JP6432023B2 - 脊髄小脳失調症31型(sca31)治療剤 - Google Patents
脊髄小脳失調症31型(sca31)治療剤 Download PDFInfo
- Publication number
- JP6432023B2 JP6432023B2 JP2014244350A JP2014244350A JP6432023B2 JP 6432023 B2 JP6432023 B2 JP 6432023B2 JP 2014244350 A JP2014244350 A JP 2014244350A JP 2014244350 A JP2014244350 A JP 2014244350A JP 6432023 B2 JP6432023 B2 JP 6432023B2
- Authority
- JP
- Japan
- Prior art keywords
- uggaa
- exp
- tdp
- rna
- drosophila
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images


























Landscapes
- Peptides Or Proteins (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Description
[1]TDP-43(transactivation responsive region DNA-binding protein 43)もしくはFUS/TLS(fused in sarcoma / translated in liposarcoma)から選択されるRNA結合タンパク質、またはこれらのRNA結合領域を含む断片を含む、脊髄小脳失調症31型(SCA31)治療剤。
[2]TDP-43が
(a)配列番号1で示されるアミノ酸配列からなるか、または、
(b)配列番号1で示されるアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなり、かつ(UGGAA)n(n=80-100)に対する結合活性を有するものである、上記[1]記載の治療剤。
[3]FUS/TLSが
(a)配列番号3で示されるアミノ酸配列からなるか、または、
(b)配列番号3で示されるアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなり、かつ(UGGAA)n(n=80-100)に対する結合活性を有するものである、上記[1]記載の治療剤。
[4]TDP-43のRNA結合領域を含む断片が、配列番号1の105〜169位のアミノ酸に相当するRRM1領域、および/または193〜257位のアミノ酸に相当するRRM2領域を必須に含む、上記[1]記載の治療剤。
[5]FUS/TLSのRNA結合領域を含む断片が、配列番号3の285〜371位のアミノ酸を必須に含む、上記[1]記載の治療剤。
(a)配列番号1で示されるアミノ酸配列からなるか、または、
(b)配列番号1で示されるアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなり、かつ(UGGAA)n(n=80-100)に対する結合活性を有するものである、脊髄小脳失調症31型(SCA31)治療剤である。
(a)配列番号3で示されるアミノ酸配列からなるか、または、
(b)配列番号3で示されるアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなり、かつ(UGGAA)n(n=80-100)に対する結合活性を有するものである、脊髄小脳失調症31型(SCA31)治療剤である。
ショウジョウバエにおける(UGGAA) exp の発現によるRNA凝集および毒性誘導の実証
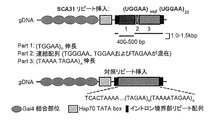
SCA31におけるプルキンエ細胞変性にin vivoで関与し得る(UGGAA)expの毒性の特徴を明らかにするために、本発明者等は、UAS/GAL4システムを用いて、疾患に特異的な(UGGAA)expリピート、または正常集団に見られるUAGAAおよびUAAAAUAGAA(配列番号5)複合体からなる対照リピートを発現するトランスジェニックショウジョウバエの系を作製するための発現構築物を作製した(図1−1)。
ショウジョウバエ成虫での(UGGAA) exp の発現による毒性誘導の実証
次に、本発明者等は、ショウジョウバエの中枢神経系(CNS)での(UGGAA)expの発現の影響を調べるためにelav-GAL4システムを用いた。elav-GAL4を用いたCNSでの(UGGAA)expの持続的発現は蛹を致死させたため、本実施例においては誘導性Gene-Switch発現系を用い、羽化後のショウジョウバエのCNSで(UGGAA)exp発現を誘導した。
in vitroおよびin vivoにおけるTDP-43とUGGAA RNAとの相互作用
in vitro RNAプルダウンアッセイによって、潜在的(UGGAA)exp結合パートナーのスクリーニングを行った。具体的には、(UGGAA)expまたは対照リピートを含むビオチン化RNAをT7 RNAポリメラーゼ(Roche)を用いてin vitro転写によって合成し、PC12細胞またはマウス脳の核画分(NE-PER抽出試薬(Pierce)を用いて細胞を溶解させて分画した)と共にインキュベートした。次いで、ストレプトアビジンコートした磁気ビーズ(Dynabeads M-280 Streptavidin (Invitrogen))を用いてプルダウンし、捕捉されたタンパク質-(UGGAA)exp複合体をSDS-PAGEによって分離し、クマシーブルーによって染色した。目的のバンドをゲルから切り出し、LC-MS分析によってタンパク質を同定した。
TDP-43による(UGGAA) exp 誘導毒性の抑制
(UGGAA)exp誘導毒性におけるTDP-43の役割を調べるために、本発明者等は(UGGAA)exp発現ショウジョウバエをヒト野生型TDP-43発現ショウジョウバエと交配した。TDP-43の発現のために使用した構築物の模式図を図4Aに示す。
(UGGAA) exp RNAのフォールディングのATP非依存型RNAシャペロンとしてのTDP-43の機能
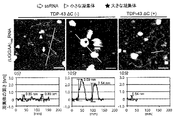
TDP-43がそのRNA結合能を介して(UGGAA)exp誘導毒性を抑制するメカニズムを解析するために、本発明者等は(UGGAA)expとTDP-43とを共発現するショウジョウバエにおける(UGGAA)exp転写産物の発現レベルおよびRNAの凝集を検討した。
in vitroにおけるTDP-43と(UGGAA) exp との相互作用
実施例5のデータから、TDP-43が(UGGAA)expと直接相互作用して異常なRNAのin vivoにおける非機能的コンフォメーションの形成を抑制する可能性が示唆された。本発明者等は次いで、TDP-43がin vitroにおける(UGGAA)exp凝集を直接抑制するか否かを原子間力顕微鏡(High-speed AFM, NanoExplorer, RIBM)を用いて試験した。
リピートがコードするペンタペプチドリピートタンパク質産生のTDP-43による抑制
イントロンTGGAAリピートがRAN翻訳メカニズムによってペンタペプチドリピート(PPR)タンパク質に異常に翻訳され得るとの仮説に基づけば、図7Aに示すように、全てのフレームで、TGGAAリピートの翻訳は1種のPPRタンパク質、ポリ-(Trp-Asn-Gly-Met-Glu)(配列番号12)を生じる。本発明者等は、[H]CMEWNGMEWMGMEWNG[OH](配列番号13)ペプチドに対するポリクローナル抗体(抗-WNGME SGJ1705およびSGJ1706)を常法により作製した。アフィニティー精製したPPR抗体の特異性を、組換えペンタペプチドを含む小脳組織切片の処理によって確認したところ、PPRタンパク質の封入量が減少し、アフィニティー精製PPR抗体はGST融合物として細菌中に生成した組換えポリ-(WNGME)ペプチドを検出した。
FUS/TLSによる(UGGAA) exp 誘導毒性の抑制効果
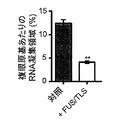
ショウジョウバエにおける(UGGAA)exp毒性に対するFUS/TLSの効果を評価した。ヒトFUS/TLSをアップレギュレーションすると、TDP-43と同様に(UGGAA)exp(S)系での複眼変性を劇的に軽減した(図8A)。また、FUS/TLSは、TDP-43と同様に、(UGGAA)expを発現する三齢幼虫におけるRNAの凝集およびリピートRNAをコードするPPRタンパク質の生成を抑制した。FUS/TLSを(UGGAA)expと共発現させても(UGGAA)exp転写産物の発現レベルには影響しなかった(図8B)が、それらの発現はRNA凝集(**p < 0.01, 複眼原基あたり12.4±0.76 から4.1±0.26%に減少、約67%の凝集阻害活性 )およびPPRタンパク質(**p < 0.001, 複眼原基あたり4.1±0.48から1.3±0.16 %に減少)の統計的に有意な減少をもたらした(図8Cおよび8D)。また、免疫ブロット解析から、FUS/TLSがPPRタンパク質の合成を阻害していることが明らかとなった(***p < 0.001, 相対PPRタンパク質レベルは複眼原基あたり1.0から0.5±0.05 %に減少)(図8E)。
Claims (5)
- TDP-43(transactivation responsive region DNA-binding protein 43)もしくはFUS/TLS(fused in sarcoma / translated in liposarcoma)から選択されるRNA結合タンパク質、またはこれらのRNA結合領域を含む断片を含む、脊髄小脳失調症31型(SCA31)治療剤。
- TDP-43が
(a)配列番号1で示されるアミノ酸配列からなるか、または、
(b)配列番号1で示されるアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなり、かつ(UGGAA)n(n=80-100)に対する結合活性を有するものである、請求項1記載の治療剤。 - FUS/TLSが
(a)配列番号3で示されるアミノ酸配列からなるか、または、
(b)配列番号3で示されるアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換または付加されたアミノ酸配列からなり、かつ(UGGAA)n(n=80-100)に対する結合活性を有するものである、請求項1記載の治療剤。 - TDP-43のRNA結合領域を含む断片が、配列番号1の105〜169位のアミノ酸に相当するRRM1領域、および/または193〜257位のアミノ酸に相当するRRM2領域を必須に含む、請求項1記載の治療剤。
- FUS/TLSのRNA結合領域を含む断片が、配列番号3の285〜371位のアミノ酸を必須に含む、請求項1記載の治療剤。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2014244350A JP6432023B2 (ja) | 2014-12-02 | 2014-12-02 | 脊髄小脳失調症31型(sca31)治療剤 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2014244350A JP6432023B2 (ja) | 2014-12-02 | 2014-12-02 | 脊髄小脳失調症31型(sca31)治療剤 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2016108249A JP2016108249A (ja) | 2016-06-20 |
| JP6432023B2 true JP6432023B2 (ja) | 2018-12-05 |
Family
ID=56123154
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2014244350A Active JP6432023B2 (ja) | 2014-12-02 | 2014-12-02 | 脊髄小脳失調症31型(sca31)治療剤 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP6432023B2 (ja) |
Families Citing this family (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2018017144A1 (en) | 2016-07-19 | 2018-01-25 | Brandeis University | Compositions and methods for identifying rna binding polypeptide targets |
| WO2019204781A1 (en) * | 2018-04-20 | 2019-10-24 | Design Therapeutics Inc. | Methods and compounds for the treatment of genetic disease |
| JP7295584B2 (ja) * | 2018-11-09 | 2023-06-21 | スンチョニャン ユニヴァーシティ インダストリー アカデミー コーペレーション ファウンデーション | 神経系退行性疾患診断用マーカーおよび治療用組成物 |
| EP3994159A4 (en) * | 2019-07-05 | 2023-08-09 | University of Florida Research Foundation, Incorporated | METHODS OF TREATING NEUROLOGICAL DISEASES ASSOCIATED WITH THE RAN PROTEIN |
| WO2023204313A1 (ja) * | 2022-04-22 | 2023-10-26 | 国立大学法人新潟大学 | Tdp-43の凝集抑制剤及び医薬組成物 |
| CN115198007A (zh) * | 2022-08-23 | 2022-10-18 | 迈基诺(重庆)基因科技有限责任公司 | 检测bean1和tk2基因tggaa重复的引物组、试剂盒及系统 |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2016088797A1 (ja) * | 2014-12-02 | 2016-06-09 | 国立大学法人東京医科歯科大学 | Alsの原因タンパク毒性を軽減する核酸 |
-
2014
- 2014-12-02 JP JP2014244350A patent/JP6432023B2/ja active Active
Also Published As
| Publication number | Publication date |
|---|---|
| JP2016108249A (ja) | 2016-06-20 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6432023B2 (ja) | 脊髄小脳失調症31型(sca31)治療剤 | |
| Gleixner et al. | NUP62 localizes to ALS/FTLD pathological assemblies and contributes to TDP-43 insolubility | |
| Saito et al. | Potent amyloidogenicity and pathogenicity of Aβ43 | |
| Kuwahara et al. | LRRK2 and RAB7L1 coordinately regulate axonal morphology and lysosome integrity in diverse cellular contexts | |
| Finley et al. | Blue cheese mutations define a novel, conserved gene involved in progressive neural degeneration | |
| Berdowski et al. | Dominant-acting CSF1R variants cause microglial depletion and altered astrocytic phenotype in zebrafish and adult-onset leukodystrophy | |
| EP2039367B1 (en) | Prophylactic/therapeutic agent for neurodegenerative disease | |
| Qiu et al. | Widespread aggregation of mutant VAPB associated with ALS does not cause motor neuron degeneration or modulate mutant SOD1 aggregation and toxicity in mice | |
| US10801027B2 (en) | Inhibitors of SRSF1 to treat neurodegenerative disorders | |
| KR101762134B1 (ko) | 이분자 형광 상보 시스템을 이용한 알파-시뉴클린 응집체의 세포간 전이 측정방법 및 이를 이용한 퇴행성 신경질환의 예방 또는 치료용 물질의 스크리닝 방법 | |
| IL194274A (en) | Mecp2 polypeptides and use thereof in manufacture of pharmaceutical compositions for neurological disease | |
| ITMI20071975A1 (it) | Prodotti e loro uso per la diagnosi prevenzione e-o cura di patologie umane e-o animali caraterizzate dalla anomala deposizione di sostanza b-amiloide e-o similamiloide in organi e tesstui umani e-o animali e metodo di screening per la determinazione | |
| He et al. | The carboxyl termini of RAN translated GGGGCC nucleotide repeat expansions modulate toxicity in models of ALS/FTD | |
| Letelier et al. | Mutation of vsx genes in zebrafish highlights the robustness of the retinal specification network | |
| KR20230159847A (ko) | 염증성 또는 활성화된 세포를 표적으로 하고 염증성 상태 및 통증을 치료 또는 개선하기 위한 조성물 및 방법 | |
| JP6879738B2 (ja) | snoRNA、組成物、及び使用 | |
| WO2016088797A1 (ja) | Alsの原因タンパク毒性を軽減する核酸 | |
| JP4613824B2 (ja) | トランスジェニック非ヒト哺乳動物 | |
| JPWO2020027311A1 (ja) | Tdp−43凝集阻害剤 | |
| JP2025148560A (ja) | 進行性核上性麻痺の治療 | |
| EP3747468A1 (en) | Therapeutic agent for frontotemporal lobar degeneration, method for screening therapeutic agent for frontotemporal lobar degeneration and method for treating frontotemporal lobar degeneration | |
| US20230365626A1 (en) | Alloferon Peptide and Method Using the Same | |
| CN118829441A (zh) | 用于治疗polyq疾病的治疗因子 | |
| Allan et al. | A commonly used Drosophila model of Alzheimer’s disease generates an aberrant species of amyloid-β with an additional N-terminal glutamine residue | |
| JP6959632B2 (ja) | 筋萎縮性側索硬化症又は前頭側頭型認知症の予防又は治療剤 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20171128 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20180523 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20180523 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20180828 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20180927 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6432023 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| S111 | Request for change of ownership or part of ownership |
Free format text: JAPANESE INTERMEDIATE CODE: R313117 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| S111 | Request for change of ownership or part of ownership |
Free format text: JAPANESE INTERMEDIATE CODE: R313115 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |