JP2004201619A - 体細胞相同組換えの誘発方法 - Google Patents
体細胞相同組換えの誘発方法 Download PDFInfo
- Publication number
- JP2004201619A JP2004201619A JP2002376555A JP2002376555A JP2004201619A JP 2004201619 A JP2004201619 A JP 2004201619A JP 2002376555 A JP2002376555 A JP 2002376555A JP 2002376555 A JP2002376555 A JP 2002376555A JP 2004201619 A JP2004201619 A JP 2004201619A
- Authority
- JP
- Japan
- Prior art keywords
- gene
- homologous recombination
- transcription
- vector
- promoter
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images



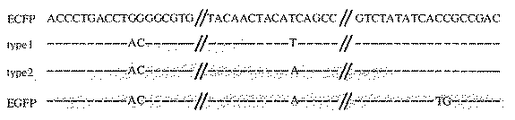
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
- C12N15/902—Stable introduction of foreign DNA into chromosome using homologous recombination
Landscapes
- Genetics & Genomics (AREA)
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Organic Chemistry (AREA)
- Biomedical Technology (AREA)
- Chemical & Material Sciences (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Mycology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
Abstract
【解決手段】任意の遺伝子座においてDNA相同組換えが起きている真核生物細胞中の該遺伝子座に存在する遺伝子の転写活性を制御することにより、該遺伝子と、転写プロモーターの上流域に存在する該遺伝子に類似するDNA配列を持つ遺伝子との間で体細胞相同組換えを誘発し、これによって、複数の遺伝情報を保持する多様な新規遺伝子の取得を可能にする。
【選択図】 なし
Description
【発明の属する技術分野】
本発明は、一般には真核生物細胞における体細胞相同組換えを誘発する技術に係り、より詳細には体細胞中の任意の遺伝子座における体細胞相同組換えを誘発する方法及びかかる方法によって体細胞相同組換えが誘発される細胞に関する。また、本発明は、前記の体細胞相同組換えの誘発方法を利用して複数の遺伝情報を保持する新規遺伝子を作製する方法並びにかかる方法によって作成された新規遺伝子にも関する。
さらに、本発明は上記新規遺伝子によってコードされる新規タンパク質にも関する。
またさらに、本発明は体細胞相同組換えを誘発するために使用して好適な構成を構築したベクターにも関する。
【0002】
【従来の技術】
真核生物にとって体細胞の相同組換えは、遺伝子の多様性を獲得し、その結果としてタンパク質の多様な活性を創出する上で最も重要なDNA代謝反応の一つである。従って、体細胞の相同組換えを制御することは、多様な遺伝子を獲得する上で、非常に重要な課題の一つである。
従来、新しい機能・属性を有する多様な遺伝子を取得する手法として、DNAシャッフリングという技術が(例えば、非特許文献1参照。)がある。この技術は、相同性のある複数の遺伝子配列を混合し、DNaseIで適当に消化して生じた小さな断片をプライマーとし、もともとの遺伝子を鋳型としてPCRを行うことで、擬似的に相同組換えをおこさせるものである。しかしながら、組換え産物の解析は、増幅された断片を発現ベクターにつなぎ、バクテリアにトランスフォームしたのちにおこなうのが一般的であり、産物が高等真核生物においてどのような性質をもつかを解析するのは直接的には困難であると思われる。また、動物細胞内での発現チェックは、新たなベクターへの移入とコドン利用率などに起因する発現適合性を逐次確認する必要がある。
【0003】
一方、動物細胞内で遺伝子組換えを誘発する系としては、細胞内での組換えを活性化する系として、部位特異的組換え酵素Cre-loxを用いた系(例えば、非特許文献2参照。)、配列特異的エンドヌクレアーゼI-SceIを用いた系(例えば、非特許文献3参照。)などが存在する。Cre-loxの系はバクテリオファージP1から得られた38kDaの部位特異的リコンビナーゼであるCreを用いて、loxPサイトと呼ばれる特定部位間で組換えを行わせるものである。また、I-SceIの系は、出芽酵母由来のエンドヌクレアーゼであるI-SceIが、その認識サイトにおいてDNA二重鎖を切断し、DNA相同組換えを誘起する活性を利用したものである。しかしながら、これらの系では、組換えが特定配列間で限定して起きることと、組換え事象が単発であることなどから、得られる組換え体は原則的に一種類のみであった。また、組換えの制御に関わる配列をあらかじめ染色体に導入し、CreあるいはI-SceIなどの組換え酵素を細胞内で発現させる必要があり、染色体の組換えを誘導するのは容易ではなかった。
