JP2024522012A - Rnaに作用する内因性アデノシンデアミナーゼ(adar)による効率的および正確なrna編集のためのアンチセンスオリゴヌクレオチド(aso) - Google Patents
Rnaに作用する内因性アデノシンデアミナーゼ(adar)による効率的および正確なrna編集のためのアンチセンスオリゴヌクレオチド(aso) Download PDFInfo
- Publication number
- JP2024522012A JP2024522012A JP2023574225A JP2023574225A JP2024522012A JP 2024522012 A JP2024522012 A JP 2024522012A JP 2023574225 A JP2023574225 A JP 2023574225A JP 2023574225 A JP2023574225 A JP 2023574225A JP 2024522012 A JP2024522012 A JP 2024522012A
- Authority
- JP
- Japan
- Prior art keywords
- chemically modified
- seq
- editing
- nucleotides
- central base
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images












Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
- C12N15/1137—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing against enzymes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/70—Carbohydrates; Sugars; Derivatives thereof
- A61K31/7088—Compounds having three or more nucleosides or nucleotides
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/28—Drugs for disorders of the nervous system for treating neurodegenerative disorders of the central nervous system, e.g. nootropic agents, cognition enhancers, drugs for treating Alzheimer's disease or other forms of dementia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P27/00—Drugs for disorders of the senses
- A61P27/02—Ophthalmic agents
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/11—Antisense
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPR]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/31—Chemical structure of the backbone
- C12N2310/315—Phosphorothioates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/32—Chemical structure of the sugar
- C12N2310/321—2'-O-R Modification
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/32—Chemical structure of the sugar
- C12N2310/322—2'-R Modification
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/32—Chemical structure of the sugar
- C12N2310/323—Chemical structure of the sugar modified ring structure
- C12N2310/3231—Chemical structure of the sugar modified ring structure having an additional ring, e.g. LNA, ENA
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/34—Spatial arrangement of the modifications
- C12N2310/345—Spatial arrangement of the modifications having at least two different backbone modifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/34—Spatial arrangement of the modifications
- C12N2310/346—Spatial arrangement of the modifications having a combination of backbone and sugar modifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y207/00—Transferases transferring phosphorus-containing groups (2.7)
- C12Y207/11—Protein-serine/threonine kinases (2.7.11)
- C12Y207/11001—Non-specific serine/threonine protein kinase (2.7.11.1), i.e. casein kinase or checkpoint kinase
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Biomedical Technology (AREA)
- Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Organic Chemistry (AREA)
- Molecular Biology (AREA)
- General Engineering & Computer Science (AREA)
- Zoology (AREA)
- Biotechnology (AREA)
- Wood Science & Technology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Public Health (AREA)
- Medicinal Chemistry (AREA)
- Pharmacology & Pharmacy (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Neurosurgery (AREA)
- Neurology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Virology (AREA)
- Epidemiology (AREA)
- Hospice & Palliative Care (AREA)
- Psychiatry (AREA)
- Ophthalmology & Optometry (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Saccharide Compounds (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
本発明は、内因性ADARによる、細胞内の標的RNAの部位特異的A-to-I編集に使用するための化学修飾オリゴリボヌクレオチドに関し、上記標的RNA中の標的配列に結合可能である、25~80ヌクレオチドの長さの配列を含み、イノシンに編集される上記標的RNA中の標的アデノシンと向かい合うセントラルヌクレオチドを有する、3個のヌクレオチドであるセントラル塩基トリプレットを含み、a)上記セントラル塩基トリプレットの外側のピリミジンヌクレオシドの少なくとも90%は、糖部分の2’位で化学修飾されているか、デオキシリボヌクレオシドであるか、またはそれらの組み合わせであり、b)連続した6個以上のヌクレオシドは、糖部分の2’位において2’-O-メチルによって化学修飾され、c)上記セントラル塩基トリプレットの3つのヌクレオシドのうち少なくとも2つは、糖部分の2’位において化学修飾されているか、デオキシリボヌクレオシドである。
Description
本発明は、標的RNAの部位特異的編集に使用する方法および化学修飾された核酸に関する。分子生物学の飛躍的な進歩により、細胞の遺伝情報を改変することが可能である。DNAの編集は通常、細胞の遺伝情報の安定した改変につながるが、DNAを変更するのではなく、(m)RNAの遺伝情報を変更することが興味深い場合もある。(m)RNAの編集がDNAに勝る主要な利点は、一方では編集収率の用量依存性であり、他方では処理の可逆性である。標的RNAが編集される細胞内の化学修飾された核酸の濃度を調節することで、編集収率、ひいては標的RNAの翻訳後の修飾タンパク質の量に依存性を持たせることができる。一方、化学修飾された核酸が関連細胞内に存在しなくなると標的RNAの編集が停止し、編集されたRNAは新たに転写された未編集のRNAと置換されるため、この処理は可逆的である。
本発明によるRNA分子の編集は、RNAに作用するアデノシンデアミナーゼ(ADAR)のファミリーに属する酵素によって媒介される。ADARは、二本鎖RNA中のアデノシン(A)からイノシン(I)への脱アミノ化(A-to-I RNA編集)を触媒する酵素ファミリーのメンバーである。この酵素触媒反応の過程で、アデノシンは水和中間体を経てイノシンに変化する。グアノシンは相補的塩基であるシチジンと3つの水素結合を形成できるが、イノシンはシチジンと2つの水素結合しか形成できない。翻訳機構はイノシンをグアノシンとして読み取る。したがって、ADARはRNAレベルで機能的なアデノシンからグアノシンへの変異を導入する効果を有する。
ADARファミリーに属するデアミナーゼがRNA、特にmRNAに作用できる要件は、二本鎖が形成されることである。したがって、ADARが作用できる二本鎖分子を形成できる相補的核酸(以下、オリゴヌクレオチドまたはオリゴリボヌクレオチド)を提供することが必要である。
本発明は、アデノシン(A)からグアノシン(G)への機能的変化を引き起こすことができる化学修飾された核酸を開示する。RNAの配列によっては、このような変異が劇的な影響をもたらすことがある。その変異は、mRNAによってコードされるタンパク質に有害な影響を及ぼす点変異を直してもよく、または、他のアミノ酸が置換によって、翻訳されたタンパク質に組み込まれてもよい。しかしながら、停止コドン(UAA、UAG、UGA)が編集されるまたはスプライス部位のときは、強い影響が観察され得る。
本発明の化学修飾オリゴヌクレオチドの実質的な利点は、このようなオフターゲット編集が可逆的であり、壊滅的な副作用の危険が少ないことである。さらに、必要に応じて治療を中止し、元に戻すことができる。より優れた安全性プロファイルのために、RNAレベルでのヒト遺伝情報の一時的で限定的な操作が広く適用できるようになってもよく、DNAレベルでのゲノム編集が予測不可能で不可逆的な副作用のために危険である可能性がある医学的適応に拡大してもよい。
ヒトの組織には、アデノシンからイノシンへの変換を可能にするいくつかのADAR酵素が発現しており、イノシンは順に翻訳時には生化学的にグアノシンとして読まれる。いくつかのADARが当技術分野で知られている。ADARはXenopus levisで見出され、ヒトおよびネズミの細胞でも見出されている。3つのヒトのADARはすべて共通のC末端デアミナーゼドメインを有するが、触媒活性を有することが明らかになったのはADAR1およびADAR2のみである。ADARは二本鎖RNA結合ドメイン(dsRBD)という共通の機能ドメインを共有する。ADAR1が3つのdsRBDを含むのに対し、ADAR2およびADAR3は2つのdsRBDしか共有しない。ADAR1について、2つのアイソフォームが知られている。構成的に発現する、短い110kDaのADAR1はp110アイソフォームであるのに対し、長い150kDaのADAR1はp150アイソフォームであり、これは別のインターフェロン誘導性プロモーターから発現する。現在の知見では、ADAR2は脳内のコード部位を主に編集する。ADAR1は非コード部位を編集する主要な酵素である。
RNAを効率的に編集するためには、ADARがmRNA転写産物上の特定の標的部位に誘導されることが必要である。先行技術におけるこれまでの試みは、ADARのデアミナーゼ活性を特定の部位に誘導し、それによってADARのデアミナーゼ活性を編集されるmRNA分子上の正しい位置にもたらすために、天然のシス作用性ADARリクルート配列に由来する、特異的なループ-ヘアピン構造のADARリクルート部位を利用していた。部位特異的RNA編集のためのこのような人工核酸は、WO2020/001793に開示されている。先行技術に開示されている人工核酸は標的配列を含み、この配列は、標的RNA中の標的配列に相補的または少なくとも部分的に相補的な核酸配列と、デアミナーゼをリクルートするためのリクルート部位とを含む。
本発明の化学修飾された核酸は、特にデアミナーゼをリクルートするためのループ-ヘアピン構造のリクルート部分を有していない点で、先行技術に開示される核酸オリゴヌクレオチドとは異なる。本発明の化学修飾された核酸は、従来技術から知られているコンストラクトとは別の戦略を使用する。さまざまなRNA消化酵素、特にRNアーゼ(RNase)AおよびRNアーゼHの偏在により、RNAが非常に不安定であることはよく知られている。先行技術によるRNA編集は、リクルート部位によってデアミナーゼをリクルートすることによってのみ達成する。リクルート部位は、所望の作用部位、すなわちイノシンに変換されるべき標的アデノシン、しかし機能的にはグアノシンに変換される、にデアミナーゼを誘導する。
これに反して、本発明の化学修飾された核酸は、デアミナーゼのためのループ-ヘアピン構造のリクルート部位を必ずしも有していない。その代わりに、本発明の化学修飾された核酸は、ADAR酵素が付着するRNA二重鎖を形成し、それにより編集効率が向上する。後者は、その全長にわたって特定の最適な化学修飾パターンである化学修飾された核酸を使用することによって達成される。ASOの化学修飾がセントラルトリプレット(central triplet)に限定されず、セントラルトリプレットに隣接する側面にまで及ぶことが本発明の重要な特徴である。
標的RNA配列のセントラルトリプレットは、両側の1つのヌクレオチドによって隣接するアデノシンを含み、セントラル塩基トリプレット(Central Base Triplet)と称する。本発明の化学修飾オリゴヌクレオチド中のセントラル塩基トリプレットに相補的な配列は、その特異的な化学修飾に関して重要である。mRNA(編集)の翻訳レベルでの機能的変化を可能にするためには、本発明のオリゴヌクレオチドがmRNAの編集を可能にすることが必要である。本発明のオリゴヌクレオチドは、一方では、オリゴヌクレオチドの化学修飾、特にオリゴヌクレオチドの糖部分の修飾によって達成され得る分解(例えばRNアーゼによって引き起こされる)に対して十分に安定化されていることが必須である。
他方では、化学修飾はRNA分子の編集を可能にしなければならない。オリゴヌクレオチドの化学修飾が広範囲に及ぶと、編集効率が許容できないレベルまで低下する。したがって、オリゴヌクレオチドの修飾は、最適な編集効率を得るために、本明細書に記載のガイドラインに従わなければならない。
EP3507366は、ADAR酵素による標的アデノシンの脱アミノ化のための化学修飾一本鎖RNA編集オリゴヌクレオチドを開示しており、これにより、3個の連続したヌクレオチドであるセントラル塩基トリプレットは、糖修飾および/または塩基修飾を含む。フランキング領域(flanking region)は、すべての実施形態(上記先行技術の図2)において、リボース単位における2’-O-メチル化のブロックと、少数の付加的な末端ホスホロチオエート連結とで一様に修飾されている。
しかしながら、上記の先行技術で使用されている核酸の上記均一でブロック状の2’-O-メチル修飾は、内在性発現レベルの天然ADAR酵素による編集活性の強い損失につながることが見出された。これは、dsRNA基質への二本鎖RNA結合ドメインの結合に対する嵩高い2’-修飾の負の効果に従っている。具体的には、2’-Fおよび2’-Fと2’-OMeとの混合物が特によく受け入れられ、核酸上のすべてのピリミジン塩基に配置されるときにヌクレアーゼ消化に対してさらに良好な安定化効果を有することを本発明で記述している。したがって、本発明のオリゴヌクレオチドは、1分子あたり少なくとも1つの2’-F、好ましくは少なくとも5つ、より好ましくは少なくとも8つの2’-F修飾を含む。オリゴヌクレオチド中のすべてのピリミジン塩基における2’-F修飾は、20%~60%、好ましくは45%~55%の範囲である。
しかしながら、セントラル塩基トリプレットの2’-F、特に2’-O-メチル化は、編集収率に強い悪影響を及ぼした。しかしながら、セントラル塩基トリプレットの3つの位置すべてにおいてデオキシリボースがよく許容され、ヌクレアーゼ消化に対して実質的な安定化を与えることを見出した。本発明の好ましい実施態様において、コントロールトリプレットに相補的なオリゴヌクレオチドの3つの糖単位はデオキシリボース単位である。
化学修飾された核酸はさらに、標的mRNAの部位特異的編集における使用に適していることを特徴とする。化学修飾された核酸は、標的アデノシンと向かい合うセントラル塩基トリプレットのセントラルヌクレオチドを除く、標的mRNA中の標的配列と完全に相補的な配列を含む。セントラル塩基トリプレットのセントラルヌクレオチドは、典型的にはシトシンまたはその誘導体であるが、核酸塩基アナログもあり得、典型的にはN-ヘテロ環化合物上に構築され、通常相補的なチミジンまたはウラシルを置換し、通常ADARによる編集部位の認識を向上させる。アデノシンデアミナーゼの作用により、標的アデノシンは転写後にグアノシンへ機能的に変化される。したがって、投与されたオリゴヌクレオチドが標的RNAとハイブリダイズするときに形成されるdsRNA内のADARによって標的とされるアデノシンの認識を向上させるために、上述の1つの例外を除いて、ヌクレオチドの配列はmRNAの標的領域と常に相補的である。さらに重要なことは、オリゴヌクレオチドの修飾、特に糖部分とその間の連結のパターンである。
本発明の原理は、化学修飾された核酸がmRNAの編集を可能にするために十分な期間安定でなければならないという事実に基づいている。通常、RNA分子は細胞内で非常に速く分解される。したがって、核酸は化学修飾されるが、化学修飾された核酸が細胞内で十分な時間存在し続け、同時に当該修飾がADARによる認識を妨げない程度に当該修飾を行われなければならない。オリゴヌクレオチドの修飾には、ヌクレオチドの糖部分が関係する。RNA塩基は修飾されていない部分である。オリゴヌクレオチドを安定化させる好ましい修飾は、デオキシリボース部分、またはリボース部分に2’-O-メチルおよび/または2’-F修飾を有するRNA塩基である。別の修飾は、糖部分間のリン酸結合をホスホロチオエート結合で置き換えることである。
本発明の過程において、人工核酸(オリゴヌクレオチド)は15~80ヌクレオチド、好ましくは25~65ヌクレオチド、より好ましくは30~60ヌクレオチドの長さを有することが見出された。このような長さの核酸を、本出願ではオリゴヌクレオチドと称する。
本発明の化学修飾された核酸(オリゴヌクレオチド)は、標的mRNA中の対応する配列とほぼ100%の相補性で相補的な配列を有する。いくつかの実施形態において、標的mRNA中の対応する配列に対する化学修飾オリゴヌクレオチドの相補性は、少なくとも85%の相補性である。完全な相補性はハイブリダイゼーションプロセスにとって最適である一方、天然のADAR基質はしばしば少数のミスマッチおよび/またはバルジを含み、二本鎖RNA結合ドメインによる基質認識またはデアミナーゼの活性部位内部での基質認識を向上させるために二本鎖基質の構造的摂動を可能にすることによって編集を補助する。
本発明の化学修飾された核酸(オリゴヌクレオチド)では、ピリミジンヌクレオシドの少なくとも90%、好ましくは95%、さらに好ましくは100%が2’-位で化学修飾されている。セントラル塩基トリプレットの外側では、修飾は好ましくは2’-アルキル、より好ましくは2’-O-メチル、または2’-O-エチル、2’-アミノ、または2’-ハロ、より好ましくは2’-フルオロ修飾である。セントラル塩基トリプレットの内側では、少なくとも2つ、より好ましくはすべての核酸塩基、ピリミジンおよびプリンが、2’-OMe、または2’-F、より好ましくは2’-デオキシで修飾されている。本発明の一実施形態において、両末端(5’/3’)に末端ブロックが存在し、好ましくは両末端に3個の2’-OMe/PSヌクレオチドまたは2’-MOE/PSが存在する。
特に好ましい実施形態において、セントラル塩基トリプレットの外側のピリミジンヌクレオシドは、任意の化学量論で、好ましくは20:80~80:20、より好ましくは70:30~30:70、さらにより好ましくは60:40~40:60の2’-フルオロと2’-O-メチル置換基との混合物で修飾されている。2’-Fおよび2’-O-メチル修飾が混合される場合、6個以下、好ましくは5個以下、さらに好ましくは4個以下の2’-O-メチル修飾が1つのブロックに結合されるべきである。このような混合物において、少なくとも1つのピリミジンヌクレオシド、好ましくは少なくとも5つ、より好ましくは少なくとも8つのピリミジンヌクレオシドが2’-フルオロ置換基で修飾されている。
本発明の実施形態の化学修飾された核酸のセントラル塩基トリプレットには、N-ヘテロ環塩基、ピリジンまたはピリミジン誘導体、より好ましくはシトシンヌクレオシドまたはその誘導体を有するヌクレオシドが存在し、これは標的(m)RNA中の標的アデノシンと向かい合う。特に好ましい実施態様において、このヌクレオシドならびに5’および3’-特異的隣接ヌクレオチドは、少なくとも1つの修飾ヌクレオシド、より好ましくは2つの修飾ヌクレオシド、さらにより好ましくは3つの修飾ヌクレオシドを含み、2’炭素原子に置換基を有し、この置換基は2’-フルオロもしくは2’-O-メチル、またはさらにより好ましくは2’-デオキシのいずれかである。
標的(m)RNAにおいて、
コドン(標的とされるAに下線を付し、N=任意の核酸塩基である)が標的とされるとき、本発明の化学修飾された核酸のセントラル塩基トリプレットは、2’-デオキシ-イノシンまたはヒポキサンチン核酸塩基もしくはその誘導体を有する任意のヌクレオチドを含み、これは標的とされるアデノシンに5’-隣接するシトシン塩基と対をなす。好ましくは、2’-デオキシ-イノシンは、2つ、より好ましくは3つの2’-デオキシヌクレオチドを含むセントラル塩基トリプレットに配置される。
コドン(標的とされるAに下線を付し、N=任意の核酸塩基である)が標的とされるとき、本発明の化学修飾された核酸のセントラル塩基トリプレットは、2’-デオキシ-イノシンまたはヒポキサンチン核酸塩基もしくはその誘導体を有する任意のヌクレオチドを含み、これは標的とされるアデノシンに5’-隣接するシトシン塩基と対をなす。好ましくは、2’-デオキシ-イノシンは、2つ、より好ましくは3つの2’-デオキシヌクレオチドを含むセントラル塩基トリプレットに配置される。
本発明の化学修飾された核酸は、分解に対する安定性が向上し、ADARと結合するのに最適な化学修飾パターンを示すので、本発明の核酸は、デアミナーゼを引き付ける特異的なループ-ヘアピン構造のリクルート部分を有していないことが好ましい。
本発明の一実施態様において、化学修飾された核酸は対称的であり、これはセントラル塩基トリプレットに隣接する2つのヌクレオチド配列が同じ長さを有することを意味する。オリゴヌクレオチドが例えば59ヌクレオチドを有するとき、セントラル塩基トリプレットの両側に28ヌクレオチドが存在する。本発明の対称的な実施形態の概略構造を図1Bに示す。
別の実施形態において、本発明の核酸は対称的ではなく、これはセントラル塩基トリプレットに隣接する2つの配列の長さが異なることを意味する。非対称設計は、標的周辺の配列空間をより柔軟に使用することを可能にする。さらに、非対称設計は、核酸が正しい末端で短縮されることを条件として、対称設計と比較して、核酸の短い配列、例えば45ntの編集収率を高めることができることが見出された。好ましくは、セントラル塩基トリプレットに対するフランキング配列(flanking sequence)5’は、非対称実施形態におけるフランキング配列3’よりも長い。好ましい実施形態は、3’フランキング配列において少なくとも4nt、より好ましくは少なくとも9ntを含み、5’フランキング配列において少なくとも19nt、より好ましくは少なくとも28nt、最も好ましくは少なくとも33ntを含む。本発明の非対称実施形態の概略構造を図1Aに示す。
好ましい実施形態において、本発明の核酸は、少なくとも30%、より好ましくは40%、さらに好ましくは50%、最も好ましくは60%超の割合でホスホロチオエート連結を介して連結される。好ましくは、少なくとも10、より好ましくは少なくとも15、最も好ましくは20以上のヌクレオシド連結が、核酸の末端(5’または3’)から出発して、ほとんど不連続になることなく、より好ましくは一切不連続になることなく配置される。本発明の別の実施態様において、好ましくは連続的なホスホロチオエート連結の上記ブロックは、核酸の両末端(5’および3’)から出発して核酸の両側面に配置される。
別の好ましい実施形態において、糖部分の修飾はホスホロチオエート連結と組み合わされる。
本発明の化学修飾された核酸は、RNアーゼによる通常もたらされる分解に対して実質的により安定であり、その結果、(m)RNAが編集されるべき細胞内でより長く存在することができる。理論に束縛されることを望むものではないが、化学修飾された核酸の寿命は細胞の環境中で増加するので、二本鎖による安定性の長期化によってmRNAにADARが作用することができるため、デアミナーゼをリクルートするためのリクルート部位は必要ない。
生物学的反応は時間依存的であることが多い。脊椎動物の細胞内には多種多様なRNA分子が存在し、それらは恒常的で迅速に入れ替わる。RNA分子は様々なRNアーゼによって頻繁に分解される。したがって、治療目的でのRNA分子の使用は、RNA分子の急速な分解によって制限されることが多い。通常、in vivoの状態は、細胞培養による試験系を使用するin vitroの状態とは異なるため、治療目的で使用される分子の安定性は、処置の成功にとって決定的な意味を有する可能性がある。
本発明の化学修飾された核酸分子は、編集能力と細胞内での十分な安定性との良好なバランスを提供し、それによりエンドソーム内の状態さえも許容し得る。本発明の化学修飾オリゴヌクレオチドは、さらにジムノティックの取り込み(gymnotic uptake)が可能であり、許容できる編集効率を示す。
本発明の化学修飾された核酸による最良の結果は、オリゴヌクレオチドにおいて以下の特徴のうち好ましくはいくつか、より好ましくは少なくとも2つ、さらに好ましくは少なくとも3つの特徴が実現されたときに達成され得る:
セントラル塩基トリプレットに隣接する領域に配置されているホスホロチオエートの含有量が高い(少なくとも30%);
少なくとも1つのDNA糖ヌクレオシドが、脱アミノ化されるアデノシンと向かい合うセントラル塩基トリプレットに存在する;
ほぼ等しい化学量論でのヌクレオシドリボース部分の2’-Fまたは2’-OMe修飾による、セントラル塩基トリプレットの外側のピリミジン塩基の安定化であり、それによって2’-OMeのブロックを避けるパターンが好ましい;
両末端は、二重修飾、すなわち糖部分の2’-OMe修飾およびホスホロチオエート連結を有する3つのヌクレオチドのブロックによって安定化されている。
セントラル塩基トリプレットに隣接する領域に配置されているホスホロチオエートの含有量が高い(少なくとも30%);
少なくとも1つのDNA糖ヌクレオシドが、脱アミノ化されるアデノシンと向かい合うセントラル塩基トリプレットに存在する;
ほぼ等しい化学量論でのヌクレオシドリボース部分の2’-Fまたは2’-OMe修飾による、セントラル塩基トリプレットの外側のピリミジン塩基の安定化であり、それによって2’-OMeのブロックを避けるパターンが好ましい;
両末端は、二重修飾、すなわち糖部分の2’-OMe修飾およびホスホロチオエート連結を有する3つのヌクレオチドのブロックによって安定化されている。
本発明の化学修飾された核酸分子(オリゴヌクレオチド)は、分子が脊椎動物の生体中で十分に安定であるという利点を有することによって、所望の効果が達成され得る。本発明の分子は、種々のRNアーゼの分解に対して十分な期間安定であることによって、効果が発揮され得る。
本発明の化学修飾された核酸の別の利点は、特定のベクターまたは特定のトランスフェクション法等の他の助力機構なしに、標的細胞に直接もたらすことができることである。本発明の化学修飾された核酸はジムノシス(gymnosis)を介して作用することができ、これはベクターまたは他の担体等の補助手段なしに標的細胞に直接適用できることを意味する。
本発明の化学修飾された核酸の別の利点は、臨床的に関連する標的において編集効率が高いことである。修飾核酸はジムノシスを介して標的細胞に導入することができ、標的細胞における翻訳レベルに対して比較的高い効果を達成し得る。
本発明の化学修飾された核酸の別の利点は、AからIへの編集が、5’UAG等の比較的編集しやすい標的だけでなく、5’CAA等のより困難なトリプレットでも達成し得ることである。
本発明およびその好ましい実施形態は、実施例および図によって説明されるが、これらは限定的なものではない。
図には、本発明の好ましい実施形態が特に示されている。
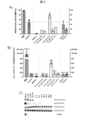
図1は、本発明の2つの好ましい実施形態の基本構造および用語を示す。図1A)は非対称修飾オリゴヌクレオチドを示し、図1B)は対称設計を示す。
図1Aは、標的とされるRNAに結合する、セントラル塩基トリプレット(NNN)の3’-隣接する4以下のヌクレオチドおよび5’-隣接する19以下のヌクレオチドを含む、本発明の非対称実施形態である。標的とされるアデノシン(A*)は、セントラル塩基トリプレットのセントラルヌクレオチドと向かい合っている。
図1Bは、標的とされるRNAに結合する、セントラル塩基トリプレットに5’-および3’-隣接する少なくとも11個のヌクレオチドを含む、本発明の対称的な実施形態である。標的とされるアデノシンは、セントラル塩基トリプレットのセントラルヌクレオチドと向かい合っている。
図2は、実施例1で実施した実験の結果を示す。GAPDHオープンリーディングフレーム(ORF)の編集の程度は、異なるオリゴヌクレオチドについてパーセントで示され、その配列は表中に提供される。実験は、インターフェロン-αの存在下(+IFN)および非存在下で行った。
図2Eは、ガイドRNAの長さが編集効率に及ぼす影響を示している。少なくとも25ヌクレオチドの長さまでは、編集効率は少なくとも許容範囲内であることが分かる。編集効率が約50%以上であるため、好ましくは、下限は30ヌクレオチドである。
図3は、リードオリゴヌクレオチドV120.2およびV117.19、ならびに、GAPDHの編集を示し、これによる実験は実施例2でより詳細に記載されている。2つの修飾オリゴヌクレオチドを図3A)(非対称)および図3B)(対称)に模式的に示す。編集は、HeLa、U2OS、SH-SY5Y、SK-N-BE、Huh7、HepG2、A549、THP-1等の種々の細胞株で試験した。実験はインターフェロン-αの存在下(+IFN)および非存在下で行った。
図4は、好ましいGADPHコンストラクトへのさらなる化学修飾の導入を示す。図4A)およびB)のより詳細な説明および詳細は、実施例3で見出すことができる。図4C)は安定性を実証し、図4D)は実施例4に開示されている編集効率を示す。
図4D)およびE)に示したグラフの詳細は、実施例5で見出すことができる。
図4Fは、セントラル塩基トリプレットの外側のピリミジンヌクレオシドの2’-フルオロおよび2’-O-メチル置換基の組み合わせを有する、「V120.21」および「V120.23」と命名された本発明の2つのオリゴヌクレオチドの編集効率の比較である。これらのオリゴヌクレオチドを、WO2019/158475の図3C(ADAR 102-2)、図7(ADAR 103-8)および図7(ADAR 102-7)で開示されたコンストラクトと比較した。本発明のコンストラクトの優れた編集効率は、図4Fから明確に見出される。
図5は、HeLa細胞における安定な化学修飾GADPH標的コンストラクトによるジムノシスを示す。実験概要は実施例6でより詳細に説明する。
図6は、実施例7(図6A))および実施例8(図6B)で得られた結果を示す。データは、STAT1 Y701編集および安定性データに関する。
図7は、MeCP2 W104Xの編集および安定性データに関する。図7A)に示した結果の詳細は実施例9で見出すことができ、図7B)のより詳細な説明は実施例10で見出すことができ、図7C)のより詳細な説明は実施例11で見出すことができる。
図8は、hIDUA W402Xの編集、α-L-イズロニダーゼアッセイ、および実験詳細の安定性データを示す。8A)については実施例12を、8B)については実施例13を、8C)については実施例14を参照されたい。
図9はmIDUA W392Xの編集および安定性データに関する。図9A)は実施例15の結果、図9B)は実施例16の結果、図9C)は実施例17の結果を示す。
図10は、PDE6A V685M編集および対応する安定性データに関する。実施例18の結果を図10A)に、実施例19の結果を図10B)に、実施例20の結果を図10C)に示す。
図11は、SERPINA1 PiZZ編集およびA1AT ELISAおよび安定性データに関する結果を示す。図11A)の詳細については実施例21を、図11B)については実施例22を、図11C)については実施例23を参照されたい。
図12は、LRRK2 G2019S標的について実施例24で得られた結果を実証する。驚くべきことに、本発明である、修飾を有するオリゴヌクレオチド(V117.19およびV120.2)は、非常に高い編集収率を示す。先行技術の教示に従った化学修飾ピリミジンヌクレオチドを導入することにより(V117.20およびV120.3)、コンストラクトの編集活性は約半分に減少する。
〔実施例1〕ヒトハウスキーピングGAPDH遺伝子のORF中のL157部位および3’UTR部位におけるコンストラクトのRNA編集活性の最適化(図2を参照)
実施例1の結果を図2に示す。2×5~2×25の連結の範囲であるホスホロチオエート(PS)連結の長いブロックは編集の改善をもたらすことが示されている。図2B)は、ホスホロチオエート連結なしでは編集が達成できないことを示している(コンストラクトV117.1)。図2C)および2D)は、オリゴヌクレオチドの長さの影響を証明している。非対称デザインを選択した場合、長さは40ヌクレオチドまで短縮できる。図2E)に示すように、オープンリーディングフレーム(ORF)の外側、例えば3’UTR領域の編集部位について、非対称デザインはオリゴヌクレオチドの長さを25ntまでも短縮できる。さらに、インターフェロン-αを加えても、編集効率の向上にはつながらないことも興味深い。
本発明の好ましい実施形態におけるホスホロチオエート連結の導入の効果、ならびに、最も好ましいコンストラクトの長さおよびセントラル塩基トリプレットの位置決めを調べた。内因性ADARをリクルートすることによって、オープンリーディングフレーム(ORF)における5’UAGコンテキストのアデノシンを編集するための内因性ハウスキーピング転写産物(GAPDH)を標的とするために、本発明の修飾オリゴヌクレオチドを使用した。概して、「IFN」として表されるインターフェロンの実験環境への追加は、コンストラクトの編集活性に大きな影響を与えなかった。コンストラクト配列の詳細を表1Aおよび1Bに示す。
結果を得るために、HeLa細胞(Cat.No.:ATCC CCL-2)をDMEM+10%FBS+P/S(100U/mLのペニシリンおよび100μg/mLのストレプトマイシン)で培養した。100μLのDMEM+10%FBS(+600ユニットのIFN-α,Merck,カタログ番号IF007,ロット番号2937858)中の5×104細胞を、96ウェルフォーマット中の0.5μLのリポフェクタミン2000と5pmolのコンストラクト/ウェルとのトランスフェクションミックスに加えた。24時間後にRNAの単離および配列決定のために細胞を回収した。実験から以下の結論が導き出された。
A)ホスホロチオエート(PS)連結の導入量が増えるにつれて、コンストラクトの編集活性は向上する。セントラル塩基トリプレットの両側にホスホチオエート連結が5つしかないコンストラクトV117.16(配列番号2)は、両側にPS連結が25個あるコンストラクトV117.19(配列番号5)と比較して、編集活性が著しく低下している。
B)ホスホロチオエート連結を全く含まないコンストラクトは、連続したブロックに配置されたPS連結を付加的に含む実施形態(V117.19,配列番号5;および、V119.4,配列番号8)とは対照的に、検出可能な編集を示さない。好ましい実施形態において、オリゴヌクレオチドは、ヌクレオチド間に連続したホスホロチオエート連結の少なくとも1つのヌクレオチドブロックを含む。コンストラクトにおけるホスホロチオエート連結の欠如(V117.1,配列番号1;および、V119.1,配列番号7)は、編集活性に対して壊滅的である。
C)本発明の好ましい実施形態の編集活性は、オリゴリボヌクレオチドの長さに依存する。最小の長さは、編集部位がORF内にあるかまたはORFの外側にあるかに依存して変化し得る。セントラル塩基トリプレットと隣り合うオリゴリボヌクレオチドアームを短くすること、例えば、全長59ntから47ntへの短縮(V117.19,配列番号5;およびV118.3,配列番号6)は、対称性の実施形態では編集活性に不利である。非対称性の実施形態(V119.4,配列番号8;V120.2、配列番号9;V121.1,配列番号10;およびV122.1,配列番号11)について、対称性の実施形態と比較して、全体としてより短いオリゴリボヌクレオチドは、高い編集効率を得るのに十分であり、例えば、40ntまで短縮すること(V121.1,配列番号10)は可能であり、一方、35ntまで短縮すると、ORF内の編集部位に対する編集活性がなくなる(V122.1,配列番号11)。ORFの外側の編集部位のオリゴヌクレオチド長に関しては、以下のコンストラクトを検討した:45ヌクレオチドを有するV120.2(配列番号63);40ヌクレオチドを有するV121.2(配列番号64);35ヌクレオチドを有するV121.3(配列番号65);30ヌクレオチドを有するV121.4(配列番号66);および、25ヌクレオチドを有するV121.5(配列番号67)について、編集効率を検討した。コンストラクトV121.2(配列番号64)は、非対称性コンストラクトの3’末端を4ヌクレオチドまで短縮できることも示している。結果を図2Eに示す。すべての特徴により、本発明の全オリゴヌクレオチド長の下限は25ヌクレオチドであると思われる。
D)同じオリゴリボヌクレオチド長のコンストラクトにおけるセントラル塩基トリプレットの好ましい位置は重要である。より長いコンストラクト、例えば本実施例では59ヌクレオチド(V117.19,配列番号5;およびV119.4,配列番号8)は、セントラル塩基トリプレットの明確な好ましい位置を示さない。しかし、より短いコンストラクト、例えば47ヌクレオチド長(V118.3,配列番号6)または45ヌクレオチド長(V120.2,配列番号9)は、V120.2(配列番号9)と同様に、セントラル塩基トリプレットをコンストラクトの3’末端側に配置する強い優先性を示し、その結果、セントラル塩基トリプレットに5’-隣接する長いヌクレオチド配列を有する本発明の非対称性の実施形態が得られる。
〔実施例2〕種々の腫瘍細胞株での、内因性ADARによるL157部位におけるヒトGAPDH転写産物の編集
異なる組織由来の細胞株における本発明のコンストラクトの適用性を調べるために、次の本発明の特に好ましい2つの実施形態を使用して細胞株スクリーニングを実施した:長く、対称性であるデザイン(V117.19,配列番号5、図3B)および短く、非対称性であるデザイン(V120.2,配列番号9、図3A)。修飾を示すコンストラクトのリストを表2に示す。
種々の腫瘍細胞株は、RPMI+10%FBSで培養したTHP-1を除き、DMEM+10%FBS+P/Sで培養した。1×105細胞/ウェル(HeLa細胞(cat. no. ATCC CCL-2)、U2OS-Flp-In T-REx32(Elmar Schiebelからの親切な寄付)、Huh7(CLS GmbH, Heidelberg, cat. no. 300156)、HepG2(DSMZ, Braunschweig, Germany, cat. no. ACC180)およびA549(European Collection of Authenticated Cell Cultures ECACC 86012804))を24ウェルプレートに播種した。SK-NBE(2)(cat. no. ATCC CRL-2271)およびSH-SY5Y(cat. no. ATCC CRL-2266)細胞株については、24ウェルあたり2.5×105細胞を播種した。24時間後に培地を交換し(IFN処理細胞には3,000U/ウェルのIFN-αを補充)、50μLのOptiMEM中の25pmolのASO/ウェルと50μLのOptiMEM中の1.5μl/ウェルのリポフェクタミンRNAiMAX試薬(ThermoFisher Scientific)とのトランスフェクションミックスで細胞をフォワードトランスフェクトした。5分間のインキュベーション後、両方の溶液を合わせ、さらに20分間インキュベートした後、トランスフェクションミックスを1つのウェルに均等に分配した。24時間後に、RNAの単離および配列決定のために細胞を回収した。
THP-1は、24ウェルプレートの3×105細胞/ウェルをRPMI+10%FBS+PMA(200nM)中で3日間分化させ、その後RPMI+10%FBSで5日間培養した後、同様にトランスフェクトした。
図Cに示すように、両方のコンストラクトとも試験したすべての細胞株で編集活性を示した。注目すべきことに、編集活性は、先行技術(WO2020/001793に開示される)の場合のようには、インターフェロンの添加(「+IFN」)によって著しく改善されることはなかった。
〔実施例3〕内因性ADARによる、内因性ヒトGAPDH転写産物のL157部位を標的とするコンストラクトへの化学修飾パターンの導入
本実施例の結果を図4A)およびB)に示す。例えばコンストラクトV120.7(配列番号13)で使用される大きな2’-O-メチルブロックの使用は、編集の実質的な減少をもたらす(図4A)。図4B)は、ピリミジンヌクレオシドの修飾に関して、2’F修飾はよく受け入れられ、2’-O-メチル修飾は多かれ少なかれ受け入れられるが、2’-MOE[2’-O-(2-メトキシエチル)]は受け入れられないことを示している。
修飾は、合成オリゴヌクレオチドが、非自己オリゴヌクレオチドに対する複数の防御線が存在する治療環境において無傷であるために不可欠であることが証明されている。これらの修飾は、ヌクレオシドの糖部分、例えば2’-ヒドロキシ基(2’OH)の代わりに2’-O-メチル、2’-フルオロ(2’F)、2’-MOEまたは2’-デオキシ基(2’H)等であるが、これらに限定されるものではない。異なる種類の修飾に加えて、コンストラクト中の修飾ヌクレオシドの位置も重要な点である。修飾の種類だけでなく、修飾ヌクレオシドの位置もコンストラクトの編集活性に強い影響を与える。試験した配列およびその修飾のリストを表3Aおよび3Bに示す。
HeLa細胞を培養し、実施例2に記載したようにフォワードトランスフェクトした。簡単に説明すると、24ウェルあたり1×105細胞を播種し、24時間後にそれぞれのコンストラクトを25pmolでフォワードトランスフェクトした。トランスフェクションしてから24時間後、RNA単離およびサンガー配列決定のために細胞を回収した。
図4Aに示すように、先行技術(例えば、EP3507366)の特徴である2’-O-メチル修飾の長いブロック(V120.7,配列番号13)の導入は、編集活性に壊滅的な影響を及ぼす。しかしながら、他の繰り返しパターンの修飾、例えば2’F/2’F/リボ(V120.13,配列番号14)、2’-O-メチル/2’F(V120.14,配列番号15)、2’-O-メチル/2’F/2’F(V120.15,配列番号16)、または2’-デオキシ/2’F/2’F(V120.16,配列番号17)もそれぞれ、編集効率にかなり強い影響を与えた。対照的に、コンストラクト中の多数の選択されたヌクレオチドの化学修飾はよく受け入れられた(図4B)。これらの実施形態では、セントラル塩基トリプレットの内側のピリミジン塩基を2’-デソキシで修飾し、セントラル塩基トリプレットの外側のすべてのピリミジンヌクレオシドで他の修飾と組み合わせた。特に、2’-F修飾は非常によく受け入れられたが(V120.17,配列番号18およびV117.28,配列番号23)、一方、2’-O-メチル修飾単独では、コンストラクトの設計によっては編集収率が低下した(V120.18,配列番号19およびV117.27,配列番号22)。よく知られている修飾である2’-MOEは、すべてのピリミジン上に置かれた場合には受け入れられなかった(V120.19,配列番号20)。
さらに、本発明によるコンストラクトの比較を、先行技術から公知の類似の構造と実施した。WO2019/158475は、塩基編集のための化学修飾ガイドRNAを開示している。図3Cにおいて、W392X変異マウスIDUA部位を標的とする2’-MOE修飾ガイドRNAが、「ADAR 102-2」という名称で開示されている。このコンストラクトは39ヌクレオチドを有する。図7には、「ADAR 103-8」および「ADAR 102-7」と称された、W392XマウスIDUA部位を標的とする他の2’-MOE修飾ガイドRNAも示されている。
上述したように、先行技術から公知の3つのコンストラクトであるADAR 102-2(配列番号68)、ADAR 103-8(配列番号69)およびADAR 102-7(配列番号70)による編集効率を測定する比較例を、本発明のコンストラクトであるV120.21(配列番号41)およびV120.23(配列番号42)と比較して実施した。コントロールは、オリゴヌクレオチドを添加しなかった(ASOなし)。
編集効率を図4Fに示す。明らかに、本発明によるオリゴヌクレオチドは、先行技術に開示されたコンストラクトの編集効率よりも約20倍高い優れた編集効率を示した。この改良された編集効率は主に、セントラル塩基トリプレットの外側のピリミジンヌクレオシドの2’-フルオロ置換基と2’-O-メチル置換基との組み合わせによるものと推測される。
全体として、化学修飾の性質およびオリゴリボヌクレオチド内の配置部位が、RNA編集収率に明確な影響を及ぼすことが示された。
〔実施例4〕内因性ADARによる、内因性L157 GAPDH遺伝子を標的とするコンストラクトの安定化
図4C)および図4D)は、10%FBS中でのヌクレアーゼに対する安定性に関する修飾の効果を示している(図4C))。図4D)は、改良されたオリゴヌクレオチドが十分に安定で、比較的短く、それにもかかわらず有効性が高いことを示している。
本発明の治療可能性を評価するためには、RNアーゼ等の非自己オリゴヌクレオチドに対する内因性防御機構に対する本発明のコンストラクトの安定性を決定することが非常に重要である。コンストラクトへの化学修飾ヌクレオシドの導入は、RNアーゼに対する安定性を改善することができる。これを決定するために、本発明の4つの好ましい実施形態である、糖部分の2’-ヒドロキシル基に化学修飾を有する2つの実施形態(V117.29,配列番号24およびV120.20,配列番号21)ならびに化学修飾がない2つの実施形態(V117.19,配列番号5およびV120.2,配列番号9)を、ウシ胎児血清中で7日間にわたって安定性を試験した。配列および修飾を表4に示す。
コンストラクトを10%ウシ胎児血清(FBS)中でインキュベートした。オリゴヌクレオチドの分解は変性尿素PAGEで可視化した。実験では、15pmolのそれぞれのASOを10μlのPBS+10%FBSで希釈した。モック(mock)サンプルはPBSのみで希釈した15pmolのASOを含んでいた。すべてのサンプルを37℃で所定の時間インキュベートした後、液体N2で凍結させ、直ちに-80℃で保存した。サンプルの変性は、それぞれ7μlのRNAローディング色素(Rotiphorese(登録商標)シークエンスゲル希釈液において、Rotiphorese(登録商標)シークエンスゲルバッファ濃縮液の1:10希釈液、Carl Roth)を加え、70℃で2分間インキュベートすることで達成した。変性したサンプルを尿素(7M)ポリアクリルアミド(15%)電気泳動(PAGE)ゲルにロードし、1×TBEバッファー中、1200Vで4~6時間泳動した。製造元の指示に従ってSYBR(商標)Gold Nucleic Acid Gel Stain(Thermo Fisher Scientific)を使用してバンドを可視化し、Fujifilm FLA-5100蛍光画像解析装置を使用して、励起波長λex=473nmでスキャンした。
コンストラクトの設計、例えばオリゴリボヌクレオチドの長さまたはセントラル塩基トリプレットの位置に関係なく、ヌクレオシド糖部分の2’-ヒドロキシル基の化学修飾を欠く2つのコンストラクト(すなわち、V117.19,配列番号5およびV120.2,配列番号9)は直ちに分解された。対照的に、ピリミジンヌクレオシドの2’-ヒドロキシル基に化学修飾を有する2つのコンストラクト(V117.29,配列番号24およびV120.20,配列番号21)は、10%ウシ胎児血清に7日間を超えて耐えることができた。したがって、選択したヌクレオシドの糖部分に化学修飾を加えることで、RNアーゼ分解に対する耐久性が増し、その結果、血清中でのコンストラクトの寿命が長くなる。
さらに、内因性HeLa ADARをリクルートすることによる内因性GAPDH転写産物の編集について、化学的に安定化された実施形態(V120.17,配列番号18およびV117.29,配列番号24)を、先行技術のオリゴリボヌクレオチド設計(V25.1,配列番号12、WO2020/001793に開示)と並べて評価した(図4D)。具体的には、化学的に安定化された実施形態の両方が、編集効率によって先行技術のオリゴヌクレオチドV25.1(配列番号12)を明らかに凌駕することが見出された。このことは、先行技術のオリゴリボヌクレオチドによる編集効率がIFN-α処理によって押し上げられたとき(「+IFN」)にも当てはまった。
〔実施例5〕種々の細胞の種類における、内因性ADARによるヒトGAPDH転写産物のL157部位の編集(図5DおよびEを参照)。
図4E)は、ヒト初代細胞で実験を実施した実施例5の結果である。これらの実験は、後の商業的使用に直結する細胞株で本発明が機能することを示すので、非常に重要である。
HeLa細胞(図4D)および種々の組織由来の初代細胞(図4E)における本発明のコンストラクトの適用性を調べるために、合計4つの本発明の特に好ましい実施形態を使用した:化学修飾ピリミジンヌクレオシドがない実施形態(V117.19,配列番号5、および、V120.2,配列番号9)、または化学修飾ピリミジンヌクレオシドを有する実施形態(V117.29,配列番号24、および、V120.17,配列番号18)。使用したコンストラクトおよび対応する配列のリストは以下の表(表5)に示す。
正常ヒトアストロサイト(NHA、Lonza cat. No.CC-2565)は、ABM Basal Medium(Lonza cat. No.CC-3187)にAGM SingleQuot Kit Supplementary & Growth Factors(Lonza cat. No.CC-4123)を添加した培地で培養した。ヒト網膜色素上皮細胞(H-RPE, Lonza cat.00194987)は、RtEBM Basal Medium(Lonza cat. No.00195407)に、RtEGM Retinal Epithelial Cell Growth Medium SingleQuots Supplements and Growth Factors(Lonza cat. No.00195407)を添加した一方、FBSを添加しなかった培地で培養した(播種のために、FBSを添加し、24時間後にFBSを含まない培地に培地を交換した)。正常ヒト気管支上皮細胞(NHBE、Lonza cat. No.CC-2540)は、BEGM Bronchial Epithelial Cell Growth Basal Medium(Lonza cat. no CC-3171)に、BEGM Bronchial Epithelial Cell Growth Medium SingleQuots Supplements and Growth Factors(CC-4175)を添加した培地で培養した。トランスフェクション手順は、1×105細胞/ウェルを播種し、25pmolのASOをトランスフェクションして、Hela細胞について実施例3に記載したように行った。IFN処理細胞(「+IFN」)には、コンストラクトのトランスフェクションの前に15,000U/ウェルのIFN-αを補充した。24時間後、RNA単離およびサンガー配列決定のために細胞を回収した。
HeLa細胞を処理し、実施例3で既に記載したように、フォワードトランスフェクトした。1×105細胞/ウェルを播種し、25pmolのASOをトランスフェクトし、対応するウェルを初代細胞について記載したようにIFNで処理した。
図DおよびEに示すように、両方のコンストラクトは、試験したすべての細胞の種類(HeLa、NHA、NHBEおよびRPE細胞)において効率的な編集活性を示した。ピリミジンヌクレオシドにさらなる化学修飾を加えることは、ほとんどの場合において良好に受け入れられた。しかしながら、化学修飾ヌクレオシドを有するコンストラクト(V117.29,配列番号24およびV120.17,配列番号18)は、化学修飾ヌクレオシドがない対応するコンストラクト(V117.19,配列番号5およびV120.2,配列番号9)と比較して、編集活性の低下を示した。注目すべきことに、本発明のすべての実施形態の編集活性は、WO2020/001793に開示された先行技術(V25.1,配列番号12)と比較して著しく改善された。このことは、インターフェロン-α(「+IFN」)を添加したV25.1オリゴリボヌクレオチドによる編集を押し上げた後でも同様であった。
〔実施例6〕内因性ADARによる、内因性ヒトGAPDH転写産物のL157部位を編集するためのコンストラクトのジムノティックの取り込み
本実施例の結果を図5に示す。本発明のコンストラクトは、いわゆる「裸の取り込み(naked uptake)」を可能にすることが示された。これは、オリゴヌクレオチドががん細胞株およびヒト初代細胞において、補助的な手段なしに標的部位に到達することを意味する。ピリミジンヌクレオシドに修飾がなければ、裸の取り込みはあまり効果的ではないことに留意すべきである。
本発明の好ましい実施形態について、トランスフェクション剤(ジムノシス)による補助なしで、ヒトハウスキーピング遺伝子(GAPDH)を編集する能力についても試験した。本実施例に使用したコンストラクトを表6に示す。
HeLa細胞はDMEM+3%FBSで培養した。500μl培地/ウェルに24ウェルあたり104のHela細胞を播種し、24時間インキュベートした。24時間後、培地を5μM、1μM、0.2μMおよび0μMのASOを含む培地に交換した。各ウェルに300μlのコンストラクト含有培地を含ませ、5日後に交換した。HeLa細胞はさらに5日後に剥離し、50%ずつ分割した。3日後、5日後、7日後および10日後にRNA単離およびサンガー配列決定のために細胞を回収した。
初代細胞については、104細胞/ウェルのRPE、NBEおよびNHAを、それぞれの培地中の24ウェルプレートに播種し、24時間後に培地を250μLの培地に交換し、OptiMEM中の50μLのASOを最終ASO濃度が5μMとなるように添加した。オリゴヌクレオチド添加後3日または5日後に、RNA単離のために細胞を回収した。
図5Aに示すように、コンストラクトv117.29(配列番号24)は処理したHeLa細胞に容易に取り込まれ、トランスフェクション剤を加えなくても高い編集活性を示した。最良の結果が得られたのは、HeLa細胞を1μMのコンストラクトで10日間処理したときであった。編集効率は、培地中のV117.29濃度(5μMまたは0.2μM)の変動によって示されるように、用量依存的であった。注目すべきことに、V117.29と比較してさらなる安定的な化学修飾を欠き、血清中で速やかに分解され(実施例4参照)、1μMのコンストラクトV117.19(配列番号5)の添加は、7日間および10日間の処理後、HeLa細胞においてほとんど編集活性を示さなかったが、V117.29はこれらの条件下で最も良好な結果を示した。このことは、本発明で開示する化学修飾パターンが、オリゴヌクレオチドのジムノティックの取り込みのもとで編集を達成することが可能であり、当該達成に必要であることを明確に示している。
処理した初代細胞(RPE、NHA、NHBE細胞、図5Bに示す)も、トランスフェクション試薬による補助なしで、編集を達成するのに十分な量のコンストラクトV117.29が取り込まれることを示している。HeLa細胞における1μMの条件と同様に、3日間の処理と比較して5日後には編集効率が増大する。結論として、安定化されたコンストラクトのジムノティックの取り込みは細胞株および初代細胞の両方で可能であり、内因性ADARを利用することで内因性転写産物の編集を達成するのに十分効率的である。
〔実施例7〕内因性ADARによる、内因性ヒトSTAT1転写産物のY701部位の編集
実施例7の結果を図6に示す。明らかな安定化が認められ、先行技術(v25.1)と比較して編集収率が大幅に向上している。初代細胞において、編集収率は約10倍向上している。オリゴヌクレオチドの化学的安定化により、高い編集収率が得られると結論づけられる。
実施例2において内因性ヒトGAPDHのL157部位を標的とするコンストラクトについて実施したように、内因性ヒトSTAT1転写産物のリン酸化部位Y701を標的とする本発明の3つの好ましいコンストラクトを、2つの異なる細胞株(Huh7およびHeLa細胞)および3つの異なる初代細胞(NHA、NHBEおよびRPE細胞)において試験した。本コンストラクトは、化学的に安定化されたもの(V117.28,配列番号27)および化学的に安定化されていないもの(V120.2,配列番号25およびV117.19,配列番号26)の両方のバージョンがあった。本実施例に使用したコンストラクトの配列を表7に示す。
腫瘍細胞株は実施例2に記載したように処理した一方、初代細胞は実施例5に記載したように処理した。簡単に説明すると、24ウェルあたり1×105の細胞を25pmolのコンストラクトでフォワードトランスフェクトし、24時間後にRNAを単離し、サンガー配列決定を行った。
図6Aに示すように、V117.19は試験したすべての細胞の種類において編集活性が高かった。コンストラクトV117.28のピリミジンヌクレオシドにさらなる安定化化学修飾を加えても、対応する安定性の低いコンストラクトV117.19と比較して編集活性を損なうことはなかった(図6B参照)。V117.19はより短く、より化学的に安定化されているにもかかわらず、両方の対称性コンストラクトの編集活性は、HeLa細胞において先行技術であるV25.1(配列番号28)と比較してほぼ3倍改善され、RPE細胞において8倍超改善された。非対称の実施形態であるV120.2も編集活性を示したが、V117.19およびV117.28の両方に比べて低かった。いずれの新規コンストラクトも、先行技術であるV25.1(WO2020/001793に開示)の場合のように、インターフェロン-α(「+IFN」)の添加によって編集活性を押し上げることはできなかった。全体として、本発明は、IFN処理とは無関係に、先行技術の解決策よりも短いものによって、明らかに優れた編集収率を与えた。
〔実施例8〕内因性ADARによる、ヒトSTAT1遺伝子の内因性Y701部位を標的とするコンストラクトの安定化(図6B参照)
実施例4で上述したように、RNアーゼ等の内因性環境における防御機構に対するコンストラクトの安定化は、本発明の治療的観点を可能にするために不可欠である。このために、本発明の好ましい実施形態である、ある場合(V117.28,配列番号27)およびない場合(V117.19,配列番号26)について、7日間にわたって10%ウシ胎児血清中でその安定性について試験した。配列および修飾は表7に示す。結果は実施例4で既に記載した結果が得られた。
図6Bに示すように、コンストラクトに修飾ピリミジンヌクレオシドを含めると(V117.28)、化学修飾ヌクレオシドを欠くコンストラクト(V117.19)と比較して、10%FBS中での寿命が強く改善される。V25.1(配列番号28、WO2020/001793に開示)と比較した実施例7のコンストラクトの優れた編集活性と合わせて、これらのコンストラクトは先行技術に対して明らかな改善である。
〔実施例9〕内因性ADARによる、マウスMECP2転写産物の疾患を引き起こすW104X変異の編集
図7は、疾患関連モデル系における高い編集効率を示している。図7は、特にFBSアッセイで示されるように、安定化後も良好な収率を示す(図7C))。図7B)は、V120.2対V120.22の比較によって示されるように、非対称の実施形態においても好ましい非対称構造が存在することを示している。
マウスMECP2遺伝子の疾患原因となるW104X変異を標的とするコンストラクトは、2つの異なるアプローチで試験された:プラスミド上の標的cDNAをトランスフェクトしたHeLa細胞、または、piggyBacトランスポザーゼ系を介して疾患原因となるMECP2 W104X変異cDNAをそのゲノムに安定に組み込まれたHeLa細胞。コンストラクトは、不安定なもの(V120.2,配列番号29)、または、修飾を加えて化学的に安定化したもの(V120.21,配列番号30)であった。本実施例に使用したコンストラクトの配列を表8に示す。
プラスミドトランスフェクションによるアプローチ(「プラスミド」)では、5×104のHeLa細胞を24ウェルプレートに播種した。24時間後、細胞をマウスMECP2 W104X cDNA遺伝子を含むプラスミドでフォワードトランスフェクトした。300ngのプラスミドおよび0.9μlのFuGENE(登録商標)6(Promega)をそれぞれ50μlのOpti-MEMで希釈し、5分間インキュベートした後、それらを合わせてさらに20分間インキュベートした。培地を交換し(IFN処理細胞には15,000U/ウェルのIFN-αを補充)、トランスフェクションミックスを1ウェルに均等に分注した。プラスミドトランスフェクションの24時間後、細胞を25pmolのコンストラクト/ウェルおよび1.5μl/ウェルのリポフェクタミンRNAiMAX試薬(ThermoFisher Scientific)によってフォワードトランスフェクトした。24時間後、RNA単離および配列決定のために細胞を回収した。
piggyBacによるアプローチ(「PiggyBac」)では、piggyBacトランスポザーゼ系を介してゲノムに安定に組み込まれたマウスMECP2 W104X cDNA遺伝子を含む1×105のHeLa細胞を24ウェルプレートに播種した。24時間後、25pmolのコンストラクト/ウェルおよび1.5μl/ウェルのリポフェクタミンRNAiMAX試薬(ThermoFisher Scientific)によって、プラスミドによるアプローチの説明と同様に細胞をフォワードトランスフェクトした。IFN処理した細胞には、コンストラクトのトランスフェクションの前に3,000U/ウェルのIFN-αを補充した。24時間後、RNA単離および配列決定のために細胞を回収した。
図7Aに示すように、不安定なコンストラクトであるV120.2、および、化学修飾したピリミジンヌクレオシドで安定化したコンストラクトであるV120.21の両方で、同等レベルの効率的な編集活性が可能であった。プラスミドを介したアプローチでは両方のコンストラクトも同じようにうまくいったが、piggyBacによるアプローチではV120.2よりもV120.21の方が、わずかに活性が低下した。上述の実施例からも分かるように、両方のコンストラクトともにインターフェロン添加後(「+IFN」)、編集活性に著しい差がなかった。
〔実施例10〕内因性ADARによる、マウスMECP2転写産物の疾患原因となるW104X変異の編集収率に対する非対称性の一般コンストラクト設計の逆位の効果(図7B参照)
非対称性コンストラクトについて、セントラル塩基トリプレットを短いコンストラクトの3’末端側に配置すると、同じ長さの対称性コンストラクトに比べて編集活性が向上することが既に示されている。非対称性コンストラクト、特に実施例3AのV120.2(配列番号29)の基本設計を反転させ、通常コンストラクトの3’末端に配置されるセントラル塩基トリプレットをコンストラクトの5’末端に配置した(V120.22、配列番号31)。本実施例に使用したコンストラクトの配列を表8に示す。HeLa細胞は、実施例9のプラスミドトランスフェクトアプローチについて上述したように処理した。
図7Bに示すように、セントラル塩基トリプレットの通常の位置(V120.2、配列番号29)を向かい合う部位(V120.22、配列番号31)に反転させると、編集活性に有害な影響を及ぼし、ほぼ半分に減少する。したがって、非対称な実施形態におけるセントラル塩基トリプレットの位置は、オリゴヌクレオチドの3’末端側にあることが好ましいことは明らかである。
〔実施例11〕内因性ADARによる、マウスMECP2転写産物の疾患を引き起こすW104X変異を標的とするコンストラクトの安定化(図7C参照)
実施例4で既に議論したように、RNアーゼなどの内因性環境における防御機構に対するコンストラクトの安定化は、本発明の治療的観点を可能にするために不可欠である。このために、本発明の好ましい実施形態である、化学修飾されたピリミジンヌクレオシドを有する実施形態(V120.21、配列番号30)、および有さない実施形態(V120.2、配列番号29)を、7日間にわたって10%ウシ胎児血清中でその安定性について試験した。配列および修飾を表8に示す。結果は実施例4で既に説明した結果が得られた。
図7Cに示すように、修飾ピリミジンヌクレオシドをコンストラクトに含めることで(V120.21)、コンストラクトは10%FBS中で7日間にわたって耐えることができるが、不安定なコンストラクト(V120.2)はほとんど直ちに分解される。
〔実施例12〕内因性ADARによる、Hurler症候群患者線維芽細胞におけるヒトIDUA転写産物の疾患原因となるW402X変異の編集(図8A)
図8は疾患関連試験系での結果を示している。標的RNAは患者の線維芽細胞で内因性に発現している。安定化は達成されているが、編集効率に関しては不利である。
IDUA遺伝子の生殖細胞系列変異は、IDUA遺伝子がコードする酵素であるα-L-イズロニダーゼが欠損し、多糖類の分解に関与できないライソゾーム病であるムコ多糖症I型の頻度の高い原因である。このため、リソソーム内にグリコサミノグリカン(GAG)が蓄積し、その重症度は様々である:軽症のScheie症候群(患者は軽度の健康問題を抱え、寿命は正常である)から、重症のHurler症候群(患者は重度の健康問題を抱え、小児期には致死的となる)までに及ぶ。IDUA遺伝子のホモ接合性W402X変異は、重篤なHurler症候群の根本的な原因の一つであり、本実施例では、病気にさせるRNAを編集し、酵素機能を回復させることを試みた、対応するコンストラクトの望ましい標的である(実施例13参照)。対応するコンストラクトの配列は表9で見出すことができる。
軽症のScheie症候群患者(「Scheie」、GM01323)、重症のHurler症候群患者(GM06214)、および健康なドナー(「健康」、GM05659)由来の線維芽細胞は、Coriell Institute for Medical Research(米国)から購入した。線維芽細胞は、15%FBSを含むDMEMで培養した。2.5mlのDMEM+15%FBS中の2.5×105細胞/ウェルを6ウェルプレートに播種した。播種24時間後に、125pmolのコンストラクトおよび7.5μlのリポフェクタミンRNAiMAX試薬(それぞれ250μlのOpti-MEMで希釈)を使用してフォワードトランスフェクションを実施した。5分間のインキュベーション後、両方の溶液を合わせ、さらに20分間インキュベートした後、トランスフェクションミックスを1つのウェルに均等に分配した。Scheie症候群および健常線維芽細胞は、コンストラクトを加えずにトランスフェクションミックスで処理した。トランスフェクションの24時間後に培地を交換した。トランスフェクションの48時間後に、RNA単離および配列決定のために線維芽細胞を回収した。
図8Aに示すように、標的アデノシンを取り囲むイントロン配列を標的とする対称的な実施形態(V117.19イントロン,配列番号35;およびV117.28イントロン,配列番号36)は、低い編集活性しか示さなかった。対照的に、標的アデノシン周辺の成熟mRNA配列を標的としたコンストラクト(V117.19エクソン,配列番号32)は非常に高い編集活性を示し、Scheie症候群線維芽細胞で見出されたものと比較して、補正されたmRNA比を上回る。しかしながら、安定化修飾をコンストラクトに加えると(V117.20エクソン,配列番号33およびV117.21エクソン,配列番号34)、安定性が低い対応するv117.19エクソンと比較して編集収率は低下した。最後に、非対称実施形態のV120.2(配列番号37)も良好な編集活性を示し、Scheie症候群線維芽細胞で見出されたレベルに近いレベルに達した。非対称な実施形態について、コンストラクトにピリミジンヌクレオシドで安定化修飾を加えることで(V120.17、配列番号38)、編集効率もある程度低下する。
〔実施例13〕ヒトIDUA転写産物の疾患原因となるW402X変異を標的とするコンストラクトで処理したHurler症候群患者線維芽細胞由来の細胞溶解物のα-L-イズロニダーゼアッセイ(図8B)
W402X変異α-L-イズロニダーゼのタンパク質機能を、コンストラクトによるRNA編集によって回復させる試みとして(実施例12に記載)、異なるコンストラクトで処理した線維芽細胞の溶解物を使用してα-L-イズロニダーゼアッセイを行った。対応するコンストラクトの配列は表9に見出すことができる。
患者の線維芽細胞を、実施例12に記載したように処理、播種およびトランスフェクトした。各試験コンストラクトについて、2つの6ウェルを使用した。トランスフェクションの48時間後、線維芽細胞を剥離し、PBSで1回洗浄した。40μlのPBS中の0.5%Triton X-100を細胞ペレットに加え、氷上で30分間インキュベートし、α-L-イズロニダーゼ酵素アッセイを行った。4-メチルウンベリフェロンの標準希釈系列を調製した(0~160μM)。標準希釈系列の各濃度について、標準溶液25μlを96ウェルプレート中の25μlの0.4Mギ酸ナトリウム緩衝液(pH3.5)に添加した。線維芽細胞サンプルについては、各溶液25μlを96ウェルに加え、基質溶液25μl(0.4Mギ酸ナトリウム緩衝液、pH3.5中、180μM 4-メチルアンベリフェリル α-L-イズノニド(4-methylumbelliferyl α-L-iduronide))と混合した。反応物を37℃で90分間インキュベートした後、ウェルに200μlのグリシン炭酸バター(pH10.4)を加えて酵素活性を停止させた。4-メチルウンベリフェロンの蛍光は、Tecan Spark 10Mプレートリーダーを使用して、励起波長λex=355nm、発光波長λem=460nmで測定した。算出された酵素活性は、タンパク質量(BCAアッセイにより決定)を基準とし、Scheie溶解物の酵素活性に標準化した。
図8Bで分かるように、結果はScheie症候群線維芽細胞の溶解物中のα-L-イズロニダーゼの酵素活性(100%に設定)に標準化され、これはまた、タンパク質の回復(protein restoration)に必要な最低の治療閾値を決定する。参考として、未処理のHurler症候群線維芽細胞(「ASOなし」)は約3倍低いα-L-イズロニダーゼ活性を示したが、健常線維芽細胞(「健康」)はScheie症候群線維芽細胞で観察された活性の約700倍高いα-L-イズロニダーゼ活性を示した。
イントロンIDUA mRNAを標的としたコンストラクト(V117.19イントロン,配列番号35およびV117.28イントロン,配列番号36)はいずれも、対応する編集活性と同様に、とても効率的なタンパク質の回復は達成できず、α-L-イズロニダーゼ活性レベルはScheie症候群線維芽細胞の半分から4分の3に達した。
非対称性コンストラクト(V120.2,配列番号37;およびV120.17,配列番号38)および安定化対称性コンストラクト(V117.20エクソン,配列番号33;およびV117.21エクソン,配列番号34)はすべて、Scheie症候群線維芽細胞で見出されたものと同程度か、わずかに上回るα-L-イズロニダーゼ活性レベルを示す。最後に、成熟mRNA、V117.19エクソンを標的とする不安定な対称性コンストラクト(配列番号32)は、Scheie症候群線維芽細胞レベルを6倍上回るα-L-イズロニダーゼ活性レベルを示す。全体として、このことは、本発明が治療効果を約束するレベルまでタンパク質機能を回復できることを示している。これは、編集効率に影響を及ぼしたとはいえ、完全な安定化化学修飾パターンを有するいくつかの実施形態にも当てはまった。
〔実施例14〕内因性ADARによる、ヒトIDUA転写産物の疾患原因となるW402X変異を標的とするコンストラクトの安定化(図8C参照)
実施例4で議論したように、RNアーゼ等の内因性環境における防御機構に対するコンストラクトの安定化は、本発明の治療的観点を可能にするために必須である。このために、本発明の好ましい実施形態である、ピリミジンヌクレオシドに化学修飾を導入して安定化させた実施形態(V120.17,配列番号38;V117.20エクソン,配列番号33;V117.21エクソン,配列番号34およびV117.28イントロン,配列番号36)ならびに安定化させていない実施形態(V120.2,配列番号37;V117.19イントロン,配列番号35;および、V117.19エクソン,配列番号32)について、7日間にわたり10%ウシ胎児血清中での安定性を試験した。配列および修飾を表9に示す。結果は実施例4で上述した結果が得られた。
図8Cに示すように、修飾ピリミジンヌクレオシドをコンストラクトに含めることで(V120.17,配列番号38;V117.20エクソン,配列番号33;V117.21エクソン,配列番号34およびV117.28イントロン,配列番号36)、コンストラクトは10%FBS中で7日間にわたって耐えることができるが、不安定なコンストラクト(V120.2,配列番号37;V117.19イントロン,配列番号35;および、V117.19エクソン,配列番号32)はほとんど直ちに分解される。全体として、これは導入した化学修飾の安定化効果を明確に実証している。
〔実施例15〕内因性ADARによる、マウスIDUA転写産物の疾患原因となるW392X変異の編集
図9は実施例15(図9A))、実施例16(図9B))および実施例17(図9C))の結果を示す。
マウスIDUA転写産物のW392X変異は、実施例12で簡単に説明したヒトIDUA遺伝子のW402X変異に相当する。マウスの疾患を引き起こす変異体を標的とするコンストラクトを、その編集活性およびタンパク質の回復について試験した(実施例16参照)。コンストラクトの配列を表10に示す。
5×104のHeLa細胞を24ウェルプレートに播種した。24時間後、細胞をマウスW392X変異IDUA cDNAを含むプラスミドでフォワードトランスフェクトした。300ngプラスミドおよび0.9μlのFuGENE(登録商標)6(Promega)をそれぞれ50μlのOpti-MEMで希釈し、5分間インキュベートし、それらを合わせて、さらに20分間インキュベートした。培地を交換し、トランスフェクションミックスを1つのウェルに均等に分配した。プラスミドトランスフェクションの24時間後、細胞を25pmolのASO/ウェルおよび1.5μl/ウェルのリポフェクタミンRNAiMAX試薬(ThermoFisher Scientific)でフォワードトランスフェクトした。24時間後、RNA単離および配列決定のために細胞を回収した。
図9Aに示すように、不安定なコンストラクトであるV120.2(配列番号40)では比較的良好な編集活性が得られる。安定化修飾を有するコンストラクト(V120.21,配列番号41;およびV120.23,配列番号42)はいずれも、V120.2と比較して編集活性の一部を失う。ピリミジンヌクレオシドの糖部分に2’-フルオロ修飾および2’-O-メチル修飾の交互の混合物を有するコンストラクトV120.23は、2’-フルオロ修飾のみを有するV120.21と比較してわずかに高い編集活性を示し、2’-Fおよび2’-O-メチルの混合が編集効率をサポートできることを示している。
〔実施例16〕マウスIDUA転写産物の疾患原因となるW392X変異を標的とするコンストラクトで処理したHeLa細胞由来の細胞溶解物のα-L-イズロニダーゼアッセイ(図9B)
編集されたIDUA RNAの、回復されたα-L-イズロニダーゼタンパク質への移行を決定するために、α-L-イズロニダーゼ酵素アッセイを行った。絶対活性の結果は、マウスIDUA野生型cDNAを含むプラスミドでトランスフェクトしたHeLa細胞からのα-L-イズロニダーゼ活性と相対化した。コンストラクトの配列は表10に示す。
HeLa細胞を、実施例15に記載したように、24ウェルあたり5×104細胞、300ngプラスミドおよび25pmolコンストラクトで処理、播種、トランスフェクトした。野生型参照データを提供するために、マウスIDUA野生型cDNAを含むプラスミドでHeLa細胞をトランスフェクトした。各設定について、1つの24ウェルを使用した。コンストラクトのトランスフェクションから24時間後、α-L-イズロニダーゼ酵素アッセイ用に、細胞を100μlのM-PERバッファー(Thermo Scientific)で溶解した。サンプルのさらなる処理は、ブラッドフォードアッセイによるタンパク質量の測定を除き、実施例13に記載したように行った。
図9Bで分かるように、α-L-イズロニダーゼアッセイの結果は、図9A(実施例15)の編集データから既に明らかなことを描写している。安定化修飾を有する両コンストラクト(V120.21,配列番号41;およびV120.23,配列番号42)は、V120.2(配列番号40)と比較してα-L-イズロニダーゼ活性が部分的に低下している。しかしながら、編集結果と比較すると、コンストラクトV120.23(ピリミジンヌクレオシドに2’-フルオロ修飾および2’-O-メチル修飾の交互のパターンを有する)は、ピリミジンヌクレオシドに2’-フルオロ修飾のみを有するV120.21と比較して、明らかに高い編集活性を示し、2’-Fと2’-O-メチルの混合が編集効率をサポートすることを示している。
〔実施例17〕内因性ADARによる、マウスIDUA転写産物の疾患原因となるW392X変異を標的とするコンストラクトの安定化(図9C)
実施例4で議論したように、例えば血清中に見出されるRNアーゼに対するコンストラクトの安定化は、本発明の治療的観点を可能にするために必須である。このために、本発明の好ましい実施形態である、ピリミジンヌクレオシドにおける化学修飾の導入により安定化された実施形態(V120.21,配列番号41)および安定化されていない実施形態(V120.2,配列番号40)を、7日間にわたる10%ウシ胎児血清中での安定性について試験した。コンストラクトの配列を表10に示す。結果は実施例4に記載した結果が得られた。
図9Cに示すように、修飾ピリミジンヌクレオシドをコンストラクト(V120.21)に含めることで、コンストラクトは10%FBS中で7日間にわたって耐えることができるが、不安定なコンストラクト(V120.2)はほとんどすぐに分解され、安定化効果が得られたことが再び示された。
〔実施例18〕内因性ADARによる、マウスPDE6A転写産物の疾患原因となるV685M変異の編集(図10A)
2つの好ましい実施形態V117.19およびV120.2は、複数の他の標的について以前に試験された(図3AおよびBに示したスキーム)。網膜色素変性症によって徐々に失明を引き起こし、5’-UAU-3’コンテキストのアデノシンによって引き起こされる、マウスPDE6A遺伝子の疾患原因となるV685M変異部位に対して設計されたコンストラクトの編集活性を調べた。コンストラクトのリストを表11に示す。
5×104のHeLa細胞を24ウェルプレートに播種した。24時間後、細胞は疾患原因となるマウスPDE6A V685M cDNAを含むプラスミドでフォワードトランスフェクトされた。300ngのプラスミドおよび0.9μlのFuGENE(登録商標)6(Promega)をそれぞれ50μlのOpti-MEMで希釈し、5分間インキュベート後、それらを合わせ、さらに20分間インキュベートした。培地を交換し、トランスフェクションミックスを1つのウェルに均等に分配した。プラスミドトランスフェクションの24時間後、細胞を25pmolのASO/ウェルおよび1.5μl/ウェルのリポフェクタミンRNAiMAX試薬(ThermoFisher Scientific)でフォワードトランスフェクトした。24時間後、RNA単離および配列決定のために細胞を回収した。
図10Aに示すように、不安定な対称性コンストラクト(V117.19,配列番号43)が最も高い編集活性を示す。しかしながら、コンストラクトが安定化修飾を有すると(V117.20,配列番号44;およびV117.21,配列番号45)、ヒトIDUA遺伝子のW402X部位を標的とする対称性コンストラクト(配列番号32~36)について、実施例12で分かることと同様に、コンストラクトの編集活性は低下する。
非対称性コンストラクトについては、正反対のことが観察される。不安定なコンストラクトV120.2(配列番号46)は、安定化されたコンストラクトV120.3(配列番号47)に比べて編集活性がわずかに低い。対称性コンストラクトのような安定化による編集活性の低下は観察されない。
〔実施例19〕内因性ADARによる、ヒトPDE6A転写産物の疾患原因となるV685M変異の編集(図10B)
実施形態V117.19およびV120.2は、既に示した多数の他の標的に対して強く好ましい。実施例18について上述したように、PDE6A遺伝子のV685M変異-今回はヒト-は、本発明が適用できる別の潜在的部位である。マウス遺伝子とは対照的に、ヒト遺伝子の標的アデノシンは5’-CAU-3’コンテキストの一部である。コンストラクトのリストを表12に示す。
結果は実施例18について記述した結果と同様に得られた。しかしながら、マウスPDE6A遺伝子を含むプラスミドでHeLa細胞をトランスフェクトする代わりに、変異ヒトPDE6A V685M cDNAを含むプラスミドをトランスフェクトした。
全体的な編集収率が実施例18のマウスコンテキストほど高くないことは明らかであるが、図10Bに示されるコンストラクトの編集活性もまた、選択された設定および遺伝子に対する本発明の適用性を証明することは明らかである。全体的に低い編集活性は、マウス遺伝子においてより好ましいコンテキスト(5’-UAU)と比較して、ヒト遺伝子(5’-CAU)においてADARが標的アデノシンを検出するためにあまり好ましくない編集コンテキストが原因である可能性がある。
V117.19(配列番号48)が最も高い全体的な編集活性を示す。興味深いことに、実施例18のマウス遺伝子で既に分かるように、安定な非対称性コンストラクトV120.17(配列番号50)は、対応する不安定コンストラクトV120.2(配列番号49)よりもわずかに高い編集活性を示す。
〔実施例20〕内因性ADARによる、マウスPDE6A転写産物の疾患原因となるV685M変異を標的とするコンストラクトの安定化(図10C)
実施例4で議論したように、血清中に見出されるRNアーゼ等の内因性防御機構に対して安定なコンストラクトが、本発明の治療的観点からは好ましい。このために、実施例18で使用した好ましい実施形態である、ピリミジンヌクレオシドにおける化学修飾の導入により安定化された実施形態(V117.20,配列番号44)および安定化されていない実施形態(V117.19,配列番号43)を、7日間にわたる10%ウシ胎児血清中での安定性について試験した。コンストラクトの配列を表11に示す。結果は実施例4に記載した結果が得られた。
図10Cに示すように、修飾ピリミジンヌクレオシドをコンストラクト(V117.20,配列番号44)に含めると、コンストラクトは10%FBS中で7日間にわたって耐えることができるが、不安定なコンストラクト(V117.19,配列番号43)はほとんどすぐに分解される。
〔実施例21〕内因性ADARによる、ヒトSERPINA1転写産物の疾患原因となるE342K変異の編集(図11A)
SERPINA1遺伝子はα-1-アンチトリプシン(A1AT)タンパク質をコードし、肝臓で合成され、特に薄い組織を自己消化から守る重要なプロテアーゼ阻害剤である。肝臓から最終的に血流に分泌され、全身に広がる。SERPINA1遺伝子の生殖細胞系列変異は、様々な重症度のA1ATの機能障害を引き起こす可能性がある。重篤なE342K変異はA1ATタンパク質のミスフォールディングを引き起こし、それによりA1ATタンパク質は血流中に適切に分泌されなくなる。これにより、A1ATの過剰蓄積による肝疾患およびA1ATの不足による肺疾患が引き起こされる。E342K変異の根底にある標的アデノシンは5’-CAA-3’にある。
ヒトSERPINA1遺伝子の疾患原因となるE342K変異を標的とするコンストラクトは、2つの異なるアプローチで試験された。すなわち、プラスミド上の標的cDNAをトランスフェクトしたHeLa細胞、または疾患原因となるE342K SERPINA1変異cDNAを、piggyBacトランスポザーゼシステムを介してゲノムに安定的に組み込んだHeLa細胞である。このコンストラクトは不安定であるか(V117.19,配列番号51;およびV120.2,配列番号55)、修飾ピリミジンヌクレオシドで化学的に安定化されていた(V117.24,配列番号52;V117.25,配列番号52;V117.25,配列番号53;V117.26,配列番号54;およびV120.10,配列番号56)。本実施例で使用したコンストラクトの配列を表13に示す。
プラスミドトランスフェクションによるアプローチ(「プラスミド」)では、2.5×104のHeLa細胞を24ウェルプレートに播種した。24時間後、細胞をヒトSERPINA1 E342K変異cDNAまたはSERPINA1健常cDNA(「野生型」)を含むプラスミドでフォワードトランスフェクトした。300ngプラスミドおよび0.9μl FuGENE(登録商標)6(Promega)をそれぞれ50μlのOpti-MEMで希釈し、5分間インキュベートした後、それらを合わせてさらに20分間インキュベートした。培地を交換し、トランスフェクションミックスを1つのウェルに均等に分配した。プラスミドトランスフェクションの24時間後、細胞を5pmolのコンストラクト/ウェルおよび1.5μl/ウェルのリポフェクタミンRNAiMAX試薬(ThermoFisher Scientific)でフォワードトランスフェクトした。24時間後、培地を交換した。トランスフェクションから48時間後、RNA単離および配列決定のために細胞を回収した。
piggyBacトランスポザーゼによるアプローチ(「PiggyBac」)について、piggyBacトランスポザーゼシステムを介してゲノムに安定的に組み込まれたヒトSERPINA1 E342K変異cDNA遺伝子またはSERPINA1野生型遺伝子(「野生型」)を含む1×105のHeLa細胞を24ウェルプレートに播種した。24時間後、細胞を25pmolのコンストラクト/ウェルおよび1.5μl/ウェルのリポフェクタミンRNAiMAX試薬(ThermoFisher Scientific)でフォワードトランスフェクトした。24時間後、RNA単離および配列決定のために細胞を回収した。
図11Aに示すように、不安定な対称性コンストラクトV117.19(配列番号51)およびV117.24(配列番号52)、ならびに安定なおよび不安定な非対称性コンストラクトV120.2(配列番号55)およびV120.10(配列番号56)の編集活性は、piggyBacおよびプラスミドアプローチのいずれにおいても中程度である。しかしながら、グアノシン塩基の代わりにイノシン塩基をセントラル塩基トリプレットの最末端3’-部位に導入すると、コンストラクト(V117.25,配列番号53;および特にV117.26,配列番号54)の編集活性は強い増加を示すが、両コンストラクトは安定化修飾を有し、通常、対応する不安定なコンストラクトと比較して編集活性の低下を引き起こしている。ここでも、実施例16(図9)で既に分かるように、ピリミジンヌクレオシドにおいて2’-Fのみではなく、2’-Fと2’-O-メチルの混合物(V117.25対117.26)は編集収率を改善した。
〔実施例22〕ヒトSERPINA1転写産物のE342K変異部位を標的とするコンストラクトで処理した細胞の細胞上清による、サンドイッチELISAで測定したα-1-アンチトリプシンの回復(図11B)
A1ATは分泌タンパク質であるため、RNA編集の成功がタンパク質の回復に及ぼす影響は、未処理の細胞と比較して、編集コンストラクトで処理した細胞から分泌されるA1ATの量を測定することによって試験することができる。これはプラスミドをトランスフェクトしたアプローチで行われた。本実施例で使用したコンストラクトの配列を表13に示す。
HeLa細胞は、24ウェルあたり2.5×104の細胞、300ngプラスミドおよび5pmolコンストラクトで、プラスミドをトランスフェクトしたアプローチの記載のように処理した。コンストラクトのトランスフェクションから48時間後、細胞から300μlの上清を除去し、300×gで5分間遠心した。上清を希釈バッファー(1×PBS+0.05%Tween-20+2%ウシ血清アルブミン(BSA))で1:30に希釈し、ウサギ抗ヒトα-1-アンチトリプシン抗体(A0012, Dako Denmark ApS)をコートしたGreiner Microlon(登録商標)High Binding 96ウェルプレートの96ウェルに100μlをアプライし、洗浄バッファー(1×PBS+0.05%Tween-20)で洗浄し、1×PBS中の5%ミルクでブロッキングした。サンプルは3連で測定し、ヒトA1AT標準物質(SRP6312, Sigma-Aldrich)とともにロードし、RTで80分間インキュベートした。その後、プレートを洗浄バッファーで洗浄し、ワサビペルオキシダーゼ標識ヤギ抗ヒトA1AT抗体(Biomol, A56-122P)100μlを、1×PBS中1:10,000希釈で各ウェルにアプライし、RTで1時間インキュベートした。プレートを洗浄バッファーで再度洗浄し、100μlのOPD基質(9mlのミリポア水、1mlのStable Peroxide Substrate Buffer 10x(34062, Thermo Scientific)および1x OPD Tablet(34006, Thermo Scientific))を各ウェルにアプライし、暗所、RTで30分間インキュベートした。反応を50μlの2.5MのH2SO4で停止し、直ちにTecan Spark(登録商標)10MプレートリーダーでOD値をλ=490nmで読み取った。タンパク質濃度は標準物質から計算し、野生型細胞上清および非標的ネガティブコントロールコンストラクト(「GAPDH V117.19」、配列番号5)をトランスフェクトした変異細胞由来の上清の値に対して標準化した。
図11Bに示すように、処理細胞から分泌されたA1AT量は、実施例21で見出された編集活性を反映している。低い編集活性を示した不安定なコンストラクト、V117.24(配列番号52)もまた、コントロールと比較してA1AT分泌を示さない。しかしながら、セントラル塩基トリプレットにイノシンヌクレオシドを導入した安定型(V117.25、配列番号53;およびV117.26、配列番号54)は、野生型陽性コントロールの半分に近いA1AT分泌量を示した。ここでも、実施例15/16(図9)および実施例21で既に分かるように、ピリミジンヌクレオシドの2’-Fのみではなく、2’-Fと2’-O-メチルの混合物(V117.25対117.26)がタンパク質の修復を改善した。
〔実施例23〕内因性ADARによる、ヒトSERPINA1転写産物の疾患原因となるE342K変異を標的とするコンストラクトの安定化(図11C)
実施例4で議論したように、血清中に見出されるRNアーゼ等の内因性防御機構に対して安定なコンストラクトが、本発明の治療的観点からは好ましい。このために、実施例22で使用した好ましい実施形態である、ピリミジンヌクレオシドにおける化学修飾の導入により安定化された実施形態(V117.25,配列番号53;およびV117.26,配列番号54)、および安定化されていない実施形態(V117.24,配列番号52)を、7日間にわたる10%ウシ胎児血清中での安定性について試験した。コンストラクトの配列を表13に示す。結果は実施例4に記載した結果が得られた。
図11Cに示すように、また以前の標的についても、V117.25およびV117.26のようにコンストラクト内のピリミジンヌクレオシドを修飾することにより、コンストラクトは10%FBS中で7日間以上耐えることができるのに対し、不安定なコンストラクトV117.24は直ちに分解される。全体として、実施例21~23は、治療への応用に関して本発明の有望な特性を示している。
先行技術から本発明を識別するために、WO2018/041973に開示された2つのオリゴヌクレオチド、ADAR60-6(ここでは配列番号57)およびADAR60-10(ここでは配列番号58)が、同じ条件下で編集活性およびタンパク質回復を比較するために選択された。両オリゴヌクレオチドは、修飾の種類、密度および位置が比較的類似していることから選ばれた。両オリゴヌクレオチドは31ヌクレオチドから構成され、そのセントラル塩基トリプレットに相当する部分は、本発明の好ましい実施形態であるv117.19(図3B)と同様に、全体構造内に対称的に配置された。両末端に連続した4個のホスホロチオエート連結から構成される先行技術のオリゴヌクレオチド骨格は、したがって、各末端に連続した25個のホスホロチオエート連結を含む、本実施例の実施形態V117.25(配列番号53)およびV117.26(配列番号54)に示されるように、より少ないホスホロチオエート連結を含んでいた。最後に、ADAR60-6はDNAヌクレオシドで完全に構成されたセントラル塩基トリプレットを有するが、ADAR60-10は、標的とされるアデノシンと向かい合う塩基に3’-隣接するイノシンヌクレオシドを含むセントラル塩基トリプレットを有する。組み合わせて、2つのオリゴヌクレオチドADAR60-6(配列番号57)およびADAR60-10(配列番号58)は、本発明で提示した好ましい実施形態V117.25(配列番号53)およびV117.26(配列番号54)と同様の修飾を示し、したがって、先行技術との比較に適していると考えられた。しかしながら、それらは、オリゴヌクレオチドの長さ、低いホスホロチオエート含有量、および2’-O-メチル化塩基の大きなブロックの使用において、本発明とは異なる。それらの正確な配列は表13に開示されている。
図11A)に示すように、ADAR60-6およびADAR60-10は、前項で記載した実施例21~23で提示した実施形態と比較して類似していたが、いずれのオリゴヌクレオチドについても、試験された条件下で追跡可能な編集活性は検出されなかった(実施例21および図11A参照)。さらに、ADAR60-6およびADAR60-10は、A1ATサンドイッチELISAを介して示されるように、A1ATタンパク質を回復させることができなかった(実施例22および図11Bを参照)が、本発明のE342K変異SERPINA1標的を標的とする優れたコンストラクトであるV117.26(配列番号54)は、野生型A1ATコントロールの平均42%を回復させた。先行技術の解決策では、内因性ADARを使用した顕著な編集を達成することはできなかった。このデータは、検出可能な編集を達成するためにADAR2の過剰発現および細胞溶解物中での編集実験の高度に人為的な実施を必要とする、元の特許WO2018/041973(図3)に提示されたデータに適合する。
全体として、本データは、生細胞内の内因性ADARによるRNA編集収率およびタンパク質の回復という点で、本発明が先行技術よりも優れていることを示している。
〔実施例24〕内因性ADARによる、ヒトLRRK2転写産物における疾患原因となるG2019S変異の編集(図12)
神経系の重要なキナーゼであるLRRK2遺伝子の変異は、神経変性疾患と関連している。病原性変異の1つであるG2019Sは、パーキンソン病の発症リスクを高める。G2019S変異の根底にある標的アデノシンは5’-CAGコンテキストにある。したがって、本発明の標的として適している。コンストラクトの配列を表14に示す。
5×104のHeLa細胞を24ウェルプレートに播種した。24時間後、細胞を、G2019S変異を有するヒトLRRK2 cDNAを含むプラスミドでフォワードトランスフェクトした。300ngのプラスミドおよび0.9μlのFuGENE(登録商標)6(Promega)をそれぞれ50μlのOpti-MEMで希釈し、5分間インキュベートした後、それらを合わせてさらに20分間インキュベートした。培地を交換し、トランスフェクションミックスを1つのウェルに均等に分配した。プラスミドトランスフェクションの24時間後、細胞を25pmolのASO/ウェルおよび1.5μl/ウェルのリポフェクタミンRNAiMAX試薬(ThermoFisher Scientific)でフォワードトランスフェクトした。24時間後、RNA単離および配列決定のために細胞を回収した。
図12に示すように、化学修飾ピリミジンヌクレオシドを含まないコンストラクト(V117.19,配列番号59;およびV120.2,配列番号61)で得られた編集収率は非常に高く、60~80%の範囲である。化学修飾されたピリミジンヌクレオシドを導入すると、すなわちV117.20(配列番号60)およびV120.3(配列番号62)は、コンストラクトの編集活性が約半分に低下する。ピリミジンヌクレオシドに化学修飾を導入すると、このように編集活性が低下することは、上述の実施例で既に注目されていた傾向である。SERPINA1標的とともに、ここで示した5’-CAGのように、ADARによってあまり好まれないコンテキストのアデノシンを標的とすることにも、先の発明が有用であることが証明できる。
本出願に記載のヌクレオチド配列は、添付の配列表にも要約されている。しかしながら、配列表はヌクレオチドの配列を示すものであり、配列の修飾は複雑であるため配列表には反映されていない。ヌクレオチド配列が標的配列と一致しなければならないのに対して、修飾は明細書に明確に示されており、修飾パターンは本発明にとって重要である。
Claims (18)
- 内因性ADARによる、細胞内の標的RNAの部位特異的A-to-I編集に使用するための化学修飾オリゴリボヌクレオチドであって、
上記標的RNA中の標的配列に結合可能である、25~80ヌクレオチドの長さの配列を含み、
イノシンに編集される上記標的RNA中の標的アデノシンと向かい合うセントラルヌクレオチドを有する、3個のヌクレオチドであるセントラル塩基トリプレットを含み、
a)上記セントラル塩基トリプレットの外側のピリミジンヌクレオシドの少なくとも90%は、糖部分の2’位で化学修飾されているか、デオキシリボヌクレオシドであるか、またはそれらの組み合わせであり、
b)連続したわずか6個のヌクレオシドは、糖部分の2’位において2’-O-メチルによって化学修飾され、
c)上記セントラル塩基トリプレットの3つのヌクレオシドのうち少なくとも2つは、糖部分の2’位において化学修飾されているか、デオキシリボヌクレオシドであるか、またはそれらの組み合わせであることを特徴とする、化学修飾オリゴリボヌクレオチド。 - 上記標的RNA中の編集される標的アデノシンと向かい合うヌクレオシドが、N-ヘテロ環塩基、より好ましくはピリジンまたはピリミジン塩基、好ましくはシトシン、ウラシル、チミンまたは5-メチルシトシンの群から選択されるヌクレオシド、最も好ましくはシトシンを有する、請求項1に記載の化学修飾オリゴリボヌクレオチド。
- ヌクレオシド間の連結の少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、最も好ましくは少なくとも60%がホスホロチオエート連結である、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 上記オリゴリボヌクレオチドが、少なくとも5個、好ましくは少なくとも10個、より好ましくは少なくとも15個、最も好ましくは少なくとも20個のホスホロチオエート連結を有するヌクレオシド間の連続したホスホロチオエート連結の少なくとも1つのヌクレオチドブロックを含む、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 上記セントラル塩基トリプレットの外側のピリミジンヌクレオシドの化学修飾が、2’-フルオロおよび2’-O-メチル置換基の組合せを構成し、それにより、化学修飾の少なくとも20%、好ましくは30~70%、より好ましくは40~60%が2’-O-メチル置換基である、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 6個以下、より好ましくは5個以下、最も好ましくは4個以下の連続したヌクレオシドが2’-O-メチル修飾を含む、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- ピリミジンの100%が、糖部分の2’位で修飾されているか、デオキシリボヌクレオシドであるか、またはそれらの組み合わせである、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 上記セントラル塩基トリプレット中の3つのヌクレオシドのそれぞれがデオキシリボヌクレオチドである、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 5’-CAN配列コンテキスト(N=G、A、U、またはC)中のアデノシンを標的とするとき、上記セントラル塩基トリプレットの3’位のヌクレオシドがイノシンまたはその誘導体からなることによって、上記オリゴリボヌクレオチド中のイノシンが標的(m)RNA中のシトシン塩基と向かい合うように存在する、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- ヘアピン-ループ構造ADARリクルート部分が存在しない、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 上記セントラル塩基トリプレットに隣り合う2つのヌクレオチド配列が同じ長さを有する、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 上記セントラル塩基トリプレットに隣り合う2つのヌクレオチド配列が異なる長さを有し、それにより、3’末端の配列が少なくとも4個のヌクレオチド、好ましくは少なくとも9個のヌクレオチドであり、5’末端の配列が少なくとも19個のヌクレオチド、好ましくは少なくとも28個のヌクレオチド、より好ましくは少なくとも33個のヌクレオチドである、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 3’末端および5’末端が、3個の連続した2’-O-メチルホスホロチオエートヌクレオチドまたは3個の2’-O-メトキシエチル(2’MOE)ホスホロチオエートヌクレオチドのブロックでそれぞれ修飾されている、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 3’末端および5’末端の両方の末端が、少なくとも5個の連続したホスホロチオエート連結を含む、先行する請求項のいずれかに記載の化学修飾オリゴヌクレオチド。
- 上記セントラル塩基トリプレットの外側のピリミジンヌクレオシドの化学修飾が、少なくとも1個、特に少なくとも5個の2’-フルオロ置換基を含む、先行する請求項のいずれかに記載の化学修飾オリゴヌクレオチド。
- 上記セントラル塩基トリプレットの外側のピリミジンヌクレオシドの化学修飾が、2’-フルオロおよび2’-O-メチル置換基の組合せを構成し、それによって化学修飾の少なくとも20%が2’-F置換基である、先行する請求項のいずれかに記載の化学修飾オリゴリボヌクレオチド。
- 先行する請求項のいずれかに記載の少なくとも1つの化学修飾オリゴリボヌクレオチドを含むことを特徴とする、ヒトの障害または疾患、好ましくは遺伝的障害または疾患の処置または予防のための医薬。
- ヒトSERPINA1中の共通のE342K変異を標的とすることによるα1-アンチトリプシン欠損症の処置もしくは予防のための、または、
ヒトLRRK2中の共通のS2019G変異を標的とすることによる神経変性疾患の処置もしくは予防のための、または、
ヒトIDUA中の共通のW402X変異を標的とすることによるムコ多糖症の処置もしくは予防のための、または、
ヒトPDE6A中のV685M変異を標的とすることによる網膜色素変性症の処置および予防のための、または、
ヒトPEX1遺伝子中のG843D変異を標的とすることによるツェルウェガースペクトラム障害の処置および予防のための、または、
ヒトABCA4中のG1961E変異を標的とすることによるスターガルト病もしくは加齢黄斑変性の処置もしくは予防のための、または、
ヒトCRB1中のC948Y変異を標的とすることによる網膜色素変性症もしくはレーバー先天性黒内障等の網膜疾患の処置もしくは予防のための、請求項1~16のいずれかに記載の化学修飾オリゴリボヌクレオチド。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP21177135.7A EP4098745A1 (en) | 2021-06-01 | 2021-06-01 | Antisense oligonucleotides (aso) for efficient and precise rna editing with endogenous adenosine deaminase acting on rna (adar) |
| EP21177135.7 | 2021-06-01 | ||
| PCT/EP2022/064714 WO2022253810A1 (en) | 2021-06-01 | 2022-05-31 | Antisense oligonucleotides (aso) for efficient and precise rna editing with endogenous adenosine deaminase acting on rna (adar) |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2024522012A true JP2024522012A (ja) | 2024-06-06 |
Family
ID=76250081
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2023574225A Pending JP2024522012A (ja) | 2021-06-01 | 2022-05-31 | Rnaに作用する内因性アデノシンデアミナーゼ(adar)による効率的および正確なrna編集のためのアンチセンスオリゴヌクレオチド(aso) |
Country Status (8)
| Country | Link |
|---|---|
| US (1) | US20250059534A1 (ja) |
| EP (2) | EP4098745A1 (ja) |
| JP (1) | JP2024522012A (ja) |
| KR (1) | KR20240016350A (ja) |
| CN (1) | CN117616122A (ja) |
| AU (1) | AU2022286618A1 (ja) |
| CA (1) | CA3221207A1 (ja) |
| WO (1) | WO2022253810A1 (ja) |
Families Citing this family (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2017220751A1 (en) | 2016-06-22 | 2017-12-28 | Proqr Therapeutics Ii B.V. | Single-stranded rna-editing oligonucleotides |
| CN110352244B (zh) | 2016-09-01 | 2023-03-21 | ProQR治疗上市公司Ⅱ | 化学修饰的编辑rna的单链寡核苷酸 |
| GB201808146D0 (en) | 2018-05-18 | 2018-07-11 | Proqr Therapeutics Ii Bv | Stereospecific Linkages in RNA Editing Oligonucleotides |
| JP2025525564A (ja) * | 2022-07-18 | 2025-08-05 | エフ. ホフマン-ラ ロシュ アーゲー | オリゴヌクレオチドの編集 |
| CN118853661A (zh) * | 2023-04-28 | 2024-10-29 | 时夕(广州)生物科技有限公司 | 编辑rna的方法、组合物及其应用 |
| WO2025015091A1 (en) * | 2023-07-11 | 2025-01-16 | Shape Therapeutics Inc. | Serpina1-targeting engineered guide rnas and polynucleotides |
| CN121794378A (zh) | 2023-08-15 | 2026-04-03 | 艾尔纳公司 | 用于rna编辑的化学修饰的反义寡核苷酸(aso)及组合物 |
| WO2025073902A1 (en) * | 2023-10-06 | 2025-04-10 | Eberhard Karls Universität Tübingen | Chemical and sequence optimisation of antisense oligonucleotides for adar-mediated rna editing |
| WO2025083268A1 (en) * | 2023-10-20 | 2025-04-24 | Airna Corporation | Chemically modified antisense oligonucleotides (asos) and compositions comprising the same for rna editing |
| WO2025155911A1 (en) * | 2024-01-18 | 2025-07-24 | Proqr Therapeutics Ii B.V. | Antisense oligonucleotides for the treatment of hurler syndrome |
| WO2025231085A1 (en) * | 2024-04-30 | 2025-11-06 | Airna Corporation | Site-directed editing of rna |
| WO2026077951A1 (en) | 2024-10-07 | 2026-04-16 | Vico Therapeutics B.V. | Improved aon for rna editing |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2019158475A1 (en) * | 2018-02-14 | 2019-08-22 | Proqr Therapeutics Ii B.V. | Antisense oligonucleotides for rna editing |
| WO2020001793A1 (en) * | 2018-06-29 | 2020-01-02 | Eberhard-Karls-Universität Tübingen | Artificial nucleic acids for rna editing |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN110352244B (zh) | 2016-09-01 | 2023-03-21 | ProQR治疗上市公司Ⅱ | 化学修饰的编辑rna的单链寡核苷酸 |
-
2021
- 2021-06-01 EP EP21177135.7A patent/EP4098745A1/en not_active Withdrawn
-
2022
- 2022-05-31 EP EP22735307.5A patent/EP4347827A1/en active Pending
- 2022-05-31 AU AU2022286618A patent/AU2022286618A1/en active Pending
- 2022-05-31 WO PCT/EP2022/064714 patent/WO2022253810A1/en not_active Ceased
- 2022-05-31 CA CA3221207A patent/CA3221207A1/en active Pending
- 2022-05-31 CN CN202280048791.8A patent/CN117616122A/zh active Pending
- 2022-05-31 JP JP2023574225A patent/JP2024522012A/ja active Pending
- 2022-05-31 KR KR1020237045375A patent/KR20240016350A/ko active Pending
- 2022-05-31 US US18/565,733 patent/US20250059534A1/en active Pending
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2019158475A1 (en) * | 2018-02-14 | 2019-08-22 | Proqr Therapeutics Ii B.V. | Antisense oligonucleotides for rna editing |
| WO2020001793A1 (en) * | 2018-06-29 | 2020-01-02 | Eberhard-Karls-Universität Tübingen | Artificial nucleic acids for rna editing |
Also Published As
| Publication number | Publication date |
|---|---|
| CN117616122A (zh) | 2024-02-27 |
| AU2022286618A9 (en) | 2023-12-14 |
| KR20240016350A (ko) | 2024-02-06 |
| WO2022253810A1 (en) | 2022-12-08 |
| EP4098745A1 (en) | 2022-12-07 |
| US20250059534A1 (en) | 2025-02-20 |
| CA3221207A1 (en) | 2022-12-08 |
| EP4347827A1 (en) | 2024-04-10 |
| AU2022286618A1 (en) | 2023-11-30 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2024522012A (ja) | Rnaに作用する内因性アデノシンデアミナーゼ(adar)による効率的および正確なrna編集のためのアンチセンスオリゴヌクレオチド(aso) | |
| US12545911B2 (en) | Single-stranded RNA-editing oligonucleotides | |
| CN110352244B (zh) | 化学修饰的编辑rna的单链寡核苷酸 | |
| JP2024544067A (ja) | Rnaに作用する内因性アデノシンデアミナーゼ(adar)による効率的および正確なrna編集のためのアンチセンスオリゴヌクレオチド(aso) | |
| EP4689107A1 (en) | Chemically modified antisense oligonucleotides for use in rna editing | |
| KR20250109237A (ko) | Rna 편집을 위한 화학적으로 변형된 안티센스 올리고뉴클레오티드(aso) 및 이를 포함하는 조성물 | |
| WO2025051946A1 (en) | Antisense oligonucleotides for the treatment of metabolic disorders | |
| CN121399260A (zh) | 用于治疗神经退行性疾病的反义寡核苷酸 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A529 | Written submission of copy of amendment under article 34 pct |
Free format text: JAPANESE INTERMEDIATE CODE: A529 Effective date: 20240130 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20250530 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20260324 |