/L) en condiciones estimuladas a 0,6 mM de calcio. (E) Respuesta inotrópica de BHM de 29 -30 días a la isoprenalina (1 pmol/L) bajo condiciones estimuladas a calcio 0,6 mM. (F) Comparación de la respuesta inotrópica a la isoprenalina (1 pmol/L) con la edad, n = 11 de 4 experimentos durante 22 días y n = 7 de 2 experimentos durante 29 - 30 días, * indica una diferencia estadísticamente significativa (p < 0,05) utilizando la prueba t de Student.
Figura 5 : El protocolo BHM se puede usar en todas las líneas de PSC probadas. (A, B, C) Datos de HES3-BHM; (D, E, F) Datos de hIPS-G1-BHM. (A, D) Tensión de contracción isométrica (fuerza de contracción) en respuesta a la concentración variable de calcio, n = 4 para cada línea. (B, E) inmunotinción whole mount. (C, F) Análisis de citometría de flujo de cardiomiocitos y células estromales, n = 3 por línea.
Figura 6: diferenciación cardiaca 2D de líneas múltiples de hPSC. Análisis de citometría de flujo de marcadores de células cardiacas (a-actinina, SIRPA) y estromales (PDGFRa, a-SMA, colágeno I).
Figura 7 : El BHM se puede usar para modelar procesos de desarrollo, utilizando la inhibición de señalización de BMP como ejemplo. (A) Análisis de qPCR de marcadores múltiples en el día 13 de la formación de BHM. (B) Análisis del ciclo celular usando clasificación por citometría de flujo en células a-actinina+. (C) Número de cardiomiocitos por BHM. (D) Tensión de contracción isométrica (fuerza de contracción) en respuesta a concentraciones de calcio variables. * indica una diferencia estadísticamente significativa (P < 0,05) en comparación con el control usando A B) prueba t de Student (n = 3 - 4 ) o D) post test de comparación múltiple ANOVA de dos vías de Sidak.
Figura 8: Resumen de los protocolos utilizados para la diferenciación cardíaca 2D y la formación de BHM. (A) Protocolo utilizado para los experimentos que se muestran en la Figura 1. (B) Protocolo utilizado para los experimentos que se muestran en la Figura 2. (C) Protocolo utilizado para los experimentos que se muestran en la Figura 3 - adición de ácido ascórbico. (D) Protocolo utilizado para los experimentos mostrados en la Figura 3 - estimulación mecánica y factores de crecimiento. (E) Protocolo utilizado para los experimentos que se muestran en la Figura 3: adición de calcio. (F) Protocolo utilizado para los experimentos que se muestran en la Figura 5. (G) Protocolo utilizado para los experimentos que se muestran en la Figura 6. (H) Protocolo utilizado para los experimentos que se muestran en la Figura 7.
Figura 9: Suplemento personalizado para reemplazar B27®. (A) Fuerza contráctil de BHM (hES2) obtenida con B27® o suplemento personalizado (CMS) a 2 mM de calcio extracelular, n = 2/grupo. (B) Número total de CM en BHM hecho con B27® o suplemento personalizado (CMS), n = 2/grupo.
Se entiende que los ejemplos que siguen ilustran más, pero no limitan la invención. Los ejemplos comprenden varias características técnicas, y se apreciará que la invención también se refiere a combinaciones de las características técnicas presentadas en esta sección de ejemplo.
Figura 10: la suplementación del medio de cultivo con TGFp-1 durante la fase de maduración cardíaca mejora la función contráctil de BHM (FOC: fuerza de contracción) de una manera dependiente de la concentración (0,3 - 10 ng/ml probado; n = 5 - 7BHMs/condición).
Ejemplos.
La diferenciación cardiaca requiere la optimización de la inducción del mesodermo cardiaco temprano. Se ha demostrado que las fracciones de células no miocíticas o las células estromales son esenciales para la función de los tejidos cardíacos de ingeniería. Por esta razón, en primer lugar se requirió un protocolo de diferenciación cardiaca que producía de forma consistente cardiomiocitos y fibroblastos/células estromales. Los autores de la presente invención optimizaron su protocolo de diferenciación cardíaca (Fig. 1a) tanto para el rendimiento como para la consistencia, basándose en un protocolo de diferenciación de hPSC 2D libre de suero previamente publicado (Hudson et al. Stem Cells Dev 21, 1513 - 1523 (2012)). Se razonó que la robustez y la eficiencia podrían mejorarse si la actividad de WNT se estabilizaba durante la fase de inducción del mesodermo. Como marcador sustituto para la expresión de MESP1, la inducción del mesodermo se analizó mediante qPCR en el día 3 de cultivo; esto fue seguido por citometría de flujo para a-actinina (marcador de cardiomiocitos) en el día 16, que se encontró que estaban muy bien correlacionados con la cantidad de actividad de latido. Los pasos más importantes para la progresión del protocolo publicado anteriormente al nuevo protocolo se resumen en la Fig. 1.
El protocolo anterior usó la inducción del mesodermo cardíaco con BMP4 y Activina-A durante los primeros 3 días, seguido de una especificación cardíaca con un inhibidor IWP4 de WNT (Hudson et al Stem Cells Dev 21, 1513 -1523 (2012)). De acuerdo con el requisito esencial de FGF2 para la formación temprana de mesodermo en estudios recientes que utilizan hPSCs y desarrollo in vivo, la adición de 5 ng/ml de FGF2 durante los primeros 3 días de diferenciación produjo una tendencia al aumento de la expresión de MESP1 (Fig. 1b) y posteriormente una tendencia a un aumento de a-actinina (Fig. 1c). Sin embargo, se aplicó la señalización WNT que ayudó a mejorar la consistencia y la diferenciación, en consonancia con su papel esencial en la inducción del mesodermo temprano. Para aplicar la señalización WNT se utilizó CHIR99021, un inhibidor de molécula pequeña de GSK3p que induce la señalización WNT incluso en presencia de inhibidores de WNT canónicos. CHIR solo o con los factores de diferenciación utilizados en la Fig. 1b,c no pudo inducir ninguna actividad de latido. Cuando se varió la concentración de BMP4, se encontró una inducción óptima y consistente de MESP1 (Fig. 1d) y la expresión de a-actinina (Fig. 1e) a una concentración de BMP4 de 5 ng/ml. Cada factor del protocolo de diferenciación se eliminó después individualmente para demostrar su requerimiento en la inducción eficiente y consistente de MESP1 (Fig. 1f) y, posteriormente, la expresión de a-actinina (Fig. 1g).
Para formar BHM también es importante que estén presentes las poblaciones de células estromales. Por tanto, se investigó si las células estromales u otros tipos de células potencialmente contaminantes estaban presentes en el protocolo de diferenciación optimizado. Se encontraron niveles muy bajos de poblaciones de células potencialmente contaminantes, analizadas utilizando qPCR para hPSCs (OCT4 también conocida como POU5F1) (Fig. 1 h), endodermo (SOX17) (Fig. 1i), neural (NEUROD1) (Fig. 1j), y mesodermo temprano (MESP1) (datos no mostrados). Se encontró una expresión muy alta de NKX2-5 y ¡5-MHC (también conocida como MYH7) en el cultivo de diferenciación cardíaca de la invención en comparación con las otras condiciones (sin factores o sin IWP4, datos no mostrados). Adicionalmente, se encontró que estaban presentes (Fig. 1m, n) tanto los cardiomiocitos (Fig. 1 k-l) como los diferentes tipos de células estromales, incluyendo: células positivas a actina del músculo liso (a-SMA+), células positivas al colágeno (COLI+), y células a-SMA+ COLI+. En conjunto, estos datos sugieren que el protocolo de diferenciación cardiaca de la invención produce eficientemente cardiomiocitos, siendo el resto de las células predominantemente células estromales, proporcionando así la composición celular requerida para aplicaciones de ingeniería de tejidos.
Formación dirigida de BHM.
Después de la optimización del protocolo de diferenciación cardíaca, se probó la hipótesis de si se podría formar BHM directamente a partir de hPSCs. El nuevo protocolo de diferenciación cardíaca libre de suero (Fig. 1) se utilizó con una etapa de maduración adicional en la que los anillos se retiraron de los moldes y se colocaron en camillas estáticas (+ 10% de la longitud de holgura) en un medio que contenía 5 ng/ml de FGF2 y 200 pM de ascorbato-2-fosfato (ASC-2-P). Se encontró que este protocolo era efectivo en la formación de BHM (Fig. 2a). El BHM comenzó a contraerse espontáneamente en diferentes áreas el día 13 y el día 15-17, las contracciones se hicieron síncronas y rítmicas, y continuaron hasta el análisis el día 22 (datos no mostrados). Los cardiomiocitos en el BHM tenían una morfología alargada y estriada (Fig. 2b) y el BHM podía estimularse eléctricamente y tenía una fuerza de contracción medible con capacidad de respuesta a la concentración de calcio (Fig. 2c).
El desarrollo del BHM siguió las rutas de desarrollo conocidas. Las hPSCs se diferenciaron en gran medida por el día 3, indicado por una disminución en las células TRA-1-60+/OCT4+ y la expresión de OCT4 (Fig. 2d, f) y una expresión concomitante de los marcadores mesodérmicos tempranos MIXL1 y MESP1 (Fig. 2f). En el día 8 hubo una pérdida de la expresión de MIXL1 y MESP1 (Fig. 2f), con un aumento simultáneo en las células a-actinina+ (Fig. 2d) y la expresión de marcadores de células progenitoras de cardiomiocitos (Fig. 2f). Hubo picos en la expresión de múltiples factores de transcripción implicados en la cardiogénesis, que incluyen: TBX5 (pico en el día 13), lSL1 (pico en el día 8) y NKX2-5 (pico en el día 13) (Fig. 2f). Esto vino seguido por la expresión de marcadores cardíacos más maduros a-MHC (también conocido como MYH6), ¡5-MHC, ANP (también conocido como NPPA) y MLC2v (también conocido como MYL2) (Fig. 2f). Curiosamente, la expresión de a-MHC alcanzó su punto máximo en el día 13, seguida por un gran
aumento en ¡3-MHC en el día 22 y, por lo tanto, un aumento de la relación 3-MHC/a-MHC (Fig. 2f). Además, hubo poca expresión de marcadores endodérmicos y neurales en el día 22 (Fig. 2f). En conjunto, estos datos sugieren no solo que el desarrollo de la BHM siguió las rutas de desarrollo conocidas, sino también que la maduración cardíaca se produce según lo indicado por la caída de la expresión del gen progenitor, el aumento de la expresión de ¡3-MHC y la relación de la expresión de ¡3-MHC/a-MHC, y aumento de la expresión de MLC2v (indica la madurez, véase Tiburcy et al. Circ Res 109, 1105- 1114 (2011)). Además, en el día 22, se encontró que el BHM estaba compuesto por 30 ± 6% (n = 4 experimentos) de cardiomiocitos y una gran proporción de células estromales (Fig. 2d). En la Fig. 5 se muestran gráficos representativos de citometría de flujo.
Optimización de la funcionalidad de BHM.
Aunque el protocolo BHM expuesto en la Fig. 2 representa el primer proceso totalmente libre de suero de expansión de hPSC y formación de miocardio de bioingeniería, los inventores plantearon la hipótesis de que la optimización podría generar tejidos con mayor funcionalidad y mayor consistencia. Para estos experimentos se usó la fuerza contráctil (tensión de contracción/fuerza de contracción) como un determinante primario de la función, ya que la tensión de contracción depende de una amplia variedad de propiedades miocárdicas que incluyen: número y fenotipo de los cardiomiocitos, número y fenotipo de fibroblastos, conectividad tisular, composición de ECM, conectividad célula-célula y conectividad ECM-célula. Como factores secundarios, los autores de la presente invención utilizaron, 1) la tensión de reposo, debido a que refleja la función de las células estromales y la biología de la matriz extracelular, 2) el tamaño de los cardiomiocitos, debido a estímulos como los farmacológicos, y 3) la composición de las células (células cardiomiocitos:estroma), que es un determinante importante para el rendimiento contráctil. Los parámetros que cambiaron se muestran en la Fig. 3.
ASC-2-P mejora la funcionalidad de BHM.
El ascorbato (vitamina C) desempeña un papel importante en la síntesis adecuada de colágeno y es un antioxidante. Por ello se planteó la hipótesis de que el ascorbato (en la forma más estable de ASC-2-P) suplementado durante el cultivo BHM temprano, los días 0 -13 (ya se agregó durante los días 13-22 en la Fig. 2), tendría una influencia positiva sobre la funcionalidad de BHM dada la importancia de los colágenos durante el desarrollo. Se encontró que ASC-2-P mejoró significativamente la tensión de contracción BHM/fuerza de contracción (Fig. 3a) e indujo una tendencia al aumento de la fracción de cardiomiocitos sin cambio en la fracción de células estromales (Fig. 3b). También se encontró que la suplementación de ASC-2-P durante todo el protocolo de diferenciación mejoró la diferenciación en el protocolo 2D del autor de la invención aumentando significativamente el número de células sin cambios en la eficiencia de la diferenciación cardíaca (datos no mostrados). Por tanto, en formatos tanto 2D como BHM, ASC-2-P puede haber aumentado la supervivencia celular y/o la proliferación progenitora como se ha propuesto en un estudio reciente (Cao et al. Cell Res 22, 219 - 236 (2012)).
La función de BHM depende del régimen de estimulación mecánica.
A continuación, se evaluó cómo el estiramiento estático y la estimulación mecánica dinámica influyeron en la función de BHM. Los dispositivos utilizados para el estiramiento estático y la estimulación mecánica dinámica se muestran en la Fig. 3d. Tanto el estiramiento estático como la estimulación mecánica dinámica aumentaron significativamente la tensión de contracción/fuerza de contracción del BHM y los regímenes de estimulación mecánica dieron como resultado una tensión de contracción del BHM similar (Fig. 3c). Ambos regímenes de estimulación mecánica mejoraron la morfología de los cardiomiocitos en el BHM, haciendo que se formasen haces de músculos compactos, alargados y estriados (Fig. 3d). La estimulación mecánica dinámica se prefirió al estiramiento estático, porque facilita las contracciones auxotónicas (Zimmermann et al., Nat Med 12, 452 - 458 (2006)).
Las propiedades de los cardiomiocitos dependen de Factores de Crecimiento Exógenos.
Había una tendencia a disminuir la tensión de contracción/fuerza de contracción al añadir FGF2 y una tendencia a aumentar la tensión de contracción al añadir TGFp1 (Fig. 3f). Por lo tanto, en los experimentos previos en los que se agregó FGF2 durante la maduración cardíaca, esto podría haber sido en realidad una función perjudicial de BHM. Se encontró que tanto el FGF2 como el TGFp1 inducían un aumento en el tamaño de los cardiomiocitos (Fig. 3g). La adición de TGFp1 dio como resultado una relación de expresión 3-MHC/a-MHC más madura (corazón humano = 9) (Fig. 3h) mientras que los marcadores de hipertrofia patológica ANP (también conocidos como NPPA) disminuyeron (Fig. 3i). La adición de FGF2 no cambió la relación de expresión 3-MHC/a-MHC (Fig. 3h), sino que indujo (de forma variable) el marcador de hipertrofia patológica ANP (Fig. 3i). En conjunto, esto indica que ambos factores inducen hipertrofia, lo que es consistente con los resultados in vivo. Sin embargo, el FGF2 puede considerarse como un inductor de hipertrofia patológica, mientras que TGFp1 puede considerarse como un inductor de hipertrofia fisiológica.
En otro conjunto de experimentos, los autores de la presente invención investigaron si la suplementación del medio de cultivo con el aumento de TGFp-1 durante la fase de maduración cardíaca tiene influencia en la función contráctil de BHM. Los autores de la presente invención observaron una mejora de la función contráctil de BHM de una manera dependiente de la concentración (Fig. 10).
En los experimentos anteriores, los autores de la presente invención encontraron una gran reducción en las células positivas para la a actina del músculo liso y para el colágeno I en el BHM en comparación con el protocolo 2D (Fig. 1 n vs. Fig. 2e). Esto puede ser un reflejo de diferencias sutiles en la diferenciación de miofibroblastos/fibroblastos en cultivos 2D y BHM. Para detectar más homogéneamente poblaciones similares a cardíacas/fibroblastos en cultivos de diferenciación cardíaca 2D y BHM derivadas de hPSC, se emplearon en subsiguientes experimentos anticuerpos contra CD90 canónica (también conocida como THY1).
El ajuste del calcio extracelular a concentraciones fisiológicas mejoraron la función de BHM.
La concentración de calcio está firmemente regulada en suero humano con concentraciones fisiológicas de calcio de 2,25 - 2,75 mM y 1,0 - 1,2 mM, para el calcio total y el calcio ionizado, respectivamente. Como la concentración de calcio en el medio RPMI es bastante baja (0,42 mM) en comparación con el calcio fisiológico, se evaluó si el ajuste de la concentración de calcio libre mejoró tanto la funcionalidad como la maduración de BHM. El ajuste del calcio a 1,2 mM (utilizando una solución de CaCh 0,2 M) aumentó considerablemente la tensión de contracción del BHM (Fig. 3j). Además, se observó un aumento en la tensión de reposo (Fig. 3k) y el módulo de elasticidad (Fig. 3l). Se ha demostrado que el módulo de elasticidad óptimo mejora el trabajo mecánico realizado por los cardiomiocitos durante las contracciones, que puede aumentar la fuerza de contracción. El mecanismo detrás del aumento del módulo de elasticidad en respuesta al aumento de calcio es actualmente desconocido. Es importante destacar que se debe tener en cuenta que no hubo cambios en las proteínas de manipulción del calcio que se evaluaron utilizando qPCR (CASQ2, PLN, ATP2A2 y RYR2, datos no mostrados, n = 3 experimentos).
BHM producido utilizando el protocolo optimizado muestra propiedades similares a in vivo. BHM contactó espontánea y coherentemente, y pudo ser estimulado eléctricamente a múltiples frecuencias hasta al menos 3 Hz (Fig. 4a), cubriendo el margen de frecuencia de latido observado durante el desarrollo del corazón. BHM también aumentó la tensión de contracción (fuerza de contracción) en respuesta al aumento de la longitud de reposo (y la tensión de reposo) consistente con el mecanismo de Frank-Starling (Fig.4b). Con el aumento del tiempo de cultivo, no se encontró ningún cambio en la tensión de contracción de BHM en comparación con los datos previos, pero se observó un aumento de EC50 de calcio de 0,2 a 0,7 mmol/L (Fig. 4c). El cultivo extendido también dio lugar a un aumento de la respuesta inotrópica a isoprenalina 1 pM, lo que indica una madurez mejorada en comparación con los formatos de cultivo tradicionales donde no se ha observado ningún aumento (Fig. 4d-f). Usando inmunotinción whole-mount, se encontró que el BHM también tiene tanto células estromales como células endoteliales presentes en los haces musculares (datos no mostrados). Es importante destacar que el cultivo de BHM también se podría mantener a largo plazo (por lo menos hasta el día 63) bajo las condiciones de maduración con una mejora observada en la apariencia morfológica (datos no mostrados).
El protocolo BHM no modificado trabaja en múltiples líneas de células madre pluripotentes humanas. A continuación, se demostró que los BHM optimizados (Fig. 5) y 2D (Fig. 6) trabajan en múltiples líneas de hPSC. Para estos análisis se utilizaron las líneas HES2, HES3 y hIPS-G1 (fibroblastos dentales reprogramados utilizando el kit de reprogramación Cytotune libre de vectores). Es importante tener en cuenta que para los protocolos 2D y BHM se encontró que la modificación del número de células de siembra o el uso de un inhibidor de la proteína quinasa asociado a Rho (10 pM, Y-27632) se requería para conseguir densidades celulares similares en todas las líneas después de la fase inicial de siembra de 24 h (datos no mostrados). Cuando se usa el protocolo de siembra requerido para una línea de células en particular, los protocolos 2D y BHM podrían usarse sin modificaciones para esas líneas.
Se encontró que tanto la línea HES3 como la línea hIPS produjeron BHM con una tensión de contracción más baja (Fig. 5a, d) en comparación con la línea HES2 (Fig. 3j). Sin embargo, tanto HES3 como el hIPS BHM tenían morfologías similares (Fig. 5b, e). Las fracciones de cardiomiocitos para el BHM HES3 fueron similares (Fig. 5c) y para BHM hIPS fueron más bajas (Fig. 5f), en comparación con BHM HES2. La disminución de funcionalidad en el b HM de estas líneas es lo más probable a partir de un número de células más bajo en el BHM HES3 (0,50 ± 0,03 x 106 células, n = 3) y BHM hIPS (0,55 ± 0,05 x 106 células, n = 3), en comparación con el BHM HES2 (0,74 ± 0,13 x 106 células, n = 6 de 3 experimentos). Por lo tanto, se debe tener cuidado de excluir las diferencias causadas por los cambios en el número y composición de las células al evaluar diferentes líneas celulares en vez de diferentes tratamientos en la misma línea.
El BHM como modelo de desarrollo revela que se requiere señalización de BPM para la diferenciación terminal de cardiomiocitos humanos.
La inhibición de la señalización de BMP es embrionariamente letal incluso cuando los efectos están restringidos (o al menos parcialmente restringidos) al corazón en desarrollo utilizando CRE impulsado por varios genes (para una revisión, véase Kruithof et al. Differentiation 84, 89-102 (2012)). En estos estudios se han atribuido múltiples procesos a la señalización del BMP, incluidos defectos estructurales, propiedades miocárdicas como la estructura de las trabéculas y el grosor de la pared, y un fenotipo celular que incluye la desregulación de los genes progenitores y la reducción de la transformación epitelial a mesenquimatosa (EMT). Para determinar el efecto de la señalización de BMP meramente en el desarrollo del miocardio sin influencias sistémicas y limitaciones anatómicas, se consideró que el BHM era un buen sistema modelo.
En estos experimentos, se añadieron 2 pmol/L del inhibidor de señalización del receptor de BMP dorsomorfina con cada cambio de medio desde el día 6 en adelante. En el día 13, el BHM tratado con dorsomorfina no logró regular el ISL1 mientras que la expresión de otros marcadores cardíacos más maduros NKX2-5 y a-MHC no se alteraron (Fig. 7a). Cuando se investigaron los genes asociados a EMT, se encontró que no había alteración en la expresión de CDH1, CHD2, SNAIL1 o TGFp2, lo que indica que EMT o los factores que regulan EMT no se modificaron en BHM después de 13 días (Fig. 7a). Utilizando citometría de flujo después de 22 días se encontró que había más cardiomiocitos en el ciclo celular activo en el grupo tratado con dorsomorfina (Fig. 7b). Sin embargo, se encontró que el tratamiento con dorsomorfina no alteraba el número de cardiomiocitos (Fig. 7c). También se encontró que el número total de células por BHM y las fracciones de cardiomiocitos (a-actinina+) y células estromales (CD90+) no cambiaron en el grupo tratado con dorsomorfina (datos no mostrados). A pesar de estas similitudes, hubo una gran reducción en la tensión de contracción/fuerza de contracción al 47% del grupo de control en el BHM tratado con dorsomorfina (Fig. 7d). Es interesante destacar que no hubo ni un cambio en la capacidad de respuesta de BHM a la isoprenalina ni un cambio en el tamaño de los cardiomiocitos en el BHM tratado con dorsomorfina (datos no mostrados).
Como el aumento de la actividad del ciclo celular no condujo a un mayor número de cardiomiocitos y redujo la tensión de contracción, se calculó si la concentración de oxígeno puede limitar el número de cardiomiocitos en el BHM. Para determinar si era este el caso, los perfiles de difusión de oxígeno modelados matemáticamente se basaron en los modelos publicados en la bibliografía y los parámetros en las diferentes condiciones de BHM (número de células, fracción y tamaño de cardiomiocitos). Se encontró que al usar los parámetros para el BHM de control no había región hipóxica, incluso aunque el número de cardiomiocitos aumentaba a 125% (datos no mostrados).
En conjunto, los presentes datos sugieren que la inhibición de BMP usando dorsomorfina produce un aumento del estado proliferativo, un resultado que es consistente con los experimentos in vivo con ratones. Sin embargo, no hubo aumento en el número de cardiomiocitos, lo que indica que hay un aumento de la apoptosis o que los cardiomiocitos son bi-nucleantes. Al margen del mecanismo, la inhibición de la señalización de BMP resultó reproducible en un fenotipo de tejido que produce menos fuerza contráctil (y también menor fuerza por cardiomiocito) y tejido miocárdico inferior.
Suplemento personalizado para reemplazar B27®.
Los BHMs se generaron a partir de hESC no diferenciadas bajo condiciones libres de suero, de acuerdo con el protocolo estándar de BHM. El protocolo estándar incluye suplemento B27®. En este experimento, el suplemento B27® fue reemplazado por un suplemento definido y personalizado (CMS, Tabla 4).
Los resultados muestran que B27® puede ser reemplazado por CMS. La fuerza es similar, también es comparable el número de cardiomiocitos generados en el BHM (Fig. 9).
Conclusión
Usando la diferenciación dirigida de PSCs en hidrogeles de colágeno I, la presente solicitud demuestra que es posible guiar el ensamblaje de BHM bajo condiciones libres de suero. El BHM tiene múltiples aplicaciones, incluyendo estudios farmacológicos, estudios de procesos de desarrollo, procesos de maduración cardíaca y también aplicaciones regenerativas potenciales.
En estos ejemplos, la robustez del protocolo recientemente desarrollado se demostró usando múltiples formatos de cultivo y múltiples líneas para la diferenciación. Sin embargo, se observará que si bien la eficiencia de diferenciación experimento-experimento fue consistente, la eficiencia varió cuando se usaron diferentes lotes de reactivos. Por lo tanto, es prudente establecer un estricto control de calidad de los reactivos con el fin de producir BHM con propiedades consistentes y definidas para aplicaciones terapéuticas tanto in vitro como potenciales.
Métodos.
Cultivo de PSC.
Se obtuvieron células HES2-ROSA26-RFP (Irion et al. Nat Biotechnol 25, 1477 - 1482 (2007)) de Gordon Keller y se obtuvieron células HES3 de Embryonic Stem Cell International (ESI, Singapur). Se generaron hIPS a partir de fibroblastos derivados de biopsia gingival humana utilizando el kit Cytotune Reprogramming (Applied Biosystems) siguiendo las instrucciones del fabricante.
Para la generación de IPS, 6 días después de la transducción viral se pusieron fibroblastos en fibroblastos embrionarios de ratón irradiados, en medio para fibroblastos (DMEM alto en glucosa, 2 mmol/L de glutamina, FBS al 10% (PAA), 100 UI/ml de penicilina, 100 pg/ml de estreptomicina, todos Gibco excepto donde se indique otra cosa). Al día siguiente, el medio se cambió por medio de PSC (DMEM knock-out (Gibco) suplementado con 20% de reemplazo de suero Knock-out (KSR, Gibco), 2 mmol/L de glutamina, 100 UI/ml de penicilina, 100 pg/ml de estreptomicina, 1% de aminoácidos no esenciales (Gibco) y 10 ng/mL de FGF2 (Miltenyi Biotec)). Las colonias iPS
emergentes se recogieron mecánicamente y se expandieron por pases semanales utilizando 1 mg/ml de colagenasa NB6 (Cresent Chemical Company).
Para los experimentos, las hPSCs se adaptaron a una sola célula y se cultivaron en fibroblastos de prepucio humano irradiado (HFF) en medio de PSC con cambios de medio diarios y pases semanales usando tratamiento de TrypLE (Gibco) de 3 minutos (Ellerstrom et al., Stem Cells 25, 1690 - 1696 (2007)). Antes de los experimentos de caracterización o diferenciación, las hPSCs se pusieron en 1:30 Matrigel (Millipore) en placas recubiertas con PBS (Gibco), a 2,5 x 104 células/cm2 para HES2 o 5 x 104 células/cm2 para líneas h ES3 y HIPs , y se cultivaron durante 3 días en medio PSC 1:1 menos FGF-2 y medio acondicionado con HFF (HFF-CM - recogido de cultivos de HFF irradiados confluentes de 5 días) con 10 ng/mL de FGF2. La línea hIPS recibió también 10 pmol/L de Y-27632 (Stemgent). Se recogieron hPSCs para experimentos pasándolos usando un tratamiento con TrypLE de 3 minutos y después se cultivaron en el formato apropiado.
Las líneas de células madre pluripotentes se analizaron regularmente en relación con el micoplasma utilizando un kit de prueba (Lonza) y se caracterizaron utilizando ensayos estándar. Los marcadores de pluripotencia se evaluaron mediante PCR (OCT4, SOX2, KLF4, MYC endógeno), qPCR (OCT4, NANOG, REX1, DNMT3B) e inmunotinción (OCT4, NANOG, TRA-1-60) (Chan y otros. Nat Biotechnol 27, 1033 - 1037 (2009)). La desmetilación del promotor de OCT4 se confirmó mediante secuenciación con bisulfito (Freberg et al. Mol Biol Cell 18, 1543 - 1553 (2007)). Se usó el cariotipado para determinar si existían anomalías genéticas (Campos et al. J Vis Exp 4 (2009)). La pluripotencia se confirmó mediante la formación de teratoma en ratones SCID mediante inyección en el flanco de 4 -6 x 106 células.
Medio de diferenciación.
Para los experimentos de diferenciación, las hPSCs se cultivaron en RPMI 1640 suplementado con 1 mmol/L de piruvato de sodio, 100 UI de penicilina, 100 pg/ml de estreptomicina y 2% de suplemento de B27 (medio SF, todos de Gibco) y diversos factores como se indica. Los factores utilizados en este estudio incluyeron: sal hidrato de ácido L-ascórbico 2 fosfato de sesquimagnesio (Sigma), BMP4 (R&D Systems), activina A (R&D Systems), FGF2 (Miltenyi Biotec), dorsomorfina (Stemgent), CHIR99021 (Stemgent), IWP4 (Stemgent), y TGF l (Peprotech).
Diferenciación cardiaca 2D.
La diferenciación cardíaca se optimizó en la línea HES2. Las líneas hPSCs de HES2 se pusieron en placas a razón de 5 x 104 células/cm2 (1 x 105 células/cm2 para las líneas HES3 y hIPS) en placas recubiertas con Matrigel/PBS 1:30 y se cultivaron en medio 1:1 de PSC menos FGF-2 y medio acondicionado con HFF (HFF-CM - recolectado de cultivos de HFF irradiados confluentes de 5 días) con 10 ng/mL de FGF2. Para la línea hIPS, se añadió Y-27632 10 pM a este medio. Después de 1 día, las células se aclararon con medio RPMI, luego se diferenciaron como se indica en cada figura con 0,5 ml de medio en cada pocillo de una placa de 24 pocillos. Los detalles del protocolo para cada figura se resumen en la Fig. 8.
Formación de BHM.
La formación de BHM fue optimizada en la línea HES2. Las hPSCs de HES2 se suspendieron 1:1 en medio PSC menos FGF-2 y medio acondicionado con HFF (HFF-CM - recogido de cultivos de HFF irradiados con confluencia de 5 días) con 10 ng/ml de FGF2 y se mezclaron con un hidrogel de colágeno I. Para HES3 y líneas HIPS se añadió también al medio Y-27632 10 pM. La matriz de colágeno I se formuló con colágeno I bovino solubilizado en ácido (Devro) con un volumen equivalente de 2X DMEM (Gibco) y se neutralizó utilizando hidróxido de sodio 0,1 M. La matriz de hPSC/colágeno I se formuló para dar una concentración final de colágeno I de 1 mg/ml y 5 x 105 hPSC por 170 pl. Para las líneas HES3 y hIPS, se utilizaron 1 x 106 y 0,5 x 106 células, respectivamente, por 170 pl. Para cada BHM, se pipetearon 170 pl de la matriz hPSC/colágeno I en moldes circulares (d. int. = 4 mm, d. ext. = 10 mm) fabricados usando poli(dimetilsiloxano) (Sylgard, Dow Corning). Después de 10 minutos de cultivo en la incubadora a 37 °C, se añadió el colágeno gelificado y 1,25 ml de medio acondicionado con fibroblastos de prepucio humano 1:1 con 10 ng/ml de FGF2 por BHM. Al día siguiente, los BHMs se aclararon con medio RPMI y luego se diferenciaron como se indica en cada figura, con 1,25 ml de medio por BHM. En el día 13, los BHM se transfirieron a estimuladores mecánicos como se indica. Los detalles del protocolo para cada figura se resumen en la Fig. 8.
Desasociación celular.
Los cultivos 2D se desasociaron mediante un aclarado con PBS seguido de incubación durante 1 h en 1 mg/ml de colagenasa tipo I (Sigma) con suero bovino fetal al 20% (FBS, Applied Bio-systems) en PBS. Las células se recogieron después en un tubo, se aclararon con PBS y se incubaron con 0,25% de tripsina-EDTA (Applied Biosystems) durante 5 min, seguido por el aclarado con un medio que contenía FBS.
Para el protocolo de digestión de BHM inicial, los BHM se desasociaron en 0,025 mg/ml de Liberase TM (Roche), 2,3-butanodiona monoxima 30 mM a 37 °C durante 60 minutos en PBS. Para preservar los marcadores de la superficie celular, el BHM se desasoció utilizando el mismo protocolo que para la digestión 2D.
PCR cuantitativa (qPCR).
Se recogieron células, BHMs o biopsias de corazón humano y se almacenaron a -80 °C hasta la extracción de ARN utilizando Trizol siguiendo las instrucciones del fabricante (Applied Biosystems). Luego se trató 1 |jg de ARN con ADNasa (Roche), seguido de síntesis de ADNc utilizando un kit de transcripción inversa de ADNc de alta capacidad (Applied Biosystems).
La qPCR se realizó utilizando Fast SYBR Green Master Mix (Applied Biosystems) en un formato de 384 pocillos AB7900 HT (Applied Biosystems). La expresión génica se normalizó utilizando 2-ñCt o 2'ññCt utilizando GAPDH como gen de mantenimiento, que se encontró que se expresaba consistentemente entre las condiciones en todos los experimentos de los autores de la presente invención. Los detalles básicos se dan en la Tabla 1 que sigue.
Tabla 1



man, et al. Cell Stem Cell 8, 228 - 240 (2011) goe, et al. Circulation 106, 1333 - 1341 (2002). , et al. Nature 451, 141 - 146 (2008).
erg, et al. Mol Biol Cell 18, 1543 - 1553 (2007).
Inmunotinción.
Se pusieron células de diferenciación cardiaca digeridas en placas sobre cubreobjetos de vidrio recubiertas con gelatina al 0,1% durante 24 h en FBS al 20% (Gibco) en RPMI 1640 suplementado con 1 mmol/L de piruvato de sodio, 100 Ul/ml de penicilina y 100 |jg/ml de estreptomicina. Las células se fijaron luego en Histofix (Roti) durante 10 minutos a temperatura ambiente. Las células se bloquearon después durante 30 minutos en FBS al 5%, albúmina de suero bovino al 1% (Sigma) y Triton X-100 al 0,5% (Sigma) en PBS (tampón de bloqueo). Luego se tiñeron las células con anticuerpos primarios en el tampón de bloqueo durante 90 minutos, seguido de los anticuerpos secundarios en el tampón de bloqueo y Hoechst durante 60 minutos a temperatura ambiente (Tabla 2). Se obtuvieron imágenes de las células teñidas utilizando un microscopio confocal Zeiss 710.
Tabla 2 Anticuerpos y tinciones (FC - Citometría de flujo, IF - Inmunofluorescencia).

Immuno-tinción Whole-Mount.
Los BHMs se fijaron en Histofix durante 2 - 4 horas a 4 °C. Los BHMs se tiñeron con anticuerpos primarios durante 2 - 3 días, seguidos de anticuerpos secundarios y Phalloidin 546/Hoechst durante 2 -3 días a 4 °C (Tabla 2). Se obtuvo la imagen de los BHMs teñidos con un microscopio confocal Zeiss 710.
Citometría de flujo.
Las células se tiñeron en vivo o fijadas usando Histofix durante 10 minutos a temperatura ambiente o en etanol. Las células se tiñeron en PBS al 5% en FBS (tampón de bloqueo de membrana) para marcadores de superficie celular (excluyendo TRA-1-60) y tampón de bloqueo para marcadores internos. Luego, las células se tiñeron con anticuerpos primarios en un tampón de bloqueo durante 45 min, seguido por anticuerpos secundarios en un tampón de bloqueo y
Hoechst durante 30 min a 4 °C (Tabla 2). Se utilizó un BD LSRII para el análisis de citometría de flujo (BD Biosystems). Las poblaciones de células vivas se clasificaron basándose en los perfiles de dispersión frontal y lateral; se dividieron las poblaciones de células fijadas basándose en la tinción de Hoechst. El software BD FACSDiva (BD Bioscience) o Cyflologic v1.2.1 (Cyflo Ltd) se usaron para el análisis.
Se realizaron experimentos de contracción en baños de órganos a 37 °C y burbujeo continuo con 5% de CO2/95% de O2 para mantener un pH fisiológico en la solución de Tyrode que contiene (todo en mM): 120 NaCl, 1 MgCh, 0,2 CaCl2, 5,4 KCl, 22,6 NaHCOa, 4,2 NaH2PO4, 5,6 glucosa y 0,56 ascorbato. El calcio se ajustó utilizando una solución de cloruro de calcio 0,2 M. Todos los BHM se analizaron primero a 3 Hz con pulsos cuadrados de 5 ms de 200 mA para latir el ritmo de aproximadamente la frecuencia cardíaca embrionaria. Los BHM se estiraron mecánicamente a intervalos de 125 pm hasta Lmax, es decir, la longitud del tejido era la tensión de contracción/fuerza de contracción máxima se registró en presencia de concentraciones de calcio máximas activas inotrópicamente (2 mmol/L; mecanismo de Frank-Starling). Posteriormente, se sometieron BHM a diferentes concentraciones de calcio (0,2, 0,4, 0,8, 1,2, 1,6, 2,0, 2,4 mM) y se registró la fuerza de contracción. Para los experimentos con isoprenalina, la concentración de calcio se ajustó a 0,6 mM y, posteriormente, la concentración de isoprenalina se ajustó a 1 pM. Perfil de difusión de oxígeno.
El perfil de difusión de oxígeno se generó usando el análisis numérico de una aproximación en estado pseudoestacionario de la difusión del cilindro con la dependencia del consumo de oxígeno en la concentración (Ec.
1). Se usaron los parámetros de la bibliografía (Brown et al. Biotechnol Bioeng 97, 962 - 075 (2007)) y como se determinó en experimentos previos (Tabla 3). El análisis numérico y las representaciones gráficas se realizaron utilizando MATLAb V12 (Mathworks) utilizando el solucionador bvp4c y la opción Singular Term.
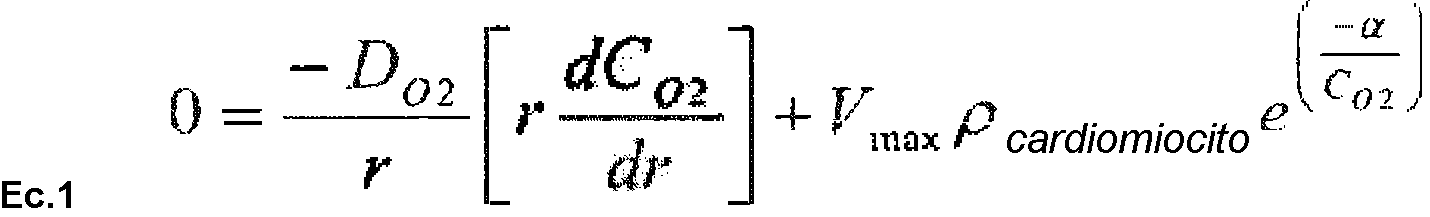
Co2 - concentración de oxígeno en función de la posición radial, r - posición radial en el cilindro, Do2 - constante de difusión de oxígeno, Vmax - velocidad de generación de oxígeno máxima por los cardiomiocitos, pcardiomiocito - densidad de cardiomiocitos, a - constante para la dependencia de la velocidad de generación de oxígeno en la concentración de oxígeno.
Tabla 3: Parámetros del modelo de difusión de oxígeno.

5. Brown, et al. Biotechnol Bioeng 97, 962 - 975 (2007).
Análisis estadístico.
Todos los datos se presentan como media ± s. e. m. Se usaron análisis estadísticos apropiados para cada conjunto de datos como se indica en las leyendas de las Figuras usando Graph Pad Prism o Microsoft Excell.
Suplemento personalizado para reemplazar B27®.
Tabla 4: Suplemento personalizado (CMS) para reemplazar B27.

Preparar 25x en agua apta para cultivo de células.
LISTA DE REFERENCIAS
Kehat, I. et al. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J Clin Invest 108, 407 -414 (2001).
Takahashi, K. et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131, 861 -872 (2007).
Zhang, J. et al. Functional cardiomyocytes derived from human induced pluripotent stem cells. Ore Res 104, e30 - 41 (2009).
Thomson, J.A. et al. Embryonic stem cell lines derived from human blastocysts. Sci- ence 282, 1145 - 1147 (1998).
Lian, X., Zhang, J., Zhu, K., Kamp, TJ. & Palecek, S.P. Insulin Inhibits Cardiac Mesoderm, not Mesendoderm, formation during Cardiac Differentiation of Human Pluripotent Stem Cells and Modulation of Canonical Wnt Signaling Can Rescue this Inhibition. Stem Cells 31{3), 447 - 457 (2013).
Schaaf, S. et al. Human engineered heart tissue as a versatile tool in basic research and preclinical toxicology. PLoS ONE 6, 20 (2011).
Carvajal-Vergara, X. et al. Patient-specific induced pluripotent stem-cell-derived models of LEOPARD syndrome. Nature 465, 808-812 (2010).
Itzhaki, I. et al. Modelling the long QT syndrome with induced pluripotent stem cells. Nature 471, 225 - 229 (2011).
Malan, D., Friedrichs, S., Fleischmann, B.K. & Sasse, P. Cardiomyocytes obtained from induced pluripotent stem cells with long-QT syndrome 3 recapitulate typical disease-specific features In Vitro. Ore Res 109(8), 841-847 (2011).
Moretti, A. et al. Patient-specific induced p!uri potent stem-cell models for long-QT syndrome. N Engl J Med363, 1397 - 1409 (2010).
Yazawa, M. et al. Using induced pluripotent stem cells to investigate cardiac phenotypes in Timothy syndrome. Nature 471, 230-234 (2010).
Bird, S.D. et al. The human adult cardiomyocyte phenotype. Cardlovasc Res 58, 423 - 434 (2003).
Eschenhagen, T. et al. Three-dimensional reconstitution of embryonic cardiomyocytes in a collagen matrix: a new heart muscle model system. FASEB J 11,683 - 694 (1997).
Zimmermann, W.H. et al. Tissue engineering of a differentiated cardiac muscle construct. Ore Res 90, 223-230 (2002). Tulloch, N.L et al. Growth of engineered human myocardium with mechanical loading and vascular coculture. Ore Res 109, 47 - 59 (2011).
Tiburcy, M. et al. Terminal differentiation, advanced organotypic maturation, and modeling of hypertrophic growth in engineered heart tissue. Ore Res 109, 1105- 1114 (2011).
Eschenhagen, T., Eder, A., Vollert, I. & Hansen, A. Physiological aspects of cardiac tissue engineering. Am J Physiol Heart Circ Physiol 303, 11 (2012).
Hudson, J.E., Brooke, G., Blair, C, Wolvetang, E. & Cooper-White, J.J. Development of myocardial constructs using modulus-matched acrylated polypropylene glycol triol substrate and different nonmyocyte cell populations. Tissue Eng Part A 17, 2279- 2289 (2011).
Hudson, J., Titmarsh, D., Hidalgo, A., Wolvetang, E. & Cooper-White, J. Primitive cardiac cells from human embryonic stem cells. Stem Cells Dev 21, 1513- 1523 (2012).
Cao, N. et al. Ascorbic acid enhances the cardiac differentiation of induced pluripotent stem cells through promoting the proliferation of cardiac progenitor cells. Cell Res 22, 219 - 236 (2012).
Kruithof, B.P., Duim, S.N., Moerkamp, AT. & Goumans, M.J. TGFbeta and BMP signaling in cardiac cushion formation: lessons from mice and chicken. Differentiation 84, 89- 102 (2012).
Brown, D.A. et al. Analysis of oxygen transport in a diffusion-limited model of engineered heart tissue. Biotechnol Bioeng 97, 962 - 975 (2007).
Ellerstrom, C, Strehl, R., Noaksson, K., Hyllner, J. & Semb, H. Facilitated expansion of human embryonic stem cells by single-cell enzymatic dissociation. Stem Cells 25, 1690 - 1696 (2007).
Chan, E.M. et al. Live cell imaging distinguishes bona fide human iPS cells from partially reprogrammed cells. Nat Biotechnol 27, 1033 - 1037 (2009).
Freberg, C.T., Dahl, J.A., Timoskainen, S. & Collas, P. Epigenetic reprogramming of OCT4 and NANOG regulatory regions by embryonal carcinoma cell extract. Mol Biol Cell 18, 1543 - 1553 (2007).
Campos, P.B., Sartore, R.C., Abdalla, S.N. & Rehen, S.K. Chromosomal spread preparation of human embryonic stem cells for karyotyping. J Vis Exp 4 (2009).
Kattman, S.J. et al. Stage-specific optimization of activin/nodal and BMP signaling promotes cardiac differentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell, 8, 228 - 240 (2011).
Neagoe, C. et al. Titin isofomn switch in ischemic human heart disease. Circulation 106, 1333 - 1341 (2002).
Park, I.H. et al. Reprogramming of human somatic cells to pluripotency with defined factors. Nature 451, 141 - 146 (2008).
Thomson, J.A., et al. Embryonic stem cell lines derived from human blastocysts. Science 282, 1145 - 1147 (1998).
Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663 - 676 (2006).
Didié et al. Parthenogenetic stem cells for tissue-engineered heart repair. J Clin Invest. 123, 1285 - 1298 (2013).
Irion, S et al. Identification and targeting of the ROSA26 locus in human embryonic stem cells. Nat Biotechnol 25, 1477 - 1482 (2007)
Naito, H. et al. Optimizing engineered heart tissue for therapeutic applications as surrogate heart muscle. Circulation 114, 172-78 (2006).
Zimmermann W.H. et al. Three-dimensional engineered heart tissue from neonatal rat cardiac myocytes. Biotechnol Bioeng 68, 106- 114 (2000).
Zimmermann W.H. et al. Engineered heart tissue grafts improve systolic and diastolic function in infarcted rat hearts. Nat Med 12, 452-458 (2006).
Claims (12)
1. Un método para producir músculo cardíaco de bioingeniería a partir de células madre pluripotentes, que comprende las etapas de
(i) cultivar células madre pluripotentes en un medio basal que comprende una cantidad efectiva de (a) BMP4, Activina A, FGF2, un inhibidor de GSK3, y (b) un suplemento libre de suero que tiene como resultado una concentración final de 0,5-50 mg/ml de albúmina, 1 -100 pg/ml de transferrina, 0,1 -10 pg/ml de etanolamina, 0,003 - 0,3 pg/ml de selenito sódico, 0,4 - 40 pg/ml de L-carnitina HCl, 0 ,1 -10 pg/ml de hidrocortisona, 0,05 - 5 pl/ml de suplemento de ácido graso, 0,0001 - 0,1 pg/ml de triyodo-L-tironina (T3), induciendo así la diferenciación mesodérmica de dichas células madre pluripotentes;
(ii) cultivar las células obtenidas en la etapa (i) en un medio basal que comprende una cantidad eficaz de un inhibidor de la ruta de señalización Wnt y un suplemento libre de suero como se define en (i), induciendo así la diferenciación cardíaca de las células; y
(iii) cultivar las células obtenidas en la etapa (ii) en un medio basal que comprende una cantidad eficaz de un suplemento libre de suero como en (i), bajo estimulación mecánica, promoviendo así la maduración cardíaca.
2. El método según la reivindicación 1, en donde las células madre pluripotentes se seleccionan entre células madre embrionarias, células madre pluripotentes inducidas, y células madre partenogenéticas; y/o en donde las células madre pluripotentes son células madre pluripotentes de origen primate, preferiblemente células madre pluripotentes humanas.
3. El método según una cualquiera de las reivindicaciones 1 -2 , en donde el medio basal en la etapa (i) comprende
1 - 20 ng/ml de BMP4, preferiblemente aproximadamente 5 ng/ml de BMP4; y
0,1 -10 ng/ml de FGF2, preferiblemente aproximadamente 5 ng/ml de FGF2; y
1 - 20 ng/ml de Activina A, preferiblemente aproximadamente 9 ng/ml de Activina A.
4. El método según una cualquiera de las reivindicaciones 1 -3 , en donde el inhibidor de GSK3 en el medio basal de la etapa (i) se selecciona entre el grupo que consiste en CHIR99021, CHIR98014, SB216763, TWS119, Tideglusib, SB415286 y LY2090314; preferiblemente en donde el medio basal de la etapa (i) comprende 0,1 -10 pM CHIR99021.
5. El método según la reivindicación 1, en donde el inhibidor de la ruta de señalización Wnt en el medio basal de la etapa (ii) se selecciona entre el grupo que consiste en IWP4, IWP2, IWR-1, IWP1, IWP3, IWR-2, IWR-3, IWR-4, IWR-5. XAV939, DKK1, quercetina, ICG-001, pirvinio, CCT031374, iCRT-3,5,14, CPG049090, NC043; preferiblemente en donde el medio basal de la etapa (ii) comprende 0,1 -10 pM IWP4.
6. El método según una cualquiera de las reivindicaciones 1 - 5, en donde el medio basal de la etapa (iii)
a) comprende además 0,1 -10 ng/ml de TGFp1, lo más preferiblemente aproximadamente 1 ng/ml de TGFp1;
y/o
b) no comprende una cantidad efectiva de FGF2; y/o
c) comprende 0,5 - 3 mM de Ca2+, lo más preferiblemente 1,2 mM Ca2+.
7. El método según una cualquiera de las reivindicaciones 1 - 6, en donde la estimulación mecánica en la etapa (iii) es estimulación mecánica dinámica o estiramiento estático, preferiblemente en donde la estimulación mecánica en la etapa (iii) es estimulación mecánica dinámica.
8. El método según una cualquiera de las reivindicaciones 1 - 7, en donde l medio basal de la etapa (i), (ii) y/o (iii) comprende comprende 10 - 1000 pM, preferiblemente 50 - 400 pM, más preferiblemente 100 - 300 pM, incluso más preferiblemente 150 - 250 pM, y lo más preferiblemente aproximadamente 200 pM de ácido ascórbico o una sal del mismo.
9. El método según una cualquiera de las reivindicaciones 1 - 8, en donde el suplemento libre de suero en la etapa (i), (ii) y/o (iii) comprende 0,1 - 10% B27 o B27 menos insulina, preferiblemente 0,5 - 8%, más preferiblemente 1 - 6%, incluso más preferiblemente 1,5 - 4%, y lo más preferiblemente aproximadamente 2% de B27 o B27 menos insulina.
10. El método según una cualquiera de las reivindicaciones 1 - 9, en donde el medio basal usado en la etapa (i), (ii) y/o (iii) es DMEM/F12, StemPro, medio de Iscove, aMEM, DMEM y RPMI, preferiblemente en donde el medio basal usado en la etapa (i), (ii) y/o (iii) es RPMI suplementado con piruvato.
11. El método según la reivindicación 1, que comprende antes de la etapa (i) una etapa de siembra, en donde dichas células madre pluripotentes se siembran en una relación de (2,5 - 6 x 106 células/1 mg de colágeno)/1 ml de medio en un molde adecuado; preferiblemente en donde la etapa de siembra se lleva a cabo 18 -30 h antes de la etapa (i).
12. El método según la reivindicación 11, en donde el medio usado en la etapa de siembre comprende además un inhibidor de ROCK, preferiblemente seleccionado entre Y27632, H-1152P, Tiazovivina, Fasudil, Hidroxifasudil, GSK429286A, y RKI-1447, más preferiblemente en donde el medio usado en la etapa de siembra comprende 1 - 50 |JM Y27632.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP13185344 | 2013-09-20 | ||
| PCT/EP2014/069951 WO2015040142A1 (en) | 2013-09-20 | 2014-09-19 | A method to direct differentiation of pluripotent stem cells into functional heart muscle |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2741806T3 true ES2741806T3 (es) | 2020-02-12 |
Family
ID=49261445
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES14767019T Active ES2741806T3 (es) | 2013-09-20 | 2014-09-19 | Un método para la diferenciación directa de células madre pluripotentes en músculo cardíaco funcional |
Country Status (10)
| Country | Link |
|---|---|
| US (3) | US10329532B2 (es) |
| EP (2) | EP3575393A1 (es) |
| JP (2) | JP6681825B2 (es) |
| CN (2) | CN105849253B (es) |
| AU (1) | AU2014323098B2 (es) |
| CA (1) | CA2924816C (es) |
| DK (1) | DK3047019T3 (es) |
| ES (1) | ES2741806T3 (es) |
| SG (3) | SG11201601798RA (es) |
| WO (1) | WO2015040142A1 (es) |
Families Citing this family (44)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2015261380B2 (en) | 2014-05-16 | 2021-04-15 | Koninklijke Nederlandse Akademie Van Wetenschappen | Improved culture method for organoids |
| HUE057135T2 (hu) * | 2015-09-01 | 2022-04-28 | Ncardia B V | In vitro módszer egy humán pluripotens õssejtpopuláció kardiomiocita sejtpopulációvá történõ differenciálására |
| KR101781693B1 (ko) | 2015-09-24 | 2017-09-26 | 서울대학교병원 | 심장내막 유래 성체줄기세포로부터 제조된 유도만능 줄기세포의 심혈관계 세포로의 분화방법 및 이의 용도 |
| GB201603569D0 (en) | 2016-03-01 | 2016-04-13 | Koninklijke Nederlandse Akademie Van Wetenschappen | Improved differentiation method |
| US20190100725A1 (en) * | 2016-03-18 | 2019-04-04 | Institute Of Biophysics, Chinese Academy Of Sciences | Cell culture medium and culture medium supplement |
| CN106244526B (zh) * | 2016-08-23 | 2020-09-11 | 北京赛贝生物技术有限公司 | 心肌分化试剂盒及心肌细胞的培养方法 |
| JP7262385B2 (ja) | 2016-08-26 | 2023-04-21 | ザ・カウンシル・オブ・ザ・クィーンズランド・インスティテュート・オブ・メディカル・リサーチ | 心筋細胞の成熟 |
| CN106244527B (zh) * | 2016-08-29 | 2019-10-08 | 广东依浦赛生物科技有限公司 | 人源iPS干细胞体外定向分化为心肌细胞的试剂盒及方法 |
| SG10202105977WA (en) * | 2016-12-04 | 2021-07-29 | Univ Health Network | Generating atrial and ventricular cardiomyocyte lineages from human pluripotent stem cells |
| GB201621594D0 (en) * | 2016-12-19 | 2017-02-01 | Univ Leuven Kath | Novel cell based methods for healing of bone, cartilage and joint disorders |
| JP7162537B2 (ja) * | 2017-01-26 | 2022-10-28 | 国立大学法人大阪大学 | 幹細胞の中胚葉系細胞への分化誘導用培地および中胚葉系細胞の製造方法 |
| CN106867961A (zh) * | 2017-03-27 | 2017-06-20 | 广州润虹医药科技有限公司 | 一种诱导多能干细胞形成中胚层细胞的诱导培养基及诱导方法 |
| US20190185816A1 (en) * | 2017-12-19 | 2019-06-20 | The Regents Of The University Of Michigan | Cardiac microtissue and uses thereof |
| GB201721615D0 (en) | 2017-12-21 | 2018-02-07 | Koninklijke Nederlandse Akademie Van Wetenschappen | Immune cell organoid co-cultures |
| CN108456654A (zh) * | 2018-02-22 | 2018-08-28 | 上海产业技术研究院 | 用于促进干细胞来源心肌细胞成熟的无血清培养体系 |
| EP3540046A1 (en) * | 2018-03-13 | 2019-09-18 | Medizinische Hochschule Hannover | Process for producing cardiac organoids |
| US12331318B2 (en) | 2018-07-13 | 2025-06-17 | Lothar Germeroth | Non-immunogenic engineered tissue and methods of producing and using the same |
| CN111214703B (zh) * | 2018-11-25 | 2021-06-15 | 中国科学院大连化学物理研究所 | 一种iPS来源心肌细胞复合补片及其制备和应用 |
| GB201819224D0 (en) | 2018-11-26 | 2019-01-09 | Koninklijke Nederlandse Akademie Van Wetenschappen | Hepatocyte expansion methods |
| CN109619087A (zh) * | 2018-11-27 | 2019-04-16 | 山东奥博森生物药业股份有限公司 | 一种小肠黏膜干细胞冻存液及冻存方法 |
| KR102639023B1 (ko) | 2019-02-21 | 2024-02-20 | 스템바이오시스, 인크. | 양수 세포-유래 ecm 상에서 심근세포 성숙을 위한 방법, 세포 구조, 및 약물 화합물의 심장 독성 및 전부정맥 스크리닝을 위한 용도 |
| KR20200102329A (ko) * | 2019-02-21 | 2020-08-31 | 고려대학교 산학협력단 | TGFβ1을 이용한 줄기세포를 성숙한 유형별 심근세포로 분화시키는 방법 |
| CN109749986B (zh) * | 2019-03-13 | 2021-04-02 | 武汉大学 | 一种由人多能干细胞分化获得胰腺前体细胞及胰岛β细胞的方法 |
| WO2020232436A1 (en) | 2019-05-16 | 2020-11-19 | Brown University | A human in vitro cardiotoxicity model |
| EP3996762B1 (en) | 2019-07-12 | 2023-11-08 | Genome Biologics UG | Method for generating cardiac tissue mimetics |
| CN110511904B (zh) * | 2019-08-21 | 2021-03-19 | 云南中科灵长类生物医学重点实验室 | 多能干细胞的培养基、培养方法及其应用 |
| DE102019127604A1 (de) * | 2019-10-14 | 2021-04-15 | Georg-August-Universität Göttingen Stiftung Öffentlichen Rechts, Universitätsmedizin | Herstellung von Skelettmuskelzellen und Skelettmuskelgewebe aus pluripotenten Stammzellen |
| EP3835403A1 (en) | 2019-12-11 | 2021-06-16 | Sartorius Stedim Biotech GmbH | Automated medium exchange strategy for suspension cells |
| KR102291265B1 (ko) | 2020-03-19 | 2021-08-18 | 연세대학교 산학협력단 | 심장 오가노이드, 이의 제조 방법 및 이를 이용한 약물 독성 평가 방법 |
| EP4129282A4 (en) * | 2020-03-31 | 2024-05-01 | Natural Medicine Institute Of Zhejiang Yangshengtang Co., Ltd. | PHARMACEUTICAL ASSOCIATION AND ITS USE |
| EP3945133A1 (en) | 2020-07-29 | 2022-02-02 | Georg-August-Universität Göttingen | Mass production of human pluripotent stem cell derived cardiac stromal cell |
| US12504423B2 (en) | 2020-10-16 | 2025-12-23 | Brown University | Human in vitro cardiotoxicity model |
| DK3988138T3 (da) | 2020-10-22 | 2024-02-26 | Georg August Univ Goettingen Stiftung Oeffentlichen Rechts Univsmedizin | Flerlaget kunstig hjertemuskel |
| CA3200169A1 (en) | 2021-01-21 | 2022-07-28 | Luis Haupt | Method of changing culture medium of a culture using spinfilters |
| CN116888470A (zh) | 2021-01-21 | 2023-10-13 | 赛多利斯斯泰帝生物技术有限公司 | 介电常数测量探针在包含细胞聚集体的悬浮培养聚集体中的应用 |
| CN113249310B (zh) * | 2021-05-17 | 2022-10-11 | 湖北大学 | 一种拓展性多能干细胞诱导分化为心肌细胞的方法及应用 |
| CN113337459B (zh) * | 2021-06-02 | 2022-09-06 | 呈诺再生医学科技(珠海横琴新区)有限公司 | 一种提高多能干细胞分化效能的方法 |
| US12076437B2 (en) | 2021-07-12 | 2024-09-03 | Brown University | Proangiogenic protein cocktails delivered in custom biomaterials to revascularize ischemic tissue |
| US20230147883A1 (en) * | 2021-11-28 | 2023-05-11 | Fatemeh Etezadi | Method for prodcing cardiomyocytes from induced pluripotent stem cells in an integrated process |
| AU2023302579A1 (en) * | 2022-07-05 | 2025-02-06 | Repairon Gmbh | Differentiation of ipscs in bioreactors |
| EP4386384A1 (en) | 2022-12-15 | 2024-06-19 | Georg-August-Universität Göttingen Stiftung Öffentlichen Rechts, Universitätsmedizin | Potency assay for pharmaceutical products |
| WO2024248017A1 (ja) | 2023-05-29 | 2024-12-05 | 国立研究開発法人理化学研究所 | 心筋細胞を成熟させる方法及び成熟した心筋細胞の製造方法 |
| CN116942798A (zh) * | 2023-08-01 | 2023-10-27 | 湖北大学 | 重组人il-37蛋白在提高人心肌组织功能中的应用 |
| WO2025040692A1 (en) | 2023-08-22 | 2025-02-27 | Fluosphera Sa | Color- and position-based identification of multicellular structures for the multiplexed and individualized analysis of multi-tissue co-cultures |
Family Cites Families (14)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20040014209A1 (en) | 2002-01-23 | 2004-01-22 | Lassar Andrew B. | Compositions and methods for modulating cell differentiation |
| EP1783206A1 (en) * | 2005-11-08 | 2007-05-09 | Universitätsklinikum Hamburg-Eppendorf | Multiring engineered heart tissue construct |
| JP2008307007A (ja) | 2007-06-15 | 2008-12-25 | Bayer Schering Pharma Ag | 出生後のヒト組織由来未分化幹細胞から誘導したヒト多能性幹細胞 |
| EP2198012B1 (en) * | 2007-07-18 | 2014-12-31 | Cellartis AB | Cardiomyocyte-like cell clusters derived from hbs cells |
| EP2100954A1 (en) * | 2008-03-10 | 2009-09-16 | Assistance Publique - Hopitaux de Paris | Method for generating primate cardiac progenitor cells for clinical use from primate embryonic stem cells, and their applications |
| EP2318520B1 (en) | 2008-07-25 | 2017-10-11 | The University Of Georgia Research Foundation, Inc. | Compositions for mesoderm derived isl1+ multipotent cells (imps), epicardial progenitor cells (epcs) and multipotent cxcr4+cd56+ cells (c56cs) and methods of use |
| US8765117B2 (en) | 2009-06-10 | 2014-07-01 | The General Hospital Corporation | Generation of vascularized human heart tissue and uses thereof |
| CN101935635A (zh) * | 2010-07-30 | 2011-01-05 | 中国人民解放军第二军医大学 | 人骨髓多能干细胞的体外扩增和向多巴胺能神经元定向分化的方法 |
| US9023340B2 (en) * | 2011-07-02 | 2015-05-05 | Chemregen, Inc. | Wnt inhibitors for human stem cell differentiation |
| WO2013013206A1 (en) * | 2011-07-21 | 2013-01-24 | The Board Of Trustees Of The Leland Stanford Junior University | Cardiomyocytes from induced pluripotent stem cells from patients and methods of use |
| US8951798B2 (en) * | 2011-10-13 | 2015-02-10 | Wisconsin Alumni Research Foundation | Generation of cardiomyocytes from human pluripotent stem cells |
| WO2013063305A2 (en) * | 2011-10-25 | 2013-05-02 | Mount Sinai School Of Medicine | Directed cardiomyocyte differentiation of stem cells |
| KR101996343B1 (ko) | 2012-01-27 | 2019-07-05 | 고쿠리츠 다이가쿠 호진 교토 다이가쿠 | 전능성 줄기세포의 심근분화 유도방법 |
| DK2840132T3 (en) * | 2013-08-22 | 2017-02-20 | Georg-August-Universität Göttingen Stiftung Öffenlichen Rechts Universitätsmedizin | Method of Manufacturing Constructed Heart Muscle (EHM) |
-
2014
- 2014-09-19 SG SG11201601798RA patent/SG11201601798RA/en unknown
- 2014-09-19 SG SG10201802167TA patent/SG10201802167TA/en unknown
- 2014-09-19 EP EP19183577.6A patent/EP3575393A1/en not_active Withdrawn
- 2014-09-19 AU AU2014323098A patent/AU2014323098B2/en active Active
- 2014-09-19 WO PCT/EP2014/069951 patent/WO2015040142A1/en not_active Ceased
- 2014-09-19 SG SG10201910753RA patent/SG10201910753RA/en unknown
- 2014-09-19 CN CN201480063250.8A patent/CN105849253B/zh active Active
- 2014-09-19 CA CA2924816A patent/CA2924816C/en active Active
- 2014-09-19 ES ES14767019T patent/ES2741806T3/es active Active
- 2014-09-19 US US14/917,103 patent/US10329532B2/en active Active
- 2014-09-19 EP EP14767019.4A patent/EP3047019B1/en active Active
- 2014-09-19 DK DK14767019.4T patent/DK3047019T3/da active
- 2014-09-19 CN CN202010891948.6A patent/CN112175900A/zh active Pending
- 2014-09-19 JP JP2016515530A patent/JP6681825B2/ja active Active
-
2018
- 2018-09-21 US US16/138,101 patent/US20190010460A1/en not_active Abandoned
-
2019
- 2019-05-20 US US16/416,580 patent/US20190300858A1/en not_active Abandoned
-
2020
- 2020-03-24 JP JP2020052526A patent/JP7548538B2/ja active Active
Also Published As
| Publication number | Publication date |
|---|---|
| SG10201802167TA (en) | 2018-04-27 |
| CN112175900A (zh) | 2021-01-05 |
| JP2016536975A (ja) | 2016-12-01 |
| AU2014323098B2 (en) | 2020-09-17 |
| DK3047019T3 (da) | 2019-08-26 |
| JP2020115866A (ja) | 2020-08-06 |
| US20190300858A1 (en) | 2019-10-03 |
| AU2014323098A1 (en) | 2016-05-05 |
| EP3047019A1 (en) | 2016-07-27 |
| EP3047019B1 (en) | 2019-07-03 |
| WO2015040142A1 (en) | 2015-03-26 |
| US10329532B2 (en) | 2019-06-25 |
| CA2924816C (en) | 2023-09-05 |
| CN105849253A (zh) | 2016-08-10 |
| EP3575393A1 (en) | 2019-12-04 |
| SG10201910753RA (en) | 2020-01-30 |
| CA2924816A1 (en) | 2015-03-26 |
| CN105849253B (zh) | 2020-10-09 |
| US20190010460A1 (en) | 2019-01-10 |
| SG11201601798RA (en) | 2016-04-28 |
| JP7548538B2 (ja) | 2024-09-10 |
| US20160215264A1 (en) | 2016-07-28 |
| JP6681825B2 (ja) | 2020-04-15 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2741806T3 (es) | Un método para la diferenciación directa de células madre pluripotentes en músculo cardíaco funcional | |
| US11680246B2 (en) | Ex vivo proliferation of epithelial cells | |
| US9347042B2 (en) | Method for producing corneal endothelial cell | |
| ES2688019T3 (es) | Métodos y composiciones para la producción acelerada de células epiteliales pigmentadas retinianas a partir de células pluripotentes | |
| ES2677319T3 (es) | Medio y método de cultivo de células madre | |
| CA2998372C (en) | EX VIVO PROLIFERATION OF EPITHELIAL CELLS | |
| US10100285B2 (en) | Ex vivo proliferation of epithelial cells | |
| US20110165130A1 (en) | Methods for Preparing Human Skin Substitutes from Human Pluripotent Stem Cells | |
| CN116769695A (zh) | 从畸胎瘤、类器官和拟胚体中产生人细胞和组织的培养基和方法 | |
| HK40017677A (en) | A method to direct differentiation of pluripotent stem cells into functional heart muscle | |
| HK40043185A (en) | A method to direct differentiation of pluripotent stem cells into functional heart muscle | |
| HK1227926B (zh) | 一种多能干细胞定向分化为功能性心肌的方法 | |
| HK1227926A1 (en) | A method to direct differentiation of pluripotent stem cells into functional heart muscle | |
| HK1249919B (en) | Ex vivo proliferation of epithelial cells |