JP6096884B2 - 完全生物体マラリアワクチンのためのプラットフォームとしての齧歯類プラスモジウム寄生虫 - Google Patents
完全生物体マラリアワクチンのためのプラットフォームとしての齧歯類プラスモジウム寄生虫 Download PDFInfo
- Publication number
- JP6096884B2 JP6096884B2 JP2015506341A JP2015506341A JP6096884B2 JP 6096884 B2 JP6096884 B2 JP 6096884B2 JP 2015506341 A JP2015506341 A JP 2015506341A JP 2015506341 A JP2015506341 A JP 2015506341A JP 6096884 B2 JP6096884 B2 JP 6096884B2
- Authority
- JP
- Japan
- Prior art keywords
- plasmodium
- human
- rodent
- parasites
- infected
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images




Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/002—Protozoa antigens
- A61K39/015—Hemosporidia antigens, e.g. Plasmodium antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P33/00—Antiparasitic agents
- A61P33/02—Antiprotozoals, e.g. for leishmaniasis, trichomoniasis, toxoplasmosis
- A61P33/06—Antimalarials
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/52—Bacterial cells; Fungal cells; Protozoal cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/52—Bacterial cells; Fungal cells; Protozoal cells
- A61K2039/523—Bacterial cells; Fungal cells; Protozoal cells expressing foreign proteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/58—Medicinal preparations containing antigens or antibodies raising an immune response against a target which is not the antigen used for immunisation
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Landscapes
- Health & Medical Sciences (AREA)
- Tropical Medicine & Parasitology (AREA)
- Life Sciences & Earth Sciences (AREA)
- Veterinary Medicine (AREA)
- Chemical & Material Sciences (AREA)
- Public Health (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Microbiology (AREA)
- Immunology (AREA)
- Mycology (AREA)
- Epidemiology (AREA)
- Organic Chemistry (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Description
本発明は、一次免疫用ベクターとしての齧歯類寄生虫の使用に基づく、マラリアに対する前赤血球期完全生物体ワクチンの開発のための代替的戦略を提案する。齧歯類プラスモジウム生物体は、齧歯類寄生虫の相同分子により生成される異種間防御能力を利用することにより、ヒト感染性プラスモジウム属種への感染に対して防御することが可能な免疫応答をヒト宿主において誘導するために使用され得る。さらに、これらの齧歯類プラスモジウム生物体の免疫能力は、選択された免疫原性抗原を遺伝子組換えにより導入することにより高められ得る。この戦略において、齧歯類プラスモジウム生物体は、進化的に保存された分子により提供されるそれらの自然の異種間防御能に加えて、ヒトプラスモジウム寄生虫に対する特異的防御免疫応答を誘発するために、ヒト感染性プラスモジウム属種の抗原を有し得る。
以下の図は、明細書を説明するための好ましい実施形態を提供するものであり、発明の範囲を限定するものと見なされるべきではない。
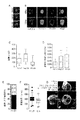
我々は、並行して、齧歯類プラスモジウム生物体である野生型P.ベルゲイによる、1つのマウス肝癌細胞系及び2つのヒト肝癌細胞系(Hepa1−6、HepG2及びHuh7)並びに1つのヒト不死化肝細胞系(HC04)のin vitro感染と、マウス一次肝細胞及びヒト一次肝細胞/線維芽細胞共培養物のex vivo感染とをモニタリングした。定量的リアルタイムPCR(qRT−PCR)及び蛍光顕微鏡検査による感染評価は、P.ベルゲイが、調査した全てのin vitro系において同様の程度まで通過(traverse)、侵入及び成長すること、並びにP.ベルゲイが、ex vivoで齧歯類及びヒト一次肝細胞内に侵入し成長することができることを示した(図1A、B、C、D)。
安全であるために、齧歯類プラスモジウム生物体ベースのマラリアワクチンは、免疫用寄生虫がヒトにおいて疾患を引き起こすことが確実にできないようにしなければならない。これは、齧歯類プラスモジウムメロゾイトが、ヒト赤血球(RBC)内に効果的に侵入し増殖することができないことを必要とする。これを確かめるために、我々は、P.ファルシパルム感染に対する薬物有効性を評価するためのモデルとして開発された特定割合のヒト赤血球が移植された血液ヒト化マウス(BHマウス)[12]の使用に基づく戦略を使用した。この系は、感染細胞と非感染細胞とを、及びヒト赤血球と齧歯類赤血球とをそれぞれ見分けるための、核SYTO−16染料及びマウス特異的TER−119染料の使用と合わされ、それにより、フローサイトメトリーによってどちらの種類の細胞の感染もモニタリングし得る(図2A1、2A2、2A3)。
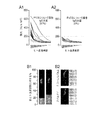
我々は、カメルーン及びタンザニアからのアフリカ人マラリア感染個体からの血清試料を、Pベルゲイ及びP.ファルシパルムCSタンパク質に対する抗体の存在について、並びにこれらの種の両方からのspzを認識するそれらの能力について評価した。我々の結果は、これらの試料はいずれもP.ベルゲイCSに対する抗体を含有していなかった(図3A1、3A2)が、それらのうちの71%が、P.ファルシパルム及びP.ベルゲイspzの両方を認識することができた(図3B1、3B2)ことを示した。これらのデータは、マラリアに対する自然獲得免疫が、CSタンパク質以外のspz上の保存されたヒトプラスモジウム寄生虫及び齧歯類プラスモジウム寄生虫エピトープに対する抗体応答を含むことを示す。全体的に見て、これらの結果は、ワクチン接種プラットフォームとしての齧歯類プラスモジウム寄生虫の使用が、現在のところ知られていない保存された抗原に対する免疫応答を惹起する可能性を有することを明らかに示す。
我々が提案するワクチン接種方法のさらなる利点は、我々は、極めて効果的な特異的免疫応答を誘発するであろうヒトプラスモジウム寄生虫の抗原の遺伝子組換えによる導入により、齧歯類プラスモジウム寄生虫によって提供される固有の異種間防御を向上させ得るという考えに依拠するものである。
放射線弱毒化スポロゾイト(RAS)及び遺伝子的に弱毒化されたスポロゾイト(GAS)により提供されるものなどの完全生物体アプローチは、そのような弱毒化スポロゾイトワクチンの製造、製剤化及び送達に関する多大な技術的課題にもかかわらず、サブユニットベースの前赤血球期ワクチン接種戦略に代わる非常に魅力的な代替法と見られている。しかしながら、これらのアプローチの両方が、それらが最も命に関わるヒトマラリア寄生虫であるP.ファルシパルムの弱毒化に基づくものであるという事実から生じる不可避の安全性の懸念を提起する。
Claims (5)
- 生きた齧歯類プラスモジウム生物体を含む、ヒトマラリアを予防するためのワクチンであって、
前記齧歯類プラスモジウム生物体が、ヒトプラスモジウム寄生虫由来のスポロゾイト周囲(CS)タンパク質を発現するように遺伝子組換えされている、ワクチン。 - 前記齧歯類プラスモジウム生物体が、自然宿主が齧歯類動物である、P.ベルゲイ(P. berghei)、P.ヨエリ(P. yoelii)、P.ビンケイ(P. vinckei)及びP.シャバウディ(P. chabaudi)を含む原生動物プラスモジウム属のいずれかのメンバーである、請求項1に記載のワクチン。
- 前記ヒトマラリアが、原生動物プラスモジウム属のP.ファルシパルム(P. falciparum)、P.ビバックス(P. vivax)、P.マラリアエ(P.malariae)、P.オヴァレ(P. ovale)及びP.ノウレシ(P. knowlesi)を含むヒトプラスモジウム寄生虫のうちのいずれかによって引き起こされる、請求項1または2に記載のワクチン。
- ヒトプラスモジウム寄生虫の前赤血球期感染を阻害するための、生きた齧歯類プラスモジウム生物体を含む、医薬組成物であって、前記齧歯類プラスモジウム生物体が、ヒトプラスモジウム寄生虫由来のスポロゾイト周囲(CS)タンパク質を発現するように遺伝子組換えされている、医薬組成物。
- 薬学的に許容可能なキャリアを含む、請求項4に記載の医薬組成物。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| PT10626212 | 2012-04-17 | ||
| PT106262 | 2012-04-17 | ||
| PCT/IB2013/053050 WO2013156949A1 (en) | 2012-04-17 | 2013-04-17 | Rodent plasmodium parasites as platforms for a whole-organism malaria vaccine |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2015514745A JP2015514745A (ja) | 2015-05-21 |
| JP6096884B2 true JP6096884B2 (ja) | 2017-03-15 |
Family
ID=48539330
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2015506341A Expired - Fee Related JP6096884B2 (ja) | 2012-04-17 | 2013-04-17 | 完全生物体マラリアワクチンのためのプラットフォームとしての齧歯類プラスモジウム寄生虫 |
Country Status (10)
| Country | Link |
|---|---|
| US (1) | US9655957B2 (ja) |
| EP (1) | EP2838554B1 (ja) |
| JP (1) | JP6096884B2 (ja) |
| CN (1) | CN104302313B (ja) |
| AU (1) | AU2013250814B2 (ja) |
| BR (1) | BR112014025823A8 (ja) |
| IN (1) | IN2014MN02275A (ja) |
| PT (1) | PT2838554T (ja) |
| WO (1) | WO2013156949A1 (ja) |
| ZA (1) | ZA201408440B (ja) |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN104771752B (zh) * | 2015-04-21 | 2018-01-05 | 中国人民解放军第三军医大学 | 疟疾红内期灭活全虫疫苗的新用途 |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7718165B2 (en) * | 2003-12-19 | 2010-05-18 | Seattle Biomedical Research Institute | Live genetically attenuated malaria vaccine |
| US7722860B2 (en) * | 2004-12-03 | 2010-05-25 | Seattle Biomedical Research Institute | Live genetically engineered protozoan vaccine |
| FR2978048B1 (fr) * | 2011-07-20 | 2014-10-10 | Univ Paris Curie | Composition vaccinale contre le paludisme |
-
2013
- 2013-04-17 BR BR112014025823A patent/BR112014025823A8/pt not_active IP Right Cessation
- 2013-04-17 WO PCT/IB2013/053050 patent/WO2013156949A1/en not_active Ceased
- 2013-04-17 IN IN2275MUN2014 patent/IN2014MN02275A/en unknown
- 2013-04-17 EP EP13726295.2A patent/EP2838554B1/en not_active Not-in-force
- 2013-04-17 CN CN201380020546.7A patent/CN104302313B/zh not_active Expired - Fee Related
- 2013-04-17 PT PT13726295T patent/PT2838554T/pt unknown
- 2013-04-17 US US14/394,964 patent/US9655957B2/en active Active
- 2013-04-17 JP JP2015506341A patent/JP6096884B2/ja not_active Expired - Fee Related
- 2013-04-17 AU AU2013250814A patent/AU2013250814B2/en not_active Expired - Fee Related
-
2014
- 2014-11-17 ZA ZA2014/08440A patent/ZA201408440B/en unknown
Also Published As
| Publication number | Publication date |
|---|---|
| WO2013156949A1 (en) | 2013-10-24 |
| US20150071966A1 (en) | 2015-03-12 |
| CN104302313A (zh) | 2015-01-21 |
| BR112014025823A2 (ja) | 2017-06-20 |
| ZA201408440B (en) | 2015-12-23 |
| BR112014025823A8 (pt) | 2017-07-25 |
| EP2838554B1 (en) | 2018-08-22 |
| IN2014MN02275A (ja) | 2015-08-07 |
| HK1206243A1 (en) | 2016-01-08 |
| EP2838554A1 (en) | 2015-02-25 |
| JP2015514745A (ja) | 2015-05-21 |
| AU2013250814B2 (en) | 2017-03-02 |
| PT2838554T (pt) | 2018-11-23 |
| US9655957B2 (en) | 2017-05-23 |
| AU2013250814A1 (en) | 2014-12-04 |
| CN104302313B (zh) | 2018-01-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Goh et al. | Vaccination with sporozoites: models and correlates of protection | |
| Vaughan et al. | Genetically engineered, attenuated whole-cell vaccine approaches for malaria | |
| Zheng et al. | Prospects for malaria vaccines: pre‐erythrocytic stages, blood stages, and transmission‐blocking stages | |
| Matuschewski et al. | Vaccines against malaria–an update | |
| Hassert et al. | Memory CD8+ T cell‐mediated protection against liver‐stage malaria | |
| EP2956166B1 (en) | Genetic attenuation of plasmodium by b9 gene disruption | |
| Mikolajczak et al. | Preerythrocytic malaria vaccine development | |
| White et al. | Potential role of vaccines in elimination of Plasmodium vivax | |
| US11529404B2 (en) | Doubly attenuated late liver stage malaria parasites and related compositions and methods | |
| Richie | High road, low road? Choices and challenges on the pathway to a malaria vaccine | |
| JP6096884B2 (ja) | 完全生物体マラリアワクチンのためのプラットフォームとしての齧歯類プラスモジウム寄生虫 | |
| Sun et al. | Vesicular stomatitis virus-based vaccine targeting plasmodium blood-stage antigens elicits immune response and protects against malaria with protein booster strategy | |
| Serrano‐Coll et al. | Plasmodium and Host Immunity: Evasion Strategies and Advances in Malaria Vaccination | |
| Azab | What do we know about the malaria vaccines? | |
| HK1206243B (en) | Rodent plasmodium parasites as platforms for a whole-organism malaria vaccine | |
| Rôla | Comparative efficacy analysis of whole-sporozoite vaccines against Malaria | |
| Manuel | Rodent Plasmodium parasites as platforms for a whole-organism malaria vaccine | |
| Reeder | Vaccine Mediated Immunity To Malaria | |
| Mo et al. | NIAID meeting report: improving malaria vaccine strategies through the application of immunological principles | |
| Nicholas III | Preliminary Characterization and Functional Validation of Pre-erythrocytic Vaccine Candidates for the Malaria Parasite Plasmodium vivax | |
| Douradinha | Immunization with mutant Plasmodium parasites: How is that achieved? | |
| MUHAMMMAD et al. | POTENTIAL ANTIGENTS FOR MALARIA VACCINE: A NEW HOPE FOR MALARIA ELIMINATION | |
| McCaffery | Development of Novel Immunogens for Malaria Vaccines and Serosurveillance | |
| WO2024173724A2 (en) | P36 and p52 as protective malaria vaccine antigens | |
| Otarigho | Malaria vaccine: hope in the future |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20151020 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20160725 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20161025 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20161219 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20170125 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20170203 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20170216 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6096884 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| LAPS | Cancellation because of no payment of annual fees |