ES2870717T3 - Anticuerpos humanos y secuencias de unión específicas del mismo para su uso en accidente cerebrovascular e isquemia y condiciones isquémicas - Google Patents
Anticuerpos humanos y secuencias de unión específicas del mismo para su uso en accidente cerebrovascular e isquemia y condiciones isquémicas Download PDFInfo
- Publication number
- ES2870717T3 ES2870717T3 ES17188389T ES17188389T ES2870717T3 ES 2870717 T3 ES2870717 T3 ES 2870717T3 ES 17188389 T ES17188389 T ES 17188389T ES 17188389 T ES17188389 T ES 17188389T ES 2870717 T3 ES2870717 T3 ES 2870717T3
- Authority
- ES
- Spain
- Prior art keywords
- seq
- antibody
- sequence
- rhigm12
- neurons
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
- A61K39/39533—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum against materials from animals
- A61K39/39541—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum against materials from animals against normal tissues, cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K45/00—Medicinal preparations containing active ingredients not provided for in groups A61K31/00 - A61K41/00
- A61K45/06—Mixtures of active ingredients without chemical characterisation, e.g. antiphlogistics and cardiaca
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K51/00—Preparations containing radioactive substances for use in therapy or testing in vivo
- A61K51/02—Preparations containing radioactive substances for use in therapy or testing in vivo characterised by the carrier, i.e. characterised by the agent or material covalently linked or complexing the radioactive nucleus
- A61K51/04—Organic compounds
- A61K51/08—Peptides, e.g. proteins, carriers being peptides, polyamino acids, proteins
- A61K51/10—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody
- A61K51/1018—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody against material from animals or humans
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P3/00—Drugs for disorders of the metabolism
- A61P3/06—Antihyperlipidemics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/02—Antithrombotic agents; Anticoagulants; Platelet aggregation inhibitors
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/10—Drugs for disorders of the cardiovascular system for treating ischaemic or atherosclerotic diseases, e.g. antianginal drugs, coronary vasodilators, drugs for myocardial infarction, retinopathy, cerebrovascula insufficiency, renal arteriosclerosis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/12—Antihypertensives
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/06—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies from serum
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
- A61K2039/507—Comprising a combination of two or more separate antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/545—Medicinal preparations containing antigens or antibodies characterised by the dose, timing or administration schedule
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/55—Medicinal preparations containing antigens or antibodies characterised by the host/recipient, e.g. newborn with maternal antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/21—Immunoglobulins specific features characterized by taxonomic origin from primates, e.g. man
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/24—Immunoglobulins specific features characterized by taxonomic origin containing regions, domains or residues from different species, e.g. chimeric, humanized or veneered
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/565—Complementarity determining region [CDR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Engineering & Computer Science (AREA)
- Immunology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Epidemiology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Biomedical Technology (AREA)
- Cardiology (AREA)
- Heart & Thoracic Surgery (AREA)
- Optics & Photonics (AREA)
- Physics & Mathematics (AREA)
- Mycology (AREA)
- Microbiology (AREA)
- Diabetes (AREA)
- Hematology (AREA)
- Obesity (AREA)
- Vascular Medicine (AREA)
- Pain & Pain Management (AREA)
- Neurosurgery (AREA)
- Neurology (AREA)
- Urology & Nephrology (AREA)
- Rheumatology (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Peptides Or Proteins (AREA)
Abstract
Un anticuerpo de IgM humano aislado o un fragmento del mismo que se une específicamente a las neuronas y protege las neuronas de la muerte celular y que no promueve la remielinización, en donde el anticuerpo de IgM humano aislado o fragmento comprende la siguiente secuencia: (a) las secuencias del dominio de CDR de aminoácidos de cadena pesada variable CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y las secuencias de la CDR de cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), como se expone en la Figura 5; o (b) las secuencias del dominio de CDR de aminoácidos de cadena pesada variable CDR1 GFTFSTYA (SEQ ID NO: 37), CDR2 INVGGVTT (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), y las secuencias de CDR de cadena ligera CDR1 QGIG ( SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42), como se establece en la Figura 6, para su uso en un método para tratar o mejorar la isquemia cerebral o accidente cerebrovascular, mejorar la función neural en un animal después de un accidente cerebrovascular o isquemia cerebral, o para proteger neuronas o células neurales del daño o muerte celular después de un accidente cerebrovascular o isquemia cerebral, en donde el método comprende administrar el anticuerpo de IgM humano aislado o fragmento del mismo en combinación con un anticuerpo remielinizante, y en donde el anticuerpo remielinizante comprende: (i) las CDR de la región variable de la cadena pesada que comprenden la secuencia de CDR1 SSGMH (SEQ ID NO: 47), la secuencia de CDR2 V(I)ISYDGSRKYYADSVKG (SEQ ID NO: 48) y la secuencia de CDR3 GVTGSPTLDY (SEQ ID NO: 49), y las CDR de la región variable de la cadena ligera que comprenden la secuencia de CDR1 SGSSSNIGNNFVS (SEQ ID NO: 50), la secuencia de CDR2 DITKRPS (SEQ ID NO: 51) y la secuencia de CDR3 G(E)TWDSSLSAW (SEQ ID NO: 52); o (ii) las CDR de la región variable de la cadena pesada que comprenden la secuencia de CDR1 SGFTFSSYW (SEQ ID NO: 53), secuencia de CDR2 IKKDGSEK (SEQ ID NO: 54) y secuencia de CDR3 ARPNCGGDCYLPWYFD (SEQ ID NO: 55), y las CDR de la región variable de la cadena ligera que comprenden la secuencia de CDR1 QSVLYSSNNKNY (SEQ ID NO: 56), la secuencia de CDR2 YWAS (SEQ ID NO: 57) y la secuencia de CDR3 QQYYNTPQA (SEQ ID NO: 58).
Description
DESCRIPCIÓN
Anticuerpos humanos y secuencias de unión específicas del mismo para su uso en accidente cerebrovascular e isquemia y condiciones isquémicas
DECLARACIÓN DE APOYO GUBERNAMENTAL
La invención descrita en el presente documento fue apoyada en su totalidad o en parte por los Institutos Nacionales de Salud (Concesión Nos. R01 NS 24180, R01 NS 32129, R01 CA104996, R01 CA096859), y la Sociedad Nacional de Esclerosis Múltiple (Concesión N° CA 1011 A8-3) El gobierno de los Estados Unidos tiene ciertos derechos sobre la invención.
CAMPO DE LA INVENCIÓN
La presente invención se refiere a anticuerpos, anticuerpos naturales humanos particularmente anticuerpos recombinantes derivados de los mismos y fragmentos de los mismos, que son capaces de unirse a y reconocer las neuronas en el SNC y provocar respuestas en las neuronas del SNC. Estos anticuerpos son útiles en el diagnóstico y tratamiento de afecciones asociadas con daños en los nervios, lesiones o degeneración, enfermedades neurodegenerativas, lesiones o daños crónicos en los nervios y lesiones o daños repentinos a los nervios. Los anticuerpos, regiones variables o secuencias del dominio CDR de los mismos, y fragmentos de los mismos de la presente invención también pueden usarse en terapia en combinación con agentes quimioterapéuticos, inmunomoduladores o agentes neuroactivos y/o con otros anticuerpos o fragmentos de los mismos. Esta invención se refiere en particular a métodos para aliviar o tratar apoplejía o isquemia, particularmente isquemia cerebral, y agentes, construcciones y composiciones asociadas para uso en apoplejía o isquemia.
ANTECEDENTES DE LA INVENCIÓN
La neuroregeneración se refiere a la regeneración o reparación de tejidos, células o productos celulares nerviosos. Dichos mecanismos pueden incluir remielinización, generación de nuevas neuronas, glia, axones, mielina o sinapsis. La neuroregeneración difiere entre el Sistema Nervioso Periférico (SNP) y el Sistema Nervioso Central (SNC) tanto por los mecanismos funcionales involucrados como por el alcance y la velocidad de la regeneración.
La regeneración de Axon en el sistema nervioso central de mamífero maduro (SNC) está muy limitada después de la lesión. En consecuencia, los déficits funcionales persisten después de la lesión de la médula espinal (LME), la lesión cerebral traumática, el accidente cerebrovascular y las afecciones relacionadas que implican la desconexión axonal. Esta situación difiere de la del sistema nervioso periférico de mamíferos (PNS), donde la regeneración axonal a larga distancia y la recuperación funcional sustancial pueden ocurrir en el adulto. Tanto las moléculas extracelulares como la capacidad de crecimiento intrínseca de la neurona influyen en el éxito regenerativo.
Axones del sistema nervioso central (SNC) no se regeneran de forma espontánea después de una lesión en los mamíferos adultos. Por el contrario, los axones del sistema nervioso periférico (SNP) se regeneran fácilmente, lo que permite la recuperación de la función después del daño del nervio periférico. Aguayo y sus colegas demostraron que al menos algunas neuronas maduras del SNC conservan la capacidad de regenerarse cuando se les proporciona un injerto de nervio periférico permisivo (Richardson PM, McGuinness UM, Aguayo AJ (1980) Nature 284: 264-265; Richardson PM, Issa VM, Aguayo AJ (1984) J Neurocytol 13: 165 - 182; David S, Aguayo a J (1981) Science 214: 931 - 933; Benfey M, Aguayo AJ (1982) Nature 296: 150 - 152). Este trabajo sugirió que el entorno de SNP es estimulante y/o que el entorno del SNC inhibe el crecimiento de axones. Estudios posteriores identificaron factores promotores del crecimiento en el SNP y factores inhibidores del crecimiento en el SNC. Los inhibidores de la regeneración incluyen proteínas específicas en la mielina del SNC y moléculas asociadas con la cicatriz astroglial. Además, un aclaramiento de residuos más lento en el sistema nervioso central con respecto al SNP puede impedir el recrecimiento axonal. La comprensión de los factores que influyen en el crecimiento del axón es fundamental para el desarrollo de terapias para promover la regeneración del SNC.
Después de la lesión del nervio periférico, los axones se regeneran fácilmente. La porción distal del axón, que está desconectada del cuerpo celular, sufre degeneración walleriana. Este proceso activo da como resultado la fragmentación y la desintegración del axón. Los restos son eliminados por las células gliales, predominantemente macrófagos. Los axones proximales pueden regenerar y re-inervar sus objetivos, permitiendo la recuperación de la función.
Las dos clases principales de inhibidores de la regeneración del SNC son los inhibidores asociados a la mielina (MAIS) y los proteoglicanos de sulfato de condroitina (CSPGs). Estas moléculas limitan la regeneración del axón y, al interferir con su función, se logra cierto grado de crecimiento en el sistema nervioso central del adulto. Los factores autónomos celulares también son determinantes importantes de la falla de regeneración del SNC. Las neuronas del SNC no regulan al alza los genes asociados al crecimiento en la misma medida que las neuronas del
SNP. En consecuencia, su capacidad de regeneración está limitada incluso en ausencia de inhibidores. El aumento de la capacidad de crecimiento intrínseco de las neuronas permite una moderada regeneración de axones dentro del SNC (Bomze HM y otros (2001) Nat Neurosci 4: 38-43; Neumann S, Woolf CJ (1999) Neuron 23: 83-91).
Los MAI son proteínas expresadas por oligodendrocitos como componentes de la mielina del SNC. Los MAI deterioran el crecimiento de neuritas in vitro y se cree que limitan el crecimiento de axones in vivo después del daño al SNC. Los MAI incluyen Nogo-A (Chen MS y otros (2000) Nature 403: 434-439; GrandPre T y otros (2000) Nature 403: 439-44), glicoproteína asociada a mielina (MAG) (McKerracher L y cols. (1994) Neuron 13: 805-811), glicoproteína de mielina de oligodendrocitos (OMgp) (Kottis V y otros (2002) J Neurochem 82: 1566-1569), efrina-B3 (Benson MD y otros (2005) Proc Nat Acad Sci USA 102: 10694 -10699) y Semaforina 4D (Sema4D) (Moreau-Fauvarque C y otros (2003) J Neurosci 23: 9229-9239). Tres de estos (Nogo-A, MAG y OMgp) interactúan con un receptor neuronal Nogo-66 1 (NgR1) para limitar el crecimiento de axones. Estos tres ligandos estructuralmente no relacionados también muestran afinidad por un segundo receptor inhibidor del crecimiento de axones, receptor emparejado tipo B de inmunoglobulina (PirB) (Atwal JK y otros (2008) Science 322: 967 - 970).
Se han identificado varias moléculas de reconocimiento que actúan como señales moleculares para la promoción y/o inhibición del crecimiento de neuritas subyacentes. Entre las neuritas que promueven la promoción de moléculas de reconocimiento, la molécula de adhesión de células neurales L1 juega un papel prominente en la mediación del crecimiento de neuritas (Schachner M (1990) Seminars in the Neurosciences 2: 497-507). El crecimiento de neuritas dependiente de L1 está mediado por la interacción homófila. L1 potencia la excrecencia de neuritas en neuritas que expresan L1 y células de Schwann, y fibroblastos transfectados en L1 (Bixby et al (1982) Proc Natl Acad. Sci. USA 84: 2555-2559; Chang et al (1987) J Cell Biol 104: 355-362 Lagenaur y otros (1987) Proc Natl Acad Sci USA 84: 7753 - 7757; Seilheimer y otros (1988) J Cell Biol 107: 341 - 351; Kadmon y otros (1990) J Cell Biol 110: 193 - 208; Williams et al (1992) J Cell Biol 119: 883 - 892).
Las lesiones del sistema nervioso afectan a más de 90,000 personas cada año, y un número mucho mayor cuando se incluyen eventos cerebrovasculares tales como apoplejía. Se estima que las lesiones de la médula espinal por sí solas afectan a 10,000 cada año. Como resultado de esta alta incidencia de lesiones neurológicas, la regeneración y reparación de nervios, un subcampo de la ingeniería de tejidos neurales se está convirtiendo en un campo en rápido crecimiento dedicado al descubrimiento de nuevas formas de recuperar la funcionalidad nerviosa después de una lesión. El sistema nervioso se divide en dos partes: el sistema nervioso central, que consiste en el cerebro y la médula espinal, y el sistema nervioso periférico, que consiste en nervios craneales y espinales junto con sus ganglios asociados. Si bien el sistema nervioso periférico tiene una capacidad intrínseca de reparación y regeneración, el sistema nervioso central tiene, de forma comparativa y en su mayor parte, restringida su capacidad de autorreparación y regeneración. Actualmente no existe un tratamiento aceptado y aprobado para recuperar la función del nervio humano después de una lesión en el sistema nervioso central.
La protección y reparación de los axones después de una lesión de la médula espinal (LME) tiene un gran potencial como estrategia efectiva para prevenir la pérdida de la neurona motora y la discapacidad permanente. La protección de neuronas se ha logrado utilizando factores tróficos dirigidos para prevenir daños y promover la reparación de axones después de una lesión. Estas moléculas se identificaron predominantemente usando estrategias de selección basadas en sistemas in vitro que se centraron en seleccionar factores neurotróficos de molécula pequeña específicos. Mientras que estas moléculas demostraron resultados neuroprotectores en modelos preclínicos, los resultados de los ensayos clínicos han sido menos favorables.
Los anticuerpos monoclonales autorreactivos naturales han demostrado funciones biológicas beneficiosas en las células del SNC usando múltiples modelos de lesión y enfermedad. La promoción mediada por anticuerpos de la supervivencia neuronal, la regeneración axonal y la recuperación funcional se ha demostrado in vivo usando la IgM monoclonal de ratón, IN-1 (Bregman BS y otros (1995) Nature 378 (6556): 498-501; Caroni P, Schwab ME (1988) Neuron 1 (1): 85 - 96). Se obtuvieron resultados similares usando la inmunización del homogenato de la médula espinal (SCH) antes del daño en el SNC (Ellezam B, Bertrand J, Dergham P, McKerracher L (2003) Neurobiol Dis 12 (1): 1-10; Huang DW et al (1999) Neuron 24 (3): 639 - 647).
La esclerosis múltiple (MS) es una enfermedad crónica, el sistema de frecuencia progresiva, inflamatoria nervioso central (SNC), caracterizada patológicamente por desmielinización primaria, habitualmente sin daño axonal inicial. La etiología y la patogénesis de la EM son desconocidas. Varias características inmunológicas de la esclerosis múltiple y su asociación moderada con ciertos alelos complejos de histocompatibilidad mayor, han llevado a la especulación de que la esclerosis múltiple es una enfermedad mediada por el sistema inmune. Una hipótesis autoinmune está respaldada por el modelo experimental de encefalomielitis (EAE) autoinmune (alérgica), donde la inyección de ciertos componentes de mielina en animales genéticamente susceptibles conduce a la desmielinización del SNC mediada por células T. Sin embargo, los autoantígenos específicos y las células T reactivas a mielina patógenas no se han identificado definitivamente en el SNC de los pacientes con EM, ni la EM está asociada con otras enfermedades autoinmunes. Una hipótesis alternativa, basada en datos epidemiológicos, es que un factor ambiental, tal vez un virus no identificado, precipita una respuesta inflamatoria en el SNC, lo que conduce a la destrucción directa o indirecta (mielógena) de la mielina, potencialmente con una composición autoinmune inducida.
Esta hipótesis está respaldada por la evidencia de que varias infecciones virales naturales, tanto en humanos como en animales, pueden causar desmielinización. Un modelo viral experimental comúnmente utilizado es inducido por el virus de la encefalomielitis murina de Theiler (TMEV) (Dal Canto, MC, y Lipton, HL, Am. J. Path., 88: 497-500 (1977)).
En el caso de la EM, la desmielinización en última instancia resulta en la muerte de las células nerviosas. Sin embargo, la muerte del axón después de la desmielinización no es inmediata. Si se proporcionan moléculas o células de soporte apropiadas, el sistema nervioso tiene una capacidad significativa de reparación. Proteger los axones del s Nc promete ser una estrategia efectiva para limitar la pérdida de axones supervivientes y prevenir la discapacidad permanente. La neuroprotección se puede lograr modulando el entorno inflamatorio potencialmente neurotóxico en las lesiones. Los reactivos diseñados para limitar la excitotoxicidad, inhibir el óxido nítrico o bloquear los canales iónicos se están estudiando como métodos para proteger a los axones en peligro (Pitt, D., P. Werner y CS Raine (2000) Nat Med 6: 67-70; Okuda, Y y otros (1997) Journal of neuroimmunology 73: 107 - 116; Waxman, SG (2002).J Rehabil Res Dev 39: 233 - 242). Muchos de estos reactivos son moléculas pequeñas con toxicidad demostrada y actúan sistémicamente en todas las células. Wootla y otros, Expert Rev Neurother. 2012 abril; 1284: 409-420, indican que los anticuerpos IgM humanos tienen potencial terapéutico para las lesiones del SNC.
La isquemia es una restricción en el suministro de sangre a los tejidos, provocando una escasez de oxígeno y glucosa necesarios para el metabolismo celular (para mantener el tejido vivo). La isquemia generalmente es causada por problemas con los vasos sanguíneos, con el consiguiente daño o disfunción del tejido. También se refiere a la anemia local en una parte determinada del cuerpo a veces como resultado de la congestión (como vasoconstricción, trombosis o embolia). La hipoxia es una afección en la cual el cuerpo como un todo o una región del cuerpo (hipoxia tisular o, con menor frecuencia, hipoxia regional) se ve privada de un suministro adecuado de oxígeno. Las variaciones en las concentraciones de oxígeno arterial pueden ser parte de la fisiología normal, por ejemplo, durante el ejercicio físico extenuante. Una falta de correspondencia entre el suministro de oxígeno y su demanda a nivel celular puede provocar una condición hipóxica. La hipoxia isquémica ocurre cuando hay una disminución del flujo sanguíneo y puede ocurrir, por ejemplo, en la insuficiencia cardíaca y el choque vasodilatador. La hipoxia cerebral ocurre cuando no llega suficiente oxígeno al cerebro. El cerebro necesita un suministro constante de oxígeno y nutrientes para funcionar.
En la hipoxia cerebral, a veces sólo se interrumpe el suministro de oxígeno. Esto puede ser causado por la inhalación de humo), intoxicación por monóxido de carbono, asfixia, enfermedades que impiden el movimiento de los músculos de la respiración, altitudes elevadas, presión sobre la tráquea y estrangulación. En otros casos, se interrumpe el suministro de oxígeno y nutrientes, como paro cardíaco, arritmia cardíaca, complicaciones de la anestesia general, ahogamiento, sobredosis de drogas, lesiones a un recién nacido durante o después del nacimiento, como parálisis cerebral, accidente cerebrovascular, presión arterial baja.
La isquemia cerebral es insuficiente flujo de sangre al cerebro, y puede ser aguda o crónica. El accidente cerebrovascular isquémico agudo es una emergencia neurológica que puede ser reversible si se trata rápidamente. La isquemia crónica del cerebro puede dar lugar a una forma de demencia llamada demencia vascular. Un breve episodio de isquemia que afecta al cerebro se denomina ataque isquémico transitorio (AIT), con frecuencia denominado mini accidente cerebrovascular.
Una apoplejía, también conocida como un accidente cerebrovascular (CVA), es la rápida pérdida de la función cerebral debido a las perturbaciones en el suministro de sangre al cerebro. Esto puede deberse a isquemia (falta de flujo sanguíneo) causada por obstrucción (trombosis, embolia arterial) o hemorragia (filtración de sangre). La isquemia cerebral, también conocida como isquemia cerebral, es una afección en la que el flujo sanguíneo al cerebro es insuficiente para satisfacer la demanda metabólica y es un tipo de accidente cerebrovascular junto con hemorragia subaracnoidea y hemorragia intracerebral. La isquemia puede ser isquemia focal, que se limita a una región específica del cerebro, o isquemia global, que abarca amplias áreas del tejido cerebral. El accidente cerebrovascular es la principal causa de discapacidad a largo plazo en los Estados Unidos. Cada año, cerca de 800,000 personas sufren un derrame cerebral. Después de una lesión isquémica, la discapacidad asociada con los déficits clínicos se debe principalmente a la disfunción de las neuronas. La falta de recuperación funcional se atribuye en parte a la restricción de la regeneración y neuroplasticidad (Walmsley, AR, y Mir, AK (2007) Current Pharmaceutical Design 13: 2470-2484). El accidente cerebrovascular puede provocar una función motora alterada, como la incapacidad para mover una o más extremidades en un lado del cuerpo, efectos cognitivos como la incapacidad de comprender o formular el habla u otros déficits, incluidos efectos visuales, como la incapacidad de ver un lado del campo visual. Actualmente no hay tratamientos efectivos para prevenir o revertir los déficits neurológicos relacionados con el accidente cerebrovascular.
Un accidente cerebrovascular isquémico se trata de vez en cuando en un hospital con trombolisis (también conocido como un "destructor de coágulos"), y algunos accidentes cerebrovasculares hemorrágicos se benefician de la neurocirugía. La prevención de la recurrencia puede implicar la administración de fármacos antiplaquetarios como la aspirina y el dipiridamol, el control y la reducción de la hipertensión y el uso de estatinas. Los pacientes seleccionados pueden beneficiarse de la endarterectomía carotídea y el uso de anticoagulantes. La terapia
intervencionista de la isquemia cerebral permanece limitada a la trombólisis enzimática mediante el uso del activador del plasminógeno tisular recombinante rtPA (Brott, TG, y cols. (1992) Stroke 23: 632-640; Haley, EC, y cols. (1992) Stroke 23: 641-645; grupo, TN (1995) New England J Med 333: 1581-1587), y a la extracción intervencionista del coágulo sanguíneo mediante cateterización directa (Gobin YP y cols. (2004) Stroke 35: 2848-2854). Sin embargo, rtPA tiene una ventana terapéutica corta de 3 horas y una mayor incidencia de isquemia cerebral hemorrágica si se infunde más allá del marco de 3 horas (Clark, WM y otros (1999) JAMA 282: 2019-2026; Hacke W et al (2008) New England J Med 359: 1317 - 1329).
Se han identificado autoanticuerpos monoclonales humanos que exhiben actividad en el sistema nervioso central, y que se asociaron particularmente con la estimulación de la remielinización. Sería deseable desarrollar anticuerpos monoclonales humanos con actividad en el SNC, particularmente anticuerpos recombinantes con estas actividades, y con la capacidad de promover la regeneración neuronal y/o proteger a las neuronas de enfermedades, lesiones, daños y/o muerte, particularmente en casos de apoplejía o isquemia cerebral/cerebral, o donde el oxígeno o el flujo sanguíneo al cerebro o al SNC se ve privado o afectado de manera aguda o crónica. En particular, sería deseable el uso y la aplicación de anticuerpos monoclonales humanos, con capacidad para acceder al SNC y tratar, prevenir o aliviar la isquemia cerebral o apoplejía. Es hacia el logro de ese objetivo que se dirige la presente invención.
RESUMEN DE LA INVENCIÓN
La invención se refiere a agentes neuromoduladores con especial eficacia en el SNC, cuyos agentes comprenden un material seleccionado del grupo que consiste de un anticuerpo del subtipo IgM, fragmentos activos de los mismos, monómeros de los mismos, agonistas de los mismos y combinaciones de los mismos. Los agentes neuromoduladores o anticuerpos tienen una o más de las siguientes características: protegen y/o estabilizan las neuronas; se dirigen a sitios en el SNC o daño, compromiso o lesión de las células nerviosas; reducen o bloquean la muerte celular, por ejemplo, muerte celular inducida por peróxido de hidrógeno. En un aspecto particular de la presente invención, los agentes, particularmente los anticuerpos monoclonales de unión a neuronas, son eficaces y tienen aplicación y uso en el tratamiento y alivio de accidentes cerebrovasculares e isquemia, particularmente isquemia cerebral, y en condiciones en las que está involucrada o ha ocurrido isquemia o hipoxia de células nerviosas o del cerebro.
La invención está definida por las reivindicaciones. La descripción proporciona a anticuerpos monoclonales de unión a neuronas con la capacidad para promover la extensión de neuritas, para actuar en neuroregeneración, y/o para proteger las neuronas de los daños y para fines de diagnóstico y terapéuticos en el sistema nervioso central. Los presentes estudios demuestran que cuando se administran anticuerpos monoclonales de unión a neuronas, particularmente anticuerpo IgM12 rHIgM12 a animales después de un accidente cerebrovascular (en este caso un modelo animal para la inducción de accidente cerebrovascular isquémico), se produce una clara mejora de la función de los animales y protege la integridad del tejido.
En particular, se proporcionan anticuerpos recombinantes, específicos, en los que dichos anticuerpos reconocen y son capaces de unir neuronas, incluyendo las neuronas corticales, las neuronas del hipocampo, las células granulares de cerebelo y células ganglionares de la retina, y que se dirigen y median neuronas y células neuronales en el evento y después de un accidente cerebrovascular, afección isquémica o isquemia o hipoxia en células nerviosas o microglía. En este documento se proporcionan anticuerpos recombinantes completamente humanos. Los anticuerpos de la presente invención tienen un uso diagnóstico y terapéutico en afecciones o enfermedades asociadas con compromiso nervioso, daño o lesión.
En un aspecto general, la presente divulgación proporciona anticuerpos dirigidos contra y capaz de unirse uno o más epítopos sobre las neuronas, incluyendo neuronas corticales, las neuronas del hipocampo, las células granulares de cerebelo y células ganglionares de la retina. En un aspecto amplio, la presente divulgación proporciona un miembro de unión específica aislada, particularmente un anticuerpo o fragmento del mismo, que incluye un anticuerpo humano recombinante, que reconoce, se une y/o se dirige a neuronas. La invención proporciona un anticuerpo, particularmente un anticuerpo humano, particularmente un anticuerpo IgM, que se une específicamente a las neuronas y protege a las neuronas de la muerte celular y que no promueve la remielinización. El anticuerpo tiene uso en el tratamiento o la mejora de enfermedades o afecciones en mamíferos en los que los nervios están comprometidos, lesionados o están en riesgo, incluso en lesión de la médula espinal (SCI), lesión traumática de brian (TBI), esclerosis lateral amolotrópica (ELA) esclerosis múltiple (EM), enfermedad de Alzheimer, apoplejía, enfermedad de Parkinson, enfermedad de Huntington, anoxia prenatal/isquemia perinatal, parálisis cerebral, encefalopatía, mielopatía o enfermedades de las neuronas motoras, y particularmente en la protección, supervivencia o mantenimiento de neuronas y capacidad neurológica en estas enfermedades.
En un aspecto particular, el anticuerpo de la invención o composiciones de los mismos, tienen uso y aplicación en el accidente cerebrovascular y en la isquemia cerebral. En un aspecto particular, los anticuerpos o composiciones de la invención tienen uso en condiciones en las que el flujo sanguíneo y/o el oxígeno son insuficientes para satisfacer la demanda metabólica o en la que existe una lesión traumática en el cerebro que
resulta en la muerte o lesión o riesgo de muerte o lesión de neuronas y células asociadas. En un aspecto particular, la presente invención proporciona un anticuerpo o fragmento del mismo, que es el anticuerpo 12 ó 42, particularmente el IgM12 o IgM42 derivado del suero o recombinante.
En un aspecto de la invención se proporciona un anticuerpo de unión a neurona recombinante o sintética que comprende las secuencias de región c Dr variables establecidas en la Figura 5 y/o 6. Anticuerpo 12 comprende secuencias de cadena pesada de CDR de CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y las secuencias CDR de cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), como se establece en la Figura 5. El anticuerpo 42 comprende secuencias CDR de cadena pesada CDR1 GfTfSTYA (SEQ ID NO: 37), INVGGVTT CDR2 (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), y cadenas ligeras CDR secuencias CDR1 QGIG (SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42), como se establece en la Figura 6. Por consiguiente, anticuerpos recombinantes que se basan en CDR del anticuerpo identificados en este documento serán útiles para dirigir y proteger neuronas, particularmente neuronas susceptibles, comprometidas, dañadas o lesionadas en enfermedades o en cánceres.
La invención proporciona de este modo el anticuerpo aislado humano IgM o fragmento del mismo que se une específicamente a las neuronas y protege a las neuronas de la muerte celular y que no promueve la remielinización, en la que el anticuerpo o fragmento comprende: (a) las secuencias de dominio de CDR de cadena pesada variable de aminoácidos CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y secuencias CDR de cadena ligera CDR1 QSISSY (SEQ IDNO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSy Ht PW (SEQ ID NO: 36), como se expone en la Figura 5; o (b) las secuencias del dominio CDR de aminoácidos de cadena pesada variables CDR1 GFTFSTYA (SEQ ID NO: 37), INVGGVTT CDR2 (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), y secuencias CDR de cadena ligera CDR1 QGIG (SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42), como se establece en la Figura 6, para uso en el tratamiento o la mejora de la isquemia cerebral o apoplejía.
En un aspecto particular adicional, el anticuerpo comprende la secuencia de aminoácidos del anticuerpo 12 o 42 incluyendo tal como se establece en la Figura 5 y/o en la Figura 6. Anticuerpo recombinante IgM12 de la presente invención comprende la secuencia de cadena pesada variable (SEQ ID NO: 1) y secuencia de cadena ligera (SEQ ID NO: 11) como se establece en la Figura 5. El anticuerpo IgM42 recombinante de la presente invención comprende la secuencia de cadena pesada variable (SEQ ID NO: 17) y la secuencia de cadena ligera (SEQ ID NO: 27) como se ha expuesto en la Figura 6. En un aspecto particular, los anticuerpos recombinantes son anticuerpos recombinantes completamente humanos, que comprenden la región variable de la cadena pesada humana, la región constante y la cadena J humana. Se proporcionan anticuerpos IgM12 recombinantes completamente humanos que comprenden cadena pesada de inmunoglobulina humana que comprende la región variable (SEQ ID NO: 1), o sus CDR, una región constante humana, particularmente una secuencia kappa (SEQ ID NO: 3, 5, 7 y 9).) y cadena J humana (SEQ ID NO: 15) como se expone en la Figura 5.
En un aspecto adicional, la presente divulgación proporciona un anticuerpo aislado o fragmento del mismo capaz de unirse a un antígeno, en el que dicho anticuerpo o fragmento del mismo comprende un dominio de unión a polipéptido que comprende una secuencia de aminoácidos sustancialmente como se establece en el presente documento y en la Figura 6. La invención proporciona un anticuerpo humano aislado o fragmento del mismo capaz de unirse a neuronas, donde dicho anticuerpo o fragmento del mismo comprende una región variable de cadena pesada (SEQ ID NO: 17), o sus CDR, y una región variable de cadena ligera (SEQ ID NO: 27), o la CDRS de la misma, o que comprende una secuencia de aminoácidos sustancialmente como se establece aquí y en la Figura 6.
En un aspecto adicional, la presente divulgación proporciona un anticuerpo completamente humano aislado o fragmento del mismo capaz de unirse a un antígeno, en el que dicho anticuerpo o fragmento del mismo comprende un dominio de unión a polipéptido que comprende una secuencia de aminoácidos sustancialmente como se expone aquí y en la Figura 5. La descripción proporciona un aislado anticuerpo completamente humano o fragmento del mismo capaz de unirse a neuronas, donde dicho anticuerpo o fragmento del mismo comprende una región variable de cadena pesada (SEQ ID NO: 1), o las CDR de la misma, y región variable de cadena ligera (SEQ ID NO: 11) o sus CDR, o que comprenden una secuencia de aminoácidos sustancialmente como se establece aquí y en la Figura 5. La divulgación proporciona particularmente un anticuerpo IgM humano aislado o fragmento activo del mismo, que comprende las secuencias del dominio CDR de aminoácidos de cadena pesada variables CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y las secuencias CDR de cadena ligera CDR1 QSISSY (SEQ IDNO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 Q QSYHTPW (SEQ ID NO: 36), como se establece en la Figura 5. En un aspecto particular, la divulgación proporciona un anticuerpo IgM recombinante completamente humano o fragmento activo del mismo, que comprende las secuencias de dominio de CDR de aminoácidos de cadena pesada variables CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y las secuencias CDR de cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), una región constante humana y una secuencia de cadena J humana como se expone en SEQ ID NO: 15.
En aspectos adicionales, la descripción proporciona un ácido nucleico aislado que comprende una secuencia que codifica un polipéptido o anticuerpo de unión como se define anteriormente neurona, y métodos de preparación de los polipéptido s o anticuerpos de la invención que comprenden expresar dicha ácidos nucleicos en condiciones de lograr expresión de dicho polipéptido o anticuerpo, y recuperación del polipéptido o anticuerpo. En uno de tales aspectos, se proporciona una secuencia de la región variable de anticuerpo que codifica ácido nucleico que tiene las secuencias de aminoácidos expuestas en las Figuras 5 o 6 o se proporciona un anticuerpo que tiene secuencias de dominio de CDR como se expone en las Figuras 5 o 6. En un aspecto, se proporciona un ácido nucleico que comprende la secuencia de ácido nucleico de la Figura 5 o 6. En un aspecto adicional, se proporciona una región variable de cadena pesada VH o un ácido nucleico que codifica VL de región variable de cadena ligera como se establece en la figura 5 o 6. La presente invención también se refiere a una molécula de ADN recombinante o un gen clonado, o una variante degenerada del mismo, que codifica un anticuerpo de la presente invención; preferiblemente una molécula de ácido nucleico, en particular una molécula de ADN recombinante o gen clonado, que codifica el anticuerpo VH y VL, particularmente las secuencias de la región CDR, que tiene una secuencia o es capaz de codificar una secuencia mostrada en la Figura 5 o 6. En aspectos preferidos la invención proporciona ácido nucleico que codifica la secuencia de la cadena pesada (SEQ ID NO: 2) y la secuencia de la región variable de la cadena ligera (SEQ ID NO: 12) de la IgM12 y la secuencia de la cadena variable de la cadena ligera (SEQ ID NO: 18) que codifica el ácido nucleico (SEQ ID NO: 28) de IgM42. En un aspecto de la invención, se proporciona un ácido nucleico que codifica un anticuerpo o fragmento del mismo que comprende las secuencias del dominio CDR de aminoácidos de cadena pesada variable CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y las secuencias de CDR de cadena ligera CDR1 QSISSY (SEQ IDNO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36). En un aspecto adicional, el ácido nucleico codifica además una región constante humana y una cadena J humana, particularmente la cadena J de SEQ ID NO: 15. En un aspecto de la invención, se proporciona un ácido nucleico que codifica un anticuerpo o fragmento del mismo que comprende las secuencias del dominio CDR de aminoácidos de cadena pesada variables CDR1 GFTFSTYA (SEQ ID NO: 37), INVGGVTT CDR2 (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), y secuencias CDR de cadena ligera CDR1 QGIG (SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42). En un aspecto adicional, el ácido nucleico codifica adicionalmente una región constante humana y una cadena J humana, particularmente la cadena J de la SEQ ID NO: 15.
Los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable se pueden usar en un método de tratamiento, prevención o diagnóstico del cuerpo humano o animal, tal como un método de neuroprotección en un mamífero que comprende administrar a dicho mamífero una cantidad eficaz de los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes de la invención. Los anticuerpos recombinantes o fragmentos de los mismos que comprenden las secuencias de la región de dominio CDR pueden usarse en un método de neuroprotección en un mamífero que comprende administrar a dicho mamífero una cantidad eficaz de los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes de la invención.
Los anticuerpos recombinantes o fragmentos de los mismos pueden ser utilizados como agentes neuroprotectores en métodos para la prevención, tratamiento o mejora de lesión del nervio, daño o compromiso y complicaciones que pueden, pueden o no resultar en daño en el SNC. Los métodos de tratamiento o prevención son aplicables cuando la pérdida de estructura, función o supervivencia de las neuronas está involucrada o asociada, incluyendo lesión o trauma cerebral, lesión de la médula espinal (SCI), lesión nerviosa, lesión en la cabeza, condiciones en las que se reduce el suministro de sangre al cerebro o enfermedades infecciosas del cerebro comprometidas, enfermedades neurodegenerativas. Ejemplos de dichas enfermedades o condiciones para el tratamiento, prevención o mejora de acuerdo con los métodos de la invención incluyen lesión de la médula espinal (SCI), lesión cerebral traumática (TBI), esclerosis lateral amolotrópica (ELA), esclerosis múltiple (MS), enfermedad de Alzheimer, accidente cerebrovascular, enfermedad de Parkinson, enfermedad de Huntington, anoxia prenatal/isquemia perinatal y/o parálisis cerebral, encefalopatía, mielopatía y enfermedades de las neuronas motoras. La presente invención se refiere así a métodos para tratar o mejorar enfermedades o afecciones en mamíferos en los que los nervios están dañados, comprometidos o escenarios donde los nervios o neuronas son susceptibles o en riesgo de compromiso, lesión o daño que comprende administrar un anticuerpo recombinante o completamente humano or fragmento del mismo seleccionado de IgM12 y IgM42.
En un aspecto particular de la invención, los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con la invención, se utilizan en métodos o se administran en composiciones para tratamiento o mejora de accidente cerebrovascular o isquemia cerebral. En un aspecto particular de la invención, los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con la invención, se pueden utilizar en métodos o se administran en composiciones para tratamiento o mejora de accidente cerebrovascular. En un aspecto particular de la invención, los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con la invención, se pueden utilizar en métodos o se administran en composiciones para tratamiento o mejora de la isquemia cerebral. En una realización de la invención, los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con la invención, se pueden utilizar en métodos o se administran en composiciones para
mejorar la función motora en condiciones tales como accidente cerebrovascular. Los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de la región variable de acuerdo con la invención, se pueden utilizar en métodos o se administran en composiciones para la mejora de la función motora en un animal que padece o se sospecha que padece un accidente cerebrovascular. Los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con la invención, se pueden utilizar en métodos o se administran en composiciones para la mejora de la función motora en un animal que ha tenido un accidente cerebrovascular o que tiene la isquemia cerebral. En un aspecto particular, IgM12, rIgM12, anticuerpo 12 que comprende la cadena pesada CDR secuencias CDR1 GGSVSlYy (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y secuencias de cadena ligera CDR de CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), tal como se establece en la Figura 5, se pueden utilizar en el tratamiento, prevención o alivio de accidente cerebrovascular o isquemia cerebral. En un aspecto de la misma, los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con la invención se pueden administrar en los casos en que se determina el accidente cerebrovascular y/o isquemia cerebral, sospechosos o en riesgo, incluso en casos en donde agentes trombolíticos enzimáticos, tales como activador del plasminógeno tisular (TPA) u otros agentes trombolíticos o antitrombóticos, se pueden administrar o utilizar.
Los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con con la invención, se pueden utilizar en métodos o se administran en composiciones para la mejora o la estabilización de la función neurológica o de la función motora en los casos, enfermedades o condiciones en que los nervios se ven comprometidos, lesionados o dañados o escenarios en que los nervios o neuronas son susceptibles o en riesgo de compromiso, lesiones o daños. En un aspecto particular, los anticuerpos o fragmentos de los mismos se utilizan o se administran en etapas tempranas o iniciales de una enfermedad, trauma, o condición, o se repiten en el curso o duración de una enfermedad, trauma, o condición, a fin de facilitar o mantener la mejora o estabilización de la función neurológica o la función motora, incluyendo o seleccionado de movimiento, como caminar, o la función cognitiva, como recuerdo o reconocimiento.
En un aspecto de la invención, los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con la invención, se pueden utilizar en métodos o administrarse en composiciones para mitigar o mejorar efectos neurológicos, defectos, daños o muerte celular neuronal en caso de un accidente cerebrovascular o isquemia cerebral. De conformidad con los métodos, el anticuerpo mejora uno o más parámetros neurológicos o motores funcionales o evaluables, incluyendo un parámetro evaluable o escalable, tal como una escala de accidente cerebrovascular o escala de función. La administración de los anticuerpos es eficaz para mejorar o aliviar uno o más de un síntoma o parámetro asociado con el accidente cerebrovascular, incluyendo por ejemplo uno o más de la función superior y la extremidad inferior del motor, ataxia de extremidad, la función sensorial, el lenguaje, la articulación, la atención, la mirada y la función visual, brazo, mano y potencia de las piernas y el movimiento.
Los métodos pueden comprender la administración de más de un anticuerpo o fragmento, incluyendo combinaciones de anticuerpo IgM12 y 42. En un aspecto de la invención, se proporcionan métodos que comprenden la administración de uno o más anticuerpos o fragmentos, en que un anticuerpo comprende (a) la secuencias de dominio variable de CDR de aminoácido de la cadena pesada CDR1 Gg Sv SLy Y (SEQ iD NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y secuencias de CDR de la cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36) o (b) el aminoácido de secuencias de dominio CDR de cadena pesada variable CDR1 GFTFSTyA (SEQ ID NO: 37), CDR2 INVGGVTT (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), y secuencias CDR de cadena ligera de CDR1 QGIG (SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42). En un tal método adicional, uno o más anticuerpos IgM12 y/o un anticuerpo IgM42 puede combinarse con otro anticuerpo activo sobre el SNC, en particular incluyendo uno o más de los anticuerpos rHIgM22 y/o rHIgM46. anticuerpo rHIgM22 comprende la secuencia de aminoácidos expuesta en la SEQ ID NO: 43 y 44, o las CDR de estas SEQ iDs. El anticuerpo rHIgM46 comprende la secuencia de aminoácidos expuesta en la SEQ ID NO: 45 y 46, o las CDR de estas SEQ IDs. Por lo tanto, uno o más de anticuerpo IgM12 y/o de anticuerpo IgM42 se puede combinar con uno o más anticuerpos remielinizantes que comprenden (a) CDRs de la región variable de cadena pesada que comprende la secuencia de CDR1 SSGMH secuencia CDR2 V(I) ISYDGSRKYYADSVKG y secuencia CDR3 GVTGSPTLDY, y la región variable de cadena luz CDR de la que comprende la secuencia de CDR1 SGSSSNIGNNFVS secuencia de CDR2 DITKRPS y secuencia CDR3 G(E)TWDSSLSAV V; o (b) región variable de cadena pesada CDR comprendiendo la secuencia de CDR1 SGFTFSSYW secuencia CDR2 IKKDGSEK y secuencia CDR3 ARPNCGGDCYLPWYFD, y región variable de cadena ligera CDR que comprende la secuencia CDR1 QSVLYSSNNKNY secuencia CDR2 YWAS y secuencia CDR3 QQYYNTPQA. Las combinaciones de anticuerpos se pueden administrar colectivamente o en serie, y en varios momentos y diferentes cantidades o concentraciones. Anticuerpos bi- o multi-específicos pueden ser utilizados. En tal aspecto particular, el anticuerpo 12 y/o 42 se administra en combinación con el anticuerpo 22 y/o 46 y/o un anticuerpo que tiene las secuencias de región CDR de CDR1, CDR2 y CDR3 de IgM22 o IgM46, por administración combinada o en serie, separados por un corto período de tiempo o una longitud de tiempo más larga, incluso por horas, días o semanas. En un tal aspecto particular, el anticuerpo 12 y/o 42 se administra en combinación con el anticuerpo 22 y/o 46, por administración combinada o en
serie para el tratamiento de mejora de una enfermedad o condición que implica la neurodegeneración, y en particular incluyendo desmielinización. En un aspecto adicional, el anticuerpo 12 y/o 42 se administra en combinación con el anticuerpo 22 y/o 46, por administración combinada o en serie para el tratamiento de mejora de una enfermedad desmielinizante o condición. En un aspecto particular, el anticuerpo 12 y/o 42 se administra en combinación con el anticuerpo 22 y/o 46, por administración combinada o en serie para el tratamiento de mejora de la esclerosis múltiple. En uno de tales aspectos, la remielinización y neuroprotección se consigue, por efectos combinados, incluidos los efectos sinérgicos, en un aspecto clínico medible de la enfermedad de MS.
En vista de las capacidades neuroprotectoras y/o neurorregenerativas de los agentes de la presente invención, está invención se refiere además a métodos de producción in vitro y estimulación de la proliferación de las neuronas, incluyendo la proliferación de las neuronas corticales, las neuronas del hipocampo, las células granulares de cerebelo y células ganglionares de la retina. Tales neuronas proliferadas pueden ser adecuadas para protocolos y métodos terapéuticos de trasplante y de nervio de células.
La utilidad de diagnóstico se extiende al uso de los anticuerpos de la presente invención en ensayos y métodos para caracterizar la lesión del nervio del SNC o daños o para detectar o evaluar la enfermedad o condiciones que implican lesiones del nervio, daños o muerte (incluyendo la muerte necrótica o apoptótica), incluyendo ensayos de diagnóstico in vitro e in vivo y de formación de imágenes. Por lo tanto, los anticuerpos pueden utilizarse en ensayos y procedimientos para evaluar accidente cerebrovascular o isquemia y/o para evaluar la lesión en isquemia o accidente cerebrovascular o isquemia o accidente cerebrovascular predicho o sospechado. En un inmunoensayo, una cantidad de control de los anticuerpos, o similares se puede preparar y marcar con una enzima, un par de unión específica, ligando, un colorante, un marcador fluorescente y/o un elemento radioactivo, y puede entonces ser introducido en una muestra celular. Después de que el material marcado o su pareja de unión ha tenido la oportunidad de reaccionar con sitios dentro de la muestra, la masa resultante puede ser examinada mediante técnicas conocidas, que pueden variar con la naturaleza de la etiqueta adjunta. En una aplicación in vivo de diagnóstico o de formación de imágenes, el anticuerpo o un fragmento de unión de la neurona de los mismos se puede preparar y marcar con una enzima, un par de unión específica, ligando, un colorante, un marcador fluorescente y/o un elemento radioactivo, y luego pueden introducirse en un animal. Después de que el material marcado o su pareja de unión ha tenido la oportunidad de reaccionar con sitios dentro del animal, el animal puede ser examinado por técnicas conocidas, que pueden variar con la naturaleza de la etiqueta adjunta. En un aspecto de la utilidad de diagnóstico de la invención, los anticuerpos o fragmentos activos de los mismos de la invención pueden utilizarse en ensayos y métodos para caracterizar la isquemia cerebral o derrame cerebral. En particular, en los casos en que se sospecha de isquemia cerebral o derrame cerebral o existe tal riesgo, los anticuerpos pueden ser utilizados para determinar o evaluar la isquemia cerebral y/o accidente cerebrovascular.
Los miembros de unión específicos radiomarcados, particularmente anticuerpos y fragmentos de los mismos, son útiles in vitro en las técnicas de diagnóstico e in vivo en técnicas de obtención de radioimágenes. En un aspecto adicional miembros radiomarcados de unión específica, particularmente anticuerpos y fragmentos de los mismos, en particular los radioinmunoconjugados, son útiles en radioinmunoterapia, particularmente como anticuerpos radiomarcados para la reparación de la lesión del nervio, neuroregeneración, la recuperación neurodegenerativa, cáncer o terapia de tumor del SNC, o alternativamente para la ablación de tejido del nervio dañado o neuronas en ciertos casos. En un aspecto in vivo, el fragmento de anticuerpo o neurona de unión del mismo se marca y se administra al animal antes de, durante o después de la cirugía o una técnica quirúrgica, incluyendo una técnica estereotáctica o mínimamente invasiva, para el propósito de localizar la lesión del nervio o para evaluar el daño que queda o lesión el tejido neural. En uno de tales aspectos, los miembros de unión específica radiomarcados, particularmente anticuerpos y fragmentos de los mismos, son útiles en técnicas de cirugía radioinmunoensayadas guiada, en las que se pueden identificar e indicar la presencia y/o ubicación de células nerviosas comprometidas lesionadas o moribundas o neuronas o tejido nervioso, antes de, durante o después de la cirugía para apuntar, identificar o eliminar tales células.
Se incluyen inmunoconjugados o proteínas de fusión de anticuerpos en las que los miembros de unión específica, particularmente anticuerpos y fragmentos de los mismos, de la presente invención están conjugados o unidos a otras moléculas o agentes que se incluyen adicionalmente, pero no se limitan a miembros de unión conjugados con un fármaco neuroactivo, agente de ablación química, toxina, inmunomodulador, citoquina, agente citotóxico, agente quimioterapéutico o fármaco.
La presente descripción incluye un sistema de ensayo que puede prepararse en forma de un kit de prueba para el análisis cuantitativo de la extensión de la presencia de, por ejemplo, neuronas dañadas, comprometidas o lesionadas o para cuantificar neuronas en una muestra. El kit de sistema o de prueba puede comprender un componente marcado preparado por una de las técnicas radiactivas y/o enzimáticas tratadas en este documento, el acoplamiento de una etiqueta para el anticuerpo, y uno o más reactivos inmunoquímicos, opcionalmente al menos uno de los cuales es un componente libre o inmovilizado o su pareja de unión.
Los anticuerpos y en una realización particular anticuerpos recombinantes que comprenden las secuencias de cadena pesada y/o ligera presentadas en las Figuras 5 y 6 en el presente documento, o fragmentos activos de los
mismos, y anticuerpos de cadena única, recombinantes o sintéticos derivados de los mismos, en particular que comprenden las secuencias de la región de CDR representadas en las Figuras 5 y 6, se pueden preparar en composiciones farmacéuticas, incluyendo un vehículo, excipiente o diluyente adecuado, para la administración en casos en los que la terapia es apropiada, tal como para tratar o mejorar las condiciones de o enfermedades que implican el compromiso de las neuronas, daños, lesiones o la muerte. Tales composiciones farmacéuticas también pueden incluir procedimientos de modulación de la vida media de los anticuerpos o fragmentos mediante métodos conocidos en la técnica tales como pegilación. Tales composiciones farmacéuticas pueden comprender además anticuerpos adicionales o agentes terapéuticos.
La presente descripción también incluye anticuerpos y fragmentos de los mismos, que están unidos de forma covalente o asociados de otro modo con otras moléculas o agentes. Estas otras moléculas o agentes incluyen, pero no se limitan a, las moléculas (incluyendo anticuerpos o fragmentos de anticuerpos) con distintas características de reconocimiento, toxinas, ligandos, agentes neuroactivos, y agentes quimioterapéuticos. En un aspecto adicional los anticuerpos o fragmentos se pueden usar para orientar o dirigir moléculas terapéuticas u otros agentes, por ejemplo a las moléculas diana o agentes a las neuronas, por ejemplo a las neuronas corticales, las neuronas del hipocampo, células ganglionares de la retina o de células granulares del cerebelo, incluyendo por los sitios de la herida, sitios isquímicos, sitios de tumor, áreas inflamatorias o lesiones neurodegenerativas. Los anticuerpos o fragmentos se pueden utilizar para orientarse a moléculas terapéuticas directos u otros agentes, por ejemplo a las moléculas diana o agentes a los sitios de isquemia cerebral o derrame cerebral.
Otros objetos y ventajas serán evidentes para los expertos en la técnica a partir de una revisión de la siguiente descripción detallada, que procede con referencia a los siguientes dibujos ilustrativos, y las reivindicaciones acompañantes.
BREVE DESCRIPCIÓN DE LOS DIBUJOS
FIGURA 1: IgM humanos se une a la superficie de múltiples tipos de neuronas. sHIgM42 (A) y rHIgM12 (C) se unen a la superficie de neuronas corticales vivas que co-expresan los neurofilamentos (NF) (B & D). Las células se mantuvieron en hielo durante una incubación de 10 min con 10 pg/ml IgM en los medios. Se fijaron las células, se lavaron y se detectó IgM humana con una cadena mu anti-humana Fab'2-FITC Ab secundaria. NF interna está presente como un patrón segmentado característico. Las células corticales se cultivaron durante 6 días en medios Neurobasal B27 en cubreobjetos de vidrio recubiertos de poliornitina. No todas las neuronas corticales expresan NF todavía, pero las IgM se unen a todas las neuronas en el cultivo. rHIgM12 (E verde) y sHIgM42 (F verde) se unen a la superficie de las neuronas del hipocampo del ratón vivo que co-expresa neurofilamentos (NF, rojo). Neuronas hippocampales se cultivaron durante 1 semana en los medios Neurobasal B27 en plástico revestido de polilisina. Las células aisladas de una resección del lóbulo temporal humano se cultivaron en DMEM/F12/N2/B27 durante 21 días. rHIgM12 unido a la superficie de las células (G) que también fueron positivos para p III tubulina (F, anti-p III de Promega).
FIGURA 2. IgM humana etiquetar la superficie de células granulares del cerebelo en patrones distintos. Rata de células granulares del cerebelo en 1 día en cultivo se incubaron in vivo con 10 pg/ml del suero humano derivado Ig Ms, sHIgM12 o sHIgM42, en medios de cultivo durante 15 min en hielo. Después del lavado y la fijación con paraformaldehído al 4%, las células se incubaron a temperatura ambiente con un anticuerpo humano anti-conjugado FITC cadena mu y la imagen usando un microscopio de inmunofluorescencia. El patrón de la etiqueta de la membrana neuronal utilizando estos dos anticuerpos es diferente. Áreas cortas de membrana de neuritas están vinculadas por sHIgM12 que resulta en un patrón claramente punteado. Las áreas más grandes de la membrana de neuritas están vinculadas por sHIgM42 que resulta en un patrón segmentado. La ampliación es 400x.
FIGURA 3. IgMs, rHIgM12 y sHIgM42 humanos de unión neuronal protegen a las neuronas corticales en cultivo a partir de peróxido inducido por la muerte celular. Las neuronas de ratón a los 3 días en cultivo se trataron con 500 nM de H2 O2 o solución salina con o sin 10 pg/ml de IgM humanos durante 30 min. Las células se lavaron con medio neurobasal B27 frescas y 24 horas más tarde la expresión de la caspasa-3 detectada por 96 pocillos triplicados tratados usando un kit de ensayo de FLICA caspasa-3 (Immunochemistry Technologies 935). Las barras indican el % de protección a partir de la activación de caspasa 3.
FIGURA 4 ilustra el vector amplificable para la producción de anticuerpo recombinante. Este es un mapa del vector usado para generar anticuerpo recombinante basado en sHIgM22 (rHIgM22), que fue modificad para la producción de IgM12 recombinante y IgM42. Para la expresión en células CHO, el promotor de VH fue reemplazado con el promotor de E1A. La región constante de cadena ligera CA se sustituyó por una región constante de la cadena ligera kappa Ck.
FIGURA 5 representa IgM12 de aminoácidos del anticuerpo y la secuencia de ácido nucleico, en el vector recombinante. La secuencia de aminoácidos de la cadena pesada y la codificación de secuencia de ácido nucleico está en gris y cada una de las Vh, y las secuencias de región constante CH1, CH2, CH3 y CH4 están señalizadas. La secuencia de aminoácidos de la cadena ligera y la codificación de secuencia de ácido nucleico está en CL púrpura y se indican las secuencias de VL tanto variables como constantes (kappa). Se observan secuencias de la región CDR de las regiones pesadas y ligeras variables de la cadena. La cadena J humana se indica en la secuencia de aminoácidos y de ácido nucleico.
FIGURA 6 representa aminoácido de anticuerpo IgM42 y la secuencia de ácido nucleico, en el vector recombinante. La secuencia de aminoácidos de la cadena pesada y la codificación de secuencia de ácido nucleico está en gris y cada una de las secuencias de región Vh y constante CH1, CH2, CH3 y CH4 se proporcionan e indican. La secuencia de aminoácidos de la cadena ligera y la codificación de secuencia de ácido nucleico está en CL púrpura y se indican las secuencias VL tanto variables como constantes (kappa). Se observan secuencias CDR de la región para las regiones pesadas y ligeras variables de la cadena.
FIGURA 7. rHIgM12 recombinante mejora los déficits en ratones con enfermedad TMEV inducida. La media de la actividad nocturna espontánea (cuando los ratones están normalmente activos) se ve alterada por 3 semanas después de una dosis ip única de 100 pg de sHIgM12. Grupos de 5 ratones SJL a los 90 días después de la infección por TMEV se trataron con rHIgM12 (barras rojas), o un control de IgM humana (barras negras) y se monitorearon durante 3 días a la semana como grupos en cajas de monitorización de la actividad AccuScan. La media 6 SEM de la actividad horizontal nocturna durante 3 períodos nocturnos se compararon con la actividad nocturna para el tratamiento 3 noches antes de IgM. Un aumento en la actividad nocturna en comparación con los niveles previos al tratamiento estaba presente a las 3 semanas a través de 7 semanas después del tratamiento con una sola dosis de rHIgM12 (P <0,01), pero no después del tratamiento con el control. Los niveles de actividad de los ratones emparejados por edad no infectados se representan gráficamente para comparación (barra verde).
FIGURA 8. Espectros MRS recogidos en el vástago de cerebro de ratón. Los paneles superiores indican el voxel desde el que se recibió la señal. El posicionamiento de esta región de interés utilizó marcas anatómicas. Una resonancia magnética muy corta se usa para obtener las imágenes en tres planos ortogonales para posicionar el 2.5 mm3 cubo sobre el tronco cerebral (panel superior). A 300 MHz, el espectro 1H del tronco cerebral de ratón (panel inferior). Colina (Cho), creatina (Cr) y picos de N-acetilo-aspartato (NAA) se identifican fácilmente. El espectro se referencia a agua a 4,7 ppm.
FIGURA 9. La disminución de los niveles de nAA en el tronco cerebral se correlaciona con el aumento de la pérdida de axón. 10 ratones SJL se estudiaron prospectivamente a partir de 0 a 270 días después de la infección con TMEV. MRS se obtuvo en el tallo cerebral de cada ratón y se determinó la concentración de NAA (parte superior). Una disminución gradual en NAA se registró en todos los ratones alcanzándose una caída significativa estadística a los 90 días después de la infección del virus, el punto en que se muestran grandes axones que se pierden por conteo directo en secciones transversales (2). Los niveles de NAA se mantuvieron bajos durante la duración de la enfermedad. Axones absolutos se contaron (parte inferior) en el nivel T6 de ratones no infectados (barra de color negro), 90 días (gris) y 270 días (blanco) después de la infección.
FIGURA 10. NAA en el tronco cerebral después de una dosis única de las IgM humanos. Grupos de 10-15 ratones SJL a los 90 días después de la infección TMEV, el tiempo cuando los niveles de NAA comienzan a caer, y la pérdida de axón grande es detectable se les dio una sola dosis de 100 pg de sHIgM12, sHIgM42, un humano IgM de control, o solución salina (PBS) MRS ip en el tronco cerebral se recogió antes y a las 5 y 10 semanas después del tratamiento. Niveles de NAA se calcularon a partir de estos espectros. En los grupos de sHIgM12 y sHIgM42 tratados NAA se aumentó en comparación con los niveles de pretratamiento después de 5 semanas (P = 0,005, P = 0,029) y a las 10 semanas (P <0,001, P = 0,037). En grupos de ratones tratados con control, los reactivos eran estables o mostraban una tendencia hacia abajo. El gráfico representa la media /- SEM de los niveles de NAA voxel de tronco cerebral.
FIGURA 11. IgM humanos que se unen a las neuronas no alteran la desmielinización, la remielinización o inflamación. Grupos de 5 ratones crónicamente infectados con TMEV se trataron con una única dosis de 100 pg de IgM humanos. Todos los grupos contienen el mismo grado de inflamación de la médula espinal (barras negras) y la desmielinización (barras rojas) a las 10 semanas después del tratamiento. Como se esperaba, los ratones tratados con unión de oligodendrocitos IgM, rHIgM22, mostraron una mayor remielinización (barras verdes). sHIgM12 y sHIgM42 no promueven la remielinización de la médula espinal. Esto sugiere que puede haber varias formas de proteger los axones in vivo.
FIGURA 12. Media vida en suero de IgM humanos de unión neuronal en la circulación de ratones CD-1 después de la inyección intravenosa. ELISA sandwich se utilizó para cuantificar la decadencia de la cadena mu humana a partir de sueros del ratón durante 48 h. La media de la DO405 de sustrato de fosfatasa alcalina (Sigma 104) se obtuvo a partir de los medios. Los sueros de 3 ratones por grupo y el error estándar se representan gráficamente.
FIGURA 13. rHIgM12 etiquetada con 35S entró en el cerebro y la médula espinal de ratones SJL infectados y no infectados con TMEV. rHIgM12 etiquetada con 35S se administró ip a ratones SJL infectados con TMEV durante 5 meses de edad igualada y no infectados. 4 h más tarde un par de ratones fueron perfundidos con solución salina, los tejidos se cosecharon de forma aguda, se pesaron, se picaron con una navaja de afeitar, y se mezclaron con 10 ml de fluido de centelleo (Ultima Gold LSC). Lo mismo se repitió a las 24 h. Después de 48 horas, permitiendo que el tejido se solubilizara, los viales se contaron durante 1 min en un contador de centelleo Beckman. Los recuentos totales de todo el cerebro y la médula espinal de cada ratón se representan gráficamente. Los recuentos de fondo con viales no tejido fueron 12 recuentos.
FIGURA 14. Diana IgM humana de unión a neuronas in vivo a las lesiones de la médula espinal. A ratones con desmielinización de médula espinal inducida por TMEV crónica se les administró 1,0 mg por vía intraperitoneal de sHIgM42 (A), rHIgM12 (B), o controlar IgM monoclonal humano (C). 4 horas más tarde los ratones fueron perfundidos con paraformaldehído al 4%, las médulas espinales se retiraron, se congelaron, se seccionaron y se inmunoetiquetaron utilizando fragmentos Fab'2 conjugados con FITC dirigidos contra la cadena
mu humana (Jackson Immunoresearch). Las médulas espinales de ratones que recibieron sHIgM42 o rHIgM12 contenían IgM humana en las lesiones. Poco control de IgM humana fue encontrado en las lesiones. Los paneles inferiores son tinción H/E para visualizar las células inflamatorias y lesiones. Llegamos a la conclusión de que ciertos IgM humanos pueden cruzar la barrera hematoencefálica en el modelo de TMEV.
FIGURA 15. IgM humanos de unión de la neurona localizan a axones de neurofilamentos (NF) en las lesiones de la médula espinal. A los ratones con desmielinización de médula espinal inducida por TMEV crónica se les administró 1,0 mg por vía intraperitoneal de IgM y fueron matados 4 horas más tarde. Las médulas espinales se congelaron, se seccionaron longitudinalmente y se inmunoetiquetaron con fragmentos FITC conjugados de cadena mu anti-humana Fab'2 (verde) y ant-NF (SMI-32 y 34, rojo) seguido de una TRICT secundario conjugado. Las imágenes confocales fusionadas confirmaron la co-localización de sHIgM42 (A) con largas NF fibras (B), se fusionaron en (C). rHIgM12 (D, verde) también se co-localizó con estructuras NF (D, rojo), tales como haces de fibras axón de cruz de corte.
FIGURA 16. rHIgM12 y sHIgM42 no exacerben las puntuaciones clínicas de EAE. Grupos de 10 ratones C57BL6 con péptido MOG indujeron EAE se les dio una sola dosis de 100 pg i.v. de rHIgM12, sHIgM42, el control de IgM humana o solución salina en el momento en que cada ratón alcanzó una puntuación clínica de 1 (cola cojera) y se controló cada dos días hasta 28 días después de la inmunización. Los gráficos son la /- SEM media de la puntuación clínica media de cada grupo de tratamiento. ANOVA sobre los rangos de las puntuaciones clínicas medias no indicó ninguna diferencia entre los grupos (P = 0,14).
FIGURA 17. rHIgM12 y sHIgM42 no exacerben la patología de la médula espinal EAE. Al final del estudio en la Figura 13 se eliminaron las médulas espinales, se cortaron en bloques de 1 mm, secciones transversales cortadas de cada tercer bloque consecutivo y se tiñeron con ericromo para visualizar patología. 10 secciones transversales por ratón (40 cuadrantes) se calificaron a ciegas. No hubo diferencias en la inflamación (p = 0,825) o la desmielinización (P = 0,766) entre los grupos.
FIGURA 18. rHIgM12 promueve la emergencia de neuritas Neuronas del hipocampo E15 se cultivaron en poli-D-lisina (PDL) (A) o rHIgM12 (B) -cubreobjetos de vidrio recubierto de nitrocelulosa. 12 horas después de la siembra de las células, estas neuronas se fijaron y se tiñeron con anticuerpo p3-tubulina (A1 y b 1, verde) y faloidina Texas-rojo (A2 y B2). A3-B3 son superposiciones de A1-A2 y B1-B2, donde los núcleos se tiñeron con DAPI (azul). C, D y E muestran la medición del total (C, p = 0,0056), el más largo (D, p <0,0001) y 2° más largo (E, p = 0,0782) longitudes de neuritas. Las neuronas que crecen en rHIgM12 tienen un menor número de neuritas (F, p <0,0001), y la mayoría son neuronas de etapa 3 (G) en comparación con las neuronas sembradas sobre sustratos PDL de control (72% vs. 18%), que tienen múltiples neuritas simétricas. Los análisis estadísticos muestran la media SEM (t-prueba no apareada).
FIGURA 19. rHlgM12 impulsa formación axón Neuronas de hipocampo 12 horas después de la siembra sobre sustratos de PDL (A), p Dl laminina (B) o rHIgM12 (C) se tiñeron con Tau1 (A1-C1, verde) o MAP2 (A2-C2, azul). A3-C3 son superposiciones de A1-C1 y A2-C2, y F-actina (rojo) se marcó con faloidina rojo Texas. DF muestran la intensidad TAU1 relativo a lo largo de las neuritas más largas de cuerpo celular a la del cono de crecimiento en A1-C1 marcada por líneas de trazos. La tinción Tau1 se enriquece de manera asimétrica en la parte distal de la neurita más larga en las neuronas de etapa 3 cultivadas en sustratos tanto rHIgM12 como laminina.
FIGURA 20. Unión de rHIgM12 reorganiza las membranas neuronales Neuronas del hipocampo DIV3 se tiñeron de modo triple con rHIgM12, la toxina de cólera B (CTB, Alexa Fluor 594) y filipina después de la fijación (A). rHIgM12 (A1) etiqueta de manera uniforme la superficie de la neurona, con un patrón similar al de GM1 (A2) o colesterol (A3) etiquetado por CTB o filipina respectivamente. Sin embargo, después del tratamiento con rHIgM12 durante 30 min a 37°C, unido a la membrana de rHIgM12 agregada en la superficie de la neurona (B1-C1). El tratamiento con resultados rHIgM12 en la agrupación de colesterol (B2) y GM1 (C2), que co-localiza con la rHIgM12 agregada (B3-C3). Los paneles inferiores en la parte inferior de B y C son un aumento mayor de las regiones en caja, mostrando colocalización de rHIgM12 agregada (verde) con el colesterol en clúster (azul) o GM1 (rojo) en la región del cono de crecimiento.
FIGURA 21. rHIgM12 co-localiza con GM1 en microdominios de membrana de la neurona A . Neuronas del hipocampo DIV1 se fijaron con paraformaldehído al 4% primero y después se tiñeron por rHIgM12. rHIgM12 etiqueta estructuras de punto más pequeñas. B. Neuronas del hipocampo vivas DIV1 se enfriaron a 4°C durante 30 min y después se marcaron con rHIgM12, seguido de tinción con anticuerpo anti p-III-tubulina después de la fijación. rHIgM12 mancha formaciones de tinción punteada mucho más grandes en la superficie de las neuronas. C. Neuronas del hipocampo vivo DIV3 fueron tratadas con metilo-pciclodextrina durante 30 min a 37°C para agotar colesterol y después se enfrió a 4°C. Las neuronas fijadas fueron teñidas con rHIgM12 (verde) y la toxina B del cólera (rojo). rHIgM12 se une a los puntos lagrimales más grande que co-localiza GM1 etiquetado por la toxina del cólera B. El panel inferior muestra un aumento mayor de las regiones en caja.
FIGURA 22. rHIgM12 se une a las balsas de lípidos Se permitió que neuronas corticales vivas DIV7 se unieran rHIgM12 o controlaran anticuerpo IgM a 4°C durante 30 min. A. Las neuronas se lavaron tres veces después de la unión del anticuerpo y se lisaron en tampón que contenía 0,5% de NP-40. Los lisados se separaron por microcentrifugación a 4°C, y tanto los sobrenadantes como los sedimentos se transfirieron con anticuerpo secundario IgM anti-humano. rHIgM12 se distribuye tanto en el sedimento como el sobrenadante en comparación con el anticuerpo IgM de control, que no se une a las neuronas y va al lavado. El panel inferior muestra un gel teñido con Coomassie separado cargado con la misma cantidad de proteína. B. Las neuronas se lisaron en tampón que contiene 1% de Triton X-100 se fraccionaron mediante ultracentrifugación en gradientes de sacarosa a 4°C. Las
fracciones resultantes se transfirieron secuencialmente para rHIgM12, caveolina-1, receptor de transferrina, p3-tubulina (B3-Tub), y beta-actina con anticuerpos específicos. rHIgM12 localiza a la fracción de baja densidad que contiene caveolina-1 y p3-tubulina. Las fracciones de mayor densidad contienen receptor de transferrina, p3-tubulina y actina. Cierta rHIgM12 localiza al sedimento insoluble detergente, que se compone principalmente de proteínas del citoesqueleto enriquecidas en la tubulina. La mayoría de actina va a fracciones de mayor densidad solubles detergentes.
FIGURA 23. rHIgM12 asociados con los microtúbulos A. Neuronas del hipocampo DIV1 fueron tratadas primero con rHIgM12 durante 30 min a 37°C, luego se fijan simultáneamente y se extraen con tampón que contenía 4% de paraformaldehído y 0,1% de Triton X-100 y seguido de tinción para rHIgM12 (verde), p3-tubulina (azul) y F-actina (rojo). rHIgM12 une a las moléculas de detergente insolubles que co-localizar con los microtúbulos fasciculadas a lo largo de los ejes de neuritas (A 2 y 4) y en el dominio central del cono de crecimiento, pero no con F-actina localizada en el crecimiento periferia cono (A 3 y 4). B. DIV7 neuronas corticales se les permitió unirse rHIgM12 primero, y luego se lisaron en 0,5% de NP-40. Los sobrenadantes se sometieron a co-inmunoprecipitación. Tanto rHIgM12 como p3-tubulina fueron co-inmunoprecipitados entre sí. FIGURA 24. Los mecanismos propuestos por los cuales rHIgM12 promueve la formación de axón A . Membranas neuronales contienen microdominios tanto de balsa como no de balsa. Los microdominios de balsa, que se distribuyen de manera uniforme en las membranas neuronales, se segregan en dos reservas. Uno de ellos se asocia con los microtúbulos B. La unión de rHIgM12 induce la agrupación de los microdominios de balsa, que promueve la estabilización de los microtúbulos y de las membranas de las neuronas de anclaje. C. Durante el crecimiento de neuritas, los conos de crecimiento exploran el medio ambiente y agrupan microdominios de balsa mediante la interacción con rHIgM12. Como resultado, los microtúbulos se estabilizan en los sitios rHIgM12 de contacto y anclan las membranas de las neuronas, lo que mejora el crecimiento de la periferia cónica a la transición de dominio central como se observa en las Figuras 3 y 6 donde el rHIgM12 aplicado por baño se agrega en el dominio central del cono de crecimiento.
FIGURA 25. rHIgM12 no se asocia con la tubulina directamente A. Células de neuroblastoma N2A se tiñeron con rHIgM12 (A1, verde) y p3-tubulina (A2, rojo). Los núcleos se marcaron con DAPI (A3, azul). Células N2a son negativas para la tinción rHIgM12 pero expresan 133-tubulina. B. Los sobrenadantes de los lisados de células N2A se incubaron con rHIgM12 y moléculas asociadas con rHIgM12 fueron derribadas por proteína-L agarosa y sujetos a transferencia de Western. rHIgM12 no está asociado con el sedimento. p3-tubulina no se tira hacia abajo por rHIgM12, aunque se detecta en los sobrenadantes.
FIGURA 26. rHIgM12 tira hacia abajo de actina, pero no colocaliza con F-actina A . Neuronas del hipocampo DIV1 in vivo se tiñeron a 4°C con rHIgM12 (A1, verde) primero, y F-actina (A2, rojo) se marcó con faloidina rojo texas después de la fijación. En la región del cono de crecimiento, F-actina se redujo y se enriqueció en el dominio central, mientras que rHIgM12 uniformemente tiñó las estructuras puntos lagrimales en la superficie del cono de crecimiento, lo que etiquetó el mismo borde del cono de crecimiento. B. Una pequeña cantidad de actina se tira hacia abajo por rHIgM12, y ninguno de los tres anticuerpos anti-actina probados funcionó en inmunoprecipitación. La banda en la misma posición como p3-tubulina es la cadena pesada de IgG indicada por la punta de flecha.
FIGURA 27 (a) Las membranas celulares se reconstituirán en una película de oro delgada perforada con miles de nanoporos (200 nm de tamaño). (b) una película de oro suspendida está en contacto con una solución de tampón en ambos lados.
FIGURA 28. Un anticuerpo humano que se une a las neuronas promueve la extensión de neuritas. Las neuronas se sembraron sobre sustratos revestidos con IgM humana, se dejaron crecer durante 24 h y se inmunoetiquetaron con anti-neurofilamentos. La sHIgM42 indujo excrecencia (A) en comparación con una IgM no permisiva humana (B).
FIGURA 29. Un anticuerpo humano que se une a las neuronas agrupa los dominios de membrana. Inmunoetiquetado uniforme sobre soma y los axones se realizó a 0°C (A). Cuando las células se calientan a 15°C durante 15 min sHIgM42 está localizada a parches más discretos en la superficie celular de los axones (B).
FIGURA 30. Extensión de neuritas promueve dianas de anticuerpos humanos in vivo a las áreas de lesión. A ratones con SCI crónica se les dio anticuerpo ip, médulas espinales se extrayeron 4 h más tarde y IgM humana se detectó en secciones transversales de la médula espinal. Áreas lesionadas de la médula espinal de un anticuerpo de ratón demuestran fibras paralelas que se unen a un IgM anti-humano fluorescentemente etiquetado (A). Áreas lesionadas de la médula espinal de un ratón con un control humano de IgM no contienen IgM humana (B).
FIGURA 31. Lesión por aplastamiento del cordón espinal. Las fotomicrografías muestran el aspecto típico de H & E manchado de la médula espinal de los ratones de control (A) o ratones 30d tras compresión de 8 g Fejota clip (B). Lesión de compresión está asociada con extensa infiltración de células inflamatorias, la pérdida de neuronas motoras.
FIGURA 32. Ejemplo de actividades nocturnas espontáneos de ratones infectados con TMEV. A) Grabaciones completas originales de la actividad horizontal durante el período de 64 días. Actividades nocturnas altas y diurnas bajas son fáciles de apreciar. Debido a que los ratones son normalmente activos durante la noche se seleccionaron actividades nocturnas para el análisis (18.00-6.00). Sin embargo, mediante inspección visual de grabaciones comprimidas sin filtrar, ya sea en actividad tanto horizontal (B) como vertical (C) puede ser difícil identificar verdaderos cambios a largo plazo.
FIGURA 33. Una dosis única ip de anticuerpo de unión a neurona-(rHIgM12) mejora la actividad horizontal en los
ratones SJL con infección crónica. Tres grupos de 5 ratones SJL en 90dpi se colocaron en cajas de monitoreo de actividad AccuScan. Mediciones de línea de base se recogieron a lo largo de 8 días. Después se monitorearon los ratones de tratamiento continuamente a lo largo de 8 semanas. Los paneles A, C y E corresponden a la actividad y B horizontal, D y F corresponden a la actividad vertical. A, B) Actividades nocturnas promedias se presentaron como contenedores para cada día, lo que sugería la mejora de la actividad horizontal sólo en el grupo tratado con rHIgM12; C) filtrado gaussiano (GB = 2 días) y E) sexto orden de ajuste polinomial de valores Z estandarizados dio lugar a una mejora clara y distinta en los ratones tratados con rHIgM12. B, D y F) La actividad vertical no se vio afectada por ningún tratamiento. Actividad (escala vertical) es el número de interrupciones del haz por hora. Parámetro GB puede ser interpretado como un valor (en días) de la mitad de la anchura del filtro en la altura media. Con el aumento del valor GB disminuye la desviación estándar. 95% de intervalos de confianza de fijación por puntos se dan para cada día.
FIGURA 34. Comparación de tres tratamientos en 90dpi utilizando valores de modelo predichos. El análisis estadístico se realizó para cada día antes y después del tratamiento. La actividad nocturna horizontal de ratones tratados con rHIgM12 en comparación con el control IgM y los ratones tratados con solución salina divergieron significativamente/se mejoraron en el día 7 (p = 0,045) y el día 11 (p = 0,042) post-tratamiento, respectivamente (indicado por flechas negras). 95% de intervalos de confianza se dan para cada día.
FIGURA 35. rHIgM12 mejora tanto la actividad horizontal como vertical en ratones SJL cuando se administra temprano en la enfermedad. Tres grupos de 5 ratones SJL en 45dpi se colocaron en cajas de monitoreo de actividad AccuScan. Las medidas de línea de base se recogieron a lo largo de 8 días. Después del tratamiento, los ratones de tratamiento se controlaron continuamente durante 8 semanas. Paneles A, C, E y G corresponden a la actividad horizontal y B, D, F y H se corresponden con la actividad vertical. A, B), grabaciones originales sin filtrar para la actividad horizontal y vertical; C, D) actividades nocturnas promedias presentadas como contenedores para cada día, E, F) filtrado gaussiano (GB = 2 días) y G, H) sexto ajuste polinómico de valores Z normalizados reveló mejora tanto en la actividad horizontal como vertical en grupo tratado con rHIgM12. Grupos de control de IgM y de salina mostraron niveles similares de actividad motora durante todo el estudio. Los intervalos de confianza del 95% de punto de fijación se dan para cada día.
FIGURA 36. Comparación de tres tratamientos en 45dpi utilizando valores de modelo predichos. El análisis estadístico se realizó para cada día antes y después del tratamiento. A, C) la actividad nocturna horizontal de ratones tratados con rHIgM12 en comparación con el control IgM y de los ratones tratados con solución salina divergieron significativamente en el día 13 (p = 0,015) y el día 9 p = 0,036) después del tratamiento, respectivamente. B, D) la actividad vertical de ratones tratados con rHIgM12 se divergió/mejoró en el día 14 (p = 0,019) y el día 6 (p = 0,037) post-tratamiento, respectivamente. Las flechas negras indican días con el inicio de la mejoría significativa. Los 95% de intervalos de confianza se dan para cada día.
FIGURA 37. Ratones normales de SJL no infectados tiene actividad espontánea altamente variable que no está influenciada por ningún tratamiento. Tres grupos de 5 ratones SJL no infectadas se colocaron en cajas de monitorización de la actividad AccuScan. Mediciones de línea de base se recogieron más de 8 días. Después del tratamiento, los ratones se controlaron continuamente durante 8 semanas. Ninguno de los tratamientos indujo cambios en actividad horizontal (A, C, E y G) o vertical (B, D, F y H). 95% de intervalos de confianza se dan para cada día.
FIGURA 38. Una sola dosis de anticuerpo humano incrementó la supervivencia en ratones transgénicos mutantes SOD1 G86R. Los ratones fueron tratados con una dosis única de rHIgM12 o PBS a los 55 días de edad. Algunos ratones se dejaron sin tratar. Los ratones fueron seguidos hasta moribundo y luego se sacrificaron para la patología. Algunos ratones murieron a causa de la enfermedad. Todos los ratones tenían una profunda pérdida de peso antes de la muerte. La supervivencia se representa gráficamente mediante las curvas de Kaplan-Meir y la significación estadística se determinó mediante la prueba de Mantel-Cox no paramétrico. Hubo un aumento en la supervivencia en ratones tratados con rHIgM12 en comparación con los ratones tratados con PBS o no tratados, P = 0,008. Todos los análisis y el tratamiento se realizaron de forma ciega de tal manera que el investigador que tome la decisión de sacrificar los ratones no tenía conocimiento de los grupos de tratamiento. Un total de 45 ratones fueron estudiados en este análisis. Ratones de tipo salvaje SOD /+ tenían una supervivencia normal y sin pérdida de peso (datos no mostrados).
FIGURA 39 representa la pérdida de peso en ratones G86R SOD1 como una función del tiempo en ratones administrados con una sola dosis de 200 pg de rHIgM12 a los 50 días de edad en comparación con los ratones de control administrados con PBS. Ratones administrados con rHIgM12 mostraron una menor pérdida de peso durante un período de 60 días después de la inyección única de anticuerpo.
FIGURA 40. Una sola dosis de anticuerpo humano disminuyó degeneración axonal de la médula espinal en ratones transgénicos mutantes SOD1 G86R. Los ratones se trataron con una sola dosis de 250 pg de rHIgM12 o PBS a los 55 días de edad. Algunos ratones se dejaron sin tratar. El número de verticilos de mielina, característico de axones en degeneración, fueron contados en la médula espinal en el nivel medio torácico. El número de axones en degeneración se redujo en ratones mutantes de la SOD1 tratados con rHIgM12 en comparación con PBS tratado (P = 0,003, prueba T) y ratones no tratados. Tenga en cuenta que el número de axones en degeneración en la médula espinal de ratones de tipo salvaje SOD -/- es mínimo en comparación con ratones mutantes SOD1 /+. En el momento del sacrificio, los ratones se perfundieron con fijador de Trump (4% de formaldehído, 0,1% de gluataraldehído) y la médula espinal en bloques de 1 mm. Los bloques fueron incrustados en plástico de araldito y secciones de 1 micra desde el nivel T6 se tiñeron con diamina para fenileno para visualizar envoltura de mielina. Se muestran secciones de la médula espinal de SOD-/- de tipo salvaje
(superior derecha) y muíante SOD1 /+ (inferior derecha).
FIGURA 41. Una sola dosis de anticuerpo humano conservó neuronas tanto anterior como posterior de la médula espinal de ratones transgénicos mutantes SOD1 G86R. Los ratones se trataron con rHIgM12 o PBS a los 55 días de edad. Algunos ratones se dejaron sin tratar. El número de células de cuerno anterior y posterior, se tiñó con un anticuerpo para NeuN que específicamente etiqueta neuronas, se contaron en las secciones de parafina de la médula espinal en el un nivel torácico medio. El número de células de cuerno anterior y posterior en ratones tratados con rHIgM12 fue significativamente mayor (P = 0,0025 y P = 0,018 por prueba T) que en los ratones tratados con PBS o aquellos no se tratan. El número de células de cuerno anterior y posterior en ratones mutantes de SOD tratados con rHgM12 fueron similares a los observados en ratones de tipo salvaje de SOD -/-.
FIGURA 42. El anticuerpo humano recombinante, rHIgM12, se une a las neuronas a través de las especies. Neuronas corticales de ratón (A, B, C, D), neuronas del hipocampo de ratón (E, F) y neuronas corticales humanas (G, H) se tiñeron in vivo con rHIgM12 (A, C, E y G) y las mismas células se co-etiquetaron con anticuerpos para los neurofilamentos (B, D), o beta tubulina (F, G). rHIgM12 se detectó con un anticuerpo secundario marcado con fluorescencia.
FIGURA 43. La isquemia cerebral en ratones CD-1. A. El barrido de imagen DWI-MRI que muestra el área de la isquemia cerebral (región hiperintensa) a las 24 h. B. daño isquémico en el cerebro del ratón después de una isquemia cerebral (región pálida circular en el hemisferio derecho del cerebro). C. Sección H&E en x4 que muestra el tejido dañado (resaltado en negro). Figura 44. Una dosis ip única de anticuerpo de unión a neurona-(rHIgM12) mejora las actividades horizontal (A) y vertical (B) en ratones CD1 sometidos a isquemia cerebral fototrombótica. Los ratones fueron tratados con una dosis única de 200 pg rHIgMl2 (n = 5) o PBS (n = 5). La actividad se mejoró del día tres en el grupo tratado rHIgM12. C. Los ratones fueron escaneados en un pequeño sistema animal MRI para medir el volumen total de la isquemia cerebral que se expresa como mm3 en el gráfico.
Figura 45. Métricas MT-MRI demuestran un mejor resultado. Los ratones fueron sometidos a isquemia cerebral y se trataron con rHIgM12 o PBS. Tres días después de la lesión isquémica, se calcularon mapas MTR (panel A) de los hemisferios isquémicos (línea roja) y contralaterales (línea verde) del cerebro. El grupo tratado con rHIgM12 demostró aumento relativo de MTR dentro de la ROI accidente cerebrovascular (panel B, barras azules representan los valores calculados en el lado de control) y la disminución de volumen sistólico (panel C) en comparación con el grupo tratado con PBS.
Figura 46. Análisis de transferencia Western de las neuronas corticales (CN) cultivadas en poli-D-lisina durante 5 días y se incubaron durante 4-44 h bajo condiciones hipóxicas (2,7% de O2 , 5% de CO2 ). Transferencia representativa que muestra los niveles de caspasa 3 escindida después del tratamiento hipóxico durante 4 h, 21 h y 44 h en comparación con el control (44 h, 0 h de incubación en la transferencia).
Figura 47 representa gráficos de barras que muestran el análisis cuantitativo de transferencias Western a partir de tres experimentos independientes con MBP marcador de mielina y el marcador de apoptosis escindidos Capasa-3. Cultivos gliales mixtos se cultivaron durante 4 días en FBS (10%) que contiene el medio y se trataron durante 7 días en medio definido químicamente con la remielinización de promoción de IgM rHIgM22, mAb rHIgM12 regenerativa, isotipo controla sHIgM116 (10 pg/ml cada uno) o solamente medio. Medio con anticuerpos frescas se intercambió cada dos días. Al final de 4 días, se evaluaron los cultivos para MBP o caspasa-3 mediante transferencia Western, se cuantificó mediante OD/área, y se representaron gráficamente.
Figura 48 proporciona transferencias de Western representativa que muestra los niveles de varias proteínas y marcadores: células progenitoras de oligodendrocitos (OPC) marcadores PDGFaR, marcadores de mielina CNPasa y MOG, marcadores microgliales CD68 y Lyn, marcador de proliferación Ki-67 y la histona H3, caspasa-3 escindido por marcador de apoptosis y marcador de ciclo celular pRbS795. Co-cultivos OPC-microgliales se cultivaron en medio de astrocitos acondicionado con remielinación que promueve mAbs A2B5, 04, 01, 78,09, 94,03, rHIgM22, regenerativa mAb rHIgM12, controles de isotipo (hOjA, 5A5, ChromPure IgM, rHIgM42, sHIgM14, sHIgM24, sHIgM26) (10 pg/ml cada uno) o medio tal como se indica.
Figura 49 representa gráficos de barras que muestran el análisis cuantitativo de transferencias Western a partir de tres experimentos independientes con CD68 marcador de microglía activada y Lyn quinasa, una quinasa altamente abundante en microglia y en un grado menor en OPCs pero no en astrocitos. Los cultivos mixtos (cocultivos OPC-microgliales) se trataron con anticuerpos como se ha indicado anteriormente para la Figura 48 y se indica.
Figura 50 proporciona imágenes de inmunofluorescencia representativas que muestran niveles de CD68 (verde) y DAPI marcador nuclear (azul) en cultivos gliales mixtos antes del tratamiento mAb en el día 4 en cultivo y en el día 11 en cultivo después de 7 días de tratamiento con rHIgM12, rHIgM22 y control de isotipo sHIgM24 (10 pg/ml cada uno).
Figura 51 proporciona transferencias representativas de Western que muestra los niveles de hipoxia-inducible factor 1 (HIF1 a), marcadores de oligodendrocitos NG2, PDGF alfa, Olig-2, marcadores de mielina de MBP y CNPasa, marcador de astrocitos GFAP, marcador microglial CD68, marcadores del ciclo celular pRbS795 y el marcador de apoptosis escinde la caspasa-3, se utilizó con Beta-actina como control de carga. Cultivos gliales mezclados se cultivaron durante 4 días en FBS (10%) que contiene el medio y se trataron durante hasta 7 días adicionales en medio químicamente definido en condiciones de hipoxia (3% 02) en comparación con condiciones de normoxia (21% 02). Las proteínas se evaluaron mediante inmunotransferencia de tipo Western en el día 1, 3, 5 y 7 como se ha señalado.
Figura 52 representa el análisis cuantitativo de las transferencias de Western de la Figura 51, para proporcionar valores cuantitativos para la expresión y la comparación de las condiciones de hipoxia y normoxia para los
marcadores de proteínas.
Figura 53 proporciona transferencias de Western representativas de marcador de proliferación Ki-67, marcadores microgliales IBA-1, CD68, y iNOS, marcador de apoptosis exfoliado caspasa-3, marcador de astrocitos GFAP, marcadores del ciclo celular pRbS795 y pRbS807, marcadores de oligodendrocitos PDGFaR, NG2, Olig-1 y Olig-2, y marcador de mielina MBP. Cultivos gliales mixtos se cultivaron durante 4 días en condiciones de normoxia en FBS (10%) que contiene el medio y se trataron posteriormente durante 1-7 días en medio químicamente definido con anticuerpo regenerativo humano rHIgM12 o medio sólo bajo la hipoxia (3% 02).
Figura 54 proporciona transferencias de Western representativas que muestran los niveles de marcadores microgliales IBA-1 y CD68, marcador de apoptosis escindido caspasa-3, astrocitos GFAP marcador, los marcadores de oligodendrocitos PDGFaR, NG2, Olig-2 y marcadores de la mielina MBP y CNPasa en cultivos mixtos en día 7 cultivados en condiciones de normoxia y condiciones de hipoxia y tratados con anticuerpos rHIgM12 o rHIgM22 frente a medio solo. Glia mixta se cultivaron como se describe antes y se trató en medio químicamente definido por 7 días con rHIgM12, rHIgM22 o medio sólo bajo condiciones hipóxicas o normóxicas. La Figura 55 proporciona un análisis RT-PCR de diversos marcadores a partir de cortezas de ratones C57/BI6 neonatas de hipoxia y normoxia en P13. Ratones neonatales C57/BI6 fueron fomentados transversalmente por presas CD1 y se expusieron a hipoxia (10% 02) durante 4 días a partir de P3 a P7. Los ratones fueron expuestos posteriormente a aire de la habitación para 6 adicional y sacrificados a P13. Se aisló ARN a partir de cortezas en ambos grupos de tratamiento y se sondó para los marcadores de mielina PLP1, MOG, Mbp1, marcadores progenitoras de oligodendrocitos PDGFaR y Olig-1, marcador astrocítico GFAP, marcadores neuronales nestina y 133-tubulina, hipoxia inducible factor 1a y la proteínasa SSPN sarcospan. Los niveles de expresión se analizaron por RT-PCR y resultados normalizados a los niveles de p-acción.
Figura 56 proporciona gráficos que indican cantidad de anticuerpo radiomarcado rIgM12 - como se indica por la radiactividad medida como cpm/100 pl - en diversos tejidos de control (simulacro manipulado) y ratones con accidente cerebrovascular (protocolo Rose Bengala) como se denotan en cada panel. En 30 a 40 min después del accidente cerebrovascular de 10 millones de cpm de 35S etiquetados rHIgM12, aproximadamente 60 ug de IgM, se administró por vía intraperitoneal a cada ratón en un volumen de 500 ul de solución salina. A las 6, 12 y 24 h después se recogió sangre de tratamiento IgM de ratones, la vasculatura se lavó con 30 ml de solución salina y tejidos de forma aguda recogida. Para el análisis de radiomarcador en el tejido accidente cerebrovascular la corteza frontal derecha del cerebro que contiene el área de la lesión se eliminó como un bloque de 75 mg de tejido (denotado cerebro en los paneles). Una parte de otros tejidos se retiraró, se pesó y se procesó. CB = cerebelo, cordón = médula espinal.
DESCRIPCIÓN DETALLADA
De acuerdo con la presente invención puede emplearse biología molecular, microbiología y técnicas de ADN recombinante convencionales dentro de la experiencia de la técnica. Tales técnicas se explican completamente en la bibliografía. Véase, por ejemplo, Sambrook et al, "Molecular Cloning: A Laboratory Manual" (1989); "Current Protocols in Molecular Biology" Volúmenes I-III [Ausubel, RM, ed. (1994)]; "Cell Biology: A Laboratoriy Handbook" Volúmenes I-III [JE Celis, ed. (1994))]; "Current Protocols in Immunology" Volúmenes I-III [Coligan, JE, ed. (1994)]; "Oligonucleotide Synthesis" (MJ de la marcha ed 1984.); "Hibridación de ácidos nucleicos" [BD Hames y SJ Higgins eds. (1985)]; "Transcription and translation" [BD Hames y SJ Higgins, eds. (1984)]; "Animal Cell Culture' [RI Freshney, ed. (1986)]; "Células inmovilizadas y las enzimas" [IRL Press, (1986)]; B. Perbal, "A Practical Guide for Molecular Cloning" (1984).
Por lo tanto, si aparecen en el presente documento, los siguientes términos tendrán las definiciones expuestas a continuación.
A. TERMINOLOGÍA
El término "miembro de unión específica" describe un miembro de un par de moléculas que tienen especificidad de unión entre sí. Los miembros de un par de unión específica pueden derivarse de manera natural o total o parcialmente producidos sintéticamente. Un miembro del par de moléculas tiene un área en su superficie, o una cavidad, o un componente en su superficie, que se une específicamente a y por lo tanto es complementaria a una organización espacial y polar particular del otro miembro del par de moléculas. Así, los miembros del par tienen la propiedad de unirse específicamente entre sí. Ejemplos de tipos de pares de unión específica son antígenoanticuerpo, biotina-avidina, hormona-receptor de hormona, receptor-ligando, enzima-sustrato. Esta aplicación se refiere a reacciones de tipo antígeno-anticuerpo.
El término "anticuerpo" describe una inmunoglobulina ya sea natural o producida parcial o totalmente sintéticamente. El término también cubre cualquier polipéptido o proteína que tiene un dominio de unión que es, o es homólogo a, un dominio de unión de anticuerpos. CDR anticuerpos injertados también se contemplan por este término. Un "anticuerpo" es cualquier inmunoglobulina, incluyendo anticuerpos y fragmentos de los mismos, que se une a un epítopo específico. El término abarca anticuerpos policlonales, monoclonales, y los anticuerpos quiméricos, el último mencionado se describe en más detalle en la Patente de Estados Unidos Nos. 4.816.397 y 4.816.567. El
término "anticuerpo (s)" incluye un tipo salvaje de inmunoglobulina (Ig) molécula, que comprende generalmente cuatro cadenas de polipéptidos de longitud completa, dos cadenas pesadas (H) y dos cadenas ligeras (L), o un homólogo Ig equivalente (por ejemplo, un nanocuerpo de camélidos, que comprende sólo una cadena pesada); incluyendo mutantes de longitud completa funcionales, variantes, o derivados de los mismos, que retienen las características de unión de una molécula de Ig epítopo esencial, y que incluye anticuerpos de dominio duales específicos, biespecíficos, multiespecíficos, y duales variables; Moléculas de inmunoglobulina pueden ser de cualquier clase (por ejemplo, IgG, IgE, IgM, IgD, IgA e IgY), o subclase (por ejemplo, IgG1, IgG2, IgG3, IgG4, IgA1, e IgA2). También se incluyen dentro del significado del término "anticuerpo" son cualquier "fragmento de anticuerpo".
Un "fragmento de anticuerpo" se refiere a una molécula que comprende al menos una cadena de polipéptido que no es de longitud completa, incluyendo (i) un fragmento Fab, que es un fragmento monovalente que consiste de la luz variable (VL), pesada variable (VH), luz constante (CL) y 1 dominios constantes de pesada (CH1); (ii) un fragmento F(ab')2, que es un fragmento bivalente que comprende dos fragmentos Fa b unidos por un puente de disulfuro en la región bisagra; (iii) una porción de la cadena pesada de un fragmento Fab (Fd), que consiste en los dominios VH y CH1; (iv) un fragmento variable (Fv) fragmento, que consiste en los dominios VL y VH de un único brazo de un anticuerpo, (v) un anticuerpo de dominio (dAb) fragmento, que comprende un único dominio variable (Ward, ES et al., Nature 341, 544-546 (1989)); (vi) un anticuerpo de camélido; (vii) una región determinante de complementariedad aislada (CDR); (viii) un fragmento Fv de cadena sencilla en el que un dominio VH y un dominio VL están unidos por un engarce peptídico que permite que los dos dominios se asocien para formar un sitio de unión de antígeno (Bird et al, Science, 242, 423-426, 1988; Huston et al, PNAS EE.UU., 85, 5879-5883, 1988); (ix) un diacuerpo, que es un anticuerpo bivalente, biespecífico en el que los dominios VH y VL se expresan en una única cadena polipeptídica, pero usando un enlazador que es demasiado corto para permitir el emparejamiento entre los dos dominios en la misma cadena, forzando de ese modo los dominios a emparejarse con los dominios de complementariedad de otra cadena y creando dos sitios de unión a antígeno (WO94/13804; P. Holliger et al Proc Natl Acad Sci EE.UU. 90 6444-6448, (1993)); y (x) un anticuerpo lineal, que comprende un par de segmentos tándem Fv (VH-CH1-VH-CH1) que, junto con polipéptido s de cadena ligera complementariedad, forman un par de regiones de unión a antígeno ; (xi) fragmentos multivalentes de anticuerpos (dímeros scFv, trímeros y/o tetrámeros (Power y Hudson, J Immunol Métodos 242:. 193-204 9 (2000)); y (xii) otras porciones de longitud no completa de cadenas pesadas y/o ligeras, o mutantes, variantes, o derivados de los mismos, solos o en cualquier combinación.
Ya que los anticuerpos pueden modificarse en un número de maneras, el término "anticuerpo" debe interpretarse por cubrir cualquier miembro específico o sustancia de unión que tenga un dominio de unión con la especificidad requerida. Por lo tanto, este término cubre fragmentos de anticuerpos, derivados, equivalentes funcionales y homólogo s de anticuerpos, incluyendo cualquier polipéptido que compren de un dominio de unión de inmunoglobulina, ya sea natural o total o parcialmente sintético. Se incluye por lo tanto, moléculas quiméricas que comprenden un dominio de unión a inmunoglobulina, o equivalente, fusionadas a otro polipéptido. La clonación y expresión de anticuerpos quiméricos se describe en EP-A-0120694 y EP-A-0.125.023 y la Patente de los Estados Unidos Nos. 4.816.397 y 4.816.567.
Un "anticuerpo sitio de combinación" es la porción estructural de una molécula de anticuerpo compuesta por la cadena ligera o variable de cadena pesada y ligera y las regiones hipervariables que se une específicamente al antígeno.
La frase "molécula de anticuerpo" en sus diversas formas gramaticales, tal como se usa en este documento, contempla tanto una molécula de inmunoglobulina intacta como una porción inmunológicamente activa de una molécula de inmunoglobulina.
moléculas de anticuerpo ejemplares son moléculas intactas de inmunoglobulina, moléculas de inmunoglobulina sustancialmente intactas y aquellas porciones de una molécula de inmunoglobulina que contiene el paratopo, incluyendo aquellas porciones conocidas en la técnica como Fab, Fab 'F(ab')2 y F(v), cuyas porciones se prefieren para uso en los métodos terapéuticos descritos en el presente documento.
Los anticuerpos también pueden ser biespecíficos, en los que un dominio de unión del anticuerpo es un miembro de unión específica de la invención, y el otro dominio de unión tiene una especificidad diferente, por ejemplo, para reclutar una función efectora o similar. Los anticuerpos biespecíficos de la presente invención incluyen, en el que un dominio de unión del anticuerpo es un miembro de unión específica de la presente invención, incluyendo un fragmento de la misma, y el otro dominio de unión es un anticuerpo distinto o fragmento del mismo, incluyendo el de un anti-cáncer distinto o anticuerpo específico anti-tumor. El otro dominio de unión puede ser un anticuerpo que reconoce o se dirige a un tipo celular particular, como en un anticuerpo específico de la célula neuronal o glial. En los anticuerpos biespecíficos de la presente invención el dominio del anticuerpo de la invención de unión puede combinarse con otros dominios de unión o moléculas que reconocen los receptores de células particulares y/o modulan las células de una manera particular, tal como por ejemplo un modulador inmune (por ejemplo, interleucina), un modulador del crecimiento o citoquina (por ejemplo, factor de necrosis tumoral (TNF) y, en particular, la modalidad de TNF biespecífico o una toxina (por ejemplo,, ricina) o anti-mitótico o agente apoptótico o factor. Por lo tanto, los anticuerpos anti-FAP de la invención pueden ser utilizados para dirigir o diana agentes,
etiquetas, otras moléculas o compuestos o anticuerpos para sitios estroma, áreas particulares de la cicatrización de heridas, la inflamación, el cáncer o tumores.
La frase "anticuerpo monoclonal" en sus diversas formas gramaticales se refiere a un anticuerpo que tiene solamente una especie de sitio de combinación de anticuerpo capaz de inmunorreaccionar con un antígeno particular. Un anticuerpo monoclonal muestra así típicamente una única afinidad de unión para cualquier antígeno con el que inmunorreacciona. Un anticuerpo monoclonal también puede contener una molécula de anticuerpo que tiene una pluralidad de sitios de combinación de anticuerpo, cada uno inmunoespecífico para un antígeno diferente; por ejemplo, un anticuerpo biespecífico anticuerpo monoclonal (quimérico).
El término "dominio de unión a antígeno " describe la parte de un anticuerpo que comprende el área que se une específicamente y es complementaria a parte o la totalidad de un antígeno. Cuando un antígeno es grande, un anticuerpo puede unirse a una parte particular de sólo el antígeno, que parte se denomina un epítope. Un dominio de unión a antígeno puede ser proporcionado por uno o más dominios variables de anticuerpos. Preferiblemente, un dominio de unión a antígeno comprende una región variable de cadena ligera de anticuerpo (VL) y una cadena pesada de la región variable de anticuerpo (VH).
Los inmunoconjugados o proteínas de fusión de anticuerpos de la presente invención, en los que los anticuerpos, moléculas de anticuerpos, o fragmentos de los mismos, de uso en la presente invención están conjugados o unidos a otras moléculas o agentes incluyen adicionalmente, pero no se limitan a tales anticuerpos, moléculas, o fragmentos conjugados con un agente de ablación química, toxina, inmunomodulador, citoquina, agente citotóxico, agente quimioterapéutico, agente antimicrobiano o un péptido, de la pared celular y/o alterador de la membrana celular o medicamento.
El término "específico" puede usarse para referirse a la situación en la que un miembro de un par de unión específica no mostrará ninguna unión significativa a moléculas distintas de su pareja de unión específica. El término es aplicable cuando también, por ejemplo. Un dominio de unión a antígeno es específico para un epítopo particular que es llevado por un número de antígeno s, en cuyo caso el miembro de unión específica que lleva el dominio de unión al antígeno será capaz de unirse a los diversos antígenos que llevan el epítopo.
El término "comprende" se utiliza generalmente en el sentido de incluir, es decir que permite la presencia de una o más características o componentes.
El término "que consta esencialmente de" se refiere a un producto, en particular una secuencia de péptido, de un número definido de residuos que no se une covalentemente a un producto más grande. En el caso del péptido de la invención mencionado anteriormente, los expertos en la técnica apreciarán que las modificaciones menores en el N- o C-término del péptido sin embargo se pueden contemplar, tales como la modificación química de la terminal para añadir un grupo protector o similar, por ejemplo, la amidación del C-terminal.
El término "aislado" se refiere al estado en el que los miembros de unión específica de la invención, o ácido nucleico que codifica tales miembros de unión estarán de acuerdo con la presente invención. Los miembros y ácido nucleico estarán libres o sustancialmente libres de material con el que están asociados naturalmente tales como otros polipéptidos o ácidos nucleicos con los que se encuentran en su ambiente natural, o el entorno en el que se preparan (por ejemplo, el cultivo celular) cuando tal preparación es mediante tecnología de ADN recombinante practicada in vitro o in vivo. Los miembros y ácido nucleico pueden formularse con diluyentes o adyuvantes y aún así ser aislados para fines prácticos - por ejemplo los miembros normalmente se mezclarán con la gelatina u otros portadores si se usan para recubrir placas de microtitulación para su uso en inmunoensayos, o serán mezclados con excipientes farmacéuticamente aceptables vehículos o diluyentes cuando se utilizan en el diagnóstico o terapia.
Como se usa en el presente documento, "pg" significa picogramo, "ng" significa nanogramo, "ug" o " pg" significa microgramos, "mg" significa miligramo, "ul" o "pl" significa microlitro, "ml" significa mililitros, "I" significa litro.
Los términos "anticuerpo", "neurona de anticuerpo de unión", "anticuerpo IgM12", "anticuerpo IgM42", "anticuerpo 12", "anticuerpo 42", "sHIgM12", "sHIgM42", "rHIgM12", "rHIgM42" y cualquier variante no enumerado específicamente, se pueden usar en el presente documento, y como se utiliza en la presente solicitud y en las reivindicaciones se refieren a material proteico incluyendo las proteínas simples o múltiples, y se extiende a aquellas proteínas que tienen los datos de la secuencia de aminoácidos descrita en el presente documento y se presentan en las Figuras 5 y 6 y el perfil de actividades expuestas en el presente documento y en las reivindicaciones. En particular el anticuerpo IgM12, el anticuerpo 12, rHIgM12 particularmente se refiere a polipéptidos o anticuerpos o fragmentos, particularmente los anticuerpos recombinantes o fragmentos, que comprenden la secuencia presentada en la Figura 5. El anticuerpo IgM12, anticuerpo 12, rHIgM12 particularmente se refieren a polipéptidos o anticuerpos o fragmentos, particularmente los anticuerpos recombinantes o fragmentos, que comprenden secuencias de CDR de la región variable de cadena pesada expuesta en la SEQ ID NOS: 31-33 de cadena ligera y secuencias de CDR de la región variable establecidas en SEQ ID NOS: 34-36. Anticuerpo IgM12, el anticuerpo 12, rHIgM12 incluye anticuerpo que tiene la secuencia de la cadena pesada de la región variable de la SEQ ID NO: 1 y la secuencia de
cadena ligera de la región variable de la SEQ ID NO: 11. En particular, el anticuerpo IgM42, el anticuerpo 42, rHIgM42 particularmente se refiere a polipéptidos o anticuerpos o fragmentos, particularmente los anticuerpos recombinantes o fragmentos, que comprenden la secuencia presentada en la Figura 6. El anticuerpo IgM42, el anticuerpo 42, rHIgM42 particularmente se refiere a polipéptidos o anticuerpos o fragmentos, particularmente los anticuerpos recombinantes o fragmentos, que comprenden secuencias de región variable de CDR de la cadena pesada expuesta en la SEQ ID NOS: 37-39 y las secuencias de CDR de la región variable de cadena ligera se exponen en SEQ ID NOS: 40-42. El anticuerpo IgM42, el anticuerpo 42, rHIgM42 incluye anticuerpo que tiene la secuencia de la cadena pesada de la región variable de la SEQ ID NO: 17 y la secuencia de la región variable de la cadena ligera de la SEQ ID NO: 27. En consecuencia, las proteínas que muestran una actividad sustancialmente equivalente o alterada están contempladas. Estas modificaciones pueden ser deliberadas, por ejemplo, tales como las modificaciones obtenidas a través de mutagénesis dirigida al sitio, o pueden ser accidentales, tales como las obtenidas a través de mutaciones en huéspedes que son productoras del complejo o sus subunidades con nombre. Además, con los términos "anticuerpo", "anticuerpo de unión neurona", "anticuerpo IgM12", "anticuerpo IgM42", "anticuerpo 12", "anticuerpo 42", "sHIgM12", "sHIgM42", "rHIgM12", "rHIgM42" se pretende incluir dentro de su alcance proteínas citadas específicamente en el presente documento, así como todos los análogos sustancialmente homólogos y las variaciones alélicas.
Residuos de aminoácido descritos en este documento se prefieren estar en la forma isomérica "L". Sin embargo, los residuos en la forma isomérica "D" pueden ser sustituidos por cualquier residuo de L-aminoácido, siempre que la propiedad funcional deseada de unión a inmunoglobulina es retenida por el polipéptido. NH2 se refiere al grupo amino libre presente en el extremo amino terminal de un polipéptido. COOH se refiere al grupo carboxi libre presente en el extremo carboxi de un polipéptido. De acuerdo con la nomenclatura estándar de los polipéptido s, J. Biol. Chem., 243: 3552-59 (1969), las abreviaturas para residuos de aminoácidos se muestran en la siguiente
Tabla de Correspondencia:
TABLA DE CORRESPONDENCIA

Hay que señalar que todas las secuencias de residuos de aminoácidos se representan en la presente memoria por las fórmulas cuya orientación izquierda y derecha está en la dirección convencional del extremo amino al extremo carboxi-terminal. Además, debe observarse que un guión al principio o al final de una secuencia de residuos de aminoácidos indica un enlace peptídico a una secuencia adicional de uno o más residuos de aminoácidos. La tabla anterior se presenta para correlacionar las notaciones de tres letras y de una letra que pueden
aparecer alternativamente en la presente memoria.
Un "replicón" es cualquier elemento genético (por ejemplo, plásmido, cromosoma, virus.) que funciona como una unidad autónoma de replicación de ADN in vivo; es decir, capaz de replicación bajo su propio control.
Un "vector" es un replicón, tal como un plásmido, fago o cósmido, al cual otro segmento de ADN puede estar unido de manera que se produzca la replicación del segmento unido.
Una "molécula de ADN" se refiere a la forma polimérica de desoxirribonucleótidos (adenina, guanina, timina, o citosina) en su forma monocatenaria, o una hélice de doble cadena. Este término se refiere sólo a la estructura primaria y secundaria de la molécula, y no la limita a ninguna forma terciaria particular. Por lo tanto, este término incluye ADN de doble cadena encontrado, entre otras cosas, en moléculas de ADN lineal (por ejemplo, fragmentos de restricción), virus, plásmidos, y cromosomas. Al discutir la estructura de moléculas de a Dn bicatenario, las secuencias pueden describirse aquí de acuerdo con la convención normal de dar sólo la secuencia en la dirección 5'-a 3' a lo largo de la cadena no transcrita de ADN (es decir, la cadena que tiene una secuencia homóloga al ARNm).
Un "origen de replicación" se refiere a aquellas secuencias de ADN que participan en la síntesis de ADN. Una "secuencia codificante" es una secuencia de ADN de doble cadena que se transcribe y traduce en un polipéptido in vivo cuando se coloca bajo el control de secuencias reguladoras apropiadas. Los límites de la secuencia codificante se determinan por un codón de inicio en el extremo 5' -terminal (amino) y un codón de parada de traducción en el extremo 3' (carboxilo). Una secuencia codificante puede incluir, pero no se limita a, secuencias procarióticas, ADNc de ARNm eucariota, secuencias de ADN genómico de ADN eucariota (por ejemplo, mamífero), e incluso secuencias de ADN sintéticas. Una secuencia de terminación de señal de poliadenilación y la transcripción por lo general se encuentra 3' de la secuencia de codificación.
Las secuencias de control transcripcional y traduccional son secuencias reguladoras de ADN, tales como promotoras, potenciadoras, señales de poliadenilación, terminadoras, y similares, que proporcionan la expresión de una secuencia codificante en una célula huésped.
Una "secuencia promotora" es una región reguladora de ADN capaz de unir la ARN polimerasa en una célula e iniciar la transcripción de un (3') aguas abajo secuencia de codificación. Para los fines de definir la presente invención, la secuencia promotora está limitada en su extremo 3' por el sitio de iniciación de la transcripción y se extiende aguas arriba (dirección 5') para incluir el número mínimo de bases o elementos necesarios para iniciar la transcripción a niveles detectables por encima del fondo. Dentro de la secuencia promotora se encontrará un sitio de iniciación de la transcripción (definido convenientemente mediante mapeo con nucleasa S1), así como dominios de unión a proteínas (secuencias consenso) responsables de la unión de la ARN polimerasa. Promotores eucariotípicos contendrán a menudo, pero no siempre, cajas "TATA" y cajas "CAT". Los promotores procarióticos contienen secuencias de Shine-Dalgarno además de las secuencias consenso -10 y -35.
Una "secuencia de control de expresión" es una secuencia de ADN que controla y regula la transcripción y traducción de otra secuencia de ADN. Una secuencia codificante está "bajo el control" de secuencias de control de transcripción y traducción en una célula cuando la ARN polimerasa transcribe la secuencia codificante en ARNm, que luego se traduce en la proteína codificada por la secuencia codificante.
Una "secuencia de señal" puede incluirse antes de la secuencia codificante. Esta secuencia codifica un péptido señal, N-terminal al polipéptido, que comunica a la célula huésped para dirigir el polipéptido a la superficie celular o secretar el polipéptido en los medios, y este péptido señal es cortado por la célula huésped antes de la proteína abandone la célula. Las secuencias señal pueden encontrarse asociadas con una variedad de proteínas nativas para procariotas y eucariotas.
El término "oligonucleótido", como se usa en este documento en referencia a la sonda de la presente invención, se define como una molécula compuesta de dos o más ribonucleótidos, preferiblemente más de tres. Su tamaño exacto dependerá de muchos factores que, a su vez, dependen de la función última y el uso del oligonucleótido.
El término "cebador' tal como se utiliza aquí, se refiere a un oligonucleótido, tanto si se producen de forma natural como en un digesto de restricción purificado o producido sintéticamente, que es capaz de actuar como un punto de iniciación de la síntesis cuando se coloca bajo condiciones en las que la síntesis de un producto de extensión del cebador, que es complementario a una cadena de ácido nucleico, es inducida, es decir, en presencia de nucleótidos y un agente inductor tal como una ADN polimerasa y a una temperatura y pH adecuado. El cebador puede ser o bien o de cadena doble y debe ser suficientemente largo para cebar la síntesis del producto de extensión deseado en presencia del agente inductor de cadena sencilla. La longitud exacta del cebador dependerá de muchos factores, incluyendo temperatura, fuente del cebador y el uso del método. Por ejemplo, para aplicaciones
de diagnóstico, dependiendo de la complejidad de la secuencia diana, el cebador oligonucleótido contiene típicamente 15-25 o más nucleótidos, aunque puede contener menos nucleótidos.
Los cebadores en este documento se seleccionan para ser "sustancialmente" complementarios a diferentes hebras de una secuencia de ADN diana particular. Esto significa que los cebadores deben ser suficientemente complementarios para hibridar con sus respectivas cadenas. Por lo tanto, la secuencia del cebador no necesita reflejar la secuencia exacta del molde. Por ejemplo, un fragmento de nucleótido no complementario puede unirse al extremo 5'-del cebador, con el resto de la secuencia del cebador complementario a la cadena. Alternativamente, bases no complementarias o secuencias más largas pueden ser intercaladas en el cebador, siempre que la secuencia del cebador tenga suficiente complementariedad con la secuencia de la cadena para hibridar con ella y de ese modo formar el molde para la síntesis del producto de extensión.
Como se usa en el presente documento, los términos "endonucleasas de restricción" y "enzimas de restricción" se refieren a enzimas bacterianas, cada una de las cuales cortan el ADN bicatenario en o cerca de una secuencia de nucleótidos específica.
Una célula ha sido "transformada" por ADN exógeno o heterólogo cuando dicho ADN se ha introducido dentro de la célula. El ADN transformante puede o puede no estar integrado (unido covalentemente) en el ADN cromosómico que constituye el genoma de la célula. En procariotas, levaduras y células de mamífero por ejemplo, el ADN transformante puede mantenerse en un elemento episomal tal como un plásmido. Con respecto a células eucariotas, una célula establemente transformada es una en la que el ADN transformante se ha integrado en un cromosoma de modo que es heredado por las células hijas a través de la replicación cromosómica. Esta estabilidad se demuestra por la capacidad de la célula eucariota para establecer líneas celulares o clones formados por una población de células hijas que contienen el ADN transformante. Un "clon" es una población de células derivadas de una única célula o ancestro común por mitosis. Una "línea celular" es un clon de una célula primaria que es capaz de crecimiento estable in vitro durante muchas generaciones.
Dos secuencias de ADN son "sustancialmente homólogas" cuando al menos aproximadamente 75% (preferiblemente al menos aproximadamente 80%, y más preferiblemente al menos aproximadamente 90 o 95%) de los nucleótidos coinciden en la longitud definida de las secuencias de ADN. Las secuencias que son sustancialmente homólogas pueden identificarse comparando las secuencias usando software estándar disponible en los bancos de datos de secuencias, o en un experimento de hibridación Southern bajo, por ejemplo, condiciones rigurosas como se definen para ese sistema en particular. La definición de condiciones de hibridación apropiadas está dentro de la experiencia de la técnica. Véase, por ejemplo, Maniatis et al, Supra; DNA Cloning, Vols. I & II, supra; Nucleic Acid Hybridization, supra.
Se debe apreciar que también dentro del alcance de la presente invención están secuencias de ADN que codifican miembros de unión específica (anticuerpos) de la invención que codifican para, por ejemplo, un anticuerpo que tiene la misma secuencia de aminoácidos proporcionada en la Figura 2 o 10, o que comprende las secuencias de la región de dominio de CDR que figuran en este documento o en la Figura 10 pero que están degeneradas a la misma. Por "degenerado a" se entiende que un codón de tres letras diferentes se utiliza para específicar un aminoácido particular. Es bien conocido en la técnica que los siguientes codones pueden utilizarse de forma intercambiable para codificar cada aminoácido específico:

Se debe entender que los codones eespecíficados anteriormente son para las secuencias de ARN. Los codones correspondientes para ADN tienen una T sustituida por U.
Las mutaciones se pueden hacer en las secuencias que codifican los aminoácidos, fragmentos de anticuerpos, secuencias de la región CDR establecidos en las figuras 5 o 6, o en las secuencias de ácidos nucleicos figuran en la figura 5 o 6, de forma que un codón particular se cambia a un codón que codifica un aminoácido diferente. Tal mutación se realiza generalmente haciendo posibles los menos cambios de nucleótido. Una mutación de sustitución de esta clase puede hacerse para cambiar un aminoácido en la proteína resultante de una manera no conservadora (por ejemplo, cambiando el codón de un aminoácido que pertenece a un grupo de aminoácidos que tienen un tamaño o característica particular a un aminoácido que pertenece a otro grupo) o de una manera conservadora (por ejemplo, cambiando el codón de un aminoácido que pertenece a un grupo de aminoácidos que tienen un tamaño o característica particular a un aminoácido que pertenece al mismo grupo). Tal cambio generalmente conduce a menos cambios en la estructura y función de la proteína resultante. Un cambio no conservador es más probable que altere la estructura, actividad o función de la proteína resultante. La presente invención debe considerarse que incluye secuencias que contienen cambios conservadores que no alteran significativamente la actividad o características de unión de la proteína resultante.
El siguiente es un ejemplo de diversos grupos de aminoácidos:
Aminoácidos con grupos R no polares
Alanina, valina, leucina, isoleucina, prolina, fenilalanina, triptófano, metionina
Aminoácidos con grupos R polares no cargados
Glicina, serina, treonina, cisteína, tirosina, asparagina, glutamina
Aminoácidos con grupos cargados polares R (cargados negativamente a pH 6,0)
Ácido aspártico, ácido glutámico
Aminoácidos básicos (cargados positivamente a pH 6,0)
Lisina, arginina, histidina (a pH 6,0)
Otra agrupación puede ser aquellos aminoácidos con grupos fenilo:
fenilalanina, triptófano, tirosina
Otra agrupación puede ser según el peso molecular (es decir, tamaño de los grupos R):

Las sustituciones particularmente preferidas son:
- Lys por Arg y viceversa de tal manera que una carga positiva se puede mantener;
- Glu para Asp y viceversa de tal manera que una carga negativa se puede mantener;
- Ser por Thr tal que un -OH libre se puede mantener; y
- Gln por Asn de tal manera que un NH2 libre se puede mantener.
Las sustituciones de aminoácidos conservativos ejemplares y preferidos incluyen cualquiera de: glutamina (Q) por ácido glutámico (E) y viceversa; leucina (L) por valina (V) y viceversa; serina (S) por treonina (T) y viceversa; isoleucina (I) para la valina (V) y viceversa; lisina (K) para glutamina (Q) y viceversa; isoleucina (I) para la metionina (M) y viceversa; serina (S) por asparagina (N) y viceversa; leucina (L) para la metionina (M) y viceversa; lisina (L) para el ácido glutámico (E) y viceversa; alanina (A) para la serina (S) y viceversa; tirosina (Y) para la fenilalanina (F) y viceversa; ácido glutámico (E) por ácido aspártico (D) y viceversa; leucina (L) para isoleucina (I) y viceversa; lisina (K) para la arginina (R) y viceversa.
Las sustituciones de aminoácidos también pueden ser introducidas para sustituir un aminoácido con una propiedad particularmente preferible. Por ejemplo, una Cys puede introducir un sitio potencial para puentes de disulfuro con otra Cys. Una His puede introducirse como un sitio particularmente "catalítico" (es decir, His puede actuar como un ácido o base y es el aminoácido más común en la catálisis bioquímica). Pro puede introducirse debido a su estructura particularmente plana, que induce p-giros en la estructura de la proteína.
Dos secuencias de aminoácidos son "sustancialmente homólogas" cuando al menos aproximadamente 70% de los residuos de aminoácidos (preferiblemente al menos aproximadamente 80%, y más preferiblemente al menos aproximadamente 90 o 95%) son idénticos o representan sustituciones conservadoras. Las regiones CDR de dos anticuerpos son sustancialmente homólogas cuando uno o más aminoácidos son sustituidos con una sustitución de aminoácidos similar o conservadores, y en el que el anticuerpo/anticuerpos tienen el perfil de unión y las actividades de una o más de IgM12 o IgM42 descrito en este documento.
Una región "heterólogo" del constructo de ADN es un segmento identificable de ADN dentro de una molécula de ADN mayor que no se encuentra en asociación con la molécula mayor en la naturaleza. Por lo tanto, cuando la región heteróloga codifica un gen de mamífero, el gen estará normalmente flanqueado por ADN que no flanquea el ADN genómico de mamífero en el genoma del organismo fuente. Otro ejemplo de una secuencia de codificación heteróloga es un constructo donde la secuencia codificante en sí no se encuentra en la naturaleza (por ejemplo, un ADNc donde la secuencia codificante genómica contiene intrones, o secuencias sintéticas que tienen codones diferentes del gen nativo). Las variaciones alélicas o eventos mutacionales de origen natural que no dan lugar a una región heteróloga de ADN como se define en el presente documento.
Una secuencia de ADN está "unida operativamente" a una secuencia de control de expresión cuando los controles de secuencia de control de expresión y regula la transcripción y traducción de esa secuencia de ADN. El término "ligado operativamente" incluye tener una señal de inicio apropiada (por ejemplo, ATG) delante de la secuencia de ADN para expresar y mantener el marco de lectura correcto para permitir la expresión de la secuencia de ADN bajo el control de la secuencia de control de expresión y la producción del producto deseado codificado por la secuencia de ADN. Si un gen que uno desea insertar en una molécula de ADN recombinante no contiene una señal de inicio apropiada, dicha señal de inicio puede insertarse frente al gen.
El término "condiciones de hibridación estándar" se refiere a condiciones de sal y temperatura sustancialmente equivalentes a 5 x SSC y 65°C tanto para la hibridación como para el lavado. Sin embargo, un experto en la técnica apreciará que tales "condiciones estándar de hibridación" son dependientes de condiciones particulares que incluyen la concentración de sodio y magnesio en el tampón, nucleótidos longitud de la secuencia y la concentración, por ciento desajuste, el por ciento de formamida, y similares. También es importante en la determinación de "condiciones de hibridación estándar" si las dos secuencias de hibridación son ARN ARN, ADN-ADN o ARN-ADN. Tales condiciones de hibridación estándar se determinan fácilmente por un experto en la técnica de acuerdo con las fórmulas bien conocidas, en las que la hibridación es típicamente 10-20°C por debajo de la Tm predicha o determinada con lavados de mayor severidad, si se desea.
El término "agente" significa cualquier molécula, incluyendo polipéptido s, anticuerpos, polinucleótidos, libras composición química y moléculas pequeñas. En particular, el término agente incluye compuestos tales como compuestos de ensayo o compuestos candidatos de fármacos.
El término "agonista" se refiere a un ligando que estimula el receptor el ligando que se une a en el sentido más amplio.
El término "ensayo" significa cualquier proceso utilizado para medir una propiedad específica de un compuesto. Un "ensayo de selección" significa un proceso utilizado para caracterizar o seleccionar compuestos en base a su actividad a partir de una colección de compuestos.
El término "prevenir" o "prevención" se refiere a una reducción en el riesgo de adquirir o desarrollar una enfermedad o trastorno (es decir, hacer que al menos uno de los síntomas clínicos de la enfermedad no se desarrollen) en un sujeto que puede estar expuesto a un agente causante de la enfermedad, o predispuesto a la enfermedad antes de la aparición de la enfermedad.
El término "profilaxis" se relaciona con y abarca el término "prevención", y se refiere a una medida o procedimiento con el propósito de evitar, en lugar de tratar o curar una enfermedad. Ejemplos de medidas profilácticas no limitantes pueden incluir la administración de vacunas; la administración de bajo peso molecular de heparina para los pacientes del hospital en riesgo de trombosis debido, por ejemplo, a la inmovilización; y la administración de un agente anti-malaria, tales como cloroquina, antes de una visita a una región geográfica donde la malaria es endémica o el riesgo de contraer malaria es alto.
"Cantidad terapéuticamente eficaz" significa la cantidad de un agente de fármaco, compuesto, agente, anticuerpo, o farmacéutico que provocará la respuesta biológica o médica de un sujeto que está siendo buscado por un médico u otro clínico. En particular, con respecto a enfermedades o condiciones neurológicas, el término "cantidad eficaz" pretende incluir una cantidad eficaz de un compuesto o agente, tal como un anticuerpo o fragmento del mismo, que traerá un cambio biológicamente significativo en un parámetro clínicamente relevante de la enfermedad. Esto puede incluir un cambio en la cantidad de lesión neurológica o la muerte observable o evaluable en formación de imágenes, un cambio en un parámetro funcional, tal como movimiento, habla o cognición, o la ausencia o reducción de un cambio de este tipo en condiciones de riesgo o susceptibilidad. La frase "cantidad terapéuticamente eficaz" se usa aquí para significar una cantidad suficiente para mejorar o prevenir, y preferiblemente reducir en al menos aproximadamente 20 por ciento, al menos aproximadamente 30 por ciento, más preferiblemente en al menos 50 por ciento, más preferiblemente en al menos 70 por ciento, más preferiblemente en al menos 80 por ciento, más preferiblemente en al menos 90 por ciento, un cambio clínicamente significativo en un fenómeno medible o evaluable, o característica de la patología, o la evaluación de la función.
El término "tratar" o "tratamiento" de cualquier enfermedad, condición, lesión o infección se refiere, en una realización, a mejorar la enfermedad, condición, lesión o infección (es decir, detener la muerte o lesión de una célula o tejido, o la reducción de la manifestación, la extensión o gravedad de al menos uno de los síntomas clínicos de la misma). En otra forma de realización “tratar” o “tratamiento” se refiere a mejorar al menos un parámetro físico, que puede no ser discernible por el sujeto. En aún otra realización, “tratar” o “tratamiento” se refiere a modular la enfermedad, afección, lesión o infección, ya sea físicamente, (por ejemplo, estabilización de un síntoma discernible), fisiológicamente, (por ejemplo, estabilización de un parámetro físico), o ambos. En una realización adicional, “tratar” o “tratamiento” se refiere a retardar la progresión de una enfermedad o reducir la cantidad de lesiones, daños, la muerte a las células o tejidos.
La frase "farmacéuticamente aceptable" se refiere a entidades moleculares y composiciones que son fisiológicamente tolerables y típicamente no producen una reacción alérgica o adversa similar, tal como molestias gástricas, mareos y similares, cuando se administra a un humano.
Como se usa en el presente documento, "PG" significa picogramo, "ng" significa nanogramo, "ug" o " pg" significa microgramos, "mg" significa miligramos, "ul" o "pl" significa microlitro, "ml" significa mililitro, "I" significa litro.
B. DIVULGACIÓN DETALLADA.
Los agentes neuroprotectores están dirigidas a la prevención y tratamiento de las complicaciones que resultan en daño en el SNC. Compuestos o agentes con capacidad para la neuroprotección tienen aplicación en los trastornos agudos tales como accidente cerebrovascular y lesiones del sistema nervioso, así como en enfermedades crónicas tales como trastornos neurodegenerativos. Muchos de los mecanismos subyacentes de daño a los tejidos neuronales son similares en estas condiciones y un compuesto o agente neuroprotector podría usarse en más de un trastorno. Enfermedades incluyen trastornos cerebrovasculares, lesión cerebral traumática, lesión de la médula espinal, la enfermedad de Alzheimer, enfermedad de Parkinson, enfermedad de Huntington, esclerosis lateral amiotrófica (ELA), esclerosis múltiple, epilepsia, enfermedad de la neurona motora y la neuropatía isquémica. Otras aplicaciones incluyen la neuroprotección durante anestesia y cirugía. Numerosos productos han sido investigados para efectos neuroprotectores incluidos los de las categorías de captadores de radicales, agentes anti-excitotóxicos, inhibidores de la apoptosis (muerte celular programada), agentes anti-inflamatorios, factores neurotróficos, quelantes de iones metálicos, moduladores de canales de iones y terapia genética. Varios productos candidatos neuroprotectores han alcanzado los ensayos clínicos y no para mostrar significado, incluyendo antagonistas de NMDA y agentes para reducir la liberación de neurotransmisores Clark WM et al (2000) Stroke 31 (6): 1234-9; Sarco RI et al (2001) JAMA 285 (13): 1719-1728; Albers, GW et al (2001) JAMA 286 (21): 2673-82). Uno de los retos para los agentes neuroprotectores es la especificidad y la orientación de modo que los efectos de un agente se dirigen contra lesión o neuronas dañadas/comprometidos sin actividad significativa contra células normales y por lo tanto desfavorable, incluso efectos secundarios potencialmente dañinos. La presente invención proporciona ahora agentes específicos y tolerables capaces de atravesar la barrera hematoencefálica y la orientación a áreas de lesiones, compremeten o dañan el efecto de protección, con efectos adversos mínimos sobre las neuronas normales.
La invención se define por las reivindicaciones. La descripción proporciona generalmente miembros de unión específica, particularmente anticuerpos que demuestran la unión de la neurona, con aplicación como agentes neuroprotectores para el uso en diagnóstico, control, mejora y tratamiento de enfermedades y condiciones donde se comprometían neuronas o células nerviosas, incluidas las condiciones neurodegenerativas, la muerte de las células nerviosas y lesiones. Los anticuerpos tienen ciertas cualidades únicas y notables que los hacen particularmente útiles y aplicables en la neuroprotección y/o neuroregeneración y en el diagnóstico y la terapéutica para la enfermedad y las condiciones neurológicas. Los nuevos anticuerpos de la invención, particularmente los anticuerpos recombinantes, se demuestran en el presente documento para cruzar la barrera de sangre del cerebro de manera eficaz y sin ninguna modificación, para proteger a las neuronas de la muerte celular, y para apuntar lesiones neuronales y los sitios de lesión en el SNC. Los anticuerpos mejoran la función nerviosa y mantienen los axones en modelos animales de lesión axonal crónica y desmielinización. Los anticuerpos tienen una toxicidad mínima en los animales y no exacerben las condiciones autoinmunes, tal como se evaluó en modelos animales.
La descripción proporciona generalmente miembros de unión específica, particularmente anticuerpos que demuestran la unión de la neurona, con aplicación como agentes terapéuticos, solos o en combinación con otros o alternativos agentes neuroactivos, para la mejora y el tratamiento de enfermedades y condiciones en las neuronas o células nerviosas están comprometidas, incluyendo las enfermedades neurodegenerativas, la muerte de las células nerviosas y lesiones. La aplicación proporciona evidencia de la capacidad y la actividad terapéuticamente relevante de anticuerpos de la presente invención, incluyendo IgM12, en modelos animales de enfermedades neurológicas o déficits. En particular, los estudios proporcionados en este documento demuestran la capacidad y la actividad en un modelo de TMEV, en un modelo animal reconocido de enfermedad de la motoneurona particularmente ELA, y proporciona estudios en modelos de lesión de la médula espinal. En particular, la presente solicitud proporciona evidencia de la capacidad y la actividad terapéuticamente relevante de anticuerpos de la presente invención, incluyendo IgM12, en modelos animales de accidente cerebrovascular y trombosis cerebral o isquemia cerebral. Los estudios proporcionados en este documento demuestran la capacidad y la actividad en un en un modelo animal reconocido de accidente cerebrovascular e isquemia cerebral. Se proporcionan composiciones para uso en la mejora o tratamiento de cualquiera de dichas enfermedades o en la prevención de la lesión del nervio o la protección de los nervios en los casos de compromiso o riesgo de compromiso.
Estudios anteriores con IgM12 y IgM42 han demostrado que el anticuerpo derivado del suero se une a las neuronas del SNC, soporta la extensión de neuritas sobre el sustrato revestido con anticuerpo, y anulan la inhibición de extensión de neuritas de los estudios de la mielina del SNC en in vitro (Warrington A et al (2004) J Neuropath Exp Neurol 63 (5): 461-473). Los estudios de sHIgM12 inyectados en los animales TMEV mostraron un aumento de la actividad nocturna espontánea, sin embargo, los títulos de virus no se evaluaron en los animales y los posibles efectos anti-virales no se pudieron determinar (Rodrigues M et al (2009) Neurology 72: 1269-1276). Los estudios descritos en el presente documento proporcionan recombinante totalmente humano 12 y la secuencia del anticuerpo 42 así como rHIgM42 recombinante. Ahora se proporcionan Los estudios que demuestran notables efectos neuroprotectores y terapéuticamente relevantes de estos anticuerpos derivados del suero y recombinantes en animales y en modelos animales de enfermedad, así como la unión/direccionamiento a los sitios de lesión neural su específico. En particular estudios usando modelos animales de isquemia cerebral y derrame cerebral proporcionan demostración de efectos terapéuticamente relevantes de estos anticuerpos derivados del suero y recombinantes, particularmente IgM12 recombinante, rHIgM12, en el accidente cerebrovascular, isquemia cerebral, y las
condiciones de o como resultado de pérdida de flujo sanguíneo en o al cerebro o las células en el SNC.
Anticuerpos y composiciones
mAbs humanos auto reactivos entran en la clasificación de los autoanticuerpos naturales humanos (NatAbs) (Cohen IR (2007) J Autoimmun 29: 246-249). NatAbs son parte de nuestro repertorio de inmunoglobulina humana (Cohen, IR, y M. Schwartz (1999) Journal of neuroimmunology 100: 111-114), de forma natural a partir de nuestros propios genes, normalmente sin mutaciones somáticas y pueden funcionar para estimular procesos celulares o eliminar restos celulares (Lutz, HU (2007) J Autoimmun 29: 287-294). NatAbs reaccionan a los antígenos propios, mientras que Abs convencionales reaccionan a los antígenos exógenos. NatAbs son de afinidad relativamente baja, por lo general poli reactiva, y con frecuencia son las IgM. El mecanismo de la acción de estas IgM humanas parece ser similar, actuando a través de la membrana micro dominio s. NatAbs son por polirreactivo definición, por lo tanto, la identificación del antígeno relevante para un específico en función vivo puede ser un desafío. Sin estar ligado por la teoría, se entiende actualmente que estas IgM se unen a la membrana microdominios de reticulación moléculas en la superficie celular. Las moléculas no normalmente en contexto se juntan el montaje de un complejo de señalización (Rodríguez, M., AE Warrington, y LR Pease (2009) Neurology 72: 1269-1276).
De acuerdo con ello, las moléculas terapéuticas se identificaron mediante cribado de autoanticuerpos reactivos humanos monoclonales (mAbs) de los sueros de los individuos que llevan los mAbs en alta concentración sin enfermedad mediada por Ab. Tales mAbs se prueban para la unión a la superficie de células del sistema nervioso sin tenerse en cuenta para el antígeno. Los mAbs se ensayan después para la capacidad modular de un modelo de enfermedad. Este método de identificación de mAbs con eficacia biológica es diferente que la metodología utilizada por la industria farmacéutica (Rodríguez, M., AE Warrington, y LR Pease (2009) Neurology 72: 1269-1276). Se ha demostrado que ciertos IgM humanos pueden promover la remielinización (Warrington AE et al (2000) Proc Natl Acad Sci EE.UU. 97: 6820-6825). Por ejemplo, uno de tales IgM es monoclonal humano recombinante rHIgM22 que promueve la remielinización (Mitsunaga YB et al (2002) Faseb J 16: 1325-1327). Anticuerpo rHIgM22 se une a oligodendrocitos y mielina y promueve la remielinización del SNC en los modelos de virus y la toxina inducida de MS (Warrington AE et al (2000) Proc Natl Acad Sci EE.UU. 97: 6820-6825; Bieber AJ et al (2002) Glia 37: 241- 249). Se induce la remielinización de la médula espinal después de una sola dosis baja de rHIgM22 (Warrington AE et al (2007) J Neurosci Res 85: 967-976).
Es notable que un tratamiento intraperitoneal (ip) con una molécula de corta duración (15 h vida media estimada en ratones) promueve la reparación de tejidos máxima dentro de 5 semanas en un modelo de MS con poca reparación espontánea inherente. Después de la inyección periférica, rHIgM22 cruza la barrera hematoencefálica (BBB) y se acumula dentro de las lesiones cerebrales y de la médula espinal de ratones con desmielinización. rHIgM22 marcado con perla ferritina se ha detectado en lesiones in vivo por MRI (Pirko I et al (2004) Faseb J 18: 1577-1579). Un anticuerpo IgM de suero humano adicional con capacidad remielinizante, sHIgM46, y su homólogo recombinante rHIgM46, también se ha sido descrito. Los anticuerpos derivados del suero sHIgM22 y sHIgM46, y versiones recombinantes de los mismos, y métodos para la remielinización se describen, por ejemplo, en el documento WO 0185797. El anticuerpo y métodos rHIgM22 del mismo está cubiertos, por ejemplo, por la patente estadounidense 7,473,423 y 7,807166.
La invención proporciona anticuerpos humanos, incluyendo anticuerpos monoclonales y anticuerpos particularmente recombinantes, que demuestran actividad en la promoción, estimulación, protección y/o regeneración de neuronas en el sistema nervioso central. Los presentes anticuerpos, incluyendo fragmentos de los mismos, demuestran actividad en la neuroprotección, preservación neuronal y axonal y la regeneración en la prevención o reducción de la muerte de células nerviosas lesión mediada en el sistema nervioso central. Los anticuerpos son aplicables en el tratamiento o mejora de enfermedades o afecciones, o en la prevención o reducción de los déficits de las células nerviosas y muerte de células nerviosas o apoptosis asociados con enfermedades o condiciones, en particular cuando los nervios están dañados, lesionados o comprometidos de otro modo. Condiciones o enfermedades para el uso o aplicación de los anticuerpos de la invención incluyen escenarios en los que la pérdida de la estructura, la función o la supervivencia de las neuronas está implicada o asociada incluyendo lesión cerebral o trauma, lesión de la médula espinal (SCI), lesión nerviosa, lesión en la cabeza, condiciones en el suministro de sangre al cerebro se reduce o se ve comprometida, las enfermedades infecciosas del cerebro, enfermedades neurodegenerativas. Los ejemplos de tales enfermedades o condiciones incluyen lesión de la médula espinal (SCI), esclerosis amiolotrópica lateral (ELA), esclerosis múltiple (MS), enfermedad, accidente cerebrovascular, enfermedad de Alzheimer de Parkinson, enfermedad de Huntington, anoxia prenatal isquemia/perinatal y/o parálisis cerebral, encefalopatía, mielopatía, y de las enfermedades de neuronas motoras.
Los agentes neuromoduladores o anticuerpos de la invención tienen una o más de las siguientes características: protegen y/o estabilizan las neuronas; se dirigen a sitios en el sistema nervioso central o el daño de las células nerviosas, compromiso o lesión; y/o muerte celular de bloque, por ejemplo, peróxido de hidrógeno induce la muerte celular. La invención proporciona cuerpos anti-monoclonales de unión a neuronas de la invención son capaces de promover la extensión de neuritas y la protección de las neuronas del daño para los propósitos de diagnóstico y terapéuticos en el sistema nervioso central. En particular, se proporcionan anticuerpos recombinantes,
en los que dichos anticuerpos reconocen y son capaces de neuronas, incluyendo neuronas corticales, las neuronas del hipocampo, las células granulares cerebelares y las células ganglionares de la retina de unión.
La presente invención proporciona anticuerpos ejemplares o fragmentos de los mismos, anticuerpo 12 y el anticuerpo 42, particularmente suero derivado o IgM12 recombinante o IgM42. En un aspecto particular adicional, el anticuerpo de la invención comprende la secuencia de aminoácidos del anticuerpo 12 o 42 incluyendo lo señalado en la Figura 5 y/o en la Figura 6. El anticuerpo recombinante IgM12 de la presente invención comprende la cadena pesada variable (SEQ ID NO: 1) y la secuencia de cadena ligera (SEQ ID NO: 11) tal como se establece en la Figura 5. El anticuerpo recombinante IgM42 de la presente invención comprende la cadena pesada variable (SEQ ID NO: 17) y la secuencia de cadena ligera (SEQ ID NO: 27) tal como se establece en la Figura 6. en un aspecto de la invención se proporciona un anticuerpo de unión a neurona recombinante o sintético que comprende las secuencias de región CDR variables establecidas en las Figuras 5 y 6. El anticuerpo 12 comprende secuencias de CDR de cadena pesada CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y secuencias de CDR de cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), tal como se establece en la Figura 5. El anticuerpo 42 comprende las secuencias de CDR de cadena pesada CDR1 GFTFSTYA (SEQ ID NO: 37), CDR2 INVGGVTT (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), secuencias de CDR de cadena ligera y CDR1 QGIG (SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42), tal como se establece en la Figura 6.
Los paneles de anticuerpos recombinantes o fragmentos de los mismos, incluyendo fragmentos Fab o bibliotecas de presentación de fagos, que son capaces de reconocer las neuronas, particularmente neuronas humanas, pueden ser seleccionadas para diversas propiedades; es decir, la afinidad, isotipo, epítopo, la estabilidad, etc. De particular interés son anticuerpos que imitan la actividad de los anticuerpos ejemplares IgM12 y IgM42, y tienen la capacidad de unirse a las neuronas y para proteger a las neuronas de la muerte celular o lesión, tales como por ejemplo muerte celular mediada por peróxido. Tales anticuerpos pueden ser fácilmente identificados y/o seleccionados en ensayos de unión y de actividad específica. Los anticuerpos recombinantes que comprenden la región de unión al antígeno o las regiones de CDR de cadena pesada y/o ligera de los presentes anticuerpos IgM12 y/o IgM42, pueden ser generados y se criban para la actividad.
En general, las regiones CDR, que comprenden secuencias de aminoácidos sustancialmente como se establecen como las regiones CDR de la Figura 5 y 6 serán realizadas en una estructura que permite la unión de las regiones CDR a la superficie o en la superficie de las neuronas y, particularmente, a las neuronas de mamífero, en particular humano, mono, babuino, rata, y/o las neuronas de ratón. Por "sustancialmente como se establece" se quiere decir que las secuencias de la región variable, y/o particularmente las secuencias/CDR, de la invención serán idénticas o altamente homólogas a las regiones determinadas de la Figura 5 y 6. Por "altamente homóloga" se contempla que sólo unas pocas sustituciones, preferiblemente de 1 a 8, preferiblemente de 1 a 5, preferiblemente de 1 a 4, o de 1 a 3, o 1 o 2 sustituciones pueden ser hechas en la secuencia de región variable y/o en secuencias de CDR. El término "sustancialmente como se establece" incluye sustituciones de aminoácidos particularmente conservadoras que no afectan materialmente o significativamente la especificidad y/o actividad de los anticuerpos instantáneos.
Las sustituciones pueden ser hechas en la secuencia de región variable fuera de las CDR a fin de retener las secuencias de CDR. Por lo tanto, los cambios en la secuencia de la región variable o secuencias de la región alternativas no homólogas variables pueden ser introducidas o utilizadas, de manera que las secuencias de CDR se mantienen y el resto de la secuencia de región variable pueden sustituirse. Alternativamente, se pueden hacer sustituciones particularmente en las CDRs. Secuencias de CDR de los anticuerpos de la presente invención se exponen y describen en la presente memoria incluyendo en la Figura 5 y 6. Anticuerpo 12 comprende CDR de cadena pesada las secuencias de CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y secuencias de CDR de la cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), tal como se establece en la Figura 5. El anticuerpo 42 comprende CDR de cadena pesada las secuencias de c Dr 1 GFTFSTYA (SEQ ID NO: 37), CDR2 INVGGVTT (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), y secuencias de CDR de cadena ligera de CDR1 QGIG (SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42), tal como se establece en la Figura 6. Los anticuerpos de la invención que tienen sustituciones como se describió anteriormente y se seleccionan para mantener las actividades y especificidad contemplada en consonancia con categorías de anticuerpos ejemplares, incluyendo anticuerpos IgM12 y IgM42 y que tienen las características como se establece en el presente documento y en las reivindicaciones.
La estructura para llevar los CDRs será generalmente de una secuencia de cadena pesada o ligera de anticuerpo o una porción sustancial de la misma en la cual las regiones CDR están situadas en posiciones correspondientes a la región CDR de origen natural de dominios variables de anticuerpos VH y VL codificados por genes de inmunoglobulina reordenados. Las estructuras y ubicaciones de los dominios variables de inmunoglobulina pueden ser determinados por referencia a Kabat, EA et al, Sequences of Proteins of Immunological Interest. 4a Edición. Departamento de Salud y Servicios Humanos. 1987, y sus actualizaciones, ahora disponibles en Internet
(immuno.bme.nwu.edu). Los dominios variables se pueden derivar de cualquier línea germinal o dominio variable humano reordenado, o pueden ser un dominio variable sintético basado en secuencias consenso de dominios variables humanos conocidos. Las secuencias derivadas de CDR de la invención, como se define en el párrafo anterior, pueden introducirse en un repertorio de dominios variables que carecen de regiones CDR, utilizando tecnología de ADN recombinante.
Por ejemplo, Marks et al (Bio/Technology, 1992, 10: 779-783) describen métodos para producir repertorios de dominios variables de anticuerpo en el que los cebadores consenso dirigidos a o adyacentes al extremo 5' de la zona de dominio variable están usados en conjunción con cebadores consenso para la tercera región de marco de genes VH humanos para proporcionar un repertorio de dominios variables VH que carecen de una CDR/CDRs. Marks et al describen además cómo este repertorio se puede combinar con una c Dr de un anticuerpo particular. El repertorio puede entonces ser representado en un sistema huésped adecuado tal como el sistema de presentación de fagos de WO92/01047 de modo miembros de unión específica adecuados se pueden seleccionar. Un repertorio puede consistir de cualquier cosa a partir de 104 miembros individuales hacia arriba, por ejemplo de 106 a 108 o 1010 miembros. El barajado análogo o técnicas combinatorias también se dan a conocer por Stemmer (Nature, 1994, 370: 389-391), que describe la técnica en relación con un gen de p-lactamasa, pero observa que el enfoque puede ser utilizado para la generación de anticuerpos.
Una alternativa adicional consiste en generar nuevas regiones VH o VL que llevan las secuencias de CDR derivadas utilizando mutagénesis aleatoria de, por ejemplo, genes Ab VH o VL para generar mutaciones dentro de todo el dominio variable. Tal técnica es descrita por Gram et al (1992, Proc Natl Acad Sci, EE.UU., 89: 3576-3580), que utiliza PCR propenso a errores. Otro método que puede usarse consiste en dirigir la mutagénesis a regiones CDR de VH o VL genes. Dichas técnicas se describen por Barbas et al, (1994, Proc Natl Acad Sci, EE.UU., 91: 3809-3813) y Schier et al (1996, J. Mol Biol. 263:. 551-567). Todas las técnicas descritas anteriormente son conocidas como tales en la técnica. La persona experta será capaz de usar tales técnicas para proporcionar miembros de unión específica de la invención usando metodología rutinaria en la técnica.
Una parte sustancial de un dominio variable de inmunoglobulina comprenderá al menos las tres regiones CDR, junto con sus regiones estructurales intermedias. Preferiblemente, la porción también incluirá al menos aproximadamente 50% de cualquiera o ambas de las regiones primera y cuarta estructurales, siendo el 50% el C-término 50% de la primera región marco y el N-terminal 50% de la cuarta región de marco. Los residuos adicionales en el extremo N-terminal o C-término de la parte sustancial del dominio variable pueden ser aquellos no asociados normalmente con regiones de origen natural de dominio variable. Por ejemplo, la construcción de miembros de unión específicos de la presente invención se realizada mediante técnicas de ADN recombinante puede resultar en la introducción de residuos N- o C-término codificados por enlazadores introducidos para facilitar la clonación u otras etapas de manipulación. Otras etapas de ma nipulación incluyen la introducción de enlazadores para unir dominios variables de la invención a otras secuencias de proteínas, incluyendo cadenas pesadas de inmunoglobulina, otros dominios variables (por ejemplo en la producción de diacuerpos) o etiquetas de proteínas como se proporciona aquí y/o conocidos para los expertos en la técnica.
Aunque en un aspecto preferido de la invención, los anticuerpos recombinantes comprenden un par de dominios basados en secuencias establecidas sustancialmente en las figuras de unión 5 y/o 6 se prefieren, dominios de unión individuales en base a cualquiera de estas secuencias forman aspectos adicionales de la invención. En el caso de los dominios de unión basados en la secuencia expuesta sustancialmente en la Figura 5 y/o 6, dichos dominios de unión pueden ser utilizados como agentes de direccionamiento para las neuronas del SNC, en particular los sitios de daño al nervio o lesión, ya que se sabe que los dominios de inmunoglobulina VH son capaces de antígenos diana de unión de una manera específica.
Los miembros de unión específicos de la presente invención pueden comprender además regiones constantes de anticuerpo o partes del mismo. Por ejemplo, los anticuerpos recombinantes en base a las secuencias de VH y VL de las figuras 5 y 6 pueden estar unidos en su extremo C-término a dominios constantes de cadena ligera del anticuerpo incluyendo CK humanos o cadenas CA, preferiblemente cadenas de CA, incluyendo dominios constantes que son distintos o variantes de los respectivos dominios constantes expuestas en la Figura 5 o 6. De manera similar, los anticuerpos recombinantes basados en las secuencias de las Figuras 5 o 6 pueden estar unidos en su extremo C-término a la totalidad o parte de una cadena pesada de inmunoglobulina derivada de cualquier anticuerpo isotipo, por ejemplo, IgG, IgA, IgE, IgD e IgM y cualquiera de los isotipos de sub-clases, particularmente IgG1, IgG2b, e IgG4, y luego ensayadas para afirmar o determinar la actividad y la capacidad comparable y/o adecuado. Se prefiere IgM.
Los anticuerpos, o cualquier fragmento del mismo, puede conjugarse con o fusionarse recombinantemente a cualquier toxina celular, bacteriana o de otro tipo, por ejemplo, exotoxina de pseudomonas, ricina, o la toxina de la difteria. La parte de la toxina utilizada puede ser toda la toxina, o cualquier dominio particular de la toxina. Tales moléculas de anticuerpo-toxina han utilizado con éxito la orientación y la terapia de diferentes tipos de cánceres, véase por ejemplo, Pastan, Biochem Biophys Acta. 1997 Oct 24; 1333 (2): C1-6; Kreitman et al., N Engl J Med. 2001 Jul 26; 345 (4): 241-7; Schnell et al., Leucemia. 2000 Ene; 14 (1): 129-35; Ghetie et al., Mol Biotechnol. 2001 Jul; 18
(3): 251-68. Multímeros bi- y tri-específicos se pueden formar por asociación de diferentes moléculas de scFv y se han diseñado como reactivos de reticulación para el reclutamiento de células T en los tumores (inmunoterapia), reorientación (terapia génica) viral y como reactivos de aglutinación de glóbulos rojos (inmunodiagnóstico), véase por ejemplo. Todorovska et al., Methods J Immunol. 2001 Feb 1; 248 (1-2): 47-66; Tomlinson et al., Methods Enzymol. 2000; 326: 461-79; McCall et al., J Immunol. Mayo de 2001 15; 166 (10): 6112-7.
Los anticuerpos de la invención pueden marcarse con una etiqueta detectable o funcional. Los marcadores detectables incluyen, pero no se limitan a, marcadores radioactivos, tales como los isótopos 3H, 14C, 32P, 35S, 36Cl, 51Cr, 57Co, 58Co, 59Fe, 90Y, 121I, 124I, 125I, 131I, 111In, 117Lu, 211a, 198Au, 67Cu, 225Ac, 213Bi, 99Tc y 186Re, que puede estar unido a los anticuerpos de la invención utilizando la química convencional conocida en la técnica de formación de imágenes de anticuerpos. Las etiquetas también incluyen marcadores fluorescentes (por ejemplo fluoresceína, rodamina, rojo de Texas) y etiquetas utilizadas convencionalmente en la técnica para formación de imágenes MRI-CT. También incluyen marcadores enzimáticos tales como peroxidasa de rábano picante, p-glucoronidasa, pgalactosidasa, ureasa. Las etiquetas incluyen además restos químicos tales como biotina que pueden detectarse mediante la unión a un resto detectable cognado específico, por ejemplo, avidina marcada. Etiquetas funcionales incluyen sustancias que están diseñadas para dirigirse al sitio de compromiso, daño o lesión para proporcionar protección de o causar la destrucción de tejido neural. Tales etiquetas funcionales incluyen fármacos citotóxicos tales como 5-fluorouracilo o ricina y enzimas tales como carboxipeptidasa bacteriana o nitrorreductasa, que son capaces de convertir profármacos en fármacos activos en el sitio. Se contemplan inmunoconjugados o proteínas de fusión de anticuerpos de la presente invención, en que los anticuerpos y fragmentos de los mismos se conjugan o se unen a otras moléculas o agentes incluyen adicionalmente, pero no se limitan a los miembros conjugados a un agente de ablación química, toxina, inmunomodulador, citoquina de unión, agente citotóxico, agente quimioterapéutico o fármaco. Cuando se utilizan etiquetas radiactivas, procedimientos de recuento disponibles actualmente conocidos se pueden utilizar para identificar y cuantificar los miembros de unión específicos. En el caso en el que la etiqueta es una enzima, la detección puede realizarse por cualquiera de las técnicas conocidas utilizadas actualmente colorimétricas, espectrofotométricas, fluoroespectrofotométricas, amperométricas o gasométricas.
Modelos animales in vivo de complicaciones que resultan en daño en el SNC, incluyendo lesiones de las células nerviosas o daño o enfermedades neurodegenerativas pueden utilizarse por el experto en la materia para promover o adicionalmente visualizar, evaluar, y/o verificar los anticuerpos o fragmentos de los mismos de la presente invención, variantes de los mismos, o combinaciones de los mismos, o combinaciones con otros anticuerpos reactivos del SNC. Tales modelos animales incluyen, pero no están limitados a los modelos de condiciones o enfermedades asociadas con el daño de la célula nerviosa, el compromiso, alteración, muerte, lesión o degeneración. Los modelos incluyen los de aspectos imitadores o de un derrame cerebral, lesión cerebral, isquemia, esclerosis múltiple, enfermedad de Alzheimer, enfermedad de Huntington, enfermedad de Parkinson, demencia, encefalitis, meningitis, ELA o enfermedad de la motoneurona. Modelos de accidente cerebrovascular que utilizan oclusión de la arteria cerebral media en primates, tales como babuinos, monos o macacos son conocidos en la técnica y pueden ser adecuados (Young A et al (1997) Stroke 28: 1471-1476; Delzoppo GJ et al (1986) Stroke 17: 1254-1265; Marshall y Ridley 91996) Neurodegeneration 5: 275-286). Modelos de MS tales como TMEV se conocen y describen y se utilizan en el presente documento. Modelos SOD de ratón para la ELA y la enfermedad de la neurona motora se conocen y describen y se utilizan en el presente documento (Gurney ME et al (1994) Science 264: 1772-1775).
Métodos de derrame y terapéuticas
Los ejemplos y estudios proporcionados en este documento demuestran la eficacia y la aplicación de los anticuerpos proporcionados, descritos y contemplados en este documento en modelos de apoplejía, isquemia y casos de hipoxia al cerebro o las células neuronales. Así, en un aspecto particular, la presente invención se refiere al uso de los anticuerpos de la intervención relativa a apoplejía, isquemia cerebral, sospechoso o presunto accidente cerebrovascular, ataques isquémicos transitorios (AIT) o movimientos de menor importancia, o en individuos con alto riesgo de accidente cerebrovascular o un evento isquémico cerebral o evento hipóxico cerebral.
En el caso de un derrame cerebral, o derrame sospechoso de accidente cerebrovascular, el tratamiento rápido es importante ya que los mayores daños de un derrame cerebral ocurre generalmente dentro de las primeras horas. Resultados más rápidos de tratamiento en un menor daño inicial y permanente o a largo plazo. El tratamiento depende de la naturaleza o causa del accidente cerebrovascular, ya sea restaurando el flujo sanguíneo en el ictus isquémico debido a un coágulo de sangre (isquemia) o mediante el control de la hemorragia durante un accidente cerebrovascular hemorrágico. Las tomografías computarizadas y resonancias magnéticas se utilizan para evaluar o diagnosticar el tipo o magnitud de un derrame cerebral. Los tratamientos actuales para el accidente cerebrovascular isquémico agudo incluyen la terapia IV trombolítica, en particular con los de tipo activador del plasminógeno tisular (tPA), y terapias endovasculares, incluyendo la terapia trombolítica intra-arterial y el uso de dispositivos de recuperación de coágulos. Sin embargo, el tPA tiene una ventana terapéutica limitada y se administra típicamente a menos de 3 h de inicio del accidente cerebrovascular. La protección de las neuronas dañadas después de la isquemia cerebral es una estrategia prometedora para limitar la discapacidad permanente. Mientras que las terapias
que modulan la respuesta inflamatoria pueden tener propiedades neuroprotectoras, en la actualidad hay muy pocos fármacos que actúan directamente sobre las neuronas para protegerlas.
Una serie de medicamentos que pueden ayudar a prevenir el accidente cerebrovascular en pacientes de alto riesgo, en particular aquellos que han tenido anteriores isquémicos transitorios (AIT) o movimientos de menor importancia, están disponibles. Estos medicamentos se dividen en dos categorías principales - anticoagulantes y agentes antiplaquetarios. Ejemplos de anticoagulantes incluyen heparina, Coumadina, warfarina, ximelagatran y exanta. Estos medicamentos actúan al adelgazamiento de la sangre para ayudar a prevenir la coagulación, incluso en casos de trombosas de vena profunda y embolias pulmonares, y son muy eficaces para ayudar a prevenir el ictus en pacientes con fibrilación auricular. Los agentes antiplaquetarios incluyen la aspirina, persantina, dipiridamol, plavix y clopidogrel. Estos agentes previenen o reducen la agregación de plaquetas y por lo tanto reducen la formación de coágulos o la formación de coágulos y pueden reducir el riesgo de ictus en pacientes que han tenido AIT o ictus isquémicos anteriores. A pesar de la disponibilidad de la terapia trombolítica, derrame cerebral sigue siendo la tercera causa principal de muerte en los Estados Unidos y la causa más común de discapacidad en los adultos. Los científicos y los médicos continúan buscando maneras de lograr una mejor recuperación funcional en pacientes con accidente cerebrovascular. Agentes neuroprotectores han generado un interés intenso y tienen el potencial de hacer el cerebro menos susceptible a los efectos dañinos de un derrame cerebral.
En la evaluación de la magnitud y los efectos de un derrame cerebral, los médicos y el personal médico examinan y consideran diversos aspectos de las capacidades y funciones del paciente. Además de las técnicas de imagen para identificar y localizar el área de un accidente cerebrovascular o coágulo y la isquemia, los pacientes se evalúan utilizando escalas de evaluación con accidente cerebrovascular y escalas de evaluación funcional. Numerosas escalas se aceptan o utilizan y serán reconocidas y conocidas para los médicos y los expertos en la técnica. Escalas de evaluación de funciones comprenden, por ejemplo, la escala de equilibrio de Berg, Lawton Instrumental Activities of Daily Living (AIVD) Escala, Rankin o escala de Rankin modificada, y la escala de derrame NIH (NIHHS). Estas escalas evalúan y determinan varios o un número de diversos parámetros, incluyendo la función superior y la extremidad inferior del motor, extremidad de ataxia, la función sensorial, el lenguaje, la articulación, la atención, la mirada y la función visual, brazo, mano y potencia de las piernas y el movimiento. Los pacientes pueden ser evaluados en una o más escalas justo después del accidente cerebrovascular o accidente isquémico, después del evento, antes y después del tratamiento, para la evaluación de posibles candidatos o tratamientos de agentes, para la comparación de tratamientos o agentes y con el tiempo, incluyendo los días, semanas, meses, años.
Los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden secuencias de la región variable pueden ser utilizados en métodos de tratamiento, prevención o diagnóstico del cuerpo humano o animal, tales como un método de neuroprotección en un mamífero, particularmente en los casos de derrame establecido, sospechoso o posible accidente cerebrovascular o isquemia cerebral, que comprende administrar a dicho mamífero una cantidad eficaz de los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes los anticuerpos recombinantes o fragmentos de los mismos que comprenden las secuencias de la región de dominio de CDR de acuerdo con la invención se pueden usar en tales métodos. Los agentes particularmente anticuerpos recombinantes o fragmentos de los mismos pueden usarse en los casos de accidente cerebrovascular establecido, sospechoso o posible o isquemia cerebral, incluyendo como agentes neuroprotectores en métodos para la prevención, tratamiento o mejora de lesión del nervio, daño o compromiso y complicaciones que pueden, o pueden no resultar en daño en el SNC. Los métodos de la invención son aplicables donde la pérdida de estructura, la función o la supervivencia de las neuronas está involucrada o asociada incluyendo lesión cerebral o trauma, lesión de la médula espinal (SCI), lesión nerviosa, lesión en la cabeza, condiciones en las que el suministro de sangre u oxígeno al cerebro se reduce o se compromete. Los ejemplos de tales enfermedades o condiciones para el tratamiento, prevención o mejora de acuerdo con los métodos de la invención incluyen el accidente cerebrovascular, ataques isquémicos transitorios, isquemia cerebral, anoxia prenatal/isquemia perinatal y/o parálisis cerebral.
Los agentes particularmente anticuerpos recombinantes o fragmentos de los mismos pueden usarse como agentes en los métodos de prevención, tratamiento o mejora de accidente cerebrovascular o isquemia cerebral. La presente invención proporciona métodos de tratamiento o mejora de accidente cerebrovascular o la isquemia cerebral que comprende administrar un anticuerpo de la presente invención, particularmente anticuerpo recombinante, particularmente anticuerpo recombinante IgM12 o IgM12, incluyendo un anticuerpo o fragmento del mismo que comprende las secuencias CDR de cadena pesada CDR1 g GsVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y secuencias de CDR de cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), tal como se establece en la Figura 5.
La presente descripción proporciona métodos de tratamiento o mejora de enfermedades o condiciones en mamíferos en los que se comprometían nervios, lesionados o dañados o escenarios donde los nervios o neuronas son susceptibles o están en riesgo del compromiso, lesiones o daños que comprenden administrar un anticuerpo recombinante o plenamente anticuerpo o fragmento humano de los mismos seleccionados de IgM12 y IgM42. Los métodos pueden comprender la administración de más de un anticuerpo o fragmento, incluyendo combinaciones de anticuerpo IgM12 y 42. En un tal método adicional, uno o más de anticuerpo IgM12 y/o de anticuerpo IgM42 puede
combinarse con otro anticuerpo activo sobre el SNC, en particular incluyendo uno o más de los anticuerpos rHIgM22 y/o rHIgM46. Las combinaciones de anticuerpos se pueden administrar colectivamente o en serie, y en varios momentos y diferentes cantidades o concentraciones. Por lo tanto, el anticuerpo 12 y/o 42 se pueden administrar en combinación con anticuerpo 22 y/o 46, por administración combinada o en serie, separados por un tramo corto de tiempo o la longitud de tiempo más largo, incluso por horas, días o semanas. Anticuerpo 12 y/o 42 pueden particularmente administrarse en combinación con anticuerpo 22 y/o 46 (sHIgM22, rHIgM22, sHIgM46 o rHIgM46), por administración combinada o en serie para el tratamiento de mejora de una enfermedad o condición que implica la neurodegeneración, y en particular incluyendo desmielinización. En un tal método, el anticuerpo 12 y/o 42 se administra en combinación con el anticuerpo 22 y/o 46, por administración combinada o en serie para el tratamiento de mejora de una enfermedad desmielinizante o condición, en particular incluyendo la esclerosis múltiple (MS). Las secuencias de cadena pesada y ligera variable del anticuerpo IgM22 se exponen en SEQ ID NO: 43 y SEQ ID NO: 44, respectivamente. Las secuencias de cadena pesada y ligera variable del anticuerpo IgM46 se exponen en SEQ ID NO: 45 y SEQ ID NO: 46, respectivamente. Uno o más de anticuerpo IgM12 y/o de anticuerpo IgM42 se puede combinar con uno o más anticuerpos remielinizantes que comprende (a) CDRs de la región variable de cadena pesada que comprende la secuencia CDR1 SSGMH secuencia CDR2 V(I)ISYDGSRKYYADSVKG y secuencia CDR3 GVTGSPTLDY, y CDR de región variable de cadena ligera comprende la secuencia CDR1 SGSSSNIGNNFVS secuencia CDR2 DITKRPS y secuencia CDR3 G(E)TWDSSLSAV V; o (b) CDR de la región variable de la cadena pesada que comprende la secuencia de CDR1 SGFTFSSYW secuencia CDR2 IKKDGSEK y secuencia CDR3 ARPNCGGDCYLPWYFD, y CDR de región variable de cadena ligera que comprende la secuencia CDR1 QSVLYSSNNKNY secuencia CDR2 secuencia YWAS y CDR3 QQYYNTPQA.
Los métodos de la invención pueden incluir la administración de los anticuerpos de la invención solo o en combinación con, en serie con, o posteriormente a la administración de accidente cerebrovascular o agentes isquémicos incluyendo uno o más de los agentes trombolíticos, agentes antiplaquetarios, moduladores de presión arterial, agentes antihipertensivos, antiinflamatorios, que alteran aminoácidos excitatorios o sus receptores, moduladores de calcio intracelulares, moduladores del canal de captadores de radicales. En un aspecto particular, los anticuerpos o fragmentos activos de los mismos, en particular anticuerpo IgM12 y/o IgM42, se administran con agentes trombolíticos y/o agentes anti-plaquetarios. En un aspecto, los anticuerpos se pueden administrar con o después de un agente trombolítico, tal como TPA.
Administrado con puede incluir más o menos al mismo tiempo, poco después, minutos después, hasta una hora después, horas después, un día después. La administración del anticuerpo, ya sea solo particularmente como anticuerpos IgM12 o IgM42, o en combinación, o con otros agentes, puede estar dentro o después de 3 horas, 4 horas, 5 horas, 6 horas, hasta 9 horas terapéuticas para administración de TPA o del agente trombolítico después de una apoplejía documentada o sospechada o evento isquémico cerebral. Anticuerpos se puede administrar en una sola dosis o de múltiples dosis. A modo de ejemplo y no de limitación, anticuerpos se pueden administrar dentro de 3 horas, después de 3 horas, hasta 4 horas, hasta 5 horas, hasta 6 horas, hasta 8 horas, hasta 12 horas, hasta 24 horas, hasta un día, hasta 2 días, hasta 3 días, hasta varios días, hasta una semana o más después de una apoplejía documentada o sospechada o evento isquémico cerebral. Anticuerpos pueden, a modo de ejemplo y no de limitación, administrarse inicialmente dentro de 3-6 horas después de una apoplejía documentada o sospechada o evento isquémico cerebral, seguido de una o más administraciones adicionales de anticuerpos hasta un día, 2 días, 3 días, 4 días, 5 días, 6 días, una semana, más de una semana, 2 semanas, más de 2 semanas, o más tarde después de la administración inicial o posterior. La dosificación puede ser modulada basándose en la respuesta o función del paciente. Por ejemplo, el paciente puede ser evaluado, por ejemplo utilizando una escala de accidente cerebrovascular tal como NihSs antes o después, antes y después de, o después de la administración inicial de anticuerpo, y la administración del anticuerpo puede ser modificada o continuada o alterada sobre la base de una escala u otra evaluación neurológica o motor del paciente. En un aspecto particular de la invención y el método, el anticuerpo son eficaces para mejorar las medidas de escala de ictus u otras escalas neurológicas o funcionales, en particular en comparación con ninguna administración del anticuerpo, la administración de TPA o de otro agente solo, o frente a un agente alternativo, incluyendo un anticuerpo alternativo o agente neuroprotector.
Por lo tanto, como se demuestra en el presente documento en modelos animales de accidente cerebrovascular, animales a los que se les administra anticuerpos de la invención demuestran una mejor actividad funcional. Los anticuerpos de la presente invención son eficaces en la mejora de la actividad locomotora, incluyendo la mejora en la función motora horizontal, mediante la administración de los mismos después de un accidente cerebrovascular o isquemia cerebral. Después de una lesión isquémica o accidente cerebrovascular, la discapacidad asociada con déficits clínicos se debe principalmente a la disfunción de las neuronas. La falta de recuperación funcional es, en parte, atribuida a la restricción de la regeneración y la neuroplasticidad (Walmsley, AR, y Mir, AK (2007) Current Pharmaceutical Design 13: 2470-2484). El accidente cardiovascular puede resultar en una función motora alterada tal como una incapacidad para mover una o más extremidades de un lado del cuerpo, los efectos cognitivos tales como una incapacidad para comprender o formular el habla, o otros déficits incluyendo efectos visuales tales como una incapacidad para ver un lado del campo visual. En un aspecto de la invención y sus métodos, los anticuerpos de la invención son eficaces para prevenir o revertir los déficits neurológicos relacionados con el derrame. Por lo tanto, los anticuerpos de la invención pueden mejorar uno o más síntomas neurológicos o déficit en los casos de accidente cerebrovascular o isquemia cerebral. Los anticuerpos de la invención son eficaces
en la mitigación de los efectos o daños neurológicos, particularmente en el SNC, en el caso o ejemplo de un accidente cerebrovascular. Los anticuerpos son de uso en métodos para reducir el daño neurológico o muerte celular neuronal, y eficaces para preservar o proteger a las neuronas en el caso de un accidente cerebrovascular o isquemia cerebral.
En un aspecto de la invención, los anticuerpos, fragmentos de los mismos y anticuerpos recombinantes que comprenden las secuencias de región variable de acuerdo con la invención, se pueden utilizar en métodos o administrarse en composiciones para la mejora o la estabilización de la función neurológica o de la función motora en los casos, particularmente durante, después o en un derrame cerebral o evento isquémico cerebral, en el que los nervios se vean comprometidos, lesionados o dañados o escenarios en que los nervios o neuronas son susceptibles o están en riesgo de compromiso, lesiones o daños. En un aspecto particular, los anticuerpos o fragmentos de los mismos se utilizan o se administran en etapas tempranas o iniciales de una enfermedad, trauma, o condición, tales como un accidente cerebrovascular o sospecha de accidente cerebrovascular, o repetirse durante el curso o duración de una enfermedad, trauma, o condición, como después de una apoplejía, a fin de facilitar o mantener la mejoría o estabilización de la función neurológica o la función motora, incluyendo o seleccionándose de movimiento, tal como caminar, o la función cognitiva, como recuerdo o reconocimiento.
Los anticuerpos de la presente invención se pueden administrar a un paciente en necesidad de tratamiento a través de cualquier vía adecuada, incluyendo por inyección intraperitoneal, en el torrente sanguíneo o CSF, o directamente en el sitio de la lesión o el compromiso. Una ventaja particular de los anticuerpos ejemplares de la presente invención es que cruzan la BBB y por lo tanto pueden dirigirse a la SNC incluso en la administración ip. La dosis precisa dependerá de un número de factores, incluyendo si el anticuerpo es para diagnóstico o para tratamiento, el tamaño o la extensión y localización de la lesión, la naturaleza precisa del anticuerpo (ya sea anticuerpo completo, fragmento, diacuerpo, etc), y la naturaleza de cualquier marca detectable o funcional unida al anticuerpo. Cuando se usa un radionúclido para la terapia, una dosis única máxima adecuada puede ser de aproximadamente 45 mCi/m2, a un máximo de aproximadamente 250 mCi/m2. La dosis preferible está en el intervalo de 15 a 40 mCi, con un mayor intervalo de dosificación preferido de 20 a 30 mCi, o de 10 a 30 mCi. Tal terapia puede requerir la médula ósea o células madre de reemplazo celular. Anticuerpos desnudos aprobados clínicamente se administran generalmente en cantidades mg, con dosis de adultos de 20 a 2000 mg de proteína por dosis, de 20 a 1500 mg de proteína por dosis, o 20 a 1000 mg de proteína por dosis, o de 20 a 500 mg de proteína por dosis, o de 20 a 100 mg de proteína por dosis. Anticuerpos monoclonales inyectables aprobados clínicamente se administran en cantidades en mg, 3-5 mg/kg, 5-10 mg/kg por dosis, 300-400mg/dosis, 300-500mg/dosis (Newsome BW y Ernstoff MS (2008) Br J Clin Macol Pharmacia 66 (1): 6-19; herceptin.net, tysabri.net, avastin.net, remicade.com). Es notable que el anticuerpo remielinizante IgM22 se ha demostrado que es eficaz en dosis comparativamente significativamente más bajas, en el rango pg, y es capaz de atravesar la BBB a ser activos en el SNC, incluso con una dosis única de anticuerpo (documento WO 2004/110355; Warrington AE et al (2007) J Neurosci Res 85 (5): 967 976). Anticuerpo IgM12 recombinante se muestra en este documento que tienen actividad terapéuticamente relevante en modelos animales a una dosis ip individual en el rango pg. Por lo tanto la dosificación de los anticuerpos de la presente invención, ya sea en dosis única o dosis múltiples y/o periódicas, en el intervalo de pgs por dosis o pg/kg por dosis, o en bajo mg por dosis (100 pg-1 mg, a menos de 1 mg, 1 mg-5 mg, 1 mg-10 mg, 5 mg-15 mg, 10 mg-20 mg por dosis), pueden ser aplicables y eficaces. Una dosis para un solo tratamiento de un paciente adulto, puede ser ajustada proporcionalmente para niños y bebés, y también ajustarse para otros formatos de anticuerpos, en proporción, por ejemplo, con el peso molecular. Los tratamientos pueden repetirse a diario, dos veces por semana, intervalos semanales o mensuales, a discreción del médico. Una ventaja de los anticuerpos a modo de ejemplo es que se cruzan los sitios de BBB y de destino de daño o lesión, facilitando así el uso de menos dosis más bajas y potencialmente para conseguir efectos adecuados.
Composiciones farmacéuticas y terapéuticas
Los anticuerpos o fragmentos de la presente invención generalmente serán administrados en forma de una composición farmacéutica, que puede comprender al menos un componente además del anticuerpo o fragmentos de la invención. Así, las composiciones farmacéuticas de acuerdo con la presente invención, y para su uso de acuerdo con la presente invención, pueden comprender, además del ingrediente activo, un excipiente farmacéuticamente aceptable, portador, tampón, estabilizador u otros materiales bien conocidos por los expertos en la técnica. Tales materiales deben ser no tóxicos y no deben interferir con la eficacia del ingrediente activo. La naturaleza precisa del portador u otro material dependerá de la vía de administración, que puede ser oral, o por inyección, por ejemplo, intravenosa, o por deposición en un sitio tumoral.
Las composiciones farmacéuticas para administración oral pueden estar en forma de tableta, cápsula, polvo o forma líquida. Un comprimido puede comprender un vehículo sólido tal como gelatina o un adyuvante. Las composiciones farmacéuticas líquidas comprenden generalmente un vehículo líquido tal como agua, petróleo, aceites animales o vegetales, aceite mineral o aceite sintético. Solución salina fisiológica, dextrosa u otra solución de sacárido o glicoles tales como etilenglicol, propilenglicol o polietilenglicol pueden ser incluidos. Para inyección intravenosa, o inyección en el sitio de la aflicción, el ingrediente activo puede estar en la forma de una solución acuosa parenteralmente aceptable que está libre de pirógenos y tiene pH, isotonicidad y estabilidad. Los expertos
relevantes en la técnica son capaces de preparar soluciones adecuadas utilizando, por ejemplo, vehículos ¡sotónicos tales como inyección de cloruro sódico, inyección de Ringer, inyección de lactato de Ringer. Conservantes, estabilizantes, tampones, antioxidantes y/u otros aditivos pueden incluirse, según sea necesario.
Una composición puede administrarse sola o en combinación con otros tratamientos, agentes terapéuticos o agentes, ya sea simultánea o secuencialmente dependiendo de la condición a ser tratada. Se contemplan composiciones que comprenden combinaciones de uno o más anticuerpos recombinantes o fragmento del mismo como se describe aquí. Además, la presente invención contempla e incluye composiciones que comprenden el anticuerpo o fragmento del mismo, otros agentes o terapéuticos tales como agentes neuroactivos o agentes terapéuticos, agentes anti-inflamatorios, la liberación de agentes moduladores neurotransmisores, ligandos neurorreceptores o agonistas o antagonistas, agentes del canal de calcio en el presente documento se describe, moduladores inmunes, u otros anticuerpos reactivos SNC. Se contemplan composiciones que comprenden combinaciones de uno o más anticuerpos recombinantes o fragmentos del mismo como se describe aquí. Otros tratamientos o agentes terapéuticos pueden incluir el modo de gestión de dosis adecuadas de medicamentos de alivio del dolor tales como fármacos no esteroides anti-inflamatorios (por ejemplo aspirina, paracetamol, ibuprofeno o ketoprofeno) o los opiáceos como la morfina, o antieméticos. Además, la composición se puede administrar con moduladores inmunes, tales como interleucinas, factor de necrosis tumoral (TNF) u otros factores de crecimiento, factores estimulantes de colonias, citoquinas u hormonas tales como dexametasona que estimulan la respuesta inmune y la reducción o eliminación de células de cáncer o tumores. La composición también puede administrarse junto con, o puede incluir combinaciones, junto con otros anticuerpos reactivos SNC, en particular incluyendo anticuerpos remielinizantes, incluyendo rHIgM22 y/o rHIgM46.
La presente descripción contempla además composiciones terapéuticas útiles en la práctica de los métodos terapéuticos de esta invención. Una composición terapéutica incluye, en mezcla, un excipiente farmacéuticamente aceptable (transportador) y uno o más de un anticuerpo, polipéptido análogo del mismo o fragmento del mismo, como se describe en el presente documento como un ingrediente activo. En una realización preferida, la composición comprende un antígeno capaz de modular la unión del miembro/anticuerpo de unión a una célula diana específica presente. La preparación de composiciones terapéuticas o farmacéuticas que contienen los anticuerpos, polipéptidos, o fragmentos activos como ingredientes activos se entiende bien en la técnica. Típicamente, tales composiciones se preparan como inyectables, bien como soluciones líquidas o suspensiones. Sin embargo, formas sólidas adecuadas para solución en, o suspensión en, líquido antes de la inyección también se pueden preparar. La preparación también se puede emulsionar. El ingrediente terapéutico activo se mezcla a menudo con excipientes que son farmacéuticamente aceptables y compatibles con el ingrediente activo. Los excipientes adecuados son, por ejemplo, agua, solución salina, dextrosa, glicerol, etanol, o similares y combinaciones de los mismos. Además, si se desea, la composición puede contener cantidades menores de sustancias auxiliares tales como agentes humectantes o emulsionantes, agentes de tamponamiento del pH que aumentan la eficacia del ingrediente activo. Un anticuerpo o fragmento activo puede formularse en la composición terapéutica como farmacéuticos que neutralizan formas de sal farmacéuticamente aceptables. Las sales farmacéuticamente aceptables incluyen las sales de adición de ácido (formadas con los grupos amino libres de la molécula de polipéptido o anticuerpo) y que se forman con ácidos inorgánicos tales como, por ejemplo, ácidos clorhídrico o fosfórico, o ácidos orgánicos tales como acético, oxálico, tartárico, mandélico, y similares. Las sales formadas a partir de los grupos carboxilo libres también se pueden derivar de bases inorgánicas tales como, por ejemplo, sodio, potasio, amonio, calcio, o hidróxidos férricos, y bases orgánicas tales como isopropilamina, trimetilamina, 2 -etilamino etanol, histidina, procaína, y similares.
Las composiciones que contienen fragmentos en anticuerpos o activos terapéuticos se administran convencionalmente intraperitonealmente o por vía intravenosa, como mediante inyección de una dosis unitaria, por ejemplo. El término "dosis unitaria" cuando se utiliza en referencia a una composición terapéutica de la presente invención se refiere a unidades físicamente discretas adecuadas como dosificación unitaria para seres humanos, conteniendo cada unidad una cantidad predeterminada de material activo calculado para producir el efecto terapéutico deseado en asociación con el diluyente requerido; es decir, portador, o vehículo.
Las composiciones se administran de una manera compatible con la formulación de dosificación, y en una cantidad terapéuticamente eficaz. La cantidad a administrar depende del sujeto a tratar, capacidad del sistema del sujeto para utilizar el ingrediente activo, y el grado de capacidad de unión de la neurona deseado o extensión de la lesión neural. Las cantidades precisas de ingrediente activo requeridas para ser administradas dependen del juicio del médico y son peculiares para cada individuo. Los regímenes adecuados para la administración inicial y seguir en la administración también son variables, y pueden incluir una administración inicial seguida de dosis repetidas a uno o más intervalos de una hora, día, semana o mes por una inyección posterior u otra administración. Alternativamente, se contemplan infusión o administración suficiente para mantener concentraciones adecuadas y suficientes en la sangre, SNC o en el sitio de terapia deseada continua.
El momento de administración puede variar y puede determinarse por el experto en la técnica o profesional médico, basado en la enseñanza de la memoria descriptiva, los parámetros clínicos del paciente o sujeto, el estado o la gravedad de la afección o enfermedad, o grado o la naturaleza de la lesión neuronal, implicación o compromiso.
Por lo tanto, la mejora en la función neural o una mayor protección de las neuronas, por ejemplo, de la muerte o compromiso, se puede mejorar mediante la administración temprana de la aparición o manifestación clínica de una enfermedad, a fin de minimizar la magnitud del daño neurológico o compromiso. En un aspecto, el momento de administración se coordina con evaluaciones de la función neurológica, determinación de la condición y/u otras evaluaciones clínicas a fin de minimizar o aliviar la progresión de la enfermedad de deterioro neurológico o daños.
Ensayos de diagnóstico
La presente descripción también se refiere a una variedad de aplicaciones de diagnóstico, incluyendo los métodos para detectar o determinar el daño de la célula nerviosa, lesión o compromiso. Los anticuerpos y fragmentos pueden ser utilizados para evaluar, cuantificar, orientarse a y/o neuronas de imagen, in vitro o in vivo. Los presentes anticuerpos, incluyendo fragmentos de los mismos, y los fármacos que modulan la producción o actividad de los miembros de unión, anticuerpos específicos y/o sus subunidades pueden poseer determinadas aplicaciones de diagnóstico y pueden, por ejemplo, utilizarse para el propósito de detectar y/o condiciones de medida o enfermedades que implican el compromiso de neuronas, degeneración, lesión, daño o la muerte. Los presentes anticuerpos incluyendo fragmentos de los mismos poseen ciertas aplicaciones diagnósticas y pueden por ejemplo, utilizarse para el propósito de detectar y/o condiciones o enfermedades que implican isquemia cerebral incluyendo accidente cerebrovascular o lesión cerebral, incluyendo lesión cerebral traumática de medición. El marcado, incluyendo radiomarcados, anticuerpos y fragmentos de los mismos, son útiles in vitro en las técnicas de diagnóstico y técnicas in vivo de obtención de radioimágenes y en radioinmunoterapia. En el caso de formación de imágenes in vivo de los anticuerpos o fragmentos de la presente invención se pueden conjugar a un agente de imagen en lugar de un radioisótopo, incluyendo, pero no limitados a una imagen de resonancia magnética para agente potenciador, en el que por ejemplo una molécula de anticuerpo es cargado con un gran número de iones paramagnéticos a través de grupos quelantes. Ejemplos de grupos quelantes incluyen EDTA, porfirinas, éteres corona y poliaminas polioximas. Los ejemplos de iones paramagnéticos incluyen gadolinio, hierro, manganeso, renio, europio, lantano, holmio y ferbio. En un aspecto adicional de la invención, los anticuerpos radiomarcados y fragmentos de los mismos, particularmente radioinmunoconjugados, son útiles en radioinmunoterapia, particularmente como anticuerpos radiomarcados para la terapia celular. En un aspecto adicional más, los miembros de unión específica radiomarcados, particularmente anticuerpos y fragmentos de los mismos, son útiles en técnicas de cirugía radioinmunoensayadas, en que se pueden identificar e indicar la presencia y/o localización de las neuronas comprometidas o dañadas o los sitios de la lesión del nervio, durante o después de la cirugía para apuntar o eliminar tales células o trasplantar o administrar células a esos sitios específicos.
La radioinmunoterapia (RAIT) ha entrado en la clínica y eficacia demostrada usando varios inmunoconjugados de anticuerpos. Anticuerpo humanizado hMN-14 de antígeno (anti-CEA) anti-carcinoembriónico etiquetado con 131I ha sido evaluado en el cáncer colorrectal (Behr TM et al (2002) Cáncer 94 (4Suppl): 1373-1381) y el mismo anticuerpo con etiqueta 90Y ha sido evaluado en carcinoma medular de tiroides (Stein R et al (2002) Cancer 94 (1): 51-61). La radioinmunoterapia utilizando anticuerpos monoclonales también ha sido evaluada y reportada para linfoma no de Hodgkin y cáncer de páncreas (Goldenberg DM (2001) Crit Rev Onco1Hematol 39 (1 2): 195-201; Oro DV et al (2001) Crit Rev Oncol Hematol 39 (1-2) 147-54). También se describen métodos de radioinmunoterapia con anticuerpos particulares en la Patente de Estados Unidos 6.306.393 y 6.331.175. Cirugía radioinmunoguiada (RIGS) también ha entrado en la clínica y demostrado eficacia y utilidad, incluyendo el uso de anticuerpos anti-CEA y anticuerpos dirigidos contra antígenos asociados a tumores (Kim JC et al (2002) Int J Cancer 97 (4): 542-7; Schneebaum S et al (2001) World J Surg 25 (12): 1495-8; Avital S et al (2000) Cancer 89 (8 ): 1692-8; Mclntosh DG et al (1997) Cancer Biother Radiopharm 12 (4): 287-94).
Los anticuerpos radiomarcados y fragmentos de los mismos, son útiles in vitro en las técnicas de diagnóstico y técnicas in vivo de obtención de radioimágenes. Los anticuerpos y fragmentos de los mismos, en particular los radioinmunoconjugados, son útiles en radioinmunoterapia, particularmente como anticuerpos radiomarcados para la reparación de la lesión del nervio, recuperación neurodegenerativa, cáncer o terapia de tumor del SNC, o alternativamente para la ablación de tejido nervioso dañado o neuronas en ciertos casos. En un aspecto in vivo, el fragmento de anticuerpo o neurona de unión del mismo se marca y se administra al animal antes de, durante o después de la cirugía o una técnica quirúrgica, incluyendo una técnica estereotáctica o mínimamente invasiva, para el propósito de localizar la lesión del nervio o para evaluar quedando dañado o lesionado el tejido neural. En uno de tales aspectos, los miembros de unión específica radiomarcados, particularmente anticuerpos y fragmentos de los mismos, son útiles en técnicas de cirugía radioinmunoguiadas, en las que se pueden identificar e indicar la presencia y/o ubicación de células nerviosas comprometidas dañadas lesionadas o moribundas o neuronas o tejido nervioso, antes de, durante o después de la cirugía para apuntar, identificar o eliminar tales células.
Aplicaciones de diagnóstico de los anticuerpos y fragmentos de los mismos de la divulgación incluyen aplicaciones in vitro e in vivo conocidas y estándar para el experto en la técnica y en base a la presente descripción. Los ensayos de diagnóstico y kits para valoración y evaluación in vitro de neuronas o de tejido nervioso, se pueden utilizar para diagnosticar, evaluar y controlar las muestras de pacientes, incluyendo aquellos conocidos por tener o que se sospecha que tienen condiciones o enfermedades neurológicas en las que se comprometían células
nerviosas, dañadas o lesionadas o determinar el grado de muerte celular o lesión o de un tumor o cáncer del SNC, en particular en una muestra de un paciente o sujeto. La evaluación y la evaluación del estado de enfermedad neurológica también es útil para determinar la idoneidad de un paciente para un ensayo clínico de un medicamento o para la administración de un agente neuroterapéutico o quimioterapéutico particular, o un anticuerpo de la presente invención, incluyendo combinaciones de los mismos, frente a un agente o un anticuerpo diferente.
La presente invención incluye un sistema de ensayo que puede prepararse en forma de un kit de prueba para el análisis cuantitativo de la extensión de la presencia de, por ejemplo, neuronas dañadas, comprometidas o lesionadas o para cuantificar neuronas en una muestra. El kit de sistema o de prueba puede comprender un componente marcado preparado por una de las técnicas radiactivas y/o enzimáticas tratadas en este documento, el acoplamiento de una etiqueta para el anticuerpo, y uno o más reactivos inmunoquímicos, al menos uno de los cuales es un componente libre o inmovilizado a determinarse o su pareja de unión.
En una realización adicional de esta invención, kits de pruebas comerciales para uso por un médico especialista pueden prepararse en base a lo anterior para determinar el estado de las neuronas en una muestra. De acuerdo con las técnicas de ensayo descritas anteriormente, una clase de tales kits contendrá por lo menos el anticuerpo marcado o su pareja de unión, por ejemplo, un anticuerpo para el mismo específico, y las instrucciones, por supuesto, dependiendo del método seleccionado, por ejemplo, "competitivo, " 'sándwich', 'DASP' y similares. Los kits también pueden contener reactivos periféricos tales como tampones, estabilizadores, etc.
Por consiguiente, un kit de prueba se puede preparar para la demostración de la presencia o la determinación de la situación de las neuronas, que comprende:
(a) una cantidad predeterminada de al menos un componente reactivo marcado inmunoquímicamente obtenido por la unión directa o indirecta del presente anticuerpo o fragmento o una pareja de unión específica a la misma, a un marcador detectable;
(b) otros reactivos; e
(c) instrucciones de uso de dicho kit.
Un kit de ensayo se puede preparar para la demostración de la presencia de la lesión de las células nerviosas, dañadas o comprometidas que comprende:
(a) una cantidad predeterminada de al menos un marcado componente inmunoquímicamente reactivo obtenido por la unión directa o indirecta del presente anticuerpo o una pareja de unión específica a la misma, a un marcador detectable;
(b) otros reactivos; e
(c) instrucciones de uso de dicho kit.
Ácidos nucleicos
La presente divulgación proporciona además un ácido nucleico aislado que codifica un anticuerpo, particularmente un anticuerpo recombinante, particularmente un anticuerpo completamente humano, de la presente invención. El ácido nucleico incluye ADN y ARN. En un aspecto preferido, la presente descripción proporciona un ácido nucleico que codifica para un polipéptido de la invención como se definieron anteriormente, que incluye un polipéptido tal como se establece en las Figuras 5 o 6 o capaces de codificar las regiones CDR de los mismos.
Los anticuerpos y moléculas de ácido nucleico codificadoras y vectores pueden proporcionarse aisladas y/o purificadas, por ejemplo, de su ambiente natural, en forma sustancialmente pura u homogénea, o, en el caso del ácido nucleico, libre o sustancialmente libre de ácido nucleico o genes de origen distinto de la secuencia que codifica un polipéptido con la función requerida. El ácido nucleico de acuerdo con la presente invención puede comprender ADN o a Rn y puede ser total o parcialmente sintético.
Los sistemas para la clonación y expresión de un polipéptido en una variedad de células huésped diferentes son bien conocidos. Las células huésped adecuadas incluyen bacterias, células de mamífero, levaduras y sistemas de baculovirus. Líneas celulares de mamífero disponibles en la técnica para la expresión de un polipéptido heterólogo incluyen células de ovario de hámster chino, células HeLa, células de riñón de cría de hámster, células cancerosas, células de cáncer de ovario y muchos otros. Un huésped bacteriano común, preferido es E. coli. La expresión de anticuerpos y fragmentos de anticuerpos en células procariotas tales como E. coli está bien establecida en la técnica. Los vectores adecuados pueden elegirse o construirse, conteniendo secuencias reguladoras apropiadas, incluyendo secuencias promotoras, secuencias terminadoras, secuencias de poliadenilación, secuencias potenciadoras, genes marcadores y otras secuencias según sea apropiado. Los vectores pueden ser plásmidos, por ejemplo, 'fago, o fagémido viral, según proceda. Para más detalles, véase, por ejemplo, Molecular Cloning: A Laboratory Manual:. 2a edición, Sambrook y otros, 1989, Cold Spring Harbor Laboratory Press. Muchas técnicas conocidas y protocolos para manipulación de ácido nucleico, por ejemplo en la preparación de construcciones de ácido nucleico, génesis mutatis, secuenciación, introducción de ADN en células y expresión
génica, y análisis de proteínas, se describen en detalle en Short Protocols in Molecular Biology, Segunda Edición, Ausubel et al. eds., John Wiley & Sons, 1992.
Por lo tanto, un aspecto adicional proporciona una célula huésped que contiene ácido nucleico como se describe aquí. Un aspecto adicional proporciona un método que comprende introducir tal ácido nucleico en una célula huésped. La introducción puede emplear cualquier disposición técnica. Para las células eucariotas, las técnicas adecuadas pueden incluir transfección con fosfato de calcio, DEAE-dextrano, electroporación, transfección mediada por liposomas y transducción usando retrovirus u otros virus, por ejemplo vaccinia o, para células de insectos, baculovirus. Para células bacterianas, las técnicas adecuadas pueden incluir transformación con cloruro de calcio, electroporación y transfección usando bacteriófagos. La introducción puede ser seguida causando o permitiendo la expresión a partir del ácido nucleico, por ejemplo, cultivando células huésped bajo condiciones para la expresión del gen. La presente descripción también proporciona un método que comprende utilizar una construcción como se ha indicado anteriormente en un sistema de expresión con el fin de expresar un miembro de unión específica o polipéptido como anteriormente.
Otra característica es la expresión de las secuencias de ADN descritas en la presente memoria. Como es bien conocido en la técnica, las secuencias de ADN pueden expresarse enlazando operativamente a una secuencia de control de la expresión en un vector de expresión apropiado y empleando ese vector de expresión para transformar un huésped unicelular apropiado. Una amplia variedad de combinaciones huésped/vector de expresión puede emplearse en la expresión de las secuencias de ADN de esta invención. Vectores de expresión útiles, por ejemplo, pueden consistir en segmentos cromosómicos, no cromosómicos y secuencias de ADN sintético. Los vectores adecuados incluyen derivados de SV40 y plásmidos bacterianos conocidos, por ejemplo, plásmidos E. coli Col El, pCR1, pBR322, pMB9 y sus derivados, plásmidos tales como RP4; fago ADN, por ejemplo, los numerosos derivados del fago A, por ejemplo, NM989, y otros ADN de fago, por ejemplo, M13 y el ADN filamentoso único fago de cadena; plásmidos de levadura tales como el plásmido 2 u o derivados de los mismos; vectores útiles en células eucariotas, tales como vectores útiles en células de insecto o de mamífero; vectores derivados de combinaciones de plásmidos y ADN de fagos, tales como plásmidos que han sido modificados para emplear ADN de fago u otras secuencias de control de la expresión; y similares. Cualquiera de una amplia variedad de secuencias de control de la expresión - secuencias que controlan la expresión de una secuencia de ADN ligada operativamente a la misma -puede usarse en estos vectores para expresar las secuencias de ADN de esta invención. Tales secuencias de control de la expresión útiles incluyen, por ejemplo, los promotores tempranos o tardíos de SV40, CMV, vaccinia, polioma o adenovirus, el sistema lac, el sistema trp, el sistema TAC, el sistema TRC, el sistema LTR, el operador principal y las regiones promotoras del fago A, las regiones de control de la proteína de recubrimiento fd, el promotor para 3-fosfoglicerato quinasa u otras enzimas glicolíticas, los promotores de la fosfatasa ácida (por ejemplo, Pho5), los promotores del factor de levadura, y otras secuencias conocidas para controlar la expresión de genes de células procariotas o eucariotas o sus virus, y diversas combinaciones de los mismos.
Una amplia variedad de células huésped unicelulares son también útiles para expresar las secuencias de ADN de esta invención. Estos huéspedes pueden incluir huéspedes eucariotas y procariotas bien conocidas, tales como cepas de E. coli, Pseudomonas, Bacillus, Streptomyces, hongos tales como levaduras, y células animales, tales como células CHO, YB/20, NSO, SP2/0, células Ri.I, B-W y L-M, de riñón de mono verde africano (por ejemplo, COS 1, COS 7, BSC1, BSC40, y BMT10), células de insecto (por ejemplo, Sf9), y células humanas y células vegetales en cultivo de tejidos.
Se entenderá que no todos los vectores, secuencias de control de expresión y huéspedes funcionarán igualmente bien para expresar las secuencias de ADN de esta invención. Ni todas las huéspedes funcionarán igualmente bien con el mismo sistema de expresión. Sin embargo, un experto en la técnica será capaz de seleccionar los vectores, secuencias de control de la expresión, y huéspedes sin experimentación indebida para realizar la expresión deseada sin apartarse del alcance de esta invención. En la selección de una secuencia de control de la expresión, normalmente se considerarán una diversidad de factores. Éstos incluyen, por ejemplo, la fuerza relativa del sistema, su controlabilidad, y su compatibilidad con la secuencia de ADN o gen particular a expresar, particularmente en lo relativo al potencial de estructuras secundarias. Los huéspedes unicelulares adecuados se seleccionarán por consideración con- de, por ejemplo, su compatibilidad con el vector elegido, sus características de secreción, su capacidad para plegar proteínas correctamente y sus requisitos de fermentación, así como la toxicidad para el huésped del producto codificado por las secuencias de ADN a expresar, y la facilidad de purificación de los productos de expresión. Teniendo en cuenta estos y otros factores un experto en la técnica será capaz de construir una variedad de vector/secuencia de control de expresión combinaciones/huésped que expresarán las secuencias de ADN de esta invención sobre la fermentación o en cultivo animal a gran escala.
Una secuencia de ADN que codifica un anticuerpo o fragmento del mismo se puede preparar sintéticamente en lugar de clonarse. La secuencia de ADN puede diseñarse con los codones apropiados para la secuencia específica de aminoácidos miembro de unión. En general, se seleccionarán codones preferidos para el huésped pretendido si la secuencia se utilizará para expresión. La secuencia completa se ensambla a partir de oligonucleótidos solapantes preparados por métodos estándar y ensamblados en una secuencia codificante completa. Véase, por ejemplo, Edge, Naturaleza, 292: 756 (1981);. Nambair et al, Science, 223: 1299 (1984); Jay et
al., J. Biol. Chem., 259: 6311 (1984). Secuencias de ADN sintético permiten la construcción conveniente de genes que expresarán análogos de miembro de unión específica o "muteínas". Alternativamente, el ADN que codifica las muteínas pueden hacerse por mutagénesis dirigida de genes o ADNc miembro de unión nativo específicos, y muteínas se pueden hacer directamente utilizando síntesis de polipéptido convencional.
EJEMPLOS
La invención puede entenderse mejor por referencia a los siguientes Ejemplos no limitantes, que se proporcionan como ejemplares de la invención. Los siguientes ejemplos se presentan con el fin de ilustrar más completamente las realizaciones preferidas de la invención y no deben en ningún modo interpretarse, sin embargo, como limitantes del amplio alcance de la invención.
Ejemplo 1: Identificación de autoanticuerpos naturales (NatAbs) que se unen a las células sistémicas del sistema nervioso
IgM humano de suero natural que se unen a las neuronas se identificaron mediante el cribado del banco de suero Mayo Clinic, que contiene más de 140.000 muestras recogidas más de 45 años, para los candidatos con un alto pico monoclonal de IgG o IgM (mayor que 10 mg/ml en la sangre) y después probando el suero para la unión a rodajas de córtex vivo cerebral y cerebelo (31) de anticuerpo. Tales IgM fueron entonces purificados a partir de muestras positivas y se ensayaron adicionalmente para 1 ) la unión a la superficie de las neuronas primarias aisladas 2) como sustratos para el apoyo de la extensión de neuritas 3) la capacidad de proteger a las neuronas de la apoptosis en la cara de las moléculas de factor estresante. Este protocolo de cribado se basa en que se utiliza para identificar IgM humanos que se unen a los oligodendrocitos y promover la remielinización en modelos de MS (22, y como se describe en el documento WO 0185797). El reconocimiento de la superficie de tejidos o células apropiadas parece ser una importante característica definitoria de IgM terapéuticos.
Dos IgM de unión de la neurona humana derivada del suero novelos distintos han sido identificados (sHIgM12 y sHIgM42). Ciertas características de estos anticuerpos derivados del suero se enumeran en la TABLA 1. sHIgM12 y sHIgM42 derivados de suero se han demostrado inicialmente in vitro para apoyar la extensión de neuritas, así como la laminina de sustrato potente y para anular la inhibición de crecimiento de neuritas de mielina del SNC (23).
Otros estudios han demostrado que derivados del suero sHIgM12 mejoran la función espontánea en ratones infectados con TMEV, según la evaluación de la actividad nocturna media espontánea por hora en ratones (21). En contraste con NatAbs IgM previamente identificados (sHIgM22 y sHIgM46), ni sHIgM12 ni sHIgM42 promueven la remielinización de médula espinal.
Tabla 1 - IgM humanos terapéuticos

A menudo, se cree que los autoanticuerpos son patógenos. En contraste, como se demuestra en el presente documento, los autoanticuerpos a neuronas (sHIgM12 y sHIgM42 y anticuerpos recombinantes basados en ellas) no matan a las neuronas. En cambio, estos IgM protegen a las neuronas de la muerte, promueven la extensión de neuritas, aumentan la NAA in vivo, protegen axones in vivo en el modelo de TMEV, y mejoran la función espontánea nocturna de ratones enfermos TMEV.
Los anticuerpos de unión a neuronas IgM12 y IgM42 se dirigen a neuronas en lesiones del SNC y son aplicables para revertir la pérdida neuronal y/o mejorar los efectos de lesión neuronal o enfermedad. IgM humanos de unión a neurona representan una nueva clase de agentes terapéuticos con aplicación única para diversos enfermedades y condiciones que impliquen la neurodegeneración, lesión neuronal o muerte de las neuronas. Dichas enfermedades incluyen, sin limitación, MS, lesión de la médula espinal, esclerosis lateral amiotrófica, enfermedad de Alzheimer, lesión cerebral traumática, eventos cerebro-vasculares o accidente cerebrovascular. Estos IgM humanos han sido mínimamente antigénicos cuando se administran sistémicamente en animales.
Identificación y Caracterización de IgM humanos que se unen a la superficie de múltiples tipos de neuronas vivas:
Las muestras de suero con una alta concentración de IgG o IgM (> 10 mg/ml) fueron seleccionadas para la unión de Ab a capas neuronales en cortes de tejido del SNC in vivo (corteza y cerebelo). De 152 sueros ensayados, 17 sueros humanos fueron positivos en cortes de tejido (23).
Los anticuerpos sHIgM12 y sHIgM42 se unen a la superficie de una amplia variedad de neuronas que co etiqueta con marcadores neuronales neurofilamentos o p III tubulina. Estos incluyen células cerebelares de gránulos (23), neuronas corticales, neuronas del hipocampo, neuronas de la biopsia del lóbulo temporal humano (Figura 1), y las células ganglionares de la retina (datos no mostrados). rHIgM12 tiñe el soma, neuritas y conos de crecimiento de las neuronas del hipocampo (datos no mostrados). Esta reactividad cruzada sugiere los IgM humanos pueden actuar sobre las células del SNC afectadas en un número de condiciones/enfermedades neurológicas, tales como MS, esclerosis lateral amiotrófica, o un derrame cerebral.
Los presentes datos indican que la unión de sHIgM12 o sHIgM42 a la superficie de las neuronas es dependiente de carbohidratos. Tratamiento de sialidasa de las neuronas en cultivo eliminó la unión de ambos IgM humanos a la superficie celular, mientras que el bloqueo de la síntesis de esfingolípidos con fumonisina B 1 o eliminación de proteínas unidas a GPI con PIPLC no eliminó unión de la IgM (23). Los gangliósidos son candidatos para estos antígenos de los IgM humanos. En los experimentos de co-etiquetado, rHIgM12 co-localiza con GM1 en la membrana neuronal (véase el Ejemplo 11 y FIGURA 21C).
A pesar de que comparten ciertas características funcionales, tales como la obtención de la excrecencia de neuritas, sHIgM12 y sHIgM42 no demostraron diferencias y son cada uno de anticuerpos únicos. Esto es evidente en estudios de unión y de inmunofluorescencia; etiquetan la superficie de células granulares del cerebelo en patrones distintos. En los estudios de inmunofluorescencia de células granulares de cerebelo de rata en cultivo, los patrones de etiquetado de membrana neuronal mediante el uso de estos dos anticuerpos son distintos (FIGURA 2). Áreas cortas de membrana de neuritas están obligadas por sHIgM12 que resulta en un patrón puntiforme. Las áreas más grandes de la membrana de neuritas están enlazadas por sHIgM42 que resulta en un patrón más segmentado.
Ejemplo 2: IgM humanos protegen las neuronas corticales a partir de peróxido inducido por la muerte
IgM humanos promotores de remielinización se han demostrado para proteger los oligodendrocitos en cultivo de la activación de peróxido inducida de la caspasa 3 (33), un marcador de la apoptosis activa. Como se expone en el presente documento, sHIgM12 o sHIgM42, se evaluaron en los protocolos análogos. El sHIgM12 o sHIgM42, respectivamente, y el peróxido se añadieron juntos a cultivos de neuronas corticales de ratón primario y el grado de activación de la caspasa 3 ensayada 24 h después (FIGURA 3).
El tratamiento de las neuronas cultivadas con rHIgM12 resultó en la protección de 80% de activación de la caspasa 3. El tratamiento de las neuronas con sHIgM42 era protectora también con aproximadamente de 40% de activación de la caspasa 3. Estos resultados fueron significativamente diferentes (P <0,01) en comparación con un control de IgM humana, lo que resultó en la protección de menos del 10% de la caspasa 3 de activación.
Por lo tanto, sHIgM12 o sHIgM42 de unión a neuronas protejeron neuronas corticales de la muerte celular inducida por peróxido. Por lo tanto, cuando se proporcionó un inductor o agente de daño de las células nerviosas (o muerte), IgM12 y IgM42 impidieron que se produjera el daño independiente y significativamente (o muerte).
Ejemplo 3: Los anticuerpos recombinantes derivados de sHIgM12
Se han construido dos formas recombinantes de sHIgM12. Cada una utilizó el mismo vector de expresión como se utilizó anteriormente para el anticuerpo IgM22 recombinante (rHIgM22) para las cadenas pesadas y ligeras (22, 28, WO0185797). El vector incluye un gen DHFR seleccionable expresado bajo el control del promotor de SV40. Una forma de anticuerpo IgM12 recombinante parcialmente humana con una cadena J de ratón se construyó inicialmente como sigue, a partir de entonces produjo anticuerpo en células de línea de hibridoma F3B6 humano/ratón.
La construcción del vector de anticuerpo IgM12 recombinante (PAD12) se realizó mediante la inserción del ADNc de región variable de cadena pesada con una secuencia líder de la base de datos de nucleótidos y de ADNc completo de la cadena ligera con la secuencia líder adjunta de la base de datos de nucleótidos de una manera similar a la descrita previamente (6 ), utilizando los cebadores descritos a continuación. El vector se representa en la FIGURA 4. La secuencia de la cadena pesada y ligera utilizada para el anticuerpo IgM12 humano recombinante, con regiones variables y constantes observadas, se muestra en la FIGURA 5.
Los cebadores usados para hacer rHIgM12VH y empalmar por extensión de solapamiento (Horton RM et al (1989), Gene 77: 61-68) a una secuencia líder con un intrón (letras minúsculas) de la base de datos (M29812) son como sigue:
(1) Cebador 5'-con BspEI sitio: TCC GGA CGG TCC GGG A
(2) TCC GGA CGG TCC G ggacctcct gtgcaagaac atgaaacatc tgtggttctt ccttctcctg gtggcagctc ccagatgtga gtatctcagg gatccagaca
(3) cagg gatccagaca tggggatatg ggaggtgcct ctgatcccag ggctcactgt gggtctctct gttcacaggg gtcctgtccc aggtgcagct gcag- gagtcg ggcccaggac
(4) Cebador 3' con el sitio PAC I: CCTTAATTAAGACCTGG AGAGGCCATTCTTACCTGAG GAGACGGTGACCAGGGTTC
Los cebadores usados para hacer rHIgM12Vk y empalmar por extensión de solapamiento (soe) a la secuencia líder de la base de datos (las letras minúsculas, adhesión X59312) son los siguientes:
(1) Cebador 5'-Ck a soe lym 12 Vk a Ck: CGA ACT GTGGCT GCA C
(2) Cebador 3' Ck con Xho 1: CCGCTCGAGTATCTAACACTCTCCCCTGTT
(3 ) 5'-Lym 12 Vk con Nhe I: AGCATTACTAGCTAGCTC AAGACTCAGCCTGGAC atggaca tgagggtccc cgctcagctc ctggggctcc tgctactctggctccgag gtgccaga tgt GAC ATC CAG ATG ACC CA
El vector con los genes de anticuerpos sintéticos se introdujo en células de hibridoma F3B6 por electroporación y metotrexato (MTX) de amplificación se realizó como se ha descrito previamente (6).
Brevemente, 8 millones de células de heterohibridoma F3B6 de ratón/humano (American Type Culture Collection: ATCC) se incubaron con 10 pg de vector PAD12 linealizado con Bgl II durante 10 minutos en 800 pl medio libre de suero momento en el que se sometieron a electroporación en 02 V en un Biorad Gene Pulser™ (Biorad, Hercules, CA, EE.UU.). Siguiendo una incubación de 10 min en hielo, las células se diluyeron a 24 ml en RPMI-1640 que contenía suero de ternera fetal al 10% (FC) (Gibco, Carlsbad, CA, EE.UU.), se sembraron en una placa de 24 pocillos y se incubaron a 37°C; 48 h más tarde, se seleccionaron las células que contienen vector usando 1 pM metotrexato (Calbiochem, La Jolla, CA, EE.UU.). Después de un período de incubación de 2 semanas, las colonias se recogieron a una nueva placa y se dejaron crecer hasta la confluencia. En este punto, el sobrenadante se recogió y se ensayó por ensayo de inmunoabsorción enzimática (ELISA) para la presencia de IgM humana. Las colonias positivas se seleccionaron adicionalmente en el transcurso de 2 meses con dosis crecientes de metotrexato que van desde 1 pM a 200 pM. De esta manera, se generaron células que producen más que 10 pg/ml Igm.
El primer anticuerpo recombinante construido como anteriormente no es totalmente humano. Para generar una forma completamente humana, células CHO (GibcoBRL n° de cat 11619) fueron co-transfectadas con el vector que codifica la cadena pesada y ligera recombinante bajo el control de un promotor de E1A y una construcción que expresa la cadena J humana en pCI (Promega). Se seleccionaron las células con dosis crecientes de metotrexato en una mezcla 50/50 de PowerCho1 y IMDM con 10% clon cósmico y los dos clones que produjeron la mayoría de anticuerpo medido por ELISA fueron sub-clonados. Los sub-clones se expandieron y viales se congelaron hacia abajo. Ambas formas IgM12 recombinantes mantienen la unión de la neurona y el carácter de eficacia in vivo del suero aislado IgM, pero tienen una vida media más corta en ratones (que puede ser debido a diferencias en la glicosilación).
Un procedimiento similar y comparable se utilizó para construir anticuerpo HIgM42 recombinante en el vector comparable. La secuencia de la cadena pesada y ligera del anticuerpo IgM42 humano, con regiones variables y constantes observadas, se muestra en la FIGURA 6.
Ejemplo 4: Dosis periférica individual de rHIgM12 mejora la función neurológica en modelo de TMEV de MS
En vista del hecho de que rHIgM12 protegió neuronas primarias en cultivo, y que la discapacidad en el modelo de TMEV de la esclerosis múltiple (MS) se correlaciona con la pérdida de axón (2), el tratamiento de ratones infectados con TMEV con rHIgM12 se utilizó para evaluar la capacidad para retardar la progresión de los déficits neurológicos.
Los grupos de 5 ratones a los 90 días después de la infección TMEV, el punto en el que los axones comienzan a ser perdidos, fueron tratados con una única dosis de 100 pg de rHIgM12 o un control de IgM humano. Cinco ratones infectados fueron seleccionados al azar para cada grupo y grabaciones funcionales de pretratamiento utilizadas para estar seguro de que la actividad de línea de base no era diferente entre los grupos. Los ratones fueron seguidos como grupos durante 3 días consecutivos más de varias semanas utilizando cajas de actividad (34, 35). Los cambios en el comportamiento nocturno son medidos sensibles de déficit neurológico en la enfermedad mediada por TMEV. Cajas de actividad son cajas de acrílico transparente con rayos infrarrojos opuestos que crean una cuadrícula en todo el recinto que registra todos los movimientos horizontales y verticales. La sensibilidad del ensayo refleja la capacidad de medir el encabritarse y caminar. En los ratones infectados con TMEV las extremidades posteriores se vuelven rígidas y se reduce el encabritarse. Sin embargo, el caminar espontáneamente por la jaula no está tan gravemente afectado. Es más fácil para los ratones con extremidades traseras rígidas para
caminar que para encabritarse, e incluso ratones con enfermedad avanzada pueden ser muy activos durante la noche.
Cada grupo de tratamiento fue ensamblado al azar y alojado en cajas de actividad durante 72 h antes del tratamiento, y luego durante 72 horas cada semana después del tratamiento. Las interrupciones medias de haz por hora se calcularon para la actividad horizontal y vertical sobre las 1 2 h de 18:00-6:00 durante el período de análisis de 72 h. No hubo diferencias entre grupos de tratamiento en la actividad horizontal o vertical durante el día, que era típicamente por debajo de 600 interrupciones/h mientras que duermen los ratones. Sin embargo, un aumento en la actividad horizontal nocturna espontánea en comparación con su línea de base de pre-tratamiento se registró en el grupo de rHIgM12 tratado (P <0,01, semanas 3 a 7) (Figura 7). Un ser humano IgM de control que no se une a las células no mejoró la actividad.
En contraste con el efecto visto con suero derivado de sHIgM12, y ahora también anticuerpo recombinante rIgM12, en un estudio similar el anticuerpo sHIgM42 humano no alteró la actividad nocturna de ratones infectados con TMEV en las mismas condiciones. Parámetros de dosificación alternativos para sHIgM42 en el mismo formato de ensayo producen resultados diferentes y más positivos en la actividad nocturna.
Una posible explicación de la mejora de la función es que las IgM eficaces interfieren con cargas de virus que se traducen en menos enfermedad. Esto no parece ser la explicación, sin embargo. Los ratones con enfermedad TMEV crónica fueron tratados con una dosis única de rHIgM12, sHIgM42 o IgM de control, el cerebro y la médula espinal se cosecharon 5 semanas más tarde y entonces el nivel de transcripciones genómicas ARN TMEV se midieron por PCR con una sonda para Proteína Viral 2. Transcripciones virales no eran diferentes grupos a través de (P <0,01) (datos no mostrados).
Ejemplo 5: Niveles de NAA dentro del tronco encefálico de los ratones con la enfermedad de la médula espinal - un indicador no invasivo de sustituto de Preservación de Axon a lo largo de la médula espinal
NAA es un metabolito asociado con la función de la neurona (36, 37). NAA es el segundo aminoácido más abundante en el cerebro y se limita casi exclusivamente a las neuronas. La preservación de los niveles de NAA en el tronco cerebral es una medida de la salud del axón de la médula espinal en general validado por nuestro grupo utilizando el modelo de ratón TMEV (8 ). Cuando los axones en los niveles inferiores de la médula espinal están dañados, las células en la base del cerebro mueren, reduciendo NAA. Niveles de NAA medidos por MRS refleja densidad principalmente de neurona. NAA se expresa en otras células neurales, pero la expresión primaria de nAa es en las neuronas. Los estudios del perfil MRS de células del SNC purificados indican que la amplitud de señal NAA se predomina en las neuronas, mientras que la amplitud de la señal NAA de los oligodendrocitos o astrocitos fueron del 5% y 10% de la señal de la neurona, respectivamente (38).
Para evaluar adicionalmente la capacidad de sHIgM12 y IgM42 para mejorar la actividad en ratones mediante la preservación de la función axón, se utilizó un ensayo de formación de imágenes no invasiva y la morfología tradicional para evaluar los axones de la médula espinal. El rastreo retrógrado se utilizó para demostrar la disfunción de axón de la médula espinal después de la desmielinización en la enfermedad mediada por TMEV (5). Se midió una reducción dramática de la etiqueta retrógrada desde los axones de nivel torácico a tronco cerebral nucleico. El tronco cerebral es donde muchos de los cuerpos celulares residen que proyectan extensiones largas de axones a lo largo de la médula espinal. A partir de entonces los niveles de N-acetilo aspartato (NAA) en el tronco cerebral se evaluaron mediante espectroscopía de resonancia magnética (MRS) en ratones de TMEV (FIGURA 8 ) infectados usando un protocolo previamente informado (8 ).
Se ha observado una reducción en el tronco cerebral NAA lo largo del tiempo en ratones con la enfermedad inducida por TMEV (Figura 9). Niveles de NAA cayeron durante los primeros 45 días de la infección y se mantuvieron bajos a 270 días después de la infección. En los ratones SJL infectados con TMEV, la extensión de la desmielinización de la médula espinal se equilira 90 días después de la infección (39). Es en este punto de tiempo que la pérdida de axon es notable por la histología en este modelo (2). Por lo tanto NAA es una medida sensible de la disfunción de axón.
Después de la última colección de MRS, los axones fueron muestreados sistemáticamente de 6 áreas de materia blanca de apariencia normal dentro de una sección transversal de la médula espinal a nivel T6 . Este nivel fue elegido porque esto proporcionó una representación global de la pérdida axonal de lesiones desmielinizadas múltiples, distribuidas al azar en toda la médula espinal (39). Nos encontramos que había 30,5% menos de axones en ratones SJL infectados con TMEV de 270 días en comparación con los controles no infectados (p<0,001). Se encontró una correlación positiva que existe entre los niveles de NAA del tronco cerebral y los recuentos de los axones en el nivel T6 de la médula espinal (r = 0,823) (8 ).
Dosis única de IgM humana de unión de neurona preserva los niveles de NAA en el tronco cerebral y los axones en la médula espinal:
La capacidad de las IgM de unión a neuronas humanas para alterar los niveles de NAA o recuentos de axones en ratones infectados con TMEV se evaluó. Se encontró que cuando los ratones TMEV infectados en el inicio de axón de médula espinal (90 días después de la infección) fueron tratados con una sola dosis de 100 pg de IgM de unión de neurona de sHIgM12 o de sHIgM42, la densidad de axones mielinizados en la materia blanca normal de unión de la médula espinal torácica fue mayor cuando se mide 1 0 semanas más tarde.
Los grupos de 10-15 ratones a los 90 días después de la infección se trataron con una única dosis de 100 pg de sHIgM12, sHIgM42, IgM de control humano o solución salina (Figura 10). Antes del tratamiento y a las 5 y 10 semanas más tarde, cada ratón se colocó en un pequeño imán diámetro y MRS recogido en el tronco cerebral. En murieron 10 sem ratones, las médulas espinales recogido, secciones de plástico incrustado y Cross Cut en el nivel T6 teñido con paraphenylamindiamine para visualizar las vainas de mielina, se recogieron 6 imágenes por sección que abarcan 400.000 pM de apariencia normal de la materia blanca y la auto axones contados. En los grupos tratados con cualquiera de los dos IgM de unión a neuronas (sHIgM12, sHIgM42), los niveles de NAA se incrementaron en 5 y 10 semanas más tarde en comparación con los niveles de pretratamiento. Los ratones tratados con IgM de control mostraron una tendencia más baja y los ratones tratados con solución salina se mantuvieron constantes.
Niveles de NAA de grupos tratados con sHIgM12 y sHIgM42 se aumentaron a 9,13 y 9,3 mM, respectivamente, cada uno de los cuales estaba bien por debajo de los niveles de 12,0 mM de ratones no infectados. Cuando los axones en el nivel T6 se compararon entre los grupos de tratamiento (Tabla 2), los ratones tratados con sHIgM12 o sHIgM42 contenían más axones que el grupo tratado con solución salina (17.303 y 17.771 axones en comparación con 15.198 axones, P = 0,008 y P <0,001), pero menos que el número de axones contados en los ratones no infectados (21.284 axones).
Estos resultados indicaron que cada uno de los anticuerpos sHIgM12 y rHIgM12 mejoraron la función mediante la preservación de los axones, como se evidencia en el modelo de TMEV. A pesar de los axones de anticuerpos sHIgM42 conservados en el modelo de TMEV, cambios demostrables en la evaluación de la actividad nocturna no se observaron en la prueba inicial. Después de los estudios de rango de dosis, se encuentra que sHIgM42 también logra mayor actividad en el ensayo de la actividad nocturna.
El uso de MRS de tronco cerebral para evaluar el estado del axón en modelos de ratón de la enfermedad de la médula espinal valida aún más la presente invención; en consecuencia, NAA en el tronco cerebral sirve como un excelente punto final para el uso de ensayos clínicos de estos anticuerpos humanos.
Los ratones tratados con sHIgM12 sHIgM42 y con mejores niveles de NAA también contenían más axones en la médula espinal torácica media. Grupos de 10-15 ratones SJL infectados con TMEV fueron tratados con una única dosis de 100 pg de rHIgM22, sHIgM42, sHIgM12, sHIgM39 de control y solución salina a los 90 días después de la infección. Diez semanas después del tratamiento, las médulas espinales fueron retiradas y una sección mediatorácica se tiñeron con PPD para visualizar la mielina. Seis áreas que abarcan 400.000 pM2 de la materia blanca se tomaron muestras de cada ratón y el número de axones mielinizados contados (1). El promedio de número absoluto de axones mielinizados por sección transversal T6 + SEM aparece en la Tabla 2.
TABLA 2 Tratamiento con IgM de unión a neurona conserva recuento de axon en la médula espinal

Ejemplo 6: IgM de unión a neurona conserva axones sin promover la remielinización
Se han identificado varios IgM humanos (por ejemplo, sHIgM22 y sHIgM46) que se unen a oligodendrocitos y promueven la remielinización (22, 40, 41). Como se muestra abajo, la neurona de unión a IgM mejora los números de neuronas y la función aún sin remielinización obvia. Sin estar ligado por la teoría, el mecanismo de acción se entiende que es debido a la activación directa de los axones (protección, extensión de neuritas) y/o mediante la activación del sistema inmune innato o adaptativo para secretar factores que protegen a las neuronas. Los resultados presentados anteriormente demuestran claramente que los anticuerpos tienen un efecto directo sobre axones/neuronas. Dentro de la misma médula espinal en la que preservación de axón mediada por sHIgM12 y sHIgM42 y/o recrecimiento se midió, desmielinización general, la remielinización y la inflamación no fueron diferentes entre los grupos de tratamiento (Figura 11).
Estos atributos se cuantificaron mediante clasificación de los cuadrantes de 10 secciones transversales de la médula espinal (6 ) que representan las muestras a lo largo de la longitud de la médula espinal. El grupo tratado con rHIgM22 (control positivo) mostró el aumento esperado en la remielinización, mientras que ratones tratados con sHIgM12 y sHIgM42 contenían poca remielinización de médula espinal. En cons ecuencia, los déficits neurológicos en el modelo de TMEV se mejoraron (por ejemplo, IgM12) sin el requisito de la remielinización significativa; por otra parte, la remielinización no es necesaria para la preservación del axón y/o recrecimiento dentro del marco de tiempo examinado.
Ejemplo 7: Vida media en suero de rHIgM12 (con la cadena J humana) y sHIgM42
Para determinar la vida media de los IgM humanos, rHIgM12 (con la cadena J humana) y sHIgM42, 100 pg de rHIgM12 que contiene la cadena J humana o 100 pg de sHIgM42 en 200 pl de solución salina se inyectó en la vena de la cola de ratones CD-1 normal (Figura 12). A intervalos definidos (15 min, 1, 4, 8 , 24, 48 h) se recogió sangre de grupos de 3 ratones por punción cardiaca. Se recogió el suero y se ensayó para la presencia de la cadena mu de IgM humana utilizando un sándwich ELISA.
Para rHIgM12 la vida media entre la primera colección en 15 min y la colección en 8 h fue 3,8 h. Para sHIgM42 la vida media entre la primera colección en 15 min y la colección a las 24 h fue de 20,5 h. Estos valores agrupan el vida media de la rHIgM22 de promoción de remielinización que tiene una vida media en ratones de 15 h (29) y en conejos de 90 h. La vida media se calculó utilizando las fórmulas Kelim = (In(cpico)-In(cbajo))/tintervalo y T1/2 = 0,693/kelim.
Ejemplo 8: IgM humano radiomarcado monoclonal curza la barrera hematoencefálica
A menudo se ha aceptado que las IgM con un peso molecular de cerca de 1 millón puede ser demasiado grande para atravesar la barrera hematoencefálica (BBB) de la circulación y de este modo entrar en el SNC (42, 43). Hay cierta evidencia, sin embargo, que algunos IgM hacen cruzar la barrera hematoencefálica.
Se midió la distribución tisular de rHIgM12 etiquetado 35S en ratones SJL normales e infectados con TMEV (FIGURA 13). 50 pg de rHIgM12 (1 X 107 cpm) se administró ip. Los ratones se perfundieron con solución salina 4 o 24 horas más tarde, los tejidos se cosecharon de forma aguda, se picaron y se disolvieron en el líquido de centelleo. El cerebro y la médula espinal de ratones no infectados contenían radiomarcador en ambos puntos temporales. El SNC de ratones infectados con TMEV contenía el doble de radiomarcador en el punto de tiempo de 4 h como ratones no infectadas - esto se aumentó a 4x a 24 h.
También se encuentra que rHIgM12 (tanto con como sin cadena J humana) y sHIgM42 son capaces de atravesar la BBB en ratones normales y SAMP8 , que presentan enfermedad de Alzheimer. En consecuencia, IgM humanos etiquetados con 125I se inyectan i.v. y cerebros recogidos dos horas más tarde. Cada uno de estos anticuerpos se acumula en el cerebro de ratones normales y enfermos. Estos anticuerpos también invierten el deterioro cognitivo en el modelo de ratón de la enfermedad de Alzheimer si la IgM es administrada por inyección intracerebral o i.v.
Ejemplo 9: IgM de unión de neurona humana administradas intraperitonealmente introducen las lesiones de la médula espinal desmielinizadas y localizar a axones positivos neurofilamentos
El estudio de la remielinización marcada isotópicamente que promueve IgM de ratón, SCH94.03, por Hunter (44) utilizó autorradiografía para demostrar radiomarcador localizado in vivo en la médula espinal de ratones infectados con TMEV, específicamente a las células que fueron identificadas ultraestructuralmente como oligodendrocitos. Los estudios de autorradiografía similares con 35S rHIgM12 se han realizado. El uso de inmunocitoquímica tradicional, hemos detectado neurona IgM humanos de unión de neurona dentro de las lesiones de la médula espinal (Figura 14).
Por consiguiente, se administraron 1,0 mg de rHIgM12 (con la cadena J humana), sHIgM42 o IgM humana de control comercial (Jackson Immuno Research) por vía intraperitoneal a ratones infectados con TMEV con desmielinización crónica. Cuatro horas más tarde los ratones fueron perfundidos con paraformaldehído, las médulas espinales congelados se seccionaron longitudinalmente y se inmunotiñeron para detectar la presencia de la cadena mu IgM humana. En los ratones que recibieron rHIgM12 o sHIgM42 se localizó la cadena mu humana en las lesiones desmielinizadas en secciones paralelas, lo que sugiere fibras de axones. No se encontró el control de IgM humana dentro de las lesiones o el cable no lesionado. Por lo tanto, estaba claro que las IgM humanas de neuronas de unión rHIgM12 o sHIgM42 cruzan la BBB en ratones infectados con TMEV.
Secciones de la médula espinal adyacentes fueron entonces immunoetiquetadas con anti-neurofilamento (NF) Abs (SMI-32 y 34, Sternberger) seguido de cadena-FITC mu Abs secundaria anti-humana fluorescente y antiratón-TRITC. Microscopía confocal demostró que rHIgM12 y sHIgM42 se co-localizaron en los axones NF dentro
de las lesiones, en pistas paralelas de fibras y, como haces de fibras de axones cortados en el extremo (FIGURA 15).
Ejemplo 10: rHIgM12 o sHIgM42 no exacerben EAE inducida por péptido MOG cuando se administra en la aparición de la enfermedad
Para abordar las preocupaciones de que la administración de IgM de unión del SNC autorreactivos a los animales con la autoinmunidad activa puede exacerbar la enfermedad, los efectos de rHIgM12 y sHIgM42 se probaron en ratones con EAE. Grupos de 10 ratones C57BL6 con EAE inducida por el péptido MOG (200 pg) se les administró una sola dosis de 100 pg de rHIgM12, sHIgM42, controlan IgM o solución salina humana por vía intravenosa. Ratones individuales se trataron en el momento que su puntuación clínica alcanzó 1 (cola cojera). Los ratones fueron seguidos por un investigador ciego a los grupos de tratamiento que registraron los pesos y las puntuaciones clínicas graduadas cada dos días hasta que los ratones alcanzaron los 28 días después de la inmunización o estaban moribundos.
No hubo diferencias entre grupos de tratamiento en los pesos, o puntuaciones clínicas medias (FIGURA 16, P = 0,14). Además 10 secciones transversales de la médula espinal que abarcan toda la médula espinal de cada ratón se clasifican a ciegas para la presencia de inflamación meníngea y desmielinización en cada cuadrante de la médula espinal (6 ). No hubo diferencias en el porcentaje de cuadrantes con inflamación meníngea (P = 0,825) o la desmielinización (P = 0,766) entre los grupos de tratamiento (Figura 17). Por lo tanto se encontró que una dosis única de IgM humanos de neurona que son eficaces en la protección de los axones en el modelo de TMEV no empeora déficits clínicos de EAE, ni acelera la progresión de déficit ni aumenta la patología de la médula espinal de unión.
REFERENCIAS
1. Howe, C. L., J. D. Adelson, y M. Rodriguez. 2007. Absence of perforin expression confers axonal protection despite demyelination. Neurobiol Dis 25:354-359.
2. McGavern, D. B., P. D. Murray, C. Rivera-Quinones, J. D. Schmelzer, P. A. Low, y M. Rodriguez. 2000. Axonal loss results in spinal cord atrophy, electrophysiological abnormalities and neurological deficits following demyelination in a chronic inflammatory model of multiple sclerosis. Brain 123 Pt 3:519-531.
3. Jones, M. V., T. T. Nguyen, C. A. Deboy, J. W. Griffin, K. A. Whartenby, D. A. Kerr, y P. A. Calabresi. 2008. Behavioral and pathological outcomes in MOG 35-55 experimental autoimmune encephalomyelitis. Journal of ne roimmunology.
4. Rodriguez, M., E. Oleszak, y J. Leibowitz. 1987. Theiler's murine encephalomyelitis: a model of demyelination and persistence of virus. Crit Rev Immunol 7:325-365.
5. Ure, D. R., y M. Rodriguez. 2002. Preservation of neurologic function during inflammatory demyelination correlates with axon sparing in a mouse model of multiple sclerosis. Neuroscience 111:399-411.
6 . Mitsunaga, Y., B. Ciric, V. Van Keulen, A. E. Warrington, M. Paz Soldan, A. J. Bieber, M. Rodriguez, y L. R. Pease. 2002. Direct evidence that a human antibody derived from patient serum can promote myelin repair in a mouse model of chronic-progressive demyelinating disease. Faseb J 16:1325-1327.
7. Van Keulen, V. P., B. Ciric, S. Radhakrishnan, K. L. Heckman, Y. Mitsunaga, K. lijima, H. Kita, M. Rodriguez, y L. R. Pease. 2006. Immunomodulation using the recombinant monoclonal human B7-DC cross-linking antibody rHIgM12. Clin Exp Immunol 143:314-321.
8. Denic, A., A. Bieber, A. Warrington, P. K. Mishra, S. Macura, y M. Rodriguez. 2009. Brainstem (1)H nuclear magnetic resonance (NMR) spectroscopy: Marker of demyelination and repair in spinal cord. Annals of neurology 66:559-564.
9. Duncan, I.D. 1996. Glial cell transplantation and remyelination of the central nervous system. Neuropathol Appl Neurobiol 22:87-100.
10. Fernandez, M., A. Giuliani, S. Pirondi, G. D'Intino, L. Giardino, L. Aloe, R. Levi-Montalcini, y L. Calza. 2004. Thyroid hormone administration enhances remyelination in chronic demyelinating inflammatory disease. Proc Natl Acad Sci U S A 101:16363-16368.
11. Craig, C. G., V. Tropepe, C. M. Morshead, B. A. Reynolds, S. Weiss, y D. van der Kooy. 1996. In vivo growth factor expansion of endogenous subependymal neural precursor cell populations in the adult mouse brain. J Neurosci 16:2649-2658.
12. Rodriguez, M. 2007. Effectors of Demyelination and Remyelination in the CNS: Implications for Multiple Sclerosis. Brain Pathol 17:219-229.
13. Pitt, D., P. Werner, y C. S. Raine. 2000. Glutamate excitotoxicity in a model of multiple sclerosis. Nat Med 6:67-70.
14. Okuda, Y., S. Sakoda, H. Fujimura, y T. Yanagihara. 1997. Nitric oxide via an inducible isoform of nitric oxide synthase is a possible factor to eliminate inflammatory cells from the central nervous system of mice with experimental allergic encephalomyelitis. Journal of neuroimmunology 73:107-116.
15. Waxman, S. G. 2002. Sodium channels as molecular targets in multiple sclerosis. J Rehabil Res Dev 39:233-242.
16. Irvine, K. A., y W. F. Blakemore. 2008. Remyelination protects axons from demyelination-associated axon
degeneration. Brain 131:1464-1477.
17. Linker, R. A., M. Maurer, S. Gaupp, R. Martini, B. Holtmann, R. Giess, P. Rieckmann, H. Lassmann, K. V. Toyka, M. Sendtner, y R. Gold. 2002. CNTF is a major protective factor in demyelinating CNS disease: a neurotrophic cytokine as modulator in neuroinflammation. Nat Med 8:620-624.
18. Chang, A., W. W. Tourtellotte, R. Rudick, y B. D. Trapp. 2002. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N Engl J Med 346:165-173.
19. McTigue, D. M., R. Tripathi, y P. Wei. 2006. NG2 colocalizes with axons and is expressed by a mixed cell population in spinal cord lesions. J Neuropathol Exp Neurol 65:406-420.
20. Franklin, R. J., G. L. Hinks, R. H. Woodruff, y M. T. O'Leary. 2001. What roles do growth factors play in CNS remyelination? Prog Brain Res 132:185-193.
21. Rodriguez, M., A. E. Warrington, y L. R. Pease. 2009. Invited Article: Human natural autoantibodies in the treatment of neurologic disease. Neurology 72:1269-1276.
22. Warrington, A. E., K. Asakura, A. J. Bieber, B. Ciric, V. Van Keulen, S. V. Kaveri, R. A. Kyle, L. R. Pease, y M. Rodriguez. 2000. Human monoclonal antibodies reactive to oligodendrocytes promote remyelination in a model of multiple sclerosis. Proc Natl Acad Sci U S A 97:6820-6825.
23. Warrington, A. E., A. J. Bieber, V. Van Keulen, B. Ciric, L. R. Pease, y M. Rodriguez. 2004. Neuron-binding human monoclonal antibodies support central nervous system neurite extension. J Neuropathol Exp Neurol 63:461-473.
24. Radhakrishnan, S., L. T. Nguyen, B. Ciric, D. Flies, V. P. Van Keulen, K. Tamada, L. Chen, M. Rodriguez, y L. R. Pease. 2004. Immunotherapeutic potential of B7-DC (PD-L2) cross-linking antibody in conferring antitumor immunity. Cancer Res 64:4965-4972.
25. Cohen, I. R. 2007. Biomarkers, self-antigens and the immunological homunculus. J Autoimmun 29:246-249.
26. Cohen, I. R., y M. Schwartz. 1999. Autoimmune maintenance and neuroprotection of the central nervous system. Journal of neuroimmunology 100:111-114.
27. Lutz, H. U. 2007. Homeostatic roles of naturally occurring antibodies: an overview. J Autoimmun 29:287-294.
28. Bieber, A. J., A. Warrington, K. Asakura, B. Ciric, S. V. Kaveri, L. R. Pease, y M. Rodriguez. 2002. Human antibodies accelerate the rate of remyelination following lysolecithin-induced demyelination in mice. Glia 37:241-249.
29. Warrington, A. E., A. J. Bieber, B. Ciric, L. R. Pease, V. Van Keulen, y M. Rodriguez. 2007. A recombinant human IgM promotes myelin repair after a single, very low dose. J Neurosci Res 85:967-976.
30. Pirko, I., B. Ciric, J. Gamez, A. J. Bieber, A. E. Warrington, A. J. Johnson, D. P. Hanson, L. R. Pease, S. I. Macura, y M. Rodriguez. 2004. A human antibody that promotes remyelination enters the CNS and decreases lesion load as detected by T2-weighted spinal cord MRI in a virus-induced murine model of MS. Faseb J 18:1577-1579.
31. Warrington, A. E., y S. E. Pfeiffer. 1992. Proliferation and differentiation of O4+ oligodendrocytes in postnatal rat cerebellum: analysis in unfixed tissue slices using anti-glycolipid antibodies. J Neurosci Res 33:338-353. 32. Ciric, B., V. Van Keulen, M. Paz Soldan, M. Rodriguez, y L. R. Pease. 2004. Antibody-mediated remyelination operates through mechanism independent of immunomodulation. Journal of neuroimmunology 146:153-161. 33. Howe, C. L., A. J. Bieber, A. E. Warrington, L. R. Pease, y M. Rodriguez. 2004. Antiapoptotic signaling by a remyelination-promoting human antimyelin antibody. Neurobiol Dis 15:120-131.
34. Rivera-Quinones, C., D. McGavern, J. D. Schmelzer, S. F. Hunter, P. A. Low, y M. Rodriguez. 1998. Absence of neurological deficits following extensive demyelination in a class I-deficient murine model of multiple sclerosis. Nature medicine 4:187-193.
35. McGavern, D. B., L. Zoecklein, K. M. Drescher, y M. Rodriguez. 1999. Quantitative assessment of neurologic deficits in a chronic progressive murine model of CNS demyelination. Exp Neurol 158:171-181.
36. De Stefano, N., P. M. Matthews, L. Fu, S. Narayanan, J. Stanley, G. S. Francis, J. P. Antel, y D. L. Arnold.
1998. Axonal damage correlates with disability in patients with relapsing-remitting multiple sclerosis. Results of a longitudinal magnetic resonance spectroscopy study. Brain 121 (Pt 8):1469-1477.
37. Tourbah, A., C. Linnington, C. Bachelin, V. Avellana-Adalid, H. Wekerle, y A. Baron-Van Evercooren. 1997. Inflammation promotes survival and migration of the CG4 oligodendrocyte progenitors transplanted in the spinal cord of both inflammatory and demyelinated EAE rats. J Neurosci Res 50:853-861.
38. Manganas, L. N., X. Zhang, Y. Li, R. D. Hazel, S. D. Smith, M. E. Wagshul, F. Henn, H. Benveniste, P. M. Djuric, G. Enikolopov, y M. Maletic-Savatic. 2007. Magnetic resonance spectroscopy identifies neural progenitor cells in the live human brain. Science 318:980-985.
39. McGavern, D. B., P. D. Murray, y M. Rodriguez. 1999. Quantitation of spinal cord demyelination, remyelination, atrophy, and axonal loss in a model of progressive neurologic injury. J Neurosci Res 58:492-504.
40. Asakura, K., D. J. Miller, L. R. Pease, y M. Rodriguez. 1998. Targeting of IgMkappa antibodies to oligodendrocytes promotes CNS remyelination. J Neurosci 18:7700-7708.
41. Asakura, K., D. J. Miller, K. Murray, R. Bansal, S. E. Pfeiffer, y M. Rodriguez. 1996. Monoclonal autoantibody SCH94.03, which promotes central nervous system remyelination, recognizes an antigen on the surface of oligodendrocytes. J Neurosci Res 43:273-281.
42. Kozlowski, G. P., I. Sterzl, and G. Nilaver. 1992. Localization patterns for immunoglobulins and albumins in the brain suggest diverse mechanisms for their transport across the blood-brain barrier (BBB). Prog Brain Res 91:149-154.
43. Banks, W. A. 2008. Developing drugs that can cross the blood-brain barrier: applications to Alzheimer's
disease. BMC Neurosci 9 Suppl 3:S2.
44. Hunter, S. F., D. J. Miller, y M. Rodriguez. 1997. Monoclonal remyelination-promoting natural autoantibody SCH 94.03: pharmacokinetics and in vivo targets within demyelinated spinal cord in a mouse model of multiple sclerosis. J Neurol Sci 150:103-113.
45. Banks, W. A., S. A. Farr, J. E. Morley, K. M. Wolf, V. Geylis, y M. Steinitz. 2007. Anti-amyloid beta protein antibody passage across the blood-brain barrier in the SAMP8 mouse model of Alzheimer's disease: an agerelated selective uptake with reversal of learning impairment. Exp Neurol 206:248-256.
46. Schenk, D., R. Barbour, W. Dunn, G. Gordon, H. Grajeda, T. Guido, K. Hu, J. Huang, K. Johnson-Wood, K. Khan, D. Kholodenko, M. Lee, Z. Liao, I. Lieberburg, R. Motter, L. Mutter, F. Soriano, G. Shopp, N. Vasquez, C. Vandevert, S. Walker, M. Wogulis, T. Yednock, D. Games, y P. Seubert. 1999. Immunization with amyloid-beta attenuates Alzheimer-disease-like pathology in the PDAPP mouse. Nature 400:173-177.
47. Bard, F., C. Cannon, R. Barbour, R. L. Burke, D. Games, H. Grajeda, T. Guido, K. Hu, J. Huang, K. Johnson-Wood, K. Khan, D. Kholodenko, M. Lee, I. Lieberburg, R. Motter, M. Nguyen, F. Soriano, N. Vasquez, K. Weiss, B. Welch, P. Seubert, D. Schenk, y T. Yednock. 2000. Peripherally administered antibodies against amyloid betapeptide enter the central nervous system and reduce pathology in a mouse model of Alzheimer disease. Nat Med 6:916-919.
48. Mikami, Y., M. Toda, M. Watanabe, M. Nakamura, Y. Toyama, y Y. Kawakami. 2002. A simple and reliable behavioral analysis of locomotor function after spinal cord injury in mice. Technical note. J Neurosurg 97:142-147.
49. Mikami, Y., H. Okano, M. Sakaguchi, M. Nakamura, T. Shimazaki, H. J. Okano, Y. Kawakami, Y. Toyama, y M. Toda. 2004. Implantation of dendritic cells in injured adult spinal cord results in activation of endogenous neural stem/progenitor cells leading to de novo neurogenesis and functional recovery. J Neurosci Res 76:453-465.
50. Gilgun-Sherki, Y., H. Panet, V. Holdengreber, R. Mosberg-Galili, y D. Offen. 2003. Axonal damage is reduced following glatiramer acetate treatment in C57/bl mice with chronic-induced experimental autoimmune encephalomyelitis. Neurosci Res 47:201-207.
51. Genain, C. P., B. Cannella, S. L. Hauser, y C. S. Raine. 1999. Identification of autoantibodies associated with myelin damage in multiple sclerosis. Nat Med 5:170-175.
52. Genain, C. P., M. H. Nguyen, N. L. Letvin, R. Pearl, R. L. Davis, M. Adelman, M. B. Lees, C. Linington, y S. L. Hauser. 1995. Antibody facilitation of multiple sclerosis-like lesions in a nonhuman primate. J Clin Invest 96:2966-2974.
53. Lennon, V. A., D. M. Wingerchuk, T. J. Kryzer, S. J. Pittock, C. F. Lucchinetti, K. Fujihara, I. Nakashima, y B. G. Weinshenker. 2004. A serum autoantibody marker of neuromyelitis optica: distinction from multiple sclerosis. Lancet 364:2106-2112.
54. Rodriguez, M., W. E. Karnes, J. D. Bartleson, y A. A. Pineda. 1993. Plasmapheresis in acute episodes of fulminant CNS inflammatory demyelination. Neurology 43:1100-1104.
55. Herrero-Herranz, E., L. A. Pardo, R. Gold, y R. A. Linker. 2008. Pattern of axonal injury in murine myelin oligodendrocyte glycoprotein induced experimental autoimmune encephalomyelitis: implications for multiple sclerosis. Neurobiol Dis 30:162-173.
56. Yednock, T. A., C. Cannon, L. C. Fritz, F. Sanchez-Madrid, L. Steinman, y N. Karin. 1992. Prevention of experimental autoimmune encephalomyelitis by antibodies against alpha 4 beta 1 integrin. Nature 356:63-66. 57. Kanwar, J. R., J. E. Harrison, D. Wang, E. Leung, W. Mueller, N. Wagner, y G. W. Krissansen. 2000. Beta7 integrins contribute to demyelinating disease of the central nervous system. Journal of neuroimmunology 103:146-152.
58. Warrington, A. E., y M. Rodriguez. 2008. Remyelination-promoting human IgMs: developing a therapeutic reagent for demyelinating disease. Curr Top Microbiol Immunol 318:213-239.
59. Jaffe, J. D., H. Keshishian, B. Chang, T. A. Addona, M. A. Gillette, y S. A. Carr. 2008. Accurate inclusion mass screening: a bridge from unbiased discovery to targeted assay development for biomarker verification. Mol Cell Proteomics 7:1952-1962.
60. Keshishian, H., T. Addona, M. Burgess, E. Kuhn, y S. A. Carr. 2007. Quantitative, multiplexed assays for low abundance proteins in plasma by targeted mass spectrometry and stable isotope dilution. Mol Cell Proteomics 6:2212-2229.
Ejemplo 11: rHIaM12 de anticuerpos de IaM humana Promueve Formación de Axon e interactúa con lípidos a microtúbulos diana
El anticuerpo sHIgM12, promueve el crecimiento de neuritas en las neuronas granulares del cerebelo en cultivo primario. En el presente ejemplo, el uso de las neuronas del hipocampo primarias completamente humanas (rHIgM12 que tiene una cadena J humana) se encontró para promover la formación del axón. rHIgM12 une a las membranas de las neuronas e induce la agrupación de colesterol y gangliósido GM1.
Por otra parte, la rHIgM12 unida a la membrana se distribuye en dos grupos, uno asociado a dominios de balsa lipídica, y el otro con el sedimento insoluble detergente enriquecido en citoesqueleto. Agregados de rHIgM12 se colocalizan sobre microtúbulos, pero no sobre actina filamentosa después de la extracción del detergente. Estudios de coinmunoprecipitación demostraron que rHIgM12 y p3-tubulina existen en un complejo. Estos resultados indican que rHIgM12 específica la formación de axón por la agrupación de los dominios de membrana para señalar
el citoesqueleto de microtúbulos.
Las neuronas desarrollan axones mediante la regulación de crecimiento de neuritas (Barnes y Polleux, 2009). La citoesqueleto de neurona, que incluye actina filamentosa (F-actina) y los microtúbulos, juega un papel crucial en el crecimiento de neuritas y búsqueda de ruta cónica de crecimiento.
Anticuerpo derivado de suero puede no ser adecuado para los estudios de mayor escala, en particular si no se puede producir en cantidad y debe estar aislado para cada uso del paciente que produce el anticuerpo, la generación de una forma recombinante que demuestra actividad comparable y capacidad es ventajosa. El presente estudio demostró que el anticuerpo IgM12 recombinante y completamente humano (rHIgM12) promovió la formación de axón y, por tanto condujo polarización neuronal en las neuronas del hipocampo en cultivo. El rHIgM12 agrupa los dominios de la membrana neuronal que contienen colesterol y gangliósido GM1.
El fraccionamiento en gradiente de densidad de sacarosa indicó que rHIgM12 neuronal unido a la membrana se segrega en dos grupos, uno asociado con la fracción más ligera resistente a los detergentes que contiene caveolina-1, y el otro con la gránulo enriquecido por citoesqueleto. La extracción con detergente de las neuronas vivas demostró que rHIgM12 se asocia con microtúbulos. rHIgM12 también se co-inmunoprecipitó con p3-tubulina. En conjunto, se entiende que rHIgM12 se une dominios de membrana asociadas con los microtúbulos. Cuando está presente como un sustrato en una superficie, rHIgM12 promueve microtúbulos que se anclan en las membranas de las neuronas, lo que facilita el crecimiento de neuritas y la formación de axón.
IgM 12 recombinante humano (rHIgM12): rHIgM12 se expresó en células CHO (GibcoBRL, n° de cat 11619). Plásmidos que expresan las secuencias de codificación de cadena pesada y ligera para el anticuerpo predominante expresado en el suero de paciente de macroglobulinemia de Waldenstrom 12 se transfectaron junto con un transgén de cadena J humana en células CHO-S. Se seleccionaron las células CHO resultantes con dosis crecientes de metotrexato y un clon estable que produce el anticuerpo según lo medido por ELISA se subclonó y se amplió. El anticuerpo rHIgM1 2 del sobrenadante del cultivo se purificó por cromatografía a 97%, medido por análisis de HPLC.
Cultivo de células y el ensayo de crecimiento de neuritas. Las neuronas del hipocampo primarias se prepararon a partir de ratones FVB. Neuronas del hipocampo embriónicas del día 15 se disociaron en tripsina-EDTA, se sembraron en poli-D-lisina (PDL), PDL con laminina o sustratos rHIgM12 revestidos sobre una capa fina de membrana de nitrocelulosa unida a cubreobjetos de vidrio y se cultivaron en Medios Neurobasales que contienen 2 % (v/v) de B27. El crecimiento de neuritas se ensayó 12 horas después de sembrarse las neuronas. Las neuronas se fijaron con paraformaldehído al 4% y se tiñeron con anticuerpo anti p3-tubulina. Actina filamentosa (F-actina) se marcó con faloidina de rojo Texas, y los núcleos con DAPI. La longitud de neuritas se midió utilizando software neurona J, procesada con Excel (Microsoft) y se analizó estadísticamente con Prism (GraphPad). Una neurona de etapa 3 se definió como una neurona con múltiples neuritas en la que la neurita más larga (Dotti et al., 1988), determinada como un axón por tinción TAU1, fue al menos dos veces la longitud de la segunda neurita más larga. TAU1 se enriqueció asimétricamente en la parte distal del axón. Por el contrario, neuronas de etapa 2 tenían múltiples neuritas simétricas.
Inmunotinción, inmunoprecipitación y fraccionamiento en gradiente de densidad de sacarosa de neuronas de cultivos primarios: Neuronas del hipocampo en cultivo de uno a tres días in vitro (DIV), se permeabilizaron con 0,2% Triton X-100 después de la fijación con paraformaldehído al 4% seguido de inmunotinción. Las imágenes se recogieron usando un microscopio Olympus en posición vertical y se procesaron usando Photoshop (Adobe). La distribución rHIgM12 se determinó mediante ultracentrifugación en gradiente de sacarosa no continua. Brevemente, se permitió que rHIgM12 se uniera a neuronas corticales vivas de DIV7 a 4°C durante 30 min y después se lisaron en tampón de lisis enfriado con hielo (50 mM Tris.HCl, pH 7,4, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100 y inhibidor de cóctel de proteasa) durante 30 min. Los lisados neuronales se mezclaron con un volumen igual de 100% (p/v) de sacarosa. La mezcla se transfirió a un tubo de centrífuga, y se recubrió secuencialmente 8 ml de 35% de sacarosa y 3,5 ml de 5% de sacarosa. Después de centrifugación a 2x105 g durante 20 h a 4°C, se recogieron seis fracciones (2 ml de cada uno) desde la parte superior del gradiente. Cada fracción y el sedimento se disolvieron en tampón de muestra de SDS y se sometieron a transferencia de Western.
Para co-inmunoprecipitación de rHIgM12 con proteína de citoesqueleto, neuronas corticales vivas de DIV7 fueron tratadas con rHIgM12 a 4°C durante 30 min y se lisaron en tampón de lisis que contiene 0,5% de NP-40. rHIgM12 fue capturado por perlas de proteína L-agarosa, y p3-tubulina por la proteína G-resina (Thermo). Por doble etiquetado, neuronas in vivo se tiñeron con rHIgM12 en hielo y se permeabilizaron con 0,2% Triton X-100 después de la fijación. La extracción neuronal viva y fijación se llevaron a cabo en tampón que contenía 60 mM Pipes, 25 mM Hepes, 5 mM EGTA, 1 mM MgCl2 , 4% de paraformaldehído y 0,1% de Triton X-100.
Anticuerpos y otros reactivos en este ejemplo: Anti-p3-tubulina (Promega); anti-actina, EDTA, poli-D-lisina, metilo-p-ciclodextrina y filipina (Sigma); anti-TAU1, anti-caveolina-1 y anti-receptor de transferrina (Millipore); Texas Red-faloidina, toxina del cólera B, medio neurobasal y B27 (Invitrogen); comprimido de inhibidor de proteasa
(Roche).
IgM humano recombinante, rHIgM12, promovió formación de axon:
Para comprender mejor el efecto de la IgM en la diferenciación de neuronas y los mecanismos moleculares subyacentes, los ensayos de desarrollo de neuritas se llevaron a cabo con las neuronas del hipocampo en cultivo utilizando un rHIgM12 recombinante. Neuronas de hipocampo se someten a tres etapas de diferenciación para formar un axón (Dotti et al., 1988). Múltiples procesos se inician desde el cuerpo celular. Una de las neuritas de crecimiento rápido se diferencia en el axón, los otros en dendritas posteriores, que puede identificarse por la distribución diferencial de TAU1 y MAP2 respectivamente (Lewis et al, 1989; (Kanai y Hirokawa, 1995). Se encontró que rHIgM12, cuando se presenta como un sustrato, promovió sustancialmente diferenciación de las neuronas. Dentro de las 1 2 horas después de la siembra, las neuronas del hipocampo que crecen en rHIgM1 2 desarrollaron múltiples neuritas, y uno de ellos era mucho más largo en comparación con las neuritas vecinas. En contraste, las neuronas en placas en poli-D-lisina (PDL) tenían múltiples neuritas simétricas (18A FIGURA, B).
Después de medir la longitud de las neuritas, se encontró que las neuronas sembradas en placas sobre rHIgM12 (n = 8 6 ) en comparación con PDL (n = 74) tenían una longitud significativamente más larga total de las neuritas (195,8 pM vs. 150,7 pM, p = 0,0056) (FIGURA 18C). La longitud neurita más larga en más del doble (117,1 pM vs. 51,8 pM, p <0,0001) (FIGURA 18D), pero ninguna diferencia significativa en la longitud de la segunda longitud de neuritas más larga (38,7 pM vs. 33,3 pM, p = 0,0782) se encontró (FIGURA 18E). Las neuronas que crecen en rHIgM12 llevaban menos neuritas primarias (2,9 vs. 4,1, p <0,0001) (FIGURA 18F), y la mayoría era de la etapa 3 de fenotipo neuronal (72% en rHIgM12 vs. 1 8 % en PDL) (FIGURA 18G). Estos resultados indicaron que rHIgM12 promovió la diferenciación de las neuronas cuando se presenta como un sustrato.
El rasgo característico de diferenciación de las neuronas es crecimiento de axón polarizado y segregación de dendritas. Hemos observado que rHIgM12 promovió la extensión de neuritas, con una neurita significativamente mayor. Para verificar además que estas neuritas más largas desde la etapa 3 neuronas se convirtieron en axones, las neuronas del hipocampo sembradas en diferentes sustratos se tiñeron con anti-tau1 y anticuerpos MAP2 (FIGURA 19). Tinción de Tau1 era débil en las neuronas cultivadas en PDL (FIGURA 19A, D), pero mucho más fuerte en las neuronas sembradas en PDL-laminina (FIGURA 19B, E) o rHIgM12 (FIGURA 19C, F). La laminina es un sustrato clásico para la diferenciación de neuronas y la formación de axón (Chen et al., 2009) y fue utilizada como control positivo.
La comparación de la intensidad tau1 de las neuronas cultivadas en PDL (FIGURA 19D), la distribución de tau1 se enriqueció asimétricamente en la parte distal de la neurita más larga y mucho mayor en las neuronas se sembraron en cualquier PDL-laminina (FiGu RA 19E) o rHIgM12 (FIGURA 19F). En contraste, MAP2 tiñó la parte proximal de todas las neuritas (Figura 19A2- C2). Este resultado indicó que las neuritas más largas desde la etapa 3 neuronas cultivadas en rHIgM12 y laminina eran axones. Estos estudios demuestran que rHIgM12 soporta el crecimiento de neuritas a partir de múltiples tipos de neuronas, y que rHIgM12 impulsa la formación de axón.
rHIgM12 indujo agrupamiento de microdominio de membrana neuronal
Las neuronas del hipocampo que crecen en rHIgM12 como sustrato mostraron diferenciación mejorada (Figura 18 y 19). Sin embargo, no observamos que rHIgM12 indujo el crecimiento de neuritas cuando rHIgM12 se aplicó a los medios de baño (datos no mostrados). Esta observación sugirió que rHIgM12, para ejercer su función, se requiere para presentarse como una molécula de matriz extracelular. Para comprender mejor las interacciones de membrana rHIgM12 de neuronas, las neuronas vivas fueron tratadas primero con rHIgM12, y a continuación, fijadas y doble teñidas con filipina o toxina B del cólera (CTB), que se unen colesterol o gangliósido, GM1, respectivamente (Shogomori y Futerman, 2001). rHIgM12, CTB y filipina superficies celulares etiquetadas uniformemente de las neuronas de control no tratadas (Figura 20A). En contraste, el tratamiento con rHIgM12 a 37°C indujo reorganización de las membranas neuronales 30 minutos (Figura 20B, C). En primer lugar, rHIgM12 agregado en estructuras de tipo "parche" (FIGURA 20B1, C1) en la membrana, que no se observaron en neuronas no tratadas (FIGURA 20A) o de las neuronas tratadas con un anticuerpo IgM de control que no se une la membrana de la neurona (datos no mostrados). En segundo lugar, tanto agrupaciones de colesterol (FIGURA 20B2) como GM1 (FIGURA 20C2) se formaron en las membranas neuronales en el cuerpo de la célula, el eje de las neuritas y el cono de crecimiento. En tercer lugar, el agregado rHIgM12 colocalizado con el colesterol en clúster (FIGURA 20b3) o GM1 (FIGURA 20C3), especialmente en el dominio central del cono de crecimiento (FIGURA 20B-C, de gran aumento), pero no en la periferia del cono de crecimiento. Estos resultados indicaron que la rHIgM12 de baño aplicado indujo reordenamiento de la membrana neuronal, a pesar de que no soporta la extensión de neuritas/axón. Es posible que rHIgM12, cuando se presenta como un sustrato, puede reclutar moléculas de la membrana a los sitios de contacto de membrana rHIgM12 neuronales y resultar en la señalización direccional.
Balsas lipídicas unidas a rHIgM12
Los estudios bioquímicos sobre la base de la extracción con detergente no iónico en condiciones de frío se
han utilizado para demostrar la existencia de balsas de lípidos que contienen colesterol y glicoesfingolípidos (Chichili y Rodgers, 2009). Sin embargo, no se limita la información morfológica sobre las balsas de lípidos debido al tamaño de nanoescala que supera la resolución del microscopio óptico (Lingwood y Simons, 2010). La observación de que rHIgM12 induce la agrupación de membrana y colocaliza con colesterol y GM1 que contiene dominios de membrana (Figura 2 0 ) planteó la posibilidad de que rHIgM1 2 se asocia con dominios de balsa lipídica.
Puesto que las membranas neuronales están enriquecidas en colesterol y glicoesfingolípidos, que tienen temperaturas de fusión más alta (Tm) (Samsonov et al., 2001), la disminución de la temperatura de la membrana por debajo de Tm puede facilitar la visualización de balsa de lípidos y sus moléculas asociadas que son más dinámicas a 37°C. Para probar esta hipótesis, las neuronas vivas se enfriaron a 4°C, seguido de tinción rHIgM12. En contraste con las neuronas fijas a 37°C donde rHIgM12 se unió de manera uniforme (FIGURA 21A), a 4°C rHIgM12 etiquetó estructuras punteadas mucho más grandes. Además, los conos de crecimiento neuronal se encogieron (FIGURA 2 1 B).
Se aprovechó el hecho de que las balsas lipídicas son dominios de membrana dependientes de colesterol y que el agotamiento de colesterol altera su estructura (Chichili y Rodgers, 2009). Las neuronas del hipocampo cultivadas se trataron primero con metilo-p-ciclodextrina (M p CD) para agotar colesterol (Ko et al., 2005) y después se enfrió a 4°C, seguido por tinción rHIgM12 y CTB después de la fijación. rHIgM12 se unió a estructuras punctadas en las membranas neuronales después de la depleción de colesterol, y se colocalizaron con GM1 marcado con CTB (FIGURA 21C). Debido a que GM1 reside dentro de microdominios de balsa de lípidos y rHIgM12 induce agrupamiento de GM1 (FIGURA 20), concluimos que rHIgM12 une microdominios que contienen GM1, que son independientes de colesterol de la membrana neuronal. Estos experimentos confirmaron que las balsas lipídicas existen como estructuras morfológicas distintas, que se insertan de manera uniforme en las membranas (FIGURA 20A y FIGURA 21A) o se agregan para formar dominios más grandes (FIGURA 20B y C, FIGURA 21B y C). Estos resultados indican que rHIgM12 se une a la molécula que se mueven entre dominios de balsa y no balsa en función de las interacciones biofísicas con otras moléculas.
Además, ensayamos la hipótesis de que rHIgM12 se une a las balsas de lípidos. Debido a las balsas de lípidos se localizan dentro de las membranas no iónicas detergentes resistentes (DRM) a temperatura fría (Samsonov et al., 2 0 0 1 ), la localización de rHIgM1 2 se analizó en los lisados de las neuronas corticales preparadas a 4°C. Transferencias de los lisados neuronales con anticuerpo IgM secundario anti-humano reveló que se encontraron unidas a la membrana rHIgM12 tanto en el sedimento como el sobrenadante, mientras que el anticuerpo IgM de control no neuronal de unión se encuentra en sólo los eluatos de lavado (FIGURA 22A). Estas observaciones indicaron que rHIgM12 se segregaba en dos reservas, una localizada en la fracción de "detergente soluble" en el sobrenadante y la otra asociada con el sedimento de "insoluble en detergente".
En un segundo experimento, las neuronas corticales se marcaron primero con rHIgM12, a continuación, se fraccionó por gradiente de densidad de sacarosa a 4°C. Transferencias de diferentes fracciones indicó que rHIgM12 localizada en la fracción más ligera que también contenía la caveolina-1 , un marcador de balsa de lípidos, pero no a la fracción no balsa enriquecida en receptor de transferrina (FIGURA 22B). Sin embargo, incluso en este estricto proceso de fraccionamiento (1% Triton X-100 y ultracentrifugación a 2x105 g), algunos rHIgM12 se detectó en el sedimento, que contiene principalmente citoesqueleto insoluble en detergente (FIGURA 22B). Aunque tanto la tubulina (Sorice et al., 2009) como la actina (Levitan y Gooch, 2007) son pre-enviadas en balsas lipídicas, solamente se detectó p3-tubulina en la balsa y la mayoría de actina fue a la fracción no balsa (FIGURA 22B). Estos datos indicaron la existencia de dos grupos de membrana unidos rHIgM12, uno presente en la balsa de lípidos, y el otro en el sedimento.
rHIgM12 asociados con los microtúbulos
Nuestros datos muestran que rHIgM12 agregada se localiza en las regiones tales como el cuerpo de la célula, el eje de las neuritas y del cono de crecimiento de dominio central (FIGURA 20), que están dominados por microtúbulos (Forscher y Smith, 1988). Juntos, con el hecho de que rHIgM12 unida a la membrana se segregó en la balsa de lípidos y las fracciones de gránulos, las cuales contenían p3-tubulina (FIGURA 22B), estos resultados sugirieron que rHIgM12 puede estar asociado con el componente citoesqueleto. Para probar esta hipótesis, las neuronas del hipocampo se trataron con rHIgM12 para inducir la transposición de la membrana, fijándose a continuación de forma simultánea y extrayéndose a 37°C con 4% de paraformaldehído que contiene 0,1% de Triton X-100. Después de la extracción, puntos de membrana insolubles de detergente marcados con rHIgM12 se alinearon con fascículos de microtúbulos en manojos en el eje de las neuritas (FIGURA 23A2). En el cono de crecimiento, rHIgM12 localiza principalmente el dominio central ocupado por microtúbulos desfasciculan, pero no en la periferia del cono de crecimiento enriquecida con F-actina (FIGURA 23A3 y A4). Estos resultados indicaron que rHIgM12 puede asociarse con los microtúbulos
Se encontró que rHIgM12 se colocalizó con GM1 (Figura 21C). GM1 ha demostrado ser anclado por tubulina de membrana en las neuronas granulares del cerebelo, y se tira hacia abajo por un anticuerpo de antitubulina después de la reticulación (Palestini et al., 2000). Por lo tanto, es probable que rHIgM12 se asocia con
microdominios de membrana que contienen tubulina. A fin de ensayar esta hipótesis, los experimentos modificados se llevaron a cabo con los lisados celulares de las neuronas corticales vivas tratadas con rHIgM12 a 4°C. Se encontró que tanto rHIgM12 como p3-tubulina se coinmunoprecipitaron entre sí (FIGURA 23B), lo que sugiere que rHIgM12 y p3-tubulina existe como un complejo. Molécula de IgM tiene una estructura de pentámero con un peso molecular de aproximadamente 900 kDa. Para descartar la posibilidad de que rHIgM12 se asocie con tubulina o microtúbulos directamente en lisados neuronales, ensayos desplegables se realizaron con células de neuroblastoma N2A de que no se unen rHIgM12, pero no expresan p3-tubulina (FIGURA 25A). rHIgM12 ni se asoció con p3-tubulina ni se presentó en el sedimento. Sólo se detectó en los sobrenadantes de los lisados de células N2A de neuroblastoma (FIGURA 25B). Juntos, estos resultados confirmaron que las balsas de lípidos median la asociación entre rHIgM12, su antígeno y los microtúbulos.
Los datos en el presente documento demostraron que una IgM recombinante totalmente humana, rHIgM12, condujo polarización neurona mediante la promoción de axón en neuronas del hipocampo de cultivo primario cuando se presenta como un sustrato (FIGURA 18 y 19). El rHIgM12 se agregó sobre las superficies de las neuronas y se indujo la agrupación de colesterol y gangliósido, GM1 (FIGURA 20). rHIgM12 se colocalizó con GM1 (FIGURA 21). Fraccionamiento en gradiente de sacarosa de los lisados de las neuronas indica que rHIgM12 unido a la membrana se segregó en dos reservas: una asociada con balsas de lípidos, la otra con el sedimento (FIGURA 22). Mediante la extracción de neuronas vivas con detergente no iónico, se demostró, además, que rHIgM12 unido a la membrana se co-localizó con microtúbulos y fue coinmunoprecipitado con p3-tubulina (FIGURA 23).
Lo anterior indica que rHIgM12 se une a e interactúa con microdominios de balsa de lípidos, y además indica que dominios de balsa asociados a rHIgM12 transmiten señalización a los microtúbulos. Como resultado, rHIgM12 media estabilización de microtúbulos, que impulsa axón. Este ejemplo demostró que rHIgM12 promueve selectivamente axón de las neuronas del hipocampo primarias (FIGURA 18 y 19).
Recientemente, se demostró que los sueros de origen humano IgM, sHIgM12, mejoró la función neurológica de un modelo animal de la esclerosis múltiple que se desarrolla una amplia degeneración axonal y la pérdida (Rodríguez et al., 2009). Estos estudios apoyan el concepto de que HIgM12 ejerce su función fomentando el crecimiento axonal.
Las neuronas específican an axones a través de un programa intrínseco controlado que regula la extensión de neuritas. Una neurona en desarrollo inicia múltiples neuritas. Una de las neuritas se diferencia en el axón, las otras dendritas. Las neuritas vecinas compiten entre sí. El neurita más larga, que también tiene la tasa de extensión más rápida, rompe primero la extensión de neuritas simétricas para convertirse en un axón, mientras que las otras neuritas se extienden mucho más lento y se desarrollan en las dendritas más tarde (Dotti et al, 1988;. Goslin y Banker, 1989). F-actina y los microtúbulos son los dos principales citoesqueletos que están implicados en la extensión de neuritas y la formación de axón. Nuestra comprensión de la especificación de axón ha ido evolucionando. F-actina, que se localiza principalmente a la periferia del cono de crecimiento, se consideraba que desempeñaba un papel importante. Los microtúbulos, que dominan el eje de neuritas y están confinados al dominio central del cono de crecimiento, se pensaba que eran secundarias a F-actina. Recientemente, se encontró que los microtúbulos también juegan un papel fundamental en el crecimiento de los axones. Los microtúbulos son capaces de explorar dinámicamente la malla de crecimiento de cono actina. La estabilización de los microtúbulos es suficiente para inducir la formación de axón (Witte y Bradke, 2008).
Nuestros resultados, que rHIgM12 y p3-tubulina co-inmunoprecipitaron, que rHIgM12 se colocalizó con los microtúbulos y que rHIgM12 estaba presente en el sedimento después de la extracción con detergente, no sólo los microtúbulos de apoyo como un jugador central. Además, estos datos proporcionan la primera evidencia en la técnica que los microtúbulos interactúan directamente con las membranas neuronales. Estos hallazgos apoyan el concepto de que rHIgM12 interactúa con o se une a una cascada de transmembrana que transporta la señal extracelular a los microtúbulos. La propiedad dinámica de los microtúbulos para crecer y retraer, por ejemplo, en los conos de crecimiento, que les permite conducir el movimiento de la membrana de la neurona (Dent y Kalil, 2 0 0 1 ). Los microtúbulos estabilizados anclan las membranas neuronales. La existencia de los microtúbulos dinámicos y estabilizados y/o la transición entre estos dos estados realizan el proceso de crecimiento de neuritas para específicar un axón. En consecuencia, IgM12 mejora la extensión de axones efectuando estabilidad de los microtúbulos.
Los lípidos y las proteínas se incorporan continuamente en las membranas celulares que luego se dividen en microdominios, las denominadas balsas de lípidos. Balsas de membrana se definen generalmente como escala nano (1 0 - 2 0 0 nm), heterogéneas, y los compartimentos de membrana dinámicos enriquecidos con esteroles y esfingolípidos (Lingwood y Simons).
Proponemos que rHIgM12 se una a las balsas de lípidos. En primer lugar, se observó que rHIgM12 se agregó sobre las membranas de las neuronas y se indujo la agrupación de colesterol o gangliósido, GM1 (FIGURA 20). Estos resultados indican que rHIgM12 se une a dominios de balsa de lípidos que contienen colesterol y GM1. Pequeñas balsas pueden interactuar con lípidos y/o proteínas. Las pequeñas balsas individuales pueden estabilizar
y fundirse en plataformas más grandes que integran señales y regulan la intensidad y la amplitud de las vías de señalización. Los anticuerpos IgM, que poseen una estructura pentamérica, pueden amplificar señales por reticulación de antígenos adyacentes (receptores) y/o llevar a los antígenos lo suficientemente cerca para mejorar la reticulación o interacción. Por lo tanto, los antígenos reticulados y las moléculas de señalización asociadas pueden agruparse. En segundo lugar, hemos demostrado que rHIgM12 unido a puntos lagrimales mucho más grandes en las membranas de las neuronas cuando las neuronas del hipocampo se enfriaron a 4°C (FIGURA 21). El colesterol y esfingolípidos tienen temperatura de fusión alta (Tm). El tratamiento de las neuronas a la temperatura por debajo de Tm puede disminuir dinámicamente de la membrana, lo que facilita la visualización de las balsas de lípidos agregados. Junto con el hallazgo de que rHIgM12 se colocalizó con GM1 incluso después de agotamiento de colesterol (FIGURA 21), estos resultados sugieren que rHIgM12 se une a e interactúa con moléculas de la membrana de la neurona que se colocaliza con GM1. La identidad exacta no está clara, aunque nuestra anterior conclusión, que el tratamiento con sialidasa de neuronas granulares de rata cultivadas de cerebelo abolió la unión a sHIgM12, sugirió que el sHIgM12 unido a epítopo es un hidrato de carbono (Warrington et al., 2004). En tercer lugar, después del fraccionamiento de sacarosa de los lisados de las neuronas, el rHIgM12 unido a la membrana s se localizó en la fracción más ligera, que también contenía el lípido marcador de balsa, caveolina-1. Tomados en conjunto, nuestros resultados apoyan la hipótesis de que rHIgM12 se une a las balsas de lípidos.
Cascadas de señales, a partir de la hoja externa de las membranas de las neuronas al citoesqueleto intracelular, son fundamentales para la específicación de axón. Agregados de rHIgM12 se distribuyeron al eje de las neuritas y el dominio central del cono de crecimiento (FIGURA 20), que están dominados por los microtúbulos, pero no actina filamentosa (FIGURA 23). Algunos de los agregados rHIgM12 eran detergentes insolubles y localizados en haces de microtúbulos fasciculados después de la extracción con detergente (FIGURA 23A). Estas observaciones indican la existencia de otro grupo de moléculas unidas a rHIgM12, que puede ser diferente de la que asocia las balsas de lípidos. El estudio de fraccionamiento de sacarosa confirmó, además, que existe un conjunto de moléculas asociadas a rHIgM12 de localización en el sedimento insoluble en detergente (FIGURA 22B), que puede ser la reserva que se localiza con microtúbulos tal como se detecta en FIGURA 23A.
Estos resultados demostraron que parte del rHIgM12 se asocia con el componente del citoesqueleto, túbulos posiblemente microorganismos. El hallazgo de que rHIgM12, que asocia con "moléculas de detergente soluble" en el sobrenadante (FIGURA 22A y FIGURA 23B), se co-inmunoprecipitó con p3-tubulina (FIGURA 23B), indica que moléculas unidas a rHIgM12 y p3-tubulina puede existir como un complejo en las balsas de lípidos. Sin embargo, no es probable que rHIgM12 interactúe con p3-tubulina directamente porque rHIgM12 no tire hacia abajo p3-tubulina en células de neuroblastoma N2A que rHIgM12 no se une (FIGURA 25). Así, tanto el citoesqueleto como el rHIgM12 asociado a balsa de lípidos se unen a las moléculas que se localizan en las proximidades de la tubulina. Este concepto se refuerza aún más por las observaciones de que rHIgM12 se colocalizó con GM1 (FIGURA 21C), y que GM1 se tira hacia abajo por un anticuerpo anti-tubulina después de una reacción de reticulación (Palestini et al., 2 0 0 0 ).
Las moléculas unidas a rHIgM12 en los dominios de balsa de lípidos se pueden incorporar constantemente en los citoesqueletos durante el proceso de crecimiento de las neuritas y la extensión de axones. Esta hipótesis está apoyada por los hallazgos de que, en la región del cono de crecimiento, rHIgM12 se agregó en el dominio central (Fig. 3 y 6 A), y era de detergente insoluble a 37°C (FIGURA 23A). Tomados en conjunto, se describe que HIgM12 se une y afecta a la dinámica de las balsas de lípidos que median la señalización desde HIgM12 a los microtúbulos.
Si F-actina está implicada en la señalización inducida por rHIgM12 no está claro. Un número de líneas de evidencias indican que tanto la actina como proteínas de unión a actina se asocian con balsas lipídicas (Levitan y Gooch, 2007). Los filamentos de actina y los microtúbulos interactúan activamente en el movimiento celular coordinado (Rodríguez et al., 2003) que también incluyen las neuronas. Así, es probable que la actina también juegue un papel en la señalización inducida por rHIgM12. Junto con p3-tubulina, una pequeña cantidad de actina se tira hacia abajo por rHIgM12 (FIGURA 26B). Por lo tanto rHIgM1 2 se une balsas lipídicas que contienen tanto la actina y la 133-tubulina.
Sin embargo, puntos lagrimales marcados con rHIgM12 permaneció en el mismo borde del cono de crecimiento, mientras que las redes de F-actina se encogieron desde la periferia del cono de crecimiento cuando las neuronas se trataron a 4°C (FIGURA 26 A). Después del fraccionamiento en gradiente de sacarosa, actina se localiza principalmente en la fracción no balsa, y sólo se detectó una pequeña cantidad de actina en el sedimento insoluble con detergente (FIGURA 22B). Estas observaciones indican que la F-actina es muy dinámica y la mayoría se despolimeriza en condiciones de frío y/o por extracción con detergente. Por lo tanto, la actina puede estar implicada en la señalización mediada rHIgM12.
Se concluye que rHIgM12 de IgM humana se une a y reorganiza las balsas de lípidos, y que los microtúbulos son una de las dianas aguas abajo. rHIgM12 promueve la extensión de axones sólo cuando se presenta como sustrato. Como un sustrato, rHIgM12 se inmovilizó y se constriñió en su interacción con la membrana neuronal. El rHIgM12 inmovilizado puede haber creado un gradiente de señales a través de las membranas neuronales como se observa con frecuencia en la señalización inducida por morfógeno (Schmitt et al., 2006). En
contraste, la aplicación del baño de rHIgM12 solamente puede facilitar agrupación aleatoria de balsa de lípidos (FIGURA 20) y puede no ser suficiente para romper el crecimiento de neuritas simétricas y mejorar la extensión de axones. En resumen, nuestros resultados apoyan el modelo propuesto en membranas Figura 24. Neurona contiene tanto balsa como microdominios no balsa. Hay dos reservas de dominios de balsa con una de ellas asociada con los microtúbulos (FIGURA 24A). 2) rHIgM12 se une a y agrupa dominios de balsa, que promueve la estabilización de los microtúbulos y el anclaje de la membrana (FIGURA 24B). 3) En el cono de crecimiento, agrupación de balsa inducida por rHIgM12 puede mejorar la periferia del cono de crecimiento a la transición de dominio central, rompiendo el crecimiento de neuritas simétricas para específicar formación de axón (FIGURA 24C).
REFERENCIAS
Allen JA, Allen JA, Halverson-Tamboli RA, Rasenick MM (2007) Lipid raft microdomains and neurotransmitter signalling. Nat Rev Neurosci 8:128-140.
Barnes AP, Polleux F (2009) Establishment of axon-dendrite polarity in developing neurons. Annu Rev Neurosci 32:347-381.
Chen ZL, Haegeli V, Yu H, Strickland S (2009) Cortical deficiency of laminin gamma1 impairs the AKT/GSK-3beta signaling pathway and leads to defects in neurite outgrowth and neuronal migration. Dev Biol 327:158-168. Chichili g R, Rodgers W (2009) Cytoskeleton-membrane interactions in membrane raft structure. Cell Mol Life Sci 66:2319-2328.
Conde C, Caceres A (2009) Microtubule assembly, organization and dynamics in axons and dendrites. Nat Rev Neurosci 10:319-332.
de Anda FC, Pollarolo G, Da Silva JS, Camoletto PG, Feiguin F, Dotti CG (2005) Centrosome localization determines neuronal polarity. Nature 436:704-708.
Dent EW, Kalil K (2001) Axon branching requires interactions between dynamic microtubules and actin filaments. J Neurosci 21:9757-9769.
Dent EW, Callaway JL, Szebenyi G, Baas PW, Kalil K (1999) Reorganization and movement of microtubules in axonal growth cones and developing interstitial branches. J Neurosci 19:8894-8908.
Dotti CG, Sullivan CA, Banker GA (1988) The establishment of polarity by hippocampal neurons in culture. J Neurosci 8:1454-1468.
Forscher P, Smith SJ (1988) Actions of cytochalasins on the organization of actin filaments and microtubules in a neuronal growth cone. J Cell Biol 107:1505-1516.
Goslin K, Banker G (1989) Experimental observations on the development of polarity by hippocampal neurons in culture. J Cell Biol 108:1507-1516.
Herdegen T, Skene P, Bahr M (1997) The c-Jun transcription factor--bipotential mediator of neuronal death, survival and regeneration. Trends Neurosci 20:227-231.
Higginbotham HR, Gleeson JG (2007) The centrosome in neuronal development. Trends Neurosci 30:276-283.
Kanai Y, Hirokawa N (1995) Sorting mechanisms of tau and MAP2 in neurons: suppressed axonal transit of MAP2 and locally regulated microtubule binding. Neuron 14:421-432.
Ko M, Zou K, Minagawa H, Yu W, Gong JS, Yanagisawa K, Michikawa M (2005) Cholesterol-mediated neurite outgrowth is differently regulated between cortical and hippocampal neurons. J Biol Chem 280:42759-42765.
Levitan I, Gooch KJ (2007) Lipid rafts in membrane-cytoskeleton interactions and control of cellular biomechanics: actions of oxLDL. Antioxid Redox Signal 9:1519-1534.
Lewis SA, Ivanov IE, Lee GH, Cowan NJ (1989) Organization of microtubules in dendrites and axons is determined by a short hydrophobic zipper in microtubule-associated proteins MAP2 and tau. Nature 342:498-505. Lingwood D, Simons K Lipid rafts as a membrane-organizing principle. Science 327:46-50.
Niethammer P, Delling M, Sytnyk V, Dityatev A, Fukami K, Schachner M (2002) Cosignaling of NCAM via lipid rafts and the FGF receptor is required for neuritogenesis. J Cell Biol 157:521-532.
Palestini P, Pitto M, Tedeschi G, Ferraretto A, Parenti M, Brunner J, Masserini M (2000) Tubulin anchoring to glycolipid-enriched, detergent-resistant domains of the neuronal plasma membrane. J Biol Chem 275:9978-9985. Rodriguez M, Warrington AE, Pease LR (2009) Invited Article: Human natural autoantibodies in the treatment of neurologic disease. Neurology 72:1269-1276.
Rodriguez OC, Schaefer AW, Mandato CA, Forscher P, Bement WM, Waterman-Storer CM (2003) Conserved microtubule-actin interactions in cell movement and morphogenesis. Nat Cell Biol 5:599-609.
Samsonov AV, Mihalyov I, Cohen FS (2001) Characterization of cholesterol-sphingomyelin domains and their dynamics in bilayer membranes. Biophys J 81:1486-1500.
Schmid RS, Maness PF (2008) L1 and NCAM adhesion molecules as signaling coreceptors in neuronal migration and process outgrowth. Curr Opin Neurobiol 18:245-250.
Schmitt AM, Shi J, Wolf AM, Lu CC, King LA, Zou Y (2006) Wnt-Ryk signalling mediates medial-lateral retinotectal topographic mapping. Nature 439:31-37.
Segal RA (2003) Selectivity in neurotrophin signaling: theme and variations. Annu Rev Neurosci 26:299-330.
Shogomori H, Futerman AH (2001) Cholera toxin is found in detergent-insoluble rafts/domains at the cell surface of hippocampal neurons but is internalized via a raft-independent mechanism. J Biol Chem 276:9182-9188.
Sorice M, Matarrese P, Tinari A, Giammarioli AM, Garofalo T, Manganelli V, Ciarlo L, Gambardella L, Maccari G, Botta M, Misasi R, Malorni W (2009) Raft component GD3 associates with tubulin following CD95/Fas ligation. Faseb J 23:3298-3308.
Warrington AE, Bieber AJ, Van Keulen V, Ciric B, Pease LR, Rodríguez M (2004) Neuron-binding human monoclonal antibodies support central nervous system neurite extension. J Neuropathol Exp Neurol 63:461-473. Witte H, Bradke F (2008) The role of the cytoskeleton during neuronal polarization. Curr Opin Neurobiol 18:479-487. Witte H, Neukirchen D, Bradke F (2008) Microtubule stabilization specifies initial neuronal polarization. J Cell Biol 180:619-632.
EJEMPLO 12: Ensayos biosensores ópticos de nanoporos para MAbs humanos beneficiosos para la lesión medular
Protección de axon y reparación después de una lesión de la médula espinal (SCI) tiene un gran potencial como una estrategia eficaz para prevenir la pérdida de la neurona motora y discapacidad permanente. La protección de las neuronas se ha logrado usando factores tróficos específicos para la prevención de daños y promover la reparación de los axones después de una lesión. Estas moléculas se identificaron predominantemente usando estrategias de selección basadas en sistemas in vitro que se centraron en factores neurotróficos de molécula pequeña de orientación específica.
Los anticuerpos monoclonales autorreactivos naturales han demostrado funciones biológicas beneficiosas en las células del SNC usando múltiples modelos de lesión y la enfermedad. Promoción mediada por anticuerpos de la supervivencia neuronal, la regeneración axonal y la recuperación funcional se ha demostrado in vivo usando el IgM de ratón monoclonal, IN-1 (Bregman 1995; Caroni y Schwab 1988). Se obtuvieron resultados similares usando inmunización de la médula espinal homogeneizada (SCH) antes de la lesión del SNC (Ellezam 2003; Huang 1999).
En base a los datos de los anticuerpos humanos de unión a neurona incluyendo IgM12 y el anticuerpo remielinizante IgM22, se describe que las interacciones de anticuerpo monoclonal humano que protegen los axones de daños mediante la promoción de la supervivencia neuronal y función, y beneficioso para el tratamiento de lesiones y/o enfermedad del SNC tal como SCI, dependen de interacciones de bicapa de proteína-lípido de membrana plasmática superficial en condiciones patológicas/fisiológicas.
Los anticuerpos humanos que promueven la supervivencia de las neuronas después de la lesión del nervio o el daño (por ejemplo, en SCI) o que protegen a las neuronas de lesiones o muerte, en cada caso que resulta en la preservación de la función neurológica, son evaluados, caracterizados y/o identificados utilizando tecnologías que mantienen bicapas de proteína-lípido de la membrana plasmática superficial patológica/fisiológica.
En este estudio, determinamos la cinética de interacción de unión a la superficie de los anticuerpos monoclonales humanos usando sensores de resonancia de plasma superficial de bicapa proteína-lípido (SPR). Los anticuerpos monoclonales humanos se caracterizan o identifican usando la unión en las lesiones de la médula espinal de secciones ex vivo de tejido de roedores después de una lesión de la médula espinal. A continuación, una médula espinal de lesión por aplastamiento de roedor se utilizó para evaluar aún más la capacidad de los anticuerpos humanos para promover la supervivencia de neuronas, extensión de neuritas o la remielinización in vivo resulta en la patología reducida y mejora la función neurológica.
Un sensor de bicapa lipídica SPR novelo proporciona una tecnología rápida, libre de marcador que permite la caracterización de cinética de unión proteína-lípido de la superficie y la afinidad de anticuerpos IgM humanos beneficiosos. Esta tecnología de sensor de resonancia de plasmón superficial (SPR) se desarrolló basándose en matrices de nanoporos periódicas en películas metálicas combinadas con bicapa lipídica plana fisiológica. La tecnología de SPR ha permitido la caracterización rápida, libre de etiqueta de cinética de unión de anticuerpos IgM y la afinidad con antígenos relevantes.
Cinética de unión importante para cuantificar pequeñas diferencias proporcionan un fundamento para la selección de moléculas de plomo durante el desarrollo y el impacto tanto de la dosificación como la potencia de una molécula terapéutica in vivo. SPR se ha convertido en un estándar aceptado en entornos industriales y académicos, en el que se caracteriza típicamente la interacción molecular entre un par de parejas de unión solubles. Sin embargo, la tecnología sólo está comenzando a ser adaptada a las necesidades de los antígenos unidos a la membrana. El sustrato de oro utilizado en la detección SPR no es susceptible a la formación de membrana lipídica compatible. Además, las proteínas de membrana en contacto directo con la superficie de oro a menudo pierden su funcionalidad. Sin embargo, el desarrollo de nuevos nanoporos de detección de la tecnología de la arquitectura que utiliza una película de oro delgada perforada con ensayos periódicas de nanoporos ha superado estos retos (véase, por ejemplo, Figura 27). Cada nanoporo se sienta en un sustrato de vidrio y forma un pocillo pequeño para confinar las membranas lipídicas soportadas, mientras que la película de oro que rodea proporciona efectos SPR para controlar dinámicamente la unión de moléculas en la membrana (FIGURA 27B). Ensayos de nanoporo molidos en una película de oro proporcionan una geometría única, ya que una bicapa lipídica delgada puede ser suspendida en los nanoporos, manteniendo la estabilidad mecánica y estando rodeada de un tampón en ambos lados (FIGURA 27B).
Las proteínas de membrana de este modo pueden ser integradas a la perfección con la detección de la
capacidad de SPR de ensayos de nanoporos de oro, manteniendo su funcionalidad en un entorno que imita más estrechamente su estado natural. Además, las proteínas de membrana integradas en la bicapa lipídica independiente se puede acceder fácilmente desde ambos lados, haciendo este enfoque más atractivo para estudiar el mecanismo por el que la unión antígeno-anticuerpo en los resultados de la superficie celular en la señalización celular (FIGURA 27B). Este sensor de bicapa lipídica SPR proporciona una tecnología rápida, libre de etiqueta que se utiliza para la evaluación, identificación y caracterización de cinética de unión proteína-lípido de la superficie y afinidad de anticuerpos IgM humanos terapéuticos.
Esta plataforma de sensor caracteriza cinética de unión proteína-lípido de la superficie y afinidad de los anticuerpos IgM terapéuticos para pruebas en un modelo animal de lesión de la médula espinal. Hemos caracterizado ampliamente ciertos anticuerpos IgM identificados mediante la unión a la superficie "in vivo" no fijada de tejido cerebelo y células en el SNC. Estos anticuerpos IgM (ejemplificados por IgM22 y IgM46) estimulan el flujo de calcio in vitro y promueven la remielinización in vivo. En base a los mismos criterios, los anticuerpos humanos se han identificado mediante el cribado de muestras de banco de suero de disproteína en la Mayo Clinic. Anticuerpos IgM humanos adicionales (ejemplificados en este documento por IgM12 y IgM42) se unen a la superficie de neuronas y se promueve el crecimiento de neuritas y se impidió la muerte de neuronas in vitro.
Los anticuerpos de unión a neuronas exhibieron inmunorreactividad específica a células específicas del sistema nervioso central, las estructuras, la lesión y la patología de enfermedad usando secciones corticales “in vivo” no fijadas de los múltiples modelos de ratón de lesión del SNC y la enfermedad.
Los anticuerpos monoclonales humanos naturales se han demostrado potenciar funciones biológicas beneficiosas en células SNC de varios modelos de lesión y enfermedad. Los anticuerpos monoclonales humanos se unen a la superficie de una amplia variedad de neuronas en cultivo, incluyendo el cerebelo, cortical, ganglionares de la retina y las neuronas de la médula espinal (por ejemplo, anticuerpos humanos IgM12 y IgM42). Estos anticuerpos inducen la extensión de neuritas a partir de neuronas del SNC (FIGURA 28) y anular la inhibición de la mielina del SNC sobre el crecimiento de neuritas (Warrington et al. 2004).
Los estudios in vitro demuestran la unión de anticuerpos monoclonales humanos a membrana plasmática de las neuronas de la superficie. La característica de unión es uniforme a 0°C mientras que los reordenamientos de superficie forman estructuras punteadas a 15°C (FIGURA 29). La superficie de unión celular es característica de la agrupación de microdominios de membrana asociada con moléculas que inician cascadas de señales de señalización (Howe et al. 2004).
Los anticuerpos monoclonales humanos entran en el tejido de la médula espinal y se acumulan en los sitios de daño 4 horas después de la administración intravenosa (FIGURA 30). La inmunocitoquímica realizada en secciones de vibratoma de médula espinal detecta IgM humana dentro de las lesiones después de la administración de anticuerpos de unión a las neuronas, pero no después de la administración de un control de IgM humano. Los ratones con la enfermedad de la médula espinal crónica (ratones infectados con TMEV) se les dio 0,5 mg de sHIgM42 intraperitoneal, las médulas espinales removidas 4 h más tarde, y la presencia de IgM humana se detectó en las secciones transversales de la médula espinal (FIGURA 30). Áreas lesionadas de la médula espinal de un ratón al que se administró sHIgM42 demuestran fibras paralelas que se unen a una IgM anti-humana fluorescentemente etiquetada (FIGURA 30A). Áreas lesionadas de la médula espinal de un ratón dado un control de IgM humana no contienen IgM humana (FIGURA 30B).
Los anticuerpos monoclonales humanos se han identificado usando ensayos de la función biológica a las muestras de visualización desde el banco de suero de disproteína de Mayo Clinic. Las muestras de suero, 115000 recogidos durante más de 40 años, contienen altas concentraciones de anticuerpo monoclonal a partir de pacientes con gammapatía monoclonal. Hemos sintetizado y probado IgM 22 humana recombinante, que tiene actividad promotora de la remielinización en modelos animales (Mitsunaga 2002; Warrington 2000).
En una realización, el presente Ejemplo genera anticuerpo monoclonal de calidad GMP útil para aplicaciones terapéuticas, pronósticas, diagnósticas y/o profilácticas en condiciones de degeneración de los nervios, la lesión del nervio y/o daños, tales como en SCI humana o en condiciones desmielinizantes que exponen las neuronas del SNC.
Estos estudios caracterizan más y evalúan anticuerpos IgM humanos que promueven la supervivencia de neuronas tras SCI traumática resultante en la preservación de la función neurológica. Pruebas de comportamiento y mediciones morfológicas de los números de los axones después de SCI se evalúan y se determinan las diferencias entre preservación endógena y mediada por anticuerpos de la función neurológica. Los tratamientos con los anticuerpos que se extienden axón-axón-protector y se compararon con controles de anticuerpos después de SCI se utilizan para caracterizar las diferencias de conservación de la función neurológica para actividades específicas mediadas por anticuerpos.
Vesículas de membrana que contiene la bicapa de proteína-lípido de la membrana plasmática se aíslan de
células en condiciones fisiológicas. Nanoporos de los sensores de SPR se recubren utilizando preparaciones de microvesículas de neuronas aisladas, glia, de sinaptosomas y las membranas de mielina de SNC normal y los tejidos de la médula espinal lesionada. Muestras de suero de anticuerpos monoclonales humanos y muestras de anticuerpos recombinantes se rastrearon para determinar la cinética de unión para tipos específicos de membrana.
Los anticuerpos monoclonales humanos que se unen específicamente a la membrana plasmática superficial de las células no congeladas no fijadas “in vivo” y tejidos en condiciones fisiológicas como se confirma por medio de sensores de SPR se prueban para la unión usando tejidos no congelados, no fijados, patológicos “vivos” tras SCI específico. Los anticuerpos monoclonales humanos que demuestran la unión a lesiones de la médula espinal son caracterizados adicionalmente para la aplicabilidad y la actividad en el tratamiento tras SCI en el roedor.
Protocolo de Diseño. La lesión de compresión se genera en el nivel de T9 en ratones de diez semanas de edad (18-22 g) C57BL/6J (Jackson Laboratories) utilizando un clip de aneurisma modificado (clip de ratón Fejota, 3G o fuerza de cierre 8 g). Este modelo de SCI incluye no sólo un impacto inicial, sino también una fase de compresión persistente que promueve la cavitación microquística, degeneración axonal, y astrogliosis robusta, todas las características de la SCI traumática humana (Joshi y FEHLINGS 2002). Pruebas de comportamiento utilizando la escala de calificación locomotora Basso, Beattie, Bresnahan (BBB) para ratas y la Escala de Ratón Basso (BMS) para la locomoción se determinan después de SCI para evaluar los déficits neurológicos. La técnica se describe en la Figura 31.
Secciones de tejido in vivo no congeladas, no fijadas ex vivo se preparan a continuación de lesiones patológicas después de una lesión por compresión de la médula espinal en roedores. La inmunorreactividad de unión de anticuerpos IgM humanos se tamiza usando tinción de inmunofluorescencia (IF) de las secciones mantenidas bajo condiciones fisiológicas.
Los ratones se tratan de forma aguda después de una lesión por compresión de la médula espinal con anticuerpos humanos, incluyendo rHIgM12 y sHIgM42 o rHIgM42, administrados por vía intraperitoneal. La capacidad de los anticuerpos humanos para promover la conservación del axón o la reparación del tejido se mide. Pruebas de comportamiento utilizando la acreditación y la escala BMS se realizan a intervalos regulares durante el período de 5 semanas después de SCI. Una dosis única de 0,5 mg de anticuerpo humano se administra ip a los ratones 30 minutos después de la SCI. Grupos de ratones (15 ratones) reciben anticuerpos humanos que promueven la protección de neuronas, el crecimiento de neuritas, el control de los anticuerpos humanos que no se unen a las células o control. Ratones individuales se alojan durante la noche una vez por semana en cajas de actividad infrarroja automatizada (Mikami et al. 2002) y encabritarse espontáneamente, una medida de la discapacidad de la extremidad posterior, y la actividad horizontal grabada (Accuscan Inc, Ohio). Puntajes tradicionales BBB también se recogen semanalmente. Las evaluaciones funcionales se realizan cegados.
Fluoro-oro y azul rápido, 4 semanas después del tratamiento, se inyecta en las médulas espinales de cuatro ratones en el nivel torácico inferior. Una semana después, los ratones se sometieron a perfusión y los cerebros y las médulas espinales se removieron. Cuerpos celulares marcados en retrógrado en núcleos reticular, vestibular y rojos del tallo cerebral se cuentan entre los grupos por microscopía fluorescente (Ure y Rodríguez 2002). El número de cuerpos celulares marcados en la (sección de vibratoma) tronco cerebral refleja el nivel de los axones funcionales a lo largo de la médula espinal. La inmunocitoquímica para acumulación de proteína de p amiloide se realiza en secciones transversales de tejido tomadas cada 1 mm a lo largo de la médula espinal. El número de agregados de amiloides p en una sección refleja alteración de transporte axonal y la disfunción de axón en ese nivel.
Cinco semanas después de lesión, los ratones restantes se sometieron a perfusión, con formaldehído/glutaraldehído y se eliminó la médula espinal. Cada otro bloque de 1 mm que comprende el sitio de la lesión de la médula espinal está incrustada en plástico de araldito y se seccionó ( 1 micra) y se tiñó con parafenileno para visualizar los axones conservados rodeados por mielina. El área de sección transversal de la lesión, el número de axón conservado y la infiltración de células inmunes se mide utilizando software Microsuite (Olympus). Frecuencia Axon se mide y se compara entre los grupos (McGavern et al 2000;. Rodríguez et al., 1987). Todos los análisis se realizan sobre muestras codificadas y cegadas sin conocimiento del grupo experimental. Axones conservados se expresan como el número de axones conservados/mm2 de la lesión. Comparación por pares de datos entre los grupos experimentales y de control utiliza la prueba de Mann-Whitney Sum.
Referencias
Rodriguez M, Warrington AE, Pease LR. Human Natural Autoantibodies in the Treatment of Neurologic Disease. Neurology 2009; 72: 1269-1276.
Im H, Lesuffleur A, Lindquist NC, Oh SH. Plasmonic nanoholes in a multichannel microarray format for parallel kinetic assays and differential sensing. Analytical Chemistry. 2009;81:2854-2859
Maynard jA, Lindquist NC, Sutherland j N, Lesuffleur A, Warrington AE, Rodriguez M, and Oh S-H. Next generation SPR technology of membrane-bound proteins for ligand screening and biomarker discovery. J Biotech 2009; 4(11):1542-1558
Nagpal P, Lindquist NC, Oh SH, Norris DJ. Ultrasmooth patterned metals for plasmonics and metamaterials. Science. 2009;325:594-597.
Bieber AJ, Warrington A, Asakura K, Ciric B, Kaveri SV, Pease LR, Rodriguez M. 2002. Human antibodies accelerate the rate of remyelination following lysolecithin-induced demyelination in mice. Glia 37(3):241-249. Bregman BS, Kunkel-Bagden E, Schnell L, Dai HN, Gao D, Schwab Me . 1995. Recovery from spinal cord injury mediated by antibodies to neurite growth inhibitors. Nature 378(6556):498-501.
Caroni P, Schwab ME. 1988. Antibody against myelin-associated inhibitor of neurite growth neutralizes nonpermissive substrate properties of CNS white matter. Neuron 1(1):85-96.
Corse AM, Bilak MM, Bilak SR, Lehar M, Rothstein JD, Kuncl rW. 1999. Preclinical testing of neuroprotective neurotrophic factors in a model of chronic motor neuron degeneration. Neurobiol Dis 6(5):335-346.
Ellezam B, Bertrand J, Dergham P, McKerracher L. 2003. Vaccination stimulates retinal ganglion cell regeneration in the adult optic nerve. Neurobiol Dis 12(1):1-10.
Estevez AG, Spear N, Manuel SM, Radi R, Henderson CE, Barbeito L, Beckman JS. 1998. Nitric oxide and superoxide contribute to motor neuron apoptosis induced by trophic factor deprivation. J Neurosci 18(3):923-931. Festoff BW, Ameenuddin S, Arnold p M, Wong A, Santacruz KS, Citron BA. 2006. Minocycline neuroprotects, reduces microgliosis, and inhibits caspase protease expression early after spinal cord injury. J Neurochem 97(5):1314-1326. Hemendinger RA, Armstrong EJ, 3rd, Persinski R, Todd J, Mougeot JL, Volvovitz F, Rosenfeld J. 2008. Huperzine A provides neuroprotection against several cell death inducers using in vitro model systems of motor neuron cell death. Neurotox Res 13(1 ):49-61. Howe CL, Bieber AJ, Warrington AE, Pease LR, Rodriguez M. 2004. Antiapoptotic signaling by a remyelination-promoting human antimyelin antibody. Neurobiol Dis 15(1):120-131.
Huang DW, McKerracher L, Braun PE, David S. 1999. A therapeutic vaccine approach to stimulate axon regeneration in the adult mammalian spinal cord. Neuron 24(3):639-647.
Joshi M, Fehlings MG. 2002. Development and characterization of a novel, graded model of clip compressive spinal cord injury in the mouse: Part 1. Clip design, behavioral outcomes, and histopathology. J Neurotrauma 19(2):175-190. McGavern DB, Murray PD, Rivera-Quinones C, Schmelzer JD, Low PA, Rodriguez M. 2000. Axonal loss results in spinal cord atrophy, electrophysiological abnormalities and neurological deficits following demyelination in a chronic inflammatory model of multiple sclerosis. Brain 123 Pt 3:519-531.
McTigue DM, Tripathi R, Wei P. 2006. NG2 colocalizes with axons and is expressed by a mixed cell population in spinal cord lesions. J Neuropathol Exp Neurol 65(4):406-420.
Mikami Y, Toda M, Watanabe M, Nakamura M, Toyama Y, Kawakami Y. 2002. A simple and reliable behavioral analysis of locomotor function after spinal cord injury in mice. Technical note. J Neurosurg 97(1 Suppl):142-147. Mitsunaga Y, Ciric B, Van Keulen V, Warrington Ae , Paz Soldan M, Bieber AJ, Rodriguez M, Pease LR. 2002. Direct evidence that a human antibody derived from patient serum can promote myelin repair in a mouse model of chronic- progressive demyelinating disease. Faseb J 16(10):1325-1327. Pannu R, Christie DK, Barbosa E, Singh I, Singh AK. 2007. Post-trauma Lipitor treatment prevents endothelial dysfunction, facilitates neuroprotection, and promotes locomotor recovery following spinal cord injury. J Neurochem 101(1 ):182-200.
Pirko I, Ciric B, Gamez J, Bieber AJ, Warrington AE, Johnson AJ, Hanson DP, Pease LR, Macura SI, Rodriguez M. 2004. A human antibody that promotes remyelination enters the CNS and decreases lesion load as detected by T2-weighted spinal cord MRI in a virus-induced murine model of MS. Faseb J 18(13):1577-1579. Rodriguez M, Lennon VA, Benveniste EN, Merrill JE. 1987. Remyelination by oligodendrocytes stimulated by an tiserum to spinal cord. J Neuropathol Exp Neurol 46(1):84-95.
Warrington AE, Asakura K, Bieber AJ, Ciric B, Van Keulen V, Kaveri SV, Kyle RA, Pease LR, Rodriguez M. 2000. Human monoclonal antibodies reactive to oligodendrocytes promote remyelination in a model of multiple sclerosis. Proc Natl Acad Sci U S A 97(12):6820-6825.
Warrington AE, BieberAJ, Ciric B, Pease LR, Van Keulen V, Rodriguez M. 2007. A recombinant human IgM promotes myelin repair after a single, very low dose. J Neurosci Res 85(5):967-976.
Warrington AE, Bieber AJ, Van Keulen V, Ciric B, Pease LR, Rodriguez M. 2004. Neuron-binding human monoclonal antibodies support central nervous system neurite extension. J Neuropathol Exp Neurol 63(5):461-473.
Ejemplo 13: Modelo de esclerosis múltiple - Dosis: evaluación de la respuesta
Los anticuerpos IgM son evaluados por su capacidad para promover la mejora funcional y aumentar los procesos de la remielinización, el crecimiento de neuritas o ambos en un modelo de esclerosis múltiple en ratones. El modelo implica el uso de Virus Murino Encefalomielitis del picornavirus Theiler (TMEV) para inducir lesiones crónicas persistentes, desmielinizantes progresivas del SNC con características similares a los encontrados en MS humano usando un método similar al Warrington et al., 2007.
Ratones hembra y macho (aproximadamente 8 w, cepa SJL/J) son inyectados con virus de encefalomielitis murinos de Theiler (2 x 105 unidades de formación de placa de cepa de Daniel, 10 pl intracerebroventricularmente). Los ratones se dejaron recuperar durante un período de 6 meses antes de la aleatorización para el tratamiento. Los ratones se examinan a continuación, para las condiciones del pelaje, la marcha y el reflejo de enderezamiento y se asignaron aleatoriamente a grupos de tratamiento (como se indica en las tablas de tratamiento). Los ratones se
trataron a continuación con una sola inyección intravenosa de vehículo (solución salina normal) o anticuerpo IgM humana recombinante (0,025 a 2,5 mg/kg). La función neurológica se supervisa a las 2 semanas, 1 mes y 2 meses después del tratamiento.
Criterios de valoración funcionales incluyen puntuación basada en las condiciones del pelaje, la marcha y el rendimiento rotarod. Para la apariencia de capa, la puntuación es como sigue: sin enfermedad (0); cambios mínimos de la capa (1); la capa desaliñada (2); la incontinencia y la capa manchada (3). Cambios en la actividad se cuantifican en una caja de actividad automatizada. Además, la marcha se analiza usando adquisición de la marcha digital (DigiGait) usando velocidades de marcha> 90 cm/s. Puntos finales cuantitativos de análisis de la marcha incluyen anchura de la postura y la duración, longitud de la zancada y frecuencia, ángulo de la pata así como oscilación, el frenado y la duración de propulsión. Cambios de la marcha de la línea de base se cuantifican. Rendimiento Rotarod (una medida de la coordinación sensorial y el equilibrio se evalúa por la capacidad del animal para caminar sobre un eje de rotación midiendo el motor en términos de velocidad y la duración en el eje rotativo) se cuantifica como la cantidad de tiempo que los animales permanecen en el aparato giratorio a través de una serie de ensayos.
En la realización del experimento, los animales son sacrificados por la sobreexposición a CO2. Los cerebros y las médulas espinales se retiran para la evaluación del 1 ) grado de mielinización (análisis de microscopía electrónica de tejido osmicado fijo de paraformaldehído/glutaraldehído incrustado) y 2 ) el crecimiento de las neuritas (evaluación de la densidad de las neuritas por la evaluación estereológica).
En la primera realización, los anticuerpos humanos recombinantes se administran solos en tres niveles de dosis como se indica en la Tabla 3A y en la Tabla 3B.
Tabla 3A. Resument de grupos de tratamiento y puntos finales - anticuerpos IgM recombinantes individuales

Tabla 3B. Resumen de resultados para grupos de tratamiento

(*) X indica una mejora, X indica una mayor mejora, y X + indica aún más mejora. Los valores de puntuación de mejora están en relación con la dosificación de un anticuerpo dado. Así, un X es una mejora mayor que X para ese mismo Ab. La mejora X para un Ab no es necesariamente el mismo que el valor X para otro Ab; el valor X de un Ab no es necesariamente el mismo que el valor X de otro Ab, y el valor X ++ de un Ab no es necesariamente el mismo que el valor X ++ de otro Ab.
En este ejemplo, los anticuerpos recombinantes IgM12, IgM 42, IgM22 y IgM46 están cada uno encontrado para producir mejoras estadísticamente significativas, dependientes de la dosis en varios puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod (p <0,05) para cada punto final en comparación con el tratamiento con vehículo). Los animales tratados con vehículo producen un déficit estable en varios puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod dentro de los 6 meses de lesión TMEV. Un cambio significativo dependiente de la dosis en los puntos finales sensoriomotores sobre el rango de dosis desde 0,025 hasta 2,5 mg/kg, donde X es una mejora, se encuentran para cada Ab, con un aumento X mejorado en puntos finales de sensoriomotor a la dosis baja, un mayor aumento de mejora X de puntos finales sensoriomotores con 0,25 mg/kg y una mejora aún mayor X ++ en puntos finales sensoriomotores en la dosis de 2,5 mg/kg. Esta tendencia se observa para tamaños de los grupos de 12 animales alimentados a un nivel de 0,8 y un nivel alfa de 0,05. Por lo tanto, cada uno de los anticuerpos IgM12, IgM 42, IgM22 y IgM46 producen mejoras estadísticamente significativas, dependientes de la dosis en varios puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod (p <0,05 para cada punto final) en comparación con el tratamiento con vehículo. No hay diferencias en la mejora basada en el género.
En este Ejemplo, se encuentra que IgM12 y IgM42 producen incrementos estadísticamente significativos, dependientes de la dosis en el crecimiento de neuritas en las secciones del cerebro y la médula espinal de ratones infectados con TMEV según la evaluación de la estereología en comparación con los controles tratados con vehículo. Los cambios significativos, dependientes de la dosis en el crecimiento de neuritas sobre el rango de dosis desde 0,025 hasta 2,5 mg/kg, donde X es una mejora, se encuentran para cada Ab, con un aumento de mejora X en el crecimiento de neuritas a la dosis baja, una mayor mejora X se aumenta en el crecimiento de neuritas con 0,25 mg/kg e incluso un mayor aumento en el crecimiento de neuritas X ++ al 2,5 mg/kg de dosis. Esta tendencia se observa para tamaños de los grupos de 12 animales alimentados a un nivel de 0,8 y un nivel alfa de 0,05.
Se encuentra que IgM22 y IgM46 producen incrementos estadísticamente significativos, dependientes de la dosis en la remielinización en secciones del cerebro y la médula espinal de ratones infectados con TMEV según la evaluación de la estereología en comparación con los controles tratados con vehículo. Los cambios significativos, dependientes de la dosis en la remielinización más el intervalo de dosis de 0,025 a 2,5 mg/kg, donde X es una mejora, se encuentran para cada Ab, con un aumento de mejora X en la remielinización a la dosis baja, un aumento de mayor mejora X en la remielinización con 0,25 mg/kg e incluso un mayor aumento en la remielinización X ++ en la dosis de 2,5 mg/kg. Esta tendencia se observa para tamaños de los grupos de 12 animales alimentados a un nivel de 0,8 y un nivel alfa de 0,05.
Ejemplo 14: Modelo de esclerosis múltiple - combinaciones Ab
En este experimento, varias combinaciones de relaciones de dosis fijas de anticuerpos IgM recombinantes se examinan para la mejora del resultado neurológico en el modelo de TMEV de MS como se describe en el ejemplo anterior. Seis meses después de la exposición a TMEV, el tratamiento con combinaciones de IgM recombinantes se inicia en ratones. En esta serie de estudios, a los ratones se les administró vehículo solo, o diversas combinaciones de las IgM, en una única administración intravenosa (mezcla). Puntos finales sensorimotores son supervisados como anteriormente. Mielinización y el crecimiento de las neuritas se evalúan para examinar los cambios producidos por combinaciones de anticuerpos. Las combinaciones bajo evaluación se muestran en la Tabla 4A y en la Tabla 4B.
Tabla 4A. Resumen de grupos de tratamiento - Combinaciones de anticuerpo IgM recombinante

Tabla 4A. Resumen de grupos de tratamiento - Combinaciones de anticuerpo IgM recombinante

(*) X indica una mejora, X indica una mayor mejoría, y X + indica aún más mejora. Los valores de puntuación de mejora están en relación con la dosificación de un anticuerpo dado. Así, un X es una mejora mayor que X para ese mismo Ab. La mejora X para un Ab no es necesariamente el mismo que el valor X para otro Ab; el valor X de un Ab no es necesariamente el mismo que el valor X de otro Ab, y el valor X ++ de un Ab no es necesariamente el mismo que el valor X ++ de otro Ab.
En este ejemplo, las combinaciones de anticuerpos recombinantes de IgM12 IgM42, IgM22 IgM46, IgM12 IgM22, IgM12 IgM46, IgM42 IgM22, y IgM42 IgM46 producen mejoras estadísticamente significativas, dependientes de la dosis en varios puntos finales sensoriomotores incluyendo condición de revestimiento, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo). No se han encontrado diferencias en la mejora basadas en el género.
Además, el grado de mejora de punto final de sensoriomotor con cada una de estas combinaciones es supraaditivo (sinergismo exhibido) de una manera estadísticamente significativa, dependiente de la dosis en comparación con las mejoras aditivas predichas de cada uno de los anticuerpos solos. Una vez más, no se encuentran diferencias en la mejora basadas en el género.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en puntos finales diferentes sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM12 IgM42 potenció de forma significativa el crecimiento de neuritas en comparación con los controles del vehículo (p <0,05). El grado de crecimiento de las neuritas es supraaditivo en comparación con el efecto aditivo de cada anticuerpo administrado solo. Por lo tanto, se entiende que estos anticuerpos provocan la excrecencia por distintos mecanismos de acción; esto es consistente con los respectivos patrones de unión diferentes de estos anticuerpos en el tejido neural.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM22 IgM 46 produjo un aumento significativo en la mielinización comparación con los controles tratados con vehículo (p <0,05). Además, el grado de remielinización es supraaditivo en comparación con el efecto aditivo predicho de IgM22 o IgM46 se administra solo. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, las células provocan respectivamente la producción paracrina que afectan positivamente el otro tipo de célula en evaluación.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM12 IgM 22 producen un aumento significativo en la mielinización así como el crecimiento de neuronas en comparación con controles tratados con vehículo en la médula espinal lesionada (p <0,05). Además, el grado de crecimiento de las neuronas y la remielinización se supraaditiva en comparación con el efecto de IgM12 o IgM22, respectivamente, administrado solo a las dosis utilizadas aquí. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, estas células provocan respectivamente la producción paracrina que afecta positivamente el otro tipo de célula en evaluación.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM12 IgM 46 producen un aumento significativo en la mielinización así como el crecimiento de neuronas en comparación con controles tratados con vehículo (p <0,05). Además, el grado de crecimiento de las neuronas y la remielinización es supraaditiva en comparación con el efecto de IgM12 o IgM46, respectivamente, administrado solo a las dosis utilizadas aquí. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, las células provocan respectivamente la producción paracrina que afectan positivamente el otro tipo de célula en evaluación.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM22 IgM 42 producen un aumento significativo en la mielinización así como el crecimiento de neuronas en comparación con controles tratados con vehículo (p <0,05). Además, el grado de crecimiento de las neuronas y la remielinización es supraaditiva en comparación con el efecto de IgM22 o IgM42, respectivamente, administrado solo a las dosis utilizadas aquí. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, las células provocan respectivamente la producción paracrina que afectan positivamente el otro tipo de célula en evaluación.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM46 IgM 42 producen un aumento significativo en la mielinización así como el crecimiento de neuronas en comparación con controles tratados con vehículo (p <0,05). Además, el grado de crecimiento de las neuronas y la remielinización es supraaditiva en comparación con el efecto de IgM46 o IgM42, respectivamente, administrado solo a las dosis utilizadas aquí. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, las células provocan respectivamente la producción paracrina que afectan positivamente el otro tipo de célula en evaluación.
Ejemplo 15: lesión de la médula espinal Modelo
Los anticuerpos IgM son evaluados por su capacidad para promover la mejora funcional y aumentar los procesos de la remielinización, el crecimiento de neuritas o ambos en un modelo de lesión de la médula espinal en ratas. El modelo implica la compresión lateral de la médula espinal entre las paletas del fórceps cubreobjetos modificado, donde la anchura de la hoja de fórceps es de 2,5 mm, a una separación predeterminada de las cuchillas a 0,9 mm durante un período de 15 segundos. La lesión resultante tiene características similares a las encontradas en SCI humana (Gruner et al., 1995)
Ratas macho y hembra (aproximadamente 200-225 g, Long Evans) se someten a cirugía para producir una lesión en la médula espinal como se ha descrito anteriormente. Anticuerpos IgM recombinantes o tratamiento con vehículo se administran por vía intravenosa 10 min después de la cirugía. La función del miembro locomotor Hind y la marcha son evaluados por un período de 1 2 semanas utilizando la escala de calificación locomotora BBB (véase, por ejemplo, Basso DM, Beattie Ms , Bresnahan JC, Anderson DK, Faden Al, Gruner JA, Holford TR, Hsu CY, Noble LJ, Nockels R, Perot PL, SK Salzman, W. Young (1996) MASCIS evaluation of open field locomotor scores: Effects of experience and teamwork on reliability. Journal of Neurotrauma., 13: 343-359; Basso MS, MS Beattie, Bresnahan JC (1995) a escala de calificación locomotor sensible y fiable para las pruebas de campo abierto en ratas. Journal of Neurotrauma, 12: 1-21) Las ratas se ensayaron a 1, 3, 5, 7 y 10 días, y luego semanalmente después de SCI durante 9-12 semanas. Se cuantifican cambios de BBB a partir de la línea de base.
En la realización del experimento, los animales son sacrificados por la sobreexposición a CO2. Los cerebros y las médulas espinales se retiran para la evaluación del 1 ) grado de mielinización (análisis de microscopía electrónica de tejido osmicado fijo maldehído/glutaraldehído incrustado en plástico) y 2 ) el crecimiento de las neuritas (evaluación de la densidad de las neuritas por la evaluación estereológica).
Ejemplo 16: El uso de anticuerpos individuales en un modelo de lesión de médula espinal
En este experimento, los anticuerpos humanos recombinantes se administran solos en tres niveles de dosis como se indica en la Tabla 5A y la Tabla 5B.
Tabla 5A. Resumen de grupos de tratamiento y puntos finales - anticuerpos IgM recombinantes individuales

Tabla 5B. Resumen de resultados para grupos de tratamiento

(*) X indica una mejora, X indica una mayor mejora, y X + indica aún más mejora. Los valores de puntuación de mejora están en relación con la dosificación de un anticuerpo dado. Así, un X es una mejora mayor que X para ese mismo Ab. La mejora X para un Ab no es necesariamente el mismo que el valor X para otro Ab; el valor X de un Ab no es necesariamente el mismo que el valor X de otro Ab, y el valor X ++ de un Ab no es necesariamente el mismo que el valor X ++ de otro Ab.
En este ejemplo, los anticuerpos recombinantes IgM12, IgM 42, IgM22 y IgM46 están cada uno encontrados para producir mejoras estadísticamente significativas, dependiendo de la dosis en función de la extremidad locomotora trasera con mejores parámetros de BBB (p <0,05 para cada punto final en comparación con el tratamiento con vehículo). Los animales tratados con vehículo dan una puntuación estable BBB dentro de las 6 semanas de un nivel moderado de lesión de la médula espinal. Un cambio significativo dependiente de la dosis en la puntuación de BBB sobre el rango de dosis desde 0,025 hasta 2,5 mg/kg, donde X es una mejora, se encuentran para cada Ab, con un incremento de mejora X en el nivel de la acreditación a la dosis baja, una mayor mejora X aumento BBB con 0,25 mg/kg e incluso un mayor BBB mejora X ++ en la dosis de 2,5 mg/kg. Esta tendencia se observa para tamaños de los grupos de 12 animales alimentados a un nivel de 0,8 y un nivel alfa de 0,05. Por lo tanto, cada uno de los anticuerpos IgM12, IgM 42, distinciones IgM22 y IgM46. No hay distinciones en la mejora sobre la base de género. Producir mejoras estadísticamente significativas, dependientes de la dosis en función locomotora de la extremidad posterior con BBB mejorado (p <0,05 para cada punto final) en comparación con el tratamiento con vehículo.
IgM12 y IgM42 se encuentran para producir incrementos estadísticamente significativos, dependientes de la dosis en el crecimiento de neuritas en las secciones del cerebro y la médula espinal de ratones infectados con TMEV según la evaluación de la estereología en comparación con los controles tratados con vehículo. Los cambios significativos, dependientes de la dosis en el crecimiento de neuritas sobre el rango de dosis desde 0,025 hasta 2,5 mg/kg, donde X es una mejora, se encuentran para cada Ab, con un aumento de mejora X en el crecimiento de neuritas a la dosis baja, una mayor mejora X aumentar en el crecimiento de neuritas con 0,25 mg/kg e incluso un mayor aumento en el crecimiento de neuritas X ++ al 2,5 mg/kg de dosis. Esta tendencia se observa para tamaños de los grupos de 12 animales alimentados a un nivel de 0,8 y un nivel alfa de 0,05.
IgM22 y IgM46 se encuentran para producir incrementos estadísticamente significativos, dependientes de la dosis en la remielinización en secciones del cerebro y la médula espinal de ratones infectados con TMEV según la evaluación de la estereología comparación con los controles tratados con vehículo. Los cambios significativos, dependientes de la dosis en la remielinización más el intervalo de dosis de 0,025 a 2,5 mg/kg, donde X es una mejora, se encuentran para cada Ab, con un aumento mejora X en la remielinización a la dosis baja, una mayor mejora X aumento en la remielinización con 0,25 mg/kg e incluso un mayor aumento en la remielinización X ++ en la dosis de 2,5 mg/kg. Esta tendencia se observa para tamaños de los grupos de 12 animales alimentados a un nivel de 0,8 y un nivel alfa de 0,05.
Ejemplo 17: Combinaciones de Anticuerpo de Lesión de Médula Espinal
En este ejemplo, varias combinaciones de relaciones de dosis fijas de anticuerpos IgM recombinantes se
examinan para la mejora del resultado neurológico en el modelo de lesión de la médula espinal como se ha descrito en el Ejemplo 14 en el presente documento. Por consiguiente, el tratamiento intravenoso con combinaciones de IgM de anticuerpos (como aditivos) o control de vehículo se le da 10 min después de la lesión. Puntos finales locomotores son supervisados como anteriormente. Además, la mielinización y el crecimiento de neuritas se evalúan para examinar los cambios producidos por combinaciones de anticuerpos. Las combinaciones evaluadas se muestran en la Tabla 6 A y en la Tabla 6 B.
Tabla 6A. Resumen de grupos de tratamiento - combinaciones de anticuerpo IgM recombinantes
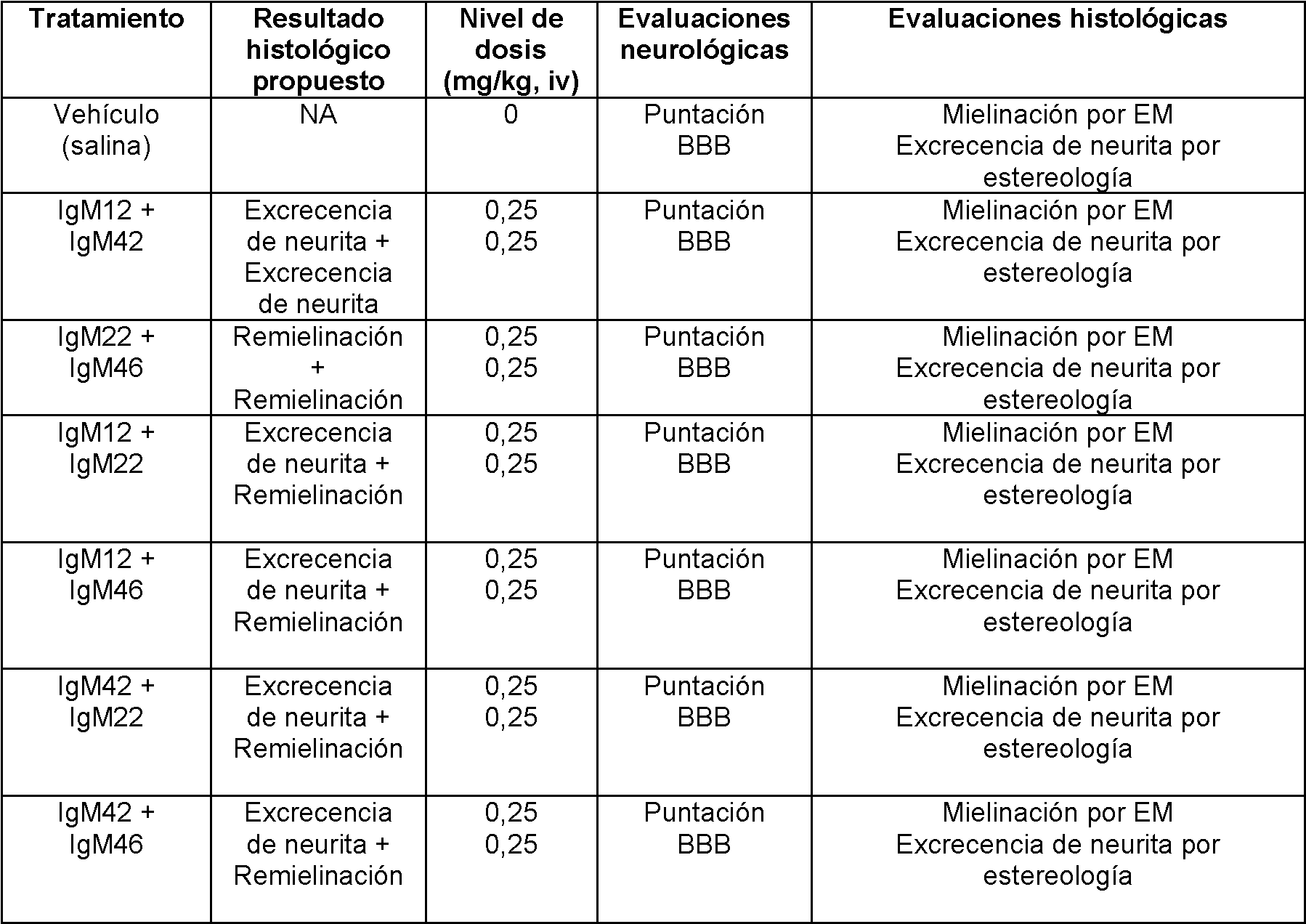
Tabla 6B. Resumen de grupos de tratamiento - Resultados

(*) X indica una mejora, X indica una mayor mejora, y X + indica aún más mejora. Los valores de puntuación de mejora están en relación con la dosificación de un anticuerpo dado. Así, un X es una mejora mayor que X para ese mismo Ab. La mejora X para un Ab no es necesariamente el mismo que el valor X para otro Ab; valor X de un Ab no es necesariamente el mismo que el valor X de otro Ab, y valor X ++ de un Ab no es necesariamente el mismo que el valor X ++ de otro Ab.
En este experimento, cada una de las combinaciones de anticuerpos recombinantes IgM12 IgM42, IgM22 IgM46, IgM12 IgM22, IgM12 IgM46, IgM42 IgM22, y IgM42 IgM46 se encuentran para producir mejoras estadísticamente significativas, dependientes de la dosis en función de las extremidades posteriores (puntos finales motriz) que se encuentran por parámetros BBB. No hay diferencias en la mejora basadas en el género.
Por otra parte, cada una de las combinaciones se encuentra para mejorar la función locomotora en un supra-aditivo (es decir sinergístico) de forma en comparación con las mejoras de cada uno de los anticuerpos solos en la misma dosis. Una vez más, no se encuentran diferencias en la mejora basadas en el género.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM12 IgM42 mejorado significativamente el crecimiento de neuritas en comparación con los controles del vehículo (p <0,05). El grado de crecimiento de las neuritas es supraaditivo en comparación con el efecto aditivo de cada anticuerpo administrado solo. Por lo tanto, se entiende que estos anticuerpos provocan la excrecencia por distintos mecanismos de acción; esto es consistente con los respectivos patrones de unión diferentes de estos anticuerpos en el tejido neural.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM22 IgM 46 produjo un aumento significativo en la mielinización en comparación con los controles tratados con vehículo (p <0,05). Además, el grado de remielinización es supraaditivo en comparación con el efecto aditivo predicho de IgM22 o IgM46 administrado solo. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, las células provocan respectivamente la producción paracrina que afectan positivamente el otro tipo de célula en evaluación.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM12 IgM 22 producen un aumento significativo en la mielinización así como el crecimiento de neuronas en comparación con controles tratados con vehículo en la médula espinal lesionada (p <0,05). Además, el grado de crecimiento de las neuronas y la remielinización es supraaditiva en comparación con el efecto de IgM12 o IgM22, respectivamente, administrado solo a las dosis utilizadas aquí. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, estas células provocan respectivamente la producción paracrina que afecta positivamente el otro tipo de célula en evaluación.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM12 IgM 46 producen un aumento significativo en la mielinización así como el crecimiento de neuronas en comparación con controles tratados con vehículo (p <0,05). Además, el grado de crecimiento de las neuronas y la remielinización es supraaditiva en comparación con el efecto de IgM12 o IgM46, respectivamente, administrado solo a las dosis utilizadas aquí. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, las células provocan respectivamente la producción paracrina que afectan positivamente el otro tipo de célula en evaluación.
Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM22 IgM 42 producen un aumento significativo en la mielinización así como el crecimiento de neuronas en comparación con controles tratados con vehículo (p <0,05). Además, el grado de crecimiento de las neuronas y la remielinización es supraaditiva en comparación con el efecto de IgM22 o IgM42, respectivamente, administrado solo a las dosis utilizadas aquí. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, las células provocan respectivamente la producción paracrina que afectan positivamente el otro tipo de célula en evaluación.
[0309] Además de producir mejoras estadísticamente significativas, dependientes de la dosis en diferentes puntos finales sensoriomotores incluyendo las condiciones del pelaje, parámetros de la marcha y la puntuación Rotarod en comparación con el tratamiento con vehículo (p <0,05 para cada punto final en comparación con el tratamiento con vehículo), la combinación de IgM46 IgM 42 producen un aumento significativo en la mielinización así como el crecimiento de neuronas en comparación con controles tratados con vehículo (p <0,05). Además, el grado de crecimiento de las neuronas y la remielinización es supraaditiva en comparación con el efecto de IgM46 o IgM42, respectivamente, administrado solo a las dosis utilizadas aquí. Sin estar ligado por la teoría, esto se entiende que es un resultado de la actividad paracrina. Por consiguiente, cuando crecen las neuronas y oligodendrocitos en crecimiento están presentes cada uno, las células provocan respectivamente la producción paracrina que afectan positivamente el otro tipo de célula en evaluación.
Ejemplo 18: Anticuerpo recombinante rHIgM12, una IgM de unión de neurona humana, protege los axones de la médula espinal
Hemos demostrado que un anticuerpo de suero humano natural, sHIgM12, se une a las neuronas in vitro y promueve el crecimiento de neuritas. Hemos generado una forma recombinante, rHIgM12, con propiedades idénticas. Infección intracerebral con virus de encefalomielitis murinos de Theiler (TMEV) de cepas de ratón susceptibles causa la enfermedad desmielinizante crónica con pérdida axonal progresiva y disfunción neurológica similar a las formas progresivas de la esclerosis múltiple. El tratamiento de este modelo con anticuerpos de clase IgM que se unen a los oligodendrocitos mejora la remielinización del SNC (Warrington et al (2007) J Neurosci Res 85: 967-976). En contraste, el anticuerpo monoclonal humano derivado del suero (sHIgM12) que se une a las
neuronas, promueve robusto crecimiento de neuritas en el mismo grado como laminina, y reduce los efectos inhibidores de la mielina del SNC sobre el crecimiento de neuritas (Warrington et al (2004) J Neuropathol Exp Neurol 63 (5): 461-473). Más recientemente una forma recombinante de sHIgM12 humano se generó (rHIgM12) con propiedades biológicas idénticas, como se describe anteriormente. Para estudiar los efectos de rHIgM12 sobre la integridad axonal en la médula espinal se realizó etiquetado retrogrado, una técnica que se basa en los axones anatómicos continuos y funcionalmente conservados.
MÉTODOS
Etiquetado retrógrado: Después de anestesiar los ratones, laminectomía dorsal se realizó en la vertebra torácica inferior (T10-11). La médula espinal se hemi-seccionó en el lado derecho y el sitio de hemisección se rellenó con una solución estéril de 4% Fluorogold. Una semana después de la cirugía, los ratones se sacrificaron, y los cerebros se retiraron. Las secciones seriadas (de 40 mm de espesor) del tronco cerebral se hicieron y se analizó cada cuarta sección. Neuronas grandes fluorescentes brillantes fueron contadas bajo 200x aumentos de 16 rebanadas del tronco cerebral.
RESULTADOS
El modelo de TMEV ofrece una plataforma para desarrollar estrategias para promover la neuroprotección y prevenir la pérdida de axones. Las etapas tempranas de la enfermedad incluyen inflamación y desmielinización; etapas posteriores se presentan con la pérdida de axones y los déficits funcionales. Como se detalla en los ejemplos anteriores, rHIgM12 entra en la médula espinal y se localiza en los axones de neurofilamentos (NF) después de una sola inyección ip 1mg, como se confirma por las imágenes confocales (FIGURA 15). rHIgM12 también se colocalizó como haces de fibras NF (FIGURA 15D) de corte transversal. En estudios con animales, rHIgM12 mejora el número de neuronas de tronco cerebral retrógrado marcado. rHIgM12 o solución salina se administraron a ratones SJL a los 90 días después de la infección (dpi). Cirugía de la médula espinal para el etiquetado retrógrado se realizó 9 semanas después del tratamiento. Una semana después de la cirugía, los ratones se sacrificaron y las neuronas marcadas con fluorescencia se cuantificaron en las secciones de la serie del tronco cerebral. rHIgM12 mejora el número de neuronas de marcado retrógrado del tronco cerebral, en comparación con solución salina de control (datos no mostrados). Por lo tanto, los ratones tratados con rHIgM12 tenía neuronas del tronco cerebral más retrógradas-etiquetadas como comparación con los controles tratados con solución salina.
Ejemplo 19: Una sola dosis de la neurona de unión a anticuerpo monoclonal humano mejora la actividad espontánea en un modelo murino de desmielinización
Nuestro laboratorio demostró que un anticuerpo de suero humano natural, sHIgM12, se une a las neuronas in vitro y promueve el crecimiento de neuritas. Hemos generado una forma recombinante, rHIgM12, con propiedades idénticas. Infección intracerebral con Virus de Encefalomielitis Murinos de Theiler (TMEV) de cepas de ratón susceptibles resulta en la enfermedad desmielinizante crónica con pérdida axonal progresiva y disfunción neurológica similar a las formas progresivas de la esclerosis múltiple. Para estudiar los efectos de rHIgM12 en la función motora de los ratones infectados con TMEV, que supervisó la actividad nocturna espontánea durante muchas semanas. Comportamiento nocturno es una medida sensible de la función neurológica roedores porque se espera que la actividad máxima cambios que se produzcan durante el período de seguimiento durante la noche normalmente activo. Los ratones se colocaron en cajas de actividad ocho días antes del tratamiento para recoger actividad basal espontánea. Después del tratamiento, la actividad en cada grupo se registró continuamente durante 8 semanas. Elegimos un largo período de seguimiento de 8 semanas por dos razones: (1) Hemos demostrado anteriormente que la remielinización inducida IgM está presente por 5 semanas de tratamiento post, y enfermedad desmielinizante inducida por TMEV (2) en esta cepa progresa muy lentamente. Debido a los largos períodos de observación y grandes conjuntos de datos, las diferencias entre los grupos de tratamiento pueden ser difíciles de apreciar el estudio de las grabaciones sin filtrar originales. Para delinear claramente los cambios en los datos originales altamente fluctuante aplicamos tres métodos diferentes: (1) de agrupación, (2) la aplicación de filtros gaussianos paso bajo (GF) y (3) ajuste polinómico. El uso de cada uno de los tres métodos que mostramos que en comparación con el control de IgM y solución salina, el tratamiento temprano con una mejora inducida rHIgM12 tanto en la función de motor horizontal y vertical, mientras que el tratamiento posterior mejoró única actividad horizontal. rHIgM12 no alteró la actividad de ratones normales, no infectadas. Este estudio apoya la hipótesis de que el tratamiento con una neurona de unión a IgM no sólo protege a las neuronas in vitro, sino que también influye mejora motor funcional.
Introducción
Seguimiento y análisis de la función neurológica en modelos de enfermedad de roedor largo plazo sigue siendo un reto. El comportamiento nocturno es una medida sensible de la función neurológica de roedor porque se espera que los cambios de actividad máxima a ocurran durante el período de seguimiento de la noche normalmente activo [1]. Sin embargo, en modelos animales de enfermedades de progresión lenta, supervisión del rendimiento a menudo se extiende durante varias semanas. Hemos informado anteriormente del anticuerpo de unión a
oligodendrocitos (rHIgM22) mejoró la remielinización por 5 semanas después del tratamiento
Publicación Duración de la sesión grabada de actividad locomotora 
[2]. Tomados en conjunto que tanto la evolución de la enfermedad y la reparación es un proceso lento y para asegurar que cualquier fluctuación en la actividad se tiene en cuenta, que supervisó la actividad prolongada con la misma densidad de muestreo utilizado en monitorización a corto plazo. Esto creó grandes y altamente fluctuantes conjuntos de datos (FIGURA 32A, C). Para delinear claramente los cambios post-tratamiento y recuperación de las tendencias generales en la actividad a largo plazo, se comparó el uso de agrupamiento de datos, filtrado gaussiano, y el ajuste de polinomio mediante el uso de Mathematica (Wolfram Research, Inc.).
La infección intracerebral con Virus de Encefalomielitis Murinos de Theiler (TMEV) de cepas de ratón susceptibles resultan en la enfermedad desmielinizante crónica con disfunción neurológica progresiva similar a las formas progresivas de esclerosis múltiple [3]. El tratamiento de este modelo con anticuerpos de clase IgM que se unen a los oligodendrocitos mejora la remielinización del SNC [4]. En contraste, un anticuerpo monoclonal derivado de suero humano (sHIgM12) que se une a las neuronas, promueve el crecimiento de neuritas robusto en el mismo grado como laminina, y reduce los efectos inhibidores de la mielina del SNC sobre el crecimiento de neuritas [5]. Más recientemente una forma recombinante de sHIgM12 humano se generó (rHIgM12) con propiedades biológicas idénticas. Nos mostró anteriormente que rHIgM12 tiene una vida media de 3,6 horas, pero todavía atraviesa la barrera hematoencefálica y se une a los tejidos nerviosos (observación no publicada). Para estudiar los efectos de rHIgM12 sobre la actividad de los ratones infectados con TMEV monitorizamos la actividad espontánea durante varias semanas utilizando cajas de actividad AccuScan (Accuscan Instruments, Inc., Columbus, OH). Elegimos un período relativamente largo de seguimiento de 8 semanas por dos razones: (1) la remielinización inducida IgM está presente por 5 semanas después del tratamiento, y enfermedad desmielinizante inducida por TMEV (2) en esta cepa progresa muy lentamente en comparación con los modelos de EAE puramente autoinmunes de MS [6]. Debido a los periodos de observación largos en comparación con los estudios publicados previamente (TABLA 7), la identificación de cambios por inspección visual de los datos en bruto resultó difícil.
TABLA 7 - Longitud de supervisión de la actividad espontánea en los estudios publicados en comparación con el estudio actual

Materiales y métodos:
Ratones: Ratones SJL/J (Jackson Laboratories, Bar Harbor, ME) fueron alojados y criados en la instalación de cuidado de los animales en Mayo Clinic. Protocolos de animales fueron aprobados por el Mayo Clinic_Institutional Animal Care and Use Committee.
Modelo de virus de Theiler de desmielinización: Enfermedad desmielinizante fue inducida en ratones hembra de 6 a 8 semanas de edad por inyección intracerebral de TMEV. Una aguja de calibre 27 administró 10 l que contenía 2,0 x 105 unidades formadoras de placas de cepa de Daniel TMEV. Esto resultó en> incidencia del 98% de la infección con víctimas mortales raras. Todos los animales desarrollaron encefalitis leve, que se resolvió dentro de 2 semanas. Animales empeoran durante los próximos 6-8 meses con enfermedad desmielinizante crónica de la médula espinal. Lesiones y la pérdida de axones comienzan tres meses después de la infección se correlaciona con disfunción neurológica [3].
Tratamiento de anticuerpo: ratones SJL (no infectados, 45 y 90 días después de la infección) fueron tratados con una sola dosis 200 pg intraperitoneal de anticuerpos (o isotipo de control rHIgM12 IgM) disuelto en 0,5 ml de PBS. El tercer grupo se trató con solamente 0,5 ml de p Bs .
Monitorización de la actividad espontánea: La actividad locomotora espontánea se registró con el aparato
Digiscan de campo abierto (OF) (Omnitech Electrónica; Columbus, OH) y el software Versamax, v.4.12-1AFE (Accuscan Instruments, Inc., Columbus, OH). El aparato consta de seis jaulas de acrílico (40x40x30.5cm) soportados por un marco de metal que sostiene dos conjuntos de células de fotos. El dispositivo mide el número de movimientos horizontales y verticales discretos tabulando el número de interrupciones del haz de infrarrojos proyectados. En todas las jaulas ratones fueron expuestos a condiciones ambientales idénticas: (a) la comida libremente accesible y agua, (b) un ciclo normal de 12 horas de luz/oscuridad, (c) 70°F de temperatura ambiente. En cada experimento hiperactivo y, en raras ocasiones, los animales con sobrepeso fueron excluidos y el resto de los ratones fueron asignados al azar. Varios grupos de 5 ratones SJL se colocaron en el centro de cada jaula y la línea de base recogió la actividad espontánea durante el período de 8 días consecutivos. Después de este período, tres grupos de ratones con la actividad de referencia más similar se trataron con rHIgM12, control de anticuerpos IgM o solución salina y luego monitoreados durante el período de 8 semanas. Los datos se recogieron como número de roturas de haz por bloques de 1 hora. Las actividades horizontales y verticales totales se registraron usando el programa Versadat, v.3.02-1AFE (Accuscan Instruments). No podíamos poner más de 5 animales por caja actividad porque este es el máximo número de animales permitidos por Institutional Animal Care and Use Committee (IACUC).
Análisis de datos: Puede ser difícil apreciar las diferencias entre los grupos de tratamiento mediante el estudio de grabaciones original sin filtrar (FIGURA 32 A-C). Para recuperar las tendencias lentas en los datos originales altamente fluctuantes aplicamos tres métodos diferentes: 1) agrupación, 2) aplicación de filtro gaussiano de paso bajo (GF), y 3) ajuste polinómico.
Agrupamiento de datos es el método más simple y sustituye a un grupo de puntos de datos dentro de un bin preseleccionado por su valor medio. En nuestro caso, hemos elegido un cubo nocturno dado que los picos de actividad murina por la noche. Por lo tanto, reemplazamos todas las lecturas nocturnas (12 h/día) por su valor medio como se muestra en las Figuras 33A, 33B, 35C, 35D, 37C y 37D. La comparación de los grupos se puede realizar mediante una simple prueba de la t para una diferencia de medios, con un tamaño efectivo de la muestra de 12 horas por tratamiento. Considerando que estas comparaciones son simples, el tamaño pequeño de la muestra en cada momento los resultados puntuales en grandes errores estándar y las comparaciones estadísticas son de uso limitado. En general, el agrupamiento de datos es un método eficaz para la visualización de datos ruidosos, pero es de uso limitado para la prueba estadística.
El filtrado gaussiano (GF) (también conocido como filtración de paso bajo, suavizado de los datos, o la mejora de sensibilidad) es un procedimiento de reducción de ruido utilizado comúnmente en espectroscopia transformada de Fourier de procesamiento e imagen [7]. GF, realizada con una selección apropiada de ensanchamiento gaussiano (GB, en días), se filtraron las fluctuaciones de alta frecuencia a un nivel deseado. La elección de filtro es arbitraria y puede ser guiada por la velocidad de los cambios de actividad esperados. GF es un método de suavizado utilizando información de puntos tomados desde ambos lados de un valor y la ponderación de la influencia de estos puntos tales que la influencia se desvanece de acuerdo con la función de Gauss. Computacionalmente, GF puede ser implementado de dos maneras equivalentes: 1) la transformación de Fourier (FT) de los datos, seguido por multiplicación con una función gaussiana e inversa FT del producto; y 2) la convolución directa de los datos con un kernel de Gauss. En alto nivel de paquetes de software (Matlab (Mathworks) o Mathematica (Wolfram)) la función de filtro de Gauss está disponible con mínima o ninguna programación por parte del usuario. Con la selección apropiada de GB el método GF detallado anterior permite la visualización simplificada de datos erógenos altamente complejos y Het. Una limitación de GF es que, mientras que permite una fácil visualización de las tendencias, las comparaciones estadísticas se complican por la elección de la GB.
Para la comparación y para demostrar un método que permite la comparación estadística simplificada, nos ajustamos polinomios a los datos. Permitidos para términos polinomiales arriba a través de cualquier grado (xn) Estos modelos, permitieron los términos polinomiales por cualquier grado (xn), y parámetros estimados de la forma por separado para cada grupo de tratamiento (términos de interacción). Nuestra elección de un polinomio de grado sexto fue arbitraria después de explorar varias opciones, pero deja la flexibilidad suficiente para modelar los efectos no lineales en el tiempo. Debido a que los diferentes grupos de tratamiento fueron ajustados de manera óptima usando diferentes polinomios de orden, decidimos permitir la mayor flexibilidad de orden. Se utilizó el Criterio de Información de Akaike (AIC) para determinar el "mejor ajuste" de líneas polinómicas a través de varios de los grupos de tratamiento [8]. AIC es un método de comparación de modelos que equilibra la ganancia en R2 mediante la adición de términos adicionales, pero penaliza para complejidad excesiva (es decir, el uso de grados de libertad). En general, la tercera orden de ajuste fue suficiente para captar la tendencia de los datos, y para algunos casos, el segundo orden o incluso el ajuste lineal fue "mejor" de acuerdo con la AIC. Sin embargo, nuestro objetivo principal cosistió en comparar los grupos de tratamiento en los puntos de tiempo observado. Debido al gran número de puntos de datos, y debido a que no estamos utilizando el ajuste polinómico para predecir (o extrapolar) los valores de tratamiento fuera de nuestros datos observados, el impacto de "sobreajuste" en los resultados es mínimo.
Una ventaja de ajuste polinómico es que la comparación estadística de los grupos de tratamiento es sencilla: tratamientos se pueden comparar en los puntos de tiempo especificados utilizando los valores de modelo predichos y respectivos errores estándar. Comparaciones directas por pares de tratamientos pueden llevarse a cabo
a intervalos regulares a través de todo el período de tiempo para determinar cuando los grupos de tratamiento han divergido significativamente. Por último, el ajuste polinómico elimina medio adicional y el ruido de baja frecuencia que se centra la atención visual en la tendencia general en todo el período de tiempo.
Sin embargo, dentro de cada uno de los experimentos algunos grupos de ratones tenían actividad horizontal y vertical de línea de base algo diferente (8 días). Por lo tanto, se utilizó primero valores de Z para normalizar la actividad de referencia de forma independiente para cada grupo y entonces realiza el filtrado gaussiano o polinomios montados en estos valores.
Análisis estadístico: los datos de intervalos y filtro gaussiano de paso bajo de suavizado de los datos se realizaron usando un macro escrito en Mathematica (Wolfram). Macro e instrucciones están disponibles en mayoresearch.mayo.edu/mayo/research/rodriguez_lab/software.cfm. Modelos de regresión polinómica y comparaciones estadísticas de los grupos de tratamiento se realizaron usando los valores de modelo predichos y los errores estándar respectivos basados en la z-estadística (SAS Institute, Inc.). Comparaciones directas por pares de tratamientos se realizaron para cada día en todo el marco de tiempo, y se determinó la significación estadística en el umbral típico a = 0,05. No se hicieron ajustes para comparaciones múltiples.
RESULTADOS
rHIgM12 mejora la actividad horizontal en los ratones SJL cuando se administra en 90dpi
Se colocaron en cajas de actividad y Basilea tres grupos de 5 ratones SJL a los 90 días después de la infección (dpi) y se midió la actividad durante 8 días consecutivos. Dos grupos de ratones fueron tratados con una única dosis de 200 pg de rHIgM12 o un control de isotipo IgM. El tercer grupo se trató con solución salina. Después del tratamiento, la actividad espontánea se registró continuamente durante 8 semanas. Dado que los datos se recogieron en bloques de 1 hora, se obtuvieron aproximadamente 900 puntos de datos para cada grupo. Estos datos brutos originales (FIGURA 32A-C) es altamente fluctuante, por lo que las diferencias entre los grupos de tratamiento pueden ser difíciles de apreciar. Todos los tres métodos (agrupamiento, filtrado gaussiano y polinómicas de montaje) revelaron que los ratones tratados con rHIgM12 mostraron una mejora en la actividad horizontal, mientras que el control IgM y los ratones tratados con solución salina no tenían cambios en la actividad durante el período de 8 semanas (FIGURA 33A, C y E). El uso de las comparaciones directas por pares de tres tratamientos después del polinomio apropiado, se determinó que la mejora en la función motora horizontal en los ratones tratados con rHIgM12 se convirtió en estadísticamente significativa en el día 7 post tratamiento (comparado con el control IgM) y en el día 11 (en comparación con solución salina) (FIGURA 34). Persistió la mejora observada en la actividad nocturna horizontal de los animales tratados con rHIgM12 durante aproximadamente 30 días y luego regresó a los niveles basales. La comparación por parejas de solución salina - versus grupos de tratamiento de control IgM mostraron significancia estadística para los días 38-52. Sin embargo, cuando se compara con grandes diferencias observadas entre los grupos de tratamiento y los controles rHIgM12-, creemos que estas diferencias entre los grupos de control no son biológicamente significativas. Por otro lado, la actividad vertical no mostró diferencias importantes y fue similar en los tres grupos (FIGURA 33B, D y F).
rHIgM12 mejora la actividad horizontal y vertical en ratones SJL cuando se administra en 45dpi
El estudio anterior utilizó actividad vertical (comportamiento de crianza) como una lectura primaria [1]. Debido a la deserción axonal en ratones crónicamente infectados con TMEV, extremidades traseras se hicieron progresivamente débiles y rígidas por lo que se redujo el encabritarse. En el primer experimento de este estudio, cuando rHIgM12 se administró en el momento de la desmielinización máxima (90dpi) y aparición de la pérdida axonal progresiva, se mejoró sólo la actividad horizontal. El comportamiento de encabritarse no fue afectado y era equivalente en los tres grupos de tratamiento. Por lo tanto, nos preguntamos si el tratamiento en un punto de tiempo anterior puede ser más beneficioso. En un segundo experimento, grupos de 5 ratones se trataron a 45 dpi con una sola dosis de 200 pg de rHIgM12, una IgM de control de isotipo o solución salina. Se usó un diseño experimental idéntico; se recogió la actividad de línea de base durante 8 días, y la actividad de post-tratamiento durante 8 semanas. En este experimento, a partir de alrededor de 2 semanas después del tratamiento, los ratones tratados con rHIgM12 mostraron mejoras claras tanto en la actividad horizontal como vertical. Esto fue evidente después de usar los tres métodos: datos de promediación, la aplicación de filtro gaussiano o ajuste polinómico (FIGURA 35C-H). Control de IgM y de los ratones tratados con solución salina mostraron niveles de actividad similares en todo el final del estudio. En los ratones tratados con rHIgM12, mejora en la actividad horizontal fue evidente hasta que terminó el experimento. Por otro lado, la mejora de la actividad vertical, duró cerca de 4 semanas y luego se redujo a los valores basales. Comparaciones directas por pares de tres tratamientos mostraron que la mejora en la función motora horizontal en los ratones tratados con rHIgM12 divergieron significativamente en el día 13 post-tratamiento (en comparación con el control de IgM) y en el día 9 (en comparación con solución salina) (FIGURA 36A y C). Del mismo modo, la función motora vertical en los ratones tratados con rHIgM12 divergieron significativamente en el día 14 post-tratamiento (comparado con el control IgM) y en el día 6 (en comparación con solución salina) (FIGURA 36B y D). Comparación de control de IgM-frente a los grupos tratados con solución salina no revelaron grandes diferencias biológicas, ya sea para la actividad horizontal o vertical (FIGURA 36E y F).
rHIaM12 no altera la actividad espontánea en ratones no infectados normales.
El tratamiento con rHIgM12 en dos experimentos anteriores mostraron claro efecto beneficioso en ratones infectados neurológicamente discapacitados. Para excluir la posibilidad de que este anticuerpo puedq tener propiedades irritables y por lo tanto provocar un aumento de la función motora, se realizó un experimento similar con los ratones no infectados. Tres grupos de edad emparejados ratones no infectados se trataron con rHIgM12, IgM control o solución salina. En comparación con la mejora de la actividad en ratones infectados, rHIgM12 así como los otros dos tratamientos no indujo ningún aumento en la función motora de ratones normales (FIGURA 37). Los tres grupos mostraron una tendencia a la baja en la actividad espontánea. Este resultado indica que ninguno de los anticuerpos tiene una influencia en el aumento de la actividad en ratones normales.
DISCUSIÓN
Existe una necesidad crítica para el desarrollo de terapias neuroprotectoras para la esclerosis múltiple, así como para otras enfermedades desmielinizantes y neurodegenerativas. Aunque hay algunos fármacos anti inflamatorios que pueden conducir indirectamente a una disminución en el daño axonal durante la enfermedad inflamatoria del s Nc , no hay fármacos que actúan directamente en el nivel de las neuronas/axones. El objetivo principal de la neuroprotección es limitar la disfunción neuronal para tratar de mantener la integridad funcional de las neuronas y los axones. Durante muchos años la desmielinización, la característica patológica de la MS, fue considerada como una de las causas de los déficits neurológicos permanentes. Está claro ahora que la desmielinización es necesaria, pero no suficiente para la pérdida axonal permanente [9]. La desmielinización solamente predispone axones denudados a la lesión secundaria causada por cualquiera de citotoxicidad de células T o pérdida de apoyo neurotrófico local a partir de oligodendrocitos muertos [10].
Observación anterior de que la unión del anticuerpo neurona-sHIgM12 promueve la extensión de neuritas robustas [5] representa un claro beneficioso in vitro de respuesta. Debido a que la versión recombinante de este anticuerpo mostró propiedades similares in vitro nos preguntamos si esto afectaría la actividad motora de ratones con enfermedad desmielinizante inducida por TMEV. Análisis de la función motora se realizó mediante el control de la actividad nocturna espontánea. En primer lugar, se trataron los ratones en el momento de la demielinación máxima y la aparición de la pérdida axonal (90dpi). Ocho semanas después del tratamiento, rHIgM12 sólo mejoró la actividad motora horizontal, mientras que la actividad vertical no se vio afectada. Sin embargo, cuando los ratones fueron tratados anteriormente en la enfermedad (en 45dpi), rHIgM12 mejora la actividad tanto horizontal como vertical. En la enfermedad crónica TMEV inducida, el comportamiento de encabritarse (actividad vertical) se ve afectado más gravemente y parece que la degeneración y pérdida de axones responsables de esta actividad es irreversible. Por el contrario, la fase temprana de la enfermedad, cuando estos axones no se lesionan de manera irreversible, parece ser el momento ideal para el tratamiento. Jones et al. utilizaron modelo de EAE y mediante el estudio de deserción axonal y la función motora propusieron un paradigma idéntico; tratamiento con fármacos neuroprotectores debe comenzar temprano en la enfermedad, incluso antes de la aparición de déficits motores [11]. En segundo lugar, debido a que la mejora funcional ocurre aproximadamente dos semanas después del tratamiento y comienza a desaparecer aproximadamente 25-30 días más tarde, puede ser que los tratamientos repetidos serán necesarios para mantener la función motora. Por desgracia, en nuestros estudios, no fue posible probar múltiples dosis de IgM humanos debido a la respuesta inmune de anticuerpos anti-humano en ratones que se traduce en la anafilaxia. A2B5 es un anticuerpo monoclonal de ratón que también promueve el crecimiento de neuritas [5] y representa un probable candidato para probar única frente a dosis múltiples en el resultado funcional y su duración de acción. Finalmente, ninguno de los tratamientos tuvo un efecto sobre la función motora de los animales no infectados normales. Independientemente del tratamiento, todos los grupos de ratones normales mostraron una disminución gradual de la actividad espontánea, lo que puede ser explicado por la habituación al medio ambiente. Esta disminución de la actividad en animales normales hace aún más impresionante el aumento inducido por rHIgM12 en la actividad de ratones enfermos.
Hemos demostrado una mejora en la función motora en un modelo crónico progresivp de la enfermedad de demielinación inflamatoria en la que ha sido generalmente muy difícil prevenir el desarrollo de déficits neurológicos. Por lo tanto, el anticuerpo monoclonal de unión neurona-rHIgM12 representa un agente muy prometedor terapéutico para el tratamiento no sólo de MS humanos, sino también otros desmielinizante o trastornos neurodegenerativos. Además, debido a que hay ejemplos de ataques de EM clínicamente silenciosos [12, 13], nosotros y otros hemos propuesto que los compuestos neuroprotectores deberían ser complementados con agentes inmunomoduladores [11]. Proponemos que el tratamiento combinado de fármacos inmunomoduladores y rHIgM12 puede resultar en una mejora significativa de la conservación y reparación del SNC después de una lesión axonal.
En conjunto, los resultados de este estudio proporcionan tres conclusiones importantes: 1) la etapa de la enfermedad cuando se administra el tratamiento es crítica (el tratamiento temprano es más beneficioso); 2) para mantener aún más la función motora mejorada, puede haber una necesidad de tratamientos repetidos y 3) rHIgM12 no es tóxico y no afecta a la función motora en animales normales no infectados. Los resultados son consistentes con la hipótesis de que un anticuerpo recombinante que se dirige a las neuronas mejora la función neurológica en un
modelo axonal crónico de desmielinización.
REFERENCIAS
1. McGavern, D.B., et al., Quantitative assessment of neurologic deficits in a chronic progressive murine model of CNS demyelination. Exp Neurol, 1999.158(1): p. 171-81.
2. Warrington, A.E., et al., Human monoclonal antibodies reactive to oligodendrocytes promote remyelination in a model of multiple sclerosis. Proc Natl Acad Sci U S A, 2000. 97(12): p. 6820-5.
3. McGavern, D.B., et al., Axonal loss results in spinal cord atrophy, electrophysiological abnormalities and neurological deficits following demyelination in a chronic inflammatory model of multiple sclerosis. Brain, 2000. 123 Pt 3: p. 519-31.
4. Warrington, A.E., et al., A recombinant human IgM promotes myelin repair after a single, very low dose. J Neurosci Res, 2007. 85(5): p. 967-76.
5. Warrington, A.E., et al., Neuron-binding human monoclonal antibodies support central nervous system neurite extension. J Neuropathol Exp Neurol, 2004. 63(5): p. 461-73.
6. Denic, A., et al., The relevance of animal models in multiple sclerosis research. Pathophysiology, 2010. 18(1): p. 21-9.
7. Hoch, J.C. y A.S. Stern, NMR Data Processing. 1 ed1996, New York: Wiley- Liss. 230.
8. Akaike, H., A new look at the statistical model identification. IEEE Transactions on Automatic Control, 1974.
19(6): p. 716-723.
9. Howe, C.L., J.D. Adelson, and M. Rodriguez, Absence of perforin expression confers axonal protection despite demyelination. Neurobiol Dis, 2007. 25(2): p. 354-9.
10. Rodriguez, M., A function of myelin is to protect axons from subsequent injury: implications for deficits in multiple sclerosis. Brain, 2003. 126(Pt 4): p. 751-2.
11. Jones, M.V., et al., Behavioral and pathological outcomes in MOG 35-55 experimental autoimmune encephalomyelitis. J Neuroimmunol, 2008.199(1-2): p. 83-93.
12. Jacobs, L., P.R. Kinkel, and W.R. Kinkel, Silent brain lesions in patients with isolated idiopathic optic neuritis. A clinical and nuclear magnetic resonance imaging study. Arch Neurol, 1986. 43(5): p.452-5.
13. Pohl, D., et al., Pediatric multiple sclerosis: detection of clinically silent lesions by multimodal evoked potentials. J Pediatr, 2006.149(1): p. 125-7.
14. Mikami, Y., et al., A simple and reliable behavioral analysis of locomotor function after spinal cord injury in mice. Technical note. J Neurosurg, 2002. 97(1 Suppl): p. 142-7.
15. Mikami, Y., et al., Implantation of dendritic cells in injured adult spinal cord results in activation of endogenous neural stem/progenitor cells leading to de novo neurogenesis and functional recovery. J Neurosci Res, 2004.
76(4): p. 453-65.
16. Melnick, S.M. y D.L. Dow-Edwards, Differential behavioral responses to chronic amphetamine in adult male and female rats exposed to postnatal cocaine treatment. Pharmacol Biochem Behav, 2001.69(1-2): p. 219-24. 17. Shen, H., et al., Inosine reduces ischemic brain injury in rats. Stroke, 2005. 36(3): p. 654-9.
18. Torres-Reveron, A. and D.L. Dow-Edwards, Prenatal cocaine dampened behavioral responses to methylphenidate in male and female adolescent rats. Neurotoxicol Teratol, 2006. 28(2): p. 165-72.
19. Zhu, N., J. Weedon, and D.L. Dow-Edwards, Oral methylphenidate improves spatial learning and memory in pre- and periadolescent rats. Behav Neurosci, 2007. 121 (6): p. 1272-9.
20. Li, X., et al., Attenuation of basal and cocaine-enhanced locomotion and nucleus accumbens dopamine in cannabinoid CB1-receptor-knockout mice. Psychopharmacology (Berl), 2009. 204(1): p. 1-11.
21. Rivera-Quinones, C., et al., Absence of neurological deficits following extensive demyelination in a class I-deficient murine model of multiple sclerosis. Nat Med, 1998. 4(2): p. 187-93.
EJEMPLO 20: Neurona-Protective mAbs humano para el tratamiento de la neurona motora enfermedad del ELA
La esclerosis lateral amiotrófica (ELA) es una enfermedad neurológica devastadora que afecta principalmente a células del asta anterior y las neuronas del tracto corticoespinal. La ELA es una enfermedad de la motoneurona caracterizada por la degeneración progresiva de las células de motor en el cerebro y la médula espinal. Las células de motor (neuronas) controlan los músculos que permiten a un individuo moverse, hablar, respirar y tragar. Sin nervios para activarlos, los músculos se debilitan y los residuos de forma gradual. A pesar de una extensa investigación, la etiología de este trastorno es en gran parte desconocida, y no existen tratamientos eficaces. Genes implicados en una minoría de pacientes con una forma genética rara de e La proporcionan una pista potencial para este trastorno. [1] Una mutación del gen identificado ha sido la mutación de So D Cu/Zn (dismutasa de superóxido de cobre/zinc), que está presente en un pequeño porcentaje de los pacientes con la forma genética de la ELA. [2] Es evidente que los pacientes portadores de las mutaciones de SOD tienen resultados neurológicos muy similares en comparación con los pacientes que desarrollan la enfermedad de forma espontánea sin una mutación genética asociada. [3] Esta enzima cataliza superóxido en oxígeno y peróxido de hidrógeno. SOD1 es la forma de la enzima asociada a la ELA. Parece que hay una ganancia de función de SOD1, que afecta el transporte axonal rápido. [4] La frecuencia de la mutación de SOD en ELA familiar varía del 12 al 23% y el gen generalmente se hereda de forma autosómica dominante.
La identificación del gen permitió el desarrollo de modelos animales con características de la enfermedad de ELA para diseñar y probar nuevos medicamentos. Debido a formas heredadas y no heredadas de ELA son enfermedades similares, la causa subyacente puede estar relacionado. Por lo tanto, los fármacos que son eficaces en los modelos de ratón de ELA heredado también pueden trabajar en las personas con las formas aleatorias más comunes de ELA. La disponibilidad de modelos de ratones transgénicos con genes de SOD1 humanos y las características patológicas de la ELA han avanzado el desarrollo de fármacos para la enfermedad. [5] Estos ratones transgénicos desarrollan la pérdida de neuronas motor progresivo y déficits neurológicos. Debido a que formas heredadas y no hereditarias de ELA son similares clínicamente, los defectos subyacentes en la función de las neuronas motoras pueden estar relacionados. Reactivos eficaces en ELA SOD1 ligado también pueden ayudar a las formas esporádicas más prevalentes de ELA. En la actualidad hay solamente un medicamento en el mercado para la ELA, el glutamato fármaco bloqueador Rilutek® (riluzol comprimidos). Sin embargo, los estudios han demostrado que este fármaco no mejora la calidad de vida del paciente y sólo puede prolongar la vida de un promedio de 2 meses. Se necesitan críticamente Claramente fármacos más eficaces.
El laboratorio ha desarrollado nuevas terapias para el tratamiento de desmielinizante y trastornos degenerativos del sistema nervioso central (SNC) [6] y se han identificado una serie de anticuerpos monoclonales humanos (mAbs) que se unen ya sea a oligodendrocitos [7] o neuronas [8], que se ha demostrado para inducir la reparación en el s Nc . Estos anticuerpos se han clonado, secuenciado [9] y producido en grandes cantidades en una fábrica BPM para futuros ensayos clínicos [10]. Anticuerpo humano recombinante IgM12 (rHIgM12) se deriva de un anticuerpo aislado de un paciente con macroglobulinemia de Waldenstrom. [11] Este anticuerpo específicamente etiqueta neuronas y los axones del SNC. [8] A pesar de ser un anticuerpo IgM, IgM12, como se muestra en los Ejemplos anteriores, cruza la barrera hematoencefálica y se une específicamente a los axones y las neuronas dentro del SNC. rHIgM12 in vitro se une a una amplia gama de neuronas, incluyendo neuronas del cerebelo granulares, las neuronas corticales, neuronas del hipocampo y neuronas de células ganglionares de la retina [12]. Los experimentos in vitro demuestran que rHIgM12 protege a las neuronas de la muerte celular. El anticuerpo monoclonal rHIgM12, se administra a pacientes en las primeras etapas de la ELA (tanto las formas genéticas como espontáneas de la enfermedad), puede actuar para proteger a las células del asta anterior y prevenir lesiones axón y de ese modo retrasar la aparición de la discapacidad y mejorar la supervivencia.
RESULTADOS
Hemos realizado experimentos para tratar los ratones con la mutación G86R hSOD1 [13] (SOD1 G86R FVBTg M1Jwg, Jackson Labs) con anticuerpo humano rHIgM12. Ratones G86R parecen normales al nacer. Sin embargo, a partir de aproximadamente 90-100 días de edad, se someten a la pérdida de peso pronunciada, desarrollan atrofia muscular significativa, y mueren de insuficiencia respiratoria en una cuestión de semanas de la pérdida de peso precipitada. ¿LUDOLPH? En un ensayo aleatorizado "cegado", estos ratones recibieron rHIgM12 purificado por g Mp a la edad de 55 días como una sola dosis intraperitoneal (200 pg). En contraste, el grupo de placebo recibió solución salina tamponada con fosfato (PBS). Unos ratones se dejaron sin tratar para determinar el curso natural de la enfermedad sin manipulación humana. Dado que los datos para los ratones tratados con PBS y no tratados eran similares, los grupos se combinaron para el análisis estadístico. Los ratones fueron asignados al azar para recibir tratamiento por un investigador en nuestro laboratorio y se observaron los déficits neurológicos por otro investigador "cegado" hasta que se convirtieron en moribundos. Los investigadores tratantes y otros investigadores que realizan los análisis patológicos no eran conscientes del protocolo de aleatorización hasta que se decifró el código.
Los animales se pesaron semanalmente para determinar si el tratamiento con los anticuerpos monoclonales humanos no sólo aumentaría la supervivencia, sino también retrasaría la aparición de la pérdida de peso. Los resultados fueron sorprendentes y mostraron un aumento estadísticamente significativo en la supervivencia en los animales que recibieron rHIgM12 en comparación con los animales que recibieron PBS. En promedio, los investigadores registraron un aumento de la supervivencia de 25 a 30 días, lo que, por las curvas de Kaplan-Meier (FIGURA 38), mostró significación estadística (P = 0,008). Además, los ratones tratados con rHIgM12 tenían pérdida menos pronunciada de peso (FIGURA 39), un parámetro comúnmente evaluado en la evaluación de la progresión de la enfermedad en los animales. [18] Para nuestro conocimiento, esta es la primera demostración de un anticuerpo monoclonal humano recombinante totalmente dirigido contra las neuronas que son eficaces para aumentar la supervivencia de animales con un fenotipo ELA.
Cuando los animales alcanzaron la fase moribunda, que fueron sacrificados y se perfundieron con fijador de Trump. Las médulas espinales se retiraron y se cortaron en bloques de 1 mm y se embebieron en plástico Araldite para 1 pM secciones gruesas. Estas secciones se examinaron histológicamente. Los animales que se desarrollaron a partir de la mutación ELA SOD mostraron significativa degeneración axonal en los tractos de sustancia blanca, de tal manera que cuando el axón se derrumba, espirales de mielina relativamente fáciles de identificar se desarrollan en la sustancia blanca de la médula espinal. Cuando se cuantificó el número de espirales de la médula espinal (degeneración de los axones) en las secciones torácicas en animales que recibieron rHIgM12 frente a PBS, hubo una disminución estadísticamente significativa en el número de espirales de mielina en animales tratados con terapia
con anticuerpos monoclonales (FIGURA 40).
Secciones de la médula espinal también se tiñeron con un anticuerpo para NeuN, un marcador que específicamente etiqueta neuronas en el SNC y define muy bien las células del asta anterior, así como las neuronas en el asta posterior. Un número de células del asta anterior se cuantificaron a nivel de la médula espinal torácica en animales que recibieron rHIgM12 en comparación con PBS (FIGURA 41, izquierda). Hubo una diferencia altamente estadística con un aumento en el número de células del asta anterior conservadas en los animales que recibieron rHIgM12 frente a PBS. Un análisis similar sobre las neuronas en las astas posteriores también reveló un aumento significativo en las neuronas de los ratones tratados con rHIgM12 (FIGURA 41, derecha).
El anticuerpo humano, rHIgM12, se une a la superficie de muchos tipos de neuronas del SNC obtenidos a partir de muchas especies, incluyendo ratones, seres humanos (FIGURA 42), conejos y primates. Esto proporciona evidencia de que la señalización de las neuronas rHIgM12 se conserva en mamíferos, incluidos los de los ratones a los seres humanos. Nuestros estudios en los ratones SOD sugieren que la protección de las células anterior y cuerno posterior de la muerte puede dar lugar a un menor número de axones en degeneración de la médula espinal y el aumento de la supervivencia del animal. Los experimentos descritos anteriormente en los Ejemplos anteriores con las neuronas corticales en cultivo demuestran que el tratamiento de neuronas con rHIgM12 puede protegerlos de la muerte celular (véase Figura 3). Las neuronas corticales cultivadas a partir de ratones neonatales fueron expuestas a concentraciones muy altas de peróxido de hidrógeno para determinar una concentración donde el 90% de las neuronas moriría. Neuronas expuestas a peróxido de hidrógeno fueron tratadas al mismo tiempo con rHIgM12 u otro IgM humano que no se une a las neuronas. El tratamiento con rHIgM12 resultó en la preservación de 80% de las neuronas. En contraste, el tratamiento con un control de IgM resultó en la muerte de 90% de las neuronas después de la exposición a peróxido de hidrógeno. Nuestra hipótesis es que rHIgM12 extiende la vida útil de los animales con ELA mediante la protección de las neuronas de la muerte celular.
Los anticuerpos humanos también se ensayaron como sustratos en placas de cultivo tisular, y se compararon para la capacidad para promover la extensión normal del proceso de la célula de las neuronas del cerebelo o corticales. rHIgM12 y sHlgM42, los anticuerpos que se unen a la superficie de las neuronas, y rHIgM22 y sHIgM39, los anticuerpos que no se unen a las neuronas, se compararon con el potente proceso de apoyo molécula de la matriz extracelular, laminina. El cultivo de neuronas sobre un sustrato de anticuerpos humanos rHIgM12 o sHIgM42 resultaron en una extensión dramática del crecimiento de la neurona, similar a la observada con laminina [8]. Este estudio también mostró que las neuronas se comportan normalmente en presencia de rHIgM12.
Se ha aceptado generalmente como dogma que las IgM con un peso molecular de cerca de 1 millón son demasiado grandes para cruzar la barrera hematoencefálica (BBB) de la circulación para entrar en el SNC [15] [16]. Sin embargo, existe cada vez más evidencia que algunos anticuerpos no cruzan la BBB. Anteriormente hemos descrito la detección de rHIgM12 dentro de la médula espinal después de la inyección periférica. 1,0 mg de rHIgM12 o un control de IgM humana se administraron por vía intraperitoneal a ratones con lesiones de la médula espinal demielinadas. Cuatro horas más tarde los ratones se sacrificaron y las secciones de la médula espinal se inmunotiñeron para detectar la presencia de la cadena humana mu IgM y anti-neurofilamentos. En los ratones que recibieron microscopía confocal rHIgM12 cadena mu humana demostrada dentro de las lesiones desmielinizadas co localizadas con antineurofilamento, un marcador de axones, en secciones paralelas y, como haces de corte en extremo (véase la Figura 15). IgM humana no se encontró dentro de las lesiones de la médula espinal de los ratones que recibieron un control de IgM.
Si bien los estudios proporcionados son en modelos animales, hay una serie de aspectos tentadores de estos resultados de investigación que indican además que este nuevo enfoque será eficaz en pacientes humanos. Es importante destacar que el anticuerpo IgM12 descrito y utilizado en el presente documento es un anticuerpo completamente humano monoclonal. Como resultado, sólo una dosis única de este anticuerpo se puede utilizar en ratones. Las dosis posteriores causan que los animales desarrollan una respuesta inmune al anticuerpo resultante, ya sea en la neutralización de rHIgM12 o la muerte del animal debido a la anafilaxia. Sin embargo, ya que estos son "verdaderos" anticuerpos humanos, pacientes humanos tratados con rHIgM12 es poco probable el desarrollo de una respuesta inmune a ellos. Además, rHIgM12 potencialmente puede darse de forma intermitente continua a los pacientes sin efecto perjudicial o desarrollo de una reacción de bloqueo de anticuerpos. Debido a que rHIgM12 es un autoanticuerpo humano natural, son poco probables efectos secundarios adversos, y por lo tanto, esperamos que la toxicidad de este agente sea mínima. Hay mucha evidencia de que rHIgM12 estará a salvo en los seres humanos. Antes de estos resultados positivos en el modelo ELA la forma de suero del anticuerpo, sHIgM12, fue probado en varios modelos de enfermedad neurológica sin toxicidad, como se describió anteriormente en el presente documento. En estos estudios hemos encontrado, 1) ningún aumento en la patología del SNC en ratones con virus mediada por desmielinización después de una dosis de 1 mg, 2) ningún aumento en la gravedad de las puntuaciones clínicas en ratones con EAE activa después de una dosis de 200 pg y 3) la sangre limpia química y el tejido patología en ratones CD-1 normales después de una dosis de 300 pg. Aun así, es notable que una sola dosis a un ratón en un modelo animal reconocido de ELA dio resultados notables.
rHIgM12 ya se ha cultivado en una instalación de GMP de acuerdo con directrices de la FDA, y, por
directrices de la FDA, líneas celulares transfectadas estables se han generado y almacenado sin infecciones adventicias. Estas líneas celulares han sido probadas contra un panel de> 50 organismos infecciosos, todos con resultados negativos. Además, las líneas celulares hacen grandes cantidades de anticuerpo en cultivo de tejidos (150 |jg/ml). Una metodología para purificar rHIgM12 a> 97% de pureza sin virus adventicios, ADN, ARN u otro material extraño de acuerdo con directrices de la FDA se ha establecido, proporcionando una fuerte base preclínica para rHIgM12 en ensayo clínico de fase inicial futuro en pacientes con las etapas tempranas de ELA.
ESTUDIOS EN CURSO
Después de haber demostrado anteriormente que el anticuerpo monoclonal humano recombinante (rHIgM12) puede prolongar la supervivencia en un modelo transgénico de e La humana mediante la prevención de la pérdida de células del asta anterior y axones, se están realizando estudios para generar datos pre-clínicos a través de la farmacocinética y toxicología para avanzar hacia una fase I de ensayos clínicos en pacientes con ELA.
Los estudios cegados versus placebo Ab: un estudio definitivo "cegado", controlado con placebo se realiza en dos cepas de ratones SOD con el fenotipo ELA (SOD1 G86R y G93A), probando anticuerpo humano recombinante rHIgM12 frente a un anticuerpo humano de control de isotipo, sHIgM39. La eficacia de una sola dosis 200 jg de anticuerpo humano recombinante, rHIgM12, se prueba para reducir la enfermedad, tanto en la G86R como G93A SOD1 [14] (B6 CG-Tg SOD1 G93A 1Gur, Jackson Labs) modelos de ratones transgénicos mutantes, en comparación con una dosis de 200 jg de un anticuerpo humano de control de isotipo, sHIgM39. El tratamiento con anticuerpo es a los 55 días de edad tras recomendación ENMC de tratamiento de pre-aparición [18], con resultados para reflejar los estudios piloto que comparan rHIgM12 a PBS (Figuras 38-41). Principales criterios de valoración son la supervivencia y la prevención de la pérdida de peso. Cada grupo experimental consta de 24 ratones, lo cual es suficiente para detectar una diferencia en base a los datos anteriores. Además, el SNC se examinó en todos los ratones después del sacrificio para determinar el número de células del asta anterior de la médula espinal torácica media usando una etiqueta para NeuN. Se cuenta el número de axones degeneradores indicados por verticilos de mielina anormales. El modelo mutante G93A SOD1 (B6.Cg-Tg) SOD1G93A) 1Gur/J # 004435, Jackson Laboratory) se incluye porque este es el primer modelo genéticamente basado, más ampliamente utilizado y mejor caracterizado de ELA y permite resultados con rHIgM12 que deben compararse a una base de datos amplia de los paradigmas de tratamiento. Ratones G86R tienen un inicio tardío de la enfermedad (7 meses) y una progresión rápida; mientras que los ratones G93A tienen un inicio más rápido (3-4 meses) con una progresión más lenta. Este estudio controla para la introducción de una proteína extraña en ratones SOD1 y también se ocupa de conceptos de mecanismo de acción. El carácter vinculante de las neuronas de rHIgM12 es crítica y una IgM que no se une a las neuronas no debería mejorar la enfermedad. El punto final primario en este estudio incrementa la supervivencia (10% o mayor P <0,05) y la disminución de la pérdida de peso (10% o mayor P <0,05). Mejoras en al menos una cepa de ratones SOD1 se considera un éxito. Todos los ratones se sacrifican en el momento que son incapaces de enderezarse dentro de 15-30 segundos si se colocan a cada lado. Los puntos finales secundarios miden el número de axones en degeneración, como se indica por verticilos de mielina anormales, y la densidad de células del asta anterior positivo NeuN a nivel torácico (T6) de la médula espinal. Los eventos adversos se consideran si los ratones tratados con rHIgM12, 1) pierden 20% más de peso corporal en comparación con los controles, 2) desarrollan convulsiones, 3) tienen una tasa de mortalidad de 20% mayor que los controles.
Estudio de titulación de dosis: Tras los resultados positivos en el estudio anterior, un estudio de titulación de dosis (una dosis única de 0, 5, 50, 100 y 250 jg se emprende por ratón dado a los 55 días de edad) para determinar la dosis mínima necesaria para proteger a los ratones SOD1 de la muerte. Espectroscopía de MR del tronco cerebral de ratón se usa para medir aspartato N-acetilo (NAA). Hemos demostrado en otros modelos de enfermedad neurológica que NAA en el tronco cerebral es un excelente indicador sustituto de la salud a lo largo del axón de la médula espinal. [17] Los datos de espectroscopia por RM ayudan a validar NAA como un marcador sustituto de la eficacia del anticuerpo en los ensayos en humanos potenciales utilizando rHIgM12. Una sola dosis de 0, 5, 50, 100 y 250 jg se administra intraperitonealmente a los 55 días de edad para grupos de 20-24 ratones SOD1. Los puntos finales primarios y secundarios y los eventos adversos son los mismos como se describió anteriormente con la adición de la espectroscopia de MR en el tronco cerebral para medir Nacetilo aspartato (NAA). Espectroscopía de MR se recoge para cada ratón en el día de tratamiento con el anticuerpo, a los 100 días de edad e inmediatamente antes del sacrificio. Un aumento del 10% (P <0,05) en la concentración de NAA de tallo cerebral en cualquier grupo tratado rHIgM12 en 100 días o ratones tratados con solución salina comparados de última exploración se considera una mejora.
Estudios farmacocinéticos y BBB: Los estudios se realizaron en ratones SOD1 G86R y G93A a 50-70 días de edad [18] después de solo una dosis intravenosa de 200 jg, usando un sistema de detección ELISA establecido que detecta específicamente IgM humana dentro del fondo de la inmunoglobulina sangre de ratón normal. IgM humano se mide en la sangre en diversos puntos temporales después de la dosificación (0, 15 min, 30 min, 1 h, 4 h, 8 h, 12 h, 18 h, 24 h, 2 días, 3 días, 5 días, y 7 días). Debido a que el volumen de sangre de los ratones es pequeño (en total <1,5 ml), tres ratones individuales se utilizan para cada punto de recogida.
Se propone rHIgM12 para actuar directamente sobre el sistema nervioso, cruzar la barrera
hematoencefálica después de la inyección periférica de interactuar con las neuronas para protegerlas de la muerte. Es posible, aunque improbable, que el anticuerpo humano no cruza la barrera hematoencefálica, pero estimula aspectos de una respuesta inmune, que luego proporciona protección de axón. Seguimiento in vivo de rHIgM1235S marcada se utiliza para abordar plenamente esta pregunta. Dos niveles de dosis de 35S etiquetados rHIgM12, 250 pg y la dosis eficaz mínima establecida anteriormente, son seguidos en ratones SOD1 en 50-70 días de edad. Después de una dosis intravenosa de 35S marcado rHIgM12, el porcentaje de 35S que cruza la barrera sangrecerebro y se encuentra en el cerebro/parénquima de la médula espinal a 4, 8, 24, 48 y 72 h después de determinarse la inyección. Además, el sitio de la localización de 35S en el cerebro/médula espinal se determina usando técnicas de autorradiografía ya publicadas. [19]
Estudios de vida media en suero: La cinética de sangre para rHIgM12 se realiza usando una dosis de 200 pg en la cepa SOD1 en la que rHIgM12 demostró una eficacia. El estudio se lleva a cabo en grupos de 3 ratones SOD1 por punto de tiempo; 13 puntos de recogida de más de 7 días después de la inyección intravenosa. Ratones SOD1 son tratados en 50-70 días de edad para entender la cinética de anticuerpos en el momento de tratamiento. suero rHIgM12 media vida, la saturación de la exposición y la presencia de neutralizar Abs IgM será determinada. Los estudios de seguimiento de rHIgM12 marcado isotópicamente (35S) requiere la determinación de la dosis eficaz mínima descrita anteriormente. A ratones SOD1 se les da una sola dosis intravenosa de 250 pg y la dosis eficaz mínima de rHIgM12 (que contiene al menos 1x107 cpm) [19] a 50-70 días de edad para abordar si rHIgM12 entra en el SNC en el momento del tratamiento terapéutico. A las 4, 8, 24, 48 y 72 h después de la inyección principales tejidos se eliminan de forma aguda, incluyendo cerebro, médula espinal, hígado, corazón, pulmón, estómago, intestino, músculo, bazo, hígado, páncreas. Una porción de 150 mg de tejido se extrae, se pesó, se solubilizó en soluble (Perkin Elmer) y cpm determinado en el líquido de centelleo (Ultima Gold, Perkin Elmer). rHIgM12 marcada isotópicamente está localizada en las secciones de la médula espinal usando autorradiografía [19]. En este experimento, el punto final primario es una acumulación de isótopo 35S en el cable de cerebro o la médula de los ratones a las 4, 8, 24, 48 y 72 h después de la inyección de rHIgM12 en comparación con los controles de punto de tiempo cero. Un aumento del 50% (P <0,05) en 35S recuento/mg en todo el cerebro o la médula espinal cualquier punto de tiempo se considera significativa y evidencia que rHIgM12 puede entrar en el SNC. Para estudios de autorradiografía, los ratones se dosifican con la etiqueta rHIgM12 y médulas espinales eliminadas en un momento en el que rHIgM12 se aumentó en el SNC. Un aumento del 50% en granos/mm2 de plata sobre las neuronas en las secciones de la médula espinal se considera evidencia significativa de anticuerpo dirigido in vivo.
Estudios de toxicidad iniciales: Los experimentos se llevaron a cabo con rHIgM12 para determinar si el anticuerpo es tóxico en ratones normales CD-1 y conejos. Grupos de 10 ratones normales de CD-1 de ambos sexos se administran 1 vez (1x), 10 veces (10x) y 20 veces (20x) la dosis eficaz mínima de rHIgM12 determinada anteriormente o solución salina por vía intravenosa una vez o diariamente durante 7 días. Dos semanas más tarde sangre y los tejidos son recogidos por una necropsia "total" y analizada. Secciones de tejidos de todos los órganos principales se examinan "cegadas" por un patólogo veterinario con experiencia en evaluaciones toxicológicas. La sangre se caracteriza en la química y hematología para el panel habitual de visualizaciones de toxicología que incluyen análisis de sangre para las enzimas hepáticas, enzimas cardíacas, electrolitos y efectos en grupo de hematología. Estudios de exposición idénticos se realizan en una especie no roedora; dos conejos de cada sexo se ensayan en cada dosis. Además, se utilizan tejidos de estudios de reactividad cruzada utilizando rHIgM12 para determinar si el anticuerpo se une a cualquier otro tejido u órganos. Un panel de secciones de tejido embebidas en parafina de ratón, conejo, primate y humano (Zimad) se immunoetiquetaron con rHIgM12 o un control de IgM humano. La intensidad y estructuras ligadas dentro de cada tejido se forman imágenes y se compararon con rHIgM12 y etiquetado IgM de control de cortes de tejido vivo cortados con un cortador de tejidos [7] ya que esto imita más estrechamente la situación in vivo. Además, la unión de rHIgM12 y un anticuerpo de control se ensaya en humano congelado y el tejido de primate como consistente con la sección de toxicología preliminar de una aplicación de fármaco nuevo investigado de la FDA. Estudios de reactividad cruzada de tejido proporcionan pistas en cuanto a qué órganos deben ser controlados en particular durante los estudios de exposición en ratones y conejos.
Unión de tejido: Después de la determinación de eficacia, estos estudios abarcan la unión a tejidos diana y no diana. Matrices comercialmente disponibles de tejido (Zimad) y secciones de mono cynomolgus y humano (Charles River) se immunoetiquetaron con 10 pg/ml de rHIgM12 y controlan IgM humana, sHIgM39. La intensidad de la etiqueta se compara mediante el uso de imágenes digitales. la unión a matrices de tejido fijado embebido en parafina y la médula espinal humana congelada y el cerebro de los primates y rHIgM12 se compara con unión observada de anticuerpo en rebanadas cerebelares de ratón vivo. La unión del anticuerpo a rebanadas cerebelosas vivas, un ensayo de potencia estándar, que hemos utilizado y de la primera visualización en la que se identificó la forma de suero de rHIgM12.
REFERENCIAS CITADAS
Gurney 1. Gurney, M.E., et al., Mutant CuZn superoxide dismutase in motor neuron disease. J Inherit Metab Dis, 1998. 21(5): p. 587-97.
2. Morrison, B.M. y J.H. Morrison, Amyotrophic lateral sclerosis associated with mutations in superoxide dis mutase: a putative mechanism of degeneration. Brain Res Rev, 1999. 29(1): p. 121-35.
3. Siddique, T., D. Nijhawan, and A. Hentati, Familial amyotrophic lateral sclerosis. J Neural Transm Suppl, 1997.
49: p. 219-33.
4. Cluskey, S. y D.B. Ramsden, Mechanisms of neurodegeneration in amyotrophic lateral sclerosis. Mol Pathol, 2001. 54(6): p. 386-92.
5. Turner, B.J. y K. Talbot, Transgenics, toxicity and therapeutics in rodent models of mutant SOD1-mediated familial ALS. Prog Neurobiol, 2008. 85(1): p. 94-134.
6. Bieber, A.J., et al., Humoral autoimmunity as a mediator of CNS repair. Trends Neurosci, 2001. 24(11 Suppl): p. S39-44.
7. Warrington, A.E., et al., Human monoclonal antibodies reactive to oligodendrocytes promote remyelination in a model of multiple sclerosis. Proc Natl Acad Sci U S A, 2000. 97(12): p. 6820-5.
8. Warrington, A.E., et al., Neuron-binding human monoclonal antibodies support central nervous system neurite extension. J Neuropathol Exp Neurol, 2004. 63(5): p. 461-73.
9. Ciric, B., et al., Clonal evolution in Waldenstrom macroglobulinemia highlights functional role of B-cell receptor. Blood, 2001. 97(1): p. 321-3.
10. Mitsunaga, Y., et al., Direct evidence that a human antibody derived from patient serum can promote myelin repair in a mouse model of chronic-progressive demyelinating disease. Faseb J, 2002. 16(10): p. 1325-7.
11. Rodriguez, M., A.E. Warrington, and L.R. Pease, Invited article: human natural autoantibodies in the treatment of neurologic disease. Neurology, 2009. 72(14): p. 1269-76.
12. Wright, B. R., et al., Cellular mechanisms of central nervous system repair by natural autoreactive monoclonal antibodies. Arch Neurol, 2009. 66(12): p. 1456-9.
13. Bruijn, L.I., et al., ALS-linked SOD1 mutant G85R mediates damage to astrocytes and promotes rapidly progressive disease with SOD1-containing inclusions. Neuron, 1997.18(2): p. 327-38.
14. Gurney, M.E., et al., Motor neuron degeneration in mice that express a human Cu, Zn superoxide dismutase mutation. Science, 1994. 264(5166): p. 1772-5.
15. Banks, W.A., Developing drugs that can cross the blood-brain barrier: applications to Alzheimer's disease. BMC Neurosci, 2008. 9 Suppl 3: p. S2.
16. Kozlowski, G.P., I. Sterzl, and G. Nilaver, Localization patterns for immunoglobulins and albumins in the brain suggest diverse mechanisms for their transport across the blood-brain barrier (BBB). Prog Brain Res, 1992. 91: p.
149-54.
17. Denic, A., et al., Brainstem 1H nuclear magnetic resonance (NMR) spectroscopy: marker of demyelination and repair in spinal cord. Ann Neurol, 2009. 66(4): p. 559-64.
18. Ludolph, A.C., et al., Guidelines for the preclinical in vivo evaluation of pharmacological active drugs for ALS/MND: report on the 142nd ENMC international workshop. Amyotroph Lateral Scler, 2007. 8(4): p. 217-23.
19. Hunter, S.F., D.J. Miller, and M. Rodriguez, Monoclonal remyelination-promoting natural autoantibody SCH 94.03: pharmacokinetics and in vivo targets within demyelinated spinal cord in a mouse model of multiple sclerosis. J Neurol Sci, 1997.150(2): p. 103-13.
EJEMPLO 21: Tratamiento de la isquemia cerebral y accidente cerebrovascular con mAbs humano protector de neurona
La incidencia de accidente cerebrovascular isquémico se aumenta con la edad. El envejecimiento de la población se traduce en un creciente número de golpes por año, produciendo una carga acumulada en nuestra infraestructura de atención de la salud y las familias. El accidente cerebrovascular es la principal causa de discapacidad a largo plazo en los Estados Unidos. Cada año, cerca de 800.000 personas sufren un accidente cerebrovascular. Después de una lesión isquémica, la discapacidad asociada con déficits clínicos se debe principalmente a la disfunción de las neuronas. La falta de recuperación funcional, en parte, está atribuida a la restricción de la regeneración y la neuroplasticidad (1). Actualmente no hay tratamientos eficaces para prevenir o revertir los déficits neurológicos relacionados con el accidente cerebrovascular.
Los anticuerpos de clase humano de origen natural monoclonales IgM se han identificado y utilizado para promover la protección y reparación del SNC, incluyendo la reparación de la mielina y recuperación axonal/neuronal (2). Se identificó un anticuerpo monoclonal humano derivado del suero IgM que se une a la superficie de las neuronas. La IgM se une a las neuronas a través de especies, incluidas los humanos, y promueve la extensión de neuritas. Este IgM protege a las neuronas de la apoptosis in vitro y preserva los axones de la médula espinal in vivo como se mide por espectroscopia de resonancia magnética y la histología de la médula espinal. Una forma recombinante, rHIgM12 ha sido sintetizado que conserva las características de la forma derivada del suero. Los neuroprotectores propiedades de rHIgM12 son similares a los observados con el gangliósido GM1. Los gangliósidos pueden ser un/el antígeno de anticuerpo IgM12 y rHIgM12. rHIgM12 se co-localiza en la membrana neuronal con GM1, el colesterol y la integrina p 1. GM1 se enriquece en microdominios de membrana, que funcionan para agrupar receptores de células y moléculas de señalización. rHIgM12 parece imitar los efectos pleiotrópicos de GM1 por la agrupación de dominios de la membrana de la neurona.
Se llevaron a cabo los presentes estudios para evaluar la eficacia de rHIgM12 para revertir los déficits neurológicos después de la lesión isquémica cerebral. Estos estudios están dirigidos a las aplicaciones de los anticuerpos en pacientes con estadios tempranos de la lesión isquémica cerebral o que sufren de un derrame
cerebral. La hipótesis central es que el anticuerpo IGM12 y los efectos neuroprotectores mediados por rHIgM12 son aplicables en la terapia para mejorar la restauración celular y tisular y para promover la recuperación global a partir del accidente cerebrovascular isquémico.
El manejo terapéutico de isquemia cerebral se basa en la prevención y la reducción de factores de riesgo, como la presión sanguínea elevada, trombosis y la aterosclerosis de la arteria carótida. La terapia de intervención de la isquemia cerebral sigue siendo limitada a la trombolisis enzimática mediante el uso de recombinante activador del rtPA plasminógeno tisular (3-5), y para inter eliminación convencional del coágulo de sangre por cateterización directa (6). Sin embargo, rtPA tiene una corta ventana terapéutica de 3 h y aumento de la incidencia de la isquemia cerebral hemorrágica si se infunde más allá del marco de tiempo de 3 h (7, 8). La protección de las neuronas dañadas después de la isquemia cerebral es una estrategia prometedora para limitar la discapacidad permanente. Mientras que las terapias que modulan la respuesta inflamatoria pueden tener propiedades neuroprotectoras, en la actualidad hay muy pocos fármacos que actúan directamente sobre las neuronas para protegerlas. Neuroprotección puede lograrse mediante la modulación de ambiente inflamatorio neurotóxico en la región isquémica. Estudios anteriores utilizando anticuerpos para tratar la isquemia cerebral no han dado un beneficio potencial: Se emplearon los antagonistas a moléculas de adhesión intercelular en un gran ensayo multicéntrico, los sujetos tratados tenían mayores tasas de mortalidad y los resultados más pobres en comparación con el grupo placebo (9). El uso de anticuerpos anti-plaquetas en un ensayo clínico de fase III en la isquemia cerebral llevó a hemorragia fatal en una pequeña cohorte de pacientes (10). El uso de anticuerpos policlonales IgG humana (IVIg) resultó en una mejora de un modelo animal de isquemia cerebral (11), sin embargo los ensayos clínicos informaron de complicaciones tromboembólicas después de la administración de IgIV, que conduce a una opinión mixta sobre el uso de IgIV para tratar la isquemia cerebral (12, 13). Un objetivo ha sido el de identificar reactivos terapéuticos que se dirigen directamente al sistema nervioso. La forma recombinante de un anticuerpo completamente humano rHIgM12 promueve la supervivencia neuronal y puede tener uso aplicable como agente terapéutico restaurador/neuroprotector. Los experimentos en este ejemplo se centran en los efectos terapéuticos de anticuerpo IgM12, rHIgM12, en un modelo murino de isquemia cerebral. El modelo utilizado es un modelo foto-trombótico no invasivo utilizando colorante foto-sensible y por lo tanto más fisiológico que los modelos invasivos.
Nuestro enfoque para proteger a las neuronas y mejorar el comportamiento en los animales después de la isquemia cerebral utilizando un anticuerpo monoclonal totalmente humano es a la vez original e innovador y el anticuerpo tiene el potencial como el primer fármaco diseñado para atacar y proteger las neuronas en la isquemia cerebral y accidente cerebrovascular. Aspectos relevantes de la molécula se enumeran a continuación:
1) rHIgM12 se une a la superficie de las neuronas (véase Figura 1), y protege a las neuronas de la muerte celular apoptótica (véase Figura 3).
2) rHIgM12 es un autoanticuerpo natural del ser humano y no son probables efectos adversos.
3) rHIgM12, a pesar de ser una IgM cruza la barrera hematoencefálica (BBB) en ratones SJL normales y en animales con inflamación (véase la Figura 13).
4) Los presentes estudios ahora demuestran que cuando rHIgM12 se administra a los animales después de la inducción del ictus isquémico, que se traduce en una clara mejora de la función de los animales (FIGURA 44) y protege la integridad del tejido (FIGURA 45).
RESULTADOS Y DISCUSIÓN
Un enfoque único se utilizó para desarrollar nuevas terapias para el tratamiento de la lesión en el sistema nervioso central (SNC) (14, 15). Una serie de anticuerpos monoclonales humanos han sido identificados que se unen ya sea a los oligodendrocitos (16) o de las neuronas (17) y que tienen aplicación en el SNC. Anticuerpo humano recombinante IgM12 (rHIgM12) se deriva de un anticuerpo aislado de un paciente con macroglobulinemia de Waldenstrom. Este anticuerpo específicamente etiqueta neuronas y los axones del SNC (17). A pesar de ser un anticuerpo IgM, que cruza la barrera hematoencefálica (BBB) (véase Figura 13) y se une específicamente a los axones y las neuronas dentro del SNC. rHIgM12 in vitro se une a una amplia gama de neuronas, incluyendo neuronas del cerebelo granulares, las neuronas corticales, neuronas del hipocampo y neuronas de células ganglionares de la retina. Nuestra hipótesis actual sobre el mecanismo de acción de los anticuerpos IgM reparativos es que las moléculas de entrecruzamiento en microdominios para estimular los eventos que se implican en la protección y reparación de señalización.
Normalización de un modelo de ratón de isquemia cerebral
Tinte fotosensible (Rose Bengal, RB) se utilizó en un modelo foto-trombótico colorante mediado de accidente cerebrovascular isquémico (isquemia cerebral foto-trombótica) en nuestro laboratorio. En 1985, Watson et al. introdujeron PT del cerebro como un modelo simple de la isquemia cerebral focal (Watson BD, et al (1985) Ann Neurol 17: 497-504). Para lograr la trombosis, un tinte fotosensible se inyecta por vía sistémica. La iluminación posterior a través del cráneo intacto conduce a la activación local del colorante con la formación de radicales libres y fotoperoxidación del endotelio. La formación del coágulo está mediada por lo tanto en los vasos iluminados y la oclusión de los vasos imita la isquemia cerebral (Dietrich WD, et al (1987) Acta Neuropathol 72: 315-325). Las
lesiones PT provocan una reacción inflamatoria fuerte, pero retrasada (Schroeter M, et al (1997) Stroke 28: 382 386). Este modelo tiene la ventaja de que la región de la isquemia puede predefinirse y altamente circunscribirse, abriendo la posibilidad para coagular áreas corticales distintas con precisión estereotáctica.
RB, se disolvió en PBS a 10 mg/ml se inyectó a través de la vena de la cola (0,02 mg/g de peso corporal). La abertura (5 mm) de la articulación de fibra óptica unida a una fuente de luz (lámpara halógena de 150 W) se colocó en una ubicación estándar en la cabeza del ratón. Un minuto después de la inyección de RB, los cerebros fueron expuestos a luz constante (para activar RB) a través del cráneo intacto durante 30 segundos. Tras la activación, RB genera radicales libres que perturban las células endoteliales dentro de la microvasculatura, lo que resulta en la agregación plaquetaria y desencadena la coagulación que conduce a la oclusión de vasos sanguíneos pequeños y causas de isquemia cerebral trombótica. Se realizó todos los experimentos en ratones hembra de 6 semanas de edad CD-1®.
En un ensayo aleatorio "cegado", los ratones recibieron rHIgM12 purificado por GMP treinta minutos después de la inducción de la isquemia cerebral (un intento de imitar la situación clínica humana), como una sola dosis intraperitoneal (ip 200 pg en 500 pl). En contraste, el grupo de placebo recibió solución salina tamponada con fosfato (PBS). Estandarizamos técnicas para visualizar y cuantificar región isquémica después de la isquemia cerebral (Figuras 43 y 45)
rHIgM12 mejora la actividad funcional en los animales después de la isquemia cerebral sin diferencias significativas en volumen del infarto definido por (DWI) de imágenes ponderadas por difusión
Déficits funcionales se calcularon utilizando un aparato de campo abierto Accuscan. El aparato consiste en jaulas de acrílico individuales con fotocélulas que miden los movimientos horizontales y verticales discretos. Grupos de 5 ratones CD-1 se colocaron en cada jaula y los valores de línea de base se registraron durante 1 semana antes de la inducción de isquemia cerebral. Los datos se recogieron como número de roturas de haz por bloques de 1 hora y se expresó como% de cambio (FIGURA 44A) vertical o horizontal (FIGURA 44B) desde la línea base. En el día uno, después de la inducción de isquemia cerebral, había déficits similares en la actividad locomotora de ambos grupos de ratones. Comparaciones por pares directos de dos tratamientos mostraron que la mejora en la función motora horizontal en los ratones tratados con rHIgM12 divergieron significativamente (p <0,05) en el día 3 post tratamiento en comparación con PBS (FIGURA 44A). Del mismo modo, la función motora vertical en los ratones tratados con rHIgM12 divergieron significativamente en el día 3 post-tratamiento en comparación con PBS (FIGURA 44B). Mejora persistió hasta por 30 días, cuando se terminó el experimento.
DWI-MRI se utilizó para definir el volumen isquémico y para correlacionar la mejora funcional a la reducción en el tamaño del infarto. Los ratones fueron escaneados en los días 1, 3, 7 y 21 en un sistema animal pequeño MRI de Bruker Avance 300 MHz (7 Tesla) y se obtuvieron imágenes en tres dimensiones (FIGURA 43A) se cuantificaron utilizando NIH software Image J. dW i no mostró significación estadística entre los dos grupos de ratones en cualquiera de los días escaneados (FIGURA 44C), una indicación de que los volúmenes isquémicos cerebrales en ambos grupos de ratones fueron similares. Por lo que sabemos, esto proporciona la primera demostración de que un anticuerpo IgM monoclonal humano recombinante totalmente natural dirigido contra las neuronas es eficaz para mejorar el resultado del comportamiento en los animales después de la isquemia cerebral experimental.
Tratamiento de rHIgM12 mejora las métricas MT-MRI y demuestra diferencias significativas en las proporciones de MT
Formación de imágenes DWI estima la presencia y el grado de edema citotóxico sobre la difusión del agua tras el accidente cerebrovascular isquémico. Con el fin de evaluar las medidas de MRI más sensibles a la integridad del tejido y la recuperación después de la lesión, se utilizó la transferencia de magnetización MRI (MT-MRI) en un experimento posterior, en 10 ratones en el día 3 después de la inducción de apoplejía y después del tratamiento con rHIgM12 o tratamiento simulado de PBS. Las imágenes obtenidas fueron importadas en FSL (23, 24) y un mapa de MTR paramétrico 3D se calculó en FSL MATHS. Se utilizó la fórmula estándar de 100 * ((Mo-Mt)/Mo), donde Mo es la imagen 3D sin necesidad de, y Mt es la imagen 3D con el pulso MT. Las imágenes se escalan apropiadamente para proporcionar una salida de mapeo de percentil de escala de Gray con valores que van de 0 a 100 (FIGURA 45A). Valores lesionales MT se analizaron usando las herramientas estándar 3D de ROI en Analyze 11,0, lo que permitió el análisis de daño de tejidos sensibles MT en las zonas de isquemia. Un ROI comparativo en el hemisferio contralateral fue utilizado para proporcionar los valores de control para la comparación. Valores de MTR en el lado de control fueron (59,9 0,9) y (58,3 0,8) en grupos rHIgM12 y p Bs , tratados respectivamente: como se esperaba, el análisis estadístico no reveló una diferencia significativa entre los dos grupos en los hemisferios de control. Curiosamente, el tratamiento rHIgM12 resultó en un aumento relativo en los valores de MTR en la carrera de sub volumen en comparación con el tratamiento simulado con PBS (46,0 0,9 vs 41,9 08, p = 0,01). Esto mostró relativa normalización de la arquitectura del tejido detectable MT-MRI en el grupo tratado con anticuerpo. Además, aunque no se alcanzó significación estadística (p = 0,39), una disminución del volumen sistólico fue visto en ratones tratados con rHIgM12 vs controles tratados con PBS (18,1 5,9 mm3 vs 26,1 6,6 mm3). La diferencia altamente significativa en MT-MRI apoya que los cambios de anticuerpos inducidos resultan en la restauración de tejido
mejorado en oposición a una reducción directa en los volúmenes de accidente cerebrovascular.
rHIgM12 se une a neuronas de ratón y humanas y protegen a las neuronas corticales de ratón de la muerte de daño oxidativo inducido
Se supone que rHIgM12 protege a las neuronas en la región isquémica mediante la unión a las neuronas. El anticuerpo humano, rHIgM12, se une a la superficie de muchos tipos de neuronas del SNC obtenidos a partir de ratones y seres humanos (véase la Figura 1). Esto apoya que neuro-protección mediada por rHIgM12 y mejora funcional se conservarán a partir de ratones a humanos. Si rHIgM12 protegería a las neuronas in vitro de la activación inducida por peróxido de la caspasa-3, un marcador de apoptosis activo se ha evaluado previamente. Los cultivos de neuronas corticales de ratón primario se trataron simultáneamente con rHIgM12 o controlar IgM humana y peróxido de hidrógeno. El grado de activación de la caspasa-3 se determinó 24 horas más tarde, utilizando un kit de ensayo caspasa-3 FLICA (Immunochemistry Technologies) (véase Figura 3). El tratamiento de las neuronas lesionadas con rHIgM12 reduce la activación de caspasa-3 en un 80%. Las neuronas lesionadas tratadas con control de IgM mostraron menos de un 10% de reducción en la activación de la caspasa-3.
En un ensayo de neurotóxico diferente, las neuronas corticales cultivadas fueron sometidas directamente a condiciones de hipoxia (2,7% O2 , 92,3% N2 , 5% de atmósfera CO2 durante 4, 21 y 44 h (44 h, las células de la cámara no hipoxia fueron consideradas como 0 h de control). Los niveles de caspasa-3 se determinaron por transferencia Western (FIGURA 46) y los niveles de caspasa-3 se han encontrado para ser regularse hacia arriba por 4 h y alcanzó su punto máximo a 21 h. A 44 h mayoría de las células estaban muertas y los niveles de caspasa-3 estaban bajos. Estos ensayos se pueden usar para demostrar que rHIgM12 es neuro-protector en condiciones de isquemia in vitro.
Un modelo de ratón de accidente cerebrovascular isquémico cerebral se utilizó en los presentes estudios para demostrar la capacidad de rIgM12 para revertir los déficits neurológicos siguientes y proporcionar protección frente a la lesión isquémica cerebral. Este modelo se puede aplicar adicionalmente para optimizar la formulación de dosis, para determinar el porcentaje de anticuerpo que cruza la barrera sangre-cerebro, y para entender mejor protección de las neuronas cultivadas mediante rHIgM12 in vitro de las condiciones de isquemia.
Dosis y biomarcadores para la mejora neurológica
Un ensayo aleatorio controlado en placebo en ratones utilizando rHIgM12 se utiliza para determinar la dosis mínima requerida para la mejora de la actividad espontánea e identificar biomarcadores metabólicos para la mejora neurológica. Los datos anteriores indican que 200 pg de rHIgM12 mejoró la función neurológica en un modelo murino de accidente cerebrovascular isquémico. La determinación de la dosis mínima efectiva es relevante para el diseño de los estudios de seguridad. Un estudio de titulación de dosis de rHIgM12 se lleva a cabo en ratones después de la isquemia cerebral.
La isquemia cerebral en ratones CD-1 hembra utilizando el modelo foto-trombótico con RB colorante se utiliza en estudios de dosificación. Grupos de 5 ratones se tratan ip con 5, 50, 100 o 500 pg de rHIgM12, o con un control comercial de IgM humana (Jackson ImmunoResearch cat 009-000-012) que no se une a las neuronas, o con solución salina, dentro de 30 minutos después de la inducción de isquemia cerebral. La función neurológica se mide antes de la inducción de isquemia o el tratamiento y semanalmente en cajas de actividad cerebral. Grupos de 5 ratones se alojan en cajas de actividad y se registra la actividad vertical y horizontal espontánea. Actividad horizontal nocturna es una medida sensible de la capacidad funcional de un animal (25). Además, todos los ratones se someten a DWI-MRI (Fig 2C) para supervisar el tamaño del infarto en todos los grupos.
La variable principal es la mejora motorada evaluada mediante la supervisión de la actividad espontánea. Un aumento del 20% en la actividad nocturna media (2000 rotura/h) grabado más de 72 h en comparación con los grupos de control de más de 5 días de un grupo rHIgM12 tratadas (P <0,05) se considera una mejora significativa biológica (25). Estableció la metodología de nuestra oratoria en laboratorio (25) se utiliza para calcular las diferencias estadísticas entre los grupos. El análisis de DWI-RM se realizó de forma ciega usando herramientas de análisis de ROI en 3D Analizar 11,0, un paquete de software de análisis de imágenes biomédicas desarrollado en la Mayo Clinic (26, 27).
Se determina si la mejora de la función neurológica tras el tratamiento con rHIgM12 correlaciona con metabolito NAA conservado (un metabolito asociado con la función neuronal (28)), y con relativa normalización de la relación de transferencia de magnetización en MT-MRI en el cerebro. MT es un marcador general de la integridad del tejido y se ha utilizado como un marcador sensible para la integridad de penumbra en el ictus isquémico humano (29). Estos ensayos son útiles como un futuro punto final de los ensayos clínicos en seres humanos de rHIgM12 en la isquemia cerebral.
Sobre la base de los cálculos de potencia obtenidos para los datos MT-MRI, 12 ratones por grupo se usan en este experimento. Técnicas se estandarizaron anteriormente para medir los niveles de N-acetilo-aspartato (NAA)
en el SNC (30). Con el fin de investigar si NAA puede ser utilizado como biomarcador de visualización de metabolito de mejora funcional mediada por rHIgM12 después de la isquemia cerebral, hemos estandarizado un voxel para realizar MRS en la región isquémica. El metabolito NAA se mide en animales tratados con la dosis mínima efectiva, o IgM de control de isotipo o solución salina. MRS se realiza utilizando el espectrómetro de RMN taladro vertical, 7T y datos MRS obtiene de un mm voxel 3x2x1.5 colocado dentro de la zona isquémica como se identifica en DWI y T2 MRI ponderada. El hemisferio contralateral sirve como un control apropiado para cada ratón. Como controles adicionales, los ratones en otros grupos son tratados con control de IgM o PBS. Datos MT-MRI se obtiene mediante la adquisición de una secuencia RARE PD ponderada estándar con 256x96x96 tamaño de la matriz, 6.4 x 1.92 x 1.92 cm FOV, TR: 1500, TE: 30, factor de rAr E de 16, con y sin un pulso de MT de un total tiempo de 270 ms con ancho de banda de pulso de excitación de 500 Hz y el desplazamiento de 1.500 Hz, antes y después del accidente cerebrovascular isquémico seguido de tratamiento con PBS o rHIgM12 MT. Cada experimento se repitió tres veces para confirmar la reproducibilidad de la utilización de biomarcadores de imágenes como criterios de valoración de la mejora funcional, para un total de 108 ratones (12 ratones por grupo para un total de tres grupos, que se repite tres veces) en este experimento.
Los espectros de MRS se cuantificaron usando software TopSpin de Bruker Biospin y a través de LCModel (31-34). Las diferencias estadísticas entre los grupos se calcula mediante ANOVA (30). MTR datos se calculan utilizando las FSL MATHS y el software Analizar 11.0. Normalización relativa de la MTR y la conservación o aumento relativo de los niveles de NAA se determina en los grupos rHIgM12 tratados en comparación con los grupos de control. Un aumento del 10% en NAA conservado en cualquier grupo tratado con rHIgM12 en comparación con los grupos de control se considera una mejora. Neurotransmisores excitatorios tales como el glutamato y glicina (35) se ha informado que están presentes en niveles elevados después de la isquemia cerebral. Ya que espectroscopia MRS es una técnica versátil, también cuantificamos otros metabolitos tales como lactato, mio-inositol, colina, creatina, glicina y glutamato.
Evaluación barrera hematoencefálica
La proporción de rHIgM12 que cruza la barrera hematoencefálica (BBB) se determina en el modelo de accidente cerebrovascular RB foto-trombótica. Ha habido un debate sobre si las grandes moléculas de IgM pueden entrar en el sistema nervioso central. Un dogma generalmente aceptada es que las IgM con un peso molecular de cerca de 1 millón son demasiado grandes para cruzar la BBB de la circulación para entrar en el SNC (36, 37). Sin embargo, hay cada vez más pruebas de que algunos IgM pueden cruzar la barrera hematoencefálica. Se ha demostrado que rHIgM12 puede entrar en el SNC de ratones no infectados normales, así como ratones infectados con TMEV (véase Figura 13). 35S-anticuerpo marcado se usa para determinar el porcentaje de anticuerpo que atraviesa la BBB y demostrando, además, que la IgM actúa directamente sobre las neuronas en la región isquémica. Evaluamos para la unión en el SNC después de la inyección iv en el modelo de isquemia cerebral de neuronas.
rHIgM12 marcado isotópicamente (35S) se produce y se validó para la unión a la superficie de las neuronas. rHIgM12 en dosis eficaz o un control de isotipo IgM o un control de IgG no específico se administran iv, a grupos de 3 ratones por punto de tiempo después de la inducción de isquemia cerebral. Ratones normales CD-1 sin isquemia sirven como control. El experimento se llevó a cabo tres veces utilizando un total de 144 ratones (3 ratones por grupo para un total de 5 puntos de tiempo y 3 grupos además de un grupo de 3 ratones no tratados, repetido 3 veces).
El porcentaje de 35S que atraviesa la BBB y se encuentra en el cerebro/parénquima de la médula espinal a 4, 8, 24, 48 y 72 h después de la inyección se determina. Además, determinamos el sitio de 35S de localización en el cerebro/médula espinal usando técnicas de autorradiografía publicadas por nuestro laboratorio (38). Hemos propuesto que rHIgM12 actúa directamente sobre el sistema nervioso, que cruza la barrera hematoencefálica después de la inyección periférica de interactuar con las neuronas y las protege de la muerte.
Cultivos de neuronas
Neuro-protección de rHIgM12 también se evalúa en ensayo de cultivo in vitro de neuronas "de tipo isquemia". La demostración de eficacia de rHIgM12 en la protección de las neuronas en un ambiente in vitro se dirigirá a la preocupación de que un autoanticuerpo de unión de neuronas puede ser tóxico para las neuronas. Nos dirigimos a este mediante el análisis de los niveles de caspasa después del tratamiento de las neuronas corticales normales, oxígeno y/o glucosa-privada con rHIgM12 o con anticuerpos de control. Además el desarrollo de un ensayo in vitro proporcionará herramientas necesarias para los estudios de señalización futuros.
Los daños después de la isquemia cerebral puede ocurrir por varios mecanismos diferentes. Reactivos que son neuro-protectores para un tipo de daño no son necesariamente de protección para todos los tipos (39). Establecemos ensayos de neurotoxicidad utilizando la privación de glucosa (GD), la privación de oxígeno (OD) y OD combinado y GD (OGD), insultos similares a los observados después de la isquemia cerebral. Hemos llevado a cabo ensayos análogos usando peróxido de hidrógeno (FIGURA 3) y la cámara hipóxica (FIGURA 46) para evaluar los efectos protectores de rHIgM12 sobre las neuronas corticales de ratón. GM1 se utiliza como control positivo, ya que
las propiedades neuroprotectoras de rHIgM12 son una reminiscencia de las observadas para GM1.
Se establecen cultivos de células disociadas de fragmentos neocorticales de embriones CD-1 de ratón de 16 días. Aproximadamente 95% de las células en estos cultivos son las neuronas y las células restantes son astrocitos. GD se induce mediante la incubación de las neuronas en medio de Locke libre de glucosa durante 24 horas. Una cámara libre de oxígeno con 95% N2 , 5% de CO2 atmósfera se utiliza para inducir OD (neuronas se incubaron en medio normal durante 12 h) y OGD (neuronas se incubaron en medio de Locke libre de glucosa durante 12 horas). Las neuronas se incubaron en presencia de medio de cultivo solo, o medio de cultivo complementado con rHIgM12 (en dosis efectiva), o IgM control o GM1. El experimento se realizó tres veces independientemente por triplicado utilizando tratamientos codificados en diferentes días para evitar el sesgo.
El punto final primario es la viabilidad celular neuronal, que se determina mediante un ensayo de la actividad mitocondrial usando solución de azul de Alamar. Cultivos neuronales se analizan para las caspasas activadas utilizando el Kit FLICA de detección de apoptosis. Se utilizan dos métodos de detección de fluorescencia diferentes: microscopía de fluorescencia para el análisis cualitativo; y de fluorometría placa de microtitulación de 96 pocillos para la cuantificación. La supervivencia celular se evaluó mediante la incubación de las células durante 30 min con mancha Hoechst y se leyó usando un filtro UV con excitación a 365 nm y emisión a 480 nm. Cuando las células se observan a través de un microscopio de fluorescencia, las células apoptóticas presentan fluorescencia roja, mientras que las células no apoptóticas aparecen sobre todo sin teñir. Las células en etapas más avanzadas de la apoptosis aparecen como rojo más brillante que las células en las etapas anteriores. El uso de un lector de placas de fluorescencia (con una placa de micro-titulación negra), la apoptosis se cuantifica como la cantidad de fluorescencia roja emitida desde las sondas FLICA unidas a las caspasas. ANOVA se utilizó para comparar las diferencias significativas entre los grupos. Los resultados de este experimento le ayudarán a determinar los eventos de señalización que están involucrados en neuro-protección.
REFERENCIAS CITADAS
1. Walmsley, A.R., y Mir, A.K. 2007. Targeting the Nogo-A signalling pathway to promote recovery following acute CNS injury. Current pharmaceutical design 13:2470-2484.
2. Rodriguez, M., Warrington, A.E., y Pease, L.R. 2009. Invited article: human natural autoantibodies in the treatment of neurologic disease. Neurology 72:1269-1276.
3. Brott, T.G., Haley, E.C., Jr., Levy, D.E., Barsan, W., Broderick, J., Sheppard, G.L., Spilker, J., Kongable, G.L., Massey, S., Reed, R., et al. 1992. Urgent therapy for stroke. Part I. Pilot study of tissue plasminogen activator administered within 90 minutes. Stroke; a journal of cerebral circulation 23:632-640.
4. Haley, E.C., Jr., Levy, D.E., Brott, T.G., Sheppard, G.L., Wong, M.C., Kongable, G.L., Torner, J.C., y Marler, J.R. 1992. Urgent therapy for stroke. Part II. Pilot study of tissue plasminogen activator administered 91-180 minutes from onset. Stroke; a journal of cerebral circulation 23:641-645.
5. Group, T.N.I.o.N.D.a.S.r.-P.S.S. 1995. Tissue plasminogen activator for acute ischemic stroke. The New England journal of medicine 333:1581-1587.
6. Gobin, Y.P., Starkman, S., Duckwiler, G.R., Grobelny, T., Kidwell, C.S., Jahan, R., Pile-Spellman, J., Segal, A., Vinuela, F., y Saver, J.L. 2004. MERCI 1: a phase 1 study of Mechanical Embolus Removal in Cerebral Ischemia. Stroke; a journal of cerebral circulation 35:2848-2854.
7. Clark, W.M., Wissman, S., Albers, G.W., Jhamandas, J.H., Madden, K.P., y Hamilton, S. 1999. Recombinant tissue-type plasminogen activator (Alteplase) for ischemic stroke 3 to 5 hours after symptom onset. The ATLANTIS Study: a randomized controlled trial. Alteplase Thrombolysis for Acute Noninterventional Therapy in Ischemic Stroke. JAMA : the journal of the American Medical Association 282:2019-2026.
8. Hacke, W., Kaste, M., Bluhmki, E., Brozman, M., Davalos, A., Guidetti, D., Larrue, V., Lees, K.R., Medeghri, Z., Machnig, T., et al. 2008. Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic stroke. The New England journal of medicine 359:1317-1329.
9. Investigators, E.A.S.T. 2001. Use of anti-ICAM-1 therapy in ischemic stroke: results of the Enlimomab Acute Stroke Trial. Neurology 57:1428-1434.
10. Adams, H.P., Jr., Effron, M.B., Torner, J., Davalos, A., Frayne, J., Teal, P., Leclerc, J., Oemar, B., Padgett, L., Barnathan, E.S., et al. 2008. Emergency administration of abciximab for treatment of patients with acute ischemic stroke: results of an international phase III trial: Abciximab in Emergency Treatment of Stroke Trial (AbESTT-II). Stroke; a journal of cerebral circulation 39:87-99.
11. Arumugam, T.V., Tang, S.C., Lathia, J.D., Cheng, A., Mughal, M.R., Chigurupati, S., Magnus, T., Chan, S.L., Jo, D.G., Ouyang, X., et al. 2007. Intravenous immunoglobulin (IVIG) protects the brain against experimental stroke by preventing complement-mediated neuronal cell death. Proceedings of the National Academy of Sciences of the United States of America 104:14104-14109.
12. Vucic, S., Chong, P.S., Dawson, K.T., Cudkowicz, M., y Cros, D. 2004. Thromboembolic complications of intravenous immunoglobulin treatment. European neurology 52:141-144.
13. Rajabally, Y.A., y Kearney, D.A. 2011. Thromboembolic complications of intravenous immunoglobulin therapy in patients with neuropathy: a two-year study. J Neurol Sci 308:124-127.
14. Bieber, A.J., Warrington, A., Pease, L.R., y Rodriguez, M. 2001. Humoral autoimmunity as a mediator of CNS repair. Trends in neurosciences 24:S39-44.
15. Warrington, A.E., Bieber, A.J., Ciric, B., Van Keulen, V., Pease, L.R., Mitsunaga, Y., Paz Soldán, M.M., y Rodríguez, M. 2001. Immunoglobulin-mediated CNS repair. J Allergy Clin Immunol 108:S121-125.
16. Warrington, A.E., Asakura, K., Bieber, A.J., Ciric, B., Van Keulen, V., Kaveri, S.V., Kyle, R.A., Pease, L.R., y Rodriguez, M. 2000. Human monoclonal antibodies reactive to oligodendrocytes promote remyelination in a model of multiple sclerosis. Proc Natl Acad Sci U S A 97:6820-6825.
17. Warrington, A.E., Bieber, A.J., Van Keulen, V., Ciric, B., Pease, L.R., y Rodriguez, M. 2004. Neuron-binding human monoclonal antibodies support central nervous system neurite extension. Journal of neuropathology and experimental neurology 63:461-473.
18. Ciric, B., VanKeulen, V., Rodriguez, M., Kyle, R.A., Gertz, M.A., y Pease, L.R. 2001. Clonal evolution in Waldenstrom macroglobulinemia highlights functional role of B-cell receptor. Blood 97:321-323.
19. Mitsunaga, Y., Ciric, B., Van Keulen, V., Warrington, A.E., Paz Soldan, M., Bieber, A.J., Rodriguez, M., y Pease, L.R. 2002. Direct evidence that a human antibody derived from patient serum can promote myelin repair in a mouse model of chronic-progressive demyelinating disease. The FASEB journal : official publication of the Fed- eration of American Societies for Experimental Biology 16:1325-1327.
20. Wright, B.R., Warrington, A.E., Edberg, D.D., y Rodriguez, M. 2009. Cellular mechanisms of central nervous system repair by natural autoreactive monoclonal antibodies. Archives of neurology 66:1456-1459.
21. Howe, C.L., Bieber, A.J., Warrington, A.E., Pease, L.R., y Rodriguez, M. 2004. Antiapoptotic signaling by a remyelination-promoting human antimyelin antibody. Neurobiol Dis 15:120-131.
22. Watzlawik, J., Holicky, E., Edberg, D.D., Marks, D.L., Warrington, A.E., Wright, B.R., Pagano, R.E., y Rodriguez, M. 2010. Human remyelination promoting antibody inhibits apoptotic signaling and differentiation through Lyn kinase in primary rat oligodendrocytes. Glia 58:1782-1793.
23. Woolrich, M.W., Jbabdi, S., Patenaude, B., Chappell, M., Makni, S., Behrens, T., Beckmann, C., Jenkinson, M., y Smith, S.M. 2009. Bayesian analysis of neuroimaging data in FSL. Neurolmage 45:S173-186.
24. Smith, S.M., Jenkinson, M., Woolrich, M.W., Beckmann, C.F., Behrens, T.E., Johansen-Berg, H., Bannister, P.R., De Luca, M., Drobnjak, I., Flitney, D.E., et al. 2004. Advances in functional and structural MR image analysis and implementation as FSL. Neurolmage 23 Suppl 1:S208-219.
25. Denic, A., Macura, S.I., Warrington, A.E., Pirko, I., Grossardt, B.R., Pease, L.R., y Rodriguez, M. 2011. A single dose of neuron-binding human monoclonal antibody improves spontaneous activity in a murine model of demyelination. PloS one 6:e26001.
26. Robb, R.A. 1999. 3-D visualization in biomedical applications. Annual review of biomedical engineering 1:377-399.
27. Robb, R.A. 2001. The biomedical imaging resource at Mayo Clinic. IEEE transactions on medical imaging 20:854-867.
28. De Stefano, N., Matthews, P.M., Fu, L., Narayanan, S., Stanley, J., Francis, G.S., Antel, J.P., y Arnold, D.L.
1998. Axonal damage correlates with disability in patients with relapsing-remitting multiple sclerosis. Results of a longitudinal magnetic resonance spectroscopy study. Brain 121 (Pt 8):1469-1477.
29. Tourdias, T., Dousset, V., Sibon, I., Pele, E., Menegon, P., Asselineau, J., Pachai, C., Rouanet, F., Robinson, P., Chene, G., et al. 2007. Magnetization transfer imaging shows tissue abnormalities in the reversible penumbra. Stroke; a journal of cerebral circulation 38:3165-3171.
30. Denic, A., Bieber, A., Warrington, A., Mishra, P.K., Macura, S., y Rodriguez, M. 2009. Brainstem 1H nuclear magnetic resonance (NMR) spectroscopy: marker of demyelination and repair in spinal cord. Ann Neurol 66:559-564.
31. Henry, P.G., Oz, G., Provencher, S., y Gruetter, R.2003. Toward dynamic isotopomer analysis in the rat brain in vivo: automatic quantitation of 13C NMR spectra using LCModel. NMR in biomedicine 16:400-412.
32. Opstad, K.S., Provencher, S.W., Bell, B.A., Griffiths, J.R., y Howe, F.A. 2003. Detection of elevated glutathione in meningiomas by quantitative in vivo 1H MRS. Magnetic resonance in medicine : official journal of the Society of Magnetic Resonance in Medicine / Society of Magnetic Resonance in Medicine 49:632-637.
33. Provencher, S.W. 2001. Automatic quantitation of localized in vivo 1H spectra with LCModel. NMR in biomedicine 14:260-264.
34. Pfeuffer, J., Tkac, I., Provencher, S.W., and Gruetter, R. 1999. Toward an in vivo neurochemical profile: quantification of 18 metabolites in short-echo-time (1)H NMR spectra of the rat brain. Journal of magnetic resonance 141:104-120.
35. Castillo, J., Davalos, A., y Noya, M. 1997. Progression of ischaemic stroke and excitotoxic aminoacids. Lancet 349:79-83.
36. Banks, W.A. 2008. Developing drugs that can cross the blood-brain barrier: applications to Alzheimer's disease. BMC Neurosci 9 Suppl 3:S2.
37. Kozlowski, G.P., Sterzl, I., and Nilaver, G. 1992. Localization patterns for immunoglobulins and albumins in the brain suggest diverse mechanisms for their transport across the blood-brain barrier (BBB). Prog Brain Res 91:149-154.
38. Hunter, S.F., Miller, D.J., y Rodriguez, M. 1997. Monoclonal remyelination-promoting natural autoantibody SCH 94.03: pharmacokinetics and in vivo targets within demyelinated spinal cord in a mouse model of multiple sclerosis. J Neurol Sci 150:103-113.
39. Ryu, B.R., Choi, D.W., Hartley, D.M., Costa, E., Jou, I., y Gwag, B.J. 1999. Attenuation of cortical neuronal apoptosis by gangliosides. The Journal of pharmacology and experimental therapeutics 290:811-816.
EJEMPLO 22: Actividad de anticuerpos y efectos en condiciones normales (Normóxico) y condiciones hipóxicas
A fin de evaluar los efectos y la capacidad del anticuerpo neuroactivo, particularmente anticuerpo IgM12, en las células y en las condiciones pertinentes y que imitan accidente cerebrovascular, infarto de miocardio o isquemia, se llevaron a cabo estudios para evaluar los anticuerpos en condiciones de normoxia y condiciones de hipoxia. Condiciones hipóxicas en cultivo sirven para imitar aspectos de la privación de oxígeno que resulta de apoplejía y la isquemia en los escenarios clínicos. Cultivos normóxicos se evaluaron primero para diversos marcadores en cultivos gliales mixtos. Marcadores relevantes se evaluaron a continuación en condiciones de hipoxia, con y sin anticuerpos neuroactivos y de control para evaluar los efectos y capacidades.
Condiciones de normoxia
Para evaluar las condiciones normales o de normoxia, se hicieron crecer cultivos gliales mixtos durante 4 días en FBS (10%) que contiene el medio y se trataron durante 7 días en medio definido químicamente con la remielinización de promoción de IgM rHIgM22, mAb rHIgM12regenerativa, isotipo controla sHIgM116 (10 pg/ml cada uno) o medio solamente. El medio con anticuerpos frescos se intercambió cada dos días. Los resultados se representan en la Figura 47. Los gráficos de barras muestran el análisis cuantitativo de transferencias Western a partir de tres experimentos independientes con MBP marcador de mielina, marcador de apoptosis escinde la caspasa-3 y CD68 como marcador de microglía activada. Estos resultados muestran que el anticuerpo regenerativo rHIgM12 estimula la diferenciación de célula progenitor de oligodendrocitos (OPC) y la supervivencia en cultivos gliales mixtos. Estos datos muestran que el anticuerpo regenerativo rHIgM12, pero no anticuerpo promotor de remielinización rHIgM22 o control de isotipo sHIgM116, estimula la diferenciación de OPC, reduce los niveles de apoptosis glial y activa la microglia. El tipo específico de célula rescatado de la apoptosis no se ha determinado.
Co-cultivos microgliales OPC se cultivaron a continuación en medio de astrocitos acondicionado con remielinización que promueve mAbs A2B5, O4, O1, 78.09, 94.03, rHIgM22, mAb rHIgM12regenerativa, controles de isotipo (LEAF, 5A5, ChromPure IgM, rHIgM42, sHIgM14, sHIgM24, sHIgM26) (10 pg/ml cada uno) o medio. Medio con anticuerpos frescos se intercambió cada dos días. Transferencias Western representativas en la Figura 48 muestran niveles de marcadores de OPC PDGFaR, marcadores de mielina CNPasa y MOG, marcadores microgliales CD68 y Lyn, la proliferación de marcador Ki-67 y la histona H3, marcador de apoptosis escindido pRbS795 caspasa-3 y marcadores de ciclo celular. Los astrocitos y microglia son necesarios para mAbs regenerativos de unión a GSL para estimular diferenciación OPC y la supervivencia en medio de astrocitos acondicionado.
Los datos de normoxia anteriores demuestran que (i) mAbs de promoción de remielinización (A2B5, O4, O1, 78.09, 94.03) o mAb rHIgM12 regenerativa no inducen la diferenciación OPC o la supervivencia en comparación con controles de isotipo en co-cultivos microgliales de OPC/OL; (ii) factores solubles en el medio de astrocitos acondicionado no eran suficientes para estimular las diferencias entre grupos de tratamiento; (iii) la promoción y la remielinización mAb regenerativas no activan la microglia (CD68, Lyn) en ausencia de astrocitos; (iv) factores astrocíticos solubles parecen ser suficientes para mediar la proliferación de OPC por rHIgM22 (Ki-67, la histona H3) después de 6 días de tratamiento de IgM.
En la siguiente serie de estudios de normoxia, los cultivos gliales mixtos fueron cultivados y tratados como se describió anteriormente (FIGURA 47 anterior). FIGURA 49 proporciona gráficos de barras del análisis cuantitativo de las transferencias de Western de tres experimentos independientes con marcador CD68 de microglia activada y quinasa de Lyn, una quinasa muy abundante en la microglía y en un grado menor en los OPC pero no en astrocitos. Imágenes representativas de inmunofluorescencia se proporcionan en la Figura 50 y muestran niveles de CD68 (verde) y marcador nuclear DAPI (azul) en glial mixto antes del tratamiento mAb en el día 4 en cultivo y en el día 11 en cultivo después de 7 días de tratamiento con control de isotipo rHIgM12, rHIgM22 y sHIgM24 (10 pg/ml cada uno). Estos datos demuestran que mAb rHIgM12 regenerativa aumenta la expresión de CD68 y quinasa de Lyn en cultivos gliales mixtos. Altos niveles de expresión de CD68 en transferencias Western (FIGURA 49) pueden correlacionarse con un mayor número de microglia activada mediante inmunocitoquímica (FIGURA 50). Por lo tanto, como se muestra en estos estudios, los anticuerpos de promoción de remielinización (ejemplos de anticuerpos A2B5 y O1) y mAb IgM12 regenerativa activan microglia (CD68) en cultivos gliales mixtos.
Condiciones hipóxicas
Los efectos moleculares de la hipoxia en cultivos gliales mixtos fueron comparados con los cultivos de normoxia. Cultivos gliales mixtos se cultivaron durante 4 días en FBS (10%) que contenía medio y se trató durante 7 días adicionales en medio químicamente definido en condiciones de hipoxia (3% 02) en comparación con condiciones de normoxia (21% 02). Transferencias de Western representativas (FIGURA 51) muestran niveles del factor inducible por hipoxia 1 (HIF1 a), los marcadores de oligodendrocitos NG2, PDGFa, Olig-2, marcadores de mielina de MBP y CNPasa, marcador de astrocitos GFAP, marcador microglial CD68, marcadores del ciclo celular pRbS795 y el marcador de apoptosis escinde la caspasa-3. Beta-actina se utilizó como control de carga. El análisis
cuantitativo de las transferencias de Western de cultivos gliales mixtos expuestos a la hipoxia y normoxia como se muestra en la Figura 51 se completó a partir de tres experimentos independientes y se muestra en la Figura 52. Estos datos demuestran la reducción de niveles de expresión de marcadores microgliales y astrocíticos y aumento de los niveles de marcadores de mielina en cultivos gliales mixtos en condiciones de hipoxia en comparación con condiciones de normoxia. La verificación del modelo hipóxico se muestra por el aumento de expresión HIF1a en condiciones de hipoxia.
Después de haber verificado el modelo hipóxico, el tratamiento de los cultivos hipóxicos con anticuerpo regenerativo IgM12 se llevó a cabo a continuación para evaluar los efectos de anticuerpos en condiciones relevantes para imitar accidente cerebrovascular, infarto y efectos de isquemia. Cultivos gliales mixtos se cultivaron durante 4 días en condiciones de normoxia en FBS (10%) que contiene el medio y se trataron posteriormente durante 1-7 días en medio químicamente definido con anticuerpo regenerativo humano rHIgM12 o medio sólo bajo la hipoxia (3% 02). transferencias Western representativas (FIGURA 53) muestran niveles de marcador de proliferación Ki-67, los marcadores microgliales IBA-1, CD68, y iNOS, marcador de apoptosis exfoliado caspasa-3, astrocitos GFAP marcador, marcadores del ciclo celular pRbS795 y pRbS807 y los marcadores de oligodendrocitos PDGFaR, NG2, Olig-1, Olig-2 y la mielina marcador MBP. Los resultados de estos datos indican la eficacia del tratamiento rHIgM12 en condiciones de hipoxia. rHIgM12 activa microglia, estimula la salida del ciclo celular de las células gliales (PRBS), reduce los niveles de apoptosis glial, y reduce niveles incrementados sobre la base de la producción de mielina en condiciones de hipoxia. Por lo tanto, el anticuerpo regenerativo de rHIgM12 estimula la supervivencia glial y reduce la diferenciación OPC en cultivos gliales mixtos hipóxicos.
El resultado molecular de rHIgM12 bajo hipoxia frente a normoxia se evaluó por comparación directa de los efectos mediados por rHIgM12 en glia mixtos en condiciones de hipoxia en comparación con la normoxia. Glia mixtos se cultivaron como se describe antes y se trataron en medio químicamente definido durante 7 días con rHIgM12, rHIgM22 o medio sólo bajo condiciones hipóxicas o normóxicas. Transferencias Western representativas Figura 54) muestran los niveles de marcadores microgliales IBA-1 y CD68, marcador de apoptosis escinde la caspasa-3, marcador de astrocitos GFAP, los marcadores de oligodendrocitos PDGFaR, NG2 y Olig-2 y los marcadores de mielina de MBP y CNPasa. rHIgM12 muestra una eficacia tanto en condiciones de normoxia como hipóxicas para estimular microgliales de marcadores de expresión CD68 y IBA-1. Más significativamente, rHIgM12 equilibra el impacto de la hipoxia sobre la expresión de marcadores de mielina MBP y CNPasa en comparación con las condiciones de normoxia.
Se evaluaron las condiciones hipóxicas en los animales para comprobar la validez y la coherencia de los marcadores de cultivo. Se realizó un análisis molecular de cortezas hipóxicas y normóxicas de ratones B16 neonatales de P13 de edad. Ratones neonatales C57/BI6 fueron fomentados transversalmente por embalses CD1 y se expusieron a hipoxia (10% 02) durante 4 días a partir de P3 a P7. Los ratones fueron posteriormente expuestos al aire ambiente durante 6 días adicionales y se sacrificaron a P13. Se aisló ARN a partir de cortezas en ambos grupos de tratamiento (ratones de normoxia e hipoxia) y se sondó para los marcadores de mielina PLP1, MOG, Mbp1, marcadores progenitores de oligodendrocitos PDGFaR y Olig-1, marcador astrocítico GFAP, marcadores neuronales nestina y 133-tubulina, hipoxia inducible factor 1a y la proteína sarcospan SSPN. Los niveles de expresión se analizaron por RT-PCR y resultaron normalizados a los niveles de p-actina. Análisis de RT-PCR del marcador se llevó a cabo en cortezas de neonatos C57/B16 de hipoxia y normoxia a P13 y los resultados se muestran en la Figura 55. Los resultados de estos datos indican que el tratamiento hipóxico de ratones recién nacidos reduce selectivamente los niveles de expresión de los marcadores de mielina pero no marcadores neuronales, de OPC o astrocíticos en el tejido cortical.
EJEMPLO 23 - Localización de anticuerpos IgM rHIgM12 en un modelo de ratón de accidente cerebrovascular
Para evaluar el acceso anticuerpo y la localización de intervención tras un accidente cerebrovascular o evento isquémico, un modelo animal de accidente cerebrovascular se utilizó con la administración IP de anticuerpo monoclonal. Un derrame cerebral fue inducido en ratones, el anticuerpo radiomarcado administrado, y después se evaluaron los tejidos animales para anticuerpo mediante la determinación del radio-etiquetado en períodos de hasta 24 horas después de la administración. Se evaluó cerebro (corteza) que contiene la lesión de accidente cerebrovascular, así como cerebelo, médula espinal, hígado, bazo, riñón y suero sanguíneo.
El accidente cerebrovascular fue inducido en la corteza cerebral derecha en tres adultos ratones CD-1 utilizando el foto activan protocolo de Rosa de Bengala descrito anteriormente y otros tres ratones recibieron un procedimiento de accidente cerebrovascular, Rosa de Bengala, pero no la activación de la luz. En 30 a 40 min después del accidente cerebrovascular, 10 millones cpm de 35S etiquetados rHIgM12, correspondientes a aproximadamente 60 ug de IgM, se administró por vía intraperitoneal a cada ratón en un volumen de 500 ul de solución salina. En IgM 6, 12 y 24 horas después de la la administración o tratamiento, se recogió sangre de los ratones, la vasculatura se lavó con 30 ml de solución salina y tejidos recogidos de forma aguda. Para el análisis de radiomarcador en el tejido, la corteza frontal derecha del cerebro que contiene el área de la lesión se eliminó como un bloque de tejido 75 mg. El tejido se disolvió en soluble (PerkinElmer) y cpm determina en cóctel de centelleo
Claims (15)
1. Un anticuerpo de IgM humano aislado o un fragmento del mismo que se une específicamente a las neuronas y protege las neuronas de la muerte celular y que no promueve la remielinización, en donde el anticuerpo de IgM humano aislado o fragmento comprende la siguiente secuencia:
(a) las secuencias del dominio de CDR de aminoácidos de cadena pesada variable CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y las secuencias de la CDR de cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), como se expone en la Figura 5; o
(b) las secuencias del dominio de CDR de aminoácidos de cadena pesada variable CDR1 GFTFSTYA (SEQ ID NO: 37), CDR2 INVGGVTT (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), y las secuencias de CDR de cadena ligera CDR1 QGIG ( SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42), como se establece en la Figura 6,
para su uso en un método para tratar o mejorar la isquemia cerebral o accidente cerebrovascular, mejorar la función neural en un animal después de un accidente cerebrovascular o isquemia cerebral, o para proteger neuronas o células neurales del daño o muerte celular después de un accidente cerebrovascular o isquemia cerebral, en donde el método comprende administrar el anticuerpo de IgM humano aislado o fragmento del mismo en combinación con un anticuerpo remielinizante, y en donde el anticuerpo remielinizante comprende:
(i) las CDR de la región variable de la cadena pesada que comprenden la secuencia de CDR1 SSGMH (SEQ ID NO: 47), la secuencia de CDR2 V(I)ISYDGSRKYYADSVKG (SEQ ID NO: 48) y la secuencia de CDR3 GVTGSPTLDY (SEQ ID NO: 49), y las CDR de la región variable de la cadena ligera que comprenden la secuencia de CDR1 SGSSSNIGNNFVS (SEQ ID NO: 50), la secuencia de CDR2 DITKRPS (SEQ ID NO: 51) y la secuencia de CDR3 G(E)TWDSSLSAW (SEQ ID NO: 52); o
(ii) las CDR de la región variable de la cadena pesada que comprenden la secuencia de CDR1 SGFTFSSYW (SEQ ID NO: 53), secuencia de CDR2 IKKDGSEK (SEQ ID NO: 54) y secuencia de CDR3 Ar PNCGGDCYLPWYFD (SEQ ID NO: 55), y las CDR de la región variable de la cadena ligera que comprenden la secuencia de CDR1 QSVLYSSNNKNY (SEQ ID NO: 56), la secuencia de CDR2 YWAS (SEQ ID NO: 57) y la secuencia de CDR3 QQYYNTPQA (SEQ ID NO: 58).
2. Un anticuerpo remielinizante que comprende:
(i) las CDR de la región variable de la cadena pesada que comprenden la secuencia de CDR1 SSGMH (SEQ ID NO: 47), la secuencia de CDR2 V(I)ISYDGSRKYYADSVKG (SEQ ID NO: 48) y la secuencia de CDR3 GVTGSPTLDY (SEQ ID NO: 49), y las CDR de la región variable de la cadena ligera que comprenden la secuencia de CDR1 SGSSSNIGNNFVS (SEQ ID NO: 50), la secuencia de CDR2 DITKRPS (SEQ ID NO: 51) y la secuencia de CDR3 G (E) TWDSSLSAW (SEQ ID NO: 52); o
(ii) las CDR de la región variable de la cadena pesada que comprenden la secuencia de CDR1 SGFTFSSYW (SEQ ID NO: 53), la secuencia de CDR2 IKKDGSEK (SEQ ID NO: 54) y la secuencia de CDR3 ARPNCGGDCYLPWYFD (SEQ ID NO: 55), y las CDR de la región variable de la cadena ligera que comprenden la secuencia de CDR1 QSVLYSSNNKNY (SEQ ID NO: 56), la secuencia de CDR2 YWAS (SEQ ID NO: 57) y la secuencia de CDR3 QQYYNTPQA (SEQ ID NO: 58),
para su uso en un método para tratar o mejorar la isquemia cerebral o accidente cerebrovascular, mejorar la función neural en un animal después de un accidente cerebrovascular o isquemia cerebral, o para proteger neuronas o células neurales del daño o muerte celular después de un accidente cerebrovascular o isquemia cerebral, en donde el método comprende administrar el anticuerpo remielinizante en combinación con un anticuerpo de IgM humano aislado o un fragmento del mismo que se une específicamente a las neuronas y protege a las neuronas de la muerte celular y que no promueve la remielinización, y en donde el anticuerpo de IgM humano aislado o fragmento comprende la siguiente secuencia:
(a) las secuencias del dominio de CDR de aminoácidos de la cadena pesada variable CDR1 GGSVSLYY (SEQ ID NO: 31), CDR2 GYIYSSGST (SEQ ID NO: 32) y CDR3 ARSASIRGWFD (SEQ ID NO: 33), y las secuencias de CDR de cadena ligera CDR1 QSISSY (SEQ ID NO: 34), CDR2 AAS (SEQ ID NO: 35) y CDR3 QQSYHTPW (SEQ ID NO: 36), como se expone en la Figura 5; o
(b) las secuencias del dominio de CDR de aminoácidos de cadena pesada variable CDR1 GFTFSTYA (SEQ ID NO: 37), CDR2 INVGGVTT (SEQ ID NO: 38) y CDR3 VRRSGPDRNSSPADF (SEQ ID NO: 39), y las secuencias de CDR de cadena ligera CDR1 QGIG (SEQ ID NO: 40), CDR2 TTS (SEQ ID NO: 41) y CDR3 QKYNSAPRT (SEQ ID NO: 42), como se expone en la Figura 6.
3. El anticuerpo de IgM humano aislado o fragmento del mismo para el uso de la reivindicación 1, o el anticuerpo remielinizante para el uso de la reivindicación 2, en donde el anticuerpo de IgM humano aislado o fragmento del mismo comprende la secuencia de aminoácidos de la cadena pesada variable expuesta en la SEQ ID NO: 1 y la
secuencia de aminoácidos de la cadena ligera variable expuesta en la SEQ ID NO: 11, o la secuencia de aminoácidos de la cadena pesada variable expuesta en la SEQ ID NO: 17 y la secuencia de aminoácidos de la cadena ligera variable expuesta en la SEQ ID NO: 27, o variantes altamente homólogas de las mismas, en donde dichas variantes retienen la unión neuronal y la actividad neuroprotectora o neurorregenerativa.
4. El anticuerpo de IgM humano aislado o fragmento del mismo para el uso de la reivindicación 1, o el anticuerpo remielinizante para el uso de la reivindicación 2, en donde el anticuerpo de IgM humano aislado o fragmento del mismo comprende además una secuencia de cadena J humana.
5. El anticuerpo de IgM humano aislado o fragmento del mismo para su uso o el anticuerpo remielinizante para el uso de la reivindicación 4, en donde la cadena J comprende la secuencia de aminoácidos expuesta en la SEQ ID NO: 15.
6. El anticuerpo de IgM humano aislado o un fragmento del mismo para su uso o el anticuerpo remielinizante para el uso de la reivindicación 5, en donde el anticuerpo de IgM humano aislado o un fragmento del mismo es un anticuerpo completamente humano o un fragmento de anticuerpo del mismo que comprende una región variable de la cadena pesada y de la cadena ligera que comprende una secuencia de aminoácidos seleccionada de las secuencias de aminoácidos SEQ ID NO: 1 y 11, o variantes altamente homólogas de las mismas, en donde dichas variantes retienen la unión de neuronas y la actividad neuroprotectora o neurorregenerativa.
7. El anticuerpo de IgM humano aislado o fragmento del mismo para su uso o el anticuerpo remielinizante para el uso de cualquiera de las reivindicaciones 1 a 6, en donde el anticuerpo remielinizante comprende:
(i) la secuencia de aminoácidos expuesta en la SEQ ID NO: 43 y 44, o
(ii) la secuencia de aminoácidos expuesta en la SEQ ID NO: 45 y 46.
8. El anticuerpo de IgM humano aislado o fragmento del mismo para su uso o el anticuerpo remielinizante para el uso de cualquiera de las reivindicaciones 1 a 7, en donde la combinación de anticuerpos se administra:
(i) colectivamente, o
(ii) en serie.
9. El anticuerpo de IgM humano aislado o fragmento del mismo para su uso o el anticuerpo remielinizante para el uso de cualquiera de las reivindicaciones 1 a 8, en donde el método comprende administrar el anticuerpo de IgM humano aislado o fragmento del mismo y el anticuerpo remielinizante en combinación con uno o más agentes que es un trombolítico, un fármaco antiplaquetario, un fármaco antihipertensivo o estatina.
10. El anticuerpo de IgM humano aislado o fragmento del mismo para su uso o anticuerpo remielinizante para el uso de cualquiera de las reivindicaciones 1 a 9, en donde el anticuerpo de IgM humano aislado o fragmento del mismo y/o el anticuerpo remielinizante está marcado con un marcador detectable o funcional.
11. El anticuerpo de IgM humano aislado o fragmento del mismo para su uso o el anticuerpo remielinizante para el uso de la reivindicación 10, en donde el marcador es una enzima, una pareja de unión específica, un ligando, un colorante, un marcador fluorescente y/o un elemento radiactivo.
12. Una composición farmacéutica para el uso como se define en cualquiera de las reivindicaciones 1 a 11, en donde la composición farmacéutica comprende un anticuerpo de IgM humano aislado o un fragmento como se define en cualquiera de las reivindicaciones 1 a 11 y un vehículo, portador o diluyente farmacéuticamente aceptable.
13. Una composición farmacéutica para el uso como se define en cualquiera de las reivindicaciones 1 a 11, en donde la composición farmacéutica comprende un anticuerpo remielinizante como se define en cualquiera de las reivindicaciones 1 a 11 y un vehículo, portador o diluyente farmacéuticamente aceptable.
14. La composición farmacéutica para su uso de acuerdo con la reivindicación 12 o la reivindicación 13, en donde el método comprende administrar la composición como una dosis única a dicho animal que se sospecha o se determina que padece de accidente cerebrovascular o isquemia cerebral.
15. Un kit para su uso en el tratamiento o la mejora de un accidente cerebrovascular o isquemia cerebral en un animal, que comprende una forma de dosificación farmacéutica de la composición farmacéutica de la reivindicación 12, y una forma de dosificación farmacéutica separada de la composición farmacéutica de la reivindicación 13, y que comprende además opcionalmente uno o más de un agente neuroactivo adicional o terapéutico, trombolítico, agente antihipertensivo, agente antiplaquetario, agente antiinflamatorio, agente modulador de la liberación de neurotransmisores, ligando o agonista o antagonista de neurorreceptores, agente de canal de calcio, modulador inmune u otro anticuerpo reactivo del SNC.
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201261625628P | 2012-04-17 | 2012-04-17 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2870717T3 true ES2870717T3 (es) | 2021-10-27 |
Family
ID=49384031
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES13778780.0T Active ES2655077T3 (es) | 2012-04-17 | 2013-04-17 | Anticuerpos humanos y secuencias de unión específicos del mismo para uso en accidente cerebrovascular e isquemia y condiciones isquémicas |
| ES17188389T Active ES2870717T3 (es) | 2012-04-17 | 2013-04-17 | Anticuerpos humanos y secuencias de unión específicas del mismo para su uso en accidente cerebrovascular e isquemia y condiciones isquémicas |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES13778780.0T Active ES2655077T3 (es) | 2012-04-17 | 2013-04-17 | Anticuerpos humanos y secuencias de unión específicos del mismo para uso en accidente cerebrovascular e isquemia y condiciones isquémicas |
Country Status (12)
| Country | Link |
|---|---|
| US (1) | US20150104460A1 (es) |
| EP (2) | EP3305323B1 (es) |
| JP (5) | JP6162219B2 (es) |
| CN (2) | CN108285483B (es) |
| BR (1) | BR112014025898A2 (es) |
| CA (1) | CA2870625C (es) |
| ES (2) | ES2655077T3 (es) |
| IL (2) | IL235118B (es) |
| IN (1) | IN2014MN02274A (es) |
| MX (1) | MX356347B (es) |
| NZ (1) | NZ701278A (es) |
| WO (1) | WO2013158748A1 (es) |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20060099203A1 (en) * | 2004-11-05 | 2006-05-11 | Pease Larry R | B7-DC binding antibody |
| PL3560954T3 (pl) * | 2014-04-03 | 2021-12-13 | Igm Biosciences, Inc. | Zmodyfikowany łańcuch J |
| WO2016073963A1 (en) * | 2014-11-07 | 2016-05-12 | Mayo Foundation For Medical Education And Research | Treatment of neonatal hypoxia including impairments or effects thereof |
| EP3957738A1 (en) | 2015-03-04 | 2022-02-23 | IGM Biosciences, Inc. | Cd20 binding molecules and uses thereof |
| EP3355913B1 (en) | 2015-09-30 | 2024-10-30 | IGM Biosciences, Inc. | Binding molecules with modified j-chain |
| CN113035298B (zh) * | 2021-04-02 | 2023-06-20 | 南京信息工程大学 | 递归生成大阶数行限制覆盖阵列的药物临床试验设计方法 |
| CN113444787A (zh) * | 2021-07-05 | 2021-09-28 | 中山大学附属第七医院(深圳) | 一种自闭症的脂质生物标志物及其应用 |
| CN114276418B (zh) * | 2021-12-28 | 2024-06-25 | 中国科学技术大学 | 一种可用于调控微管、治疗脱髓鞘病变靶向治疗的rna |
Family Cites Families (19)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB8308235D0 (en) | 1983-03-25 | 1983-05-05 | Celltech Ltd | Polypeptides |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| US5776093A (en) | 1985-07-05 | 1998-07-07 | Immunomedics, Inc. | Method for imaging and treating organs and tissues |
| GB9015198D0 (en) | 1990-07-10 | 1990-08-29 | Brien Caroline J O | Binding substance |
| ES2156149T3 (es) | 1992-12-04 | 2001-06-16 | Medical Res Council | Proteinas de union multivalente y multiespecificas, su fabricacion y su uso. |
| US7473423B2 (en) * | 1994-04-29 | 2009-01-06 | Mayo Foundation For Medical Education And Research | Human IgM antibodies, and diagnostic and therapeutic uses thereof particularly in the central nervous system |
| US6306393B1 (en) | 1997-03-24 | 2001-10-23 | Immunomedics, Inc. | Immunotherapy of B-cell malignancies using anti-CD22 antibodies |
| IL152738A0 (en) * | 2000-05-10 | 2003-06-24 | Mayo Foundation | Human igm antibodies with the capability of inducing remyelination, and diagnostic and therapeutic uses thereof particularly in the central nervous system |
| CA2525917C (en) | 2003-05-16 | 2017-07-04 | Mayo Foundation For Medical Education & Research | Compositions and methods including a recombinant human mab that promotes cns remyelination |
| DE602004030451D1 (de) * | 2003-07-15 | 2011-01-20 | Chugai Pharmaceutical Co Ltd | Produktion von igm durch transformierte zellen und verfahren zur quantifizierung dieser igm-produktion |
| WO2006004988A2 (en) * | 2004-06-30 | 2006-01-12 | Mayo Foundation For Medical Education And Research | B7-dc binding antibody |
| WO2006029184A2 (en) * | 2004-09-08 | 2006-03-16 | Expression Diagnostics, Inc. | Genes useful for diagnosing and monitoring inflammation related disorders |
| AU2007227251A1 (en) * | 2006-03-20 | 2007-09-27 | Amgen Fremont, Inc. | Antibodies directed to angiopoietin-like protein 4 and uses thereof |
| US8796429B2 (en) * | 2006-05-31 | 2014-08-05 | Lpath, Inc. | Bioactive lipid derivatives, and methods of making and using same |
| CA2734173C (en) * | 2007-10-18 | 2019-04-23 | Mayo Foundation For Medical Education And Research | Igm-mediated receptor clustering |
| WO2009140361A1 (en) * | 2008-05-13 | 2009-11-19 | Genentech, Inc. | ANTI-PirB ANTIBODIES |
| EP3828205A1 (en) * | 2010-10-01 | 2021-06-02 | Oxford BioTherapeutics Ltd | Anti-ror1 antibodies |
| CA2815046C (en) * | 2010-10-19 | 2019-05-14 | Mayo Foundation For Medical Education And Research | Human antibodies and diagnostic and therapeutic uses thereof for the treatment of neurological disease |
| WO2016073963A1 (en) * | 2014-11-07 | 2016-05-12 | Mayo Foundation For Medical Education And Research | Treatment of neonatal hypoxia including impairments or effects thereof |
-
2013
- 2013-04-17 CN CN201810021751.XA patent/CN108285483B/zh not_active Expired - Fee Related
- 2013-04-17 JP JP2015507147A patent/JP6162219B2/ja not_active Expired - Fee Related
- 2013-04-17 WO PCT/US2013/036947 patent/WO2013158748A1/en not_active Ceased
- 2013-04-17 ES ES13778780.0T patent/ES2655077T3/es active Active
- 2013-04-17 US US14/395,141 patent/US20150104460A1/en not_active Abandoned
- 2013-04-17 CA CA2870625A patent/CA2870625C/en active Active
- 2013-04-17 EP EP17188389.5A patent/EP3305323B1/en not_active Not-in-force
- 2013-04-17 NZ NZ701278A patent/NZ701278A/en not_active IP Right Cessation
- 2013-04-17 ES ES17188389T patent/ES2870717T3/es active Active
- 2013-04-17 IN IN2274MUN2014 patent/IN2014MN02274A/en unknown
- 2013-04-17 EP EP13778780.0A patent/EP2838562B1/en not_active Not-in-force
- 2013-04-17 BR BR112014025898A patent/BR112014025898A2/pt not_active Application Discontinuation
- 2013-04-17 MX MX2014012651A patent/MX356347B/es active IP Right Grant
- 2013-04-17 CN CN201380027368.0A patent/CN104379167B/zh not_active Expired - Fee Related
-
2014
- 2014-10-19 IL IL235118A patent/IL235118B/en active IP Right Grant
-
2017
- 2017-04-28 JP JP2017089783A patent/JP2017127329A/ja not_active Withdrawn
-
2019
- 2019-01-04 JP JP2019000205A patent/JP2019050832A/ja not_active Withdrawn
- 2019-05-15 IL IL266658A patent/IL266658B/en active IP Right Grant
-
2020
- 2020-09-23 JP JP2020158408A patent/JP2021000120A/ja not_active Withdrawn
-
2022
- 2022-05-16 JP JP2022080204A patent/JP2022107016A/ja active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| MX356347B (es) | 2018-05-23 |
| JP2021000120A (ja) | 2021-01-07 |
| EP3305323B1 (en) | 2021-02-17 |
| JP2019050832A (ja) | 2019-04-04 |
| NZ701278A (en) | 2016-12-23 |
| IN2014MN02274A (es) | 2015-08-07 |
| CA2870625C (en) | 2023-06-20 |
| HK1258380A1 (zh) | 2019-11-08 |
| ES2655077T3 (es) | 2018-02-16 |
| US20150104460A1 (en) | 2015-04-16 |
| MX2014012651A (es) | 2015-05-07 |
| JP2017127329A (ja) | 2017-07-27 |
| IL266658B (en) | 2021-04-29 |
| IL266658A (en) | 2019-07-31 |
| BR112014025898A2 (pt) | 2017-11-07 |
| EP3305323A1 (en) | 2018-04-11 |
| CN108285483B (zh) | 2022-04-26 |
| HK1206997A1 (en) | 2016-01-22 |
| CA2870625A1 (en) | 2013-10-24 |
| JP2015520608A (ja) | 2015-07-23 |
| CN104379167B (zh) | 2018-02-13 |
| HK1207565A1 (en) | 2016-02-05 |
| EP2838562A1 (en) | 2015-02-25 |
| EP2838562B1 (en) | 2017-08-30 |
| IL235118B (en) | 2019-06-30 |
| IL235118A0 (en) | 2014-12-31 |
| JP2022107016A (ja) | 2022-07-20 |
| EP2838562A4 (en) | 2015-09-30 |
| CN104379167A (zh) | 2015-02-25 |
| CN108285483A (zh) | 2018-07-17 |
| JP6162219B2 (ja) | 2017-07-12 |
| WO2013158748A1 (en) | 2013-10-24 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2870717T3 (es) | Anticuerpos humanos y secuencias de unión específicas del mismo para su uso en accidente cerebrovascular e isquemia y condiciones isquémicas | |
| US20200397898A1 (en) | Human antibodies and diagnostic and therapeutic uses thereof for the treatment of neurological disease | |
| HK1207565B (en) | Human antibodies and specific binding sequences thereof for use in stroke and ischemia or ischemic conditions | |
| HK1258380B (en) | Human antibodies and specific binding sequences thereof for use in stroke and ischemia or ischemic conditions | |
| HK1188731A (en) | Human antibodies and diagnostic and therapeutic uses thereof for the treatment of neurological disease | |
| HK1188731B (en) | Human antibodies and diagnostic and therapeutic uses thereof for the treatment of neurological disease | |
| HK1211593B (zh) | 用於治疗神经疾病的人抗体及其诊断和治疗用途 | |
| HK1206997B (en) | Human antibodies and specific binding sequences thereof for use in stroke and ischemia or ischemic conditions |