【0004】
【非特許文献1】
Crameri等, 1998. Nature 391:288-291
【非特許文献2】
DiSanto等, 1995. Proc. Natl. Acad. Sci. USA 92:377-381
【非特許文献3】
Rouet 等, 1994. Proc. Natl. Acad. Sci. USA 91:6064-6068
【0005】
【発明が解決しようとする課題】
本発明者等は、上記事情に鑑みて、所望の体細胞相同組換えを制御された状況下で誘発させる方法がないかについて鋭意研究した結果、意外にも、組換えが望まれる遺伝子座における転写を制御することで、類似した塩基配列との間で体細胞相同組換えを誘発させることが可能になることを見出した。
よって、本発明は、一般には、体細胞中の遺伝子座における体細胞相同組換えを誘発する方法を提供することを目的とする。
また、本発明は上記方法によって体細胞相同組換えが誘発される細胞を提供することを目的とする。
さらに、本発明は、細胞において誘発された体細胞相同組換えを利用して複数の遺伝情報を保持する新規遺伝子及び該新規遺伝子を作製する方法を提供することを目的とする。
またさらに、本発明は該新規遺伝子によってコードされる新規タンパク質を提供することを目的とする。
また、本発明は体細胞相同組換えを誘発するために使用して好適な転写制御のための構成を構築したベクターを提供することを目的とする。
【0006】
【課題を解決するための手段】
しかして、本発明においては、任意の遺伝子座においてDNA相同組換えが起きうる真核生物細胞の体細胞相同組換えを、該遺伝子座における遺伝子の転写制御を行うことによって、該遺伝子と類似する塩基配列との間でDNA相同組換えを誘発することを特徴とする体細胞相同組換えの誘発方法が提供される。また、かかる方法によって体細胞相同組換えが誘発された細胞も提供される。
また、本発明においては、任意の遺伝子座においてDNA相同組換えが起きうる真核生物細胞の体細胞相同組換えを、該遺伝子座における遺伝子の転写制御を行うことによって、該遺伝子と類似する塩基配列との間でDNA相同組換えを誘発して複数の遺伝情報を保持する新規遺伝子を取得する方法が提供される。また、かかる方法によって作成される新規遺伝子も提供される。
またさらに、該新規遺伝子によってコードされる新規タンパク質も提供される。
また、本発明は体細胞相同組換えを誘発するために使用して好適な転写制御に必要な構成を構築したベクターが提供される。
本発明の明細書中に開示された技術を用いれば、DNA相同組換えが色々な箇所で継続的に行われるため、得られる組換体の多様性を高めることが可能である。また、本発明の明細書中に開示された技術を用いれば、DNAシャッフリング技術において必要とされる組換え用のDNAライブラリーを作製することなく、目的の細胞内で遺伝的多様性をもった目的遺伝子が自発的に形成されるため、所望の新規遺伝子の産生について該細胞内にてスクリーニングを行うことができる。従って、多様な新規遺伝子の取得を従来技術に比べて短縮された時間で達成することができる。
【0007】
本発明において使用できる真核生物細胞としては、任意の遺伝子座において体細胞相同組換えが生じうる細胞であれば如何なるものでも使用可能であると思われるが、好適にはニワトリ由来のDT40培養細胞が使用される。
体細胞相同組換えを誘発させる遺伝子は、内在性の遺伝子であっても、外来性の遺伝子であってもよい。また、該遺伝子と類似する塩基配列には、例えば、天然に存在する遺伝子配列、遺伝子としての形態を具備していない天然に存在する塩基配列、人為的な配列などが含まれる。
内在性の遺伝子の場合は、該遺伝子の転写活性を促進する転写プロモーターの上流近傍に該遺伝子と類似する配列が存在する遺伝子座に存在していれば利用可能であり、例えば、抗体軽鎖又は重鎖遺伝子座などが利用可能である。
また、外来性の遺伝子の場合は、相同組換えを引き起こす遺伝子の上流に転写プロモーターを含み、さらにその上流近傍に該遺伝子との類似配列を含むような構成で適当なベクター上に構築させ、対象の細胞の染色体上に組込ませるのであれば、如何なる遺伝子も利用可能である。
ここで、対象の遺伝子と類似する塩基配列は、対象の遺伝子の転写開始位置から1 bps〜100kbps程度上流域に配置されればよく、より好ましくは500bps〜50kbps程度、さらに好ましくは3kbps〜25kbps程度上流域に配置されるのがよい。外来性の遺伝子として、限定はしないが、蛍光タンパク質遺伝子(例えば、GFP遺伝子、CFP遺伝子など)、薬剤耐性遺伝子(例えば、ハイグロマイシン耐性遺伝子、又はピューロマイシン耐性遺伝子)などが含まれる。
本発明において使用される、「体細胞相同組換えを誘発させる遺伝子と類似する塩基配列」とは、該遺伝子の全体もしくは一部と類似する塩基配列のことである。ここで、類似するとは、体細胞相同組換えを誘発させる遺伝子の全体もしくは一部と、60%以上の配列同一性、好ましくは70%以上の配列同一性、さらに好ましくは80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、を含む80%〜99%の配列同一性を有することである。
【0008】
本発明において使用される転写制御の方法は、当業者において周知の方法であれば使用可能であるが、内在性の転写プロモーターを利用することによる方法、又は外来性の誘導的な転写プロモーターを用いて制御する方法などが好ましい。また、転写活性を促進させるために、目的の遺伝子に適するエンハンサー、核マトリックス結合領域(MAR)を用いてもよい。転写制御を行うためには、転写を促進する対象遺伝子に対して、転写プロモーター、エンハンサー、核マトリックス結合領域(MAR)が作用可能に配置される必要がある。ここで、「作用可能に」とは、転写プロモーター、エンハンサー及び核マトリックス結合領域(MAR)がそれぞれの機能を発揮することができるようにとの意味であり、対象遺伝子が所望の状態に転写制御されることを意味する。例えば、転写プロモーターの場合は、特に限定はしないが、対象の遺伝子の転写開始位置より上流側100bps以内程度の領域に配置され、エンハンサーの場合は、限定はしないが、通常、転写開始点から100bps以上、あるいは数キロbps以上離れて存在してもよく、対象遺伝子の5’側であっても、3'側であってもよい。また、核マトリックス結合領域(MAR)はエンハンサーと隣接して配置され、エンハンサーの3'側でも5'側でもよい。
【0009】
【発明の実施の形態】
強化青緑色蛍光タンパク質(ECFP)遺伝子の相同組換え方法は、本発明に係る体細胞相同組換えの誘発方法を一部に利用するものであるので、以下では強化青緑色蛍光タンパク質(ECFP)遺伝子の相同組換え方法について詳細に説明する。前述のように、本発明の強化青緑色蛍光タンパク質(ECFP)遺伝子の相同組換え方法においては、DNA相同組換えが起きている細胞を選択して培養し、相同組換えを誘発するにあたり、該細胞のDNA相同組換えを誘導的なプロモーターによって調節し、ECFPと類似した塩基配列を持つ強化緑色蛍光タンパク質(EGFP)遺伝子配列との間で相同組換えを誘発させる。
よって、以下では、細胞の選択及び培養、誘導的転写制御のためのベクターの構築、相同組換えが誘発された新規遺伝子の発現確認、相同組換えが誘発された新規遺伝子配列の確認について順に説明する。
【0010】
1.体細胞相同組換えを誘発させる細胞の選択
本発明における「真核生物細胞」とは、体細胞相同組換えがある程度の頻度で生じているものを指し、好ましくは、ヒト、マウス、ヒツジ、ラット、ウサギ、ニワトリなどのB細胞又はその細胞株、より好ましくはヒトバーキットリンパ腫由来のラモス細胞(Ramos cell)株、又はニワトリB細胞由来のDT40細胞株であり、最も好ましくはDT40細胞株である。
本発明において用いられる「DT40細胞株」は、当該細胞の保有する染色体に何らかの修飾(例えば、特定の遺伝子の組換え、挿入、削除等)が加えられた、誘導体株、サブライン(Subline)なども含む。
本発明で用いる細胞の培養条件は当該技術分野において周知の方法によって行われるが、選択される細胞に適した培地、培養条件(培養温度、CO2濃度)下で行われることは言うまでもない。 培地としては、一般に使用されているRPMI1640培地、DMEM培地又はこれらの培地にウシ胎児血清等を添加した培地等が挙げられる。培養は、通常、5%CO2存在下、用いる細胞にとっての適温(例えば、25℃〜40℃)で1〜30日行う。培養中は必要に応じてカナマイシン、ペニシリン等の抗生物質を培地に添加してもよい。また、転写制御を誘導的なプロモーターを用いて行う場合は、転写を誘導するための薬剤(例えば、テトラサイクリンなど)を添加してもよい。しかして、選択される免疫細胞がDT40細胞である場合、例えば、培地はIMDM(Invitrogen社)を用い、培養温度は例えば39.5℃、5%のCO2濃度条件下で行う。
【0011】
2.転写制御のためのベクターの構築
(1)シスに作用する領域
本発明で用いられる転写制御の方法は、当業者にとって周知の技術であれば、如何なる方法を用いてもよいが、好ましくは、転写制御においてシスに作用する領域を適当なベクターに構築し、該ベクターを細胞内へ導入することで、染色体上の特定の遺伝子座に組込ませる方法が用いられる。ここで、「シスに作用する領域」とは、特定の遺伝子の転写制御に必要なDNA配列であって、該遺伝子に隣接して存在する領域のことを指す。「シスに作用する領域」には、転写プロモーター、エンハンサー、核マトリックス結合領域(MAR)、その他転写活性を制御するのに必要な任意の配列が含まれる。
転写プロモーターは、構成的プロモーター又は誘導的プロモーターのいずれも使用可能であるが、転写制御が容易であることから、誘導的プロモーターが好ましい。誘導的プロモーターとしては、α−インターフェロン、ヒートショック、重金属イオン、及びグルココルチコイドなどのステロイド(Kaufman, 1990)及びテトラサイクリンに応答するものが含まれる。他の望ましい誘導可能なプロモーターとしては、プロモーターの活性化因子が外から添加された場合に、細胞内で応答するものが含まれる。好適には、例えば、テトラサイクリン誘導プロモーターなどが使用可能である。
エンハンサー及び核マトリックス結合領域(MAR)は選択された細胞中での転写活性の制御に機能するものであれば、如何なるものも使用可能であるが、DT40細胞を用いる場合には、ニワトリ抗体軽鎖遺伝子エンハンサー(3' enhancer)、ニワトリ由来の核マトリックス結合領域を用いるのが好ましい。
【0012】
シスに作用する領域の構成成分のうち、誘導的プロモーター、エンハンサー及び核マトリックス結合領域(MAR)は、配列が公知である場合には、その配列に基づいてPCR法等により該配列を増幅するためのプライマーを作製し、適当なcDNAライブラリー又はゲノムライブラリーから直接取得することも可能である。ここで用いるプライマーは、クローニングの便宜上、クローン化するためのベクター上に存在する制限酵素サイトを付加させるように設計するのが望ましい。取得されたエンハンサー及び核マトリックス結合領域(MAR)は相同組換えを誘発させる遺伝子及び該遺伝子と類似したDNA配列が構築されているベクター上、該遺伝子と類似したDNA配列の間にプロモーター、該遺伝子の下流、3'方向に向かって、核マトリックス結合領域(MAR)、エンハンサーの順番あるいはその逆の順番で、適切に付加された制限酵素サイトを用いてクローニングすることができる。
また、誘導的プロモーターが予めクローニングされた市販のベクター、例えば、pTRE2hyg(Clontech社)などに、目的の遺伝子、該遺伝子と類似の塩基配列、エンハンサー、核マトリックス結合領域(MAR)等をクローニングしてもよい。
【0013】
(2)遺伝子(ECFP)の取得
相同組換えを誘発させる遺伝子は、内在性の遺伝子あってもよいが、単離された外来性の遺伝子でもよい。転写制御が容易であることから、外来性の遺伝子が好ましい。
相同組換えを誘発させる遺伝子は、当業者において周知の方法によりクローニングされるが、該遺伝子が存在するcDNAライブラリー又はゲノムDNAに対し、該遺伝子を増幅するのに使用可能なプライマーセット(例えば、該遺伝子の5'端側及び3'端側に該遺伝子を増幅可能に設計した1対のプライマーセット)を用いて、正確性の高いDNAポリメラーゼ(例えば、Pyrobest DNA Polymerase(TaKaRa))により該遺伝子を増幅してもよい。
得られた遺伝子は、適当なクローニング用のベクター(例えば、pBlueScriptII、pUC19など)にクローニングすることができる。
相同組換えを誘発させる遺伝子がECFP遺伝子の場合は、市販のベクター(pECFP-C1)からクローニングを行ってもよい。
同様な方法により、ECFP遺伝子の全体又は一部と類似する塩基配列を得ることが可能である。ECFP遺伝子と類似する配列であれば、如何なる配列でも利用可能であるが、例えば、EGFP遺伝子配列などが好ましい。
取得されたECFP遺伝子及びECFP遺伝子と類似する配列は、前述したように、転写プロモーターを間に挟んで、ECFP遺伝子の上流側(即ち、5'側)になるように、「シスに作用する領域」を構築したベクター上に作用可能に挿入する。類似するDNA配列の挿入する方向は、順方向でも逆方向でもよい。「順方向」とは、対象のDNA配列が天然に存在するものであれば、天然に存在する方向のことであり、人為的な配列の場合は、任意の方向を「順方向」と決めてもよい。
【0014】
3.転写因子発現ベクターの構築
遺伝子の体細胞相同組換えを誘発するためには、転写プロモーターの活性を促進又は抑制するのに必要な転写因子の発現が必要である。例えば、テトラサイクリン誘導プロモーターの場合は、テトラサイクリン応答配列に結合して転写を調節する転写因子(例えば、TetリプレッサーとVP16転写活性化ドメインを融合したもの)が必要である。これら転写因子を発現させるためのベクターは、例えば、該転写因子の発現に対して作用可能に連結されたプロモーター(例えば、ヒトサイトメガロウィルス最初期プロモーターなど)、適当な選択マーカー(例えば、ネオマイシン耐性マーカー、ハイグロマイシン耐性マーカーなど)などを含んでもよい。
また、プロモーターを制御するためのベクターは、市販のもの、例えば、pTet-Off又はpTet-Onベクター(Clontech社)などを用いてもよい。
【0015】
4.相同組換えが誘発された新規遺伝子の発現確認
(1)選択した遺伝子の相同組換えの誘発
選択した遺伝子(ここではECFP遺伝子)の相同組換えを誘発させるためには、ECFP遺伝子に作用可能に連結されたプロモーターに対する転写因子の活性を、さらに調節する因子が必要である。該因子は選択された転写プロモーターに依存して決定され、当業者であれば過度の実験等をすることなく選択することができる。例えば、選択されたプロモーターがテトラサイクリン誘導プロモーターである場合、該因子として、テトラサイクリン又はドキソサイクリンなどが利用可能である。
また、転写制御のためのベクター及び転写因子発現ベクターを選択された細胞に導入する必要がある。ベクターを細胞に導入する方法としては、当業者に周知の方法を用いることができるが、例えば、リン酸カルシウム法(Chen及びOkayama, 1988)、カチオン性脂質による方法(Elroy-Stein及びMoss, 1990)、エレクトロポレーション法(Neumann等, 1982)等が利用可能である。
前記2種類のベクターを細胞に導入したのち、転写プロモーターの活性化に必要な因子(例えば、テトラサイクリン、ドキソサイクリンなど)を培地に添加又は除去することで目的の転写プロモーターを活性化させる。これにより、選択した遺伝子の相同組換えが誘発される。
(2)相同組換えが誘発された新規遺伝子の発現確認
相同組換えが誘発されることにより生じる新規遺伝子の発現は、組換えが誘発される前の遺伝子産物の特性に応じて、当業者により周知の技術を用いて確認することができる。例えば、選択された遺伝子が特定の薬剤耐性を示すものである場合には、相同組換え誘発前の細胞と誘発後の細胞における特定の薬剤に対する許容濃度等を比較することで、新規遺伝子の発現を予測することができる。また、 ECFP遺伝子の場合には、発する蛍光波長の変化等で、新規遺伝子の発現を確認することができる。
【0016】
5.相同組換えが誘発された新規遺伝子の配列解析
新規遺伝子が創出されたことは、相同組換えが行われた新規遺伝子の配列解析を行い、実際に相同組換えが起こったことによりさらに確認することができる。相同組換えに関し確認を行う細胞のゲノムDNAを既知の方法により抽出し、確認すべき遺伝子の増幅に使用可能な特異的なDNAプライマー(例えば、目的の遺伝子領域全体を含むように、該遺伝子の5’側近傍に正方向のプライマーを設計し、該遺伝子の3’側に逆方向のプライマーを設計する)を用いてPCR法により目的の遺伝子を増幅させる。
増幅に使用されるDNAポリメラーゼは市販のものを用いることができるが、正確性の高いものを使用することが望ましい。増幅を行うための条件は、使用するDNAプライマーのアニール温度、使用するDNAポリメラーゼの性質等に依存するが、例えば、98℃2分間の反応後、98℃30秒、58℃30秒、72℃1分を26サイクル、さらに72℃で5分間反応させる。反応後の増幅産物は、アガロースゲル電気泳動で分離し、目的の遺伝子を含むDNAのバンドを切り出し、DNAを回収後、配列決定用のベクターへ組込む。配列決定用のベクターは当該技術分野で用いられる如何なるベクターであってもよいが、例えば、pCR2.1-TOPO(Invitrogen社)などが用いられる。上記調製された配列決定用のベクター中の遺伝子のDNA配列を、定法に従い決定することができる。
【0017】
【実施例】
以下に実施例を示すが、本発明はこれに限定されるものではない。
【0018】
実施例1:誘導的転写制御のためのベクターの構築
遺伝子の発現を誘導的に制御するためのプロモーターを細胞内に導入するために、転写誘導に必要なシスに作用する領域をベクター上に構築した(図1)。
テトラサイクリン誘導プロモーター(TRE及びヒトサイトメガロウィルスミニプロモーター)の下流にECFP遺伝子をつなぎ、さらに下流にMAR、エンハンサー(3' enhancer)をつないだ。上流には、約3kbのスペーサー配列(酵母Arg4遺伝子、3kbのPstI断片)を配置し、さらにECFPとは逆向きにEGFPをつないだ。
MAR及び3' enhancerのクローニングは、PCR法を利用した。PCRは、Roche社製、Expand High Fidelity PCR systemを用い、DT40ゲノムDNAを鋳型とし、以下のようにデザインしたプライマーにより行った。
MAR:
正方向プライマー
5'-GCTGCAGTGTCCTTGGGGGTGAAATTCAG-3'(配列番号:1)
逆方向プライマー
5'-GCTCTAGAACTGCCCCCATTAAAAACTTTC-3' (配列番号:2)
3' enhancer:
正方向プライマー
5'-GCTCTAGAAGGCACAGCGCTGTCAGGGTGC-3' (配列番号:3)
逆方向プライマー
5'-CCGCGGCCGCGTGGTGGGAGCGGGCAGGGG-3' (配列番号:4)
PCR産物は、一度pCR2.1ベクターにTAクローニングし、その後適切な制限酵素で消化(MAR:PstI, XbaI, 3' enhancer:XbaI, NotI)した後、pBluescriptIIにクローニングし直した。テトラサイクリン誘導プロモーターは、pTRE2hyg(Clontech社)の、TREを含むXhoI, EcoRI断片をpBlueScriptIIにクローニングした(pHS305)。この、TREを含むプラスミドに、ECFP遺伝子を挿入した。ECFP遺伝子は、pECFP-C1プラスミド(Clontech社)のECFPを含むNheI, MulI断片を、EcoRI消化したpHS305にブラントエンドライゲーションした(pHS344)。さらに、pBluescriptIIにクローニングしてあるMARをpHS344のECFPの下流部分に挿入した(pHS345)。前述のように、スペーサー配列には酵母ARG4遺伝子を用いた。これは、PstIにより生じる3kb断片をpUC119つないだものであるが、このプラスミドのSnaBI断片に、EGFP断片(pEGFP-C1のNheI, BspEI断片)をブラントエンドライゲーションした。ここから、PstIによりEGFP, ARG4遺伝子を含む配列を切り出し、pHS345のXhoI断片にブランドエントライゲーションすることで、pHS346を得た。さらに、pBlueScriptIIにクローニングしてある3’enhancerをXbaI およびNotIで消化したpHS346に挿入してpHS347とした。
【0019】
実施例2:DNA相同組換えが誘発される細胞の調製
(1)細胞培養
DT40細胞は、CO2恒温槽にて5%CO2、39.5℃で培養した。培地は、IMDM培地(Invitrogen社)を用い、10%FBS、1%ニワトリ血清、ペニシリン 100単位/ml、ストレプトマイシン 100μg/ml、2-メルカプトエタノール 55μMを加えて使用した。
(2)トランスフェクション
テトラサイクリン誘導転写因子を発現する発現ベクターpTA-Hyg(Clontech社) 30μgをHindIIIで、また、60μgのpHS347をXmnIで直線化し、既知の手法(Buerstedde,等, 1991)に従ってDT40細胞に感染させた。その際、2.5mg/mlハイグロマイシン、100ng/mlドキシサイクリンを培地に加えてセレクションを行った。その結果、ハイグロマイシン耐性クローンのうち、ドキシサイクリン非存在下でECFPを発現するクローンを得ることができた(HS101株)。
【0020】
実施例3:新規遺伝子産物の確認
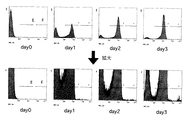
HS101をドキシサイクリン非存在下で培養し、蛍光強度をEPICS ELITE ESP (ベックマン-コールター社)にて測定した結果、ECFPと思われる蛍光を発する集団が観測された(図2:ゲートE)。培養を続けると、ECFPよりもさらに蛍光強度の強い集団が観測された(図2:ゲートF)。EとFの比をとったところ、誘導開始からの培養時間に依存してFの存在比が上昇することが観測された(図3)。さらに、誘導後培養5日目の細胞を蛍光顕微鏡で観察すると、ECFPの蛍光を発する細胞の中に、GFPと思われる蛍光を発するものが観測された(図4)。
【0021】
実施例4:新規遺伝子配列の解析
(1)ゲノムDNAの抽出
FACSにおいてゲートF近くの細胞をソートし、EPICS ELITE ESP (ベックマン-コールター社)により生細胞5000個をシリコンコートした1.5mlチューブに集めた。シース液に懸濁された細胞を遠心(1000g, 10min)により回収し、ペレットに直接100μlのゲノム抽出バッファー(100 mM Tris-HCl(pH8.0), 5mM EDTA, 0.2%SDS, 200 mM NaCl及び100μg/mlプロテイナーゼK)を加え、50℃で一晩消化した。翌日、250μlのエタノールを加え、穏やかに上下し反転させて混ぜた。ゲノムを遠心(1000 g, 10 min)により回収し、70%エタノールで洗い、風乾した。ここに100μlのTE(10 mM Tris-HCl(pH8.0), 1 mM EDTA)を加え、50℃で30分放置した後、4℃にて一晩かけて溶解させた。
(2)TRE直下の配列の解析
抽出したゲノムから、PCR(Perkin Elmer 9600)でテトラサイクリン誘導プロモーターの直下の配列を増幅したものをTAクローニングして配列を解析した。
ゲノムDNA溶液5μl(細胞5000個に相当)を鋳型とし、プライマーは上流(5'-CCATAGAAGACACCGGGACCGATCC-3')(配列番号:5)、下流(5'-TGCACGCTGCCGTCCTCGATGTTG-3')(配列番号:6)それぞれ10 pmolを使用した。Pyrobest DNA Polymerase(TaKaRa)を用いて、50μlスケールで反応させた。反応条件は、98℃2分の後、98℃30秒、58℃30秒、72℃1分を26サイクル行い、最後に72℃5分間反応させた。その後、ExTaq DNA Plymerase(TaKaRa)を1μl加え、72℃15分反応させた後、全反応液の20μl分をアガロースゲル電気泳動で分離した。軽鎖遺伝子可変領域に相当するバンドを切り出し、Gel Extraction kit(Qiagen社)によりDNAを回収後、TOPO TA Cloning kit(Invitrogen社)にてpCR2.1-TOPOベクターに組込み、大腸菌にトランスフォーメーションした。プラスミドを抽出し、ABI PRISM 377 DNA Sequencer(Perkin Elmer社)により配列を解析した。その結果、図5に示すように、ECFPとEGFPのキメラが生じていることが明らかとなった。二種類のキメラのうち、type1は22クローン、type2は1クローン得られた。
【0022】
【発明の効果】
本発明ではライブラリー作製を経ずに、自発的に遺伝的多様性を持った目的遺伝子が形成され、その後すぐに動物細胞内でのスクリーニングが可能になるので、目的遺伝子の取得までの時間を大幅に短縮することが可能となる。
また、本発明では、様々な箇所において、継続的に相同組換えが誘発されるため、得られる組換体の多様性を上昇させることも原理的に可能であり、新規な活性及び機能を持つ遺伝子を創出する技術としても利用可能である。
【0023】
参考文献
Buerstedde, J.M.及びTakeda, S. 1991. Cell 67:179-88
Chen, C. 及び Okayama, H. 1988. BioTechniques 6:632-638.
Crameri等, 1998. Nature 391:288-291
DiSanto等, 1995. Proc. Natl. Acad. Sci. USA 92:377-381
Elroy-Stein, O.及び B. Moss. 1990. Proc. Natl. Acad. Sci. USA. 87:6743-6747.
Kaufman, R.J. 1990. Methods Enzymol. 185:487-511.
Neumann, E., M.等, 1982. EMBO J. 1:841-845.
Rouet 等, 1994. Proc. Natl. Acad. Sci. USA 91:6064-6068
【0024】
【配列表】

【図面の簡単な説明】
【図1】転写誘導に必要なシスに作用する領域をベクター上に構築する方法の概略を示す。
【図2】FACSによる蛍光強度の経時的変化について示す。ECFPと思われるゲートE、さらに蛍光強度の強い集団をゲートFとした。
【図3】FACSによる蛍光強度の経時的変化について示す。Eの蛍光強度とFの蛍光強度との比をとり、定量化した。
【図4】ECFP遺伝子とEGFP遺伝子との間で相同組換えを誘発させた細胞の蛍光顕微鏡による観察像を示す。ECFPの転写誘導5日後の蛍光顕微鏡像である。CFPの蛍光を発する細胞(A)の中に、GFPと思われる蛍光を発する細胞(B)が観測された(C)。
【図5】新規遺伝子配列の解析結果を示す。蛍光強度の強い細胞集団のTRE直下の配列を解析した結果、type1、type2の新規な二種類の組換体が見出された。
Claims (16)
- 任意の遺伝子座においてDNA相同組換えが起きている真核生物細胞の体細胞相同組換えを誘発する方法であって、該遺伝子座の遺伝子の転写制御を行うことによって、該遺伝子の塩基配列と該遺伝子と類似する塩基配列との間でDNA相同組換えを誘発することを特徴とする体細胞相同組換えの誘発方法。
- 前記細胞がDT40細胞であることを特徴とする請求項1に記載の方法。
- 前記転写制御のための転写プロモーターが、前記遺伝子と類似する塩基配列の下流3’側に配置され、前記遺伝子と作用可能に隣接することによって、該遺伝子の転写を制御することを特徴とする請求項1又は2に記載の方法。
- 前記転写制御のためのシスに作用する領域が、エンハンサー、核マトリックス結合領域(MAR)のいずれか一つ又は両方を含むことを特徴とする請求項1ないし3のいずれか一項に記載の方法。
- 前記遺伝子及び前記遺伝子と類似する塩基配列が外来性である場合、
(a)該遺伝子に類似する塩基配列、転写プロモーター及び該遺伝子のベクター上における順番が、5’側から該遺伝子と類似する塩基配列、転写プロモーター、該遺伝子の順番であって、該転写プロモーターが該遺伝子と作用可能となるように挿入する段階、
(b)該ベクターを細胞内へ導入して、該遺伝子に類似する塩基配列、転写プロモーター及び該遺伝子を染色体上に組込む段階、
を含むことを特徴とする請求項1ないし4のいずれか一項に記載の方法。 - 前記ベクター上にエンハンサー、核マトリックス結合領域(MAR)のいずれか一つ又は両方を前記転写プロモーターに対して作用可能に挿入することを特徴とする請求項5に記載の方法。
- 前記転写プロモーターが誘導的プロモーターであることを特徴とする請求項5又は6に記載の方法。
- 前記誘導的プロモーターがテトラサイクリン誘導プロモーターであることを特徴とする請求項7に記載の方法。
- 前記遺伝子が強化青緑色蛍光タンパク質(EGFP)遺伝子であることを特徴とする請求項5ないし8のいずれか一項に記載の方法。
- 前記遺伝子と類似する塩基配列が強化緑色蛍光タンパク質(EGFP)遺伝子配列であることを特徴とする請求項5ないし9のいずれか一項に記載の方法。
- 前記エンハンサーがニワトリ抗体軽鎖遺伝子エンハンサー(3' enhancer)であって、前記核マトリックス結合領域(MAR)がニワトリ由来であることを特徴とする請求項4ないし10のいずれか一項に記載の方法。
- 請求項1ないし11のいずれか一項に記載の方法により、DNA相同組換えが誘発される細胞。
- 請求項1ないし11のいずれか一項に記載の方法により、相同組換えが誘発される遺伝子。
- 請求項13に記載の相同組換えが誘発される遺伝子によってコードされるタンパク質。
- 相同組換えを誘発する遺伝子及び該遺伝子の転写制御のための転写プロモーターが配置されたベクターにおいて、該転写プロモーターの5'側上流域に該遺伝子に類似する塩基配列が配置され、該遺伝子の相同組換えを誘発するために構築されるベクター。
- エンハンサー、核マトリックス結合領域(MAR)のいずれか一つまたは両方をさらに作用可能に挿入した請求項15に記載のベクター。
Priority Applications (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2002376555A JP5319863B2 (ja) | 2002-12-26 | 2002-12-26 | 体細胞相同組換えの誘発方法 |
| CN200380110005XA CN1756840B (zh) | 2002-12-26 | 2003-12-22 | 诱导体细胞同源重组的方法 |
| US10/540,302 US9587251B2 (en) | 2002-12-26 | 2003-12-22 | Method of inducing homologous recombination of somatic cell |
| EP03781013.2A EP1584678B1 (en) | 2002-12-26 | 2003-12-22 | Method of inducing homologous recombination of somatic cell |
| PCT/JP2003/016496 WO2004058964A1 (ja) | 2002-12-26 | 2003-12-22 | 体細胞相同組換えの誘発方法 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2002376555A JP5319863B2 (ja) | 2002-12-26 | 2002-12-26 | 体細胞相同組換えの誘発方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2004201619A true JP2004201619A (ja) | 2004-07-22 |
| JP5319863B2 JP5319863B2 (ja) | 2013-10-16 |
Family
ID=32677367
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002376555A Expired - Lifetime JP5319863B2 (ja) | 2002-12-26 | 2002-12-26 | 体細胞相同組換えの誘発方法 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US9587251B2 (ja) |
| EP (1) | EP1584678B1 (ja) |
| JP (1) | JP5319863B2 (ja) |
| CN (1) | CN1756840B (ja) |
| WO (1) | WO2004058964A1 (ja) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2008516637A (ja) | 2004-10-20 | 2008-05-22 | ザ スクリップス リサーチ インスティテュート | 真正細菌へのn−アセチルガラクトサミンアミノ酸のインビボ部位特異的組込み |
| US20100291142A1 (en) * | 2007-10-18 | 2010-11-18 | Roger Maes | Bacterial Artificial Chromosome Containing Feline Herpes Virus Type 1 Genome and Uses Thereof |
| JP6156144B2 (ja) | 2011-09-21 | 2017-07-05 | 富士レビオ株式会社 | 親和性複合体に対する抗体 |
-
2002
- 2002-12-26 JP JP2002376555A patent/JP5319863B2/ja not_active Expired - Lifetime
-
2003
- 2003-12-22 CN CN200380110005XA patent/CN1756840B/zh not_active Expired - Lifetime
- 2003-12-22 US US10/540,302 patent/US9587251B2/en not_active Expired - Fee Related
- 2003-12-22 EP EP03781013.2A patent/EP1584678B1/en not_active Expired - Lifetime
- 2003-12-22 WO PCT/JP2003/016496 patent/WO2004058964A1/ja not_active Ceased
Non-Patent Citations (10)
| Title |
|---|
| JPN6008057472, NICKOLOFF,J.A., ""Transcription Enhances Interchromosomal Homologous Recombination in Mammalian Cells"", Mol. Cell. Biol., 1992, Vol.12, No.12, pp.5311−5318 * |
| JPN6008057473, SLEBOS, R.J.C. and TAYLOR, J.A., ""A novel host cell reactivation assay to assess homologous recombination capacity in human cancer ce", Biochem. Biophys. Res. Commun., 2001, Vol.281, No.1, pp.212−219 * |
| JPN6008057475, PHI−VAN, L. and STRATLING, W.H., ""Dissection of the ability of the chicken lysozyme gene 5’ matrix attachment region to stimulate tra", Biochemistry, 1996, Vol.35, No.33, pp.10735−10742 * |
| JPN6008057477, BUERSTEDDE, J. and TAKEDA, S., Cell, 1991, Vol.67, No.1, pp.179−188 * |
| JPN6008057480, LAHTI, J.M., Methods, 1999, Vol.17, No.4, pp.305−312 * |
| JPN6008057482, BULFONE−PAUS,S. et al., Nucleic Acids Res., 1995, Vol.23, No.11, pp.1997−2005 * |
| JPN6008057485, LAUSTER,R., et al., EMBO J., 1993, Vol.12, No.12, pp.4615−4623 * |
| JPN6008057488, ISRAEL, D.I. and KAUFMAN, R.J., Nucleic Acids Res., 1989, Vol.17, No.12, pp.4589−4604 * |
| JPN6010014501, KANAYAMA, N. et al., Nucleic Acids Research, 2006, Vol.34, No.2, e10 * |
| JPN6010014504, REYNAUD, C−A. et al., ""A hyperconversion mechanism generates the chicken light chain preimmune repertoire"", Cell, 1987, Vol.48, pp.379−388 * |
Also Published As
| Publication number | Publication date |
|---|---|
| CN1756840B (zh) | 2010-08-04 |
| WO2004058964A1 (ja) | 2004-07-15 |
| EP1584678A1 (en) | 2005-10-12 |
| JP5319863B2 (ja) | 2013-10-16 |
| US9587251B2 (en) | 2017-03-07 |
| US20060099678A1 (en) | 2006-05-11 |
| EP1584678A4 (en) | 2006-06-21 |
| CN1756840A (zh) | 2006-04-05 |
| EP1584678B1 (en) | 2016-08-31 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US11535871B2 (en) | Optimized gene editing utilizing a recombinant endonuclease system | |
| JP2024536853A (ja) | 環状rnaおよびその調製方法 | |
| US8501454B2 (en) | Homologous recombination-based DNA cloning compositions | |
| CN107794272B (zh) | 一种高特异性的crispr基因组编辑体系 | |
| US7244609B2 (en) | Synthetic genes and bacterial plasmids devoid of CpG | |
| CN103224947A (zh) | 一种基因打靶系统 | |
| JP7852942B2 (ja) | 活性dnaトランスポゾンシステム及びその使用方法 | |
| JP4487068B2 (ja) | 細胞の遺伝子変異機能の制御による変異タンパク質の作製方法 | |
| JP6713691B2 (ja) | 動物細胞ゲノム部位特異的外来dna挿入方法及び前記挿入方法を用いて得られる細胞 | |
| JP2007502129A (ja) | 短鎖干渉rnaライブラリーならびに合成および使用の方法 | |
| CN113260700A (zh) | 基于可检测标签与靶蛋白的CRISPR/Cas控制的整合而选择细胞的方法 | |
| JP5319863B2 (ja) | 体細胞相同組換えの誘発方法 | |
| JP2003532381A (ja) | 二重選択ベクター | |
| CN113474454A (zh) | 可控的基因组编辑系统 | |
| Buchholz et al. | LoxP-directed cloning: use of Cre recombinase as a universal restriction enzyme | |
| JP6779513B2 (ja) | インビボクローニング可能な細胞株をスクリーニングするための方法、インビボクローニング可能な細胞株の製造方法、細胞株、インビボクローニング方法、及びインビボクローニングを行うためのキット | |
| TWI843749B (zh) | PaCas9核酸酶 | |
| JP2004141025A (ja) | 細胞発現用合成dna断片及び作成方法 | |
| JP7018198B2 (ja) | インビボクローニング可能な細胞株をスクリーニングするための方法、インビボクローニング可能な細胞株の製造方法、細胞株、インビボクローニング方法、及びインビボクローニングを行うためのキット | |
| JP7041571B2 (ja) | 組換え哺乳動物細胞および目的物質の生産方法 | |
| US11473066B2 (en) | Flp-TAL recombinases | |
| TW201840849A (zh) | 編輯核酸序列之組成物及方法 | |
| KR20250163962A (ko) | Crispr 간섭 및 활성화를 위한 개선된 방법 및 조성물 | |
| JP4581838B2 (ja) | Dna断片連結方法及びキット | |
| HK40088155A (zh) | 活性dna转座子系统及其使用方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20040825 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20040825 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20050707 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20050707 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20081202 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20090129 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20100406 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20100706 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20100720 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20101021 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20101029 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20130612 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20130712 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 5319863 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| S533 | Written request for registration of change of name |
Free format text: JAPANESE INTERMEDIATE CODE: R313533 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |