KR20170008151A - 메틸말로닐-CoA 리덕테이즈 코딩 유전자를 함유하는 미생물 변이체 및 이의 용도 - Google Patents
메틸말로닐-CoA 리덕테이즈 코딩 유전자를 함유하는 미생물 변이체 및 이의 용도 Download PDFInfo
- Publication number
- KR20170008151A KR20170008151A KR1020160075640A KR20160075640A KR20170008151A KR 20170008151 A KR20170008151 A KR 20170008151A KR 1020160075640 A KR1020160075640 A KR 1020160075640A KR 20160075640 A KR20160075640 A KR 20160075640A KR 20170008151 A KR20170008151 A KR 20170008151A
- Authority
- KR
- South Korea
- Prior art keywords
- val
- gly
- leu
- ala
- ile
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images






Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/40—Preparation of oxygen-containing organic compounds containing a carboxyl group including Peroxycarboxylic acids
- C12P7/42—Hydroxy-carboxylic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y101/00—Oxidoreductases acting on the CH-OH group of donors (1.1)
- C12Y101/01—Oxidoreductases acting on the CH-OH group of donors (1.1) with NAD+ or NADP+ as acceptor (1.1.1)
- C12Y101/01031—3-Hydroxyisobutyrate dehydrogenase (1.1.1.31)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y102/00—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2)
- C12Y102/01—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2) with NAD+ or NADP+ as acceptor (1.2.1)
- C12Y102/01075—Malonyl CoA reductase (malonate semialdehyde-forming)(1.2.1.75)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y504/00—Intramolecular transferases (5.4)
- C12Y504/99—Intramolecular transferases (5.4) transferring other groups (5.4.99)
- C12Y504/99001—Methylaspartate mutase (5.4.99.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y504/00—Intramolecular transferases (5.4)
- C12Y504/99—Intramolecular transferases (5.4) transferring other groups (5.4.99)
- C12Y504/99002—Methylmalonyl-CoA mutase (5.4.99.2)
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Health & Medical Sciences (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Molecular Biology (AREA)
- Plant Pathology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description
도 2는 MMCR 후보 유전자들을 PCR을 이용하여 복제한 결과를 나타낸 것이다.
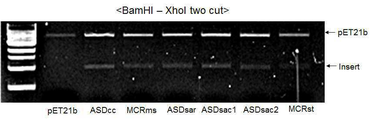
도 3은 MMCR 후보 유전자들을 ligation하여 벡터에 삽입한 결과를 나타낸 것이다.
도 4는 MMCR 후보 유전자들이 형질전환된 균주를 배양하여 각 효소의 발현량을 확인한 결과를 나타낸 것이다.
도 5는 메틸말로닐-CoA (methylmalonyl-CoA)를 반응 기질로 하였을 때 MMCR 후보의 역가 측정 결과를 나타낸 것이다.
도 6은 메틸말로닐-CoA를 반응 기질로 하였을 때 반응산물을 MS로 분석한 결과를 나타낸 것이다.
도 7은 3-HIBA 생산 균주 배양액을 HPLC로 분석하여 3-HIBA 생산을 확인한 결과를 나타낸 것이다.




Claims (8)
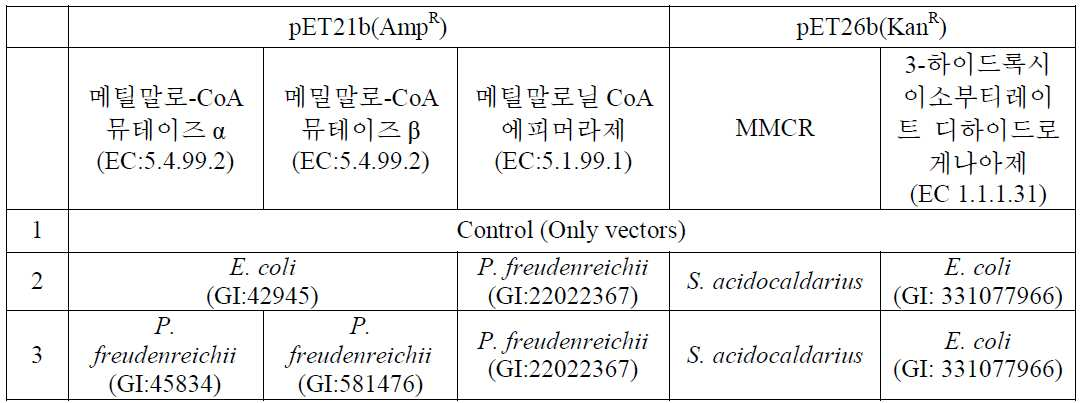
- 탄소원으로부터 숙시닐-CoA 생성능을 가지는 미생물에서 다음의 효소를 코딩하는 유전자를 함유하고 3-HIBA (3-hydroxyisobutyric acid) 생성능을 가지는 미생물 변이체에 있어서,
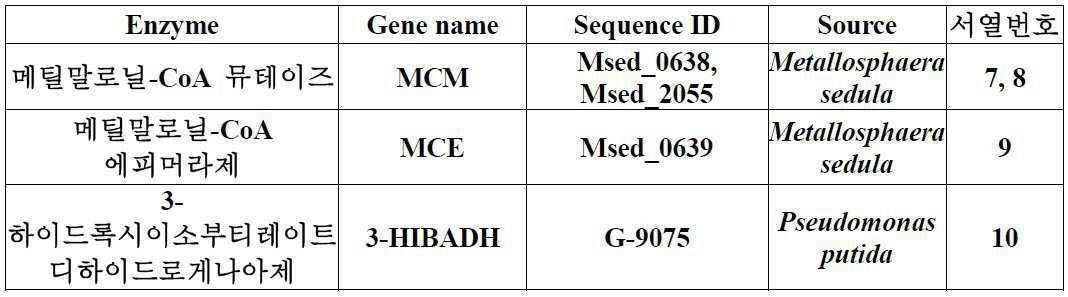
(i) 메틸말로닐-CoA 뮤테이즈 (methylmalonyl-CoA mutase);
(ii) 메틸말로닐-CoA 에피머라제 (methylmalonyl-CoA epimerase);
(iii) 메틸말로닐-CoA 리덕테이즈 (methylmalonyl-CoA reductase, MMCR); 및
(iv) 3-하이드록시이소부티레이트 디하이드로게나아제 (3-hydroxyisobutyrate dehydrogenase),
상기 (iii)의 효소는 말로닐-CoA 리덕테이즈 (malonyl-CoA reductase, MCR) 활성을 가진 효소 중 메틸말로닐-CoA 리덕테이즈 (methylmalonyl-CoA reductase, MMCR) 활성을 나타내는 것임을 특징으로 하는 미생물 변이체.
- 제1항에 있어서,
상기 (iii)의 효소는 말로닐-CoA 리덕테이즈 활성을 가진 효소 중 메틸말로닐-CoA를 메틸말로네이트 세미알데하이드로 전환하는 단일 관능성 (monofunctional)의 메틸말로닐-CoA 리덕테이즈 활성을 나타내는 것임을 특징으로 하는 미생물 변이체.
- 제1항에 있어서,
상기 말로닐-CoA 리덕테이즈 활성을 가진 효소 중 메틸말로닐-CoA 리덕테이즈 활성을 나타내는 효소는 아키아 (Archaea)계 유래인 것을 특징으로 하는 미생물 변이체.
- 제3항에 있어서,
상기 (iii)의 효소는 Candidatus Caldiarchaeum subterraneum, Sulfolobales archaeon Acd1 및 Sulfolobus acidocaldarius Ron12/I로 이루어진 군에서 선택된 아키아계 유래인 것을 특징으로 하는 미생물 변이체.
- 제1항에 있어서,
상기 (iii)의 효소는 서열번호 23 (IKVLGDAYDAKTVKEVTRILSEVKRNVPGTM-DELTLSATTHRIA)으로 표시되는 서열과 60% 이상의 상동성 또는 80% 이상의 유사성을 나타내는 서열을 가지는 것을 특징으로 하는 미생물 변이체.
- 제1항에 있어서,
상기 (iii)의 효소는 서열번호 3 내지 5로 이루어진 군에서 선택되는 서열 중 어느 하나와 60% 이상의 상동성을 나타내는 서열을 가지는 것을 특징으로 하는 미생물 변이체.
- 제1항에 있어서,
상기 (iii)의 효소는 서열번호 3 내지 5로 이루어진 군에서 선택되는 서열 중 어느 하나의 서열을 가지는 것을 특징으로 하는 미생물 변이체.
- 제1항 내지 제7항 중 어느 한 항에 따른 미생물 변이체를 배양하여 3-HIBA를 생성하는 단계; 및 상기 생성된 3-HIBA를 회수하는 단계를 포함하는 3-HIBA의 제조방법.
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| DE102016212497.8A DE102016212497B4 (de) | 2015-07-13 | 2016-07-08 | Mutanter Mikroorganismus, der ein Gen umfasst, das Methylmalonyl-CoA-Reductase codiert, und seine Verwendung |
| US15/207,953 US20170016033A1 (en) | 2015-07-13 | 2016-07-12 | Mutant Microorganism Comprising Gene Encoding Methylmalonyl-CoA Reductase and Use Thereof |
| CN201610552723.1A CN106350474B (zh) | 2015-07-13 | 2016-07-13 | 包含编码甲基丙二酰-CoA还原酶的基因的突变微生物及其用途 |
| KR1020240055415A KR20240060569A (ko) | 2015-07-13 | 2024-04-25 | 메틸말로닐-CoA 리덕테이즈 코딩 유전자를 함유하는 미생물 변이체 및 이의 용도 |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020150099352 | 2015-07-13 | ||
| KR20150099352 | 2015-07-13 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020240055415A Division KR20240060569A (ko) | 2015-07-13 | 2024-04-25 | 메틸말로닐-CoA 리덕테이즈 코딩 유전자를 함유하는 미생물 변이체 및 이의 용도 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20170008151A true KR20170008151A (ko) | 2017-01-23 |
Family
ID=57989968
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020160075640A Ceased KR20170008151A (ko) | 2015-07-13 | 2016-06-17 | 메틸말로닐-CoA 리덕테이즈 코딩 유전자를 함유하는 미생물 변이체 및 이의 용도 |
Country Status (1)
| Country | Link |
|---|---|
| KR (1) | KR20170008151A (ko) |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US11680270B2 (en) | 2019-10-08 | 2023-06-20 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast with inhibited lactate metabolism and alcohol production and method of producing lactic acid using the same |
| US11898173B2 (en) | 2020-06-24 | 2024-02-13 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast having improved lactic-acid-producing ability |
| US12084665B2 (en) | 2018-10-08 | 2024-09-10 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast in which alcohol production is inhibited and method for producing lactic acid by using same |
| US12157902B2 (en) | 2020-04-17 | 2024-12-03 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast with suppressed glycerol production and method of producing lactic acid using the same |
-
2016
- 2016-06-17 KR KR1020160075640A patent/KR20170008151A/ko not_active Ceased
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US12084665B2 (en) | 2018-10-08 | 2024-09-10 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast in which alcohol production is inhibited and method for producing lactic acid by using same |
| US11680270B2 (en) | 2019-10-08 | 2023-06-20 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast with inhibited lactate metabolism and alcohol production and method of producing lactic acid using the same |
| US12157902B2 (en) | 2020-04-17 | 2024-12-03 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast with suppressed glycerol production and method of producing lactic acid using the same |
| US11898173B2 (en) | 2020-06-24 | 2024-02-13 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast having improved lactic-acid-producing ability |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US12071645B2 (en) | Enzymes and applications thereof | |
| US8124388B2 (en) | Production of 3-hydroxypropionic acid using beta-alanine/pyruvate aminotransferase | |
| US8043841B2 (en) | Nucleic acid molecules encoding alanine 2,3-aminomutases | |
| JP2014506466A (ja) | イソ酪酸を製造するための細胞及び方法 | |
| KR20170008151A (ko) | 메틸말로닐-CoA 리덕테이즈 코딩 유전자를 함유하는 미생물 변이체 및 이의 용도 | |
| Bäumchen et al. | D‐mannitol production by resting state whole cell biotransformation of D‐fructose by heterologous mannitol and formate dehydrogenase gene expression in Bacillus megaterium | |
| CN112908417B (zh) | 功能序列和结构模拟相结合的基因挖掘方法、nadh偏好型草铵膦脱氢酶突变体及应用 | |
| JP5142268B2 (ja) | 改良型没食子酸合成酵素および没食子酸の製造法 | |
| Yamamoto et al. | Synthesis of ethyl (S)-4-chloro-3-hydroxybutanoate using fabG-homologues | |
| KR20240060569A (ko) | 메틸말로닐-CoA 리덕테이즈 코딩 유전자를 함유하는 미생물 변이체 및 이의 용도 | |
| CN117903007B (zh) | 一种亚精胺生物合成途径关键酶 | |
| KR20130091080A (ko) | meso-2,3-부탄다이올 제조방법 | |
| JP5381113B2 (ja) | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 | |
| KR102028161B1 (ko) | 형질전환 미생물을 이용한 2,3-부탄디올 생산방법 | |
| JP6034332B2 (ja) | 組換え微生物及び当該組換え微生物を用いた物質製造方法 | |
| KR102424603B1 (ko) | 알코올 탈수소효소 변이체, 및 이를 이용한 알데히드 화합물의 생산 방법 | |
| KR20130058236A (ko) | meso-2,3-부탄다이올 제조방법 | |
| JPWO2006043555A1 (ja) | 生体高分子生成のための還元酵素変異体 | |
| US20020187537A1 (en) | Recombinantly producing levodione reductase |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PA0109 | Patent application |
Patent event code: PA01091R01D Comment text: Patent Application Patent event date: 20160617 |
|
| PG1501 | Laying open of application | ||
| PA0201 | Request for examination |
Patent event code: PA02012R01D Patent event date: 20210401 Comment text: Request for Examination of Application Patent event code: PA02011R01I Patent event date: 20160617 Comment text: Patent Application |
|
| E902 | Notification of reason for refusal | ||
| PE0902 | Notice of grounds for rejection |
Comment text: Notification of reason for refusal Patent event date: 20230222 Patent event code: PE09021S01D |
|
| E902 | Notification of reason for refusal | ||
| PE0902 | Notice of grounds for rejection |
Comment text: Notification of reason for refusal Patent event date: 20230823 Patent event code: PE09021S01D |
|
| E90F | Notification of reason for final refusal | ||
| PE0902 | Notice of grounds for rejection |
Comment text: Final Notice of Reason for Refusal Patent event date: 20240226 Patent event code: PE09021S02D |
|
| E601 | Decision to refuse application | ||
| PE0601 | Decision on rejection of patent |
Patent event date: 20240723 Comment text: Decision to Refuse Application Patent event code: PE06012S01D |