JP2004503252A - Hiv−1リポーターウイルスおよび抗ウイルス性化合物をアッセイするうえでのその使用 - Google Patents
Hiv−1リポーターウイルスおよび抗ウイルス性化合物をアッセイするうえでのその使用 Download PDFInfo
- Publication number
- JP2004503252A JP2004503252A JP2002510723A JP2002510723A JP2004503252A JP 2004503252 A JP2004503252 A JP 2004503252A JP 2002510723 A JP2002510723 A JP 2002510723A JP 2002510723 A JP2002510723 A JP 2002510723A JP 2004503252 A JP2004503252 A JP 2004503252A
- Authority
- JP
- Japan
- Prior art keywords
- hiv
- gene
- genome
- vector
- replication
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images









Classifications
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/502—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics for testing non-proliferative effects
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5044—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics involving specific cell types
- G01N33/5047—Cells of the immune system
- G01N33/505—Cells of the immune system involving T-cells
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/575—Immunoassay; Biospecific binding assay; Materials therefor for cancer
- G01N33/57515—Immunoassay; Biospecific binding assay; Materials therefor for cancer of the breast
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2740/00—Reverse transcribing RNA viruses
- C12N2740/00011—Details
- C12N2740/10011—Retroviridae
- C12N2740/16011—Human Immunodeficiency Virus, HIV
- C12N2740/16041—Use of virus, viral particle or viral elements as a vector
- C12N2740/16043—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/005—Assays involving biological materials from specific organisms or of a specific nature from viruses
- G01N2333/08—RNA viruses
- G01N2333/15—Retroviridae, e.g. bovine leukaemia virus, feline leukaemia virus, feline leukaemia virus, human T-cell leukaemia-lymphoma virus
- G01N2333/155—Lentiviridae, e.g. visna-maedi virus, equine infectious virus, FIV, SIV
- G01N2333/16—HIV-1, HIV-2
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Immunology (AREA)
- Biomedical Technology (AREA)
- Chemical & Material Sciences (AREA)
- Molecular Biology (AREA)
- Hematology (AREA)
- Urology & Nephrology (AREA)
- Biotechnology (AREA)
- Cell Biology (AREA)
- Biochemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Physics & Mathematics (AREA)
- Microbiology (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Analytical Chemistry (AREA)
- General Physics & Mathematics (AREA)
- Pathology (AREA)
- Genetics & Genomics (AREA)
- Food Science & Technology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Toxicology (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Virology (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
本発明は、複製コンピテントなHIV−1リポーターウイルスを生成する、HIV−1プロウイルス構築物に関する。本発明はまた、抗ウイルス性化合物の高処理量スクリーニングにおけるこれらHIV−1リポーターウイルスの使用にも関する。
Description
【0001】
(技術分野)
本発明は、リポーター遺伝子、好ましくはレニラ(renilla)ルシフェラーゼか分泌胎盤アルカリホスファターゼ(secreted placental alkaline phosphatase)(SEAP)のいずれかのリポーター遺伝子をコードする、複製コンピテントな(replication competent)HIV−1プロウイルス構築物に関する。本発明はさらに、HIV−1の複製を抑制する化合物の高処理量スクリーニングにおけるこれらプロウイルス構築物ビリオンの使用に関する。
【0002】
(背景技術)
1型ヒト免疫不全ウイルス(HIV−1)の細胞培養中での複製は、一般に、ウイルスの逆転写酵素(RT)活性の産生を定量するか、細胞培養液中の成熟ウイルスカプシドタンパク質(すなわち、p24)を定量するか、または逆転写酵素によって指令された複製連鎖反応(RT−PCR)によるウイルスRNAコピー数を定量することによって測定される。あるいは、HIV−1複製は、XTTやMTTなどの色素還元(dye reduction)法を用いて感染細胞に対するウイルスによって誘発された細胞変性効果(cpe)を定量することによって間接的に測定することができる(ワイスロウ(Weislow)ら、1989)。
【0003】
HIV−1 RTアッセイまたはp24アッセイの場合、これら方法の厄介な性質はこれら方法を高処理量の抗ウイルスアッセイに適用するのを排除している。一方、XTTなどの色素還元法によるHIV複製の分析は単一工程のプロセスであり、それゆえ高処理量が可能である。しかしながら、色素還元法はしばしば培養液中の大部分の細胞が感染し死滅した後に行われるので、一層有効性の少ないHIV−1インヒビターの同定の感度の低減という結果となる。さらに、色素還元法は有意の細胞変性効果に依存しているので、シンシチウム非誘導性のHIV−1株のように培養細胞株中で攻撃的な(aggressive)複製動力学を示さないHIV株の複製を正確に測定するのに用いることはできない。
【0004】
(発明の開示)
(発明が解決しようとする技術的課題)
HIV−1リポーターウイルスの生成は、HIV−1複製の測定の従来法に代わる方法を提供する。リポーター遺伝子としてルシフェラーゼをコードする単サイクル感染性HIV−1リポーターウイルスが記載されている(チェン(Chen)ら、1994)。これらウイルスは、ウイルスの侵入、逆転写、組込み、および遺伝子発現を含むHIV−1複製サイクルのほぼ最初の半分の簡単かつ迅速な分析を可能にする。しかしながら、感染細胞でのHIV遺伝子発現の後の工程、たとえばビリオンの成熟に必要なウイルス前駆体ポリペプチドのHIVプロテアーゼ媒体プロセシングなどは、これらウイルスを用いては容易に測定できない。
【0005】
HIVの完全な複製サイクルを分析し、細胞集団中のHIV−1の伝播を追跡するには、複製コンピテントなリポーターウイルスが必要である。HIV−1の複製の分析に用いることのできる幾つかの複製コンピテントなHIV−1リポーターウイルスが作製されている(ターウィリガー(Terwilliger)ら、1989;マリム(Malim)ら、1992;ヘ(He)およびランダウ(Landau)、1995;プラネレ(Planelles)ら、1995;チェン(Chen)ら、1996;ページ(Page)ら、1997;ジャミエソン(Jamieson)およびザック(Zack)、1998)。しかしながら、これらリポーターウイルスがコードするリポーター遺伝子産物は簡単かつ迅速なアッセイによって測定できないため、これらリポーターウイルスは高容量の抗ウイルスアッセイには有用ではない。たとえば、クロラムフェニコールアセチルトランスフェラーゼ(CAT)リポーター遺伝子は厄介な酵素アッセイによって制限された範囲の感度でしか測定することができない。同様に、マーカー遺伝子をコードするリポーターウイルスは、HIV感染細胞を同定するために蛍光活性化セルソーティングを必要とする。
【0006】
(その解決方法)
本発明は、複製コンピテントなHIV−1リポーターウイルスを生成する、HIV−1プロウイルスに関する。これらプロウイルスは、ウイルス複製に必須でない領域がリポーター遺伝子、好ましくはレニラルシフェラーゼ、レニラルシフェラーゼ遺伝子の誘導体または分泌胎盤アルカリホスファターゼ(SEAP)リポーター遺伝子によって置換されたHIV−1ゲノムをコードしている。レニラルシフェラーゼ遺伝子の誘導体としては、機能的なレニラルシフェラーゼ酵素という結果となる野生型レニラルシフェラーゼ配列中でのアミノ酸またはヌクレオチド置換(すなわち、cys−ala置換)が挙げられる(リウ(Liu)ら、(1999))。リポーター遺伝子の発現は真正の(bona fide)ウイルス複製に依存しており、簡単かつ迅速なアッセイを用いて検出することができる。
【0007】
本発明の複製コンピテントなHIV−1プロウイルスは、あらゆるHIV−1ウイルス株からのいかなる複製コンピテントなHIV−1プロウイルスクローンを用いても構築することができる。一つの態様において、複製コンピテントなHIV−1プロウイルスは、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換することによって構築される。得られたプロウイルスは、ウイルス感染のためのコレセプターとしてCXCR4を利用するT細胞親和性の(T−tropic)ウイルスを生成する。使用できるリポーター遺伝子の他の例としては、SEAP遺伝子、CAT遺伝子またはグリーン蛍光タンパク質遺伝子(GFP)が挙げられるが、これらに限られるものではない。
【0008】
他の態様において、複製コンピテントなHIV−1プロウイルスの構築は、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換し、さらにpNL4−3のエンベロープ遺伝子をHIV−1プロウイルスJRFLのエンベロープ遺伝子で置換することによって行う。得られたプロウイルスは、ウイルス感染のためのコレセプターとしてCCR5を利用するマクロファージ親和性の(macrophage−tropic)ウイルスを生成する。
【0009】
さらに他の態様において、複製コンピテントなHIV−1プロウイルスは、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をSEAPリポーター遺伝子で置換し、さらにpNL4−3のエンベロープ遺伝子をHIV−1プロウイルスクローンJRFLのエンベロープ遺伝子で置換することによって生成する。得られたプロウイルス(「JRFNSEAP」と称する)は、ウイルス感染のためのコレセプターとしてCCR5を利用するマクロファージ親和性のウイルスを生成する。
【0010】
本発明のHIV−1プロウイルスを生成するのに用いることのできるエンベロープ遺伝子の他の例としては、CCまたはCXCケモカインレセプターのいずれかに対して親和性(tropism)を示すHIV−1エンベロープまたは真核生物細胞の感染を開始することのできるHIV−1単離物エンベロープが挙げられるが、これらに限られるものではない。
本発明はまた、HIV−1の複製を抑制する化合物の高処理量インビトロスクリーニングにおける本発明のHIV−1複製コンピテントリポーターウイルスの使用にも関する。
【0011】
本発明は複製コンピテントなHIV−1プロウイルスに関する。これらの複製コンピテントなHIV−1プロウイルスは、ウイルスの複製に必須でないHIV−1ウイルスゲノムの領域をリポーター遺伝子、好ましくはレニラルシフェラーゼ遺伝子、レニラルシフェラーゼ遺伝子の誘導体または分泌胎盤アルカリホスファターゼ(SEAP)リポーター遺伝子で置換することによって生成される。レニラルシフェラーゼ遺伝子の誘導体としては、機能的なレニラルシフェラーゼ酵素という結果となる野生型レニラルシフェラーゼ配列中のアミノ酸またはヌクレオチド置換(すなわち、cys−ala置換)が挙げられる(リウ(Liu)ら、(1999))。リポーター遺伝子の発現は真正のウイルス複製に依存しており、それゆえウイルス複製の定量法として用いることができる。リポーター遺伝子の発現は、簡単かつ迅速なアッセイを用いて検出することができる。
【0012】
本発明の複製コンピテントなHIV−1プロウイルスを構築するため、衰弱しない(non−debilitating)リポーター遺伝子、たとえばレニラルシフェラーゼリポーター遺伝子を、HIV−1ウイルスの複製を妨害しない限りにおいて該ウイルスゲノム中のどこにでも挿入することができる。好ましくは、ウイルスの複製に必須でないHIV−1ゲノムの領域を置換することによって該ウイルスゲノム中にリポーター遺伝子を挿入する。そのような領域としては、nef遺伝子、vpr遺伝子、およびnef遺伝子やvpr遺伝子のフラグメントが挙げられるが、これらに限られるものではない。nefおよびvpr遺伝子のフラグメントは、一般に1〜300ヌクレオチドの長さである。
【0013】
本発明の複製コンピテントなHIV−1プロウイルスは、あらゆるHIV−1ウイルス株からのいかなる複製コンピテントなHIV−1プロウイルスクローンを用いても構築することができる。そのようなクローンの例としては、pNL4−3(アダチ(Adachi)ら、1986)、pYU−2(ライ(Lai)、1991)、p89.6(コルマン(Collman)ら、1992)およびHIV−1 Lai(ウエイン−ホブソン(Wain−Hobson)ら、1985)が挙げられるが、これらに限られるものではない。
【0014】
一つの態様において、複製コンピテントなHIV−1プロウイルスはHIV−1プロウイルスクローンpNL4−3のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換することによって生成される。得られたプロウイルス(「NL4Rluc」と称する)は、ウイルス感染のためのコレセプターとしてCXCR4を利用するT細胞親和性のウイルスを生成する。
【0015】
他の態様において、複製コンピテントなHIV−1プロウイルスは、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換し、さらにpNL4−3のエンベロープ遺伝子をHIV−1プロウイルスクローンJRFLのエンベロープ遺伝子で置換することによって生成される。得られたプロウイルス(「JRFNRluc」と称する)は、ウイルス感染のためのコレセプターとしてCCR5を利用するマクロファージ親和性のウイルスを生成する。
【0016】
さらに他の態様において、複製コンピテントなHIV−1プロウイルスは、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をSEAPリポーター遺伝子で置換し、さらにpNL4−3のエンベロープ遺伝子をHIV−1プロウイルスクローンJRFLのエンベロープ遺伝子で置換することによって生成される。得られたプロウイルス(「JRFNSEAP」と称する)は、ウイルス感染のためのコレセプターとしてCCR5を利用するマクロファージ親和性のウイルスを生成する。
【0017】
本発明のHIV−1ウイルスに感染した細胞中でのレニラルシフェラーゼリポーター遺伝子、レニラルシフェラーゼ遺伝子の誘導体またはSEAPリポーター遺伝子の発現は細胞培養中での真正のウイルス複製に依存しており、当該技術分野で知られた方法を用いて検出することができる。一つの態様において、レニラルシフェラーゼ活性の測定は、該プロウイルスに感染した細胞にレニラルシフェラーゼ基質であるコエレンテラミン(coelenteramine)(Promega Corp.、マジソン、ウイスコンシン)を加えることによって行う。レニラルシフェラーゼはコエレンテラミンをコエレンテラミド(coelenteramide)に変換し、同時にCO2および光を産生するので、レニラルシフェラーゼ活性は光の放射によって検出され、光の放射はシンチレーションカウンターまたはルミノメーターを用いて定量することができる。
【0018】
他の態様において、SEAP活性の測定は、感染細胞培養からの上澄み液にSEAP基質であるCSPD(Tropix PE Applied Systems、ベッドフォード、マサチューセッツ)、ルミネセンスエンハンサー、および内生の非胎盤アルカリホスファターゼ活性を阻害する緩衝液系を加えることによって行う。SEAPによる基質の脱リン酸後に準安定なフェノラートアニオン中間体が生成し、該中間体は分解して光を放射する。光の放射はシンチレーションカウンターまたはルミノメーターを用いて測定することができる。
【0019】
レニラルシフェラーゼ活性またはSEAP活性を測定するため、当該技術分野で知られたレニラルシフェラーゼまたはSEAPの他の基質を使用できることが理解される。
細胞培養中での本発明のHIV−1ウイルスの複製は簡単かつ迅速なアッセイを用いて容易に測定することができるので、本発明はまたHIV−1複製を抑制する化合物の高容量スクリーニングにおけるこれらHIV−1ウイルスの使用にも関する。
【0020】
一つの態様において、スクリーニング法は、本発明の複製コンピテントなプロウイルスのウイルスストックに感染した細胞に被験化合物を加え、ついで2〜7日、好ましくは6日間のインキュベーション後にレニラルシフェラーゼまたはSEAP活性を測定することを含む。他の態様において、ウイルスのストックを感染させるべき細胞とともに被験化合物に加えてもよい。
抗ウイルス性の化合物のスクリーニングに用いるのに適した細胞または細胞株としては、MT−2細胞、MT−2#18細胞、PM−1、CEM SSおよびジャーカット(Jurkat)細胞が挙げられるが、これらに限られるものではない。
【0021】
被験化合物で処理した細胞でのレニラルシフェラーゼまたはSEAP活性のレベルを感染コントロール細胞のレベルと比較する。コントロール細胞でのレベルと比較して被験化合物で処理した細胞でのレニラルシフェラーゼまたはSEAP活性レベルの低減は、該被験化合物がHIV−1ウイルスの複製を抑制する能力を有していることを示している。好ましくは、抑制は50〜90%、さらに好ましくは90%以上である。
【0022】
本発明のHIV−1ウイルスのウイルスストックは当業者によく知られた方法により生成することができる。一つの態様において、ウイルスストックは、本発明のプロウイルスクローンを細胞培養中にトランスフェクションし、ついでトランスフェクションした細胞の上澄み液を回収することによって調製される。トランスフェクションは、リン酸カルシウム、エレクトロポレーションまたはリポソーム媒体トランスフェクションなどの当該技術分野で知られた方法を用いて行うことができる。トランスフェクションに用いる細胞の例としては、HEK293細胞、ジャーカット細胞およびCEM細胞が挙げられるが、これらに限られるものではない。
被験化合物で処理した細胞でのレニラルシフェラーゼまたはSEAP活性レベルの変化を測定することにより、被験化合物がウイルスの複製を抑制するうえでの有効性並びに有効性の程度を決定することができる。
【0023】
つぎに本発明を実施例により記載するが、これら実施例は本発明を説明するためのものであって本発明の範囲を限定することを意図するものではない。
実施例
材料および方法
細胞株および細胞培養条件
NIH AIDSリサーチ・アンド・レファレンス・リージェント・プログラム(NIH AIDS Research and Reference Reagent Program)(ベセスダ、メリーランド)から得たCD4+ヒト悪性T細胞株であるMT−2細胞株を、10%ウシ胎仔血清(FBS、Life Technologies)を含有するロスウエル・パーク・メモリアル・インスティチュート(Roswell Park Memorial Institute)(RPMI)1640培地(Life Technologies、ゲイサーズバーグ、メリーランド)中で増殖させた。この細胞株は本来、CD4およびCXCR4を発現するが、形質転換してマクロファージ親和性のHIV−1コレセプターであるヒトCCR5を発現するようにした。形質転換した細胞株をそのクローン集団の番号によりMT−2#18と名付け、その親細胞株MT−2と同様に0.5mg/mlのジェネティシン(Life Technologies、ゲイサーズバーグ、メリーランド)を加えて培養した。MT−2#18細胞は、T細胞親和性のウイルスに対してはCXCR4コレセプターの使用を介して、またはマクロファージ親和性のウイルスに対してはCCR5コレセプターの使用を介して、HIV−1感染を許容する。
【0024】
HEK293−Tは、SV−40ラージT抗原で形質転換したヒト胚性腎細胞株である。HEK293−T細胞を、10%ウシ胎仔血清を含有するダルベッコ改変イーグル培地(DMEM)(Life Technologies、ゲイサーズバーグ、メリーランド)中で増殖させた。
【0025】
リポーターウイルスの構築
HIV−1プロウイルスクローンpNL4−3(アダチ(Adachi)ら、1986)を用いてプロウイルスを構築した。NL4Rlucプロウイルスクローン(図1)を構築するため、pNL4−3中のエンベロープコード配列の直ぐ3’側に唯一のXbaIエンドヌクレアーゼ制限部位を導入した。導入したXbaI部位とNef中のBglIIエンドヌクレアーゼ制限部位との間の191塩基対のセグメントを欠失させ、唯一のXhoIエンドヌクレアーゼ制限部位で置換した。ついで、導入したXbaIおよびXhoI部位を用いて947塩基対のレニラルシフェラーゼリポーター遺伝子(RLuc)(マシューズ(Matthews)ら、およびローレンツ(Lorenz)ら)を改変pNL4−3中に挿入した。得られた構築物NL4RlucはpNL4−3構築物よりも756塩基対だけ長かった。
【0026】
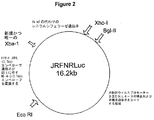
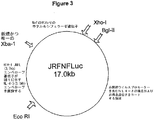
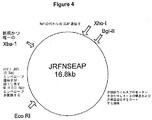
プロウイルスクローンJRFNRLuc(図2)を構築するため、NL4Rlucのエンベロープコード領域(約3,100塩基対)をエンベロープコード配列の直ぐ5’側に位置するEcoRI部位および上記唯一のXbaI部位により除去した。この断片をHIV−1 JRFLのエンベロープコード領域(約3,700塩基対)(オブライエン(O’Brien)ら、1990)で置換した。JRFLクローンのエンベロープコード領域は、この領域のPCR増幅およびEcoRIおよびXbaI部位での組換えクローニングにより調製した。JRFNFLuc cDNAクローン(図3)の構築は、JRFNRLuc中のレニラルシフェラーゼ遺伝子(947塩基対)をホタルルシフェラーゼ遺伝子(1,706塩基対)で置換することにより行った。プロウイルスクローンJRFNSEAPの構築は、XbaIおよびXhoI部位を用いてJRFNRLuc中のレニラルシフェラーゼ遺伝子をSEAPコード遺伝子(1,557塩基対)で置換することにより行った(図4)。
【0027】
ウイルスストックの調製
LipofectAMINE plus(Life Technologies、ゲイサーズバーグ、メリーランド)を用い、製造業社の指示に従ってHEK293細胞をNL4Rluc、JRFNRLuc、JRFNFLucまたはJRFNSEAPプロウイルスクローンでトランスフェクションした。各トランスフェクションに約3×107細胞を用いた。トランスフェクションの72時間後にトランスフェクションした細胞の上澄み液を回収し、低速遠心分離により清澄化し、−70℃で貯蔵した。感染性のウイルスを解凍し、MT−2#18細胞を用いて力価測定した。
【0028】
細胞の感染およびp24アッセイ
MT−2#18細胞をペレット化し、0.01の感染多重度(MOI)にてウイルスストックで37℃で2時間感染させた。ウイルスを除去し、最終濃度10,000細胞/100μLで再浮遊する前に細胞を1×PBSで2回洗浄した。感染性アッセイおよび薬剤感受性アッセイを96ウエル組織培養プレートで行い、動力学試験をT−25フラスコで行った。細胞の上澄み液を第0日目、第2日目、第3日目、第5日目または第6日目に回収し、継代したウイルスを第2日目または第3日目に回収した。ゼプトメトリックス・レトロテック(Zeptometrix Retrotek)製品(Zeptometrix Incorporated、バッファロー、ニューヨーク)を用い、製造業社のプロトコールに従ってカプシドタンパク質p24の活性を分析した。
【0029】
高処理量スクリーニングアッセイ
MT−2#18細胞の大容量培養液を調製し、10%ウシ胎仔血清を加えたRPMI培地中で0.5×106細胞/ml未満の濃度で保持した。被験化合物を96ウエル組織培養処理した透明底ブラックプレート(Corning Incorporated、コーニング、ニューヨーク)に20μlの脱イオン水および5%ジメチルスルホキシド中で3mMの濃度にてプレーティングした。MT−2#18細胞を低速遠心分離によりペレット化し、新たな培地に再浮遊させた。この新たな培地および細胞にリポーターウイルスを加えて約0.01のMOIおよび160μl中に1×104の細胞濃度とした。ついで、20μl/ウエルの2×完全改変イーグル培地を脱イオン水の容量を補正するために加えた。ついで、この混合物180μlを各被験化合物のウエルおよびコントロールのウエルに加える。37℃で5〜6日間インキュベーション後、12チャンネルマニホルドを用いてウエルから培地を吸引した。
【0030】
レニラルシフェラーゼ活性の測定は、プロメガ・デュアル・ルシフェラーゼ・リポーター・キット(Promega Dual Luciferase Reporter kit)(Promega Corp.、マジソン、ウイスコンシン)を用いて行った。マルチドロップ装置(Multidrop instrument)(Labsystems、フランクリン、マサチューセッツ)を用い、プレートの各ウエル中の露出した細胞に基質を10のバッチにて直接加えた。基質添加後、プレートを直ちに密封し、ウォラック・マイクロベータ(Wallac Microbeta)1450(Perkin−Elmer Wallac Incorporated、ゲイサーズバーグ、メリーランド)をルミネセンスモードで用いてウエル当たり5秒読み取った。
【0031】
分泌胎盤アルカリホスファターゼ(SEAP)活性の測定は、CSPDと呼ばれる1,2ジオキセタンアルカリホスファターゼ基質(Tropix PE Applied Biosystems、ベッドフォード、マサチューセッツ)を用いて行った。アッセイには、65℃にて5〜30分間、好ましくは30分間のインキュベーション、ついでL−ホモアルギニンを加えて5〜20分間、好ましくは20分間のインキュベーションを含む2つのインキュベーション工程が含まれる。光の放射はシンチレーションカウンターまたはルミノメーターを用いて測定することができる。
【0032】
実施例1
レニラルシフェラーゼリポーターウイルスの複製
よく特徴付けられたCXCR4親和性のHIV−1プロウイルスクローンであるpNL4−3構築物をHIV−1プロウイルスの構築のためのプロウイルスcDNA骨格として用いた。レニラルシフェラーゼ遺伝子をコードする3つのプロウイルスクローン、NL4Rluc、JRFNRluc、およびJRFNFLucを構築した。NL4Rlucクローンは、HIV−1ゲノム中のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換することによって構築した。NL4Rlucの子孫ビリオンは、CXCR−4をウイルス感染のためのコレセプターとして利用する。JRFNRlucクローンは、NL4Rlucクローンのエンベロープ領域をHIV−1株JRFLのエンベロープ領域で置換することによって構築した。JRFNFLucクローンは、JRFNRlucクローンのレニラルシフェラーゼリポーター遺伝子をホタルルシフェラーゼ遺伝子で置換することによって構築した。JRFNRlucおよびJRFNFLucの子孫ビリオンは、CCR5をウイルス感染のためのコレセプターとして利用する。
【0033】
これらクローンに由来するウイルスがインビトロで複製する能力を試験した。感染5日後、NL4Rlucに感染したMT−2#18細胞でのレニラルシフェラーゼ活性は非感染細胞で観察されるものと比較して1000倍以上であった(図5)。同様に、JRFNRLucに感染したMT−2#18細胞でのレニラルシフェラーゼ活性は非感染細胞で観察されるものと比較して800倍以上であった(図6)。いずれのウイルスに感染した細胞でのレニラルシフェラーゼ活性の動力学もp24(成熟ビリオン中に存在するカプシドタンパク質)の産生と相関関係があった(図5および6)。このことは、検出されたレニラルシフェラーゼ活性がHIV−1のウイルス複製の結果であることを示唆している。NL4RlucおよびJRFNRlucウイルスによるp24産生のレベルが野生型pNL4−3の子孫ウイルスのレベルと実質的に同等であるとの知見は、レニラルシフェラーゼ遺伝子によるnef遺伝子の置換がHIV−1のウイルス複製に対して有害な作用をもたらさないことを証明している(図12)。
【0034】
細胞培養のNL4RlucおよびJRFNRLuc感染の後に生成するHIV−1ウイルスが細胞培養において新たな感染の繰り返しを開始できるか否かを決定するため、MT−2#18細胞をNL4RlucまたはJRFNRLuc上澄み液に感染させ、p24活性およびレニラルシフェラーゼ活性を測定した。図7に示すように、T細胞株を6日間感染させた後に生成されるNL4Rlucウイルスはリポーター遺伝子を新たなMT−2#18細胞に形質導入することができた。同様に、T細胞株を6日間感染させた後に生成されるJRFNRLucビリオンはリポーター遺伝子を新たなMT−2#18細胞に形質導入することができた(図8)。これら結果は、NL4RlucおよびJRFNRLucウイルスが感染細胞培養から伝播して新たな細胞培養において多数の複製の繰り返しを開始できること、それゆえ複製コンピテントであることを証明している。
【0035】
HIV−1プロテアーゼインヒビターのアンプレナビル(Amprenavir)(Glaxo Wellcome、リサーチ・トライアングル・パーク、ノースカロライナ)および非ヌクレオシド逆転写酵素インヒビターのエファビレンツ(Efavirenz)(Dupont Pharmaceuticals、ウイルミントン、デラウエア)を用い、NL4RlucおよびJRFNRLucウイルスの複製特性を調べた。アンプレナビルおよびエファビレンツはHIV−1感染の介入のために臨床的に用いられているものである。逆転写酵素インヒビターであるエファビレンツはHIV−1複製サイクルの初期で作用するのに対し、プロテアーゼインヒビターであるアンプレナビルはHIV−1複製サイクルの後期で作用する。
【0036】
表1に示すように、NL4RlucおよびJRFNRLucによるMT−2#18細胞の感染はエファビレンツ並びにアンプレナビルによって抑制された。アンプレナビルはNL4RlucおよびJRFNRLucの複製をそれぞれ19.4nMおよび6.9nMの50有効濃度(EC50)値で抑制し、エファビレンツはNL4RlucおよびJRFNRLucの複製をそれぞれ0.36nMおよび0.12nMのEC50で抑制した。HIV−1プロテアーゼインヒビターであるアンプレナビルは複製サイクルの後期にビリオンの成熟に際して作用し、単一サイクルの感染は抑制しない。それゆえ、これら結果はさらに、観察されたレニラルシフェラーゼリポーター遺伝子活性がNL4RlucまたはJRFNRLucの真正の複製の結果であることを証明している。
【0037】
表1
アンプレナビルおよびエファビレンツによるウイルス複製の抑制
 【0038】
【0038】
ついで、NL4RlucおよびJRFNRLucウイルスの複製能をJRFNFLucウイルスの複製能と比較した。JRFNFLucはJRFNRLucとレニラルシフェラーゼ遺伝子の代わりにホタルルシフェラーゼ遺伝子をコードしている点で異なる。JRFNFLucの構築は以前に報告されたのと同様の仕方で行った(チェン(Chen)ら、1994)。JRFNFLucによるT細胞株の感染は、NL4RlucおよびJRFNRLucウイルスと全く同様にして行った。
【0039】
図9に示すように、JRFNFLucによる細胞の感染は、感染3日後に非感染細胞で観察されるものと比較して200倍以上のホタルルシフェラーゼ活性の産生という結果となった。しかしながら、この活性レベルはこの時点以後に増大することはなく、p24活性も6日間の感染期間を通じてバックグラウンドレベルを超えることはなかった。これら結果は、成熟ウイルスコア粒子が生成されなかったことを示している。6日間感染後のJRFNFLuc上澄み液を非感染MT−2#18細胞に継代したところ、接種7日後までホタルルシフェラーゼ活性もp24産生も検出できなかった(データは示していない)。これら結果は、これら上澄み液が新たな複製の繰り返しを開始できるウイルスを含んでいなかったことを示唆している。
【0040】
最後に、非ヌクレオシド逆転写酵素インヒビターであるエファビレンツはJRFNFLucを0.18nMの50有効濃度(EC50)値で抑制したが、HIV−1プロテアーゼインヒビターであるアンプレナビルは複製コンピテントなJRFNRLucの1000倍以上に相当する濃度の10μMでも該ウイルスを抑制できなかった(表1)。このウイルスが後期HIV−1複製サイクルインヒビターによって抑制することができないとの知見は、このウイルスが複製できないことのさらなる証拠である。以上を総合すると、これらデータは、JRFNFLucウイルスが感染の単一の繰り返しを行うことができるのみであり、複製コンピテントではないのに対し、NL4RlucおよびJRFNRLucは感染の多数の繰り返しを行うことができ、真正に複製コンピテントなプロウイルスであることを強く示唆している。
【0041】
実施例2
SEAPリポーターウイルスの複製
JRFNRLuc中のレニラルシフェラーゼ遺伝子をSEAPをコードする遺伝子で置換することによりJRFNSEAPプロウイルス構築物を生成した。このプロウイルスクローンによって生成したウイルスがインビトロで複製する能力を試験した。感染6日後、JRFNSEAPに感染したMT−2#18細胞でのSEAP活性は非感染細胞で観察される活性の150倍以上であった。このウイルスに感染した細胞でのSEAP活性の動力学はp24活性と相関関係があった(図10)。このことは、検出されたSEAP活性がHIV−1のウイルス複製の結果であることを示唆している。JRFNSEAPウイルスによるp24産生のレベルが野生型pNL4−3の子孫ウイルスのレベルと実質的に同等であるとの知見は、SEAP遺伝子によるnef遺伝子の置換がHIV−1のウイルス複製に対して有害な作用をもたらさないことを証明している(図12)。
【0042】
ついで、細胞培養の感染後に生成するJRFNSEAPウイルスが細胞培養において新たな感染の繰り返しを開始できるか否かを決定するための試験を行った。図11に示すように、T細胞株を6日間感染させた後に生成されるJRFNSEAPウイルスはリポーター遺伝子を新たなMT−2#18細胞に形質導入することができた。これら結果は、JRFNSEAPウイルスが感染細胞培養から伝播して新たな細胞培養において多数の複製の繰り返しを開始できること、それゆえ複製コンピテントであることを証明している。
【0043】
上記HIV−1プロテアーゼインヒビターであるアンプレナビルおよび非ヌクレオシド逆転写酵素インヒビターであるエファビレンツを用い、JRFNSEAPウイルスの複製特性を調べた。表1に示すように、JRFNSEAPによるMT−2#18細胞の感染はエファビレンツ並びにアンプレナビルによって抑制された。アンプレナビルはJRFNSEAPの複製を21.5nMの50有効濃度(EC50)値で抑制し、エファビレンツはJRFNSEAPの複製を0.10nMのEC50で抑制した。HIV−1プロテアーゼインヒビターであるアンプレナビルは複製サイクルの後期にビリオンの成熟に際して作用し、単一サイクルの感染は抑制しない。それゆえ、この知見は、観察されたSEAPリポーター遺伝子活性が真正のウイルス複製の結果であることをさらに証明している。
【0044】
参照文献
1.チェン(Chen)ら、1994, J. Virol. 68: 654−660
2.ターウィリガー(Terwilliger)ら、1989, PNAS 86: 3857−3861
3.マリム(Malim)ら、1992, J. Exp. Med. 176: 1197−1201
4.ヘ(He)およびランダウ(Landau)、1995, J. Virol. 69: 4587−4592
5.プラネレ(Planelles)ら、1995, Gene Therapy 2: 369−376
6.チェン(Chen)ら、1996, J. Virol. 70: 6044−6053
7.ページ(Page)ら、1997, Aids Research and Human Retro. 13: 1077−1081
8.ジャミエソン(Jamieson)およびザック(Zack)、1998, J. Virol. 72: 6520−6526
9.ワイスロウ(Weislow)ら、1989, J. of Nat. Cancer Inst. 81: 577−586
【0045】
10.アダチ(Adachi)ら、1986, J. Virol. 59: 284−291
11.オブライエン(O’Brien)ら、1990, Nature 348: 69−73
12.マシューズ(Matthews)ら、1977, Biochemistry 16: 85
13.エディンガー(Edinger)ら、1998, J. Virol. 72: 7934−7940
14.ウエイン−ホブソン(Wain−Hobson)ら、1985, Cell 40: 9−17
15.リ(Li)ら、1991, J. Virol. 65: 3973−3985
16.コルマン(Collman)ら、1992, J. Virol. 66: 7517−7521
17.ローレンツ(Lorenz)ら、1991, P.N.A.S. USA 88: 4438
18.リウ(Liu)ら、1999, Gene 237: 153−159
【0046】
本明細書に引用した全ての特許、特許出願、公開文献、参照マニュアル、出典および要約の内容は、本発明が関連する技術分野の技術の状態を一層完全に記載するためにその全体が参照のため引用される。
上記に記載した組成物および方法には本発明の範囲から逸脱しない限り種々の変更を施すことが可能なので、上記記載に含まれ、添付の図面に示され、または請求の範囲で定められる事項は全て例示のものとして解釈されるのであって限定的な意味でないことが意図される。
【図面の簡単な説明】
【図1】NL4RLucプロウイルス構築物の模式図である。
【図2】JRFNRLucプロウイルス構築物の模式図である。
【図3】JRFNFLucプロウイルス構築物の模式図である。
【図4】JRFNSEAPプロウイルス構築物の模式図である。
【図5】HIV−1プロウイルスpNL4RLucによって生成したHIV−1ウイルスの複製の動力学を示す。第0日目、第2日目および第5日目に測定したレニラルシフェラーゼ活性および成熟ウイルスカプシドタンパク質p24レベルを示す。
【図6】HIV−1プロウイルスJRFNRlucによって生成したHIV−1ウイルスの複製の動力学を示す。第0日目、第3日目および第6日目に測定したレニラルシフェラーゼ活性および成熟ウイルスカプシドタンパク質p24レベルを示す。
【図7】MT−2#18細胞のpNL4Rluc感染後に生成したHIV−1ウイルスはMT−2#18細胞において新たな感染の繰り返しを開始することができたことを示す。
【図8】MT−2#18細胞のJRFNRluc感染後に生成したHIV−1ウイルスはMT−2#18細胞において新たな感染の繰り返しを開始することができたことを示す。
【図9】HIV−1プロウイルスJRFNFLucによって生成したHIV−1ウイルスの複製の動力学を示す。
【図10】HIV−1プロウイルスJRFNSEAPによって生成したHIV−1ウイルスの複製の動力学を示す。第0日目、第3日目および第6日目に測定したSEAP活性および成熟ウイルスカプシドタンパク質p24レベルを示す。
【図11】MT−2#18細胞のJRFNSEAP感染後に生成したHIV−1ウイルスはMT−2#18細胞において新たな感染の繰り返しを開始することができたことを示す。
【図12】種々のHIV−1リポーターウイルスおよびpNL4−3ウイルスのp24活性を示す。
(技術分野)
本発明は、リポーター遺伝子、好ましくはレニラ(renilla)ルシフェラーゼか分泌胎盤アルカリホスファターゼ(secreted placental alkaline phosphatase)(SEAP)のいずれかのリポーター遺伝子をコードする、複製コンピテントな(replication competent)HIV−1プロウイルス構築物に関する。本発明はさらに、HIV−1の複製を抑制する化合物の高処理量スクリーニングにおけるこれらプロウイルス構築物ビリオンの使用に関する。
【0002】
(背景技術)
1型ヒト免疫不全ウイルス(HIV−1)の細胞培養中での複製は、一般に、ウイルスの逆転写酵素(RT)活性の産生を定量するか、細胞培養液中の成熟ウイルスカプシドタンパク質(すなわち、p24)を定量するか、または逆転写酵素によって指令された複製連鎖反応(RT−PCR)によるウイルスRNAコピー数を定量することによって測定される。あるいは、HIV−1複製は、XTTやMTTなどの色素還元(dye reduction)法を用いて感染細胞に対するウイルスによって誘発された細胞変性効果(cpe)を定量することによって間接的に測定することができる(ワイスロウ(Weislow)ら、1989)。
【0003】
HIV−1 RTアッセイまたはp24アッセイの場合、これら方法の厄介な性質はこれら方法を高処理量の抗ウイルスアッセイに適用するのを排除している。一方、XTTなどの色素還元法によるHIV複製の分析は単一工程のプロセスであり、それゆえ高処理量が可能である。しかしながら、色素還元法はしばしば培養液中の大部分の細胞が感染し死滅した後に行われるので、一層有効性の少ないHIV−1インヒビターの同定の感度の低減という結果となる。さらに、色素還元法は有意の細胞変性効果に依存しているので、シンシチウム非誘導性のHIV−1株のように培養細胞株中で攻撃的な(aggressive)複製動力学を示さないHIV株の複製を正確に測定するのに用いることはできない。
【0004】
(発明の開示)
(発明が解決しようとする技術的課題)
HIV−1リポーターウイルスの生成は、HIV−1複製の測定の従来法に代わる方法を提供する。リポーター遺伝子としてルシフェラーゼをコードする単サイクル感染性HIV−1リポーターウイルスが記載されている(チェン(Chen)ら、1994)。これらウイルスは、ウイルスの侵入、逆転写、組込み、および遺伝子発現を含むHIV−1複製サイクルのほぼ最初の半分の簡単かつ迅速な分析を可能にする。しかしながら、感染細胞でのHIV遺伝子発現の後の工程、たとえばビリオンの成熟に必要なウイルス前駆体ポリペプチドのHIVプロテアーゼ媒体プロセシングなどは、これらウイルスを用いては容易に測定できない。
【0005】
HIVの完全な複製サイクルを分析し、細胞集団中のHIV−1の伝播を追跡するには、複製コンピテントなリポーターウイルスが必要である。HIV−1の複製の分析に用いることのできる幾つかの複製コンピテントなHIV−1リポーターウイルスが作製されている(ターウィリガー(Terwilliger)ら、1989;マリム(Malim)ら、1992;ヘ(He)およびランダウ(Landau)、1995;プラネレ(Planelles)ら、1995;チェン(Chen)ら、1996;ページ(Page)ら、1997;ジャミエソン(Jamieson)およびザック(Zack)、1998)。しかしながら、これらリポーターウイルスがコードするリポーター遺伝子産物は簡単かつ迅速なアッセイによって測定できないため、これらリポーターウイルスは高容量の抗ウイルスアッセイには有用ではない。たとえば、クロラムフェニコールアセチルトランスフェラーゼ(CAT)リポーター遺伝子は厄介な酵素アッセイによって制限された範囲の感度でしか測定することができない。同様に、マーカー遺伝子をコードするリポーターウイルスは、HIV感染細胞を同定するために蛍光活性化セルソーティングを必要とする。
【0006】
(その解決方法)
本発明は、複製コンピテントなHIV−1リポーターウイルスを生成する、HIV−1プロウイルスに関する。これらプロウイルスは、ウイルス複製に必須でない領域がリポーター遺伝子、好ましくはレニラルシフェラーゼ、レニラルシフェラーゼ遺伝子の誘導体または分泌胎盤アルカリホスファターゼ(SEAP)リポーター遺伝子によって置換されたHIV−1ゲノムをコードしている。レニラルシフェラーゼ遺伝子の誘導体としては、機能的なレニラルシフェラーゼ酵素という結果となる野生型レニラルシフェラーゼ配列中でのアミノ酸またはヌクレオチド置換(すなわち、cys−ala置換)が挙げられる(リウ(Liu)ら、(1999))。リポーター遺伝子の発現は真正の(bona fide)ウイルス複製に依存しており、簡単かつ迅速なアッセイを用いて検出することができる。
【0007】
本発明の複製コンピテントなHIV−1プロウイルスは、あらゆるHIV−1ウイルス株からのいかなる複製コンピテントなHIV−1プロウイルスクローンを用いても構築することができる。一つの態様において、複製コンピテントなHIV−1プロウイルスは、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換することによって構築される。得られたプロウイルスは、ウイルス感染のためのコレセプターとしてCXCR4を利用するT細胞親和性の(T−tropic)ウイルスを生成する。使用できるリポーター遺伝子の他の例としては、SEAP遺伝子、CAT遺伝子またはグリーン蛍光タンパク質遺伝子(GFP)が挙げられるが、これらに限られるものではない。
【0008】
他の態様において、複製コンピテントなHIV−1プロウイルスの構築は、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換し、さらにpNL4−3のエンベロープ遺伝子をHIV−1プロウイルスJRFLのエンベロープ遺伝子で置換することによって行う。得られたプロウイルスは、ウイルス感染のためのコレセプターとしてCCR5を利用するマクロファージ親和性の(macrophage−tropic)ウイルスを生成する。
【0009】
さらに他の態様において、複製コンピテントなHIV−1プロウイルスは、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をSEAPリポーター遺伝子で置換し、さらにpNL4−3のエンベロープ遺伝子をHIV−1プロウイルスクローンJRFLのエンベロープ遺伝子で置換することによって生成する。得られたプロウイルス(「JRFNSEAP」と称する)は、ウイルス感染のためのコレセプターとしてCCR5を利用するマクロファージ親和性のウイルスを生成する。
【0010】
本発明のHIV−1プロウイルスを生成するのに用いることのできるエンベロープ遺伝子の他の例としては、CCまたはCXCケモカインレセプターのいずれかに対して親和性(tropism)を示すHIV−1エンベロープまたは真核生物細胞の感染を開始することのできるHIV−1単離物エンベロープが挙げられるが、これらに限られるものではない。
本発明はまた、HIV−1の複製を抑制する化合物の高処理量インビトロスクリーニングにおける本発明のHIV−1複製コンピテントリポーターウイルスの使用にも関する。
【0011】
本発明は複製コンピテントなHIV−1プロウイルスに関する。これらの複製コンピテントなHIV−1プロウイルスは、ウイルスの複製に必須でないHIV−1ウイルスゲノムの領域をリポーター遺伝子、好ましくはレニラルシフェラーゼ遺伝子、レニラルシフェラーゼ遺伝子の誘導体または分泌胎盤アルカリホスファターゼ(SEAP)リポーター遺伝子で置換することによって生成される。レニラルシフェラーゼ遺伝子の誘導体としては、機能的なレニラルシフェラーゼ酵素という結果となる野生型レニラルシフェラーゼ配列中のアミノ酸またはヌクレオチド置換(すなわち、cys−ala置換)が挙げられる(リウ(Liu)ら、(1999))。リポーター遺伝子の発現は真正のウイルス複製に依存しており、それゆえウイルス複製の定量法として用いることができる。リポーター遺伝子の発現は、簡単かつ迅速なアッセイを用いて検出することができる。
【0012】
本発明の複製コンピテントなHIV−1プロウイルスを構築するため、衰弱しない(non−debilitating)リポーター遺伝子、たとえばレニラルシフェラーゼリポーター遺伝子を、HIV−1ウイルスの複製を妨害しない限りにおいて該ウイルスゲノム中のどこにでも挿入することができる。好ましくは、ウイルスの複製に必須でないHIV−1ゲノムの領域を置換することによって該ウイルスゲノム中にリポーター遺伝子を挿入する。そのような領域としては、nef遺伝子、vpr遺伝子、およびnef遺伝子やvpr遺伝子のフラグメントが挙げられるが、これらに限られるものではない。nefおよびvpr遺伝子のフラグメントは、一般に1〜300ヌクレオチドの長さである。
【0013】
本発明の複製コンピテントなHIV−1プロウイルスは、あらゆるHIV−1ウイルス株からのいかなる複製コンピテントなHIV−1プロウイルスクローンを用いても構築することができる。そのようなクローンの例としては、pNL4−3(アダチ(Adachi)ら、1986)、pYU−2(ライ(Lai)、1991)、p89.6(コルマン(Collman)ら、1992)およびHIV−1 Lai(ウエイン−ホブソン(Wain−Hobson)ら、1985)が挙げられるが、これらに限られるものではない。
【0014】
一つの態様において、複製コンピテントなHIV−1プロウイルスはHIV−1プロウイルスクローンpNL4−3のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換することによって生成される。得られたプロウイルス(「NL4Rluc」と称する)は、ウイルス感染のためのコレセプターとしてCXCR4を利用するT細胞親和性のウイルスを生成する。
【0015】
他の態様において、複製コンピテントなHIV−1プロウイルスは、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換し、さらにpNL4−3のエンベロープ遺伝子をHIV−1プロウイルスクローンJRFLのエンベロープ遺伝子で置換することによって生成される。得られたプロウイルス(「JRFNRluc」と称する)は、ウイルス感染のためのコレセプターとしてCCR5を利用するマクロファージ親和性のウイルスを生成する。
【0016】
さらに他の態様において、複製コンピテントなHIV−1プロウイルスは、HIV−1プロウイルスクローンpNL4−3のnef遺伝子をSEAPリポーター遺伝子で置換し、さらにpNL4−3のエンベロープ遺伝子をHIV−1プロウイルスクローンJRFLのエンベロープ遺伝子で置換することによって生成される。得られたプロウイルス(「JRFNSEAP」と称する)は、ウイルス感染のためのコレセプターとしてCCR5を利用するマクロファージ親和性のウイルスを生成する。
【0017】
本発明のHIV−1ウイルスに感染した細胞中でのレニラルシフェラーゼリポーター遺伝子、レニラルシフェラーゼ遺伝子の誘導体またはSEAPリポーター遺伝子の発現は細胞培養中での真正のウイルス複製に依存しており、当該技術分野で知られた方法を用いて検出することができる。一つの態様において、レニラルシフェラーゼ活性の測定は、該プロウイルスに感染した細胞にレニラルシフェラーゼ基質であるコエレンテラミン(coelenteramine)(Promega Corp.、マジソン、ウイスコンシン)を加えることによって行う。レニラルシフェラーゼはコエレンテラミンをコエレンテラミド(coelenteramide)に変換し、同時にCO2および光を産生するので、レニラルシフェラーゼ活性は光の放射によって検出され、光の放射はシンチレーションカウンターまたはルミノメーターを用いて定量することができる。
【0018】
他の態様において、SEAP活性の測定は、感染細胞培養からの上澄み液にSEAP基質であるCSPD(Tropix PE Applied Systems、ベッドフォード、マサチューセッツ)、ルミネセンスエンハンサー、および内生の非胎盤アルカリホスファターゼ活性を阻害する緩衝液系を加えることによって行う。SEAPによる基質の脱リン酸後に準安定なフェノラートアニオン中間体が生成し、該中間体は分解して光を放射する。光の放射はシンチレーションカウンターまたはルミノメーターを用いて測定することができる。
【0019】
レニラルシフェラーゼ活性またはSEAP活性を測定するため、当該技術分野で知られたレニラルシフェラーゼまたはSEAPの他の基質を使用できることが理解される。
細胞培養中での本発明のHIV−1ウイルスの複製は簡単かつ迅速なアッセイを用いて容易に測定することができるので、本発明はまたHIV−1複製を抑制する化合物の高容量スクリーニングにおけるこれらHIV−1ウイルスの使用にも関する。
【0020】
一つの態様において、スクリーニング法は、本発明の複製コンピテントなプロウイルスのウイルスストックに感染した細胞に被験化合物を加え、ついで2〜7日、好ましくは6日間のインキュベーション後にレニラルシフェラーゼまたはSEAP活性を測定することを含む。他の態様において、ウイルスのストックを感染させるべき細胞とともに被験化合物に加えてもよい。
抗ウイルス性の化合物のスクリーニングに用いるのに適した細胞または細胞株としては、MT−2細胞、MT−2#18細胞、PM−1、CEM SSおよびジャーカット(Jurkat)細胞が挙げられるが、これらに限られるものではない。
【0021】
被験化合物で処理した細胞でのレニラルシフェラーゼまたはSEAP活性のレベルを感染コントロール細胞のレベルと比較する。コントロール細胞でのレベルと比較して被験化合物で処理した細胞でのレニラルシフェラーゼまたはSEAP活性レベルの低減は、該被験化合物がHIV−1ウイルスの複製を抑制する能力を有していることを示している。好ましくは、抑制は50〜90%、さらに好ましくは90%以上である。
【0022】
本発明のHIV−1ウイルスのウイルスストックは当業者によく知られた方法により生成することができる。一つの態様において、ウイルスストックは、本発明のプロウイルスクローンを細胞培養中にトランスフェクションし、ついでトランスフェクションした細胞の上澄み液を回収することによって調製される。トランスフェクションは、リン酸カルシウム、エレクトロポレーションまたはリポソーム媒体トランスフェクションなどの当該技術分野で知られた方法を用いて行うことができる。トランスフェクションに用いる細胞の例としては、HEK293細胞、ジャーカット細胞およびCEM細胞が挙げられるが、これらに限られるものではない。
被験化合物で処理した細胞でのレニラルシフェラーゼまたはSEAP活性レベルの変化を測定することにより、被験化合物がウイルスの複製を抑制するうえでの有効性並びに有効性の程度を決定することができる。
【0023】
つぎに本発明を実施例により記載するが、これら実施例は本発明を説明するためのものであって本発明の範囲を限定することを意図するものではない。
実施例
材料および方法
細胞株および細胞培養条件
NIH AIDSリサーチ・アンド・レファレンス・リージェント・プログラム(NIH AIDS Research and Reference Reagent Program)(ベセスダ、メリーランド)から得たCD4+ヒト悪性T細胞株であるMT−2細胞株を、10%ウシ胎仔血清(FBS、Life Technologies)を含有するロスウエル・パーク・メモリアル・インスティチュート(Roswell Park Memorial Institute)(RPMI)1640培地(Life Technologies、ゲイサーズバーグ、メリーランド)中で増殖させた。この細胞株は本来、CD4およびCXCR4を発現するが、形質転換してマクロファージ親和性のHIV−1コレセプターであるヒトCCR5を発現するようにした。形質転換した細胞株をそのクローン集団の番号によりMT−2#18と名付け、その親細胞株MT−2と同様に0.5mg/mlのジェネティシン(Life Technologies、ゲイサーズバーグ、メリーランド)を加えて培養した。MT−2#18細胞は、T細胞親和性のウイルスに対してはCXCR4コレセプターの使用を介して、またはマクロファージ親和性のウイルスに対してはCCR5コレセプターの使用を介して、HIV−1感染を許容する。
【0024】
HEK293−Tは、SV−40ラージT抗原で形質転換したヒト胚性腎細胞株である。HEK293−T細胞を、10%ウシ胎仔血清を含有するダルベッコ改変イーグル培地(DMEM)(Life Technologies、ゲイサーズバーグ、メリーランド)中で増殖させた。
【0025】
リポーターウイルスの構築
HIV−1プロウイルスクローンpNL4−3(アダチ(Adachi)ら、1986)を用いてプロウイルスを構築した。NL4Rlucプロウイルスクローン(図1)を構築するため、pNL4−3中のエンベロープコード配列の直ぐ3’側に唯一のXbaIエンドヌクレアーゼ制限部位を導入した。導入したXbaI部位とNef中のBglIIエンドヌクレアーゼ制限部位との間の191塩基対のセグメントを欠失させ、唯一のXhoIエンドヌクレアーゼ制限部位で置換した。ついで、導入したXbaIおよびXhoI部位を用いて947塩基対のレニラルシフェラーゼリポーター遺伝子(RLuc)(マシューズ(Matthews)ら、およびローレンツ(Lorenz)ら)を改変pNL4−3中に挿入した。得られた構築物NL4RlucはpNL4−3構築物よりも756塩基対だけ長かった。
【0026】
プロウイルスクローンJRFNRLuc(図2)を構築するため、NL4Rlucのエンベロープコード領域(約3,100塩基対)をエンベロープコード配列の直ぐ5’側に位置するEcoRI部位および上記唯一のXbaI部位により除去した。この断片をHIV−1 JRFLのエンベロープコード領域(約3,700塩基対)(オブライエン(O’Brien)ら、1990)で置換した。JRFLクローンのエンベロープコード領域は、この領域のPCR増幅およびEcoRIおよびXbaI部位での組換えクローニングにより調製した。JRFNFLuc cDNAクローン(図3)の構築は、JRFNRLuc中のレニラルシフェラーゼ遺伝子(947塩基対)をホタルルシフェラーゼ遺伝子(1,706塩基対)で置換することにより行った。プロウイルスクローンJRFNSEAPの構築は、XbaIおよびXhoI部位を用いてJRFNRLuc中のレニラルシフェラーゼ遺伝子をSEAPコード遺伝子(1,557塩基対)で置換することにより行った(図4)。
【0027】
ウイルスストックの調製
LipofectAMINE plus(Life Technologies、ゲイサーズバーグ、メリーランド)を用い、製造業社の指示に従ってHEK293細胞をNL4Rluc、JRFNRLuc、JRFNFLucまたはJRFNSEAPプロウイルスクローンでトランスフェクションした。各トランスフェクションに約3×107細胞を用いた。トランスフェクションの72時間後にトランスフェクションした細胞の上澄み液を回収し、低速遠心分離により清澄化し、−70℃で貯蔵した。感染性のウイルスを解凍し、MT−2#18細胞を用いて力価測定した。
【0028】
細胞の感染およびp24アッセイ
MT−2#18細胞をペレット化し、0.01の感染多重度(MOI)にてウイルスストックで37℃で2時間感染させた。ウイルスを除去し、最終濃度10,000細胞/100μLで再浮遊する前に細胞を1×PBSで2回洗浄した。感染性アッセイおよび薬剤感受性アッセイを96ウエル組織培養プレートで行い、動力学試験をT−25フラスコで行った。細胞の上澄み液を第0日目、第2日目、第3日目、第5日目または第6日目に回収し、継代したウイルスを第2日目または第3日目に回収した。ゼプトメトリックス・レトロテック(Zeptometrix Retrotek)製品(Zeptometrix Incorporated、バッファロー、ニューヨーク)を用い、製造業社のプロトコールに従ってカプシドタンパク質p24の活性を分析した。
【0029】
高処理量スクリーニングアッセイ
MT−2#18細胞の大容量培養液を調製し、10%ウシ胎仔血清を加えたRPMI培地中で0.5×106細胞/ml未満の濃度で保持した。被験化合物を96ウエル組織培養処理した透明底ブラックプレート(Corning Incorporated、コーニング、ニューヨーク)に20μlの脱イオン水および5%ジメチルスルホキシド中で3mMの濃度にてプレーティングした。MT−2#18細胞を低速遠心分離によりペレット化し、新たな培地に再浮遊させた。この新たな培地および細胞にリポーターウイルスを加えて約0.01のMOIおよび160μl中に1×104の細胞濃度とした。ついで、20μl/ウエルの2×完全改変イーグル培地を脱イオン水の容量を補正するために加えた。ついで、この混合物180μlを各被験化合物のウエルおよびコントロールのウエルに加える。37℃で5〜6日間インキュベーション後、12チャンネルマニホルドを用いてウエルから培地を吸引した。
【0030】
レニラルシフェラーゼ活性の測定は、プロメガ・デュアル・ルシフェラーゼ・リポーター・キット(Promega Dual Luciferase Reporter kit)(Promega Corp.、マジソン、ウイスコンシン)を用いて行った。マルチドロップ装置(Multidrop instrument)(Labsystems、フランクリン、マサチューセッツ)を用い、プレートの各ウエル中の露出した細胞に基質を10のバッチにて直接加えた。基質添加後、プレートを直ちに密封し、ウォラック・マイクロベータ(Wallac Microbeta)1450(Perkin−Elmer Wallac Incorporated、ゲイサーズバーグ、メリーランド)をルミネセンスモードで用いてウエル当たり5秒読み取った。
【0031】
分泌胎盤アルカリホスファターゼ(SEAP)活性の測定は、CSPDと呼ばれる1,2ジオキセタンアルカリホスファターゼ基質(Tropix PE Applied Biosystems、ベッドフォード、マサチューセッツ)を用いて行った。アッセイには、65℃にて5〜30分間、好ましくは30分間のインキュベーション、ついでL−ホモアルギニンを加えて5〜20分間、好ましくは20分間のインキュベーションを含む2つのインキュベーション工程が含まれる。光の放射はシンチレーションカウンターまたはルミノメーターを用いて測定することができる。
【0032】
実施例1
レニラルシフェラーゼリポーターウイルスの複製
よく特徴付けられたCXCR4親和性のHIV−1プロウイルスクローンであるpNL4−3構築物をHIV−1プロウイルスの構築のためのプロウイルスcDNA骨格として用いた。レニラルシフェラーゼ遺伝子をコードする3つのプロウイルスクローン、NL4Rluc、JRFNRluc、およびJRFNFLucを構築した。NL4Rlucクローンは、HIV−1ゲノム中のnef遺伝子をレニラルシフェラーゼリポーター遺伝子で置換することによって構築した。NL4Rlucの子孫ビリオンは、CXCR−4をウイルス感染のためのコレセプターとして利用する。JRFNRlucクローンは、NL4Rlucクローンのエンベロープ領域をHIV−1株JRFLのエンベロープ領域で置換することによって構築した。JRFNFLucクローンは、JRFNRlucクローンのレニラルシフェラーゼリポーター遺伝子をホタルルシフェラーゼ遺伝子で置換することによって構築した。JRFNRlucおよびJRFNFLucの子孫ビリオンは、CCR5をウイルス感染のためのコレセプターとして利用する。
【0033】
これらクローンに由来するウイルスがインビトロで複製する能力を試験した。感染5日後、NL4Rlucに感染したMT−2#18細胞でのレニラルシフェラーゼ活性は非感染細胞で観察されるものと比較して1000倍以上であった(図5)。同様に、JRFNRLucに感染したMT−2#18細胞でのレニラルシフェラーゼ活性は非感染細胞で観察されるものと比較して800倍以上であった(図6)。いずれのウイルスに感染した細胞でのレニラルシフェラーゼ活性の動力学もp24(成熟ビリオン中に存在するカプシドタンパク質)の産生と相関関係があった(図5および6)。このことは、検出されたレニラルシフェラーゼ活性がHIV−1のウイルス複製の結果であることを示唆している。NL4RlucおよびJRFNRlucウイルスによるp24産生のレベルが野生型pNL4−3の子孫ウイルスのレベルと実質的に同等であるとの知見は、レニラルシフェラーゼ遺伝子によるnef遺伝子の置換がHIV−1のウイルス複製に対して有害な作用をもたらさないことを証明している(図12)。
【0034】
細胞培養のNL4RlucおよびJRFNRLuc感染の後に生成するHIV−1ウイルスが細胞培養において新たな感染の繰り返しを開始できるか否かを決定するため、MT−2#18細胞をNL4RlucまたはJRFNRLuc上澄み液に感染させ、p24活性およびレニラルシフェラーゼ活性を測定した。図7に示すように、T細胞株を6日間感染させた後に生成されるNL4Rlucウイルスはリポーター遺伝子を新たなMT−2#18細胞に形質導入することができた。同様に、T細胞株を6日間感染させた後に生成されるJRFNRLucビリオンはリポーター遺伝子を新たなMT−2#18細胞に形質導入することができた(図8)。これら結果は、NL4RlucおよびJRFNRLucウイルスが感染細胞培養から伝播して新たな細胞培養において多数の複製の繰り返しを開始できること、それゆえ複製コンピテントであることを証明している。
【0035】
HIV−1プロテアーゼインヒビターのアンプレナビル(Amprenavir)(Glaxo Wellcome、リサーチ・トライアングル・パーク、ノースカロライナ)および非ヌクレオシド逆転写酵素インヒビターのエファビレンツ(Efavirenz)(Dupont Pharmaceuticals、ウイルミントン、デラウエア)を用い、NL4RlucおよびJRFNRLucウイルスの複製特性を調べた。アンプレナビルおよびエファビレンツはHIV−1感染の介入のために臨床的に用いられているものである。逆転写酵素インヒビターであるエファビレンツはHIV−1複製サイクルの初期で作用するのに対し、プロテアーゼインヒビターであるアンプレナビルはHIV−1複製サイクルの後期で作用する。
【0036】
表1に示すように、NL4RlucおよびJRFNRLucによるMT−2#18細胞の感染はエファビレンツ並びにアンプレナビルによって抑制された。アンプレナビルはNL4RlucおよびJRFNRLucの複製をそれぞれ19.4nMおよび6.9nMの50有効濃度(EC50)値で抑制し、エファビレンツはNL4RlucおよびJRFNRLucの複製をそれぞれ0.36nMおよび0.12nMのEC50で抑制した。HIV−1プロテアーゼインヒビターであるアンプレナビルは複製サイクルの後期にビリオンの成熟に際して作用し、単一サイクルの感染は抑制しない。それゆえ、これら結果はさらに、観察されたレニラルシフェラーゼリポーター遺伝子活性がNL4RlucまたはJRFNRLucの真正の複製の結果であることを証明している。
【0037】
表1
アンプレナビルおよびエファビレンツによるウイルス複製の抑制
ついで、NL4RlucおよびJRFNRLucウイルスの複製能をJRFNFLucウイルスの複製能と比較した。JRFNFLucはJRFNRLucとレニラルシフェラーゼ遺伝子の代わりにホタルルシフェラーゼ遺伝子をコードしている点で異なる。JRFNFLucの構築は以前に報告されたのと同様の仕方で行った(チェン(Chen)ら、1994)。JRFNFLucによるT細胞株の感染は、NL4RlucおよびJRFNRLucウイルスと全く同様にして行った。
【0039】
図9に示すように、JRFNFLucによる細胞の感染は、感染3日後に非感染細胞で観察されるものと比較して200倍以上のホタルルシフェラーゼ活性の産生という結果となった。しかしながら、この活性レベルはこの時点以後に増大することはなく、p24活性も6日間の感染期間を通じてバックグラウンドレベルを超えることはなかった。これら結果は、成熟ウイルスコア粒子が生成されなかったことを示している。6日間感染後のJRFNFLuc上澄み液を非感染MT−2#18細胞に継代したところ、接種7日後までホタルルシフェラーゼ活性もp24産生も検出できなかった(データは示していない)。これら結果は、これら上澄み液が新たな複製の繰り返しを開始できるウイルスを含んでいなかったことを示唆している。
【0040】
最後に、非ヌクレオシド逆転写酵素インヒビターであるエファビレンツはJRFNFLucを0.18nMの50有効濃度(EC50)値で抑制したが、HIV−1プロテアーゼインヒビターであるアンプレナビルは複製コンピテントなJRFNRLucの1000倍以上に相当する濃度の10μMでも該ウイルスを抑制できなかった(表1)。このウイルスが後期HIV−1複製サイクルインヒビターによって抑制することができないとの知見は、このウイルスが複製できないことのさらなる証拠である。以上を総合すると、これらデータは、JRFNFLucウイルスが感染の単一の繰り返しを行うことができるのみであり、複製コンピテントではないのに対し、NL4RlucおよびJRFNRLucは感染の多数の繰り返しを行うことができ、真正に複製コンピテントなプロウイルスであることを強く示唆している。
【0041】
実施例2
SEAPリポーターウイルスの複製
JRFNRLuc中のレニラルシフェラーゼ遺伝子をSEAPをコードする遺伝子で置換することによりJRFNSEAPプロウイルス構築物を生成した。このプロウイルスクローンによって生成したウイルスがインビトロで複製する能力を試験した。感染6日後、JRFNSEAPに感染したMT−2#18細胞でのSEAP活性は非感染細胞で観察される活性の150倍以上であった。このウイルスに感染した細胞でのSEAP活性の動力学はp24活性と相関関係があった(図10)。このことは、検出されたSEAP活性がHIV−1のウイルス複製の結果であることを示唆している。JRFNSEAPウイルスによるp24産生のレベルが野生型pNL4−3の子孫ウイルスのレベルと実質的に同等であるとの知見は、SEAP遺伝子によるnef遺伝子の置換がHIV−1のウイルス複製に対して有害な作用をもたらさないことを証明している(図12)。
【0042】
ついで、細胞培養の感染後に生成するJRFNSEAPウイルスが細胞培養において新たな感染の繰り返しを開始できるか否かを決定するための試験を行った。図11に示すように、T細胞株を6日間感染させた後に生成されるJRFNSEAPウイルスはリポーター遺伝子を新たなMT−2#18細胞に形質導入することができた。これら結果は、JRFNSEAPウイルスが感染細胞培養から伝播して新たな細胞培養において多数の複製の繰り返しを開始できること、それゆえ複製コンピテントであることを証明している。
【0043】
上記HIV−1プロテアーゼインヒビターであるアンプレナビルおよび非ヌクレオシド逆転写酵素インヒビターであるエファビレンツを用い、JRFNSEAPウイルスの複製特性を調べた。表1に示すように、JRFNSEAPによるMT−2#18細胞の感染はエファビレンツ並びにアンプレナビルによって抑制された。アンプレナビルはJRFNSEAPの複製を21.5nMの50有効濃度(EC50)値で抑制し、エファビレンツはJRFNSEAPの複製を0.10nMのEC50で抑制した。HIV−1プロテアーゼインヒビターであるアンプレナビルは複製サイクルの後期にビリオンの成熟に際して作用し、単一サイクルの感染は抑制しない。それゆえ、この知見は、観察されたSEAPリポーター遺伝子活性が真正のウイルス複製の結果であることをさらに証明している。
【0044】
参照文献
1.チェン(Chen)ら、1994, J. Virol. 68: 654−660
2.ターウィリガー(Terwilliger)ら、1989, PNAS 86: 3857−3861
3.マリム(Malim)ら、1992, J. Exp. Med. 176: 1197−1201
4.ヘ(He)およびランダウ(Landau)、1995, J. Virol. 69: 4587−4592
5.プラネレ(Planelles)ら、1995, Gene Therapy 2: 369−376
6.チェン(Chen)ら、1996, J. Virol. 70: 6044−6053
7.ページ(Page)ら、1997, Aids Research and Human Retro. 13: 1077−1081
8.ジャミエソン(Jamieson)およびザック(Zack)、1998, J. Virol. 72: 6520−6526
9.ワイスロウ(Weislow)ら、1989, J. of Nat. Cancer Inst. 81: 577−586
【0045】
10.アダチ(Adachi)ら、1986, J. Virol. 59: 284−291
11.オブライエン(O’Brien)ら、1990, Nature 348: 69−73
12.マシューズ(Matthews)ら、1977, Biochemistry 16: 85
13.エディンガー(Edinger)ら、1998, J. Virol. 72: 7934−7940
14.ウエイン−ホブソン(Wain−Hobson)ら、1985, Cell 40: 9−17
15.リ(Li)ら、1991, J. Virol. 65: 3973−3985
16.コルマン(Collman)ら、1992, J. Virol. 66: 7517−7521
17.ローレンツ(Lorenz)ら、1991, P.N.A.S. USA 88: 4438
18.リウ(Liu)ら、1999, Gene 237: 153−159
【0046】
本明細書に引用した全ての特許、特許出願、公開文献、参照マニュアル、出典および要約の内容は、本発明が関連する技術分野の技術の状態を一層完全に記載するためにその全体が参照のため引用される。
上記に記載した組成物および方法には本発明の範囲から逸脱しない限り種々の変更を施すことが可能なので、上記記載に含まれ、添付の図面に示され、または請求の範囲で定められる事項は全て例示のものとして解釈されるのであって限定的な意味でないことが意図される。
【図面の簡単な説明】
【図1】NL4RLucプロウイルス構築物の模式図である。
【図2】JRFNRLucプロウイルス構築物の模式図である。
【図3】JRFNFLucプロウイルス構築物の模式図である。
【図4】JRFNSEAPプロウイルス構築物の模式図である。
【図5】HIV−1プロウイルスpNL4RLucによって生成したHIV−1ウイルスの複製の動力学を示す。第0日目、第2日目および第5日目に測定したレニラルシフェラーゼ活性および成熟ウイルスカプシドタンパク質p24レベルを示す。
【図6】HIV−1プロウイルスJRFNRlucによって生成したHIV−1ウイルスの複製の動力学を示す。第0日目、第3日目および第6日目に測定したレニラルシフェラーゼ活性および成熟ウイルスカプシドタンパク質p24レベルを示す。
【図7】MT−2#18細胞のpNL4Rluc感染後に生成したHIV−1ウイルスはMT−2#18細胞において新たな感染の繰り返しを開始することができたことを示す。
【図8】MT−2#18細胞のJRFNRluc感染後に生成したHIV−1ウイルスはMT−2#18細胞において新たな感染の繰り返しを開始することができたことを示す。
【図9】HIV−1プロウイルスJRFNFLucによって生成したHIV−1ウイルスの複製の動力学を示す。
【図10】HIV−1プロウイルスJRFNSEAPによって生成したHIV−1ウイルスの複製の動力学を示す。第0日目、第3日目および第6日目に測定したSEAP活性および成熟ウイルスカプシドタンパク質p24レベルを示す。
【図11】MT−2#18細胞のJRFNSEAP感染後に生成したHIV−1ウイルスはMT−2#18細胞において新たな感染の繰り返しを開始することができたことを示す。
【図12】種々のHIV−1リポーターウイルスおよびpNL4−3ウイルスのp24活性を示す。
Claims (23)
- 複製コンピテントなHIV−1ウイルスをコードするベクターであって、ウイルスの複製に必須でない領域がリポーター遺伝子によって置換されているHIV−1ゲノムを含むことを特徴とするベクター。
- 該リポーター遺伝子が、レニラルシフェラーゼリポーター遺伝子、SEAPリポーター遺伝子、CAT遺伝子、およびグリーン蛍光タンパク質遺伝子よりなる群から選ばれる、請求項1に記載のベクター。
- 該リポーター遺伝子がレニラルシフェラーゼリポーター遺伝子およびSEAPリポーター遺伝子よりなる群から選ばれる、請求項2に記載のベクター。
- ウイルスの複製に必須でない領域がnef遺伝子またはnef遺伝子のフラグメントをコードする、請求項1、2または3に記載のベクター。
- ウイルスの複製に必須でない領域がvpr遺伝子またはvpr遺伝子のフラグメントをコードする、請求項1、2または3に記載のベクター。
- 該HIV−1ゲノムがpNL4−3プロウイルスクローンのゲノムである、請求項1、2または3に記載のベクター。
- 該HIV−1ゲノムがpYU−2プロウイルスクローンのゲノムである、請求項1、2または3に記載のベクター。
- 該HIV−1ゲノムがp89.6プロウイルスクローンのゲノムである、請求項1、2または3に記載のベクター。
- 該HIV−1ゲノムがHIV−1 Laiプロウイルスクローンのゲノムである、請求項1、2または3に記載のベクター。
- 請求項1、2または3に記載のベクターを含む細胞。
- HIV−1に対する抗ウイルス活性を示す化合物のスクリーニング法であって、
(a)請求項1、2または3に記載のベクターで感染したまたは感染させるべき哺乳動物細胞に被験化合物を加え、ついで
(b)該被験化合物に暴露した細胞でのリポーター遺伝子の活性をコントロールの細胞での発現レベルと比較する
ことを含み、その際、リポーター遺伝子の発現レベルの低減が該被験化合物がHIV−1の複製を抑制したことを示すことを特徴とする方法。 - 該哺乳動物細胞がMT−2#18細胞である、請求項8に記載の方法。
- 複製コンピテントなHIV−1ウイルスをコードするベクターであって、ウイルスの複製に必須でない領域が機能的なレニラルシフェラーゼ酵素をコードする核酸配列によって置換されているHIV−1ゲノムを含むことを特徴とするベクター。
- 該レニラルシフェラーゼ遺伝子が、機能的なレニラルシフェラーゼ酵素という結果となるアラニンへのシステイン置換を含む、請求項13に記載のベクター。
- ウイルスの複製に必須でない領域がnef遺伝子またはnef遺伝子のフラグメントをコードする、請求項13に記載のベクター。
- ウイルスの複製に必須でない領域がvpr遺伝子またはvpr遺伝子のフラグメントをコードする、請求項13に記載のベクター。
- 該HIV−1ゲノムがpNL4−3プロウイルスクローンのゲノムである、請求項13に記載のベクター。
- 該HIV−1ゲノムがpYU−2プロウイルスクローンのゲノムである、請求項13に記載のベクター。
- 該HIV−1ゲノムがp89.6プロウイルスクローンのゲノムである、請求項13に記載のベクター。
- 該HIV−1ゲノムがHIV−1 Laiプロウイルスクローンのゲノムである、請求項13に記載のベクター。
- 請求項13に記載のベクターを含む細胞。
- HIV−1に対する抗ウイルス活性を示す化合物のスクリーニング法であって、
(a)請求項13に記載のベクターで感染したまたは感染させるべき哺乳動物細胞に被験化合物を加え、ついで
(b)該被験化合物に暴露した細胞でのリポーター遺伝子の活性をコントロールの細胞での発現レベルと比較する
ことを含み、その際、リポーター遺伝子の発現レベルの低減が該被験化合物がHIV−1の複製を抑制したことを示すことを特徴とする方法。 - 該哺乳動物細胞がMT−2#18細胞である、請求項13に記載の方法。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US21062600P | 2000-06-09 | 2000-06-09 | |
| PCT/US2001/018438 WO2001096610A1 (en) | 2000-06-09 | 2001-06-07 | Hiv-1 reporter viruses and their use in assaying anti-viral compounds |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2004503252A true JP2004503252A (ja) | 2004-02-05 |
Family
ID=22783626
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002510723A Pending JP2004503252A (ja) | 2000-06-09 | 2001-06-07 | Hiv−1リポーターウイルスおよび抗ウイルス性化合物をアッセイするうえでのその使用 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20030013078A1 (ja) |
| EP (1) | EP1294952A1 (ja) |
| JP (1) | JP2004503252A (ja) |
| AU (2) | AU7535701A (ja) |
| CA (1) | CA2412217A1 (ja) |
| WO (1) | WO2001096610A1 (ja) |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20110212434A1 (en) * | 2004-05-10 | 2011-09-01 | Jose Alcami Pertejo | Novel hiv-based recombinant viral clones and use thereof in analytical methods |
| ES2244332B1 (es) | 2004-05-10 | 2007-02-16 | Fundacion Para La Investigacion Y La Prevencion Del Sida En España | Nuevos clones virales recombinantes basados en vih y su utilizacion en metodos analiticos. |
| US8247167B2 (en) * | 2005-12-05 | 2012-08-21 | University Of Medicine And Dentistry Of New Jersey | HIV-1 latency model for high throughput screening |
| EP2571998B1 (en) * | 2010-05-19 | 2017-06-07 | Icosagen Cell Factory OÜ | Method and kit for identifying compounds capable of inhibiting human papilloma virus replication |
| US10106818B2 (en) | 2013-08-16 | 2018-10-23 | The J. David Gladstone Institutes | Dual-color HIV reporter system for the detection of latently-infected cells |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ATE118247T1 (de) * | 1988-09-27 | 1995-02-15 | Dana Farber Cancer Inst Inc | Vektor, der ein replikationskompetentes hiv-i- provirus und ein heterologes gen enthält. |
-
2001
- 2001-06-07 AU AU7535701A patent/AU7535701A/xx active Pending
- 2001-06-07 WO PCT/US2001/018438 patent/WO2001096610A1/en not_active Ceased
- 2001-06-07 CA CA002412217A patent/CA2412217A1/en not_active Abandoned
- 2001-06-07 US US09/876,680 patent/US20030013078A1/en not_active Abandoned
- 2001-06-07 EP EP01942062A patent/EP1294952A1/en not_active Withdrawn
- 2001-06-07 AU AU2001275357A patent/AU2001275357B2/en not_active Ceased
- 2001-06-07 JP JP2002510723A patent/JP2004503252A/ja active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| EP1294952A1 (en) | 2003-03-26 |
| CA2412217A1 (en) | 2001-12-20 |
| WO2001096610A1 (en) | 2001-12-20 |
| US20030013078A1 (en) | 2003-01-16 |
| AU7535701A (en) | 2001-12-24 |
| AU2001275357B2 (en) | 2006-06-08 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Gelderblom et al. | Viral complementation allows HIV-1 replication without integration | |
| Chen et al. | Distinct modes of human immunodeficiency virus type 1 proviral latency revealed by superinfection of nonproductively infected cell lines with recombinant luciferase-encoding viruses | |
| US5837464A (en) | Compositions and methods for determining anti-viral drug susceptibility and resistance and anti-viral drug screening | |
| Freed et al. | Role of the basic domain of human immunodeficiency virus type 1 matrix in macrophage infection | |
| Zhou et al. | Small-molecule inhibition of human immunodeficiency virus type 1 replication by specific targeting of the final step of virion maturation | |
| CA2216126C (en) | Compositions and methods for determining anti-viral drug susceptibility and resistance and anti-viral drug screening | |
| US6942969B2 (en) | Compositions and methods for determining anti-viral drug susceptibility and resistance and anti-viral drug screening | |
| US9631220B2 (en) | Method for designing a drug regime for HIV-infected patients | |
| Kristbjörnsdóttir et al. | The vif gene of maedi-visna virus is essential for infectivity in vivo and in vitro | |
| Klimkait et al. | Dissecting the mode of action of various HIV-inhibitor classes in a stable cellular system | |
| JP2004503252A (ja) | Hiv−1リポーターウイルスおよび抗ウイルス性化合物をアッセイするうえでのその使用 | |
| AU2001275357A1 (en) | HIV-1 reporter viruses and their use in assaying anti-viral compounds | |
| AU4827899A (en) | Cell-based assay for immunodeficiency virus infectivity and sensitivity | |
| US6797462B1 (en) | Cell-based assay for immunodeficiency virus infectivity and sensitivity | |
| CN106800603B (zh) | 检测抗hiv抗体的adcc活性的方法 | |
| EP1963495B1 (en) | An hiv-1 latency model for high throughput screening | |
| US7807346B2 (en) | High-throughput assay for virus entry and drug screening | |
| Ma et al. | Establishment of an indicator cell line to quantify bovine foamy virus infection | |
| Deminie et al. | [12] Quantitation of virus stocks produced from cloned human immunodeficiency virus DNA | |
| Münk et al. | Production and Use of HIV‐1 Luciferase Reporter Viruses | |
| JP2002345457A (ja) | T細胞株およびその用途 | |
| Agosto | HIV Can Establish Latency by Direct Infection of Resting CD4+ T Cells | |
| Omar et al. | P39. 07 Real Time Fitness Assay of Two CRF01_A/E HIV-1 Transmitted Founder Variants | |
| MXPA98006104A (en) | Compositions and methods to determine the susceptibility and resistance to antiviral drug and the antivi drug examination | |
| JP2002191399A (ja) | Hiv薬剤耐性試験方法 |