JP2017525355A - バクテリオファージ・ラムダ・インテグラーゼの変異体 - Google Patents
バクテリオファージ・ラムダ・インテグラーゼの変異体 Download PDFInfo
- Publication number
- JP2017525355A JP2017525355A JP2017506963A JP2017506963A JP2017525355A JP 2017525355 A JP2017525355 A JP 2017525355A JP 2017506963 A JP2017506963 A JP 2017506963A JP 2017506963 A JP2017506963 A JP 2017506963A JP 2017525355 A JP2017525355 A JP 2017525355A
- Authority
- JP
- Japan
- Prior art keywords
- nucleic acid
- sequence
- lambda integrase
- amino acid
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images












Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases [RNase]; Deoxyribonucleases [DNase]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
- C12N15/902—Stable introduction of foreign DNA into chromosome using homologous recombination
- C12N15/907—Stable introduction of foreign DNA into chromosome using homologous recombination in mammalian cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1241—Nucleotidyltransferases (2.7.7)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2795/00—Bacteriophages
- C12N2795/00011—Details
- C12N2795/10011—Details dsDNA Bacteriophages
- C12N2795/10311—Siphoviridae
- C12N2795/10322—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Molecular Biology (AREA)
- General Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Mycology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Cell Biology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
Description
以下の語及びここで使用される用語は、表示された意味を有するものとする。
43位、319位及び336位で少なくとも1つのアミノ酸変異を含むラムダ・インテグラーゼの例示的、非限定的な実施形態がこれから開示される。
ベストモード及び比較例を含む、本発明の非限定的な例が、具体例を参照することによってより詳細に更に記載されるが、それらは本発明の範囲を多少なりとも限定するものであるとは解釈されるべきではない。
配列番号1
Int−h/218
MGRRRSHERRDLPPNLYIRNNGYYCYRDPRTGKEFGLGRDRRIAITEAIQANIELFSGHKHKPLTARINSDNSVTLHSWLDRYEKILASRGIKQKTLINYMSKIKAIRRGLPDAPLEDITTKEIAAMLNGYIDEGKAASAKLIRSTLSDAFREAIAEGHITTNHVAATRAAKSKVRRSRLTADEYLKIYQAAESSPCWLRLAMELAVVTGQRVGDLCKMKWSDIVDGYLYVEQSKTGVKIAIPTALHIDALGISMKETLDKCKEILGGETIIASTRREPLSSGTVSRYFMRARKASGLSFEGDPPTFHELRSLSARLYEKQISDKFAQHLLGHKSDTMASQYRDDRGREWDKIEIK。
C2インテグラーゼ変異体:
MGRRRSHERRDLPPNLYIRNNGYYCYRDPRTGKEFGLGRDRRIAITEAIQANIELFSGHKHKPLTARINSDNSVTLHSWLDRYEKILASRGIKQKTLINYMSKIKAIRRGLPDAPLEDITTKEIAAMLNGYIDEGKAASAKLIRSTLSDAFREAIAEGHITTNHVAATRAAKSKVRRSRLTADEYLKIYQAAESSPCWLRLAMELAVVTGQRVGDLCKMKWSDIVDGYLYVEQSKTGVKIAIPTALHIDALGISMKETLDKCKEILGGETIIASTRREPLSSGTVSRYFMRARKASGLSFEGDPPTFHELRSLSARLYEKQISDKFAQHLLGHKSVTMASQYRDDRGREWDKIEIK。
C3インテグラーゼ 変異体:
MGRRRSHERRDLPPNLYIRNNGYYCYRDPRTGKEFGLGRDRRFAITEAIQANIELFSGHKHKPLTARINSDNSVTLHSWLDRYEKILASRGIKQKTLINYMSKIKAIRRGLPDAPLEDITTKEIAAMLNGYIDEGKAASAKLIRSTLSDAFREAIAEGHITTNHVAATRAAKSKVRRSRLTADEYLKIYQAAESSPCWLRLAMELAVVTGQRVGDLCKMKWSDIVDGYLYVEQSKTGVKIAIPTALHIDALGISMKETLDKCKEILGGETIIASTRREPLSSGTVSRYFMRARKASGLSFEGDPPTFHELRSLSARLYGKQISDKFAQHLLGHKSVTMASQYRDDRGREWDKIEIK。
ラムダ・インテグラーゼ:
ATGGGAAGAAGGCGAAGTCATGAGCGCCGGGATTTACCCCCTAACCTTTATATAAGAAACAATGGATATTACTGCTACAGGGACCCAAGGACGGGTAAAGAGTTTGGATTAGGCAGAGACAGGCGAATCGCAATCACTGAAGCTATACAGGCCAACATTGAGTTATTTTCAGGACACAAACACAAGCCTCTGACAGCGAGAATCAACAGTGATAATTCCGTTACGTTACATTCATGGCTTGATCGCTACGAAAAAATCCTGGCCAGCAGAGGAATCAAGCAGAAGACACTCATAAATTACATGAGCAAAATTAAAGCAATAAGGAGGGGTCTGCCTGATGCTCCACTTGAAGACATCACCACAAAAGAAATTGCGGCAATGCTCAATGGATACATAGACGAGGGCAAGGCGGCGTCAGCCAAGTTAATCAGATCAACACTGAGCGATGCATTCCGAGAGGCAATAGCTGAAGGCCATATAACAACAAACCATGTCGCTGCCACTCGCGCAGCAAAATCAAAGGTAAGGAGATCAAGACTTACGGCTGACGAATACCTGAAAATTTATCAAGCAGCAGAATCATCACCATGTTGGCTCAGACTTGCAATGGAACTGGCTGTTGTTACCGGGCAACGAGTTGGTGATTTATGCAAAATGAAGTGGTCTGATATCGTAGATGGATATCTTTATGTCGAGCAAAGCAAAACAGGCGTAAAAATTGCCATCCCAACAGCATTGCATATTGATGCTCTCGGAATATCAATGAAGGAAACACTTGATAAATGCAAAGAGATTCTTGGCGGAGAAACCATAATTGCATCTACTCGTCGCGAACCGCTTTCATCCGGCACAGTATCAAGGTATTTTATGCGCGCACGAAAAGCATCAGGTCTTTCCTTCGAAGGGGATCCGCCTACCTTTCACGAGTTGCGCAGTTTGTCTGCAAGACTCTATGAGAAGCAGATAAGCGATAAGTTTGCTCAACATCTTCTCGGGCATAAGTCGGACACCATGGCATCACAGTATCGTGATGACAGAGGCAGGGAGTGGGACAAAATTGAAATCAAATAA。
attB: CTGCTTTTTT ATACTAACTT G。
attP: CAGCTTTTTT ATACTAAGTT G。
attH: CTGCTTTCTT ATACCAAGTG G。
attPH: CAGCTTTCTT ATACCAAGTT G。
attP4X: CAGCTTTATT TCATTAAGTT G。
petF2: CATCGGTGATGTCGGCGAT。
petR: CGGATATAGTTCCTCCTTTCAGCA。
attP−F: cacagaattcCGT CTG TTA CAG GTC ACT AAT ACC ATC T。
attPSOE−R: ACA TTT CCC CGA AAA GTG CCA CCT GAA CAT CAC CGG GAA ATC AAA TAA TGA T。
TEM1prom−F: TTC AGG TGG CAC TTT TCG GGG AAA TGT。
TEM1prom−R: TGT GGA ATT CCT ACA CTA GAA GGA CAG TAT TTG GTA TCT GC。
EcoliAttB−F: CTG AAA ATG TGT TCA CAG GTT GCT。
EcoliattB−R: GCA ATG CCA TCT GGT ATC ACT。
C2遺伝子配列:
ATGGGAAGAAGGCGAAGTCATGAGCGCCGGGATTTACCCCCTAACCTTTATATAAGAAACAATGGATATTACTGCTACAGGGACCCAAGGACGGGTAAAGAGTTTGGATTAGGCAGAGACAGGCGAATCGCAATCACTGAAGCTATACAGGCCAACATTGAGTTATTTTCAGGACACAAACACAAGCCTCTGACAGCGAGAATCAACAGTGATAATTCCGTTACGTTACATTCATGGCTTGATCGCTACGAAAAAATCCTGGCCAGCAGAGGAATCAAGCAGAAGACACTCATAAATTACATGAGCAAAATTAAAGCAATAAGGAGGGGTCTGCCTGATGCTCCACTTGAAGACATCACCACAAAAGAAATTGCGGCAATGCTCAATGGATACATAGACGAGGGCAAGGCGGCGTCAGCCAAGTTAATCAGATCAACGCTGAGCGATGCATTCCGAGAGGCAATAGCTGAAGGCCATATAACAACAAACCATGTCGCTGCCACTCGCGCAGCAAAGTCAAAGGTAAGGAGATCAAGACTTACGGCTGACGAATACCTGAAAATTTATCAAGCAGCAGAATCATCACCATGTTGGCTCAGACTTGCAATGGAACTGGCTGTTGTTACCGGGCAACGAGTTGGTGACTTGTGCAAAATGAAGTGGTCTGATATCGTAGATGGATATCTTTATGTCGAGCAAAGCAAAACAGGCGTAAAAATTGCCATCCCAACAGCATTGCATATTGATGCTCTCGGAATATCAATGAAGGAAACACTTGATAAATGCAAAGAGATTCTTGGCGGAGAAACCATAATTGCATCTACTCGTCGCGAACCGCTTTCATCCGGCACAGTATCAAGGTATTTTATGCGCGCACGAAAAGCATCAGGTCTTTCCTTCGAAGGGGATCCGCCTACCTTTCACGAGTTGCGCAGTTTGTCTGCAAGACTCTATGAGAAGCAGATAAGCGATAAGTTTGCTCAACATCTTCTCGGGCATAAGTCGGTCACCATGGCATCACAGTATCGTGATGACAGAGGCAGGGAGTGGGACAAAATTGAAATCAAATAA。
C3遺伝子配列:
ATGGGAAGAAGGCGAAGTCATGAGCGCCGGGATTTACCCCCTAACCTTTATATAAGAAACAATGGATATTACTGCTACAGGGACCCAAGGACGGGTAAAGAGTTTGGATTAGGCAGAGACAGGCGATTCGCAATCACTGAAGCTATACAGGCCAACATTGAGTTATTTTCAGGACACAAACACAAGCCTCTGACAGCGAGAATCAACAGTGATAATTCCGTTACGTTACATTCATGGCTTGATCGCTACGAAAAAATCCTGGCCAGCAGAGGAATCAAGCAGAAGACACTCATAAATTACATGAGCAAAATTAAAGCAATAAGGAGGGGTCTGCCTGATGCTCCACTTGAAGACATCACCACAAAAGAAATTGCGGCAATGCTCAATGGATACATAGACGAGGGCAAGGCGGCGTCAGCCAAGTTAATCAGATCAACGCTGAGCGATGCATTCCGAGAGGCAATAGCTGAAGGCCATATAACAACAAACCATGTCGCTGCCACTCGCGCGGCAAAGTCAAAGGTAAGGAGATCAAGACTTACGGCTGACGAATACCTGAAAATTTATCAAGCAGCAGAATCATCACCATGTTGGCTCAGACTTGCAATGGAACTGGCTGTTGTTACCGGGCAACGAGTTGGTGACTTGTGCAAAATGAAGTGGTCTGATATCGTAGATGGATATCTTTATGTCGAGCAAAGCAAAACAGGCGTAAAAATTGCCATCCCAACAGCATTGCATATTGATGCTCTCGGAATATCAATGAAGGAAACACTTGATAAATGCAAAGAGATTCTTGGCGGAGAAACCATAATTGCATCTACTCGTCGCGAACCGCTCTCATCCGGCACAGTATCAAGGTATTTTATGCGCGCACGAAAAGCATCAGGTCTTTCCTTCGAAGGGGATCCGCCTACCTTTCACGAGTTGCGCAGTTTGTCTGCAAGACTCTATGGGAAGCAGATAAGCGATAAGTTTGCTCAACATCTTCTCGGGCATAAGTCGGTCACCATGGCATCACAGTATCGTGATGACAGAGGCAGGGAGTGGGACAAAATTGAAATCAAATAA。
C3ミニサークル:
CATCGGTGATGTCGGCGATATAGGCGCCAGCAACCGCACCTGTGGCGCCGGTGATGCCGGCCACGATGCGTCCGGCGTAGAGGATCGAGATCTCGATCCCGCGAAATTAATACGACTCACTATAGGGGAATTGTGAGCGGATAACAATTCCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACATATGGGAAGAAGGCGAAGTCATGAGCGCCGGGATTTACCCCCTAACCTTTATATAAGAAACAATGGATATTACTGCTACAGGGACCCAAGGACGGGTAAAGAGTTTGGATTAGGCAGAGACAGGCGATTCGCAATCACTGAAGCTATACAGGCCAACATTGAGTTATTTTCAGGACACAAACACAAGCCTCTGACAGCGAGAATCAACAGTGATAATTCCGTTACGTTACATTCATGGCTTGATCGCTACGAAAAAATCCTGGCCAGCAGAGGAATCAAGCAGAAGACACTCATAAATTACATGAGCAAAATTAAAGCAATAAGGAGGGGTCTGCCTGATGCTCCACTTGAAGACATCACCACAAAAGAAATTGCGGCAATGCTCAATGGATACATAGACGAGGGCAAGGCGGCGTCAGCCAAGTTAATCAGATCAACGCTGAGCGATGCATTCCGAGAGGCAATAGCTGAAGGCCATATAACAACAAACCATGTCGCTGCCACTCGCGCGGCAAAGTCAAAGGTAAGGAGATCAAGACTTACGGCTGACGAATACCTGAAAATTTATCAAGCAGCAGAATCATCACCATGTTGGCTCAGACTTGCAATGGAACTGGCTGTTGTTACCGGGCAACGAGTTGGTGACTTGTGCAAAATGAAGTGGTCTGATATCGTAGATGGATATCTTTATGTCGAGCAAAGCAAAACAGGCGTAAAAATTGCCATCCCAACAGCATTGCATATTGATGCTCTCGGAATATCAATGAAGGAAACACTTGATAAATGCAAAGAGATTCTTGGCGGAGAAACCATAATTGCATCTACTCGTCGCGAACCGCTCTCATCCGGCACAGTATCAAGGTATTTTATGCGCGCACGAAAAGCATCAGGTCTTTCCTTCGAAGGGGATCCGCCTACCTTTCACGAGTTGCGCAGTTTGTCTGCAAGACTCTATGGGAAGCAGATAAGCGATAAGTTTGCTCAACATCTTCTCGGGCATAAGTCGGTCACCATGGCATCACAGTATCGTGATGACAGAGGCAGGGAGTGGGACAAAATTGAAATCAAACATCATCACCATCACCACTAATGAGAATTCgagctccgtcgacaagcttgcggccgcactcgagcaccaccaccaccaccactgagatccggctgctaacaaagcccgaaaggaagctgagttggctgctgccaccgctgagcaataactagcataaccccttggggcctctaaacgggtcttgaggggttttttgctgaaaggaggaactatatccg。
attP−TEM1:
cacagaattcCGtctgttacaggtcactaataccatctaagtagttgattcatagtgactgcatatattgtgttttacagtattatgtagtctgttttttatgcaaaatctaatttaatatattgatatttatatcattttacgtttctcgttcagcttttttatactaagttggcattataaaaaagcattgcttatcaatttgttgcaacgaacaggtcactatcagtcaaaataaaatcattatttgATTTCCCGGTGATGttcaggtggcacttttcggggaaatgtgcgcggaacccctatttgtttatttttctaaatacattcaaatatgtatccgctcatgagacaataaccctgataaatgcttcaataatattgaaaaaggaagagtatgagtattcaacatttccgtgtcgcccttattcccttttttgcggcattttgccttcctgtttttgctcacccagaaacgctggtgaaagtaaaagatgctgaagatcagttgggtgcacgagtgggttacatcgaactggatctcaacagcggtaagatccttgagagttttcgccccgaagaacgttttccaatgatgagcacttttaaagttctgctatgtggcgcggtattatcccgtattgacgccgggcaagagcaactcggtcgccgcatacactattctcagaatgacttggttgagtactcaccagtcacagaaaagcatcttacggatggcatgacagtaagagaattatgcagtgctgccataaccatgagtgataacactgcggccaacttacttctgacaacgatcggaggaccgaaggagctaaccgcttttttgcacaacatgggggatcatgtaactcgccttgatcgttgggaaccggagctgaatgaagccataccaaacgacgagcgtgacaccacgatgcctgcagcaatggcaacaacgttgcgcaaactattaactggcgaactacttactctagcttcccggcaacaattaatagactggatggaggcggataaagttgcaggaccacttctgcgctcggcccttccggctggctggtttattgctgataaatctggagccggtgagcgtgggtctcgcggtatcattgcagcactggggccagatggtaagccctcccgtatcgtagttatctacacgacggggagtcaggcaactatggatgaacgaaatagacagatcgctgagataggtgcctcactgattaagcattggtaactgtcagaccaagtttactcatatatactttagattgatttaaaacttcatttttaatttaaaaggatctaggtgaagatcctttttgataatctcatgaccaaaatcccttaacgtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactgtccttctagtgtagccgtagttagg。
HOP’:ATGCTTTATTTCATTAAGTTG。
attL:
GCATTATAAAAAAGCATTGCTTATCAATTTGTTGCAACGAACAGGTCACTATCAGTCAAAATACAATCATTATTTGATTTCAATTTTGTCCCACTCCCTCCCG。
PGKプロモーター:
AATTCTACCGGGTAGGGGAGGCGCTTTTCCCAAGGCAGTCTGGAGCATGCGCTTTAGCAGCCCCGCTGGGCACTTGGCGCTACACAAGTGGCCTCTGGCCTCGCACACATTCCACATCCACCGGTAGGCGCCAACCGGCTCCGTTCTTTGGTGGCCCCTTCGCGCCACCTTCTACTCCTCCCCTAGTCAGGAAGTTCCCCCCCGCCCCGCAGCTCGCGTCGTGCAGGACGTGACAAATGGAAGTAGCACGTCTCACTAGTCTCGTGCAGATGGACAGCACCGCTGAGCAATGGAAGCGGGTAGGCCTTTGGGGCAGCGGCCAATAGCAGCT。
HOP’attH4X_F1: GAGTGTTTTCCAACTTGGTTCCATT。
PuroRev24: CACCGTGGGCTTGTACTCGGTC。
pLIR−F1: CTGCATCGATTCAGCTAGCTG。
pLIR−R1: CTGATAGTGACCTGTTCGTTGC。
pPGKssPuro−attP4x(対象ベクター):
gaattcctctgttacaggtcactaataccatctaagtagttgattcatagtgactgcatatgttgtgttttacagtattatgtagtctgttttttatgcaaaatctaatttaatatattgatatttatatcattttacgtttctcgttcagctttatttcattaagttggcattataaaaaagcattgcttatcaatttgttgcaacgaacaggtcactatcagtcaaaataaaatcattatttgatttcaattttgtcccactccctcccgaattctaccgggtaggggaggcgcttttcccaaggcagtctggagcatgcgctttagcagccccgctggcacttggcgctacacaagtggcctctggcctcgcacacattccacatccaccggtagcgccaaccggctccgttctttggtggccccttcgcgccacttctactcctcccctagtcaggaagtttcccccccgccccgcagctcgcgtcgtgcaggacgtgacaaatggaagtagcacgtctcactagtctcgtgcagatggacagcaccgctgagcaatggaagcgggtaggcctttggggcagcggccaatagcagctttgctccttcgctttctgggctcagaggctgggaaggggtgggtccgggggcgggctcaggggcgggctcaggggcggggcgggcgcccgaaggtcctccggaggcccggcattctgcacgcttcaaaagcgcacgtctgccgcgctgttctcctcttcctcatctccgggcctttcgaccaattcgctgtctgcgagggccagctgttggggtgagtactccctctcaaaagcgggcatgacttctgcgctaagattgtcagtttccaaaaacgaggaggatttgatattcacctggcccgcggtgatgcctttgagggtggccgcgtccatctggtcagaaaagacaatctttttgttgtcaagcttgaggtgtggcaggcttgagatctggccatacacttgagtgacaatgacatccactttgcctttctctccacaggtgtccactcccaggtccaactgcagatgaccgagtacaagcccacggtgcgcctcgccacccgcgacgacgtcccccgggccgtacgcaccctcgccgccgcgttcgccgactaccccgccacgcgccacaccgtcgacccggaccgccacatcgagcgggtcaccgagctgcaagaactcttcctcacgcgcgtcgggctcgacatcggcaaggtgtgggtcgcggacgacggcgccgcggtggcggtctggaccacgccggagagcgtcgaagcgggggcggtgttcgccgagatcggcccgcgcatggccgagttgagcggttcccggctggccgcgcagcaacagatggaaggcctcctggcgccgcaccggcccaaggagcccgcgtggttcctggccaccgtcggcgtctcgcccgaccaccagggcaagggtctgggcagcgccgtcgtgctccccggagtggaggcggccgagcgcgccggggtgcccgccttcctggagacctccgcgccccgcaacctccccttctacgagcggctcggcttcaccgtcaccgccgacgtcgaggtgcccgaaggaccgcgcacctggtgcatgacccgcaagcccggtgcctgatctagagctcgctgatcagcctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctggggatgcggtgggctctatggcttctgaggcggaaagaaccagctggggctcgagatccactagttctagcctcgaggctagagcggccgccaccgcggtggagctccaattcgccctatagtgagtcgtattacgcgcgctcactggccgtcgttttacaacgtcgtgactgggaaaaccctggcgttacccaacttaatcgccttgcagcacatccccctttcgccagctggcgtaatagcgaagaggcccgcaccgatcgcccttcccaacagttgcgcagcctgaatggcgaatgggacgcgccctgtagcggcgcattaagcgcggcgggtgtggtggttacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttcccttcctttctcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatttagtgctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggccatcgccctgatagacggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttccaaactggaacaacactcaaccctatctcggtctattcttttgatttataagggattttgccgatttcggcctattggttaaaaaatgagctgatttaacaaaaatttaacgcgaattttaacaaaatattaacgcttacaatttaggtggcacttttcggggaaatgtgcgcggaacccctatttgtttatttttctaaatacattcaaatatgtatccgctcatgagacaataaccctgataaatgcttcaataatattgaaaaaggaagagtatgagtattcaacatttccgtgtcgcccttattcccttttttgcggcattttgccttcctgtttttgctcacccagaaacgctggtgaaagtaaaagatgctgaagatcagttgggtgcacgagtgggttacatcgaactggatctcaacagcggtaagatccttgagagttttcgccccgaagaacgttttccaatgatgagcacttttaaagttctgctatgtggcgcggtattatcccgtattgacgccgggcaagagcaactcggtcgccgcatacactattctcagaatgacttggttgagtactcaccagtcacagaaaagcatcttacggatggcatgacagtaagagaattatgcagtgctgccataaccatgagtgataacactgcggccaacttacttctgacaacgatcggaggaccgaaggagctaaccgcttttttgcacaacatgggggatcatgtaactcgccttgatcgttgggaaccggagctgaatgaagccataccaaacgacgagcgtgacaccacgatgcctgtagcaatggcaacaacgttgcgcaaactattaactggcgaactacttactctagcttcccggcaacaattaatagactggatggaggcggataaagttgcaggaccacttctgcgctcggcccttccggctggctggtttattgctgataaatctggagccggtgagcgtgggtctcgcggtatcattgcagcactggggccagatggtaagccctcccgtatcgtagttatctacacgacggggagtcaggcaactatggatgaacgaaatagacagatcgctgagataggtgcctcactgattaagcattggtaactgtcagaccaagtttactcatatatactttagattgatttaaaacttcatttttaatttaaaaggatctaggtgaagatcctttttgataatctcatgaccaaaatcccttaacgtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactgtccttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcctacatacctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgtgcacacagcccagcttggagcgaacgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccgaagggagaaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtcaggggggcggagcctatggaaaaacgccagcaacgcggcctttttacggttcctggccttttgctggccttttgctcacatgttctttcctgcgttatcccctgattctgtggataaccgtattaccgcctttgagtgagctgataccgctcgccgcagccgaacgaccgagcgcagcgagtcagtgagcgaggaagcggaagagcgcccaatacgcaaaccgcctctccccgcgcgttggccgattcattaatgcagctggcacgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagctcactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggaattgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgccaagcgcgcaattaaccctcactaaagggaacaaaagctgggtaccgggccccccctcgaggtcgacggtatcgataagcttgatatc。
pCMVssKZ−IntC3−CNLS(インテグラーゼ発現プラスミド):
gaattcctcacatgttctttcctgcgttatcccctgattctgtggataaccgtattaccgccatgcattagttattaatagtaatcaattacggggtcattagttcatagcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggctgaccgcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttccattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgccaagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgaccttatgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatggtgatgcggttttggcagtacatcaatgggcgtggatagcggtttgactcacggggatttccaagtctccaccccattgacgtcaatgggagtttgttttggcaccaaaatcaacgggactttccaaaatgtcgtaacaactccgccccattgacgcaaatgggcggtaggcgtgtacggtgggaggtctatataagcagagctggtttagtgaaccgtcagatccgctagcaattcgctgtctgcgagggccagctgttggggtgagtactccctctcaaaagcgggcatgacttctgcgctaagattgtcagtttccaaaaacgaggaggatttgatattcacctggcccgcggtgatgcctttgagggtggccgcgtccatctggtcagaaaagacaatctttttgttgtcaagcttgaggtgtggcaggcttgagatctggccatacacttgagtgacaatgacatccactttgcctttctctccacaggtgtccactcccaggtccaactgcagctcgaggtccaccatgggaagaaggcgaagtcatgagcgccgggatttaccccctaacctttatataagaaacaatggatattactgctacagggacccaaggacgggtaaagagtttggattaggcagagacaggcgattcgcaatcactgaagctatacaggccaacattgagttattttcaggacacaaacacaagcctctgacagcgagaatcaacagtgataattccgttacgttacattcatggcttgatcgctacgaaaaaatcctggccagcagaggaatcaagcagaagacactcataaattacatgagcaaaattaaagcaataaggaggggtctgcctgatgctccacttgaagacatcaccacaaaagaaattgcggcaatgctcaatggatacatagacgagggcaaggcggcgtcagccaagttaatcagatcaacgctgagcgatgcattccgagaggcaatagctgaaggccatataacaacaaaccatgtcgctgccactcgcgcggcaaagtcaaaggtaaggagatcaagacttacggctgacgaatacctgaaaatttatcaagcagcagaatcatcaccatgttggctcagacttgcaatggaactggctgttgttaccgggcaacgagttggtgacttgtgcaaaatgaagtggtctgatatcgtagatggatatctttatgtcgagcaaagcaaaacaggcgtaaaaattgccatcccaacagcattgcatattgatgctctcggaatatcaatgaaggaaacacttgataaatgcaaagagattcttggcggagaaaccataattgcatctactcgtcgcgaaccgctctcatccggcacagtatcaaggtattttatgcgcgcacgaaaagcatcaggtctttccttcgaaggggatccgcctacctttcacgagttgcgcagtttgtctgcaagactctatgggaagcagataagcgataagtttgctcaacatcttctcgggcataagtcggtcaccatggcatcacagtatcgtgatgacagaggcagggagtgggacaaaattgaaatcaaatccggaggcggccctaagaagaagagaaaggtatgataatctagagctcgctgatcagcctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctggggatgcggtgggctctatggcttctgaggcggaaagaaccagctggggctcgagatccactagttctagcctcgaggctagagcggccgccaccgcggtggagctccaattcgccctatagtgagtcgtattacgcgcgctcactggccgtcgttttacaacgtcgtgactgggaaaaccctggcgttacccaacttaatcgccttgcagcacatccccctttcgccagctggcgtaatagcgaagaggcccgcaccgatcgcccttcccaacagttgcgcagcctgaatggcgaatgggacgcgccctgtagcggcgcattaagcgcggcgggtgtggtggttacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttcccttcctttctcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatttagtgctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggccatcgccctgatagacggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttccaaactggaacaacactcaaccctatctcggtctattcttttgatttataagggattttgccgatttcggcctattggttaaaaaatgagctgatttaacaaaaatttaacgcgaattttaacaaaatattaacgcttacaatttaggtggcacttttcggggaaatgtgcgcggaacccctatttgtttatttttctaaatacattcaaatatgtatccgctcatgagacaataaccctgataaatgcttcaataatattgaaaaaggaagagtatgagtattcaacatttccgtgtcgcccttattcccttttttgcggcattttgccttcctgtttttgctcacccagaaacgctggtgaaagtaaaagatgctgaagatcagttgggtgcacgagtgggttacatcgaactggatctcaacagcggtaagatccttgagagttttcgccccgaagaacgttttccaatgatgagcacttttaaagttctgctatgtggcgcggtattatcccgtattgacgccgggcaagagcaactcggtcgccgcatacactattctcagaatgacttggttgagtactcaccagtcacagaaaagcatcttacggatggcatgacagtaagagaattatgcagtgctgccataaccatgagtgataacactgcggccaacttacttctgacaacgatcggaggaccgaaggagctaaccgcttttttgcacaacatgggggatcatgtaactcgccttgatcgttgggaaccggagctgaatgaagccataccaaacgacgagcgtgacaccacgatgcctgtagcaatggcaacaacgttgcgcaaactattaactggcgaactacttactctagcttcccggcaacaattaatagactggatggaggcggataaagttgcaggaccacttctgcgctcggcccttccggctggctggtttattgctgataaatctggagccggtgagcgtgggtctcgcggtatcattgcagcactggggccagatggtaagccctcccgtatcgtagttatctacacgacggggagtcaggcaactatggatgaacgaaatagacagatcgctgagataggtgcctcactgattaagcattggtaactgtcagaccaagtttactcatatatactttagattgatttaaaacttcatttttaatttaaaaggatctaggtgaagatcctttttgataatctcatgaccaaaatcccttaacgtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactgtccttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcctacatacctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgtgcacacagcccagcttggagcgaacgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccgaagggagaaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtcaggggggcggagcctatggaaaaacgccagcaacgcggcctttttacggttcctggccttttgctggccttttgctcacatgttctttcctgcgttatcccctgattctgtggataaccgtattaccgcctttgagtgagctgataccgctcgccgcagccgaacgaccgagcgcagcgagtcagtgagcgaggaagcggaagagcgcccaatacgcaaaccgcctctccccgcgcgttggccgattcattaatgcagctggcacgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagctcactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggaattgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgccaagcgcgcaattaaccctcactaaagggaacaaaagctgggtaccgggccccccctcgaggtcgacggtatcgataagcttgatatc。
attH4X: acgctttatttcattaagttg。
本例は、図2に描かれた手順に従う。C3INT−HIS−PET22b(+)は、petF2(配列番号10)及びpetR(配列番号11)で増幅し、その後、PCR産物を分子内でライゲーションし、C3INT−HISミニサークルを作出した。attP−PET22b(+)は、attP−F(配列番号12)及びattPSOE−R(配列番号13)で増幅し、一方でPET22b(+)は、TEM1prom−F(配列番号14)及びTEM1promR(配列番号15)で増幅し、それぞれattP及びアンピシリン耐性遺伝子をコードするPCR産物を生成した。スプライス・オーバーラップ・エクステンション・PCR(SOE−PCR)をこれら2つのPCR産物を用いてattP−F(配列番号12及びTEM1prom−R(配列番号15)を使用して行った。次いでPCR産物を分子内でライゲーションし、attP−TEM1ミニサークルを作出した。100ngのC3INT−HISミニサークルと100ngのattP−TEM1ミニサークルとを組み合わせて、25μLのエレクトロコンピテントTG1細胞にエレクトロポレーションした。細胞は、多様な濃度(0.01mg/mL、0.02mg/mL、0.05mg/mL、0.07mg/mL及び0.1mg/mL)のアンピシリン−LB寒天プレート上への播種の前に1時間に亘って回復させた。インキュベーションを37℃で12〜14時間に亘って行い、C3インテグラーゼの発現及びC3インテグラーゼによりアンピシリン耐性カセットの染色体統合を可能にした。コロニーPCRは、EcoliAttB−F(配列番号16)及びEcoliAttB−R(配列番号17)、TEM1prom−F(配列番号14)及びEcoliAttB−F(配列番号16)、又はTEM1prom−F(配列番号14)及びEcoliAttB−R(配列番号17)で行い、アンピシリン耐性カセットの染色体統合の存在を検証した。PCR産物はまた、同じプライマーによって配列を決定し、結果を確認した。シーケンシングは、染色体attB部位への正確な統合イベントを示した(図3C)。
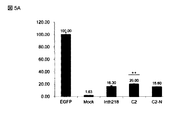
本例は、親Int−h/218及び選択された変異体(C2、C3及びその示されたバリアント)の組み換え活性を実証した。図4Aは、インビトロ転写/翻訳により産生されたインテグラーゼを使用するインビトロ分子内組み換え反応からの結果を図示する。それぞれインテグラーゼ(Int−h/218、C2、C3又はそのバリアント)をコードするプラスミドをプライマーIntRBS−F及びINTstop−Rを用いて増幅し、PCR産物をプライマーユニバーサル及びINTstop−Rを用いて再増幅して、インビトロ転写−翻訳(IVT)において必要なT7プロモーター及びリボソーム結合部位を有するインテグラーゼアンプリコンを得た。20gの各インテグラーゼアンプリコンを総容量9μLのPURエクスプレス(登録商標)・インビトロ・タンパク質・シンセーシス・キット(PURExpressR In Vitro Protein Synthesis Kit)を使用して30℃1時間に亘って発現させた。次に、attB/attP部位、attPH/attH部位又はattH4x/attP4x部位(図1)の何れかを含む10ngのプラスミド基質を総容量10μLまで添加することによって分子内組み換えを行った。混合物を37℃で2時間インキュベートさせた。その後、組み換え効率のリアルタイムPCR定量のために1μL採取する前に反応液を1/10に希釈した。リアルタイムPCR定量は、250nMの各プライマーpLIR−F1(配列番号27)及びpLIR−R1(配列番号28)を用いて終容量20μLのエッソアドバンスドTM・ユニバーサル・SYBR(登録商標)グリーン・スーパーミックス中で行った。組み換えインテグラーゼ・タンパク質の活性は、attB/attPプラスミド基質(値を1に設定)の相対的活性で示す。エラーバーは、2つの独立した実験の標準偏差を示す。
HT1080細胞株における内因性ターゲティングのために、トランスフェクションの1日前に10cmの細胞培養皿当たり3×106細胞をダルベッコ改変イーグル培地[10%のFBS、1%のL−グルタミン及び各100ユニット/mLのペニシリン/ストレプトマイシンが補われたDMEM(ライフ・テクノロジー)]中に播種し、トランスフェクション時には70〜90%のコンフルエントを達成した。トランスフェクションは、リポフェクタミン2000試薬(ライフ・テクノロジー)を使用して行った。プラスミドDNA−脂質複合体は、5ngの対象ベクター(pPGKssPuro−attP4x(配列番号29))及び75μLのOpti−MEM培地(ライフ・テクノロジー)中に希釈された2.5μLのリポフェクタミン2000試薬を含む、75μLのOpti−MEM培地中に希釈された100ngのインテグラーゼ発現プラスミド(pCMVssKZ−IntC3−CNLS(配列番号30))を混合し、20分間室温でインキュベートすることにより調製された。トランスフェクションミックスを(抗生物質を含まないDMEM下で)細胞に対して添加し、4〜6時間に亘ってトランスフェクションを進めて、それに続いて、新しい培地で置き換えることによって複合体を取り除いた。トランスフェクションの48時間後、細胞を1mL当たり3μgのピューロマイシンを含む殖培地中で増殖させ、ピューロマイシン耐性コロニーを選択した。選択の3週間後、ピューロマイシン耐性コロニーを収穫し、展開した。DNeasy・ブラッド・アンド・ティッシュ・キット(DNeasy Blood & Tissue Kit(キアゲン))を用いてゲノムDANを抽出した。
HT1080細胞株でのエピソーム性分子内組み換えアッセイのために、トランスフェクションの1日前に6ウェルプレートの1ウェル当たりに3×105細胞をダルベッコ改変イーグル培地[10%のFBS、1%のL−グルタミン及び各100ユニット/mLのペニシリン/ストレプトマイシンが補われたDMEM(ライフ・テクノロジー)]中に播種し、トランスフェクション時には70〜90%のコンフルエントが達成された。トランスフェクションは、リポフェクタミン2000試薬を使用して行った。ウェル毎の全てのトランスフェクションについて、1.5μgのpLIRと、100μLのOpti−MEM培地中に希釈された6μLのリポフェクタミン2000試薬を含む100μLのOpti−MEM中に希釈された1.5μgのλインテグラーゼ発現プラスミドとを混合し、室温で20分間インキュベートすることにより、プラスミドDNA−脂質複合体を調製した。トランスフェクションミックスを(抗生物質を含まないDMEM下で)細胞に対して滴下によって添加し、トランスフェクションを4〜6時間に亘って進めて、それに続いて、新しいDMEM培地で置き換えることによって複合体を取り除いた。トランスフェクションの48〜72時間後、細胞をトリプシン処理し、DMEMでエッペンドルフチューブに回収し、遠心分離(5分間に亘り1000×rcf)により沈殿させ、1mLの新しいDMEMに再懸濁した。BD FACSキャリブル(登録商標)マシーン(BD FACSCalibur(登録商標) machine、ベクトン−ディッキンソン)上でのFACSによって、GFP陽性細胞を定量化した。
GoTaq Flexi DNAポリメラーゼ(プロメガ)を使用し、プライマーHOP’ attH4X_F1(配列番号25)及びPuroRev24(配列番号26)、並びに200ngのゲノムDNAをテンプレートとして用いて、PCR反応当たり50μL容量中でPCRを行った。以下のサーマル・サイクリング・パラメータをPCRのために使用した:95℃5分間の初期段階、95℃1分間、57℃30秒間及び72℃1分間を35サイクル、並びに72℃5分間の最終段階。PCRサンプルは、Tris−ホウ酸−EDTAバッファー中0.8%アガロースゲルでの電気泳動によって分析した。
本発明のインテグラーゼバリアント、並びにattH/attPH及びattH4X/attP4X基質対の使用によって改善されたインビトロ組み換えが、ここで記載されるインテグラーゼバリアントは生物工学的応用、例えば、組み換えを基礎にしたクローニングへの応用などのための有用な試薬手段であり得ることを示す。
Claims (37)
- 配列番号1に記載のラムダ・インテグラーゼの336位、319位及び43位の少なくとも1つでアミノ酸変異を含むラムダ・インテグラーゼ。
- 請求項1に記載のラムダ・インテグラーゼであって、配列番号1に記載の前記ラムダ・インテグラーゼの43位、319位及び336位でアミノ酸変異を含むラムダ・インテグラーゼ。
- 請求項1又は2に記載のラムダ・インテグラーゼであって、配列43位でアミノ酸残基イソロイシンが芳香族アミノ酸により置換されているラムダ・インテグラーゼ。
- 請求項3に記載の当該ラムダ・インテグラーゼであって、前記芳香族アミノ酸が、フェニルアラニン、チロシン及びトリプトファンからなる群より選択されるラムダ・インテグラーゼ。
- 請求項1〜4の何れか1項に記載のラムダ・インテグラーゼであって、配列319位のアミノ酸残基グルタメート(glutamate)がグリシンにより置換されているラムダ・インテグラーゼ。
- 請求項1〜5の何れか1項に記載のラムダ・インテグラーゼであって、配列336位のアミノ酸残基アスパルテート(aspartate)が疎水性アミノ酸により置換されているラムダ・インテグラーゼ。
- 請求項6に記載のラムダ・インテグラーゼであって、前記疎水性アミノ酸が脂肪族アミノ酸であるラムダ・インテグラーゼ。
- 請求項7に記載のラムダ・インテグラーゼであって、前記脂肪族アミノ酸が、イソロイシン、ロイシン及びバリンからなる群より選択されるラムダ・インテグラーゼ。
- 請求項1〜8の何れか1項に記載のラムダ・インテグラーゼであって、配列番号3に記載のラムダ・インテグラーゼにアミノ酸変異I43F、E319G及びD336Vを含むラムダ・インテグラーゼ。
- 請求項1に記載のラムダ・インテグラーゼであって、配列番号1に記載のラムダ・インテグラーゼの336位にアミノ酸変異を含むラムダ・インテグラーゼ。
- 請求項10に記載のラムダ・インテグラーゼであって、配列336位のアミノ酸残基アスパルテート(aspartate)が疎水性アミノ酸により置換されているラムダ・インテグラーゼ。
- 請求項11に記載のラムダ・インテグラーゼであって、前記疎水性アミノ酸が脂肪族アミノ酸であるラムダ・インテグラーゼ。
- 請求項12に記載のラムダ・インテグラーゼであって、前記脂肪族アミノ酸がイソロイシン、ロイシン及びバリンからなる群より選択されるラムダ・インテグラーゼ。
- 請求項10〜13の何れか1項に記載のラムダ・インテグラーゼであって、配列番号2に記載のラムダ・インテグラーゼにおいてアミノ酸変異D336Vを含むラムダ・インテグラーゼ。
- 請求項10〜14の何れか1項に記載のラムダ・インテグラーゼであって、配列319位のアミノ酸残基グルタメート(glutamate)がグリシンにより置換されているラムダ・インテグラーゼ。
- 請求項15に記載のラムダ・インテグラーゼであって、配列番号3に記載のラムダ・インテグラーゼにおいてアミノ酸変異E319G及びD336Vを含むラムダ・インテグラーゼ。
- 請求項1〜16の何れか1項に記載のラムダ・インテグラーゼをコードするヌクレオチド配列を含む核酸分子。
- 請求項15に記載の核酸分子であって、当該核酸分子の発現を可能にする調節配列に稼働可能に連結されている核酸分子。
- 請求項16に記載の核酸分子であって、前記調節配列がプロモーター配列を含む核酸分子。
- 請求項17〜19の何れか1項に記載の核酸分子であって、ベクター内に位置している核酸分子。
- 請求項17〜20の何れか1項に記載の当該核酸分子を含むホスト細胞。
- 標的核酸に対して興味のある核酸を組み換える方法であって、請求項1〜16の何れか1項に記載のラムダ・インテグラーゼの存在下で前記興味のある核酸を含む対象核酸と前記標的核酸とを接触させることを含む方法。
- 請求項22に記載の方法であって、前記標的核酸がDNAを含む方法。
- 請求項22に記載の方法であって、前記標的核酸がゲノムDNAを含む方法。
- 請求項22〜24の何れか1項に記載の方法であって、前記標的核酸が、attH配列(配列番号7)及びattH4X配列(配列番号31)からなる群より選択される配列を含む方法。
- 請求項22〜25の何れか1項に記載の方法であって、前記対象核酸がベクターである方法。
- 請求項26に記載の方法であって、前記対象核酸が、attPH配列(配列番号8)及びattP4X配列(配列番号9)からなる群より選択される配列を含む方法。
- 請求項22〜27の何れか1項に記載の方法であって、前記配列特異的組み換えが1種類以上のコファクターの存在下で実行される方法。
- 請求項28に記載の方法であって、前記コファクターがXIS、FIS及びIHFからなる群より選択される方法。
- 請求項24〜29の何れか1項に記載の方法であって、前記ゲノムDNAが細胞に含まれる方法。
- 配列特異的組み換えキットであって、
a.興味のある核酸が挿入できる対象核酸、及び
b.請求項1〜16の何れか1項に記載のラムダ・インテグラーゼ、又は請求項17〜20の何れか1項に記載の核酸
を含むキット。 - 請求項31に記載のキットであって、更に、前記対象核酸に前記興味のある核酸を挿入するための少なくとも1種類の試薬を含むキット。
- 請求項31又は32に記載のキットであって、前記対象核酸が、attPH配列(配列番号8)及びattP4X配列(配列番号9)からなる群より選択される配列を含むキット。
- 請求項31又は32に記載のキットであって、前記対象核酸がattH配列(配列番号7)及びattH4X配列(配列番号31)からなる群より選択される配列を含むキット。
- 請求項31〜34の何れか1項に記載のキットであって、更に、バッファー(複数若しくは単数)及び/又は前記興味のある核酸を問題の標的核酸と組み換えるための説明書を含むキット。
- 請求項31〜35の何れか1項に記載のキットであって、更に、達成された配列特異的組み換えイベントを測定するための少なくとも1種類の試薬を含むキット。
- 請求項36に記載のキットであって、前記試薬がプライマー対であるキット。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| GBGB1414130.3A GB201414130D0 (en) | 2014-08-08 | 2014-08-08 | Mutants of the bacteriophage lambda integrase |
| GB1414130.3 | 2014-08-08 | ||
| PCT/SG2015/050255 WO2016022075A1 (en) | 2014-08-08 | 2015-08-11 | Mutants of the bacteriophage lambda integrase |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2017525355A true JP2017525355A (ja) | 2017-09-07 |
| JP2017525355A5 JP2017525355A5 (ja) | 2018-09-13 |
| JP6616822B2 JP6616822B2 (ja) | 2019-12-04 |
Family
ID=51629549
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2017506963A Active JP6616822B2 (ja) | 2014-08-08 | 2015-08-11 | バクテリオファージ・ラムダ・インテグラーゼの変異体 |
Country Status (7)
| Country | Link |
|---|---|
| US (2) | US10344301B2 (ja) |
| EP (1) | EP3177717A4 (ja) |
| JP (1) | JP6616822B2 (ja) |
| CN (1) | CN108064279B (ja) |
| GB (1) | GB201414130D0 (ja) |
| SG (2) | SG11201700955SA (ja) |
| WO (1) | WO2016022075A1 (ja) |
Families Citing this family (14)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB201414130D0 (en) * | 2014-08-08 | 2014-09-24 | Agency Science Tech & Res | Mutants of the bacteriophage lambda integrase |
| GB201603577D0 (en) * | 2016-03-01 | 2016-04-13 | Oxford Genetics Ltd | Promoter |
| US11078493B2 (en) | 2016-06-21 | 2021-08-03 | Nanyang Technological University | Site-specific DNA recombination |
| FI3638776T3 (fi) | 2017-06-14 | 2025-12-09 | Univ Dresden Tech | Menetelmiä ja välineitä genomien geneettiseen muokkaamiseen hyödyntäen suunnittelu-DNA:ta rekombinoivia entsyymejä |
| WO2021021964A2 (en) | 2019-07-29 | 2021-02-04 | Chondrial Therapeutics, Inc. | Methods of quantifying frataxin and frataxin fusion proteins |
| BR112022001657A2 (pt) * | 2019-07-29 | 2022-03-22 | Univ Indiana Trustees | Polipeptídeo de fusão, sequência de ácidos nucleicos, cassete de expressão, vetor de expressão, célula hospedeira, composição farmacêutica, e, métodos para tratar a ataxia de friedreich e para tratar uma doença associada à ataxia de friedreich |
| KR20220108094A (ko) | 2019-11-25 | 2022-08-02 | 라리마 테라퓨틱스, 인코포레이티드 | 프라탁신 활성의 정량화 방법 |
| US11976100B2 (en) | 2020-04-30 | 2024-05-07 | Larimar Therapeutics, Inc. | Methods for treating myelin associated diseases and mitochondria associated diseases |
| CN116867506A (zh) | 2020-12-12 | 2023-10-10 | 拉利玛生物医药公司 | 包括弗里德赖希共济失调蛋白融合蛋白的药物组合物及其使用方法 |
| CN117178056A (zh) * | 2021-03-12 | 2023-12-05 | 南洋理工大学 | 用于生产无缝dna载体的方法 |
| WO2023234866A1 (en) * | 2022-05-31 | 2023-12-07 | Nanyang Technological University | Method for production of a eukaryotic host cell or cell line for lambda-integrase-mediated recombination |
| CN118546903A (zh) * | 2023-02-24 | 2024-08-27 | 尧唐(上海)生物科技有限公司 | 能介导大片段dna整合到哺乳动物基因组的酶及其应用 |
| WO2024181932A1 (en) * | 2023-03-02 | 2024-09-06 | Nanyang Technological University | Lambda integrase mutants and methods of their use |
| CN121693564A (zh) | 2023-06-26 | 2026-03-17 | 夏威夷大学 | 进化的整合酶和其用于基因组编辑的方法 |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20130017578A1 (en) * | 2009-11-06 | 2013-01-17 | Farid John Ghadessy | Muteins of the bacteriophage lambda integrases |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| HUP0402659A3 (en) * | 2001-05-30 | 2008-07-28 | Glaxo Group Ltd | Chromosome-based platforms |
| CA2522166C (en) | 2001-05-30 | 2011-05-03 | Chromos Molecular Systems, Inc. | Lambda integrase mutein for use in recombination |
| US20100192985A1 (en) * | 2008-11-11 | 2010-08-05 | Wolfgang Aehle | Compositions and methods comprising serine protease variants |
| GB201414130D0 (en) * | 2014-08-08 | 2014-09-24 | Agency Science Tech & Res | Mutants of the bacteriophage lambda integrase |
-
2014
- 2014-08-08 GB GBGB1414130.3A patent/GB201414130D0/en not_active Ceased
-
2015
- 2015-08-11 SG SG11201700955SA patent/SG11201700955SA/en unknown
- 2015-08-11 EP EP15829802.6A patent/EP3177717A4/en active Pending
- 2015-08-11 JP JP2017506963A patent/JP6616822B2/ja active Active
- 2015-08-11 CN CN201580054345.8A patent/CN108064279B/zh active Active
- 2015-08-11 WO PCT/SG2015/050255 patent/WO2016022075A1/en not_active Ceased
- 2015-08-11 SG SG10202005089XA patent/SG10202005089XA/en unknown
- 2015-08-11 US US15/502,783 patent/US10344301B2/en active Active
-
2019
- 2019-06-03 US US16/430,410 patent/US11414681B2/en active Active
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20130017578A1 (en) * | 2009-11-06 | 2013-01-17 | Farid John Ghadessy | Muteins of the bacteriophage lambda integrases |
Non-Patent Citations (2)
| Title |
|---|
| J. MOL. BIOL., vol. 252, JPN6019024965, 1995, pages 178 - 188, ISSN: 0004066973 * |
| NUCLEIC ACIDS RESEARCH, vol. 38(4), JPN6019024964, 2010, pages 25, ISSN: 0004066972 * |
Also Published As
| Publication number | Publication date |
|---|---|
| CN108064279B (zh) | 2022-03-11 |
| SG11201700955SA (en) | 2017-03-30 |
| US20170327847A1 (en) | 2017-11-16 |
| GB201414130D0 (en) | 2014-09-24 |
| CN108064279A (zh) | 2018-05-22 |
| EP3177717A4 (en) | 2017-12-27 |
| JP6616822B2 (ja) | 2019-12-04 |
| SG10202005089XA (en) | 2020-07-29 |
| EP3177717A1 (en) | 2017-06-14 |
| WO2016022075A1 (en) | 2016-02-11 |
| US11414681B2 (en) | 2022-08-16 |
| US10344301B2 (en) | 2019-07-09 |
| US20200063165A1 (en) | 2020-02-27 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6616822B2 (ja) | バクテリオファージ・ラムダ・インテグラーゼの変異体 | |
| US12612650B2 (en) | Methods for improved homologous recombination and compositions thereof | |
| US20220220508A1 (en) | Engineered casx systems | |
| JP7109547B2 (ja) | 真核ゲノム修飾のための操作されたCas9システム | |
| JP6688231B2 (ja) | 標的遺伝子座を修飾するための方法及び組成物 | |
| EP3222728B1 (en) | Method for regulating gene expression using cas9 protein expressed from two vectors | |
| US20240336905A1 (en) | Class ii, type v crispr systems | |
| JP2024009934A (ja) | 細胞ゲノムにおける、相同組換え修復(hdr)の効率を上昇させるための方法 | |
| CN109804066A (zh) | 可编程cas9-重组酶融合蛋白及其用途 | |
| BR112019028146A2 (pt) | rna guia sintético para sistemas ativadores de crispr/cas | |
| CN110520163A (zh) | 独立于dna双链断裂的靶向基因编辑平台及其用途 | |
| JP6892721B2 (ja) | 新規ヌクレアーゼドメインおよびその利用 | |
| JP2016523084A (ja) | 標的組込み | |
| EA038500B1 (ru) | Термостабильные нуклеазы cas9 | |
| KR20250166280A (ko) | Cas단백질 및 이의 돌연변이체, 이에 대응하는 유전자 편집 시스템 및 응용 | |
| JP2013094148A (ja) | 新規dna結合ドメインおよびそれを含む新規dna切断酵素 | |
| JP2025508794A (ja) | 融合タンパク質 | |
| WO2025160202A1 (en) | Engineered large serine recombinases | |
| JP2026500173A (ja) | シトシン塩基エディターゲノム編集特異性及び効率を改善するための方法及び組成物 | |
| EP2772539B1 (en) | Bari promoter sequences and its uses | |
| EA048319B1 (ru) | СКОНСТРУИРОВАННЫЕ СИСТЕМЫ CasX | |
| HK40040051A (en) | Engineered cas9 systems for eukaryotic genome modification | |
| Akopian | Site-specific DNA recombinases with novel sequence recognition | |
| HK40027119A (en) | Methods for improved homologous recombination and compositions thereof | |
| Abil | Study and engineering of nucleic acid-binding repeat proteins |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20170316 Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20170410 |
|
| RD01 | Notification of change of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7426 Effective date: 20170316 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20180802 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20180802 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20190626 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20190702 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20191001 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20191023 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20191108 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6616822 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |