JP2012530735A - 眼疾患の治療におけるCD44v6の使用 - Google Patents
眼疾患の治療におけるCD44v6の使用 Download PDFInfo
- Publication number
- JP2012530735A JP2012530735A JP2012516550A JP2012516550A JP2012530735A JP 2012530735 A JP2012530735 A JP 2012530735A JP 2012516550 A JP2012516550 A JP 2012516550A JP 2012516550 A JP2012516550 A JP 2012516550A JP 2012530735 A JP2012530735 A JP 2012530735A
- Authority
- JP
- Japan
- Prior art keywords
- peptide
- cd44v6
- seq
- vegfr
- vegf
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images








Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/04—Peptides having up to 20 amino acids in a fully defined sequence; Derivatives thereof
- A61K38/08—Peptides having 5 to 11 amino acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- A61K38/17—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- A61K38/177—Receptors; Cell surface antigens; Cell surface determinants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P27/00—Drugs for disorders of the senses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P27/00—Drugs for disorders of the senses
- A61P27/02—Ophthalmic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/10—Drugs for disorders of the cardiovascular system for treating ischaemic or atherosclerotic diseases, e.g. antianginal drugs, coronary vasodilators, drugs for myocardial infarction, retinopathy, cerebrovascula insufficiency, renal arteriosclerosis
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Veterinary Medicine (AREA)
- Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- General Health & Medical Sciences (AREA)
- Public Health (AREA)
- Immunology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Epidemiology (AREA)
- Gastroenterology & Hepatology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Organic Chemistry (AREA)
- Cell Biology (AREA)
- Zoology (AREA)
- Vascular Medicine (AREA)
- Cardiology (AREA)
- Heart & Thoracic Surgery (AREA)
- Urology & Nephrology (AREA)
- Ophthalmology & Optometry (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Peptides Or Proteins (AREA)
Abstract
【選択図】なし
Description
(a)配列番号2又は配列番号1で示されるようなアミノ酸配列を含むか、又はそれらで構成されるペプチド、
(b)配列番号2又は配列番号1のフラグメントで構成され、かつCD44と、VEGFR−2との間の複合体形成を阻害する活性を有するペプチド、
(c)異種アミノ酸配列に融合された(a)若しくは(b)によるペプチドを含むか、又はそれらで構成される異種融合ペプチド、及び
(d)CD44と、VEGFR−2との間の複合体形成を阻害する活性を有する(a)、(b)又は(c)によるペプチドの誘導体
からなる群から選択される。
(a)配列番号2若しくは配列番号1で示されるようなアミノ酸配列を含むか、又はそれらで構成されるペプチド、
(b)下記アミノ酸配列:
配列番号2又は配列番号1のアミノ酸2〜14、
配列番号2又は配列番号1のアミノ酸2〜13、
配列番号2又は配列番号1のアミノ酸2〜12、
配列番号2又は配列番号1のアミノ酸2〜11、
配列番号2又は配列番号1のアミノ酸3〜14、
配列番号2又は配列番号1のアミノ酸3〜13、
配列番号2又は配列番号1のアミノ酸3〜12、
配列番号2又は配列番号1のアミノ酸3〜11、
配列番号2又は配列番号1のアミノ酸4〜14、
配列番号2又は配列番号1のアミノ酸4〜13、
配列番号2又は配列番号1のアミノ酸4〜12、
配列番号2又は配列番号1のアミノ酸4〜11、
配列番号2又は配列番号1のアミノ酸5〜14、
配列番号2又は配列番号1のアミノ酸5〜13、
配列番号2又は配列番号1のアミノ酸5〜12、
配列番号2又は配列番号1のアミノ酸5〜11、
配列番号2又は配列番号1のアミノ酸6〜14、
配列番号2又は配列番号1のアミノ酸6〜13、
配列番号2又は配列番号1のアミノ酸6〜12、
配列番号2又は配列番号1のアミノ酸6〜11、
配列番号2又は配列番号1のアミノ酸7〜14、
配列番号2又は配列番号1のアミノ酸7〜13、
配列番号2又は配列番号1のアミノ酸7〜12、
配列番号2又は配列番号1のアミノ酸7〜11
のいずれか1つを含むか、又はそれらで構成されるペプチド、及び
(c)異種アミノ酸配列に融合された(a)又は(b)を含む異種融合ペプチド
からなる群から選択される。
実験手順:
ヒト臍帯静脈内皮細胞(HUVEC、Provitri GmbH、ドイツ、ベルリン)は、SupplementMix(Provitro GmbH、ドイツ、ベルリン)を添加した内皮細胞成長培地中で成長させた。ヒト大動脈内皮細胞(HAOEC、Promocell、ドイツ、ハイデルベルク)及びヒト心臓微小血管内皮細胞(HCMEC、Promocell、ドイツ、ハイデルベルク)は、製造業者の取扱説明書に従ってサプリメントを備えた内皮細胞成長培地MV(Prmocell、ハイデルベルク)中で成長させた。EC細胞は全て、9回を超えて継代培養させなかった(not passaged)。HEK293細胞(American Tissue Culture Collection、ATCC、ドイツ、ウェーゼル)及びエストロゲン非依存的ヒト乳がん細胞株MDA−MB231(American Type Culture Collection、メリーランド州ロックビル)は、10%FCS(PAA Laboratories、ドイツ、コエルバ(Coelbe))を添加したDMEM(Invitrogen、ドイツ、カールスルーエ)中で成長させた。ラット膵癌細胞株BSp73AS及びそのトランスフェクタント(Bsp73ASs6)は、RMPI(Invitrogen、ドイツ、カールスルーエ)+10%FCS中で成長させた。ドイツのミュンヘン大学のC. Bruns氏によりご厚意で提供されたヒト膵がん細胞株L3.6plは、10%FCS、ピルビン酸ナトリウム、非必須アミノ酸、L−グルタミン及び2倍ビタミン溶液(Invitrogen、ドイツ、カールスルーエ)を添加したDMEM中に維持した。
CD44v6に対するヒトモノクローナル抗体VFF18(14mer:KEQWFGNRWHEGYR、配列番号2)は、Bender(オーストリア、ウィーン)から入手した。汎CD44抗体IM7は、BD Biosciences(カリフォルニア州サンディエゴ)製のものであり、抗Erk 1(K−23)抗体は、Santa Cruz(ドイツ、ハイデルベルグ)製であった。リンVEGFR−2(Tyr1175)及びリンErk(リンp44/42)は、Cell Signaling Technology(イングランド、ベバリー)から購入した。VEGFR−2に対する抗体は、R&D Systems(ドイツ、ウィースバーデン)から又はSanta Cruz(ハイデルベルク)(クローンA−3)から入手した。HRPで標識した二次抗体は、Dako(ドイツ、ハンブルク)から購入した。VEGF−A165及びVEGF−A121は、ピチア・パストリス(Pichia pastoris)で産生された。HGFは、George Vande Woude(Van Andel Institute、米国、グランド・ラピッズ)のご厚意による贈呈であった。v6ヒトペプチド(14mer)及び対照ペプチドは、Matzke他、「5−アミノ酸ペプチドはMet及びRon依存的細胞遊走を阻止する(A 5-amino-acid peptide blocks Met and Ron dependent cell migration)」(Cancer research 65: 6105-6110)に記載されている。v6マウスペプチドの配列は、QETWFQNGWQGKNP(配列番号3)であった。
VEGFR−2発現プラスミドpBE hVEGFR−2は、フォワードプライマー:
GCTCTTCGGGGAGCAGCGATGGAGAGCAAGGTGCTGCTG(配列番号4)及びリバースプライマー:
GGAGGTTTTTTAAAGCAAGTAAAACCTTTATCACAGATCCTCTTCTGAGAGAG(配列番号5)を用いてPCRサブクローニング法によりヒトVEGFR−2をコードする配列を導入して、GFPリーディングフレームを除去することによって、Clontech(カリフォルニア州マウンテンビュー)製のpEGFP−C1ベクターから得た。
エクソン特異的RT−PCRは、Konig他、1996年、「トランス作動性因子はCD44スプライス変異体の発現を調節する(Trans-acting factors regulate the expression of CD44 splice variants)」(EMBO J. 15:4030-4039)に記載されるように、同じヒトプライマーを使用して実施した。
HEK293細胞を、製造業者のプロトコルに従って、6ウェルプレート中でリポフェクトアミン 2000(Invitrogen、ドイツ、カールスルーエ)で一過的にトランスフェクトした。BSp73AS細胞及びBSp73ASs6細胞のトランスフェクションを電気穿孔により実施した。簡潔に述べると、3×106個の細胞を、氷上でベクターDNA 5μgと混合した。Gene Pulser(Bio-Rad、ドイツ、ミュンヘン)を使用して、250μF、0.28kVで4mm電気穿孔キュベット中で、電気穿孔を実施した。血清を含有する予め温めた培地を添加して、細胞を6ウェルプレート中に分配した。次に、細胞を24時間成長させ、血清を24時間飢餓状態にさせて、実験に使用した。
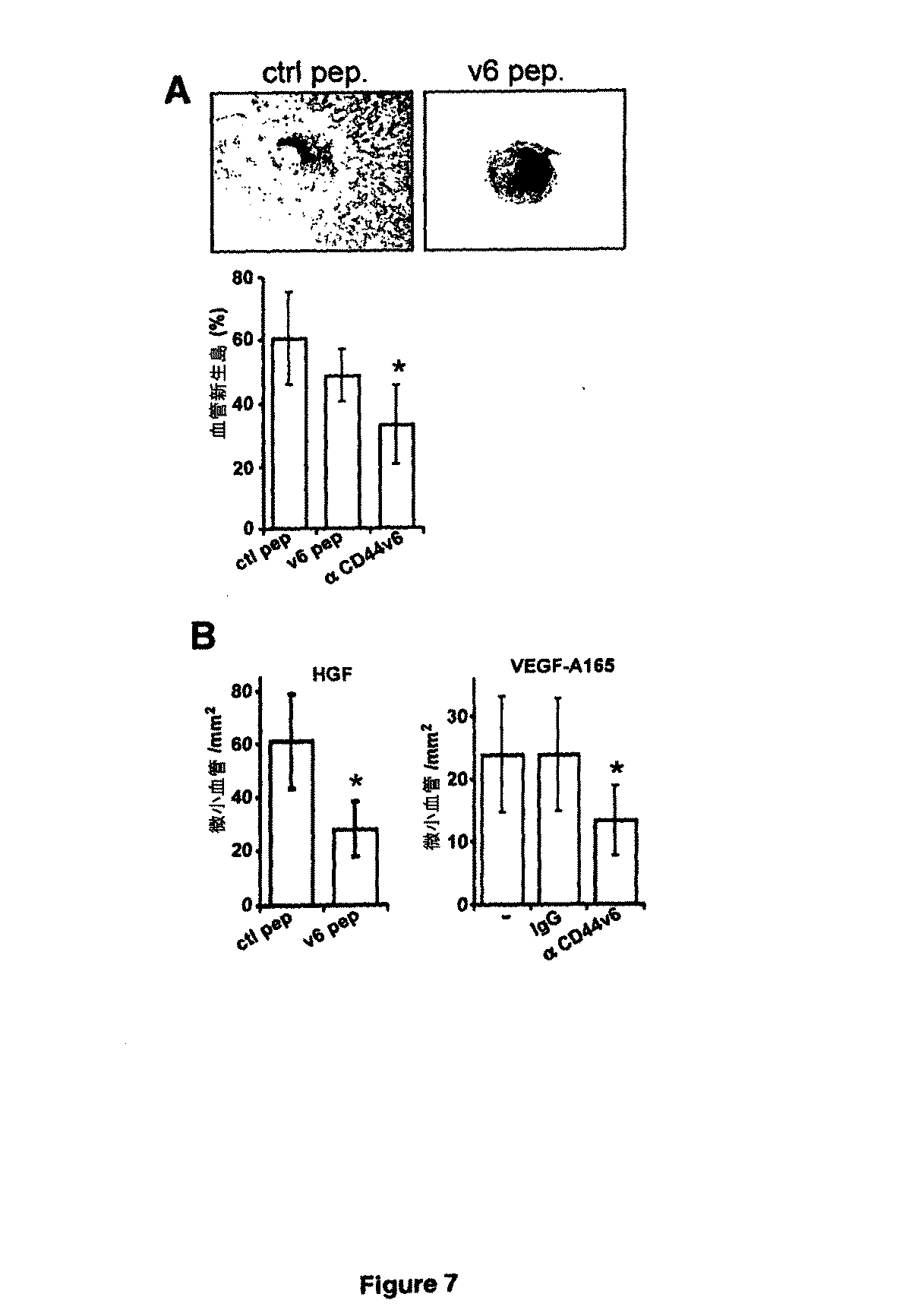
MDA−MB231細胞(動物1匹当たり1×106個)を亜集密的に収集して、PBSで洗浄して、PBS 50μl中で再懸濁した。皮膚切開を介して、インスリンシリンジを用いて、麻酔をかけた雌SCIDマウス(株C.B−17/Ztm−scid)の第4乳腺複合体の乳腺脂肪体へ非常にゆっくりと細胞を移植した。皮膚用の断続バイクリル縫合(5/0、Ethicon、ドイツ、ノルダーシュテット)を使用して、切開を閉塞した。腫瘍を1週間成長させ、続いて動物をグループに分割して(1グループ当たり6匹〜7匹の動物)、抗マウスCD44v6(クローン 9A4)、対照IgG(NatuTec、ドイツ、フランクフルト)、マウスv6ペプチド又は対照ペプチド 20μgで、1週間に3回、非経口処理した。腫瘍成長は、ノギスを使用して、又はfpVCT画像化により1週間に1回、モニタリングした(以下を参照)。腫瘍細胞移植の5週後に、動物を屠殺して、腫瘍を切除して、測定して、半分に分割して、免疫組織化学的分析用に4%ホルマリン又は亜鉛固定剤(0.1M Tris(pH7.4)中に0.5g/lの酢酸カルシウム、5g/lの酢酸亜鉛、5g/lの塩化亜鉛)中で24時間固定させた。L3.6plヒト膵がん細胞を同所に注入した。簡潔に述べると、小さな左腹部脇腹切開を行い、脾臓を露出させた。5×105個の細胞/40μl(ハンクス緩衝塩溶液、Invitrogen、ドイツ、カールスルーエ)を、30ゲージ針を使用して脾臓の真下の膵臓の被膜領域へ注入した。注入の傍らに綿棒を1分間保持して、漏出を防いだ。出現してくる流動的な泡は、被膜膵臓内注入の成功の徴候とみなした。腫瘍細胞の移植の7日後、マウスをそれぞれ5匹のマウスのグループへと無作為に割り当てた:(1)PBS、(2)対照ペプチド(20μg/ml)、(3)CD44v6ペプチド(20μg/ml)を1週に3回注入。処理の開始の21日後に、動物を屠殺した。腫瘍体積は、長さ×高さ×幅÷2として算出した。組織は上述のように加工処理した。動物は全て、動物実験に関するドイツの規制に従って収容し、実験は全て、欧州及びドイツの法的規制に従って実施した。マウスは全て、Harlan(ドイツ)から入手した。
マウスは、ノンクリニカルボリュームコンピュータ断層撮影法プロトタイプフラットパネル検出器(GE Global Research、米国ニューヨーク州ニスカユナ)により画像化した。簡潔に述べると、マウスを0.8%〜1%の気化イソフルランで麻酔して、系のz軸に対して垂直に配置した。スキャニングの30秒前に、ヨウ素含有造影剤Isovist 300(マウス1匹当たり150μl、Bayer-Schering、ドイツ、ベルリン)を静脈内に適用した。小さな血管のより良好な表示のために、Isovist 300を血液プール剤eXia 160(Binitio Biomedical Inc., カナダ、オタワ)で置き換え、それを解剖の当日にスキャンの90秒前に使用した。データ組は全て、同じプロトコルで取得した。1回転につき500回の表示(view)、回転時間4秒、360の使用検出器列、管電圧80kVp及び電流100mA。修正Feldkampアルゴリズムが画像再構成に使用され、等方性高解像度ボリュームデータ組(512×512マトリクス、およそ100£gmの等方性ボクセルサイズで)。腫瘍区分け及び体積推定に関して、voxtools 3.0.64 Advantage Workstation 4.2(GE Healthcare、英国)を用いて、データ組を解析した。
血清飢餓細胞(24時間)を、成長因子HGF(20ng/ml)を用いて37℃にて5分間、又はVEGF−A165又はVEGF−A121(40ng/ml)を用いて37℃にて8分間誘導した。示されている場合は、37℃で10分間の誘導(100μg/mlの抗CD44v6、100ng/mlのv6ペプチド又は100ng/mlの対照ペプチド、0.5μg/mlのCD44v6ECD)の前に、細胞をブロッキング試薬で処理した。細胞を氷冷PBSで洗浄した。活性化Erkを検出するために、100mM ジチオスレイトール(DTT)を含有する沸騰SDSサンプル緩衝液中に細胞を溶解させて、リン酸化Erkに対する抗体を使用して、ウェスタンブロット分析に付した。Erk負荷対照は、剥脱した同じブロット上で実施して(62.5mM Tris(pH6.8)、2%SDS、0.8%DTT)、Erk抗体でプローブした。活性化VEGFR−2を検出するために、細胞を還元性サンプル緩衝液中に溶解させて、SDS−PAGEゲルのブロットを、リン酸化VEGFR−2に対する抗体でプローブした。代替的には、溶解液(20mM Tris(pH 7.4)、1mM EDTA、1mM EGTA、1mM DTT、25mM NaCl、1.5% Triton X−100、10mM NaF、1mM PMSF、1mM オルトバナジン酸ナトリウム、1mM アポロチニン及びロイペプチン)を調製した。遠心分離後、清澄な溶解液(12000rpmで15分間)をVEGFR−2抗体(クローン A−3、Santa Cruz、ドイツ、ハイデルベルク)又はマウスIgG対照(Santa Cruz、ドイツ、ハイデルベルク)とともに4℃で一晩インキュベートした後、プロテインA/Gアガロースビーズ(Merck、ドイツ、ダルムシュタット)とともに4℃で2時間、インキュベートした。ビーズを3回洗浄して、サンプル緩衝液中で沸騰させて、リン特異的VEGFR−2抗体を使用してウェスタンブロット分析に付した。負荷対照を得るために、ブロットを剥脱して、VEGFR−2抗体で再プローブした。ブロットは、増強した化学発光系(ECL、Thermo FIshcer Scientific Inc.,マサチューセッツ州ウォルサム)を使用して染色した。ウェスタンブロット分析におけるバンドは、プログラムImage Jで定量化した。
共免疫沈降に関して、HUVEC(10cmプレート中に1.5×106個)は、上述するように各々のリガンドによって誘導させた。細胞を溶解緩衝液(25mM ヘペス(pH7.5)、100mM NaCl、10mM MgCl2、1mM EDTA、10%グリセロール、1% Igepal、10mM NaF、1mM PMSF、1mM オルトバナジン酸ナトリウム、1mMアポロチニン及びロイペプチン)中で氷上にて30分間インキュベートして、続いて12000rpmで20分間遠心分離した。免疫沈降に関して、清澄な溶解液を汎CD44に対する抗体(IM7)とともに4℃で一晩インキュベートした後、プロテインA/Gアガロースビーズ(Merck、ドイツ、ダルムシュタット)を用いて沈降させた。沈降物を溶解緩衝液中で3回洗浄して、ウェスタンブロット分析に付した。
HUVEC及びHAOECをウェル1つ当たり2.5×105個の細胞の濃度で12ウェルプレート中に播種させた。24時間後、滅菌ピペットチップを使用して集密的な細胞層にスクラッチを作製した。培地を変えて、新鮮な培地又は100μg/mlの抗CD44v6、100ng/mlのv6ペプチド若しくは100ng/ml対照ペプチドを含有する培地と交換した。37℃で10分後に、成長因子(HGF(20ng/ml)、VEGF−A165又はVEGF−A121(40ng/ml))による誘導を実施した。Canon Power Shot S620デジタルカメラを使用して、誘導の24時間後に細胞の写真を撮った。コンピュータプログラムImage Jを定量的評価に使用した。スクラッチ中の細胞を含まない領域を測定した。スクラッチの閉塞の効率は、細胞を含まない領域のパーセントとして表わされる。
HUVECのスフェロイドを懸滴中に生成させた。0.25%(wt/vol)メチルセルロースを含有する内皮細胞成長培地(Sigma、ドイツ、ハンブルク)中に懸濁させて、一晩インキュベートした(スフェロイド1個当たり:25μl中に750個の細胞)。穏やか遠心分離(5分、800rpm)によりスフェロイドを収集して、1mg/mlのラット尾コラーゲンI(BD Biosciences、マサチューセッツ州ベッドフォード)及び0.6%(wt/vol)メチルセルロースを含有する内皮細胞成長培地中で再懸濁させた。スフェロイド/マトリックス混合物を48ウェルプレート中に分配した(1ウェルあたり30個のスフェロイド)。37℃でのコラーゲンの凝固後に、スフェロイド/マトリックス混合物を内皮細胞成長培地で覆った。ブロッキング試薬(100μg/mlの抗CD44v6、100ng/mlのv6ペプチド又は100ng/mlの対照ペプチド)を添加して、スフェロイドをVEGF(40ng/ml)によって誘導した。48時間後に写真を撮った。
1:1の比で内皮細胞成長培地と混合した成長因子低減マトリゲル(BD Biosciences、マサチューセッツ州ベッドフォード)で、48ウェルプレートをコーティングした。HUVECは、ウェル1つ当たり2.5×104個の細胞の濃度で、これらのマトリゲルでコーティングしたプレート上に播種した。ブロッキング試薬(100μg/mlの抗CD44v6、100ng/mlのv6ペプチド又は100ng/mlの対照ペプチド)を37℃で10分間添加した後、成長因子で誘導した。24時間後に写真を撮った。管形成の定量化は、コンピュータプログラムImage Jを使用して、1視野当たりの分岐点又は総血管長を計数することにより実施した。
8週〜9週齢のRip1Tag2マウスから得られた血管新生過形成性ランゲルハンス島は、コラゲナーゼ灌流により単離した。HUVECはブロッキング試薬と混合して、24ウェルプレートにおいて三次元コラーゲンマトリックス中でこれらの島と共培養した(ウェル1つ当たり:コラーゲンマトリックス 350μl中に4×104個の細胞、5個の血管新生島)。3日に1回、新たなブロッキング試薬を添加した。5日〜7日後、血管新生島に対するECの応答を求めた。1つの条件当たりおよそ60個の島を分析した。
このアッセイは、過去に記載されるように実施した(Alajati他、2008年、「マウスにおけるヒト脈管構造のスフェロイドベースの操作(Speroid-based engineering of a human vasculature in mice」(Nat Methods 5:439-445))。HUVECのスフェロイドを、100個の細胞を含有する懸滴(0.25%(wt/vol)メチルセルロースを含有する成長培地(Sigma、ドイツ、ハンブルク)25ml)中で生成させて、一晩インキュベートした。スフェロイドは、穏やか遠心分離(5分、800rpm)により収集して、内皮細胞成長培地で洗浄して、マトリゲル(成長因子は低減されている;BD Sciences、ドイツ、ハイデルベルク)600μl及び500ng/mlの濃度で成長因子HGF又はVEGF−A165を含有するフィブリノーゲン(最終濃度 2ml/ml、Calbiochem、英国、ビーストン)と混合した。CD44v6抗体(VFF18)又はペプチド20μgを混合物へ添加した後、トロンビン(0.4U、Carbiochem、英国、ビーストン)を添加した。混合物を、腹部正中部に対して側面の各側面上に4週〜6週齢のSCIDマウスへ皮下注入した。2日に1回、ブロッキング試薬(マウス1匹当たり20μg)を、マトリゲル/フィブリンプラグの近くに注入した。移植の21日後にマウスを屠殺して、プラグを回収して、免疫組織学的分析用に4%ホルムアルデヒド中で一晩固定した。
固定後、マトリゲル/フィブリンプラグ又は腫瘍組織を加工処理して、パラフィン中に包埋した。パラフィンブロックの7μm切片を脱パラフィン処理して、再度水和させた。マトリゲル/フィブリンプラグ切片を、10%ヤギ血清(Dako、ドイツ、ハンブルク)で60分間ブロッキングした後、マウス抗ヒトCD34抗体(クローンQBEND/10、20μg/ml、2時間;Novocastra、英国、ニューカッスルアポンタイン)で染色した。次に、切片を、ヤギ抗マウスAlexa Fluor 488(Invitrogen、ドイツ、カールスルーエ)とともに45分間インキュベートした。核をHoechst色素 33258(Sigma Aldrich、ドイツ、ハンブルク)で染色した。OlympusIX50倒立顕微鏡を使用して、完全マトリゲル/フィブリン領域の画像を撮影した。完全マトリックス領域中の蛍光構造を計数して、1mm2当たりの血管数として算出した。腫瘍切片において、内因性ペルオキシダーゼを、PBS中の3%H2O2でブロックした後、アビジン/ビオチン(Dako、ドイツ、ハンブルク)とともにインキュベートした。ラット抗マウスCD31抗体とのインキュベーション(0.5μg/ml 4℃で一晩)の前に、非特異的結合を10%ウサギ血清(BD Biosciences、ドイツ、ハイデルベルク)で60分間ブロックした。続いて、切片をビオチン化ウサギ抗ラット抗体(2μ/ml、45分)とともにインキュベートした後、ストレプトアビジンペルオキシダーゼ結合体(conjugate)(Dako、ドイツ、ハンブルク)処理を行い、DAB基質系(3,3’−ジアミノベンジジン、Biozol、ドイツ、エヒング)で発色させた。コンピュータプログラムImage Jを用いて、染色した構造全てを、1mm2当たりの血管数及び平均血管サイズに関して解析した。
定量化は全て、平均値±標準偏差(s.d.)として与えられる。各種条件間の差は、対応のあるスチューデントのt検定(ttest)により解析されて、p<0.05は、統計学的に有意であるとみなした。
実施例1:CD44v6は、VEGFR−2の活性化及び下流のシグナル伝達を制御する。
実施例2:CD44v6ペプチド及び抗体は、VEGFに対するECの応答を阻止する。
実施例3:Rip1Taq2マウスから単離した過形成性ランゲルハンス島により誘導される血管新生応答は、CD44v6に依存的である。
実施例4:移植したECスフェロイドから血管のin vivoでの発達は、CD44v6ブロッキング試薬による処理時に損なわれる。
実施例5:乳腺腫瘍及び膵臓腫瘍の血管新生化はCD44v6を必要とする。
表1:CD44v6ペプチド及び抗体による腫瘍誘導性の血管の阻害
SCIDマウスへのMDA−MB231細胞の注入、及び続く処理は、図7Aに記載するように実施した。腫瘍切片をCD31特異的抗体で染色して、血管数及びサイズを計数した。数は、5匹の動物それぞれのカウントを反映する。
配列番号5:プライマー
Claims (10)
- 個体における眼疾患の予防及び/又は治療における使用のための配列番号2又は配列番号1のアミノ酸7〜11により表示されるアミノ酸配列を含むペプチド化合物、又はその機能的に活性な誘導体、又はその薬学的に許容される塩。
- 前記ペプチド化合物が配列番号2を含む、請求項1に記載のペプチド化合物。
- 前記ペプチド化合物が配列番号1を含む、請求項1に記載のペプチド化合物。
- 前記眼疾患が、VEGFR−2の過剰発現に関連する、請求項1〜3のいずれか一項に記載のペプチド化合物。
- 前記眼疾患が、内皮細胞の過剰増殖に関連する、請求項1〜4のいずれか一項に記載のペプチド化合物。
- 前記眼疾患が、黄斑変性症又は糖尿病性網膜症である、請求項1〜5のいずれか一項に記載のペプチド化合物。
- 前記個体が哺乳動物である、請求項1〜6のいずれか一項に記載のペプチド化合物。
- 前記ペプチド化合物が、配列番号2のアミノ酸7〜11により表示されるアミノ酸配列を含み、前記個体がヒトである、請求項1、請求項2及び請求項4〜7のいずれか一項に記載のペプチド化合物。
- 前記ペプチド化合物が、配列番号1のアミノ酸7〜11により表示されるアミノ酸配列を含み、前記個体がラットである、請求項1及び請求項3〜7のいずれか一項に記載のペプチド化合物。
- 前記ペプチド化合物が、環状ペプチド又はその薬学的に許容される塩である、請求項1〜9のいずれか一項に記載のペプチド化合物。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP09008272.8 | 2009-06-24 | ||
| EP09008272A EP2266593A1 (en) | 2009-06-24 | 2009-06-24 | Use of CD44v6 in the treatment of ophthalmic diseases |
| PCT/EP2010/003523 WO2010149281A1 (en) | 2009-06-24 | 2010-06-11 | Use of cd44v6 in the treatment of ophthalmic diseases |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2012530735A true JP2012530735A (ja) | 2012-12-06 |
Family
ID=41338473
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2012516550A Pending JP2012530735A (ja) | 2009-06-24 | 2010-06-11 | 眼疾患の治療におけるCD44v6の使用 |
Country Status (4)
| Country | Link |
|---|---|
| US (2) | US20120115794A1 (ja) |
| EP (1) | EP2266593A1 (ja) |
| JP (1) | JP2012530735A (ja) |
| WO (1) | WO2010149281A1 (ja) |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20140045764A1 (en) * | 2011-03-24 | 2014-02-13 | Neurim Pharmaceuticals (1991) Ltd. | Neuroprotective peptides |
| GB201220889D0 (en) | 2012-11-21 | 2013-01-02 | Kit Karlsrusher Inst Fuer Technologie Und Inst Fuer Toxikologie Und Genetik And Amcure Gmbh | CD44v6-derived peptides for treating metastasizing cancers |
| GB201220901D0 (en) * | 2012-11-21 | 2013-01-02 | Kit Karlsrusher Inst Fuer Technologie Und Inst Fuer Toxikologie Und Genetik And Amcure Gmbh | CD44v6-derived peptides for treating pancreatic cancer |
| GB201220899D0 (en) * | 2012-11-21 | 2013-01-02 | Kit Karlsrusher Inst Fuer Technologie Und Inst Fuer Toxikologie Und Genetik And Amcure Gmbh | CD44v6-derived pegylated peptides |
| GB201220891D0 (en) * | 2012-11-21 | 2013-01-02 | Kit Karlsrusher Inst Fuer Technologie Und Inst Fuer Toxikologie Und Genetik And Amcure Gmbh | CD44v6-derived peptides for treating breast cancers |
| GB201421647D0 (en) | 2014-12-05 | 2015-01-21 | Amcure Gmbh And Ruprecht-Karls-Universitat And Karlsruher Institut F�R Technologie | CD44v6-derived cyclic peptides for treating cancers and angiogenesis related diseases |
| US10604586B2 (en) | 2016-05-09 | 2020-03-31 | Industrial Technology Research Institute | Humanized monoclonal antibody and uses thereof |
| JP2023516080A (ja) * | 2020-03-06 | 2023-04-17 | ジーオー セラピューティクス,インコーポレイテッド | 抗グリコcd44抗体およびその使用 |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP1647556A1 (en) * | 2004-10-14 | 2006-04-19 | TopoTarget Germany AG | Peptidic compounds and derivatives thereof for the treatment of human diseases through inhibition of signaling via growth factors |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| UA58482C2 (uk) * | 1994-06-08 | 2003-08-15 | Бьорінгер Інгельхайм Інтернаціональ Гмбх | Моноклональне антитіло vff-18 проти сd44v6 і його фрагменти |
| WO2009023411A1 (en) * | 2007-08-09 | 2009-02-19 | Bausch & Lomb Incorporated | Compositions and methods for treating or controlling anterior- and posterior-segment ophthalmic diseases |
-
2009
- 2009-06-24 EP EP09008272A patent/EP2266593A1/en not_active Withdrawn
-
2010
- 2010-06-11 US US13/378,574 patent/US20120115794A1/en not_active Abandoned
- 2010-06-11 JP JP2012516550A patent/JP2012530735A/ja active Pending
- 2010-06-11 WO PCT/EP2010/003523 patent/WO2010149281A1/en not_active Ceased
-
2012
- 2012-12-13 US US13/713,594 patent/US8883738B2/en not_active Expired - Fee Related
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP1647556A1 (en) * | 2004-10-14 | 2006-04-19 | TopoTarget Germany AG | Peptidic compounds and derivatives thereof for the treatment of human diseases through inhibition of signaling via growth factors |
Also Published As
| Publication number | Publication date |
|---|---|
| US20120115794A1 (en) | 2012-05-10 |
| EP2266593A1 (en) | 2010-12-29 |
| US20130109632A1 (en) | 2013-05-02 |
| US8883738B2 (en) | 2014-11-11 |
| WO2010149281A1 (en) | 2010-12-29 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Ferrara | Vascular endothelial growth factor: basic science and clinical progress | |
| US8883738B2 (en) | Use of a peptide fragment of CD44v6 in the treatment of ophthalmic diseases | |
| ES2525669T3 (es) | Inhibidores de la vascularización | |
| PT1140969E (pt) | Fragmentos do factor de crescimento de tecido conectivo e os seus métodos e utilizações | |
| JP2006527198A (ja) | Vegfインヒビターを用いる腫瘍退縮方法 | |
| CA2592292A1 (en) | Sustained delivery of pdgf using self-assembling peptide nanofibers | |
| WO2017174758A1 (en) | Combination of an apelin antagonist and an angiogenesis inhibitor for the treatment of cancer | |
| US8420780B2 (en) | Mutated netrin 4, fragments thereof and uses thereof as drugs | |
| CN103228295B (zh) | 一种限制急性心肌缺血后微血管损伤的方法 | |
| JP5400006B2 (ja) | 虚血性疾患の予防または治療剤 | |
| WO2005087954A2 (en) | Roles of foxc2, pdgfr, vegfr3 in lymphatic vessel development | |
| KR102384795B1 (ko) | 전이성 암의 치료 및 예방을 위한 제약 조성물 및 방법 | |
| US9718878B2 (en) | Kit for detecting Cthrc1 in a sample | |
| JP2024150757A (ja) | ペプチドおよびその医学的使用 | |
| US8506965B2 (en) | R-RAS activity in vascular regulation | |
| JP7232537B2 (ja) | c-METアゴニスト抗体及びその使用 | |
| JP2003529370A (ja) | 血管浸透性に対する有害な作用を有さないve−カドヘリンに対する拮抗薬抗体 | |
| JPWO2002100441A1 (ja) | 血管再生療法 | |
| WO2015110802A1 (en) | Abl1 inhibitor for treating and preventing ocular neovascularisation | |
| Brkić | Molecular Regulation of Intussusceptive Angiogenesis by EphrinB2/EphB4 Signaling and Its Therapeutic Potential | |
| KR20030081402A (ko) | Vegf-x 또는 그의 길항제에 의한 평활근 세포 증식의조절 | |
| JP2016501856A (ja) | 乳癌を処置するためのcd44v6由来ペプチド | |
| CN1681525A (zh) | 一种抑制血管渗漏和组织水肿的方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20130121 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20140304 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20140521 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20141007 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20141209 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20150203 |