JP2004121180A - ポリペプチドの生体内安定化配列及びその利用 - Google Patents
ポリペプチドの生体内安定化配列及びその利用 Download PDFInfo
- Publication number
- JP2004121180A JP2004121180A JP2002294200A JP2002294200A JP2004121180A JP 2004121180 A JP2004121180 A JP 2004121180A JP 2002294200 A JP2002294200 A JP 2002294200A JP 2002294200 A JP2002294200 A JP 2002294200A JP 2004121180 A JP2004121180 A JP 2004121180A
- Authority
- JP
- Japan
- Prior art keywords
- polypeptide
- vivo
- fish
- gonadotropin
- dna
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images








Classifications
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A40/00—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production
- Y02A40/80—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production in fisheries management
- Y02A40/81—Aquaculture, e.g. of fish
Landscapes
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
- Farming Of Fish And Shellfish (AREA)
Abstract
【解決手段】胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるポリペプチド、その変異体、及び該ペプチドをコードする遺伝子を用いて、生体内ポリペプチドの生体内における安定化を図ることよりなる。本発明の方法は、魚類における性腺刺激ホルモンや成長ホルモン等の生体内導入に適用して、魚類の産卵誘発や産卵調節のような成熟調節、或いは魚類の成長調節における投入ポリペプチドの安定化を図る。
【選択図】なし
Description
【発明の属する技術分野】
本発明は、生体内ポリペプチドの生体内での安定化機能を付与するポリペプチド、該ポリペプチドをコードするDNA、及び該ポリペプチド及び該ポリペプチドをコードするDNAを利用した生体内安定化ポリペプチドによる生体内生理機能の調節、特に魚類の成熟調節及び成長調節のような生体内生理機能の調節への利用に関する。
【0002】
【従来の技術】
従来より、ヒトをはじめとして、種々の分野の種々の動物において、生体内に存在するポリペプチドを、生体に投与して、生体の種々の生理機能の調節や、種々の薬理作用を利用した生体の防御などが行われている。
魚類においては、その養殖に際して、性腺刺激ホルモン(生殖腺刺激ホルモン)を用いて、該ポリペプチドを、生体に投与し、魚類の成熟を調節することが行われている。例えば、魚類の養殖における卵の人口孵化に際しては、ウナギやハマチのような通常、産卵を行わない魚に対しては、性腺刺激ホルモンを投与することで、産卵を誘発し、卵や精子を採る方法が行われている。また、マダイやヒラメのような産卵を行う魚に対しては、性腺刺激ホルモンを投入することで、産卵期等を調節し、養殖業者が望む時期に望む量・質の卵や精子を採る方法が行われている。
【0003】
また、養殖している魚に、成長ホルモン(ポリペプチド)を投与して、魚の成長を促進させたり、魚に抵抗力を付与するポリペプチドを投与して、病気に対する耐性を高める方法も知られている。
このように、例えば、魚類の養殖に際しては、種々のポリペプチドの魚体内への投与が行われているが、その場合に、投与されたポリペプチドの生体内での安定性が問題になる。すなわち、魚類の場合には、上記のようにポリペプチドを投与するには、生け簀の中の魚を捕まえ、その魚に性腺刺激ホルモンを注射器で注射することにより行うのが普通である。この場合、魚に注入されたポリペプチドは、魚体内に注入された後、吸収→代謝→分布→排出の過程を経るが、この生体内の代謝の過程で分解が起こる。したがって、通常、魚体内に注射されたポリペプチドの濃度を一定レベル以上に保つためには、ポリペプチドが減衰する前の所定の期間で、連続的に行わなければならない。
【0004】
このような、ポリペプチドの魚体への連続投与の作業は、養殖業者にとって、作業にかかる時間とコストの増大の問題をひき起こすだけでなく、ポリペプチドを投与される魚体自体にとっても、例えば、ポリペプチド投与のために魚が弱って食欲を失ったり、魚の成長が阻害されたり、また魚が死亡したりするような、魚への負荷がかかり、大きな影響が及ぼされる。これらの問題を解決するために、養殖業者の間では、魚体内に投与したポリペプチドの延命化(生体内での半減期の延長効果)を図る方策が期待されている。
【0005】
上記のように、例えば、魚類の養殖の分野では、性腺刺激ホルモンや成長ホルモンのようなポリペプチドを魚体内に投与することが行われており、そのような場合に、投与したポリペプチドの安定性の問題が提起されているが、一般に、生体内にポリペプチドを投与した場合には、その生体内におけるポリペプチドの分解の問題がある。そこで、投与したポリペプチドの本来の効果を有効に利用できるようにするためには、投与したポリペプチドの生体内での安定化を図ることが必要である。したがって、生体内に投与したポリペプチドの安定化は、ペプチドを生体機能の調整に用いる分野の一般的な課題でもある。
【0006】
【特許文献1】
特開平10−36285号公報
【特許文献2】
特開平10−36398号公報
【特許文献3】
特開平10−36399号公報
【特許文献4】
WO98/21238
【非特許文献1】
Mol Endocrinol 6, 951−959, 1992
【0007】
【発明が解決しようとする課題】
本発明の課題は、ホルモンのような生体内ポリペプチドの生体内での安定化機能を付与するポリペプチド、該ポリペプチドをコードするDNA、及び該ポリペプチド及び該ポリペプチドをコードするDNAを利用した生体内安定化ポリペプチによる生体内生理機能の調節方法を提供すること、及び特別には、生体内でのポリペプチド安定化機能を付与するポリペプチド及び該ポリペプチドをコードするDNAを利用した魚類の成熟調節及び成長調節のような生体内生理機能の調節を行う方法を提供することにある。
【0008】
【課題を解決するための手段】
通常、霊長類やウマは、(1)胎盤から分泌される胎盤性性腺刺激ホルモン(絨毛性ゴナドトロピン)(chorionic gonadotropin:CG)、(2)下垂体から分泌される濾胞刺激ホルモン(follicle stimulating hormone:FSH)及び(3)黄体形成ホルモン(luteinizing hormone:LH)の3種の性腺刺激ホルモンを持つ。これらのホルモンのうち、CGは、FSH及びLHと比較してホルモンの半減期が長いことが知られている。本発明者は、この原因究明のため、ホルモンの構造に着目して研究したところ、CGには、FSH及びLHには存在しないカルボキシ末端ペプチド(CTP)が付加していることが確認された。このカルボキシ末端ペプチド(CTP)が、ホルモンの半減期の延長に関与するかどうか調べた結果、CTPを、FSH及びLHに付加したものは、従来のFSH及びLHに比較して、ホルモンの半減期が著しく延長することが判った。また、FSH及びLHの本来のホルモンの活性には変化はないことが判った。
【0009】
本発明は、上記知見に基づいてなされたもので、胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるポリペプチドを、ホルモンのような生体内ポリペプチドの生体内安定化機能を付与するポリペプチドとして用いて、生体内におけるポリペプチドの安定化を図ることよりなるものである。本発明は、また、生体内安定化機能を付与するポリペプチドをコードするDNA、該ポリペプチド及びDNAの変異体、及び生体内ポリペプチドと生体内安定化機能を付与するポリペプチドとを結合した生体内安定化ポリペプチドからなる。
【0010】
更に、本発明は、生体内ポリペプチドの生体内安定化機能を付与するポリペプチド及び該ポリペプチドをコードするDNAを、魚類における性腺刺激ホルモンや成長ホルモン等の生体内導入に適用して、魚類の産卵誘発や産卵調節のような成熟調節、或いは魚類の成長調節等を行う方法を含むものである。本発明の生体内ポリペプチドの生体内安定化機能を付与するポリペプチドのアミノ酸配列は、配列表の配列番号2に、該ポリペプチドをコードする塩基配列は、配列表の配列番号1に、例示される。
【0011】
すなわち本発明は、胎盤性性腺刺激ホルモンのカルボキシ末端領域からなる、生体内ポリペプチドの生体内安定化機能を付与するポリペプチドをコードするcDNA(請求項1)や、配列表の配列番号1に示される塩基配列からなる、請求項1記載の生体内ポリペプチドの生体内安定化機能を付与するポリペプチドをコードするDNA(請求項2)や、以下の(a)又は(b)のポリペプチドをコードするDNA、すなわち、(a)配列表の配列番号2に示されるアミノ酸配列からなるポリペプチド、又は(b)配列表の配列番号2に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ生体内ポリペプチドの生体内安定化機能を付与するポリペプチド(請求項3)や、請求項2記載のDNAとストリンジェントな条件下でハイブリダイズし、かつ生体内ポリペプチドの生体内安定化機能を付与するポリペプチドをコードすることを特徴とするDNA(請求項4)からなる。
【0012】
また本発明は、配列表の配列番号2に示されるアミノ酸配列からなることを特徴とする生体内ポリペプチドの生体内安定化機能を付与するポリペプチド(請求項5)や、配列表の配列番号2に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ生体内ポリペプチドの生体内安定化機能を付与することを特徴とするポリペプチド(請求項6)や、生体内ポリペプチドが、性腺刺激ホルモン又は成長ホルモンであることを特徴とする請求項1〜6のいずれか記載のDNA又はポリペプチド(請求項7)や、生体内ポリペプチドのカルボキシ末端側に、請求項5又は6記載のポリペプチドを結合したことを特徴とする生体内安定化ポリペプチド(請求項8)や、生体内ポリペプチドが、性腺刺激ホルモン又は成長ホルモンであることを特徴とする請求項8記載の生体内安定化ポリペプチド(請求項9)や、サブユニット構造を備えた生体内ポリペプチドの一方のサブユニットのカルボキシ末端側に、胎盤性性腺刺激ホルモンのカルボキシ末端領域ペプチドを結合したことを特徴とする請求項8又は9記載の生体内安定化ポリペプチド(請求項10)や、魚類の生体内ポリペプチドのカルボキシ末端側に、胎盤性性腺刺激ホルモンのカルボキシ末端領域ペプチドを結合したことを特徴とする請求項8〜10のいずれか記載の生体内安定化ポリペプチド(請求項11)や、魚類の生体内ポリペプチドが、性腺刺激ホルモン又は成長ホルモンであることを特徴とする請求項11記載の生体内安定化ポリペプチド(請求項12)からなる。
【0013】
さらに本発明は、請求項12記載の生体内安定化性腺刺激ホルモンを魚類に導入することを特徴とする魚類の成熟調節方法(請求項13)や、魚類の成熟調節が、魚類の産卵誘発或いは産卵調節であることを特徴とする請求項13記載の魚類の成熟調節方法(請求項14)や、請求項12記載の生体内安定化成長ホルモンを魚類に導入することを特徴とする魚類の成長調節方法(請求項15)や、生体内ポリペプチドをコードするDNAの3´末端に、請求項1〜4のいずれか記載のDNAを結合した遺伝子を、生体内で発現することを特徴とする生体内発現ポリペプチドの生体内安定化方法(請求項16)や、魚類で発現するポリペプチドをコードするDNAの3´末端に、請求項1〜4のいずれか記載のDNAを結合した遺伝子を、魚類生体内で発現することを特徴とする請求項16記載の生体内発現ポリペプチドの生体内安定化方法(請求項17)や、魚類で発現する性腺刺激ホルモンのポリペプチドをコードするDNAの3´末端に、胎盤性性腺刺激ホルモンのカルボキシ末端領域ペプチドをコードするDNAを結合した遺伝子を、魚類生体内で発現することを特徴とする魚類の成熟調節方法(請求項18)や、魚類の成熟調節が、魚類の産卵誘発或いは産卵調節であることを特徴とする請求項18記載の魚類の成熟調節方法(請求項19)や、魚類で発現する成長ホルモンのポリペプチドをコードするDNAの3´末端に、胎盤性性腺刺激ホルモンのカルボキシ末端領域ペプチドをコードするDNAを結合した遺伝子を、魚類生体内で発現することを特徴とする魚類の成長調節方法(請求項20)からなる。
【0014】
【発明の実施の形態】
本発明は、胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるペプチドを、ホルモンのような生体内ポリペプチドのカルボキシ末端に結合して、該生体内ポリペプチドに生体内安定化機能を付与することよりなる。胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるポリペプチドのアミノ酸配列は、配列表の配列番号2で示され、胎盤性性腺刺激ホルモンのカルボキシ末端35アミノ酸残基からなる。また、本発明は、該胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるペプチドをコードするcDNAからなる。該cDNAの塩基配列は、配列表の配列番号1で示され、胎盤性性腺刺激ホルモンのカルボキシ末端35アミノ酸残基をコードする105の塩基からなる。
【0015】
更に、本発明は、上記胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるペプチドのペプチド変異体、及び該胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるペプチドをコードするcDNAのDNA変異体を含む。DNA変異体としては、配列番号2に示されるアミノ酸配列からなる生体内安定化機能を付与するポリペプチドをコードするDNAや、配列番号2に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなる生体内安定化機能を付与するポリペプチドをコードするDNAを挙げることができ、これらの胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるペプチドをコードするcDNAやその変異体は、そのDNA配列情報等に基づき、例えばヒトやウマの遺伝子ライブラリーやcDNAライブラリーなどから公知の方法により調製することができ、また、そのDNA変異体は、公知の遺伝子操作の方法により、そのDNA配列を変更して、調製することができる。
【0016】
また、本発明の対象となる生体内安定化機能を付与するポリペプチドをコードするDNAとしては、配列番号1に示されるDNAとストリンジェントな条件下でハイブリダイズし、かつ生体内安定化機能を付与するポリペプチドをコードするDNAを挙げることができる。これらは例えば、ヒトやウマ由来のDNAライブラリーに対してストリンジェントな条件下でハイブリダイゼーションを行うことにより、取得することができる。かかるDNAを取得するためのハイブリダイゼーションの条件としては、例えば、42℃でのハイブリダイゼーション、及び1×SSC、0.1%のSDSを含む緩衝液による42℃での洗浄処理を挙げることができ、65℃でのハイブリダイゼーション、及び0.1×SSC,0.1%のSDSを含む緩衝液による65℃での洗浄処理をより好ましく挙げることができる。
【0017】
本発明の胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるペプチドのペプチド変異体としては、配列番号2に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなる生体内安定化機能を付与するポリペプチドを挙げることができる。かかるポリペプチドは、該ポリペプチドをコードするDNA配列を用い、そのDNA配列情報等に基づき公知の遺伝子工学的方法により調製することができる。
【0018】
本発明の生体内安定化機能を付与するポリペプチドを用いて生体内ポリペプチドの安定化を図るには、例えば、性腺刺激ホルモンや成長ホルモンのような生体内ポリペプチドをコードするDNA配列の3´末端に、本発明の生体内安定化機能を付与するポリペプチドをコードするDNA配列を結合し、該遺伝子を公知の真核生物を用いた遺伝子の発現系を用いて発現させて、生体内ポリペプチドのカルボキシ末端に本発明の生体内安定化機能を付与するポリペプチドを結合したポリペプチドを取得する。該生体内安定化機能を付与するポリペプチドを結合したポリペプチドの形で、ポリペプチドを生体内へ投与することにより、生体内ポリペプチドの生体内における安定化を図ることができる。
【0019】
また、本発明の生体内安定化機能を付与するポリペプチドを用いて生体内ポリペプチドの安定化を図る方法としては、生体内ポリペプチドをコードするDNA配列の3´末端に、本発明の生体内安定化機能を付与するポリペプチドをコードするDNA配列を結合し、該遺伝子を生体内に導入し、発現させて、安定化したポリペプチドとして利用することができる。
【0020】
本発明の生体内安定化機能を付与したポリペプチドをコードする遺伝子を生体内へ導入するには、公知の動物生体組織や細胞内への遺伝子を導入する方法を使用することができる。例えば、ガラス針を動物細胞又は組織細胞にさして該細胞内導入物質を注入するマイクロインジェクション法(発生工学実験マニュアル、講談社)や、電気パルスを印加することによって細胞膜に穴を開け、遺伝子を動物生体組織や細胞内に導入するエレクトロポーレーション法のような方法を用いることができる(EMBO J. 1, 841−845 ,1982、Gene 96, 23−28, 1990)。これらの生体内ポリペプチドをコードする遺伝子を生体内へ導入する方法において、通常は、その操作が直接的であり、かつ簡便であることから、通常マイクロインジェクション法が便利に利用することができる。
【0021】
更に、本発明は、生体内ポリペプチドの生体内安定化機能を付与するポリペプチド及び該ポリペプチドをコードするDNAを、魚類における性腺刺激ホルモンや成長ホルモン等の生体内導入に適用して、魚類の産卵誘発や産卵調節のような成熟調節、或いは魚類の成長調節等を行う方法を含む。本発明の生体内ポリペプチドの生体内安定化機能を付与するポリペプチドを用いて、魚類の産卵誘発や産卵調節のような成熟調節、或いは魚類の成長調節等を行うには、例えば性腺刺激ホルモンや成長ホルモン等のアミノ酸配列をコードするDNAの3´末端に、配列表の配列番号1に示されるDNA配列を結合した遺伝子を、魚類の受精卵へ注入することにより、生体内へ該遺伝子を導入して行うことができる。
【0022】
また、上記のような生体内ポリペプチドとポリペプチドの生体内安定化機能を付与するポリペプチドを結合したポリペプチドをコードする遺伝子を、別途、遺伝子発現系を用いて発現させ、該結合ポリペプチドを取得して、それを魚類の生体内へ導入して、魚類の産卵誘発や産卵調節のような成熟調節や、或いは魚類の成長調節等を行うことができる。従来、魚類の養殖に際しては、種々のポリペプチドを魚体内へ投与することが行われており、その場合に、投与されたポリペプチドの生体内での安定性が問題になっていたが、本発明の方法を用いることにより、ポリペプチドの生体内での安定化を図ることが可能となり、養殖業者にとって、魚体へのポリペプチドの繰り返し投与を行う必要がなくなるため、作業にかかる時間とコストの増大の問題を解消する。それと共に、ポリペプチドの生体内での効果の持続を図ることが可能となり、かつ魚体自体にとっても、その投与に対する魚への負荷の軽減を図ることが可能となる。
【0023】
本発明の方法が適用される生体内ポリペプチドとしては、特に限定されないが、上記のように、αサブユニットとβサブユニットを備えた生体内ポリペプチドである性腺刺激ホルモンや成長ホルモンのような、生体内へ投与されるホルモンが挙げられる。
従来、魚類の産卵誘発や産卵調節のような成熟調節に際して魚の生体内へ投与されていた濾胞刺激ホルモン(FSH)や、黄体形成ホルモン(LH)は、生体内に投与した場合に、比較的短いホルモンの半減期しか得られなかったが、本発明の安定化機能を付与するペプチドを用いることによって、その半減期を著しく延長することが可能となる。
【0024】
【実施例】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
[材料と方法]
(1本鎖GtH発現コンストラクト・pB−SCF、pB−SCL+の構築)
タンパク質をコードする領域として、小林ら(Gen Comp Endocrinol 105, 372−378, 1997)及び吉浦ら(Gen Comp Endocrinol 105, 3, 379−389, 1997)によってクローン化されたキンギョ(goldfish)性腺刺激ホルモン(GtH)α鎖(以下、「α1」という。)のcDNA配列(配列表の配列番号3)、及び、濾胞刺激ホルモン(FSH)β鎖(以下、「Fβ」という。)(配列表の配列番号5)、若しくは、黄体形成ホルモン(LH)β鎖(以下、「Lβ」という。)(配列表の配列番号7)の、いずれか1種類のβ鎖のcDNA配列を用いた。
ここで、キンギョGtHα鎖は、小林ら(Gen Comp Endocrinol 105, 372−378, 1997)により、脳下垂体からα1とα2の2種類のcDNA配列がクローン化されているが、α1が常に優位に発現していることから、本発明においてはα1を用いることとした。
【0025】
β−アクチンは生物の細胞骨格を構成する主要なタンパク質で、その遺伝子は種を通じてよく保存されているうえ、いずれの種においてもその発現量が極めて高いことが知られており、そのメダカβ−アクチン遺伝子プロモーター/エンハンサー(以下、「β−アクチンP/E配列」という。)はメダカにおいて外来遺伝子の発現を組織非特異的に強く促すことが示されている(Mol Mar Biol Biotechnol 3, 192−199, 1998、Mol Mar Biol Biotechnol 7, 173−180, 1998)ことから、本発明においては、遺伝子発現を制御する領域として、メダカβ−アクチンP/E配列(Mol Mar Biol Biotechnol 3, 192−199, 1998)を用いることとした。また、mRNAを安定化させる目的で、発現コンストラクトにはGtH−cDNA配列の下流に、ウシ成長ホルモン遺伝子ポリA付加シグナル(BGH polyA配列)を組み込んだ。
【0026】
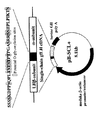
1本鎖GtH発現コンストラクトは、PCR法により作製した。すなわち、タンパク質をコードする領域として、キンギョβ及びα各鎖cDNAを融合し、更に、Minら(J Reprod Dev 40, 301−305, 1994)によってクローン化されているウマ黄体形成ホルモン/絨毛性ゴナドトロピン(eLH/CG、ウマではLHβ鎖遺伝子とCGβ遺伝子が同一であるため、「LH/CG」と表記する。)cDNA(配列表の配列番号9)由来のC末端ペプチド領域(eCTP;5’− tcctcttcctctaaggatcccccatcccaacctctcacatccacatccaccccaactcctggggccagcagacgttcctctcatcccctcccaataaagacttct −3’:配列表の配列番号1)、アミノ酸35残基分(配列表の配列番号2)をコードしている。以下、「CTP」と表記する。)のcDNA配列を挿入した1本鎖FSH−cDNA(以下、「SCF」と表記する。)(配列表の配列番号11)及び1本鎖LH−cDNA(以下、「SCL+」と表記する。)(配列表の配列番号13)を用いた(図1)。そして、遺伝子発現を制御する領域としてのメダカβ−アクチンP/E配列及びBGH polyAを、pBluescriptII SK+に組み込んだもの(以下、「pB−SCF」、及び、「pB−SCL+」と表記する。)を用いた(図2及び図3)。
【0027】
ここで、α鎖、β鎖cDNAのタンパク質コード領域を一致させて融合することにより、α鎖サブユニットとβ鎖サブユニットの会合効率を上昇させることができるため、遺伝子導入細胞内において組換えGtH分子が効率的に正確な立体構造をとり、生理活性を有するGtHが生産される。α鎖とβ鎖の連結部分には、CTPのcDNAを挿入した。従来、CTPのcDNA配列は、α鎖及びβ鎖の立体構造及びin vitroにおける生物活性を変化させることなく連結できる配列として、1本鎖GtH生産の際一般的に用いられてきた。したがって、本発明の実施例においても、LH/CG由来のCTP−cDNA配列をα鎖とβ鎖の連結配列として利用した。
【0028】
上記pB−SCFを構築するため、クローニングベクター・pBluescriptII SK+又はpGEM T−EasyにクローニングされているキンギョGtH各サブユニット(α鎖サブユニット:α1、及びβ鎖サブユニット:Fβ)及びpUC119にクローニングされているCTPのcDNA配列をPCR法によって増幅し(得られたPCR増幅産物;Fβは440bp、α1は670bp、eCTPは130bp)、SCF断片を作製した。
GtHサブユニット及びCTPを増幅するため、Fβ鎖を増幅するプライマーとしてP−1及びP−2を、CTPを増幅するプライマーとしてP−3及びP−4を、α1鎖を増幅するプライマーとしてP−5及びP−6をそれぞれ設計し、PCRの反応に用いた(表1及び図4)。その際、P−2は5´側にCTPのDNA配列を、P−3は5´側にFβ鎖のDNA配列を、P−4は5´側にα1鎖のDNA配列を、P−5は5´側にCTPの配列をそれぞれ含むように設計した(図4)。
【0029】
【表1】

まず、P−1とP−2、P−3とP−4、P−5とP−6を用いてPCRを行い、それぞれFβ鎖、CTP及びα1鎖を増幅した(図4:反応1、反応2、反応3)。PCRはGtHα1、Fβ、CTPそれぞれのcDNAが組み込まれたpGEM T−Easy、pBluescriptII SK+(約0.5ng)、又はpUC119を鋳型として、1μLの10×LA PCR buffer、1μMの各サブユニットに特異的なプライマー、400μM dNTP solution、2.5mM MgCl2、2.5unitのLA Taq polymeraseを含む10μLの反応混合液中で行った。PCRの反応サイクルは一連の反応(94℃で30秒間熱変性させ、60℃で30秒間アニーリングし、72℃で30秒間伸長反応させる)を30回繰り返して行った。
【0031】
次に、P−1とP−4を用いたPCRによりDNA断片FβとCTPを融合し、Fβ−CTP(570bp)を作製した(図4:反応4)。PCRの反応条件としては、反応1で得たFβ鎖断片0.5μLと、反応2で得たCTP0.05μLを鋳型として、LA Taq polymeraseを含む上述と同様の反応混合液中で行った。PCRの反応サイクルは一連の反応(94℃で30秒間熱変性させ、60℃で30秒間アニーリングし、72℃で40秒間伸長反応させる)を30回繰り返して行った。更に、P−1とP−6を用いたPCRによりDNA断片Fβ−CTPとα1鎖を融合し、1本鎖Fβ−CTP−α1(SCF;1240bp)を作製した(図4:反応5)。PCRの反応条件としては、Fβ−CTP断片0.5μLとα1断片0.5μLを鋳型として、LA Taq polymeraseを含む上述の反応混合液中で行った。PCRの反応サイクルは一連の反応(94℃で30秒間熱変性させ、65℃で30秒間アニーリングし、72℃で60秒間伸長反応させる)を30回繰り返して行った。PCRによる増幅産物は制限酵素Sal Iで消化し、ベクターの構築に用いた。
【0032】
pB−SCL+を構築するため、キンギョGtH各サブユニット(α鎖サブユニット:α1、及びβ鎖サブユニット:Lβ)及びCTPのcDNA配列をPCR法によって増幅し(得られたPCR増幅産物;Lβは440bp、α1は670bp、eCTPは130bp)、SCL+断片を作製した。
GtHサブユニット及びCTPを増幅するため、LHβ鎖を増幅するプライマーとしてP−1´及びP−2´を、CTPを増幅するプライマーとしてP−3´をそれぞれ設計した(表1及び図4)。その際、P−2´は5´側にCTPのDNA配列を、P−3´は5´側にLβ鎖のDNA配列を含むように設計した(図4)。
なお、CTPの3´側プライマーであるP−4、α1鎖を増幅するP−5及びP−6はSCF作製の際に用いたプライマーと同一のものを用いた(表1)。以上のプライマーを用いてSCF作製の際と同様の条件でPCR反応を行い、ベクター構築を行った(図4)。
【0033】
β−アクチンP/E配列(約3800bp)については、まずβ−アクチンP/E配列に特異的なプライマー(表1)を用いてPCRを行い、目的のDNA断片を増幅すると同時にその両端に制限酵素Eco RI(TaKaRa社製)の認識配列を付加した。PCR反応はpOBAを鋳型として10×Ex PCR buffer 1μL、1μMの各プライマー、200μMの各dNTP solution、1.25unitのEx Taq polymerase(TaKaRa社製)を含む50μLの混合液中で行った。
PCRの反応サイクルは一連の反応(93℃で45秒間熱変性させ、60℃で60秒間アニーリングし、72℃で5分間伸長反応させる)を30回繰り返して行った。PCRで増幅されたDNA断片は電気泳動で展開後、回収してEco RIで消化した。
BGH polyA(280bp)は、Rc/RSVベクター(Invitrogen社製)から制限酵素Xho I(TaKaRa社製)で切り離し、ベクター構築に用いた。
【0034】
それぞれのDNA配列をpBluescriptII SK+ベクターの制限酵素クローニングサイト、即ちメダカβアクチンP/E遺伝子配列をEco RIサイトへ、1本鎖GtH−cDNA(SCF又はSCL+)配列をSal Iサイトへ、BGH polyAをXho Iサイトへ、それぞれライゲーション反応によって組み込み、2種類の発現コンストラクト・pB−SCF及びpB−SCL+を構築した(図5)。
また、pB−SCL+についてはCTPの効果を確認するためにpB−SCL+からCTP配列を除去したpB−SCL−も同時に構築した。これらの発現コンストラクトはマイクロインジェクションに用いるためTEバッファーに溶解し、50μg/mLの濃度に調整した。
【0035】
(マイクロインジェクション法による外来遺伝子の導入)
マイクロインジェクションには、東京水産大学大泉実験実習場で飼育され成熟した3才又は4才のニジマス(Oncorhynchus mykiss)雌から卵を採取し、1才のニジマス雄から搾出した精子を用いて媒精した。その後、卵膜硬化抑制のため1mM還元型グルタチオン溶液(pH8.0)中にて10℃で培養したものを用いた(Nippon Suisan Gakkaishi 55, 369, 1989)。マイクロインジェクションは受精後1時間から4時間の間にマイクロマニピュレーター(ナリシゲ化学社製)を用いて行った。ニジマス受精卵を等張液(NaCl 9.04g、KCl 0.24g、CaCl2・2H2O 0.34g/L)中に微針で固定し、DNA溶液2nLをマイクロピペットによって受精卵の胚盤へ注入した(Nippon Suisan Gakkaishi 57, 819−824, 1991)。これらの作業は、室温をニジマス飼育水の温度である10℃に保った状態で行った。
【0036】
(遺伝子導入ニジマス初期胚を用いたウエスタン・ブロット解析)
pB−SCL+を導入した後、4日経過したニジマス胚(pB−SCL+導入ニジマス初期胚)とキンギョ脳下垂体から全タンパク質を抽出しウエスタン・ブロット法による解析を行った。pB−SCL+導入ニジマス初期胚の胚盤を剥がし10個の胚をプールしてpB−SCL+導入区として3サンプルをサンプルバッファー(50mM Tris−HCl(pH6.8)、2%SDS、5%β−メルカプトエタノール、10%グリセロール)中でホモジナイズし、タンパク質を抽出した。一方、キンギョ雌1尾(BW 81.5g、GSI 11.7%)から脳下垂体を摘出し、上記ニジマス初期胚の際と同様の方法によりタンパク質の抽出を行った。
【0037】
pB−SCL+導入ニジマス初期胚及びキンギョ脳下垂体から抽出したタンパク質のうち、それぞれ1個分及び100分の1個分を97℃で5分間熱処理を施した後、アクリルアミド濃度15%のゲルを用いてSDS−ポリアクリルアミドゲル電気泳動(SDS−PAGE)を行った。
SDS−PAGEによってタンパク質を分子量の差で展開した後、アクリルアミドゲル中のタンパク質をミニバッファートランスファー装置(BIOCRAFT社製)を用いてHybond ECL nitrocellulose membrane(Pharmacia社製)へ転写した。メンブレンフィルターは、小林らによって作製された抗キンギョ合成α1ペプチド抗体又は抗キンギョLHβ鎖抗体とともに室温で1.5時間インキュベートした。
【0038】
その後、メンブレンフィルターをPBS−T(80mM Na2HPO4、20mM NaH2PO4、100mM NaCl、0.1% Tween20)で洗浄し、horseradish peroxidase−conjugated goat anti−rabbit IgG antibody(Pharmacia社製)とともに1時間室温でインキュベートし、この抗体の結合をECL Western blotting detecting reagents(Pharmacia社製)によって検出した。ゲルからメンブレンへのタンパク質の転写、メンブレンの抗体とのインキュベーション、PBS−Tでの洗浄、タンパク質の検出は、ECL Western Blotting detecting reagentsに添付のプロトコールに従って行った。
また、pB−SCF導入ニジマス初期胚についても、上記pB−SCL+導入ニジマス初期胚と同様の手法でタンパク質を抽出し、ウエスタン・ブロット法による解析を行った。
【0039】
(キンギョ生殖腺を用いた生物検定)
pB−SCF導入ニジマス初期胚から剥離した胚盤25粒を回収し、培養培地中でホモジナイズしたものをサンプルとし、それぞれ2サンプルずつ用意した。その後、キンギョ雌2尾(平均BW 24.6g、平均GSI 0.74%)から卵巣を摘出した。摘出した卵巣は培養培地中で小片に切り分け、1区あたり約30mgを用いた。培養培地としては、ストレプトマイシン100mg/l、ペニシリン60mg/l、HEPES 2.5g/lを加え、2N NaOHでpH7.6に調整したLeibovitz’s L−15 Medium(L−15培地;GIBCO社製)0.5mlにニジマス初期胚25粒分のホモジネートを加えたものを用いた。
【0040】
実験区としては、培地にニジマス初期胚を加えず、L−15培地のみで精巣を培養した「培地のみ」、遺伝子を導入しなかったニジマス胚を加えた「野生型」、pB−SCF導入ニジマス胚を加えた「SCF」及びニジマス初期胚を加えず、hCG(三共株式会社製)5IUを添加した「hCG」の4区を設け、それぞれ2連の実験で行った。キンギョ卵巣片の培養は20℃で20時間行った。卵巣片を培養後、L−15培地0.5mlを回収し、エストラジオール(E2)量の測定を行った。E2は卵胞顆粒膜細胞から分泌されるホルモンであり、体内でテストステロンから合成され、卵子形成を促進する雌性ステロイドホルモンである。したがって本研究ではE2量を測定することにより、卵胞機能を正確に把握する。つまりニジマス初期胚内において産生された組換えタンパク質がin vitroにおけるGtHとしての活性の有無を判定した。
【0041】
E2量の測定は、Aidaら(Jpn. Soc. Sci. Fish. 50, 565−571, 1984)による放射免疫測定法(radioimmunoassay;以下、「RIA法」という。)を用いて行った。まず、回収した培地400μLからジエチルエーテルによって3回のステロイドホルモン抽出を行い、40℃の恒温槽中においてジエチルエーテル画分を乾固させた。そこに200μlの0.1%Gel−PBS溶液(Gelatin−10mM PBS(pH7.5)、140mM NaCl)を加えて攪拌し、ステロイドホルモンを再溶解した。次に[3H]テストステロン(アマシャムバイオサイエンス社製)100μl(約10000cpm)、及び抗エストラジオール抗体(COSMO BIO社製)200μlを加え、抗原抗体反応を進行させるため4℃で12時間放置した。
【0042】
続いて、デキストランチャーコール溶液(デキストラン(Sigma社製)5g/l PBS、チャーコール(Norit“Plus”社製)0.5g/l PBS)を250μl加えた後、3000rpmで20分間、冷却遠心を行い、抗体と結合したステロイドホルモン(bound)と結合していないステロイドホルモン(free)を分離した。さらに、boundのみを含む上清にシンチレーター(オムニフロー;Packard社製)3mLを加え、攪拌後12時間放置した。最後に、液体シンチレーションカウンター(liquid scintillation analyzer 1600TR;Packard社製)を用いて5分間放射活性を測定した。測定した値と濃度既知のスタンダード(60〜3840pg/mL)より得られた標準曲線からL−15培地中に分泌されたE2量をキンギョ卵巣1mg当たりの量に換算後、算出した。
【0043】
pB−SCL+導入ニジマス初期胚から剥離した胚盤25粒を回収し、培養培地中でホモジナイズしたものをサンプルとし、それぞれ2サンプルずつ用意した。その後、キンギョ雄2尾(平均BW 16.0g、平均GSI 1.1%)から精巣を摘出し、摘出した精巣を培養培地中で小片に切り分け、1区あたり約30mgを用いた。また、培養培地はpB−SCF導入ニジマス初期胚を用いた研究と同様のものを使用した。実験区はpB−SCL+導入ニジマス胚を加えたものを「SCL+」とし、その他3区はpB−SCF導入ニジマス初期胚を用いた研究と同様に設け4区とし、2連の実験を行った。キンギョの精巣片の培養はキンギョ卵巣片の培養と同様の手法で行った。
【0044】
精巣片を培養後、L−15培地0.5mLを回収し、テストステロン(T)量の測定を行った。Tは精子形成を促進する雄性ステロイドホルモンである11−ケトテストステロンの前駆ホルモンで、GtHの作用によって精巣内で多量に産生されることが知られている(長浜 1991 生殖:配偶子形成の制御機構.“魚類生理学” 板沢 靖男・羽生 功 編.pp.243−286.恒星社厚生閣,東京. p20)。したがって、本研究ではT量を測定することにより、ニジマス初期胚内において産生された組換えタンパク質がin vitroにおけるGtHとしての活性の有無を判定した。
測定方法は、pB−SCF導入ニジマス初期胚を用いた場合と同様の手法であるRIA法を採用した。測定後、測定値と濃度既知のスタンダード(60〜3840pg/ml)より得られた標準曲線からL−15培地中に分泌されたT量をキンギョ精巣1mg当たりの量に換算後、算出した。
【0045】
(キンギョ個体を用いた生物検定)
pB−SCL+及びCTPを含まないpB−SCL−導入ニジマス胚を淡水魚用リンゲル液(0.75% NaCl、0.02% CaCl2、0.2% KCl)中でホモジナイズし、体重1g当たり20粒の胚盤に相当する量を雄キンギョ(平均体重5.0g)の腹腔内に注射した。注射後キンギョは水温20℃の循環濾過水槽(60L)に収容した。注射後6時間目、12時間目及び24時間目に尾柄部から約50μLの血液を採取した。採取した血液を4℃、3000rpmにて15分間遠心分離後、上清を用いて血中テストステロン量を測定した。テストステロン量の測定には前述のRIA法を用いた。なお、これらの測定は3個体のキンギョを用いて行った。
【0046】
[実験結果]
(ニジマス初期胚におけるキンギョ1本鎖LHの発現)
キンギョ1本鎖GtH遺伝子発現コンストラクト・pB−SCL+導入初期胚より抽出したタンパク質をサンプルとし、ウエスタン・ブロット法によって抗キンギョ合成α1鎖又は抗組換えLβ鎖抗体と反応するタンパク質を検出した。その結果、キンギョ脳下垂体のα1鎖及びLβ鎖と同様の抗原性を示す分子量約36kDaのタンパク質が検出された。この分子量は本発現コンストラクトから産生されるタンパク質に糖鎖が付加した場合の予想分子量と一致したことから、ニジマス初期胚内においてキンギョ1本鎖LHが産生されたことを示している(図6)。
【0047】
(遺伝子導入ニジマス初期胚で生産した組換えタンパク質の生物活性)
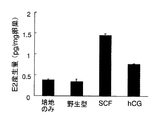
pB−SCF導入ニジマス初期胚ホモジネートを添加した培養液中でキンギョ卵巣を培養し、キンギョ卵巣が培養液中に産生したE2量をRIA法により測定した。その結果、「hCG」区、及び「SCF」区においてE2量の増加が見られた(図7)。また、「SCF」区のエストラジオール量の平均値は、「培養のみ」区、及び「野生型」区の約4倍の1.4pg/mg卵巣片であった。
pB−SCL+導入ニジマス初期胚ホモジネートを添加した培養液中でキンギョ精巣を培養し、キンギョ精巣が培養液中に産生したT量をRIA法により測定した。その結果、「hCG」区、及び「SCL+」区においてT量の増加が見られた(図8)。また、「SCL+」区のテストステロン量の平均値は、「培養のみ」区、及び「野生型」区の約5倍の4.6pg/mg精巣片であった。
【0048】
(キンギョ個体内で生産した組換えタンパク質の生物活性)
キンギョ個体内にpB−SCL+及びpB−SCL−導入ニジマス初期胚ホモジネートを注射し、注射後12時間目の各区における血中T量の結果を図9に示した。「SCL+」区及び「SCL−」区とも有意に血中T量の上昇を促したものの、「SCL+」区では「SCL−」区の約3倍ほど血中T量が高かった。
【0049】
(キンギョ個体内で生産した組換えタンパク質の経時変化)
キンギョ個体内にpB−SCL+及びpB−SCL−導入ニジマス初期胚ホモジネートを注射した場合の、注射後の血中テストステロン量の経時変化を図10に示す。
注射後6時間目には、両区とも同程度の血中テストステロン量を示した。注射後12時間目には、両区とも血中テストステロン量の減少が見られたが、減少量は「SCL−」区が「SCL+」区より3倍程度大きかった。さらに注射後24時間目にも注射後12時間目と同様の減少量の差が顕著に見られた。このことから、pB−SCL+がキンギョ個体内において長期間安定的に効力を発揮したと考えられる。
【0050】
[評価]
本発明においては、1本鎖GtH−cDNA作製の際、α鎖とβ鎖の間に、Minら(J Reprod Dev 40, 301−305, 1994)によってクローン化されたLH/CGβ鎖由来のCTP配列を挿入した。Galetら(J Endocrinol 167, 117−124, 2000)によってLH/CGβ鎖のCTP配列の有無で、組換えLH/CGのin vitroにおける生物活性に変化はないことが示されているが、hCGのCTP配列を用いた研究から、CTP配列がin vivoにおいてGtH分子の半減期を延長する役割を持つことが示唆されている(Proc Natl Acad Sci USA 89, 4304−4308, 1992)。これは、LH、FSHやCGは、分子量の約3割が付加されている糖鎖によるものであることから、これらのホルモンを比較した場合、生体内でのホルモンの寿命が長いCGのカルボキシ末端ポリペプチド(CTP)に特異的に結合している糖鎖によるものと推測される。
【0051】
従って、このようなCTP配列を付加した(付加型)LHやFSHを用いて魚類の催熟を行った場合、親魚体内に投与した付加型LH及び付加型FSHは、これら性腺刺激ホルモン(GtH)が長期間体内に留まることとなり、その催熟効率が上昇する。上記のことから、本発明による発現系で産生された1本鎖LHに含まれるeLH/CGβ鎖由来のCTP配列も組換えGtH分子への半減期延長効果を有することが示される。
【0052】
【発明の効果】
本発明により、胎盤性性腺刺激ホルモンのカルボキシ末端領域からなるポリペプチド、その変異体、或いは該ポリペプチドをコードする遺伝子を用いて、ホルモンのような生体内ポリペプチドの生体内安定化を図ることにより、広く、生体に投与されるポリペプチド薬剤の生体内での安定化を図り、その効果を飛躍的に増大することを可能とする。また、特に、本発明の生体内安定化機能を付与するポリペプチド及び該ポリペプチドをコードするDNAを、魚類における性腺刺激ホルモンや成長ホルモン等のようなペプチドの、魚の生体内導入に適用することにより、魚類の産卵誘発や産卵調節のような成熟調節、或いは魚類の成長調節等を効果的に行うことを可能とし、近年多くの種類の栽培漁業が行われている魚類の養殖技術に、その技術の効率化と確実性とをもたらすことができる。したがって、本発明の技術は、実用化技術としてこの分野の技術に大きな進歩をもたらすものである。
【0053】
【配列表】
【図面の簡単な説明】
【図1】本発明の実施例における、一本鎖SCF−cDNA及びSCL+−cDNAの構造を示す図である。
【図2】本発明の実施例における、一本鎖SCF−cDNAを含む発現コンストラクトを示す図である。
【図3】本発明の実施例における、一本鎖SCL+−cDNAを含む発現コンストラクトを示す図である。
【図4】本発明の実施例における、一本鎖SCF−cDNA及びSCL+−cDNAの作製について示す図である。
【図5】本発明の実施例における、一本鎖SCF−cDNA及びSCL+−cDNAを含む発現コンストラクトの構造を示す図である。
【図6】本発明の実施例における、pB−SCL+導入ニジマス初期胚を用いたウエスタン・ブロット解析を示す図である。
【図7】本発明の実施例において、SCFの生物活性について示す図である。
【図8】本発明の実施例において、SCL+の生物活性について示す図である。
【図9】本発明の実施例において、SCL−及びSCL+のin vivo生物活性について示す図である。
【図10】本発明の実施例において、SCL+及びSCL−の血中T量経時変化について示す図である。
Claims (20)
- 胎盤性性腺刺激ホルモンのカルボキシ末端領域からなる、生体内ポリペプチドの生体内安定化機能を付与するポリペプチドをコードするcDNA。
- 配列表の配列番号1に示される塩基配列からなる、請求項1記載の生体内ポリペプチドの生体内安定化機能を付与するポリペプチドをコードするDNA。
- 以下の(a)又は(b)のポリペプチドをコードするDNA。
(a)配列表の配列番号2に示されるアミノ酸配列からなるポリペプチド
(b)配列表の配列番号2に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ生体内ポリペプチドの生体内安定化機能を付与するポリペプチド。 - 請求項2記載のDNAとストリンジェントな条件下でハイブリダイズし、かつ生体内ポリペプチドの生体内安定化機能を付与するポリペプチドをコードすることを特徴とするDNA。
- 配列表の配列番号2に示されるアミノ酸配列からなることを特徴とする生体内ポリペプチドの生体内安定化機能を付与するポリペプチド。
- 配列表の配列番号2に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ生体内ポリペプチドの生体内安定化機能を付与することを特徴とするポリペプチド。
- 生体内ポリペプチドが、性腺刺激ホルモン又は成長ホルモンであることを特徴とする請求項1〜6のいずれか記載のDNA又はポリペプチド。
- 生体内ポリペプチドのカルボキシ末端側に、請求項5又は6記載のポリペプチドを結合したことを特徴とする生体内安定化ポリペプチド。
- 生体内ポリペプチドが、性腺刺激ホルモン又は成長ホルモンであることを特徴とする請求項8記載の生体内安定化ポリペプチド。
- サブユニット構造を備えた生体内ポリペプチドの一方のサブユニットのカルボキシ末端側に、胎盤性性腺刺激ホルモンのカルボキシ末端領域ペプチドを結合したことを特徴とする請求項8又は9記載の生体内安定化ポリペプチド。
- 魚類の生体内ポリペプチドのカルボキシ末端側に、胎盤性性腺刺激ホルモンのカルボキシ末端領域ペプチドを結合したことを特徴とする請求項8〜10のいずれか記載の生体内安定化ポリペプチド。
- 魚類の生体内ポリペプチドが、性腺刺激ホルモン又は成長ホルモンであることを特徴とする請求項11記載の生体内安定化ポリペプチド。
- 請求項12記載の生体内安定化性腺刺激ホルモンを魚類に導入することを特徴とする魚類の成熟調節方法。
- 魚類の成熟調節が、魚類の産卵誘発或いは産卵調節であることを特徴とする請求項13記載の魚類の成熟調節方法。
- 請求項12記載の生体内安定化成長ホルモンを魚類に導入することを特徴とする魚類の成長調節方法。
- 生体内ポリペプチドをコードするDNAの3´末端に、請求項1〜4のいずれか記載のDNAを結合した遺伝子を、生体内で発現することを特徴とする生体内発現ポリペプチドの生体内安定化方法。
- 魚類で発現するポリペプチドをコードするDNAの3´末端に、請求項1〜4のいずれか記載のDNAを結合した遺伝子を、魚類生体内で発現することを特徴とする請求項16記載の生体内発現ポリペプチドの生体内安定化方法。
- 魚類で発現する性腺刺激ホルモンのポリペプチドをコードするDNAの3´末端に、胎盤性性腺刺激ホルモンのカルボキシ末端領域ペプチドをコードするDNAを結合した遺伝子を、魚類生体内で発現することを特徴とする魚類の成熟調節方法。
- 魚類の成熟調節が、魚類の産卵誘発或いは産卵調節であることを特徴とする請求項18記載の魚類の成熟調節方法。
- 魚類で発現する成長ホルモンのポリペプチドをコードするDNAの3´末端に、胎盤性性腺刺激ホルモンのカルボキシ末端領域ペプチドをコードするDNAを結合した遺伝子を、魚類生体内で発現することを特徴とする魚類の成長調節方法。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2002294200A JP4027766B2 (ja) | 2002-10-07 | 2002-10-07 | ポリペプチドの生体内安定化配列及びその利用 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2002294200A JP4027766B2 (ja) | 2002-10-07 | 2002-10-07 | ポリペプチドの生体内安定化配列及びその利用 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2004121180A true JP2004121180A (ja) | 2004-04-22 |
| JP4027766B2 JP4027766B2 (ja) | 2007-12-26 |
Family
ID=32284868
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002294200A Expired - Fee Related JP4027766B2 (ja) | 2002-10-07 | 2002-10-07 | ポリペプチドの生体内安定化配列及びその利用 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP4027766B2 (ja) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2008528027A (ja) * | 2005-01-26 | 2008-07-31 | ユニヴェルシテイト レイデン | 移植細胞により産生されるホルモンを用いた魚の卵及び/又は魚の精子の発生及び成熟を改善するための手段及び方法 |
-
2002
- 2002-10-07 JP JP2002294200A patent/JP4027766B2/ja not_active Expired - Fee Related
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2008528027A (ja) * | 2005-01-26 | 2008-07-31 | ユニヴェルシテイト レイデン | 移植細胞により産生されるホルモンを用いた魚の卵及び/又は魚の精子の発生及び成熟を改善するための手段及び方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| JP4027766B2 (ja) | 2007-12-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Pursel et al. | Genetic engineering of livestock | |
| Hossner | Hormonal regulation of farm animal growth | |
| CN113930447B (zh) | 对非人动物进行基因改造和构建免疫缺陷动物模型的方法 | |
| Chen et al. | Isolation and characterization of tilapia (Oreochromis mossambicus) insulin-like growth factors gene and proximal promoter region | |
| Hoelker et al. | Quantification of DNA binding, uptake, transmission and expression in bovine sperm mediated gene transfer by RT-PCR: effect of transfection reagent and DNA architecture | |
| JP2001501482A (ja) | トランスジェニック魚による組換えペプチドの製造 | |
| Morita et al. | Fish eggs as bioreactors: the production of bioactive luteinizing hormone in transgenic trout embryos | |
| US20020013955A1 (en) | Production of recombinant protein in transgenic fish | |
| CN102482678B (zh) | 利用蚯蚓的性腺再生能力的转基因蚯蚓的养殖方法,通过该养殖方法养殖的转基因蚯蚓以及从转基因蚯蚓的体液生产重组蛋白质的方法 | |
| WO2004028552A1 (en) | Method for inducing mammary epithelial cell differentiation | |
| Chang et al. | Production of transgenic rats and mice by the testis-mediated gene transfer | |
| Su et al. | Increased vibrissa growth in transgenic mice expressing insulin-like growth factor 1 | |
| KR101292894B1 (ko) | 루시퍼라아제 유전자를 과발현하는 형질전환 동물 및 이의 제조 방법 | |
| US8633348B2 (en) | Genetic vasectomy by overexpression of PRML-EGFP fusion protein in spermatids | |
| JP4027766B2 (ja) | ポリペプチドの生体内安定化配列及びその利用 | |
| YOSHIZAKI | Gene transfer in salmonidae: applications to aquaculture | |
| US7579515B2 (en) | Increasing gamete production with a gene switch | |
| Funkenstein et al. | α2-Macroglobulin in the marine fish Sparus aurata | |
| JPWO2018190333A1 (ja) | トランスジェニックカイコ、および該カイコを用いた非天然アミノ酸含有タンパク質の製造方法 | |
| WO2001033957A1 (en) | Transgenic non-human mammals for monitoring change in calcium ion concentration in cells | |
| JPWO2001033957A1 (ja) | 細胞内のカルシウムイオンの濃度変化をモニターするトランスジェニック非ヒト哺乳動物 | |
| WO2024204445A1 (ja) | 魚類の製造方法、魚類および魚類の配偶子の製造方法 | |
| Nguyen et al. | Red Fluorescent Protein Expression in Transgenic Founder of Angelfish (Pterophyllum sp) Driven by Zebrafish Myosin Light Chain 2 Promoter | |
| JP2001086992A (ja) | トランスジェニック魚類を用いた多量体糖タンパク質の生産方法 | |
| WO2001072119A2 (en) | Non-human animal model for growth deficiency and information processing or cognitive function defects and use thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20040129 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20040908 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20070219 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070420 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20070703 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070828 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20071009 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20071010 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101019 Year of fee payment: 3 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20111019 Year of fee payment: 4 |
|
| LAPS | Cancellation because of no payment of annual fees |