JP5975886B2 - Fc融合タンパク質を含む、免疫応答を増強するための方法および組成物 - Google Patents
Fc融合タンパク質を含む、免疫応答を増強するための方法および組成物 Download PDFInfo
- Publication number
- JP5975886B2 JP5975886B2 JP2012557275A JP2012557275A JP5975886B2 JP 5975886 B2 JP5975886 B2 JP 5975886B2 JP 2012557275 A JP2012557275 A JP 2012557275A JP 2012557275 A JP2012557275 A JP 2012557275A JP 5975886 B2 JP5975886 B2 JP 5975886B2
- Authority
- JP
- Japan
- Prior art keywords
- lda
- tumor
- cells
- fcγ2a
- mice
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images









Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/52—Cytokines; Lymphokines; Interferons
- C07K14/521—Chemokines
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/0005—Vertebrate antigens
- A61K39/0011—Cancer antigens
- A61K39/001102—Receptors, cell surface antigens or cell surface determinants
- A61K39/001116—Receptors for cytokines
- A61K39/001121—Receptors for chemokines
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/10—Cellular immunotherapy characterised by the cell type used
- A61K40/11—T-cells, e.g. tumour infiltrating lymphocytes [TIL] or regulatory T [Treg] cells; Lymphokine-activated killer [LAK] cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/10—Cellular immunotherapy characterised by the cell type used
- A61K40/19—Dendritic cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/20—Cellular immunotherapy characterised by the effect or the function of the cells
- A61K40/24—Antigen-presenting cells [APC]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/40—Cellular immunotherapy characterised by antigens that are targeted or presented by cells of the immune system
- A61K40/41—Vertebrate antigens
- A61K40/42—Cancer antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/40—Cellular immunotherapy characterised by antigens that are targeted or presented by cells of the immune system
- A61K40/41—Vertebrate antigens
- A61K40/42—Cancer antigens
- A61K40/4256—Tumor associated carbohydrates
- A61K40/4258—Gangliosides, e.g. GM2, GD2 or GD3
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/57—Medicinal preparations containing antigens or antibodies characterised by the type of response, e.g. Th1, Th2
- A61K2039/572—Medicinal preparations containing antigens or antibodies characterised by the type of response, e.g. Th1, Th2 cytotoxic response
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2239/00—Indexing codes associated with cellular immunotherapy of group A61K40/00
- A61K2239/31—Indexing codes associated with cellular immunotherapy of group A61K40/00 characterized by the route of administration
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2239/00—Indexing codes associated with cellular immunotherapy of group A61K40/00
- A61K2239/38—Indexing codes associated with cellular immunotherapy of group A61K40/00 characterised by the dose, timing or administration schedule
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/30—Non-immunoglobulin-derived peptide or protein having an immunoglobulin constant or Fc region, or a fragment thereof, attached thereto
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Epidemiology (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Medicinal Chemistry (AREA)
- Pharmacology & Pharmacy (AREA)
- Biophysics (AREA)
- Immunology (AREA)
- Molecular Biology (AREA)
- Mycology (AREA)
- Microbiology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Genetics & Genomics (AREA)
- Biochemistry (AREA)
- Gastroenterology & Hepatology (AREA)
- Zoology (AREA)
- Toxicology (AREA)
- Oncology (AREA)
- Cell Biology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Description
KCKRQCGPGPGAKFVAAWTLKAAAGPGPGCKRKIHIGPGQAFYTGPGPGEDPSHSLGLDAALFM
配列番号2の1〜6位アミノ酸は、スペーサーを構成する。7〜11および25〜29位のアミノ酸は、リンカーを構成する。どのTヘルパー・エピトープも本発明との関連で使用できると考えられるが、好適なTヘルパー・エピトープの非限定的な実施形態は、非天然panDRエピトープ(PADRE)[AKFVAAWTLKAAA:配列番号3]およびHIVgp120糖タンパク質のV3ループ[CKRKIHIGPGQAFYT:配列番号4]と称される普遍的なThペプチドを含む。これらの代表的なTヘルパー・エピトープのそれぞれは、配列番号Xにも示される。
ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSSGLYSLSSVVT VPSSSLGTQT YICNVNHKPS NTKVDKKVEP KSCDKTHTCP PCPAPELLGGPSVFLFPPKP KDTLMISRTP EVTCVVVDVS HEDPEVKFNW YVDGVEVHNA KTKPREEQYN STYRVVSVLT VLHQDWLNGK EYKCKVSNKA LPAPIEKTIS KAKGQPREPQ VYTLPPSRDE LTKNQVSLTC LVKGFYPSDI AVEWESNGQP ENNYKTTPPV LDSDGSFFLY SKLTVDKSRW QQGNVFSCSV MHEALHNHYT QKSLSLSPGK(配列番号7)
6〜8週齢のメスのA/Jマウスを、Jackson Laboratory(メイン州、バーハーバー)から入手した。実験手順は、ロズウェル・パークがん協会の施設内実験動物委員会によって承認されたプロトコルに従って行われた。マウスのNXS2神経芽細胞腫株化細胞(カリフォルニア州、ラ・ホーヤ、スクリプス研究所のDr.R.A.Reisfeldによって提供された)は、GD2-ネガティブC1300マウスの神経芽細胞腫(A/Jバックグラウンド)とGD2-ポジティブマウスの後根神経節腫細胞とのハイブリッドである。前記ハイブリッド株化細胞は、そのH-2Kk-ポジティブ/H-2Kbネガティブ表現型によってA/Jマウスと同系のMHCクラスIであることが示された(Lode, H. N., R. Xiang, T. Dreier, N. M. Varki, S. D. Gillies, and R. A. Reisfeld. 1998. Natural killer cell-mediated eradication of neuroblastoma metastases to bone marrow by targeted interleukin-2 therapy. Blood 91: 1706-1715.)。GD2-特異的mAbを分泌する14G2aハイブリドーマ株化細胞(Mujoo, K., T. J. Kipps, H. M. Yang, D. A. Cheresh, U. Wargalla, D. J. Sander, and R. A. Reisfeld. 1989. Functional properties and effect on growth suppression of human neuroblastoma tumors by isotype switch variants of monoclonal antiganglioside GD2 antibody 14.18. Cancer Res 49: 2857-2861)は、Dr.R.A.Reisfeld(スクリプス研究所)によって提供された。
組織プラスミノーゲン活性化因子分泌性の(tPA)シグナル配列を有する47-LDA発現ベクターの作成 2つの普遍的なThペプチドPADRE[AKFVAAWTLKAAA:配列番号3](Alexander, J., M. F. del Guercio, A. Maewal, L. Qiao, J. Fikes, R. W. Chesnut, J. Paulson, D. R. Bundle, S. DeFrees, and A. Sette. 2000. Linear PADRE T helper epitope and carbohydrate B cell epitope conjugates induce specific high titer IgG antibody responses. J Immunol 164: 1625-1633)、および、HIV gp120 糖タンパク質のV3ループ[CKRKIHIGPGQAFYT:配列番号4](Ahluwalia, A., K. Gokulan, I. Nath, and D. N. Rao. 1997. Modification of delivery system enhances MHC nonrestricted immunogenicity of V3 loop region of HIV-1 gp120. Microbiol Immunol 41: 779-784)が、47-LDA模倣ペプチドとともに他で報告されている(Bolesta, E., A. Kowalczyk, A. Wierzbicki, P. Rotkiewicz, B. Bambach, C. Y. Tsao, I. Horwacik, A. Kolinski, H. Rokita, M. Brecher, X. Wang, S. Ferrone, and D. Kozbor. 2005. DNA vaccine expressing the mimotope of GD2 ganglioside induces protective GD2 cross-reactive antibody responses. Cancer Res 65: 3410-3418.)。マウスの47-LDA-Fcγ2a融合タンパク質は、EcoRIおよびBglII制限酵素切断部位を使用して、pFUSE-mIgG2Aa-Fc1ベクター(カリフォルニア州、サンディエゴ、InvivoGen社)のhEF1-HTLVプロモーターとマウスIgG2aFc領域の間に、47-LDAポリペプチドコード配列を挿入することによって作成された。293T細胞の安定な形質移入は、47-LDA-Fcγ2a融合タンパク質コンストラクト、またはリポフェクタミン試薬を使用する偽のベクター(カリフォルニア州、カールズバッド、Invitrogen社)を用いて行われ、続いてゼオシン(zeocin)含有培地で選択を行った。47-LDA-Fcγ2a融合タンパク質は、トランスフェクト細胞の培養上清から、プロテインGカラム(ニュージャージー州、ピスカタウェイ、GE Healthcare, Bio-Sciences社)で精製された。融合タンパク質は、製造者のプロトコルに従い、SDS-15%PAGEと14G2a mAbを用いた免疫ブロット法によって、続いてECL+ウエスタンブロット検出システム(Amersham Pharmacia Biotech社)によって分析された。
DCsは、前述の通り、BM前駆物質からインビトロで産生した(Brinker, K. G., H. Garner, and J. R. Wright. 2003. Surfactant protein A modulates the differentiation of murine bone marrow-derived dendritic cells. Am J Physiol Lung Cell Mol Physiol 284: L232-241)。簡単に説明すると、BM細胞は、6〜8週齢のメスのA/Jマウス(n=5)の脛骨および大腿骨から収集し、その後、10ng/mlのGM-CSFを追加した完全培地中で37℃で6日間培養した。前記培地は2〜3日ごとに補充された。7日目に、ほとんどの非付着性細胞は、DC形態を獲得し、フローサイトメトリー分析によって特定されるように、CD11cが高く、CD80、CD86、CD40およびMHCクラスIIが低かった。DCsは5時間、10μ/mlの47-LDA-Fcγ2a融合タンパク質または47-LDAポリペプチド(マサチューセッツ州、ガードナー、New England Peptide社)でパルスされ、成熟を誘発するために1時間、LPS(1.0μg/ml)とともにインキュベートされ、洗浄され、マウスに静脈内注射(2×106)された(図1A)。20μgのDNAプラスミド-コード化IL-15およびIL-21サイトカインは、ワクチン接種時および5日後に、それぞれ筋肉内注射された。
未熟DCsは、ビオチン標識47-LDA-Fcγ2a融合タンパク質または47-LDAポリペプチドとともに、続いてストレプトアビジン-結合PEとともにインキュベートされた。いくつかの実験で、DCsは以下のmAbsを用いて染色された:抗-CD11c-FITC、抗-CD80-PE、抗-CD86-PE、抗-CD40-PEまたは抗-MHCクラスII-PE(カリフォルニア州、サンノゼ、BD Bioscience社)。脾細胞は、抗-CD4-PE、抗-CD8-PE、抗-CD25-FITC(BD Bioscience社)あるいは関連するアイソタイプ・コントロールを用いて標識された。特異的な抗体染色の前に、細胞はFcブロッカー(抗-CD16/CD32mAb)とともに10分間インキュベートされた。前記細胞は、0.01%のアジ化ナトリウムと1%のFCSを含むHBSS中で二度洗浄され、1%のパラホルムアルデヒドで固定され、分析前は暗所で4℃にて貯蔵された。バックグラウンド染色は、適切な蛍光色素-結合もしくは非結合マウスIgG1、IgG2aまたはIgG2b(BD Bioscience社)を含むアイソタイプ・コントロールを用いて評価された。腋窩、上腕リンパ節および脾臓内のTreg細胞の数は、製造者のプロトコルに従って、抗-FoxP3-AlexaFluor 647 mAb(カリフォルニア州、サンディエゴ、eBioscience社)を用いたCD4+CD25+リンパ球の細胞内染色によって決定された。INF-γあるいはTNF-αを分泌するCD4+およびCD8+脾細胞の数は、抗-IFN-γ-PEあるいは抗-TNF-α-PE mAb(BD Bioscience社)と協同した抗-CD4-FITCおよび抗-CD8-FITC mAbsを用いて決定された。LPS-刺激DCsにおけるIL-12p70の細胞内発現は、IL-12p35サブユニット内のエピトープを検出し、IL-12p40と交差反応しないラット抗マウスIL-12p70 mAb(R&D Systems社)を用いた染色によって測定した。LPS-刺激DCsにおけるCCL22の細胞内発現は、ラット抗マウスCCL22 mAb(R&D Systems社)を用いた染色によって決定した。47-LDA-Fcγ2a+、47-LDAポリトープ+あるいはCD86+DCsの選別は、BD FACSAria(登録商標)フローサイトメーター(ニュージャージー州、フランクリン・レイクス、BD社)を用いて実施された。全てのフローサイトメトリー評価は、FACScanもしくはFACSCaliburフローサイトメーターを用いて実施された。前方および側方散乱パラメーターにゲーティング(gating)後、少なくとも10,000のゲート化イベント(gated events)がルーチン的に獲得され、CellQuestソフトウェア(BD Bioscience社)を用いて分析された。
A/Jマウス(1グループあたりn=8〜10)は、図1Aに描写されるように、2×106のNXS2神経芽細胞腫細胞を皮下注射され、15日後、47-LDA-Fcγ2a-もしくは47-LDAポリトープ-DC-免疫化マウスから単離したCD8+強化脾細胞を用いた静脈内注射で治療された。CD8+脾細胞は、製造者の説明書に従って、抗マウスCD4(L3T4)および抗マウスCD45R(B220)mAbsと結合した常磁性ミクロビーズ(Miltenyi Biotec社のMACS)を使用してネガティブに選択された。単離されたCD8+脾細胞(2×107)は、LPS-成熟47-LDAポリペプチド-もしくは47-LDA-Fc融合タンパク質-被覆DCsとともにインキュベートされた。T細胞およびDCsの混合物(比率20:1)は、下記の文献に記載されるように、rmIL-2(1μg/dose)の存在下、リンパ球枯渇NXS2担がんマウスに静脈内注射された(Zeng, R., R. Spolski, S. E. Finkelstein, S. Oh, P. E. Kovanen, C. S. Hinrichs, C. A. Pise-Masison, M. F. Radonovich, J. N. Brady, N. P. Restifo, J. A. Berzofsky, and W. J. Leonard. 2005. Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function. J Exp Med 201: 139-148)。担がんマウスにおけるリンパ球減少症は、ACTおよびワクチン接種の一日前に、5Gyの骨髄非破壊的なTBIもしくは9Gyの骨髄破壊的なTBIおよびBM移入(107細胞)によって誘発された(図1B)。確立された皮下NXS2腫瘍を有するコントロールマウスは、5 Gyもしくは9Gyで照射され、且つBM 移植された。マウスは、2週間間隔で、下記の文献に記載されるように、IL-15およびIL-21ベクターの存在下、DCワクチンで免疫化された(Kowalczyk, A., A. Wierzbicki, M. Gil, B. Bambach, Y. Kaneko, H. Rokita, E. Repasky, R. Fenstermaker, M. Brecher, M. Ciesielski, and D. Kozbor. 2007. Induction of protective immune responses against NXS2 neuroblastoma challenge in mice by immunotherapy with GD2 mimotope vaccine and IL-15 and IL-21 gene delivery. Cancer Immunol Immunother 56: 1443-1458)。腫瘍増殖は、週に一度から三度、マイクロキャリパーを用いて皮下腫瘍を測定し、腫瘍体積(幅×長さ×幅/2=mm3)を決定することによってモニターされた。
47-LDA-Fcγ2a-もしくは47-LDAポリペプチド-被覆DCsで免疫化されたA/Jマウスから得られた脾細胞は、20:1の比率にて47-LDA発現DCsで一晩刺激された後、INF-γおよびTNF-α発現について分析された。偽ベクター免疫化マウスから単離された細胞は、コントロールとして利用された。47-LDA-DCおよび47-LDA-Fcγ2a-DCワクチンで免疫化されたマウスにおけるTreg細胞の誘発を研究するために、脾臓、腋窩および上腕リンパ節が最後の免疫化の3週間後に免疫化マウスから除去され、抗-CD4-PE、抗-CD25-FITCおよび抗-FoxP3-Alexa Fluor 647 mAbs、または関連するアイソタイプ・コントロールを用いた染色によってTreg細胞の数が分析された。Teff細胞増殖へのTreg細胞の効果を分析するために、47-LDA-DCもしくは47-LDA-Fcγ2a-DCワクチンで免疫化されたマウス由来のCD8+T細胞は、37℃にて10分間、25μMのカルボキシフルオセインサクシニミジルエステル(CFSE:オレゴン州、ユージーン、Molecular Probes社)を負荷され、Treg細胞の存在下もしくは不存在下(比率1:1)、72時間、47-LDA-発現同系DCs(比率20:1)とともに培養された。細胞は、PE-結合抗-CD8 mAbsで染色され、フローサイトメトリーで分析された。Treg細胞集団は、製造者のプロトコルに従って、CD4+CD25+調節性T細胞単離キットを用いて単離された(Miltenyi社)。
脾細胞は、外因性IL-2のソースとして、15%のT細胞刺激因子(T-STIM[登録商標] カルチャー・サプリメント;マサチューセッツ州、ベッドフォード、Collaborative Biomedical Products社)中で20:1の比率にて47-LDA発現DCsとともに培養された。刺激の3日後、細胞は分裂され、マウスのrIL-2(0.3ng/ml)(BD Bioscience社)を追加した培地中で培養された。刺激前に、CD8+T細胞は、製造者のプロトコルに従ってT細胞富化カラム(Miltenyi社)を使用したネガティブ・セレクションによって単離された。NXS2腫瘍細胞に対するCTLsの細胞溶解活性は、スタンダード4-h51Cr-リリース・アッセイによって5日後に分析された。特異的溶解のパーセントは、次のように算定された:([cpm実験的リリース−cpm自発的リリース]/[cpm最大リリース−cpm自発的リリース])×100。最大リリースは、5%のTriton X-100の添加によって、溶解された細胞の上清から定量された。自発的リリースは、培地のみとインキュベートされた標的細胞から定量された。
本発明における使用に好適なrOVVsを作成できる種々の方法が存在する。ある特定の実演において、rOVVs発現EGFPおよび47-LDA-Fcγ2a融合タンパク質が、VSC20ワクシニアウイルスおよびワクシニア・シャトル・プラスミドpSEL-EGFP並びにpCB023-47-LDA-Fcγ2aをそれぞれ使用してCV-1細胞における相同組換えによって産生された。親ワクシニアウイルスは、VGF遺伝子の代わりにクローン化されたLacZ遺伝子を有するVSC20であった。VSC20ワクシニアウイルスは、米国特許公開公報2007/0154458号に記述されており、そのVSC20ウイルスに関する記述は参照により本明細書に組み込まれる。
グループ間の差異の統計的有意性が、等しい分散を想定する両側スチューデントt検定を用いて実施された。平均腫瘍増殖の対(ペアワイズ)グループ比較のためのP値は、ノンパラメトリック・ウィルコクソン順位和検定を使用してコンピューターで計算された。カプラン・マイヤー生存プロットが作成され、生存期間中央値が、マウスのNXS2-チャレンジグループについて決定された。複数のグループにわたる生存時間の統計的有意差は、ログランクMantel-Cox法を用いて評価された。データは、算術的な平均値±標準偏差として表され、ウィンドウズ・ベースのプラットフォーム上でJMPプログラム(ノースカロライナ州、カリー、SAS Institute社)を用いて分析された。
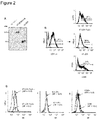
マウスの 47-LDA-Fcγ2a 融合タンパク質は、EcoRIおよび BglII制限酵素切断部位を使用して、pFUSE-mIgG2Aa-Fc1ベクターのhEF1-HTLVプロモーターと、マウスIgG2a Fc領域との間のインフレームに、関連するコード配列[組織プラスミノーゲン活性化因子分泌性(tPA)シグナル配列、2つの普遍的なThペプチド、および47-LDAミモトープを含む]を挿入することによって産生された。無損傷のオープン・リーディング・フレームの存在を確認するための47-LDA-Fcγ2a構築物の配列決定後、14G2a mAbを用いた47-LDA-Fcγ2-トランスフェクト293T細胞の免疫蛍光染色と、それに続くフローサイトメトリー分析が、融合タンパク質の発現を判定するためにトランスフェクションから48時間後に行われた(図示せず)。ゼオシン-含有培地中で選択された、47-LDA-Fcγ2a安定発現株からの融合タンパク質の分泌は、14G2a mAbを用いて培養上清の免疫ブロット法によって確認された。図2Aに示すように、31.9kDaの顕著なバンドが、47-LDA-Fcγ2aトランスフェクタントから収集された培養上清中で、14G2a mAbを用いて検出された。偽プラスミドをトランスフェクトされた細胞から調製された上清にはバンドは存在しなかった。ポジティブ・コントロールとして分析に含まれた、6.6kDaの分子量の47-LDAポリペプチドは、14G2a mAbによってはっきりと認識された。ようするに、これらの結果は、47-LDA-Fcγ2a 融合タンパク質と関連して発現されたGD2ミモトープが、14G2a mAbによって認識される合成ペプチドの生来の抗原決定基を保持することを示した。
LPS-刺激47-LDA-Fcγ2a+DCおよび47-LDA+DCsの間のIL-12とCCL22の発現レベルの差は、これらの細胞が、インビボでTeffおよびTreg細胞と相互作用する能力が異なる可能性が有ることを示唆する。それゆえ、我々は、47-LDA-Fcγ2a-および47-LDA-DCワクチンを用いたA/Jマウスの免疫化後の、腫瘍特異的なTh1およびCTL応答の誘発を研究した(図1A)。各DCワクチンは、2週間間隔で3回静脈内注射によって送達され、これとともにDNAプラスミド-コード化IL-15およびIL-21がワクチン接種時および5日後にそれぞれ筋肉内に送達された(Kowalczyk, A., A. Wierzbicki, M. Gil, B. Bambach, Y. Kaneko, H. Rokita, E. Repasky, R. Fenstermaker, M. Brecher, M. Ciesielski, and D. Kozbor. 2007. Induction of protective immune responses against NXS2 neuroblastoma challenge in mice by immunotherapy with GD2 mimotope vaccine and IL-15 and IL-21 gene delivery. Cancer Immunol Immunother 56: 1443-1458)。最後の免疫化の3週間後、免疫化されたマウス由来のCD4+脾細胞は、47-LDA発現DCsで一晩刺激した後、細胞内染色およびフローサイトメトリーによってINF-γおよびTNF-α産生について分析された。DCsおよびIL-15ならびにIL-21ベクターを用いて免疫化されたマウスから単離された細胞は、コントロールとして含まれた。図3Cに示すように、IFN-γ-およびTNF-α-産生CD4+細胞の数は、47-LDA-Fcγ2a融合タンパク質-被覆DCsで免疫化されたマウスにおいて、コントロールの動物より4倍以上高く、47-LDA-DCワクチンによって誘発された応答より2倍以上高かった(それぞれ、P=0.002およびP=0.003)。E:T比の広範囲にわたって、47-LDA-Fcγ2a融合タンパク質ワクチン後のIFN-γ-産生CD4+T細胞応答の増加は、NXS2神経芽細胞腫瘍に対するより高いCD8+T細胞性細胞傷害活性(47-LDAポリトープ-被覆DCsで誘発されたものと比較して)と関連した(図3D)。
47-LDA-および47-LDA-Fcγ2a-DCワクチンの治療的有効性を比較するために、我々は、NXS2担癌A/Jマウスにおいて、抗原経験CD8+脾細胞を用いたACT実験を行った(図1B)。CD8+T細胞が、Teff細胞の増殖を誘発するために、およびリンパ球枯渇ホストへの養子移入による二次応答を開始するために、47-LDA-もしくは47-LDA-Fcγ2a-DCワクチンとともに、NXS2-担持マウスに静脈内注射された。図5Aは、ACTおよび47-LDAポリペプチド-被覆DCワクチンに先立つ骨髄非破壊的な(5 Gy)投与計画でのリンパ球枯渇が、腫瘍増殖の抑制にバックグラウンド作用を持つことを示す(P=0.28)。養子導入CD8+T細胞の抗腫瘍有効性は、治療マウスの全生存期間が25日から70日に延長したことから判定されるように、骨髄破壊的な(9 Gy)TBI後に増強された(図5A;P=0.007)。我々は、骨髄破壊的な投与計画が、7-LDA-Fcγ2a-DCsとともに送達された養子導入CD8+脾細胞との組み合わせで特に効果的であることを見い出した。図5Bに示すように、後者の治療は、未処置マウス由来の同系BMの移植を伴う骨髄破壊的な量のTBIを受けたNXS2-担持動物の22%において腫瘍増殖の完全寛解をもたらした(P=0.0003)。骨髄非破壊的な状況において47-LDA-Fcγ2a-DCワクチンで刺激されたCD8+T細胞を用いた治療は、完全な腫瘍の消散を示さなかったが、コントロール群と比較した際、治療マウスにおいて生存期間の有意な延長があったことは注目に値する(図5B;P=0.015)。全てのコントロールマウスが、進行性に増大する腫瘍を発症し、25日までに屠殺されなければならなかったので、ACTの抗腫瘍効力は特異的であった。
原発腫瘍の切除後、自然転移を起こすNXS2神経芽細胞腫の能力は、播種性疾患をコントロールするために、養子導入CD8+脾細胞の有効性を研究するためのモデルを提供した(図1C)。Treg細胞の枯渇と協同した根治目的の手術が、持続的な腫瘍保護の発展とCD8+細胞記憶を可能にするという累積証拠を考慮して(9)、NXS2担癌マウスは、腫瘍切除および抗原経験CD8+脾細胞のACT時に骨髄非破壊的な(5 Gy)もしくは骨髄破壊的な(9 Gy)TBIを受けた。図5Cに示すように、47-LDAポリペプチド-被覆DCsによって増殖されたCD8+T細胞の移入は、骨髄破壊マウスにおいてのみ観察された転移性疾患の進行を遅延した(P=0.024)。無腫瘍生存率は、この動物のグループでは観察されなかった。
我々は、次に、治療開始前に原発腫瘍が切除された骨髄破壊マウスへの未処置脾細胞の養子移入後に、転移性疾患を抑制できる抗腫瘍性免疫応答を誘発する、47-LDA-および47-LDA-Fcγ2a-DCワクチンの能力を研究した(図5E)。CD4+T細胞は、増殖因子を有するCD8+T細胞を提供するため、および抗腫瘍性応答を誘発するためにも要求されるので、これらの実験では、未処置脾細胞の全集団が、ACTのために使用された。図5Eは、NXS2-担持マウスにおいてインビボで刺激した未処置のT細胞の測定可能な治療的影響は、抗原経験CD8+T細胞によって媒介されたものと比較して低かったものの、20%の治療マウスは、47-LDA-Fcγ2a-DCワクチンを用いたACT後100日間、腫瘍が無いままであったことを示す(P=0.007)。47-LDA+DCワクチンは、たとえより小さい程度でも、抗腫瘍性T細胞応答を誘発する能力もあった。原発腫瘍が切除されたコントロールマウスは、治療の最初の二ヵ月以内に転移性疾患を発症し、NXS2-担持マウスにおける47-LDA-Fcγ2a-DCワクチンを用いた能動免疫によって誘発される腫瘍特異的T細胞の有意な効果を示した。
我々は、次に、ACTおよびDCワクチン後の、担がんマウスおよび腫瘍の無いマウスにおける細胞応答を研究した。治療的な47-LDA-および47-LDA-Fcγ2a-DCワクチンの両方が、養子導入される未処置脾細胞と一緒に、原発性NXS2腫瘍の切除後、骨髄破壊マウスに送達され、Teff細胞の誘発を分析された。ACTおよび47-LDA+DCワクチンでの免疫化後、無腫瘍生存率は観察されなかったので、リンパ節に可視の転移巣を有する動物は、脾臓における抗腫瘍性免疫応答について試験された。図6Aに示すように、担がんマウス中のCD4+あるいはCD8+脾細胞の15%未満が、47-LDA発現DCsでの72時間の刺激後、インビトロで分裂し、NXS2細胞へのCTL応答は、バックグラウンドレベルであった(図6B)。対照的に、CD4+およびCD8+脾細胞の力強い増殖(それぞれ、55%から69%および39%から46%の範囲)が、ACTおよび47-LDA-Fcγ2a-DCワクチンを受けた後で腫瘍の無いマウスにおいて検出された(図6C)。増殖応答は、NXS2細胞に対する抗腫瘍性CTL活性も伴っていた(図6D)。
47-LDA-Fcγ2a-DCワクチンの抗腫瘍性効果が照射された担がんホストにおけるACTに限定されないことを決定するために、我々は次に、rOVVワクシニアウイルスによって発現された47-LDA-Fcγ2a融合タンパク質の治療的な有効性を試験した。ワクシニアrOVV-47-LDA-Fcγ2aの腫瘍退縮性TK-,VGF-変異体が腫瘍特異的な複製を示すため、融合タンパク質が感染細胞から分泌され、腫瘍退縮部位で、腫瘍浸潤DCsによって交差提示されるであろうことが予測された。並行研究において、NXS2担がんマウスの追加グループが、ウイルス感染したNXS2細胞からリリースされる腫瘍関連抗原によって誘発される保護の程度を決定するために、特異性コントロールとして、rOVV-EGFPベクターを注射された。動物は、一週間に一度〜三度、皮下腫瘍を測定することによって腫瘍増殖を試験された。図7Aに示すように、コントロールrOVV-EGFPベクターを用いて治療されたNXS2担持マウスの約25%が、腫瘍が無い状態になった。対照的に、ウイルスベクターの代わりにPBSを用いて治療された全ての担がんマウスは、進行性に増大する腫瘍を発症し、20日までに屠殺されなければならなかった。最も高い抗腫瘍有効性は、rOVV-47-LDA-Fcγ2aベクターを用いた腫瘍退縮性免疫ウイルス療法後、担がんマウスで観察された、100日の間60%を超える無腫瘍生存率によって特徴付けられる(図7A;P=0.0004)。
メスのBALB/c(n=8〜12)は、胸部の乳房脂肪体中に、50μlのPBS中7×104の4T1細胞を接種された。100〜150mm3の腫瘍体積を有する4T1-担持マウスは、OVV-EGFPもしくはOVV-CXCR4A-Fc(108PFU)単独で、または前記ウイルスの24時間後腹腔内に送達される最大耐用量のASA404(20mg/kg)との組み合わせで、静脈内に注射された。今回のASA404送達は、前記ウイルスの4時間前または同時に送達された薬物で治療された動物と比較して最適な抗腫瘍効率を示した(データは示さず)。図10Aは、OVV-CXCR4-A-Fcもしくは ASA404を用いた単独療法は、腫瘍増殖を有意に抑制したが(P<0.05)、無腫瘍生存率は、併用療法で治療された動物でのみ達成されたことを示す(図10B)。OVV-EGFPとASA404を用いた治療は、13%の生存者をもたらし、一方、OVV-CXCR4-FcとASA404は、42%の腫瘍の無い動物をもたらした(図10B)。転移性疾患の治療のために、4T1担持マウス(V=〜200mm3)は、腫瘍ウイルス療法を単独でもしくはASA404との併用で受け、腫瘍は8日後(ウイルス感染が、ホストの自然および獲得免疫が原因で除去された時点;データは示さない)に切除された。コントロールマウスは、治療なしで原発腫瘍を切除された。図10Cに示すように、転移性疾患に対する最も高い保護は、OVV-CXCR4-FcとASA404の組み合わせで治療された動物で観察された(80%生存率)。腫瘍ウイルス療法またはASA404治療単独では、保護効果はより小さかった。生存特性における変化に対応して、治療の4日後に切除された4T1腫瘍中のGr1+CD11b+(Ly6G+)MDSCsの数は、コントロール動物での数とは異なった。
Claims (2)
- 融合タンパク質をコードするポリヌクレオチドを含む組成物であって、
前記融合タンパク質が、免疫グロブリンFc部および腫瘍細胞によって発現されるレセプターのアンタゴニスト・ペプチドを含むこと、
前記アンタゴニスト・ペプチドが、配列 KGVSLSYR(配列番号6)からなること、
前記融合タンパク質の発現後、2つの融合タンパク質のFc部の間にジスルフィド結合が生じ、2つの前記アンタゴニスト・ペプチドの単量体ユニットに空間的な近接をもたらすこと、
前記単量体ユニットの間に連結アミノ酸が存在しないこと
を特徴とする組成物。 - 請求項1に記載のポリヌクレオチドによってコードされるタンパク質を含む組成物。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US31298210P | 2010-03-11 | 2010-03-11 | |
| US61/312,982 | 2010-03-11 | ||
| PCT/US2011/028070 WO2011112915A2 (en) | 2010-03-11 | 2011-03-11 | Methods and compositions containing fc fusiong proteins for enhancing immune responses |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2013529178A JP2013529178A (ja) | 2013-07-18 |
| JP2013529178A5 JP2013529178A5 (ja) | 2014-05-01 |
| JP5975886B2 true JP5975886B2 (ja) | 2016-08-23 |
Family
ID=44564143
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2012557275A Active JP5975886B2 (ja) | 2010-03-11 | 2011-03-11 | Fc融合タンパク質を含む、免疫応答を増強するための方法および組成物 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US9296803B2 (ja) |
| EP (1) | EP2545174B1 (ja) |
| JP (1) | JP5975886B2 (ja) |
| CN (1) | CN102884194B (ja) |
| WO (1) | WO2011112915A2 (ja) |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2010226726B8 (en) | 2009-03-16 | 2014-08-14 | Pangu Biopharma Limited | Compositions and methods comprising histidyl-tRNA synthetase splice variants having non-canonical biological activities |
| ES2708565T3 (es) | 2013-03-15 | 2019-04-10 | Atyr Pharma Inc | Conjugados de Fc-histidil-ARNt sintetasa |
| EP3548070A4 (en) * | 2016-11-29 | 2020-07-22 | Health Research, Inc. | METHODS AND COMPOSITIONS FOR ANTI-CANCER THERAPY |
| CA3060514A1 (en) | 2017-04-20 | 2018-10-25 | Atyr Pharma, Inc. | Compositions and methods for treating lung inflammation |
| RU2676317C1 (ru) * | 2017-08-30 | 2018-12-27 | федеральное государственное автономное образовательное учреждение высшего образования Первый Московский государственный медицинский университет имени И.М. Сеченова Министерства здравоохранения Российской Федерации (Сеченовский университет) | ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pCXCRs-Fc ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО СЛИТОГО БЕЛКА, ОБЛАДАЮЩЕГО АНТИ-IL-8 АКТИВНОСТЬЮ |
| CN116375881A (zh) * | 2021-12-31 | 2023-07-04 | 广州国家实验室 | 融合蛋白疫苗 |
| CN116059388A (zh) * | 2022-07-25 | 2023-05-05 | 国家纳米科学中心 | 一种多肽-化疗药物偶联物、其制备方法和应用 |
| CN117003890B (zh) * | 2023-08-09 | 2024-06-04 | 成都新诺明生物科技有限公司 | 一种含有P2和两个Fc的gE融合蛋白及其制备方法和应用 |
Family Cites Families (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP0906954A1 (en) | 1997-09-29 | 1999-04-07 | Applied Research Systems ARS Holding N.V. | Amino-terminal truncated c-c chemokines as chemokine antagonist |
| CA2245224A1 (en) * | 1998-08-14 | 2000-02-14 | Jiang-Hong Giong | Chemokine receptor antagonists and chemotherapeutics |
| CA2405290C (en) * | 2000-04-13 | 2011-06-21 | Bio Life Science Forschungs Und Entwicklungsgesellschaft Mbh | Vaccine against cancerous diseases |
| AU5943201A (en) * | 2000-05-03 | 2001-11-12 | Amgen Inc | Modified peptides as therapeutic agents |
| US7465790B2 (en) * | 2000-10-09 | 2008-12-16 | Isis Innovation, Inc. | Therapeutic antibodies |
| US20050002939A1 (en) * | 2002-12-23 | 2005-01-06 | Albert Zlotnik | Tumor killing/tumor regression using CXCR4 antagonists |
| US20080108664A1 (en) * | 2005-12-23 | 2008-05-08 | Liu Belle B | Solid-state form of AMG 706 and pharmaceutical compositions thereof |
| FR2915102B1 (fr) * | 2007-04-23 | 2014-05-16 | Pf Medicament | Utilisation d'un anticorps anti-cxcr4 pour le traitement du cancer |
| CA2696460A1 (en) | 2007-08-13 | 2009-02-19 | Pfizer Inc. | Combination motif immune stimulatory oligonucleotides with improved activity |
-
2011
- 2011-03-11 CN CN201180013504.1A patent/CN102884194B/zh active Active
- 2011-03-11 US US13/577,990 patent/US9296803B2/en active Active
- 2011-03-11 EP EP11754148.2A patent/EP2545174B1/en active Active
- 2011-03-11 JP JP2012557275A patent/JP5975886B2/ja active Active
- 2011-03-11 WO PCT/US2011/028070 patent/WO2011112915A2/en not_active Ceased
Also Published As
| Publication number | Publication date |
|---|---|
| US20130011436A1 (en) | 2013-01-10 |
| WO2011112915A2 (en) | 2011-09-15 |
| EP2545174A2 (en) | 2013-01-16 |
| JP2013529178A (ja) | 2013-07-18 |
| EP2545174A4 (en) | 2013-08-21 |
| CN102884194B (zh) | 2015-07-22 |
| WO2011112915A3 (en) | 2012-01-19 |
| HK1178935A1 (zh) | 2013-09-19 |
| EP2545174B1 (en) | 2015-11-11 |
| US9296803B2 (en) | 2016-03-29 |
| CN102884194A (zh) | 2013-01-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5975886B2 (ja) | Fc融合タンパク質を含む、免疫応答を増強するための方法および組成物 | |
| JP2021531013A (ja) | Il2アゴニスト | |
| CA2821080C (en) | Universal anti-tag chimeric antigen receptor-expressing t cells and methods of treating cancer | |
| US7563445B2 (en) | CD40 binding molecules and CTL peptides for treating tumors | |
| US8968720B2 (en) | Heat shock protein GP96 vaccination and methods of using same | |
| CN118546959A (zh) | Cd19定向性嵌合抗原受体及其在免疫疗法中的用途 | |
| Rashidijahanabad et al. | Recent advances in tumor associated carbohydrate antigen based chimeric antigen receptor T cells and bispecific antibodies for anti-cancer immunotherapy | |
| WO2010030002A1 (ja) | 外来性gitrリガンド発現細胞 | |
| KR20040098048A (ko) | 4-1bb 및/또는 cd40의 생체내 격발을 통한 항종양ctl 면역성 유발 | |
| US10934331B2 (en) | Methods for enhancing immune responsiveness in an individual toward a target cancer cell population comprising apoptotic cells | |
| CN104284680A (zh) | 使用对受体的亲和力增加的突变型light分子用于癌症治疗的方法和组合物 | |
| JP2011513399A (ja) | 同種癌細胞による免疫療法 | |
| Sin et al. | Combined stimulation of TLR9 and 4.1 BB augments Trp2 peptide vaccine-mediated melanoma rejection by increasing Ag-specific CTL activity and infiltration into tumor sites | |
| JP7271037B2 (ja) | 免疫シナプスを安定化させるキメラ抗原受容体(car)t細胞 | |
| Kaufman et al. | Strategies for cancer therapy using carcinoembryonic antigen vaccines | |
| Huang et al. | Improved immunogenicity of a self tumor antigen by covalent linkage to CD40 ligand | |
| JP2022539543A (ja) | Il2アゴニスト | |
| KR20220145337A (ko) | 질병 치료용 다중특이적 항체 | |
| CN116472060A (zh) | 抗体疗法 | |
| Lou et al. | Tumour immunity and T cell memory are induced by low dose inoculation with a non-replicating adenovirus encoding TAP1 | |
| Chung et al. | Generation of tumor-specific cytotoxic T lymphocyte and prolongation of the survival of tumor-bearing mice using interleukin-18-secreting fibroblasts loaded with an epitope peptide | |
| HK1178935B (en) | Methods and compositions containing fc fusion proteins for enhancing immune responses | |
| WO2021118997A1 (en) | Bcg car constructs and methods of their manufacture and use | |
| WO2012018260A1 (en) | Epidermal growth factor receptor targeted immune therapy | |
| CD | OX40 Costimulation Synergizes with |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20140311 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20140311 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20150311 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20150609 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20150615 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20151216 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20160208 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20160629 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20160719 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 5975886 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |