JP2022019308A - Fadgdhおよびその製造方法 - Google Patents
Fadgdhおよびその製造方法 Download PDFInfo
- Publication number
- JP2022019308A JP2022019308A JP2020123071A JP2020123071A JP2022019308A JP 2022019308 A JP2022019308 A JP 2022019308A JP 2020123071 A JP2020123071 A JP 2020123071A JP 2020123071 A JP2020123071 A JP 2020123071A JP 2022019308 A JP2022019308 A JP 2022019308A
- Authority
- JP
- Japan
- Prior art keywords
- bipyridine
- fadgdh
- seq
- amino acid
- lutenium
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images




Landscapes
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
【課題】
メディエーターに対する反応性向上したフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを提供すること。
【解決手段】
配列番号1と70%以上の同一性を有するアミノ酸配列を有し、配列番号1の322位に相当するアミノ酸残基がトレオニン、セリン、アスパラギンまたはグルタミンである、フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
【選択図】なし
メディエーターに対する反応性向上したフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを提供すること。
【解決手段】
配列番号1と70%以上の同一性を有するアミノ酸配列を有し、配列番号1の322位に相当するアミノ酸残基がトレオニン、セリン、アスパラギンまたはグルタミンである、フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
【選択図】なし
Description
本発明は、フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ(以下、「FADGDH」とも称する)に関する。より詳しくは、本発明は、メディエーターに対する反応性が改善されたムコール(Mucor)属由来の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ、該FADGDHをコードするDNA、該DNAを有する発現ベクター、該発現ベクターで形質転換した形質転換体、該変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの製造方法、該変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを使用したグルコースの測定方法、および、グルコースセンサに関する。
自己血糖測定(SMBG:Self-Monitoring of Blood Glucose)は糖尿病患者が自己の血糖値を管理し、治療に活用するために重要である。SMBGに関しては、多くの方法で実用化が進んでおり、検体の微量化、測定時間の短縮、装置の小型化の点で電気化学的なセンシングが有利である。血糖測定技術におけるセンシングの手法としては血液中のグルコースを基質とする酵素が利用される。そのような酵素の例としては、グルコースデヒドロゲナーゼ(EC 1.1.1.47)が挙げられる。近年、グルコースデヒドロゲナーゼ(以下、GDHとも表す。)の中でもアスペルギルス・オリゼ(Aspergillus oryzae)由来のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼは、グルコースに対する選択性の高さや熱に対する安定性が高いという利点から、グルコースセンサ用酵素として実用化されている。特許文献1および特許文献2には、アスペルギルス・オリゼ由来FADGDHの酵素特性が報告されている。
FADGDHを利用したグルコースセンサは、グルコースを酸化してD-グルコノ-δ-ラクトンに変換する過程で生じる電子がメディエーターを介して電極に流れることで測定される。FADGDHと組み合わせて使用されるメディエーターとしては、フェリシアン化カリウムが一般的である。しかしながら、フェリシアン化カリウムは血液中の還元物質の影響を受けやすく、加えて環境の水分の影響で容易に還元型に変化しやすいという問題がある。
より安定なメディエーターとしては、例えばルテニウム錯体が挙げられる。ルテニウム錯体は保存安定性が優れている反面、アスペルギルス・オリゼ由来FADGDHはルテニウム錯体に電子を伝達することができないことが問題であった。この問題を克服するために、特許文献3にはアスペルギルス・オリゼ由来FADGDHとルテニウム錯体にフェナジンメトサルフェート(以下、PMSとも表す。)を加えたグルコースセンサが報告されている。このセンサではグルコースとFADGDHの酸化反応で得られた電子を、PMSを介してルテニウム錯体に伝達することができるので、電流応答値を測定することでグルコース濃度を定量することができる。しかしながら、PMSは環境中の光の影響や水分によって還元しやすい性質があるので、依然として保存安定性に課題がある。
一方、酵素の改良も報告されている。非特許文献1には大腸菌発現系で作製したアスペルギルス・フラバス(Aspergillus flavus)由来FADGDHの特定のアミノ酸に変異を導入することで、ルテニウム錯体に直接電子を伝達することができるグルコースデヒドロゲナーゼ変異体が報告されている。この方法では、403位のヒスチジンをアスパラギン酸に置換したグルコースデヒドロゲナーゼ変異体とルテニウム錯体を用いたグルコースセンサが報告されている。
特許文献4、アスペルギルス属由来FADGDHよりも基質特異性の優れたムコール属由来FADGDHが報告されているが、当該FADGDHもルテニウム錯体に対する反応性は低く、実用化には至っていない。
Bioelectrochemistry,2018,Vol.123:p62-69
本発明は、かかる従来技術の現状に鑑み創作されたものである。その目的の1つは、メディエーターに対する反応性が改良されたFADGDHを提供することにある。
上記課題を解決するために、本発明者らは、鋭意検討を重ねた結果、FADGDHの特定のアミノ酸残基を他のアミノ酸に置換することでルテニウム錯体に対する反応性が大幅に改善することを見出した。本発明は係る知見に基づき、更なる研究と改良を重ねた結果完成したものであり、代表的な本発明は、以下の通りである。
項1.
配列番号1と70%以上の同一性を有するアミノ酸配列を有し、配列番号1の322位に相当するアミノ酸残基がトレオニン、セリン、アスパラギンまたはグルタミンである、フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項2.
配列番号1と80%以上の同一性を有するアミノ酸配列を有する、項1に記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項3.
配列番号1と90%以上の同一性を有するアミノ酸配列を有する、項1または2に記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項4.
配列番号1と95%以上の同一性を有するアミノ酸配列を有する、項1~3のいずれかに記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項5.
配列番号1の322位に相当する位置以外の位置において、1若しくは数個のアミノ酸が欠失、置換、付加若しくは挿入されている、項1~4のいずれかに記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項6.
項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼをコードするDNA。
項7.
項6に記載のDNAを含むベクター。
項8.
項7に記載のベクターで形質転換された形質転換体。
項9.
形質転換体がクリプトコッカス属の微生物である、項8に記載の形質転換体。
項10.
項8または9に記載の形質転換体を培養し、培養液中からグルコースデヒドロゲナーゼ活性を有するタンパク質を採取する工程を含む、変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの製造方法。
項11.
項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを使用するグルコース測定方法。
項12.
項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを含むグルコースセンサ。
項13.
メディエーターとして、ルテニウム化合物を含む、項12に記載のグルコースセンサ。
項14.
ルテニウム化合物が、塩化ヘキサアミンルテニウム(III)、(ビピリジン)ルテニウム(II)、(4,4’-ジメチル-2,2’-ビピリジン)ルテニウム(II)、(4,4’-ジフェニル-2,2’-ビピリジン)ルテニウム(II)、(4,4’ -ジアミノ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジヒドロキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジカルボキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジブロモ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジメチル-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジフェニル-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジアミノ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジヒドロキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジカルボキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、および、(5,5’-ジブロモ-2,2’-ビピリジン)ルテニウム(II)からなる群から選択された少なくとも一種である、項12又は13に記載のグルコースセンサ。
配列番号1と70%以上の同一性を有するアミノ酸配列を有し、配列番号1の322位に相当するアミノ酸残基がトレオニン、セリン、アスパラギンまたはグルタミンである、フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項2.
配列番号1と80%以上の同一性を有するアミノ酸配列を有する、項1に記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項3.
配列番号1と90%以上の同一性を有するアミノ酸配列を有する、項1または2に記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項4.
配列番号1と95%以上の同一性を有するアミノ酸配列を有する、項1~3のいずれかに記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項5.
配列番号1の322位に相当する位置以外の位置において、1若しくは数個のアミノ酸が欠失、置換、付加若しくは挿入されている、項1~4のいずれかに記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
項6.
項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼをコードするDNA。
項7.
項6に記載のDNAを含むベクター。
項8.
項7に記載のベクターで形質転換された形質転換体。
項9.
形質転換体がクリプトコッカス属の微生物である、項8に記載の形質転換体。
項10.
項8または9に記載の形質転換体を培養し、培養液中からグルコースデヒドロゲナーゼ活性を有するタンパク質を採取する工程を含む、変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの製造方法。
項11.
項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを使用するグルコース測定方法。
項12.
項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを含むグルコースセンサ。
項13.
メディエーターとして、ルテニウム化合物を含む、項12に記載のグルコースセンサ。
項14.
ルテニウム化合物が、塩化ヘキサアミンルテニウム(III)、(ビピリジン)ルテニウム(II)、(4,4’-ジメチル-2,2’-ビピリジン)ルテニウム(II)、(4,4’-ジフェニル-2,2’-ビピリジン)ルテニウム(II)、(4,4’ -ジアミノ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジヒドロキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジカルボキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジブロモ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジメチル-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジフェニル-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジアミノ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジヒドロキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジカルボキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、および、(5,5’-ジブロモ-2,2’-ビピリジン)ルテニウム(II)からなる群から選択された少なくとも一種である、項12又は13に記載のグルコースセンサ。
ルテニウム錯体に対する反応性に優れたFADGDHが提供される。ルテニウム錯体はFADGDHを利用したグルコースセンサのメディエーターとして機能するため、そのようなセンサの提供が可能となる。
以下に、本発明の態様について、詳細に説明する。
FADGDHはフラビンアデニンジヌクレオチド(以下、FADとも記載する。)を補酵素として使用し、グルコースを選択的に酸化する酵素である。
本発明において、アミノ酸配列はアルファベット1文字または3文字で表記する。また、アミノ酸の変異の位置については次のように表記する。例えば、「H322T」は322位のヒスチジン(His)がトレオニン(Thr)に置換することを意味する。なお、配列番号1におけるアミノ酸残基の番号はN末端のメチオニン(Met)を1として番号付けされている。
一実施形態において、FADGDHは、配列番号1と70%以上の同一性を有するアミノ酸配列を有し、配列番号1の322位に相当するアミノ酸残基が、トレオニン、セリン、アスパラギン、グルタミン、グリシン、アラニン、バリン、ロイシン、イソロイシン、システイン、メチオニン、プロリン、フェニルアラニン、チロシン、トリプトファン、アスパラギン酸、またはグルタミン酸であることが好ましい。配列番号1の322位に相当するアミノ酸残基が、特定のアミノ酸残基であることにより、ルテニウム化合物がFADGDHのメディエーターとして機能する活性部位近傍にアクセスし易くなり、FADGDHとルテニウム化合物との反応性が向上すると考えられる。
配列番号1は、Mucor hiemalis由来の野生型FADGDHのアミノ酸配列である。一実施形態において、FADGDHのアミノ酸配列は、配列番号1のアミノ酸配列に対して、60%以上、70%以上、80%以上、85%以上、90%以上、91%以上、92%以上、93%以上、94%以上、95%以上、96%以上、97%以上、98%以上、または99%以上の同一性を有することが好ましい。
他の実施形態において、FADGDHは、配列番号20または21と70%以上の同一性を有するアミノ酸配列を有し、配列番号1の322位に相当するアミノ酸残基がトレオニン、セリン、アスパラギンまたはグルタミンであることが好ましい。配列番号20または21との同一性は、60%以上、70%以上、80%以上、85%以上、90%以上、91%以上、92%以上、93%以上、94%以上、95%以上、96%以上、97%以上、98%以上、または99%以上であることが好ましい。配列番号20は、Circinella simplex由来の野生型FADGDHのアミノ酸配列であり、配列番号21は、Mucor prainii由来の野生型FADGDHのアミノ酸配列である。
本発明において、アミノ酸配列の同一性は、GENETYXソフトで比較した値を意味する。また、あるアミノ酸配列における配列番号1の322位に相当する位置は、GENETYXソフト(GENETYX CORPORATION)で配列の一次構造(例えばアラインメント)を比較したとき、配列番号1の322位と対応する位置をもって相当する判断する。必要に応じて、さらに立体構造の知見などを参照しても良い。なお、本発明においては、GENETYXソフトとしては、GENETYX WIN Version 6.1を使用した。
一実施形態において、FADGDHは、グルコースデヒドロゲナーゼ活性を有する範囲で、配列番号1の322位に相当する位置以外の位置において、1若しくは数個のアミノ酸が欠失、置換、付加若しくは挿入されていてもよい。「数個」とは、例えば、50個以下、45個以下、30個以下、25個以下、20個以下、15個以下、10個以下、5個以下、3個以下、又は2個以下である。
1又は数個のアミノ酸残基が置換されている場合、置換の種類は、特に制限されないが、ポリペプチドの高次構造、表現形又は特性に顕著な負の影響を与えないという観点から保存的アミノ酸置換が好ましい。「保存的アミノ酸置換」とは、あるアミノ酸残基を、同様の性質の側鎖を有するアミノ酸残基に置換することをいう。アミノ酸残基はその側鎖によって塩基性側鎖(例えば、リシン、アルギニン、ヒスチジン)、酸性側鎖(例えば、アスパラギン酸、グルタミン酸)、非荷電極性側鎖(例えば、アスパラギン、グルタミン、セリン、スレオニン、チロシン、システイン)、非極性側鎖(例えば、グリシン、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン)、分岐側鎖(例えば、スレオニン、バリン、イソロイシン)、芳香族側鎖(例えば、チロシン、フェニルアラニン、トリプトファン、ヒスチジン)のように、同様の性質を有するファミリーに分類することができる。よって、アミノ酸置換は、置換前のアミノ酸残基と同一の上記カテゴリーに属する他のアミノ酸残基間で置換されることが好ましい。
一実施形態において、FADGDH活性を維持するために、FADGDHのアミノ酸配列は、FADGDHにおいて保存性の高いアミノ酸残基及び領域を保持していることが好ましい。保存性の高いアミノ酸残基及び領域は、例えば、複数種FADGDHのアミノ酸配列を比較することで特定することができる。
一実施形態において、FADGDHは、ムコール属またはシルシネラ属の微生物に由来することが好ましく、ムコール属微生物に由来することが好ましい。菌種については、特に限定されないが、ムコール・ヒエマリス(Mucor hiemalis)、ムコール・プライニ(Mucor prainii)、シルシネラ・シンプレックス(Circinella simplex)などが挙げられる。
一実施形態において、FADGDHは、クリプトコッカス属の微生物を宿主として生産されたFADGDHであることが好ましい。一実施形態において、FADGDHは、その糖鎖を含む分子量(SDS-PAGEで測定)は、約80~110kDa、好ましくは約85~105kDa、より好ましくは約90~100kDaである。宿主として用いられるクリプトコッカス属の微生物は特に限定されないが、クリプトコッカスsp.S-2(Cryptococcus sp.S-2)(受託番号FERM BP-10961)またはその変異体が好ましい。
FADGDHの製造方法は、特に限定されないが、以下に示すような手順で製造することが可能である。
FADGDHを構成するアミノ酸配列を改変する方法としては、通常行われている遺伝情報を改変する手法が用いられる。すなわち、タンパク質の遺伝情報を有するDNAの特定の塩基を変換することにより、或いは特定の塩基を挿入または欠失させることにより、改変タンパク質の遺伝情報を有するDNAが作製される。DNA中の塩基配列を変換する具体的な方法としては、例えば市販のキット(Transformer Mutagenesis Kit;Clonetech社,EXOIII/Mung Bean Deletion Kit;Stratagene製,Quick Change Site Directed Mutagenesis Kit;Stratagene製など)の使用、或いはポリメラーゼ連鎖反応(PCR)の利用が挙げられる。
一実施形態において、FADGDHをコードするDNAは、配列番号2に記載の塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グルコースデヒドロゲナーゼ活性を有するタンパク質をコードするDNAである。ここでストリンジェントな条件とは、同一性が高い核酸同士、例えば完全にマッチしたハイブリッドのTmから該Tmより15℃、好ましくは10℃低い温度までの範囲の温度でハイブリダイズする条件をいう。具体的には、例えば一般的なハイブリダイゼーション用緩衝液(5×SSC(0.75M NaCl,0.075Mクエン酸-Na;pH7.0),1%ブロッキング試薬,0.02% SDS,0.1% N-ラウリルサルコシン)中で、68℃、20時間の条件でハイブリダイズする条件をいう。本発明において、FADGDHをコードするDNAは、配列番号2の塩基配列と60%以上であり、好ましくは65%以上、より好ましくは70%以上、さらに好ましくは80%以上、さらに好ましくは90%以上、さらに好ましくは95%以上の同一性を有することが好ましい。配列番号2は、Mucor hiemalis由来の野生型FADGDHをコードする塩基配列である。
塩基配列の同一性は、市販の又は電気通信回線(インターネット)を通じて利用可能な解析ツールを用いて算出することができ、例えば、FASTA、BLAST、PSI-BLAST、SSEARCH等のソフトウェアを用いて計算される。具体的には、BLAST検索に一般的に用いられる主な初期条件は、以下の通りである。即ち、Advanced BLAST 2.1において、プログラムにblastnを用い、各種パラメータはデフォルト値に設定して検索を行うことにより、塩基配列の相同性の値(%)を算出することができる。
一実施形態において、FAGDHをコードするDNAは、単離された状態で存在するDNAであることが好ましい。ここで「単離されたDNA」とは、天然状態において共存するその他の核酸やタンパク質等の成分から分離された状態であることをいう。但し、天然状態において隣接する核酸配列(例えばプロモーター領域の配列やターミネーター配列など)など一部の他の核酸成分を含んでいてもよい。cDNA分子など遺伝子工学的手法によって調製されるDNAの場合の「単離された」状態は、好ましくは、細胞成分や培養液などを実質的に含まない。同様に、化学合成によって調製されるDNAの場合の「単離されたDNA」は、好ましくは、dNTPなどの前駆体(原材料)や合成過程で使用される化学物質等を実質的に含まないこと。一実施形態において、DNAはcDNAであることが好ましい。
DNAは、配列番号2の塩基配列に基づいて、化学的なDNAの合成法(例えば、フォスフォアミダイト法)や遺伝子工学的手法を用いて容易に取得することができる。
本発明の一実施形態は、上記のDNAを含むベクター、該ベクターにより形質転換された形質転換体、該形質転換体を培養し、GDH活性を有するタンパク質を採取する方法、GDH活性を有するタンパク質を生産する方法である。以下、これらについて説明する。
ベクターは、上記FADGDHをコードするDNAを発現可能な様式で含むことが好ましい。ベクターの種類は、宿主細胞の種類に応じて適宜選択することができる。例えば、プラスミドベクター、コスミドベクター、ファージベクター、ウイルスベクター(アデノウイルスベクター、レトロウイルスベクター、ヘルペスウイルスベクター等)等を挙げることができる。例えば、宿主と指定大腸菌を用いる場合のベクターとしては、pBR322、pUC19、pGEM-T、pCR-Blunt、pTA2、pETなどが挙げられる。
一実施形態において、ベクターは、栄養要求性マーカー、薬剤耐性マーカー、発現プロモーターDNA配列、及び/又は発現ターミネーター配列を含むことが好ましく、クリプトコッカスsp.S-2由来のorotate phosphoribosyl transferase遺伝子、キシラナーゼプロモーター、及びキシラナーゼターミネーターを含むことが好ましい。宿主に組換え発現ベクターを導入する方法としては、特に限定するものではないが、エレクトロポレーションなどの方法が挙げられる。
一実施形態において、タンパク質の発現効率及び発現成功率を高めるために、タンパク質(FADGDHのアミノ酸配列)のN末満に分泌シグナルペプチドを結合させた状態で発現させることが好ましい。一般的に、分泌タンパク質のN末端には、分泌シグナルペプチドが存在しており、この既存の分泌シグナルペプチドを高効率な分泌シグナルペプチドと置換することで、これら分泌タンパク質の発現効率及び発現成功率を上げることができることが報告されている。
使用できる分泌シグナルペプチドとしては、例えば、クリプトコッカスsp.S-2が生産する酸性キシラナーゼに由来する分泌シグナルペプチド配列であり、配列番号3で示されるアミノ酸配列は分泌シグナルペプチド配列を開始コドンから17アミノ酸までとした配列である。一実施形態において、ベクターは、FADGDHのDNAに加え、シグナルペプチドをコードする塩基配列を含むことが好ましい。
形質転換体は、FADGDHをコードするDNAで形質転換されていることが好ましい。形質転換に使用する宿主細胞は、特に制限されず、原核細胞及び真核細胞のいずれでもよく、目的に応じて適宜選択できる。発現ベクターと宿主微生物の組み合わせは、特に限定されないが、例えば、遺伝子を組み込む宿主由来の栄養要求性マーカー遺伝子または薬剤耐性マーカー遺伝子と、遺伝子を組み込む宿主由来の発現プロモーターDNA配列と、遺伝子を組み込む宿主由来のターミネーターDNA配列とを含む発現ベクターと、栄養要求性変異宿主または薬剤感受性宿主との組み合わせが挙げられる。好ましくは、クリプトコッカスsp.S-2由来のorotate phosphoribosyl transferase遺伝子と、キシラナーゼプロモーターDNA配列と、キシラナーゼターミネーターDNA配列とを含む発現ベクターと、クリプトコッカスsp.S-2との組み合わせが挙げられる。
形質転換体の培養形態は、宿主の栄養生理的性質を考慮して適宜選択すればよく、多くの場合は液体培養で行うが、工業的には通気攪拌培養を行うのが有利である。なお、培養に先立って、高FADGDH生産細胞株を予め選抜しておくことが有利である。
培養に用いる窒素源は、特定のアミノ酸成分に欠失があるなど特殊なN源を除いて、宿主微生物が利用可能な窒素化合物であれば使用できる。これらは主として有機窒素源であり、例えばペプトン、肉エキス、酵母エキス、カゼイン加水分解物、大豆粕アルカリ分解物などが使用される。特に、酵母エキスや大豆蛋白質が好ましいが、これに限定されるものではなく、カゼインポリペプトン、発酵麹エキス、麦芽抽出物などを用いることもできる。
その他の栄養源としては、微生物の培養に通常用いられるものが広く使用される。炭素源としては、資化可能な炭素化合物であればよく、例えば、グルコース、シュークロース、ラクトース、マルトース、キシロース、糖蜜、ピルビン酸などが使用される。その他、リン酸塩、炭酸塩、硫酸塩、マグネシウム、カルシウム、カリウム、鉄、マンガン、亜鉛などの塩類、特定のアミノ酸、特定のビタミンなどが必要に応じて使用される。
培養温度は、菌が発育してFADGDHを生産する範囲で適宜設計できる。例えば、クリプトコッカスsp.S-2の場合、通常は20~25℃程度である。培養時間は、条件によって多少異なるが、FADGDHが最高収量に達する時期を見計らって適当な時期に培養を終了することが好ましく、例えば60~120時間程度である。培地pHは、菌が発育しFADGDHを生産する範囲で適宜設計でき、通常はpH3.0~9.0程度である。
培養物中のFADGDHを生産する菌体を含む培養液をそのまま採取し、利用することもできるが、一般には、常法に従って、FADGDHが培養液中に存在する場合はろ過、遠心分離などにより、FADGDH含有溶液と微生物菌体とを分離した後に利用される。FADGDHが菌体内に存在する場合には、得られた培養物からろ過または遠心分離などの手段により菌体を採取し、次いで、この菌体を機械的方法またはリゾチームなどの酵素的方法で破壊し、必要に応じて、EDTA等のキレート剤及びポリエチレングリコールモノ-p-イソオクチルフェニルエーテル(Triton-100)、ポリオキシエチレンソルビタンモノラウレート(Tween-20)等の界面活性剤を添加してFADGDHを可溶化し、水溶液として分離採取する。
上記のようにして得られたFADGDH含有溶液を、例えば減圧濃縮、膜濃縮、さらに硫酸アンモニウム、硫酸ナトリウムなどの塩析処理、あるいは親水性有機溶媒、例えばメタノール、エタノール、アセトンなどによる分別沈殿法により沈殿せしめればよい。また、加熱処理や等電点処理も有効な精製手段である。その後、吸着剤あるいはゲルろ過剤などによるゲルろ過、吸着クロマトグラフィー、イオン交換クロマトグラフィー、アフィニティクロマトグラフィーを行うことにより、精製されたFADGDHを得ることができる。
例えば、セファデックス(Sephadex)ゲル(GEヘルスケア・バイオサイエンス社製)などによるゲルろ過、DEAEセファロースCL-6B(GEヘルスケア・バイオサイエンス社製)、オクチルセファロースCL-6B(GEヘルスケア・バイオサイエンス社製)等のカラムクロマトグラフィーにより分離、精製し、精製酵素標品を得ることができる。該精製酵素標品は、電気泳動(SDS-PAGE)的に単一のバンドを示す程度に純化されていることが好ましい。
これらは、例えば、以下の文献に従って進めることができる。
(a)西村善文,大野茂男 監修;タンパク質実験プロトコール 第1巻 機能解析編,第2巻 構造解析編(秀潤社)p50-52
(b)竹縄忠臣 編集;タンパク質ハンドブック p22-47 また、以下に例示する方法によって進めることもできる。
(a)西村善文,大野茂男 監修;タンパク質実験プロトコール 第1巻 機能解析編,第2巻 構造解析編(秀潤社)p50-52
(b)竹縄忠臣 編集;タンパク質ハンドブック p22-47 また、以下に例示する方法によって進めることもできる。
本発明において、DCPIPを用いたGDHの活性測定は以下の条件で行う。
[試験例]
<試薬>
50mM PIPES緩衝液pH6.5(0.1% TritonX-100を含む)
2.0mM 2,6-ジクロロフェノールインドフェノール(DCPIP)溶液
1M D-グルコース溶液
<試薬>
50mM PIPES緩衝液pH6.5(0.1% TritonX-100を含む)
2.0mM 2,6-ジクロロフェノールインドフェノール(DCPIP)溶液
1M D-グルコース溶液
上記PIPES緩衝液20.5ml、DCPIP溶液3.0ml、D―グルコース溶液5.9mlを混合して、反応試薬とする。
<測定条件>
反応試薬3mlを37℃で5分間予備加温し、FADGDH溶液0.1mlを添加し、ゆるやかに混和後、水を対照として、37℃に制御された分光光度計で600nmの吸光度変化を5分記録し、直線部分から1分間あたりの吸光度変化(ΔODTEST)を測定する。盲検としては、FADGDH溶液の代わりにFADGDHを溶解する溶媒を試薬混液に加えて、同様に1分間あたりの吸光度変化(ΔODBLANK)を測定する。これらの値から、次の式(1)に従ってGDH活性を求める。ここでGDH活性における1単位(U)とは、濃度200mMのD-グルコース存在下で、1分間に1マイクロモルのDCPIPを還元する酵素量として定義している。
反応試薬3mlを37℃で5分間予備加温し、FADGDH溶液0.1mlを添加し、ゆるやかに混和後、水を対照として、37℃に制御された分光光度計で600nmの吸光度変化を5分記録し、直線部分から1分間あたりの吸光度変化(ΔODTEST)を測定する。盲検としては、FADGDH溶液の代わりにFADGDHを溶解する溶媒を試薬混液に加えて、同様に1分間あたりの吸光度変化(ΔODBLANK)を測定する。これらの値から、次の式(1)に従ってGDH活性を求める。ここでGDH活性における1単位(U)とは、濃度200mMのD-グルコース存在下で、1分間に1マイクロモルのDCPIPを還元する酵素量として定義している。

なお、式中の3.0は反応試薬+酵素溶液の液量(ml)、16.3は本活性測定条件におけるミリモル分子吸光係数(cm2/マイクロモル)、0.1は酵素溶液の液量(ml)、1.0はセルの光路長(cm)を示す。
本発明において、塩化ヘキサアミンルテニウム(III)/臭化3-(4,5-ジメチル-2-チアゾリル)-2,5-ジフェニルテトラゾリウム(以下、Ru/MTTとも表す。)を用いたGDHの活性測定は以下の条件で行う。
[試験例]
<試薬>
50mM リン酸緩衝液pH6.5(0.1% TritonX-100を含む)
30mM 塩化ヘキサアミンルテニウム(III)溶液
10mM 臭化3-(4,5-ジメチル-2-チアゾリル)-2,5-ジフェニルテトラゾリウム溶液
1M D-グルコース溶液
上記リン酸緩衝液10.5ml、塩化ヘキサアミンルテニウム(III)溶液10.0ml、臭化3-(4,5-ジメチル-2-チアゾリル)-2,5-ジフェニルテトラゾリウム溶液 3ml
D―グルコース溶液5.9ml
を混合して、反応試薬とする。
<試薬>
50mM リン酸緩衝液pH6.5(0.1% TritonX-100を含む)
30mM 塩化ヘキサアミンルテニウム(III)溶液
10mM 臭化3-(4,5-ジメチル-2-チアゾリル)-2,5-ジフェニルテトラゾリウム溶液
1M D-グルコース溶液
上記リン酸緩衝液10.5ml、塩化ヘキサアミンルテニウム(III)溶液10.0ml、臭化3-(4,5-ジメチル-2-チアゾリル)-2,5-ジフェニルテトラゾリウム溶液 3ml
D―グルコース溶液5.9ml
を混合して、反応試薬とする。
<測定条件>
反応試薬3mlを、37℃で5分間予備加温する。FADGDH溶液0.1mlを添加し、ゆるやかに混和後、水を対照に37℃に制御された分光光度計で、565nmの吸光度変化を5分記録し、直線部分から1分間あたりの吸光度変化(ΔODTEST)を測定する。盲検はFADGDH溶液の代わりに、FADGDHを溶解する溶媒を試薬混液に加えて、同様に1分間あたりの吸光度変化(ΔODBLANK)を測定する。これらの値から、次の式(2)に従ってGDH活性を求める。ここで、GDH活性における1単位(U)とは、濃度200mMのD-グルコース存在下で、1分間に1マイクロモルの臭化3-(4,5-ジメチル-2-チアゾリル)-2,5-ジフェニルテトラゾリウムを還元する酵素量として定義している。
反応試薬3mlを、37℃で5分間予備加温する。FADGDH溶液0.1mlを添加し、ゆるやかに混和後、水を対照に37℃に制御された分光光度計で、565nmの吸光度変化を5分記録し、直線部分から1分間あたりの吸光度変化(ΔODTEST)を測定する。盲検はFADGDH溶液の代わりに、FADGDHを溶解する溶媒を試薬混液に加えて、同様に1分間あたりの吸光度変化(ΔODBLANK)を測定する。これらの値から、次の式(2)に従ってGDH活性を求める。ここで、GDH活性における1単位(U)とは、濃度200mMのD-グルコース存在下で、1分間に1マイクロモルの臭化3-(4,5-ジメチル-2-チアゾリル)-2,5-ジフェニルテトラゾリウムを還元する酵素量として定義している。

なお、式中の3.0は反応試薬+酵素溶液の液量(ml)、20.0は本活性測定条件におけるミリモル分子吸光係数(cm2/マイクロモル)、0.1は酵素溶液の液量(ml)、1.0はセルの光路長(cm)を示す。
グルコースセンサ
グルコースセンサは、上述のFADGDHを含むことが好ましい。グルコースセンサが備える電極としては、カーボン電極、金電極、白金電極などを用いることができる。参照極と作用極上にキャピラリー構造を形成し、キャピラリー内に酵素とメディエーターを充填させるのが一般的である。電極上にFADGDHを固定化する方法としては、架橋試薬を用いる方法、高分子マトリックス中に封入する方法、透析する方法、光架橋性ポリマー、導電性ポリマー、酸化還元ポリマーなどを用いる方法があり、フェロセンあるいはその誘導体に代表される電子メディエーターとともにポリマー中に固定あるいは電極上に吸着固定してもよく、またこれらを組み合わせて用いてもよい。該ポリマーとしては、親水性ポリマーが好ましく、例えばカルボキシメチルセルロース、ポリビニルアルコール、ポリスチレンスルホン酸、コンドロイチン硫酸、ヒアルロン酸、アラビアガム、カラギーナンなどが挙げられる。典型的には、グルタルアルデヒドを用いてFADGDHをカーボン電極上に固定化した後、アミン基を有する試薬で処理してグルタルアルデヒドをブロッキングする。
グルコースセンサは、上述のFADGDHを含むことが好ましい。グルコースセンサが備える電極としては、カーボン電極、金電極、白金電極などを用いることができる。参照極と作用極上にキャピラリー構造を形成し、キャピラリー内に酵素とメディエーターを充填させるのが一般的である。電極上にFADGDHを固定化する方法としては、架橋試薬を用いる方法、高分子マトリックス中に封入する方法、透析する方法、光架橋性ポリマー、導電性ポリマー、酸化還元ポリマーなどを用いる方法があり、フェロセンあるいはその誘導体に代表される電子メディエーターとともにポリマー中に固定あるいは電極上に吸着固定してもよく、またこれらを組み合わせて用いてもよい。該ポリマーとしては、親水性ポリマーが好ましく、例えばカルボキシメチルセルロース、ポリビニルアルコール、ポリスチレンスルホン酸、コンドロイチン硫酸、ヒアルロン酸、アラビアガム、カラギーナンなどが挙げられる。典型的には、グルタルアルデヒドを用いてFADGDHをカーボン電極上に固定化した後、アミン基を有する試薬で処理してグルタルアルデヒドをブロッキングする。
グルコース濃度の測定は、以下のようにして行うことができる。恒温セルに緩衝液を入れ、一定温度に維持する。メディエーターとしては、FADGDHの補酵素であるFADから電子を受け取り、発色物質や電極に電子を供与しうるものが挙げられる。例えばフェリシアン化物塩、フェナジンエトサルフェート、フェナジンメトサルフェート、フェニレンジアミン、N,N,N’,N’-テトラメチルフェニレンジアミン、1-メトキシ-フェナジンメトサルフェート、2,6-ジクロロフェノールインドフェノール、2,5-ジメチル-1,4-ベンゾキノン、2,6-ジメチル-1,4-ベンゾキノン、2,5-ジクロロ-1,4-ベンゾキノン、ニトロソアニリン、フェロセン誘導体、オスミウム化合物、ルテニウム化合物等が例示されるが、中でも、ルテニウム化合物が好ましい。
上記のルテニウム化合物としては、塩化ヘキサアミンルテニウム(III)、(ビピリジン)ルテニウム(II)、(4,4’-ジメチル-2,2’-ビピリジン)ルテニウム(II)、(4,4’-ジフェニル-2,2’-ビピリジン)ルテニウム(II)、(4,4’-ジアミノ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジヒドロキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジカルボキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジブロモ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジメチル-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジフェニル-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジアミノ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジヒドロキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジカルボキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジブロモ-2,2’-ビピリジン)ルテニウム(II)などが例示される。一実施形態において、塩化ヘキサアミンルテニウム(III)が好ましい。
グルコースセンサは、作用電極がFADGDHを固定化した電極であり、対極(例えば白金電極)および参照電極(例えばAg/AgCl電極)を備えることが好ましい。カーボン電極に一定の電圧を印加して、電流が定常になった後、血液、尿などのグルコースを含む試料を加えて電流の増加を測定するアンペロメトリーによる方法でグルコース濃度を測定することが好ましい。標準濃度のグルコース溶液により作製したキャリブレーションカーブに従い、試料中のグルコース濃度を計算することができる。
以下に、本発明を実施例により具体的に説明する。実施例により、本発明が特に限定されるものではない。
実施例1:大腸菌発現用ベクターの作製
ムコール・ヒエマリス由来FADGDHのアミノ酸配列である配列番号1のアミノ酸配列のN末端にメチオニンを付加したアミノ酸配列をコードするDNA配列(配列番号2)を組み換えプラスミドpBluescript SK(+)のマルチクローニング部位に挿入した。これで市販の大腸菌コンピテントセル(E.coli DH5α;TOYOBO製)を形質転換し、アンピシリンを含んだLB寒天培地(1.0%ポリペプトン、0.5%酵母エキス、1.0%NaCl、1.5%寒天;pH7.3)に塗布し、30℃で一晩培養した。寒天培地上にコロニーを形成した形質転換体を、アンピシリン(50mg/ml;ナカライテスク製)を含んだLB液体培地(1%ポリペプトン、0.5%酵母エキス、1.0%NaCl;pH6.5)に接種し、30℃で一晩振とう培養した。得られた菌体からMagExtractor(登録商標)-Plasmid-(東洋紡製)を用いてプラスミドをミニプレップ抽出・精製し、pBluescript SK(+)のマルチクローニング部位に配列番号2の塩基配列が挿入されたプラスミドpMhGDHプラスミドを得た。
ムコール・ヒエマリス由来FADGDHのアミノ酸配列である配列番号1のアミノ酸配列のN末端にメチオニンを付加したアミノ酸配列をコードするDNA配列(配列番号2)を組み換えプラスミドpBluescript SK(+)のマルチクローニング部位に挿入した。これで市販の大腸菌コンピテントセル(E.coli DH5α;TOYOBO製)を形質転換し、アンピシリンを含んだLB寒天培地(1.0%ポリペプトン、0.5%酵母エキス、1.0%NaCl、1.5%寒天;pH7.3)に塗布し、30℃で一晩培養した。寒天培地上にコロニーを形成した形質転換体を、アンピシリン(50mg/ml;ナカライテスク製)を含んだLB液体培地(1%ポリペプトン、0.5%酵母エキス、1.0%NaCl;pH6.5)に接種し、30℃で一晩振とう培養した。得られた菌体からMagExtractor(登録商標)-Plasmid-(東洋紡製)を用いてプラスミドをミニプレップ抽出・精製し、pBluescript SK(+)のマルチクローニング部位に配列番号2の塩基配列が挿入されたプラスミドpMhGDHプラスミドを得た。
実施例2:変異型FADGHD発現プラスミドの調製
実施例1で得たpMhGDHプラスミドを鋳型として、58位のグルタミンをアスパラギン酸に置換するよう設計した、配列番号4の合成オリゴヌクレオチドとそれに相補的な配列番号5の合成オリゴヌクレオチドを使用して、QuickChange(登録商標)Site-Directed Mutagenesis Kit(STRATAGENE製)を用いてPCRを行った。続いて、PCR産物で市販の大腸菌コンピテントセル(E.coli DH5α;TOYOBO製)を形質転換し、アンピシリンを含んだLB寒天培地で、37℃、16時間培養した。その後、シングルコロニーを、アンピシリンを含んだLB液体培地に接種し、30℃で一晩振とう培養した。その後、1mlの培養液を接種し、常法によりプラスミドを抽出した。抽出したプラスミドの置換された部位を、DNAシークエンサー(ABI PRISM(登録商標)3700DNA Analyzer;Perkin-Elmer製)を用いて特定した。配列番号1の58位のグルタミンをアスパラギン酸に置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-Q58Dを得た。同様の手法で配列番号6と配列番号7の合成オリゴヌクレオチドを用いて配列番号1の79位のアルギニンをグルタミンに置換したプラスミドpMhGDH-R79Qを得た。配列番号8と配列番号9の合成オリゴヌクレオチドを用いて配列番号1の320位のヒスチジンをグルタミンに置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-H320Qを得た。配列番号10と配列番号11の合成オリゴヌクレオチドを用いて配列番号1の322位のヒスチジンをトレオニンに置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-H322Tを得た。配列番号12と配列番号13の合成オリゴヌクレオチドを用いて配列番号1の326位のバリンをアルギニンに置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-V326Rを得た。配列番号16と配列番号17の合成オリゴヌクレオチドを用いて配列番号1の438位のトレオニンをアスパラギン酸に置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-T438Dを得た。配列番号18と配列番号19の合成オリゴヌクレオチドを用いて配列番号1の458位のアスパラギンをアスパラギン酸に置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-N458Dを得た。
実施例1で得たpMhGDHプラスミドを鋳型として、58位のグルタミンをアスパラギン酸に置換するよう設計した、配列番号4の合成オリゴヌクレオチドとそれに相補的な配列番号5の合成オリゴヌクレオチドを使用して、QuickChange(登録商標)Site-Directed Mutagenesis Kit(STRATAGENE製)を用いてPCRを行った。続いて、PCR産物で市販の大腸菌コンピテントセル(E.coli DH5α;TOYOBO製)を形質転換し、アンピシリンを含んだLB寒天培地で、37℃、16時間培養した。その後、シングルコロニーを、アンピシリンを含んだLB液体培地に接種し、30℃で一晩振とう培養した。その後、1mlの培養液を接種し、常法によりプラスミドを抽出した。抽出したプラスミドの置換された部位を、DNAシークエンサー(ABI PRISM(登録商標)3700DNA Analyzer;Perkin-Elmer製)を用いて特定した。配列番号1の58位のグルタミンをアスパラギン酸に置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-Q58Dを得た。同様の手法で配列番号6と配列番号7の合成オリゴヌクレオチドを用いて配列番号1の79位のアルギニンをグルタミンに置換したプラスミドpMhGDH-R79Qを得た。配列番号8と配列番号9の合成オリゴヌクレオチドを用いて配列番号1の320位のヒスチジンをグルタミンに置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-H320Qを得た。配列番号10と配列番号11の合成オリゴヌクレオチドを用いて配列番号1の322位のヒスチジンをトレオニンに置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-H322Tを得た。配列番号12と配列番号13の合成オリゴヌクレオチドを用いて配列番号1の326位のバリンをアルギニンに置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-V326Rを得た。配列番号16と配列番号17の合成オリゴヌクレオチドを用いて配列番号1の438位のトレオニンをアスパラギン酸に置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-T438Dを得た。配列番号18と配列番号19の合成オリゴヌクレオチドを用いて配列番号1の458位のアスパラギンをアスパラギン酸に置換した変異型FADGDHをコードする塩基配列を含むプラスミドpMhGDH-N458Dを得た。
実施例3:大腸菌発現系を用いた変異型FADGDHを含む粗酵素液の調製
実施例2で取得した7種の変異型FADGDH発現プラスミドで形質転換した大腸菌JM109を、アンピシリンを含んだLB寒天培地で30℃、16時間培養しシングルコロニーを取得した。その後、該形質転換体のシングルコロニーを、アンピシリンを含んだ5mlLB液体培地に接種し、30℃で16時間振とう培養した。その培養液の一部から遠心分離によって得られた菌体を回収し、50mMのリン酸緩衝液(pH6.0)中でガラスビーズを用いて該菌体を破砕することにより粗酵素液を調製した。
実施例2で取得した7種の変異型FADGDH発現プラスミドで形質転換した大腸菌JM109を、アンピシリンを含んだLB寒天培地で30℃、16時間培養しシングルコロニーを取得した。その後、該形質転換体のシングルコロニーを、アンピシリンを含んだ5mlLB液体培地に接種し、30℃で16時間振とう培養した。その培養液の一部から遠心分離によって得られた菌体を回収し、50mMのリン酸緩衝液(pH6.0)中でガラスビーズを用いて該菌体を破砕することにより粗酵素液を調製した。
実施例4:大腸菌発現系を用いた変異体を含む粗酵素液の反応性
実施例3で調製した粗酵素液について、上述したRu/MTTを用いた活性測定法によりGDH活性を測定した。表1にその結果を示す。GDH活性は、野生型(MhGDH)のGDH活性を1.0とした場合、H322T変異体は3.7倍に増加した。一方、他の変異体いずれも1.0未満であった。この結果からH322T変異体はルテニウムに対する反応性が大幅に向上することが判明した。
実施例3で調製した粗酵素液について、上述したRu/MTTを用いた活性測定法によりGDH活性を測定した。表1にその結果を示す。GDH活性は、野生型(MhGDH)のGDH活性を1.0とした場合、H322T変異体は3.7倍に増加した。一方、他の変異体いずれも1.0未満であった。この結果からH322T変異体はルテニウムに対する反応性が大幅に向上することが判明した。

実施例5:クリプトコッカス発現系を用いたH322T変異体の発現
特許第6461793号に記載のpCsUXXs3MhFADGLD(opt)を鋳型として、実施例2と同様の手法で配列番号10と配列番号11の合成オリゴヌクレオチドを用いて322位のヒスチジンをトレオニンに置換したpCsUXXs3MhFADGLD(opt)-H322Tプラスミドを作製した。組換え宿主にはクリプトコッカスsp.S-2. DA25株を使用した。本菌株は、特許第5588578号に記載されており且つ、FERM BP-11482に国際寄託された、クリプトコッカスsp.S-2株の変異体に、クリプトコッカスsp.S-2株に由来するade1遺伝子を導入した菌株である。本菌は、形質転換体の選択のために、ウラシル要求性となっており、pCsUXXs3MhFADGLD(opt)-H322Tプラスミドを形質転換することにより、ウラシル要求性によって形質転換体を選抜することが可能である。
特許第6461793号に記載のpCsUXXs3MhFADGLD(opt)を鋳型として、実施例2と同様の手法で配列番号10と配列番号11の合成オリゴヌクレオチドを用いて322位のヒスチジンをトレオニンに置換したpCsUXXs3MhFADGLD(opt)-H322Tプラスミドを作製した。組換え宿主にはクリプトコッカスsp.S-2. DA25株を使用した。本菌株は、特許第5588578号に記載されており且つ、FERM BP-11482に国際寄託された、クリプトコッカスsp.S-2株の変異体に、クリプトコッカスsp.S-2株に由来するade1遺伝子を導入した菌株である。本菌は、形質転換体の選択のために、ウラシル要求性となっており、pCsUXXs3MhFADGLD(opt)-H322Tプラスミドを形質転換することにより、ウラシル要求性によって形質転換体を選抜することが可能である。
形質転換は、Infect Immun. 1992 Mar;60(3):1101-8.(Varmaら)に記載の方法で実施した。クリプトコッカスsp.S-2. DA-25株を20mlYM培地(酵母エキス0.3%、麦芽エキス0.3%、ポリペプトン0.5%、グルコース1.0%)で25℃、48時間培養した。得られた培養液の吸光度(OD660nm)を測定した。次に、この培養液を200ml液体培地(酵母エキス0.3%、麦芽エキス0.3%、ポリペプトン0.5%、グルコース1.0%)に培養液の吸光度(OD660nm)が0.1Absになるように植菌して25℃で培養し、培養液の吸光度(OD660nm)が約1.0Absになるまで培養を行った。次に、この培養液を遠心分離して菌体を回収し、Wash buffuer(270mMシュークロース、1mM塩化マグネシウム、4mM DTT、10mM Tris-HCl pH7.6)で菌体を2回洗浄し、Electroporation buffer(270mMシュークロース、1mM塩化マグネシウム、10mM Tris-HCl pH7.6)に吸光度OD660=50Absとなるように懸濁した。得られた懸濁液100μlに、予めSbfIによって制限酵素処理し、直鎖化したプラスミド10μg(~5μl)を添加し、エレクトロポレーション用のキュベットに移した後、Gene Pulser Xcell (BIO-RAD社)を用いて通電した。通電条件は、C=25μF; V=0.47kVで行った。通電後の液中に600μl Electroporation buffer(270mMシュークロース、1mM塩化マグネシウム、10mM Tris-HCl pH7.6)を加え、選択プレート上に塗り広げた。選択プレートはYNB-ura寒天培地(0.67%Yeast Nitrogen Base W/O amino acid、0.078% -ura DO supplement、2%グルコース、1%寒天粉末)を用いた。植菌したプレートを25℃で1週間静置培養し、生育コロニーを選抜した。
該形質転換体を、200ml液体培地(2%酵母エキス、5%キシロース)で25℃、600rpm、48時間培養し、培養液上清のGDH活性を確認した。
実施例6:H322T変異体の精製
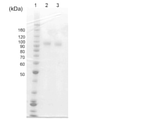
実施例5でGDH活性を有することを確認した培養液上清を濾布で濾過し、濾過液を回収した。濾過液を分画分子量30,000のUF膜(ミリポア製)を用いて濃縮し、濃縮液に連続してリン酸緩衝液(50mM、pH6.0)を添加することによってバッファーを置換した。続いて、濃縮液に硫酸アンモニウムを40%(w/v)徐々に添加して、室温で30分間攪拌した後、ろ過助剤を用いて余分な沈殿を除去した。次に、予め40%(w/v)の硫酸アンモニウムを含む50mMリン酸カリウム緩衝液(pH6.0)で平衡化した200mLのPSセファロースFastFlow(GEヘルスケア製)カラムに濾過液をチャージし、50mMリン酸緩衝液(pH6.0)に段階的に置換してタンパク質を溶出させた。そして、溶出された画分を、分画分子量10,000の中空糸膜(スペクトラムラボラトリーズ製)で濃縮し、濃縮液に連続してリン酸緩衝液(50mM,pH6.0)を添加することによってバッファーを置換した。続いて、50mMリン酸緩衝液(pH6.0)で平衡化したDEAEセファロースFast Flow(GEヘルスケア製)カラム酵素液を通液して、精製酵素を得た。図1に、得られた精製酵素に関するSDS-PAGEの結果を示す。SDS-PAGEの結果、H322T変異体の分子量は、約90-100kDaであり野生型と同等であった。レーン1は分子量マーカー(BenchMarkTM Protein Ladder Marker)であり、レーン2はH322T変異体であり、レーン3は野生型MhGDHである。
実施例5でGDH活性を有することを確認した培養液上清を濾布で濾過し、濾過液を回収した。濾過液を分画分子量30,000のUF膜(ミリポア製)を用いて濃縮し、濃縮液に連続してリン酸緩衝液(50mM、pH6.0)を添加することによってバッファーを置換した。続いて、濃縮液に硫酸アンモニウムを40%(w/v)徐々に添加して、室温で30分間攪拌した後、ろ過助剤を用いて余分な沈殿を除去した。次に、予め40%(w/v)の硫酸アンモニウムを含む50mMリン酸カリウム緩衝液(pH6.0)で平衡化した200mLのPSセファロースFastFlow(GEヘルスケア製)カラムに濾過液をチャージし、50mMリン酸緩衝液(pH6.0)に段階的に置換してタンパク質を溶出させた。そして、溶出された画分を、分画分子量10,000の中空糸膜(スペクトラムラボラトリーズ製)で濃縮し、濃縮液に連続してリン酸緩衝液(50mM,pH6.0)を添加することによってバッファーを置換した。続いて、50mMリン酸緩衝液(pH6.0)で平衡化したDEAEセファロースFast Flow(GEヘルスケア製)カラム酵素液を通液して、精製酵素を得た。図1に、得られた精製酵素に関するSDS-PAGEの結果を示す。SDS-PAGEの結果、H322T変異体の分子量は、約90-100kDaであり野生型と同等であった。レーン1は分子量マーカー(BenchMarkTM Protein Ladder Marker)であり、レーン2はH322T変異体であり、レーン3は野生型MhGDHである。
実施例7:H322T変異体の電気化学式センサにおける応答値の評価
グルコース測定用試薬として、以下の組成からなる溶液(pH7.0)を作製した。
1mM クエン酸ナトリウム(pH7.0)
45mM 塩化ヘキサアミンルテニウム(III)
0.4mg/ml FADGDH
グルコース測定用試薬として、以下の組成からなる溶液(pH7.0)を作製した。
1mM クエン酸ナトリウム(pH7.0)
45mM 塩化ヘキサアミンルテニウム(III)
0.4mg/ml FADGDH
0.5%のCMC(カルボキシメチルセルロース)5μLを、3電極を有するディスポーサブルチップ(DEP-CHIP;サイズ:12.5×4×0.3mm、カーボン:EP-PP、バイオデバイステクノロジーズ社製)の作用極・対極・参照極上に滴下し、50℃、10分の加温処理を行って乾燥させた。続いて、上記組成液5μLを、CMCを固定化した箇所に滴下し、50℃、10分の加温処理を行って、センサチップとした。このセンサチップを、専用ソケットを介してポテンショ/ガルバノスタット(装置名:HSV-110、HOKUTO DENKO社製)に接続し、電極上の組成物に0~500mg/dlの標準グルコース溶液5μLを添加して+0.42Vの電圧を印加して電流値をモニタリングし、印加後3秒時点の電流値を応答値とした。グルコース標準液の濃度と応答値の関係を図2に示す。
測定の結果、H322T変異体はグルコース濃度依存的な応答値が得られ、塩化ヘキサアミンルテニウム(III)をメディエーターとして利用できることが判明した。一方で、対照として用いた野生型は各グルコース濃度での応答値が得られず、塩化ヘキサアミンルテニウム(III)をメディエーターとして利用できないことが判明した。
実施例8:FADGDHの配列相同性比較
GENETYXソフト(GENETYX CORPORATION)を使用して、Mucor hiemalis、Mucor prainii、Circinella simplex由来FADGDHのアライメントを実施した結果を図3に示す。Mucor hiemalis由来FADGDHのH322と相同のある部位はMucor prainiiはH343、Circinella simplexはH342であることを見出した。Circinella simplex由来FADGDHのアミノ酸配列を配列番号20に示す。Mucor prainii由来FADGDHのアミノ酸配列を配列番号21に示す。
GENETYXソフト(GENETYX CORPORATION)を使用して、Mucor hiemalis、Mucor prainii、Circinella simplex由来FADGDHのアライメントを実施した結果を図3に示す。Mucor hiemalis由来FADGDHのH322と相同のある部位はMucor prainiiはH343、Circinella simplexはH342であることを見出した。Circinella simplex由来FADGDHのアミノ酸配列を配列番号20に示す。Mucor prainii由来FADGDHのアミノ酸配列を配列番号21に示す。
図4にムコール・ヒエマリス由来FADGDHの立体構造、及び、H322の位置を示す。図4から、メディエーターとなるルテニウム化合物の活性中心への経路上にH322が位置することが分かる。+にチャージするヒスチジンを中性のトレオニンに置換したことで、+にチャージした塩化ヘキサアミンルテニウム(III)が活性中心にアクセスしやすくなったと考えられる。非特許文献1に記載されるアルペルギルス・フラバス由来FADGDHにおけるH403は、ムコール・ヒエマリス由来FADGDHではT438であるが、このアミノ酸残基について非特許文献1と同様にT438D置換に置換しても、表1に示すとおり、ルテニウム化合物の反応性に改善は見られなかった。これは、ムコール・ヒエマリス由来FADGDHは、図4の左上に示される大きなαヘリックス構造が存在し、これがT438近傍を通る活性中心への経路を塞いでいるためと推測される。一方、アルペルギルス・フラバス由来FADGDHには、そのような大きなαヘリックス構造は存在しない。図5に、アルペルギルス・フラバス由来FADGDHの立体構造を示す。これに対し、図3に示す、Mucor prainii由来FADGDH及びCircinella simplex由来FADGDHは、いずれもムコール・ヒエマリス由来FADGDHと同様の前記αヘリックス構造を有する。図3において当該αヘリックス構造に相当する領域を四角で囲んだ。
本発明の変異型FADGDHは、血糖値測定のための試薬やセンサに適用されることにより、医療、診断の分野において広く用いられることが期待される。
Claims (14)
- 配列番号1と70%以上の同一性を有するアミノ酸配列を有し、配列番号1の322位に相当するアミノ酸残基がトレオニン、セリン、アスパラギンまたはグルタミンである、フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
- 配列番号1と80%以上の同一性を有するアミノ酸配列を有する、請求項1に記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
- 配列番号1と90%以上の同一性を有するアミノ酸配列を有する、請求項1または2に記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
- 配列番号1と95%以上の同一性を有するアミノ酸配列を有する、請求項1~3のいずれかに記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
- 配列番号1の322位に相当する位置以外の位置において、1若しくは数個のアミノ酸が欠失、置換、付加若しくは挿入されている、請求項1~4のいずれかに記載のフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ。
- 請求項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼをコードするDNA。
- 請求項6に記載のDNAを含むベクター。
- 請求項7に記載のベクターで形質転換された形質転換体。
- 形質転換体がクリプトコッカス属の微生物である、請求項8に記載の形質転換体。
- 請求項8または9に記載の形質転換体を培養し、培養液中からグルコースデヒドロゲナーゼ活性を有するタンパク質を採取する工程を含む、変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの製造方法。
- 請求項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを使用するグルコース測定方法。
- 請求項1~5のいずれかに記載の変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを含むグルコースセンサ。
- メディエーターとして、ルテニウム化合物を含む、請求項12に記載のグルコースセンサ。
- ルテニウム化合物が、塩化ヘキサアミンルテニウム(III)、(ビピリジン)ルテニウム(II)、(4,4’-ジメチル-2,2’-ビピリジン)ルテニウム(II)、(4,4’-ジフェニル-2,2’-ビピリジン)ルテニウム(II)、(4,4’ -ジアミノ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジヒドロキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジカルボキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(4,4’-ジブロモ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジメチル-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジフェニル-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジアミノ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジヒドロキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、(5,5’-ジカルボキシ-2,2’-ビピリジン)ルテニウム(II)ルテニウム(II)、および、(5,5’-ジブロモ-2,2’-ビピリジン)ルテニウム(II)からなる群から選択された少なくとも一種である、請求項12又は13に記載のグルコースセンサ。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2020123071A JP2022019308A (ja) | 2020-07-17 | 2020-07-17 | Fadgdhおよびその製造方法 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2020123071A JP2022019308A (ja) | 2020-07-17 | 2020-07-17 | Fadgdhおよびその製造方法 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2022019308A true JP2022019308A (ja) | 2022-01-27 |
Family
ID=80203620
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020123071A Pending JP2022019308A (ja) | 2020-07-17 | 2020-07-17 | Fadgdhおよびその製造方法 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP2022019308A (ja) |
-
2020
- 2020-07-17 JP JP2020123071A patent/JP2022019308A/ja active Pending
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5398004B2 (ja) | 改変型グルコース脱水素酵素遺伝子 | |
| JP5169991B2 (ja) | 改変型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ | |
| JP5350762B2 (ja) | フルクトシルアミノ酸オキシダーゼ、およびその利用法 | |
| JP5243878B2 (ja) | フルクトシルバリルヒスチジン測定用酵素、およびその利用法 | |
| JP2013138677A (ja) | グルコース測定用バイオセンサ | |
| US9506042B2 (en) | Glucose dehydrogenase | |
| JP6311270B2 (ja) | 耐熱性に優れたフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ | |
| JP5408125B2 (ja) | 糸状菌由来フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ(fadgdh) | |
| JP5465415B2 (ja) | フルクトシルアミノ酸測定用酵素、およびその利用法 | |
| JP6455134B2 (ja) | 新規なグルコースデヒドロゲナーゼ | |
| JP6460513B2 (ja) | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有するタンパク質 | |
| JP5465427B2 (ja) | フルクトシル−l−バリルヒスチジン測定用酵素、およびその利用法 | |
| JP7775826B2 (ja) | 変異型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼおよびその製造方法 | |
| JP5811521B2 (ja) | フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの比活性を向上するための方法 | |
| JP7842407B2 (ja) | 変異グルコースデヒドロゲナーゼ | |
| JP2022019308A (ja) | Fadgdhおよびその製造方法 | |
| JP5793835B2 (ja) | フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼを用いるグルコース測定方法 | |
| JP6342174B2 (ja) | 新規なグルコースデヒドロゲナーゼ | |
| JP5799534B2 (ja) | フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの安定性を向上するための方法 | |
| JP2011217731A (ja) | 改変型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ | |
| JP5793841B2 (ja) | フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの比活性を向上する方法 | |
| JP2012029677A (ja) | フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの基質特異性を改善するための方法 | |
| JP6390776B2 (ja) | 耐熱性に優れたフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ | |
| JP6619152B2 (ja) | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有するキメラタンパク質 | |
| JP2016116470A (ja) | 変異型ピロロキノリンキノン依存性グルコース脱水素酵素、変異型ピロロキノリンキノン依存性グルコース脱水素酵素をコードする単離核酸分子、変異型ピロロキノリンキノン依存性グルコース脱水素酵素を固定した酵素電極、酵素電極を備えたバイオ電池、及びバイオセンサー |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20230407 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20240319 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20240514 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20240716 |