이하, 본 발명을 실시하기 위한 적합한 형태에 대해서 도면을 참조하면서 설명한다. 또한, 이하에 설명하는 실시형태는, 본 발명의 대표적인 실시형태의 일례를 나타낸 것이고, 이에 따라 본 발명의 범위가 좁게 해석되는 경우는 없다.
(정의) 본 명세서 중, 「암」 과「종양」 은 동일한 의미로 사용되며, 각각, 특별히 한정되지 않는 한, 고형암, 비고형암 또는 그들 양방을 의도하는 경우에 사용된다.
본 명세서 중, 「유전자」 라고 하는 말에는, DNA 뿐만 아니라 그 mRNA, cDNA 및 그 cRNA 도 포함된다.
본 명세서 중, 「폴리뉴클레오티드」 또는 「뉴클레오티드」 는, 핵산과 동일한 의미로 사용하고 있고, DNA, RNA, 프로브, 올리고 뉴클레오티드, 및 프라이머도 포함된다.
본 명세서 중, 「폴리펩티드」 와「단백질」 은 구별하지 않고 사용하고 있다.
본 명세서 중, 「세포」 에는, 동물 개체 내의 세포, 배양 세포도 포함하고 있다.
본 명세서 중, 「CD147」 은, CD147 단백질과 동일한 의미로 사용하고 있다.
본 명세서 중, 「항체의 기능성 단편」 이란, 「항체의 항원 결합 단편」 이라고도 불리며, 항원과의 결합 활성을 갖는 항체의 부분 단편을 의미하고 있고, Fab, F(ab')2, Fv, scFv, 디아바디 (diabody), 선상 (線狀) 항체 및 항체 단편으로 형성된 다특이성 항체 등을 포함한다. 또, F(ab')2 를 환원 조건하에서 처리한 항체의 가변 영역의 1 가의 단편인 Fab' 도 항체의 항원 결합 단편에 포함된다. 단, 항원과의 결합능을 갖고 있는 한 이들 분자에 한정되지 않는다. 또, 이들 항원 결합 단편에는, 항체 단백질의 전체 길이 분자를 적당한 효소로 처리한 것 뿐만 아니라, 유전자 공학적으로 개편된 항체 유전자를 사용하여 적당한 숙주 세포에 있어서 산생된 단백질도 포함된다.
본 명세서 중, 「이펙터 활성」 이란, 항체 의존성 세포 상해 (Antibody-Dependent-Cellular-Cytotoxicity, 이하, ADCC) 활성, 보체 의존성 세포 상해 (Complement-dependent cytotoxicity, 이하, CDC) 활성, 또는 항체 의존성 세포 탐식 (Antibody-dependent cellular phagocytosis, 이하, ADCP) 활성 중 어느 하나 이상을 말한다.
본 명세서 중, 「이펙터 기능」 이란, 각 「이펙터 활성」 이 발휘되는 것을 말한다.
항체 의존성 세포 상해 (ADCC) 활성은, 이펙터 활성을 갖는 면역 세포, 항체 및 51Cr 표지한 표적 세포를 접촉시킨 경우에 생기는 세포사를 측정하는 51Cr 릴리스 어세이 방법으로 측정할 수 있다. 본 발명의 인간 CD147 항체의 ADCC 활성은, 다음과 같이 측정된다. 평가하는 인간 CD147 항체의 ADCC 활성에 대해, 이펙터 세포로서 인간의 말초혈 단핵구 (PBMC), ADCC 표적 세포로서 CD147 양성의 인간 암 세포주 (예를 들어, 췌장암주 MIA PaCa-2) 를 사용한다. 방사선 동위체 51Cr 로 표지한 암 세포와 평가 대상의 항체를 0.5 또는, 5 ㎍/㎖ 의 농도로 4 ℃, 30 분간 처리한 후, 인간의 말초혈로부터 분리한 PBMC 를 암 세포의 20 배의 비율로 첨가하고, 4 시간, 37 도, 5 % CO2 존재하에서 배양한다. 상청 중에 방출된 51Cr 을 TopCount NXT v2.53 을 사용하여 측정하고 총 방출 (Total release) 값을 얻는다. 51Cr 로 표지한 암 세포를 트리톤-100 으로 처리하여 방출된 51Cr 의 측정값을 최대 방출 (Maximum release) 값, PBMC 를 첨가하지 않는 항체 처리 세포로부터 처리하여 방출된 51Cr 의 측정값을 자연 방출 (Spontaneous release) 값으로 하여, 하기의 식으로부터, % 특이적 방출 (% specific release) 을 산출한다. 음성 대조 샘플로서, 인간 IgG (hIgG, ChromPure Human IgG, Jackson I㎜unoResearch Laboratories 사, Cat. 009-000-003) 를 사용한다. 측정은 삼중으로 실시하고, 평균값, 표준 편차를 산출한다.
% 특이적 방출 = (총 방출 ― 자연 방출) / 최대 방출
보체 의존성 세포 상해 (CDC) 활성은, 혈액에 포함되는 보체, 항체 및 표적 세포를 접촉시켰을 경우에 생기는 세포사를 측정함으로써 평가를 할 수 있다. 본 발명의 인간 CD147 항체의 CDC 활성은, 다음과 같이 측정된다. 평가하는 인간 CD147 항체에 의한 보체 의존적인 살세포 활성 (CDC 활성) 을 표적 세포로 하여 인간 췌장주 MIA PaCa-2 를 사용하여 평가한다. 보체로서 시판되는 토끼 보체 (Low Tox-M Rabbit Complement, CEDARLANE LABORATORIES LIMITED, Cat. CL3051) 를 사용한다. CDC 활성 음성의 대조 항체로서 인간 IgG (hIgG, ChromPure Human IgG, Jackson I㎜unoResearch Laboratories 사, Cat. 009-000-003) 를 사용한다. 평가하는 항체 및 음성의 대상 항체를, 각각, 0, 0.1, 1 또는, 10 ㎍/㎖ 의 농도로 1 시간, 4 ℃ 에서 처리한 후, 토끼 보체를 종농도 7.5 % 가 되도록 첨가하고, 37 ℃, 5 % CO2 존재하에서, 3 시간 가온 후, 생 세포에 포함되는 세포내 ATP 를 트리톤-100Titer-Glo Lumimescent 트리톤-100 Viability Assay (Promega 사, Cat. G7572) 를 사용하여 측정하였다. 트리톤-100Titer-Glo Lumimescent 트리톤-100 Viability Assay 를 사용하여 얻어지는 발광 시그널에 대해, EnVision 2104 Multilabel Reader (Perkin Elmer 사) 를 사용하여 정량한다. 측정은 삼중으로 실시하고, 평균값과 표준 편차를 산출한다. 무처리의 세포로부터 얻어진 발광 시그널을 100 % 로 하여, 항체와 보체 의존적으로 감소한 발광 시그널을 CDC 활성으로 한다.
항체 의존성 세포 탐식 (ADCP) 활성은, 탐식 작용을 갖는 면역 세포, 항체 및 표적 세포를 접촉시킴으로써 일어나는 탐식을 2 중 형광 표지법으로 측정할 수 있다.
본 발명의 인간 CD147 항체의 ADCP 활성은, 다음과 같이 측정된다. 인간 IgG 항체는 마우스의 Fcγ 수용체와의 상호 작용을 통해서, 항체 의존적인 단구, 매크로파지에 의한 탐식 작용 (ADCP) 을 유도함으로써, 암 세포에 대한 살세포 활성을 나타내는 것이 보고되어 있다 (Overdijk et al., Journal of I㎜unology, 1-9, 2012). 본 발명의 인간 키메라 항체의 ADCP 활성에 대해, 이펙터 세포로서 RAW264.7 (ATCC, TIB-71), ADCP 표적 세포로서 인간 췌장주 PANC-1 또는 MIA PaCa-2 를 사용하여 평가한다. PKH67 Green Fluorescent Cell Linker Mini Kit for General Cell Membrane Labeling (SIGMA, Cat. MINI67-1KIT) 으로 표지한 ADCP 표적 세포라고 평가하는 항체와, 20 ㎍/㎖ 의 농도의 평가하는 항체로 4 ℃, 1 시간 처리한 후, PKH26 Red Fluorescent Cell Linker Kit for General Cell Membrane Labeling (SIGMA, Cat. PKH26GL-1KT) 으로 표지한 RAW264.7 세포를 ADCP 표적 세포의 5 배 첨가하고, 3 시간, 37 ℃, 5 % CO2 존재하에서 가온한다. 플로우 사이토미터 (BD 사, CantoII) 를 사용하여, 탐식 작용에 의해 PKH67 시그널 양성으로 이행한 PKH26 양성 세포의 비율을 측정한다. 음성 대조 샘플로서, 인간 IgG (hIgG, ChromPure Human IgG, Jackson I㎜unoResearch Laboratories 사, Cat. 009-000-003) 를 처리한 샘플에 대해 동일하게 측정을 실시한다. 측정은 삼중으로 실시하고, 평균값, 표준 편차를 산출한다.
본 명세서 중, 「이펙터 활성을 실질적으로 갖지 않는다」 또는 「이펙터 활성이 저하 또는 결실하였다」 라는 것은, 당해 항체가, ADCC 활성, CDC 활성 또는 ADCP 활성 중의 적어도 1 개가, 활성을 나타내지 않거나, 혹은, 그들의 기능이 충분하게는 발휘되지 않을 정도로 낮은 정도의 활성인 것을 말한다. 「이펙터 활성을 실질적으로 갖지 않는다」 또는 「이펙터 활성이 저하 또는 결실하였다」 라는 것은, 예를 들어, 상기의 활성 평가 방법에 있어서, 평가하는 항체의 활성이 네거티브 컨트롤과 동일한 정도의 활성인 것을 말한다.
본 명세서 중, 「ADCC 활성이 저하 또는 결실하였다」 라는 것은, 평가하는 항체가, ADCC 활성을 나타내지 않거나, 혹은, 그들의 기능이 충분하게는 발휘되지 않을 정도로 낮은 정도의 활성인 것을 말한다. 「ADCC 활성이 저하 또는 결실하였다」 라는 것은, 예를 들어, 상기의 활성 평가 방법에 있어서, 평가하는 항체의 활성이 네거티브 컨트롤과 동일한 정도의 활성인 것을 말한다.
본 명세서 중, 「CDC 활성이 저하 또는 결실하였다」 라는 것은, 평가하는 항체가, CDC 활성을 나타내지 않거나, 혹은, 그들의 기능이 충분하게는 발휘되지 않을 정도로 낮은 정도의 활성인 것을 말한다. 「CDC 활성이 저하 또는 결실하였다」 라는 것은, 예를 들어, 상기의 활성 평가 방법에 있어서, 평가하는 항체의 활성이 네거티브 컨트롤과 동일한 정도의 활성인 것을 말한다.
본 명세서 중, 「ADCP 활성이 저하 또는 결실하였다」 라는 것은, 평가하는 항체가, ADCP 활성을 나타내지 않거나, 혹은, 그들의 기능이 충분하게는 발휘되지 않을 정도로 낮은 정도의 활성인 것을 말한다. 「ADCP 활성이 저하 또는 결실하였다」 라는 것은, 예를 들어, 상기의 활성 평가 방법에 있어서, 평가하는 항체의 활성이 네거티브 컨트롤과 동일한 정도의 활성인 것을 말한다.
「ADCC 활성이 저하 또는 결실하였다」, 「CDC 활성이 저하 또는 결실하였다」 또는 「ADCP 활성이 저하 또는 결실하였다」 라는 것은, 예를 들어, 각각, 상기의 활성 평가 방법에 있어서, 평가하는 항체의 활성이 네거티브 컨트롤과 동일한 정도의 활성인 것을 말한다.
본 명세서 중, 「CD147 을 개재한 시그널 전달을 활성화한다」, 「CD147 을 개재한 관련 시그널 분자의 활성화」, 「CD147 의 활성화」 또는 「CD147 을 활성화」 란, CD147 을 개재한 세포 시그널 전달계의 활성화를 말하며, CD147 의 하류의 관련 시그널 분자 중의 적어도 어느 것을 활성화시키는 것을 말한다. CD147 을 개재한 시그널 전달의 활성화란, CD147 시그널의 하류에 있는 유전자의 발현이 항진 또는 저하되거나, 단백질의 발현이 항진 또는 저하되거나, 혹은, 단백질의 인산화가 항진 또는 저하되거나 하는 것을 말한다. CD147 의 하류의 관련 시그널 분자로는, 예를 들어, FAK, MEK, Erk, JAK/STAT, AKT 또는 MAP 키나아제 (MAPK), 혹은 이들의 더 하류의 시그널 분자의 활성화를 들 수 있다. MAPK 로는, 예를 들어, ERK1/2, JNK 또는 p38MAPK 를 들 수 있고, 보다 바람직하게는 p38MAPK 이다. MAPK 의 더 하류의 시그널 분자로는, 예를 들어, HSP27, cxcl8 또는 SMAD (예를 들어, SMAD2, SMAD3 및/또는 SMAD4) 를 들 수 있다. 「CD147 의 활성화」 로는, 예를 들어, p38MAPK 의 mRNA 발현량의 증가, p38MAPK 의 단백질 발현량의 증가, p38MAPK 의 인산화, HSP27 의 인산화 (예를 들어, HSP27 의 Ser82 의 인산화 또는 HSP27 의 Ser15 의 인산화), cxcl8 mRNA 발현량의 증가, cxcl8 단백질 발현량의 증가, SMAD 시그널 활성화를 개재한 rhoB mRNA 발현량의 증가 또는 rhoB 단백질 발현량의 증가, 혹은, KLF5 mRNA 의 저하 또는 KLF5 단백질 발현량의 저하를 들 수 있다.
본 명세서 중, 「에피토프」 란, 특정한 항 CD147 항체가 결합하는 CD147 의 부분 펩티드 또는 부분 입체 구조를 의미한다. 상기의 CD147 의 부분 펩티드인 에피토프는 면역 어세이법 등 당업자에게는 잘 알려져 있는 방법에 의해 결정할 수 있다. 먼저, 항원의 다양한 부분 구조를 제조한다. 부분 구조의 제조에 있어서는, 공지된 올리고 뉴클레오티드 합성 기술을 사용할 수 있다. 예를 들어, CD147 의 C 말단 또는 N 말단으로부터 적당한 길이로 순차 짧게 한 일련의 폴리펩티드를 당업자에게 주지의 유전자 조작 기술을 사용하여 제조한 후, 그것들에 대한 항체의 반응성을 검토하고, 대략적인 인식 부위를 결정한 후에, 더욱 짧은 펩티드를 합성하여 그들의 펩티드와의 반응성을 검토함으로써, 에피토프를 결정할 수 있다. 또, 복수의 세포외 도메인으로 이루어지는 막 단백질에 결합하는 항체가, 복수의 도메인으로 이루어지는 입체 구조를 에피토프로 하고 있는 경우에는, 특정한 세포외 도메인의 아미노산 서열을 개변함으로써, 입체 구조를 개변함으로써 어느 도메인과 결합할지를 결정할 수 있다. 특정한 항체가 결합하는 항원의 부분 입체 구조인 에피토프는, X 선 구조 해석에 의해 상기의 항체와 인접하는 항원의 아미노산 잔기를 특정함으로써도 결정할 수 있다.
제 1 항체가 결합하는 부분 펩티드 또는 부분 입체 구조에 제 2 항체가 결합하면, 제 1 항체와 제 2 항체가 공통의 에피토프를 갖는 것으로 판정할 수 있다. 또, 제 1 항체의 항원에 대한 결합에 제 2 항체가 교차 경합하는 (즉, 제 2 항체가 제 1 항체와 항원의 결합을 방해한다) 것을 확인함으로써, 구체적인 에피토프의 서열 또는 구조가 결정되어 있지 않아도, 제 1 항체와 제 2 항체의 동일한 에피토프에 결합하는 것으로 판정할 수 있다. 제 1 항체와 제 2 항체가 동일한 에피토프에 결합하고, 또한 제 1 항체가 항종양 활성 등의 특수한 효과를 갖는 경우, 제 2 항체도 동일한 활성을 갖는 것을 기대할 수 있다.
항체 분자의 중쇄 및 경쇄에는 각각 3 개 지점의 상보성 결정 영역 (CDR : Complementarity determining region) 이 있는 것이 알려져 있다. 상보성 결정 영역은, 초가변 영역 (hypervariable domain) 이라고도 불리며, 항체의 중쇄 및 경쇄의 가변 영역 내에 있어, 1 차 구조의 변이성이 특히 높은 부위이며, 중쇄 및 경쇄의 폴리펩티드 사슬의 1 차 구조 상에 있어서, 각각 3 개 지점으로 분리되어 있다. 본 명세서 중에 있어서는, 항체의 상보성 결정 영역에 대해, 중쇄의 상보성 결정 영역을 중쇄 아미노산 서열의 아미노 말단측으로부터 CDRH1, CDRH2, CDRH3 이라고 표기하고, 경쇄의 상보성 결정 영역을 경쇄 아미노산 서열의 아미노 말단측으로부터 CDRL1, CDRL2, CDRL3 이라고 표기한다. 이들 부위는 입체 구조 상에서 상호 근접하고, 결합하는 항원에 대한 특이성을 결정하고 있다.
본 명세서에 있어서, 「1 내지 수 개」 및 「1 혹은 수 개」 라는 기재가 있는 경우의 「수 개」 란, 2 내지 10 개를 나타내고 있다. 바람직하게는 10 개 이하, 보다 바람직하게는 5 혹은 6 개 이하, 보다 더 바람직하게는 2 혹은 3 개이다.
(CD147)
CD147 은, 이뮤노글로불린형 도메인을 2 ∼ 3 갖는 1 회 막관통 단백질이며, CD147 끼리의 상호 작용, CD44, 인테그린 패밀리 분자, CD98, VEGFR, CypA/B, MCT1/3/4 와 같은 증식, 침윤, 염증에 관여하는 세포외나 세포막 표면의 분자와 상호 작용함으로써, 하류 시그널 관련 분자, FAK, MEK, Erk, JAK/STAT, AKT, MAPK 패밀리 분자를 활성화시키고, MMP 를 비롯한 프로테아제 산생, 암의 증식, 전이, 침윤을 촉진하는 것이 알려져 있다.
인간 CD147 은, 3 개의 베어리언트가 알려져 있다. 베어리언트 1 은, 망막 특이적으로 발현하고 3 개의 이뮤노글로불린형 도메인 (각각, 본 명세서 중, D0, D1 및 D2 라고 하는 경우가 있다.) 을 갖는 1 회 막관통 단백질이고, 베어리언트 2 는, T 세포나 다양한 정상 세포에서 발현하고, 또한, 다양한 암 조직에서 발현 증가가 보고되어 있는 2 개의 이뮤노글로불린형 도메인 (D1, D2) 을 갖는 1 회 막관통 단백질이며, 베어리언트 3 은, 1 개의 이뮤노글로불린형 도메인을 갖는 1 회 막관통 단백질이다.
인간 CD147 의 베어리언트 1 의 아미노산 서열 및 뉴클레오티드 서열은, GenBank 액세션 번호 NP_001719.2, NM_001728.3 을 참조함으로써 입수 가능하고, 본 명세서 중, 아미노산 서열은 서열 번호 1, 뉴클레오티드 서열은 서열 번호 2 로서도 개시되어 있다. 베어리언트 1 의 3 개의 이뮤노글로불린형 도메인은, 서열 번호 1 로 말하면, 각각, 아미노산 번호 22 ∼ 138 (D0), 아미노산 번호 140 ∼ 218 (D1) 및 아미노산 번호 223 ∼ 323 (D2) 이다 (Redzic, J., J. Mol. Biol., 2011, 68-82) (Grass et al., Biosol. Rep, 2016, 1-16). 또, 베어리언트 1 의 막관통 영역은, 서열 번호 1 로 말하면, 아미노산 번호 324 ∼ 344 이다.
인간 CD147 의 베어리언트 2 의 아미노산 서열 및 뉴클레오티드 서열은, GenBank 액세션 번호 NP_940991.1, NM_198589.2 를 참조함으로써 입수 가능하고, 본 명세서 중, 아미노산 서열은 서열 번호 3, 뉴클레오티드 서열은 서열 번호 4 로서도 개시되어 있다. 베어리언트 2 의 2 개의 이뮤노글로불린형 도메인 (D1, D2) 은, 서열 번호 3 으로 말하면, 각각, 아미노산 번호 24 ∼ 102 (D1) 및 아미노산 번호 107 ∼ 207 (D2) 이다. 또, 베어리언트 2 의 막관통 영역은, 서열 번호 3 으로 말하면, 아미노산 번호 208 ∼ 228 이다 (Grass et al., Biosol. Rep, 2016, 1-16).
인간 CD147 의 베어리언트 3 의 아미노산 서열 및 뉴클레오티드 서열은, GenBank 액세션번호 NP_940992.1, NM_198590.2 를 참조함으로써 입수 가능하다. 또, 인간 CD147 유전자는, 시판원으로부터 입수하는 것도 가능하다.
게잡이 원숭이 CD147 (본 명세서 중, 원숭이 CD147 이라고 기재하는 경우도 있다.) 의 아미노산 서열 및 뉴클레오티드 서열은, GenBank 액세션 번호 XP_005587354.1, XM_005587297.1 을 참조함으로써 입수 가능하다. 또, 원숭이 CD147 유전자는, 시판원으로부터 입수하는 것도 가능하다. 마우스 CD147 의 아미노산 서열 및 뉴클레오티드 서열은, GenBank 액세션 번호 NP_001070652.1, NM_001077184.1 을 참조함으로써 입수 가능하다. 또, 마우스 CD147 유전자는, 시판원으로부터 입수하는 것도 가능하다.
본 발명에서 사용하는 CD147 은, in vitro 에서 합성하거나, 혹은 유전자 조작에 의해 숙주 세포에 산생시킴으로써 얻을 수 있다. 구체적으로는, CD147 cDNA 를 발현 가능한 벡터에 짜넣은 후, 전사와 번역에 필요한 효소, 기질 및 에너지 물질을 포함하는 용액 중에서 합성하거나, 혹은 다른 원핵 생물, 또는 진핵 생물의 숙주 세포를 형질 전환시킴으로써 CD147 을 발현시킴으로써, 그 단백질을 얻을 수 있다.
CD147 의 cDNA 는, 예를 들어, CD147 의 cDNA 를 발현하고 있는 cDNA 라이브러리를 주형 (鑄型) 으로 하여, CD147 의 cDNA 를 특이적으로 증폭하는 프라이머를 사용하여 폴리머라아제 연쇄 반응 (이하 「PCR」 이라고 한다) (Saiki, R. K., et al., Science, (1988) 239, 487-49) 을 실시하는, 이른바 PCR 법에 의해 취득할 수 있다.
또한, 인간, 원숭이 또는 마우스의 CD147 을 코드하는 뉴클레오티드 서열과 상보적인 뉴클레오티드 서열로 이루어지는 폴리뉴클레오티드와 스트린젠트한 조건으로 하이브리다이즈 하고, 또한, CD147 과 동등한 생물 활성을 갖는 단백질을 코드하는 폴리뉴클레오티드도 CD147 의 cDNA 에 포함된다. 또한, 인간, 원숭이 혹은 마우스 CD147 유전자좌로부터 전사되는 스플라이싱 베어리언트 또는 이것에 스트린젠트한 조건으로 하이브리다이즈 하는 폴리뉴클레오티드로서, 또한, CD147 과 동등한 생물 활성을 갖는 단백질을 코드하는 폴리뉴클레오티드도 CD147 의 cDNA 에 포함된다.
또, 인간, 원숭이 또는 마우스의 CD147 의 아미노산 서열, 또는 이들 서열로부터 시그널 서열이 제외된 아미노산 서열에 있어서, 1 혹은 수 개의 아미노산이 치환, 결실, 또는 부가된 아미노산 서열로 이루어지고, CD147 과 동등한 생물 활성을 갖는 단백질도 CD147 에 포함된다. 또한, 인간, 원숭이 혹은 마우스 CD147 유전자좌로부터 전사되는 스플라이싱 베어리언트에 코드되는 아미노산 서열 또는 그 아미노산 서열에 있어서, 1 혹은 수 개의 아미노산이 치환, 결실, 또는 부가된 아미노산 서열로 이루어지고, 또한, CD147 과 동등한 생물 활성을 갖는 단백질도 CD147 에 포함된다.
(항 CD147 항체의 제조)
본 발명의 CD147 에 대한 항체는, 비인간 동물을 목적 항원으로 면역하고, 면역 성립 후의 동물로부터 림프액, 림프 조직, 혈구 시료 또는 골수 유래의 세포를 채취하고, 공지된 방법 (예를 들어, Kohler and Milstein, Nature (1975) 256, p. 495-497, Kennet, R. ed., Monoclonal Antibodies, p. 365-367, Plenum Press, N. Y. (1980)) 에 따라서, CD147 에 대한 항체를 산생하는 항체 산생 세포와 미엘로마 세포를 융합시킴으로써 하이브리도마를 수립하고, 모노클로날 항체를 얻을 수 있다. 이와 같은 방법의 구체적인 예는, WO2009/48072 (2009년 4월 16 일 공개) 및 WO2010/117011 (2010년 10월 14 일 공개) 에 기재되어 있다. 이와 같이 하여 얻어진 모노클로날 항체의 예로는, 예를 들어, LN22R8, 2P10F2, 래트_CD147_#84, 래트_CD147_#101, 래트_CD147_#110 또는 래트_CD147_#131 을 들 수 있다. 그러나, 모노클로날 항체를 취득하는 방법은, 이미 확립된 분야에 해당하고, 상기의 구체예에 한정되는 것은 아니다.
본 발명의 항체에는, 상기 CD147 에 대한 모노클로날 항체에 더하여, 인간에 대한 이종 항원성을 저하시키는 것 등을 목적으로 하여 인위적으로 개변한 유전자 조작형 항체, 예를 들어, 키메라 (Chimeric) 항체, 인간화 (Humanized) 항체, 인간 항체 등도 포함된다. 이들 항체는, 이미 알려진 방법을 사용하여 제조할 수 있다.
키메라 항체로는, 항체의 가변 영역과 정상 영역이 서로 이종인 항체, 예를 들어 마우스 또는 래트 유래 항체의 가변 영역을 인간 유래의 정상 영역에 접합한 키메라 항체를 들 수 있다 (Proc. Natl. Acad. Sci. U.S.A., 81, 6851-6855, (1984) 참조). LN22R8 유래의 키메라 항체의 일례로서, 서열표의 서열 번호 33 의 20 내지 471 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 서열 번호 35 의 20 내지 467 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄 혹은 서열 번호 37 의 20 내지 468 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄 및 서열 번호 31 의 21 내지 234 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 경쇄로 이루어지는 항체를 들 수 있다. 2P10F2 유래의 키메라 항체의 일례로서, 서열표의 서열 번호 43 의 20 내지 466 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 서열표의 서열 번호 45 의 20 내지 462 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄 혹은 서열 번호 47 의 20 내지 463 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄 및 서열 번호 41 의 21 내지 234 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 경쇄로 이루어지는 항체를 들 수 있다.
래트_CD147_#84 유래의 키메라 항체의 일례로서, 서열표의 서열 번호 92 의 20 내지 470 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 서열표의 서열 번호 94 의 20 내지 466 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 서열 번호 96 의 20 내지 467 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 서열 번호 98 의 20 내지 470 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄 혹은 서열 번호 100 의 20 내지 467 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 및, 서열 번호 90 의 21 내지 234 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 경쇄로 이루어지는 항체를 들 수 있다.
래트_CD147_#101 유래의 키메라 항체의 일례로서, 서열표의 서열 번호 104 의 20 내지 463 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 서열표의 서열 번호 106 의 20 내지 464 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄 혹은 서열 번호 108 의 20 내지 464 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 및, 서열 번호 102 의 21 내지 234 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 경쇄로 이루어지는 항체를 들 수 있다.
래트_CD147_#110 유래의 키메라 항체의 일례로서, 서열표의 서열 번호 112 의 20 내지 462 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 서열표의 서열 번호 114 의 20 내지 463 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄 혹은 서열 번호 116 의 20 내지 463 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 및, 서열 번호 110 의 21 내지 234 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 경쇄로 이루어지는 항체를 들 수 있다.
래트_CD147_#131 유래의 키메라 항체의 일례로서, 서열표의 서열 번호 120 의 20 내지 464 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄 혹은 서열 번호 122 의 20 내지 465 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 중쇄, 및, 서열 번호 118 의 21 내지 234 번째의 아미노산 잔기로 이루어지는 아미노산 서열을 갖는 경쇄로 이루어지는 항체를 들 수 있다.
인간화 항체로는, CDR 만을 인간 유래의 항체에 짜넣은 항체 (Nature (1986) 321, p. 522-525 참조), CDR 이식법에 의해, CDR 의 서열에 더하여 일부의 프레임 워크의 아미노산 잔기도 인간 항체에 이식한 항체 (국제 공개 제WO90/07861호 팸플릿) 를 들 수 있다.
래트_CD147_#84 항체 유래의 인간화 항체로는, 래트_CD147_#84 의 6 종 모두의 CDR 서열을 유지하고, CD147 에 대한 결합 활성을 갖고, 또한, CD147 을 활성화하는 항체인 한, 본 발명의 항체에 포함된다. 또한, 래트_CD147_#84 항체의 중쇄 가변 영역은, 서열 번호 55 에 나타내는 아미노산 서열로 이루어지는 CDRH1, 서열 번호 56 에 나타내는 아미노산 서열로 이루어지는 CDRH2, 및 서열 번호 57 에 나타내는 아미노산 서열로 이루어지는 CDRH3 을 보유하고 있다. 또, 래트_CD147_#84 항체의 경쇄 가변 영역은, 서열 번호 52 에 나타내는 아미노산 서열로 이루어지는 CDRL1, 서열 번호 53 에 나타내는 아미노산 서열로 이루어지는 CDRL2, 및 서열 번호 54 에 나타내는 아미노산 서열로 이루어지는 CDRL3 을 보유하고 있다. 래트_CD147_#84 항체의 경쇄 가변 영역 또는 중쇄 가변 영역의 아미노산 서열 및 뉴클레오티드 서열, CDR 의 아미노산 서열은, 도 33a 및 도 33b 에도 기재되어 있다.
래트_CD147_#101 항체 유래의 인간화 항체로는, 래트_CD147_#101 의 6 종 모두의 CDR 서열을 유지하고, CD147 에 대한 결합 활성을 갖고, 또한, CD147 을 활성화하는 항체인 한, 본 발명의 항체에 포함된다. 또한, 래트_CD147_#101 항체의 중쇄 가변 영역은, 서열 번호 65 에 나타내는 아미노산 서열로 이루어지는 CDRH1, 서열 번호 66 에 나타내는 아미노산 서열로 이루어지는 CDRH2, 및 서열 번호 67 에 나타내는 아미노산 서열로 이루어지는 CDRH3 을 보유하고 있다. 또, 래트_CD147_#101 항체의 경쇄 가변 영역은, 서열 번호 62 에 나타내는 아미노산 서열로 이루어지는 CDRL1, 서열 번호 63 에 나타내는 아미노산 서열로 이루어지는 CDRL2, 및 서열 번호 64 에 나타내는 아미노산 서열로 이루어지는 CDRL3 을 보유하고 있다. 래트_CD147_#101 항체의 경쇄 가변 영역 또는 중쇄 가변 영역의 아미노산 서열 및 뉴클레오티드 서열, CDR 의 아미노산 서열은, 도 34a 및 도 34b 에도 기재되어 있다.
래트_CD147_#110 항체 유래의 인간화 항체로는, 래트_CD147_#110 의 6 종 모두의 CDR 서열을 유지하고, CD147 에 대한 결합 활성을 갖고, 또한, CD147 을 활성화하는 항체인 한, 본 발명의 항체에 포함된다. 또한, 래트_CD147_#110 항체의 중쇄 가변 영역은, 서열 번호 75 에 나타내는 아미노산 서열로 이루어지는 CDRH1, 서열 번호 76 에 나타내는 아미노산 서열로 이루어지는 CDRH2, 및 서열 번호 77 에 나타내는 아미노산 서열로 이루어지는 CDRH3 을 보유하고 있다. 또, 래트_CD147_#110 항체의 경쇄 가변 영역은, 서열 번호 72 에 나타내는 아미노산 서열로 이루어지는 CDRL1, 서열 번호 73 에 나타내는 아미노산 서열로 이루어지는 CDRL2, 및 서열 번호 74 에 나타내는 아미노산 서열로 이루어지는 CDRL3 을 보유하고 있다. 래트_CD147_#110 항체의 경쇄 가변 영역 또는 중쇄 가변 영역의 아미노산 서열 및 뉴클레오티드 서열, CDR 의 아미노산 서열은, 도 35a 및 도 35b 에도 기재되어 있다.
래트_CD147_#131 항체 유래의 인간화 항체로는, 래트_CD147_#131 의 6 종 모두의 CDR 서열을 유지하고, CD147 에 대한 결합 활성을 갖고, 또한, CD147 을 활성화하는 항체인 한, 본 발명의 항체에 포함된다. 또한, 래트_CD147_#131 항체의 중쇄 가변 영역은, 서열 번호 85 에 나타내는 아미노산 서열로 이루어지는 CDRH1, 서열 번호 86 에 나타내는 아미노산 서열로 이루어지는 CDRH2, 및 서열 번호 87 에 나타내는 아미노산 서열로 이루어지는 CDRH3 을 보유하고 있다. 또, 래트_CD147_#131 항체의 경쇄 가변 영역은, 서열 번호 82 에 나타내는 아미노산 서열로 이루어지는 CDRL1, 서열 번호 83 에 나타내는 아미노산 서열로 이루어지는 CDRL2, 및 서열 번호 84 에 나타내는 아미노산 서열로 이루어지는 CDRL3 을 보유하고 있다. 래트_CD147_#131 항체의 경쇄 가변 영역 또는 중쇄 가변 영역의 아미노산 서열 및 뉴클레오티드 서열, CDR 의 아미노산 서열은, 도 36a 및 도 36b 에도 기재된다.
또, 또한 각 CDR 중의 1 내지 3 개의 아미노산 잔기를 다른 아미노산 잔기로 치환한 CDR 개변 인간화 항체도, CD147 에 대한 결합 활성을 갖고, 또한, CD147 을 활성화하는 항체인 한, 본 발명의 항체에 포함된다.
래트_CD147_#84 항체 유래의 인간화 항체로는, 이하의 (a) 및 (b) 를 갖는, 인간 CD147 항체 또는 그 항체의 항원 결합 단편을 들 수 있다 :
(a) 이하의 (a1) 내지 (a4) 로 이루어지는 군에서 선택되는 어느 하나에 기재된 중쇄 가변 영역 :
(a1) 서열 번호 123 에 나타내는 아미노산 서열의 20 내지 140 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 ;
(a2) 서열 번호 125 에 나타내는 아미노산 서열의 20 내지 140 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 ;
(a3) (a1) 또는 (a2) 의 서열에 있어서 각 CDR 서열 이외의 프레임 워크 영역의 서열에 대하여 적어도 95 % 이상의 서열 동일성을 갖는 아미노산 서열 ; 및
(a4) (a1) 내지 (a3) 중 어느 하나에 기재된 서열에 있어서의 각 CDR 서열 이외의 프레임 워크 영역의 서열에 있어서 1 또는 수 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열, 그리고,
(b) 이하의 (b1) 내지 (b3) 으로 이루어지는 군에서 선택되는 어느 하나에 기재된 경쇄 가변 영역 :
(b1) 서열 번호 127 에 나타내는 아미노산 서열의 21 내지 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역 ;
(b2) (b1) 의 서열에 있어서 각 CDR 서열 이외의 프레임 워크 영역의 서열에 대하여 적어도 95 % 이상의 서열 동일성을 갖는 아미노산 서열 ; 및
(b3) (b1) 또는 (b2) 의 서열에 있어서의 각 CDR 서열 이외의 프레임 워크 영역의 서열에 있어서 1 또는 수 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열.
래트_CD147_#84 항체 유래의 인간화 항체의 바람직한 예로는, 서열 번호 125 에 나타내는 아미노산 서열의 20 내지 140 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 및 서열 번호 127 에 나타내는 아미노산 서열의 21 내지 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역을 포함하는 항체, 또는, 서열 번호 123 에 나타내는 아미노산 서열의 20 내지 140 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 및 서열 번호 127 에 나타내는 아미노산 서열의 21 내지 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역을 포함하는 항체를 들 수 있다.
래트_CD147_#84 항체 유래의 인간화 항체의 보다 바람직한 예로는, 서열 번호 125 에 나타내는 아미노산 서열의 20 내지 467 번째의 아미노산 잔기로 이루어지는 중쇄 및 서열 번호 127 에 나타내는 아미노산 서열의 21 내지 234 번째의 아미노산 잔기로 이루어지는 경쇄를 포함하는 항체, 또는, 서열 번호 123 에 나타내는 아미노산 서열의 20 내지 466 번째의 아미노산 잔기로 이루어지는 중쇄 및 서열 번호 127 에 나타내는 아미노산 서열의 21 내지 234 번째의 아미노산 잔기로 이루어지는 경쇄를 포함하는 항체를 들 수 있다.
래트_CD147_#101 항체 유래의 인간화 항체로는, 이하의 (e) 및 (f) 를 갖는, 인간 CD147 항체 또는 그 항체의 항원 결합 단편을 들 수 있다 :
(e) 이하의 (e1) 내지 (e4) 로 이루어지는 군에서 선택되는 어느 하나에 기재된 중쇄 가변 영역 :
(e1) 서열 번호 129 에 나타내는 아미노산 서열의 20 내지 137 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 ;
(e2) 서열 번호 131 에 나타내는 아미노산 서열의 20 내지 137 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 ;
(e3) (e1) 또는 (e2) 의 서열에 있어서 각 CDR 서열 이외의 프레임 워크 영역의 서열에 대하여 적어도 95 % 이상의 서열 동일성을 갖는 아미노산 서열 ; 및
(e4) (e1) 내지 (e3) 중 어느 하나에 기재된 서열에 있어서의 각 CDR 서열 이외의 프레임 워크 영역의 서열에 있어서 1 또는 수 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열, 그리고,
(f) 이하의 (f1) 내지 (f3) 으로 이루어지는 군에서 선택되는 어느 하나에 기재된 경쇄 가변 영역 :
(f1) 서열 번호 133 에 나타내는 아미노산 서열의 21 내지 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역 ;
(f2) (f1) 의 서열에 있어서 각 CDR 서열 이외의 프레임 워크 영역의 서열에 대하여 적어도 95 % 이상의 서열 동일성을 갖는 아미노산 서열 ; 및
(f3) (f1) 또는 (f2) 의 서열에 있어서의 각 CDR 서열 이외의 프레임 워크 영역의 서열에 있어서 1 또는 수 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열.
래트_CD147_#101 항체 유래의 인간화 항체의 바람직한 예로는, 서열 번호 129 에 나타내는 아미노산 서열의 20 내지 137 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 및 서열 번호 133 에 나타내는 아미노산 서열의 21 내지 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역을 포함하는 항체, 또는, 서열 번호 131 에 나타내는 아미노산 서열의 20 내지 137 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 및 서열 번호 133 에 나타내는 아미노산 서열의 21 내지 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역을 포함하는 항체를 들 수 있다.
래트_CD147_#101 항체 유래의 인간화 항체의 보다 바람직한 예로는, 서열 번호 129 에 나타내는 아미노산 서열의 20 내지 463 번째의 아미노산 잔기로 이루어지는 중쇄 및 서열 번호 133 에 나타내는 아미노산 서열의 21 내지 234 번째의 아미노산 잔기로 이루어지는 경쇄를 포함하는 항체, 또는, 서열 번호 131 에 나타내는 아미노산 서열의 20 내지 464 번째의 아미노산 잔기로 이루어지는 중쇄 및 서열 번호 133 에 나타내는 아미노산 서열의 21 내지 234 번째의 아미노산 잔기로 이루어지는 경쇄를 포함하는 항체를 들 수 있다.
래트_CD147_#110 항체 유래의 인간화 항체로는, 이하의 (c) 및 (d) 를 갖는, 인간 CD147 항체 또는 그 항체의 항원 결합 단편을 들 수 있다 :
(c) 이하의 (c1) ∼ (c4) 로 이루어지는 군에서 선택되는 어느 하나에 기재된 중쇄 가변 영역 :
(c1) 서열 번호 135 에 나타내는 아미노산 서열의 20 ∼ 136 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 ;
(c2) 서열 번호 147 에 나타내는 아미노산 서열의 20 ∼ 136 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 ;
(c3) (c1) 또는 (c2) 의 서열에 있어서 각 CDR 서열 이외의 프레임 워크 영역의 서열에 대하여 적어도 95 % 이상의 서열 동일성을 갖는 아미노산 서열 ; 및
(c4) (c1) ∼ (c3) 중 어느 하나에 기재된 서열에 있어서의 각 CDR 서열 이외의 프레임 워크 영역의 서열에 있어서 1 또는 수 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열, 그리고,
(d) 이하의 (d1) ∼ (d5) 로 이루어지는 군에서 선택되는 어느 하나에 기재된 경쇄 가변 영역 :
(d1) 서열 번호 137 에 나타내는 아미노산 서열의 21 ∼ 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역 ;
(d2) 서열 번호 149 에 나타내는 아미노산 서열의 21 ∼ 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역 ;
(d3) 서열 번호 151 에 나타내는 아미노산 서열의 21 ∼ 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역 ;
(d4) (d1) ∼ (d3) 중 어느 하나에 기재된 서열에 있어서 각 CDR 서열 이외의 프레임 워크 영역의 서열에 대하여 적어도 95 % 이상의 서열 동일성을 갖는 아미노산 서열 ; 및
(d5) (d1) ∼ (d4) 중 어느 하나에 기재된 서열에 있어서의 각 CDR 서열 이외의 프레임 워크 영역의 서열에 있어서 1 또는 수 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열.
래트_CD147_#110 항체 유래의 인간화 항체의 바람직한 예로는, 서열 번호 135 에 나타내는 아미노산 서열의 20 내지 136 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 및 서열 번호 137 에 나타내는 아미노산 서열의 21 내지 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역을 포함하는 항체 ; 서열 번호 147 에 나타내는 아미노산 서열의 20 ∼ 136 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 및 서열 번호 149 에 나타내는 아미노산 서열의 21 ∼ 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역을 포함하는 항체 ; 또는, 서열 번호 147 에 나타내는 아미노산 서열의 20 ∼ 136 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 및 서열 번호 151 에 나타내는 아미노산 서열의 21 ∼ 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역을 포함하는 항체를 들 수 있다.
래트_CD147_#110 항체 유래의 인간화 항체의 보다 바람직한 예로는, 서열 번호 135 에 나타내는 아미노산 서열의 20 내지 463 번째의 아미노산 잔기로 이루어지는 중쇄 및 서열 번호 137 에 나타내는 아미노산 서열의 21 내지 234 번째의 아미노산 잔기로 이루어지는 경쇄를 포함하는 항체 ; 서열 번호 147 에 나타내는 아미노산 서열의 20 ∼ 463 번째의 아미노산 잔기로 이루어지는 중쇄, 및, 서열 번호 149 에 나타내는 아미노산 서열의 21 ∼ 234 번째의 아미노산 잔기로 이루어지는 경쇄를 포함하는 항체 ; 또는, 서열 번호 147 에 나타내는 아미노산 서열의 20 ∼ 463 번째의 아미노산 잔기로 이루어지는 중쇄, 및, 서열 번호 151 에 나타내는 아미노산 서열의 21 ∼ 234 번째의 아미노산 잔기로 이루어지는 경쇄를 포함하는 항체를 들 수 있다.
래트_CD147_#131 항체 유래의 인간화 항체로는, 이하의 (g) 및 (h) 를 갖는, 인간 CD147 항체 또는 그 항체의 항원 결합 단편을 들 수 있다 :
(g) 이하의 (g1) ∼ (g3) 으로 이루어지는 군에서 선택되는 어느 하나에 기재된 중쇄 가변 영역 :
(g1) 서열 번호 139 에 나타내는 아미노산 서열의 20 ∼ 138 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 ;
(g2) (g1) 의 서열에 있어서 각 CDR 서열 이외의 프레임 워크 영역의 서열에 대하여 적어도 95 % 이상의 서열 동일성을 갖는 아미노산 서열 ; 및
(g3) (g1) 또는 (g2) 의 서열에 있어서의 각 CDR 서열 이외의 프레임 워크 영역의 서열에 있어서 1 또는 수 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열, 그리고,
(h) 이하의 (h1) ∼ (h3) 으로 이루어지는 군에서 선택되는 어느 하나에 기재된 경쇄 가변 영역 :
(h1) 서열 번호 141 에 나타내는 아미노산 서열의 21 ∼ 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역 ;
(h2) (h1) 의 서열에 있어서 각 CDR 서열 이외의 프레임 워크 영역의 서열에 대하여 적어도 95 % 이상의 서열 동일성을 갖는 아미노산 서열 ; 및
(h3) (h1) 또는 (h2) 의 서열에 있어서의 각 CDR 서열 이외의 프레임 워크 영역의 서열에 있어서 1 또는 수 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열.
래트_CD147_#131 항체 유래의 인간화 항체의 바람직한 예로는, 서열 번호 139 에 나타내는 아미노산 서열의 20 내지 138 번째의 아미노산 잔기로 이루어지는 중쇄 가변 영역 및 서열 번호 141 에 나타내는 아미노산 서열의 21 내지 128 번째의 아미노산 잔기로 이루어지는 경쇄 가변 영역을 포함하는 항체를 들 수 있다.
래트_CD147_#131 항체 유래의 인간화 항체의 보다 바람직한 예로는, 서열 번호 139 에 나타내는 아미노산 서열의 20 내지 464 번째의 아미노산 잔기로 이루어지는 중쇄 및 서열 번호 141 에 나타내는 아미노산 서열의 21 내지 234 번째의 아미노산 잔기로 이루어지는 경쇄를 포함하는 항체를 들 수 있다.
상기의 래트_CD147_#84 항체 유래의 인간화 항체, 래트_CD147_#101 항체 유래의 인간화 항체, 래트_CD147_#110 항체 유래의 인간화 항체 또는 래트_CD147_#131 항체 유래의 인간화 항체는, 바람직하게는, CD147 을 개재한 p38MAPK 시그널 전달 및/또는 SMAD4 의 시그널 전달을 활성화한다.
본 발명의 항체로는, 또한, 인간 항체를 들 수 있다. 항 CD147 인간 항체란, 인간 염색체 유래의 항체의 유전자 서열만을 갖는 인간 항체를 의미한다. 항 CD147 인간 항체는, 예를 들어, 인간 항체의 중쇄와 경쇄의 유전자를 포함하는 인간 염색체 단편을 갖는 인간 항체 산생 마우스를 사용한 방법 (Tomizuka, K. et al., Nature Genetics (1997) 16, p. 133-143 ; Kuroiwa, Y. et. al., Nucl. Acids Res. (1998) 26, p. 3447-3448 ; Yoshida, H. et. al., Animal Cell Technology : Basic and Applied Aspects vol. 10, p. 69-73 (Kitagawa, Y., Matsuda, T. and Iijima, S. eds.), Kluwer Academic Publishers, 1999 ; Tomizuka, K. et. al., Proc. Natl. Acad. Sci. USA (2000) 97, p. 722-727 등을 참조.) 에 의해 취득할 수 있다.
이와 같은 인간 항체 산생 마우스는, 구체적으로는, 내재성 면역 글로불린 중쇄 및 경쇄의 유전자좌가 파괴되고, 대신에 인간 인공 염색체 (Human artificial chromosome, HAC) 벡터나 마우스 인공 염색체 (Mouse artificial chromosome, MAC) 벡터 등의 벡터를 개재하여 인간 면역 글로불린 중쇄 및 경쇄의 유전자좌가 도입된 유전자 조작 동물을, 녹아웃 동물 및 트랜스제닉 동물의 제조, 및 이들 동물끼리를 교배시킴으로써 만들어 낼 수 있다.
또, 유전자 조작 기술에 의해, 그러한 인간 항체의 중쇄 및 경쇄의 각각을 코드하는 cDNA, 바람직하게는 그 cDNA 를 포함하는 벡터에 의해 진핵 세포를 형질 전환하고, 유전자 조작 인간 모노클로날 항체를 산생하는 형질 전환 세포를 배양함으로써, 이 항체를 배양 상청 중으로부터 얻을 수도 있다. 여기서, 숙주로는 예를 들어 진핵 세포, 바람직하게는 CHO 세포, 림프구나 미엘로마 등의 포유 동물 세포를 사용할 수 있다.
또, 인간 항체 라이브러리에서 선별한 파지 디스플레이 유래의 인간 항체를 취득하는 방법 (Wormstone, I. M. et. al, Investigative Ophthalmology & Visual Science. (2002) 43 (7), p. 2301-2308 ; Carmen, S. et. al., Briefings in Functional Genomics and Proteomics (2002), 1 (2), p. 189-203 ; Siriwardena, D. et. al., Ophthalmology (2002) 109 (3), p. 427-431 등 참조.) 도 알려져 있다.
예를 들어, 인간 항체의 가변 영역을 1 본쇄 항체 (scFv) 로서 파지 표면에 발현시켜, 항원에 결합하는 파지를 선택하는 파지 디스플레이법 (Nature Biotechnology (2005), 23, (9), p. 1105-1116) 을 사용할 수 있다. 항원에 결합함으로써 선택된 파지의 유전자를 해석함으로써, 항원에 결합하는 인간 항체의 가변 영역을 코드하는 DNA 서열을 결정할 수 있다. 항원에 결합하는 scFv 의 DNA 서열이 밝혀지면, 당해 서열을 갖는 발현 벡터를 제조하고, 적당한 숙주에 도입하여 발현시킴으로써 인간 항체를 취득할 수 있다 (WO92/01047, WO92/20791, WO93/06213, WO93/11236, WO93/19172, WO95/01438, WO95/15388, Annu. Rev. I㎜unol (1994) 12, p. 433-455, Nature Biotechnology (2005) 23 (9), p. 1105-1116).
본 발명이 제공하는 항체와 동일한 에피토프를 갖는 항체도 본 발명의 항체에 포함된다. 예를 들어, LN22R8, 2P10F2, 래트_CD147_#84, 래트_CD147_#101, 래트_CD147_#110 또는 래트_CD147_#131 중의 적어도 어느 1 개와 동일한 에피토프를 갖는 항체를 들 수 있다.
본 발명의 LN22R8 및 2P10F2 는, 도 3 의 mu3 으로 나타내는 에피토프 (인간 CD147V1 : DALPGQKTEFKVDSDDQ (서열 번호 143), 원숭이 CD147 : DTLPGQKTDFEVDSDDL (서열 번호 144)) 를 인식하였다. 본 발명의 항체에는, 서열 번호 143 또는 서열 번호 144, 혹은 서열 번호 143 또는 서열 번호 144 의 서열에 있어서 1 또는 수 개, 바람직하게는 1 ∼ 3 개, 보다 바람직하게는 1 또는 2 개의 아미노산이 결실, 치환 또는 부가된 아미노산 서열을 포함하는 서열을 인식 또는 결합하는 항체도, 그 범위에 포함된다. 이 항체는, CD147 을 개재한 시그널 전달을 활성화하는 것이 바람직하다.
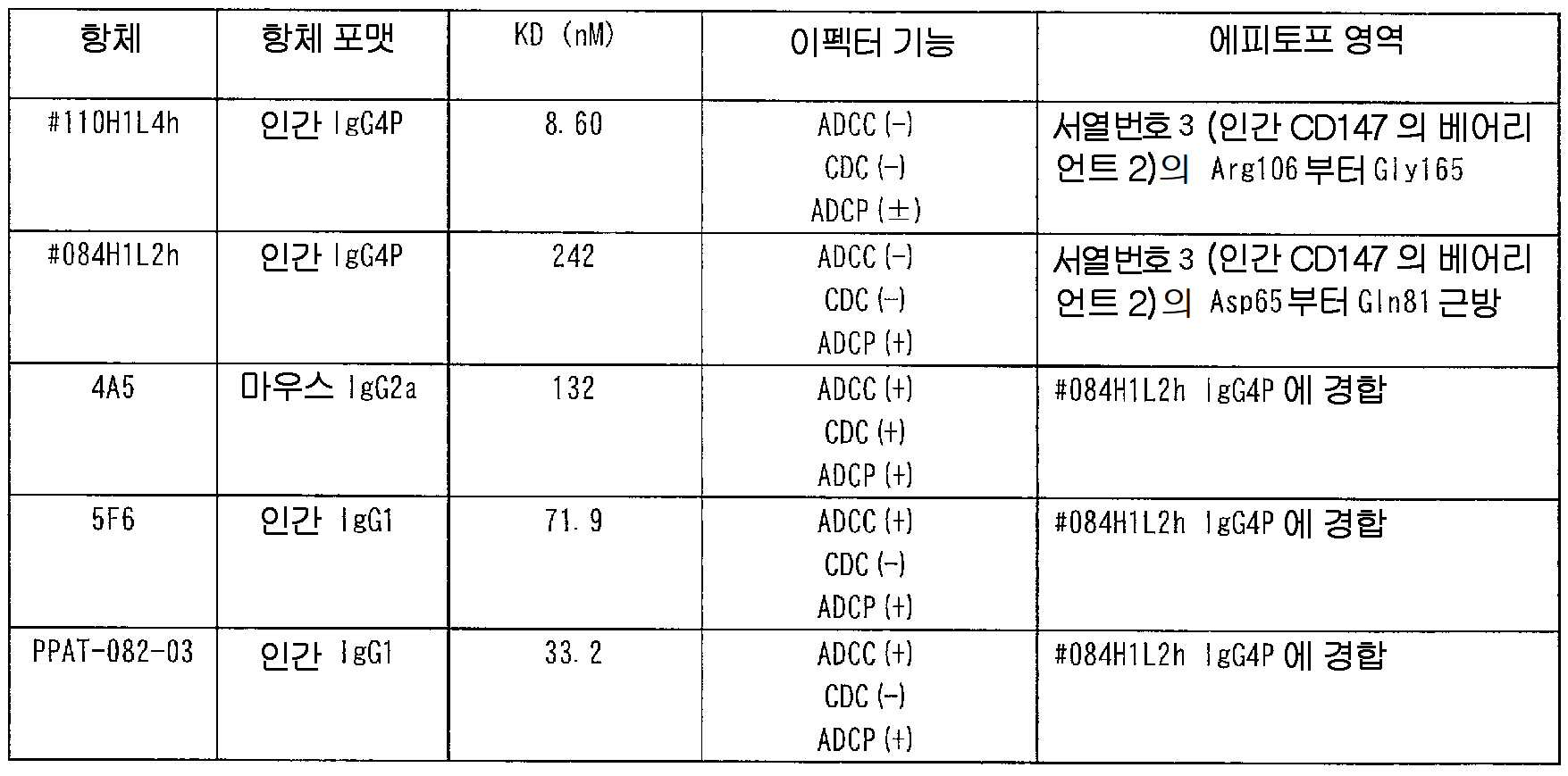
또, 본 발명의 항체에는, 래트_CD147_#110, 바람직하게는, 인간화 #110H1L4 와 동일한 에피토프를 인식하는 항체를 포함한다. 이 항체는, CD147 을 개재한 시그널 전달을 활성화하는 것이 바람직하다. 인간화 #110H1L4 의 에피토프 해석의 결과는, 실시예 17 에 나타난다.
적합한 항체는, 당업자에게 주지인 항원에 대한 결합성을 평가함으로써 선발할 수 있다. 항체와 항원 (CD147) 의 해리 정수 (定數) 는 표면 플라즈몬 공명 (SPR) 을 검출 원리로 하는 비아코어 T200 (GE Healthcare Bioscience 사) 를 사용하여 측정할 수 있다. 예를 들어, 리간드로서 고상화한 항원에 대하여, 적당한 농도로 설정한 항체를 아날라이트와 반응시키고, 그 결합 및 해리를 측정함으로써, 결합 속도 정수 ka1, 해리 속도 정수 kd1 및 해리 정수 (KD ; KD = kd1/ka1) 를 얻을 수 있다.
CD147 에 대한 결합성 평가는, 비아코어 T200 의 사용에 한정되지 않고, 표면 플라즈몬 공명 (SPR) 을 검출 원리로 하는 기기, 결합 평형 제외법 (Kinetic Exclusion Assay) 을 검출 원리로 하는 KinExA (Sapidyne Instruments 사), 바이오레이어 간섭법 (Bio-Layer Interferometry) 을 검출 원리로 하는 BLItz 시스템 (Pall 사) 혹은 ELISA (Enzyme-Linked I㎜unoSorbent Assay) 법 등에 의해서도 가능하다.
항체의 성질을 비교할 때의 다른 지표의 일례로는, 항체의 안정성을 들 수 있다. 시차 주사 칼로리메트리 (DSC) 는, 단백의 상대적 구조 안정성이 좋은 지표가 되는 열 변성 중점 (Tm) 을 재빠르게, 또 정확하게 측정할 수 있는 방법이다. DSC 를 사용하여 Tm 값을 측정하고, 그 값을 비교함으로써, 열안정성의 차이를 비교할 수 있다. 항체의 보존 안정성은, 항체의 열안정성과 어느 정도의 상관을 나타내는 것이 알려져 있으며 (Lori Burton, et. al., Pharmaceutical Development and Technology (2007) 12, p. 265-273), 열안정성을 지표로 적합한 항체를 선발할 수 있다. 항체를 선발하기 위한 다른 지표로는, 적절한 숙주 세포에 있어서의 수량 (收量) 이 높은 것, 및 수용액 중에서의 응집성이 낮은 것을 들 수 있다. 예를 들어 수량이 가장 높은 항체가 가장 높은 열안정성을 나타낸다고는 할 수 없기 때문에, 이상에 서술한 지표에 기초하여 종합적으로 판단하여, 인간에 대한 투여에 가장 적합한 항체를 선발할 필요가 있다.
또, 항체의 중쇄 및 경쇄의 전체 길이 서열을 적절한 링커를 사용하여 연결하고, 1 본쇄 이뮤노글로불린 (single chain i㎜unoglobulin) 을 취득하는 방법도 알려져 있다 (Lee, H-S, et. al., Molecular I㎜unology (1999) 36, p. 61-71 ; Shirrmann, T. et. al., ㎃bs (2010), 2, (1) p. 1-4). 이와 같은 1 본쇄 이뮤노글로불린은 2량체화 함으로써, 본래는 4량체인 항체와 유사한 구조와 활성을 유지하는 것이 가능하다. 또, 본 발명의 항체는, 단일의 중쇄 가변 영역을 갖고, 경쇄 서열을 갖지 않는 항체여도 된다. 이와 같은 항체는, 단일 도메인 항체 (single domain antibody : sdAb) 또는 나노보디 (nanobody) 라고 불리고 있으며, 실제로 낙타 또는 라마에서 관찰되고, 항원 결합능이 유지되고 있는 것이 보고되어 있다 (Muyldemans S. et. al., Protein Eng. (1994) 7 (9), 1129-35, Hamers-Casterman C. et. al., Nature (1993) 363 (6428) 446-8). 상기의 항체는, 본 발명에 있어서의 항체의 항원 결합 단편의 1 종이라고 해석하는 것도 가능하다.
또, 본 발명의 항체에 결합하고 있는 당사슬 수식을 조절함으로써, 항체 의존성 세포 장애 활성을 증강하는 것이 가능하다. 항체의 당사슬 수식의 조절 기술로는, WO99/54342, WO2000/61739, WO2002/31140 등이 알려져 있지만, 이들에 한정되는 것은 아니다.
항체 유전자를 일단 단리한 후, 적당한 숙주에 도입하여 항체를 제조하는 경우에는, 적당한 숙주와 발현 벡터의 조합을 사용할 수 있다.
항체 유전자의 구체예로는, 본 명세서에 기재된 항체의 중쇄 서열을 코드하는 유전자, 및 경쇄 서열을 코드하는 유전자를 조합한 것을 들 수 있다. 숙주 세포를 형질 전환할 때에는, 중쇄 서열 유전자와 경쇄 서열 유전자는, 동일한 발현 벡터에 삽입되어 있는 것이 가능하고, 또 별개의 발현 벡터에 삽입되어 있는 것도 가능하다. 진핵 세포를 숙주로서 사용하는 경우, 동물 세포, 식물 세포, 진핵 미생물을 사용할 수 있다. 동물 세포로는, 포유류 세포, 예를 들어, 원숭이의 세포인 COS 세포 (Gluzman, Y. Cell (1981) 23, p. 175-182, ATCC CRL-1650), 마우스 선유아세포 NIH3T3 (ATCC No. CRL-1658) 이나 차이니즈·햄스터 난소 세포 (CHO 세포, ATCC CCL-61) 의 디하이드로 엽산 환원 효소 결손주 (Urlaub, G. and Chasin, L. A. Proc. Natl. Acad. Sci. U.S.A. (1980) 77, p. 4126-4220) 를 들 수 있다. 또, 원핵 세포를 사용하는 경우에는, 예를 들어, 대장균, 고초균을 들 수 있다. 이들 세포에 목적으로 하는 항체 유전자를 형질 전환에 의해 도입하고, 형질 전환된 세포를 in vitro 에서 배양함으로써 항체가 얻어진다. 이상의 배양법에 있어서는 항체의 서열에 의해 수량이 상이한 경우가 있고, 동등한 결합 활성을 갖는 항체 중에서 수량을 지표로 의약으로서의 생산이 용이한 것을 선별하는 것이 가능하다.
본 발명의 항체의 아이소타입으로서의 제한은 없고, 예를 들어 IgG (IgG1, IgG2, IgG3, IgG4), IgM, IgA (IgA1, IgA2), IgD 혹은 IgE 등을 들 수 있지만, 바람직하게는 IgG 또는 IgM, 더욱 바람직하게는 IgG 를 들 수 있다.
인간 IgG1 이란, 5 종 존재하는 인간 IgG 서브 클래스 중에서, 보체 결합을 통한 CDC 활성, 항체 의존적인 세포 장애 활성과 같은 이펙터 기능이 매우 강하고 (Bruggemann et al., J. Exp. Med., 1351-1361, 1987), 치료용 항체로 암에 고발현하는 분자를 표적으로 하는 경우에, 이펙터 기능을 통한 세포 장애에 의한 암 세포의 세포사 유도를 촉진함으로써 치료 효과를 나타내는 IgG 포맷으로서 이용된다 (트라스투주맙, 리툭시맙 등). HLA-DR 을 표적으로 한 인간 IgG1 항체에서는, 인간 IgG1 에 구비된 항체의 CDC 활성에 의존하고, 투여 후의 게잡이 원숭이가 사망한 것이 보고되어 있어, 정상 장기에서도 발현하고 있는 분자를 표적으로 하는 항체 의약에서는, 이펙터 기능이 심각한 부작용을 일으킬 염려가 있다 (Tawara, T., J. I㎜unology, 2008, 2294-2298). 본 발명의 항체의 아이소타입으로서 IgG1 을 사용하는 경우에는, IgG1 항체는 변이를 갖고 있어도 되고, 정상 영역의 아미노산 잔기의 일부를 치환함으로써, 이펙터 기능을 조정하는 것이 가능하다 (WO88/007089, WO94/28027, WO94/29351 참조). 이펙터 기능을 감약시킨 IgG1 의 변이체로는, IgG1 LALA (IgG1-L234A, L235A), 1gG1 LAGA (IgG1-L235A, G237A) 등을 들 수 있다.
인간 IgG2 란, 5 종 존재하는 인간 IgG 서브 클래스 중에서, 보체 결합을 통한 CDC 활성, 항체 의존적인 세포 장애 활성과 같은 이펙터 기능이 매우 약하고 (Bruggemann et al., J. Exp. Med., 1351-1361, 1987), 치료용 항체로 정상 장기에 발현하는 분자를 표적으로 하는 경우에 이펙터 기능을 통한 세포 장애에 의한 독성을 회피하기 위한 IgG 포맷의 하나로서 이용된다 (데노스맙, 에볼로쿠맙, 브로달루맙).
인간 IgG4 란, 5 종 존재하는 인간 IgG 서브 클래스 중에서, 보체 결합을 통한 CDC 활성, 항체 의존적인 세포 장애 활성과 같은 이펙터 기능이 매우 약하고 (Bruggemann et al., J. Exp. Med., 1351-1361, 1987), 치료용 항체로 정상 장기에 발현하는 분자를 표적으로 하는 경우에 이펙터 기능을 통한 세포 장애에 의한 독성을 회피하기 위한 IgG 포맷의 하나로서 이용된다 (옵디보). 본 발명의 항체의 아이소타입으로서 IgG4 를 사용하는 경우에는, 정상 영역의 아미노산 잔기의 일부를 치환함으로써, IgG4 특유의 분할이 억제되고, 반감기를 연장하는 것이 가능하다 (Molecular I㎜unology, 30, 1 105-108 (1993) 참조).
본 발명의 항체의 아이소타입으로서 IgG4 를 사용하는 경우, IgG4 항체는 변이를 갖고 있어도 된다. IgG4 의 변이체로는, EU 인덱스 (Proc Natl Acad Sci US A. 1969, 63 (1), 78-85 ; Kabat et. al., Sequences of proteins of i㎜unological interest, 1991 Fifth edition) 에 의해 나타내는 234 위치의 페닐알라닌의 알라닌으로의 치환 (F234A) 및 235 위치의 류신의 알라닌으로의 치환 (L235A) 을 들 수 있다 (Parekh et al., ㎃bs, 310-318, 2012). 이와 같은 항체의 변이를 FALA 변이라고 부른다. IgG4PFALA 는, CH2 도메인에 존재하는 FcγRs (예를 들어, FcγRI, FcγRII 또는 FcγRIII 등) 와의 상호 작용에 필요한 아미노산 잔기 2 개를 알라닌으로 치환함으로써, 더욱 이펙터 기능을 감약시키고 있다. 또, IgG4 는 항체 중쇄간의 SS 결합의 형성이 안정되어 있지 않기 때문에, 안정성을 높이기 위해서, 항체 중쇄간의 SS 결합의 형성을 촉진시키는 변이를 도입한다. 이와 같은 변이로서, EU 인덱스에 의해 나타내는 228 위치의 세린의 프롤린으로의 치환 (S228P) 을 들 수 있다 (ANGAL et. al., Molecular I㎜unology, 105-108, 1993). 이 항체의 변이를 Pro 변이라고 부른다. 본 발명의 항체의 정상 영역에는, 상기의 FALA 변이 및 Pro 변이가 동시에 도입되어 있어도 된다 (Vafa et. al., Methods, 65, 114-126, 2014). FALA 변이를 갖는 IgG4 중쇄를 「IgG4FALA」 타입 중쇄라고 부르고, Pro 변이를 갖는 IgG4 중쇄를 「IgG4P」 타입 중쇄라고 부르고, FALA 변이 및 Pro 변이의 양방의 변이를 갖는 IgG4 중쇄를 「IgG4PFALA」 타입 중쇄라고 부른다.
항체 중쇄 정상 영역은 CH1, 힌지, CH2 및 CH3 영역으로 이루어지고, CH1 은, EU 인덱스 118 내지 215, 힌지는 EU 인덱스 216 내지 230, CH2 는 EU 인덱스 231 내지 340, CH3 은 EU 인덱스 341 내지 446 으로 정의된다. EU 인덱스에 의해 나타내는, 228 위치의 세린으로부터 치환된 프롤린, 234 위치의 페닐알라닌으로부터 치환된 알라닌 및 235 위치의 류신으로부터 치환된 알라닌은, 각각, 인간 키메라 래트_CD147_#84 중쇄 IgG4PFALA 의 아미노산 서열을 나타내는 서열 번호 100 에 있어서 제 248 번째의 프롤린, 제 254 번째의 알라닌 및 제 255 번째의 알라닌, 인간 키메라 래트_CD147_#101 중쇄 IgG4PFALA 의 아미노산 서열을 나타내는 서열 번호 108 에 있어서 제 245 번째의 프롤린, 제 251 번째의 알라닌 및 제 252 번째의 알라닌, 인간 키메라 래트_CD147_#110 중쇄 IgG4PFALA 의 아미노산 서열을 나타내는 서열 번호 108 에 있어서 제 244 번째의 프롤린, 제 250 번째의 알라닌 및 제 251 번째의 알라닌에 상당한다.
본 발명의 항체의 바람직한 아이소타입으로는, IgG1, IgG2, IgG4, IgG4P 또는, IgG4PFALA 를 들 수 있고, 특히 바람직하게는, IgG2, IgG4P 또는, IgG4PFALA 를 들 수 있고, 보다 더 바람직하게는, IgG2 또는 IgG4P 를 들 수 있다.
또 본 발명의 항체는, 항체의 항원 결합부를 갖는 항체의 항원 결합 단편 또는 그 수식물이어도 된다. 항체를 파파인, 펩신 등의 단백질 분해 효소로 처리하거나, 혹은 항체 유전자를 유전자 공학적 수법에 의해 개변하여 적당한 배양 세포에 있어서 발현시킴으로써, 그 항체의 단편을 얻을 수 있다. 이와 같은 항체 단편 중에서, 항체 전체 길이 분자가 갖는 기능의 전부 또는 일부를 유지하고 있는 단편을 항체의 항원 결합 단편이라고 부를 수 있다. 항체의 기능으로는, 항원에 관련하는 시그널 전달의 활성화를 들 수 있다.
CD147 은, 적혈구를 비롯한 혈액 세포나 생존에 필수적인 정상 장기에서도 발현하고 있기 때문에 (Spring, et al., Eur. J. I㎜unol., 1997, 891-897), 항체에 부수하는 이펙터 기능을 이용한 항종양 효과는 부작용의 리스크가 높은 것으로 생각된다. 실제로 적혈구는, 항체 결합에 의해 생기는 이펙터 기능 (ADCC, CDC, ADCP) 에 감수성이 있는 것이 보고되어 있고 (Flegel, W., Transfusion, 2015, S47-S58), 적혈구에 대한 항체가 체내에서 증가함으로써 자기 면역성 용혈성 빈혈이 되는 것이 알려져 있다 (Gibson, J., Aust. N. Z. J. Med., 1988. 625-637). 본 발명의 항체는, 정상 세포에도 발현하는 CD147 을 표적으로 한 치료용 항체에 있어서, 심각한 부작용의 원인이 되는 ADCC 활성, ADCP 활성 또는 CDC 활성 중 어느 것, 혹은, 전부가 낮거나 또는 거의 검출되지 않는 것을 특징으로 한다.
본 발명자들은, 항체의 이펙터 기능에 따르지 않고, CD147 의 세포 시그널 전달을 활성화함으로써 항종양 활성을 나타내는 인간 CD147 항체를 처음으로 알아냈다. 본 발명에 있어서의 항체가 유지하는 기능은, CD147 에 대한 결합 활성 및/또는 CD147 을 활성화하는 기능이다. 본 발명의 항체는, 바람직하게는 CD147 을 개재하는 하류의 관련 시그널 분자, 예를 들어, FAK, MEK, Erk, JAK/STAT, AKT 또는 MAP 키나아제 (MAPK) 혹은 이들의 더 하류의 관련 시그널 분자를 활성화한다. 본 발명의 항체는, 보다 바람직하게는, MAPK 또는 MAPK 의 하류의 분자를 활성화한다. MAPK 로서, 바람직하게는 p38MAPK 를 들 수 있다. MAPK 의 더 하류의 시그널 분자로는, 예를 들어, HSP27, cxcl8 또는 SMAD (예를 들어, SMAD2, SMAD3 또는 SMAD4, 바람직하게는, SMAD4) 를 들 수 있다. 「CD147 의 활성화」 로는, 예를 들어, p38MAPK 의 mRNA 발현량의 증가, p38MAPK 의 단백질 발현량의 증가, p38MAPK 의 인산화, HSP27 의 인산화 (예를 들어, HSP27 의 Ser82 의 인산화 또는 HSP27 의 Ser15 의 인산화), cxcl8 mRNA 발현량의 증가, cxcl8 단백질 발현량의 증가 또는 SMAD 시그널 활성화를 개재한 rhoB mRNA 발현량의 증가 또는 rhoB 단백질 발현량의 증가를 들 수 있다. 「CD147 의 활성화」 로서 바람직하게는, p38MAPK 의 단백질 발현량의 증가, p38MAPK 의 인산화, HSP27 의 인산화 (예를 들어, HSP27 의 Ser82 의 인산화 또는 HSP27 의 Ser15 의 인산화), cxcl8 mRNA 발현량의 증가 또는 SMAD 시그널 활성화를 개재한 rhoB mRNA 발현량의 증가를 들 수 있다.
SMAD2 또는 SMAD3 은, TGFβ 가 TGFβ 수용체 (TGFBR1/2) 에 결합했을 때에, TGFβ 수용체에 의해 인산화 되고, SMAD4 와 헤테로 3량체를 형성하여, 핵내로 이행하고, 염색체 상의 SMAD DNA 결합 서열 (Smad binding element : SBE) 을 갖는 전사 조절 영역에 결합하고, 하류 유전자의 mRNA 발현을 정 (正) 혹은 부 (負) 로 제어하는 것이 알려져 있다 (미야조노, 일본 노년 의약회 잡지, 1999, 162-166). 따라서, SMAD4 의 활성화에는, SMAD2 또는 SMAD3 의 존재가 필요한 것으로 생각된다. SMAD2, SMAD3 및 SMAD4 는, TGFb 의존적으로 KLF5 의 발현량을 부로 제어하고 있다 (David et al., Cell, 2016, 164 (5), 1015-1030). SMAD4 가 손실되어 있는 췌장암 세포에서는, KLF5 유전자의 SMAD2, SMAD3 및 SMAD4 에 의한 억제 시그널이 해제되고, KLF5 단백질이 발현한다. SMAD4 가 손실되고, KLF5 가 발현하고 있으면 TGFb 의존적인 세포사의 시그널 (SOX4 의존적) 이 억제되는 것이 알려져 있다 (전술, David et al, Cell).
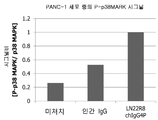
본 발명자들은, 본 발명의 인간 CD147 항체가, p38MAPK 를 인산화하고 (도 21), HSP27 을 인산화하고 (도 20), cxcl8 의 발현을 증가시키는 것을 알아냈다 (도 22(b) 및 도 23(b)). 따라서, 본 발명의 항체 투여 전후로, 이들 분자 중의 적어도 어느 1 개의 유전자 발현 혹은 단백질 발현, 또는 인산화 상태가 변화하는지를 확인함으로써, 본 발명의 항체가, CD147 의 활성화를 개재하고 있는지 여부를 확인할 수 있다.
또, 본 발명자들은, 본 발명의 인간 CD147 항체는, SMAD4 의 단백질이 발현하고 있는 췌장암 모델에 대해 약효를 나타내는 것 (도 25) ; 본 발명의 인간 CD147 항체는, SMAD4 에 유전자 변이가 있고, SMAD4 를 발현하고 있지 않는 BxPC-3 등의 췌장암 모델에서는, 항종양 효과는 부분적 (∼ 30 % 정도) 인 것 (도 24) ; 그리고, 본 발명의 인간 CD147 항체를 투여한 후의 종양에서는, SMAD2, SMAD3 및 SMAD4 의 하류의 분자 (rhoB, 도 22(c) 및 도 23(c)) 가 유도되는 것을 알아냈다. 따라서, 본 발명의 항체 투여 전후로, rhoB 의 유전자 발현 또는 단백질 발현이 변화하는지 여부를 확인함으로써, 본 발명의 항체가, CD147 의 활성화를 개재하고 있는지 여부를 확인할 수 있다. 또, 환자 샘플에 있어서의 SMAD4 의 게놈 서열, 유전자 발현 또는 단백질 발현을 당업자에게 주지의 방법을 사용하여 측정하고, SMAD4 를 발현하고 있는 환자를 본 발명의 항체를 투여하는 대상 환자로서 선택하고, 본 발명의 항체를 투여할 수 있다.
또, BxPC-3 등의 SMAD4 음성의 모델에서는, KLF5 의 발현이 높은 것이 알려져 있다 (David et al., Cell, 2016, 164 (5), 1015-1030). 본 발명의 인간 CD147 항체는, KLF5 를 발현시킨 MIA PaCa-2 의 모델에서, 감수성이 91 % 에서 20 % 로 저하되었다 (실시예 26). 이로부터, KLF5 가 발현함으로써, CD147 항체에 의해 유도되는 SMAD2, SMAD3 및 SMAD4 의존적인 세포사의 시그널이 억제되는 것으로 본 발명자들은 생각한다. 간암, ALL, 림프종, 소화관 간질 종양 (GIST), 피부암, 육종, AML 또는, 신장암은 KLF5 의 발현량이 낮고, 본 발명의 인간 CD147 항체가 효과가 있는 환자가 많은 것이 예상된다. 또, 환자 샘플에 있어서의 KLF5 의 유전자 발현 또는 단백질 발현을 당업자에게 주지의 방법을 사용하여 측정하고, KLF5 의 발현이 저하 또는 결실되어 있는 환자를 본 발명의 항체를 투여하는 대상 환자로서 선택하여, 본 발명의 항체를 투여할 수 있다. 이 경우의 KLF5 발현의 저하의 정도는, 당업자에 의해 주지의 방법 및 적절한 임상 시험의 실시에 의해 결정될 수 있으며, 예를 들어, 효과가 얻어진 환자와 효과가 얻어지지 않은 환자에 있어서의 KLF5 의 발현량을 비교하여 적절한 임계값이 설정된다.
예를 들어, 항체의 단편으로는, Fab, F(ab')2, Fv, 또는 중쇄 및 경쇄의 Fv 를 적당한 링커로 연결시킨 싱글 체인 Fv (scFv), diabody (diabodies), 선상 항체, 및 항체 단편으로 형성된 다특이성 항체 등을 들 수 있다. 또, F(ab')2 를 환원 조건하에서 처리한 항체의 가변 영역의 1 가의 단편인 Fab' 도 항체의 단편에 포함된다.
또한, 본 발명의 항체는 적어도 2 종류의 상이한 항원에 대하여 특이성을 갖는 다특이성 항체여도 된다. 통상적으로 이와 같은 분자는 2 종류의 항원에 결합하는 것이지만 (즉, 이중 특이성 항체 (bispecific antibody)), 본 발명에 있어서의 「다특이성 항체」 는, 그 이상 (예를 들어, 3 종류) 의 항원에 대하여 특이성을 갖는 항체를 포함하는 것이다.
본 발명의 다특이성 항체는, 전체 길이로 이루어지는 항체, 또는 그러한 항체의 단편 (예를 들어, F(ab')2 이중 특이성 항체) 이어도 된다. 이중 특이성 항체는 2 종류의 항체의 중쇄와 경쇄 (HL 쌍) 를 결합시켜 제조할 수도 있고, 상이한 모노클로날 항체를 산생하는 하이브리도마를 융합시켜, 이중 특이성 항체 산생 융합 세포를 제조하는 것에 의해서도, 제조할 수 있다 (Millstein et al., Nature (1983) 305, p. 537-539).
본 발명의 항체는 1 본쇄 항체 (scFv 라고도 기재한다) 여도 된다. 1 본쇄 항체는, 항체의 중쇄 가변 영역과 경쇄 가변 영역을 폴리펩티드의 링커로 연결함으로써 얻어진다 (Pluckthun, The Pharmacology of Monoclonal Antibodies, 113 (Rosenberg 및 Moore 편, Springer Verlag, New York, p. 269-315 (1994), Nature Biotechnology (2005), 23, p. 1126-1136). 또, 2 개의 scFv 를 폴리펩티드 링커로 결합시켜 제조되는 BiscFv 단편을 이중 특이성 항체로서 사용할 수도 있다.
1 본쇄 항체를 제조하는 방법은 당 기술 분야에 있어서 주지이다 (예를 들어, 미국 특허 제4,946,778호, 미국 특허 제5,260,203호, 미국 특허 제5,091,513호, 미국 특허 제5,455,030호 등을 참조). 이 scFv 에 있어서, 중쇄 가변 영역과 경쇄 가변 영역은, 콘쥬게이트를 만들지 않는 링커, 바람직하게는 폴리펩티드 링커를 개재하여 연결된다 (Huston, J. S. et al., Proc. Natl. Acad. Sci. U.S.A. (1988), 85, p. 5879-5883). scFv 에 있어서의 중쇄 가변 영역 및 경쇄 가변 영역은, 동일한 항체에서 유래해도 되고, 별개의 항체에서 유래해도 된다. 가변 영역을 연결하는 폴리펩티드 링커로는, 예를 들어 12 ∼ 19 잔기로 이루어지는 임의의 1 본쇄 펩티드가 사용된다.
scFv 를 코드하는 DNA 는, 상기 항체의 중쇄 또는 중쇄 가변 영역을 코드하는 DNA, 및 경쇄 또는 경쇄 가변 영역을 코드하는 DNA 중, 그들의 서열 중 전부 또는 원하는 아미노산 서열을 코드하는 DNA 부분을 주형으로 하고, 그 양단을 규정하는 프라이머쌍을 사용하여 PCR 법에 의해 증폭하고, 이어서, 또한 폴리펩티드 링커 부분을 코드하는 DNA, 및 그 양단이 각각 중쇄, 경쇄와 연결되도록 규정하는 프라이머쌍을 조합하여 증폭함으로써 얻어진다.
또, 일단 scFv 를 코드하는 DNA 가 제조되면, 그것들을 함유하는 발현 벡터, 및 그 발현 벡터에 의해 형질 전환된 숙주를 통상적인 방법에 따라서 얻을 수 있고, 또, 그 숙주를 사용함으로써, 통상적인 방법에 따라서 scFv 를 얻을 수 있다. 이들의 항체 단편은, 상기와 동일하게 하여 유전자를 취득하여 발현시키고, 숙주에 의해 산생시킬 수 있다.
본 발명의 항체는, 다량화하여 항원에 대한 친화성을 높인 것이어도 된다. 다량화하는 항체로는, 1 종류의 항체여도 되고, 동일한 항원의 복수의 에피토프를 인식하는 복수의 항체여도 된다. 항체를 다량화하는 방법으로는, IgG CH3 도메인과 2 개의 scFv 의 결합, 스트렙토아비딘과의 결합, 헬릭스 턴-헬릭스 모티프의 도입 등을 들 수 있다.
본 발명의 항체는, 아미노산 서열이 상이한 복수 종류의 항 CD147 항체의 혼합물인, 폴리클로날 항체여도 된다. 폴리클로날 항체의 일례로는, CDR 이 상이한 복수 종류의 항체의 혼합물을 들 수 있다. 그러한 폴리클로날 항체로는, 상이한 항체를 산생하는 세포의 혼합물을 배양하고, 그 배양물로부터 정제된 항체를 사용할 수 있다 (WO2004/061104 참조).
본 발명의 항체는, 상기의 항체의 중쇄 및/또는 경쇄와 비교하여 80 % 내지 99 % 라는 동일성 (또는 상동성) 을 갖는 항체여도 된다. 상기의 중쇄 아미노산 서열 및 경쇄 아미노산 서열과 높은 상동성을 나타내는 서열을 조합함으로써, 상기의 각 항체와 동등한 항원 결합능, CD147 의 활성화, 바람직하게는, MAPK 의 활성화, MAPK의 하류 시그널 분자의 활성화를 갖는 항체를 선택하는 것이 가능하다. 이와 같은 상동성은, 일반적으로는 80 % 이상의 상동성이고, 바람직하게는 90 % 이상의 상동성이고, 보다 바람직하게는 95 % 이상의 상동성이며, 가장 바람직하게는 99 % 이상의 상동성이다. 또, 중쇄 및/또는 경쇄의 아미노산 서열에 1 내지 수 개의 아미노산 잔기가 치환, 결실 및/또는 부가된 아미노산 서열을 조합함으로써도, 상기의 각 항체와 동등한 각종 작용을 갖는 항체를 선택하는 것이 가능하다. 치환, 결실 및/또는 부가되는 아미노산 잔기수는, 일반적으로는 10 아미노산 잔기 이하이고, 바람직하게는 5 내지 6 아미노산 잔기 이하이고, 보다 바람직하게는 2 내지 3 아미노산 잔기 이하이고, 가장 바람직하게는 1 아미노산 잔기이다. 또한, 포유류 배양 세포에서 생산되는 항체의 중쇄의 카르복실 말단의 리신 잔기가 결실하는 것이 알려져 있음 (Tsubaki et. al., Int. J. Biol. Macromol, 139-147, 2013). 그러나, 이들 중쇄 서열의 결실 및 수식은, 항체의 항원 결합능 및 이펙터 기능 (보체의 활성화나 항체 의존성 세포 장애 작용 등) 에는 영향을 미치지 않는다. 따라서, 본 발명에는 당해 수식을 받은 항체도 포함되고, 중쇄 카르복실 말단에 있어서 1 또는 2 개의 아미노산이 결실된 결실체, 및 아미드화 된 당해 결실체 (예를 들어, 카르복실 말단 부위의 프롤린 잔기가 아미드화 된 중쇄) 등을 들 수 있다. 단, 항원 결합능 및 CD147 의 하류의 관련 시그널 분자를 활성화하는 기능이 유지되고 있는 한, 본 발명에 관련된 항체의 중쇄의 카르복실 말단의 결실체는 상기의 종류에 한정되지 않는다. 본 발명에 관련된 항체를 구성하는 2 개의 중쇄는, 완전 길이 및 상기의 결실체로 이루어지는 군에서 선택되는 중쇄 중 어느 1 종이어도 되고, 어느 2 종을 조합한 것이어도 된다. 각 결실체의 양비 (量比) 는 본 발명에 관련된 항체를 산생하는 포유류 배양 세포의 종류 및 배양 조건에 영향을 받을 수 있지만, 본 발명에 관련된 항체의 주성분으로는 2 개의 중쇄의 쌍방에서 카르복실 말단의 하나의 아미노산 잔기가 결실하고 있는 경우를 들 수 있다.
2 종류의 아미노산 서열간의 상동성은, Blast algorithm version 2.2.2 (Altschul, Stephen F., Thomas L. Madden, Alejandro A. Schaffer, Jinghui Zhang, Zheng Zhang, Webb Miller, and David J. Lipman (1997), 「Gapped BLAST and PSI-BLAST : a new generation of protein database search programs」, Nucleic Acids Res. 25 : 3389-3402) 의 디폴트 패러미터를 사용함으로써 결정할 수 있다. Blast algorithm 은, 인터넷에서 www. ncbi. nlm. nih. gov/blast 에 액세스 함으로써도 사용할 수 있다. 또한, 상기의 Blast algorithm 에 의해 Identity (또는 Identities) 및 Positivity (또는 Positivities) 의 2 종류의 퍼센티지의 값이 계산된다. 전자는 상동성을 요구해야 할 2 종류의 아미노산 서열 사이에서 아미노산 잔기가 일치했을 경우의 값이며, 후자는 화학 구조가 유사한 아미노산 잔기도 고려한 수치이다. 본 명세서에 있어서는, 아미노산 잔기가 일치하고 있는 경우의 Identity (동일성) 의 값을 가지고 상동성의 값으로 한다.
항체의 수식물로서, 폴리에틸렌글리콜 (PEG) 등의 각종 분자와 결합한 항체를 사용할 수도 있다.
본 발명의 항체는, 또한 이들 항체와 다른 약제가 콘쥬게이트를 형성하고 있는 것 (I㎜unoconjugate) 이어도 된다. 이와 같은 항체의 예로는, 그 항체가 방사성 물질이나 약리 작용을 갖는 화합물과 결합하고 있는 것을 들 수 있다 (Nature Biotechnology (2005) 23, p. 1137-1146).
얻어진 항체는, 균일하게까지 정제할 수 있다. 항체의 분리, 정제는 통상적인 단백질에서 사용되고 있는 분리, 정제 방법을 사용하면 된다. 예를 들어 칼럼 크로마토그래피, 필터 여과, 한외 여과, 염석, 투석, 조제용 폴리아크릴아미드 겔 전기 영동, 등전점 전기 영동 등을 적절히 선택, 조합하면, 항체를 분리, 정제할 수 있지만 (Strategies for Protein Purification and Characterization : A Laboratory Course Manual, Daniel R. Marshak et al. eds., Cold Spring Harbor Laboratory Press (1996) ; Antibodies : A Laboratory Manual. Ed Harlow and David Lane, Cold Spring Harbor Laboratory (1988)), 이들에 한정되는 것은 아니다.
크로마토그래피로는, 어피니티 크로마토그래피, 이온 교환 크로마토그래피, 소수성 크로마토그래피, 겔 여과 크로마토그래피, 역상 크로마토그래피, 흡착 크로마토그래피 등을 들 수 있다.
이들 크로마토그래피는, HPLC 나 FPLC 등의 액체 크로마토그래피를 사용하여 실시할 수 있다.
어피니티 크로마토그래피에 사용하는 칼럼으로는, 프로테인 A 칼럼, 프로테인 G 칼럼을 들 수 있다.
예를 들어 프로테인 A 칼럼을 사용한 칼럼으로서, Hyper D, POROS, Sepharose F. F. (GE 헬스케어) 등을 들 수 있다.
또 항원을 고정화한 담체를 사용하여, 항원에 대한 결합성을 이용하여 항체를 정제하는 것도 가능하다.
(항 CD147 항체를 함유하는 의약)
상기 서술한 「항 CD147 항체의 제조」 의 항에 기재된 방법으로 얻어지는 항 CD147 항체 중에서, 본 발명의 항 CD147 항체를 얻을 수 있다. 이와 같이 하여 얻어진 항체는, 종양 및/또는 암의 치료 및/또는 예방제로서 사용할 수 있다. 본 발명의 항 CD147 항체는, 우수한 항종양 활성을 갖고, 종양 또는 암의 치료약으로서 유용하다. 본 발명의 항 CD147 항체는, 겜시타빈 내성 암 세포나 솔라페닙 저감수성 암 세포에 대해서도 우수한 항종양 효과를 나타내었다. 본 발명의 항 CD147 항체는, 만성 골수성 백혈병 세포에 있어서, 이마티닙보다 현저하게 강한 약효를 나타내었다.
본 발명의 항 CD147 항체 또는 그 항체를 함유하는 의약에 의해 치료될 수 있는 종양으로는, CD147 을 발현하는 종양이면 특별히 한정되지 않지만, 바람직하게는, 췌장암, 간암, 위암, 대장암, 신암, 유방암, 자궁암, 난소암, 폐암, 갑상선암, 피부암, 두경부암, 육종, 전립선암, 방광암, 뇌종양, 소화관 간질 종양 (GIST), 백혈병 (예를 들어, 급성 골수성 백혈병 (AML), 만성 골수성 백혈병 (CML), 또는 만성 림프성 백혈병 (CLL) 또는 급성 림프성 백혈병 (ALL)), 림프종 또는 악성 림프종 (예를 들어, B 세포 림프종, 비호지킨 림프종 또는 미만성 대세포형 B 세포 림프종 (diffuse large B-cell lymphoma, DLBCL)) 을 들 수 있고, 보다 바람직하게는, 췌장암, 간암, 위암, 대장암, 신암, 백혈병, 급성 골수성 백혈병 (AML), 만성 골수성 백혈병 (CML), 만성 림프성 백혈병 (CLL), 급성 림프성 백혈병 (ALL), 악성 림프종, B 세포 림프종, 비호지킨 림프종 또는 미만성 대세포형 B 세포 림프종 (DLBCL) 을 들 수 있다.
또, 본 발명의 항체 또는 그 항체를 함유하는 의약에 의해 치료될 수 있는 종양으로는, SMAD 양성의 CD147 을 발현하는 종양을 들 수 있고, SMAD 양성의 CD147 을 발현하는 종양으로는, 예를 들어, SMAD 양성의 CD147 을 발현하는 간암 또는 췌장암을 들 수 있다. 본 발명의 항 CD147 항체 또는 그 항체를 함유하는 의약은, 바람직하게는, CD147 및/또는 SMAD 의 발현이 확인된 환자에게 투여된다. SMAD 로는, 바람직하게는, SMAD2, SMAD3 및/또는 SMAD4 를 들 수 있고, 보다 바람직하게는 SMAD4 를 들 수 있다. 바람직하게는, SMAD4 의 발현 확인에 맞추어, SMAD2 또는 SMAD3 중의 적어도 어느 1 개가 발현하고 있는 것이 확인된다.
혹은, 본 발명의 항체 또는 그 항체를 함유하는 의약에 의해 치료될 수 있는 종양으로는, KLF5 의 발현이 저하 또는 결실된 종양을 들 수 있고, KLF5 의 발현이 저하 또는 결실된 종양으로는, 간암, ALL, 림프종, 소화관 간질 종양 (GIST), 피부암, 육종, AML 또는 신장암을 들 수 있다. 본 발명의 항 CD147 항체 또는 그 항체를 함유하는 의약은, 바람직하게는, KLF5 의 발현이 저하 또는 결실이 확인된 환자에게 투여된다.
본 발명의 항 CD147 항체는, 치료 목적에 따라, 2, 3 혹은 그 이상의 다른 치료제를 투여할 수도 있고, 그들의 다른 치료제는 동일한 제제 중에 봉입함으로써 동시에 투여할 수 있다. 다른 치료제와 항 CD147 항체는 동일한 제제 중에 봉입함으로써 동시에 투여할 수도 있다. 또, 항 CD147 항체와 다른 치료제를 별개의 제제에 봉입하여 동시에 투여할 수도 있다. 또한, 다른 약제와 항 CD147 항체를 잇달아 따로 따로 투여할 수도 있다. 즉, 다른 치료제를 투여한 후에 항 CD147 항체 또는 그 항체의 항원 결합 단편을 유효 성분으로서 함유하는 치료제를 투여하거나, 혹은 항 CD147 항체 또는 그 항체의 항원 결합 단편을 유효 성분으로서 함유하는 치료제를 투여한 후에 다른 치료제를 투여해도 된다.
본 발명은, 치료 및/또는 예방에 유효한 양의 항 CD147 항체와 약학상 허용되는 희석제, 담체, 가용화제, 유화제, 보존제 및/또는 보조제를 포함하는 의약 조성물도 제공한다.
본 발명은, 치료 및/또는 예방에 유효한 양의 항 CD147 항체와 치료 및/또는 예방에 유효한 양의 적어도 하나의 항종양 치료제와 약학상 허용되는 희석제, 담체, 가용화제, 유화제, 보존제 및/또는 보조제를 포함하는 의약 조성물도 제공한다.
본 발명의 의약 조성물에 있어서 허용되는 제제에 사용하는 물질로는 바람직하게는 투여량이나 투여 농도에 있어서, 의약 조성물이 투여되는 인간에 대하여 비독성의 것이 바람직하다.
본 발명의 의약 조성물은, pH, 삼투압, 점도, 투명도, 색, 등장성, 무균성, 안정성, 용해율, 서방율, 흡수율, 침투율을 바꾸거나, 유지하거나 하기 위한 제제용의 물질을 포함할 수 있다. 제제용의 물질로서 이하의 것을 들 수 있지만, 이들에 제한되지 않는다 : 글리신, 알라닌, 글루타민, 아스파라긴, 아르기닌 또는 리신 등의 아미노산류, 항균제, 아스코르브산, 황산나트륨 또는 아황산수소나트륨 등의 항산화제, 인산, 시트르산, 붕산 버퍼, 탄산수소나트륨, 트리스-염산 (Tris-Hcl) 용액 등의 완충제, 만니톨이나 글리신 등의 충전제, 에틸렌디아민4아세트산 (EDTA) 등의 킬레이트제, 카페인, 폴리비닐피롤리딘, β-시클로덱스트린이나 하이드록시프로필-β-시클로덱스트린 등의 착화제, 글루코오스, 만노오스 또는 덱스트린 등의 증량제, 단당류, 이당류 등의 다른 탄수화물, 착색제, 향미제, 희석제, 유화제나 폴리비닐피롤리딘 등의 친수 폴리머, 저분자량 폴리펩티드, 염 형성 카운터 이온, 염화벤즈알코늄, 벤조산, 살리실산, 티메로살, 페네틸알코올, 메틸파라벤, 프로필파라벤, 클로렉시딘, 소르빈산 또는 과산화수소 등의 방부제, 글리세린, 프로필렌·글리콜 또는 폴리에틸렌글리콜 등의 용매, 만니톨 또는 소르비톨 등의 당 알코올, 현탁제, 소르비탄에스테르, 폴리소르베이트 20 이나 폴리소르베이트 80 등의 폴리소르베이트, 트리톤 (triton), 트로메타민 (tromethamine), 레시틴 또는 콜레스테롤 등의 계면 활성제, 수크로오스나 소르비톨 등의 안정화 증강제, 염화나트륨, 염화칼륨이나 만니톨·소르비톨 등의 탄성 증강제, 수송제, 부형제, 및/또는 약학상의 보조제. 이들 제제용 물질의 첨가량은, 항 CD147 항체의 중량에 대하여 0.01 ∼ 100 배, 특히 0.1 ∼ 10 배 첨가하는 것이 바람직하다. 제제 중의 적합한 의약 조성물의 조성은 당업자에 의해, 적용 질환, 적용 투여 경로 등에 따라 적절히 결정할 수 있다.
의약 조성물 중의 부형제나 담체는 액체여도 되고 고체여도 된다. 적당한 부형제나 담체는 주사용의 물이나 생리 식염수, 인공 뇌척수액이나 비경구 투여에 통상적으로 사용되고 있는 다른 물질이어도 된다. 중성의 생리 식염수나 혈청 알부민을 포함하는 생리 식염수를 담체에 사용할 수도 있다. 의약 조성물에는 pH 7.0-8.5 의 Tris 버퍼, pH 4.0-5.5 의 아세트산 버퍼, pH 3.0-6.2 의 시트르산 버퍼를 포함할 수 있다. 또, 이들 버퍼에 소르비톨이나 다른 화합물을 포함할 수도 있다. 본 발명의 의약 조성물에는 항 CD147 항체를 포함하는 의약 조성물 그리고, 항 CD147 항체 및 적어도 하나의 항종양 치료제를 포함하는 의약 조성물을 들 수 있으며, 본 발명의 의약 조성물은 선택된 조성과 필요한 순도를 갖는 약제로서, 동결 건조품 혹은 액체로서 준비된다. 항 CD147 항체를 포함하는 의약 조성물 그리고, 항 CD147 항체 및 적어도 하나의 항암제 치료제를 포함하는 의약 조성물은 수크로오스와 같은 적당한 부형제를 사용한 동결 건조품으로서 성형될 수도 있다.
본 발명의 의약 조성물은 비경구 투여용으로 조제할 수도 있고, 경구에 의한 소화관 흡수용으로 조제할 수도 있다. 제제의 조성 및 농도는 투여 방법에 의해 결정할 수 있다. 본 발명의 항체를 인간에 대해 투여할 때에는, 약 0.1 ∼ 100 ㎎/㎏ 을 1 ∼ 180 일간에 1 회 또는 복수 회 투여하면 된다. 그러나, 투여량이나 투여 횟수는, 일반적으로, 환자의 성별, 체중, 연령, 증상, 위중도, 부작용 등을 고려하여 결정되어야 할 것이므로, 상기의 용량이나 용법에는 한정되지 않는 것으로 한다.
본 발명의 의약 조성물의 형태로는, 점적 (点滴) 을 포함하는 주사제, 좌제, 경비제, 설하제, 경피 흡수제 등을 들 수 있다. 투여 경로는, 경구 경로 또는 비경구 경로이며, 비경구 경로에는, 예를 들어, 정맥내, 동맥내, 근육내, 직장내, 경점막내, 피내 (皮內) 등의 경로를 들 수 있다.
본 발명의 항체 혹은 당해 항체의 항원 결합 단편, 이들을 포함하는 약물 복합체, 이들을 포함하는 바이스페시픽 항체 또는 이들을 함유하는 의약 조성물은, 이들이 투여되는 환자 선별을 위한 바이오 마커와 조합하여 제공될 수 있다. 이들의 항체나 의약 조성물은, 바이오 마커를 검출하는 수단과 조합하여 키트로서 제공되고 있어도 되고, 항체나 의약 조성물의 제공과 바이오 마커의 제공은 별개여도 된다. 바이오 마커를 사용함으로써, 본 발명의 항체나 의약 조성물은, 본 발명의 항체의 효과가 보다 높게 기대되는 환자군에게 투여될 수 있다.
본 발명은, 암환자 유래의 생물학적 시료를 사용하고, 그 생물학적 시료 중에 포함되는 SMAD4 의 발현 또는 KLF5 의 발현을 측정하고, SMAD4 가 검출된 환자 또는 KLF5 의 발현 저하 혹은 결실이 검출된 환자를, 본 발명의 항체 혹은 당해 항체의 기능성 단편 또는 본 발명의 의약 조성물에 의한 암의 치료에 대한 응답성이 있다고 판정하는 것을 포함하는, 암의 치료에 대한 응답성을 예측하는 방법 ; 암환자 유래의 생물학적 시료를 사용하고, 그 생물학적 시료 중에 있어서의 SMAD4 의 발현 또는 KLF5 의 발현을 검출하고, SMAD4 가 검출된 환자 또는 KLF5 의 발현 저하 혹은 결실이 검출된 환자를, 본 발명의 항체 혹은 당해 항체의 기능성 단편 또는 본 발명의 의약 조성물에 의한 암 치료 대상자로서 선별하는 것을 포함하는, 암의 치료 대상을 선별하는 방법 ; 암환자 유래의 생물학적 시료를 사용하고, 그 생물학적 시료 중에 포함되는 SMAD4 의 발현 또는 KLF5 의 발현을 검출하고, SMAD4 가 검출된 환자 또는 KLF5 의 발현 저하 혹은 결실이 검출된 환자에 대하여, 본 발명의 항체 혹은 당해 항체의 기능성 단편 또는 본 발명의 의약 조성물을 투여하는 것을 포함하는, 암의 치료 방법 ; 혹은, 본 발명의 항체 혹은 당해 항체의 기능성 단편 또는 본 발명의 의약 조성물에 의한 암의 치료에 대한 응답성을 판정하기 위한 키트로서, 암환자 유래의 생물학적 시료 중의 SMAD4 의 발현 또는 KLF5 의 발현을 검출하는 수단을 적어도 포함하는, 키트에 관한 것이다.
본 명세서에 있어서 「생물학적 시료」 는, 개체로부터 단리된 조직, 액체, 세포, 및 그들의 혼합물을 말하며, 예를 들어 종양 생검, 수액 (髓液), 흉강내액, 복강내액, 림프액, 피부 절편, 혈액, 요 (尿), 분변, 담, 호흡기, 장관, 요 생식기관, 타액, 젖, 소화 기관, 및 이들로부터 채취된 세포를 들 수 있지만, 이들에 한정되지 않는다. 「생물학적 시료」 는, 암 세포를 포함하는 시료인 것이 바람직하고, 보다 바람직하게는 절제나 생검에 의해 얻어진 조직이나 세포, 혹은 흉강 내액이나 복강 내액에서 유래하는 세포를 예시할 수 있다. 더욱 바람직한 생물학적 시료는, 암 세포 또는 암 조직을 포함하는 시료이다.
「SMAD4 의 발현」 의 검출이나 측정은, SMAD4 의 게놈 서열, 유전자 발현 또는 단백질 발현을 당업자에게 주지의 방법을 사용하여 실시할 수 있고, 예를 들어, RNA 시퀀스, 마이크로 어레이, 게놈 시퀀스, 이뮤노 어세이를 들 수 있다.
「KLF5 의 발현」 의 검출이나 측정은, KLF5 의 게놈 서열, 유전자 발현 또는 단백질 발현을 당업자에게 주지의 방법을 사용하여 검출할 수 있으며, 예를 들어, IHC, RNA 시퀀스, 마이크로 어레이, 게놈 시퀀스, 이뮤노 어세이를 들 수 있다. 「KLF5 의 발현 저하」 란, 대조 (예를 들어, 정상인이나 동일 환자의 비암 조직에 있어서의 발현 레벨) 와의 비교에 있어서, 그것보다 발현 레벨이 낮은 것을 말한다. 혹은, 본 발명의 항체 또는 의약 조성물에 의한 암의 치료에 대한 응답성이 있다고 판정할 수 있는 KLF5 발현의 저하의 정도는, 당업자에 의해 주지의 방법 및 적절한 임상 시험의 실시에 의해 결정될 수 있으며, 예를 들어, 효과가 얻어진 환자와 효과가 얻어지지 않은 환자에 있어서의 KLF5 의 발현량을 비교하여 적절한 임계값이 설정된다. 따라서, 「KLF5 의 발현 저하」 란, 예를 들어, 이와 같이 하여 설정된 임계값보다 아래인 것을 말한다.
실시예
이하에 나타내는 실시예에 의해 본 발명을 구체적으로 설명하는데, 본 발명은 이들에 한정되는 것은 아니다. 또, 이들은 어떠한 의미에 있어서도 한정적으로 해석되는 것은 아니다. 또한, 하기 실시예에 있어서 유전자 조작에 관한 각 조작은 특별히 명시가 없는 한, 「몰레큘러 클로닝 (Molecular Cloning)」 (Sambrook, J., Fritsch, E. F. 및 Maniatis, T. 저, Cold SpringHarbor Laboratory Press 에서 1989년 발간) 에 기재된 방법 및 그 밖의 당업자가 사용하는 실험서에 기재된 방법에 의해 실시하거나, 또는, 시판되는 시약이나 키트를 사용하는 경우에는 시판품의 지시서에 따라서 실시하였다. 또, 본 명세서에 있어서, 특별히 기재된 없는 시약, 용매 및 출발 재료는, 시판되는 공급원으로부터 용이하게 입수 가능하다. 본 실시예에 있어서, 인간 췌장주 MIA PaCa-2 는, ATCC, Cat. CRL-1420 을 사용하고, 인간 췌장암 세포주 PANC-1 은, ATCC, Cat. CRL-1469 를 사용하였다.
(실시예 1) 세포 면역에 의한 마우스·래트 항체의 제조
1)-1 CD147 발현 벡터의 제조
시판되는 인간 CD147 유전자 (BSG variant2/CD147v2) 의 클론 IOH3378 (인비트로젠사) 과 포유류 세포용 발현 벡터 pcDNA-DEST40 (인비트로젠사) 을 Gateway LR 클로나아제를 사용하여 반응시켜, 인간 CD147v2 발현 벡터 (pcDNA-DEST40-CD147v2) 를 제조하였다.
시판되는 인간 CD147 유전자 (BSG variant1/CD147v1) 의 포유류 세포용 발현 벡터 pCMV6-XL5-hBSGv1 (Origene 사, Cat. SC303059) 을 구입하고, 인간 CD147v1 발현 벡터로 하였다.
게잡이 원숭이 CD147 발현 벡터로서, pCMV3-cynoBSG (Sino Biological Inc., Cat. CG90636-UT) 를 구입하였다.
마우스 CD147v2 발현 벡터로서, pCMV3-mBSGv2 (Sino Biological Inc., Cat. MG50332-UT) 를 구입하였다.
1)-2 마우스 하이브리도마의 제조
4 ∼ 6 주령의 BALB/cAnNCrlCrlj 마우스 (닛폰 찰스·리버사) 를 사용하였다. 0 일째, 7 일째, 15 일째 및 24 일째에 베르센 (Thermo Fisher Scientific 사) 으로 벗긴 5 × 106 개의 LNCaP 세포 (ATCC, CRL-1740) 를 PBS 에 현탁하여 등부 피하에 투여하였다. 31 일째에 동일한 세포를 5 × 106 개 정맥 투여하고, 같은 날에 비장을 채취하고 하이브리도마 제조에 사용하였다. 비장 세포와 마우스 미엘로마 P3X63Ag8U.1 세포 (ATCC, CRL-1597) 를 PEG4000 (IBL 사) 을 사용하여 세포 융합하여 하이브리도마를 제조하였다. 하이브리도마의 단리, 배양에는, ClonaCell-HY Medium D (STEMCELL TECHNOLOGIES 사), ClonaCell-HY Medium E (STEMCELL TECHNOLOGIES 사) 를 사용하였다.
1)-3 래트 하이브리도마의 제조
7 주령의 WKY/Izm (닛폰 에스엘씨 주식회사) 을 사용하였다. 인간 췌장암 세포주 PANC-1 을 1 × 107 개, 둔부에 면역하고 13 일 후에 장골 림프절 세포를 채취하고 하이브리도마 제조에 사용하였다. 래트 비장 세포와 마우스 미엘로마 SP2/0-Ag14 세포 (ATCC, CRL-1581) 를 LF301 세포 융합 장치 (주식회사 벡스) 를 사용하여 세포 융합하고 하이브리도마를 제조하였다. 하이브리도마의 단리, 배양에는, ClonaCell-HY Medium D (STEMCELL TECHNOLOGIES 사), ClonaCell-HY Medium E (STEMCELL TECHNOLOGIES 사) 를 사용하였다.
1)-4 ELISA 에 의한 항원 동정
인간 CD147Fc 융합 단백질 (Sino Biological 사, Cat. 10186-H02H) 과 마우스 CD147Fc 융합 단백질 (Sino Biological 사, Cat. 50332-M03H) 을 사용하였다. 인간 CD147Fc 융합 단백질과 마우스 CD147Fc 융합 단백질은, PBS 완충액을 첨가하여, 빙상 (氷上) 에서 용해하고, 1 ㎍/㎖ 로 조제하였다. 동 (同) 단백질 용해액을 96 웰 플레이트 (NUNC 사, Cat. 442404) 에 100 ㎕ 첨가하고, 4 ℃ 하룻밤 보존하고, 웰을 CD147Fc 융합 단백질로 코트하였다. 단백질 용해액을 제거하고, 1 % BSA (Research Organics 사, Cat. 1334A) 를 포함하는 PBS 완충액으로 웰을 4 ℃, 2 시간 블로킹하였다. 0.05 % 트윈20 (ATTO 사, Cat. WSE-7235) 을 포함하는 PBS 완충액으로 웰을 3 회 세정한 후, 실시예 1)-2, 1)-3 에서 조제한 하이브리도마 배양 상청을 PBS 완충액으로 20 배로 희석하고 각 웰에 첨가하고 실온에서 1 시간 가온하였다. 0.05 % 트윈20 (ATTO 사, Cat. WSE-7235) 을 포함하는 PBS 완충액으로 웰을 3 회 세정한 후, 1 % BSA 를 포함하는 PBS 완충액으로 50000 배로 희석한 항 래트-Fab2-igG-HRP (Jackson I㎜unoResearch 사, Cat. 112-036-072) 를 100 ㎕ 첨가하고 30 분간, 실온에서 흔들었다. 0.05 % 트윈20 (ATTO 사, Cat. WSE-7235) 을 포함하는 PBS 완충액으로 웰을 5 회 세정한 후, 100 ㎕ 의 HRP 효소 발색 시약 (eBioscience 사, Super AquaBlue ELISA substrate, Cat. 00-4203) 을 첨가하고, 10 ∼ 20 분 실온에서 가온하고, 플레이트 리더 (Envision, 퍼킨엘머사) 로 405 ㎚ 의 흡광도를 측정하였다. 2 ∼ 3 웰의 흡광도의 측정값에 대해 평균값을 산출하고, 1 차 항체 없는 대조 웰에서의 측정값의 2 배 이상의 흡광도가 관찰된 항체에 대해 결합성 있음 (+), 2 배에 미치지 않는 것을 결합성 없음 (-) 으로 판정하고 표 1 에 정리하였다. LN22R8, 2P1A6, 2P3A9, 2P3G8, 2P8C12, 2P10F2, 2P2D7, 2P2D10, 2P1B7 의 배양 상청에 대해, 인간 CD147Fc 융합 단백질 코트 웰 특이적인 발색을 확인하였다. LN24R7, 2P5F5, 2P6A2, 2P3G8 에 대해서는, 인간 및 마우스 CD147Fc 융합 단백질 코트 웰 특이적인 발색을 확인하였다.
1)-5 모노클로날 항체의 조제와 항체 아이소타입의 결정
실시예 1)-4 에서 항인간 CD147 항체 산생이 확인된 하이브리도마에서, 안정적으로 배양 가능한 것에 대해, 시판되는 Isotyping 키트를 사용하여, 배양 상청에 포함되는 항체의 아이소타입을 결정하고 표 2 에 나타내었다. CL-1000 플라스크 (닛폰 벡톤·디킨슨 주식회사) 를 사용하여, 이들 하이브리도마를 배양하고, 모노클로날 항체를 포함하는 하이브리도마 배양 상청을 조제하였다.
1)-6 모노클로날 항체의 정제
실시예 1)-5 에서 제조한 배양 상청으로부터 항체를 정제하였다. 항인간 CD147 마우스 모노클로날 항체에 대해서는, rProtein A 어피니티 크로마토그래피 (4 ∼ 6 ℃ 하) 1 단계 공정으로 정제하였다. rProtein A 어피니티 크로마토그래피 정제 후의 버퍼 치환 공정은 4 ∼ 6 ℃ 하에서 실시하였다. 최초로 배양 상청을 PBS 로 평형화한 MabSelectSuRe (GE Healthcare Bioscience 사 제조) 가 충전된 칼럼에 어플라이하였다. 배양액이 칼럼에 모두 들어간 후, 칼럼 용량 2 배 이상의 PBS 로 칼럼을 세정하였다. 다음으로 2M 아르기닌염산염 용액 (pH 4.0) 으로 용출하고, 항체가 포함되는 획분을 모았다. 그 획분을 투석 (Thermo Scientific 사, Slide-A-Lyzer Dialysis Cassette) 에 의해 HBSor (25 mM Histidine/5 % Sorbitol/pH 6.0) 으로의 액 치환을 실시하였다. Centrifugal UF Filter Device VIVASPIN20 (분획 분자량 UF10K, Sartorius 사, 4 ℃ 하) 으로 농축하고, IgG 농도를 4.9 ㎎/㎖ 로 조제하였다. 마지막으로 Minisart-Plus filter (Sartorius 사) 로 여과하고, 정제 샘플로 하였다.
항인간 CD147 래트 모노클로날 항체에 대해서는, Protein G 어피니티 크로마토그래피 (4 ∼ 6 ℃ 하) 1 단계 공정으로 정제하였다. Protein G 어피니티 크로마토그래피 정제 후의 버퍼 치환 공정은 4 ∼ 6 ℃ 하에서 실시하였다. 최초로, PBS 로 평형화한 ProteinG (GE Healthcare Bioscience 사) 가 충전된 칼럼에 하이브리도마의 배양 상청을 어플라이하였다. 배양 상청액이 칼럼에 모두 들어간 후, 칼럼 용량 2 배 이상의 PBS 로 칼럼을 세정하였다. 다음으로 0.1 M 글리신/염산 수용액 (pH 2.7) 으로 용출하고, 항체가 포함되는 획분을 모았다. 모은 획분에 1M Tris-HCl (pH 9.0) 을 첨가하여 pH 7.0 ∼ 7.5 로 조정한 후에, Centrifugal UF Filter Device VIVASPIN20 (분획 분자량 UF30K, Sartorius 사, 4 ∼ 6 ℃ 하) 으로 HBSor (25 mM Histidine/5 % Sorbitol/pH 6.0) 로의 버퍼 치환을 실시함과 함께 농축을 실시하고, 항체 농도를 1 ㎎/㎖ 이상으로 조제하였다. 마지막으로 Minisart-Plus filter (Sartorius 사) 로 여과하고, 정제 샘플로 하였다.
1)-7 in vivo 항종양 활성 측정에 의한 항체 스크리닝
1 × 107 개의 인간 췌장주 PANC-1 을 PBS 로 현탁하고, NOD-scid 마우스 (닛폰 찰스·리버사, NOD. CB17-Prkdc<scid>/J) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 군나누기를 하고, 이식 27, 34, 41 일 후에 마우스 항 CD147 항체 (LN22R8), 래트 항 CD147 항체 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2) 를 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 6). 이식 27, 34 일 후에 래트 항 CD147 항체 (2P2D6) 를 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 6). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 1(a) ∼ (c) 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다. 2P2D6 항체, 2P3G8 항체 및 2P2D10 항체의 결과를 도 1(a) 에 나타내었다. 최종 측정일인 이식 48 일 후에 있어서의 종양 증식 억제율은, 10 ㎎/㎏ 투여군에서, 각각, 10 %, 45 % 및 40 % 였다. LN22R8 항체, 2P1A6 항체 및 2P1B7 항체의 결과를 도 1(b) 에 나타내었다. 최종 측정일인 이식 48 일 후에 있어서의 종양 증식 억제율은, 10 ㎎/㎏ 투여군에서, 각각, 50 %, 26 % 및 24 % 였다. 2P8C12 항체 및 2P10F2 항체의 결과를 도 1(c) 에 나타내었다. 최종 측정일인 이식 48 일 후에 있어서의 종양 증식 억제율은, 10 ㎎/㎏ 투여군에서, 각각, ―2 % 및 62 % 였다.
1)-8 CD147 항체의 종 교차성의 해석
CHO-K1 세포 (ATCC, CCL-61) 에 Lipofectamine 2000 (Thermofishers scientific 사, Cat. 11668-019) 을 사용하여, 실시예 1)-1 에서 제조한 pcDNA-DEST40-CD147v2, 또는 pCMV3-cynoBSG 를 도입하고, 1 일 후에 마우스 항인간 CD147 항체 (LN22R8), 또는 래트 항인간 CD147 항체 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2, 2P2D6), 10 ㎍/㎖ 로 처리하고, 항마우스 IgG-FITC (MP Biomedical 사, Cat. 554936) 또는, 항래트 IgG-PE (BD Biosciences 사, Cat. 550767) 를 사용하여 각 항체의 CD147 발현 CHO-K1 세포에 대한 결합을 형광 검출 가능하게 하였다. CHO-K1 세포의 인간과 게잡이 원숭이 CD147 발현은, 시판되는 항 CD147 항체 (MEM-M6/1, Ab serotec 사, Cat. MCA28822) 의 결합에 의해 형광 검출 가능하게 하였다. 상기의 세포에 대하여, 플로우 사이토미터 (CantoII, BD Bioscience 사) 의 측정을 실시하고, 도 2a ∼ 2c 에 결과를 정리하였다. 도면의 세로축은 세포수를 나타내고, 가로축은 형광 시그널의 강도를 나타낸다.
시판되는 항 CD147 항체 (MEM-M6/1), 마우스 항인간 CD147 항체 (LN22R8), 래트 항 CD147 항체 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2, 2P2D6), 어느 항 CD147 항체도 인간 CD147 발현 CHO-K1 세포에 결합성을 나타냈다 (도 2a ∼ 2c).
시판되는 항 CD147 항체 (MEM-M6/1) 는 게잡이 원숭이 CD147 발현 CHO-K1 세포에 결합성을 나타냈지만, 마우스 항인간 CD147 항체 (LN22R8), 래트 항인간 CD147 항체 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2, 2P2D6), 어느 CD147 항체도 게잡이 원숭이 CD147 발현 CHO-K1 세포에는 결합성을 나타내지 않았다 (도 2a ∼ 2c).
또한, 어느 항 CD147 항체도 마우스 CD147 발현 CHO-K1 세포에는 결합성을 나타내지 않았다 (데이터 미기재).
1)-9 에피토프 해석
에피토프 해석용의 인간 CD147 변이체 발현 벡터의 제조
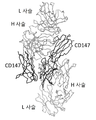
게잡이 원숭이와 인간의 CD147 의 아미노산 서열은, BLAST 검색 (https : /blast. ncbi. nlm. nih. gov/Blast. cgi) 의 결과, 81 % 가 동일하고, 한정된 아미노산의 차이가, CD147 항체 결합성의 에피토프 인식에 직접 영향을 미치고 있는 것으로 상정하고, 종간에서 상이한 아미노산 서열을 부분적으로 이식한 변이체를 사용한 항종양 에피토프의 추정을 진행시키기로 하였다. hCD147v1, v2 공통으로 포함되는 아미노산 서열을 대상으로 하여, 게잡이 원숭이와 인간의 CD147 에서의 서열 비교를 실시하고, 종간에서 상이한 아미노산 영역의 9 영역으로 분류하고, mu1 ∼ mu9 로 하였다 (도 3). CD147 변이체 발현 벡터 제조와 세포막 상에 대한 발현 확인용으로 N 말단에 FLAG 서열을 도입한 인간 CD147variant2 의 cDNA 서열을 인공 합성하고, pcDNA3.1 벡터에 도입한 플라스미드, Signal-N-Flag-hCD147v2_pcDNA3.1 (Genscript 사에서 제조) 을 제조하였다. 또한 동 (同) 플라스미드의 인간 CD147 유전자에 게잡이 원숭이의 CD147 아미노산 서열 mu1 ∼ mu9 에 대해, DNA 치환에 의한 아미노산 치환 변이로서 도입한 인간-게잡이 원숭이 키메라 CD147 발현 벡터 9 종, hCD147-mu1_pcDNA3.1, hCD147-mu2_pcDNA3.1, hCD147-mu3_pcDNA3.1, hCD147-mu4_pcDNA3.1, hCD147-mu5_pcDNA3.1, hCD147-mu6_pcDNA3.1, hCD147-mu7_pcDNA3.1, hCD147-mu8_pcDNA3.1, hCD147-mu9_pcDNA3.1 제조하였다 (Genscript 사에서 제조).
1)-10 변이체를 사용한 항종양 에피토프 영역의 특정
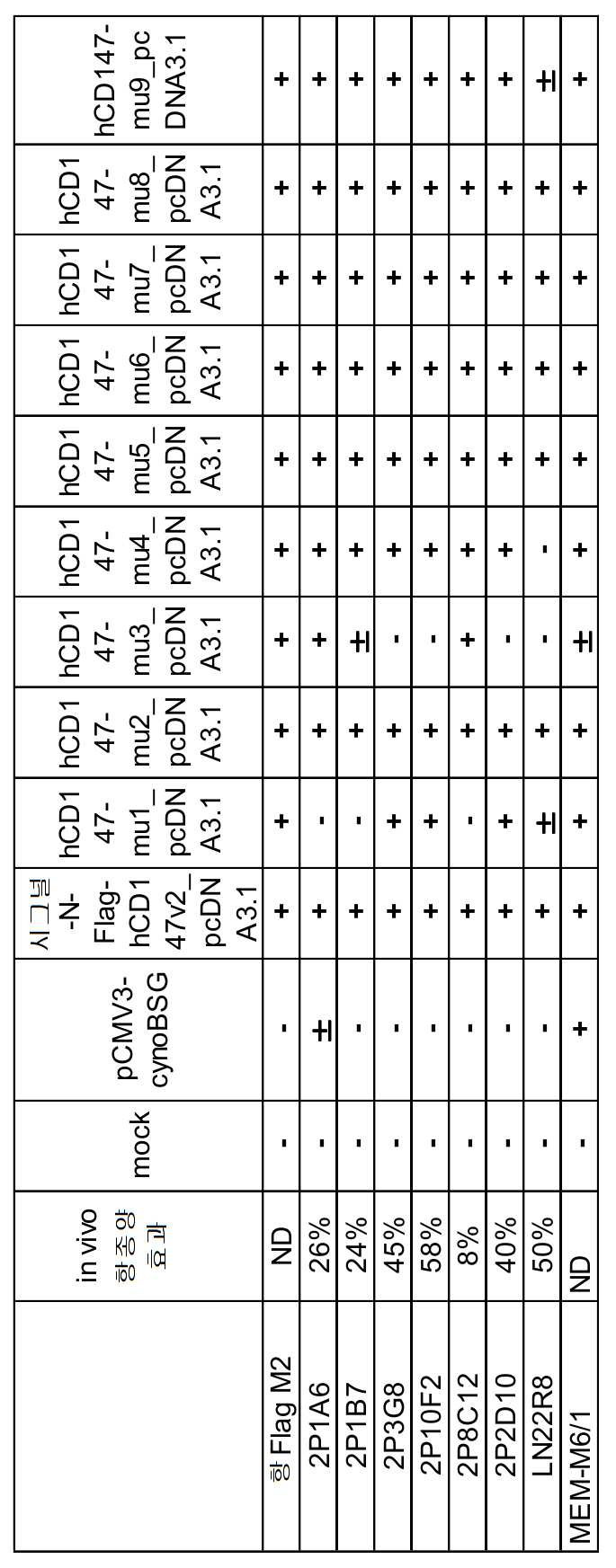
인간 CD147, 게잡이 원숭이 CD147 또는, 인간-게잡이 원숭이 키메라 CD147 발현 벡터 9 종의 발현 벡터를 CHO-K1 세포 (ATCC, CCL-61) 에 Lipofectamine 2000 (서모 사이언티픽사, Cat. 11668-019) 을 사용하여, 도입하고, 1 일 후에 항인간 CD147 마우스 항체 (LN22R8), 래트 항인간 CD147 항체 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2, 2P2D6), 10 ㎍/㎖ 로 처리하고, 항마우스 IgG-PE (DAKO사, Cat. R480), 항래트 IgG-PE (BD 사, #550767) 를 사용하여 항 CD147 항체의 CD147 발현 CHO-K1 세포에 대한 결합을 조사하였다. CD147 단백질의 발현은, 시판되는 항FLAG 항체 (항 Flag M2, SIGMA 사, Cat. F4049-.2MG) 를 사용하여 확인하였다. 플로우 사이토미터 (CantoII, BD Bioscience 사) 의 측정을 실시하고, 표 3 에 결과를 정리하였다. 1 차 항체 미처리의 대조 세포와 비교하여, 10 배 이상의 형광 시그널의 증가가 확인된 샘플에 대해, 결합 양성 (+) 으로 판정하였다. 10 배 미만의 부분적인 형광 시그널의 증가가 확인된 샘플에 대해, 결합약 양성 (±) 으로 판정하였다. 1 차 항체 미처리의 대조 세포와 비교하여, 형광 시그널의 증가가 확인되지 않은 샘플에 대해, 결합 음성 (―) 으로 판정하였다.
실시예 1)-7 에서 40 % 이상의 항종양 효과가 관찰된 항체 2P3G8, 2P10F2, 2P2D10, LN22R8 의 항체는, 모두 mu3 의 변이를 갖는 CD147 에 대한 결합성이 소실되어 있었다. 항종양 효과에 중요한 에프토프는 m3 영역인 것이 시사되었다.
1)-11 LN22R8 과 2P10F2 항체의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 결정
1)-11-1 LN22R8 항체의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 결정
1)-11-1-1 LN22R8 항체 생산 하이브리도마의 총 RNA 의 조제
LN22R8 항체의 가변 영역을 코드하는 cDNA 를 증폭하기 위해서, LN22R8 항체 산생 하이브리도마로부터 TRIzol Reagent (Ambion 사) 를 사용하여 총 RNA 를 조제하였다.
1)-11-1-2 5'-RACE PCR 에 의한 LN22R8 항체의 경쇄 가변 영역을 코드하는 cDNA 의 증폭과 서열의 결정
경쇄 가변 영역을 코드하는 cDNA 의 증폭은, 실시예 1)-11-1-1 에서 조제한 총 RNA 의 약 1 ㎍ 과 SMARTer RACE 5'/3' Kit (Clontech 사) 를 사용하여 실시하였다. LN22R8 항체의 경쇄 유전자의 가변 영역을 코드하는 cDNA 를 PCR 로 증폭하기 위한 프라이머로서, UPM (Universal Primer A Mix : SMARTer RACE 5'/3' Kit 에 부속), 및 공지된 마우스 경쇄의 정상 영역의 서열로부터 설계한 프라이머를 사용하였다.
5'-RACE PCR 로 증폭한 경쇄의 가변 영역을 코드하는 cDNA 를 플라스미드에 클로닝하고, 다음으로 경쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 시퀀스 해석을 실시하였다.
결정된 LN22R8 항체의 경쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 7 에 나타내고, 아미노산 서열을 서열 번호 8 에 나타낸다. LN22R8 항체의 경쇄 가변 영역의 CDRL1, CDRL2 및 CDRL3 을, 각각, 서열 번호 11, 12 및 13 에 나타낸다.
1)-11-1-3 5'-RACE PCR 에 의한 LN22R8 항체의 중쇄 가변 영역을 코드하는 cDNA 의 증폭과 서열의 결정
중쇄 가변 영역을 코드하는 cDNA 의 증폭은, 실시예 1)-11-1-1 에서 조제한 총 RNA 의 약 1 ㎍ 과 SMARTer RACE 5'/3' Kit (Clontech 사) 를 사용하여 실시하였다. LN22R8 항체의 중쇄 유전자의 가변 영역을 코드하는 cDNA 를 PCR 로 증폭하기 위한 프라이머로서, UPM (Universal Primer A Mix : SMARTer RACE 5'/3' Kit 에 부속), 및 공지된 마우스 중쇄의 정상 영역의 서열로부터 설계한 프라이머를 사용하였다.
5'-RACE PCR 로 증폭한 중쇄의 가변 영역을 코드하는 cDNA 를 플라스미드에 클로닝하고, 다음으로 중쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 시퀀스 해석을 실시하였다.
결정된 LN22R8 항체의 중쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 9 에 나타내고, 아미노산 서열을 서열 번호 10 에 나타낸다. LN22R8 항체의 중쇄 가변 영역의 CDRH1, CDRH2 및 CDRH3 을, 각각, 서열 번호 14, 15 및 16 에 나타낸다.
1)-11-2 2P10F2 항체의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 결정
실시예 1)-11-1 과 동일한 방법으로 실시하였다. 단, 경쇄 유전자의 가변 영역을 코드하는 cDNA 를 PCR 로 증폭하기 위한 프라이머로서, UPM (Universal Primer A Mix : SMARTer RACE 5'/3' Kit 에 부속), 및 공지된 래트 경쇄의 정상 영역의 서열로부터 설계한 프라이머를 사용하고, 중쇄 유전자의 가변 영역을 코드하는 cDNA 를 PCR 로 증폭하기 위한 프라이머로서, UPM (Universal Primer A Mix : SMARTer RACE 5'/3' Kit 에 부속), 및 공지된 래트 중쇄의 정상 영역의 서열로부터 설계한 프라이머를 사용하였다.
결정된 2P10F2 항체의 경쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 17 에 나타내고, 아미노산 서열을 서열 번호 18 에 나타낸다. 2P10F2 항체의 경쇄 가변 영역의 CDRL1, CDRL2 및 CDRL3 을, 각각, 서열 번호 21, 22 및 23 에 나타낸다. 중쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 19 에 나타내고, 아미노산 서열을 서열 번호 20 에 나타낸다. 2P10F2 항체의 중쇄 가변 영역의 CDRH1, CDRH2 및 CDRH3 을, 각각, 서열 번호 24, 25 및 26 에 나타낸다.
1)-12 LN22R8 의 인간 키메라 항체 발현 벡터의 제조
1)-12-1 인간 키메라 및 인간화 경쇄 발현 벡터 pCMA-LK 의 구축
플라스미드 pcDNA3.3-TOPO/LacZ (Invitrogen 사) 를 제한 효소 XbaI 및 PmeI로 소화하여 얻어지는 약 5.4 kb 의 프래그먼트와, 서열 번호 27 에 나타내는 인간 경쇄 시그널 서열 및 인간 κ 사슬 정상 영역을 코드하는 DNA 서열을 포함하는 DNA 단편을 In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여 결합하여, pcDNA3.3/LK 를 제조하였다.
pcDNA3.3/LK 로부터 네오마이신 발현 유닛을 제거함으로써 pCMA-LK 를 구축하였다.
1)-12-2 인간 키메라 및 인간화 IgG1 타입 중쇄 발현 벡터 pCMA-G1 의 구축
pCMA-LK 를 XbaI 및 PmeI 로 소화하여 경쇄 시그널 서열 및 인간 κ 사슬 정상 영역을 없앤 DNA 단편과, 서열 번호 28 로 나타내는 인간 중쇄 시그널 서열 및 인간 IgG1 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편을 In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여 결합하여, pCMA-G1 을 구축하였다.
1)-12-3 인간 키메라 및 인간화 IgG2 타입 중쇄 발현 벡터 pCMA-G2 의 구축
서열 번호 29 로 나타내는 인간 중쇄 시그널 서열 및 인간 IgG2 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편을 사용하여, 실시예 1)-12-2 와 동일한 방법으로 pCMA-G2 를 구축하였다.
1)-12-4 인간 키메라 LN22R8 의 경쇄 발현 벡터의 구축
실시예 1)-11-1-2 에서 얻어진 LN22R8 의 경쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로 하여, In-fusion 클로닝용으로 설계한 프라이머로 PCR 을 실시함으로써 경쇄의 가변 영역을 코드하는 cDNA 를 포함하는 DNA 단편을 증폭하였다. pCMA-LK 를 제한 효소 BsiWI 로 절단한 지점에, In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, 증폭한 DNA 단편을 삽입함으로써 인간 키메라 LN22R8 의 경쇄 발현 벡터를 구축하였다. 인간 키메라 LN22R8 의 경쇄의 뉴클레오티드 서열 및 그 경쇄의 아미노산 서열을, 서열 번호 30 및 서열 번호 31 에 각각 나타낸다.
1)-12-5 인간 키메라 LN22R8 의 IgG1 타입 중쇄 발현 벡터의 구축
실시예 1)-11-1-3 에서 얻어진 LN22R8 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로 하여, In-fusion 클로닝용으로 설계한 프라이머로 PCR 을 실시함으로써 중쇄의 가변 영역을 코드하는 cDNA 를 포함하는 DNA 단편을 증폭하였다. pCMA-G1 을 제한 효소 BlpI 로 절단한 지점에, In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, 증폭한 DNA 단편을 삽입함으로써 인간 키메라 LN22R8 의 IgG1 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 LN22R8 의 IgG1 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 32 및 서열 번호 33 에 각각 나타낸다.
1)-12-6 인간 키메라 LN22R8 의 IgG2 타입 중쇄 발현 벡터의 구축
실시예 1)-11-1-3 에서 얻어진 LN22R8 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로 하여, In-fusion 클로닝용으로 설계한 프라이머로 PCR 을 실시함으로써 중쇄의 가변 영역을 코드하는 cDNA 를 포함하는 DNA 단편을 증폭하였다. pCMA-G2 를 제한 효소 BlpI 로 절단한 지점에, In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, 증폭한 DNA 단편을 삽입함으로써 인간 키메라 LN22R8 의 IgG2 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 LN22R8 의 IgG2 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 34 및 서열 번호 35 에 각각 나타낸다.
1)-12-7 인간 키메라 LN22R8 의 IgG4P 타입 중쇄 발현 벡터의 구축
서열 번호 36 에 나타내는 인간 키메라 LN22R8 의 IgG4P 타입 중쇄의 아미노산 서열을 코드하는 DNA 서열을 포함하는 DNA 단편을 합성하였다 (GENEART 사). 합성한 DNA 단편을 사용하여, 실시예 1)-12-2 와 동일한 방법으로 인간 키메라 LN22R8 의 IgG4P 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 LN22R8 의 IgG4P 타입 중쇄의 아미노산 서열을 서열 번호 37 에 나타낸다.
1)-13 2P10F2 의 인간 키메라 항체 발현 벡터의 제조
1)-13-1 인간 키메라 및 인간화 IgG1LALA 타입 중쇄 발현 벡터 pCMA-G1LALA 의 구축
서열 번호 38 로 나타내는 인간 중쇄 시그널 서열 및 인간 IgG1LALA 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편을 사용하여, 실시예 1)-12-2 와 동일한 방법으로 pCMA-G1LALA 를 구축하였다.
1)-13-2 인간 키메라 및 인간화 IgG4P 타입 중쇄 발현 벡터 pCMA-G4P 의 구축
서열 번호 39 로 나타내는 인간 중쇄 시그널 서열 및 인간 IgG4P 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편을 사용하여, 실시예 1)-12-2 와 동일한 방법으로 pCMA-G4P 를 구축하였다.
1)-13-3 인간 키메라 2P10F2 의 경쇄 발현 벡터의 구축
실시예 1)-11-2 에서 얻어진 2P10F2 의 경쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-4 와 동일한 방법으로 인간 키메라 2P10F2 의 경쇄 발현 벡터를 구축하였다. 인간 키메라 2P10F2 의 경쇄의 뉴클레오티드 서열 및 그 경쇄의 아미노산 서열을, 서열 번호 40 및 서열 번호 41 에 각각 나타낸다.
1)-13-4 인간 키메라 2P10F2 의 IgG1LALA 타입 중쇄 발현 벡터의 구축
실시예 1)-11-2 에서 얻어진 2P10F2 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, In-fusion 클로닝용으로 설계한 프라이머로 PCR 을 실시함으로써 중쇄의 가변 영역을 코드하는 cDNA 를 포함하는 DNA 단편을 증폭하였다. pCMA-G1LALA 를 제한 효소 BlpI 로 절단한 지점에, In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, 증폭한 DNA 단편을 삽입함으로써 인간 키메라 2P10F2 의 IgG1LALA 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 2P10F2 의 IgG1LALA 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 42 및 서열 번호 43 에 각각 나타낸다.
1)-13-5 인간 키메라 2P10F2 의 IgG2 타입 중쇄 발현 벡터의 구축
실시예 1)-11-2 에서 얻어진 2P10F2 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-6 과 동일한 방법으로 인간 키메라 2P10F2 의 IgG2 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 2P10F2 의 IgG2 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 44 및 서열 번호 45 에 각각 나타낸다.
1)-13-6 인간 키메라 2P10F2 의 IgG4P 타입 중쇄 발현 벡터의 구축
실시예 1)-11-2 에서 얻어진 2P10F2 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, In-fusion 클로닝용으로 설계한 프라이머로 PCR 을 실시함으로써 중쇄의 가변 영역을 코드하는 cDNA 를 포함하는 DNA 단편을 증폭하였다. pCMA-G4P 를 제한 효소 BlpI 로 절단한 지점에, In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, 증폭한 DNA 단편을 삽입함으로써 인간 키메라 2P10F2 의 IgG4P 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 2P10F2 의 IgG4P 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 46 및 서열 번호 47 에 각각 나타낸다.
1)-14 LN22R8, 2P10F2 의 인간 키메라 항체의 생산과 조제
1)-14-1 LN22R8, 2P10F2 의 인간 키메라 항체의 생산
FreeStyle 293F 세포 (Invitrogen 사) 는 매뉴얼에 따라서, 계대, 배양을 실시하였다. 대수 증식기의 1.2 × 109 개의 FreeStyle 293F 세포 (Invitrogen 사) 를 3 L Fernbach Erle㎚eyer Flask (CORNING 사) 에 파종하고, FreeStyle293 expression medium (Invitrogen 사) 으로 희석하여 2.0 × 106 세포/㎖ 로 조제하였다. 40 ㎖ 의 Opti-Pro SFM 배지 (Invitrogen 사) 에 0.24 ㎎ 의 중쇄 발현 벡터와 0.36 ㎎ 의 경쇄 발현 벡터와 1.8 ㎎ 의 Polyethyleneimine (Polyscience #24765) 을 첨가하여 완만하게 교반하고, 또한 5 분간 방치한 후에 FreeStyle 293F 세포에 첨가하였다. 37 ℃, 8 % CO2 인큐베이터로 4 시간, 90 rpm 으로 진탕 배양 후에 600 ㎖ 의 EX-CELL VPRO 배지 (SAFC Biosciences 사), 18 ㎖ 의 GlutaMAX I (GIBCO 사), 및 30 ㎖ 의 Yeastolate Ultrafiltrate (GIBCO 사) 를 첨가하고, 37 ℃, 8 % CO2 인큐베이터로 7 일간, 90 rpm 으로 진탕 배양하여 얻어진 배양 상청을 Disposable Capsule Filter (Advantec #CCS-045-E1H) 로 여과하였다.
인간 키메라 LN22R8 의 IgG1 타입 중쇄 발현 벡터와 인간 키메라 LN22R8 의 경쇄 발현 벡터의 조합으로 취득한 LN22R8 의 인간 키메라 항체를 「LN22R8chIgG1」 이라고 명명하였다. 인간 키메라 LN22R8 의 IgG2 타입 중쇄 발현 벡터와 인간 키메라 LN22R8 의 경쇄 발현 벡터의 조합으로 취득한 LN22R8 의 인간 키메라 항체를 「LN22R8chIgG2」 라고 명명하였다. 인간 키메라 LN22R8 의 IgG4P 타입 중쇄 발현 벡터와 인간 키메라 LN22R8 의 경쇄 발현 벡터의 조합으로 취득한 LN22R8 의 인간 키메라 항체를 「LN22R8chIgG4P」 라고 명명하였다. 인간 키메라 2P10F2 의 IgG1LALA 타입 중쇄 발현 벡터와 인간 키메라 2P10F2 의 경쇄 발현 벡터의 조합으로 취득한 L2P10F2 의 인간 키메라 항체를 「2P10F2chIgG1LALA」 라고 명명하였다. 인간 키메라 2P10F2 의 IgG2 타입 중쇄 발현 벡터와 인간 키메라 2P10F2 의 경쇄 발현 벡터의 조합으로 취득한 L2P10F2 의 인간 키메라 항체를 「2P10F2chIgG2」 라고 명명하였다. 인간 키메라 2P10F2 의 IgG4P 타입 중쇄 발현 벡터와 인간 키메라 2P10F2 의 경쇄 발현 벡터의 조합으로 취득한 L2P10F2 의 인간 키메라 항체를 「2P10F2chIgG4P」 라고 명명하였다.
1)-14-2 LN22R8, 2P10F2 의 인간 키메라 항체의 정제
실시예 1)-14-1 에서 얻어진 배양 상청으로부터 항체를 rProtein A 어피니티 크로마토그래피의 1 단계 공정으로 정제하였다. 배양 상청을 PBS 로 평형화한 MabSelectSuRe 가 충전된 칼럼 (GE Healthcare Bioscience 사 제조) 에 어플라이한 후에, 칼럼 용량의 2 배 이상의 PBS 로 칼럼을 세정하였다. 다음으로 2M 아르기닌염산염 용액 (pH 4.0) 으로 용출하고, 항체가 포함되는 획분을 모았다. 그 획분을 투석 (Thermo Scientific 사, Slide-A-Lyzer Dialysis Cassette) 에 의해 HBSor (25 mM 히스티딘/5 % 소르비톨, pH 6.0) 로의 버퍼 치환을 실시하였다. Centrifugal UF Filter Device VIVASPIN20 (분획 분자량 UF10K, Sartorius 사) 으로 항체를 농축하고, IgG 농도를 1 ㎎/㎖ 이상으로 조제하였다. 마지막으로 Minisart-Plus filter (Sartorius 사) 로 여과하고, 정제 샘플로 하였다.
1)-15 인간 키메라 항체의 ADCC 활성
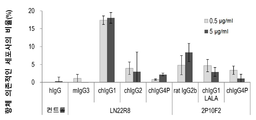
인간 키메라 항체의 ADCC 활성에 대해, 이펙터 세포로서 인간의 말초혈 단핵구 (PBMC), ADCC 표적 세포로서 인간 췌장주 MIA PaCa-2 를 사용하여 평가하였다. 방사선 동위체 51Cr 로 표지한 MIA PaCa-2 세포와 마우스 항체 (LN22R8), 래트 항체 (2P10F2), 또는, 인간 키메라 항체 (LN22R8chIgG1, LN22R8chIgG2, LN22R8chIgG4P, 2P10F2chIgG1LALA, 2P10F2chIgG4P) 를 0.5 또는, 5 ㎍/㎖ 의 농도로 4 ℃ 30 분간 처리한 후, 인간의 말초혈로부터 분리한 PBMC 를 MIA PaCa-2 세포의 20 배의 비율로 첨가하고, 4 시간, 37 도, 5 % CO2 존재하에서 배양하였다. 상청 중에 방출된 51Cr 을 TopCount NXT v2.53 을 사용하여 측정하고 총 방출 (Total release) 값을 얻었다. 51Cr 로 표지한 MIA PaCa-2 세포를 트리톤-100 으로 처리하여 방출된 51Cr 의 측정값을 최대 방출 (Maximum release) 값, PBMC 를 첨가하지 않는 항체 처리 세포로부터 처리하여 방출된 51Cr 의 측정값을 자연 방출 (Spontaneous release) 값으로서, 하기의 식으로부터, % 특이적 방출 (% specific release) 을 산출하고, 도 6 에 정리하였다. 음성 대조 샘플로서, 인간 IgG (hIgG, ChromPure Human IgG, Jackson I㎜unoResearch Laboratories 사, Cat. 009-000-003) 를 처리한 샘플에 대해 동일하게 측정을 실시하고, 아울러 나타내었다. 측정은 삼중으로 실시하고, 평균값, 표준 편차를 산출하여 아울러 나타내었다.
% 특이적 방출 = (총 방출 ― 자연 방출) / 최대 방출
인간 IgG (hIgG) 와 LN22R8 의 마우스 항체는, ADCC 활성을 나타내지 않았던 데 반해, LN22R8chIgG1 은 0.5 ㎍/㎖ 에서 17.4 %, 5 ㎍/㎖ 에서 18.1 % 로 ADCC 활성을 나타내었다. LN22R8chIgG2, LN22R8chIgG4P 의 ADCC 활성은, LN22R8chIgG1 보다 낮고, 5 ㎍/㎖ 에서도 각각 3.0 %, 2.2 % 였다.
2P10F2 래트 항체는, 0.5 ㎍/㎖ 에서 4.8 %, 5 ㎍/㎖ 에서 8.4 % 로 ADCC 활성을 나타내었다. 2P10F2chIgG1LALA 는, 0.5 ㎍/㎖ 에서 4.7 %, 5 ㎍/㎖ 에서 2.9 % 로 ADCC 활성을 나타내었다. 2P10F2chIgG4P 는, 0.5 ㎍/㎖ 에서 3.4 %, 5 ㎍/㎖ 에서 1.1 % 로 2P10F2 래트 항체, 2P10F2chIgG1LALA 보다 낮은 ADCC 활성을 나타내었다. 문헌 (Bruggemann et al., J. Exp. Med., 1351-1361, 1987) 보고와 같이, IgG1 서브타입을 이용한 인간 키메라 항체가 가장 높은 ADCC 활성을 나타내었다.
1)-16 인간 키메라 항체의 CDC 활성
항인간 CD147 항체에 의한 보체 의존적인 살세포 활성 (CDC 활성) 을 표적 세포로서 인간 췌장주 MIA PaCa-2 를 사용하여 평가하였다. 보체로서 시판되는 토끼 보체 (Low Tox-M Rabbit Complement, CEDARLANE LABORATORIES LIMITED, Cat. CL3051) 를 사용하였다. 항인간 CD147 항체로서, 마우스 항체 (LN22R8), 래트 항체 (2P10F2), 또는, 인간 키메라 항체 (LN22R8chIgG1, LN22R8chIgG2, LN22R8chIgG4P, 2P10F2chIgG1LALA, 2P10F2chIgG4P) 를 사용하였다. CDC 활성 음성의 대조 항체로서 인간 IgG (hIgG, ChromPure Human IgG, Jackson I㎜unoResearch Laboratories 사, Cat. 009-000-003) 를 사용하였다. 상기 항체를 0, 0.1, 1 또는, 10 ㎍/㎖ 의 농도로 1 시간, 4 ℃ 에서 처리한 후, 토끼 보체를 종농도 7.5 % 가 되도록 첨가하고, 37 ℃, 5 % CO2 존재하에서, 3 시간 가온 후, 생 세포에 포함되는 세포내 ATP 를 CellTiter-Glo Lumimescent Cell Viability Assay (Promega 사, Cat. G7572) 를 사용하여 측정하였다. CellTiter-Glo Lumimescent Cell Viability Assay 를 사용하여 얻어지는 발광 시그널에 대해, EnVision 2104 Multilabel Reader (Perkin Elmer 사) 를 사용하여 정량하였다. 측정은 삼중으로 실시하고, 평균값과 표준 편차를 산출하였다. 무처리의 세포로부터 얻어진 발광 시그널을 100 % 로서, 항체와 보체 의존적으로 감소한 발광 시그널을 CDC 활성으로서, 도 7 에 정리하였다.
마우스 항체 (LN22R8), 래트 항체 (2P10F2) 에서만, 음성 대조의 hIgG 에 대하여 농도 의존적인 CDC 활성이 관찰되고, LN22R8 에서는 10 ㎍/㎖ 에서 최대 41.1 % 까지 생 세포가 저하되었다. 2P10F2 에서는, 10 ㎍/㎖ 에서 최대 53.5 % 까지 생 세포가 저하되었다.
인간 키메라 항체 (LN22R8chIgG1, LN22R8chIgG2, LN22R8chIgG4P, 2P10F2chIgG1LALA, 2P10F2chIgG4P) 에서는, 음성 대조의 hIgG 에 대하여 명확한 CDC 활성은 관찰되지 않았다.
1)-17 인간 키메라 항체의 ADCP 활성
인간 IgG 항체는 마우스의 Fcγ 수용체와의 상호 작용을 통해서, 항체 의존적인 단구, 매크로파지에 의한 탐식 작용 (ADCP) 을 유도함으로써, 암 세포에 대한 살세포 활성을 나타내는 것이 보고되어 있다 (Overdijk et al., Journal of I㎜unology, 1-9, 2012). 인간 키메라 항체의 ADCP 활성에 대해, 이펙터 세포로서 RAW264.7 (ATCC, TIB-71), ADCP 표적 세포로서 인간 췌장주 PANC-1 또는 MIA PaCa-2 를 사용하여 평가하였다. PKH67 Green Fluorescent Cell Linker Mini Kit for General Cell Membrane Labeling (SIGMA, Cat. MINI67-1KIT) 으로 표지한 ADCP 표적 세포와 인간 키메라 항체 (LN22R8chIgG1, LN22R8chIgG2, LN22R8chIgG4P) 를 20 ㎍/㎖ 의 농도로 4 ℃, 1 시간 처리한 후, PKH26 Red Fluorescent Cell Linker Kit for General Cell Membrane Labeling (SIGMA, Cat. PKH26GL-1KT) 로 표지한 RAW264.7 세포를 ADCP 표적 세포의 5 배 첨가하고, 3 시간, 37 ℃, 5 % CO2 존재하에서 가온하였다. 플로우 사이토미터 (BD 사, CantoII) 를 사용하여, 탐식 작용에 의해 PKH67 시그널 양성으로 이행한 PKH26 양성 세포의 비율을 측정하였다. 음성 대조 샘플로서, 인간 IgG (hIgG, ChromPure Human IgG, Jackson I㎜unoResearch Laboratories 사, Cat. 009-000-003) 를 처리한 샘플에 대해 동일하게 측정을 실시하였다. 측정은 삼중으로 실시하고, 평균값, 표준 편차를 산출하여 도 8(a) PANC-1 의 결과를, 도 8(b) MIA PaCa-2 의 결과를 각각 나타내었다.
PANC-1 세포를 ADCP 의 표적으로 한 경우, LN22R8chIgG1 은 9.2 %, LN22R8chIgG4P 는 9.0 % 로 인간 IgG (5.5 %) 에 대해, 높은 ADCP 활성을 나타내었다. LN22R8chIgG2 는, 5.9 % 이고, ADCP 활성이었다.
MIA PaCa-2 세포를 ADCP 의 표적으로 한 경우도, 동일한 경향을 나타내고, LN22R8chIgG1 은 6.6 %, LN22R8chIgG4P 는 6.1 % 로 인간 IgG (3.6 %) 에 대해, 높은 ADCP 활성을 나타내었다. LN22R8chIgG2 는, 3.6 % 이고, ADCP 활성이었다.
1)-18 인간 키메라 항체의 in vivo 항종양 활성 측정
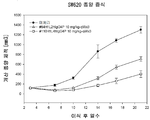
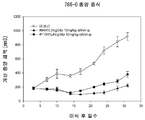
5 × 106 세포의 인간 췌장주 MIA PaCa-2 를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 을 포함하는 PBS 로 현탁하고, 4 ∼ 5 주령 암컷 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 찰스·리버로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 5 ∼ 7 일 후에 군나누기를 실시하고, 항인간 CD147 항체 LN22R8 의 마우스 항체 (LN22R8), 인간 키메라 항체 3 종 (LN22R8chIgG1, LN22R8chIgG2, LN22R8chIgG4P) 을 1 ㎎/㎏, 3 ㎎/㎏, 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 항인간 CD147 항체 2P10F2 의 래트 항체 (2P10F2), 인간 키메라 항체 2 종 (2P10F2chIgG2, 2P10F2chIgG4P) 을 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5 ∼ 6). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 9a (a) ∼ (d), 도 9b (e) ∼ (g) 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
LN22R8 은, 마우스 항체, 3 종의 인간 키메라 항체도 용량 의존적인 항종양 효과를 나타내었다. 인간 키메라 항체 LN22R8chIgG4P 10 ㎎/㎏ 투여군에서는, 이식 후 18 일 후에 5 마리 중 5 마리의 마우스에서 종양의 완전 축퇴가 관찰되고, 실험 종료 시의 이식 후 41 일 후에도 종양의 재증식은 보이지 않았다. 다른 LN22R8 항체 투여군에서는, 일부 혹은 모든 마우스에서 종양의 재증식이 관찰되었다.
2P10F2 는, 래트 항체, 2 종의 인간 키메라 항체에서 항종양 효과가 확인되었다. 2P10F2chIgG4P 10 ㎎/㎏ 투여군에서는, 이식 21 일 후에 6 마리 중 6 마리의 마우스에서 종양의 완전 축퇴가 관찰되었다.
동일한 에피토프 부분을 인식하는 항인간 CD147 항체인 마우스 항체 LN22R8 과 래트 항체 2P10F2 는, ADCC 활성, ADCP, CDC 활성과 같은 마우스 면역계에 의존한 이펙터 기능을 갖는 인간 키메라 항체 chIgG1 이나 ADCP 활성을 갖는 인간 키메라 항체 chIgG4P 뿐만 아니라, 어느 이펙터 기능도 거의 나타내지 않는 인간 키메라 항체 chIgG2 에서도 90 % 이상의 항종양 효과가 유지되고 있기 때문에, 마우스의 면역에 의존하지 않고, CD147 에 작용하는 신규의 작용 기전에 의해 항종양 효과를 나타내고 있는 것이 시사되었다.
1)-19 NOG 마우스에서의 CD147 인간 키메라 항체의 항종양 효과
NOG (NOD/Shi-scid, IL-2Rγnull) 마우스에서는, 마우스 T 세포, B 세포를 결실하고 있는 NOD-scid 마우스에 사이토카인 리셉터 공통 도메인인 IL-2 리셉터 γ 사슬 녹아웃을 교배시킴으로써, 마우스 T, B 세포에 더하여, NK 세포, 보체 활성을 결실하고 있고, 매크로파지나 수상 세포의 기능 저하가 보이고, 매우 중증인 면역 부전 상태에 있다 (Ito, Blood, 3175-3182, 2002). 이들 마우스 면역계의 중증인 부전 상태에서, CD147 항체의 항종양 효과가 영향을 받거나 MIA PaCa-2 의 피하 이식 모델로 검증하였다.
5 × 106 세포의 인간 췌장주 MIA PaCa-2 를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 을 포함하는 PBS 로 현탁하고, 7 주령 암컷 NOG 마우스 (NOD/Shi-scid, IL-2RγKO Jic, In-Vivo Science 사부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 6 일 후에 군나누기를 실시하고, 항 CD147 인간 키메라 항체 (LN22R8chIgG4P) 를 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 10 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
도 10 에 나타낸 인간 키메라 항체 LN22R8chIgG4P 의 결과는, 이식 17 일 후에 있어서의 종양 증식 억제율은, 10 ㎎/㎏ 투여군에서 99 % 이고, 5 마리 중 3 마리의 마우스에서 종양의 완전 축퇴가 관찰되었다.
항 CD147 인간 키메라 항체 (LN22R8chIgG4P) 는, 마우스 T, B 세포에 더하여, NK 세포, 보체 활성을 결실한 NOG 마우스에 형성시킨 췌장암 종양에 대해서도, 강한 항종양 효과를 나타내었기 때문에, 마우스의 면역 세포에 의존하지 않고 항종양 효과를 나타낼 가능성이 시사되었다.
(실시예 2) CD147 단백의 면역에 의한 원숭이 교차성 래트 항체의 제조
실시예 1 에서 얻어진 강한 항종양 효과를 나타내는 항인간 CD147 항체는, 마우스, 래트, 게잡이 원숭이의 CD147 에 교차성을 나타내지 않았다. 실시예 1 에서 얻어진 항체를 사용하여 게잡이 원숭이 CD147 교차성을 나타내는 CD147 항체의 취득을 시도하였다.
2)-1 면역
면역에는 WKY/Izm 래트의 암컷 (닛폰 에스엘씨사) 을 사용하였다. 리콤비넌트 인간 BSG, His tagged (CREATIVE BIOMART 사 제조) 항원 단백과 Freund's Comple te Adjuvant (와코 쥰야쿠사 제조) 를 혼합한 것을 미근부 (尾根部) 에 투여한 래트의 림프절 및 비장을 채취하고 하이브리도마 제조에 사용하였다.
2)-2 하이브리도마의 제조
림프절 세포 혹은 비장 세포와 마우스 미엘로마 SP2/0-ag14 세포 (ATCC, No. CRL-1581) 를 LF301 Cell Fusion Unit (BEX 사) 을 사용하여 전기 세포 융합하고, ClonaCell-HY Selection Medium D (StemCell Technologies 사) 에 희석하여 배양하였다. 출현한 하이브리도마 콜로니를 회수함으로써 모노 클론 하이브리도마를 제조하였다. 회수된 각 하이브리도마 콜로니를 배양하고, 얻어진 하이브리도마 배양 상청을 사용하여 항 CD147 항체 산생 하이브리도마의 스크리닝을 실시하였다.
2)-3 플로우 사이토메트리법에 의한 항체 스크리닝
인간 암 세포에 결합하고, 인간 CD147, 게잡이 원숭이 CD147 에 대한 결합성을 나타내는 항체 산생 하이브리도마를 선택하기 위해서 플로우 사이토미터를 사용한 항체 결합성 스크리닝을 실시하였다. 인간 암 세포로서, CD147 양성의 인간 췌장주 MIA PaCa-2 를 사용하였다. 실시예 1)-8 과 마찬가지로 인간, 또는, 게잡이 원숭이의 CD147 을 발현한 CHO-K1 세포 (CHO-K1-hCD147v2, CHO-K1-cynoCD147) 를 인간, 또는 게잡이 원숭이 CD147 에 대한 결합성 확인에 사용하였다. MIA PaCa-2, CHO-K1-hCD147v2, CHO-K1-cynoCD147 의 현탁액에 하이브리도마 배양 상청을 등량 첨가하고, 4 ℃ 에서 1 시간 이상 반응시킨 후, 5 % FBS 를 포함하는 PBS 로 세포를 세정하고, 항래트 IgG-PE (BD Biosciences 사, Cat. 550767) 를 사용하여 각 항체의 세포에 대한 결합을 형광 검출 가능하게 하였다. 플로우 사이토미터 (CantoII, BD Bioscience 사) 를 사용하여 세포의 형광 시그널의 측정을 실시하고, 음성 대조 샘플 (하이브리도마 배양액을 첨가하지 않은 세포) 에 대한 형광 시그널의 비를 산출하고, 표 2 에 결과의 일부를 정리하였다.
시판되는 래트 항체 Isotyping 키트 (Bio-Rad Laboratories 사, RMT1) 를 사용하여, 배양 상청에 포함되는 항체의 아이소타입 (Isotype) 을 결정하고 아울러 표 4 에 나타내었다.
실시예 1 에서 취득한 2P10F2 는 MIA PaCa-2, CHO-K1-hCD147v2 에 결합성을 나타내고, CHO-K1-cynoCD147 에는 결합성을 나타내지 않았다. 래트_CD147_#84 (본 명세서 중, r#84 로 표기하는 경우도 있다), 래트_CD147_#131 (본 명세서 중, r#131 로 표기하는 경우도 있다), 래트_CD147_#110 (본 명세서 중, r#110 으로 표기하는 경우도 있다), 래트_CD147_#101 (본 명세서 중, r#101 로 표기하는 경우도 있다) 은, MIA PaCa-2, CHO-K1-hCD147v2, CHO-K1-cynoCD147 에 결합성을 나타내고, 게잡이 원숭이 교차성을 나타내는 항인간 CD147 래트 항체를 취득할 수 있었다.
2)-4 저(低) IgG 혈청을 사용한 래트 모노클로날 항체의 조제
게잡이 원숭이 교차성을 나타내는 항인간 CD147 모노클로날 항체는, 하이브리도마 배양 상청으로부터 정제하였다. 먼저, 래트_CD147_#131 의 항체 산생 하이브리도마를 ClonaCell-HY Selection Medium E 에서 충분량까지 증식시킨 후, Ultra Low IgG FBS (Life Technologies 사) 를 20 % 첨가한 Hybridoma SFM (Life Technologies 사) 에 배지 교환하고, 7 일간 배양하였다. 본 배양 상청을 회수하고 0.45 ㎛ 의 필터를 통과시켜 멸균하였다.
2)-5 고밀도 배양에 의한 래트 모노클로날 항체의 조제
CL-1000 플라스크 (닛폰 벡톤·디킨슨 주식회사) 를 사용하여, 래트_CD147_#84, 래트_CD147_#101, 래트_CD147_#110 하이브리도마를 배양하고, 모노클로날 항체를 포함하는 하이브리도마 배양 상청을 조제하였다.
2)-6 모노클로날 항체의 정제
실시예 2)-4 및 실시예 2)-5 에서 제조한 배양 상청으로부터 실시예 1)-6 과 동일한 방법으로 항체를 정제하였다.
2)-7 in vivo 항종양 활성 측정에 의한 항체 스크리닝
5 × 106 세포의 인간 췌장주 MIA PaCa-2 를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 을 포함하는 PBS 로 현탁하고, 5 주령 암컷 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 찰스·리버로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 6 ∼ 8 일 후에 군나누기를 실시하고, 게잡이 원숭이 교차성 항 CD147 래트 항체 #84, #101 또는 #110 을 이식 후, 8 일 후, 15 일 후에 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 대조군의 마우스에는, PBS 를 동일하게 복강내에 투여하였다. 게잡이 원숭이 교차성 항 CD147 래트 항체 #131 을 이식 후, 6 일 후에 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
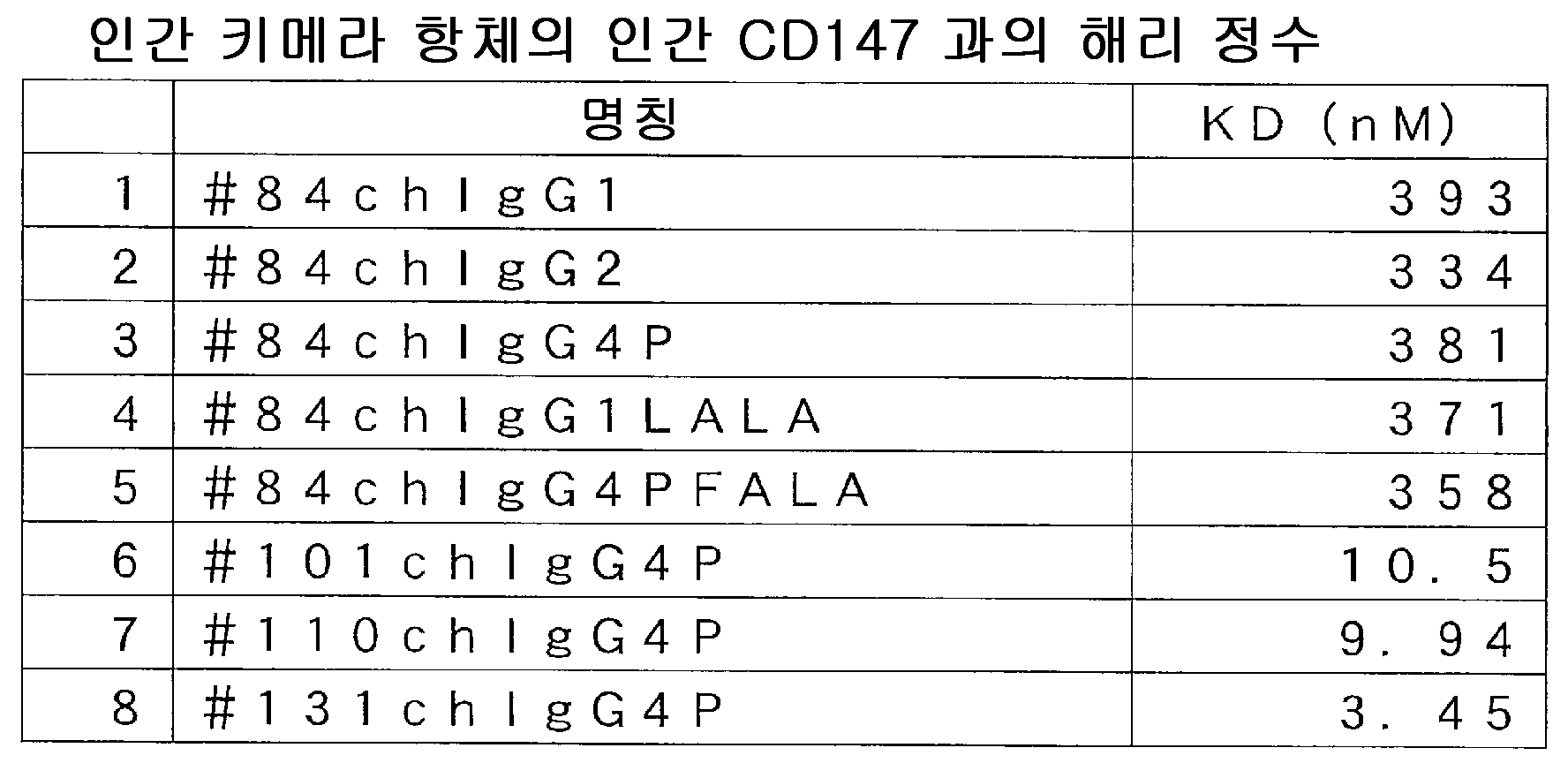
결과를 도 11(a) ∼ (d) 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다. r#84 는, 이식 28 일 후에 있어서의 종양 증식 억제율이, 10 ㎎/㎏ 투여군에서 95 % 였다 (도 11(a)). r#101 은, 이식 15 일 후에 있어서의 종양 증식 억제율이 10 ㎎/㎏ 투여군에서 37 % 였지만, 이식 28 일 후에는 종양의 재증식이 보였다 (도 11(b)). r#110 은, 이식 15 일 후에 있어서의 종양 증식 억제율이 10 ㎎/㎏ 투여군에서 51 % 였지만, 이식 28 일 후에는 종양의 재증식이 보였다 (도 11(c)). r#131 은, 이식 16 일 후에 있어서의 종양 증식 억제율은, 10 ㎎/㎏ 투여군에서 50 % 였다 (도 11(d)). MIA PaCa-2 의 종양 증식을 강하게 저해하는 래트 항체, r#84, 부분적인 저해를 하는 r#101, r#110, r#131 이 얻어졌다.
결과를 도 11(a) ∼ (d) 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다. r#84 는, 이식 28 일 후에 있어서의 종양 증식 억제율이 10 ㎎/㎏ 투여군에서 95 % 였다 (도 11(a)). r#101 은, 이식 15 일 후에 있어서의 종양 증식 억제율이 10 ㎎/㎏ 투여군에서 37 % 였지만, 이식 28 일 후에는 종양의 재증식이 보였다 (도 11(b)). r#110 은, 이식 15 일 후에 있어서의 종양 증식 억제율이 10 ㎎/㎏ 투여군에서 51 % 였지만, 이식 28 일 후에는 종양의 재증식이 보였다 (도 11(c)). r#131 은, 이식 16 일 후에 있어서의 종양 증식 억제율이 10 ㎎/㎏ 투여군에서 50 % 였다 (도 11(d)).
2)-8 에피토프 해석 : 2P10F2-경합 ELISA (2P10F2-competitive ELISA)
원숭이 교차성 래트 CD147 항체의 에피토프 해석을 목적으로 하여, 2P10F2chIgG4P 의 CD147 리콤비넌트 단백질에 대한 결합성을 저해하는지 경합 ELISA 에 의해 조사하였다. 인간 CD147-Fc 융합 단백질 (Sino Biological Inc., 10186-H02H) 을 PBS 로 용해하고, 20 ㎍/㎖ 로 조제하고, 96 웰 플레이트 (Thermofisher 사, Cat. 43454) 에 50 ㎕ 첨가하고 4 ℃ 에서 보관하였다. 단백 용액을 제거하고, 300 ㎕ 의 1 % BSA 함유 PBS 를 첨가하여, 실온에서 1 시간 가온하였다. 1 % BSA 함유 PBS 로 희석한 25 ㎕ 의 20 혹은 60 ㎍/㎖ 의 CD147 래트 항체 r#84, r#101, r#110, r#131 또는 2P10F2, 또는 1 % BSA 함유 PBS 를 경합 항체로서 96 웰 플레이트에 첨가하고, 실온에서 2 시간 가온하였다. 1 % BSA 함유 PBS 로 희석한 20 ng/㎖ 의 2P10F2chIgG4P 를 25 ㎕, 96 웰 플레이트에 첨가하고, 실온에서 2 시간 가온하였다. 0.05 % 트윈 20 (BIO RAD, Cat. 170-6531) 을 포함하는 PBS 로 2 회, 96 웰을 세정하였다. 1 % BSA 함유 PBS 로 2000 배로 희석한 마우스 모노클로날 HP6025 항 인간 IgG4 Fc (HRP) (abcam 사, Cat. ab99823) 를 50 ㎕, 96 웰 플레이트에 첨가하고, 실온에서 1 시간 가온하였다. 0.05 % 트윈 20 (BIO RAD, Cat. 170-6531) 을 포함하는 PBS 로 3 회, 96 웰을 세정하였다. 50 ㎕ 의 Super AquaBlue ELISA Substrate (eBioscience 사, 00-4203-58) 를 96 웰 플레이트에 첨가하고, 실온에서 20 분간 가온하였다. EnVision 2104 Multilabel Reader (Perkin Elmer 사) 로, 96 웰 플레이트의 405 ㎚ 의 흡광도를 측정하였다. 경합 항체를 포함하지 않는 웰에서의 측정값을 대조로서, 경합 항체에 의해 저하된 흡광도를 % 로 산출하고 도 12 에 나타내었다. 측정은 3 웰로 실시하고, 평균값을 나타내었다.
r#84, r#101, r#131 은, 2P10F2 래트 항체와 마찬가지로 2P10F2chIgG4P 의 결합성을 90 % 이상 저해하고, 항체 인식 부위가, 2P10F2 와 가까운 것이 시사되었다. r#110 은, 2P10F2 래트 항체의 결합을 저해하지 않았다. 항체 인식 부위는, 2P10F2 와 떨어져 있거나, 혹은 결합성이 약하기 때문에, 2P10F2 래트 항체의 결합을 저해할 수 없을 가능성이 생각되었다.
(실시예 3) 원숭이 교차성 래트 항체의 클로닝과 인간 키메라 항체의 제조
3)-1 래트 항 CD147 항체의 가변 영역을 코드하는 cDNA 의 클로닝 뉴클레오티드 서열의 결정
3)-1-1 래트_CD147_#84 항체의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 결정
실시예 1)-11-2 와 동일한 방법으로 실시하였다. 결정된 래트_CD147_#84 항체의 경쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 48 에 나타내고, 아미노산 서열을 서열 번호 49 에 나타낸다. 중쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 50 에 나타내고, 아미노산 서열을 서열 번호 51 에 나타낸다. 래트_CD147_#84 항체의 경쇄 가변 영역의 CDRL1, CDRL2 및 CDRL3 을, 각각, 서열 번호 52, 53 및 54 에 나타낸다. 래트_CD147_#84 항체의 중쇄 가변 영역의 CDRH1, CDRH2 및 CDRH3 을, 각각, 서열 번호 55, 56 및 57 에 나타낸다.
3)-1-2 래트_CD147_#101 항체의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 결정
실시예 1)-11-2 와 동일한 방법으로 실시하였다. 결정된 래트_CD147_#101 항체의 경쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 58 에 나타내고, 아미노산 서열을 서열 번호 59 에 나타낸다. 중쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 60 에 나타내고, 아미노산 서열을 서열 번호 61 에 나타낸다. 래트_CD147_#101 항체의 경쇄 가변 영역의 CDRL1, CDRL2 및 CDRL3 을, 각각, 서열 번호 62, 63 및 64 에 나타낸다. 래트_CD147_#101 항체의 중쇄 가변 영역의 CDRH1, CDRH2 및 CDRH3 을, 각각, 서열 번호 65, 66 및 67 에 나타낸다.
3)-1-3 래트_CD147_#110 항체의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 결정
실시예 1)-11-2 와 동일한 방법으로 실시하였다. 결정된 래트_CD147_#110 항체의 경쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 68 에 나타내고, 아미노산 서열을 서열 번호 69 에 나타낸다. 중쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 70 에 나타내고, 아미노산 서열을 서열 번호 71 에 나타낸다. 래트_CD147_#110 항체의 경쇄 가변 영역의 CDRL1, CDRL2 및 CDRL3 을, 각각, 서열 번호 72, 73 및 74 에 나타낸다. 래트_CD147_#110 항체의 중쇄 가변 영역의 CDRH1, CDRH2 및 CDRH3 을, 각각, 서열 번호 75, 76 및 77 에 나타낸다.
3)-1-4 래트_CD147_#131 항체의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열의 결정
실시예 1)-11-2 와 동일한 방법으로 실시하였다. 결정된 래트_CD147_#131 항체의 경쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 78 에 나타내고, 아미노산 서열을 서열 번호 79 에 나타낸다. 중쇄의 가변 영역을 코드하는 cDNA 의 뉴클레오티드 서열을 서열 번호 80 에 나타내고, 아미노산 서열을 서열 번호 81 에 나타낸다. 래트_CD147_#131 항체의 경쇄 가변 영역의 CDRL1, CDRL2 및 CDRL3 을, 각각, 서열 번호 82, 83 및 84 에 나타낸다. 래트_CD147_#131 항체의 중쇄 가변 영역의 CDRH1, CDRH2 및 CDRH3 을, 각각, 서열 번호 85, 86 및 87 에 나타낸다.
3)-2 인간 키메라 항체 발현 벡터의 제조
3)-2-1 래트_CD147_#84 의 인간 키메라 항체 발현 벡터의 제조
3)-2-1-1 인간 키메라 및 인간화 IgG4PFALA 타입 중쇄 발현 벡터 pCMA-G4PFALA 의 구축
서열 번호 88 로 나타내는 인간 중쇄 시그널 서열 및 인간 IgG4PFALA 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편을 사용하여, 실시예 1)-12-2 와 동일한 방법으로 pCMA-G4PFALA 를 구축하였다.
3)-2-1-2 인간 키메라 래트_CD147_#84 의 경쇄 발현 벡터의 구축
실시예 3)-1-1 에서 얻어진 래트_CD147_#84 의 경쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-4 와 동일한 방법으로 인간 키메라 래트_CD147_#84 의 경쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#84 의 경쇄의 뉴클레오티드 서열 및 그 경쇄의 아미노산 서열을, 서열 번호 89 및 서열 번호 90 에 각각 나타낸다.
3)-2-1-3 인간 키메라 래트_CD147_#84 의 IgG1 타입 중쇄 발현 벡터의 구축
실시예 3)-1-1 에서 얻어진 래트_CD147_#84 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-5 와 동일한 방법으로 인간 키메라 래트_CD147_#84 의 IgG1 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#84 의 IgG1 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 91 및 서열 번호 92 에 각각 나타낸다.
3)-2-1-4 인간 키메라 래트_CD147_#84 의 IgG2 타입 중쇄 발현 벡터의 구축
실시예 3)-1-1 에서 얻어진 래트_CD147_#84 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-6 과 동일한 방법으로 인간 키메라 래트_CD147_#84 의 IgG2 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#84 의 IgG2 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 93 및 서열 번호 94 에 각각 나타낸다.
3)-2-1-5 인간 키메라 래트_CD147_#84 의 IgG4P 타입 중쇄 발현 벡터의 구축
실시예 3)-1-1 에서 얻어진 래트_CD147_#84 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-13-6 과 동일한 방법으로 인간 키메라 래트_CD147_#84 의 IgG4P 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#84 의 IgG4P 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 95 및 서열 번호 96 에 각각 나타낸다.
3)-2-1-6 인간 키메라 래트_CD147_#84 의 IgG1LALA 타입 중쇄 발현 벡터의 구축
실시예 3)-1-1 에서 얻어진 래트_CD147_#84 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-13-4 와 동일한 방법으로 인간 키메라 래트_CD147_#84 의 IgG1LALA 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#84 의 IgG1LALA 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 97 및 서열 번호 98 에 각각 나타낸다.
3)-2-1-7 인간 키메라 래트_CD147_#84 의 IgG4PFALA 타입 중쇄 발현 벡터의 구축
실시예 3)-1-1 에서 얻어진 래트_CD147_#84 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, In-fusion 클로닝용으로 설계한 프라이머로 PCR 을 실시함으로써 중쇄의 가변 영역을 코드하는 cDNA 를 포함하는 DNA 단편을 증폭하였다. pCMA-G4PFALA 를 제한 효소 BlpI 로 절단한 지점에, In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, 증폭한 DNA 단편을 삽입함으로써 인간 키메라 래트_CD147_#84 의 IgG4PFALA 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#84 의 IgG4PFALA 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 99 및 서열 번호 100 에 각각 나타낸다.
3)-2-2 래트_CD147_#101 의 인간 키메라 항체 발현 벡터의 제조
3)-2-2-1 인간 키메라 래트_CD147_#101 의 경쇄 발현 벡터의 구축
실시예 3)-1-2 에서 얻어진 래트_CD147_#101 의 경쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-4 와 동일한 방법으로 인간 키메라 래트_CD147_#101 의 경쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#101 의 경쇄의 뉴클레오티드 서열 및 그 경쇄의 아미노산 서열을, 서열 번호 101 및 서열 번호 102 에 각각 나타낸다.
3)-2-2-2 인간 키메라 래트_CD147_#101 의 IgG2 타입 중쇄 발현 벡터의 구축
실시예 3)-1-2 에서 얻어진 래트_CD147_#101 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-6 과 동일한 방법으로 인간 키메라 래트_CD147_#101 의 IgG2 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#101 의 IgG2 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 103 및 서열 번호 104 에 각각 나타낸다.
3)-2-2-3 인간 키메라 래트_CD147_#101 의 IgG4P 타입 중쇄 발현 벡터의 구축
실시예 3)-1-2 에서 얻어진 래트_CD147_#101 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-13-6 과 동일한 방법으로 인간 키메라 래트_CD147_#101 의 IgG4P 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#101 의 IgG4P 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 105 및 서열 번호 106 에 각각 나타낸다.
3)-2-2-4 인간 키메라 래트_CD147_#101 의 IgG4PFALA 타입 중쇄 발현 벡터의 구축
실시예 3)-1-2 에서 얻어진 래트_CD147_#101 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 3)-2-1-7 과 동일한 방법으로 인간 키메라 래트_CD147_#101 의 IgG4PFALA 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#101 의 IgG4PFALA 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 107 및 서열 번호 108 에 각각 나타낸다.
3)-2-3 래트_CD147_#110 의 인간 키메라 항체 발현 벡터의 제조
3)-2-3-1 인간 키메라 래트_CD147_#110 의 경쇄 발현 벡터의 구축
실시예 3)-1-3 에서 얻어진 래트_CD147_#110 의 경쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-4 와 동일한 방법으로 인간 키메라 래트_CD147_#110 의 경쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#110 의 경쇄의 뉴클레오티드 서열 및 그 경쇄의 아미노산 서열을, 서열 번호 109 및 서열 번호 110 에 각각 나타낸다.
3)-2-3-2 인간 키메라 래트_CD147_#110 의 IgG2 타입 중쇄 발현 벡터의 구축
실시예 3)-1-3 에서 얻어진 래트_CD147_#110 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-6 과 동일한 방법으로 인간 키메라 래트_CD147_#110 의 IgG2 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#110 의 IgG2 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 111 및 서열 번호 112 에 각각 나타낸다.
3)-2-3-3 인간 키메라 래트_CD147_#110 의 IgG4P 타입 중쇄 발현 벡터의 구축
실시예 3)-1-3 에서 얻어진 래트_CD147_#110 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-13-6 과 동일한 방법으로 인간 키메라 래트_CD147_#110 의 IgG4P 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#110 의 IgG4P 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 113 및 서열 번호 114 에 각각 나타낸다.
3)-2-3-4 인간 키메라 래트_CD147_#110 의 IgG4PFALA 타입 중쇄 발현 벡터의 구축
실시예 3)-1-3 에서 얻어진 래트_CD147_#110 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 3)-2-1-7 과 동일한 방법으로 인간 키메라 래트_CD147_#110 의 IgG4PFALA 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#110 의 IgG4PFALA 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 115 및 서열 번호 116 에 각각 나타낸다.
3)-2-4 래트_CD147_#131 의 인간 키메라 항체 발현 벡터의 제조
3)-2-4-1 인간 키메라 래트_CD147_#131 의 경쇄 발현 벡터의 구축
실시예 3)-1-4 에서 얻어진 래트_CD147_#131 의 경쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-4 와 동일한 방법으로 인간 키메라 래트_CD147_#131 의 경쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#131 의 경쇄의 뉴클레오티드 서열 및 그 경쇄의 아미노산 서열을, 서열 번호 117 및 서열 번호 118 에 각각 나타낸다.
3)-2-4-2 인간 키메라 래트_CD147_#131 의 IgG2 타입 중쇄 발현 벡터의 구축
실시예 3)-1-4 에서 얻어진 래트_CD147_#131 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-12-6 과 동일한 방법으로 인간 키메라 래트_CD147_#131 의 IgG2 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#131 의 IgG2 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 119 및 서열 번호 120 에 각각 나타낸다.
3)-2-4-3 인간 키메라 래트_CD147_#131 의 IgG4P 타입 중쇄 발현 벡터의 구축
실시예 3)-1-4 에서 얻어진 래트_CD147_#131 의 중쇄의 가변 영역을 코드하는 cDNA 를 템플릿으로서 사용하여, 실시예 1)-13-6 과 동일한 방법으로 인간 키메라 래트_CD147_#131 의 IgG4P 타입 중쇄 발현 벡터를 구축하였다. 인간 키메라 래트_CD147_#131 의 IgG4P 타입 중쇄의 뉴클레오티드 서열 및 그 중쇄의 아미노산 서열을, 서열 번호 121 및 서열 번호 122 에 각각 나타낸다.
3)-3 인간 키메라 항체의 조제
3)-3-1 원숭이 교차성 래트 항체의 인간 키메라 항체의 생산
실시예 1)-14-1 과 동일한 방법으로 생산하였다.
인간 키메라 래트_CD147_#84 의 IgG1 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#84 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#84 의 인간 키메라 항체를 「#84chIgG1」 이라고 명명하였다. 인간 키메라 래트_CD147_#84 의 IgG2 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#84 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#84 의 인간 키메라 항체를 「#84chIgG2」 라고 명명하였다. 인간 키메라 래트_CD147_#84 의 IgG4P 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#84 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#84 의 인간 키메라 항체를 「#84chIgG4P」 라고 명명하였다. 인간 키메라 래트_CD147_#84 의 IgG1LALA 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#84 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#84 의 인간 키메라 항체를 「#84chIgG1LALA」 라고 명명하였다. 인간 키메라 래트_CD147_#84 의 IgG4PFALA 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#84 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#84 의 인간 키메라 항체를 「#84chIgG4PFALA」 라고 명명하였다.
인간 키메라 래트_CD147_#101 의 IgG2 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#101 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#101 의 인간 키메라 항체를 「#101chIgG2」 라고 명명하였다. 인간 키메라 래트_CD147_#101 의 IgG4P 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#101 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#101 의 인간 키메라 항체를 「#101chIgG4P」 라고 명명하였다. 인간 키메라 래트_CD147_#101 의 IgG4PFALA 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#101 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#101 의 인간 키메라 항체를 「#101chIgG4PFALA」 라고 명명하였다.
인간 키메라 래트_CD147_#110 의 IgG2 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#110 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#110 의 인간 키메라 항체를 「#110chIgG2」 라고 명명하였다. 인간 키메라 래트_CD147_#110 의 IgG4P 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#110 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#110 의 인간 키메라 항체를 「#110chIgG4P」 라고 명명하였다. 인간 키메라 래트_CD147_#110 의 IgG4PFALA 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#110 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#110 의 인간 키메라 항체를 「#110chIgG4PFALA」 라고 명명하였다.
인간 키메라 래트_CD147_#131 의 IgG2 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#131 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#131 의 인간 키메라 항체를 「#131chIgG2」 라고 명명하였다. 인간 키메라 래트_CD147_#131 의 IgG4P 타입 중쇄 발현 벡터와 인간 키메라 래트_CD147_#131 의 경쇄 발현 벡터의 조합으로 취득한 래트_CD147_#131 의 인간 키메라 항체를 「#131chIgG4P」 라고 명명하였다.
3)-3-2 원숭이 교차성 래트 항체의 인간 키메라 항체의 정제
실시예 3)-3-1 에서 얻어진 배양 상청으로부터, 실시예 1)-14-2 와 동일한 방법으로 정제하였다.
(실시예 4) 인간 키메라 항체의 in vivo 항종양 활성
5 × 106 세포의 인간 췌장주 MIA PaCa-2 를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 을 포함하는 PBS 로 현탁하고, 5 ∼ 6 주령 암컷 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 찰스·리버로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 5 ∼ 6 일 후에 군나누기를 실시하고, 게잡이 원숭이 교차성 항 CD147 인간 키메라 항체 (#84chIgG1, #84chIgG1LALA, #84chIgG2, #84chIgG4P, #84chIgG4PFALA) 를 군나누기 후 당일에 1, 3, 또는 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 게잡이 원숭이 교차성 항 CD147 인간 키메라 항체 (#101chIgG2, #101chIgG4P, #101chIgG4PFALA, #110chIgG2, #110chIgG4P, #110chIgG4PFALA, #131chIgG2, #131chIgG4P) 를 군나누기 후 당일에 3, 또는 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 13a (a) ∼ (d), 도 13b (e) ∼ (h), 도 13c (i) ∼ (l), 도 13d (m) 및 (n) 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
인간 키메라 항체 #84 는, 어느 IgG 서브타입도 용량 의존적인 항종양 효과를 나타내었다. 가장 강한 항종양 효과가 확인된 #84chIgG4P 10 ㎎/㎏ 투여군에서는, 이식 20 일 후에 5 예 중 5 예의 마우스에서 종양의 완전한 퇴축이 확인되었다.
인간 키메라 항체 #101 은, 어느 IgG 서브타입도 용량 의존적인 항종양 효과를 나타내었다. 가장 강한 항종양 효과가 확인된 101chIgG4P 10 ㎎/㎏ 투여군에서는, 이식 20 일 후에 5 예 중 4 예의 마우스에서 종양의 완전한 퇴축이 확인되었다.
인간 키메라 항체 #110 은, 어느 IgG 서브타입도 용량 의존적인 항종양 효과를 나타내었다. 가장 강한 항종양 효과가 확인된 110chIgG4P 10 ㎎/㎏ 투여군에서는, 이식 22 일 후에 5 예 중 3 예의 마우스에서 종양의 완전한 퇴축이 확인되었다.
인간 키메라 항체 #131 은, IgG4P 서브타입에서 용량 의존적인 항종양 효과를 나타내었다. #131chIgG2 에서는, 3 ㎎/㎏, 10 ㎎/㎏ 투여군에서 보인 항종양 효과에 차는 없었다.
어느 인간 키메라 항체도, 항체의 인간 IgG subtype 에 의존하지 않고, 10 ㎎/㎏ 의 투여량에서는, 68 ∼ 100 % 의 종양 증식 억제 효과를 나타내었다. 게잡이 원숭이 교차성 항인간 CD147 인간 키메라 항체 #084, #101, #110, #131 에 대해서도, 세포의 면역으로부터 취득된 항인간 CD147 항체, LN22R8 이나 2P10F2 와 동일하게, 마우스의 면역계에 의존하지 않고, CD147 에 작용하는 신규의 작용 기전에 의해 항종양 효과를 나타내고 있는 것이 시사되었다.
(실시예 5) 인간 키메라 항체의 CD147 결합성 평가
실시예 3)-3-1 에서 제조한 #84chIgG1, #84chIgG2, #84chIgG4P, #84chIgG1LALA, #84chIgG4PFALA, #101chIgG4P, #110chIgG4P 의 인간 CD147 에 대한 해리 정수 측정은, Biacore T200 (GE Healthcare Bioscience 사 제조) 을 사용하고, Human Antibody Capture Kit (GE Healthcare Bioscience 사 제조) 를 사용하여 고정화한 항 인간 IgG (Fc) 항체에 항체를 리간드로서 포착 (캡처) 하고, 항원을 아날라이트로서 측정하는 캡처법으로 실시하였다. 러닝 버퍼로서 HBS-EP+ (GE Healthcare Bioscience 사 제조), 센서 칩으로서 CM5 (GE Healthcare Bioscience 사 제조) 를 사용하였다. 칩 상에 1 ㎍/㎖ 또는 2 ㎍/㎖ 의 인간 키메라 항체를 10 ㎕/분으로 60 초간 첨가한 후, 항원으로서 CD147 단백질의 희석 계열 용액 (#131chIgG4P 에 대해 0.25 ∼ 4 ㎍/㎖, #84chIgG1, #84chIgG2, #84chIgG4P, #84chIgG1LALA, #84chIgG4PFALA, #101chIgG4P, #110chIgG4P 에 대해 0.5 ∼ 8 ㎍/㎖) 을 유속 30 ㎕/분으로 120 초간 첨가하고, 계속해서 120 초간, 300 초간 또는 600 초간의 해리상을 모니터하였다. 여기서, CD147 단백질은, 대장균에서 발현시키고, Ni 어피니티 (Ni affinity), SEC 의 2 스텝 (steps) 정제 후, 태그 절단 정제한 것을 사용하였다. 재생 용액으로서, 3M 마그네슘클로라이드 (GE Healthcare Bioscience 사 제조) 를 유속 20 ㎕/분으로 30 초간 첨가하였다. 데이터의 해석에는 1 : 1 결합 모델을 사용하여, 결합 속도 정수 ka, 해리 속도 정수 kd 및 해리 정수 (KD ; KD = kd/ka) 를 산출하였다. 결과를 표 5 에 나타낸다.
(실시예 6) 인간화 항체의 제조
6)-1 인간화 항체의 설계
6)-1-1 가변 영역의 분자 모델링
호몰로지 모델링으로서 공지된 방법 (Methods in Enzymology, 203, 121-153, (1991)) 을 이용하였다. 시판되는 단백질 입체 구조 해석 프로그램 Discovery Studio (다쏘·시스템사 제조) 를 사용하여, 가변 영역에 대하여 높은 서열 동일성을 갖는 Protein Data Bank (Nuc. Acids Res. 35, D301-D303 (2007)) 에 등록되어 있는 구조를 검색하였다. 히트한 중쇄, 경쇄 및 중쇄 경쇄 인터페이스 구조를 주형으로 하여, 삼차원 모델이 작성되었다.
6)-1-2 인간화 항체의 설계 방법
CDR 그래프팅 (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)) 에 의해 인간화하였다. 래트_CD147_#84 의 프레임 워크 영역은, KABAT et al. (Sequences of Proteins of I㎜unological Interest, 5th Ed. Public Health Service National Institutes of Health, Bethesda, MD. (1991)) 에 있어서 기정 되는 인간 카파 (kappa) 사슬 서브그룹 1 의 콘센서스 서열과, 인간 감마 (ga㎜a) 사슬 서브그룹 3 의 콘센서스 서열에 높은 상동성을 갖는다는 점에서, 그것들이 래트_CD147_#84 의 경쇄와 중쇄의 억셉터로서 각각 선택되었다. 래트_CD147_#101 의 프레임 워크 영역은, KABAT et al. 에 있어서 기정되는 인간 카파 (kappa) 사슬 서브그룹 1 의 콘센서스 서열과, 인간 감마 (ga㎜a) 사슬 서브그룹 3 및 IMGT (THE INTERNATIONAL IMMUNOGENETICS INFORMATION SYSTEM (등록상표)) 에 있어서 규정되는 인간 감마 (ga㎜a) 사슬의 IGHV3-30*05 와 IGHJ3*01 에, 높은 상동성을 갖는다는 점에서, 그것들이 래트_CD147_#101 의 경쇄와 중쇄의 억셉터로서 각각 선택되었다. 래트_CD147_#110 의 프레임 워크 영역은, IMGT 에 있어서 규정되는 인간 카파 (kappa) 사슬의 IGKV1-39*01 과 IGKJ4*01 과, 인간 감마 (ga㎜a) 사슬의 IGHV1-2*02 와 IGHJ6*01 에, 높은 상동성을 갖는다는 점에서, 그것들이 래트_CD147_#110 의 경쇄와 중쇄의 억셉터로서 각각 선택되었다. 래트_CD147_#131 의 프레임 워크 영역은, IMGT 에 있어서 규정되는 인간 카파 (kappa) 사슬의 IGKV1-39*01 과 IGKJ2*01 과, 인간 감마 (ga㎜a) 사슬의 IGHV3-30*05 와 IGHJ6*01 에, 높은 상동성을 갖는다는 점에서, 래트_CD147_#131 의 경쇄와 중쇄의 억셉터로서 각각 선택되었다. 억셉터 상에 이입해야 할 도너 잔기는, Queen et al. (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)) 에 의해 부여되는 기준 등을 참고로, 삼차원 모델을 분석함으로써 선택되었다.
6)-1-3 래트_CD147_#84 중쇄의 인간화
래트_CD147_#84 중쇄의 가변 영역에 대해, 도 14(a) 에 나타내는 억셉터의 아미노산 잔기를 이입함으로써, 인간화 항체 중쇄 #84H1h 의 가변 영역이 설계되었다.
설계된 가변 영역에, 인간의 IgG2 및 IgG4P 의 감마 (ga㎜a) 사슬 정상 영역을 접속시킨 인간화 항체 중쇄가 설계되고, 각각 #84H1hIgG2, #84H1hIgG4P 라고 명명하였다. #84H1hIgG2 의 전체 길이 아미노산 서열을 서열 번호 123 에 기재한다. 서열 번호 123 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 124 에 기재한다. #84H1hIgG4P 의 전체 길이 아미노산 서열을 서열 번호 125 에 기재한다. 서열 번호 125 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 126 에 기재한다.
6)-1-4 래트_CD147_#84 경쇄의 인간화
래트_CD147_#84 경쇄의 가변 영역에 대해, 도 14(b) 에 나타내는 억셉터의 아미노산 잔기를 이입함으로써, 인간화 항체 경쇄 #84L2h 의 가변 영역이 설계되었다.
설계된 가변 영역에, 인간의 κ 사슬 정상 영역을 접속시킨 인간화 항체 경쇄가 설계되고, #84L2h 라고 명명하였다. #84L2h 의 경쇄 전체 길이 아미노산 서열을 서열 번호 127 에 기재한다. 서열 번호 127 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 128 에 기재한다.
6)-1-5 #84 인간화 항체
상기로부터 설계된 중쇄 #84H1hIgG2 와 경쇄 #84L2h 를 조합하고, 인간화 항체 #84H1L2hIgG2 를 설계하였다. 또, 중쇄 #84H1hIgG4P 와 경쇄 #84L2h 를 조합하고, 인간화 항체 #84H1L2hIgG4P 를 설계하였다.
6)-1-6 래트_CD147_#101 중쇄의 인간화
래트_CD147_#101 중쇄의 가변 영역에 대해, 도 15(a) 에 나타내는 억셉터의 아미노산 잔기를 이입함으로써, 인간화 항체 중쇄 #101H1h 의 가변 영역이 설계되었다.
설계된 가변 영역에, 인간의 IgG2 및 IgG4P 의 감마 (ga㎜a) 사슬 정상 영역을 접속시킨 인간화 항체 중쇄가 설계되고, 각각 #101H1hIgG2, #101H1hIgG4P 라고 명명하였다. #101H1hIgG2 의 전체 길이 아미노산 서열을 서열 번호 129 에 기재한다. 서열 번호 129 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 130 에 기재한다. #101H1hIgG4P 의 전체 길이 아미노산 서열을 서열 번호 131 에 기재한다. 서열 번호 131 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 132 에 기재한다.
6)-1-7 래트_CD147_#101 경쇄의 인간화
래트_CD147_#101 경쇄의 가변 영역에 대해, 도 15(b) 에 나타내는 억셉터의 아미노산 잔기를 이입함으로써, 인간화 항체 경쇄 #101L2h 의 가변 영역이 설계되었다.
설계된 가변 영역에, 인간의 κ 사슬 정상 영역을 접속시킨 인간화 항체 경쇄가 설계되고, #101L2h 라고 명명하였다. #101L2h 의 경쇄 전체 길이 아미노산 서열을 서열 번호 133 에 기재한다. 서열 번호 133 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 134 에 기재한다.
6)-1-8 #101 인간화 항체
상기로부터 설계된 중쇄 #101H1hIgG2 와 경쇄 #101L2h 를 조합하고, 인간화 항체 #101H1L2hIgG2 를 설계하였다. 또, 중쇄 #101H1hIgG4P 와 경쇄 #101L2h 를 조합하고, 인간화 항체 #101H1L2hIgG4P 를 설계하였다.
6)-1-9 래트_CD147_#110 중쇄의 인간화
래트_CD147_#110 중쇄의 가변 영역에 대해, 도 16(a), (b) 에 나타내는 억셉터의 아미노산 잔기를 이입함으로써, 인간화 항체 중쇄 #110H1h, #110H13h 의 가변 영역이 설계되었다.
설계된 가변 영역에, IgG4P 의 감마 (ga㎜a) 사슬 정상 영역을 접속시킨 인간화 항체 중쇄가 설계되고, 각각 #110H1hIgG4P, #110H13hIgG4P 라고 명명하였다. #110H1hIgG4P 의 전체 길이 아미노산 서열을 서열 번호 135, #110H13hIgG4P 의 전체 길이 아미노산 서열을 서열 번호 147 에 기재한다. 서열 번호 135 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 136, 서열 번호 147 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 148 에 기재한다.
6)-1-10 래트_CD147_#110 경쇄의 인간화
래트_CD147_#110 경쇄의 가변 영역에 대해, 도 16(c), (d), (e) 에 나타내는 억셉터의 아미노산 잔기를 이입함으로써, 인간화 항체 경쇄 #110L4h, #110L2h, #110L12h 의 가변 영역이 설계되었다.
설계된 가변 영역에, 인간의 κ 사슬 정상 영역을 접속시킨 인간화 항체 경쇄가 설계되고, 각각 #110L4h, #110L2h, #110L12h 라고 명명하였다. #110L4h 의 경쇄 전체 길이 아미노산 서열을 서열 번호 137, #110L2h 의 경쇄 전체 길이 아미노산 서열을 서열 번호 149, #110L12h 의 경쇄 전체 길이 아미노산 서열을 서열 번호 151 에 기재한다. 서열 번호 137 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 138, 서열 번호 149 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 150, 서열 번호 151 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 152 에 기재한다.
6)-1-11 #110 인간화 항체
상기로부터 설계된 중쇄 #110H1hIgG4P 와 경쇄 #110L4h 를 조합하고, 인간화 항체 #110H1L4hIgG4P 를 설계하였다. 또, 중쇄 #110H13hIgG4P 와 경쇄 #110L2h 를 조합하고, 인간화 항체 #110H13L2hIgG4P 를 설계하고, 중쇄 #110H13hIgG4P 와 경쇄 #110L12h 를 조합하고, 인간화 항체 #110H13L12hIgG4P 를 설계하였다.
6)-1-12 래트_CD147_#131 중쇄의 인간화
래트_CD147_#131 중쇄의 가변 영역에 대해, 도 17(a) 에 나타내는 억셉터의 아미노산 잔기를 이입함으로써, 인간화 항체 중쇄 #131H2h 의 가변 영역이 설계되었다.
설계된 가변 영역에, 인간의 IgG2 의 감마 (ga㎜a) 사슬 정상 영역을 접속시킨 인간화 항체 중쇄가 설계되고, #131H2hIgG2 라고 명명하였다. #131H2hIgG2 의 전체 길이 아미노산 서열을 서열 번호 139 에 기재한다. 서열 번호 139 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 140 에 기재한다.
6)-1-13 래트_CD147_#131 경쇄의 인간화
래트_CD147_#131 경쇄의 가변 영역에 대해, 도 17(b) 에 나타내는 억셉터의 아미노산 잔기를 이입함으로써, 인간화 항체 경쇄 #131L2h 의 가변 영역이 설계되었다.
설계된 가변 영역에, 인간의 κ 사슬의 정상 영역을 접속시킨 인간화 항체 경쇄가 설계되고, #131L2h 라고 명명하였다. #131L2h 의 경쇄 전체 길이 아미노산 서열을 서열 번호 141 에 기재한다. 서열 번호 141 의 아미노산 서열을 코드하는 뉴클레오티드 서열을 서열 번호 142 에 기재한다.
6)-1-14 #131 인간화 항체
상기로부터 설계된 중쇄 #131H2hIgG2 와 경쇄 #131L2h 를 조합하고, 인간화 항체 #131H2L2hIgG2 를 설계하였다.
6)-2 인간화 항체의 경쇄 발현 벡터의 구축
6)-2-1 #84L2h 발현 벡터의 구축
서열 번호 128 에 나타내는 #84L2h 의 뉴클레오티드 서열의 뉴클레오티드 번호 37 내지 402 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, pCMA-LK 를 제한 효소 BsiWI 로 절단한 지점에 합성한 DNA 단편을 삽입함으로써 #84L2h 발현 벡터를 구축하였다.
6)-2-2 #101L2h 발현 벡터의 구축
서열 번호 134 에 나타내는 #101L2h 의 뉴클레오티드 서열의 뉴클레오티드 번호 37 내지 402 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). 실시예 6)-2-1 과 동일한 방법으로 #101L2h 발현 벡터를 구축하였다.
6)-2-3 #110L4h, #110L2h 및 #110L12h 발현 벡터의 구축
서열 번호 138, 서열 번호 150 및 서열 번호 152 에 나타내는 #110L4h, #110L2h 및 #110L12h 의 뉴클레오티드 서열의 뉴클레오티드 번호 37 내지 402 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). 실시예 6)-2-1 과 동일한 방법으로 #110L4h, #110L2h 및 #110L12h 발현 벡터를 구축하였다.
6)-2-4 #131L2h 발현 벡터의 구축
서열 번호 142 에 나타내는 #131L2h 의 뉴클레오티드 서열의 뉴클레오티드 번호 37 내지 402 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). 실시예 6)-2-1 과 동일한 방법으로 #131L2h 발현 벡터를 구축하였다.
6)-3 인간화 항체의 중쇄 발현 벡터의 구축
6)-3-1 #84H1hIgG2 발현 벡터의 구축
서열 번호 124 에 나타내는 #84H1hIgG2 의 뉴클레오티드 서열의 뉴클레오티드 번호 36 내지 437 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, pCMA-G2 를 제한 효소 BlpI 로 절단한 지점에 합성한 DNA 단편을 삽입함으로써 #84H1hIgG2 발현 벡터를 구축하였다.
6)-3-2 #84H1hIgG4P 발현 벡터의 구축
서열 번호 126 에 나타내는 #84H1hIgG4P 의 뉴클레오티드 서열의 뉴클레오티드 번호 36 내지 437 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). In-Fusion HD PCR 클로닝 키트 (Clontech 사) 를 사용하여, pCMA-G4P 를 제한 효소 BlpI 로 절단한 지점에 합성한 DNA 단편을 삽입함으로써 #84H1hIgG4P 발현 벡터를 구축하였다.
6)-3-3 #101H1hIgG2 발현 벡터의 구축
서열 번호 130 에 나타내는 #101H1hIgG2 의 뉴클레오티드 서열의 뉴클레오티드 번호 36 내지 428 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). 실시예 6)-3-1 과 동일한 방법으로 #101H1hIgG2 발현 벡터를 구축하였다.
6)-3-4 #101H1hIgG4P 발현 벡터의 구축
서열 번호 132 에 나타내는 #101H1hIgG4P 의 뉴클레오티드 서열의 뉴클레오티드 번호 36 내지 428 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). 실시예 6)-3-2 와 동일한 방법으로 #101H1hIgG4P 발현 벡터를 구축하였다.
6)-3-5 #110H1hIgG4P 및 #110H13hIgG4P 발현 벡터의 구축
서열 번호 136 및 서열 번호 148 에 나타내는 #110H1hIgG4P 및 #110H13hIgG4P 의 뉴클레오티드 서열의 뉴클레오티드 번호 36 내지 425 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). 실시예 6)-3-2 와 동일한 방법으로 #110H1hIgG4P 및 #110H13hIgG4P 발현 벡터를 구축하였다.
6)-3-6 #131H2hIgG2 발현 벡터의 구축
서열 번호 140 에 나타내는 #131H2hIgG2 의 뉴클레오티드 서열의 뉴클레오티드 번호 36 내지 431 에 나타내는 DNA 단편을 합성하였다 (GENEART 사). 실시예 6)-3-1 과 동일한 방법으로 #131H2hIgG2 발현 벡터를 구축하였다.
6)-4 인간화 항체의 조제
6)-4-1 인간화 항체의 생산
실시예 1)-14-1 과 동일한 방법으로 생산하였다. 실시예 6)-1-5, 실시예 6)-1-8, 실시예 6)-1-11, 및 실시예 6)-1-14 에 나타낸 H 사슬과 L 사슬의 조합에 대응한 H 사슬 발현 벡터와 L 사슬 발현 벡터의 조합으로, 각종 인간화 항체를 취득하였다.
6)-4-2 인간화 항체의 1 단계 정제
실시예 6)-4-1 에서 얻어진 배양 상청으로부터 항체를 rProtein A 어피니티 크로마토그래피의 1 단계 공정으로 정제하였다. 배양 상청을 PBS 로 평형화한 MabSelectSuRe 가 충전된 칼럼 (GE Healthcare Bioscience 사 제조) 에 어플라이한 후에, 칼럼 용량의 2 배 이상의 PBS 로 칼럼을 세정하였다. 다음으로 2M 아르기닌 염산염 용액 (pH 4.0) 으로 용출하고, 항체가 포함되는 획분을 모았다. 그 획분을 투석 (Thermo Scientific 사, Slide-A-Lyzer Dialysis Cassette) 에 의해 PBS 로의 버퍼 치환을 실시하였다. Centrifugal UF Filter Device VIVASPIN20 (분획 분자량 UF10K, Sartorius 사) 로 항체를 농축하고, IgG 농도를 1 ㎎/㎖ 이상으로 조제하였다. 마지막으로 Minisart-Plus filter (Sartorius 사) 로 여과하고, 정제 샘플로 하였다.
6)-4-3 인간화 항체의 2 단계 정제
실시예 6)-4-1 에서 얻어진 배양 상청을 rProtein A 어피니티 크로마토그래피와 세라믹 하이드록시아파타이트의 2 단계 공정으로 정제하였다. 배양 상청을 PBS 로 평형화한 MabSelectSuRe 가 충전된 칼럼 (GE Healthcare Bioscience 사 제조) 에 어플라이한 후에, 칼럼 용량의 2 배 이상의 PBS 로 칼럼을 세정하였다. 다음으로 2 M 아르기닌 염산염 용액 (pH 4.0) 으로 항체를 용출하였다. 항체가 포함되는 획분을 투석 (Thermo Scientific 사, Slide-A-Lyzer Dialysis Cassette) 에 의해 PBS 로의 버퍼 치환을 실시하고, 5 mM 인산나트륨/50 mM MES/pH 7.0 의 버퍼로 5 배 희석한 후에, 5 mM NaPi/50 mM MES/30 mM NaCl/pH 7.0 의 버퍼로 평형화한 세라믹 하이드록시아파타이트 칼럼 (닛폰 바이오랏드, Bio-Scale CHT Type-1 Hydroxyapatite Column) 에 어플라이하였다. 염화나트륨에 의한 직선적 농도 구배 용출을 실시하고, 항체가 포함되는 획분을 모았다. 그 획분을 투석 (Thermo Scientific 사, Slide-A-Lyzer Dialysis Cassette) 에 의해 HBSor (25 mM 히스티딘/5 % 소르비톨, pH 6.0) 로의 버퍼 치환을 실시하였다. Centrifugal UF Filter Device VIVASPIN20 (분획 분자량 UF10K, Sartorius 사) 으로 항체를 농축하고, IgG 농도를 25 ㎎/㎖ 로 조제하였다. 마지막으로 Minisart-Plus filter (Sartorius 사) 로 여과하고, 정제 샘플로 하였다.
(실시예 7) 인간화 항체의 활성 측정
7)-1 인간화 항체의 CD147 항체 결합성 평가
실시예 6)-4-3 에서 제조한 인간화 항인간 CD147 항체 #84H1L2hIgG2, #84H1L2hIgG4P, #101H1L2hIgG2, #101H1L2hIgG4P, #110H1L4hIgG4P 및 #131H2L2hIgG2 와 인간 CD147 의 해리 정수 측정은, Biacore T200 (GE Healthcare Bioscience 사 제조) 을 사용하고, Human Antibody Capture Kit (GE Healthcare Bioscience 사 제조) 를 사용하여 고정화한 항 인간 IgG (Fc) 항체에 항체를 리간드로서 포착 (캡처) 하고, 항원을 아날라이트로서 측정하는 캡처법으로 실시하였다. 러닝 버퍼로서 HBS-EP+ (GE Healthcare Bioscience 사 제조), 센서 칩으로서 CM5 (GE Healthcare Bioscience 사 제조) 를 사용하였다. 칩 상에 1 ㎍/㎖ 의 인간화 항체를 10 ㎕/분으로 60 초간 첨가한 후, 실시예 5) 에서 사용한 항원의 희석 계열 용액 (#101H1L2hIgG2 및 #101H1L2hIgG4P 에 대해 0.0625 ∼ 1 ㎍/㎖, #84H1L2hIgG2, #84H1L2hIgG4P, #110H1L4hIgG4P, #110H13L2hIgG4P, #110H13L12hIgG4P 및 #131H2L2hIgG2 에 대해 0.25 ∼ 4 ㎍/㎖) 을 유속 30 ㎕/분으로 120 초간 첨가하고, 계속해서 300 초간의 해리상을 모니터하였다. 재생 용액으로서, 3M 마그네슘클로라이드 (GE Healthcare Bioscience 사 제조) 를 유속 20 ㎕/분으로 30 초간 첨가하였다. 데이터의 해석에는 1 : 1 결합 모델을 사용하여, 결합 속도 정수 ka, 해리 속도 정수 kd 및 해리 정수 (KD ; KD = kd/ka) 를 산출하였다.
결과를 표 6 에 나타낸다.
7)-2 췌장암 담암 모델에 있어서의 인간화 CD147 항체의 항종양 효과
5 × 106 세포의 인간 췌장주 MIA PaCa-2 를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 을 포함하는 PBS 로 현탁하고, 4 주령 암컷 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 찰스·리버로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 군나누기를 하고, 이식 7 일 후에 실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#84H1L2hIgG2, #84H1L2Ig4P, #101H1L2hIg2, #101H1L2hIg4P, #110H1L4hIg4P, #131H2L2hIg2) 를 3 ㎎/㎏, 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 대조약으로서 췌장암의 치료약인 겜시타빈 (닛폰 일라이릴리사로부터 구입) 을 이식 7, 14 일 후에 400 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
결과를 도 18(a) ∼ (d) 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
대조약의 종양 증식 억제율은, 71 % 인, 종양의 소실은 확인되지 않았던 데 반해, 어느 인간화 CD147 항체도, 3 혹은 10 ㎎/㎏ 투여군에서 겜시타빈보다 우수한 항종양 효과를 나타내었다. #84H1L2hIg4P, #101H1L2hIg2, #101H1L2hIg4P, #110H1L4hIG4P 의 10 ㎎/㎏ 투여군에서는, 종양의 소실을 수반하는 강한 항종양 효과가 확인되었다.
(실시예 8) CD147 항체의 in vitro p38MAPK 시그널의 활성화
8)-1 췌장암 세포 PANC-1 에 있어서의 항인간 CD147 인간 키메라 항체에 의한 p38MAPK 인산화의 유도
CD147 은 활성화에 의해 p38MAPK 의 인산화를 촉진하는 것이 보고되어 있다 (Lim et al., FEBS Letters, 88-92, 1998) (Li et., al. J. Hepatology, 1378-1389, 2015). 항종양 효과를 나타내는 항 CD147 이, p38MAPK 의 시그널에 미치는 영향을 조사하기 위해서, CD147 인간 키메라 항체 LN22R8chIgG4P 를 10 ㎍/㎖ 로 15 분간 처리한 PANC-1 세포에 대해, p38MAPK 인산화를 Simple Western assays 법 (프로테인 심플 재팬 주식회사, Wes) 을 사용하여 평가하였다. 대조 샘플로서, PANC-1 세포를 인간 IgG (Jackson I㎜unoResearch 사, 009-000-003) 10 ㎍/㎖ 를 동일하게 처리하여 해석에 제공하였다. p38MAPK 의 검출에는, p38 MAPK 토끼 ㎃b (Cell signaling technology 사, Cat. #9212) 를 사용하였다. 인산화 p38MAPK 의 검출에는, P-p38 MAPK (T180/Y182) (D3F9) XP 토끼 ㎃b (Cell signaling technology 사, #4511S) 를 사용하였다. 검출된 p38MAPK 시그널에 대한, 인산화 p38MAPK 시그널의 비를 도 19 에 나타내었다. 항체 처리 샘플은 이중으로 실시하고, 평균값을 도면에 나타내었다.
LN22R8chIgG4P 항체 처리군에서는, 인산화 MAPK 시그널의 증가가 관찰되었다.
8)-2 CD147 항체의 in vitro p38 시그널 하류 분자의 HSP27 의 인산화
p38MAPK 는 활성화에 의해, HSP27 을 인산화하는 것이 보고되어 있다 (Landry et al., Biochem. Cell Biol., 703-707, 1995). CD147 인간 키메라 항체 LN22R8 에 의해 유도되는 p38 의 인산화가, 실제의 p38 시그널 활성화를 촉진하고 있는지 확인하기 위해서 8)-1 의 항체 처리 샘플에 대해, 인산화를 Simple Western assays 법 (프로테인 심플 재팬 주식회사, Wes) 을 사용하여 평가하였다. HSP27 의 검출에는, HSP27 항체 (R&D systems 사, Cat. AF15801) 를 사용하였다. 인산화 HSP27 의 검출에는, Phospho-HSP27 (Ser82) antibody (Cell signaling technology 사, 2401S), 또는 항 HSP27 (phospho Ser15) 항체 (abcam 사, Cat. ab76313) 를 사용하였다.
검출된 HSP27 시그널에 대한, Ser82 또는 Ser15 인산화 HSP27 시그널의 비를, 각각, 도 20(a) 또는 도 20(b) 에 나타내었다. 항체 처리 샘플은 이중으로 실시하고, 평균값을 도면에 나타내었다.
LN22R8chIgG4P 항체 처리군에서는, 인산화 HSP27 시그널의 증가가 관찰되었다. CD147 인간 키메라 항체에 의해 유도된 p38 의 인산화에 의해 하류의 HSP27 의 인산화 활성화가 유도되고 있는 것으로부터, 항인간 CD147 인간 키메라 항체 LN22R8IgG4P 가 p38MAPK 시그널의 활성화를 유도하는 것을 알 수 있었다. 또, 췌장암 세포 MIA PaCa-2 에 있어서도 동일하게, p38MAPK 시그널의 활성화가 확인되었다.
(실시예 9) CD147 항체의 종양 내 p38 시그널의 활성화
in vitro 에서 관찰된 CD147 항체에 의한 p38MAPK 시그널의 활성화가, 마우스 피하에 형성한 종양 내에서도 일어나는지를 MIA PaCa-2 의 마우스 피하 종양으로 조사하였다. 5 × 106 세포의 인간 췌장주 MIA PaCa-2 를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 을 포함하는 PBS 로 현탁하고, 5 주령 암컷 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 찰스·리버로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 5 일 후에 군나누기를 실시하고, 항인간 CD147 인간 키메라 항체 LN22R8chIgG4P 를 군나누기 후 당일에 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다. 항체 투여 후, 6, 24, 48, 72 시간 후에 종양을 채재 (採材) 하고, 드라이아이스로 동결 후, 냉동 보존하였다. 동결한 종양 조직으로부터의 샘플 조제 (n = 3) 에는, gentleMACS Octo Dissociator with Heaters (밀테니사) 를 사용하였다. 조제한 종양 라이세이트에 대해, Simple Western assays 법 (프로테인 심플 재팬 주식회사, Wes) 을 사용하여 해석하였다.
검출된 p38MAPK 시그널의 평균값을 표준 오차와 아울러 도 21(a) 에 나타내었다. 검출된 인산화 p38MAPK 시그널의 평균값을 표준 오차와 아울러 도 21(b) 에 나타내었다. p38MAPK 시그널에 대한, 인산화 p38MAPK 시그널의 비의 평균값을 표준 오차와 아울러 도 21(c) 에 나타내었다. LN22R8chIgG4P 는, 투여 후 6 시간 내지 72 시간에서, 인산화 p38MAPK 의 비율을 증가시켰다. 24 시간 내지 72 시간에서는, p38MAPK 가 증가함으로써, 인산화 p38MAPK 의 시그널은 6 시간을 피크로 감소하고 있었다.
마우스의 피하 종양 내에서도 CD147 키메라 항체 투여에 의한 p38 인산화의 상승, HSP27 인산화의 증가가 관찰되고, p38 시그널의 활성화가 일어나고 있는 것을 알 수 있었다.
(실시예 10) p38MAPK 하류의 분자 CXCL8 의 유도
p38MAPK 의 활성화에 의해, CXCL8 의 mRNA 안정화를 통한 유도 (Hoffmann et al., J. Leukoc. Biol., 847-855, 2002), SMAD3/4 시그널의 활성화가 보고되어 있다 (Leovonen et al., PLOS ONE, e57474, 2013). MIA PaCa-2 종양 내에서, 항체 투여 후에 CXCL8 의 유전자 발현이 유도되는지 항체 투여 후의 마우스 피하 종양으로부터 추출한 RNA 를 정량적 PCR 법에 의해 조사하였다. 마찬가지로 SMAD 시그널 활성화에 의해 유도되는 것이 보고되어 있는 RHOB 유전자 (Vasilaki et al., FASEB Journal, 891-905, 2010) 에 대해서도 CD147 항체 투여에 의한 변동을 조사하였다. 내재 대조 유전자로서, importin (ipo8) 유전자와 TATA box binding protein (tbp) 유전자의 발현을 측정하였다. 항인간 CD147 인간 키메라 항체를 투여하고, 72 시간 후의 MIA PaCa-2 종양을 채재하고, RNA later (QIAGEN 사, Cat. 76104) 로 처리한 후, RNeasy Mini Kit (250) (QIAGEN 사, Cat. 74106) 을 사용하고, RNA 를 추출하였다. 항인간 CD147 인간 키메라 항체로서, 실시예 1)-14 에서 제조한 LN22R8chIgG1, LN22R8chIgG2 또는 LN22R8chIgG4P 를 사용하였다. 정량적 RT-PCR 에는, EXPRESS One-Step SuperScript qRT-PCR kit Universal (Thermofisher scienticic, Cat. 11781-01K) 을 사용하고, 유전자 정량 프로브로서, importin (ipo8) (Thermo Fisher Scientific, Cat. Hs00183533_m1), TATA box binding protein (tbp) (Thermo Fisher Scientific, Cat. Hs00427621_m1), rashomolog family member B (rhoB) (Thermo Fisher Scientific, Cat. hs00269660_s1), interleukin 8 (Thermo Fisher Scientific, Cat. Hs00174103_m1) 을 각각 사용하였다. PCR 반응에 수반하는 유전자 특이적 형광 시그널의 증가를 ABIPrism7500 (Applied Biosystems 사) 를 사용하여 측정하였다.
도 22(a) 에 ipo8/tbp 유전자의 발현비의 평균값 (n = 3) 을 표준 편차와 아울러 나타내었다.
도 22(b) 에 cxcl8/tbp 유전자의 발현비의 평균값 (n = 3) 을 표준 편차와 아울러 나타내었다.
도 22(c) 에 rhoB/tbp 유전자의 발현비의 평균값 (n = 3) 을 표준 편차와 아울러 나타내었다.
LN22R8chIgG1, LN22R8chIgG2 또는 LN22R8chIgG4P 투여 후에, ipo8 유전자의 발현은 변동하지 않았던 데 반해, 항인간 CD147 인간 키메라 항체 투여군에서의 cxcl8, rhoB 의 발현 유도가 확인되었다. cxcl8, rhoB 의 발현 유도는, 실시예 1)-18 에서 나타낸 인간 키메라 항체의 항종양 효과의 강도에 순응하고 있고, 양 유전자의 발현 유도가 항종양 효과와 상관할 가능성이 시사되었다.
(실시예 11) 항인간 CD147 래트 항체의 인간 키메라 항체화에 의한 항종양 효과의 증강에 있어서의, cxcl8, rhoB 유전자의 유도
항인간 CD147 래트 항체 래트_CD147_#110 은, 실시예 2, 실시예 7 에 나타낸 바와 같이, 인간 키메라화에 의해 항종양 효과가 증강하였다. 인간 키메라 항체 LN22R8chIgG1, LN22R8chIgG2 또는 LN22R8chIgG4P 투여 후의 종양 중에서 관찰된 cxcl8, rhoB 의 유도에 대해, 14)-2 와 마찬가지로 래트_CD147_#110 과 키메라 항체 #110chIgG4P 를 비교하였다.
도 23(a) 에 ipo8/tbp 유전자의 발현비의 평균값 (n = 3) 을 표준 편차와 아울러 나타내었다.
도 23(b) 에 cxcl8/tbp 유전자의 발현비의 평균값 (n = 3) 을 표준 편차와 아울러 나타내었다.
도 23(c) 에 rhoB/tbp 유전자의 발현비의 평균값 (n = 3) 을 표준 편차와 아울러 나타내었다.
ipo8 유전자의 발현은, 항체 투여에 의해 변동하지 않았다. cxcl8 유전자, rhoB 유전자의 발현은 래트_CD147_#110 의 투여에 의해, 항체 미투여군과 마찬가지로 변화가 보이지 않았던 데 반해, #110chIgG4P 에서의 cxcl8, rhoB 의 유도가 확인되었다. 양 유전자의 유도가 CD147 항체에 의한 항종양 효과와 상관하는 파라미터인 것이 시사되었다.
(실시예 12) SMAD4 음성 췌장암 담암 모델에 의한 평가
SMAD 시그널의 활성화에 중요한 분자의 하나로서, 전사 인자, SMAD4 가 알려져 있다 (Zang, et al., Current Biology, 270-276, 1997). 일부의 췌장암에서는, SMAD4 의 유전자적인 결손에 의해 SMAD 시그널이 부분적인 저해를 받고 있는 것이 알려져 있다 (Hahn, et al., Science, 350-353, 1996). SMAD4 가 결손하고, SMAD 시그널이 부분적으로 저해되어 있는 췌장암 세포주 BxPC-3 에 대해 (Yasutome et al., Clin. Exp. Metastasis, 461-473, 2005), CD147 항체가 항종양 효과를 나타내는지 조사하였다. 2.5 × 106 세포의 인간 췌장주 BxPC-3 (ATCC, Cat. CRL-1687) 을 100 % Matrigel (Corning 사, Cat. 354234) 을 포함하는 PBS 로 현탁하고, 6 주령 암컷 BALB/c-nu마우스 (CAnN. Cg-Foxn1nu/CrlCrlj, 닛폰 찰스·리버로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 후 8 일 후에 군나누기를 실시하고, 이식 8, 15, 22 일 후에 마우스 항인간 CD147 항체 LN22R8R8, 또는 래트 항인간 CD147 항체 2P10F2 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 대조약으로서 췌장암의 치료약인 겜시타빈 (닛폰 일라이릴리사, 젬자르 (등록상표)) 을 이식 8, 15, 22 일 후에 200 ㎎/㎏ 으로 담암 마우스의 꼬리 정맥내 투여하였다 (n = 5). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 24 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다. BxPC-3 종양은, LN22R8 항체, 2P10F2 항체 및 겜시타빈에 내성이었다.
(실시예 13) SMAD4 도입에 의한 CD147 항체 감수성의 획득
복수의 췌장암 세포주에서 SMAD4 를 도입함으로써 SMAD 시그널이 회복하는 것이 보고되어 있다. SMAD4 의 도입에 의해 회복한 SMAD 시그널에 의해 CD147 항체의 감수성이 증가하는지 조사하였다.
13)-1 SMAD4 안정 발현 세포의 제조
Retro-XTM Q 벡터 키트를 이용하여 BxPC-3 의 SMAD4 안정 발현주를 제조하였다. 레트로바이러스 벡터 (다카라 바이오사, Retro-XTM Q Vector Set, Cat. 631516) 로서 키트에 포함되는 pQCXIP 의 클로닝 부위에 인공 합성에 의해 제조한 인간 SMAD4 유전자를 도입하고, SMAD4 발현 레트로바이러스 벡터로 하였다. Retro-X Universal Packaging System (다카라 바이오사, Cat. 631530) 을 이용하고, BxPC-3 에 SMAD4 발현 레트로바이러스 벡터를 도입하고, 퓨로마이신 (다카라 바이오사, Cat. 631306) 에 의해 바이러스 감염에 의해 염색체에 레트로바이러스가 삽입되고, 퓨로마이신 내성, SMAD4 양성이 된 BxPC-3 세포를 선택하고, SMAD4 양성 BxPC-3 세포, BxPC-3-SMAD4 로 하였다. 레트로바이러스 벡터 pQCXIP 를 동일하게 감염시키고, 퓨로마이신 내성이 된 BxPC-3 세포를 BxPC-3-mock 로 하였다. 레트로바이러스 감염 실험은 2 회 실시하고, BxPC-3-mock, BxPC-3-SMAD4 에 대해, lot. 1, lot. 2 를 제조하였다.
13)-2 CD147 과 SMAD4 발현의 확인
BxPC-3 (ATCC, Cat. CRL-1687), 실시예 13)-1 에서 제조한 BxPC-3-mock, BxPC-3-SMAD4 에 대해, Simple Western assays 법 (프로테인 심플 재팬 주식회사, Wes) 을 사용하여 해석하였다. SMAD4 양성 대조 샘플로서, MIA PaCa-2 를 이용하였다. SMAD4 의 검출에는 항SMAD4 항체 (R&D systems, Cat. AF2097) 를 이용하였다. GAPDH 의 검출에는, 항 GAPDH 항체 (Abfrontier, Cat. LF-MA0026) 를 이용하였다. CD147 의 검출에는, 항 CD147 항체 (Abcam, Cat. Ab108317) 를 이용하였다.
결과를 도 25(a) 에 나타내었다. MIA PaCa-2 는 SMAD4 양성, CD147 양성이었다. BxPC-3 은 SMAD4 음성, CD147 양성이었다. BxPC-3-mock 는, 레트로바이러스 감염에 의한 영향은 보이지 않고, SMAD4 음성, CD147 양성이었다. SMAD4 발현 레트로바이러스 벡터를 도입한, BxPC-3-SMAD4 는, lot. 1, lot. 2 모두 SMAD4 양성이었다. Lot. 2 의 SMAD4 발현이 높아, 후의 실험에는, lot. 2 를 BxPC-3-SMAD4 로서 이용하였다.
13)-3 SMAD4 안정 발현한 BxPC-3 종양의 CD147 인간 키메라 항체에 대한 감수성
2.5 × 106 세포의 BxPC-3-mock, 또는 BxPC-3-SMAD4 를 100 % Matrigel (Corning 사, Cat. 354234) 을 포함하는 PBS 로 현탁하고, 5 주령 암컷 BALB/c-nu마우스 (CAnN. Cg-Foxn1nu/CrlCrlj, 닛폰 찰스·리버로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 BxPC-3-mock 이식 후 6 일 후에, BxPC-3-SMAD4 는 이식 후 3 일 후에 군나누기를 각각 실시하고, 군 나누기 실시 당일, 7, 14, 21, 28 일 후에 인간 키메라 항인간 CD147 항체 LN22R8R8chIgG2, 또는 LN22R8R8chIgG4P 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 25(b), 도 25(c) 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다. BxPC-3-mock 종양은, LN22R8R8chIgG2, 또는 LN22R8R8chIgG4P 항체에 내성 또는, 저감수성이었다. BxPC-3-SMAD4 종양은, LN22R8R8chIgG2, 또는 LN22R8R8chIgG4P 항체에 부분적인 감수성을 나타내었다.
13)-4 SMAD4 발현에 의한 종양 중 p38 시그널의 변화
SMAD4 음성의 췌장암 세포에서는, SMAD4 를 발현시킴으로써, p38 시그널이 증강하는 것이 보고되어 있다 (Chen et al., B.M.C., 1471-2407, 2014). BxPC-3-SMAD4 종양 중의 p38MAPK 와 인산화 p38MAPK 에 대해, 항인간 CD147 인간 키메라 항체 투여 (항체 투여 후 72 시간 후) 에 의한 변동에 대해, 실시예 13)-1 과 마찬가지로 Simple Western assays 법 (프로테인 심플 재팬 주식회사, Wes) 을 사용하여 해석하였다. 실시예 1)-18 의 항과 마찬가지로, 항인간 CD147 인간 키메라 항체로서, LN22R8chIgG4P 를 10 ㎎/㎏, MIA PaCa-2 피하 종양을 생착한 마우스에 투여하였다. 종양 조직으로부터의 샘플 조제 (n = 3) 에는, gentleMACS Octo Dissociator with Heaters (밀테니사) 를 사용하였다.
도 26(a) 에 p38 MAPK 측정값에 대해, 3 개의 종양 샘플의 측정값의 평균값과 표준 오차를 아울러 그래프에 기재하였다. 도 26(b) 에 인산화 p38 MAPK 측정값에 대해, 3 개의 종양 샘플의 측정값의 평균값과 표준 오차를 아울러 그래프에 기재하였다.
BxPC-3-SMAD4 종양 중에서는, p38 과 인산화 p38 이 2 배 이상으로 증가하고 있었다. CD147 인간 키메라 항체 투여 후 72 시간 후에서는, p38 의 부분적인 감소, 인산화 p38 의 부분적인 증가가 관찰되었다.
췌장암 세포 BxPC-3 의 종양에서는, SMAD4 의 발현에 의해, p38 시그널의 증강이 일어나는 것을 알 수 있었다. CD147 인간 키메라 항체 LN22R8chIgG4P 의 감수성 증가에, SMAD4 의존적인 p38 시그널의 증강이 기여하고 있을 가능성이 생각된다.
(실시예 14) 겜시타빈 내성 췌장암에 있어서의 키메라 CD147 항체의 항종양 효과
겜시타빈 내성의 췌장암 종양 모델에서의 CD147 항체의 항종양 효과를 조사하였다. 5 × 106 세포의 인간 췌장주 MIA PaCa-2 를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 을 포함하는 PBS 로 현탁하고, 5 주령 암컷 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 찰스·리버) 의 액와부 피하에 이식하였다. 이식 6 일 후에 췌장암의 치료약인 겜시타빈 (닛폰 일라이릴리사) 을 400 ㎎/㎏ 복강내 투여하고, 1 주일 후에 증식이 확인된 겜시타빈 내성 종양을 형성한 마우스를 종양의 사이즈에 기초하여 군나누기를 실시하고, 대조군 (n = 5) 에는, 겜시타빈을 이식 후 13 일째 (군나누기일), 20 일째에 400 ㎎/㎏ 복강내 투여하였다. CD147 항체와 겜시타빈 병용 투여군 (n = 5) 으로서, 겜시타빈을 이식 후 13 일째 (군나누기일), 20 일째에 400 ㎎/㎏ 복강내 투여하는 것에 더하여, CD147 인간 키메라 항체 LN22R8chIgG4P 를 이식 후 13 일째 (군나누기일), 20 일째에 10 ㎎/㎏ 복강내 투여하였다. 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
결과를 도 27 에 나타낸 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다. 겜시타빈 (Gemcitabin) 과 항체를 투여하고 있지 않은 미투여군 (non treatment) 의 종양 증식을 아울러 기재하였다 (n = 5).
대조약의 겜시타빈의 투여군의, 종양의 평균 사이즈는, 이식 후 28 일 후에 1269 ㎣ 이고 퇴축하는 종양은 확인되지 않았던 데 반해, CD147 인간 키메라 항체 LN22R8chIgG4P 를 병용 투여한 군의 종양의 평균 사이즈는, 15 ㎣ 이고, 5 예 중 3예에서의 종양의 퇴축이 확인되었다. 이 결과로부터, 겜시타빈에 내성을 나타내고 증식하는 췌장암 종양에 대해서도 CD147 항체에 대한 감수성을 나타낼 가능성이 나타났다.
(실시예 15) 간장암 모델에 있어서의 인간화 CD147 항체의 항종양 효과
15)-1 Hep G2 세포에 있어서의 CD147 과 SMAD4 의 발현
실시예 13)-2 와 마찬가지로, 간장암 세포주 HepG2 세포 (ATCC, Cat. HB-8065) 의 CD147 과 SMAD4 발현에 대해 확인하였다. CD147 양성의 대조 검체로서, MIA PaCa-2, BxPC-3 (ATCC, Cat. CRL-1687) 을 사용하였다. SMAD4 양성의 대조 검체로서, MIA PaCa-2, SMAD4 음성의 대조 검체로서, BxPC-3 을 각각 사용하였다.
검출된 시그널에 대해, GAPDH 에 대한 시그널비를 구하고, 하기에 정리하였다. Hep G2 세포는, CD147, SMAD4 양성인 것을 알 수 있었다.
CD147/GAPDH 의 시그널비
MIA PaCa-2 0.481
BxPC-3 0.481
Hep G2 2.944
SMAD4/GAPDH 의 시그널비
MIA PaCa-2 0.329
BxPC-3 0.003
Hep G2 0.723
15)-2 Hep G2 세포에 있어서의 CD147 의 플로우 사이토미터에 의한 발현 확인
HepG2 세포 (ATCC, Cat. HB-8065) 의 세포 표면에 발현하는 CD147 을 플로우 사이토미터에 의해 해석하였다. 인간 CD147 의 발현 확인에는, 시판되는 항인간 CD147 항체로서 APC 라벨된 항인간 CD147 마우스 IgG1 항체 MEM-M6/1-APC (Thermofisher, Cat. MA1-10104) 를 사용하였다. 마우스 IgG1 아이소타입 컨트롤 항체로서 mIgG1-APC (밀테니바이오사, Cat. 130-092-214) 를 사용하였다. HepG2 세포의 현탁액에, 1/10 량의 MEM-M6/1-APC 를 첨가하고, 4 ℃, 30 분간 처리하였다. 5 % FBS 를 포함하는 PBS 완충액으로 세포를 세정 후, 플로우 사이토미터 (CantoII, BD Bioscience 사) 의 측정을 실시하고, 도 28(a) 에 결과를 정리하였다.
MEM-M6/1-APC 로 처리한 Hep G2 세포는, 강한 형광 시그널을 나타내고, CD147 을 발현하고 있는 것을 알 수 있었다.
15)-3 간장암 세포에 있어서의 인간화 CD147 항체에 의한 p38 활성화
간장암 세포 HepG2 에 있어서, 항 CD147 항체가, p38MAPK 에 미치는 영향을 조사하기 위해서, 항인간 CD147 인간 키메라 항체 (LN22R8chIgG4P) 또는, 실시예 6)-4-2 에서 제조한 항인간 CD147 인간화 항체 (#84H1L2hIgG2, #84H1L2hIg4P, #101H1L2hIgG2, #101H1L2hIgG4P, #110H1L4hIg4P, #131H2L2hIgG2) 를 10 ㎍/㎖ 로 15 분간 처리한 HepG2 세포 (ATCC, Cat. HB-8065) 에 대해, P38 인산화를 Simple Western assays 법 (프로테인 심플 재팬 주식회사, Wes) 을 사용하여 평가하였다. 대조 샘플로서, HepG2 세포를 인간 IgG (hIgG, Jackson I㎜unoResearch 사, 009-000-003) 10 ㎍/㎖ 를 동일하게 처리하고 해석에 제공하였다. p38 MAPK 의 검출에는, p38 MAPK 토끼 ㎃b (Cell signaling technology 사, Cat. #9212) 를 사용하였다. 인산화 p38MAPK 의 검출에는, P-p38 MAPK (T180/Y182) (D3F9) XP 토끼 ㎃b (Cell signaling technology 사, #4511S) 를 사용하였다. 검출된 p38MAPK 시그널에 대한, 인산화 p38MAPK 시그널의 비를 도 28(b) 에 나타내었다.
LN22R8chIgG4P, #84H1L2hIgG2, #84H1L2hIg4P, #101H1L2hIgG2, #101H1L2hIgG4P, #110H1L4hIg4P 또는 #131H2L2hIgG2 의 처리에 의해, 인간 IgG 처리군으로부터 2 배 이상의 인산화 p38 MAPK 시그널의 증가가 관찰되고, 간장암 세포에서도 췌장암 세포와 동일하게 항인간 CD147 항체에 의해 p38MAPK 의 인산화가 유도되고 있는 것을 알 수 있었다.
15)-4 간장암에 있어서의 인간화 CD147 항체와 솔라페닙의 항종양 효과의 비교
CD147 과 SMAD4 양성에서, 항인간 CD147 항체에 의한 p38 의 인산화가 확인된 인간 간장주 HepG2 (ATCC, Cat. HB-8065) 에 대해, 항인간 CD147 인간 키메라 항체와 인간화 항체의 항종양 효과를 검토하였다.
5 × 106 세포의 인간 간장주 HepG2 를 50 % Matrigel (Corning 사, Cat. 354234) 을 포함하는 PBS 로 현탁하고, 4 주령 암컷 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 찰스·리버로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 후 9 일 후에 군나누기를 실시하고, 인간 키메라 CD147 항체 (LN22R8chIgG4P), 실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#84H1L2hIgG2, #84H1L2hIg4P, #110H1L4hIg4P) 를 1 ㎎/㎏, 10 ㎎/㎏ 으로 담암 마우스의 복강내에 투여하였다 (n = 5). 대조약으로서 간장암의 치료약인 솔라페닙 (넥사바정 200 mg, 바이엘 약품 주식회사) 을 넥사바정 첨부 문서의 참고 문헌 (Chang et. al., Cancer Chem. Thera. Pharm., 2007) 에 따라서 PEG-35 피마자유 (Cremophor EL, 나칼라이테스크사, Cat. 09727-14) 에탄올 용매에 용해하고, 이식 9, 10, 11, 12, 13, 16, 17, 18 일 후에 30 ㎎/㎏, 90 ㎎/㎏ 으로 담암 마우스에 경구 투여하였다 (n = 5). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 표 7 및 도 29(a) ∼ (d) 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
솔라페닙 투여는, 종양 증식의 부분적인 저해가 관찰되었지만, 종양의 소실은 확인되지 않았다. LN22R8chIgG4P, #84H1L2hIgG2, #84H1L2hIg4P, #110H1L4hIg4P 는, 모두 종양의 소실을 수반하는 우수한 항종양 효과를 나타내었다.
(실시예 16) T 세포 및 PBMC 에 대한 작용
CD147 은 T 세포의 활성화에 수반하여 CD4 양성, CD8 양성의 T 세포에서 발현이 상승하고 (Hu et al., J. Cell. Mol Med., 2132-2143, 2010), 일부의 CD147 항체에서는, T 세포의 활성화 유도능이나 증식을 저해하는 효과가 보고되어 있다 (Koch et al., Int. I㎜unology, 777-786, 1999 ; Chiampanichayakul et al., I㎜unology 167-178, 2006). 이펙터 기능에 의존하지 않고, 강한 항종양 효과를 나타내는 항인간 CD147 항체가, T 세포를 포함하는 말초혈 림프구 (PBL) 에 미치는 영향을 조사하였다.
16)-1 PBMC 의 CD3/CD28 자극에 의한 CD147 의 발현 상승
CD147 의 발현이 T 세포의 활성화에 수반하여 상승하는지, 인간 PBMC 를 사용하여 조사하였다. 인간 PBMC 를 10 % FBS 를 포함하는 RPMI1640 배지에서, 37 ℃, 5 % CO2 존재하에서 배양하였다. 배양 시에, Dynabeads Human T-Activator CD3/CD28 (CD3/CD28 beads, Thermofishers scientific 사, Cat. 1131D) 을 첨가하여 증식을 유도하고, 4 일 후에 플로우 사이토미터의 해석을 실시하고, CD147 의 발현이 변화하는지 조사하였다. 인간 CD147 의 발현 확인에는, 시판되는 항인간 CD147 항체로서 APC 라벨된 항인간 CD147 마우스 IgG1 항체 MEM-M6/1-APC (CD147-APC, Thermofisher, Cat. MA1-10104) 를 사용하였다. 마우스 IgG1 아이소타입 컨트롤 항체로서 mIgG1-APC (밀테니바이오사, Cat. 130-092-214) 를 사용하였다. 인간 PBMC 에 포함되는 T 세포의 CD3, CD4, CD8 을 검출하기 위해서, APC/FireTM 750 항 인간 CD3 항체 (BioLegend 사 제조, Cat. 344840), PerCP/Cy5.5 항 인간 CD4 (BioLegend 사 제조, Cat. 344608), Brilliant Violet 510 항 인간 CD8 (BioLegend 사 제조, Cat. 344732) 을 사용하였다.
CD3, CD4 양성의 세포 및 CD3, CD8 양성의 세포에 있어서의 CD147-APC 의 결합을 도 30 에 정리하였다. CD3, CD4 양성의 세포 및 CD3, CD8 양성의 세포에 있어서, CD3/CD28 비드 자극했을 경우에, CD147 의 발현 상승이 확인되고, T 세포의 활성화에 수반하여 CD147 의 T 세포막 표면 상의 발현이 상승하는 것이 확인되었다.
16)-2 인간 말초혈 단핵구의 증식에 있어서의 항인간 CD147 항체 작용의 평가
인간 말초혈 단핵구 (PBMC) 의 증식에 있어서의 항인간 CD147 항체 작용을 해석하였다. 항인간 CD147 항체로서, 2P10F2chIgG4P 를 사용하였다. CellVue Claret Far Red Fluorescent Cell Linker Kit (sigma, Cat. MIDCLARET-1 KT) 를 사용하여, PBMC 를 형광 라벨한 후에, 10 % FBS 를 포함하는 RPMI1640 배지에서, 37 ℃, 5 % CO2 존재하에서 배양하였다. 배양 시에, IL-2, Dynabeads Human T-Activator CD3/CD28 (CD3/CD28 beads, Thermofishers scientific 사, Cat. 1131D) 을 첨가하고 증식을 유도했을 때에 2P10F2chIgG4P (10 ㎍/㎖) 를 첨가하여, 증식에 대한 영향을 조사하였다. 배양 3 일, 5 일째에, 세포 분열에 의해 감소한 PBMC 세포 형광 시그널에 대해 플로우 사이토미터 (CantoII, BD Bioscience 사) 를 사용한 측정을 실시하고, 도 31 에 결과를 정리하였다.
CD3/CD28 비드의 첨가에 의해, 세포 분열에 의해 감소한 PBMC 세포 형광 시그널이, 배양 3 일 후, 5 일 후에 관찰되었다. IL-2, 2P10F2chIgG4P, 또는 IL-2 와 2P10F2chIgG4P 를 배양 시에 첨가했을 경우에, 형광 시그널의 감소에 변화는 보이지 않았다. 실시예 1)-18 에서 나타낸 인간 췌장암의 마우스 종양 모델에 있어서 강한 항종양 효과를 나타낸 항인간 CD147 인간 키메라 항체 2P10F2chIgG4P 는, PBMC 의 증식에는, 영향을 주지 않는 것이 시사되었다.
16)-3 인간 말초혈 림프구의 사이토카인 산생에 있어서의 항인간 CD147 항체의 평가
항인간 CD147 항체로서, 인간 키메라 항체 #84chIgG1, #84chIgG2, #84chIgG4P, #84chIgG1LALA, #84chIgG4PFALA, #101chIgG4P 또는 #110chIgG4P 를 사용하였다. 인간 말초혈로부터 Ficoll-Paque PLUS (GE 헬스케어·재팬 주식회사) 를 사용하여, PBL 을 조제하였다. 96 웰 플레이트에 10 ㎍/㎖ 의 인간 키메라 항체를 코트하였다. 음성 컨트롤 항체로서 인간 IgG (hIgG, Jackson I㎜unoResearch 사, 009-000-003), T 세포의 활성화나 사이토카인 유도를 유도하는 양성 컨트롤 항체로서 Dynabeads Human T-Activator CD3/CD28 (CD3/CD28-beads, Thermofishers scientific 사, Cat. 1131D) 을 사용하였다. 항체를 코트한 웰에 1 x 106 의 PBL 을 첨가하고, 24 시간 후에, 배지 중의 사이토카인 (IL2, TNFα, INFγ) 을 측정하였다. Dynabeads Human T-Activator CD3/CD28 은, PBL 을 첨가한 웰에 직접 첨가하고, 동일하게 24 시간 후에, 배지 중의 사이토카인 (IL2, TNFα, INFγ) 을 측정하였다. IL2 의 측정에는, Quantikine ELISA Human IL-2 (R&D systems, Cat. D2050) 를 사용하였다. TNFα 의 측정에는, Amersham TNF-α Human, Biotrak Easy ELISA (GE 헬스케어·재팬 주식회사, Cat. RPN5967) 를 사용하였다. INFγ 의 측정에는, Human IFN-γ ELISA development kit (MABTECH 사, Cat. 3420-1H-6) 를 사용하였다. 측정은 삼중으로 실시하고, 검출된 흡광도의 평균값과 표준 편차를 산출하고, 도 32 에 정리하였다.
측정한 PBL 로부터 산생된 사이토카인 (IL2, TNFα, INFγ) 은, 모두 양성 컨트롤인 CD3/CD28-비드를 첨가한 웰에서의 배양에서만 상승이 확인되고, 어느 항인간 CD147 인간 키메라 항체 코트한 웰에서의 배양에서도, 음성 컨트롤인 hIgG 와 마찬가지로 사이토카인의 상승은 확인되지 않았다.
(실시예 17) 인간화 #110Fab'-CD147 복합체의 X 선 결정 구조 해석
17)-1 복합체의 결정화
인간화 #110H1L4hIgG4P 를 펩신 (Pepsin) 에 의해 절단하여 얻어진 Fab'2 를, 디티오트레이톨 (Dithiothreitol) 로 환원한 후, 아이오도아세타마이드 (Iodoacetamide) 에 의해 알킬화함으로써 Fab' 단편을 취득하였다. 이 Fab' 단편과 실시예 5) 에서 사용한 hCD147v2(22-205) 의 혼합물을 Superdex 10/300GL Increase 칼럼 (GE Healthcare) 을 사용한 겔 여과 크로마토그래피에 공여하고, 복합체 획분을 취득하였다. 복합체는 AmiconUltra15 MWCO 10K (밀리포어사 제조) 로 완충액 (10 mM Tris HCl pH 7.5, 50 mM NaCl) 으로 치환하고, 13 g/ℓ 로 농축하였다. 복합체 용액을 증기 확산법에 의해 결정화하였다. 단백질 용액 0.5 ㎕ 에 침전제 용액 (0.1 M NaMalonate pH 7.0, 12 % (w/v) 폴리에틸렌 글리콜 3350) 을 등량 첨가한 용액을, 0.05 ㎖ 의 침전제 용액을 넣은 밀폐 용기에 양쪽 용액이 접촉하지 않도록 넣고, 25 ℃ 에서 정치 (靜置) 하였다. 약 1 주일 후에 얻어진 0.15 ㎜ × 0.15 ㎜ × 0.3 ㎜ 의 결정을 30 % (w/v) 가 되도록
폴리에틸렌 글리콜 400 (Polyethylene Glycol 400) 을 첨가한 침전제 용액에 담그고 나서 액체 질소로 동결하였다. 방사광 시설 포톤 팩토리 (이바라키현, 일본) 의 빔 라인 PF-BL17A 로 X 선 회절 데이터를 수집하였다. 얻어진 회절상으로부터 소프트웨어 mosflm (CCP4 : Collaborative Computational Project No.4) 를 사용하여 회절 강도를 수치화하고, 결정 구조 인자를 구하였다. 결정의 공간군은 P21, 결정의 단위격자는 (a = 64.96 Å, b = 93.37 Å, c = 98.31 Å, 알파 (alpha) = 감마 (ga㎜a) = 90, 베타 (beta) = 90.89) 였다.
17)-2 복합체의 구조 해석
얻어진 구조 인자와 Fab' 단편의 호몰로지 모델 및 인간 CD147 의 이미 알려진 구조 (PDBID : 3b5h) 의 삼차원 구조 좌표를 사용하여 분자 치환법을 실시하고, 위상을 결정하였다. 계산에는 소프트웨어 phaser (CCP4 : Collaborative Computational Project No.4) 를 사용하였다. 결정은 비대칭 단위에 2 개의 복합체를 포함하고 있었다. 소프트웨어 Refmac5 (CCP4 : Collaborative Computational Project No.4) 를 사용하여 구조의 정밀화를 실시하고, 소프트웨어 coot 를 사용하여 모델의 수정을 실시하였다. 이 조작을 반복 실시하고, 2.3 Å 분해능으로 최종의 R 값 23 %, 프리 R (free R) 값 28 % 를 얻었다. 최종 모델은 인간화 #110H1L4 의 Fab' 단편을 2 개와, 각각에 결합하는 hCD147v2 를 포함한다. 또한, 편방의 hCD147v2 에 대해서는 아미노산 잔기 23-203 에 상당하는 전자 밀도가 확인되었지만, 다른 편방에 대해서는 도메인 1 에 상당하는 전자 밀도는 명료하지 않고, 아미노산 잔기 103-202 에 상당하는 전자 밀도만 확인되었다.
17)-3 에피토프의 특정
비대칭 단위에 포함되는 2 개의 복합체에 있어서 공통되어 인간화 #110H1L4 의 Fab' 단편의 결합면으로부터 4 Å 이내에 있는 hCD147v2 의 아미노산 잔기는 이하와 같다 : Arg106, Lys108, Ala109, Val110, Lys127, Ser128, Glu129, Ser130, Val131, Pro132, Pro133, Val134, Gln164, Gly165. 도 41 에 복합체 전체의 리본 모델과 표면을, 도 42 에 hCD147v2 와 인간화 #110H1L4 의 상호 작용을 나타내었다.
(실시예 18) 위암 모델에 있어서의 인간화 CD147 항체의 항종양 효과
플로우 사이토미터에 의해 CD147 양성이 확인된 인간 위암 세포주 KATO III 세포 (ATCC, Cat. HTB-103) 에 대해, 항인간 CD147 인간 키메라 항체와 인간화 항체의 항종양 효과를 검토하였다.
5 × 106 세포의 인간 위암주 KATO III 을 100 % Matrigel (Corning 사, Cat. 354234) 로 현탁하고, 5 주령 암컷의 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 쿠레아로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 후 3 일 후에 군나누기를 실시하고, 인간 키메라 CD147 항체 (LN22R8chIgG4P), 실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#110H1L4hIg4P) 를 10 ㎎/㎏ 으로 담암 마우스의 복강내에, 군나누기 후 7 일마다 투여하였다 (n = 6). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 43 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
미처리군의 마우스의 종양의 평균 체적은, 이식으로부터 24 일 후에 290 ㎣ 인 데 반해, LN22R8chIgG4p 투여군에서는, 199 ㎣, #110H1L4hIgG4P 투여군의 것에서는, 134 ㎣ 이고, 산출된 항종양 효과는, LN22R8chIgG4p 투여군에서 31 %, #110H1L4hIgG4P 투여군에서는, 54 % 였다.
(실시예 19) 만성 골수성 백혈병 모델에 있어서의 인간화 CD147 항체의 항종양 효과
플로우 사이토미터에 의해 CD147 양성이 확인된 인간 만성 골수성 백혈병 세포주 KU812 세포 (ATCC, Cat. CRL-2099) 에 대해, 항인간 CD147 인간화 항체의 항종양 효과를 검토하였다.
5 × 106 세포의 만성 골수성 백혈병 세포주 KU812 를 50 % Matrigel (Corning 사, Cat. 354234) 을 포함하는 PBS 로 현탁하고, 5 주령 암컷의 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 쿠레아로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 후 3 일 후에 군나누기를 실시하고, 실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#110H1L4hIg4P) 를 10 ㎎/㎏ 으로 담암 마우스의 복강내에, 군나누기 후 7 일마다 투여하였다 (n = 5). 대조약으로서 만성 골수성 백혈병의 치료약인 이마티닙 (ASTATECH 사, Cat. #63168) 을 9 ㎎/㎖ 용액에 증류수로 조제하고, 90 ㎎/㎏ 으로 담암 마우스에 경구 투여하였다 (토일 제외한 연속 투여, 이식 후 4, 7, 8, 9, 10, 11, 14, 15, 16, 17, 18, 21, 22, 23, 24 일 후에 투여). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 44 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
미처리군의 마우스의 종양의 평균 체적은, 이식으로부터 25 일 후에 627 ㎣ 인 데 반해, 이마티닙 투여군에서는, 328 ㎣, #110H1L4hIgG4P 투여군의 것에서는, 6 ㎣ 이고, 산출된 항종양 효과는, 이마티닙 투여군에서 48 %, #110H1L4hIgG4P 투여군에서는, 97 % 였다. #110H1L4hIgG4P 투여군에서만, 5 예 중 4 예에서 종양의 완전 축퇴가 관찰되었다.
(실시예 20) 대장암 모델에 있어서의 인간화 CD147 항체의 항종양 효과
플로우 사이토미터에 의해 CD147 양성이 확인된 인간 대장암 세포주 SW620 세포 (ATCC, Cat. CCL-227) 에 대해, 인간화 항체의 항종양 효과를 검토하였다.
5 × 106 세포의 인간 대장암주 SW620 을 100 % Matrigel (Corning 사, Cat. 354234) 로 현탁하고, 5 주령 암컷의 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 쿠레아로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 후 3 일 후에 군나누기를 실시하고, 인간 키메라 CD147 항체 (LN22R8chIgG4P), 실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#084H1L2hIg4P 혹은 #110H1L4hIg4P) 를 10 ㎎/㎏ 으로 담암 마우스의 복강내에, 군나누기 후 7 일마다 투여하였다 (n = 5). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 45 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
미처리군의 마우스의 종양의 평균 체적은, 이식으로부터 21 일 후에 1302 ㎣ 인 데 반해, #084H1L2hIg4P 투여군에서는, 709 ㎣, #110H1L4hIgG4P 투여군의 것에서는, 403 ㎣ 이고, 산출된 항종양 효과는, #084H1L2hIg4P 투여군에서 46 %, #110H1L4hIgG4P 투여군에서는, 69 % 였다.
(실시예 21) 신장암 786-O 모델에 있어서의 인간화 CD147 항체의 항종양 효과
플로우 사이토미터에 의해 CD147 양성이 확인된 인간 신장암 786-O 에 대해, 인간화 항체의 항종양 효과를 검토하였다.
5 × 106 세포의 인간 신장암 786-O 를 50 % Matrigel (Corning 사, Cat. 354234) 로 현탁하고, 5 주령 암컷의 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 쿠레아로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 후 3 일 후에 군나누기를 실시하고, 인간 키메라 CD147 항체 (LN22R8chIgG4P), 실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#084H1L2hIg4P 혹은 #110H1L4hIg4P) 를 10 ㎎/㎏ 으로 담암 마우스의 복강내에, 군나누기 후 7 일마다 합계 4 회 투여하였다 (n = 6). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 46 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
미처리군의 마우스의 종양의 평균 체적은, 이식으로부터 31 일 후에 918 ㎣ 인 데 반해, #084H1L2hIg4P 투여군에서는, 224 ㎣, #110H1L4hIgG4P 투여군의 것에서는, 379 ㎣ 이고, 산출된 항종양 효과는, #084H1L2hIg4P 투여군에서 76 %, #110H1L4hIgG4P 투여군에서는, 59 % 였다.
(실시예 22) 급성 골수성 백혈병 (AML) 모델에 있어서의 인간화 CD147 항체의 항종양 효과
플로우 사이토미터에 의해 CD147 양성이 확인된 인간 AML 세포주 OCI-AML3 세포 (DSMZ, Cat. ACC 582) 에 대해, 인간화 항체의 항종양 효과를 검토하였다.
5 × 106 세포의 인간 AML 세포주 OCI-AML3 세포를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 로 현탁하고, 5 주령 암컷의 NOD-scid 마우스 (NOD. CB17-Prkdc<scid>/J, 닛폰 쿠레아로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 이식 후 3 일 후에 군나누기를 실시하고, 실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#110H1L4hIg4P) 를 꼬리 정맥 주사에 의해, 10 ㎎/㎏ 으로 담암 마우스로 군나누기 후 7 일마다 투여하였다 (n = 6). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
종양 체적 (㎣) = 1/2 × 단경 (㎜) × 단경 (㎜) × 장경 (㎜)
결과를 도 47 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
미처리군의 마우스의 종양의 평균 체적은, 이식으로부터 21 일 후에 1533 ㎣ 인 데 반해, #110H1L4hIgG4P 투여군 것에서는, 394 ㎣ 이고, 산출된 항종양 효과는, #110H1L4hIgG4P 투여군에서는, 74 % 였다.
(실시예 23) 결합성이 상이한 인간화 항 CD147 항체의 활성 측정
5 × 106 세포의 인간 췌장주 MIA PaCa-2 를 50 % GFR-Matrigel (Corning 사, Cat. 354230) 을 포함하는 PBS 로 현탁하고, 4 주령 암컷의 Nude 마우스 (BALB/c Slc-nu/nu, 닛폰 에스엘씨 (주) 로부터 구입) 의 액와부 피하에 이식하였다. 종양 체적을 기초로 군나누기를 하고, 이식 7 일 후에 실시예 6)-4-2 에서 제조한 CD147 단백질에 대한 결합성이 상이한 3 종의 인간화 CD147 항체 (#110H1L4hIgG4P, #110H13L02hIgG4P 및 #110H13L12hIgG4P, 표 6 에 결합성을 기재) 를 10 ㎎/㎏ 으로 담암 마우스에 꼬리 정맥 투여하였다 (n = 6). 대조약으로서 췌장암의 치료약인 겜시타빈 (닛폰 일라이릴리사로부터 구입) 을 이식 3, 10 일 후에 400 ㎎/㎏ 으로 담암 마우스에 꼬리 정맥 투여하였다 (n = 6). 이식 종양의 장경 및 단경을 주 2 회, 전자 디지털 노기스 (주식회사 미츠토요 제조) 를 사용하여 측정하고, 이하에 나타내는 계산식에 의해 종양 체적을 산출하였다.
결과를 도 48 에 나타내었다. 그래프에는 종양 체적의 변화에 대해, 평균값과 표준 오차를 아울러 기재하였다.
대조약의 겜시타빈의 종양 증식 억제율은, 66 % 였던 데 반해, 어느 인간화 CD147 항체도, 10 ㎎/㎏ 투여군에서 겜시타빈보다 우수한 항종양 효과를 나타내었다.
(실시예 24) CD147 항체의 비교
24)-1 기존의 항 CD147 항체의 항원 결합성 평가
경합 항체로서, WO2010/036460 의 4A5, 5F6 항체, WO2017/061602 의 PPAT-082-03 의 특허 기재 서열에 기초하여, 정제 항체를 조제하였다. CD147 단백질과의 해리 정수 측정은, Biacore T200 (GE Healthcare Bioscience 사 제조) 을 사용하고, Human Antibody Capture Kit (GE Healthcare Bioscience 사 제조) 를 사용하여 고정화한 항 인간 IgG(Fc) 항체에 항체를 리간드로서 포착 (캡처) 하고, 항원을 아날라이트로서 측정하는 캡처법으로 실시하였다. 러닝 버퍼로서 HBS-EP+ (GE Healthcare Bioscience 사 제조), 센서 칩으로서 CM5 (GE Healthcare Bioscience 사 제조) 를 사용하였다. 칩 상에 1 ㎍/㎖ 의 경합 항체를 10 ㎕/분으로 60 초간 첨가한 후, 실시예 2)-5 에서 사용한 항원의 희석 계열 용액 (0.5 ∼ 8 ㎍/㎖) 을 유속 30 ㎕/분으로 120 초간 첨가하고, 계속해서 300 초간의 해리상을 모니터하였다. 재생 용액으로서, 3M 마그네슘클로라이드 (GE Healthcare Bioscience 사 제조) 를 유속 20 ㎕/분으로 30 초간 첨가하였다. 데이터의 해석에는 1 : 1 결합 모델을 사용하여, 결합 속도 정수 ka, 해리 속도 정수 kd 및 해리 정수 (KD ; KD = kd/ka) 를 산출하였다. 인간화 CD147 항체 (#084H1L2hIgG4P 혹은 #110H1L4hIgG4P) 의 해리 정수는, 실시예 7)-1 의 방법으로 산출하였다.
표 8 에는, 산출한 해리 정수, 이펙터 기능, 에피토프 영역에 관한 정보를 나타낸다.
24)-2 기존의 CD147 항체와의 경합 ELISA
실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#084H1L2hIg4P 혹은 #110H1L4hIg4P) 를 사용하고 리콤비넌트 인간 CD147/Fc (Sino Biological 사, Cat. 10186-H02H) 에 대한 결합성을 경합 ELISA 로 평가하였다. 경합 항체로서, 24)-1 에서 조제한 4A5, 5F6항체, PPAT-082-03 항체를 사용하였다. 경합 음성 대조 항체로서, 인간 IgG (잭슨사, Cat. 130093) 를 사용하였다. 경합 양성 대조 항체로서, #084H1L2hIg4P 항체에 대해, #84H1L2hIgG2 항체, #110H1L4hIg4P 에 대해 #110chIgG2 항체를 사용하였다.
96 웰 플레이트 (서모 사이언티픽사, Cat. 43454) 에 PBS 로 희석한 리콤비넌트 인간 CD147/Fc 를 2 ug/㎖, 50 ㎕/well 로 첨가하고, 4 ℃ 에서 하룻밤 보존하였다. 단백 용액을 제거한 후에 1 % BSA 를 포함하는 PBS 를 300 ㎕ 첨가하고, 1 시간 실온에서 가온하였다. 용액을 제거하고, 25 ㎕ 의 기존의 항체액 (0, 0.2, 2, 20 ug/㎖) 을 첨가하고, 2 시간 실온에서 가온하였다. 20 ng/㎖ 농도의 25 ㎕ 의 인간화 CD147 항체 (#084H1L2hIg4P 혹은 #110H1L4hIg4P) 를 각 웰에 첨가하고, 2 시간 실온에서 가온하였다. 0.05 % 트윈20 (BIO RAD 사, Cat. 170-6531) 을 포함하는 PBS 로 2 회 세정한 후, 50 ㎕ 의 1 % BSA 를 포함하는 PBS 로 2000 배 희석한 항인간 IgG4-HRP (압캠사, Cat. ab99823) 를 첨가하고, 1 시간 실온에서 가온하였다. 0.05 % 트윈20 (BIO RAD 사, Cat. 170-6531) 을 포함하는 PBS 로 3 회 세정한 후, 충분히 세정액을 없애고, HRP 기질액 (eBioscience 사, Cat. 00-4203-58) 을 50 ㎕ 첨가하고, 15 ∼ 20 분 실온에서 가온 후, 405 ㎚ 의 흡광도를 플레이트 리더 (퍼킨엘머사, 모델명 EnVision2104) 로 측정하였다.
결과를 표 8 및 도 49(a), (b) 에 나타낸다.
#084H1L2hIG4P 의 리콤비넌트 인간 CD147/Fc 로의 결합은, hIgG 존재하에서는 1 ∼ 10 ug/㎖ 의 기존의 CD147 항체 존재하에서 저해되었다. #110H1L4hIG4P 의 리콤비넌트 인간 CD147/Fc 로의 결합은, hIgG 존재하에서는, 0.1 ∼ 10 ug/㎖ 의 기존의 CD147 항체 존재하에서 저해되지 않고, #110chIgG2 항체의 존재하에서 저해되었다. #084H1L2hIG4P 의 에피토프는, 기존의 항 CD147 항체의 결합에 의해 경합을 받지만, #110H1L4hIG4P 의 에피토프는 기존의 항 CD147 항체의 결합에 의해 영향을 받지 않는 것을 알 수 있었다.
실시예 17)-3 에서 나타낸 H110H1L4h 항체의 에피토프 정보를 표 8 에 기재하였다. 실시예 1)-9 의 CD147 변이체를 사용한 해석으로 에피토프가 추정된 2P10F2 항체와의 결합 경합성 시험 결과 (실시예 2)-8) 로부터, #084H1L2h 의 에피토프 영역을 추정하고, 표 8 에 기재하였다.
24)-3 항 CD147 항체의 ADCC 평가
실시예 1)-15 의 수법에 따라서, 항 CD147 항체의 ADCC 활성을 평가하였다. 실시예 1)-15 의 수법과 상이한 조건으로서, ADCC 표적 세포로서 HepG2 세포 (ATCC, Cat. HB-8065) 를 사용하고, CD147 항체로서 #110H1L4hIgG4P, #084H1L2hIg4P, 4A5, 5F6, PPAT-082-03 을 1 ㎍/㎖ 의 농도로 평가하였다. 측정은 삼중으로 실시하고, 평균값, 표준 편차를 산출하였다. 5 % 이상의 세포의 51Cr 이 검출되었을 경우에, ADCC 양성 (+) 으로 하였다. 5 % 미만의 경우를 ADCC 활성 음성 (―) 으로 하였다. 결과를 표 8 에 나타내었다.
#110H1L4hIgG4P, #084H1L2hIg4P 는, ADCC 활성이 검출되지 않고, ADCC (―) 로 판정하였다. 4A5, 5F6, PPAT-082-03 은, 5 % 이상의 세포의 51Cr 이 검출되고, ADCC 양성 (+) 으로 하였다. #110H1L4hIgG4P, #084H1L2hIg4P 는, ADCC 음성이고, 인간의 생체내에서 CD147 을 발현하고 있는 혈액 세포 등의 정상 세포에 대하여 세포사를 유도할 가능성이, ADCC 활성 양성의 4A5, 5F6, PPAT-082-03 보다 낮은 것이 예측된다.
24)-4 항 CD147 항체의 CDC 평가
실시예 1)-16 의 수법에 따라서, 항인간 CD147 항체에 의한 보체 의존적인 살세포 활성 (CDC 활성) 을 평가하였다. 실시예 1)-16 의 수법과 상이한 조건으로서, 표적 세포로서 인간 간장주 HepG2 세포 (ATCC, Cat. HB-8065) 를 사용하고, 항인간 CD147 항체로서, #110H1L4hIgG4P, #084H1L2hIg4P, 4A5, 5F6 및 PPAT-082-03 을 사용하고, 토끼 보체를 종농도 8 % 가 되도록 첨가하고, 측정하였다. 측정은 삼중으로 실시하고, 평균값과 표준 편차를 산출하였다. 30 % 이상의 항체 의존적인 CDC 활성이 확인된 항체에 대해서는, CDC 활성 양성으로 하고, 표에 CDC (+) 라고 기재하였다. 결과를 표 8 에 나타내었다. 항인간 CD147 항체는, 4A5 만 CDC 활성 양성을 나타내었다. #110H1L4hIgG4P, #084H1L2hIg4P, 5F6, PPAT-082-03 은, CDC 음성이고, 인간의 생체내에서 CD147 을 발현하고 있는 혈액 세포 등의 정상 세포에 대하여 세포사를 유도할 가능성이, CDC 활성 양성의 4A5 보다 낮은 것이 예측된다.
24)-5 항 CD147 항체의 ADCP 평가
실시예 1)-17 의 수법에 따라서, 항 CD147 항체의 ADCP 활성을 측정하였다. 실시예 1)-17 의 수법과 상이한 조건으로서, 표적 세포로서 인간 간장주 HepG2 세포 (ATCC, Cat. HB-8065) 를 사용하고, 항 CD147 항체로서 #110H1L4hIgG4P, #084H1L2hIg4P, 4A5, 5F6, PPAT-082-03 을 1 ㎍/㎖ 의 농도로 첨가하고, 표지한 RAW264.7 세포는 ADCP 표적 세포의 등량 첨가하고, ADCP 활성을 측정하였다. 측정은 삼중으로 실시하고, 평균값, 표준 편차를 산출하고, 인간 IgG 처리군에서 10 % 미만의 ADCP 활성 상승을 약양성 (±), 10 % 이상의 활성 상승을 양성 (+) 으로서 표 8 에 나타내었다.
#110H1L4hIgG4P 의 ADCP 활성은 10 % 미만이고 ADCC 활성 ± 로 판정하였다. #084H1L2hIgG4P, 4A5, 5F6, PPAT-082-03 항체의 ADCP 활성은 10 % 이상이고, ADCP 활성 + 로 판정하였다. CD147-D2 를 인식하는 #110H1L4hIgG4P 는, CD147-D1 을 인식하는 다른 CD147 항체보다 ADCP 활성이 낮았다. ADCP 는, CD147 항체가 CD147 에 결합한 후에, 매크로파지나 단핵구 세포 상에 발현하는 Fcγ 수용체에 의해, 항체의 Fc 부분이 인식되는 것이 필요하지만, #110 항체의 에피토프가 세포 표면에 가까워, 항원인 CD147 에 결합한 항체의 Fc 부분이, 다른 CD147-D1 을 인식하는 항체보다 Fcγ 수용체에 의해 잘 인식되지 않아, ADCP 활성이 낮을 가능성이 있다. #110H1L4hIgG4P, ADCP 활성이 약양성이며, 인간의 생체내에서 CD147 을 발현하고 있는 혈액 세포 등의 정상 세포에 대하여 세포사를 유도할 가능성이, ADCP 활성 양성의 #084H1L2hIg4P, 4A5, 5F6, PPAT-082-03 보다 낮은 것이 예측된다.
(실시예 25)
25)-1 항 CD147 항체에 의한 혈액계 세포 응집
일부의 항 CD147 항체는, 혈구계 세포의 응집을 유도하는 것이 보고되어 있다 (Kasinrerk 등 I㎜unology 1999, 96 (2) p184-192). 혈구계 세포의 응집은, 심각한 혈액 독성을 일으킬 가능성이 있어 (Doll, C., et al., 1994, Curr. Opin. Oncol., 345-350), 치료용 항체로는 바람직하지 않은 성질이다. 항 CD147 항체에 대해, 세포 집적의 활성의 차이를 조사하였다. CD147 항체로서 #110H1L4hIgG4P, #084H1L2hIg4P, 4A5, 5F6, PPAT-082-03 을 평가하였다. 음성의 대조 항체로서 인간 IgG (hIgG, ChromPure Human IgG, Jackson I㎜unoResearch Laboratories 사, Cat. 009-000-003) 를 사용하였다. HEL92.1.7 세포 (ATCC 로부터 구입, Cat. #TIB-180) 를 96 웰 U 보텀 플레이트 (스미토모 베이크라이트사, Cat. MS-9096U) 에 1 웰당, 1600 세포/80 ㎕ 10 % FBS (하이클론사, Cat. SH30084.03) 를 포함하는 RPMI1640 배지 (서모 피셔 사이언티픽사, Cat. 11875-093) 를 첨가하고, 4 시간, 5 % CO2, 습도 95 %, 37 도의 조건으로 배양하였다. 각 웰에 20 ㎕ 의 항 CD147 항체 용액 (150 ug/㎖, 50 ug/㎖) 을 첨가하고, 종농도 30, 10 ug/㎖ 로 하였다. 5 % CO2, 습도 95 %, 37 도의 조건으로 2 일간 배양하고, 현미경 관찰을 실시하였다.
인간 IgG, #110H1L4hIgG4P 의 첨가에 의한 세포의 집적은 관찰되지 않았던 데 반해, #084H1L2hIg4P, 4A5, 5F6, PPAT-082-03 항체의 존재하에서는, 세포가 집적하고, 플레이트의 중앙에 차례차례 겹쳐진 세포덩어리가 관찰되었다. CD147-D2 를 인식하는 #110H1L4hIgG4P 는, CD147-D1 도메인을 인식하는 다른 CD147 항체와는 달리, 혈액 세포의 응집 활성이 없는 것이 나타났다. #084 를 비롯한 세포 응집 활성이 있는 CD147 항체는, 인간에게 투여했을 경우에, 혈액 세포의 응집을 통한 혈전 등의 독성을 일으킬 우려가 있으므로, 혈전의 치료에 사용되는 용량의 헤파린 내지 저분자 헤파린의 피하주 (皮下注), 또는 항혈소판약의 병용 등에 의해 부작용을 회피 혹은 경감하는 것이 바람직하다.
25)-2 사이토카인 방출 증후군의 리스크 평가
치료용 항체의 투여에 의해, OKT3, TGN1412 등 일부의 항체에서는, 면역 세포를 활성화시킴으로써, 혈중의 사이토카인이 상승하고, 심각한 사이토카인 방출 증후군을 일으킨다 (Gaston, R., Kidney International, 1991, 141-148 ; Suntharalingam, G., et al., N. Engl. J. Med. 2006, 1018-1028). 일부의 CD147 항체는, 면역 세포에 작용하고, 인터페론 감마나 인터류킨-4 의 산생을 증가하는 작용이 보고되어 있다 (Hu, J., et al., J. Cell. Mol. Med., 2010, 2132-2143). 이 사이토카인 방출 증후군에 의한 항체 의약의 독성은, 말초혈을 사용한 사이토카인 릴리스 어세이에 의해 예측할 수 있다 (Vessillier, S. et al., J. I㎜unolol. Methods, 2015, 43-52). 동일한 인간 말초혈 사이토카인 릴리스 어세이에 의해 사이토카인 방출 증후군의 리스크를 평가하였다. CD147 항체로서 #110H1L4hIgG4P, #110chIgG4ProFALA, #084H1L2hIg4P, #084H1L2hIg2 를 사용하고, 비교 항체로서, 베바시주맙 (Genentech, Inc.), 트라스투주맙 (Roche Pharma AG), 알렘투주맙 (사노피 주식회사), 항인간 CD3 항체 (BioLegend Cat. No317326) 를 사용하였다. 평가한 모든 CD147 항체에 대해서는, 인간 말초혈 단핵구 (6 도너분을 각각 평가) 에 대해, 세포 증식의 항진은 확인되지 않고, 사이토카인의 방출 (TNFα, INF-γ, IL-2, IL-6, IL-8, IL-10, MIP-1α) 에 대한 영향은, 사이토카인 방출 증후군의 리스크가 낮은 베바시주맙보다 약하였다. 항인간 CD3 항체 (OKT3) 에서는, 세포 증식의 항진, 사이토카인의 방출 (TNFα, INF-γ, IL-2, IL-6, IL-8, IL-10, MIP-1α) 의 항진이 관찰되었다. #110H1L4hIgG4P, #110chIgG4ProFALA, #084H1L2hIg4P, #084H1L2hIg2 는, 사이토카인 방출 증후군의 가능성이 되는 사이토카인 릴리스를 유도하지 않는 것이 나타났다.
25)-3 항 CD147 항체의 원숭이의 안전성 평가
일부의 항마우스 CD147 항체는, 마우스에 투여했을 때에, CD147 의 기능을 저해하고, 비장에서의 적혈구의 집적을 유도함으로써, 말초혈 중의 적혈구량이 저하되고, 빈혈을 일으키는 것이 보고되어 있다 (Coste, I. et al., Blood, 2001, 3984-3988). 본 발명 중에서 취득한 #110H1L4hIgG4P 등의 CD147 항체는, 마우스의 CD147 에 결합을 나타내지 않기 때문에, 마우스에서의 안전성 평가는 적절하지 않다. 그래서, 플로우 사이토미터를 사용한 실험에 의해 인간 및 원숭이 CD147 에 대한 결합성이 확인된 항 CD147 항체로서, #110H1L4hIgG4P 를 게잡이 원숭이에 투여하고, 안전성을 평가하였다. 게잡이 원숭이 (자웅 각 1 예) 에 #110H1L4hIgG4P 를 투여 가능 최대량인 99.2 ㎎/㎏ 으로 단회 정맥내 투여한 결과, 투여 후 15 일까지의 관찰 기간과 관찰 기간 종료 시의 병리 조직학적 검사에서는, 심각한 독성 (체중 및 섭이량의 변화, 병리 조직학적 변화) 은 관찰되지 않았다. #110H1L4hIgG4P 는, 게잡이 원숭이에 대해 독성을 나타내지 않고, 인간의 암 치료에 이용할 수 있는 가능성이 나타났다.
(실시예 26) KLF5 의 CD147 항체 감수성에 대한 영향
암 세포에 있어서, SMAD2/SMAD3/SMAD4 의존적인 세포사의 시그널로서 Lethal EMT 시그널이 알려져 있으며, 이 Lethal EMT 시그널은, SMAD4 음성의 암 세포에서는, 통상적으로 SMAD 시그널에 의해 억제되고 있는 전사 인자 KLF5 단백질의 발현이 증가하고, 치사성의 시그널을 억제하는 것이 보고되어 있다 (David, C. Cell, 2016, 1015-1030). 본 발명의 CD147 항체는, SMAD 시그널을 활성화하고, SMAD4 양성의 세포에 항종양 효과를 나타내기 때문에, SMAD 시그널 의존적인 세포사를 유도하는 것으로 생각하였다. KLF5 가 CD147 항체 의존적인 항종양 효과의 감수성에 관여하는지 조사하였다.
26)-1 KLF5 발현주의 제조
실시예 13 의 방법에 준하여, MIA PaCa-2 세포의 KLF5 안정 발현주를 제조하였다. 인간 KLF5 의 아미노산 서열 및 뉴클레오티드 서열을, 각각, 서열 번호 145 및 146 에 나타낸다. KLF5 유전자로서, (Genscript 사, Cat. OHu21278C) 에 포함되는 서열 (Ref seq. ID : NM_001730.4) 을 짜넣은 레트로바이러스 벡터 pQCXIP 를 제조하고, 레트로바이러스를 제조에 사용하였다. 바이러스 감염에 의해 염색체에 레트로바이러스가 삽입되고, 퓨로마이신 내성, KLF5 양성이 된 MIA PaCa-2 세포를 선택하고, KLF5 양성 MIA PaCa-2 세포, MIA PaCa-2-KLF5 로 하였다. 레트로바이러스 벡터 pQCXIP 를 동일하게 감염시키고, 퓨로마이신 내성이 된 MIA PaCa-2 세포를 MIA PaCa-2-mock 로 하였다.
26)-2 CD147 과 SMAD4 발현의 확인
플로우 사이토미터에 의해, MIA PaCa-2-mock 와 MIA PaCa-2-KLF5 의 CD147 의 발현을 확인하였다. 실시예 13-2 의 방법에 준하여, MIA PaCa-2-KLF5 의 KLF5 의 발현량이 MIA PaCa-2-mock 로부터 증가하고 있는 것을 확인하였다. KLF5 의 검출에는, KLF5 항체 (CST 사, Cat. #51586) 를 사용하였다.
26)-3 KLF5 를 발현한 MIA PaCa-2 종양의 인간화 CD147 항체에 대한 감수성
실시예 7)-2 의 방법에 준하여, MIA PaCa-2-KLF5 와 MIA PaCa-2-mock 의 종양의 인간화 CD147 항체 #110H1L4hIgg4P 에 대한 감수성을 비교하였다. 세포 이식 3 일 후에 실시예 6)-4-2 에서 제조한 인간화 CD147 항체 (#110H1L4hIg4P) 를 1 ㎎/㎏ 으로 담암 마우스에 꼬리 정맥 투여하였다 (n = 6). 7 일 후에 동일하게 항체를 투여하였다. 대조군의 담암 마우스에는, PBS 완충액을 동일하게 꼬리 정맥 투여하였다 (n = 6). 결과를 도 50 에 나타낸다.
인간화 CD147 항체를 투여하여 14 일 후에는, MIA PaCa-2-mock 의 종양은 대조군의 9 % 까지 종양의 평균 체적이 감소하고 있고, CD147 항체에 감수성을 나타내었다. MIA PaCa-2-KLF5 의 종양의 평균 체적은, 대조군의 80 % 로 되어 있고, CD147 항체에 저감수성을 나타내었다. CD147 항체에 의한 SMAD 시그널 의존적인 항종양 효과는, KLF5 의 발현에 의해 억제되는 것을 알 수 있었다.
산업상 이용가능성
본 발명에 의해 CD147 을 활성화하고, 높은 항종양 효과를 나타내는 CD147 특이적 항체가 제공된다. 본 발명에 의해, 이펙터 기능에 의존하지 않고 높은 항종양 효과를 나타내는 항체가 제공된다. 본 발명의 항체는, 간장암 세포에 있어서, 간장암의 표준 치료약 중 하나로서 사용되는 솔라페닙보다 현저하게 강한 약효를 나타낸다. 본 발명의 항체는, 췌장암 세포에 있어서, 췌장암의 표준 치료약 중 하나로서 사용되는 겜시타빈보다 현저하게 강한 약효를 나타낸다. 또, 본 발명의 항체는, 만성 골수성 백혈병 세포에 있어서, 만성 골수성 백혈병의 표준 치료약 중 하나로서 사용되는 이마티닙보다 현저하게 강한 약효를 나타낸다. 본 발명의 항체는, 그 항종양 효과가 이펙터 기능에 의존하지 않고, 그 밖의 안전성 평가 항목에 있어서도 염려가 적고 안전성이 우수한 항 CD147 항체가 제공된다. CD147 은 종양 세포 뿐만 아니라 혈액 세포에도 발현하고 있지만, 본 발명의 항체는, T 세포 및 PBMC 에 대해 작용하지 않고 또한 이펙터 기능에도 의존하지 않기 때문에, 항종양제로서의 개발에 있어서, 안전성의 염려가 적다는 이점을 갖는다. 본 발명에 의해 상기 항체를 포함하는 의약 조성물, 그리고, 그 항체 및/또는 그 의약 조성물을 사용한 종양의 치료 방법이 제공된다.
서열표 프리 텍스트
서열 번호 1 : 인간 CD147 의 베어리언트 1 의 아미노산 서열
서열 번호 2 : 인간 CD147 의 베어리언트 1 의 뉴클레오티드 서열
서열 번호 3 : 인간 CD147 의 베어리언트 2 의 아미노산 서열
서열 번호 4 : 인간 CD147 의 베어리언트 2 의 뉴클레오티드 서열
서열 번호 5 : 인간 SMAD4 의 아미노산 서열
서열 번호 6 : 인간 SMAD4 의 뉴클레오티드 서열
서열 번호 7 : LN22R8 의 경쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 8 : LN22R8 의 경쇄의 가변 영역의 아미노산 서열
서열 번호 9 : LN22R8 의 중쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 10 : LN22R8 의 중쇄의 가변 영역의 아미노산 서열
서열 번호 11 : LN22R8 의 CDRL1 의 아미노산 서열
서열 번호 12 : LN22R8 의 CDRL2 의 아미노산 서열
서열 번호 13 : LN22R8 의 CDRL3 의 아미노산 서열
서열 번호 14 : LN22R8 의 CDRH1 의 아미노산 서열
서열 번호 15 : LN22R8 의 CDRH2 의 아미노산 서열
서열 번호 16 : LN22R8 의 CDRH3 의 아미노산 서열
서열 번호 17 : 2P10F2 의 경쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 18 : 2P10F2 의 경쇄의 가변 영역의 아미노산 서열
서열 번호 19 : 2P10F2 의 중쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 20 : 2P10F2 의 중쇄의 가변 영역의 아미노산 서열
서열 번호 21 : 2P10F2 의 CDRL1 의 아미노산 서열
서열 번호 22 : 2P10F2 의 CDRL2 의 아미노산 서열
서열 번호 23 : 2P10F2 의 CDRL3 의 아미노산 서열
서열 번호 24 : 2P10F2 의 CDRH1 의 아미노산 서열
서열 번호 25 : 2P10F2 의 CDRH2 의 아미노산 서열
서열 번호 26 : 2P10F2 의 CDRH3 의 아미노산 서열
서열 번호 27 : 인간 경쇄 시그널 서열 및 인간 κ 사슬 정상 영역을 코드하는 DNA 서열을 포함하는 DNA 단편
서열 번호 28 : 인간 중쇄 시그널 서열 및 인간 IgG1 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편
서열 번호 29 : 인간 중쇄 시그널 서열 및 인간 IgG2 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편
서열 번호 30 : 인간 키메라 LN22R8 의 경쇄의 뉴클레오티드 서열
서열 번호 31 : 인간 키메라 LN22R8 의 경쇄의 아미노산 서열
서열 번호 32 : 인간 키메라 LN22R8 의 중쇄 IgG1 타입의 뉴클레오티드 서열
서열 번호 33 : 인간 키메라 LN22R8 의 중쇄 IgG1 타입의 아미노산 서열
서열 번호 34 : 인간 키메라 LN22R8 의 중쇄 IgG2 타입의 뉴클레오티드 서열
서열 번호 35 : 인간 키메라 LN22R8 의 중쇄 IgG2 타입의 아미노산 서열
서열 번호 36 : 인간 키메라 LN22R8 의 중쇄 IgG4P 타입의 아미노산 서열을 코드하는 DNA 서열을 포함하는 DNA 단편
서열 번호 37 : 인간 키메라 LN22R8 의 중쇄 IgG4P 타입의 아미노산 서열
서열 번호 38 : 인간 중쇄 시그널 서열 및 인간 IgG1LALA 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편
서열 번호 39 : 인간 중쇄 시그널 서열 및 인간 IgG4P 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편
서열 번호 40 : 인간 키메라 2P10F2 의 경쇄의 뉴클레오티드 서열
서열 번호 41 : 인간 키메라 2P10F2 의 경쇄의 아미노산 서열
서열 번호 42 : 인간 키메라 2P10F2 의 중쇄 IgG1LALA 타입의 뉴클레오티드 서열
서열 번호 43 : 인간 키메라 2P10F2 의 중쇄 IgG1LALA 타입의 아미노산 서열
서열 번호 44 : 인간 키메라 2P10F2 의 중쇄 IgG2 타입의 뉴클레오티드 서열
서열 번호 45 : 인간 키메라 2P10F2 의 중쇄 IgG2 타입의 아미노산 서열
서열 번호 46 : 인간 키메라 2P10F2 의 중쇄 IgG4P 타입의 뉴클레오티드 서열
서열 번호 47 : 인간 키메라 2P10F2 의 중쇄 IgG4P 타입의 아미노산 서열
서열 번호 48 : 래트_CD147_#84 (r#84) 의 경쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 49 : 래트_CD147_#84 (r#84) 의 경쇄의 가변 영역의 아미노산 서열
서열 번호 50 : 래트_CD147_#84 (r#84) 의 중쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 51 : 래트_CD147_#84 (r#84) 의 중쇄의 가변 영역의 아미노산 서열
서열 번호 52 : 래트_CD147_#84 (r#84) 의 CDRL1 의 아미노산 서열
서열 번호 53 : 래트_CD147_#84 (r#84) 의 CDRL2 의 아미노산 서열
서열 번호 54 : 래트_CD147_#84 (r#84) 의 CDRL3 의 아미노산 서열
서열 번호 55 : 래트_CD147_#84 (r#84) 의 CDRH1 의 아미노산 서열
서열 번호 56 : 래트_CD147_#84 (r#84) 의 CDRH2 의 아미노산 서열
서열 번호 57 : 래트_CD147_#84 (r#84) 의 CDRH3 의 아미노산 서열
서열 번호 58 : 래트_CD147_#101 (r#101) 의 경쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 59 : 래트_CD147_#101 (r#101) 의 경쇄의 가변 영역의 아미노산 서열
서열 번호 60 : 래트_CD147_#101 (r#101) 의 중쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 61 : 래트_CD147_#101 (r#101) 의 중쇄의 가변 영역의 아미노산 서열
서열 번호 62 : 래트_CD147_#101 (r#101) 의 CDRL1 의 아미노산 서열
서열 번호 63 : 래트_CD147_#101 (r#101) 의 CDRL2 의 아미노산 서열
서열 번호 64 : 래트_CD147_#101 (r#101) 의 CDRL3 의 아미노산 서열
서열 번호 65 : 래트_CD147_#101 (r#101) 의 CDRH1 의 아미노산 서열
서열 번호 66 : 래트_CD147_#101 (r#101) 의 CDRH2 의 아미노산 서열
서열 번호 67 : 래트_CD147_#101 (r#101) 의 CDRH3 의 아미노산 서열
서열 번호 68 : 래트_CD147_#110 (r#110) 의 경쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 69 : 래트_CD147_#110 (r#110) 의 경쇄의 가변 영역의 아미노산 서열
서열 번호 70 : 래트_CD147_#110 (r#110) 의 중쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 71 : 래트_CD147_#110 (r#110) 의 중쇄의 가변 영역의 아미노산 서열
서열 번호 72 : 래트_CD147_#110 (r#110) 의 CDRL1 의 아미노산 서열
서열 번호 73 : 래트_CD147_#110 (r#110) 의 CDRL2 의 아미노산 서열
서열 번호 74 : 래트_CD147_#110 (r#110) 의 CDRL3 의 아미노산 서열
서열 번호 75 : 래트_CD147_#110 (r#110) 의 CDRH1 의 아미노산 서열
서열 번호 76 : 래트_CD147_#110 (r#110) 의 CDRH2 의 아미노산 서열
서열 번호 77 : 래트_CD147_#110 (r#110) 의 CDRH3 의 아미노산 서열
서열 번호 78 : 래트_CD147_#131 (r#131) 의 경쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 79 : 래트_CD147_#131 (r#131) 의 경쇄의 가변 영역의 아미노산 서열
서열 번호 80 : 래트_CD147_#131 (r#131) 의 중쇄의 가변 영역의 뉴클레오티드 서열
서열 번호 81 : 래트_CD147_#131 (r#131) 의 중쇄의 가변 영역의 아미노산 서열
서열 번호 82 : 래트_CD147_#131 (r#131) 의 CDRL1 의 아미노산 서열
서열 번호 83 : 래트_CD147_#131 (r#131) 의 CDRL2 의 아미노산 서열
서열 번호 84 : 래트_CD147_#131 (r#131) 의 CDRL3 의 아미노산 서열
서열 번호 85 : 래트_CD147_#131 (r#131) 의 CDRH1 의 아미노산 서열
서열 번호 86 : 래트_CD147_#131 (r#131) 의 CDRH2 의 아미노산 서열
서열 번호 87 : 래트_CD147_#131 (r#131) 의 CDRH3 의 아미노산 서열
서열 번호 88 : 인간 중쇄 시그널 서열 및 인간 IgG4PFALA 정상 영역의 아미노산을 코드하는 DNA 서열을 포함하는 DNA 단편
서열 번호 89 : 인간 키메라 래트_CD147_#84 의 경쇄의 뉴클레오티드 서열
서열 번호 90 : 인간 키메라 래트_CD147_#84 의 경쇄의 아미노산 서열
서열 번호 91 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG1 타입의 뉴클레오티드 서열
서열 번호 92 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG1 타입의 아미노산 서열
서열 번호 93 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG2 타입의 뉴클레오티드 서열
서열 번호 94 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG2 타입의 아미노산 서열
서열 번호 95 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG4P 타입의 뉴클레오티드 서열
서열 번호 96 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG4P 타입의 아미노산 서열
서열 번호 97 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG1LALA 타입의 뉴클레오티드 서열
서열 번호 98 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG1LALA 타입의 아미노산 서열
서열 번호 99 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG4PFALA 타입의 뉴클레오티드 서열
서열 번호 100 : 인간 키메라 래트_CD147_#84 의 중쇄 IgG4PFALA 타입의 아미노산 서열
서열 번호 101 : 인간 키메라 래트_CD147_#101 의 경쇄의 뉴클레오티드 서열
서열 번호 102 : 인간 키메라 래트_CD147_#101 의 경쇄의 아미노산 서열
서열 번호 103 : 인간 키메라 래트_CD147_#101 의 중쇄 IgG2 의 뉴클레오티드 서열
서열 번호 104 : 인간 키메라 래트_CD147_#101 의 중쇄 IgG2 의 아미노산 서열
서열 번호 105 : 인간 키메라 래트_CD147_#101 의 중쇄 IgG4P 의 뉴클레오티드 서열
서열 번호 106 : 인간 키메라 래트_CD147_#101 의 중쇄 IgG4P 의 아미노산 서열
서열 번호 107 : 인간 키메라 래트_CD147_#101 의 중쇄 IgG4PFALA 의 뉴클레오티드 서열
서열 번호 108 : 인간 키메라 래트_CD147_#101 의 중쇄 IgG4PFALA 의 아미노산 서열
서열 번호 109 : 인간 키메라 래트_CD147_#110 의 경쇄의 뉴클레오티드 서열
서열 번호 110 : 인간 키메라 래트_CD147_#110 의 경쇄의 아미노산 서열
서열 번호 111 : 인간 키메라 래트_CD147_#110 의 중쇄 IgG2 의 뉴클레오티드 서열
서열 번호 112 : 인간 키메라 래트_CD147_#110 의 중쇄 IgG2 의 아미노산 서열
서열 번호 113 : 인간 키메라 래트_CD147_#110 의 중쇄 IgG4P 의 뉴클레오티드 서열
서열 번호 114 : 인간 키메라 래트_CD147_#110 의 중쇄 IgG4P 의 아미노산 서열
서열 번호 115 : 인간 키메라 래트_CD147_#110 의 중쇄 IgG4PFALA 의 뉴클레오티드 서열
서열 번호 116 : 인간 키메라 래트_CD147_#110 의 중쇄 IgG4PFALA 의 아미노산 서열
서열 번호 117 : 인간 키메라 래트_CD147_#131 의 경쇄의 뉴클레오티드 서열
서열 번호 118 : 인간 키메라 래트_CD147_#131 의 경쇄의 아미노산 서열
서열 번호 119 : 인간 키메라 래트_CD147_#131 의 중쇄 IgG2 의 뉴클레오티드 서열
서열 번호 120 : 인간 키메라 래트_CD147_#131 의 중쇄 IgG2 의 아미노산 서열
서열 번호 121 : 인간 키메라 래트_CD147_#131 의 중쇄 IgG4P 의 뉴클레오티드 서열
서열 번호 122 : 인간 키메라 래트_CD147_#131 의 중쇄 IgG4P 의 아미노산 서열
서열 번호 123 : #84H1hIgG2 의 아미노산 서열
서열 번호 124 : #84H1hIgG2 의 뉴클레오티드 서열
서열 번호 125 : #84H1hIgG4P 의 아미노산 서열
서열 번호 126 : #84H1hIgG4P 의 뉴클레오티드 서열
서열 번호 127 : #84L2h 의 아미노산 서열
서열 번호 128 : #84L2h 의 뉴클레오티드 서열
서열 번호 129 : #101H1hIgG2 의 아미노산 서열
서열 번호 130 : #101H1hIgG2 의 뉴클레오티드 서열
서열 번호 131 : #101H1hIgG4P 의 아미노산 서열
서열 번호 132 : #101H1hIgG4P 의 뉴클레오티드 서열
서열 번호 133 : #101L2h 의 아미노산 서열
서열 번호 134 : #101L2h 의 뉴클레오티드 서열
서열 번호 135 : #110H1hIgG4P 의 아미노산 서열
서열 번호 136 : #110H1hIgG4P 의 뉴클레오티드 서열
서열 번호 137 : #110L4h 의 아미노산 서열
서열 번호 138 : #110L4h 의 뉴클레오티드 서열
서열 번호 139 : #131H2hIgG2 의 아미노산 서열
서열 번호 140 : #131H2hIgG2 의 뉴클레오티드 서열
서열 번호 141 : #131L2h 의 아미노산 서열
서열 번호 142 : #131L2h 의 뉴클레오티드 서열
서열 번호 143 : 인간 CD147v1 의 mu3 영역
서열 번호 144 : 게잡이 원숭이 CD147 의 mu3 영역
서열 번호 145 : 인간 KLF5 의 아미노산 서열
서열 번호 146 : 인간 KLF5 의 뉴클레오티드 서열
서열 번호 147 : #110H13hIgG4P 의 아미노산 서열
서열 번호 148 : #110H13hIgG4P 의 뉴클레오티드 서열
서열 번호 149 : #110L2h 의 아미노산 서열
서열 번호 150 : #110L2h 의 뉴클레오티드 서열
서열 번호 151 : #110L12h 의 아미노산 서열
서열 번호 152 : #110L12h 의 뉴클레오티드 서열