JP4031033B2 - T細胞の改良トランスフェクション法 - Google Patents
T細胞の改良トランスフェクション法 Download PDFInfo
- Publication number
- JP4031033B2 JP4031033B2 JP53351096A JP53351096A JP4031033B2 JP 4031033 B2 JP4031033 B2 JP 4031033B2 JP 53351096 A JP53351096 A JP 53351096A JP 53351096 A JP53351096 A JP 53351096A JP 4031033 B2 JP4031033 B2 JP 4031033B2
- Authority
- JP
- Japan
- Prior art keywords
- cells
- cell
- nucleic acid
- acid molecule
- transfection
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
Images






















Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2818—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against CD28 or CD152
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2809—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against the T-cell receptor (TcR)-CD3 complex
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K48/00—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- General Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Zoology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Animal Behavior & Ethology (AREA)
- Plant Pathology (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Pharmacology & Pharmacy (AREA)
- Oncology (AREA)
- Communicable Diseases (AREA)
- Virology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
Description
本明細書に記載した研究は、NMRDC認可61153NAE.4120.001.1402により一部支持されている。従って、米国政府は、本発明の権利を有する。
発明の背景
真核細胞における外来DNAの発現は、遺伝子発現の調節から遺伝子導入に基づく治療法まで広範囲にわたる生物学的主題の研究を可能にするものである。哺乳動物細胞へ遺伝子導入する多くの方法が考案されている。これらには、クローン化レトロウイルスベクターによるインビボおよびインビトロ感染(Shimotohno,K.,およびTemin,H.M.(1981)Cell 26:67-77;Cone,R.D.Mおよびulligan,R.C.(1984)Proc.Natl.Acad.Sci.USA 81:6349-6353;Dubensky,T.W.,Campbell,B.A.,およびVillareal,L.P.(1984)Proc.Natl.Acad.Sci.USA 81:7529-7533;Seeger,C.,Ganem.D.およびVarmus,H.E.(1984)Proc.Natl.Acad.Sci.USA 81:5849-5852)、リン酸カルシウムによるDNAの共沈(Chu,G.,およびSharp,P.(1981)Gene 13;197-202;Benvenistry,N.およびReshef,L.(1986)Proc Natl.Acad.Sci.USA 83:9551-9555)、DANのリポソーム封入(Felgner,P.I.,およびRingold,G.M.(1989)Nature 337:387-388;Kaneda,Y.,Iwai,K.,およびUchida,T.(1989)Science 243:375-378)、プラスミドDNAの直接注入(Walff,J.A.,Malone,R.W.,Williams,P.,Chong,W.,Acsadi,G.,Jani,A.,およびFelgner,P.L.(1991)Science 247:1465-1468)、DEAE−デキストラン(McCutchan,J.H.,およびPagano,J.S.(1968)J.Natl.Cancer Inst 41:351-357)、エレクトロポレーション(Neumann,E.,Schaefer-Ridder,M.Wang,Y.,およびHofschneider,P.H.(1982)EMBO J.1:841-845;Cann,A.J.,Koyanagi,Y.,およびChen,I.S.Y.(1988)Oncogene 3:123-128)、および細胞および組織のDNA−被覆粒子ボンバードメント(Yang,N-S.,Burkholder,J.,Roberts,B.,Martinell,B.,およびMcCabe,D.(1990)Proc.Natl.Acad.Sci.USA 87:9568-9572)がある。
遺伝子を含む外来核酸分子での様々な細胞タイプのトランスフェクションは、外来遺伝子の発現には有効であるが、個体から得られた一次Tリンパ球類、例えば、末梢血Tリンパ球は、外来DNAのトランスフェクションおよび発現に対して不応であることが分かっている。一次Tリンパ球類はまた、増殖のために第1刺激を与えたときに、導入した核酸の発現に対して不応であることも分かっている。そのため、外来DNAを効率良く一次T細胞へ導入でき、T細胞中で外来DNAを発現させるシステムに対する要望が依然としてある。
概要
本発明は、遺伝子のT細胞中での発現を古典的なトランスフェクション技術と比較して高めた、遺伝子を含む核酸分子でのT細胞の改良トランスフェクション法を提供する。本発明の方法は、特に、古典的なトランスフェクション技術には不応である一次T細胞のトランスフェクションに有用である。この方法は、増殖T細胞をその増殖T細胞を刺激する1またはそれ以上の作用物質と、核酸分子をそのT細胞に導入する前に、接触させることを含む。本発明の一実施態様では、T細胞を、T細胞に対する一次活性化シグナルを提供する第1作用物質およびコスティミュラトリー・シグナル(costimulatory signal)を提供する第2作用物質を組み合わせたもので刺激する。本発明の方法は、特に、遺伝子治療における様々な応用を有する。
【図面の簡単な説明】
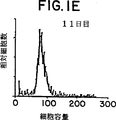
図1は、1μg/mlの抗CD3被覆プレートおよび抗CD28で刺激した(0日目)後、1、5、7、9および11日目のCD28+T細胞の相対細胞数および細胞容量グラフ表示を表す。
図2は、抗CD28mAb9.3(αCD28)の存在下、飽和量の固定化抗CD3mAbG19−4(αCD3)で刺激した、新たに単離したCD28+T細胞の成長曲線を表す。
図3は、飽和量の固定化抗CD3mAbG19−4(αCD3)および抗CD28mAb9.3(αCD28)による第1刺激(1日目)または第2刺激(8日目)後、0、6、24および72時間培養した一次CD28+T細胞中のEts−1(ETS−1)およびヒト白血球抗原(HLA)mRNAのレベルを示すノーザン・ブロットである。
図4は、まず休止T細胞(Rest)を抗CD3および抗CD28(αCD3+αCD28)と共に2日間インキュベーションし、次いで、抗CD28単独(αCD28)で3日間インキュベーションし、トランスフェクションの10時間前に刺激(第1刺激)し、T=0でトランスフェクションし、トランスフェクション後30時間で再刺激(第2刺激)し、10時間後に収集するというトランスフェクションプロトコールの一例の略式表示である。
図5は、トランスフェクションの10時間前にホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)または、ならし培地単独(MED)で刺激し、トランスフェクション後40時間で収集した、RSV−CATでトランスフェクションさせた、指数増殖しているT細胞由来の細胞抽出物で実施したCATアッセイの結果を表す。正規化したCAT活性は(アセチル化%/タンパク質mg)×50として表す。
図6(パネルA〜D)は、アクリジンオレンジ染色した一次T細胞のフロー・サイトメトリー分析および下記の条件で培養した一次T細胞の増殖アッセイの結果を示す:未処理休止一次T細胞(パネルA)、抗CD3および抗CD28で3日間刺激したT細胞(パネルB)、抗CD3および抗CD28で3日間刺激し、その後、新たな培地で更に3日間インキュベーションしたT細胞(パネルC)、または抗CD3および抗CD28で3日間刺激し、ホルボール−12,13−ジブチレート(PDBU)とイオノマイシンで10時間刺激し、その後、新たな培地で更に2日間と14時間インキュベーションしたT細胞(パネルD)。アクリジンオレンジ染色した細胞のフロー・サイトメトリー分析のグラフ表示は、細胞のDNAおよびRNA含量を示し、これは、細胞周期のG0(%G0)、G1(%G1)、およびS/G2M(%S/G2M)期における細胞数の指標である。
図7は、RSV−CATでトランスフェクションさせたT細胞で、トランスフェクションの10時間前(1°)に培地単独(MED)またはホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)で刺激し、トランスフェクション後30時間(2°)で培地単独(MED)、ホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)、抗CD3および抗CD28抗体(αCD3+αCD28)または、ならし培地(COND MED)で刺激し、10時間後に収集した指数増殖しているT細胞由来の細胞抽出物を用いて実施したCATアッセイの結果を表す。正規化したCAT活性は(アセチル化%/タンパク質mg)×50として表す。
図8は、HIV−1−CAT発現構築物でトランスフェクションさせたT細胞で、トランスフェクションの10時間前(1°)に培地単独(MED)、ホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)または抗CD3および抗CD28抗体(αCD3+αCD28)で刺激し、トランスフェクション後30時間(2°)で培地単独(MED)、ホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)、抗CD3および抗CD28抗体(αCD3+αCD28)または、ならし培地(COND MED)で刺激し、10時間後に収集した指数増殖しているT細胞由来の細胞抽出物を用いて実施したCATアッセイの結果を表す。正規化したCAT活性は(アセチル化%/タンパク質mg)×50として表す。
図9は、ノーザン・ブロット分析により測定した、培地単独(MED)で12時間または抗CD3抗体(CD3)で1、6、12および24時間培養したCD28+T細胞中の総RNA含量(パネルA)またはIL−2の場合のmRNAレベル(パネルB)およびHLAの場合のmRNAレベル(パネルC)を表す。パネルDは、培地単独(MED)と共に12時間または抗CD3抗体(CD3)と共に1、6、12、24または48時間インキュベーションしたT細胞により産生されたIL−2の量、および抗CD3と共に培養後48時間での細胞周期のS期、G2またはM期の細胞割合を示す。
図10は、IL2−CAT発現構築物でトランスフェクションさせたT細胞で、トランスフェクションの10時間前(1°)に培地単独(MED)またはホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)で刺激し、トランスフェクション後30時間(2°)で培地単独(MED)、ホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)、抗CD3および抗CD28抗体(αCD3+αCD28)または、ならし培地(COND MED)で刺激し、10時間後に収集した指数増殖しているT細胞由来の細胞抽出物を用いて実施したCATアッセイの結果を表す。正規化したCAT活性は(アセチル化%/タンパク質mg)×50として表す。
図11は、RSV−CATでトランスフェクションさせた休止T細胞で、トランスフェクションの10時間前に培地単独(1日目:MED)、抗CD28(1日目:αCD28)、スタフィロコッカスエンテロトキシンA(1日目:SEA)、またはホルボール−12,13−ジブチレートおよびイオノマイシン(1日目:PDBU+IONO)で、または、抗CD3および抗CD28で5日間処理し、次いで、トランスフェクション前の10時間、ならし培地(6日目:MED)またはホルボール−12,13−ジブチレートおよびイオノマイシン(6日目:PDBU+IONO)で処理した休止T細胞由来の細胞抽出物を用いて実施したCATアッセイの結果を表す。正規化したCAT活性は(アセチル化%/タンパク質mg)×50として表す。
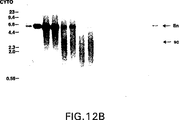
図12は、増殖T細胞をRSV−CATで、またはプラスミドなし(MOCK)でトランスフェクションさせた後0、6、24または48時間の増殖T細胞で、トランスフェクションの10時間前にホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)または、ならし培地単独(MED)で刺激し、RSV−CATのCATコード領域由来のEcoRIフラグメントとハイブリダイズさせたT細胞から抽出した核DNA(NUCLEAR、パネルA)または細胞質DNA(CYTO、パネルB)のサザン・ブロットのオートラジオグラムを示す。1(1c)、10(10c)、100(100c)、および1000(1000c)コピーのRSV−CAT/細胞に相当するプラスミドDNAを対照として使用した。(lin)鎖状プラスミド;(sc)スーパーコイル状プラスミド。分子量マーカーからのフラグメントのサイズ(キロベース、kb)は、サザン・ブロットの左側に表す。
図13は、RSV−CATでトランスフェクションさせたT細胞で、トランスフェクションの10時間前(1°)に培地単独(MED)またはホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)で刺激し、トランスフェクション後30時間(2°)で培地単独(MED)、ホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)、抗CD3および抗CD28抗体(αCD3+αCD28)または、ならし培地(COND MED)で刺激し、RSV−CATのCATコード領域由来のEcoRIフラグメントとハイブリダイズさせた指数増殖しているT細胞から抽出した核DNAのサザン・ブロットのオートラジオグラムを示す。1(1c)、10(10c)、100(100c)、および1000(1000c)コピーのRSV−CAT/細胞に相当するプラスミドDNAを対照として使用した。(lin)鎖状プラスミド;(sc)スーパーコイル状プラスミド。M:分子量マーカー。
図14は、IL2−CAT発現構築物でトランスフェクションさせたT細胞で、トランスフェクションの10時間前(1°)に培地単独(MED)またはホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)で刺激し、トランスフェクション後30時間(2°)で培地単独(MED)、ホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)、抗CD3および抗CD28抗体(αCD3+αCD28)または、ならし培地(COND MED)で刺激し、IL2−CATのCATコード領域由来のEcoRI/BamHIフラグメントとハイブリダイズさせた指数増殖しているT細胞から抽出した核DNAのサザン・ブロットのオートラジオグラムを示す。1、10、100、および1000コピーのIL2−CAT/細胞に相当するプラスミドDNAを対照として使用した。(lin)鎖状プラスミド;(sc)スーパーコイル状プラスミド。M:分子量マーカー。
図15は、32P−放射性標識した鎖状化RSV−CATでトランスフェクションさせたT細胞で、トランスフェクションの10時間前にホルボール−12,13−ジブチレートおよびイオノマイシン(PDBU+IONO)または、ならし培地単独(MED)で刺激し、トランスフェクション直後(0)、トランスフェクション後6、24または48時間で収集したT細胞の核または細胞質から回収した毎分のカウント数(cpm)の割合を表す。毎分のカウント数の割合は、トランスフェクション細胞足す毎分のカウント数の総数に比例して計算される。
図16は、RSV−CATまたはHIV−1−CATでトランスフェクションさせた増殖T細胞で、培地単独(MED)、単球(MONO)、スタフィロコッカスエンテロトキシンA(SEA)、または単球およびスタフィロコッカスエンテロトキシンA(MONO+SEA)でトランスフェクション前時間処理し、トランスフェクション後40時間で収集した増殖T細胞由来の細胞抽出物を用いて実施したCATアッセイの結果を表す。正規化したCAT活性は(アセチル化%/タンパク質mg)×50として表す。
詳細な説明
本発明は、遺伝子をT細胞中で発現させるための、遺伝子を含む核酸分子によるT細胞の改良トランスフェクション法を提供する。本発明の方法は、増殖T細胞をその増殖T細胞を刺激する少なくとも1つの作用物質と、核酸分子をそのT細胞に導入する前に接触させ、そうしてその遺伝子をそのT細胞中で発現させることを含む。
本発明の方法は、少なくともその一部において、遺伝子を含む核酸により一次T細胞をトランスフェクションしても一次T細胞が増殖しておらず、また更に、T細胞中で一次活性化シグナルおよびコスティミュラトリー・シグナルを誘起する作用物質などの刺激性作用物質で刺激しない場合にはその遺伝子はほとんど発現されないという観察に基づくものである。T細胞は、好ましくは、核酸をT細胞に導入する約10時間前に刺激性作用物質と接触させる。こうして、増殖T細胞を、遺伝子を含む核酸をそのT細胞に導入する前に刺激することにより、T細胞中での外来遺伝子の有意な発現を達成できる。
従って、本発明は、遺伝子をT細胞中で発現させるための、遺伝子を含む核酸分子によるT細胞の改良トランスフェクション法を提供する。古典的なT細胞トランスフェクション法に優る本発明方法により提供される改良点は、核酸分子を増殖T細胞に導入する前(例えば、数時間前)にT細胞を刺激性作用物質と接触させることを含む。本発明の方法により、在来のトランスフェクション技術よりもずっと高い、T細胞に導入した遺伝子の発現が可能である。本発明の方法は、特に、対象となる遺伝子を一次T細胞に導入し、かつ発現させるのに有用である。そのため、本発明の具体的な実施態様では、T細胞を被験体から入手し、本発明の方法に従い核酸分子によりインビトロでトランスフェクションさせ、宿主に再度与える。対象となる遺伝子は、タンパク質をコードする遺伝子、またはアンチセンス分子またはリボザイムなどの機能性RNA分子をコードする遺伝子であり得る。対象となる遺伝子は、T細胞を保護するタンパク質類、T細胞にとって毒性であるタンパク質類、またはT細胞から分泌されて他の細胞に作用するタンパク質類を含む対象となるあらゆるタンパク質をコードし得る。従って、本発明の方法は、例えば、遺伝子療法、T細胞機能の変更およびT細胞中でのタンパク質の産生に応用できる。
1.本発明の方法に従いトランスフェクションさせることができるT細胞
本発明は、遺伝子を含む核酸分子でT細胞をトランスフェクションさせ、その遺伝子をT細胞中で発現させる方法に関する。用語“T細胞”は、当分野で定義されているTリンパ球を表し、胸腺細胞、未熟Tリンパ球、成熟Tリンパ球、休止Tリンパ球、または活性化Tリンパ球を含むことを意図する。T細胞は、CD4+T細胞、CD8+T細胞、CD4+CD8+T細胞、またはCD4-CD8-T細胞であることができる。T細胞はまた、Tヘルパー1(Th1)またはTヘルパー2(Th2)細胞などのTヘルパー細胞であることもできる。CD4などの少なくとも1つのマーカーにより互いに異なるT細胞は、本明細書ではT細胞の“サブセット”と称する。
T細胞は、純粋なT細胞集団であることができ、あるいは、T細胞は、B細胞および/または他の末梢血細胞などの異なるタイプの細胞をもつ集団中にあることもできる。T細胞は、CD4+T細胞などのT細胞サブセットの純粋な集団であることができ、またT細胞の異なるサブセットを含むT細胞集団であることもできる。本発明の他の実施態様では、T細胞は、長期間培養維持されるT細胞クローンである。T細胞クローンは、様々な度合で形質転換させることができる。特定の実施態様では、T細胞は、培養中無期限に増殖するT細胞クローンである。
本発明の好ましい実施態様では、T細胞は、一次T細胞である。用語“一次T細胞”は、長期間培養維持されるT細胞とは反対に、個体から得られたT細胞を含むことを意図する。そのため、一次T細胞は、好ましくは、被験体から得られた末梢血T細胞である。一次T細胞集団は、ほとんど一種のT細胞サブセットから構成され得る。あるいは、一次T細胞集団は、異なるT細胞サブセットから構成されていてもよい。
T細胞は、健常個体由来のものであることができ、またあるいは、T細胞は、疾病をもった個体由来のものであってもよい。この疾病とは、ウイルス感染、細菌感染またはその他微生物による感染から起こるような感染性疾患であり得る。特定の実施態様では、T細胞は、ヒト免疫不全ウイルス(HIV)に感染した個体由来のものである。本発明のまた別の実施態様ではT細胞は、自己免疫疾患を罹病している被験体、またはこれを受け易い被験体由来のものである。T細胞は、ヒト起源、ハツカネズミ起源、またはその他あらゆる哺乳動物種のものであり得る。
本発明の方法によれば、活発に増殖しているT細胞に核酸分子を導入する。T細胞を様々な作用物質、例えば、T細胞に一次活性化シグナルおよびコスティミュラトリー・シグナルを与える作用物質類の組み合わせ、と接触させることにより、T細胞を刺激して、増殖させることができる。T細胞を刺激して増殖させるのに使用され得る作用物質類は、当分野ではよく知られており、下記2章に記載している。増殖のために刺激されるT細胞は、細胞拡張、クランピング(clumping)、および培養培地の酸性化を特徴とする。このため、T細胞増殖は、例えば、コウルター計数器(Coulter Counter)などによるT細胞のサイズ検査または容量測定により立証できる。休止T細胞は、平均直径約6.8ミクロンを有する。初期活性化および刺激後のT細胞平均直径は、4日目に12ミクロンに上昇し、およそ6日目に減少し始めるであろう。更に、T細胞増殖は、トリチウム化チミジン摂取などの当分野の標準技術によっても評価できる。
2.刺激性作用物質
本発明の方法は、核酸分子を増殖T細胞に導入する前に、増殖T細胞を少なくとも1つの刺激性作用物質と接触させることを含む。用語“刺激性作用物質”は、T細胞中へトランスフェクションした核酸分子に含まれる遺伝子の発現レベルが、核酸分子をT細胞に導入する前にT細胞を刺激性作用物質と接触させた場合に、核酸分子を導入する前に刺激性作用物質と接触させなかったT細胞におけるよりも高いように、T細胞にシグナルを提供する作用物質類を含むことを意図する。
本発明の具体的な実施態様では、刺激性作用物質は、T細胞に一次活性化シグナルを提供する作用物質である。用語“一次活性化シグナル”は、典型的には、TCR/CD3複合体を介して誘発される、T細胞の活性化を誘起するシグナル類を含むことを意図する。T細胞の活性化は、コスティミュラトリー・シグナルなどの第2シグナル受容時にT細胞を増殖および分化させるようにT細胞を誘導するようなT細胞の修飾を含むことを意図する。具体的な実施態様では、一次活性化シグナルは、T細胞受容体またはT細胞受容体と関連するCD3複合体を接触させる作用物質により提供される。好ましい実施態様では、この作用物質は、CD3に反応性の抗体、例えば、モノクローナル抗体OKT3(American Type Culture Collection, Rockville,MDから入手;番号CRL8001)である。本発明のその他の実施態様では、刺激性作用物質は、T細胞上でCD2複合体を刺激する作用物質、例えば、抗体の組み合わせ、例えば、T11.3+T11.1またはT11.3+T11.2である(例えば、Meuer,S.C.等,(1984)Cell36:897-906参照)。
本発明の好ましい実施態様では、T細胞中で一次活性化シグナルおよびコスティミュラトリー・シグナルの両方を刺激する作用物質の組み合わせでT細胞を刺激する。用語“共刺激性(コスティミュラトリー)作用物質(costimulatory agent)”は、一次活性化シグナルを受けたT細胞(例えば、活性化T細胞)を刺激して増殖させるかまたはIL−2、IL−4、またはインターフェロン−γなどのサイトカインを分泌させるようなコスティミュラトリー・シグナルをT細胞に提供する作用物質類を含むことを意図する。特定の実施態様では、共刺激性作用物質はCD28またはCTLA4分子とT細胞表面で相互作用する。更に一層具体的な実施態様では、コスティミュラトリー・シグナルは、CD28またはCTLA4のリガンド、例えば、Bリンパ球抗原B7−1またはB7−2である。用語“CD28の天然リガンドの刺激性形態”は、B7−1およびB7−2分子、それらのフラグメント、またはそれらの修飾形を含むことを意図し、これらは、T細胞にコスティミュラトリー・シグナルを提供する能力がある。CD28の天然リガンドの共刺激性形態は、例えば、活性化末梢血リンパ球をCD28の天然リガンドの一形態と接触させ、標準T細胞増殖アッセイを実施することにより、同定できる。従って、CD28の天然リガンドの刺激性形態は、T細胞の増殖を刺激する能力がある。CD28/CTLA4の天然リガンドの刺激性形態は、例えば、PCT公開公報WO95/03408に記載されている。
核酸分子をT細胞に導入する前に、T細胞刺激のために使用できるその他の作用物質類には、T細胞活性化および/または共刺激(コスティミュレーション)に関連する1またはそれ以上の細胞内シグナル伝達経路を刺激する作用物質類がある。本発明の好ましい実施態様では、刺激性作用物質はイオノマイシンまたはA23187などのカルシウムイオノホアである。あるいは、刺激性作用物質は、ホルボールエステルなどのタンパク質キナーゼCを刺激する作用物質であることもできる。好ましいホルボールエステルは、ホルボール−12,13−ジブチレートである。本発明の更に一層好ましい実施態様では、核酸分子でのトランスフェクションの前に、T細胞をカルシウムイオノホアとホルボールエステルとの組み合わせと接触させる。刺激性作用物質は、タンパク質チロシンキナーゼを活性化する作用物質であってもよい。タンパク質チロシンキナーゼを刺激する好ましい作用物質はペルバナデートである(O'Shea,J.J.,等,(1992)Proc.Natl.Acad.Sci.USA 89:10306)。
本発明の更にその他の実施態様では、刺激性作用物質はポリクローナルアクチベーターである。ポリクローナルアクチベーターには、T細胞の原形質膜上で発現する糖タンパク質に結合する作用物質類があり、フィトヘマグルチニン(PHA)、コンカナバリン(Con A)およびアメリカヤマゴボウマイトジェン(PWM)などのレクチン類がある。
クローンに特異的活性化シグナルを提供することにより、T細胞集団においてT細胞のあるクローンのみを選択的にトランスフェクションさせることが可能である。T細胞クローンの特異的活性化は、例えば、抗原提示細胞により与えられる特異的抗原を用いて達成できる。
本方法の更にその他の実施態様では、刺激性作用物質は、IL−2などのリンホカインである。リンホカインは、好ましくは、T細胞を刺激するその他の作用物質、例えば、T細胞に一次活性化シグナルを提供する作用物質、と組み合わせて使用する。従って、本発明の好ましい実施態様では、核酸分子がT細胞中で発現するよう、核酸分子でT細胞をトランスフェクションさせる前に、T細胞を、T細胞に一次活性化シグナルを提供する作用物質(例えば、抗CD3抗体)と有効量のIL−2を組み合わせたものと接触させる。
使用できるその他の刺激性作用物質には、スーパー抗原類がある。本明細書で定義した用語“スーパー抗原”は、T細胞増殖を刺激する能力がある細菌エンテロトキシン類またはその他の細菌タンパク質を含むことを意図する。スーパー抗原類には、SEA、SEB、SEC、SED、およびSEEなどのスタフィロコッカスエンテロトキシン(SE)がある。スーパー抗原類はまた、レトロウイルススーパー抗原のようにウイルス起源であることができる。
当分野で既知の、または本明細書に記載したT細胞刺激アッセイを用いて同定できる、単独またはその他の作用物質と併用するかのいずれかでT細胞を刺激する能力のある別の作用物質類もまた、本発明の範囲内である。核酸分子をT細胞に導入する前にT細胞を刺激するために、上記作用物質類のあらゆる組み合わせが使用できる。
刺激性作用物質は、溶液で、または固体表面に結合させて使用できる。この固体表面は、例えば、組織培養皿またはビーズの表面であり得る。刺激性作用物質の性質にもよるが、固体表面への結合は、当分野でよく知られている方法により実施できる。例えば、市販の架橋剤(Pierce,Rockford IL)を用いてタンパク質を細胞表面に化学的に架橋させることもでき、また、4℃で一晩インキュベーションすることによりプラスチック上に固定化することもできる。数種の作用物質を用いてT細胞を刺激する場合、その幾つかが溶液状であってもよく、またその幾つかが固体支持体に結合させたものであってもよい。好ましい実施態様では、固相結合抗CD3抗体と可溶性抗体CD28抗体を組み合わせたものでT細胞を刺激する。
T細胞に添加される刺激性作用物質の特定用量は、刺激性作用物質の種類によって変わる。典型的には、当分野で報告されているように、T細胞を刺激して増殖およびサイトカインを分泌させるのに使用される用量と同量で刺激性作用物質を使用する。
3.トランスフェクション前刺激のプロトコール
本発明の特定の実施態様では、遺伝子を含む核酸分子をT細胞に導入する前にT細胞を刺激性作用物質と接触させる。本発明の好ましい実施態様では、核酸分子をT細胞に導入する少なくとも2時間前にT細胞を刺激性作用物質と接触させる。本発明のその他の実施態様では、核酸分子をT細胞に導入する少なくとも4時間前にT細胞を刺激性作用物質と接触させる。本発明の別の実施態様では、核酸分子をT細胞に導入する少なくとも6時間前にT細胞を刺激性作用物質と接触させる。本発明の別の実施態様では、核酸分子をT細胞に導入する少なくとも8時間前にT細胞を刺激性作用物質と接触させる。別の実施態様では、トランスフェクションの多くとも約2時間前、トランスフェクションの多くとも約4時間前、トランスフェクションの多くとも約6時間前、トランスフェクションの多くとも約8時間前、トランスフェクションの多くとも約10時間前、トランスフェクションの多くとも約12時間前、トランスフェクションの多くとも約14時間前、トランスフェクションの多くとも約16時間前、トランスフェクションの多くとも約18時間前、トランスフェクションの多くとも約20時間前、トランスフェクションの多くとも約22時間前、トランスフェクションの多くとも約24時間前に、T細胞を刺激性作用物質と接触させる。
本発明のより一層好ましい実施態様では、核酸分子をT細胞に導入する約1時間から24時間前にT細胞を刺激性作用物質と接触させる。本発明の最も好ましい実施態様では、対象となる遺伝子を含む核酸分子をトランスフェクションする約5から15時間前、例えば約10時間前に増殖T細胞を少なくとも1つの刺激性作用物質と接触させる。
本方法の一実施態様では、増殖T細胞を少なくとも1つの刺激性作用物質と接触させ、更に核酸分子でトランスフェクションさせる。本方法のその他の実施態様では、非増殖T細胞を増殖させるために刺激し、それから、本発明の方法に従いトランスフェクションさせる前に少なくとも1つの刺激性作用物質と接触させる。非増殖T細胞は、上記“刺激性作用物質類”のところで記載のような当分野でよく知られている作用物質を用いて刺激し、増殖させる。好ましい作用物質類には、一次活性化シグナルを提供する作用物質およびコスティミュラトリー・シグナルを提供する作用物質の組み合わせがある。T細胞の増殖を刺激するその他の好ましい作用物質類の組み合わせには、ホルボールエステルとカルシウムイオノホア、またはPMAとIL−2の組み合わせがある。
一次T細胞のトランスフェクション法の最も好ましい実施態様によれば、休止一次T細胞を、まず、T細胞の増殖を刺激する少なくとも1つの作用物質、例えば、カルシウムイオノホアとホルボールエステルの組合わせ物と接触させる。T細胞増殖誘起後、およそ3から8時間の間で、好ましくはおよそ5時間で、増殖T細胞を、そのT細胞を刺激する少なくとも1つの作用物質と接触させる。最後に、刺激性作用物質と接触後、2から15時間、好ましくは約10時間で、T細胞を対象となる遺伝子を含む核酸分子でトランスフェクションさせ、そうしてその遺伝子をT細胞中で発現させる。その他の実施態様では、T細胞のトランスフェクション後に、T細胞を更に、T細胞を刺激する作用物質と接触させる。
被験体から一次T細胞を得るために、末梢血単核細胞を被験体から単離し、密度勾配遠心分離、例えば、Ficoll/Hypaqueにより精製できる。具体的な実施態様では、次いで、精製した末梢血細胞を本発明の方法に従い核酸分子でトランスフェクションさせる。本方法の他の実施態様では、トランスフェクションさせる前に、更に特定の細胞タイプ中で末梢血単核細胞を豊富化する。単球は、例えば、プラスチック上に付着させることにより除去できる。所望ならば、更に、CD4+T細胞集団を、市販のモノクローナル抗体(mAb)(例えば、抗CD14(Mo2)、抗CD11b(Mo1)、抗CD20(B1)、抗CD16(3G8)および抗CD8(7PT 3F9)mAb)を用いるモノクローナル抗体(mAb)および抗マウス−Ig被覆化磁気ビーズを用いて残りの単球、B細胞、NK細胞、およびCD8+T細胞から分離することにより、豊富化できる。本発明の方法は、また、CD4+T細胞のサブセット、例えば、CD4+CD45RA+(天然CD4+T細胞)およびCD4+CD45RO+(記憶T細胞)T細胞サブセットにも応用できる。これらは、上記のようにして製造でき、CD4+CD45RA+細胞の製造には更に抗CD45RO抗体(UCHL1)を使用し、またCD4+CD45RO+T細胞の製造には更に抗CD45RA抗体(2H4)を使用する。
精製効率は、T細胞の特定のサブセットを認識する抗CD3、抗CD4、抗CD8、抗CD14mAb、または更なる抗体、その後、フルオレセインイソチオシアナートコンジュゲート化ヤギ抗マウス免疫グロブリン(Fisher,Pittsburgh,PA)、または他の第2抗体を用いるフロー・サイトメトリー(Coulter, EPICS Elite)により分析できる。
本発明の好ましい実施態様では、本発明の方法は、実施例の章で記載のように、更に、T細胞のトランスフェクション後にT細胞を刺激してインビトロで拡張(expand)させることができる。具体的な実施態様では、T細胞を、一次活性化シグナルを提供する作用物質、例えば抗CD3、およびコスティミュラトリー・シグナルを提供する作用物質、例えば抗CD28抗体と共にインキュベーションする。2日後、細胞を新しい培地で希釈し、コスティミュラトリー・シグナルを提供する作用物質を培養物に加える。次いで、T細胞を、そのサイズ分布が休止細胞プロフィール近くに戻る(約10日)まで2日毎にカウントし、サイズ測定し、新たな培地で希釈する。その後、T細胞を、一次活性化シグナルを提供する作用物質およびコスティミュラトリー・シグナルを提供する作用物質で再刺激できる。T細胞サイズ測定およびカウントは、本明細書に記載のように、コウルター計数器を用いて実施できる。
更に一層好ましい実施態様では、T細胞は一次T細胞である。従って、T細胞を被験体から入手し、本発明の方法に従いトランスフェクションさせ、インビトロで拡張させることができる。本発明のその他の実施態様では、トランスフェクションさせ、かつ拡張させたT細胞を被験体に再度投与する。被験体に投与する前に、例えば、勾配遠心分離により更にT細胞を精製するのが好ましい場合もある。
4.T細胞のトランスフェクション
本発明は、遺伝子を含む核酸でT細胞をトランスフェクションさせ、その遺伝子をT細胞中で発現させる方法に関する。“T細胞をトランスフェクションさせる”の用語は、核酸分子をT細胞に導入できるあらゆる手段を含むことを意図する。用語“トランスフェクション”は、エレクロトポレーション、リン酸カルシウム沈殿、DEAE−デキストラン処理、リポフェクション、マイクロインジェクション、およびウイルス感染を含む、核酸の哺乳動物細胞への導入に有用な様々な技術を包含する。哺乳動物細胞をトランスフェクションさせる適切な方法は、Sambrook等(Molecular Cloning;A Laboratory Manual,2nd Edition,Cold Spring Harbor Laboratory press(1989))およびその他の実験書に見ることができる。
本発明の好ましい実施態様では、対象となる遺伝子をコードする核酸分子をウイルスベクターを用いてT細胞に導入する。このようなウイルスベクターには、例えば、組換えレトロウイルス、アデノウイルス、アデノ随伴ウイルス、および単純ヘルペスウイルス1がある。レトロウイルスベクターおよびアデノ随伴ウイルスベクターは、インビボでの特にヒトへの外来遺伝子導入の優れた組換え遺伝子運搬システムであると一般に理解されている。あるいは、それらは、外来遺伝子をエクスビボでT細胞に導入するのにも使用できる。これらのベクターは、遺伝子をT細胞へ効率良く運ぶものであり、導入された核酸は、宿主細胞の染色体DNAに安定に組込まれる。レトロウイルスの使用に当たっての重要な必須要件は、特に、細胞集団中で野生型ウイルスが蔓延する可能性について、その使用の安全性を確実にすることである。複製欠損性レトロウイルスのみを産生する分化した細胞系列(“パッケージング細胞”と称する)の開発は、遺伝子治療目的の遺伝子導入における使用について十分に特徴付けられている(検討のためMiller,A.D.(1990)Blood 76:271参照)。従って、レトロウイルスコード配列(gag、pol、env)の一部を対象となる遺伝子で置き換えてレトロウイルスを複製欠損性にすることにより組換えレトロウイルスを構築できる。この場合、複製欠損性レトロウイルスは、ビリオン中にパッケージされるので、これを、標準技術によりヘルパーウイルスを使用して、標的細胞を感染させるのに使用できる。組換えレトロウイルスを産生し、かかるウイルスを用いてインビトロまたはインビボで細胞を感染させるプロトコールは、Current Protocols in Molecular Biology,Ausubel,F.M.等(編)Greene Publishing Associates,(1989),Sections 9.10-9.14およびその他の標準実験マニュアルに見ることができる。適切なレトロウイルスの例には、当分野ではよく知られているpLJ、pZIP、pWEおよびpEMがある。同種指向性および両種指向性レトロウイルス系の両方を製造するのに適切なパッケージングウイルス系列の例には、ΨCrip、ΨCre、Ψ2およびΨAmがある。
更に、ウイルス粒子表面のウイルスパッケージングタンパク質を修飾してレトロウイルスの感染スペクトル、更にレトロウイルス基盤のベクターの感染スペクトルを制限することが可能であることが示されている(例えば、PCT公開公報WO93/25234およびWO94/06920参照)。例えば、レトロウイルスベクターの感染スペクトルの修飾法は、細胞表面抗原に特異的な抗体をウイルスのenvタンパク質に結合させる(Roux等(1989)PNAS 86:9079-9083;Julan等(1992)J.Gen.Virol 73:3251-3255;およびGoud等(1983)Virology 163:251-254);または細胞表面受容体リガンド類をウイルスenvタンパク質に結合させる(Neda等(1991)J.Biol.Chem.266:14143-14146)ことを含む。結合は、タンパク質またはその他の様々なもの(例えば、envタンパク質をアジアロ糖タンパク質に変換するラクトース)との化学的架橋形態であることができ、同じく、融合タンパク質(例えば、単鎖抗体/env融合タンパク質)を作成することによる。従って、本発明の具体的な実施態様では、適切な調節配列に作動可能に連結させた対象となる遺伝子を含む核酸分子を含有するウイルス粒子を、例えば、上記の方法に従い修飾することにより、それらはT細胞のサブセットを特異的に標的とすることができる。例えば、ウイルス粒子を、あるタイプのT細胞に特異的な、表面分子に対する抗体で被覆できる。特に、T細胞のCD4分子を認識するウイルス粒子抗体に連結させることにより選択的にCD4+T細胞を標的とすることが可能である。そのため、CD4+T細胞の感染は、CD8+T細胞の感染よりも優先的に起こるであろう。この方法は、特に、T細胞の特定のサブセットのみをトランスフェクションさせたい場合に有用である。一次T細胞を含むT細胞に対象となる遺伝子を含む核酸分子を導入し、かつ発現させるための更なるレトロウイルス系は、Kasid,A.等(1990)Proc.Natl.Acad.Sci.USA.87,473;Morecki,S.等(1991)Cancer Immunol.Immunother.32,342;Culver,K.等(1991)Proc.Natl.Acad.Sci.USA.,88,3155;およびFiner,M.H.等(1994)Blood,83,43に記載されている。
本発明の有用なその他のウイルス遺伝子運搬システムは、アデノウイルス由来ベクターである。アデノウイルスのゲノムは、それが対象となる遺伝子産物をコードし発現するが、正常な細胞溶解ウイルス生活環で見られる複製能力については不活性化されているように、操作することができる。例えば、Berkner等(1988)Bio Techniques 6:616;Rosenfeld等(1991)Science 252:431-434;およびRosenfeld等(1992)Cell 68:143-155参照。アデノウイルス株Adタイプ5 dl324またはその他のアデノウイルス株(例えば、Ad2、Ad3、Ad7等)に由来する適切なアデノウイルスベクターは当業者にはよく知られている。組換えアデノウイルスは、細胞分裂していない細胞には感染する能力がない点である環境では有利であり得る。更に、ウイルス粒子は、精製および濃縮に対して相対的に安定かつ従順であり、また上記のように、感染スペクトルに作用するように修飾することもできる。更に、導入したアデノウイルスDNA(およびそれに含有される外来DNA)は、宿主細胞のゲノムに組込まれないが、エピソームの状態であり、よって、導入したDNAが宿主ゲノム(例えば、レトロウイルスDNA)に組込まれるに至る状況で挿入突然変異誘発の結果として発生し得る潜在的な問題を回避できる。更に、アデノウイルスゲノムの外来DNA収容力は、その他の遺伝子運搬ベクターと比べて大きい(8キロベースまで)(Berkner等、前掲、Haj-Ahmand and Graham(1986)J.Virol.57:267)。現在使用されており、故に本発明でも好ましい多くの複製欠損性アデノウイルスベクターは、ウイルスE1およびE3遺伝子の全部または部分を欠失しているが、アデノウイルス遺伝物質の80%ほどを保持している(例えば、Jones等(1979)Cell 16:683;Berkner等、前掲;およびGraham等, in Methods inMolecular Biology,E.J.Murray,Ed.(Humana,Clifton,NJ,1991)vol7,pp.109-127)。核酸分子に含まれる対象となる遺伝子の発現は、例えば、E1Aプロモーター、主要後期プロモーター(MLP)および関連リーダー配列、E3プロモーター、または外来的に付加したプロモーター配列の制御下にあり得る。
対象となる遺伝子を含む核酸分子の運搬に有用な更に別のウイルスベクターシステムは、アデノ随伴ウイルス(AAV)である。アデノ随伴ウイルスは、アデノウイルスまたはヘルペスウイルスなど、効率良い複製および生産性生活環のためにヘルパーウイルスなどのその他のウイルスを必要とする天然の欠損ウイルスである(例えば、Muzyczka等、Curr.Topics in Micro. and Immunol.(1992)158:97-129参照)。アデノ随伴ウイルスは、高頻度の安定な組込み(インテグレーション)を示す(例えば、Flotte等(1992)Am.J.Respir.Cell,Mol.Biol.7:349-356;Samulski等(1989)J.Virol.63:3822-3828;およびMcLaughlin等(1989)J.Virol.62:1963-1973)。AAVの300塩基対程を含むベクターをパッケージすることもでき、組込むことができる。外来DNAのためのスペースは、約4.5kbに限られている。Tratschin等(1985)Mol.Cell.Biol.5:3251-3260に記載されているようにAAVベクターを用いてDNAをT細胞中に導入できる。様々な核酸がAAVベクターを用いて異なる細胞タイプに導入されている(例えば、Hermonat等(1984)Proc.Natl.Acad.Sci.USA 81:6466-6470;Tratschin等(1985)Mol.Cell.Biol.4:2072-2081;Wondisford等(1988)Mol.Endocrinol.2:32-39;Tratschin等(1984)J.Virol.51:611-619;およびFlotte等(1993)J.Biol.Chem.268:3781-3790参照)。遺伝子治療に応用できるその他のウイルスベクターシステムは、ヘルペスウイルス、ワクシニアウイルス、および数種のRNAウイルスから誘導されている。
本発明のその他の実施態様では、対象となる遺伝子を含む核酸分子を、当分野ではよく知られている非ウイルス媒介トランスフェクション法によりT細胞に導入する。これらの方法には、エレクトロポレーション、リン酸カルシウム沈澱、およびDEAEデキストラントランスフェクションがある。
更にその他の実施態様では、対象となる遺伝子を含む核酸分子を細胞運搬ベヒクルに載せてT細胞に輸送する。このようなベヒクルには、例えば、カチオン性リポソーム類(Lipofection(登録商標))または誘導体化(例えば、抗体コンジュゲート化)ポリリジンコンジュゲート類、グラミシジンS、人工ウイルスエンベロープ類がある。これらのベヒクルは、プラスミド、ベクター、またはウイルスDNA中に取り込まれた核酸を運搬できる。具体的な実施態様では、Philip,R.等(1994)Mol.Cell.Biol.14,2411に記載のように、一次Tリンパ球を、カチオン性リポソームと複合体形成させたアデノ随伴ウイルスプラスミドDNAでトランスフェクションさせることにより、一次Tリンパ球に核酸分子を効果的に導入できる。
本発明のその他の実施態様では、対象となる遺伝子を含む核酸分子を可溶性分子複合体の形態で特定の細胞中に運搬する。この複合体は、核酸結合剤および特異的T細胞の表面分子に結合する細胞特異的結合剤からなるキャリアーに解離可能に結合した核酸を含有し、続いて細胞にインターナリゼーションされ得るサイズのものである。このような複合体類は、米国特許第5,166,320号に記載されている。
本発明のその他の実施態様では、Yang,N.S.およびSun,W.H.(1995)Nature Medicine 1,481に記載のように粒子ボンバードメントにより核酸をT細胞に導入する。
5.対象となる遺伝子を含む核酸分子
本発明は、遺伝子を含む核酸分子をT細胞に導入する改良法に関する。用語“核酸分子”は、DNAおよびRNAを含むこと、および一本鎖または二本鎖であり得ることを意図する。用語“遺伝子”は、RNAに転写され得るDNAヌクレオチド配列、あるいは、少なくとも1つのタンパク質に翻訳され得るRNA分子を含むことを意図する。
本発明の具体的な実施態様では、本発明の方法によりT細胞にトランスフェクションした際に、少なくとも1つのタンパク質がT細胞中で合成されるように、その遺伝子は、1またはそれ以上のオープン・リーディング・フレーム(転写解読枠)を含有するヌクレオチド配列、即ち、ペプチドをコードする配列からなる。少なくとも1つのタンパク質をコードするその遺伝子は、サイトカインをコードする遺伝子のようなあらゆる遺伝子であることができる。その遺伝子は1つのペプチドをコードしていてもよく、また数種のペプチド類をコードすることもできる。
本発明の別の実施態様では、その遺伝子は、本発明方法に従いT細胞に導入した際に1またはそれ以上の機能性RNA分子(例えば、アンチセンスRNA分子)中で発現される、ヌクレオチド配列である。本発明の好ましい実施態様では、機能性RNA分子は、1またはそれ以上の外来遺伝子のT細胞中での発現を阻害または少なくとも低減する。そのため、本発明の方法は、選択した遺伝子のT細胞中での発現を低減するのに有用である。例えば、外来RNAの翻訳を低減させるために、T細胞をアンチセンスRNAをコードする遺伝子を含む核酸分子でトランスフェクションさせる。“アンチセンス”核酸は、ワトソンおよびクリックの塩基対形成規則に従い構築される“センス”核酸に相補的なヌクレオチド配列、例えば、タンパク質をコードするmRNA配列に相補的なヌクレオチド配列を含む。従って、アンチセンス核酸は、センス核酸に水素結合し得る。mRNAの配列に相補的なアンチセンス配列は、mRNAのコード領域にみられる配列に相補的であることができるか、またはmRNAの5’または3’非翻訳領域に相補的であることができる。好ましくは、アンチセンス核酸は、開始コドンの前にあるかまたは開始コドンにまたがる領域に、またはmRNAの3’非翻訳領域内の領域に相補的である。アンチセンス遺伝子を用いる遺伝子発現の調節については、Weintraub,H等, Antisense RNA as amolecular tool for genetic analysis,Reviews-Trends in Genetics,Vol.1(1)1986参照。
本発明のその他の実施態様では、本発明の方法に従い、T細胞にリボザイムをコードする核酸を導入することにより、外来遺伝子のT細胞中での発現を低減させる。リボザイムは、mRNAなどの一本鎖核酸に対する相補的領域を有し、その一本鎖核酸を切断する能力のあるリボヌクレアーゼ活性を有する触媒RNA分子である。対象となる核酸に対する特異性を有するリボザイムは、核酸のヌクレオチド配列に基づき設計できる。例えば、テトラヒメナL−19IVS RNAの誘導体を、その活性部位の塩基配列が対象となるmRNA中の切断される塩基配列と相補的であるように構築することができる。例えば、Cech等、米国特許第4,987,071号;およびCech等、米国特許第5,116,742号参照。
遺伝子を含む“核酸分子”は、DNA分子またはRNA分子であることができる。その核酸分子は、天然核酸分子の一部であってもよく、また合成的に作成したものであってもよい。核酸分子は、典型的には、その遺伝子を作動可能に連結させた調節配列を含有する。“作動可能に連結させた”とは、遺伝子のヌクレオチド配列を、T細胞中で遺伝子が発現(即ち、転写)できるような手段で、少なくとも1つの調節配列に連結させたという意味を意図する。調節配列は、当分野では認められており、適切なT細胞中での遺伝子の発現を目的として選択される。従って、調節配列の用語は、プロモーター類、エンハンサー類、およびその他の発現制御配列類を包含する。このような調節配列は、当業者には知られており、Goeddel,Gene Expression Technology:Methods in Enzymology 185, Academic Press,San Diego,CA(1990)にも記載されている。
これらの調節配列には、遺伝子の転写および翻訳に必要とされるものがあり、プロモーター類、エンハンサー類、ポリアデニル化シグナル、および適切な細胞コンパートメントに分子を輸送するのに必要な配列があり得る。核酸が組換え発現ベクター中のcDNAであるとき、cDNAの転写および/または翻訳を担う調節機能は、ウイルス配列により与えられることが多い。通常使用されるウイルスプロモーターの例には、ポリオーマ、アデノウイルス2、サイトメガロウイルスおよびサルウイルス40、およびレトロウイルスLTRsに由来するものがある。
cDNAに連結させる調節配列を選択して、例えば、誘導性エンハンサーの使用により構成的または誘導的転写を与えることができる。そのため、本発明の具体的な実施態様では、対象となる遺伝子を含む核酸分子は誘導性制御配列の制御下にあり、そうして、その核酸を含有するT細胞をその誘導性制御配列に影響を与える作用物質と接触させるか、または接触させないことにより、それぞれ、遺伝子の発現をオンまたはオフにすることができる。
具体的な実施態様では、核酸分子は、誘導性制御配列の制御下にある。哺乳動物細胞に使用する誘導性調節システムは、当分野では知られており、例えば、遺伝子発現を重金属イオン(Mayo等(1982)Cell 29:99-108;Brinster等(1982)Nature 296:39-43;Searle等(1985)Mol.Cel..Biol.5:1480-1489)、熱ショック(Nouer等(1991)in Heat Shock Response,e.d.Nouer,L.,CRC,Boca Raton,FL,pp167-220)、ホルモン(Lee等(1981)Nature 294:228-232;Hynes等(1981)Proc.Natl.Acad.Sci.USA 78:2038-2042;Klock等(1987)Nature 329:734-736;Israel & Kaufman(1989)Nucl.Acids Res.17:2589-2604およびPCT公開公報WO93/23431)、テトラサイクリン(Gossen,M.およびBujard,H.(1992)Proc.Natl.Acad.Sci.USA 89:5547-5551およびPCT公開公報WO94/29442)またはFK506関連分子(PCT公開公報WO94/18317)により調節するシステムがある。
誘導性制御配列は、全てのT細胞において、あるいは、T細胞の特定のサブセットにおいてのみ、例えば、CD4+T細胞、CD8+T細胞、Tヘルパー1(Th1)、Tヘルパー2(Th2)細胞において誘導性であることができる。誘導性制御配列は、1タイプのT細胞(例えば、CD4+T細胞)中の一要因によって誘導可能であり、かつその他のタイプのT細胞(例えば、CD8+T細胞)中の他の要因によって誘導可能である配列であることもできる。
本発明のその他の実施態様では、対象となる遺伝子を含む核酸分子は、核酸分子の発現を構成的に起こす調節配列の制御下にある。核酸分子の構成的発現を起こすために核酸分子に作動可能に連結させた調節配列は、ウイルスの配列であり得る(例えば、ポリオーマ、アデノウイルス2、サイトメガロウイルス、シミアンウイルス40、またはレトロウイルスに由来する)。あるいは、T細胞受容体エンハンサーなどの構成性T細胞エンハンサーを使用できる(例えば、WinotoおよびBaltimore(1989)EMBO J.8:729-733)。
調節配列に作動可能に連結させた対象となる遺伝子を含む核酸分子は、典型的には、細菌中でのベクターのインビトロ選択および増幅に必要な配列を含むベクター(例えば、プラスミドまたはウイルスDNA)内にある。そのベクターに乗せた遺伝子を発現させることができるベクターを本明細書では“発現ベクター”と称する。
6.本発明方法の応用性
本発明は、核酸分子中に含まれる遺伝子をT細胞に導入し発現させる改良法に関する。本発明の好ましい実施態様では、T細胞は、一次T細胞である。従って、本発明の方法により、従来の一次T細胞トランスフェクション法に比べて、一次T細胞に導入した遺伝子を高レベルで発現させることが可能である。遺伝子を含む核酸分子で一次T細胞をトランスフェクションさせて、その遺伝子をT細胞中で発現させる能力は、特に遺伝子治療において、多大の応用性を有する。
具体的な実施態様では、末梢血T細胞を、対象から入手し、対象となるタンパク質をコードする遺伝子を含む核酸分子でエクスビボでトランスフェクションさせ、そのタンパク質をT細胞中で合成させる。そのT細胞を更に、対象に再投与してもよい。具体的な実施態様では、T細胞中で合成させた外来タンパク質をT細胞により分泌させる。従って、本発明は、個体中で分泌可能タンパク質を産生する方法を提供するものである。本発明の範囲内のタンパク質は、例えば、サイトカイン類、リンホカイン類、成長因子類がある。そのため、トランスフェクション化T細胞により産生されるタンパク質は、Tリンパ球自身よりも他の細胞を優先的に標的とすることができる。
あるいは、トランスフェクション化T細胞により産生されるタンパク質は、細胞内タンパク質または膜タンパク質である。具体的な実施態様では、外来タンパク質は、例えばウイルスにより感染からT細胞を防御するタンパク質である。かかる方法は、その一部がウイルス、例えば、ヒト免疫不全ウイルス(HIV)に感染してるいT細胞集団を拡張(イクスパンド)するのに有用である。そのため、T細胞集団を、全ての細胞に感染を付随蔓延させることなく、展開させることができる。
その他の実施態様では、外来タンパク質は、毒素などの特定のT細胞サブセットを殺すタンパク質である。そのタンパク質は、特定のT細胞サブセットに特異的な調節制御配列の制御下で遺伝子を持つことにより、選択的にそのT細胞サブセットを標的とすることができる。また、あるT細胞サブセットの選択的トランスフェクションを可能にするトランスフェクション法を用いることにより、外来遺伝子は特異的にあるタイプのT細胞を標的とすることも可能である。例えば、その膜上にT細胞サブセット特異的分子のリガンドを含有するリポソームでT細胞をトランスフェクションさせることができる。本発明方法によりT細胞に導入された遺伝子は、アデニンデアミナーゼ欠損に起因する深刻な複合免疫不全症などの遺伝子疾患を処置するために遺伝子設計することもできる。例えば、アデノシンデアミナーゼをコードする遺伝子は、本発明の方法を用いて、一次Tリンパ球に導入し、そこで発現させることができる。本発明のその他の実施態様は、T細胞上のCD40リガンド(gp39)の突然変異を特徴とし、かつヘルパーT細胞依存性抗体応答の欠如を特徴とする遺伝子疾患である、高IgM症候群の処置に関する。高IgM症候群の患者由来のT細胞を、gp39をコードする核酸で、好ましくはそれ自身のプロモーターの制御下、エクスビボでトランスフェクションさせ、その後、患者にそのT細胞を投与できる。T細胞中の機能不全遺伝子、例えばT細胞シグナル伝達経路に関連するタンパク質をコードする遺伝子、に起因するその他の遺伝子疾患も、本発明の方法により処置できる。
続く発明は、下記実施例により更に例示説明するが、これは、本発明を限定するものと解釈されるべきではない。本出願に引用した全ての文献、継続中特許出願、および公開特許の内容は、出典明示により本明細書の一部とする。
実施例
実施例1−CD28 + 末梢血Tリンパ球のインビトロ長期培養
直接的トランスフェクション実験は、生体内の推定上の調節配列の機能的重要性を証明することがしばしば要求される。この種の研究は、典型的には形質転換もしくは不滅化した細胞系を使用する。しかしながら、対象とする一次細胞の調節DNA配列を検討するのが好ましい。細胞サイクル調節、特にG0−特異的遺伝子発現の予測的研究のためには、生理学的背景の維持が要求される。これらの実施例は、一次T細胞にトランスフェクションされた外来DNAの発現を促す条件の開発を行うようにデザインされている。
末梢血T細胞は、次のようにして調製した。軟膜を、21−31才の健康なドナーの白血球フォレシスによりまたは赤十字から入手した。末梢血の単核細胞(PBMCs)は、Ficoll−Hypaque(Pharmacia)クッションによる密度勾配遠心分離または白血球分離培地(Litton Bionetics)で取得した。T細胞のCD28+サブセットは、文献(June, C.H., Ledbetter, J.A., Gillespic, M.M., Lindste, T., and Tyhompson, C.B.(1987)Mol.Cell.Biol. 11:5435-5445)のヤギ抗マウスIgをコートした磁気粒子(Dynal Inc)による免疫吸着を利用するネガティブ分離によって、PBMCsから単離した。細胞精製は、流速血球計および組織化学により、機械的に監視した。得られた細胞群は、フルオレセイン・イソチオシアネート(FITC)をコンジュゲイトしたmAbsを用いる蛍光活性化細胞ソーター(FACS)分析によれば、>99%CD2+で、>98%CD28+であった。単球、B細胞および大顆粒白血球は免疫蛍光分析では検出できなかった。別法としては、休止T細胞を、文献(Thompson, C.B., Ryan, J.J., Sieckmann, D.G., Finkelman, F.D., Mond, J.J. and Scher, I.(1983)J. Immunol. Methods 63:299-307)の遠心溶出法により単核細胞画分から調製した。これらの細胞は、流速血球計の測定では、>95%CD2+であった。両方法とも、細胞生存率は、トリパン・ブルー・エクスクルージョンによる測定では、>99%であった。
上記のようにして得られたヒト末梢血T細胞は、>99%CD2+、>98%CD28+であり、休止期にあることが示された。この研究に使用された精製T細胞は、アクセサリー細胞を含まず、フィトヘマグルチニン(PHA)、フォルボール・ミリステート・アセテート(PMA)またはイオノマイシン単独で刺激しても、試験管内で増殖しなかった。しかしながら、これらのT細胞は、固定化モノクローナル抗体(mAb)でTCK-CD3架橋させることにより、また適量のPMAとイオノマイシン(Lindsten, T., June, C.H., and Thompson, C.B.(1988)EMBO J.,7:2787-2794)により刺激されて分裂し得た。この条件下で、休止T細胞の>90%が活性化され、細胞の大多数が同期的に細胞分裂の1回を行った。
休止T細胞を活性化し、培養による長期拡張(long-term expansion)を促進するには、上記のようにして新たに単離した休止T細胞を2×106個/mlの濃度で、完全培地(RPMI 1640(GIBCO)に10%加熱不活化ウシ胎児血清(GIBCO)、2mML−グルタミン、ペニシリンG(100U/ml)、ストレプトマイシン(100mg/ml)および15ml Hepes(N−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸;pH7.4;GIBCO)を添加し、37℃で5%CO2とともに1夜静置したもの)中で培養した。1夜培養後、拡張プロトコールの第1日に、休止T細胞を、June et al.,(1987)Mol.Cell.Biol.11;5433-5445に記載の可溶化抗CD28抗体(αCD28)mAb 9.3(1μg/ml)の存在下にCD3ε鎖に対する固定化抗CD3抗体(αCD3)mAbG19−4の飽和量で刺激した。CD3mAbG19-4(IgG1)は、文献(Ledbettger, J.A., Martin, P.J., Spooner, C.E., Wofsy, D., Tsu, T.T., Beatty, P.G. and Gladstone, P.(1985)J. Immunol.135:2331-2336)の方法で調製され、精製された。mAbG19-4は、文献(Geppert, T.D., and Lipsky, P.F.,(1987)J. Immunol. 138:1660-1666)の記載によってプラスチック組織培養フラスコ/プレートの表面に増殖に適当な量を吸着させた。これを実施したのは、架橋(Williams, J.M., Ransil, B.J., Shapiro, H.M., and Strom, T.B.(1984)J. Immunol. 133;2986-2995)が必要とされるからであり、CD3複合体のインターナリゼーション(Lebetter, J.A., Hune, C.H., Martin, P.J., Spooner, C.E., Hansen, J.A. and Meier, K.E.(1986)J. Immunol. 136:3945-3952)を阻止するためであった。CD28mAb9.3(IgG2a)はプロテインA−セファローズで精製し、PBSに対して透析し、0.22μm滅菌フィルターで濾過し、遠心分離(100,000×g、45分間)で凝集物を除き、1μg/mlで使用した(Ledbetter, J.A., Martin, P.J., Spooner, C.E., Wofsy, D., Tsu, T.T., Beaty, P.G. and Gladstone, O.(1985)J. Immunol., 135:2331-2336)。
2日後、つまり第3日目に、活性化T細胞を計数し、サイジングし、新しい完全培地で0.5×106個/mlの濃度に希釈した。サイズの分布が休止細胞のプロフィールに近くなるまで、細胞の計数、サイジング、希釈を2日毎に繰返した。そこで、T細胞を2×106個/mlの濃度で完全培地に再懸濁し、上記のようにして再刺激する(第1回の再刺激は通常第10日頃)。
細胞は、コールターZMカウンター(Coulter Electronics)を用いて計数した。細胞は、円筒状の70μmアパーチャーを備えたコールター・計数器ZMモデルとIBMのPCコンピューター接続したチャネライザーC−256モデルで、直線目盛り上にサイジングした。細胞はイソトンに懸濁し、一定の径を持つラテックス・ビーズを用いてキャリブレイションした。
マイトジェンまたは抗T細胞レセプター(抗TCR)刺激T細胞をαCD28で処理するとT細胞増殖の相乗的増加を誘発した。休止T細胞をαCD3およびαCD28で共刺激(コスティミュレーション)すると、初期に激しい指数的増殖が起り、細胞の肥大、集合および培地の酸性化によって特徴づけられる代謝が生じる。細胞は、培養の初期7−8日に数回の細胞分裂を行い、細胞数は6−8培となった。この時点で、細胞の増殖率は低下した。培養第10−11日には細胞分裂は停止し、細胞はそのサイズについて休止細胞に類似してきた(図1)。拡張プロトコールのこの点で細胞はαCD28の存在下に固定化したαCD3で再刺激し、細胞の集合と肥大で特徴づけられる再度の指数増殖期を経過した。
図2はこの方法で培養した細胞の増殖特性を示している。ここに示されているように、細胞はαCD3/αCD28共刺激の繰返しによって多週にわたって(3か月以上)指数的状態に維持され、増殖した。流速血球計分析を種々の時点で行い、これらの細胞の表現型進化を追跡した。時間とともに、こうして拡張された細胞は、表現型におけるナイーブなTヘルパー(Th)細胞から記憶細胞への転換を反映して、次第に、よりCD4+45RO+となった。これは、αCD3/αCD28共刺激の後に外来IL−2の存在下に増殖させた細胞と直接対照的である。これらの細胞は、時間と共に次第によりCD8+となり、最終的には培地中で増殖できなくなった。これらの観察は、CD4+細胞によって生産される何らかの因子が連続的なT細胞の増殖に必要とされることを示している。
実施例2 細胞増殖は外来DNAの発現に充分ではない。
一次T細胞の長期クローン拡張のための反復αCD3/αCD28共刺激を確立し、このプロトコールを用いて増殖させた細胞について、ノーザン・ブロット分析により、内生ets−1 mRNAの発現を分析した。
RNAの抽出のために、細胞を遠心分離により採取し、グアニジン・イソチオシアネート(Chirgwin et al, 1979)を用いて総細胞RNAを抽出した。試料をrRNAに関して均等化し、均等化を、文献(June, C.H., Ledbetter, J.A., Gillespie, M.M., Lindsten, T., and Thompson, C.B.(1987)Mol.Cell.Biol. 11:5435-5445)の記載により、非変性1%アガロース・ゲル上で分離したRNA試料の均等量のエチジウム・ブロミド染色によって確認した。これらの均等化したRNA試料(5−10μg)を1%アガローズ/フォルムアルデヒド・ゲル上で分離し、ニトロセルロースに移した。DNAプローブは、3−9×108cpm/μgの比活性にニック・トランスレーションで標識した。IL−2特異的プローブは、pTCGF5プラスミド(Clark, S.C., Wong−Staal, F., Matsumoto−Kobayashi, M., Kay,R.M., Kaufman, R.J., Brown, E.L., Shoemaker, C., Copeland, T., and Oroszian, S. et al,(1984)Proc.Natl.Acad.Sci.USA 81:2543-2547)由来の1.0kb PstIcDNAインサートであった。HLAB7プローブはpHLA−B7から単離された1.4kb PstLフラグメントであった(Sood, A.K., Pereira, D., and Weissman, S.M.(1981)Proc. Natl.Acad.Sci. USA 78;616-620)。est−1DNAプローブは、1.9kb est−1 cDNAからの442塩基対EcoRI/XbaIフラグメントからなるものであった(Ho, I−C., Bhat, N.K. Gottschalk, L.R., Lindsten, T., Thompson, C.B., Papas, T.S. and Leiden, J.M.(1990)Science 250:814-817)。膜は洗浄後、X線フィルム(Kodak XAR−2またはXAR-5)に、増感スクリーンを使用して、−70℃で4時間ないし7日間暴露した。
ノーザン・ブロット作成のために、RNAは、第1日のαCD3/αCD28共刺激の後または第8日の再刺激の後の種々の時点で、上記の一次T細胞の長期クローン拡張のプロトコールに従って培養された末梢血T細胞から抽出された。ノーザン・ブロット分析の結果は図3に示されている。休止ヒトT細胞はest−1 mRNAおよびタンパクを高度に発現している。休止T細胞および第8日の“活性化”細胞は、est−1 mRNAを高度に発現した。αCD28の存在下に抗原レセプター架橋を行った後は、est−1 mRNAは6時間で検出できない程に減少した。第1日および第8日の細胞は両方共にest−1 mRNAを再発現し、刺激後24ないし72時間維持された。
どのシス作用性調節配列がest−1遺伝子の発現を調節しているかを決定するために、対数生育期にある一次細胞を、CAT遺伝子に結合したest−1プロモーター(EST−1-CAT-2)を含むプラスミドで、長期培養プロトコールの第6日にトランスフェクションした。この構成はジャーカットT細胞で強力なレポーター活性を発揮した。一次T細胞の、いずれかのDEAE−デキストランによる1μg DNAのトランスフェクション後に(Ho, I−C., Bhat, N.K., Gottschalk, L.R., Lindsten, T., Thompson, C.B., Papas, T.S., and Leiden, J.M.(1990)Science 250:814-817)、細胞を反復して洗浄し、完全培地に再懸濁した。トランスフェクションの40時間後に、細胞を採取し、CAT活性を測定した。
DEAE−デキストランでのトランスフェクション・プロトコールでは、細胞はPBSで1回、TS緩衝液pH7.4で1回洗浄した。2度目の洗浄の後、細胞はDEAE−デキストラン(分子量500,000)500μgとスーパーコイル状プラスミド1−10μgを含むTS緩衝液中に107/mlで再懸濁させた。この混合物を、時々かき混ぜながら室温で12−15分間放置した。RPMI1640 10mlに20%熱不活化ウシ胎児血清、2mML−グルタミンおよび15mMHepesを添加して(RPMI 1640/20%FCS/G/H)、細胞に加えた。細胞を組織培養フラスコに移し、5%CO2中37℃で30分間培養した。細胞をペレット化し、RPMI 1640/20%FCS/G/Hで1回洗浄し、RPMI 1640/20%FCS/G/H 10ml中に37℃、5%CO2で再懸濁した。
ここに記載した他の例では、一次T細胞をエレクトロポレーションによりトランスフェクションした。エレクトロポレーションプロトコールでは、細胞を氷冷したPBSで2回洗浄し、氷冷したRPMI 1640/20%FCS/G/H中に、20×106細胞/mlで再懸濁した。300μl中に6×106細胞を含む懸濁液を滅菌した0.4cmのエレクトロポレーション用キュベット(BioRad)に移した。レポーター・プラスミド1−10μgを加え、ジーン・パルサー(BioRad)を使用して250V、960マイクロファラッドで細胞にエレクトロポレーションを行った。細胞を氷上で10分間培養し、RPMI 1640/20%FCS/G/H 10mlで希釈し、5%CO2中37℃に保持した。
等容積の細胞抽出液を16時間の反応でCAT活性を測定した。EDTAを最終濃度が5mMになるように添加し、抽出液は65℃で10分間アセチルCo−Aの加水分解とクロラムフェニコールの脱アセチル化を阻害するように加熱した(Crabb, D.W., and Dixon, J.E.(1987)Anal. Biochem. 163:88-92)。CAT測定の薄層クロマトグラフィー上でのオートラジオグラフィーの前に、[14C]クロラムフェニコールおよびそのアセチル化誘導体のスポットをベータスコープ(Betagen)またはフォスフォリメイオジャー(Molecular Dynamics)をもちいて定量した。アセチル化のパーセントは、アセチル化および非アセチル化の実験値からバックグラウンド値を差し引いて計算した。もし、トランスフェクションの与えられた一つのセットについてのアセチル化のパーセントが測定の直線領域を超えたときは、等容量の細胞抽出液を希釈して、CAT測定を再試行した。CAT活性は、結いで反応中のタンパク量に対して正規化した。正規化したCAT活性は(パーセント・アセチル化/mgタンパク)×50として表す。すべてのレポーター活性の比較は、同時に、かつ同一の試薬により刺激され、トランスフェクションさせ、採取され、測定された細胞について行う。
本方法でトランスフェクションさせた一次細胞には、EST-1-CAT-2レポーター活性は検出されなかった。内生のest−1 mRNAは休止細胞で優先的に発現し、T細胞刺激の約2−3日後に再誘発されるので、EST-1-CAT-2レポーター発現は、T細胞が休止期に再度入ったかどうかに係わりなく期待された。驚くべきことに、RSV-CAT、HIV-1-CATおよびHILV-1-CATのような陽性対照による細胞のトランスフェクションもまた証明できるようなレポーター活性を与えなかった。細胞数の増加により、長期培養プロトコールの第6日での細胞は対数増殖期にあった(図2)。しかしながら、構成的レポーター構築物のこれら細胞へのトランスフェクションは検出可能なレポーター活性を与えず、効率的な導入遺伝子の発現には、増殖だけでは不十分であるのとを示唆した。
“活性な”シグナル伝達がレポーター遺伝子の発現に必要かどうかを決定するために、培養プロトコールの第5日の対数増殖期にある一次T細胞を、フォルボール・エステル(PDBU)とカルシウム・イオノホア(イオノマイシン)でトランスフェクションの10時間前に刺激した。
図4はこの、そしてその後のトランスフェクションに用いた時刻表を示している。休止しているT細胞を、飽和量の固定化した抗CD3抗体と1μg/mlの抗CD28とともに培養して増殖するように刺激した。2日後、第3日に活性化されたT細胞を計数し、サイズ分けし、新鮮な完全培地で0.5×106細胞/mlの濃度に希釈した。第5日に、細胞をホルボール・12,13−ジブチレート(PDBU、LC Services Corp.)10ng/mlとイオノマイシン(Calbiochem)0.4μg/mlで刺激した。第6日、刺激後10時間で、細胞は、実質的に文献(Ho, I−C., Bhat, N.K., Gottschalk, L.R., Lindsten, T., Thompson, C.B., Papas, T.S. and Leiden, J.M.(1990)Science 250:814-817)に記載されているように、DEAE−デキストランを使用し、構成的発現レポーター構築物RSV-CAT 10μgでトランスフェクトした。pRSV-CAT(RSV-CAT)は、CATをコードする配列の5’末端にRSVLTRが融合している構成である(Gorman, C.M., Merlino, G.T., Willingham, M.C., Pastan, I., and Hpward, B.H.(1982)Proc.Natl.Acad.Sci. USA 79:6777-6781)。細胞を40時間後に採取し、CAT活性を測定した。
結果は、図5に示されている。増殖中の一次細胞のPDBU+イオノマイシンによる前刺激は、ならし培地のみで処理した細胞に比して、RSV−CATレポーターの発現が67倍増大した。このRSV−CATレポーター活性の劇的な相異が、刺激された細胞と刺激されていない細胞との増殖能力の違いによるものかどうかを決めるために、これらの細胞の増殖状態を次の方法で調べた:1)細胞周期の分析のためのアクリジン・オレンジ染色、2)DNA合成の指標としてのトリチル化チミジン[3H]TdRの取り込み、および3)細胞活性化の一般的指標としての細胞のサイジング。自家休止の一次細胞と長期培養プロトコールの第3日のαCD3/αCD28刺激細胞とを、静止期(G0/G1境界)と旺盛な増殖期のコントロールとして同時に測定した。
精製した休止T細胞(第1日)は、αCD28mAb 9.3(1μg/ml)の存在下にαCD3 mAb G19−4の飽和量で刺激した。第3日に活性化T細胞は新鮮な完全培地で0.5×106/mlの濃度に希釈し、mAb 9.3を最終濃度0.5μg/mlとなるように加えた。第6日に、指数増殖期の細胞をホルボール12,13−ジブチレート(PDBU、LC Services Corp.)10ng/mlとイオノマイシン(Calbiochem)0.4μg/mlまたはならし培地のみで10時間処理した。第3日および第6日の細胞を、下記のようにして細胞周期分析のためにアクリジン・オレンジで染色した。非刺激の細胞(第1日)は、同時にG0/G1境界の決定のために分析した。
細胞は、文献(Darzynkiewicz(1990)Methods Cell Biol. 33:285-298)の方法によりアクリジン・オレンジ(Polyscience)で染色した後FACスキャン流速血球計(Becton−Dickinson)でRNAおよびRNA含量を分析した。1−5×106個の細胞をPBSで2回洗浄し、70%冷エタノール中に2×106/mlの濃度に固定した。細胞を遠心分離し、洗浄し、2×106/ml以下の濃度で完全培地に再懸濁した。この細胞懸濁液0.2mlをアクリジン・オレンジで染色し、FACスキャンによって分析した。増量したRNA含量と変化のないDNA含量を示す細胞はG1期の細胞と考えられた。増量したRNAおよびDNA含量の細胞はSまたはG2M期と考えられた。
第1、3および6日のT細胞のトリチル化チミジン[3H]TdRの取り込みを決定するために、細胞は、平底96穴マイクロタイター・プレート(Costar)上、5×105細胞/ウェルで、4重試料として培養した。1μCiのトリチル化チミジン[3H]TdR(ICN)を各ウェルに加え、5%CO2中37℃で6時間培養した。6時間後、細胞は、PHD細胞採取器(Cambridge Technologies)を使用してガラスのマイクロフィルター・ストリップ(Whatman)上に採取し、液体シンチレーション・カウンター(LKB)で計測した。全ての数値は、4重試料について平均cpm±標準偏差として表される。
図6に示すように、αCD3/αCD28による休止T細胞の刺激は、細胞の拡張の第3日までに92%以上の細胞が細胞周期のG0からG1もしくはS/G2M相に進んでいた。これはトリチル化チミジンの207倍の増加および平均細胞容積の増加に対応していた。培養第6日までに、ならし培地のみに培養した91%以上の細胞が細胞周期のG1またはS/G2M相にあった。PDBU+イオノマイシン処理し、刺激の10時間後にRNAおよびDNAの含量の測定に供した細胞の92%以上が、細胞周期のG1またはS/G2M相にあることが見出された。これらのデータは、トランスフェクション時の細胞の活性な周期性とPDBU/IONO刺激細胞と無刺激細胞との同等の増殖性を示している。事実、PDBU/IONO刺激細胞は、非刺激の対応細胞と比べたとき、DNA合成率([3H]TdRの取り込みで35×103cpm対52×103cpm)および平均細胞容積において如何なる増加も示さなかった。従って、これら二つの細胞群でトランスフェクション時には増殖力に差はなく、これはRSV-CATレポーター遺伝子の発現の差を説明している。
ラウス肉腫ウイルスLTRは、カルシウム/カルモジュリン依存性プロテインキナーゼ(CaM−kinase)応答要素を含んでおり、これは増量したカルシウム・イオンの存在下に、活性化されたCaM−kinaseによる転写の選択的誘導を付与することができる(Kaipiloff., M.S., Mathis, J.M., Nelson., C.A., Lin, C.R., and Rosenfeld,M.G.(1991)Proc.Natl.Acad. Sci. USA 88:3710-3714)。PDBU/IONO刺激細胞と非刺激細胞との間のRSV−CAT発現の差異が、刺激後のRSV LTRの特異的トランス活性化から生じるかどうかを決定するために、細胞をPDBU/IONOで前刺激するかならし培地のみで処理するか、トランスフェクションし、次いでならし培地または完全培地中で培養した。トランスフェクションの30時間後完全培地で培養した細胞をPDBU/IONOまたはαCD3/αCD28で刺激するか、単に培地で処理した。10時間後に、CAT活性を見るために細胞を採取した。もしシグナル伝達の唯一の役割がRSV LTRからの転写の活性化であれば、トランスフェクション後に刺激された細胞は増加したレポーター活性をも示さなければならない。図7に示すように、PDBU/IONOで前刺激した細胞は、非刺激の増殖中のコントロールに比してRSV−CATレポーター活性に関し345倍の増加が生じた。トランスフェクション30時間後にαCD3/αCD28またはPDBU/IONOで刺激した細胞は、RSV−CATレポーター活性に関し僅か3倍または3.5倍の増加をそれぞれ示した。トランスフェクションさせた細胞を成長コンピーテントならし培地で培養するとCAT活性が2.5倍増加した。さらに、トランスフェクションの直後にPDBU/IONOまたはαCD3/αCD28で刺激した細胞はRSV−CAT活性を僅か4−5倍増加させた。これらのデータは、トランスフェクション前の刺激はRSV−CAT活性に必要であり、トランスフェクション時のシグナル形質導入は、トランスフェクションの効率を上げるか、導入遺伝子を発現可能にすることにより、レポーター遺伝子の発現を促進することを示唆している。このレポーター構築物を用いて、トランスフェクション後に細胞を刺激しても、レポーター遺伝子の発現の明らかな増加は起らなかった。
次の例では、一次T細胞の抗CD3および抗CD28による、トランスフェクション10時間後の刺激もレポーター構築物の大幅に増加した発現を起こした。先に報告されているHIV-1-CAT(Nabel,G.,and Baltimore,D.(1987)Nature 326:711-713)をこの例で使用した。増殖中のT細胞はPDBU/IONOまたはαCD3/αCD28で刺激するかならし培地のみで処理し、HIV-1-CATでトランスフェクトし、次いでならし培地中で直ちに培養するか、トランスフェクションの30時間後にPDBU/IONOまたはαCD3/αCD28で刺激するか培地のみで処理した。10時間後に細胞を採取し、CAT活性を測定した。CAT測定の結果は、図8に示されている。この結果は、トランスフェクションの10時間前に一次T細胞を前刺激すると、前刺激なしにトランスフェクションしたのに比して、HIV-1-CATレポーター構築物の発現を大幅に増加させることを示している。従って、レポーター構築物の発現増加は、この構築物のプロモーターやエンハンサーのタイプに依存しない。さらに、これらの結果はトランスフェクションしたレポーター構築物の発現を増加させるための一次T細胞の前刺激は、抗CD3または抗CD28の組み合わせ物によってもなされ得ることを示している。
実施例3 外来DNAの発現はトランスフェクション時にTCR依存のシグナル伝達を必要とする。
一次T細胞における効果的な導入遺伝子の発現の要件をさらに吟味するために、広範に研究され、よく特性付けされている細胞性IL−2遺伝子のプロモーター/エンハンサーがトランスフェクションの実験に使用された。ノーザン・ブロット分析が、至適量の固定化αCD3によるTCR−CD3複合体での刺激後のIL−2遺伝子発現の速度論的検討に使用された。さらに、これらの培養物の上清でIL−2含量を分析し、細胞で細胞周期進度を分析した。
結果は、図9に示されている。パネルBは至適のαCD3刺激により培養6時間でIL−2 mRNAの発現はピークに達し、培養12時間までにIL−2 mRNAのレベルは検出不能なまでに低下した。この一過性のIL−2 mRNAの発現は、培地上清中の少量のIL−2(24時間で5U/ml)と活発な増殖(48時間で41%の細胞が細胞周期のS/G2M相)(パネルD)を伴っていた。要するに、TCR−CD3複合体での刺激は、6時間でピークに達し、刺激後12時間で検出不能なまでに低下するIL−2遺伝子転写の一過性の誘導を起こした。IL−2 mRNAの転写が誘導性であり、一過性であるので、IL-2-CATレポーター遺伝子の発現のためのトランスフェクション時におけるシグナル伝達の必要性を調べた。IL-2-CATプラスミド(IL-2-CAT)は、文献(Bielinska, AA., Shivdasani, R.A., Zhang, L., and Nabel, G.J.(1990)Science 250:997-1000)に記載のように、クロラムフェニコール・アセチルトランスフェラーゼ(CAT)遺伝子の5’に接してIL−2プロモーター/エンハンサー(−585ないし+18)を含んでいる。拡張プロトコールの第5日、増殖中の一次T細胞はPDBU/IONOで前刺激されるか、ならし培地のみで処理され、IL−2−CATレポータープラスミドでトランスフェクションさせ、直ちに増殖可能ならし培地または完全培地で培養した。トランスフェクションの30時間後に、完全培地中で培養された細胞はPDBU/IONOまたはαCD3/αCD28で刺激するか、培地のみで処理した。10時間後、細胞の数および生存能力をトリパン・ブルー・エクスクルージョンによって調べ、細胞をCAT活性測定のために採取した。
CATの測定結果は図10に示されており、その要点は次の通りである:1)前刺激をしない場合、増殖中の一次細胞は極低いレベルのIL−2−CATレポーターを発現した。2)トランスフェクション30時間後の細胞の刺激はこの低レベルのIL−2−CATレポーター遺伝子発現を増加させなかった。3)PDBU/IONOで前刺激し、トランスフェクション後、細胞をリンフォカイン富化条件培地または完全培地に再懸濁した細胞もまた極低いレベルのIL−2−CATレポーターを発現した。4)PDBU/IONOで前刺激し、かつトランスフェクションの30時間後にTCRによる刺激を行うと、IL−2CATレポーター活性は、前刺激のみの細胞に対して79倍、無刺激増殖のコントロールに対して約20倍に増加した。これらのデータは、IL−2CAT DNAレポーター構築物の発現可能コンパートメントへの導入モデルまたはトランスフェクション前のTCR−依存的シグナル伝達を必要とする状態と一致する。しかしながら、IL−2CATレポータープラスミドが発現と競合していも、最初のシグナル伝達がトランスフェクション10時間前にされているため、IL−2CAT転写が僅かに起こるか起こらず、上記ノーザン分析(図9)により、IL−2mRNA転写がこの時点までに低レベルに減少する。トランスフェクション後のTCR/CD3−依存性刺激は、発現成分IL−2プロモーター/エンハンサーおよびCAT mRNA転写を、得られるレポーター活性と共にもたらす。
実施例4 − 細胞性増殖はトランスジェニック発現に必要である
トランスジーン発現における細胞性増殖の役割に取り掛かるために、新たに単離した休止T細胞をαCD28(1μg/ml)、SEA(10ng/ml、Toxin Technologies)、PDBU(10ng/ml)/IONO(0.4μg/ml)または培地単独で10時間刺激した。これらの細胞をRSV−CATでトランスフェクションさせ、40時間後CAT活性のために回収した。活性化T細胞はマイトジェン刺激24−36時間後に細胞分裂した。従って、トランスフェクションの時、非常にまれに、もしあれば、T細胞はM期を通して進行している。トランスフェクション40時間後(刺激後約50時間)、SEAまたはPDBU/IONOで刺激したT細胞は増殖に関して(細胞性拡大、凝集)、表現型の変化は示さなかった。しかしながら、これらの集団は細胞数は増加しなかった。図11に示されるように、全てのこれらのトランスフェクション条件に関する正常化CAT活性は、相対的に低かった(0.07−0.26)。PDBU/IONOで刺激した休止細胞は、休止対照と比較して、RSV−CAT活性において、少ない3.7倍の増加を示した。PDBU/IONO刺激および休止細胞の間の低いCAT活性の増加は、単に細胞性活性の効果およびRSV LTR転写の増殖を単に反映し得、一度この受容体プラスミドが最初のシグナル伝達の結果発現可能である。注目すべきは、αCD28またはSEAで刺激された細胞のCAT活性が休止細胞と異ならなかった。αCD28またはSEA単独で、完全なマイトジェン刺激は構築されなかった。SEAによるT細胞増殖の導入に、アクセサリー細胞(主要組織適合性複合体(MHC)クラスII提示が必要)またはCD28のモノクローナル抗体の刺激により提供される第2のコスティミュラトリー・シグナルの添加が必要であった(Green, J.M., Turka, L.A., Junc, C. H.,およびThompson, C.B.(1992)Cell. Immunol. 145(1):11-20)。休止T細胞は、MHCクラスII(HLA−Dr)をその表面に発現しない(Mayforth, R. D.(1991)Ph.D. Dissertation, Univ. of Chicago)。従って、増殖刺激非存在下で、休止T細胞は構成性活性受容体プラスミドRSV−CATの発現を証明しない。
これらの細胞がRSV−CATを発現できることを証明するために、同じ細胞の他のアリコートをαCD3/αCD28で刺激するか、またはならし培地単独で10時間で処理し、RSV−CATでトランスフェクションさせ、次いで40時間後CAT活性について回収した。PDBU/IONO予備刺激有りまたは無しの一日目(休止細胞)または6日目(増殖細胞)の細胞の相対的CAT活性の比較を図11に示す。増殖細胞のPDBU/IONO予備刺激は、増殖非刺激対照と比較して13.4倍のおよびPDBU/IONO刺激休止細胞より23.7倍のCAT活性の増加をもたらした。従って、トランスフェクションの時の細胞性増殖は、RSV−CATレポーター発現を非常に促進する。上記のトランスフェクションの結果を一緒にして、これらのデータは、トランスジーン発現に増殖が必要であるが、十分ではないことを示す。
実施例5 − レポーター遺伝子活性の差はトランスフェクション効果の差によるものではない
上に証明するように、トランスフェクション時前のTCR−依存性シグナル伝達は、トランスジーン発現に必要である。トランスフェクション後の刺激は、殆どの場合、レポーター活性を明確に増加させない。シグナル伝達の多くの役割のいくつかが予測できる。最も単純なモデルにおいて、トランスフェクション時の増殖細胞のシグナル伝達は、トランスフェクション効果を増加でき、従って、核に到達するレポータープラスミドの量を増加できる。あるいは、シグナル伝達は、トランスフェクションDNAの非転可能コンパートメントから転写可能コンパートメント、例えば、細胞質から核への移動を促進する。両方のシナリオは、シグナル伝達に続く核へのレポータープラスミドの増加の量に関して同じ結果をもたらす。これらの可能性の試験のために、DNA侵入および局在化の動態を、トランスフェクション後の種々の時間の核および細胞質コンパートメント由来のトランスフェクトDNAの救済および定量により試験した。
5日目の増殖した一次T細胞を、PDBU/IONOで刺激するかまたはならし培地単独で処理し、RSV−CATレポータープラスミドでトランスフェクトし、細胞数および生存能力をコウルター計数器およびトリパンブルー・エクスクルージョンでそれぞれ測定した。
Dounceホモジナイゼーションに続く核および細胞質への分画に続いて、DNAをこれらの二つのフラクションから、SDS可溶化およびプロテイナーゼK消化後に連続酢酸アンモニア/イソプロパノール沈殿を使用して両方の分画から抽出した。DNA単離プロトコールは、低および高分子量DNAの両方の回収に関して定量的である。トランスフェクション後の種々の時点の核および細胞質コンパートメントのトランスジーンの相対的複製数を評価するために、105全細胞相当物の核および細胞質DNAを、先に記載のように、1.0%アガロースゲルでサイズ分画し、ニトロセルロースに移した(Thompson, C. B. およびNeiman, P. E.(1987)Cell 48:369-378)。ブロットをpRSC−CATのCATコード領域のEcoRIフラグメントまたはpIL2CATのCATコード領域のEcoRI/BamHIフラグメントのいずれかとハイブリダイズした。
サザンブロット分析の結果を図12に示す。プラスミド/細胞の約105コピーを、DNA/DEAEデキストラン複合体の暴露の最初の30分以内に取り込んだ。これは、トランスフェクトした全DNAの約10%に対応する。この量で、核画分に約90%が局在化する。トランスジーンの同じかわずかに増加した量が、トランスフェクション6、24および48時間後に核および細胞質に存在した。明らかに、PDBU/IONO前刺激および非刺激細胞の間で、いかなる時点でも核または細胞質でDNAの量に明白な差はなかった。要するに、サザン分析は、シグナル伝達および非シグナル伝達細胞の間で核区画に到達するDNAの量に証明可能な差のないことを確認した。
これらの発見を確認するために、一次T細胞をRSV−CATまたはIL−2CAT受容体プラスミドで、それぞれ図7および図10に記載のようにトランスフェクトした。図13および14に示すように、トランスフェクション40時間後、RSV−CATプラスミドの約105コピー/細胞が核に存在し、全トランスフェクトDNAの10%である。上記のように、PDBU/IONO前刺激細胞、刺激細胞の核、またはPDBU/IONO、αCD3/αCD28またはならし培地でトランスフェクションした後刺激した細胞の各に存在するDNAの量に明確な差がない。注目すべきは、細胞の低張融解に続いて単離されたDNAのサザン分析でトランスフェクション40時間後のプラスミド分解の証拠は見られなかった。
二つの異なるDNA単離法を使用してこの発見を総括すると、依然としてプラスミドDNAの回収が完全に定量的でなかった可能性を考えないわけにはゆかなかった。これらの結果を個別に確認するために、PDBU/IONO前刺激および非刺激細胞を32P放射標識化鎖状RSV−CATでトランスフェクションした。これらの細胞を次いで核および細胞質分画にトランスフェクション0、6、24および48時間後に分離した。次いで、分画を液体シンチレーションカウンターで計数した。
図15に示す結果は、トランスフェクション30分以内に、全cpmの15.2%がPDBU/IONO前刺激細胞に取り込まれることを示した。こでは標識刺激細胞の11.9%と同等である。これらの計数で、前刺激細胞の核分画で92%が、非刺激細胞で84%が回収された。続く時点で、核分画から回収された全cpmの%は、PDBU/IONO前刺激細胞で0時間の14.0から48時間の17.1%に、非刺激細胞で0時間の10.0%から48時間の13.9%に増加した。この核計数における増加は、前刺激および非刺激細胞の両方の細胞質分画で回収された全cpmの割合の少しの減少と対応し、恐らく、DNAの細胞質から核への移動を反映する。しかしながら、非シグナル伝達およびPDBU/IONOシグナル伝達細胞の各分画からの全cpmの回収の割合の少しの差は、これらの二つの集団の間でRSC−CATレポーター遺伝子発現の劇的な67から345倍増加を担わなかった。トランスフェクト放射標識DNAの計数結果は、細胞内に入った全プラスミド量および続くDNAの核および細胞質区画への分散の両方でサザンブロット分析と非常に密接している。要約すると、細胞のPDBU/IONO前刺激は、トランスフェクション効率の増加により、またはDNAの細胞質から核への移動を促進することにより、レポーター遺伝子発現を増加するとは思えない。
実施例6 − スーパー抗原誘導TCR活性化はレトロウイルスLTRを含むDNAの核破壊を変える
上記の実施例は、TCR媒介シグナル伝達により抑制可能な細胞性機構の存在を示し、それはインビトロ外来DNAの暴露から、休止T細胞および非シグナル伝達増殖一次T細胞を防御する。レポーター遺伝子発現のためのTCR依存性シグナル伝達の必要性から、スーパー抗原がトランスジーン発現の十分な刺激物として働くかを調査した。
トランスフェクション10時間前、指数増殖期の一次ヒトT細胞をならし培地単独、5×106放射自己単球、SEA(10ng/ml)またはSEA(10ng/ml)+5×106放射自己単球で処理した。細胞をRSC−CATまたはIIIV−1−CATでトンスフェクションさせ、トランスフェクション40時間後に回収し、CAT活性をアッセイした。
CATアッセイの結果を図16に示す。HIV−1−CATでのトランスフェクションにおいて、PDBU/IONO前刺激は非刺激増殖コントロールと比較して、CAT活性の15倍の増加をもたらした。T細胞と5×106放射自己単球の組み合わせは、CAT活性の2.3倍の増加をもたらした。SEA 10ng/mlでの処置は、CAT活性の32倍の増加をもたらしたが、T細胞とSEA(10ng/ml)+5×106放射自己単球の組み合わせは、16倍の増加をもたらした。従って、SEAは単独またはMHCクラスII(HLA−DR)との組み合わせのいずれでも、HIV−1−CATレポーター遺伝子発現増加をもたらす。完全に同様な結果がRSC−CATでのトランスフェクションでも見られる(図16)。SEAおよびSEA+MONO刺激細胞の増殖状態は、トリチル化チミジン取り込みおよび細胞サイジングで測定して、非刺激増殖細胞と測定可能には差がなかった。従って、HIV−1−CATおよびRSC−CATレポーター発現は、シグナル伝達におけるスーパー抗原の作用からもたらされ、増殖それ自体からではない。
このような機構およびその発現は、レトロウイルス発現に関する事象に非常に重要であり得る。例えば、休止T細胞のHIV−1の感染後、続くT細胞活性化がHIV−1ゲノムの宿主ゲノムへの組込みおよび感染ウイルスの製造に必要である(Stevenson, M., Stanwick, T.L., Dempsey, M. P.およびLamonica, C. A.(1990)EMBO J. 9(5):1551-1560;Zack, J. A., Arrigo, S. J., Weitsman, S. R., Go, A. A., Haislip. A.,およびChen, I. S. Y.(1990)Cell 61:213-222:Bukrinsky, M. L., Stanwick, T. L. Dempsey, M. P., およびStevenson, M.(1991)Science 254:423-427)。これは、HIV−1が、休止T細胞が続く抗原またはマイトジェン誘発T細胞活性化まで、非製造的染色体外状態で存続するモデルで示される。最近、休止T細胞でのHIVの複製に、HIV−1マトリックスタンパク質のチロシンホスホリレーションが必要であることが報告されている(Gallay, P. et al.,(1995)Cell 80:379)。
MHCクラスIIおよびTCRを架橋する特異的TCR Vβ遺伝子産物を発現するT細胞により認識される分子であるスーパー抗原は、細胞活性化、枯渇またはアネルギーを種々に導く。このタンパク質抗原グループは、多数の末梢血T細胞を活性化できる能力により特徴付けられる。哺乳動物レトロウイルスは、細胞性免疫反応性を阻止するために、または直接細胞活性の結果の複製を容易にするために、スーパー抗原をコードし得る。最近の報告は、HIV−1スーパー抗原がHIV−1感染に見られるT細胞枯渇を媒体し得ることを示している(Imberti, I., Sottini, A., Bettinardi, A. Pouti, M., およびPrimi, D.(1991)Science 254:860-862;Cameron, P. U., Frudentlial, P. S., Barker, J. M., Gezelter, S. Inaba, K., およびSteinman, R. M.(1992)Science 257:383-387;Laurence, J., Hodtsev, A. S., およびPosnett, D. N.(1992)Int. Conf. AIDS Jul 19-24;8(1):Th72;Pantaleo, G., Rebai, N., Graziosi, C., Lane, H. C., Sekaly, R. P., およびFauci, A. S.(1992)Int. Conf. AIDS Jul 19-24;8(1):Th71)。T細胞のスーパー抗原活性化により誘発されるシグナル伝達経路の性質はまだ議論の余地がある(Liu, H. Lampe, M. A., Iregui, M. V., およびCantor, H.(1991)Proc. Natl. Acad. Sci. USA 88(19):8705-8709;Kanner, S. B., Odum, N., Grosmaire, L., Mascwicz, S., Svejgaard, A., およびLedbetter, J. A.(1992)J. Immunol. 149(11):3482-3488;Oyaizu, N., Chirmule, N., Yagura, H., Pahwa, R. Good, R. A., およびPahwa, S.(1992)Proc. Natl. Acad. Sci. USA 89(17):8035-8039)。
実施例は、静止および増殖Tリンパ球を外来DNAの暴露から防御する、TCR−媒介シグナル伝達により抑制可能な活性化機構の存在を示す。T細胞は、トランスフェクション前のシグナル伝達に続いて、外来DNAのみを発現する。この発見は、細胞性増殖のみでは外来DNAの十分な発現をするのに不十分であり得るため、体細胞遺伝子治療の分野で意味を有する。従って、本発明は、増殖T細胞に導入された遺伝子の十分な発現の方法を提供する。本発明の一つの態様において、T細胞を個体から得、刺激してエクスビボで増殖させ、本発明の方法により遺伝子的に形質導入し、個体に再投与する。この具体的態様において、T細胞を、抗CD3抗体、ホルボールエステルとイオノマイシンの組み合わせのようなT細胞受容体媒体シグナル伝達を刺激する、抗CD3抗体のような作用物質、または作用物質の組み合わせまたはT細胞受容体を迂回する作用物質と接触させる。
本発明はまたウイルスDNAのような外来DNAの発現を阻止または減少させる方法を提供する。従って、ウイルスDNAのような外来DNAを含む一次T細胞は、刺激または増殖できるが、ウイルス複製は、例えば、T細胞の増殖を外来遺伝子発現に必要な機構を活性化しない本明細書に記載の作用物質で刺激することにより阻害する。
同等物
当業者は、慣用の実験のみを使用して、本明細書に記載の具体的態様の多くの同等物ができることを認識するか、気づく。このような同等物は以下の請求の範囲に含まれると意図する。
Claims (19)
- T細胞中で外来核酸分子の発現を増加させる方法であって:
(a)T細胞を少なくとも1つの刺激性作用物質とインビトロで接触させること(ここで、該刺激性作用物質は、スーパー抗原、ホルボールエステルとカルシウムイオノホアの組み合わせ、及び抗CD3抗体と抗CD28抗体の組み合わせからなる群から選択され、該T細胞は該少なくとも1つの刺激性作用物質と接触する前に増殖している)、及び
(b)外来核酸分子を段階(a)由来のT細胞に、該T細胞を接触させてから24時間以内にインビトロで導入すること、
を含み、それにより、T細胞における外来核酸分子の発現が、該少なくとも1つの刺激性作用物質と接触しないT細胞における発現と比較して増加する、方法。 - T細胞を少なくとも1つの刺激性作用物質と接触させる前に、T細胞の増殖を刺激する少なくとも1つの増殖作用物質とインビトロで接触させる、請求の範囲第1項に記載の方法であって、該増殖作用物質が、スーパー抗原、ホルボールエステルとカルシウムイオノホアの組み合わせ、及び抗CD3抗体と抗CD28抗体の組み合わせからなる群から選択される、方法。
- T細胞が一次T細胞である、請求の範囲第1項または第2項に記載の方法。
- 抗CD3抗体および抗CD28抗体がビーズ、組織培養皿または細胞表面に結合している、請求の範囲第1−3項のいずれかに記載の方法。
- T細胞が、該T細胞に核酸分子を導入する約1−24時間前に、少なくとも1つの刺激性作用物質と接触する、請求の範囲第1−4項のいずれかに記載の方法。
- T細胞が、該T細胞に核酸分子を導入する約10時間前に少なくとも1つの刺激性作用物質と接触する、請求の範囲第5項に記載の方法。
- T細胞がインビトロで更に刺激されその数が増加する、請求の範囲第1−6項のいずれかに記載の方法。
- 外来核酸分子を、エレクトロポレーション、リン酸カルシウム沈殿、DEAE−デキストラン処理、リポフェクション、マイクロインジェクション、細胞運搬ベヒクル、または可溶性分子複合体の形態によって、導入する、請求の範囲第1項に記載の方法。
- T細胞が、CD4+T細胞、CD8+T細胞、CD4+CD8+T細胞、CD4-CD8-T細胞、Th1細胞、及びTh2細胞からなる群から選択される、請求の範囲第1項に記載の方法。
- T細胞が、一次T細胞、T細胞の純粋な集団、T細胞クローン、健常個体由来のT細胞、感染性疾患に罹患した個体由来のT細胞、及び自己免疫疾患に罹患しているまたは罹患しやすい患者由来のT細胞からなる群から選択される、請求の範囲第1項に記載の方法。
- T細胞が、哺乳動物由来である、請求の範囲第1項に記載の方法。
- 外来核酸分子がDNA分子またはRNA分子である、請求の範囲第1項に記載の方法。
- 外来核酸分子が、天然核酸分子または合成核酸分子である、請求の範囲第1項に記載の方法。
- 外来核酸分子が、少なくとも1つのタンパク質をコードする、請求の範囲第1項に記載の方法。
- タンパク質が、分泌可能タンパク質である、請求の範囲第14項に記載の方法。
- 分泌可能タンパク質がサイトカイン、リンホカイン、または成長因子である、請求の範囲第15項に記載の方法。
- 外来核酸分子が、アデノシンデアミナーゼまたはgp39をコードする、請求の範囲第1項に記載の方法。
- 外来核酸分子が、1またはそれ以上の機能性RNA分子を発現する、請求の範囲第1項に記載の方法。
- 機能性RNA分子が、アンチセンスRNA分子またはリボザイムである、請求の範囲第18項に記載の方法。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US43509595A | 1995-05-04 | 1995-05-04 | |
| US08/435,095 | 1995-05-04 | ||
| US08/475,136 | 1995-06-07 | ||
| US08/475,136 US6692964B1 (en) | 1995-05-04 | 1995-06-07 | Methods for transfecting T cells |
| PCT/US1996/006200 WO1996034970A1 (en) | 1995-05-04 | 1996-05-02 | Improved methods for transfecting t cells |
Related Child Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005372607A Division JP2006109844A (ja) | 1995-05-04 | 2005-12-26 | T細胞の改良トランスフェクション法 |
| JP2007028118A Division JP4189007B2 (ja) | 1995-05-04 | 2007-02-07 | T細胞の改良トランスフェクション法 |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JPH11505419A JPH11505419A (ja) | 1999-05-21 |
| JPH11505419A5 JPH11505419A5 (ja) | 2005-05-12 |
| JP4031033B2 true JP4031033B2 (ja) | 2008-01-09 |
Family
ID=27030423
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP53351096A Expired - Lifetime JP4031033B2 (ja) | 1995-05-04 | 1996-05-02 | T細胞の改良トランスフェクション法 |
| JP2005372607A Withdrawn JP2006109844A (ja) | 1995-05-04 | 2005-12-26 | T細胞の改良トランスフェクション法 |
| JP2007028118A Expired - Lifetime JP4189007B2 (ja) | 1995-05-04 | 2007-02-07 | T細胞の改良トランスフェクション法 |
Family Applications After (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005372607A Withdrawn JP2006109844A (ja) | 1995-05-04 | 2005-12-26 | T細胞の改良トランスフェクション法 |
| JP2007028118A Expired - Lifetime JP4189007B2 (ja) | 1995-05-04 | 2007-02-07 | T細胞の改良トランスフェクション法 |
Country Status (7)
| Country | Link |
|---|---|
| US (1) | US20060246587A1 (ja) |
| EP (1) | EP0824594B1 (ja) |
| JP (3) | JP4031033B2 (ja) |
| AT (1) | ATE292689T1 (ja) |
| CA (1) | CA2220226C (ja) |
| DE (1) | DE69634563T2 (ja) |
| WO (1) | WO1996034970A1 (ja) |
Families Citing this family (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB9907366D0 (en) * | 1999-03-30 | 1999-05-26 | Medical Res Council | Method for expressing proteins |
| US7572631B2 (en) | 2000-02-24 | 2009-08-11 | Invitrogen Corporation | Activation and expansion of T cells |
| US7541184B2 (en) | 2000-02-24 | 2009-06-02 | Invitrogen Corporation | Activation and expansion of cells |
| EP1312670A4 (en) * | 2000-08-16 | 2005-11-30 | Takara Bio Inc | METHOD FOR THE EXTENSIVE CULTURE OF ANTIGEN-SPECIFIC CYTOTOXIC T CELLS |
| US6627442B1 (en) * | 2000-08-31 | 2003-09-30 | Virxsys Corporation | Methods for stable transduction of cells with hiv-derived viral vectors |
| KR100888915B1 (ko) | 2001-08-15 | 2009-03-16 | 다카라 바이오 가부시키가이샤 | 항원특이적 세포상해성 t 세포 확대배양방법 |
| JP4406566B2 (ja) | 2002-03-25 | 2010-01-27 | タカラバイオ株式会社 | 細胞傷害性リンパ球の製造方法 |
| KR101331746B1 (ko) | 2003-08-22 | 2013-11-20 | 다카라 바이오 가부시키가이샤 | 세포 상해성 림프구의 제조 방법 |
| EP1663308A1 (en) * | 2003-09-22 | 2006-06-07 | Xcyte Therapies, Inc | Compositions and methods to accelerate hematologic recovery |
| WO2006101205A1 (ja) * | 2005-03-25 | 2006-09-28 | Kaneka Corporation | T細胞の活性化方法及び活性化t細胞製造用キット |
| EP1916302A4 (en) | 2005-08-17 | 2009-10-21 | Takara Bio Inc | PROCESS FOR THE PRODUCTION OF LYMPHOCYTES |
| JP5156382B2 (ja) * | 2005-09-30 | 2013-03-06 | タカラバイオ株式会社 | T細胞集団の製造方法 |
| JP6754761B2 (ja) * | 2014-07-11 | 2020-09-16 | セルジーン コーポレイション | Tリンパ球へのベクター導入効率の改善方法 |
| CN107980059B (zh) * | 2015-04-13 | 2022-11-11 | 美克斯细胞有限公司 | 用于修饰基因组dna的方法和组合物 |
| US10294454B2 (en) | 2016-08-24 | 2019-05-21 | General Electric Company | Methods and kits for cell activation |
Family Cites Families (19)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1988000970A2 (en) * | 1986-08-08 | 1988-02-11 | Regents Of The University Of Minnesota | Method of culturing leukocytes |
| CA2003455C (en) * | 1988-11-23 | 2000-02-22 | Craig B. Thompson | Immunotherapy involving cd28 stimulation |
| US5858358A (en) * | 1992-04-07 | 1999-01-12 | The United States Of America As Represented By The Secretary Of The Navy | Methods for selectively stimulating proliferation of T cells |
| US5399346A (en) * | 1989-06-14 | 1995-03-21 | The United States Of America As Represented By The Department Of Health And Human Services | Gene therapy |
| US5272082A (en) * | 1992-03-30 | 1993-12-21 | The Wistar Institute Of Anatomy & Biology | Cytotoxic T-ALL cell lines and uses therefor |
| WO1994012196A1 (en) * | 1992-11-25 | 1994-06-09 | Tanox Biosystems, Inc. | Conjugates and constructs including anti-cd28 and anti-cd3 binding molecules |
| KR960701988A (ko) * | 1993-04-20 | 1996-03-28 | 윌리엄 에스. 로빈슨 | 세포내 감염원에 감염된 개체의 치료방법 및 치료물질(methods and materials for treatment of individuals infected with intracellular in-fectious agents) |
| ATE293686T1 (de) * | 1993-06-04 | 2005-05-15 | Us Navy | Verfahren zur selektiven stimulierung der t- zellproliferation. |
| FR2707091B1 (fr) * | 1993-06-30 | 1997-04-04 | Cohen Haguenauer Odile | Vecteur rétroviral pour le transfert et l'expression de gènes dans des cellules eucaryotes. |
| CA2167091A1 (en) * | 1993-07-26 | 1995-02-02 | Gordon J. Freeman | B7-2: ctl a4/cd 28 counter receptor |
| US5733543A (en) * | 1994-04-29 | 1998-03-31 | Nabel; Gary J. | Introduction of HIV-protective genes into cells by particle-mediated gene transfer |
| EP0764203A1 (en) * | 1994-06-03 | 1997-03-26 | THE UNITED STATES OF AMERICA as represented by THE SECRETARY OF THE NAVY | Methods for selectively stimulating proliferation of t cells |
| JPH10501815A (ja) * | 1994-06-07 | 1998-02-17 | リージェンツ・オブ・ザ・ユニバーシティ・オブ・ミネソタ | 抗原特異的t細胞応答の阻害方法 |
| US6576236B1 (en) * | 1994-07-01 | 2003-06-10 | Dana Farber Cancer Institute | Methods for stimulating T cell responses by manipulating a common cytokine receptor γ chain |
| US5795790A (en) * | 1994-07-20 | 1998-08-18 | Cytotherapeutics, Inc. | Method for controlling proliferation and differentiation of cells encapsulated within bioartificial organs |
| US5827642A (en) * | 1994-08-31 | 1998-10-27 | Fred Hutchinson Cancer Research Center | Rapid expansion method ("REM") for in vitro propagation of T lymphocytes |
| CA2198891A1 (en) * | 1994-08-31 | 1996-03-07 | Hartmut Wekerle | Genetically modified t cells |
| US6692964B1 (en) * | 1995-05-04 | 2004-02-17 | The United States Of America As Represented By The Secretary Of The Navy | Methods for transfecting T cells |
| US7067318B2 (en) * | 1995-06-07 | 2006-06-27 | The Regents Of The University Of Michigan | Methods for transfecting T cells |
-
1996
- 1996-05-02 CA CA002220226A patent/CA2220226C/en not_active Expired - Lifetime
- 1996-05-02 WO PCT/US1996/006200 patent/WO1996034970A1/en not_active Ceased
- 1996-05-02 JP JP53351096A patent/JP4031033B2/ja not_active Expired - Lifetime
- 1996-05-02 AT AT96913327T patent/ATE292689T1/de active
- 1996-05-02 DE DE69634563T patent/DE69634563T2/de not_active Expired - Lifetime
- 1996-05-02 EP EP96913327A patent/EP0824594B1/en not_active Expired - Lifetime
-
2005
- 2005-12-26 JP JP2005372607A patent/JP2006109844A/ja not_active Withdrawn
-
2006
- 2006-06-28 US US11/476,288 patent/US20060246587A1/en not_active Abandoned
-
2007
- 2007-02-07 JP JP2007028118A patent/JP4189007B2/ja not_active Expired - Lifetime
Also Published As
| Publication number | Publication date |
|---|---|
| CA2220226A1 (en) | 1996-11-07 |
| US20060246587A1 (en) | 2006-11-02 |
| WO1996034970A1 (en) | 1996-11-07 |
| JP2006109844A (ja) | 2006-04-27 |
| ATE292689T1 (de) | 2005-04-15 |
| DE69634563T2 (de) | 2006-02-16 |
| DE69634563D1 (de) | 2005-05-12 |
| EP0824594B1 (en) | 2005-04-06 |
| JPH11505419A (ja) | 1999-05-21 |
| CA2220226C (en) | 2008-10-21 |
| JP2007125041A (ja) | 2007-05-24 |
| JP4189007B2 (ja) | 2008-12-03 |
| EP0824594A1 (en) | 1998-02-25 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4189007B2 (ja) | T細胞の改良トランスフェクション法 | |
| US6692964B1 (en) | Methods for transfecting T cells | |
| US7172869B2 (en) | Methods for transfecting T cells | |
| Jensen et al. | CD20 is a molecular target for scFvFc: zeta receptor redirected T cells: implications for cellular immunotherapy of CD20+ malignancy | |
| EP3360961B1 (en) | Method for preparing genetically-modified t cells which express chimeric antigen receptor | |
| CN109880802B (zh) | 一种基于cd19和cd70的双重嵌合抗原受体基因修饰的免疫细胞及其应用 | |
| CN112218889B (zh) | 对人T细胞受体α恒定区基因具有特异性的优化的工程化核酸酶 | |
| Osterroth et al. | Stimulation of cytotoxic T cells against idiotype immunoglobulin of malignant lymphoma with protein-pulsed or idiotype-transduced dendritic cells | |
| Schakowski et al. | Novel non-viral method for transfection of primary leukemia cells and cell lines | |
| CN110462040A (zh) | 通过新基因开关表达系统调节多肽的表达 | |
| Berger et al. | Pharmacologically regulated Fas-mediated death of adoptively transferred T cells in a nonhuman primate model | |
| WO2020108644A1 (en) | Cd19-and cd22-based combined car-t immunotherapy | |
| JP2001517425A (ja) | T細胞共同刺激性ポリペプチド、モノクローナル抗体、ならびにその製法および使用 | |
| Berger et al. | CD28 costimulation and immunoaffinity-based selection efficiently generate primary gene-modified T cells for adoptive immunotherapy | |
| EP0463080A1 (en) | Cd8-based pharmaceuticals | |
| US5688690A (en) | Human cytotoxic lymphocyte signal transduction surface protein (P38) and monoclonal antibodies thereto | |
| EP1384781B1 (en) | Methods for inhibiting T-cell survival, by inhibiting BCL-XL protein level | |
| Cheng et al. | Increased lymphocyte apoptosis in Fas ligand transgenic mice | |
| JPH11504814A (ja) | Bcl−x▲下l▼タンパク質レベルを調節することによる、t細胞生存の調節法 | |
| Gardner et al. | Robust, but transient expression of adeno-associated virus-transduced genes during human T lymphopoiesis | |
| AU762356B2 (en) | Improved methods for transfecting T cells | |
| Pullen et al. | Bone marrow-derived macrophage expression of endogenous and transfected class II MHC genes during differentiation in vitro. | |
| Strair et al. | Retroviral mediated transfer and expression of exogenous genes in primary lymphoid cells: assaying for a viral transactivator activity in normal and malignant cells | |
| Rosenzweig et al. | In vitro T lymphopoiesis: a model system for stem cell gene therapy for AIDS | |
| US6268214B1 (en) | Vectors encoding a modified low affinity nerve growth factor receptor |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20040810 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20060425 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20060725 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20061010 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070207 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20070510 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20071002 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20071018 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101026 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20111026 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121026 Year of fee payment: 5 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121026 Year of fee payment: 5 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20131026 Year of fee payment: 6 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |