본 발명은 T 세포를 활성화시키는 CD96-결합제를 제공한다. 바람직한 구현예에서, CD96-결합제는 항체 또는 이의 단편이다. 그러나, 대안적인 CD96-결합제는 또한 본 발명의 범위 내에서, CD96, 바람직하게는 인간 CD96에 결합하는, 예를 들어, 앱타머, 올리고뉴클레오타이드, 모방체, 펩타이드 또는 저분자량 화합물 또는 이들의 조합이다. 일 구현예에서, CD96-결합제는 CD96에 대한 가용성 수용체, 예를 들어 CD96에 결합하는 CD155의 가용성 단편(단량체 또는 올리고머 형태)이다.
CD96 결합제는 T 세포의 활성 및/또는 증식을 자극한다. 이는 T 세포(또는 이의 서브셋)가 시험관내 또는 생체내에서 CD96-결합제의 존재하에 증가된 활성 및/또는 증식을 나타내는 것을 의미한다. T 세포 활성 및/또는 증식은 예를 들어, 하기 실시예에 기재된 바와 같이 표준 분석에 의해 측정될 수 있다. 예를 들어, T 세포 활성화는 T 세포 활성 마커, 예를 들어 CD25, CD69, CD137 또는 CD107a의 증가된 발현에 의해 측정될 수 있으며, T 세포 증식은 형광 염료, 예를 들어 CFSE로 미리 표지된 분열 T 세포에 있어서 염료 희석의 분석에 의해 측정될 수 있다. 일부 구현예에서, T 세포는 예를 들어 헬퍼 또는 세포독성 T 세포(CD4+ 또는 CD8+ T 세포)일 수 있다.
T 세포의 증가된 활성 및/또는 증식은 예를 들어 생체내 종양 환경에서와 같이 CD96-결합제 단독의 존재하에, 또는 추가 작용제와 조합하여 CD96-결합제의 존재하에 발생할 수 있다. 예를 들어, 일 구현예에서 CD96-결합제는 T 세포 활성/증식을 공동-자극하며, 즉, CD96-결합제는 추가 작용제(T 세포 공동-자극제)와 조합하여 T 세포 활성 및/또는 증식을 자극한다.
바람직하게는 T 세포 활성 및/또는 증식(예를 들어, T 세포 활성화 마커의 발현 및/또는 형광 염료 희석에 의해 결정됨)은 CD96-결합제의 부재 하에서(예를 들어, T 세포 공동-자극제 단독의 존재하에서) T 세포 활성 및/또는 증식과 비교하여, CD96 결합제의 존재하에서 (예를 들어, CD96-결합제 및 T 세포 공동-자극제의 존재하에) 적어도 5%, 적어도 10%, 적어도 15%, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 75% 또는 적어도 100% 증가된다. 바람직하게는 CD96-결합제는 100 nM 미만, 10 nM 미만, 5 nM 미만 또는 1 nM 미만의 EC50 값으로 T 세포 활성 및/또는 증식을 자극한다.
바람직한 구현예에서, CD96-결합제는 CD96 효능제이다. CD96 효능제는 CD96(바람직하게는 인간 CD96)에 결합하고 CD96을 통한 신호전달을 자극하여 T 세포 활성 및/또는 증식을 촉진하는 결합제이다. 예를 들어, 효능제는 CD96에 결합하고 CD96 단백질에서 형태적 변화를 생성하여, CD96을 통한 세포내 신호전달을 촉진할 수 있다. 일부 구현예에서, CD96-결합제는 CD96의 이량체화 또는 다량체화를 유도할 수 있으며, 예를 들어 결합제의 CD96으로의 결합은 CD96과 동종이량체 또는 다량체로의 회합, 또는 CD226과 같은 추가 세포 표면 단백질과 조합하여 CD96의 이종이량체 또는 다량체로의 회합을 초래할 수 있다. CD96의 이량체화 또는 다량체화는 표준 기술, 예를 들어 하기 실시예에 기재된 바와 같이 형광 공명 에너지 전이(FRET)를 사용하여 검출될 수 있다.
일부 구현예에서, CD96-결합제는 항체, 바람직하게는 단클론성 항체이다. 일 양태에서, 본 발명은 CD96에 특이적으로 결합할 수 있는 마우스, 키메라, 인간화 또는 완전 인간 단클론성 항체, 및 이러한 항체를 생성하는 세포주에 관한 것이다.
일부 다른 양태에서, CD96-결합제는 항체 모방체 또는 항체 모방체 단편이다. 항체 모방체는 다양한 항원에 특이적으로 결합할 수 있는 유기 화합물 또는 스캐폴드 단백질이다. 항체 모방체의 예는 아피바디 분자, 아필린, 아피머, 아피틴, 알파바디, 안티칼린, 아비머, DARPins, 피노머, 모노바디, 압타머, 베타-헤어핀 모방체, 비-면역글로불린 스캐폴드 또는 융합 단백질을 포함한다.
용어 "항체"는 가장 넓은 의미로 사용되며, 항체 단편이 원하는 생물학적 활성을 나타내는 한 구체적으로 단클론성 항체(전장 단클론성 항체 포함), 다클론성 항체, 다중-특이적 항체(예를 들어, 이중-특이적 항체), 및 항체 단편을 포함한다. "항체"(Abs) 및 "면역글로불린"(Igs)은 동일한 구조적 특성을 갖는 당 단백질이다. 항체가 특정 항원에 대한 결합 특이성을 나타내지만, 면역글로불린은 항체 및, 항원 특이성이 없는 다른 항체-유사 분자 둘 다를 포함한다. 후자의 종류의 폴리펩타이드는 예를 들어 림프계에 의해 낮은 수준으로, 골수종에 의해 증가된 수준으로 생성된다.
본 발명에 사용된 바와 같이, 용어 "에피토프"는 항체의 파라토프가 결합하는 항원상의 임의의 항원 결정기를 의미한다. 에피토프 결정기는 일반적으로 아미노산 또는 당 측쇄와 같은 분자의 화학적 활성 표면 그룹으로 구성되며, 일반적으로 특정 3차원 구조적 특성뿐만 아니라 특정 전하 특성을 갖는다.
"천연 항체 및 면역글로불린"은 일반적으로 2개의 동일한 경쇄(L) 및 2개의 동일한 중쇄(H)로 구성된 약 150,000 달톤의 이종사량체 당단백질이다. 각각의 경쇄는 하나의 공유 이황화 결합에 의해 중쇄에 연결되는 반면, 이황화 결합의 수는 상이한 면역글로불린 이소타입의 중쇄 사이에서 다양하다. 각각의 중쇄 및 경쇄는 또한 규칙적으로 이격된 쇄내 이황화 브릿지를 갖는다. 각각의 중쇄는 한 말단에 가변 도메인(VH)을 갖고, 뒤이어 다수의 불변 도메인을 갖는다. 각각의 경쇄는 한 말단에 가변 도메인(VL)을 갖고 다른 말단에 불변 도메인을 가지며; 경쇄의 불변 도메인은 중쇄의 제1 불변 도메인과 정렬되고, 경쇄 가변 도메인은 중쇄의 가변 도메인과 정렬된다. 특정 아미노산 잔기는 경쇄 및 중쇄 가변 도메인 사이의 계면을 형성하는 것으로 여겨진다(Chothia C, Novotny J, Bruccoleri R, and Karplus M., 면역글로불린 분자내 도메인 결합. 가변 도메인의 패킹, J. Mol. Biol. 186:651-63 (1985); Novotny J. and Haber E., 항원 결합의 구조적 불변체: 면역글로불린 VL-VH 및 VL-VL 도메인 이량체의 비교, Proc. Natl. Acad. Sci. U.S.A. 82:4592-96 (1985)).
용어 "가변"은 가변 도메인의 특정 절편이 서열에서 항체마다 광범위하게 상이하고, 이의 특정 항원에 대한 각 특정 항체의 결합 및 특이성에 사용된다는 사실을 지칭한다. 그러나, 가변성은 항체의 가변 도메인의 전체 범위에 걸쳐 균등하게 분포되지 않는다. 대신에, 이는 경쇄 및 중쇄 가변 도메인 둘 다에서 상보성 결정 영역(CDR) 또는 초가변 영역으로 불리는 3개의 절편에 집중되어 있다. 가변 도메인의 보다 고도로 보존된 부분은 프레임워크(FR)로 불린다. 천연 중쇄 및 경쇄 각각의 가변 도메인은, 주로 β-시트 배위를 채택하여 3개의 CDR에 의해 연결되어 있는 4개의 FR 영역을 포함하며, 이는 β-시트 구조를 연결하고 일부 경우에는 그의 일부를 형성하는 루프를 형성한다. 각각의 쇄 내의 CDR은 FR 영역에 의해 근접하게 함께 유지되고, 다른 쇄로부터의 CDR과 함께 항체의 항원-결합 부위의 형성에 기여한다(KABAT 주석에 대해서는 Kabat E.A. 면역학적 관심 단백질의 서열, Fifth Edition, National Institutes of Health, Bethesda, MD (1991)를 참고하거나, 또는 IMGT 주석에 대해서는 http://www.imgt.org를 참조함). 불변 도메인은 항체의 항원에 대한 결합에 직접 관여하지 않지만, 다양한 효과기 기능, 예컨대 항체-의존성 세포독성(ADCC)에서 항체의 참여를 나타낸다.
항체의 파파인 소화는 각각 단일 항원-결합 부위를 갖는 "Fab" 단편으로 불리는 2개의 동일한 항원-결합 단편, 및 잔류 "Fc"단편을 생성하며, 이의 명칭은 쉽게 결정화하는 능력을 반영한다. 펩신 처리는 2개의 항원-결합 부위를 갖고 여전히 항원을 가교시킬 수 있는 F(ab')2 단편을 생성한다.
"Fv"는 완전한 항원 인식 및 결합 부위를 함유하는 최소 항체 단편이다. 2-쇄 Fv 종에서, 이 영역은 타이트한 비-공유 결합에서 하나의 중쇄 및 하나의 경쇄 가변 도메인의 이량체로 구성된다. 단일-쇄 Fv 종(scFv)에서, 하나의 중쇄 및 하나의 경쇄 가변 도메인은 가용성 펩타이드 링커에 의해 공유적으로 연결될 수 있으며, 그에 따라 경쇄 및 중쇄는 2-쇄 Fv 종의 구조와 유사한 "이량체" 구조와 연관될 수 있다. 이러한 구성에서, 각각의 가변 도메인의 3개의 CDR이 상호작용하여 VH-VL 이량체의 표면상에 항원-결합 부위를 정의한다. 총괄적으로, 6개의 CDR은 항체에 항원-결합 특이성을 부여한다. 그러나, 단일 가변 도메인(또는 항원에 특이적인 3개의 CDR만을 포함하는 Fv의 절반)조차도 전체 결합 부위보다 낮은 친화성에서도 항원을 인식하고 결합하는 능력을 갖는다. scFv의 검토에 대해서는 Pluckthun A. “단클론성 항체의 약리학”에서 대장균으로부터의 항체, by Rosenburg and Moore eds., Springer-Verlag, New York, vol. 113, pp. 269-315 (1994)를 참고한다.
Fab 단편은 또한 경쇄의 불변 도메인 및 중쇄의 제1 불변 도메인을 함유한다. Fab' 단편은 항체 힌지(hinge) 영역으로부터의 1개 이상의 시스테인을 포함하는 중쇄 CH1 도메인의 카르복시 말단에 수개의 추가의 잔기를 갖는다는 점에서 Fab 단편과 상이하다. Fab'-SH는 본원에서 불변 도메인의 시스테인 잔기(들)가 유리 티올기를 보유하는 Fab'에 대한 명칭이다. F(ab')2 항체 단편은 본래 이들 사이에 힌지 시스테인을 갖는 Fab' 단편의 쌍으로서 생산되었다. 항체 단편의 다른 화학적 커플링도 공지되어 있다.
면역글로불린에는 5가지 주요 부류의 면역글로불린이 있으며: IgA, IgD, IgE, IgG, 및 IgM, 이들 중 일부는 서브클래스(이소타입), 예를 들어 IgG1, lgG2, lgG3, lgG4, IgA1, 및 lgA2로 추가로 세분될 수 있다. 상이한 부류의 면역글로불린에 상응하는 중쇄 불변 도메인을 각각 α, δ, ε, γ 및 μ라고 한다. 상이한 부류의 면역글로불린의 서브유닛 구조 및 3차원 구성이 잘 알려져 있다.
본원에 사용된 "항체 단편" 및 이의 모든 문법적 변이체는 온전한 항체의 항원 결합 부위 또는 가변 영역을 포함하는 온전한 항체의 일부로서 정의되며, 여기서 상기 부분은 온전한 항체의 Fc 영역의 불변 중쇄 도메인(즉, 항체 이소형에 따라, CH2, CH3 및 CH4)이 없다. 항체 단편의 예는 Fab, Fab', Fab'-SH, F(ab')2 및 Fv 단편; 디아바디; (1) 단일-쇄 Fv(scFv) 분자 (2) 1개의 경쇄 가변 도메인만을 함유하는 단일 쇄 폴리펩타이드, 또는 연관된 중쇄 모이어티 없이 경쇄 가변 도메인의 3개의 CDR을 함유하는 이의 단편 및 (3) 1개의 중쇄 가변 영역만을 함유하는 단일 쇄 폴리펩타이드, 또는 연관된 경쇄 모이어티없이 중쇄 가변 영역의 3개의 CDR을 함유하는 이의 단편; 및 항체 단편으로부터 형성된 다중특이적 또는 다가 구조를 제한없이 포함하는, 연속된 아미노산 잔기의 하나의 연속된 서열로 구성된 1차 구조를 갖는 폴리펩타이드인 임의의 항체 단편(본원에서 "단쇄 항체 단편" 또는 "단쇄 폴리펩타이드"로 지칭됨)을 포함한다. 1개 이상의 중쇄를 포함하는 항체 단편에서, 중쇄(들)는 온전한 항체의 비-Fc 영역에서 발견되는 임의의 불변 도메인 서열(예를 들어, IgG 이소타입에서 CH1)을 함유할 수 있고/있거나 온전한 항체에서 발견되는 임의의 힌지 영역 서열을 함유할 수 있고/있거나 중쇄(들)의 힌지 영역 서열 또는 불변 도메인 서열에 융합되거나 위치된 류신 지퍼 서열을 함유할 수 있다.
본원에 사용된 바와 같이, 용어 "단클론성 항체"(mAb)는 실질적으로 균질한 항체 집단으로부터 수득된 항체를 지칭하며, 즉 집단을 포함하는 개별 항체는 소량으로 존재할 수 있는 가능한 자연 발생 돌연변이를 제외하고는 동일하다. 단클론성 항체는 단일 항원 부위에 대해 매우 특이적이다. 각 mAb는 항원의 단일 결정인자를 대상으로 한다. 이들의 특이성에 더하여, 단클론성 항체는 다른 면역글로불린에 의해 오염되지 않은, 하이브리도마 배양 또는 포유동물 세포주에 의해 합성될 수 있다는 점에서 유리하다. 수식어 "단클론성"은 실질적으로 균질한 항체 집단으로부터 수득된 항체의 특성을 나타내며, 임의의 특정 방법에 의한 항체의 생산을 요구하는 것으로 해석되지 않는다. 예를 들어, 본 발명에 따라 사용되는 단클론성 항체는 불멸화된 B 세포 또는 이의 하이브리도마에서 제조될 수 있거나 재조합 DNA 방법에 의해 제조될 수 있다.
본원의 단클론성 항체는 항체 단편(예를 들어, Fab, F(ab')2 및 Fv)뿐만 아니라 기원 종 또는 면역글로불린 클래스 또는 서브클래스 지정에 관계없이, 원하는 생물학적 활성을 나타내는 한, 항-CD96 항체의 가변(초가변 포함) 도메인을 불변 도메인(예를 들어, "인간화된" 항체)으로, 또는 경쇄를 중쇄로, 또는 한 종으로부터의 쇄를 다른 종으로부터의 쇄로, 또는 융합체를 이종 단백질로 스플라이싱함으로써 생산되는 하이브리드 및 재조합 항체를 포함한다.
본원의 단클론성 항체는 구체적으로 "키메라" 항체(면역글로불린)를 포함하는데, 여기서 중쇄 및/또는 경쇄의 일부는 특정 종으로부터 유래되거나 특정 항체 클래스 또는 서브클래스에 속하는 항체의 상응하는 서열과 동일하거나 상동성인 반면, 사슬(들)의 나머지는 원하는 생물학적 활성을 나타내는 한, 다른 종으로부터 유래되거나 다른 항체 클래스 또는 서브클래스에 속하는 항체 뿐만 아니라 상기 항체들의 단편에서 상응하는 서열과 동일하거나 상동성이다.
본 발명의 CD96-결합제(예를 들어, 항체)는 단리된 형태로 사용될 수 있다. "단리된" 항체는 자연 환경의 성분으로부터 동정 및 분리 및/또는 회수된 것이다. 자연 환경의 오염 성분은 항체의 진단 또는 치료 용도를 방해하는 물질이며, 효소, 호르몬 및 기타 단백질성 또는 비-단백질성 용질을 포함할 수 있다. 일부 구현예에서, 항체는 (1) 로리(Lowry) 법에 의해 결정된 바와 같이 항체의 75 중량% 초과, 및 가장 바람직하게는 80 중량%, 90 중량% 또는 99 중량% 초과, 또는 (2) 쿠마시 블루 또는 바람직하게는 은 염색을 사용하여 환원 또는 비 환원 조건 하에서 SDS-PAGE에 의해 균질성으로 정제될 것이다. 단리된 항체는 항체의 자연 환경의 적어도 하나의 성분이 존재하지 않을 것이기 때문에 재조합 세포 내의 계내 항체를 포함한다. 그러나, 일반적으로 단리된 항체는 적어도 하나의 정제 단계에 의해 제조될 것이다.
일 양태에서, 본 발명은 0.01 내지 50 nM, 예를 들어 0.18 내지 42.3 nM 범위의 해리 상수(KD)로 인간 CD96에 결합하는 마우스 및 키메라 또는 인간화된 마우스-인간 단클론성 항체를 제공한다. 일부 항체는 인간 CD4+ 및 CD8+ T 세포의 활성화 및 증식을 공동-자극한다. 이러한 효능제 항체는 암 또는 감염성 질환 환자에서 면역 반응을 자극하는데 사용된다. 개시된 항체 중 일부는 CD96-CD155 상호작용을 방해하거나 억제하며, T 세포 기능의 조절에 기능적 역할을 할 수 있다.
hCD96을 암호화하는 DNA로 프라이밍하고 인간 CD96을 표시하는 세포주로 증폭시 마우스의 면역화에 의해 항-인간 CD96(hCD96) 항체가 생성되었다. 이어서 비장 및 림프절로부터 단리된 B 세포를 ISAAC(ImmunoSpot Array Assay on Chip)로 스크리닝하여 hCD96에 결합하는 큰 항체 패널을 단리하였다. 생성된 항체의 광범위한 다양성으로 인해, 이전에 개시되지 않은 신규한 생물학적 활성을 갖는 항-hCD96 후보가 동정되었다.
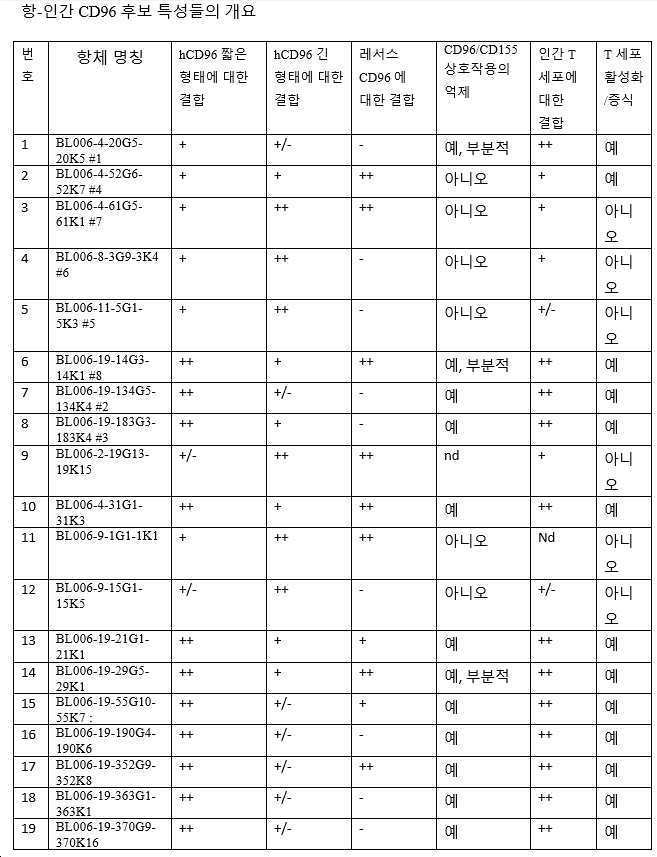
19개의 항-인간 CD96 항체가 시험관내에서 생성되고 특성화되었다. 9개의 후보 물질은 5.92 내지 19.42 nM 범위의 IC50 및 98 내지 100%의 최대 억제 용량을 가지고 CD96 및 CD155 사이의 상호작용을 강하게 억제하였다. 후보 BL006-4-20을 포함하는 3개의 mAb는 50%보다 낮은 최대 억제 용량(부분 억제제)과의 CD96/CD155 상호작용을 억제하였다.
인간 CD4+ 및 CD8+ T 세포 활성화 및 증식에 대한 강력한 공동-자극 활성을 갖는 항-CD96 후보가 동정되었다. 13개의 후보(표 4 참조)는 최적 이하의 농도의 항-CD3 항체 OKT3으로 활성화된 말초 혈액 T 세포의 공동 자극을 나타냈다. 중요하게도, 다른 그룹에 의해 사용된 벤치마크 항-hCD96 NK92.39 항체(Fuchs et al., 2004; WO2015024060)는 동일한 시험관내 분석에서 유의한 공동-자극 활성을 나타내지 않았다.
본 발명의 일부 항-CD96 항체는 인간 및 레서스 CD96에 결합하는 반면, 다른 항체들은 인간으로 제한된다. 교차-반응성은 전임상 개발 측면에서 이점, 예를 들어 동일한 모델에서 효능 및 안전성 테스트를 수행하는 능력을 나타낸다.
따라서, 인간 CD96-발현 T 세포에 대한 특이적 활성화 활성을 갖는 신규한 항체가 본 발명에서 동정되었다. 따라서, 이들 항체는 암을 치료하기 위해 T 세포 반응을 단독으로 또는 체크포인트 억제제 또는 다른 치료제와 함께 자극하는 치료제로서 사용될 수 있다.
본 발명의 항체는 하기 특성들 중 적어도 하나를 가질 수 있다: T 세포 활성 및/또는 증식의 자극(공동-자극 포함), (예를 들어, 인간 또는 레서스) CD96에 결합, (예를 들어, 인간) CD96의 효능, T 세포상의 (예를 들어, 인간) CD96의 이량체화 또는 다량체화 유도, CD96에 대한 (예를 들어, 인간) CD155의 결합 억제, 및 예를 들어, 이종이식 마우스 모델에서 종양 성장의 억제.
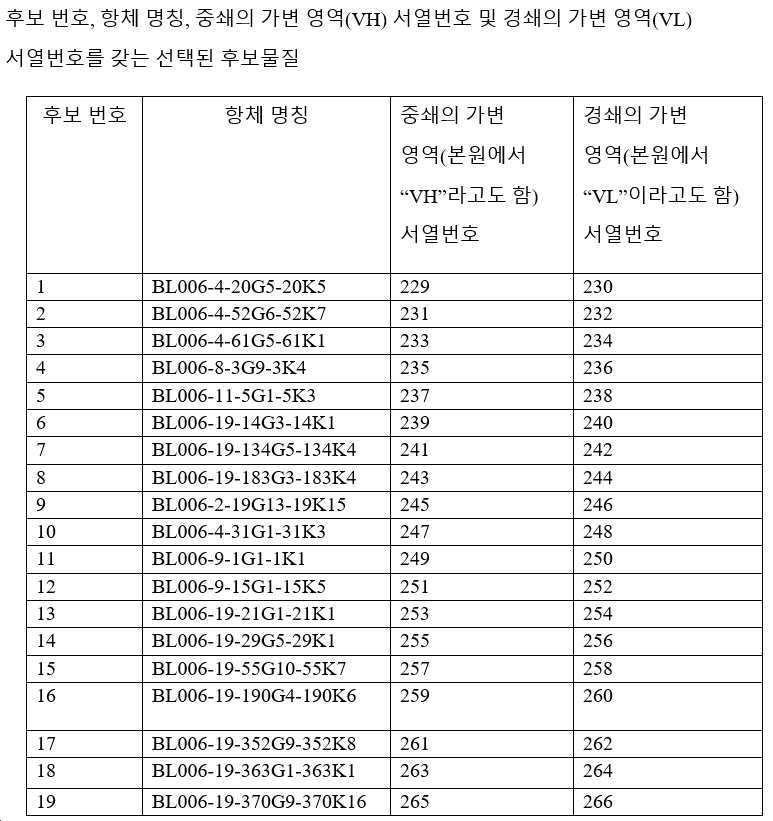
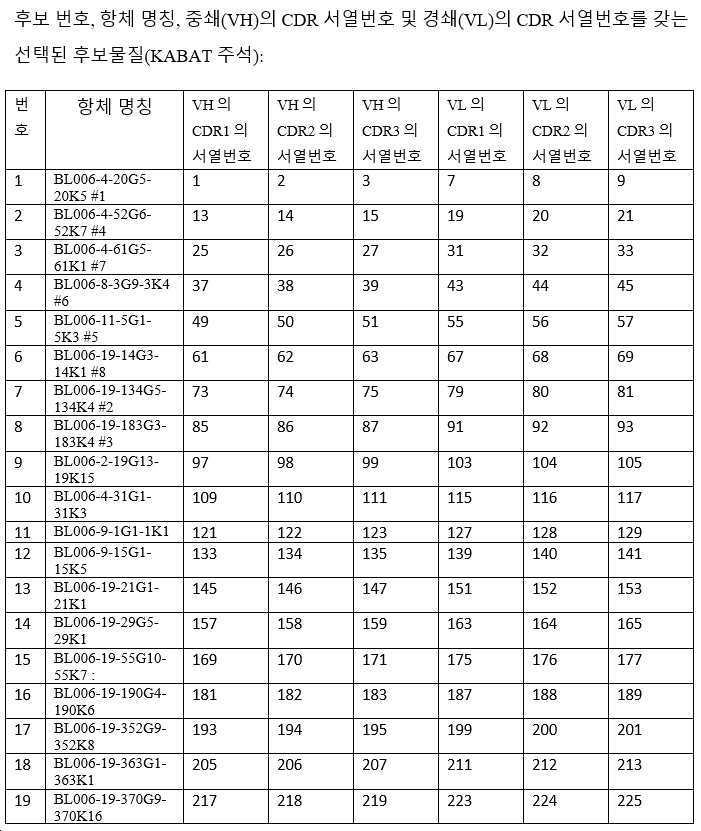
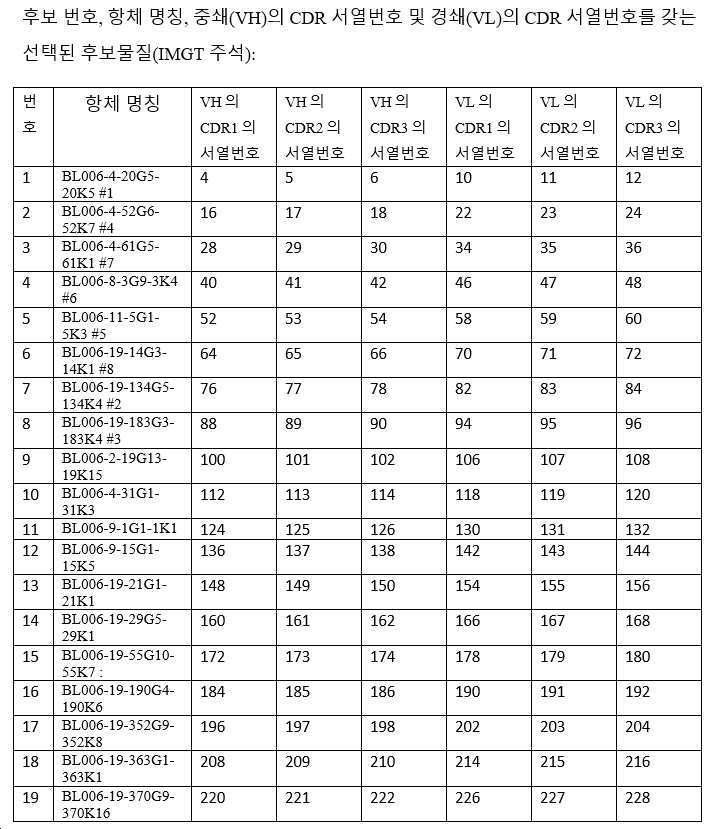
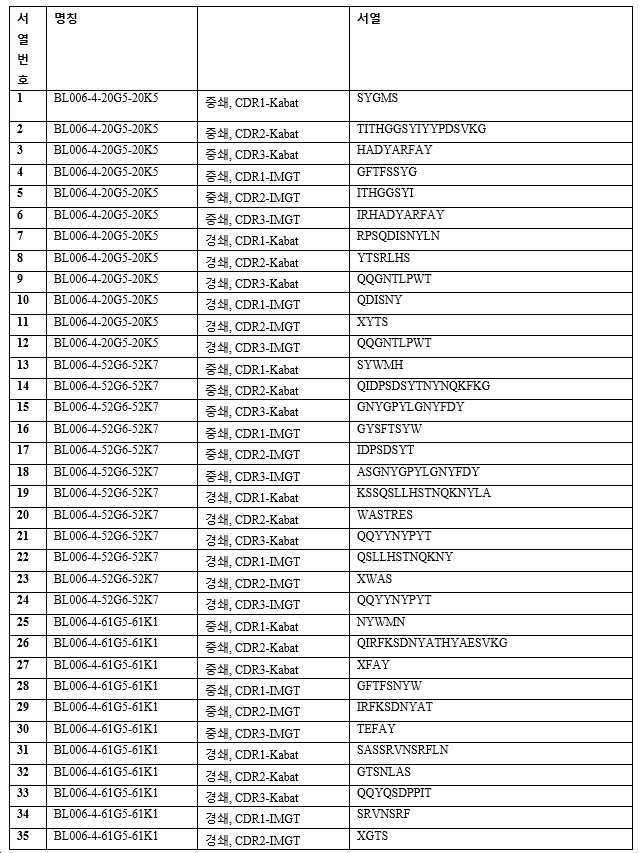
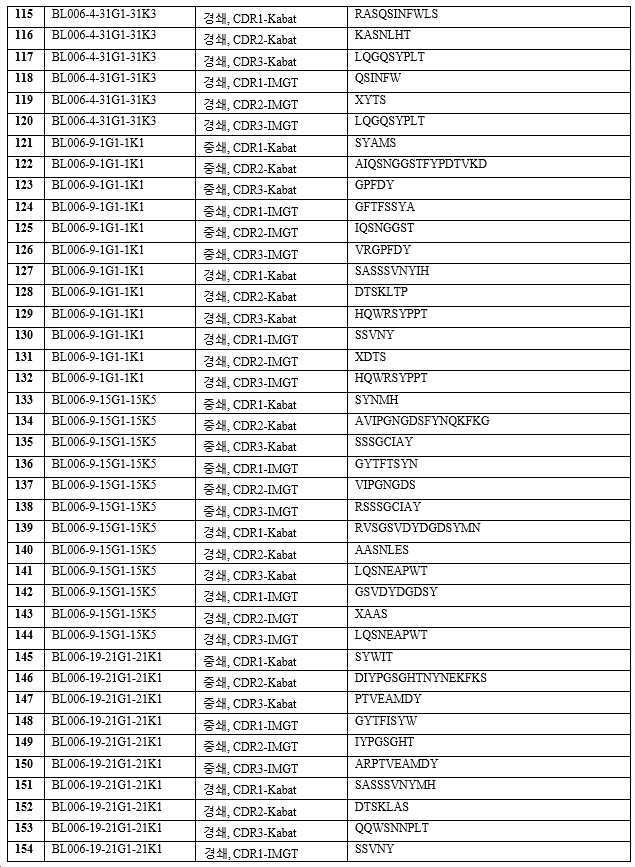
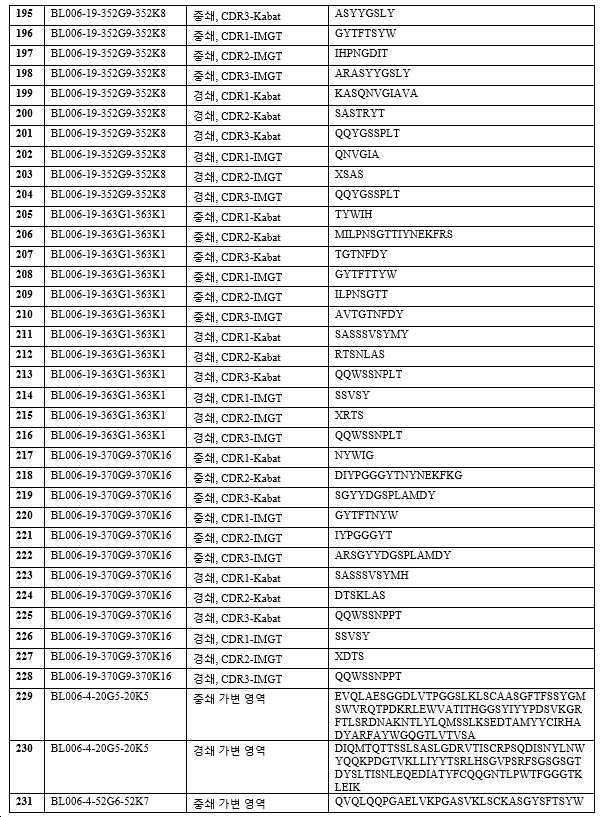
하기 단락에서, 후보 항체는 표 1 내지 3에 주어진 명칭으로 지칭되며, 이는 경우에 따라 편의를 위해 약칭될 수 있으며, 예를 들어 BL006-4-31G1-31K3은 BL006-4-31로 지칭될 수 있고, BL006-19-183G3-183K4는 BL006-19-183 등으로 지칭될 수 있다. 일부 구현예에서, 항체는 표 1 내지 3에 기재된 후보 중 하나 또는 그의 변이체, 예를 들어 상기 항체 중 하나로부터의 하나 이상의 CDR 서열 또는 가변 영역을 포함하는 항체이다.
바람직한 구현예에서, 항체는 인간 CD96에 결합하기 위한 높은 친화도를 갖는다. 예를 들어, 일 구현예에서 항체는 예를 들어, 10 nM 미만의 EC50 값으로 인간 CD96(예를 들어, 변이체 2, 짧은 이소형, 서열번호: 268)에 결합한다. EC50 값은, 예를 들어 인간 CD96 변이체 2로 형질감염된 CHO 세포에 대한 유세포 측정법에 의해 결정될 수 있다. 11개의 후보가 CD96에 강력하게 결합하는 것으로 밝혀졌다(BL006-4-31, BL006-19-183, BL006-19-190, BL006-19-134, BL006-19-21, BL006-19-55, BL006-19-352, BL006-19-363, BL006-19-370, BL006-19-14, BL006-19-29). 따라서, 일부 구현예에서, 항체는 상기 후보 중 하나 또는 이의 변이체, 예를 들어 하나 이상의 CDR 서열을 포함하는 항체 또는 상기 항체 중 하나 유래의 가변 영역이다. 바람직한 항체는 서열번호: 109~120, 247, 248(BL006-4-31); 85~96, 243, 244(BL006-19-183); 181~192, 259, 260(BL006-19-190); 73~84, 241, 242(BL006-19-134); 145~156, 253, 254(BL006-19-21); 169~180, 257, 258(BL006-19-55); 193~204, 261, 262(BL006-19-352); 205~216, 263, 264(BL006-19-363); 217~228, 265, 266(BL006-19-370); 61~72, 239, 240(BL006-19-14); 및 157~168, 255, 256(BL006-19-29)로부터 선택된 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 항체는 긴 형태의 인간 CD96, 즉 변이체 1, 서열번호: 267에 대한 높은 친화도를 가지며, 예를 들어, 항체는 인간 CD96 변이체 1로 형질감염된 CHO 세포에 대한 유세포 측정법에 의해 결정된 10 nM 미만의 EC50 값으로 결합한다. 8개의 후보들은 또한, 인간 CD96(CD96v1)의 긴 형태에 강력하게 결합하는 것으로 밝혀졌다(BL006-19-183, BL006-19-14, BL006-19-29, BL006-2-19, BL006-11-5, BL006-4-61, BL006-9-1, BL006-8-3, BL006-9-15). 따라서, 일부 구현예에서, 항체는 상기 후보 중 하나 또는 이의 변이체, 예를 들어 상기 항체 중 하나로부터의 하나 이상의 CDR 서열 또는 가변 영역을 포함하는 항체이다. 바람직한 항체는 서열번호: 85~96, 243, 244(BL006-19-183); 61~72, 239, 240(BL006-19-14); 157~168, 255, 256(BL006-19-29); 97~108, 245, 246(BL006-2-19); 49~60, 237, 238(BL006-11-5); 25~36, 233, 234(BL006-4-61); 121~132, 249, 250(BL006-9-1); 37~48, 235, 236(BL006-8-3); 133~144, 251, 252(BL006-9-15)로부터 선택된 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 항체는 인간 T 세포 및/또는 NK 세포에 강하게 결합한다. 따라서, 일부 구현예에서, 항체는 상기 후보 중 하나 또는 이의 변이체, 예를 들어 상기 항체 중 하나로부터의 하나 이상의 CDR 서열 또는 가변 영역을 포함하는 항체이다. 바람직한 항체는 서열번호: 1~12, 229, 230(BL006-4-20G5), 13~24, 231, 232(BL006-4-52G6), 109~120, 247, 248(BL006-4-31); 85~96, 243, 244(BL006-19-183); 181~192, 259, 260(BL006-19-190); 73~84, 241, 242(BL006-19-134); 145~156, 253, 254(BL006-19-21); 169~180, 257, 258(BL006-19-55); 193~204, 261, 262(BL006-19-352); 205~216, 263, 264(BL006-19-363); 217~228, 265, 266(BL006-19-370); 61~72, 239, 240(BL006-19-14); 및 157~168, 255, 256(BL006-19-29)로부터 선택되는 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 항체는 레서스 CD96에 대해 높은 친화도를 가지며, 예를 들어 항체는 레서스 CD96(서열번호: 272)으로 형질감염된 CHO 세포에서 유세포 측정법에 의해 측정된 10 nM 미만의 EC50 값으로 결합한다. 8개의 항체가 레서스 CD96을 강하게 인식한다(BL006-4-31, BL006-19-352, BL006-19-14, BL006-19-29, BL006-4-52, BL006-2-19, BL006-4-61, BL006-9-1). 따라서, 일부 구현예에서, 항체는 상기 후보 중 하나 또는 그의 변이체, 예를 들어 상기 항체 중 하나로부터의 가변 영역 또는 하나 이상의 CDR 서열을 포함하는 항체이다. 바람직한 항체는 서열번호: 109~120, 247, 248(BL006-4-31); 193~204, 261, 262(BL006-19-352); 61~72, 239, 240(BL006-19-14); 157~168, 255, 256(BL006-19-29); 13~24, 231, 232(BL006-4-52); 97~108, 245, 246(BL006-2-19); 25~36, 233, 234(BL006-4-61); 121~132, 249, 250(BL006-9-1)로부터 선택되는 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
따라서, 바람직한 구현예에서, 항체는 하나 이상의 형태의 CD96, 예를 들어 인간 CD96 변이체 1 및 2 또는 인간 CD96 및 레서스 CD96에 결합한다. 인간 및 레서스 CD96과 교차-반응하는 바람직한 항체는 서열번호: 61~72, 239, 240(BL006-19-14); 109~120, 247, 248(BL006-4-31); 157~168, 255, 256(BL006-19-29); 및 193~204, 261, 262(BL006-19-352)로부터 선택된 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다
또 다른 구현예에서, 항체는 CD155와 CD96의 결합을 억제한다. 예를 들어, 항체는 유세포 측정법에 의해 결정된 바와 같이 20 nM 미만의 IC50 값으로 CHO 세포상에서 발현된 hCD96v2에 대한 hCD155의 결합을 억제할 수 있다. 9개의 후보는 5.9 내지 19.4 nM 범위의 IC50 값으로 hCD155와 hCD96v2의 결합을 강하게 억제하였다(후보 BL006-4-31, BL006-19-21, BL006-19-183, BL006-19-190, BL006-19-134, BL006-19-55, BL006-19-352, BL006-19-363, BL006-19-370). 이들 9개의 항-CD96 후보는 이 분석에서 클론 628211 및 NK92.39 항체와 유사하였다. 따라서, 일부 구현예에서, 항체는 상기 후보 중 하나 또는 그의 변이체, 예를 들어 상기 항체 중 하나로부터의 가변 영역 또는 하나 이상의 CDR 서열을 포함하는 항체이다. 바람직한 항체는 서열번호: 109~120, 247, 248(BL006-4-31); 145~156, 253, 254(BL006-19-21); 85~96, 243, 244(BL006-19-183); 181~192, 259, 260(BL006-19-190); 73~84, 241, 242(BL006-19-134); 169~180, 257, 258(BL006-19-55); 193~204, 261, 262(BL006-19-352); 205~216, 263, 264(BL006-19-363); 217~228, 265, 266(BL006-19-370)로부터 선택되는 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
대안적인 구현예에서, 항체는 CD155와 CD96의 결합을 부분적으로 억제한다. "부분적으로 억제" 또는 "부분 억제제"는 항체가 항체의 과잉 농도에서 이용가능한 CD155/CD96 결합 부위의 100% 미만을 억제하는 것, 예를 들어, 항체가 CD155/CD96 결합의 95% 미만, 90% 미만, 80% 미만, 70% 미만, 60% 미만 또는 50% 미만을 최대로 억제하는 것을 의미한다. 예를 들어, 3개의 후보(BL006-4-20, BL006-19-14 및 BL006-19-29)는 CD96에 대한 CD155 결합의 부분적 억제제인 것으로 나타났으며, 최대 억제 능력은 50% 이하지만, 2.67 내지 5.24 nM 사이의 상대 IC50 값을 갖는다. 따라서, 일부 구현예에서, 항체는 서열번호: 1~12, 229, 230(BL006-4-20); 61~72, 239, 240(BL006-19-14); 및 157~168, 255, 256(BL006-19-29)으로부터 선택된 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
대안적인 구현예에서, 항체는 CD155와 CD96의 결합을 억제하지 않는다. 예를 들어, 6개의 후보(BL006-4-52, BL006-4-61, BL006-11-5, BL006-8-3, BL006-9-1 및 BL006-9-15)는 CD96/CD155 상호작용을 억제하지 않았다. 바람직한 항체는 서열번호: 13~24, 231, 232(BL006-4-52); 25~36, 233, 234(BL006-4-61); 49~60, 237, 238(BL006-11-5); 37~48, 235, 236(BL006-8-3); 121~132, 249, 250(BL006-9-1) 및 133~144, 251, 252(BL006-9-15)로부터 선택된 하나 이상의 CDR 서열(예를 들어 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 항체는 CD96 단백질의 D1 도메인에 결합한다. 예를 들어, 1개의 후보(BL006-19-183)는 CD96의 D1 도메인에 결합한다. 바람직한 항체는 서열번호: 85~96, 243, 244(BL006-19-183)로부터 선택된 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 항체는 CD96 단백질의 D1 및 D2 도메인에 결합한다. 예를 들어, 9개의 후보(BL006-4-31, BL006-4-20, BL006-19-190, BL006-19-21, BL006-19-55, BL006-19-370, BL006-19-363, BL006-19-352 및 BL006-19-134)는 CD96의 D1 및 D2 도메인에 결합한다. 바람직한 항체는 서열번호: 109~120, 247, 248(BL006-4-31); 1~12, 229, 230(BL006-4-20); 181~192, 259, 260(BL006-19-190); 145~156, 253, 254(BL006-19-21); 169~180, 257, 258(BL006-19-55); 217~228, 265, 266(BL006-19-370); 205~216, 263, 264(BL006-19-363); 193~204, 261, 262(BL006-19-352) 및 73~84, 241, 242(BL006-19-134)로부터 선택된 하나 이상의 CDR 서열(예를 들어 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 항체는 CD96 단백질의 D2 및 D3 도메인에 결합한다. 예를 들어, 2개의 후보(BL006-19-14 및 BL006-19-29)은 CD96의 D2 및 D3 도메인에 결합한다. 바람직한 항체는 서열번호: 61~72, 239, 240(BL006-19-14) 및 157~168, 255, 256(BL006-19-29)으로부터 선택된 하나 이상의 CDR 서열(예를 들어, 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 항체는 CD96 단백질의 D3 도메인에 결합한다. 예를 들어, 2개의 후보(BL006-4-52 및 BL006-2-19)은 CD96의 D3 도메인에 결합한다. 바람직한 항체는 서열번호: 13~24, 231, 232(BL006-4-52) 및 97~108, 245, 246(BL006-2-19)로부터 선택된 하나 이상의 CDR 서열(예를 들어 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 항체는 CD96 단백질의 D4 도메인에 결합한다. 예를 들어, 5개의 후보(BL006-4-61, BL006-8-3, BL006-9-1, BL006-9-15 및 BL006-11-5)은 CD96의 D4 도메인에 결합하지만 T 세포 활성화 또는 증식을 나타내지 않았다. 바람직한 항체는 서열번호: 25~36, 233, 234(BL006-4-61); 37~48, 235, 236(BL006-8-3); 121~132, 249, 250(BL006-9-1); 133~144, 251, 252(BL006-9-15) 및 49~60, 237, 238(BL006-11-5)로부터 선택된 하나 이상의 CDR 서열(예를 들어 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
또 다른 구현예에서, 본 발명의 구현예 중 어느 하나에 따른 항체는 T 세포의 증식을 공동-활성화할 수 있고 D1 도메인에, D1 및 D2 도메인에, 또는 D2 및 D3 도메인에, 또는 D3 도메인에 존재하는 에피토프에 결합할 수 있다. 예를 들어, 이러한 항체는 최적 이하의 농도의 T 세포 자극제 또는 증식제, 예를 들어 항-CD3 항체로 CD4+ 및/또는 CD8+ T 세포를 공-활성화시킬 수 있다. 활성화된 세포의 백분율은 예를 들어, CD25와 같은 T 세포 마커의 증가된 발현, CFSE 염색의 희석 및/또는 분열 세포의 수와 상관되는 3[H]-티미딘의 혼입을 측정함으로써 측정될 수 있다. 바람직하게는 항체의 공동-자극 활성은 저농도의 항-CD3 항체(예를 들어, 0.1 ng/ml 이하)에서 관찰된다. BL006-4-31, BL006-4-20, BL006-19-14, BL006-19-190, BL006-19-21, BL006-19-55, BL006-19-370, BL006-19-363, BL006-19-352, BL006-19-29, BL006-19-183, BL006-19-134, 및 BL006-4-52는 T 세포 자극 활성을 나타내는 것으로 밝혀졌다. 따라서, 일부 구현예에서, 항체는 상기 후보 중 하나의 키메라, 인간화 또는 조작된 버전이다. 또 다른 구현예에서, 항체는 가용성 형태로 사용된다. 따라서, 일부 구현예에서, 항체는 상기 후보 중 하나 또는 그의 변이체, 예를 들어 상기 항체 중 하나로부터의 하나 이상의 CDR 서열 또는 가변 영역을 포함하는 항체이다. 바람직한 항체는 서열번호: 109~120, 247, 248(BL006-4-31); 1~12, 229, 230(BL006-4-20); 61~72, 239, 240(BL006-19-14); 181~192, 259, 260(BL006-19-190); 145~156, 253, 254(BL006-19-21); 169~180, 257, 258(BL006-19-55); 217~228, 265, 266(BL006-19-370); 205~216, 263, 264(BL006-19-363); 193~204, 261, 262(BL006-19-352); 157~168, 255, 256(BL006-19-29); 85~96, 243, 244(BL006-19-183); 73~84, 241, 242(BL006-19-134); 및 13~24, 231, 232(BL006-4-52)로부터 선택된 하나 이상의 CDR 서열(예를 들어 3개의 중쇄 및 3개의 경쇄 CDR 서열) 또는 중쇄 및/또는 경쇄 가변 도메인을 포함한다.
바람직한 후보는 이로부터 유래된 인간화되고 추가로 조작된 항체를 포함하여, 상기 정의된 항체들 중 하나 이상이다. 예시적인 항체(본 명세서에서 일반적으로 후보 NO 또는 항체 이름으로 지칭됨)의 CDR(KABAT-(표 2) 및 IMGT-(표 3) 주석) 및 가변 영역(표 1)이 제공된다(IMGT 주석이 바람직함). 관심 항체는 상기 제공된 조합들 뿐만 아니라, 예를 들어 F(ab)' 항체를 생성하기 위한, 적절한 불변 영역 또는 불변 영역의 단편에 대한 가변 영역의 융합을 포함한다. 관심있는 가변 영역은 제공된 항-CD96 항체의 적어도 하나의 CDR 서열을 포함하고, 여기서 CDR은 3, 4, 5, 6, 7, 8, 9, 10, 11, 12개 이상의 아미노산일 수 있다. 대안적으로, 관심 항체는 제공된 항체에 제시된 가변 영역, 또는 본원에 제시된 가변 영역 서열 쌍을 포함한다. 이들 항체는 예를 들어 임의의 이소타입의 인간 면역글로불린 불변 영역, 예를 들어 lgG1, lgG2, lgG3, lgG4, IgA를 갖는 전장 항체일 수 있다. 바람직하게는 항체는 IgG1이며, 즉 항체는 인간 IgG1 불변 영역, 예컨대 본 발명의 항체의 작용제 및 기타 효과기 기능을 향상시키는 Fc 부분을 포함한다(Zhang D., Goldberg M.V. and Chiu M.L., 항-OX40 항체의 작용제 및 효과기 기능을 향상시키기 위한 Fc 공학적 접근법, The journal of biological chemistry, 291(53):27134-27146 (2016); Saxena A. and Wu, D., 치료 Fc 조작 발전 - IgG-관련 효과기 기능 및 혈청 반감기의 조절, Frontiers in Immunology, 12(7):580 (2016)). 본 발명의 다른 바람직한 항체는 항체의 다량체화를 부여하는 항체이다.
CD96-결합제(예를 들어, 항체)는 추가의 활성제, 예를 들어 접합체 형태로 커플링될 수 있다. 접합체는 예를 들어, 하나 이상의 활성제(예를 들어, 항체 단편(들))를 하나 이상의 중합체 분자(들)에 공유 부착함으로써 형성된 이종 분자일 수 있으며, 여기서 이종 분자는 수용성, 즉 혈액과 같은 생리학적 유체에 가용성이고, 여기서 이종 분자에는 임의의 구조화된 응집체가 없다. 관심 접합체는 PEG이다. 상기 정의와 관련하여, 용어 "구조화된 응집체"는 (1) 이종 분자가 미셀 또는 다른 에멀젼 구조에 있지 않도록, 및 지질 이중층, 소포 또는 리포좀에 고정되지 않도록, 타원체 또는 타원체 쉘 구조를 갖는 수용액 중의 분자의 임의의 응집체; 및 (2) 수성 상과 접촉시 이종 분자를 용액으로 방출하지 않는 크로마토그래피 비드 매트릭스와 같은 고체 또는 불용화 형태의 분자의 임의의 응집체를 지칭한다. 따라서, 본원에 정의된 바와 같이, 용어 "접합체"는 고체의 수화시 이종 분자를 수용액으로 방출할 수 있는 침전물, 침강물, 생-침식성 매트릭스 또는 다른 고체 중의 상기 언급된 이종 분자를 포함한다.
CD96-결합제(예를 들어, 항체)는 에피토프 태그될 수 있다. 본원에서 사용될 때 용어 "에피토프 태그된"은 "에피토프 태그"에 융합된 항-CD96 항체를 지칭한다. 에피토프 태그 폴리펩타이드는 항체가 제조될 수 있는 에피토프를 제공하기에 충분한 잔기를 갖지만, CD96 항체의 활성을 방해하지 않을 정도로 충분히 짧다. 에피토프 태그는 바람직하게는 에피토프에 특이적인 항체가 다른 에피토프와 실질적으로 교차-반응하지 않도록 충분히 고유하다. 적합한 태그 폴리펩타이드는 일반적으로 적어도 6개의 아미노산 잔기 및 일반적으로 약 8~50개의 아미노산 잔기(바람직하게는 약 9~30개의 잔기)를 갖는다. 예는 c-myc 태그 및 이에 대한 8F9, 3C7, 6E10, G4, B7 및 9E10 항체를 포함한다(Evan G.I., Lewis G.K., Ramsay G., et al. "인간 c-myc 원-종양유전자 생성물에 특이적인 단클론성 항체의 단리", Mol. Cell. Biol. 5(12):3610-3616 (1985)); 및 헤르페스 심플렉스 바이러스 당 단백질 D(gD) 태그 및 그의 항체(Paborsky L.R., Fendly B.M., Fisher K.L., et al. "항원 생산을 위한 조직 인자의 포유류 세포 일시적 발현", Protein Engineering 3(6):547-553 (1990)).
추가의 표지가 CD96-결합제에 부착될 수 있다. 본원에서 사용될 때 단어 "표지"는 작용제(예를 들어, 항체)에 직접 또는 간접적으로 접합된 검출가능한 화합물 또는 조성물을 지칭한다. 표지 자체는 그 자체로(예를 들어, 방사성동위원소 표지 또는 형광 표지) 검출될 수 있거나, 효소 표지의 경우 검출가능한 기질 화합물 또는 조성물의 화학적 변경을 촉매할 수 있다.
일부 구현예에서, CD96-결합제는 고체상에 부착될 수 있다. "고체상"은 본 발명의 작용제(예를 들어, 항체)가 부착될 수 있는 비-수성 매트릭스를 의미한다. 본원에 포함된 고체상의 예는 유리(예를 들어, 제어된 세공 유리), 다당류(예를 들어, 아가로스), 폴리아크릴아미드, 폴리스티렌, 폴리비닐 알코올 및 실리콘으로 부분적으로 또는 전체적으로 형성된 것들을 포함한다. 특정 구현예에서, 문맥에 따라, 고체상은 분석 플레이트의 웰을 포함할 수 있으며; 다른 경우에는 정제 컬럼(예를 들어, 친화성 크로마토그래피 컬럼)이다. 이 용어는 또한 불연속 입자의 불연속 고체상, 예컨대 미국 특허 제4,275,149호에 기술된 것들을 포함한다.
본원에 개시된 CD96-결합제(예를 들어, 항체)는 질환의 치료, 특히 포유동물(예를 들어, 인간)에서 사용될 수 있다. "치료"는 치료적 치료 및 예방적 또는 방지적 조치 둘 다를 지칭한다. 치료를 필요로 하는 환자는 이미 장애가 있는 환자 및 장애를 예방해야하는 환자를 포함한다. 치료 목적의 "포유류"는 인간, 가축 및 농장 동물을 포함하여 포유동물로 분류된 임의의 동물, 및 동물원, 스포츠 또는 애완 동물, 예컨대 개, 말, 고양이, 소 등을 지칭한다. 바람직하게는, 포유동물은 인간이다.
본 발명의 CD96-결합제(예를 들어, 단클론성 항체)는 예를 들어 암과 감염성 질환의 치료에 있어서, T 세포 활성 및/또는 증식을 촉진하는데 사용될 수 있다. 예를 들어, CD96-결합제(예를 들어, 항체) 조성물은 암 면역요법에서 T 세포 활성 및/또는 증식을 증가시키기 위해 투여될 수 있다.
또 다른 구현예에서, 항-CD96 결합제는 암 또는 만성 감염성 질환 관련 항원에 대한 T 세포 반응을 추가로 자극하기 위해 예방 또는 치료 백신에 대한 보조제로서 사용될 수 있다. 백신의 예는 펩타이드 백신, 단백질성 백신, 바이러스-기반 백신, DNA-기반 백신, RNA-기반 백신, 자가 수지상 세포-기반 백신, 동종이계 종양 세포 백신 및 수지상 세포-기반 백신을 포함하지만, 이에 제한되지 않는다(Melero et al. 암에 대한 치료 백신: 임상 시험의 개요. Nature reviews. Clinical oncology. 11(9):509-524 (2014)).
본 발명의 CD96-결합제(예를 들어, 인간화 또는 키메라 단클론성 항체) 및 선택적으로 약제학적으로 적합한 부형제 또는 담체를 포함하는 암 치료에 사용하기 위한 약제학적 조성물이 또한 제공된다.
바람직한 구현예에서, 본 발명의 CD96-결합제(예를 들어, 항체)는 암 또는 다른 신생물 병태의 증상의 치료, 진행 지연, 재발 방지 또는 증상 완화에, 단독요법 또는 다른 항암제(들)와 조합하여(병용 요법) 사용될 수 있다. 본원에 사용된 바와 같이, 용어 "암" "신생물" 및 "종양"은 상호교환 가능하다. 암의 예는 위암, 유방암, 폐암, 난소암, 자궁경부암, 전립선암, 방광암, 결장직장암, 췌장암, 간암, 신장암, 갑상선암, 뇌암, 두경부암, 혈액암, 암종, 흑색종, 평활근종, 평활근육종, 신경교종, 교모세포종 등을 제한없이 포함한다. "혈액암"은 혈액의 암을 지칭하며, 특히 백혈병, 림프종 및 골수종을 포함한다. 고형 종양은 예를 들어 위 종양, 유방 종양, 폐 종양, 난소 종양, 전립선 종양, 방광 종양, 결장직장 종양, 췌장 종양, 간 종양, 신장 종양, 갑상선 종양, 뇌 종양, 두경부 종양, 식도 종양 및 흑색종 종양 등을 포함한다. 암 및 다른 신생물 장애와 관련된 증상은 염증, 열, 전반적인 불쾌감, 통증, 식욕 부진, 체중 감소, 부종, 두통, 피로, 발진, 빈혈, 근육 약화 및 근육 피로를 포함하지만, 이에 제한되지는 않는다.
병용 요법은 하나 이상의 추가 치료제, 예를 들어 화학요법제 또는 항신생물제, 예컨대 시토카인 및 성장 인자 억제제, 면역억제제, 항염증제, 대사 억제제, 효소 억제제 및/또는 세포독성 또는 세포정지제와 함께 공동-제형화된, 및/또는 공동-투여된 본 발명의 하나 이상의 CD96-결합제(예를 들어, 항체)를 포함할 수 있다. 이와 관련하여 용어 "조합"은 결합제가 실질적으로 같은 시기에, 동시에 또는 순차적으로 제공됨을 의미한다. 예시적인 화학치료제는 알데스류킨, 알트레타민, 아미포스틴, 아스파라기나제, 블레오마이신, 카페시타빈, 카보플라틴, 카무스틴, 클라드리빈, 시스아프리드, 시스플라틴, 시클로포스파미드, 시타라빈, 다카바진(DTIC), 닥티노마이신, 도세탁셀, 독소루비신, 드로나비놀, 듀오카르마이신, 에토포시드, 필그라스팀, 플루다라빈, 플루오로우라실, 젬시타빈, 그라니세트론, 히드록시우레아, 이다루비신, 이포스파미드, 인터페론 알파, 이리노테칸, 란소프라졸, 레바미솔, 류코보린, 메게스트롤, 메스나, 메토트렉세이트, 메토클로프라미드, 미토마이신, 미토탄, 미톡산트론, 오메프라졸, 온단세트론, 파클리탁셀(Taxol™), 필로카르핀, 프로클로로페라진, 사프로인, 타목시펜, 탁솔, 토포테칸 히드로클로라이드, 빈블라스틴, 빈크리스틴 및 비노렐빈 타르트레이트를 포함하지만, 이에 제한되지는 않는다.
본 발명의 CD96-결합제(예를 들어, 항체)는 환자 헤마토크릿을 증가시키는 유효 용량의 결합제, 예를 들어 에리트로포이에틴 자극제(ESA)와 조합될 수 있다. 이러한 결합제는 예를 들어 Aranesp®, Epogen®NF/Procrit®NF, Omontys®, Procrit® 등을 포함하여 당 업계에 공지되어 사용된다.
다른 구현예에서, 본 발명의 CD96-결합제(예를 들어, 항체)는 하기 FDA 승인 단클론성 항체를 포함하지만, 이에 제한되지 않는 암 치료에 사용된 유효 용량의 다른 항체와 조합될 수 있다: 리툭시맙(Rituxan®, CD20: 키메라 IgG1), 트라스투주맙(Herceptin®, HER2: 키메라 IgG1), 알렘투주맙(Campath®, CD52: 인간화 IgG1), 이브리투모맙 티욱세탄(Zevalin®, CD20: 뮤린, IgG1, 방사성표지된(이트륨(Yttrium) 90), 토시투모맙-I-131(Bexxar®: CD20, 뮤린, IgG2a, 방사성표지된(요오드(Iodine) 131)), 세툭시맙(Erbitux®, EGFR: 키메라, IgG1), 베바시주맙(Avastin®, VEGF: 인간화, IgG4), 파니투무맙(Vectibix®, EGFR: 인간 IgG2), 오파투무맙(Arzerra®, CD20: 인간 IgG1), 이필리무맙(Yervoy®, CTLA-4: 인간 IgG1), 브렌툭시맙 베도틴(Adcetris®, CD30: 키메라, IgG1, 약물-접합체), 퍼투주맙(Perjeta®, HER2: 인간화 IgG1), 아도트라스투주맙 엠탄신(Kadcyla®, HER2: 인간화, IgG1, 약물-접합체), 오비누투주맙(Gazyva®, CD20: 인간화 및 글리콜-조작된), 니볼루맙 및 펨브롤리주맙(항-PD-1), 등. 트라스투주맙은 HER-2 항원을 표적으로 한다. 이 항원은 유방암의 25 ~ 35%와 전이성 위암에서 나타난다. 트라스투주맙은 HER2-과발현 유방암의 치료 및 HER2-과발현 전이성 위 및 위식도 접합 선암종에 대해 승인되었다. 세툭시맙은 전이성 결장직장암, 전이성 비-소세포 폐암 및 두경부암의 치료에 사용된다. 니볼루맙 및 펨브롤리주맙은 최근에 전이성 흑색종 및 비-소세포 폐암을 치료하는 것으로 승인되었다. 그들은 폐암, 신장세포 암, 림프종 및 중피종에 대한 임상 시험에서 테스트된다. 현재 임상 시험에서 시험되거나 연구된 다른 암 약물도 병용될 수 있다.
일부 구현예에서, 항-CD96 항체는 T 세포 자극제 또는 증식제, 예를 들어 항-CD3 항체와 조합하여 사용될 수 있다. 일부 다른 구현예에서, 항-CD96 항체는 T 세포 공동-자극제, 예를 들어 CD28과 같은 T 세포 공동-자극 분자에 결합하는 결합제와 조합하여 사용될 수 있다. 예를 들어, T 세포 공동-자극제는 항-CD28, 항-ICOS, 항-CD226, 항-CD40, 항-OX40, 항-CD137, 항-GITR 또는 항-CD27 항체일 수 있다.
일부 구현예에서, 항-CD96 항체는 체크포인트 억제제와 조합될 수 있다. 예를 들어, 체크포인트 억제제는 프로그래밍된 사멸-1(PD-1) 경로의 억제제, 예를 들어 항-PD1 항체, 예컨대 니볼루맙, 아테졸리주맙, 아벨루맙 또는 펨브롤리주맙일 수 있다. 또 다른 구현예에서, 체크포인트 억제제는 항-세포독성 T-림프구-관련 항원 4(CTLA4) 항체, 예를 들어 이필리무맙 또는 트레멜리무맙이다.
면역 체크포인트는 면역 반응을 늦추거나 정지시키고 면역 세포의 제어되지 않은 활동으로 인한 과도한 조직 손상을 방지하는 억제 경로이다. "체크포인트 억제제"는 억제 경로를 억제하여 보다 광범위한 면역 활성을 허용하는 임의의 소분자 화학 화합물, 항체, 핵산 분자 또는 폴리펩타이드 또는 이의 단편을 의미한다. 특정 구현예에서, 체크포인트 억제제는 프로그래밍된 사멸-1(PD-1) 경로의 억제제, 예를 들어, 항-PD1 항체, 예컨대, 이에 제한되지 않는, 니볼루맙, 아테졸리주맙, 아벨루맙 또는 펨브롤리주맙이다. 다른 구현예에서, 체크포인트 억제제는 항-세포독성 T-림프구-관련 항원(CTLA-4) 항체이다. 추가의 구현예에서, 체크포인트 억제제는 CD28/CTLA4 Ig 수퍼패밀리의 또 다른 구성원, 예컨대 BTLA, LAG3, ICOS, PDL1 또는 KIR을 표적으로 한다(Page D.B., Postow M.A., Callahan M.K., Allison J.P. and Wolchok, J., 항체에 의해 암에서 면역 조절, Annual Review of Medicine, 65:185-202 (2014)). 추가의 부가적인 구현예에서, 체크포인트 억제제는 TNFR 수퍼패밀리의 구성원, 예컨대 CD40, OX40, 4-1BB, GITR 또는 CD27을 표적으로 한다. 일부 경우에, 체크포인트 억제제를 표적으로 하는 것은 억제 항체 또는 유사한 분자로 달성된다. 다른 경우에는 표적에 대한 효능제로 달성되며; 이 부류의 예는 자극성 표적 4-1BB, OX40 및 GITR을 포함한다.
바람직한 조합은 본 발명의 CD96 항체 및 i) 면역 체크-포인트 억제제 또는 ii) 종양 관련 항원에 대한 항체의 조합이다. Herceptin® 및 Erbitux®에 대한 예시적인 조합이 본원에 기술되어 있으며, 여기서 Herceptin®과의 조합은 그의 부가적, 협력적 또는 가능하게는 상승 효과로 인해 바람직하다. 다른 결합제들도 결합하는데 유용할 수 있다.
본 발명의 CD96-결합제(예를 들어, 단클론성 항체)는 액상에서 이용되거나 고상 담체에 결합될 수 있는 면역분석에서 시험관내에서 사용될 수 있다. 또한, 이들 면역분석에서 단클론성 항체는 다양한 방식으로 검출가능하게 표지될 수 있다. 본 발명의 단클론성 항체를 이용할 수 있는 면역분석 유형의 예는 유세포 측정법, 예를 들어 직접 또는 간접 형식의 FACS, MACS, 면역조직화학, 경쟁 및 비-경쟁 면역분석 등이다. 본 발명의 단클론성 항체를 사용한 항원의 검출은 생리학적 샘플에 대한 면역조직화학 분석을 포함하여 정방향, 역방향 또는 동시 모드로 실행되는 면역분석을 이용하여 수행될 수 있다. 당업자는 과도한 실험없이 다른 면역분석 형식을 알고 있거나 쉽게 식별할 수 있다.
본 발명의 CD96-결합제(예를 들어, 단클론성 항체)는 많은 상이한 담체에 결합될 수 있고 CD96-발현 세포의 존재를 검출하는데 사용될 수 있다. 잘 알려진 담체의 예는 유리, 폴리스티렌, 폴리프로필렌, 폴리에틸렌, 덱스트란, 나일론, 아밀라제, 천연 및 변형 셀룰로오스, 폴리아크릴아미드, 아가로스 및 마그네타이트를 포함한다. 담체의 성질은 본 발명의 목적상 가용성 또는 불용성일 수 있다. 당해 분야의 숙련가는 일상적인 실험을 사용하여 단클론성 항체에 결합하기 위한 다른 적합한 담체를 알 수 있거나, 이를 확인할 수 있을 것이다.
당해 분야의 숙련가에게 공지된 많은 상이한 표지 및 표지화 방법이 있으며, 이는 치료 방법에서 추적자로서, 진단 방법 등에 사용하기 위해 사용된다. 진단 목적을 위해, 표지는 CDR 서열로 구성되거나 CDR 서열을 포함하는 단편을 포함하여, 본 발명의 항체 또는 이의 단편에 공유적으로 또는 비-공유적으로 부착될 수 있다. 본 발명에 사용될 수 있는 표지 유형의 예는 효소, 방사성동위원소, 형광 화합물, 콜로이드성 금속, 화학발광 화합물 및 생물-발광 화합물을 포함한다. 당업자는 본 발명의 CD96-결합제(예를 들어, 단클론성 항체)에 결합하기 위한 다른 적합한 표지를 알 수 있거나, 일상적인 실험을 이용하여 이를 확인할 수 있을 것이다. 또한, 본 발명의 CD96-결합제(예를 들어, 단클론성 항체)에 대한 이들 표지의 결합은 당해 분야의 숙련가에게 일반적인 표준 기술을 사용하여 수행될 수 있다.
일부 구현예에서, CD96-결합제(예를 들어, 단클론성 항체 또는 이의 단편)는 예를 들어, 이미징에 사용하기 위해, 나노입자에 부착된다. 유용한 나노입자는 예를 들어 라만-실리카-금-나노입자(R-Si-Au-NP)를 포함하지만, 이에 제한되지 않는 당 업계에 공지된 것들이다. R-Si-Au-NP는 협-대역 스펙트럼 시그니처를 갖는 라만 유기 분자로 구성되며, 금 코어에 흡착된다. 라만 유기 분자가 변경될 수 있기 때문에, 각각의 나노입자는 고유한 시그니처를 가질 수 있어, 다중화에 의해 다수의 나노입자가 동시에 독립적으로 검출될 수 있게 한다. 전체 나노입자는 실리카 쉘에 캡슐화되어 금 나노코어 상에 라만 유기 분자를 보유한다. R-Si-Au-NP의 선택적인 폴리에틸렌 글리콜(PEG)-일화는 생체이용률을 증가시키고 표적 부분을 부착하기 위한 기능적 "핸들"을 제공한다(Thakor A.S., Luong R., Paulmurugan R., et al. et al., 마우스에서 라만-활성 실리카-금 나노입자의 운명 및 독성, Sci. Transl. Med. 3(79):79ra33 (2011); Jokerst J.V., Miao Z., Zavaleta C., Cheng Z., and Gambhir S.S., 표피 성장 인자 수용체의 라만 분자 이미징을 위한 아피바디-작용화된 금-실리카 나노입자, Small. 7(5):625-633 (2011); Gao J., Chen K., Miao Z., Ren G., Chen X., Gambhir S.S. and Cheng Z., HER2-발현 세포 및 종양 이미징을 위한 아피바디-기반 나노프로브, Biomaterials 32(8):2141-2148 (2011) 참고; 각각 본원에 구체적으로 참고로 포함됨).
본 발명의 목적을 위해, CD96은 생체내 또는 시험관내 생물학적 유체 및 조직에 존재할 때 본 발명의 CD96-결합제(예를 들어, 단클론성 항체)에 의해 검출될 수 있다. 검출가능한 양의 CD96을 함유하는 임의의 샘플이 사용될 수 있다. 샘플은 소변, 타액, 뇌척수액, 혈액, 혈청 등과 같은 액체, 또는 조직, 대변 등과 같은 고체 또는 반고체, 또는 대안적으로 조직학적 진단에 일반적으로 사용되는 것과 같은 고체 조직일 수 있다.
더 큰 민감도를 초래할 수 있는 다른 표지 기술은 항체를 저분자량 합텐에 커플링하는 것으로 구성된다. 이러한 합텐은 이어서 제2 반응에 의해 구체적으로 검출될 수 있다. 예를 들어, 아비딘과 반응하는 바이오틴, 또는 특정 항-합텐 항체와 반응할 수 있는, 디니트로페놀, 피리독살 또는 플루오레세인과 같은 합텐을 사용하는 것이 일반적이다.
편의상, 본 발명의 CD96-결합제(예를 들어, 항체)는 키트, 즉 진단 분석을 수행하기 위한 설명서와 함께 미리 결정된 양의 시약의 패키지된 조합으로 제공될 수 있다. CD96-결합제(예를 들어, 항체)가 효소로 표지된 경우, 키트는 효소에 의해 요구되는 기질 및 보조인자(예를 들어, 검출가능한 발색단 또는 형광단을 제공하는 기질 전구체)를 포함할 것이다. 또한, 안정화제, 완충제(예를 들어, 블록 완충제 또는 용해 완충제) 등과 같은 다른 첨가제가 포함될 수 있다. 다양한 시약의 상대적인 양은 분석의 민감도를 실질적으로 최적화하는 시약의 용액에 농도를 제공하도록 광범위하게 변할 수 있다. 특히, 시약은 용해시 적절한 농도를 갖는 시약 용액을 제공할 부형제를 포함하여, 보통 동결건조된 건조 분말로서 제공될 수 있다.
본 발명의 하나 이상의 CD96-결합제(예를 들어, 항체)를 포함하는 치료용 제형은 원하는 순도를 갖는 CD96-결합제(예를 들어, 항체)를 동결건조 제형 또는 수용액 형태의, 임의로 생리학적으로 허용가능한 담체, 부형제 또는 안정화제(Remington's Pharmaceutical Sciences, 16th edition, Osol, A. Ed. (1980))와 혼합함으로써 저장을 위해 제조된다. 상기 조성물은 우수한 의학적 실무와 일치하는 방식으로 제형화, 투약 및 투여될 것이다. 이러한 맥락에서 고려할 요소는 치료되는 특정 장애, 치료되는 특정 포유동물, 개별 환자의 임상 상태, 장애의 원인, 결합제의 전달 부위, 투여 방법, 투여 스케줄 및 의료 종사자에게 알려진 기타 요인을 포함한다. 투여될 CD96-결합제(예를 들어, 항체)의 "치료적 유효량"은 이러한 고려 사항에 의해 지배될 것이며, 질병을 예방하는데 필요한 최소량이다.
치료 용량은 체중 kg 당 적어도 약 0.01 mg, 체중 kg 당 적어도 약 0.05 mg; 체중 kg 당 적어도 약 0.1 mg, 체중 kg 당 적어도 약 0.5 mg, 체중 kg 당 적어도 약 1 mg, 체중 kg 당 적어도 약 2.5 mg, 체중 kg 당 적어도 약 5 mg, 체중 kg 당 적어도 약 10 mg, 및 체중 kg 당 약 100 mg 이하일 수 있으며, 체중 kg 당 0.1 내지 20 mg이 바람직할 수 있다. 당업자는 이러한 가이드라인이 예를 들어 항체 단편의 사용시에, 또는 항체 접합체의 사용시에, 활성제의 분자량에 따라 조정될 것임을 이해할 것이다. 투여량은 또한, 국소 투여, 예를 들어 비강내, 흡입 등을 위해, 또는 전신 투여, 예를 들어 i.m., i.p., i.v., s.c. 등을 위해 변할 수 있다.
CD96-결합제(예를 들어, 항체)는 활성을 강화시키거나 치료 효과를 증가시키는 하나 이상의 결합제와 함께 임의로 배합될 수 있다. 이들은 일반적으로 이전에 사용된 것과 동일한 투여량 및 투여 경로, 또는 이전에 사용된 투여량의 약 1 내지 99%로 사용된다.
허용가능한 담체, 부형제 또는 안정화제는 사용된 투여량 및 농도에서 수용자에게 무독성이며, 포스페이트, 시트레이트 및 기타 유기산과 같은 완충제; 아스코르브 산 및 메티오닌을 포함하는 항산화제; 방부제(예컨대, 옥타데실디메틸벤질 암모늄 클로라이드; 헥사메토늄 클로라이드; 벤즈알코늄 클로라이드, 벤제토늄 클로라이드; 페놀, 부틸 또는 벤질 알코올; 메틸 또는 프로필 파라벤과 같은 알킬 파라벤; 카테콜; 레조르시놀; 시클로헥사놀; 3-펜탄올; 및 m-크레졸); 저분자량(약 10 잔기 미만) 폴리펩타이드; 혈청 알부민, 젤라틴 또는 면역글로불린과 같은 단백질; 친수성 중합체, 예컨대 폴리비닐피롤리돈; 글리신, 글루타민, 아스파라긴, 히스티딘, 아르기닌 또는 리신과 같은 아미노산; 단당류, 이당류 및 글루코스, 만노스 또는 덱스트린을 포함한 다른 탄수화물; 킬레이트제, 예컨대 EDTA; 수크로스, 만니톨, 트레할로스 또는 소르비톨과 같은 당; 나트륨과 같은 염-형성 반대-이온; 금속 착물(예를 들어, Zn-단백질 착물); 및/또는 비-이온성 계면활성제, 예컨대 TWEEN™, PLURONICS™ 또는 폴리에틸렌 글리콜(PEG)을 포함한다. 생체내 투여에 사용될 제제는 멸균되어야 한다. 이것은 멸균 여과막을 통한 여과에 의해 용이하게 달성된다.
활성 성분은 또한 예를 들어 코아세르베이션 기술 또는 계면중합, 예를 들어 히드록시메틸셀룰로스 또는 젤라틴-마이크로캡슐 및 폴리-(메틸메타실레이트) 마이크로캡슐에 의해 제조된 마이크로캡슐에, 각각 콜로이드 약물 전달 시스템(예를 들어, 리포좀, 알부민 마이크로스피어, 마이크로에멀젼, 나노입자 및 나노캡슐) 또는 마크로에멀젼에 포획될 수 있다. 이러한 기술은 Remington's Pharmaceutical Sciences, 16th edition, Osol, A. Ed. (1980)에 개시되어 있다.
CD96-결합제(예를 들어, 항체)는 비경구, 피하, 복강내, 폐내 및 비강을 포함한 임의의 적합한 수단에 의해 투여된다. 비경구 주입은 근육내, 정맥내, 동맥내, 복강내 또는 피하 투여를 포함한다. 또한, CD96-결합제(예를 들어, 항체)는 펄스 주입에 의해, 특히 결합제의 감소된 용량으로 적절하게 투여된다.
질환의 예방 또는 치료를 위해, 적절한 투여량의 CD96-결합제(예를 들어, 항체)는 상기 정의된 바와 같이 치료될 질환의 유형, 질병의 중증도 및 과정, 예방 목적을 위해 결합제가 투여되는지 여부, 이전 요법, 환자의 임상 이력 및 결합제에 대한 반응, 및 주치의의 재량에 따라 달라질 것이다. 결합제는 한번에 또는 일련의 치료에 걸쳐 환자에게 적절하게 투여된다.
본 발명의 또 다른 구현예에서, 상기 기재된 장애의 치료에 유용한 물질을 함유하는 제조 물품이 제공된다. 제조 물품은 컨테이너 및 표지를 포함한다. 적합한 컨테이너는 예를 들어 병, 바이알, 주사기 및 시험관을 포함한다. 컨테이너는 유리 또는 플라스틱과 같은 다양한 재료로 형성될 수 있다. 컨테이너는 병태를 치료하는데 효과적인 조성물을 보유하며, 멸균 접근 포트를 가질 수 있다(예를 들어, 컨테이너는 정맥내 용액 백 또는 피하 주사 바늘에 의해 관통될 수 있는 스토퍼를 갖는 바이알일 수 있음). 조성물 중의 활성제는 CD96-결합제(예를 들어, 항체)이다. 컨테이너상의 또는 컨테이너와 관련된 표지는 조성물이 선택된 병태를 치료하는데 사용됨을 나타낸다. 제조 물품은 약제학적으로-허용가능한 완충제, 예컨대 포스페이트-완충 식염수, 링거액 및/또는 덱스트로스 용액을 포함하는 제2 컨테이너를 추가로 포함할 수 있다. 다른 완충제, 희석제, 필터, 바늘, 주사기 및 사용 설명서가 들어 있는 포장 삽입물을 포함하여 상업적 및 사용자 관점에서 바람직한 다른 물질을 추가로 포함할 수 있다.
본 발명은 이제 완전히 설명되었지만, 본 발명의 사상 또는 범위를 벗어나지 않으면서 다양한 변경 및 변형이 이루어질 수 있음이 당해 분야의 숙련가에게 명백할 것이다.
실시예
1. 인간 CD96의 특징분석
1.1 T, NK 및 다른 조혈 세포 집단에서 CD96 발현
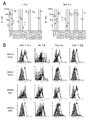
인간 PBMC의 상이한 집단상에서의 인간 CD96의 발현은 항-CD96 클론 628211(R&D Biotech) 및 NK92.39(Fuchs, 2004) 항체를 사용하여 시험되었다. CD4+ 및 CD8+ 세포, NK 세포 및 소수의 B 세포에서의 CD96 발현은 도 1A 및 1B에서 입증된다. NK, CD4+ 및 CD8+ T 세포는 또한 CD226을 발현한다(도 1C). NK 세포 및 T 세포의 약 40%가 CD226 및 CD96을 공-발현한다(도 1D). NK 세포와 비교하여 CD226을 발현하는 높은 백분율의 T 세포는 CD96을 공-발현한다(도 1E).
보다 구체적으로, 건강한 공여체의 말초 혈액 단핵 세포(PBMC) 중에서 NK 세포의 38.5%(중간 값, 55~22.5% 범위, n=8), CD4+ T 세포의 39.1%(53.2~29%, n=4) 및 CD8+ T 세포의 45.6%(53.8~26.2%, n=4)는 CD96 및 CD226을 공-발현한다. 흥미롭게도, NK 세포의 51%(75~25%, n=8), CD4+ T 세포의 96.8%(99.6~96.5%, n=4) 및 CD226을 발현하는 CD8+ T 세포의 73.1%(87.7~68.1%, n=4)는 CD96을 공-발현하는 것으로 밝혀졌다. NK 세포의 27%(45~20%, n=4), CD4+ T 세포의 15.5%(20~8%, n=4), CD226 및 CD96을 발현하는 CD8+ T 세포의 28%(30~16%, n=4)는 또한 TIGIT를 공-발현한다(데이터는 나타내지 않음).
FCM 분석은 CD226과 마찬가지로 CD96이 휴지기의 나이브 T 세포보다 CD45RO+ 기억 T 세포에서 더 강력하게 발현됨을 보여주었다(도 2).
CD226에 대해 관찰(Lozano et al. TIGIT/CD226 축 조절은 인간 T 세포 기능을 조절한다. J. Immunol. 188:3869-3875 (2012))된 바와 같이, CD96의 발현은 항-CD3 항체(OKT3)로 활성화된 CD4+ 및 CD8+ 기억 T 세포 모두에서 증가되었지만, 이 활성화 조건에서 나이브 T 세포에서는 그렇지 않았다(도 3).
마지막으로, CD96의 발현은 CD25의 높은 발현 및 CD127 마커의 낮은 발현에 의해 확인되는, 순환 CD4+ 조절 T 세포(Treg)상에서 FCM에 의해 조사되었다. 도 4에 도시된 바와 같이, CD96은 CD226과 유사하게 종래의 CD4+ T 세포(CD25lo/CD127+ 세포)와 비교하여 CD4+ Treg 세포에서 CD96이 덜 발현되는 것으로 밝혀졌다(Fuhrman et al. 수용체 TIGIT 및 CD226을 발현하는 인간 조절 T 세포의 다른 표현형. J Immunol. 195(1):145-155 (2015)). 따라서, 효능성 항-CD96 치료 항체의 사용은 조절 T 세포보다는 통상적인 T 세포를 우선적으로 공동-자극할 것으로 예상된다.
1.2 CD96은 CD226으로 동종이량체 및 이종이량체를 형성할 수 있다
CD226은 CD226과 CD155의 효율적인 상호작용 및 CD226을 발현하는 세포의 활성화에 필요한 시스-동종이량체를 형성할 수 있는 것으로 설명되었다. CD226은 또한 CD226 동종이량체화를 방해하여 아마도 CD226/CD155 상호작용의 억제를 통해 CD226 활성화를 방해하는 TIGIT와 시스-이종이량체를 형성할 수 있다(Johnston et al. 면역수용체 TIGIT는 항종양 및 항바이러스 CD8+ T 세포 효과기 기능을 조절한다. Cancer cell 26(6):923-937 (2014)). 그러나, CD96이 CD226과 동종이량체 및/또는 이종이량체를 형성할 수 있는지 여부는 알려져 있지 않다. 따라서, 본 발명자들은 피코에리트린(PE - 488 nm 레이저에 의해 여기됨)과 알로피코시아닌(APC - 방출 필터 670 LP를 사용하여 검출됨)으로 태그된 항체들 사이의 형광 공명 에너지 전이(FRET)를 사용하여 hCD96과 hCD226에 의한 hCD96의 동종 및 이종-이량체화를 조사하였다(Batard et al. 유세포 측정법에 의해 분석된 형광 공명 에너지 전이를 위한 피코에리트린 및 알로피코시아닌의 용도: 장점 및 한계. Cytometry 48(2):97-105 (2002)). 본 발명자들은 플라스미드 HG10565-NH, HG11202-NY, HG10917-NM(SinoBiological) 각각에 의한 형질감염에 의해 hCD96-HA 및/또는 hTIGIT-Myc 태그된 단백질과의 조합으로 또는 없이, CHO 세포에서 hCD226-His 태그된 단백질을 일시적으로 발현하였다. CD226, CD96 및 TIGIT는 PE 및 APC에 커플링된 항-hCD226, -hCD96, -hTIGIT, -His, -HA 또는 -Myc mAb로 표지되었다. 대조군으로서, 세포는 항-태그 mAbs-PE 또는 항-태그 mAbs-APC로 개별적으로 표지된 후 풀링된다. 형질감염된 세포를 mAbs와 함께 +4℃에서 30분 동안 인큐베이션한 후, 여러 번 세척한 후, FACSCanto II 유세포 분석기(BD biosciences)에서 유세포 측정법에 의해 분자들 사이의 상호작용(PE-접합된 mAbs와 APC-접합된 mAbs 사이의 FRET)을 분석하였다. 주목할 점은, 형질감염된 각각의 조건에 대해, mAb의 4가지 조합, 즉 항-태그-PE + 항-태그-APC; 항-태그-PE + 항-수용체-APC, 항-수용체-PE + 항-수용체-APC 및 항-수용체-PE + 항-태그-APC가 시험되었다. Batard et al에 의해 기술된 바와 같이, mAb의 일부 조합 만이 에너지 전달을 허용할 수 있다(데이터 미도시)(Batard, 2002).
hCD226이 hCD226-hCD226 동종이량체(도 5A) 및 hTIGIT를 갖는 이종이량체(도 5B)를 형성함을 확인하였다. 중요하게도, hCD96이 hCD226을 갖는 이종이량체(도 5D)뿐만 아니라 hCD96-hCD96 동종이량체(도 5C)를 형성할 수 있음을 관찰하였다. 따라서, TIGIT와 같이, CD96은 CD226과 이종이량체를 형성하여, CD226-CD155 상호작용의 새로운 억제 메커니즘을 야기하거나, CD155와의 상호작용을 통해 CD226 및 CD96을 발현하는 세포의 새로운 활성화 메커니즘(공동-자극)을 나타낼 수 있다.
문헌에 이미 기술된 바와 같이, CD96의 발현은 건강한 개체의 순환 NK, T 및 NKT 세포에서 확인되었다. 또한, CD96은 대다수의 NK, T 및 NKT 세포에서 CD226과 공-발현되는 것으로 밝혀졌다. 순환 NK 세포에서, 본 발명자들은 휴지기의 및 활성화된 CD56dim 및 CD56bright/CD16lo 서브셋에서 CD96이 발현되는 반면, TIGIT는 CD56bright/CD16lo 서브셋에서 발현되지 않는 것으로 관찰하였다(데이터는 나타내지 않음). 순환 T 세포에서, 본 발명자들은 CD96이 CD4+ 기억 T 세포 및 CD226과 같은 나이브 및 기억 CD8+ T 세포 둘 다에서 주로 발현됨을 관찰하였다. CD96의 발현은 CD226과 같은 항-CD3-활성화된 기억 T 세포에서 추가로 증가된다. 마지막으로, 본 발명자들은 CD96의 발현이 통상적인 CD4+ T 세포와 비교하여 순환 CD4+ Treg 집단에서 더 약한 것을 관찰하였다.
또한, 본 발명자들은 인간 CD96이 인간 CD226과의 이종이량체뿐만 아니라 동종이량체를 형성할 수 있음을 입증하였다. 이 이량체화는 CD96 및 CD226을 발현하는 세포에서 CD96의 기능적 활성에 중요한 역할을 할 수 있다.
공개된 항-CD96 항체 중 일부가 인간 T 세포를 공동-자극함(하기 참조)을 나타내는 이러한 관찰 결과와 함께, 효능제 항-CD96 항체가 종래의 CD8+ 및 CD4+ T 세포를 우선적으로 자극하여 암과 같은 질환이 있는 환자의 세포 면역 반응을 증가시키기 위한 용도를 찾음을 제시한다. CD96의 자극 활성은 단량체 또는 동종이량체로서, 및/또는 활성화 수용체 CD226을 갖는 이종이량체로서, CD96을 유발함으로써 매개될 수 있다.
2. 마우스 항-인간 CD96 항체의 생성
2.1 달리 언급되지 않는 한, 분석에 사용된 인간 CD96은 CD96의 더 짧은 형태(엑손 4가 없는 이소형 v2)이며, 이는 인간 림프구에서 발현되는 주요 형태이다.
인간 CD96에 대해 마우스 항체를 생성하기 위해, 마우스를 2주 간격으로 4회의 DNA 주사, 이 후 1주 간격으로 2회의 최종 부스트를 포함하는 프로토콜을 적용하여 면역화하고 혈청 스크리닝 결과에 따라 동물 희생하였다:
- 5WT 마우스: 프라임 인간CD96 DNA + 부스트 인간CD96 DNA
- 5WT 마우스: 프라임 인간CD96 DNA + 부스트 인간CD96 CHO(hCD96 발현 벡터로 형질감염된 CHO 세포)
인간 CD96에 대한 IgG 결합의 존재 및 역가는 ELISA(hCD96-humanIgG1 융합 단백질, hCD96-hFc의 코팅, 사내에서 생산 및 정제) 및 hCD96을 발현하는 CHO 세포의 유세포 측정법에 의해 면역화된 동물의 혈청에서 모니터링되었다. CD155-CD96 상호작용을 억제하는 mAb의 존재는 또한 간섭 ELISA를 사용하여 시험하였다. 강한 항-CD96 IgG 역가를 나타내는 동물을 희생시켰다. 그들의 비장 및 림프절을 추출하고, 단핵 세포(MNC)를 정제하고 동결시켰다. 인간CD96-CHO로 부스트된 5마리 마우스로부터의 세포를 풀링하였다.
2.2 ISAAC 기술을 이용한 단일 B 세포 스크리닝 및 재조합 항-CD96 항체 생성
WO2009/017226에 기재된 ISAAC 기술은 마이크로어레이 칩을 사용하여 개별 항체 분비 세포를 검출하는 독특한 방법으로, 단일-세포 단위로 살아있는 세포를 분석할 수 있고, 관련 세포를 식별하고 회수하기 위한 빠르고 효율적이며 높은 처리량(최대 234,000개의 개별 세포) 시스템을 제공한다.
면역화된 마우스의 비장 및 림프절로부터 미리 정제된 마우스 MNC를 마이크로어레이 칩에 적용함으로써 단일의 살아있는 세포 어레이를 제조하였다. 칩 표면은 미리 표적 항원(hCD96-hFc)으로 코팅되었고, 항체 분비 세포에 의해 분비된 항-CD96 마우스 항체는 웰 주위 표면에 코팅된 CD96에 의해 포획되었다. 세척 후, 고정화된 CD96에 결합된 마우스 IgG의 존재는 Cy3 및 형광 현미경에 커플링된 항-마우스 IgG 항체에 의해 검출되었다. 특이적 항체에 대한 항원의 결합은 비특이적 신호와 쉽게 구별되는 별개의 원형 스팟을 형성하였다. 이어서, 단일 세포를 분비하는 CD96-특이적 항체를 미세조작에 의해 검색하고, mRNA를 단일 세포로부터 회수하고, IgG의 중쇄(VH) 및 경쇄(VL)의 가변 영역을 암호화하는 cDNA 서열을 RT-PCR에 의해 증폭시켰다. 이어서 VH 및 VL 서열을 각각 마우스 감마-2a 불변 영역(Fcγ2a) 및 카파 불변 영역을 함유하는 발현 벡터에서 클로닝하였다. Fcγ 수용체에 대한 mIgG2a의 친화성을 감소시키는 D265A 돌연변이(mIgG2a-D265A)를 함유하는 mIgG2a 포맷으로 항체를 또한 클로닝하였다.
CHO 세포에서 H 및 L 쇄 발현 벡터의 공동-형질감염 후, hCD96-hFc로 코팅된 플레이트 또는 hFc에 융합된 비관련 인간 단백질로 코팅된 플레이트에서 ELISA에 의해 CD96 인식 및 특이성을 확인하기 위해 재조합 항체를 테스트하고, hCD96-hFc 단백질에 관해서 생산 및 정제하였다. CD155-CD96 상호작용의 억제는 ELISA에 의해 모니터링하였다. 천연 CD96의 인식 및 CD96-CD155 상호작용의 억제는 hCD96 또는 비관련 단백질로 형질감염된 CHO 세포에 대한 유세포 측정법에 의해 확인하였다.
총 160쌍의 VH 및 VL을 증폭시키고, 생성된 IgG 항체를 인간 CD96에 대한 특이적 결합에 대해 확인하였다. 그중 19개의 항체가 생산되어 정제되었다. 이들 19개 항체로부터 12개가 hCD155-hCD96 상호작용을 억제하였다.
3. 재조합 마우스 항-CD96 항체의 시험관내 특징분석
3.1 인간 CD96을 발현하는 CHO 세포 상의 FACS 결합
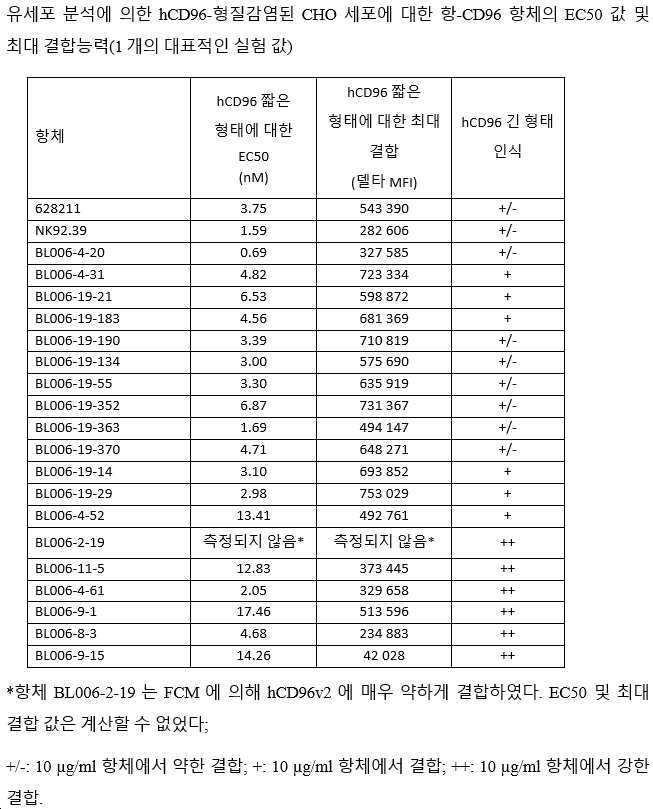
세포막 발현된 인간 CD96을 인식하는 동정된 마우스 항-CD96 항체의 능력은 CD96 짧은 이소형 v2(CD96v2; Genbank 수탁 번호 NM_005816, HA-태그가 있는 C-말단에 플랭킹됨)(서열번호: 268)로 안정적으로 형질감염된 CHO 세포를 사용하여 유세포 분석(FCM)에 의해 추가로 분석되었다. 인간 CD96(v1 이소형; CD96v1)의 긴 형태(서열번호: 267)의 인식은 또한 인간 CD96v1(Genbank 수탁 번호 NM_198196.2)을 일시적으로 발현하는 CHO 세포에 의한 유세포 측정법에 의해 시험하였다. 19개 후보의 결합능력을 2개의 mIgG1 벤치마크 항체 NK92.39(Fuchs, 2004; Biolegend) 및 클론 628211(R&D System)의 결합능력과 비교하였다.
마우스 항-CD96 항체 및 이소타입 대조군 항체를 +4℃에서 30분 동안 인간 CD96을 발현하는 CHO 세포와 함께 10 ㎍/mL 내지 4.6 ng/mL의 다양한 농도로 인큐베이션하였다. 2회 세척한 후, 세포막 CD96에 결합된 항체의 존재는 PE-커플링된 항-마우스 항체와의 인큐베이션 및 Accuri-C6 유세포 분석기(BD-Biosciences)에 대한 분석에 의해 밝혀졌다. 각각의 항체 농도에 대해 수득된 평균 형광 강도(MFI)와 일차 항체가 없는 상태에서 수득된 강도 사이의 차이(델타-MFI)를 계산하고 항체 농도에 대해 플롯팅하였다. CD96을 인식하지 않은 적절한 이소타입의 음성 대조군 항체를 동일한 조건에서 시험하여 항체 염색(비특이적 염색)의 배경을 측정하였다.
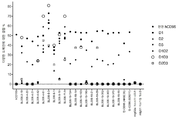
19개의 마우스 항-hCD96 항체에 대해 수득된 결합 결과는 도 6에 도시되고, 표 6에 요약되어 있으며, 벤치마크 항체에 대해 수득된 결과와 비교된다.
시험된 19개의 항체 중 11개의 후보가 hCD96v2에 강력하게 결합하는 것으로 밝혀졌다(BL006-4-31, BL006-19-183, BL006-19-190, BL006-19-134, BL006-19-21, BL006-19-55, BL006-19-352, BL006-19-363, BL006-19-370, BL006-19-14, BL006-19-29). 이들 후보의 hCD96v2에 대한 결합은 클론 628211의 것과 유사하거나 우수하였다. 6개의 후보들(BL006-4-20, BL006-4-52, BL006-4-61, BL006-11-5, BL006-8-3, BL006-9-1)은 벤치마크 NK92.39와 마찬가지로 CD96v2에 더 약하게 결합하였다. 후보 BL006-9-15는 CD96v2에 더욱더 약하게 결합하였다. 최종적으로, 후보 BL006-2-19는 FCM에 의해 CD96v2에 매우 약하게 결합하는 것으로 나타났다(도시되지 않음).
시험된 19개의 항체 중, 11개의 후보가 FCM에 의해 CHO 세포상에서 발현된 긴 형태의 인간 CD96(CD96v1)에 강하게 결합하는 것으로 밝혀졌다(BL006-4-31; BL006-19-21; BL006-19-183, BL006-19-14, BL006-19-29, BL006-4-52; BL006-2-19, BL006-11-5, BL006-4-61, BL006-9-1, BL006-8-3, BL006-9-15). 나머지 후보들은 2개의 벤치마크 NK92.39 및 628211에서와 같이, 긴 형태의 hCD96(CD96v1)에 약하게 결합하였다.
3.2 마우스와 레서스 CD96과의 교차-반응성
다른 종으로부터의 CD96 단백질을 인식하는 동정된 마우스 항-인간 CD96 항체의 능력을 마우스 또는 레서스 CD96을 발현하는 CHO 세포를 사용하여 유세포 측정법에 의해 추가로 분석하였다. CHO 세포 표면에서 종-특이적 CD96의 발현은 적절한 항-CD96 항체에 의한 염색 및 비-고정 세포상에서의 유세포 측정법을 사용하여 확인하였다. 랫트 항-mCD96 항체 클론 #630612(R&D Systems) 및 후보 BL006-9-1(레서스 CD96 상에서의 ELISA에 의해 양성)을 사용하여 레서스 CD96의 발현을 확인한 반면, 클론 6A6(랫트 IgG2a, e-Bioscience)를 사용하여 마우스 CD96의 발현을 확인하였다.
마우스 항-CD96 항체 및 이소타입 대조군 항체를 +4℃에서 30분 동안 상이한 CD96 종을 발현하는 CHO 세포와 함께 10 또는 1 ㎍/mL에서 인큐베이션하였다. 2회 세척 후, 세포막 CD96에 결합된 항체의 존재는 PE-커플링된 항-마우스 항체와 함께 인큐베이션하고 Accuri-C6 유세포 측정기(BD-Biosciences)에서 분석함으로써 밝혀졌다.
19개의 마우스 항-CD96 항체에 대해 얻은 결과가 도 7에 도시되어 있고, 벤치마크 항체에 의해 얻어진 결과와 비교된다. 시험된 19개의 후보들 중 8개의 항체가 레서스 CD96을 강하게 인식했다(BL006-4-31, BL006-19-352, BL006-19-14, BL006-19-29, BL006-4-52, BL006-2-19, BL006-4-61, BL006-9-1). 2개의 후보 BL006-19-21 및 BL006-19-55는 항체 628211과 같이 레서스 CD96에 약하게 결합하였다. 9개의 나머지 후보 및 벤치마크 NK92.39는 레서스 CD96에 결합하지 않았다. 참고로, 19개의 후보들 중 어느 것도 FCM에 의해 마우스 CD96을 인식하지 않았다(데이터는 나타내지 않음).
3.3 CHO 세포에 대한 유세포 측정법에 의해 측정된 CD96/CD155 상호작용의 억제
FCM에 의해 hCD96v2에 매우 약하게 결합된 BL006-2-19를 제외한 모든 후보들은 CHO 세포상에서 안정적으로 발현된 hCD96v2에 대한 인간 CD155의 결합을 억제하는 능력에 대해 시험되었다. 인간 CD96-형질감염된 CHO 세포(3x105 세포/96-웰 플레이트의 웰)를 4 ㎍/mL의 비오티닐화 hCD155-Fc(BliNK에서 생성 및 비오티닐화됨)의 존재하에 항-CD96 항체 또는 이소타입 대조군 항체(mIgG2a 또는 mIgG2aD265A 형식)의 연속 희석으로 +4℃에서 30분 동안 인큐베이션하였다. 이어서, 세포를 2회 세척하고, FITC-접합된 스트렙타비딘(BD Biosciences)과의 인큐베이션 및 Accuri-C6 유세포 측정기(BD biosciences)에 대한 유세포 분석으로 CD155-hFc의 CHO 세포에 대한 결합이 밝혀졌다.
hCD96에 대한 hCD155 결합의 억제 백분율은 하기와 같이 계산되었다:
% 억제 = (1-(MFI_wAb&CD155 - MFI_CHO)/(MFI_wCD155 - MFI_CHO))x100, 여기서 MFI_wAb&CD155는 시험된 항체 및 hCD155-hFc와 함께 인큐베이션된 hCD96-CHO 세포로 얻은 평균 강도 형광(MFI)이고; MFI_CHO는 hCD155 및 mAbs의 부재하에 얻은 MFI이고; MFI_wCD155는 hCD155에 의한 형광이지만 항체와의 사전-배양이 없는 형광(0% CD155 결합 억제)이다. 억제율을 항체 농도에 대해 플롯팅하고, GraphPad Prism 소프트웨어의 비선형 회귀 분석 모델을 사용하여 IC50 값을 계산하였다.
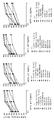
도 8 및 표 7에 도시된 결과는 9개의 후보가 CHO 세포상에서 발현된 hCD155와 hCD96v2의 결합을 5.9 내지 19.4 nM 범위의 IC50 값으로 강력하게 억제함을 나타낸다(후보 BL006-4-31, BL006-19-21, BL006-19-183, BL006-19-190, BL006-19-134, BL006-19-55, BL006-19-352, BL006-19-363, BL006-19-370). 이들 9개의 항-CD96 후보는 클론 628211 및 NK92.39 항체와 유사하였다. 후보 BL006-4-20, BL006-19-14 및 BL006-19-29는 또한 CD96v2에 대한 CD155 결합을 억제했지만, 30 ㎍/mL(200 nM)에서 결합 억제의 50%에 도달하지 않았기 때문에 더 낮은 효능을 가졌다. 마지막으로, 6개의 후보 BL006-4-52, BL006-4-61, BL006-11-5, BL006-8-3, BL006-9-1 및 BL006-9-15는 30 μg/mL(200 nM)에서 CD96v2/CD155 상호작용을 억제하지 않았다.
3.4 인간 PBMC에서 항-CD96 후보의 결합
항-CD96 mAb 후보를 인간 PBMC에 대한 그의 결합 능력에 대해 테스트하였다. mAb를 미리 비오티닐화시켰다(EZ-링크 설포-NHS-LC-비오틴). 3개의 건강한 공여체로부터의 PBMC를 항-CD3-PerCP(클론 SK7, BD biosciences), CD56-FITC(클론 REA196 Miltenyi) 및 항-CD96-비오티닐화(10 ㎍/ml)로 염색하였다. T 세포 집단은 CD3pos, CD56neg으로 게이팅하고(평균: 64.9%, SD: 림프구 집단의 9.4%), NK 세포를 CD3neg, CD56pos로 게이팅하고(평균: 11.1%, SD: 림프구 집단의 1.7%), 및 NKT 세포를 CD3pos, CD56pos로 게이팅하였다(평균: 4.2%, SD: 림프구 집단의 2.9%). 스트렙타비딘-APC(Abcam)를 사용하여 항-CD96-비오티닐화된 mAb를 검출하였다.
테스트한 18개 항체(BL006-9-1은 테스트하지 않음) 중 9개 후보(BL006-4-20, BL006-4-31, BL006-19-29, BL006-19-14, BL006-19-55, BL006-19-183, BL006-19-190, BL006-19-134, BL006-19-21)은 T 및 NK 세포(도 9A)에서 뿐만 아니라 NKT 세포(도시되지 않음)상에 강한 결합을 나타냈다. 7개의 후보(BL006-2-19, BL006-8-3, BL006-4-61, BL006-19-352, BL006-19-363, BL006-19-370, BL006-4-52)는 NK, T 및 NKT 세포 상에 약한 결합을 나타냈으며(도 9A 및 미도시), 및 2개(BL006-11-5 및 BL006-9-15)는 NK, T 및 NKT 세포에서 매우 약한 결합을 나타냈다.
CD96의 발현은 두경부 종양으로부터 수득된 인간 종양 침윤 림프구(TIL)에서 시험되었다. TIL을 추출하기 위해, 두경부 종양을 무혈청 RPMI 배지로 2회 세척하고, 혈전을 족집게로 제거하였다. 멸균된 메스 블레이드를 사용하여 종양 샘플을 먼저 기계적으로 분해하여 작은 조각으로 절단한 다음, 연속 교반하면서 37℃에서 20분 동안 DNAse I(800 U/mL) 및 리버라제 디스파제 로우(Liberase Dispase Low)(0.15 WU/mL)를 사용하여 효소적으로 소화시켰다. 효소 반응을 5 ml RMPI 10% FCS로 켄칭하고, 분해된 종양 샘플은 100 μm 세포 스트레이너를 통과시켰다. 계수하기 전에, 적혈구를 실온에서 5분 동안 적혈구 용해 완충제를 사용하여 제거하였다. 4명의 환자로부터 얻은 TIL은 항-CD3-eFluor450(UCHT1, ebioscience), 항-CD56-PC7(CMSSB, ebioscience), 항-CD4-APC-eFluor780(RPA-T4, ebioscience), 항-CD8-FITC(RPA-T8, Biolegend), 항-FoxP3-PE(PCH101(CL), ebioscience) 및 항-CD96-APC(FAB6199A, R&D Systems)로 염색되었다. CD96은 4명의 환자 유래의 CD3+CD8+ T 세포(공여체 HN306 제외), CD3+CD4+ 통상적인 T 세포, CD3-CD56+ NK 세포 및 CD3+CD4+FoxP3+ 세포에서 현저하게 발현되었다(도 9B). 주목할만한 것은, 시험된 다양한 림프구 서브셋에서 CD96의 존재는 이들 종양의 HPV 상태와 관련이 없다는 것이다.
3.5 CD96은 인간 혈소판에서 발현되지 않으며, 항-CD96 후보는 혈소판에 결합하지 않는다
CD226은 혈소판에서 발현되는 것으로 기술되었으며, 이는 치료적 항-CD226 항체의 사용에 대한 잠재적 부작용을 나타낼 수 있다. 대조적으로, CD96은 인간 혈소판에서 발현되지 않는다(Wang, 1992). 혈소판상의 19개 항-CD96 후보의 잠재적 결합을 FCM에 의해 추가로 시험하였다. 벤치마크 NK92.39 및 628211뿐만 아니라 후보들 중 어느 것도 인간 혈소판에 대한 유의한 결합을 보이지 않는다(데이터는 나타내지 않음). 따라서, CD226과 달리, CD96은 인간 혈소판에서 발현되지 않으며, 본 발명의 항-CD96 후보는 혈소판에서 반응성을 나타내지 않는다. 결론적으로, CD226과 반대로, 혈소판 응집을 유발할 위험을 유발하지 않으면서 CD96에 대한 작용성 항체가 안전하게 개발될 수 있다.
3.6 hCD96 절단 단백질에서 FCM에 의한 에피토프 맵핑
hCD96v2의 절단된 세포외 도메인(짧은 이소형 v2)을 암호화하는 발현 벡터는 hCD96v2의 3개의 기재된 면역글로불린 도메인 내에서 항-hCD96 후보에 의해 인식되는 에피토프를 맵핑하도록 구성되었다. 구축된 모든 발현 벡터는 다양한 작제물의 형질감염 효율을 모니터링하기 위해 신호 펩타이드, D4 도메인, 막관통, hCD6의 세포내 쇄 및 HA-태그를 함유하였다. 발현 벡터는 D1, D2, D3 또는 이들 3개의 도메인(D1D2, D1D3, D2D3)의 조합만을 함유하였다. 항-hCD96 항체를 CHO-S 세포에서 일시적으로 발현된 절단된 CD96 단백질을 인식하는 능력에 대해 FCM에 의해 시험하였다. 도 10에 도시된 바와 같이, 시험된 모든 항-CD96 항체는 CHO-S 세포에서 일시적으로 발현된 천연 hCD96v2 단백질을 인식하였다. 시험된 19개의 항체 중 9개의 후보 및 2개의 참조 항-CD96 항체(NK92.39 및 628211)는 D1 및 D2 도메인의 융합 작제물을 발현하는 CHO-S 세포에 대한 특이적 결합을 나타냈으며(BL006-4-20, BL006-4-31, BL006-19-134, BL006-19-190, BL006-19-21, BL006-19-352, BL006-19-363, BL006-19-370 및 BL006-19-55), 2개의 후보는 D1 및 D2 도메인의 융합 및 D2 및 D3 도메인의 융합의 작제물을 발현하는 CHO-S 세포에 대한 특이적 결합을 나타냈으며(BL006-19-14 및 BL006-19-29), 1개의 후보는 D1 도메인, D1 및 D2 도메인의 융합 및 D1 및 D3 도메인의 융합의 작제물을 발현하는 CHO-S 세포에 대한 특이적 결합을 나타내고(BL006-19-183), 2개의 후보는 D3 도메인, D1 및 D3 도메인의 융합 및 D2 및 D3 도메인의 융합의 작제물을 발현하는 CHO-S 세포에 대한 특이적 결합을 나타내고(BL006-4-52 및 BL006-2-19), 5개의 후보는 시험된 모든 작제물을 발현하는 CHO-S 세포에 대한 특이적 결합을 나타냈다(BL006-4-61, BL006-8-3, BL006-9-1, BL006-9-15 및 BL006-11-5). 이들 결과는 (i) 레퍼런스 항-CD96 항체 및 후보 BL006-4-20, BL006-4-31, BL006-19-134, BL006-19-190, BL006-19-21, BL006-19-352, BL006-19-363, BL006-19-370 및 BL006-19-55는 D1 및 D2 도메인 내에 포함된 에피토프를 인식하고; (ii) 후보 BL006-19-14 및 BL006-19-29는 D2 도메인 내에 포함된 에피토프를 인식하고; (iii) 후보 BL006-19-183은 D1 도메인 내에 포함된 에피토프를 인식하고; (iv) 후보 BL006-4-52 및 BL006-2-19는 D3 도메인 내에 포함된 에피토프를 인식하고; (v) 반면 후보 BL006-4-61, BL006-8-3, BL006-9-1, BL006-9-15 및 BL006-11-5는 D4 도메인 내에서 에피토프를 인식할 가능성이 있음을 나타낸다.
3.7 최상의 마우스 항-CD96 후보의 친화도
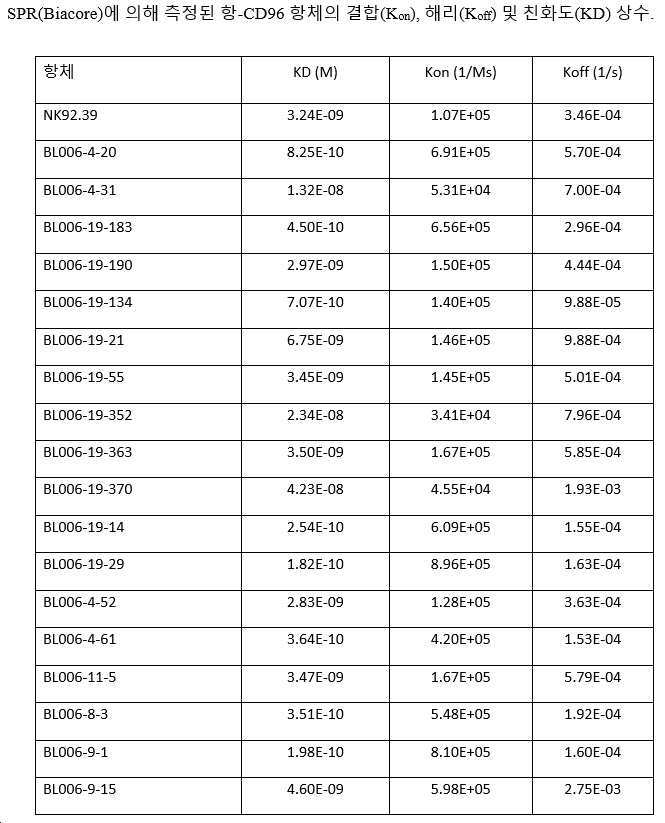
항-hCD96 후보의 운동 상수(Kon 및 Koff) 및 해리 상수(KD)는 칩 상에 코팅된 항-huFc 항체에 의해 포획된 hCD96-hFc 단백질(자체 제조)을 사용하여 Biacore 상의 표면 플라즈몬 공명(SPR)에 의해 측정하고, 벤치마크와 비교했다. 결과는 하기 표 8에 제시되어 있다. Biacore에 의한 이 설정에서 결합을 나타내지 않은 후보 BL006-2-19를 제외하고, 대부분의 후보는 0.18 내지 42.3 nM의 KD 값의 nM 범위의 친화도를 나타냈다. 10개의 후보는 NK92.39 클론보다 양호한 친화도를 가진다.
3.8 항-CD96 항체 후보는 인간 CD4+ 및 CD8+ T 세포 활성화 및 증식을 공동-자극할 수 있다
항-CD3 항체 OKT3으로 공동-자극된 인간 T 세포에 대해 항-CD96 후보의 생물학적 활성을 추가로 평가하였다. PBMC를 건강한 개체로부터 단리하고, CFSE 형광 염료로 염색하고, 상이한 농도의 가용성 항-CD3 항체(OKT3, mIgG2a)의 존재 또는 부재하에 가용성 또는 플레이트-결합된 항-CD96 항체(mIgG2a-D265A 형식)와 배양하였다. 세포를 상이한 배양 시간에 수집하고, CD25 활성화 마커의 발현 및 CFSE 형광 희석을 각각 측정함으로써 CD4+ 및 CD8+ T 세포의 활성화 및 증식을 FCM에 의해 분석하였다.
하기의 도 11 내지 17에 보고된 결과는 고정화된 항-hCD96 후보가 최적 이하의 농도의 항-CD3 항체로 CD4+ 및 CD8+ T 세포 둘 다를 공동-활성화시킬 수 있음을 보여준다. 활성화된 세포의 백분율은 CD25의 증가된 발현(도 11 및 12) 및 분열 세포의 수와 연관된 CFSE 염색의 희석(도 13 내지 17)에 의해 측정하였다.
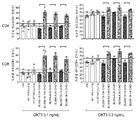
T 세포에서 증가된 CD25 발현의 예는 0.1 또는 1 ng/mL OKT3와 조합된 후보 BL006-4-20에 대해 도 11에 도시되어 있다. 항-CD96 후보의 공동-자극 활성은 이미 매우 낮은 농도의 항-CD3 항체(0.1 ng/mL)에 대해 관찰되었다. mIgG2a 이소타입 대조군으로 얻은 결과와 비교하여, 첫 번째 실험 세트(BL006-4-20, BL006-19-183, BL006-19-134, BL006-4-52, BL006-11-5, BL006-8-3)에서 테스트된 6개의 후보 모두가 항-CD3으로 공동-자극된 CD4+ 및 CD8+ T 세포 모두에서 CD25의 발현을 증가시키는 것으로 밝혀졌다(도 12), BL006-4-20, BL006-19-183 및 BL006-19-134 후보는 항-CD3으로 공동-자극된 CD4+ 및 CD8+ T 세포 모두에서 CD25의 발현을 증가시키는 데 가장 강력했다(도 12). 2개의 추가 건강한 공여체로부터 정제된 PBMC를 사용하여 유사한 결과를 얻었다(도시되지 않음).
두 번째 실험 세트에서, 8개의 항-CD96 후보를 2개의 공여체(# 2 및 #3)로부터의 T 세포를 공동-자극하는 능력에 대해 시험하고, 벤치마크 항-hCD96 항체(NK92.39)와 비교하였다. FCM에 의해 측정된 CFSE 희석의 예는 0.1, 1 또는 10 ng/mL OKT3와 함께 후보 BL006-4-20에 대해 도 13에 도시되어 있다. 대조군과 비교하여(OKT3 단독 또는 + mIgG2a 이소타입 대조군), 시험된 8개의 후보 모두는 저농도의 OKT3으로 최적-이하로 활성화된 분열 CD4+ 및 CD8+ T 세포의 백분율을 증가시키는 것으로 밝혀졌다(도 14). 후보 BL006-19-183, BL006-19-134 및 BL006-4-20은 이러한 조건에서 분열 세포의 %를 증가시키는 데 가장 강력했다(도 14). 4, 5 또는 6배로 분열된 세포의 상대적 비율이 대조군과 비교하여 후보 BL-006-4-20의 존재하에 더 높았지만(도 13), (FCM에 의해 결정된 바와 같이) CFSE 희석을 나타내는 CD4+ 및 CD8+ T 세포의 %로 보고된 공동-자극 수준은 더 높은 OKT3 농도에 대해 뚜렷하지 않았다 (도시되지 않음). 중요하게도, 벤치마크 항체 NK92.39는 두 공여체 모두에서 시험된 OKT3 농도 중 어느 것으로도 공동-자극 활성을 나타내지 않았다(도 14). NK92.39가 아닌 항-CD96 후보에 의한 CD4+ 및 CD8+ T 세포 증식의 공동-자극이 3일 내지 5일째에 시험된 모든 시점에 대해 관찰되었다(도 15).
BL006-2-19를 제외한 모든 후보 항체를 OKT3-최적-이하로 활성화된 T 세포 증식에 대한 공동-자극 효과에 대해 병행하여 시험하였다. 형광 강도의 CFSE 중간값(중간 패널)뿐만 아니라 분열 세포(CFSE 염색 감소를 나타내는 세포, 상부 패널)의 백분율이 보고되었다. 하부 패널은 최적 이하의 자극에 사용된 OKT3의 농도에 따라 각 항체 후보에 대한 분열 세포의 백분율을 보고한다. 결과는 후보 BL006-4-20, BL006-4-31, BL006-19-183, BL006-19-134, BL006-19-14, BL006-19-190, BL006-19-21, BL006-19-55, BL006-19-370, BL006-19-363, BL006-19-352, BL006-19-29 및 BL006-4-52가 CD4+(도 16) 및 CD8+ T 세포(도 17) 증식을 공동-자극함을 나타내어, 후보 BL006-19-183, BL006-19-134 및 BL006-4-20에 대한 이전 결과를 확인시켜준다. 대조군(OKT3 단독 또는 이소타입 대조군의 존재하에서)에서 관찰된 결과에 따르면, 공동-자극 효과는 CD4+ T 세포의 경우 0.1 및 0.3 ng/mL에서(도 16), 그리고 CD8+ T 세포의 경우 0.3 ng/mL에서(도 17) 가장 잘 관찰되었다. 벤치마크 항-hCD96 항체 NK92.39에 의해서는 유의한 공동-자극이 다시 관찰되지 않았다. 주목할 점은, 항-hCD96 후보는 항-CD28 항체로 얻은 것과 유사한 수준으로 T 세포 증식을 공동-자극하는 것으로 밝혀졌다.
관찰된 현상에서 PBMC에 존재하는 보조 세포의 역할을 평가하기 위해, 3개의 후보의 공동-자극 특성을 정제된 CD4+ 및 CD8+ T 세포상에서 시험하였다(자기 세포 분류). CFSE-염색된 정제된 T 세포를 2 μL/mL의 사량체 CD3/CD28 복합체(ImmunoCult™, Stemcell)로 최적-이하로 활성화시켰다. 이들 T 세포에서 항-CD96 후보(10 ㎍/mL)의 활성을 5일 후에 FCM으로 평가했다. 결과는 후보 BL006-4-20, BL006-19-134 및 BL006-4-31이 정제된 CD4+ 및 CD8+ T 세포를 공동-자극했음을 나타낸다(도 18). 최적-이하의 예비-활성화의 부재하에 이들 3개의 후보에 의한 CD96 결합은 정제된 T 세포 증식을 유발할 수 없었다. 이 관찰은 인간 림프구에서 CD96 신호전달의 공동-자극 특성을 확인한다.
CD96 및 CD226(DNAM-1)은 모두 CD155와 상호작용할 수 있다. CD155에 의한 CD226의 관여는 NK 및 T 세포에 대한 자극 신호를 제공한다. CD155는 단핵구 및 활성화된 T 세포 상에서 발현된다(데이터는 나타내지 않음). 후보 항체에 의한 CD96 마스킹이 CD226과 CD155 사이의 상호작용을 선호할 가능성을 배제하기 위해, PBMC의 공동-자극 실험을 CD226에 대한 차단 항체 또는 CD155에 대한 차단 항체의 존재하에 수행하였다. 공동-자극 4일 후 CFSE 형광의 중간값으로 보고된 결과는 후보 BL006-19-134가 CD226과 CD155 사이의 상호작용과 무관하게 CD4+ 및 CD8+ T 세포 증식을 유도할 수 있음을 나타낸다(도 19).
후보 BL006-4-20, BL006-19-134 및 BL006-4-31에 의해 유도된 공동-자극에 대한 최적의 타이밍을 평가하였다. 정제된 CD4+ 또는 CD8+ T 세포를 2 μL/mL의 사량체 CD3/CD28 복합체(ImmunoCult™, Stemcell)로 최적-이하로 활성화시켰다. 항-CD96 항체에 의한 공동-자극은 동시에 도입되거나 24시간, 48시간 또는 72시간 지연되었다. 결과는 CD96 관여가 최대 공동-자극 효과를 얻기 위해 최적-이하의 CD3 자극과 함께 제공되어야 함을 나타낸다(도 20).
인간 IgG1 골격(CHG1)에서 후보 BL006-4-20, BL006-19-134 및 BL006-19-14에 대해 키메라 항체가 생성되었다. 각 후보에 대해, 가용성 항-CD96 후보 항체의 활성에 대한 Fc 수용체와의 상호작용의 영향을 평가하기 위해 Fc 침묵 버전(CHS1)을 또한 작제하였다. 최적-이하로 활성화된(0.1 및 0.3 ng/mL OKT3) CD4+ 및 CD8+ T 세포의 증식에 대한 이들 다양한 가용성 작제물(1 ㎍/mL)의 효과를 전술한 바와 같이 평가하였다. 결과는 공동-자극 실험에서 가용성 인간 IgG1로서 도입된 후보 BL006-4-20, BL006-19-134 및 BL006-19-14가 CD4+ 및 CD8+ T 세포를 공동-자극할 수 있음을 나타낸다(도 21). 이 효과는 가장 낮은 농도의 OKT3으로 더욱 두드러진다. 각 후보의 Fc-침묵 버전은 CD4+ 및 CD8+ T 세포의 공동-자극을 유도할 수 없었으며, 이는 PBMC에서 보조 세포 상에 존재하는 인간 Fc 수용체에 의한 가교-결합이 관찰된 효과에 필수적이라는 것을 나타낸다.
고지에 입각한 동의를 얻은 2명의 환자로부터 수술적으로 제거된 두경부암으로부터 종양 침윤 림프구를 추출하였다. 이 작업은 영국 사우샘프턴 병원의 지역 윤리위원회의 승인을 받았다. 최적-이하로 활성화된(3 μL/mL ImmunoCult™) TIL의 증식에 대한 플레이트-결합된 BL006-19-134CHG1 후보(5 μg/mL)의 효과는 3[H] 티미딘의 16-시간 긴 펄스에 의해 5-일 공동-배양 후 평가하였다. 결과는 공동-자극 실험에서 플레이트-결합된 인간 IgG1로서 도입된 후보 BL006-19-134CHG1이 두경부 종양 세포로부터 생체외 TIL을 공동-자극 및 재-활성화시킬 수 있음을 나타낸다(도 22). 효과는 TIL의 기저 최적-이하의 증식이 낮은 두 실험 중 하나에서 더욱 두드러졌다. 이 경우에, BL006-19-134CHG1-유도된 공동-자극은 최적 용량의 ImmunoCult™(25 μL/mL)의 것과 비슷한 증식을 유도했다.
본 명세서에 인용 또는 참조된 모든 문헌들("본 명세서에 인용된 문헌") 및 본 명세서에 인용된 문헌들에 인용 또는 참조된 모든 문헌들은 본 명세서 또는 본 명세서에서 참고로 포함된 임의의 문헌에 언급된 임의의 제품에 대한 임의의 제조업체 지침, 설명, 제품 사양 및 제품 시트와 함께, 본 명세서에서 참고로 포함되며, 본 발명의 실시에 사용될 수 있다. 보다 구체적으로, 모든 참고문헌들은 각각의 개별 문서가 구체적이고 개별적으로 참고로 포함되는 것으로 표시된 것과 동일한 정도로 참고로 포함된다.
본 발명 및 이의 이점이 상세히 설명되었지만, 첨부된 청구범위에 정의된 바와 같이 본 발명의 사상 및 범위를 벗어나지 않으면서 다양한 변형, 치환 및 변경이 본 명세서에서 이루어질 수 있음을 이해해야 한다.
참고문헌
Batard P., Szollosi J., Luescher I., Cerottini J.-C., MacDonald R. and Romero P., 유세포 측정법에 의해 분석된 형광 공명 에너지 전달을 위한 피코에리트린 및 알로피코시아닌의 사용: 이점 및 한계, Cytometry, 2002, 48:97-105
Blake S.J., Dougall W.C., Miles J.J., Teng M.W.L, and Smyth M.J., 분자 경로: 암 면역요법을 위한 CD96 및 TIGIT 표적화, Clinical Cancer Research, 2016, 22(21): 5183-8
Chan C.J., Martinet L., Gilfillan S., Souza-Fonseca-Guimaraes F., Chow M., Town L., Ritchie D.S., Colonna M., Andrews D.M. & Smyth M.J., 수용체 CD96 및 CD226은 자연 살해 세포 기능의 조절에서 서로 반대된다, Nature Immunology, 2014, 15(5):431-8
Chothia C., Novotny J., Bruccoleri R. and Karplus M., 면역글로불린 분자내 도메인 결합. 가변 도메인의 패킹, J. Mol. Biol., 1985, 186:651-63
Evan G.I., Lewis G.K., Ramsay G., et al., 인간 c-myc 원-종양유전자 생성물에 특이적인 단클론성 항체의 단리, Mol. Cell. Biol., 1985, 5(12):3610-6
Fuchs A., Cella M., Giurisato E., Shaw A.S., and Colonna M. 최첨단: CD96(촉각)은 폴리오바이러스 수용체(CD155)와 상호작용하여 NK 세포-표적 세포 부착을 촉진한다, Journal of Immunology, 2004, 172:3994-8
Fuhrman C.A., Yeh W.-I., Seay H.R., Saikumar Lakshmi P., Chopra G., Zhang L., Perry D.J., McClymont S.A., Yadav M., Lopez M.-C., Baker H.V., Zhang Y., Li Y., Whitley M., von Schack D., Atkinson M.A., Bluestone J.A. and Brusko T.M., 수용체 TIGIT 및 CD226을 발현하는 인간 조절 T 세포의 다양한 표현형, The Journal of Immunology, 2015, 195:145-55
Gao J., Chen K., Miao Z., Ren G., Chen X., Gambhir S.S. and Cheng Z., HER2 발현 세포 및 종양 영상화를 위한 아피바디-기반 나노프로브, Biomaterials, 2011, 32(8):2141-8
Johnston R.J., Comps-Agrar L., Hackney J., Yu X., Huseni M., Yang Y., Park S., Javinal V., Chiu H., Irving B., Eaton D.L. and Grogan J.L., 면역 수용체 TIGIT는 항종양 및 항바이러스 CD8+ T 세포 효과기 기능을 조절한다, Cancer Cell, 2014, 26:923-37
Jokerst J.V., Miao Z., Zavaleta C., Cheng Z., and Gambhir S.S., 표피 성장 인자 수용체의 라만 분자 이미징을 위한 아피바디-기능화된 금-실리카 나노입자, Small., 2011, 7(5):625-33
Kennedy J., Vicari A. P., Saylor V., Zurawski, S.M., Copeland N.G., Gilber D.J., Jenkins N.A. and Zlotnik A., NKT 세포의 분자 분석: 클래스-I 제한 T 세포-관련 분자(CRTAM)의 동정, Journal of Leukocyte Biology, 2000, 67:725-34
Novotny J. and Haber E., 항원 결합의 구조적 불변량: 면역글로불린 VL-VH 및 VL-VL 도메인 이량체의 비교., Proc. Natl. Acad. Sci. U.S.A., 1985, 82:4592-96
Lozano E., Dominguez-Villar M., Kuchroo V. and Hafler D.A., TIGIT/CD226 축은 인간 T 세포 기능을 조절한다, The Journal of Immunology, 2012, 188:3869-75
Melero et al. 암 치료 백신: 임상 시험 개요. Nature reviews. Clinical oncology. 11(9):509-524(2014)
Paborsky L.R., Fendly B.M., Fisher K.L., et al., 항원 생성을 위한 조직 인자의 포유류 세포 일시적 발현, Protein Engineering, 1990, 3(6):547-53
Page D.B., Postow M.A., Callahan M.K., Allison J.P. and Wolchok, J., 항체를 이용한 암의 면역조절, Annual Review of Medicine, 2014, 65:185-202
Sanmamed et al. CD137, OX40, GITR, CD27, CD28 및 ICOS에 대한 암 면역요법의 공동-자극 효능제. Seminars in oncology. 42(4):640-655(2015)
Saxena A. and Wu, D., 치료 Fc 공학의 발전 - IgG-관련 효과기 기능 및 혈청 반감기 조절, Frontiers in Immunology, 2016, 12(7):580
Seth S., Maier M.K., Qiu Q., Ravens I., Kremmer E., Forster, R. and Bernhardt G., 뮤린 팬 T 세포 마커 CD96은 CD155 및 넥틴-1에 대한 부착 수용체이다, Biochemical and Biophysical Research Communications, 2007, 364:959-65
Shibuya A., Campbell D., Hannum C., Yssel H., Franz-Bacon K., McClanahan T., Kitamura T., Nicholl J., Sutherland G.R., Lanier L.L. and Phillips J.H., T 림프구의 세포분해 기능에 관여하는 새로운 부착 분자인, DNAM-1, Immunity, 1996, 4:573-81
Stanietsky N., Simic H., Arapovic J., Toporik A., Levy O., Novik A., Levine Z., Beiman M., Dassa L., Achdout H., Stern-Ginossar N., Tsukerman P., Jonjic S. and Mandelboim O., PVR 및 PVRL2와 TIGIT의 상호작용은 인간 NK 세포 세포독성을 억제한다, PNAS, 2009, 106(42):17858-63
Thakor A.S., Luong R., Paulmurugan R., et al. et al., 마우스에서 라만-활성 실리카-금 나노입자의 운명 및 독성, Sci. Transl. Med., 2011, 3(79):79ra33
Wang P., O'Farrell S., Clayberger C., Krensky A. 촉각의 식별 및 분자 클로닝. Ig 유전자 수퍼패밀리의 구성원인 새로운 인간 T 세포 활성 항원, Journal of Immunology, 1992, 148:2600-8
Yu X., Harden K., Gonzales L.C., Francesco M., Chiang E., Irving B., Tom I., Ivejla S., Refino C.J., Clark H., Eaton D. and Grogan J.L., 표면 단백질 TIGIT는 성숙한 면역조절 수지상 세포의 생성을 촉진함으로써 T 세포 활성화를 억제한다, Nature Immunology, 2009, 10(1):48-57