JP6525963B2 - 血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 - Google Patents
血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 Download PDFInfo
- Publication number
- JP6525963B2 JP6525963B2 JP2016514978A JP2016514978A JP6525963B2 JP 6525963 B2 JP6525963 B2 JP 6525963B2 JP 2016514978 A JP2016514978 A JP 2016514978A JP 2016514978 A JP2016514978 A JP 2016514978A JP 6525963 B2 JP6525963 B2 JP 6525963B2
- Authority
- JP
- Japan
- Prior art keywords
- cells
- stem cells
- hematopoietic stem
- vpa
- cell
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images




































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0652—Cells of skeletal and connective tissues; Mesenchyme
- C12N5/0662—Stem cells
- C12N5/0665—Blood-borne mesenchymal stem cells, e.g. from umbilical cord blood
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0647—Haematopoietic stem cells; Uncommitted or multipotent progenitors
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
- A61P35/02—Antineoplastic agents specific for leukemia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5044—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics involving specific cell types
- G01N33/5073—Stem cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/90—Serum-free medium, which may still contain naturally-sourced components
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/065—Modulators of histone acetylation
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/999—Small molecules not provided for elsewhere
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Chemical & Material Sciences (AREA)
- Hematology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- Organic Chemistry (AREA)
- Immunology (AREA)
- Cell Biology (AREA)
- Developmental Biology & Embryology (AREA)
- General Health & Medical Sciences (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Genetics & Genomics (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Molecular Biology (AREA)
- Urology & Nephrology (AREA)
- Medicinal Chemistry (AREA)
- General Engineering & Computer Science (AREA)
- Toxicology (AREA)
- Physics & Mathematics (AREA)
- General Physics & Mathematics (AREA)
- Pathology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Analytical Chemistry (AREA)
- Food Science & Technology (AREA)
- Rheumatology (AREA)
- Animal Behavior & Ethology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Public Health (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Veterinary Medicine (AREA)
- Pharmacology & Pharmacy (AREA)
- General Chemical & Material Sciences (AREA)
- Oncology (AREA)
- Diabetes (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Description
本発明は、濃縮されかつ増殖したヒト臍帯血幹細胞、その作製方法、及び治療方法に関する。
臍帯血(「CB」)造血幹細胞(「HSC」)は、幹細胞とその成熟細胞とを区別する多くの表現型及び機能的特性を有する(Cairo et al., "Placental and/or Umbilical Cord Blood: An Alternative Source of Hematopoietic Stem Cells for Transplantation," Blood 90:4665-4678 (1997)(非特許文献1);Dahlberg et al., "Ex vivo Expansion of Human Hematopoietic Stem and Progenitor Cells," Blood 117:6083-6090 (2011)(非特許文献2);Delaney et al., "Cord Blood Graft Engineering," Biol. Blood Marrow Transplant 19(1): S74-S78 (2013)(非特許文献3);Navarrete et al., "Cord Blood Banking: A Historical Perspective," Br. J. Haematol. 147:236-245 (2009)(非特許文献4);Stanevsky et al., "Umbilical Cord Blood Transplantation: Pros, Cons and Beyond," Blood Rev. 23:199-204 (2009)(非特許文献5))。CB CD34+細胞が、より未分化であると考えられている理由は、その広範囲な増殖能、インビトロで造血コロニーを生成する高い能力、胎児ヘモグロビンを含有する赤血球細胞を形成する能力、及びこのような細胞の若干が持つ骨髄破壊的同種レシピエントを再構築する能力にある(Cairo et al., "Placental and/or Umbilical Cord Blood: An Alternative Source of Hematopoietic Stem Cells for Transplantation," Blood 90:4665-4678 (1997)(非特許文献1))。CB細胞を、血液悪性腫瘍及び遺伝性疾患に罹患した同種幹細胞レシピエントへのHSC移植片として使用することは、単一のCB収集物中に存在する幹細胞数に限りがあるため、小児または若年成人レシピエントに限定されてきた(Cairo et al., "Placental and/or Umbilical Cord Blood: An Alternative Source of Hematopoietic Stem Cells for Transplantation," Blood 90:4665-4678 (1997)(非特許文献1);Navarrete et al., "Cord Blood Banking: A Historical Perspective," Br. J. Haematol. 147:236-245 (2009)(非特許文献4);Stanevsky et al., "Umbilical Cord Blood Transplantation: Pros, Cons and Beyond," Blood Rev. 23:199-204 (2009)(非特許文献5))。これらの制約から、成人レシピエントにおける移植失敗及び生着速度(kinetics)遅延の割合が容認できないほど高い結果となった(Cairo et al., "Placental and/or Umbilical Cord Blood: An Alternative Source of Hematopoietic Stem Cells for Transplantation," Blood 90:4665-4678 (1997)(非特許文献1);Dahlberg et al., "Ex vivo Expansion of Human Hematopoietic Stem and Progenitor Cells," Blood 117:6083-6090 (2011)(非特許文献2);Delaney et al., "Cord Blood Graft Engineering," Biol. Blood Marrow Transplant 19(1): S74-S78 (2013)(非特許文献3);Navarrete et al., "Cord Blood Banking: A Historical Perspective," Br. J. Haematol. 147:236-245 (2009)(非特許文献4);Stanevsky et al., "Umbilical Cord Blood Transplantation: Pros, Cons and Beyond," Blood Rev. 23:199-204 (2009)(非特許文献5);Barker et al., "Combined Effect of Total Nucleated Cell Dose and HLA Match on Transplantation Outcome in 1061 Cord Blood Recipients with Hematologic Malignancies," Blood 115:1843-1849 (2010)(非特許文献6);Delaney et al., "Strategies to Enhance Umbilical Cord Blood Stem Cell Engraftment in Adult Patients," Expert Rev. Hematol. 3:273-283 (2010)(非特許文献7))。これらの障害を克服する試みとして、2つの異なるCB移植片注入、またはCB CD34+細胞のHSC周期進行及びそれに続く分裂を促進できる様々なサイトカインの組み合わせを使用するCD34+細胞のエクスビボでの増殖など、いくつかの異なる方法が挙げられてきた(2、6−9)。エクスビボでの幹細胞増殖におけるこれらの初期の試みでは、より多数の造血始原及び前駆細胞を産生したが、骨髄再構築細胞数は減少してしまった。HSCの大半は、インビボでの細胞周期が遅い静止細胞である。(Giebel et al., "Primitive Human Hematopoietic Cells Give Rise to Differentially Specified Daughter Cells Upon their Initial Cell Division," Blood 107(5):2146-2152 (2006)(非特許文献8);Ho et al., "The Beauty of Asymmetry: Asymmetric Divisions and Self-Renewal in the Haematopoietic System," Curr. Opin. Hematol. 14(4):330-336 (2007)(非特許文献9);Huang et al., "Symmetry of Initial Cell Divisions Among Primitive Hematopoietic Progenitors Is Independent of Ontogenic Age and Regulatory Molecules," Blood 94(8):2595-2604 (1999)(非特許文献10);Srour et al., "Modulation of In vitro Proliferation Kinetics and Primitive Hematopoietic Potential of Individual Human CD34+CD38-/lo Cells in G0," Blood 105(8):3109-3116 (2005)(非特許文献11))。このようなサイトカインの組み合わせの存在下で生じるCB CD34+細胞のエクスビボ細胞周期進行及び分裂を早めることで、HSCへのコミットメントをもたらしたが、静止状態を維持したまたは細胞分裂回数の少なかったわずかな幹細胞画分に、骨髄再構築能が残されていた(10−13)。ごく最近では、移植可能なCB HSCの数を増やすことを目的として、これらのサイトカインの組み合わせに対して、間葉系細胞フィーダー層、または固定化したnotchリガンド、銅キレート剤、ヒストンデアセチラーゼ阻害剤(HDACI)、オールトランスレチノイン酸、アリールハイドロカーボン受容体のアンタゴニスト、プロスタグランジンE2(PGE2)、もしくはc−MPLアゴニストなどの多数の分子を添加してきた(Dahlberg et al., "Ex vivo Expansion of Human Hematopoietic Stem and Progenitor Cells," Blood 117:6083-6090 (2011)(非特許文献2);Delaney et al., "Strategies to Enhance Umbilical Cord Blood Stem Cell Engraftment in Adult Patients," Expert Rev. Hematol. 3:273-283 (2010)(非特許文献7);Boitano et al., "Aryl Hydrocarbon Receptor Antagonists Promote the Expansion of Human Hematopoietic Stem Cells," Science 329:1345-1348 (2010)(非特許文献12);De Felice et al., "Histone Deacetylase Inhibitor Valproic Acid Enhances the Cytokine-Induced Expansion of Human Hematopoietic Stem Cells," Cancer Res. 65:1505-1513 (2005)(非特許文献13);Himburg et al., "Pleiotrophin Regulates the Expansion and Regeneration of Hematopoietic Stem Cells," Nat. Med. 16:475-482 (2010)(非特許文献14);Milhem et al., "Modification of Hematopoietic Stem Cell Fate by 5aza 2'deoxycytidine and Trichostatin A," Blood 103:4102-4110 (2004)(非特許文献15);Nishino et al., "Ex vivo Expansion of Human Hematopoietic Stem Cells by a Small-Molecule Agonist of c-MPL," Exp. Hematol. 37:1364-1377 e1364. (2009)(非特許文献16);North et al., "Prostaglandin E2 Regulates Vertebrate Haematopoietic Stem Cell Homeostasis," Nature 447:1007-1011 (2007)(非特許文献17))。これらのアプローチのいくつかは、臨床試験で評価されたが、長期ではなく短期の骨髄再構築細胞が多数産生される結果となった(Dahlberg et al., "Ex vivo Expansion of Human Hematopoietic Stem and Progenitor Cells," Blood 117:6083-6090 (2011)(非特許文献2);de Lima et al., "Transplantation of Ex vivo Expanded Cord Blood Cells Using the Copper Chelator Tetraethylenepentamine: A Phase I/II Clinical Trial," Bone Marrow Transplant 41:771-778 (2008)(非特許文献18);de Lima et al., "Cord-Blood Engraftment with Ex Vivo Mesenchymal-Cell Coculture," N. Engl. J. Med. 367(24):2305-2315 (2012)(非特許文献19);Delaney et al., "Notch-Mediated Expansion of Human Cord Blood Progenitor Cells Capable of Rapid Myeloid Reconstitution," Nat. Med. 16(2):232-236 (2010)(非特許文献20))。あるいは、同種CB移植の有効性を向上させるために、CB CD34+細胞のホーミング及び生着効率を促進する方法も進められている(Goessling et al., "Prostaglandin E2 Enhances Human Cord Blood Stem Cell Xenotransplants and Shows Long-Term Safety in Preclinical Nonhuman Primate Transplant Models," Cell Stem Cell 8(4):445-458 (2011)(非特許文献21);Hoggatt et al., "Differential Stem and Progenitor-Cell Trafficking by Prostaglandin E2," Nature 495(7441):365-369 (2013)(非特許文献22);O'Leary et al., "The Role of Dipeptidyl Peptidase 4 in Hematopoiesis and Transplantation," Curr. Opin. Hematol. 20(4):314-319 (2013)(非特許文献23))。
[本発明1001]
単離されかつ増殖したヒト臍帯血幹細胞の濃縮された集団であって、該幹細胞が、CD34 + 、CD90 + 、CD184 + 、CD117 + 、CD49f + 、ALDH + 、かつCD45RA − でありかつ多能性遺伝子SOX2、OCT4、NANOG、及びZIC3を発現する、前記濃縮された集団。
[本発明1002]
前記幹細胞の少なくとも約60%が、アルデヒドデヒドロゲナーゼ活性に対して陽性である、本発明1001の濃縮された集団。
[本発明1003]
前記幹細胞が、胚性幹細胞の多能性遺伝子hTERTの発現レベルにおいて上方制御を示さない、本発明1001または本発明1002の濃縮された集団。
[本発明1004]
前記幹細胞の少なくとも約95%が、SOX2、OCT4、NANOG、及びZIC3に対して陽性である、前記本発明のいずれかの濃縮された集団。
[本発明1005]
前記幹細胞が、SCF、Flt3、TPO、IL3、及びこれらの組み合わせからなる群より選択されるサイトカインと接触している、前記本発明のいずれかの濃縮された集団。
[本発明1006]
前記幹細胞が、ヒストンデアセチラーゼ阻害剤と接触している、前記本発明のいずれかの濃縮された集団。
[本発明1007]
前記ヒストンデアセチラーゼ阻害剤が、VPA、SCR、LBH589、TSA、SAHA、Cay10433、及びCay10398からなる群より選択される、本発明1006の濃縮された集団。
[本発明1008]
前記幹細胞が、ヒストンデアセチラーゼ阻害剤と接触していない単離されかつ増殖したヒト臍帯血幹細胞と比較して減少した、HDAC1、HDAC3、及びHDAC5の発現を有する、本発明1006または本発明1007の濃縮された集団。
[本発明1009]
前記幹細胞の少なくとも約18.0%±1.2%が、G2/M期にある、前記本発明のいずれかの濃縮された集団。
[本発明1010]
前記幹細胞の少なくとも約23.2%±13.8%が、G0/G1期にある、前記本発明のいずれかの濃縮された集団。
[本発明1011]
以下の工程を含む、単離されかつ増殖したヒト臍帯血幹細胞の濃縮された集団の作製方法:
単離されたヒト臍帯血幹細胞の集団を提供する工程、ならびに
単離されかつ増殖したヒト臍帯血幹細胞の濃縮された集団を作製するのに有効な条件下において、無血清培養系においてヒストンデアセチラーゼ阻害剤の存在下にて、ヒト臍帯血幹細胞の単離された集団を処理する工程であって、該増殖したヒト臍帯血幹細胞が、CD34 + 、CD90 + 、CD184 + 、CD117 + 、CD49f + 、ALDH + 、かつCD45RA − でありかつ多能性遺伝子SOX2、OCT4、NANOG、及びZIC3を発現する、工程。
[本発明1012]
前記単離されたヒト臍帯血幹細胞の集団が、前記処理を行うと約20,202倍に増殖する、本発明1011の方法。
[本発明1013]
前記処理が7日間実施される、本発明1011または本発明1012の方法。
[本発明1014]
前記増殖した幹細胞の少なくとも約88.3%±5.9%が、アルデヒドデヒドロゲナーゼ活性に対して陽性である、本発明1011〜1013のいずれかの方法。
[本発明1015]
前記増殖した幹細胞が、胚性幹細胞の多能性遺伝子hTERTの発現レベルにおいて上方制御を示さない、本発明1011〜1014のいずれかの方法。
[本発明1016]
前記増殖した幹細胞の少なくとも約95%が、SOX2、OCT4、NANOG、及びZIC3に対して陽性である、本発明1011〜1015のいずれかの方法。
[本発明1017]
前記無血清培養系が、SCF、Flt3、TPO、IL3、及びこれらの組み合わせからなる群より選択されるサイトカインを更に含む、本発明1011〜1016のいずれかの方法。
[本発明1018]
前記ヒストンデアセチラーゼ阻害剤が、VPA、SCR、LBH589、TSA、SAHA、Cay10433、及びCay10398からなる群より選択される、本発明1011〜1017のいずれかの方法。
[本発明1019]
前記増殖した幹細胞が、前記処理前の前記単離された幹細胞と比較して減少した、HDAC1、HDAC3、及びHDAC5の発現を有する、本発明1011〜1018のいずれかの方法。
[本発明1020]
前記増殖した幹細胞の少なくとも約95%が、SOX2、OCT4、NANOG、及びZIC3に対して陽性である、本発明1011〜1019のいずれかの方法。
[本発明1021]
前記増殖した幹細胞の少なくとも約18.0%±1.2%が、G2/M期にある、本発明1011〜1020のいずれかの方法。
[本発明1022]
前記増殖した幹細胞の少なくとも約23.2%±13.8%が、G0/G1期にある、本発明1011〜1021のいずれかの方法。
[本発明1023]
以下の工程を含む、対象の血液疾患を治療する方法:
該対象における該血液疾患を治療するために、本発明1001〜1010のいずれかの単離されかつ増殖したヒト臍帯血幹細胞の濃縮された集団を該対象に投与する工程。
[本発明1024]
前記投与する工程が、濃縮または精製された調製物に由来する前記幹細胞を前記対象内に移植することによって実施される、本発明1023の方法。
[本発明1025]
前記増殖した幹細胞が、前記対象由来である、本発明1023または本発明1024の方法。
[本発明1026]
前記増殖した幹細胞の少なくとも約60%が、アルデヒドデヒドロゲナーゼ活性に対して陽性である、本発明1023〜1025のいずれかの方法。
[本発明1027]
前記増殖した幹細胞が、胚性幹細胞の多能性遺伝子hTERTの発現レベルにおいて全く上方制御を示さない、本発明1023〜1026のいずれかの方法。
[本発明1028]
前記増殖した幹細胞の少なくとも約95%が、SOX2、OCT4、NANOG、及びZIC3に対して陽性である、本発明1023〜1027のいずれかの方法。
[本発明1029]
前記増殖した幹細胞の少なくとも約23%が、G2/M期にある、本発明1023〜1028のいずれかの方法。
[本発明1030]
前記増殖した幹細胞の少なくとも約18%が、G0/G1期にある、本発明1023〜1029のいずれかの方法。
[本発明1031]
以下の工程を含む、造血幹細胞に対する化合物の効果を決定する方法:
本発明1001〜1010のいずれかの単離されかつ増殖したヒト臍帯血幹細胞の濃縮された集団を提供する工程;
該幹細胞を、試験される化合物に接触させる工程;ならびに
該幹細胞に対する該化合物の効果を決定するために、該接触後に該幹細胞を解析する工程。
本発明は、ヒト臍帯血由来の単離されかつ増殖した幹細胞の濃縮された集団、そのエクスビボにおける増殖、並びに単離されかつ増殖した幹細胞の濃縮された集団を対象に投与する工程を含む治療方法に関する。本発明にしたがって、造血幹細胞を無血清培地内及びヒストンデアセチラーゼ阻害剤の存在下にてエクスビボで培養することにより造血幹細胞を増殖させて、特定の表現型を発現するユニークな幹細胞集団を得ることが可能である。
方法
CB CD34+細胞の単離及びそのエクスビボ培養
CB収集物は、ニューヨーク血液センター(New York Blood Center)のPlacental Blood Programから購入した。CB−MNCは、フィコール・ハイパック(Ficoll−Hypaque)密度遠心分離法によって単離し、CD34+細胞は、先に記載した通り免疫磁性選別法により精製した(Chaurasia et al., "Chromatin-Modifying Agents Promote the Ex vivo Production of Functional Human Erythroid Progenitor Cells," Blood 117:4632-4641 (2011),その内容全体が、本明細書に参考として組み込まれる)。高純度(90%〜98%)PC(4.0〜5.0×104)を、SF Stemline II(Sigma−Aldrich社)培地または30%FBS(HyClone Laboratories社)含有IMDM(Lonza社)中で、150ng/mlのSCF、100ng/mlのfms様チロシンキナーゼ受容体3(FLT3リガンド)、100ng/mlのトロンボポエチン(TPO)、及び50ng/mlのインターロイキン3(IL−3)(R&D Systems社)を補充して培養し、37℃にて5%CO2を維持した加湿インキュベーター内でインキュベートした。16時間インキュベートした後、更に7日間、サイトカインの非存在下または持続的存在下のいずれかにて、トリコスタチンA(TSA)、スベロイルアニリドヒドロキサム酸(SAHA)、VPA(Sigma−Aldrich社)、SCR、及びCAY10433(C433、BML−210とも称される)、CAY10398(MD85としても知られる)、及びCAY10603(分子式:C22H30N4O6)(Cayman Chemicals社)を含む種々の濃度の各HDACIに、細胞を曝露した(図1A)。検討した細胞集団及びその集団を培養した各種条件は、表1に記載された特定の用語により示される。
PC、またはHDACIの存在下または非存在下にて増殖させた培養細胞は、抗ヒトCD34 mAbまたはアイソタイプ適合コントロールmAbで染色し、FACSCanto II(BD社)を使用して解析した。CD34+細胞は、更なる表現型及び機能解析のために、先に記載した通りCD34+細胞単離キットを使用して再単離された。すべてのmAbは、BD Biosciences社及びCell Signaling Technology社から購入した。コントロール培養、またはHDACIを加えたサイトカイン中で7日間エクスビボにおいて増殖させた細胞の表現型解析(CD34−APC、CD90−FITC、CD184−PE、CD117−PE、CD49f−PE、及びCD45RA−PECy7)は、先に記載した通り実施した(Chaurasia et al., "Chromatin-Modifying Agents Promote the Ex vivo Production of Functional Human Erythroid Progenitor Cells," Blood 117:4632-4641 (2011),その内容全体が、本明細書に参考として組み込まれる)。
CB CD34+細胞の遊走挙動は、直径6.5mm、5μm孔のTranswellプレート(Costar社)を使用して、先に記載した通り評価した(Shivtiel et al., "CD45 Regulates Homing and Engraftment of Immature Normal and Leukemic Human Cells in Transplanted Immunodeficient Mice," Exp. Hematol. 39(12):1161-1170 (2011),その内容全体が、本明細書に参考として組み込まれる)。Transwellの下層区画に、100ng/mlのストローマ細胞由来因子1(SDF1)(R&D Systems社)を補充したStemLine II SF培地を充填した。Transwellフィルターは、30分間37℃にてマトリゲルでコーティングした。コントロール及びVPA含有培養物に由来する再単離されたCD34+細胞(1×105)は、100μlのStemline II培地中にて、マトリゲルコーティングしたフィルター上へプレーティングした。16時間後及び48時間後、下層区画へと遊走した細胞を数えて、遊走割合(%)を以下の通り計算した:(遊走した細胞数/プレーティングした細胞の総数)×100。
コントロール条件下またはVPAでの処理後に再単離されたCD34+細胞(5×105/マウス)のホーミングを、先に記載した通り実施した(Shivtiel et al., "CD45 Regulates Homing and Engraftment of Immature Normal and Leukemic Human Cells in Transplanted Immunodeficient Mice," Exp. Hematol. 39(12):1161-1170 (2011),その内容全体が、本明細書に参考として組み込まれる)。レシピエントNSGマウスは、Jackson Laboratory社から購入し、4時間亜致死線量の放射線を照射(300cGy)をして、再単離されたCD34+細胞を尾静脈から注入した。注入後16時間及び48時間で、各レシピエントマウスの大腿骨2本及び脛骨2本からBM細胞を採取し、ヒトCD34−APC mAbを使用して、ヒトCD34+細胞の存在をフローサイトメトリーによって解析した。これらの細胞集団のホーミングは、レシピエントマウスのBM中において、得られた106イベント当たりのCD34+細胞数を定量化して測定した。4匹のマウスでは細胞が生着せず、実験サンプルからのバックグラウンドを差し引くために同様に解析した(その全体が参照として本明細書に組み込まれるColvin et al., "Allogeneic In vivo Stem Cell Homing,". J. Cell. Physiol. 211(2):386-391 (2007))。コントロール条件下で培養したCD34+細胞が8匹のNSGマウスで生着し、VPA含有培養物に由来するCD34+細胞が10匹のマウスで生着した。
培養されたCD34+CD90+細胞の細胞周期状態は、メーカーの取扱説明書に従ってFITC−BrdU Kit(BD Pharmingen)を使用して評価した。VPAの存在下でコントロール条件下において7日間培養したCB CD34+細胞を、続いて2.5時間BrdUでパルスした。次いで、細胞を染色バッファー(3%FBS添加PBS)で洗浄し、CD34−APC及びCD90−PE mAbで染色し、Cytofix/Cytopermバッファーで固定及び透過処理を行い、Perm/Washバッファー(両方ともBD Pharmingen)で洗浄した。透過処理後、細胞を30μgのDNAseで30分間37℃にて処理し、次いで、FITC共役抗BrdU抗体及び7AADで染色した。次いで、CD34+CD90+でゲートした細胞の細胞周期状態を、FAC−SCanto IIフローサイトメーターでFACSDivaソフトウエア(BD Biosciences社)を使用して記録した。
全細胞抽出物は、SCR(8μM)、C433(80μM)、及びVPA(1.25mM)の存在下で培養した、新たに単離されたCB−MNC及びヒト胚性腎臓293(HEK293)細胞から調製した。細胞タンパク質は、Novex(Invitrogen社)を使用してSDS−PAGEによって分離し、iBlot(Invitrogen社)で転写した。メンブレンを、各ヒストンデアセチラーゼ(HDAC1、HDAC2、HDAC3、HDAC4、HDAC5、HDAC6、及びβ−アクチン;Cell Signaling Technology社)に対するmAbで調査し、メーカーの取扱説明書に従ってHRP共役二次抗体(Amersham Biosciences社)を用いた化学発光系を使用して現像した。ウェスタンブロッティングの濃度解析は、ImageJソフトウエア(NIH)で実施した。
高いALDH活性は、未分化造血細胞及び癌幹細胞の特性である(その内容全体が、本明細書に参考として組み込まれる、Hess et al., Functional Characterization of Highly Purified Human Hematopoietic Repopulating Cells Isolated According to Aldehyde Dehydrogenase Activity," Blood 104:1648-1655 (2004);Lioznov et al., "Aldehyde Dehydrogenase Activity as a Marker for the Quality of Hematopoietic Stem Cell Transplants," Bone Marrow Transplant 35:909-914 (2005);Storms et al., "Distinct Hematopoietic Progenitor Compartments are Delineated by the Expression of Aldehyde Dehydrogenase and CD34," Blood 106:95-102 (2005);Aguila et al., "SALL4 is a Robust Stimulator for the Expansion of Hematopoietic Stem Cells," Blood 118:576-585 (2011))。高ALDH活性で細胞集団を同定するために、Aldefluorキット(StemCell Technologies社)をメーカーの取扱説明書に従って使用した。細胞(1×106個/ml)をアッセイバッファー中に懸濁し、次いで細胞の半分をAldefluor基質(テストサンプル)に添加して、残りの半分をDEAB阻害剤(コントロールサンプル)に添加した。テストサンプル及びコントロールサンプルは、40分間37℃にてインキュベートした。その後、細胞をCD34−APC及び/もしくはCD117−PE mAbまたはアイソタイプ適合IgGで、更に20分間染色した。細胞を洗浄し、BD FACSCanto IIフローサイトメーターによって解析した。
全RNAは、ヒトES細胞株H9(WA09;WiCell Research Institute社、マディソン、ウィスコンシン州、アメリカ)、PC、SF及びSC培地でのコントロール培養物またはVPA含有培養物に由来する再単離されたCD34+細胞から、TRIzol及びQIAGEN社(CA)製RNeasyキットを使用して抽出した。全RNA(0.5〜1.0μg)は、RNA to cDNA EcoDry Premixキット(Clontech社)を使用して、cDNAへと逆転写した。プライマー配列は、表6に記載されている。qPCRは、SYBR Green(サーモフィッシャーサイエンティフィック社)及びRealplex thermocycler(Eppendorf社)を使用して実施した。すべての実験は3回実施し、非鋳型コントロール(cDNA鋳型無し)を各アッセイに含めた。GAPDHは、内部標準としての役割を果たした。アンプリコンは、50bpサイズ(DNAラダー)マーカーを用いて2%アガロースゲル上で泳動した。
PC並びにコントロール培養物及びVPA含有培養物に由来する再単離された培養CD34+細胞は、メタノールフリーのホルムアルデヒド(2.8%)で10分間37℃にて固定して、少しの間氷上で冷却し、氷冷した100%メタノールで透過処理を行った。更に、細胞を氷上で20分間インキュベートし、インキュベーションバッファー(0.5%BSA含有PBS)で10分間ブロッキングして、FITC共役mAbまたはアイソタイプコントロールで1時間室温にて、SOX2及びOCT4に対して染色した。細胞はまた、NANOGに対してウサギmAb、続いてFITC共役抗ウサギ二次抗体でも染色し、細胞を洗浄して、フローサイトメトリーによって解析した。
ES細胞及びVPAで7日間処理したCD34+細胞を、RIPAバッファー(50mM Tris−HCl、pH7.4、150mM NaCl、1%Triton、0.1%NP40、及び1.5mM EDTA)中に溶解させた。ES細胞またはVPAで処理された細胞に由来する細胞溶解物(1.0mg)を、6μgのIgG(コントロール)、20μl(6μg)のNANOG pAb(カタログ AF1997;R&D Systems社)でインキュベートして、4℃にて回転プラットフォーム上に一晩置いた。翌日、Protein Gビーズ(50μl)(Cell Signaling Technology社)を添加し、更に4時間サンプルを回転させ続けた。ビーズを溶解バッファーで3回洗浄し、煮沸して、結合タンパク質を溶出させた。ES細胞に由来する全細胞溶解物(25μg)及びVPAで処理された細胞に由来する全細胞溶解物(125μg)を、SDS−PAGEによって分画し、NANOG mAb(Cell Signaling Technology社)を使用してウェスタンブロッティングによって解析した。IP実験からのタンパク質をSDS−PAGEによって分離し、iBlot(Invitrogen社)を使用して転写し、ヤギpAb抗OCT4で免疫ブロットして、洗浄し、ECL検出キットを使用して現像した。
CB CD34+細胞は、SF培養条件下においてVPAで処理した。VPAで処理された細胞に、SOX2、OCT4、NANOG、GAPDH、及びスクランブルsiRNAそれぞれ、またはSOX2、OCT4、及びNANOGの組み合わせであるSilencer Select siRNA(Invitrogen社、CA)をトランスフェクションした。GFPプラスミドは、Neon transfection system(Invitrogen社)のメーカー取扱説明書に従ってトランスフェクション効率を測定するために含めた。
前述の通り、NSGマウスに、4時間300cGyで亜致死線量の放射線を照射をして、PC(2×105)を注入し、サイトカインの存在下もしくは非存在下でのコントロール条件下での培養またはVPA含有培養から1週間後に再単離されたCD34+細胞を、NSGマウスに尾静脈から注入した(その内容全体が、本明細書に参考として組み込まれる、Milhem et al., "Modification of Hematopoietic Stem Cell Fate by 5aza 2'deoxycytidine and Trichostatin A," Blood 103:4102-4110 (2004);Araki et al., "Expansion of Human Umbilical Cord Blood SCID-Repopulating Cells Using Chromatin-Modifying Agents," Exp. Hematol. 34:140-149 (2006);Araki et al., "Chromatin-Modifying Agents Permit Human Hematopoietic Stem Cells to Undergo Multiple Cell Divisions While Retaining Their Repopulating Potential," Blood 109:3570-3578 (2007))。移植後13〜14週間でマウスを犠牲死させた。各マウスのBM細胞を、ヒトCD45−PECy7またはAPC、CD34−APCまたはFITC、CD36−APC、CD33−PECy7、CD14−FITC、CD19−PE、CD41−FITC、CD71−FITC、及びグリコフォリンA−PE(GPA−PE)を発現する細胞の存在について解析した。各レシピエントマウスの骨髄中に、ドナー由来ヒト造血細胞の生着を示す、少なくとも0.1%のヒトCD45+細胞の存在が確認された(その内容全体が、本明細書に参考として組み込まれる、Milhem et al., "Modification of Hematopoietic Stem Cell Fate by 5aza 2'deoxycytidine and Trichostatin A," Blood 103:4102-4110 (2004);Araki et al., "Expansion of Human Umbilical Cord Blood SCID-Repopulating Cells Using Chromatin-Modifying Agents," Exp. Hematol. 34:140-149 (2006);Araki et al., "Chromatin-Modifying Agents Permit Human Hematopoietic Stem Cells to Undergo Multiple Cell Divisions While Retaining Their Repopulating Potential," Blood 109:3570-3578 (2007);Chaurasia et al., "Chromatin-Modifying Agents Promote the Ex vivo Production of Functional Human Erythroid Progenitor Cells," Blood 117:4632-4641 (2011))。一次レシピエントNSGマウスのBM細胞(2×106個)を、亜致死線量の放射線を照射した二次NSGレシピエントマウスに再注入した。移植後15〜16週間でマウスを犠牲死させ、BM細胞をmAbで染色し、上記の通りヒト細胞キメリズムの証拠をフローサイトメトリーによって解析した。
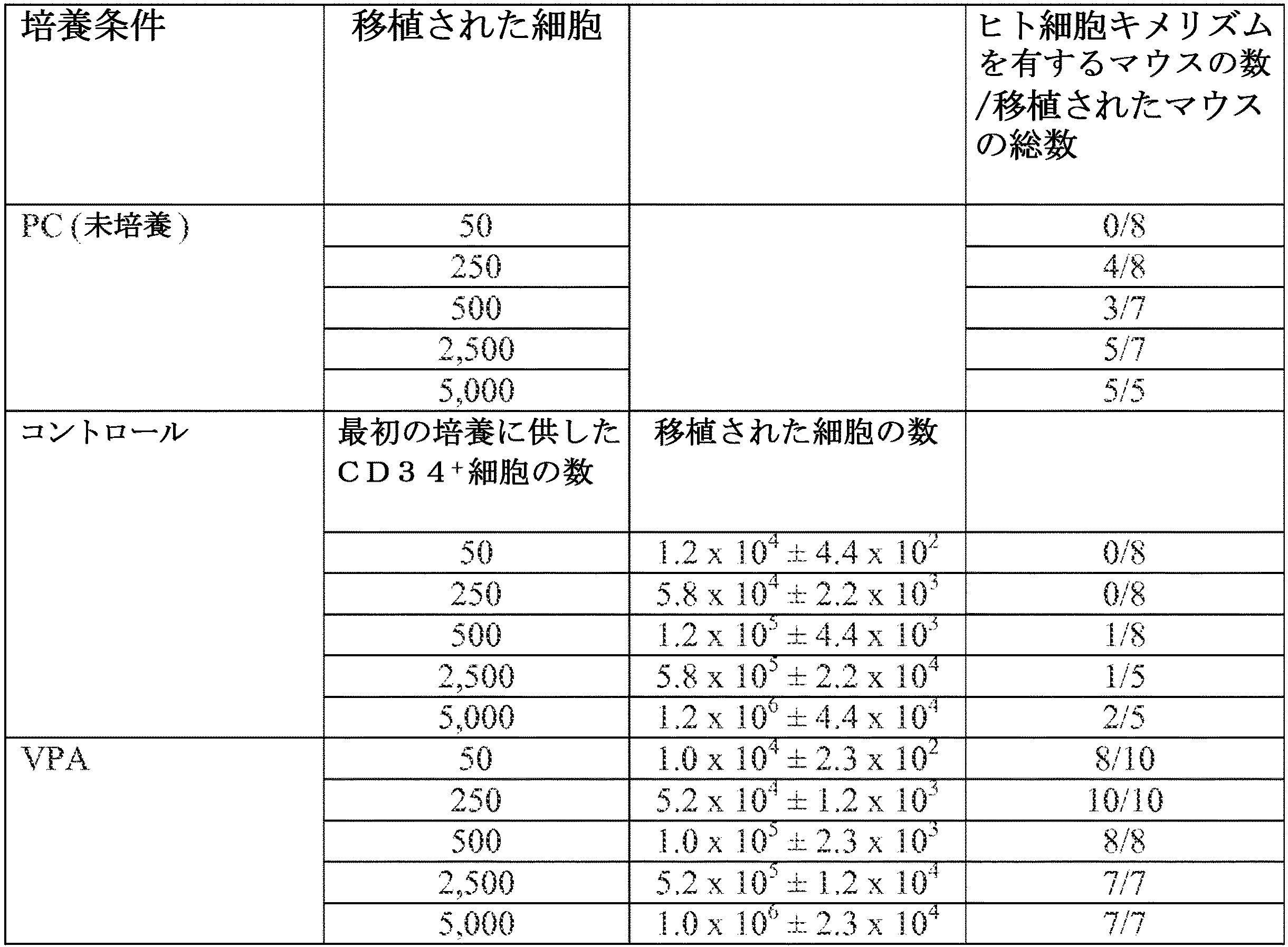
PC及びコントロール条件下またはVPAの存在下で増殖させた当量数のCD34+細胞の子孫におけるヒトSRCの頻度を、前述の通り限界希釈解析によって解析した(その内容全体が、本明細書に参考として組み込まれるBoitano et al., "Aryl Hydrocarbon Receptor Antagonists Promote the Expansion of Human Hematopoietic Stem Cells," Science 329:1345-1348 (2010))。数を増加させたPC(50、250、500、2,500、5,000)またはVPAで7日間もしくはコントロール条件下で培養した当量数のPCの子孫を、NSGマウスに注入した(n=111)。限界希釈実験の結果データを収集し、シングルヒットモデルに対してポアソン統計を適用して解析した(n=111)。頻度は、L−Calcソフトウエア(StemCell Technologies社)を使用して計算し、ウォルター・アンド・エリザ・ホール医学研究所(Walter and Eliza Hall Bioinformatics Institute of Medical Research)で入手可能なELDAソフトウエア(bioinf.wehi.edu.au/software/elda/)を使用してプロットした。以下の式を使用して、未反応値のlogフラクションをネガティブマウスの割合(%)に変換した:ネガティブマウスの割合(%)=e logフラクション。
コントロール培養物もしくはVPA含有培養物から再単離されたCD34+細胞(1×106個)またはほぼ同等の数のES細胞を、100μlのPBS中に懸濁した。これらの細胞を、等容積の氷冷マトリゲルと混合し、グループ(コントロール培養物、VPAで処理された培養物から再単離されたCD34+細胞、及びES細胞)当たり3匹のNSGマウスの右後肢へと皮下注射した。マウスの奇形腫形成を週毎に観察し、8週間後に犠牲死させた。奇形腫を切開、固定、薄切りし、H&Eで染色して形態学的に調査した(その内容全体が、本明細書に参考として組み込まれるO'Connor et al., "Functional Assays for Human Embryonic Stem Cell Pluripotency," Methods Mol Biol. 690:67-80 (2011))。
結果は、各実験における様々な数値の平均±SDまたは平均±SEMとして表した。統計差は、別段の指定がない限り、スチューデント両側t検定を使用して評価した。テューキーの検定による対比較を併用した一元配置ANOVA並びに/または等分散性のためのバートレット検定及び分散比較のためのF検定も使用した。0.05以下のP値は、有意であると判断した。
すべての動物実験は、アイカーン医科大学(Icahn School of Medicine)の動物実験委員会(animal care and use committee)によって承認された。少量の匿名CB単位は、ニューヨーク血液センター(New York Blood Center)から購入したので、インフォームドコンセントまたは対象承認は、本研究では不要であった。
CB CD34+細胞及びCD34+CD90+細胞のエクスビボにおける増殖に対するHDACI及びSF培地の効果
HSCのエクスビボにおける増殖を可能にする培養条件の決定は、多数の研究の主題であった(その内容全体が、本明細書に参考として組み込まれる、Dahlberg et al., "Ex vivo Expansion of Human Hematopoietic Stem and Progenitor Cells," Blood 117:6083-6090 (2011);Delaney et al., "Strategies to Enhance Umbilical Cord Blood Stem Cell Engraftment in Adult Patients," Expert Rev. Hematol. 3:273-283 (2010);Boitano et al., "Aryl Hydrocarbon Receptor Antagonists Promote the Expansion of Human Hematopoietic Stem Cells," Science 329:1345-1348 (2010);De Felice et al., "Histone Deacetylase Inhibitor Valproic Acid Enhances the Cytokine-Induced Expansion of Human Hematopoietic Stem Cells," Cancer Res. 65:1505-1513 (2005);Himburg et al., "Pleiotrophin Regulates the Expansion and Regeneration of Hematopoietic Stem Cells," Nat. Med. 16:475-482 (2010);Milhem et al., "Modification of Hematopoietic Stem Cell Fate by 5aza 2'deoxycytidine and Trichostatin A," Blood 103:4102-4110 (2004);Nishino et al., "Ex vivo Expansion of Human Hematopoietic Stem Cells by a Small-Molecule Agonist of c-MPL," Exp. Hematol. 37:1364-1377 e1364. (2009);North et al., "Prostaglandin E2 Regulates Vertebrate Haematopoietic Stem Cell Homeostasis," Nature 447:1007-1011 (2007))。SC培養条件並びにDNAメチルトランスフェラーゼ阻害剤(DNMTI)及びHDACIを用いた一連の処理を使用すると、CB HSC数の増加が制限される可能性があると、これまでに実証されている(その内容全体が、本明細書に参考として組み込まれる、Milhem et al., "Modification of Hematopoietic Stem Cell Fate by 5aza 2'deoxycytidine and Trichostatin A," Blood 103:4102-4110 (2004);Araki et al., "Expansion of Human Umbilical Cord Blood SCID-Repopulating Cells Using Chromatin-Modifying Agents," Exp. Hematol. 34:140-149 (2006);Araki et al., "Chromatin-Modifying Agents Permit Human Hematopoietic Stem Cells to Undergo Multiple Cell Divisions While Retaining Their Repopulating Potential," Blood 109:3570-3578 (2007);Chaurasia et al., "Chromatin-Modifying Agents Promote the Ex vivo Production of Functional Human Erythroid Progenitor Cells," Blood 117:4632-4641 (2011))。CB HSC増殖をより促進させる培養条件を更に最適化するために、まずCD34+細胞、次いでCD34+CD90+細胞の産生を、本明細書においてはコントロール条件と称する、サイトカインを補充したSF培養及びSC培養において評価した。CB CD34+細胞を増殖させるために使用したエクスビボにおける増殖方法の略図及び検討した細胞集団を表すために使用した用語は、図1A及び表1で提供される。種々のHDACI(VPA、スクリプタイド[SCR]、トリコスタチンA[TSA]、スベロイルアニリドヒドロキサム酸[SAHA]、BML−210としても知られるCAY10433[C433]、MD85としても知られるCAY10398、及びCAY10603[分子式:C22H30N4O6])を、様々な添加量及びインキュベーション期間(5〜9日間)で、添加した。SCまたはSF培養条件下において、インビトロで産生させたCD34+細胞数を増加させる、HDACIの能力を評価した。検討した8つのHDACIの内、VPA、SCR、及びC433で7日間処理することが、本目的に対して最も有効であると分かった。コントロール条件(16.2%±9.2%)と比較して、これらの剤の各々で処理すると、ほぼ同等の割合(%)のCD34+CD90+細胞(SCR:73.4%±13.9%、C433:70.1%±18.4%、及びVPA:75.2%±10.7%)が産生された(ANOVA、P<0.0001)。しかしながら、7日間を超えて細胞を維持すると、その割合(%)は次第に減少した。同様に、これらのHDACIの各々は、CD34+CD90+細胞によるCXCR4発現(CD184)を促進するだけでなく、CB収集物当たりのCD34+細胞及びCD34+CD90+細胞の産生絶対数を高め(ANOVA、P≦0.0007)(図1B及び1C)、コントロール条件と比較してCD34+CD90+CD184+細胞の産生絶対数を高める(ANOVA、P<0.0001)(図1D)のに有効であったことが見出された。最適濃度及び半最適濃度で混ぜ合わせた場合、VPA、SCR、及びC433の効果は、付加的ではなかった。
HDACは、多タンパク質複合体のサブユニットとして細胞内に存在し、遺伝子発現を制御する。クラスI HDAC(HDAC1、2、3、及び8)は、酵母の転写制御因子RPD3に対して配列相同性を有する。しかしながら、クラスII HDAC(HDAC4、5、6、7、9、及び10)は、酵母内で見られるデアセチラーゼHDA1と類似したドメインを共有する(その全体が参照として本明細書に組み込まれるDelcuve et al., "Roles of Histone Deacetylases in Epigenetic Regulation: Emerging Paradigms from Studies with Inhibitors," Clin. Epigenetics 4(1):5 (2012))。クラスI及びII HDACは、転写コリプレッサーmSIN3、NCoR、及びSMRTと相互作用し、それによりHDACを転写因子へとリクルートする(その内容全体が、本明細書に参考として組み込まれるDelcuve et al., "Roles of Histone Deacetylases in Epigenetic Regulation: Emerging Paradigms from Studies with Inhibitors," Clin. Epigenetics 4(1):5 (2012);Kramer et al., "The Histone Deacetylase Inhibitor Valproic Acid Selectively Induces Proteasomal Degradation of HDAC2," EMBO J. 22:3411-3420 (2003))。HDACIは、CB CD34+細胞におけるH3アセチル化を促進させることが、これまでに分かっている(その内容全体が、本明細書に参考として組み込まれるChaurasia et al., "Chromatin-Modifying Agents Promote the Ex vivo Production of Functional Human Erythroid Progenitor Cells," Blood 117:4632-4641 (2011))。しかしながら、HDAC活性は、触媒ドメインでのこれらの阻害剤との結合によるだけでなく、ユビキチン/プロテアソーム経路を介するHDAC分解の微調整によっても調節することができる。E2ユビキチンコンジュガーゼ(conjugase)Ubc8及びE3ユビキチンリガーゼRLIMの量を制限することで、VPAによる調節を受けやすいHDACのバランスのとれた定常状態のタンパク質レベルが維持されると報告されてきた(その内容全体が、本明細書に参考として組み込まれる、Kramer et al., "The Histone Deacetylase Inhibitor Valproic Acid Selectively Induces Proteasomal Degradation of HDAC2," EMBO J. 22:3411-3420 (2003);Cedar et al., "Epigenetics of Haematopoietic Cell Development," Nat. Rev. Immunol. 11:478-488 (2011);Cunliffe, "Eloquent Silence: Developmental Functions of Class I Histone Deacetylases," Curr. Opin. Genet. Dev. 18(5):404-410 (2008))。HDACがHDACIの影響を受けることを決定するために、クラスI及びIIの両方のHDACタンパク質レベルに対するSCR、C433、及びVPAの効果を、CB単核細胞(CB−MNC)及びヒト胚性腎臓293(HEK293)細胞において、処理の2時間及び24時間後に評価した。CB−MNCの処理から2時間後、SCR、C433、及びVPAはHDACの発現を阻害しなかったが、処理から24時間後、異なる割合でクラスI(HDAC1、2、及び3)、クラスIIa(HDAC4及び5)、並びにクラスIIb(HDAC6)HDACを阻害した。SCR及びC433は、各HDACに対して最も有効な阻害剤であった(図6及び表7)。
サイトカインを用いてインビトロで増殖させたCB及び骨髄細胞の表現型は、必ずしも機能と関連するとは限らないので、ALDH活性をHSCの機能マーカーとして使用した(その内容全体が、本明細書に参考として組み込まれる、Hess et al., Functional Characterization of Highly Purified Human Hematopoietic Repopulating Cells Isolated According to Aldehyde Dehydrogenase Activity," Blood 104:1648-1655 (2004);Lioznov et al., "Aldehyde Dehydrogenase Activity as a Marker for the Quality of Hematopoietic Stem Cell Transplants," Bone Marrow Transplant 35:909-914 (2005);Spangrude et al., "Long-Term Repopulation of Irradiated Mice with Limiting Numbers of Purified Hematopoietic Stem Cells: In vivo Expansion of Stem Cell Phenotype but not Function," Blood 85(4):1006-1016 (1995);Storms et al., "Distinct Hematopoietic Progenitor Compartments are Delineated by the Expression of Aldehyde Dehydrogenase and CD34," Blood 106:95-102 (2005);Veeraputhiran et al., "Aldehyde Dehydrogenase as an Alternative to Enumeration of Total and Viable CD34(+) Cells in Autologous Hematopoietic Progenitor Cell Transplantation," Cytotherapy 13:1256-1258 (2011))。サイトカインの存在下にて、SC培養中で培養された細胞よりも、SF培養中で培養された細胞において、より多くのALDH活性を有する細胞画分が観察された。更に、SF培養条件下でサイトカインを含有する培養にVPAを添加すると、SC培養中で観察されたのと比較して、ALDH+細胞の比率がより高くなった(図7A及び表2)。
VPAを添加したSF培養において産生されたALDH+CD34+CD117+細胞の絶対数は、SC培養中で得られた数よりも多かった(P=0.009)(図7B)。
転写因子SOX2、OCT4、及びNANOGは、そのプロモーターを含む標的遺伝子を共同占有し、それによって、自己複製及び多能性の維持に必要な調節及び自己調節フィードバックループの両方で協調することにより、胚性幹細胞及び人工多能性幹細胞(iPS)の両方の運命決定を確定させる調節機能の中核を担う(その内容全体が、本明細書に参考として組み込まれる、Boyer et al., "Core Transcriptional Regulatory Circuitry in Human Embryonic Stem Cells," Cell 122:947-956 (2005);Loh et al., "The Oct4 and Nanog Transcription Network Regulates Pluripotency in Mouse Embryonic Stem Cells," Nat. Genet. 38(4):431-440 (2006))。VPA媒介HSC増殖における、このようなマスター転写因子の役割は、コントロール条件下での培養またはSF及びSC培地中にてVPAで処理された培養の7日後に再単離されたCD34+細胞における、SOX2、OCT4、及びNANOGの発現を調査することで探求した。RT−PCRは、VPA含有SF培養物に由来するCD34+細胞におけるSOX2、OCT4、及びNANOG転写の発現を明らかにしたが、コントロール培養またはVPAが添加されたSC培養中では、OCT4及びSOX2転写がほとんど検出できなかった(図8A)。定量PCR(qPCR)によって、これらの多能性遺伝子の発現は、SC培養中で観察されたのと比較して、SF培養中におけるVPAの存在下で増加したことが実証された(ANOVA、P=0.0001)(図8B)。成体幹細胞におけるOCT4の発現は、機能型のOCT4ではなく、実際には不活性な偽遺伝子の発現によるものであるという可能性が、他の研究者によって報告されている(その内容全体が、本明細書に参考として組み込まれる、Redshaw et al., "Human Haematopoietic Stem Cells Express Oct4 Pseudogenes and Lack the Ability to Initiate Oct4 Promoter-Driven Gene Expression," J. Negat. Results Biomed. 9(1):2-8 (2010);Zangrossi et al., "Oct-4 Expression in Adult Human Differentiated Cells Challenges its Role as a Pure Stem Cell Marker," Stem Cells 25:1675-1680 (2007))。RT−PCRを使用して、1つのOCT4偽遺伝子が、VPAで処理されたCD34+細胞においては存在しなかったことが見出された(図8A)。qPCR解析によって、2つの偽遺伝子の転写は存在しなかったことが示された。OCT4、SOX2、及びNANOGとは異なり、ES細胞における別の既知の多能性マーカーであるテロメラーゼ逆転写酵素(hTERT)(その内容全体が、本明細書に参考として組み込まれるTakahashi et al., "Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors," Cell 131:861-872 (2007))は、VPAで処理されたCD34+細胞において増加しなかった。クロマチンリモデリングにおいて重要な役割を担うSET、SMARCAD1、及びMYST3を含む、SOX2、OCT4、及びNANOGの下流標的遺伝子を、追加の多能性遺伝子ZIC3も加えて調査した(その内容全体が、本明細書に参考として組み込まれる、Boyer et al., "Core Transcriptional Regulatory Circuitry in Human Embryonic Stem Cells," Cell 122:947-956 (2005);Takahashi et al., "Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors," Cell 131:861-872 (2007);Lim et al., "Zic3 is Required for Maintenance of Pluripotency in Embryonic Stem Cells," Mol. Biol. Cell 18:1348-1358 (2007))。SMARCAD1、MYST3、及びZIC3、ただしSETは除く、もまたSF培養中のVPAで処理されたCD34+細胞において劇的に増加した(ANOVA、P=0.04)。ZIC3 mRNAは、血清の存在下でVPAを補充したコントロール培養中では検出されなかったが、VPAを補充したSF培養中においてのみ増加した(図8C)。ZIC3遺伝子は、ES細胞においてOCT4、SOX2、及びNANOGの標的として同定されてきた。OCT4、NANOG、及びSOX2の転写ネットワークと一部重なり合うZIC3は、多能性の維持に重要であり、NANOGの発現を直接調節することができる(その内容全体が、本明細書に参考として組み込まれる、Boyer et al., "Core Transcriptional Regulatory Circuitry in Human Embryonic Stem Cells," Cell 122:947-956 (2005);Lim et al., "Zic3 is Required for Maintenance of Pluripotency in Embryonic Stem Cells," Mol. Biol. Cell 18:1348-1358 (2007);Declercq et al., "Zic3 Enhances the Generation of Mouse Induced Pluripotent Stem Cells," Stem Cells Dev. (2013))。
次いで、NANOG及びOCT4間の物理的相互作用を、VPAで処理された細胞に由来するタンパク質のco−IPによって記録した(図9C)。更に、ウェスタンブロット解析によって、VPAで処理された細胞における内因性OCT4及びNANOGの発現は、ES細胞の場合よりも多くなかったことが明らかとなった(図9C)。
VPAで処理された培養物における多能性遺伝子の上方制御とCD34+CD90+細胞の増殖との間の機能的関連性を立証するために、CD34+細胞に各siRNAまたはSOX2、OCT4、及びNANOG(SON)に対するsiRNAの複合プールのいずれかをトランスフェクションした。初めに、異なる濃度のsiRNAを、各遺伝子について、及びコントロール培養またはVPAで処理された培養中の細胞に対する遺伝子の潜在的な毒性作用について試験した。SON siRNAで処理された細胞の形態学的外観は、スクランブルsiRNAで処理されたのと比較して、変化はなかった。スクランブル、SON、及びGAPDH siRNAのトランスフェクション後、コントロール培養及びVPA含有培養において産生された細胞の総数に、有意な減少は観察されなかった(表8)。多能性遺伝子の発現は、qPCR及びRT−PCRを使用して、VPAで処理された培養物におけるsiRNAトランスフェクション後に観察した(図10A及び10B)。siRNAによるノックダウンによって、SOX2、OCT4、NANOG、及びZIC3転写の発現(80%〜84%)が著しく減少した(ANOVA、P<0.0001)。共焦点顕微鏡及びmAb染色によっても、SOX2、OCT4、及びNANOGタンパク質の発現が著しく減少したことが明らかとなった。OCT4、SOX2、及びNANOG調節ネットワークの下流であるZIC3タンパク質の発現は、より少ない程度に減少した(図10C)。CD34+細胞(47.8%±4.4%対22.8%±8.6%)及びCD34+CD90+細胞(20.5%±6.1%対11.7%±4.5%)(ANOVA、P=0.0005)の割合(%)における有意な減少が、多能性遺伝子各々について特異的な各siRNAまたはスクランブルsiRNAと比較して、SON siRNAでの処理後に観察された(図10D)。更に、VPAで処理されたCD34+細胞をSON siRNAで処理した後、CB収集物当たりのCD34+細胞及びCD34+CD90+細胞の絶対数が、それぞれ89.1%及び88.7%(ANOVA、P=0.0008)減少したことが観察された(図10E)。
下部パネル:VPAで処理された培養物は、方法セクションにて先に記載したとおりにスクランブルsiRNA及びGAPDH siRNAをトランスフェクションした。CD34+CD90+細胞の総細胞数及び割合(%)における有意差は、GAPDH siRNAトランスフェクションから72時間後では観察されなかった。(n=3)(*p<0.25、ns)
PC並びにコントロール条件下並びにVPA及びサイトカインで培養されたCB CD34+細胞の骨髄再構築能を、NSGレシピエントマウスの骨髄内での細胞生着を確認することによって評価した。すべてのレシピエントマウスにおいて、移植された移植片の種類に関係なく、ヒトCD45+細胞及びCD45+CD34+細胞が検出された。移植から13〜14週間後(図11A及び11B)、PCが生着したマウス内における骨髄細胞の19.4%±4.9%がドナー由来のCD45+細胞であり、それに比較して、コントロール培養物に由来する細胞が生着したマウス内では13.2%±6.4%であった。一方、VPAで処理されたCB CD34+細胞の移植では、ヒトCD45+細胞キメリズム(32.2%±11.3%)及びCD45+CD34+細胞(13.0%±8.7%)の割合が、コントロール細胞で得られたのと比較して、より高い割合となった(それぞれ、P=0.0008及びP=0.004)(図11A及び11B)。VPA移植片でのCD45+細胞キメリズムの割合もまた、PCで得られた割合よりも統計上高い割合となった(P=0.006)。
(平均±SE、*p<0.05、(ANOVA P<0.0001)。NSGマウスレシピエント(n=15))。
HSCの多能性は、HSCのクロマチン構造及びエピジェネティックな可塑性が動的に維持されることによって説明することができる。クロマチン構造の変換は、主に特異的なヒストンの翻訳後修飾により制御されて、結果として得られたクロマチン構造が、許容的か抑制的かを決定する(その内容全体が、本明細書に参考として組み込まれる、Cedar et al., "Epigenetics of Haematopoietic Cell Development," Nat. Rev. Immunol. 11:478-488 (2011);Kouzarides, "Chromatin Modifications and Their Function," Cell 128:693-705 (2007);Oh et al., "Concise Review: Multidimensional Regulation of the Hematopoietic Stem Cell State," Stem Cells 30:82-88 (2012))。CB CD34+細胞による幹細胞機能がSC培養条件及びサイトカインの組み合わせを使用したインビトロでの培養後に次第に失われることは、依然として、移植可能なHSC数のインビボでの増加に対する障壁である(その内容全体が、本明細書に参考として組み込まれる、Giebel et al., "Primitive Human Hematopoietic Cells Give Rise to Differentially Specified Daughter Cells Upon their Initial Cell Division," Blood 107(5):2146-2152 (2006);Ho et al., "The Beauty of Asymmetry: Asymmetric Divisions and Self-Renewal in the Haematopoietic System," Curr. Opin. Hematol. 14(4):330-336 (2007);Huang et al., "Symmetry of Initial Cell Divisions Among Primitive Hematopoietic Progenitors Is Independent of Ontogenic Age and Regulatory Molecules," Blood 94(8):2595-2604 (1999);Srour et al., "Modulation of In vitro Proliferation Kinetics and Primitive Hematopoietic Potential of Individual Human CD34+CD38-/lo Cells in G0," Blood 105(8):3109-3116 (2005))。この幹細胞機能の減退は、十分に機能的なHSCが、宿主内に存在する許容的環境から分離されて不利なエクスビボ環境へと配置され、それにより、自己複製能、骨髄再構築能、及び多系統分化能を含む、幹細胞の重要な機能を決定する遺伝子発現プログラムを変更するようなエピジェネティックな変化がもたらされることに起因すると考えられる。このHSC機能の喪失は、使用された培養条件に対して生じる細胞周期の急激な進行及び細胞分裂に起因している(その内容全体が、本明細書に参考として組み込まれる、Declercq et al., "Zic3 Enhances the Generation of Mouse Induced Pluripotent Stem Cells," Stem Cells Dev. (2013);Sauvageau et al., "In vitro and In vivo Expansion of Hematopoietic Stem Cells," Oncogene 23:7223-7232 (2004);Walasek et al., "Hematopoietic Stem Cell Expansion: Challenges and Opportunities," Ann. NY Acad. Sci. 1266:138-150 (2012))。エクスビボでの幹細胞増殖における先の試みは、造血微小環境と類似させ、それによって幹細胞の機能的完全性の保持を促す環境の構築に重点を置いたことが恐らく部分的な要因となり、ある程度の成果を得てきた(その内容全体が、本明細書に参考として組み込まれる、Dahlberg et al., "Ex vivo Expansion of Human Hematopoietic Stem and Progenitor Cells," Blood 117:6083-6090 (2011);Delaney et al., "Cord Blood Graft Engineering," Biol. Blood Marrow Transplant 19(1): S74-S78 (2013);Delaney et al., "Strategies to Enhance Umbilical Cord Blood Stem Cell Engraftment in Adult Patients," Expert Rev. Hematol. 3:273-283 (2010))。エクスビボでのこのような微小環境構築の難しさは、十分理解されており、クロマチン構造に影響を及ぼす剤を使用してHSCのエピジェネティック特性を直接維持する試みである別のアプローチが取られてきた。このアプローチは、クロマチン状態の動的変化が、ヌクレオソームリモデリング、DNAメチル化、及びヒストンアセチル化によって媒介されるという理解に基づいている(その内容全体が、本明細書に参考として組み込まれる、Cedar et al., "Epigenetics of Haematopoietic Cell Development," Nat. Rev. Immunol. 11:478-488 (2011);Kouzarides, "Chromatin Modifications and Their Function," Cell 128:693-705 (2007);Oh et al., "Concise Review: Multidimensional Regulation of the Hematopoietic Stem Cell State," Stem Cells 30:82-88 (2012);Elizalde et al., "Histone Deacetylase 3 Modulates the Expansion of Human Hematopoietic Stem Cells," Stem Cells Dev. 21:2581-2591 (2012))。このために、本研究では、細胞分裂を繰り返した後でも幹細胞の機能保持に関与する遺伝子を持つクロマチン構造の脱凝縮を図るために、いくつかのHDACIについて、クラスI及びII HDACの両方を阻害することが可能であるかを評価した。HDAC1、HDAC3、及びHDAC5タンパク質は、3種の最も活性なHDACIによって均一に減少し、このことは、これら3種のHDACの組み合わせは、インビトロでの分裂後も幹細胞の機能を保持することを支持する幹細胞の運命決定において重要な役割を担うことを示唆した。
VPA補充SF培養を用いて産生させた精製ALDH+CD34+細胞では、産生したBFU−E及びCFU−Mixの絶対数(8.4x107±6.7x107/CB収集物)が、SC培養(1.4x107±0.88x107)と比較して、より多かった(ANOVA,p=0.001)(図20A)。一方、VPA補充SC培養物に由来するALDH−CD34+細胞では、産生したBFU−E及びCFU−Mixの数(2.7x106±1.0x106)が、SF培養(3.5x105±0.78x105)と比較して、より多かった(ANOVA,p=0.01)(図20B)。これらのデータは、血清が、ALDH+細胞及びALDH−細胞の集団のインビトロでの運命に対して、特異的に影響を与えることを示している。
増殖させたHSC生成物は、移植センターへの配送前に凍結保存すると予想されるので、HSC表現型を発現する生存可能な細胞の回復に対する凍結保存の効果を調査した。CB−CD34+細胞をVPAで7日間処理し、細胞数を数えて、解凍前後の両方で表現型解析を実施した。凍結保存は、Invitrogen社(Life Technologies社、グランドアイランド、NY)製Synth−a−Freezeを使用して実施した。
VPAにより増殖したHSCは多数の長期再構築細胞を含むことを示した。これらの細胞は、全能性細胞の特性である多能性遺伝子を発現するので、このような移植片には生着遅延を伴う可能性があるという妥当な懸念がある。Dick研究室では、短期並びに中間時点後及び長期間後にNSGマウスにおいて再構築されるHSCの階層を定義し、その表現型階層特性を同定した(その全体が参照として本明細書に組み込まれるNotta et al., "Isolation of Single Human Hematopoietic Stem Cells Capable of Long-term Multilineage Engraftment," Science 333(6039):218-221 (2011))。短期SCID再構築細胞(R−SRC)は移植後2〜4週間でNSGマウスから検出され、その表現型はCD34+CD90−CD49f−であり、中期SRC(IT−SRC)は12〜14週間後に再構築され、その表現型はCD34+CD90+CD49f−であり、及び長期SRC(LT−SRC)は>24週間後に再構築され、その表現型はCD34+CD90+CD49f+である。これらの様々なHSCクラスの分布を、VPAにより増殖したHSC生成物3つについて評価した。図19A〜Cに見ることができる通り、HSCのこれらのクラスは各々、CB−CD34+初代細胞と比較して、VPAで処理された細胞に存在する数が多かった。このことは、これらの増殖移植片が、未処理のCB移植片と比較して恐らくより短期で、持続的な生着パターンをもたらし、加えて移植失敗の発生率を低下させるだろうことを示唆した。興味深いことに、SF条件下にてサイトカインのみの存在下で処理したCB−CD34+細胞が含む、R−SRC数が最も高い(図19C)。この興味深い結果は、第1相試験の完了後にVPAで処理された移植片の注入が、造血回復に対して短期と関連がない場合、重要であり得る。これらの結果に基づいて、R−SRC数をより大幅に増加させることで更に短期での生着を促進させるために、CD34+初代細胞のごく一部をサイトカインのみの存在下で増殖し、VPAで処理された細胞生成物と混ぜ合わせることが可能となった。
Claims (30)
- 単離されかつ増殖したヒト造血臍帯血幹細胞の濃縮された集団であって、該造血幹細胞が、サイトカインSCF、Flt3、TPO、及びIL3と接触しており、該造血幹細胞が、CD34+、CD90+、CD184+、CD117+、CD49f+、ALDH+、かつCD45RA−でありかつ多能性遺伝子SOX2、OCT4、NANOG、及びZIC3を発現する、前記濃縮された集団。
- 前記造血幹細胞の少なくとも60%が、アルデヒドデヒドロゲナーゼ活性に対して陽性である、請求項1に記載の濃縮された集団。
- 前記造血幹細胞が、通常のヒト造血幹細胞と比較して、胚性幹細胞の多能性遺伝子hTERTの発現レベルにおいて上方制御を示さない、請求項1または請求項2に記載の濃縮された集団。
- 前記造血幹細胞の少なくとも95%が、SOX2、OCT4、NANOG、及びZIC3に対して陽性である、請求項1〜3のいずれか一項に記載の濃縮された集団。
- 前記造血幹細胞が、ヒストンデアセチラーゼ阻害剤と接触している、請求項1〜4のいずれか一項に記載の濃縮された集団。
- 前記ヒストンデアセチラーゼ阻害剤が、VPA、SCR、LBH589、TSA、SAHA、Cay10433、及びCay10398からなる群より選択される、請求項5に記載の濃縮された集団。
- 前記造血幹細胞が、ヒストンデアセチラーゼ阻害剤と接触していない単離されかつ増殖したヒト造血臍帯血幹細胞と比較して減少した、HDAC1、HDAC3、及びHDAC5の発現を有する、請求項5または請求項6に記載の濃縮された集団。
- 前記造血幹細胞の少なくとも18.0%±1.2%が、G2/M期にある、請求項1〜7のいずれか一項に記載の濃縮された集団。
- 前記造血幹細胞の少なくとも23.2%±13.8%が、G0/G1期にある、請求項1〜8のいずれか一項に記載の濃縮された集団。
- 以下の工程を含む、単離されかつ増殖したヒト造血臍帯血幹細胞の濃縮された集団の作製方法:
単離されたヒト造血臍帯血幹細胞の集団を提供する工程であって、該造血幹細胞が、サイトカインSCF、Flt3、TPO、及びIL3と接触している、工程、ならびに
単離されかつ増殖したヒト造血臍帯血幹細胞の濃縮された集団を作製するために、無血清培養系においてヒストンデアセチラーゼ阻害剤の存在下にて、ヒト造血臍帯血幹細胞の単離された集団を処理する工程であって、該増殖したヒト造血臍帯血幹細胞が、CD34+、CD90+、CD184+、CD117+、CD49f+、ALDH+、かつCD45RA−でありかつ多能性遺伝子SOX2、OCT4、NANOG、及びZIC3を発現する、工程。 - 前記単離されたヒト造血臍帯血幹細胞の集団が、前記処理を行うと20,202倍に増殖する、請求項10に記載の方法。
- 前記処理が7日間実施される、請求項10または請求項11に記載の方法。
- 前記増殖した造血幹細胞の少なくとも88.3%±5.9%が、アルデヒドデヒドロゲナーゼ活性に対して陽性である、請求項10〜12のいずれか一項に記載の方法。
- 前記増殖した造血幹細胞が、通常のヒト造血幹細胞と比較して、胚性幹細胞の多能性遺伝子hTERTの発現レベルにおいて上方制御を示さない、請求項10〜13のいずれか一項に記載の方法。
- 前記増殖した造血幹細胞の少なくとも95%が、SOX2、OCT4、NANOG、及びZIC3に対して陽性である、請求項10〜14のいずれか一項に記載の方法。
- 前記ヒストンデアセチラーゼ阻害剤が、VPA、SCR、LBH589、TSA、SAHA、Cay10433、及びCay10398からなる群より選択される、請求項10〜15のいずれか一項に記載の方法。
- 前記増殖した造血幹細胞が、前記処理前の前記単離された幹細胞と比較して減少した、HDAC1、HDAC3、及びHDAC5の発現を有する、請求項10〜16のいずれか一項に記載の方法。
- 前記増殖した造血幹細胞の少なくとも95%が、SOX2、OCT4、NANOG、及びZIC3に対して陽性である、請求項10〜17のいずれか一項に記載の方法。
- 前記増殖した造血幹細胞の少なくとも18.0%±1.2%が、G2/M期にある、請求項10〜18のいずれか一項に記載の方法。
- 前記増殖した造血幹細胞の少なくとも23.2%±13.8%が、G0/G1期にある、請求項10〜19のいずれか一項に記載の方法。
- 対象の血液疾患を治療するための、請求項1〜9のいずれか一項に記載の単離されかつ増殖したヒト造血臍帯血幹細胞の濃縮された集団を含む薬学的組成物であって、該対象に投与される、薬学的組成物。
- 前記投与する工程が、濃縮または精製された調製物に由来する前記造血幹細胞を前記対象内に移植することによって実施される、請求項21に記載の薬学的組成物。
- 前記増殖した造血幹細胞が、前記対象由来である、請求項21または請求項22に記載の薬学的組成物。
- 前記増殖した造血幹細胞の少なくとも60%が、アルデヒドデヒドロゲナーゼ活性に対して陽性である、請求項21〜23のいずれか一項に記載の薬学的組成物。
- 前記増殖した造血幹細胞が、通常のヒト造血幹細胞と比較して、胚性幹細胞の多能性遺伝子hTERTの発現レベルにおいて全く上方制御を示さない、請求項21〜24のいずれか一項に記載の薬学的組成物。
- 前記増殖した造血幹細胞の少なくとも95%が、SOX2、OCT4、NANOG、及びZIC3に対して陽性である、請求項21〜25のいずれか一項に記載の薬学的組成物。
- 前記増殖した造血幹細胞の少なくとも23%が、G2/M期にある、請求項21〜26のいずれか一項に記載の薬学的組成物。
- 前記増殖した造血幹細胞の少なくとも18%が、G0/G1期にある、請求項21〜27のいずれか一項に記載の薬学的組成物。
- 以下の工程を含む、造血幹細胞に対する化合物の効果を決定する方法:
請求項1〜9のいずれか一項に記載の単離されかつ増殖したヒト造血臍帯血幹細胞の濃縮された集団を提供する工程;
該造血幹細胞を、試験される化合物に接触させる工程;ならびに
該造血幹細胞に対する該化合物の効果を決定するために、該接触後に該造血幹細胞を解析する工程。 - 前記処理が、単一のエピジェネティックな制御因子の存在下にて実施され、該単一のエピジェネティックな制御因子がヒストンデアセチラーゼ阻害剤である、請求項10に記載の方法。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201361825354P | 2013-05-20 | 2013-05-20 | |
| US61/825,354 | 2013-05-20 | ||
| US201461983805P | 2014-04-24 | 2014-04-24 | |
| US61/983,805 | 2014-04-24 | ||
| PCT/US2014/038361 WO2014189781A1 (en) | 2013-05-20 | 2014-05-16 | Enriched and expanded human cord blood stem cells for treatment of hematological disorders |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2019087708A Division JP2019150040A (ja) | 2013-05-20 | 2019-05-07 | 血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2016524468A JP2016524468A (ja) | 2016-08-18 |
| JP6525963B2 true JP6525963B2 (ja) | 2019-06-05 |
Family
ID=51933996
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2016514978A Active JP6525963B2 (ja) | 2013-05-20 | 2014-05-16 | 血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 |
| JP2019087708A Pending JP2019150040A (ja) | 2013-05-20 | 2019-05-07 | 血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2019087708A Pending JP2019150040A (ja) | 2013-05-20 | 2019-05-07 | 血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US11261429B2 (ja) |
| EP (1) | EP2999343A4 (ja) |
| JP (2) | JP6525963B2 (ja) |
| CN (2) | CN105705021B (ja) |
| CA (1) | CA2912688C (ja) |
| WO (1) | WO2014189781A1 (ja) |
Families Citing this family (19)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP6525963B2 (ja) | 2013-05-20 | 2019-06-05 | アイカーン スクール オブ メディシン アット マウント サイナイ | 血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 |
| WO2017053729A1 (en) * | 2015-09-25 | 2017-03-30 | The Board Of Trustees Of The Leland Stanford Junior University | Nuclease-mediated genome editing of primary cells and enrichment thereof |
| WO2017079744A1 (en) * | 2015-11-05 | 2017-05-11 | President And Fellows Of Harvard College | Enhancement of stem cell engraftment with oncostatin m |
| EP3493820A4 (en) * | 2016-08-04 | 2020-04-22 | Icahn School of Medicine at Mount Sinai | PRODUCTION OF MEGACARYOCYTE COMPOSITIONS AND THROMBOCYTOPENIA TREATMENT THERAPIES |
| CN108456658A (zh) * | 2017-02-22 | 2018-08-28 | 苏州方舟基因药业有限公司 | 促进造血干细胞扩增并抑制其分化的化合物及细胞因子组合物 |
| US20210380937A1 (en) | 2017-04-04 | 2021-12-09 | The Board Of Trustees Of The Leland Stanford Junior University | Preparation , expansion, and uses of adult pluripotent stem cells |
| GB201706544D0 (en) * | 2017-04-25 | 2017-06-07 | Plasticell Ltd | Method |
| US11458122B2 (en) | 2017-08-09 | 2022-10-04 | Children's Hospital Medical Center | Methods for treating diseases and nerve injuries |
| CN110205283B (zh) * | 2018-04-18 | 2021-04-23 | 浙江大学 | 一种诱导人羊膜上皮细胞向视网膜色素上皮细胞分化的方法及其应用 |
| CN112739208A (zh) * | 2018-09-21 | 2021-04-30 | 艾普斯丹医疗公司 | 人类多能成体干细胞 |
| GB201817385D0 (en) * | 2018-10-25 | 2018-12-12 | Plasticell Ltd | Therapy |
| CA3117459A1 (en) * | 2018-10-31 | 2020-05-07 | Mesoblast International Sarl | Expansion of hematopoietic stem cells |
| CN110468103B (zh) * | 2019-07-11 | 2023-03-31 | 中国医学科学院血液病医院(中国医学科学院血液学研究所) | 一种在体外维持造血干细胞自我更新能力的细胞因子组合 |
| CN110373387B (zh) * | 2019-08-01 | 2021-03-30 | 中国医学科学院血液病医院(中国医学科学院血液学研究所) | 组蛋白去乙酰化酶抑制剂在制备促进多能干细胞分化为造血干祖细胞的产品中的用途 |
| CN112980789B (zh) * | 2019-12-16 | 2024-11-29 | 深圳市海伦海生物科技有限公司 | 扩增造血干细胞的小分子化合物及其组合 |
| US20230027247A1 (en) * | 2019-12-16 | 2023-01-26 | Edigene (Guangzhou) Inc. | Small molecule compounds for amplifying hematopoietic stem cells, and combination thereof |
| US20220280574A1 (en) * | 2021-03-04 | 2022-09-08 | Therapeutic Solutions International, Inc. | Therapeutic monocytes for prevention of suicidal ideation |
| CN114045265A (zh) * | 2021-11-17 | 2022-02-15 | 深圳知因细胞生物科技有限公司 | 一种慢病毒体外转染人造血干细胞的试剂和试剂盒 |
| GB202303026D0 (en) | 2023-03-01 | 2023-04-12 | Plasticell Ltd | Therapy |
Family Cites Families (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2004046312A2 (en) * | 2002-11-15 | 2004-06-03 | The Board Of Trustees Of The University Of Illinois | Methods for in vitro expansion of hematopoietic stem cells |
| US7622108B2 (en) * | 2004-04-23 | 2009-11-24 | Bioe, Inc. | Multi-lineage progenitor cells |
| US7670596B2 (en) * | 2004-04-23 | 2010-03-02 | Bioe, Inc. | Multi-lineage progenitor cells |
| WO2006088867A2 (en) * | 2005-02-15 | 2006-08-24 | Medistem Laboratories, Incorporated | Method for expansion of stem cells |
| GB0503918D0 (en) * | 2005-02-25 | 2005-04-06 | Erasmus University | Cell |
| US9388382B2 (en) | 2005-10-05 | 2016-07-12 | The Board Of Trustees Of The University Of Illinois | Isolation of CD14 negative, CD45 positive and CD117 positive embryonic-like stem cells free of monocytes from human umbilical cord blood mononuclear cells |
| US20090252711A1 (en) * | 2006-05-11 | 2009-10-08 | Andrew Craig Boquest | Stem Cells And Methods Of Making And Using Stem Cells |
| US20090028834A1 (en) | 2007-07-27 | 2009-01-29 | Hal Siegel | Methods and compositions for stimulating the proliferation or differentiation of stem cells with substance P or an analog thereof |
| US20120301438A1 (en) * | 2009-09-14 | 2012-11-29 | The Johns Hopkins University | Reprogramming Blood Cells to Pluripotent and Multipotent Stem Cells |
| US9050315B2 (en) * | 2010-04-12 | 2015-06-09 | The Trustees Of The University Of Pennsylvania | In vivo and ex vivo expansion of hematopoietic stem cells with a targeted combination of clinically tested, FDA approved drugs |
| CN102465112A (zh) * | 2010-11-01 | 2012-05-23 | 张正前 | 人源脐带血造血干细胞体外高效扩增技术 |
| WO2013086029A1 (en) * | 2011-12-05 | 2013-06-13 | Primorigen Biosciences Inc. | Compositions and methods for differentiating pluripotent stem cells into primitive blood cells and uses thereof |
| JP6525963B2 (ja) | 2013-05-20 | 2019-06-05 | アイカーン スクール オブ メディシン アット マウント サイナイ | 血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 |
-
2014
- 2014-05-16 JP JP2016514978A patent/JP6525963B2/ja active Active
- 2014-05-16 CN CN201480034563.0A patent/CN105705021B/zh active Active
- 2014-05-16 CA CA2912688A patent/CA2912688C/en active Active
- 2014-05-16 US US14/892,772 patent/US11261429B2/en active Active
- 2014-05-16 WO PCT/US2014/038361 patent/WO2014189781A1/en not_active Ceased
- 2014-05-16 EP EP14801860.9A patent/EP2999343A4/en active Pending
- 2014-05-16 CN CN201911214820.XA patent/CN111019894A/zh active Pending
-
2019
- 2019-05-07 JP JP2019087708A patent/JP2019150040A/ja active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| CA2912688A1 (en) | 2014-11-27 |
| EP2999343A1 (en) | 2016-03-30 |
| WO2014189781A1 (en) | 2014-11-27 |
| CN111019894A (zh) | 2020-04-17 |
| US20160097036A1 (en) | 2016-04-07 |
| CN105705021B (zh) | 2020-01-03 |
| JP2019150040A (ja) | 2019-09-12 |
| CN105705021A (zh) | 2016-06-22 |
| JP2016524468A (ja) | 2016-08-18 |
| US11261429B2 (en) | 2022-03-01 |
| CA2912688C (en) | 2021-07-13 |
| EP2999343A4 (en) | 2016-12-28 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6525963B2 (ja) | 血液疾患を治療するための、濃縮されかつ増殖したヒト臍帯血幹細胞 | |
| Chaurasia et al. | Epigenetic reprogramming induces the expansion of cord blood stem cells | |
| US12497593B2 (en) | Generation of hematopoietic progenitor cells from human pluripotent stem cells | |
| EP2956538B1 (en) | Bioengineered liver constructs and methods relating thereto | |
| CN110050061A (zh) | 从具有MeCP2破坏的诱导多能干细胞生成成熟谱系 | |
| WO2005007799A2 (en) | Methods for ex-vivo expanding stem/progenitor cells | |
| JP2002527101A (ja) | ヒストンデアセチラーゼ阻害剤による造血幹細胞への遺伝子形質導入の改良及び自己再生の促進 | |
| AU2025271540A1 (en) | Expansion of hematopoietic stem cells | |
| Yang et al. | MiR-17 partly promotes hematopoietic cell expansion through augmenting HIF-1α in osteoblasts | |
| US20260103680A1 (en) | Pluripotent stem cell-derived megakaryocytes and platelets | |
| Deng et al. | Advances in hematopoietic stem cells ex vivo expansion associated with bone marrow niche | |
| Gross et al. | Prospects and challenges of reprogrammed cells in hematology and oncology | |
| EP4733383A1 (en) | Culture medium and blood cell clone, and method for producing same | |
| WO2026014176A1 (ja) | 骨髄間質オルガノイドの製造方法、骨髄間質オルガノイド、及びその利用 | |
| EP4644539A1 (en) | Method for regulating degree of cell differentiation | |
| HK40067001A (zh) | 扩增造血干细胞的方法及其组合物 | |
| Garcia | Defining the Role of SOX17 in Human Hematopoietic Development | |
| US20150258084A1 (en) | Stem cell compositions and methods of their use | |
| de Soure | Ex vivo expansion of human hematopoietic stem/progenitor cells from the umbilical cord blood for Cellular Therapy |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20170515 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20180314 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20180402 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20180628 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20181001 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20190304 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20190402 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20190507 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6525963 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |