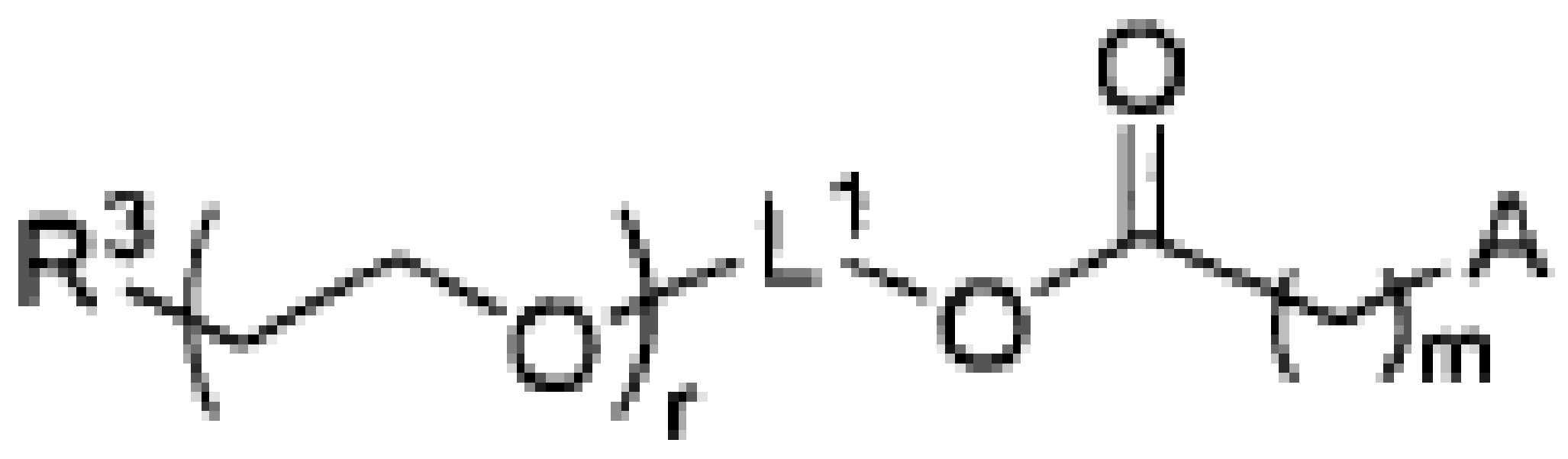I. 정의
본 명세서에서 달리 정의되지 않는 한, 본 출원과 관련하여 사용된 과학 및 기술 용어는 본 발명이 속하는 기술 분야의 숙련가에 의해 일반적으로 이해되는 의미를 가질 것이다. 본 발명은 본원에 기술된 특정 방법론, 프로토콜 및 시약 등에 제한되지 않으며, 다양할 수 있음을 이해해야 한다. 본원에 사용된 용어는 단지 특정한 구현예를 설명하기 위한 것으로, 본 발명의 범위를 한정하려는 의도가 아니며, 본 발명의 범위는 청구범위에 의해서만 정의된다. 면역학 및 분자 생물학에서 일반적인 용어의 정의는 하기에서 찾을 수 있다: The Merck Manual of Diagnosis and Therapy, 19th Edition, published by Merck Sharp & Dohme Corp., 2011 (ISBN 978-0-911910-19-3); Robert S. Porter et al. (eds.), Fields Virology, 6th Edition, published by Lippincott Williams & Wilkins, Philadelphia, PA, USA (2013), Knipe, D.M. and Howley, P.M. (ed.), The Encyclopedia of Molecular Cell Biology and Molecular Medicine, published by Blackwell Science Ltd., 1999-2012 (ISBN 9783527600908); and Robert A. Meyers (ed.), Molecular Biology and Biotechnology: a Comprehensive Desk Reference, published by VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8); Immunology by Werner Luttmann, published by Elsevier, 2006; Janeway's Immunobiology, Kenneth Murphy, Allan Mowat, Casey Weaver (eds.), Taylor & Francis Limited, 2014 (ISBN 0815345305, 9780815345305); Lewin's Genes XI, published by Jones & Bartlett Publishers, 2014 (ISBN-1449659055); Michael Richard Green and Joseph Sambrook, Molecular Cloning: A Laboratory Manual, 4th ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., USA (2012) (ISBN 1936113414); Davis et al., Basic Methods in Molecular Biology, Elsevier Science Publishing, Inc., New York, USA (2012) (ISBN 044460149X); Laboratory Methods in Enzymology: DNA, Jon Lorsch (ed.) Elsevier, 2013 (ISBN 0124199542); Current Protocols in Molecular Biology (CPMB), Frederick M. Ausubel (ed.), John Wiley and Sons, 2014 (ISBN047150338X, 9780471503385), Current Protocols in Protein Science (CPPS), John E. Coligan (ed.), John Wiley and Sons, Inc., 2005; and Current Protocols in Immunology (CPI) (John E. Coligan, ADA M Kruisbeek, David H Margulies, Ethan M Shevach, Warren Strobe, (eds.) John Wiley and Sons, Inc., 2003 (ISBN 0471142735, 9780471142737) (이들의 내용은 그 전문이 본원에 참고로 포함됨).
본원에 사용된 용어 "이종 뉴클레오티드 서열" 및 "이식유전자"은 상호 교환적으로 사용되며, 본 명세서에 개시된 바와 같은 ceDNA 벡터에 통합되고 ceDNA 벡터에 의해 전달되고 발현될 수 있는 관심 핵산 (캡시드 폴리펩티드를 인코딩하는 핵산 이외)을 지칭한다.
본원에 사용된 용어 "발현 카세트" 및 "전사 카세트"는 상호 교환적으로 사용되며, 하나 이상의 프로모터 또는 이식유전자의 전사를 지시하기에 충분한 다른 조절 서열에 작동 가능하게 연결되지만, 캡시드-인코딩 서열, 다른 벡터 서열 또는 역 말단 반복 영역을 포함하지 않는 이식유전자를 포함하는 핵산의 선형 스트레치를 지칭한다. 발현 카세트는 하나 이상의 시스-작용 서열 (예를 들어, 프로모터, 인핸서 또는 리프레서), 하나 이상의 인트론 및 하나 이상의 전사 후 조절 요소를 추가로 포함할 수 있다.
본원에서 상호 교환적으로 사용되는 용어 "폴리뉴클레오티드" 및 "핵산"은 리보뉴클레오티드 또는 데옥시리보뉴클레오티드인, 임의의 길이의 뉴클레오티드의 폴리머 형태를 지칭한다. 따라서, 이 용어는 단일, 이중 또는 다중 가닥 DNA 또는 RNA, 게놈 DNA, cDNA, DNA-RNA 하이브리드, 또는 퓨린 및 피리미딘 염기 또는 다른 천연, 화학적 또는 생화학적으로 변형된, 비-천연 또는 유도체화된 뉴클레오티드 염기를 포함하는 폴리머를 포함한다. "올리고뉴클레오티드"는 일반적으로 단일 가닥 또는 이중 가닥 DNA의 약 5개 내지 약 100개의 뉴클레오티드의 폴리뉴클레오티드를 지칭한다. 그러나, 본 개시의 목적상, 올리고뉴클레오티드의 길이에 대한 상한은 없다. 올리고뉴클레오티드는 또한 "올리고머" 또는 "올리고"로 공지되어 있으며, 유전자로부터 단리되거나, 당 업계에 공지된 방법에 의해 화학적으로 합성될 수 있다. 용어 "폴리뉴클레오티드" 및 "핵산"은, 기술된 구현예에 적용 가능한, 단일 가닥 (예컨대 센스 또는 안티센스) 및 이중 가닥 폴리뉴클레오티드를 포함하는 것으로 이해되어야 한다.
본원에 사용된 용어 "핵산 작제물"은 단일 가닥 또는 이중 가닥의 핵산 분자를 말하며, 이는 천연 발생 유전자로부터 단리되거나 또는 달리 자연적으로 존재하지 않거나 합성되는 방식으로 핵산 세그먼트를 함유하도록 변형된다. 핵산 작제물이라는 용어는 핵산 작제물이 본 개시내용의 코딩 서열의 발현에 필요한 조절 서열을 함유할 때 "발현 카세트"라는 용어와 동의어이다. "발현 카세트"는 프로모터에 작동 가능하게 연결된 DNA 코딩 서열을 포함한다.
"하이브리드화 가능" 또는 "상보적" 또는 "실질적으로 상보적"이란 핵산 (예를 들어, RNA)이 적절한 시험관내 및/또는 생체내 조건의 온도 및 용액 이온 강도 하에서 서열 특이적 역평행 방식으로 (즉, 핵산은 상보적인 핵산에 특이적으로 결합함) 또 다른 핵산에 비-공유적으로 결합하거나, 즉 왓슨-크릭(Watson-Crick) 염기쌍 및/또는 G/U 염기쌍을 형성하거나, "어닐링"하거나 "하이브리드화"될 수 있는 뉴클레오티드 서열을 포함함을 의미한다. 당 업계에 공지된 바와 같이, 표준 왓슨-크릭 염기쌍은 티미딘 (T)과 쌍을 이루는 아데닌 (A), 우라실 (U)과 쌍을 이루는 아데닌 (A) 및 시토신 (C)과 쌍을 이루는 구아닌 (G)을 포함한다. 또한, 2개의 RNA 분자 (예를 들어, dsRNA) 간의 하이브리드화, 우라실 (U)과 구아닌 (G)의 염기쌍에 대해 당 업계에 공지되어 있다. 예를 들어, G/U 염기 짝짓기는 mRNA 내 코돈과 tRNA 항-코돈 염기 짝짓기의 맥락에서 유전자 코드의 축퇴 (즉 , 중복)를 부분적으로 담당한다. 본 개시의 맥락에서, 대상 DNA-표적화 RNA 분자의 단백질-결합 세그먼트 (dsRNA 듀플렉스)의 구아닌 (G)은 우라실 (U)과 상보적인 것으로 간주되고, 그 반대도 마찬가지이다. 이와 같이, G/U 염기쌍이 주어진 뉴클레오티드 위치 대상 DNA-표적화 RNA 분자의 단백질-결합 세그먼트 (dsRNA 듀플렉스)에서 만들어질 수 있는 경우, 위치는 비-상보적인 것으로 간주되지 않고, 대신에 상보적인 것으로 간주된다.
용어 "펩티드", "폴리펩티드" 및 "단백질"은 본원에서 상호 교환적으로 사용되며, 코딩된 및 코딩되지 않은 아미노산, 화학적으로 또는 생화학적으로 변형되거나 유도체화된 아미노산, 및 변형된 펩티드 골격을 갖는 폴리펩티드를 포함할 수 있는, 임의의 길이의 아미노산의 폴리머 형태를 지칭한다.
특정 RNA 또는 단백질 유전자 산물을 "인코딩"하는 DNA 서열은 특정 RNA 및/또는 단백질로 전사되는 DNA 핵산 서열이다. DNA 폴리뉴클레오티드는 단백질로 번역되는 RNA (mRNA)를 인코딩할 수 있거나, DNA 폴리뉴클레오티드는 단백질로 번역되지 않는 RNA (예를 들어, tRNA, rRNA, 또는 DNA-표적화 RNA; "비-코딩" "RNA 또는 "ncRNA"라고도 함)를 인코딩 할 수 있다.
본원에 사용된 용어 "유전자 편집 분자"는 단백질 또는 단백질을 인코딩하는 핵산 중 하나 이상을 지칭하며, 여기서 단백질은 트랜스포사제, 뉴클레아제, 인테그라제, 가이드 RNA (gRNA), 가이드 DNA, 리보핵단백질 (RNP) 또는 활성화제 RNA를 포함하는 군으로부터 선택된다. 뉴클레아제 유전자 편집 분자는 뉴클레아제 활성을 갖는 단백질이며, 비제한적인 예로서, CRISPR 단백질 (Cas), CRISPR 관련 단백질 9 (Cas9); 타입 IIS 제한 효소; 전사 활성화제-유사 이펙터 뉴클레아제 (TALEN); 및 아연 핑거 뉴클레아제 (ZFN), 메가뉴클레아제, 조작된 부위-특이적 뉴클레아제 또는 CRISPRi 또는 CRISPRa 시스템을 위한 불활성화된 CAS가 포함된다. 유전자 편집 분자는 또한 DNA-결합 도메인 및 뉴클레아제를 포함할 수 있다. 특정 구현예에서, 유전자 편집 분자는 DNA-결합 도메인 및 뉴클레아제를 포함한다. 특정 구현예에서, DNA-결합 도메인은 가이드 RNA를 포함한다. 특정 구현예에서, DNA-결합 도메인은 TALEN의 DNA-결합 도메인을 포함한다. 특정 구현예에서, 적어도 하나의 유전자 편집 분자는 하나 이상의 전이성 요소(들)를 포함한다. 특정 구현예에서, 하나 이상의 전이성 요소(들)는 원형 DNA를 포함한다. 특정 구현예에서, 하나 이상의 전이성 요소(들)는 플라스미드 벡터 또는 미니서클 DNA 벡터를 포함한다. 특정 구현예에서, DNA-결합 도메인은 아연-핑거 뉴클레아제의 DNA-결합 도메인을 포함한다. 특정 구현예에서, 적어도 하나의 유전자 편집 분자는 하나 이상의 전이성 요소(들)를 포함한다. 특정 구현예에서, 하나 이상의 전이성 요소(들)는 선형 DNA를 포함한다. 트랜스포손을 인코딩하는 선형 재조합 및 비-천연 발생 DNA 서열은 시험관내에서 생성될 수 있다. 본 개시내용의 선형 재조합 및 비-천연 발생 DNA 서열은 원형 DNA의 제한 분해 산물일 수 있다. 특정 구현예에서, 원형 DNA는 플라스미드 벡터 또는 미니서클 DNA 벡터이다. 본 개시내용의 선형 재조합 및 비-천연 발생 DNA 서열은 중합효소 연쇄 반응 (PCR)의 산물일 수 있다. 본 개시내용의 선형 재조합 및 비-천연 발생 DNA 서열은 이중 가닥 doggybone™ DNA 서열일 수 있다. 본 개시내용의 doggybone™ DNA 서열은 항원 발현 카세트, 컴프리신 항원, 프로모터, 폴리-A 테일 및 텔로머 말단만을 인코딩하는 효소 공정에 의해 생성될 수 있다.
본원에서 사용되는 용어 "유전자 편집 기능"은 기능의 손실 또는 이득을 갖는 게놈의 특정 부위에서 DNA의 삽입, 결실 또는 대체를 의미한다. 특정 부위에서 DNA의 삽입, 결실 또는 대체는 예를 들어 상동성 지정 복구 (HDR) 또는 비-동종 말단 결합 (NHEJ), 또는 단일 염기 변화 편집에 의해 달성될 수 있다. 일부 구현예에서, 공여체 주형은, 예를 들어, HDR에 사용되어, 공여체 주형 내의 원하는 서열이 상동성 재조합 사건에 의해 게놈에 삽입되도록 한다. 일 구현예에서, "공여체 주형" 또는 "복구 주형"은 숙주 게놈으로 도입될 핵산 서열에서 원하는 돌연변이 또는 삽입을 포함하는 공여체 서열의 양쪽에 플랭킹된 2개의 상동성 아암 (예를 들어, 5' 상동성 아암 및 3' 상동성 아암)을 포함한다. 5' 및 3' 상동성 아암은 엔도뉴클레아제 매개 절단 부위에서 표적 유전자의 게놈 서열과 실질적으로 상동성이다. 3' 상동성 아암은 일반적으로 엔도뉴클레아제가 절단되거나 (예를 들어, 이중 가닥 DNA 절단), 또는 일부 구현예에서 DNA를 닉킹하는 프로토스페이서 인접 모티프 (protospacer adjacent motif; PAM) 부위의 바로 하류에 있다.
본원에 사용된 용어 "유전자 편집 시스템"은 세포에서 게놈 편집을 수행하는 데 필요한 최소 성분을 지칭한다. 예를 들어, 아연 핑거 뉴클레아제 또는 TALEN 시스템은 표적 유전자의 서열에 상보적인 핵산에 융합된 엔도뉴클레아제의 발현만을 필요로 할 수 있는 반면, CRISPR/Cas 유전자 편집 시스템의 경우 최소 성분은 예를 들어, Cas 엔도뉴클레아제 및 가이드 RNA를 필요로 할 수 있다. 유전자 편집 시스템은 원하는 경우 단일 ceDNA 벡터 또는 다수의 벡터에서 인코딩될 수 있다. 당업자는 유전자 편집 시스템에 필요한 성분(들)을 쉽게 이해할 것이다.
본원에 사용된 용어 "염기 편집 모이어티"는 서열 내 단일 뉴클레오티드를, 예를 들어 시토신/구아닌 뉴클레오티드 쌍 "G/C"를 아데닌 및 티민 "T"/우리딘 "U" 뉴클레오티드 쌍 (A/T,U)으로 (예를 들어, Shevidi et al. Dev Dyn 31 (2017) PMID:28857338; Kyoungmi et al. Nature Biotechnology 35:435-437 (2017) 참조, 이들 각각의 내용은 그 전문이 본원에 참고로 포함됨) 또는 아데닌/티민 "A/T" 뉴클레오티드 쌍을 구아닌/시토신 "G/C" 뉴클레오티드 쌍으로(예를 들어, Gaudelli et al. Nature (2017), in press doi:10.1038/nature24644 참조, 이의 내용은 그 전문이 본원에 참고로 포함됨) 변경할 수 있는 효소 또는 효소 시스템을 지칭한다.
본원에 사용된 용어 "게놈 세이프 하버 유전자" 또는 "세이프 하버 유전자"는 내인성 유전자 활성에 중대한 부정적인 영향 없이 또는 암의 촉진 없이 서열이 예측 가능한 방식으로 통합되고 기능할 수 있도록 (예를 들어, 관심 단백질을 발현하도록) 핵산 서열이 삽입될 수 있는 유전자 또는 유전자좌를 지칭한다. 일부 구현예에서, 세이프 하버 유전자는 또한 삽입된 핵산 서열이 비-세이프 하버 부위보다 효율적으로 그리고 더 높은 수준으로 발현될 수 있는 유전자좌 또는 유전자이다.
본원에 사용된 용어 "유전자 전달"은 외래 DNA가 유전자 요법 적용을 위해 숙주 세포로 전달되는 과정을 의미한다.
본원에 사용된 용어 "CRISPR"은 CRISPR-Cas9 게놈 편집 기술의 기초를 형성하는 박테리아 방어 시스템의 특징인, 클러스터된 규칙적으로 간격을 둔 짧은 회문 구조 반복(Clustered Regularly Interspaced Short Palindromic Repeats)을 의미한다.
본원에 사용된 용어 "아연 핑거"는 접힘을 안정화시키기 위해 하나 이상의 아연 이온의 배위를 특징으로 하는 작은 단백질 구조 모티프를 의미한다.
본원에 사용된 용어 "상동성 재조합"은 뉴클레오티드 서열이 2개의 유사하거나 동일한 DNA 분자 사이에서 교환되는 유형의 유전자 재조합을 의미한다. 상동성 재조합은 또한 DNA 서열의 새로운 조합을 생성한다. 이 새로운 DNA 조합은 유전자 변이를 나타낸다. 상동성 재조합은 또한 상이한 균주와 바이러스 종 사이에 유전 물질을 교환하기 위해 수평 유전자 전달에 사용된다.
본원에 사용된 용어 "말단 반복부" 또는 "TR"은 적어도 하나의 최소 요구된 복제 기원 및 회문 헤어핀 구조를 포함하는 영역을 포함하는 임의의 바이러스 말단 반복 또는 합성 서열을 포함한다. Rep-결합 서열 ("RBS") (RBE (Rep-결합 요소)로도 지칭됨) 및 말단 분해 부위 ("TRS")는 함께 "최소 요구된 복제 기원"을 구성하므로 TR은 적어도 하나의 RBS 및 적어도 하나의 TRS를 포함한다. 주어진 폴리뉴클레오티드 서열의 스트레치 내에서 서로의 역 보체인 TR은 전형적으로 각각 "역 말단 반복" 또는 "ITR"로 지칭된다. 바이러스와 관련하여, ITR은 복제, 바이러스 패키징, 통합 및 프로바이러스 구조를 중재한다. 본 발명에서 예기치 않게 발견된 바와 같이, 전장에 걸쳐 역 보체가 아닌 TR은 여전히 ITR의 전통적인 기능을 수행할 수 있으므로, ITR이라는 용어는 본원에서 ceDNA 벡터의 복제를 매개할 수 있는 ceDNA 게놈 또는 ceDNA 벡터 내의 TR을 지칭하는데 사용된다. 복잡한 ceDNA 벡터 구성에서 2개 이상의 ITR 또는 비대칭 ITR 쌍이 존재할 수 있음을 당업자는 이해할 것이다. ITR은 AAV ITR 또는 비-AAV ITR일 수 있거나, AAV ITR 또는 비-AAV ITR로부터 유도될 수 있다. 예를 들어, ITR은 파보바이러스 및 데펜도바이러스 (예를 들어, 개 파보바이러스, 소 파보바이러스, 마우스 파보바이러스, 돼지 파보바이러스, 인간 파보바이러스 B-19)를 포함하는 파보비리다에 패밀리로부터 유래될 수 있거나, SV40 복제 기원으로 사용되는 SV40 헤어핀을 ITR로 사용할 수 있으며, 이는 절단, 치환, 결실, 삽입 및/또는 첨가에 의해 추가로 변형될 수 있다. 파보비리다에 패밀리 바이러스는 척추동물을 감염시키는 파보비리나에(Parvovirinae), 및 무척추동물을 감염시키는 덴소비리나에(Densovirinae)의 2개의 하위패밀리로 구성된다. 데펜도파보바이러스는 인간, 영장류, 소, 개, 말 및 양 종을 포함하지만 이에 제한되지 않는 척추동물 숙주에서 복제할 수 있는 아데노-관련 바이러스 (AAV)의 바이러스 패밀리를 포함한다. 본원의 편의를 위해, ceDNA 벡터에서 발현 카세트의 5'에 (상류에) 위치한 ITR은 "5' ITR" 또는 "좌측 ITR"로 지칭되고, ceDNA 벡터에서 발현 카세트의 3'에 (하류에) 위치한 ITR은 "3' ITR" 또는 "우측 ITR"로 지칭된다.
"야생형 ITR" 또는 "WT-ITR"은, 예를 들어, Rep 결합 활성 및 Rep 닉킹 능력을 보유하는 AAV 또는 다른 데펜도바이러스에서 천연 발생 ITR 서열의 서열을 지칭한다. 임의의 AAV 혈청형으로부터의 WT-ITR의 뉴클레오티드 서열은 유전자 코드 또는 드리프트의 축퇴로 인해 정규 천연 발생 서열과 약간 다를 수 있으므로, 본원에 사용하기 위해 포함된 WT-ITR 서열은 생성 과정 중에 발생하는 천연 발생 변화 (예를 들어, 복제 오류)의 결과로서 WT-ITR 서열을 포함한다.
본원에 사용된 용어 "실질적으로 대칭인 WT-ITR" 또는 "실질적으로 대칭인 WT-ITR 쌍"은 둘 다 전체 길이에 걸쳐 역 보체 서열을 갖는 야생형 ITR인 단일 ceDNA 게놈 또는 ceDNA 벡터 내의 한 쌍의 WT-ITR을 지칭한다. 예를 들어, ITR은 변화가 서열의 특성 및 전체 3차원 구조에 영향을 미치지 않는 한, 정규 천연 발생 서열로부터 벗어난 하나 이상의 뉴클레오티드를 갖는 경우에도 야생형 서열로 간주될 수 있다. 일부 측면에서, 변형 뉴클레오티드는 보존적 서열 변화를 나타낸다. 하나의 비제한적인 예로서, 정규 서열과 적어도 95%, 96%, 97%, 98% 또는 99%의 서열 동일성을 갖고 (예를 들어, 디폴트 설정에서 BLAST를 사용하여 측정됨), 또한 3D 구조가 기하학적 공간에서 동일한 형상이 되도록 다른 WT-ITR에 대해 대칭 3차원 공간 구성을 갖는 서열. 실질적으로 대칭인 WT-ITR은 3D 공간에서 동일한 A, C-C' 및 B-B' 루프를 갖는다. 실질적으로 대칭인 WT-ITR은 그것이 적절한 Rep 단백질과 쌍을 이루는 작동 가능한 Rep 결합 부위 (RBE 또는 RBE') 및 말단 분해 부위 (trs)를 갖는 것으로 결정함으로써 WT로서 기능적으로 확인될 수 있다. 허용 조건 하에서 이식유전자 발현을 포함하여, 다른 기능을 선택적으로 시험할 수 있다.
본원에 사용된 "변형된 ITR" 또는 "모드-ITR" 또는 "돌연변이 ITR"의 문구는 본원에서 상호 교환적으로 사용되며, 동일한 혈청형으로부터의 WT-ITR과 비교하여 적어도 하나 이상의 뉴클레오티드 내의 돌연변이를 갖는 ITR을 지칭한다. 돌연변이는 ITR에서 A, C, C', B, B' 영역 중 하나 이상에서의 변화를 초래할 수 있고, 동일한 혈청형의 WT-ITR의 3D 공간 구성과 비교하여 3차원 공간 구성 (즉, 기하학적 공간에서의 3D 구조)의 변화를 초래할 수 있다.
본원에서 사용되는 용어 "비대칭 ITR"은 "비대칭 ITR 쌍"으로도 지칭되고, 전체 길이에 걸쳐 역 보체가 아닌 단일 ceDNA 게놈 또는 ceDNA 벡터 내의 한 쌍의 ITR을 지칭한다. 하나의 비제한적인 예로서, 비대칭 ITR 쌍은 3D 구조가 기하학적 공간에서 상이한 형상이 되도록 동족 ITR에 대해 대칭인 3차원 공간 구성을 갖지 않는다. 다르게 말하면, 비대칭 ITR 쌍은 전체적인 기하학적 구조가 다르며, 즉, 3D 공간에서 A, C-C' 및 B-B' 루프의 구성이 다르다 (예를 들어, 하나의 ITR은 동족 ITR과 비교하여 짧은 C-C' 아암 및/또는 짧은 B-B' 아암을 가질 수 있음). 두 ITR 사이의 서열 차이는 하나 이상의 뉴클레오티드 첨가, 결실, 절단 또는 점 돌연변이에 기인할 수 있다. 일 구현예에서, 비대칭 ITR 쌍의 하나의 ITR은 야생형 AAV ITR 서열일 수 있고, 다른 ITR은 본원에 정의된 변형된 ITR (예를 들어, 비-야생형 또는 합성 ITR 서열)일 수 있다. 또 다른 구현예에서, 비대칭 ITR 쌍의 모든 ITR은 야생형 AAV 서열이 아니며, 2개의 ITR은 기하학적 공간에서 상이한 형상 (즉, 상이한 전체 기하학적 구조)을 갖는 변형된 ITR이다. 일부 구현예에서, 비대칭 ITR 쌍의 하나의 모드-ITR은 짧은 C-C' 아암을 가질 수 있고 다른 ITR은 상이한 변형 (예를 들어, 단일 암 또는 짧은 B-B' 아암 등)을 가질 수 있으므로 동족 비대칭 모드-ITR과 비교하여 상이한 3차원 공간 구성을 갖는다.
본원에 사용된 용어 "대칭 ITR"은 야생형 데펜도바이러스 ITR 서열에 대해 돌연변이되거나 변형되고 전장에 걸쳐 역 보체인 단일 ceDNA 게놈 또는 ceDNA 벡터 내의 한 쌍의 ITR을 지칭한다. ITR은 모두 야생형 ITR AAV2 서열이 아니며 (즉, 돌연변이 ITR이라고도 하는 변형된 ITR임), 뉴클레오티드 첨가, 결실, 치환, 절단 또는 점 돌연변이로 인해 야생형 ITR과 서열상 차이가 있을 수 있다. 본원의 편의를 위해, ceDNA 벡터에서 발현 카세트의 5'에 (상류에) 위치한 ITR은 "5' ITR" 또는 "좌측 ITR"로 지칭되고, ceDNA 벡터에서 발현 카세트의 3'에 (하류에) 위치한 ITR은 "3' ITR" 또는 "우측 ITR"로 지칭된다.
본원에 사용된 용어 "실질적으로 대칭인 변형된-ITR" 또는 "실질적으로 대칭인 모드-ITR 쌍"은 둘 다 전체 길이에 걸쳐 역 보체 서열을 갖는 단일 ceDNA 게놈 또는 ceDNA 벡터 내의 한 쌍의 변형된-ITR을 지칭한다. 예를 들어, 변형된 ITR은 변화가 특성 및 전체 형상에 영향을 미치지 않는 한, 역 보체 서열로부터 벗어난 일부 뉴클레오티드 서열을 가지더라도 실질적으로 대칭인 것으로 간주될 수 있다. 하나의 비제한적인 예로서, 정규 서열과 적어도 85%, 90%, 95%, 96%, 97%, 98% 또는 99% 서열 동일성을 갖고 (디폴트 설정에서 BLAST를 사용하여 측정됨), 또한 3D 구조가 기하학적 공간에서 동일한 형상이 되도록 동족 변형된 ITR에 대해 대칭 3차원 공간 구성을 갖는 서열. 다르게 말하면, 실질적으로 대칭인 변형된-ITR 쌍은 3D 공간에서 구성된 동일한 A, C-C' 및 B-B' 루프를 갖는다. 일부 구현예에서, 모드-ITR 쌍으로부터의 ITR은 상이한 역 보체 뉴클레오티드 서열을 가질 수 있지만 여전히 동일한 대칭 3차원 공간 구성을 가질 수 있으며 - 즉, 두 ITR은 모두 동일한 전체 3D 형상을 초래하는 돌연변이를 갖는다. 예를 들어, 모드-ITR 쌍에서 하나의 ITR (예를 들어, 5' ITR)은 하나의 혈청형으로부터 유래될 수 있고, 다른 ITR (예를 들어, 3' ITR)은 상이한 혈청형으로부터 유래될 수 있지만, 둘 모두 동일한 상응하는 돌연변이를 가질 수 있으므로 (예를 들어, 5'ITR이 C 영역에서 결실을 갖는 경우, 상이한 혈청형으로부터의 동족 변형된 3'ITR은 C '영역 내 상응하는 위치에서 결실을 가짐) 변형된 ITR 쌍은 동일한 대칭 3차원 공간 구성을 갖는다. 이러일 구현예에서, 변형된 ITR 쌍의 각각의 ITR은 AAV2 및 AAV6의 조합과 같은 다른 혈청형 (예를 들어 AAV1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 및 12)으로부터 유래할 수 있으며, 하나의 ITR에서의 변형은 다른 혈청형으로부터의 동족 ITR의 상응하는 위치에 반영된다. 일 구현예에서, 실질적으로 대칭인 변형된 ITR 쌍은 ITR 사이의 뉴클레오티드 서열의 차이가 특성 또는 전체 형상에 영향을 미치지 않고 3D 공간에서 실질적으로 동일한 형상을 갖는 한 변형된 ITR (모드-ITR)의 쌍을 지칭한다. 비제한적인 예로서, BLAST(기본 지역 정렬 검색 도구(Basic Local Alignment Search Tool)) 또는 디폴트 설정의 BLASTN과 같이 당 업계에 잘 알려진 표준 수단에 의해 결정된 정규 모드-ITR과 적어도 95%, 96%, 97%, 98% 또는 99% 서열 동일성을 갖고, 또한 3D 구조가 기하학적 공간에서 동일한 형상이 되도록 대칭 3차원 공간 구성을 갖는, 모드-ITR. 실질적으로 대칭인 모드-ITR 쌍은 3D 공간에서 동일한 A, C-C' 및 B-B' 루프를 가지며, 예를 들어, 실질적으로 대칭인 모드-ITR 쌍의 변형된 ITR이 C-C' 아암의 결실을 갖는다면, 동족 모드-ITR은 상응하는 C-C' 루프의 결실을 가지며 또한 동족 모드-ITR의 기하학적 공간에서 동일한 형태로 남은 A 및 B-B' 루프의 유사한 3D 구조를 갖는다.
용어 "플랭킹"은 또 다른 핵산 서열에 대한 하나의 핵산 서열의 상대적 위치를 지칭한다. 일반적으로, 서열 ABC에서, B는 A 및 C에 의해 플랭킹된다. 배열 AxBxC에 대해서도 동일하다. 따라서, 플랭킹 서열은 플랭킹된 서열에 선행하거나 후속하지만 플랭킹된 서열에 근접하거나 또는 바로 인접할 필요는 없다. 일 구현예에서, 용어 플랭킹은 선형 듀플렉스 ceDNA 벡터의 각 말단에서의 말단 반복부를 지칭한다.
본원에 사용된 용어 "ceDNA 게놈"은 적어도 하나의 역 말단 반복 영역을 추가로 포함하는 발현 카세트를 지칭한다. ceDNA 게놈은 하나 이상의 스페이서 영역을 추가로 포함할 수 있다. 일부 구현예에서, ceDNA 게놈은 DNA의 분자간 듀플렉스 폴리뉴클레오티드로서 플라스미드 또는 바이러스 게놈에 통합된다.
본원에 사용된 용어 "ceDNA 스페이서 영역"은 ceDNA 벡터 또는 ceDNA 게놈에서 기능적 요소를 분리하는 개재 서열을 지칭한다. 일부 구현예에서, ceDNA 스페이서 영역은 최적의 기능성을 위해 2개의 기능성 요소를 원하는 거리로 유지한다. 일부 구현예에서, ceDNA 스페이서 영역은 예를 들어, 플라스미드 또는 바쿨로바이러스 내에서 ceDNA 게놈의 유전자 안정성을 제공하거나 추가한다. 일부 구현예에서, ceDNA 스페이서 영역은 클로닝 부위 등을 위한 편리한 위치를 제공함으로써 ceDNA 게놈의 준비된 유전자 조작을 용이하게 한다. 예를 들어, 특정 측면에서, 몇몇 제한 엔도뉴클레아제 부위를 함유하는 올리고뉴클레오티드 "폴리링커", 또는 공지된 단백질 (예를 들어, 전사 인자) 결합 부위를 갖지 않도록 설계된 비-오픈 리딩 프레임 서열은, 예를 들어, 말단 분해 부위와 상류 전사 조절 요소 사이에 6mer, 12mer, 18mer, 24mer, 48mer, 86mer, 176mer 등을 삽입하여 시스 - 작용 인자를 분리하도록 ceDNA 게놈에 위치할 수 있다. 유사하게, 스페이서는 폴리아데닐화 신호 서열과 3'-말단 분해 부위 사이에 포함될 수 있다.
본원에 사용된 용어 "Rep 결합 부위, "Rep 결합 요소, "RBE" 및 "RBS"는 상호 교환적으로 사용되며, Rep 단백질에 의해 결합될 때 Rep 단백질이 RBS를 포함하는 서열에서 이의 부위-특이적 엔도뉴클레아제 활성을 수행할 수 있도록 하는 Rep 단백질 (예를 들어, AAV Rep 78 또는 AAV Rep 68)에 대한 결합 부위를 지칭한다. RBS 서열 및 이의 역 보체는 함께 단일 RBS를 형성한다. RBS 서열은 당 업계에 공지되어 있으며, 예를 들어, AAV2에서 확인된 RBS 서열인, 5'-GCGCGCTCGCTCGCTC-3' (서열 번호: 531)를 포함한다. 다른 공지된 AAV RBS 서열 및 다른 자연적으로 알려진 또는 합성 RBS 서열을 포함하는, 임의의 공지된 RBS 서열이 본 발명의 구현예에서 사용될 수 있다. 이론에 구속되지 않고, Rep 단백질의 뉴클레아제 도메인은 듀플렉스 뉴클레오티드 서열 GCTC에 결합하고, 따라서 공지된 2개의 AAV Rep 단백질은 듀플렉스 올리고뉴클레오티드, 5'-(GCGC)(GCTC)(GCTC)(GCTC)-3' (서열 번호: 531)에 직접 결합하고 안정적으로 조립된다고 생각된다. 또한, 가용성 응집된 컨포머 (즉, 정의되지 않은 수의 상호 연관된 Rep 단백질)는 해리되어 Rep 결합 부위를 함유하는 올리고뉴클레오티드에 결합한다. 각각의 Rep 단백질은 각 가닥에서 질소 염기 및 포스포디에스테르 골격 둘 다와 상호 작용한다. 질소 염기와의 상호 작용은 서열 특이성을 제공하는 반면, 포스포디에스테르 골격과의 상호 작용은 서열에 특이적이지 않거나 덜 특이적이고, 단백질-DNA 복합체를 안정화시킨다.
본원에 사용된 용어 "말단 분해 부위" 및 "TRS"는 본원에서 상호 교환적으로 사용되며, Rep가 DNA 중합 효소, 예를 들어, DNA pol 델타 또는 DNA pol 엡실론을 통해 DNA 확장을 위한 기질로 작용하는 3' OH를 생성하는 5' 티미딘과 티로신-포스포디에스테르 결합을 형성하여 영역을 지칭한다. 대안적으로, Rep-티미딘 복합체는 조정된 결찰 반응에 참여할 수 있다. 일부 구현예에서, TRS는 비-염기쌍을 이루는 티미딘을 최소로 포함한다. 일부 구현예에서, TRS의 닉킹 효율은 RBS로부터 동일한 분자 내에서의 거리에 의해 적어도 부분적으로 제어될 수 있다. 수용체 기질이 상보적 ITR인 경우, 생성된 생성물은 분자내 듀플렉스이다. TRS 서열은 당 업계에 공지되어 있으며, 예를 들어, AAV2에서 확인된 헥사뉴클레오티드 서열인, 5'-GGTTGA-3' (서열 번호: 45)를 포함한다. 다른 공지된 AAV TRS 서열 및 다른 자연적으로 알려진 또는 합성 TRS 서열 예컨대 AGTT (서열 번호: 46), GGTTGG (서열 번호: 47), AGTTGG (서열 번호: 48), AGTTGA (서열 번호: 49), 및 RRTTRR (서열 번호: 50)과 같은 다른 모티프를 포함하는, 임의의 공지된 TRS 서열이 본 발명의 구현예에서 사용될 수 있다.
본원에 사용된 용어 "ceDNA-플라스미드"는 분자간 듀플렉스로서 ceDNA 게놈을 포함하는 플라스미드를 지칭한다.
본원에 사용된 용어 "ceDNA-박미드"는 이. 콜라이에서 플라스미드로서 증식할 수 있는 분자간 듀플렉스로서 ceDNA 게놈을 포함하는 감염성 바쿨로바이러스 게놈을 지칭하며, 따라서 바쿨로바이러스에 대한 셔틀 벡터로서 작용할 수 있다.
본원에 사용된 용어 "ceDNA- 바쿨로바이러스"는 바쿨로바이러스 게놈 내의 분자간 듀플렉스로서 ceDNA 게놈을 포함하는 바쿨로바이러스를 지칭한다.
본원에 사용된 용어 "ceDNA-바쿨로바이러스 감염된 곤충 세포" 및 "ceDNA-BIIC"는 상호 교환적으로 사용되며, ceDNA-바쿨로바이러스로 감염된 무척추동물 숙주 세포 (곤충 세포 (예를 들어, Sf9 세포)를 포함하지만 이에 제한되지 않음)를 지칭한다.
본원에 사용된 용어 "폐쇄 말단 DNA 벡터", "ceDNA 벡터" 및 "ceDNA"는 상호 교환적으로 사용되고 적어도 하나의 공유적으로 폐쇄된 말단을 갖는 비-바이러스 캡시드 비함유 DNA 벡터 (즉, 분자내 듀플렉스)를 지칭한다. 일부 구현예에서, ceDNA는 2개의 공유적으로 폐쇄된 말단을 포함한다.
본원에 정의된 바와 같이, "리포터"는 검출 가능한 판독을 제공하는데 사용될 수 있는 단백질을 지칭한다. 리포터는 일반적으로 형광, 색 또는 발광과 같은 측정 가능한 신호를 생성한다. 리포터 단백질 코딩 서열은 세포 또는 유기체에서의 존재가 쉽게 관찰되는 단백질을 인코딩한다. 예를 들어, 형광 단백질은 특정 파장의 빛으로 여기될 때 세포가 형광을 일으키고, 루시퍼라제는 세포가 빛을 생성하는 반응을 촉매하게 하며, β-갈락토시다제와 같은 효소는 기질을 착색된 생성물로 전환시킨다. 실험 또는 진단 목적에 유용한 예시적인 리포터 폴리펩티드는 β-락타마제, β-갈락토시다제 (LacZ), 알칼리 포스파타제 (AP), 티미딘 키나제 (TK), 녹색 형광 단백질 (GFP) 및 기타 형광 단백질, 클로람페니콜 아세틸트랜스퍼라제 (CAT), 루시퍼라제 및 당 업계에 잘 알려진 다른 것들을 포함하지만, 이에 제한되지는 않는다.
본원에 사용된 용어 "이펙터 단백질"은, 예를 들어, 리포터 폴리펩티드로서 또는 보다 적절하게는, 세포를 사멸시키는 폴리펩티드, 예를 들어 독소, 또는 세포를 선택된 제제로의 사멸 또는 이의 결핍에 민감하게 하는 제제로서 검출 가능한 판독을 제공하는 폴리펩티드를 지칭한다. 이펙터 단백질은 숙주 세포의 DNA 및/또는 RNA를 직접 표적화하거나 손상시키는 임의의 단백질 또는 펩티드를 포함한다. 예를 들어, 이펙터 단백질은 숙주 세포 DNA 서열 (게놈 또는 염색체외 요소에 관계없이)을 표적화하는 제한 엔도뉴클레아제, 세포 생존에 필요한 폴리펩티드 표적을 분해하는 프로테아제, DNA 자이레이스 억제제, 및 리보뉴클레아제-타입 독소를 포함할 수 있지만, 이에 제한되지는 않는다. 일부 구현예에서, 본원에 기재된 합성 생물학적 회로에 의해 제어되는 이펙터 단백질의 발현은 또 다른 합성 생물학적 회로의 인자로서 참여하여 생물학적 회로 시스템의 반응성의 범위 및 복잡성을 확장시킬 수 있다.
전사 조절제는 관심 유전자의 전사를 활성화시키거나 억제하는 전사 활성화제 및 억제제를 지칭한다. 프로모터는 특정 유전자의 전사를 개시하는 핵산의 영역이다. 전사 활성화제는 전형적으로 전사 프로모터 인근에 결합하고 전사를 직접 개시하기 위해 RNA 폴리머라제를 동원한다. 억제제는 전사 프로모터에 결합하고 RNA 폴리머라제에 의한 전사 개시를 입체적으로 방해한다. 다른 전사 조절제는 이들이 결합하는 위치 및 세포 및 환경 조건에 따라 활성화제 또는 억제제로서 작용할 수 있다. 전사 조절제 부류의 비제한적인 예는 호메오도메인 단백질, 아연-핑거 단백질, 윙드(winged)-헬릭스 (포크헤드) 단백질 및 류신-지퍼 단백질을 포함하지만, 이에 제한되지는 않는다.
본원에 사용된 "억제 단백질" 또는 "유도 단백질"은 조절 서열 요소에 결합하고 조절 서열 요소에 작동적으로 연결된 서열의 전사를 각각 억제하거나 활성화시키는 단백질이다. 본원에 기술된 바람직한 억제 및 유도 단백질은 적어도 하나의 투입 제제 또는 환경 투입의 존재 또는 부재에 민감하다. 본원에 기재된 바와 같은 바람직한 단백질은, 예를 들어, 분리 가능한 DNA-결합 및 투입 제제-결합 또는 반응성 요소 또는 도메인을 포함하는 형태의 모듈이다.
본원에 사용된 "담체"는 임의의 및 모든 용매, 분산 매질, 비히클, 코팅, 희석제, 항균제 및 항진균제, 등장제 및 흡수 지연제, 완충제, 담체 용액, 현탁액, 콜로이드 등을 포함한다. 약제학적 활성 물질을 위한 이러한 매질 및 제제의 사용은 당 업계에 잘 알려져 있다. 보충 활성 성분이 또한 조성물에 혼입될 수 있다. 문구 "약제학적으로 허용되는"은 숙주에 투여될 때 독성, 알레르기성 또는 유사한 유해 반응을 일으키지 않는 분자 엔티티 및 조성물을 지칭한다.
본원에 사용된 "입력 제제 반응 도메인"은 연결된 DNA 결합 융합 도메인이 그 조건 또는 입력의 존재에 반응하도록 하는 방식으로 조건 또는 입력 제제에 결합하거나 달리 반응하는 전사 인자의 도메인이다. 일 구현예에서, 조건 또는 투입의 존재는 전사 인자의 전사-조절 활성을 변형시키는, 투입 제제 반응 도메인 또는 그것이 융합된 단백질에서 형태적 변화를 초래한다.
"생체내"라는 용어는 유기체, 예컨대 다세포 동물에서 또는 내에서 발생하는 분석 또는 과정을 지칭한다. 본원에 기재된 일부 측면에서, 박테리아와 같은 단세포 유기체가 사용될 때 방법 또는 사용이 "생체내"에서 발생한다고 말할 수 있다. "생체외"라는 용어는 다세포 동물 또는 식물의 몸체 외부, 예를 들어, 외식편, 일차 세포 및 세포주를 비롯한 배양 세포, 형질전환 세포주, 및 추출된 조직 또는 특히 혈액 세포를 포함하는 세포의 외부인 온전한 막을 갖는 살아있는 세포를 사용하여 수행되는 방법 및 사용을 지칭한다. "시험관내"라는 용어는 세포 추출물과 같은 온전한 막을 갖는 세포의 존재를 필요로 하지 않는 분석 및 방법을 지칭하고, 비-세포 시스템, 예컨대 세포 추출물과 같이 세포 또는 세포 시스템을 포함하지 않는 배지에서 프로그램 가능한 합성 생물학적 회로의 도입을 지칭할 수 있다.
본원에 사용된 용어 "프로모터"는 단백질 또는 RNA를 인코딩하는 이종 표적 유전자일 수 있는, 핵산 서열의 전사를 유도함으로써 또 다른 핵산 서열의 발현을 조절하는 임의의 핵산 서열을 지칭한다. 프로모터는 구성적, 유도성, 억제성, 조직-특이적, 또는 이들의 임의의 조합일 수 있다. 프로모터는 핵산 서열의 나머지 부분의 개시 및 전사 속도를 제어하는 핵산 서열의 제어 영역이다. 프로모터는 또한 조절 단백질 및 분자가 결합할 수 있는 유전자 요소, 예컨대 RNA 폴리머라제 및 다른 전사 인자를 함유할 수 있다. 본원에 기재된 측면의 일부 구현예에서, 프로모터는 프로모터 자체의 발현을 조절하는 전사 인자의 발현을 구동할 수 있다. 프로모터 서열 내에서 전사 개시 부위 및 RNA 폴리머라제의 결합을 담당하는 단백질 결합 도메인이 발견될 것이다. 진핵생물 프로모터는 종종 "TATA" 박스 및 "CAT" 박스를 포함하지만, 항상 그런 것은 아니다. 유도성 프로모터를 포함하는 다양한 프로모터가 본원에 개시된 ceDNA 벡터에서 이식유전자의 발현을 유도하는데 사용될 수 있다. 프로모터 서열은 전사 개시 부위에 의해 이의 3' 말단에 결합될 수 있고, 배경 이상의 검출 가능한 수준으로 전사를 개시하는데 필요한 최소 수의 염기 또는 요소를 포함하도록 상류 (5' 방향)로 연장된다.
본원에 사용된 용어 "인핸서"는 핵산 서열의 전사 활성화를 증가시키기 위해 하나 이상의 단백질 (예를 들어, 활성화제 단백질 또는 전사 인자)에 결합하는 시스-작용 조절 서열 (예를 들어, 50-1,500 염기쌍)을 지칭한다. 인핸서는 이들이 조절하는 유전자 개시 부위의 상류 또는 유전자 개시 부위의 하류에서 최대 1,000,000개의 염기 파스(base pars)에 위치할 수 있다. 인핸서는 인트론 영역 내에 또는 관련되지 않은 유전자의 엑손 영역에 위치할 수 있다.
프로모터는 그것이 조절하는 핵산 서열의 발현 또는 전사를 유도한다고 할 수 있다. "작동 가능하게 연결된", "작동적으로 위치된", "작동적으로 연결된", "제어 하" 및 "전사적 제어 하"라는 문구는 프로모터가 핵산 서열과 관련하여 해당 서열의 전사 개시 및/또는 발현을 제어하도록 조절하는 올바른 기능적 위치 및/또는 배향에 있음을 나타낸다. 본원에 사용된 "역 프로모터"는 핵산 서열이 역 배향으로 되어, 코딩 가닥이 이제 비-코딩 가닥이 되거나 그 반대인 프로모터를 지칭한다. 역 프로모터 서열은 스위치의 상태를 조절하기 위해 다양한 구현예에서 사용될 수 있다. 또한, 다양일 구현예에서, 프로모터는 인핸서와 함께 사용될 수 있다.
프로모터는 주어진 유전자 또는 서열의 코딩 세그먼트 및/또는 엑손의 상류에 위치한 5' 비-코딩 서열을 단리함으로써 수득될 수 있는 바와 같이 유전자 또는 서열과 자연적으로 관련된 것일 수 있다. 이러한 프로모터는 "내인성"으로 지칭될 수 있다. 유사하게, 일부 구현예에서, 인핸서는 그 서열의 하류 또는 상류에 위치한 핵산 서열과 자연적으로 관련된 것일 수 있다.
일부 구현예에서, 코딩 핵산 세그먼트는 "재조합 프로모터" 또는 "이종 프로모터"의 제어 하에 위치되며, 이들 둘 모두는 천연 환경에서 작동 가능하게 연결되는 인코딩된 핵산 서열과 정상적으로 연관되지 않은 프로모터를 지칭한다. 재조합 또는 이종 인핸서는 천연 환경에서 주어진 핵산 서열과 정상적으로 관련되지 않은 인핸서를 지칭한다. 이러한 프로모터 또는 인핸서는 다른 유전자의 프로모터 또는 인핸서; 임의의 다른 원핵생물, 바이러스 또는 진핵생물 세포로부터 단리된 프로모터 또는 인핸서; 및 "천연 발생"이 아닌 합성 프로모터 또는 인핸서를 포함할 수 있으며, 즉, 상이한 전사 조절 영역의 상이한 요소, 및/또는 당 업계에 공지된 유전 공학 방법을 통해 발현을 변경시키는 돌연변이를 포함한다. 프로모터 및 인핸서의 핵산 서열을 합성적으로 생성하는 것에 더하여, 프로모터 서열은 본원에 개시된 합성 생물학적 회로 및 모듈과 관련하여, PCR을 포함하는 재조합 클로닝 및/또는 핵산 증폭 기술을 사용하여 생성될 수 있다 (예를 들어, 미국 특허 번호 4,683,202, 미국 특허 번호 5,928,906 참조, 각각 본원에 참고로 포함됨). 또한, 미토콘드리아, 엽록체 등과 같은 비-핵 소기관 내에서 서열의 전사 및/또는 발현을 지시하는 제어 서열도 사용될 수 있는 것으로 고려된다.
본원에 기재된 바와 같이, "유도성 프로모터"는 유도제 또는 유도 제제의 존재하, 이에 영향을 받거나 이에 의해 접촉될 때 전사 활성을 개시 또는 향상시키는 것을 특징으로 하는 것이다. 본원에 정의된 "유도제" 또는 "유도 제제"는 유도성 프로모터로부터 전사 활성을 유도하는데 활성이 있는 방식으로 투여되는 내인성, 또는 일반적으로 외인성 화합물 또는 단백질일 수 있다. 일부 구현예에서, 유도제 또는 유도 제제, 즉, 화학 물질, 화합물 또는 단백질 자체는 핵산 서열의 전사 또는 발현 결과일 수 있으며 (즉, 유도제는 또 다른 성분 또는 모듈에 의해 발현된 유도 단백질일 수 있음), 이는 그 자체로 제어 하에 있거나 유도성 프로모터일 수 있다. 일부 구현예에서, 유도성 프로모터는 특정 작용제, 예컨대 억제제의 부재 하에 유도된다. 유도성 프로모터의 예는 테트라사이클린, 메탈로티오닌, 엑디손, 포유동물 바이러스 (예를 들어, 아데노바이러스 후기 프로모터; 및 마우스 유방 종양 바이러스 긴 말단 반복부 (MMTV-LTR)) 및 다른 스테로이드-반응성 프로모터인, 라파마이신 반응성 프로모터 등을 포함하지만, 이에 제한되지는 않는다.
본원에서 상호 교환적으로 사용되는 용어 "DNA 조절 서열", "제어 요소" 및 "조절 요소"는 전사 및 번역 제어 서열, 예컨대 프로모터, 인핸서, 폴리아데닐화 신호, 종결자, 단백질 분해 신호 등을 지칭하고, 비-코딩 서열 (예를 들어, DNA-표적화 RNA) 또는 코딩 서열 (예를 들어, 부위 지향적 변형 폴리펩티드, 또는 Cas9/Csn1 폴리펩티드)의 전사를 제공 및/또는 조절하고/하거나 인코딩된 폴리펩티드의 번역을 조절한다.
"작동 가능하게 연결된"은 설명된 구성 요소들이 의도된 방식으로 기능할 수 있게 하는 관계에 있는 근접부위(juxtaposition)를 지칭한다. 예를 들어, 프로모터가 이의 전사 또는 발현에 영향을 미치는 경우, 프로모터는 코딩 서열에 작동 가능하게 연결된다. "발현 카세트"는 ceDNA 벡터에서 이식유전자의 전사를 지시하기에 충분한 프로모터 또는 다른 조절 서열에 작동 가능하게 연결된 외인성 DNA 서열을 포함한다. 적합한 프로모터는, 예를 들어, 조직 특이적 프로모터를 포함한다. 프로모터는 또한 AAV 기원일 수 있다.
본원에 사용된 용어 "대상체"는 예방적 치료를 포함하여, 본 발명에 따른 ceDNA 벡터를 사용한 치료가 제공되는 인간 또는 동물을 지칭한다. 일반적으로 동물은 영장류, 설치류, 가축 또는 게임 동물과 같은 척추동물이다. 영장류는 침팬지, 사이노몰구스 원숭이(cynomologous monkey), 거미 원숭이 및 마카크(macaque), 예를 들어 레소스(Rhesus)를 포함하지만 이에 제한되지 않는다. 설치류에는 마우스, 랫트, 우드척(woodchuck), 흰 족제비, 토끼 및 햄스터가 포함된다. 가축 및 게임 동물에는 소, 말, 돼지, 사슴, 들소, 버팔로, 고양이과 종, 예를 들어, 집고양이, 개과 종, 예를 들어, 개, 여우, 늑대, 조류 종, 예를 들어, 닭, 에뮤, 타조 및 어류, 예를 들어, 송어, 메기 및 연어가 포함되지만 이에 제한되지 않는다. 본원에 기재된 측면의 특정 구현예에서, 대상체는 포유동물, 예를 들어, 영장류 또는 인간이다. 대상체는 남성 또는 여성일 수 있다. 또한, 대상체는 유아 또는 소아일 수 있다. 일부 구현예에서, 대상체는 신생아 또는 태어나지 않은 대상체일 수 있으며, 예를 들어, 대상체는 자궁 내에 있다. 바람직하게는, 대상체는 포유동물이다. 포유동물은 인간, 비인간 영장류, 마우스, 랫트, 개, 고양이, 말 또는 소일 수 있지만, 이들 예에 제한되지는 않는다. 인간 이외의 포유동물이 질환 및 장애의 동물 모델을 나타내는 대상체로서 유리하게 사용될 수 있다. 또한, 본원에 기술된 방법 및 조성물은 길들여진 동물 및/또는 애완 동물을 위해 사용될 수 있다. 인간 대상체는 임의의 연령, 성별, 인종 또는 민족 그룹, 예를 들어, 코카서스(백인), 아시아인, 아프리카인, 흑인, 아프리카계 미국인, 아프리카계 유럽인, 히스패닉계, 중동인 등일 수 있다. 일부 구현예에서, 대상체는 임상 환경의 환자 또는 다른 대상체일 수 있다. 일부 구현예에서, 대상체는 이미 치료 중이다. 일부 구현예에서, 대상체는 배아, 태아, 신생아, 유아, 소아, 청소년 또는 성인이다. 일부 구현예에서, 대상체는 인간 태아, 인간 신생아, 인간 유아, 인간 소아, 인간 청소년 또는 인간 성인이다. 일부 구현예에서, 대상체는 동물 배아, 또는 비인간 배아 또는 비인간 영장류 배아이다. 일부 구현예에서, 대상체는 인간 배아이다.
본원에 사용된 용어 "숙주 세포"는 본 개시내용의 핵산 작제물 또는 ceDNA 발현 벡터에 의한 형질전환, 형질감염, 형질도입 등에 민감한 임의의 세포 유형을 포함한다. 비제한적인 예로서, 숙주 세포는 단리된 1차 세포, 다능성 줄기세포, CD34+ 세포), 유도된 다능성 줄기세포, 또는 임의의 다수의 불멸화 세포주 (예를 들어, HepG2 세포)일 수 있다. 대안적으로, 숙주 세포는 조직, 기관 또는 유기체 내의 동일계 또는 생체내 세포일 수 있다.
용어 "외인성"은 천연 공급원이 아닌 세포에 존재하는 물질을 지칭한다. 본원에서 사용될 때 용어 "외인성"은 인간의 손을 포함하는 과정에 의해 세포 또는 유기체와 같은 생물학적 시스템으로 도입된 핵산 (예를 들어, 폴리펩티드를 인코딩하는 핵산) 또는 폴리펩티드를 지칭할 수 있으며, 이는 정상적으로는 발견되지 않으며, 그러한 세포 또는 유기체 내로 핵산 또는 폴리펩티드를 도입하고자 하는 것이다. 대안적으로, "외인성"은 인간의 손을 포함하는 과정에 의해 세포 또는 유기체와 같은 생물학적 시스템으로 도입된 핵산 또는 폴리펩티드를 지칭할 수 있으며, 이는 비교적 적은 양으로 발견되고 세포 또는 유기체에서 핵산 또는 폴리펩티드의 양을 증가시키고자 하는 것, 예를 들어, 이소성 발현 또는 수준을 생성하고자 하는 것이다. 대조적으로, 용어 "내인성"은 생물학적 시스템 또는 세포에 자생하는 물질을 지칭한다.
용어 "서열 동일성"은 두 뉴클레오티드 서열 사이의 관련성을 지칭한다. 본 개시의 목적을 위해, 2개의 데옥시리보뉴클레오티드 서열 사이의 서열 동일성 정도는 EMBOSS 패키지 (EMBOSS: 유럽 분자 생물학 개방 소프트웨어 스위트, Rice et al., 2000, 상기)의 니들 프로그램, 바람직하게는 버전 3.0.0 이상에서 구현된 바와 같이 니들맨-운쉬 알고리즘 (Needleman and Wunsch, 1970, 상기)을 사용하여 결정된다. 사용되는 선택적 파라미터는 갭 오픈 페널티(gap open penalty) 10, 갭 연장 페널티(gap extension penalty) 0.5, 및 EDNAFULL (NCBI NUC4.4의 EMBOSS 버전) 치환 매트릭스이다. "가장 긴 동일성"으로 표시된 니들의 출력 (-nobrief 옵션을 사용하여 획득)은 백분율 동일성으로 사용되며 다음과 같이 계산된다: (동일한 데옥시리보뉴클레오티드.횟수.100)/(정렬의 길이-정렬에서의 총 갭의 수). 정렬의 길이는 바람직하게는 적어도 10개의 뉴클레오티드, 바람직하게는 적어도 25개의 뉴클레오티드, 보다 바람직하게는 적어도 50개의 뉴클레오티드, 가장 바람직하게는 적어도 100개의 뉴클레오티드이다.
본원에 사용된 용어 "상동성" 또는 "상동"은 서열을 정렬하고, 필요한 경우, 최대 퍼센트 서열 동일성을 달성하기 위해 갭을 도입한 후 표적 염색체 상의 상응하는 서열의 뉴클레오티드 잔기와 동일한 상동성 아암의 뉴클레오티드 잔기의 백분율로 정의된다. 뉴클레오티드 서열 상동성 백분율을 결정하기 위한 정렬은, 예를 들어, BLAST, BLAST-2, ALIGN, ClustalW2 또는 Megalign (DNASTAR) 소프트웨어와 같은 공개적으로 이용 가능한 컴퓨터 소프트웨어를 사용하여 당 업계의 기술 내에 있는 다양한 방식으로 달성될 수 있다. 당업자는 비교되는 서열의 전체 길이에 걸쳐 최대 정렬을 달성하는데 필요한 임의의 알고리즘을 포함하여, 서열을 정렬하기 위한 적절한 파라미터를 결정할 수 있다. 일부 구현예에서, 서열이 숙주 세포의 상응하는 천연 또는 편집되지 않은 핵산 서열 (예를 들어, 게놈 서열)과 적어도 70%, 적어도 75%, 적어도 80%, 적어도 85%, 적어도 90%, 적어도 91%, 적어도 92%, 적어도 93%, 적어도 94%, 적어도 95%, 적어도 96%, 적어도 97%, 적어도 98%, 적어도 99% 이상 동일한 경우, 예를 들어 복구 주형의 상동성 아암의 핵산 서열 (예를 들어, DNA 서열)은 "상동성"인 것으로 간주된다.
본원에 사용된 "상동성 아암"은 상동성 재조합을 통해 공여체 서열을 게놈에 표적화하는데 적합한 폴리뉴클레오티드를 지칭한다. 전형적으로, 2개의 상동성 아암은 공여체 서열의 측면에 위치하고, 여기서 각각의 상동성 아암은 통합 유전자좌의 상류 및 하류에 있는 게놈 서열을 포함한다.
본원에 사용된 "공여체 서열"은 숙주 세포 게놈에 삽입되거나 숙주 세포 게놈에 대한 복구 주형으로서 사용될 폴리뉴클레오티드를 지칭한다. 공여체 서열은 유전자 편집 동안 일어나는 것이 바람직한 변형을 포함할 수 있다. 혼입될 서열은 표적 서열에서 상동성 지정 복구를 통해 표적 핵산 분자 내로 도입될 수 있고, 이로써 표적 서열이 원래 표적 서열에서 공여체 서열에 의해 포함된 서열로 변경된다. 따라서, 공여체 서열에 의해 포함된 서열은, 표적 서열과 관련하여, 삽입, 결실, 인델, 점 돌연변이, 돌연변이의 복구 등일 수 있다. 공여체 서열은, 예를 들어, 단일 가닥 DNA 분자; 이중 가닥 DNA 분자; DNA/RNA 하이브리드 분자; 및 DNA/modRNA (변형된 RNA) 하이브리드 분자일 수 있다. 일 구현예에서, 공여체 서열은 상동성 아암과 이질적이다. 편집은 DNA 편집뿐만 아니라 RNA일 수 있다. 공여체 서열은 원하는 유전자 편집의 성질에 따라 숙주 세포 게놈에 대해 내생적이거나 또는 외인성일 수 있다.
본원에 사용된 용어 "이종"은 각각 고유 핵산 또는 단백질에서 발견되지 않는 뉴클레오티드 또는 폴리펩티드 서열을 의미한다. 예를 들어, 키메라 Cas9/Csn1 단백질에서, 천연 발생 박테리아 Cas9/Csn1 폴리펩티드 (또는 이의 변이체)의 RNA-결합 도메인은 이종 폴리펩티드 서열 (즉, Cas9/Csn1 이외의 단백질로부터의 폴리펩티드 서열 또는 또 다른 유기체로부터의 폴리펩티드 서열)에 융합될 수 있다. 이종 폴리펩티드 서열은 키메라 Cas9/Csn1 단백질에 의해 또한 나타날 활성 (예를 들어, 효소 활성)을 나타낼 수 있다 (예를 들어, 메틸트랜스퍼라제 활성, 아세틸트랜스퍼라제 활성, 키나제 활성, 유비퀴틴화 활성 등). 이종 핵산 서열은 (예를 들어, 유전 공학에 의해) 천연 발생 핵산 서열 (또는 이의 변이체)에 연결되어 키메라 폴리펩티드를 인코딩하는 키메라 뉴클레오티드 서열을 생성할 수 있다. 또 다른 예로서, 융합 변이체 Cas9 부위 지향적 폴리펩티드에서, 변이체 Cas9 부위 지향적 폴리펩티드는 이종 폴리펩티드 (즉, Cas9 이외의 폴리펩티드)에 융합될 수 있으며, 이는 융합 변이체 Cas9 부위 지향적 폴리펩티드에 의해 또한 나타날 활성을 나타낸다. 이종 핵산 서열은 (예를 들어, 유전 공학에 의해) 변이체 Cas9 부위 지향적 폴리펩티드에 연결되어 융합 변이체 Cas9 부위 지향적 폴리펩티드를 인코딩하는 뉴클레오티드 서열을 생성할 수 있다.
"벡터" 또는 "발현 벡터"는 또 다른 DNA 세그먼트, 즉 "삽입물"이 세포에 부착된 세그먼트의 복제를 야기하도록 부착될 수 있는 플라스미드, 박미드, 파지, 바이러스, 비리온 또는 코스미드와 같은 레플리콘이다. 벡터는 숙주 세포로의 전달 또는 상이한 숙주 세포 사이의 이동을 위해 설계된 핵산 작제물일 수 있다. 본원에 사용된 벡터는 최초 및/또는 최종 형태의 바이러스성 또는 비-바이러스성일 수 있지만, 본 개시의 목적상, "벡터"는 일반적으로 그 용어가 본원에 사용된 바와 같이 ceDNA 벡터를 지칭한다. 용어 "벡터"는 적절한 제어 요소와 관련될 때 복제될 수 있고 유전자 서열을 세포로 전달할 수 있는 임의의 유전자 요소를 포함한다. 일부 구현예에서, 벡터는 발현 벡터 또는 재조합 벡터일 수 있다.
본원에 사용된 용어 "발현 벡터"는 벡터 상의 전사 조절 서열에 연결된 서열로부터 RNA 또는 폴리펩티드의 발현을 지시하는 벡터를 지칭한다. 발현된 서열은 종종 세포에 대해 이종성일 수 있지만, 반드시 그런 것은 아니다. 발현 벡터는 추가 요소를 포함할 수 있으며, 예를 들어, 발현 벡터는 2개의 복제 시스템을 가질 수 있고, 따라서 2개의 유기체, 예를 들어 발현을 위한 인간 세포 및 클로닝 및 증폭을 위한 원핵생물 숙주에서 유지될 수 있도록 한다. 용어 "발현"은, 적용 가능한 경우, 예를 들어, 전사, 전사체 프로세싱, 번역 및 단백질 폴딩, 변형 및 프로세싱을 포함하지만, 이에 제한되지 않는, RNA 및 단백질 및 적절한 경우, 분비 단백질을 생산하는데 관여하는 세포 과정을 지칭한다. "발현 산물"은 유전자로부터 전사된 RNA, 및 유전자로부터 전사된 mRNA의 번역에 의해 수득된 폴리펩티드를 포함한다. 용어 "유전자"는 적절한 조절 서열에 작동 가능하게 연결된 경우 시험관내 또는 생체내에서 (DNA가) RNA로 전사되는 핵산 서열을 의미한다. 유전자는 코딩 영역의 앞뒤에 있는 영역, 예를 들어, 5' 비번역 (5'UTR) 또는 "리더" 서열 및 3' UTR 또는 "트레일러" 서열뿐만 아니라 개별 코딩 세그먼트 (엑손) 간의 개재 서열 (인트론)을 포함하거나 포함하지 않을 수 있다.
"재조합 벡터"는 이종 핵산 서열, 또는 생체내에서 발현될 수 있는 "이식유전자"을 포함하는 벡터를 의미한다. 본원에 기술된 벡터는, 일부 구현예에서, 다른 적합한 조성물 및 요법과 조합될 수 있음을 이해해야 한다. 일부 구현예에서, 벡터는 에피솜이다. 적합한 에피솜 벡터의 사용은 대상체에서 관심 뉴클레오티드를 높은 카피 수의 추가 염색체 DNA로 유지하여 염색체 통합의 잠재적 영향을 제거하는 수단을 제공한다.
본원에 사용된 용어 "교정", "게놈 편집" 및 "복원"은 전장 기능성 또는 부분적으로 전장 기능성 단백질 발현이 얻어지도록 절단된 단백질을 인코딩하거나 전혀 단백질을 코딩하지 않는 돌연변이 유전자를 변화시키는 것을 지칭한다. 돌연변이 유전자를 교정 또는 복원하는 것은 돌연변이를 갖는 유전자의 영역을 대체하거나 전체 돌연변이 유전자를 상동성 지정 복구 (HDR)와 같은 복구 메커니즘으로 돌연변이를 갖지 않는 유전자의 카피로 대체하는 것을 포함할 수 있다. 돌연변이 유전자를 교정 또는 복원하는 것은 또한 유전자에 이중 가닥 파괴물을 생성한 후 비-동종 말단 결합 (NHEJ)을 사용하여 복구함으로써 조기 정지 코돈, 이상 스플라이스 수용체 부위 또는 이상 스플라이스 공여체 부위를 유발하는 프레임시프트 돌연변이를 복구하는 것을 포함할 수 있다. NHEJ는 복구 동안, 적절한 판독 프레임을 복원하고 조기 정지 코돈을 제거할 수 있는 적어도 하나의 염기쌍을 추가 또는 결실시킬 수 있다. 돌연변이 유전자를 교정 또는 복원하는 것은 또한 비정상 스플라이스 수용체 부위 또는 스플라이스 공여체 서열을 파괴하는 것을 포함할 수 있다. 돌연변이 유전자를 교정 또는 복원하는 것은 또한 2개의 뉴클레아제 표적 부위 사이의 DNA를 제거하고 NHEJ에 의해 DNA 파괴를 복구함으로써 적절한 판독 프레임을 복원하기 위해 동일한 DNA 가닥 상에서 2개의 뉴클레아제의 동시 작용에 의해 비-필수 유전자 세그먼트를 결실시키는 것을 포함할 수 있다.
본원에 사용된 문구 "유전 질환"은 게놈에서 하나 이상의 이상, 특히 출생시 존재하는 상태에 의해 부분적으로 또는 전적으로, 직접적으로 또는 간접적으로 유발되는 질환을 지칭한다. 이상은 돌연변이, 삽입 또는 결실일 수 있다. 이상은 유전자의 코딩 서열 또는 이의 조절 서열에 영향을 줄 수 있다. 유전 질환은 DMD, 혈우병, 낭포성 섬유증, 헌팅턴 무도병, 가족성 고콜레스테롤혈증 (LDL 수용체 결함), 간모세포종, 윌슨병(Wilson's disease), 선천성 간성 포르피린증, 간 대사의 유전성 장애, 레쉬 니한 증후군(Lesch Nyhan syndrome), 겸상 적혈구 빈혈, 지중해빈혈, 색소피부건조증, 판코니 빈혈(Fanconi's anemia), 망막색소변성증, 모세혈관확장성 실조증, 블룸 증후군(Bloom's syndrome), 망막모세포종 및 테이-삭스병(Tay-Sachs disease)일 수 있지만, 이에 제한되지는 않는다.
본원에 사용된 문구 "비-동종 말단 결합 (NHEJ) 경로"는 상동성 주형을 필요로 하지 않고 파괴 말단을 직접 결찰함으로써 DNA에서 이중 가닥 파괴물을 복구하는 경로를 지칭한다. NHEJ에 의한 DNA 말단의 주형 독립적 재결찰은 DNA 파괴점에서 임의의 미세-삽입 및 미세-결실 (인델)을 도입하는, 확률적이며, 오류가 발생하기 쉬운 복구 과정이다. 이 방법은 표적화된 유전자 서열의 판독 프레임을 의도적으로 파괴, 결실 또는 변경시키기 위해 사용될 수 있다. NHEJ는 전형적으로 복구를 유도하기 위해 미세상동성(microhomology)이라고 하는 짧은 상동 DNA 서열을 사용한다. 이러한 미세상동성은 종종 이중 가닥 파괴물의 끝에 있는 단일 가닥 오버행(overhang)에 존재한다. 오버행이 완벽하게 호환되는 경우, NHEJ는 일반적으로 파괴물을 정확하게 복구하지만, 뉴클레오티드 손실로 이어지는 부정확한 복구도 발생할 수 있지만, 오버행이 호환되지 않을 때 훨씬 더 일반적이다. 본원에서 사용되는 "뉴클레아제 매개 NHEJ"는 cas9 또는 다른 뉴클레아제와 같은 뉴클레아제가 이중 가닥 DNA를 절단한 후에 개시되는 NHEJ를 지칭한다. CRISPR/CAS 시스템에서, NHEJ는 단일 가이드 RNA 서열을 사용하여 표적화될 수 있다.
본원에서 상호 교환적으로 사용되는 문구 "상동성 지정 복구" 또는 "HDR"은 상동성 DNA 조각이 핵에 존재할 때 이중 가닥 DNA 병변을 복구하기 위한 세포 내 메카니즘을 지칭한다. HDR은 공여체 DNA 주형을 사용하여 복구를 유도하고, 전체 유전자의 표적화된 첨가를 포함하여, 게놈에 대한 특정 서열 변화를 생성하는데 사용될 수 있다. 공여체 주형이 CRISPR/Cas9-기반 시스템과 같은 부위 특이적 뉴클레아제와 함께 제공된다면, 세포 기구는 상동성 재조합에 의해 파괴를 복구할 것이며, 이는 DNA 절단의 존재하에 10의 몇 승 정도(several orders of magnitude) 향상된다. 상동 DNA 조각이 없으면, 비-동종 말단 결합이 대신 일어날 수 있다. CRISPR/Cas 시스템에서 HDR에는 하나의 가이드 RNA 또는 두 개의 다른 가이드 RNA가 사용될 수 있다.
본원에서 상호 교환적으로 사용되는 문구 "반복 가변 이잔기(diresidue)" 또는 "RVD"는 TALE DNA 결합 도메인의 33-35개 아미노산을 포함하는, DNA 인식 모티프 ("RVD 모듈"로도 알려짐) 내의 한 쌍의 인접한 아미노산 잔기를 지칭한다. RVD는 RVD 모듈의 뉴클레오티드 특이성을 결정한다. RVD 모듈은 RVD 어레이를 생성하기 위해 조합될 수 있다. 본원에 사용된 "RVD 어레이 길이"는 TALEN에 의해 인식되는 TALEN 표적 영역, 즉, 결합 영역 내의 뉴클레오티드 서열의 길이에 상응하는 RVD 모듈의 수를 지칭한다.
본원에 사용된 용어 "부위-특이적 뉴클레아제" 또는 "서열 특이적 뉴클레아제"는 DNA 서열을 특이적으로 인식하고 절단할 수 있는 효소를 지칭한다. 부위-특이적 뉴클레아제는 조작될 수 있다. 조작된 부위-특이적 뉴클레아제의 예는 다양한 천연 및 비천연 Cas 효소를 사용하는 아연 핑거 뉴클레아제 (ZFN), TAL 이펙터 뉴클레아제 (TALEN) 및 CRISPR/Cas 기반 시스템을 포함한다.
본원에 사용된 용어 "포함하는" 또는 "포함하다"는 방법 또는 조성물에 필수적인 조성물, 방법 및 이의 각각의 성분(들)을 언급하는데 사용되지만, 필수적인지 여부에 관계없이, 특정되지 않은 요소의 포함에도 개방적이다.
본원에 사용된 용어 "본질적으로 구성되는"은 주어진 구현예에 필요한 요소들을 지칭한다. 상기 용어는 그 구현예의 기본 및 신규 또는 기능적 특성(들)에 실질적으로 영향을 미치지 않는 요소의 존재를 허용한다. "포함하는"의 사용은 제한이 아니라 포함을 나타낸다.
"구성되는"이라는 용어는 구현예의 설명에서 언급되지 않은 임의의 요소를 배제한, 본원에 기재된 조성물, 방법 및 이들의 각각의 구성 요소를 지칭한다.
본원에 사용된 용어 "본질적으로 구성되는"은 주어진 구현예에 필요한 요소들을 지칭한다. 상기 용어는 본 발명의 구현예의 기본 및 신규 또는 기능적 특성(들)에 실질적으로 영향을 미치지 않는 추가 요소의 존재를 허용한다.
본 명세서 및 첨부된 청구범위에 사용된 바와 같이, 단수 형태 "a", "an" 및 "the"는 문맥상 명백하게 달리 지시되지 않는 한 복수의 지시대상을 포함한다. 따라서, 예를 들어, "방법"에 대한 언급은 하나 이상의 방법, 및/또는 여기에 설명된 유형의 단계를 포함하고/하거나 본 개시내용 등을 읽을 때 당업자에게 명백할 것이다. 유사하게, 용어 "또는"은 문맥상 명백하게 달리 나타내지 않는 한 "및"을 포함하는 것으로 의도된다. 본 명세서에 기술된 것과 유사하거나 동등한 방법 및 재료가 본 개시의 실시 또는 시험에 사용될 수 있지만, 적합한 방법 및 재료가 후술된다. 약어 "예를 들어(e.g.)"는 라틴어 예컨대(Latin exempli gratia)로부터 유래되며, 비제한적인 예를 나타내기 위해 본원에서 사용된다. 따라서, 약어 "예를 들어(e.g.)"는 "예를 들어(for example)"라는 용어와 동의어이다.
작동 실시예 이외의, 또는 달리 지시된 경우에, 본원에 사용된 성분 또는 반응 조건의 양을 나타내는 모든 수는 모든 경우에 용어 "약"에 의해 수식되는 것으로 이해되어야 한다. 백분율과 관련하여 사용될 때 "약"이라는 용어는 ±1%를 의미할 수 있다. 이하의 실시예에 의해 본 발명을 더욱 상세하게 추가로 설명하지만, 본 발명의 범위가 이것에 한정되는 것은 아니다.
본원에 개시된 본 발명의 대안적인 요소 또는 구현예의 그룹핑은 제한으로서 해석되지 않아야 한다. 각각의 그룹 구성원은 개별적으로 또는 그룹의 다른 구성원 또는 본원에서 발견된 다른 요소와의 임의의 조합으로 언급되고 청구될 수 있다. 편의성 및/또는 특허성의 이유로 그룹의 하나 이상의 구성원이 그룹에 포함되거나 그룹에서 삭제될 수 있다. 그러한 포함 또는 삭제가 발생할 때, 본 명세서는 본원에서 수정된 그룹을 포함하는 것으로 간주되어 첨부된 청구범위에 사용된 모든 마쿠시 그룹의 서면 설명을 충족시킨다.
임의의 측면의 일부 구현예에서, 본원에 기재된 개시내용은 인간을 클로닝하는 과정, 인간의 생식선 유전자 동일성을 변형시키는 공정, 산업 또는 상업적 목적을 위한 인간 배아의 사용 또는 사람이나 동물에게 어떠한 실질적인 의학적 이익 없이 고통을 유발할 수 있는 동물의 유전적 정체성을 변경시키는 과정, 및 그러한 과정으로 얻어진 동물에 관한 것이 아니다.
다른 용어는 본 발명의 다양한 측면의 설명 내에서 본원에 정의된다.
문헌 참조, 발행된 특허, 공개된 특허 출원 및 동시 계류중인 특허 출원을 포함하고; 본원 전체에 걸쳐 인용된 모든 특허 및 기타 출판물은, 예를 들어, 본원에 설명된 기술과 관련하여 사용될 수 있는 그러한 공보에 기술된 방법론을 기술하고 개시할 목적으로 명백히 본원에 참고로 포함된다. 이들 간행물은 본 출원의 출원일 이전에 그들의 공개만을 위해 제공된다. 이와 관련하여 어떠한 것도 발명자가 선행 발명에 의해 또는 임의의 다른 이유로 그러한 개시를 앞당길 자격이 없다는 것을 인정하는 것으로 해석되어서는 안된다. 이 문서의 날짜 또는 내용에 대한 표현에 관한 모든 진술은 출원인이 이용할 수 있는 정보를 기반으로 하며 이 문서의 날짜 또는 내용의 정확성에 대한 어떠한 입장을 의미하지 않는다.
본 개시의 구현예의 설명은 완전하거나 본 개시내용을 개시된 정확한 형태로 제한하려는 것이 아니다. 본 개시의 특정 구현예 및 본 개시에 대한 예가 예시적인 목적으로 본 명세서에 기술되었지만, 관련 기술 분야의 숙련가들이 인식할 수 있는 바와 같이, 본 개시의 범위 내에서 다양한 등가의 변형들이 가능하다. 예를 들어, 방법 단계 또는 기능이 주어진 순서로 제시되지만, 대안적인 구현예가 다른 순서로 기능을 수행할 수 있거나, 기능이 실질적으로 동시에 수행될 수 있다. 본원에 제공된 개시내용의 교시는 다른 절차 또는 방법에 적절하게 적용될 수 있다. 본원에 기재된 다양한 구현예는 추가 구현예를 제공하기 위해 조합될 수 있다. 본 개시의 측면은, 필요하다면, 본 개시의 추가 구현예를 제공하기 위해 상기 참조 및 적용의 조성물, 기능 및 개념을 사용하도록 변형될 수 있다. 또한, 생물학적 기능적 동등성 고려 사항으로 인해, 생물학적 또는 화학적 작용의 종류 또는 양에 영향을 미치지 않으면서 단백질 구조에서 약간의 변화가 이루어질 수 있다. 상세한 설명에 비추어 본 개시내용에 대한 이들 및 다른 변경이 이루어질 수 있다. 이러한 모든 변형은 첨부된 청구범위의 범위 내에 포함되도록 의도된다.
임의의 전술한 구현예의 특정 요소는 조합되거나 다른 구현예의 요소를 대체할 수 있다. 또한, 본 개시내용의 특정 구현예와 관련된 이점이 이들 구현예의 맥락에서 설명되었지만, 다른 구현예도 그러한 이점을 나타낼 수 있으며, 모든 구현예가 본 개시내용의 범위 내에 있도록 반드시 이러한 이점을 나타낼 필요는 없다.
본원에 기재된 기술은 하기 실시예에 의해 추가로 설명되며, 이는 결코 추가로 제한되는 것으로 해석되어서는 안된다.
본 발명은 본원에 기재된 특정 방법론, 프로토콜 및 시약 등에 제한되지 않으며, 다양할 수 있음을 이해해야 한다. 본원에서 사용된 용어는 단지 특정한 구현예를 설명하기 위한 것으로, 본 발명의 범위를 한정하려는 의도가 아니며, 본 발명의 범위는 청구범위에 의해서만 정의된다.
II. 유전자 편집용 ceDNA 벡터
본 발명의 구현예는 숙주 세포에서 유전자 편집 분자인 이식유전자를 발현하고 (예를 들어, 이식유전자는 뉴클레아제, 예컨대 ZFN, TALEN, Cas; 하나 이상의 가이드 RNA; CRISPR; 리보핵단백질 (RNP), 또는 이들의 임의의 조합임), 보다 효율적인 게놈 편집을 초래할 수 있는 폐쇄 말단 선형 이중화 (ceDNA) 벡터를 포함하는 방법 및 조성물에 기초한다. 본원에 기재된 ceDNA 벡터는 크기에 의해 제한되지 않으므로, 예를 들어, 단일 벡터로부터 유전자 편집 시스템 (예를 들어, CRISPR/Cas 유전자 편집 시스템 (예를 들어, Cas9 또는 변형된 Cas9 효소, 가이드 RNA 및/또는 상동성 지정 복구 주형), 또는 TALEN 또는 아연 핑거 시스템)에 필요한 모든 성분의 발현을 허용한다. 그러나, 하나의 벡터 상에 인코딩된 이러한 성분 중 하나 또는 둘만을 갖는 반면, 나머지 성분(들)은 별도의 ceDNA 벡터 또는 예를 들어 전통적인 플라스미드에서 발현될 수 있는 것으로 또한 고려된다.
본원의 일 측면은, 예를 들어, 하나 이상의 외인성 공여체 서열을 세포 염색체 상의 특정 표적 부위에 고효율로 도입하기 위한 DNA 녹인 방법(들)에 대한 신규 ceDNA 벡터에 관한 것이다. 유전자 편집을 위한 하나 이상의 ceDNA를 사용하는 것에 이외에, ceDNA 벡터가 (i) 적어도 하나의 WT ITR 및 적어도 하나의 변형된 AAV 역 말단 반복부 (모드-ITR) (예를 들어, 비대칭인 변형된 ITR); (ii) 모드-ITR 쌍이 서로 상이한 3차원 공간 구성을 갖는 2개의 변형된 ITR (예를 들어, 비대칭 변형된 ITR), 또는 (iii) 대칭 또는 실질적으로 대칭인 WT-WT ITR 쌍 (여기서 각각의 WT-ITR은 동일한 3차원 공간 구성을 가짐), 또는 (iv) 대칭 또는 실질적으로 대칭인 변형된 ITR 쌍 (여기서 각각의 모드-ITR은 동일한 3차원 공간 구성을 가짐) 중 어느 것으로부터 선택된 ITR 서열을 포함하며, 본원에 개시된 방법 및 조성물은, 비제한적으로, 리포좀 나노입자 전달 시스템과 같은 전달 시스템을 추가로 포함할 수 있다. 사용을 위해 포함되는 비제한적인 예시적인 리포좀 나노입자 시스템이 본원에 개시되어 있다. 일부 측면에서, 본 개시내용은 유전자 편집을 위한 ceDNA 및 이온화 가능한 지질을 포함하는 지질 나노입자를 제공한다. 예를 들어, 공정에 의해 수득된 유전자 편집 ceDNA로 제조되고 로딩된 지질 나노입자 제형은 2018년 9월 7일자로 출원된 국제 출원 PCT/US2018/050042에 개시되어 있으며, 이는 본원에 포함된다.
공유적으로 폐쇄된 말단 (ceDNA)을 갖는 신규한 비-바이러스성, 캡시드-비함유 ceDNA 분자가 본원에 제공된다. 이들 비-바이러스성 캡시드 비함유 ceDNA 분자는 2개의 상이한 역 말단 반복 (ITR) 서열 사이에 위치한 이종 유전자 (이식유전자)를 함유하는 발현 작제물 (예를 들어, ceDNA-플라스미드, ceDNA-박미드, ceDNA- 바쿨로바이러스 또는 통합된 세포주)로부터 허용 숙주 세포에서 생성될 수 있으며, 여기서 ITR은 서로 상이하다. 일부 구현예에서, ITR 중 하나는 야생형 ITR 서열 (예를 들어 AAV ITR)과 비교하여 결실, 삽입 및/또는 치환에 의해 변형되고; ITR 중 적어도 하나는 기능적 말단 분해 부위 (trs) 및 Rep 결합 부위를 포함한다. ceDNA 벡터는 바람직하게는 발현 카세트와 같이, 분자의 적어도 일부에 대해 듀플렉스, 예를 들어 자가 상보적이다 (예를 들어, ceDNA는 이중 가닥 원형 분자가 아님). ceDNA 벡터는 공유적으로 폐쇄된 말단을 가지며, 따라서 37℃에서 한 시간 이상에 대한 엑소뉴클레아제 소화 (예를 들어 엑소뉴클레아제 I 또는 엑소뉴클레아제 III)에 저항성이 있다.
본원에 개시된 유전자 편집용 ceDNA 벡터는 바이러스 캡시드 내의 제한 공간에 의해 부과되는 패키징 제약이 없다. ceDNA 벡터는 캡슐화된 AAV 게놈과 대조적으로, 원핵생물 생산 플라스미드 DNA 벡터에 대한 생존가능 진핵생물적으로 생산된 대안을 나타낸다. 이는 제어 요소, 예를 들어, 본원에 개시된 바와 같은 조절 스위치, 대형 이식유전자, 다수의 이식유전자 등의 삽입을 허용한다.
일 측면에서, 유전자 편집용 ceDNA 벡터는 5'에서 3' 방향으로, 제1 아데노 관련 바이러스 (AAV) 역 말단 반복부 (ITR), 관심 뉴클레오티드 서열 (예를 들어 본원에 기재된 발현 카세트) 및 제2 AAV ITR을 포함한다. 일부 구현예에서, 제1 ITR (5' ITR)과 제2 ITR (3' ITR)은 서로 비대칭이며, 즉, 서로 다른 3D 공간 구성을 갖는다. 예시적인 구현예로서, 제1 ITR은 야생형 ITR일 수 있고, 제2 ITR은 돌연변이되거나 변형된 ITR일 수 있으며, 반대로, 제1 ITR이 돌연변이되거나 변형된 ITR이고, 제2 ITR이 야생형 ITR일 수 있다. 또 다른 구현예에서, 제1 ITR 및 제2 ITR은 모두 변경되었지만 상이한 서열이거나, 또는 상이한 변형을 갖거나, 또는 동일한 변형된 ITR이 아니며, 상이한 3D 공간 구성을 갖는다. 다르게 말하면, 비대칭 ITR을 이용한 유전자 편집용 ceDNA 벡터는 WT-ITR에 대한 하나의 ITR의 임의의 변화가 다른 ITR에 반영되지 않는 ITR을 가지며; 또는 대안적으로, 비대칭 ITR이 변형된 비대칭 ITR 쌍을 갖는 경우 서로 상이한 서열 및 상이한 3차원 형상을 가질 수 있다. ceDNA 벡터에서 및 ceDNA-플라스미드를 생성하기 위해 사용하기 위한 예시적인 비대칭 ITR은 하기 "비대칭 ITR"이라는 제목의 섹션에서 논의된다.
또 다른 측면에서, 유전자 편집용 ceDNA 벡터는 5'에서 3' 방향으로, 제1 아데노 관련 바이러스 (AAV) 역 말단 반복부 (ITR), 관심 뉴클레오티드 서열 (예를 들어 본원에 기재된 발현 카세트) 및 제2 AAV ITR을 포함하며, 여기서 제1 ITR (5' ITR)과 제2 ITR (3' ITR)이 서로 대칭이거나 실질적으로 대칭이고, 즉, 유전자 편집 ceDNA 벡터는 구조가 기하학적 공간에서 동일한 형상이거나 3D 공간에서 동일한 A, C-C' 및 B-B' 루프를 갖도록 대칭인 3차원 공간 구성을 갖는 ITR 서열을 포함할 수 있다. 이러한 구현예에서, 대칭 ITR 쌍, 또는 실질적으로 대칭인 ITR 쌍은 야생형 ITR이 아닌 변형된 ITR (예를 들어, 모드-ITR)일 수 있다. 모드-ITR 쌍은 야생형 ITR로부터 하나 이상의 변형을 갖고 서로의 역 보체 (역상)인 동일한 서열을 가질 수 있다. 대안적인 구현예에서, 변형된 ITR 쌍은 본 명세서에 정의된 바와 같이 실질적으로 대칭이고, 즉, 변형된 ITR 쌍은 상이한 서열을 가질 수 있지만 상응하는 또는 동일한 대칭 3차원 형상을 가질 수 있다. 일부 구현예에서, 대칭 ITR 또는 실질적으로 대칭인 ITR은 본원에 기재된 바와 같은 야생형 (WT-ITR)일 수 있다. 즉, 두 ITR은 모두 야생형 서열을 갖지만 반드시 동일한 AAV 혈청형으로부터의 WT-ITR일 필요는 없다. 즉, 일부 구현예에서, 하나의 WT-ITR은 하나의 AAV 혈청형으로부터 유래될 수 있고, 다른 WT-ITR은 상이한 AAV 혈청형으로부터 유래될 수 있다. 이러한 구현예에서, WT-ITR 쌍은 본원에 정의된 바와 같이 실질적으로 대칭이며, 즉, 대칭인 3차원 공간 구성을 여전히 유지하면서 하나 이상의 보존적 뉴클레오티드 변형을 가질 수 있다.
대칭 ITR 또는 실질적으로 대칭인 ITR은 아래의 "대칭 ITR 쌍"이라는 제목의 섹션에서 논의된다.
본원에 제공된 야생형 또는 돌연변이된 또는 달리 변형된 ITR 서열은 ceDNA 벡터의 생성을 위한 발현 작제물 (예를 들어, ceDNA-플라스미드, ceDNA 박미드, ceDNA-바쿨로바이러스)에 포함된 DNA 서열을 나타낸다. 따라서, ceDNA-플라스미드 또는 다른 발현 작제물로부터 생성된 ceDNA 벡터에 실제로 함유된 ITR 서열은 생산 과정 동안 일어나는 자연적으로 발생하는 변화 (예를 들어, 복제 오류)의 결과로 본 명세서에 제공된 ITR 서열과 동일하거나 동일하지 않을 수 있다.
일부 구현예에서, 유전자 편집 분자, 또는 유전자 편집 핵산 서열인 이식유전자를 갖는 발현 카세트를 포함하는 본원에 기재된 ceDNA 벡터는 이식유전자의 발현을 허용하거나 제어하는 하나 이상의 조절 서열(들)에 작동적으로 연결될 수 있다. 일 구현예에서, 폴리뉴클레오티드는 제1 ITR 서열 및 제2 ITR 서열을 포함하고, 여기서 관심 뉴클레오티드 서열은 제1 및 제2 ITR 서열에 의해 플랭킹되고, 제1 및 제2 ITR 서열은 서로 비대칭이거나 서로 대칭이다.
이들 각각의 측면에서 하나의 구현예에서, 발현 카세트는 이식유전자에 작동 가능하게 연결된 프로모터, 전사 후 조절 요소, 및 폴리아데닐화 및 종결 신호 중 하나 이상을 갖는 다음 순서로 구성된 2개의 ITR 사이에 위치된다. 일 구현예에서, 프로모터는 조절 가능하고, 즉 유도 가능하거나 억제 가능하다. 프로모터는 이식유전자의 전사를 용이하게 하는 임의의 서열 일 수 있다. 일 구현예에서 프로모터는 CAG 프로모터 (예를 들어, 서열 번호: 03) 또는 이의 변형이다. 전사 후 조절 요소는 이식유전자의 발현을 조절하는 서열, 비제한적인 예로서, 유전자 편집 분자, 또는 유전자 편집 핵산 서열인 이식유전자의 발현을 향상시키는 3차 구조를 생성하는 임의의 서열이다.
일 구현예에서, 전사 후 조절 요소는 WPRE (예를 들어, 서열 번호: 08)를 포함한다. 일 구현예에서, 폴리아데닐화 및 종결 신호는 BGH폴리A (예를 들어, 서열 번호: 09)를 포함한다. 당해 기술 분야에 공지된 임의의 시스 조절 요소, 또는 이들의 조합, 예를 들어, SV40 후기 폴리 A 신호 상류 인핸서 서열 (USE), 또는 단순 포진 바이러스 또는 B형 간염 바이러스 (HBV)의 티미딘 키나제 유전자를 포함하지만, 이에 제한되지 않는, 다른 전사 후 처리 요소가 추가로 사용될 수 있다. 일 구현예에서, 5'에서 3' 방향으로의 발현 카세트 길이는 AAV 비리온에 캡슐화되는 것으로 알려진 최대 길이보다 더 크다. 일 구현예에서, 길이는 4.6 kb 초과, 5 kb 초과, 또는 6 kb 초과, 또는 7 kb 초과이다. 다양한 발현 카세트가 본원에 예시된다.
발현 카세트는 4000개 초과의 뉴클레오티드, 5000개 뉴클레오티드, 10,000개 뉴클레오티드 또는 20,000개 뉴클레오티드, 또는 30,000개 뉴클레오티드, 또는 40,000개 뉴클레오티드 또는 50,000개 뉴클레오티드, 또는 약 4000-10,000개 뉴클레오티드 또는 10,000-50,000개 뉴클레오티드 사이의 임의의 범위, 또는 50,000개 초과의 뉴클레오티드를 포함할 수 있다. 일부 구현예에서, 발현 카세트는 500 내지 50,000개 뉴클레오티드 길이 범위의 유전자 편집 분자, 또는 유전자 편집 핵산 서열인 이식유전자를 포함할 수 있다. 일부 구현예에서, 발현 카세트는 500 내지 75,000개 뉴클레오티드 길이 범위의 유전자 편집 분자, 또는 유전자 편집 핵산 서열인 이식유전자를 포함할 수 있다. 일부 구현예에서, 발현 카세트는 500 내지 10,000개 뉴클레오티드 길이 범위의 유전자 편집 분자, 또는 유전자 편집 핵산 서열인 이식유전자를 포함할 수 있다. 일부 구현예에서, 발현 카세트는 1000 내지 10,000개 뉴클레오티드 길이 범위의 유전자 편집 분자, 또는 유전자 편집 핵산 서열인 이식유전자를 포함할 수 있다. 일부 구현예에서, 발현 카세트는 500 내지 5,000개 뉴클레오티드 길이 범위의 유전자 편집 분자, 또는 유전자 편집 핵산 서열인 이식유전자를 포함할 수 있다. ceDNA 벡터는 캡슐화된 AAV 벡터의 크기 제한을 갖지 않으므로, 대형 발현 카세트를 전달하여 유전자 편집 분자, 또는 유전자 편집 핵산 서열인 효율적인 이식유전자를 제공할 수 있다. 일부 구현예에서, ceDNA 벡터는 원핵생물-특이적 메틸화가 없다.
발현 카세트는 또한 내부 리보솜 진입 부위 (IRES) 및/또는 2A 요소를 포함할 수 있다. 시스 조절 요소는 프로모터, 리보스위치, 절연체, 미르-조절 가능 요소, 전사 후 조절 요소, 조직- 및 세포 유형-특이적 프로모터 및 인핸서를 포함하지만, 이에 제한되지는 않는다. 일부 구현예에서, ITR은 이식유전자의 프로모터로서 작용할 수 있다. 일부 구현예에서, ceDNA 벡터는 이식유전자의 발현을 조절하기 위한 추가 성분, 예를 들어, 본원에서 이식유전자의 발현을 제어 및 조절하기 위한 "조절 스위치"라는 제목의 섹션에 기재되어 있는 조절 스위치를 포함하고, 원하면, ceDNA 벡터를 포함하는 세포의 제어된 세포 사멸을 가능하게 하는 사멸 스위치인 조절 스위치를 포함할 수 있다.
도 1a-1e는 비제한적, 예시적인 ceDNA 벡터 또는 상응하는 ceDNA 플라스미드 서열의 개략도를 보여준다. ceDNA 벡터는 캡시드가 없고, 제1 ITR, 발현 가능한 이식유전자 카세트 및 제2 ITR의 순서로 인코딩된 플라스미드로부터 수득될 수 있으며, 여기서 제1 및/또는 제2 ITR 서열 중 적어도 하나는 상응하는 야생형 AAV2 ITR 서열에 대해 돌연변이된다. 카세트는 바람직하게는 다음 순서로: 인핸서/프로모터, ORF 리포터 (이식유전자), 전사 후 조절 요소 (예를 들어, WPRE), 및 폴리아데닐화 및 종결 신호 (예를 들어, BGH 폴리A) 중 하나 이상을 포함한다.
발현 카세트는 유전자 편집 분자 또는 유전자 편집 핵산 서열인 임의의 이식유전자를 포함할 수 있다. 유전자 편집 ceDNA 벡터는 폴리펩티드를 인코딩하는 핵산, 또는 비-코딩 핵산 (예를 들어, RNAi, miR 등), 뿐만 아니라 대상체의 게놈 내의 바이러스 서열, 예를 들어, HIV 바이러스 서열 등을 포함하는 외인성 유전자 및 뉴클레오티드 서열을 포함하지만, 이에 제한되지 않는, 대상체에서의 임의의 관심 유전자를 편집한다. 바람직하게는 본원에 개시된 유전자 편집 ceDNA 벡터는 치료 목적 (예를 들어, 의학적, 진단 또는 수의학적 용도) 또는 면역원성 폴리펩티드에 사용된다. 특정 구현예에서, 유전자 편집 ceDNA 벡터는 하나 이상의 폴리펩티드, 펩티드, 리보자임, 펩티드 핵산, siRNA, RNAis, 안티센스 올리고뉴클레오티드, 안티센스 폴리뉴클레오티드, 항체, 항원 결합 단편, 또는 이들의 임의의 조합을 포함하는, 대상체에서의 임의의 관심 유전자를 편집할 수 있다.
ceDNA 발현 카세트는, 예를 들면, 수용 대상체에서 부재하거나, 비활성이거나 활성이 불충분한 단백질을 인코딩하는 발현 가능한 외인성 서열 (예를 들어, 오픈 리딩 프레임) 또는 원하는 생물학적 또는 치료적 효과를 갖는 단백질을 인코딩하는 유전자를 포함할 수 있다. 공여체 서열과 같은 외인성 서열은 결함 유전자 또는 전사체의 발현을 교정하는 기능을 할 수 있는 유전자 산물을 인코딩할 수 있다. 발현 카세트는 또한 교정 DNA 가닥을 인코딩하고, 폴리펩티드, 센스 또는 안티센스 올리고뉴클레오티드를 인코딩하거나, 또는 RNA (코딩 또는 비-코딩; 예를 들어, siRNA, shRNA, 마이크로-RNA 및 이들의 안티센스 대응물 (예를 들어, antagoMiR))를 인코딩할 수 있다. 발현 카세트는 β-락타마제, β-갈락토시다제 (LacZ), 알칼리성 포스파타제, 티미딘 키나제, 녹색 형광 단백질 (GFP), 클로람페니콜 아세틸트랜스퍼라제 (CAT), 루시퍼라제 및 당 업계에 잘 알려진 다른 것들과 같은 실험 또는 진단 목적으로 사용되는 리포터 단백질을 인코딩하는 외인성 서열을 포함할 수 있다.
원칙적으로, 발현 카세트는 돌연변이로 인해 감소되거나 부재하거나 과발현될 때 치료적 이점을 전달하는 단백질, 폴리펩티드 또는 RNA를 인코딩하는 임의의 유전자를 포함할 수 있으며, 이는 본 개시의 범위 내에 있는 것으로 간주된다. ceDNA 벡터는 뉴클레아제에 의해 제공된 이중 가닥 파괴 (또는 닉) 후에 삽입되는 교정 DNA 가닥으로서 사용되는 주형 또는 공여체 뉴클레오티드 서열을 포함할 수 있다. ceDNA 벡터는 가이드 RNA 뉴클레아제, 메가뉴클레아제 또는 아연 핑거 뉴클레아제에 의해 제공된 이중 가닥 파괴 (또는 닉) 후에 삽입되는 교정 DNA 가닥으로서 사용되는 주형 뉴클레오티드 서열을 포함할 수 있다. 바람직하게는, 비-삽입된 박테리아 DNA는 존재하지 않으며 바람직하게는 박테리아 DNA가 본원에 제공된 ceDNA 조성물에 존재하지 않는다. 일부 경우에, 단백질은 닉 없이 코돈을 변화시킬 수 있다.
본원에 기재된 ceDNA 벡터의 발현 카세트, 발현 작제물 또는 공여체 서열에 제공된 서열은 숙주 세포에 대해 코돈 최적화될 수 있다. 본원에 사용된 용어 "코돈 최적화된" 또는 "코돈 최적화"는 천연 서열 (예를 들어, 원핵생물 서열)의 적어도 하나, 하나 초과 또는 상당한 수의 코돈을 척추동물의 유전자에서 보다 빈번하게 또는 가장 빈번하게 사용되는 코돈으로 대체함으로써 관심 척추동물 세포, 예를 들어 마우스 또는 인간의 세포에서 발현을 향상시키기 위해 핵산 서열을 변형시키는 과정을 지칭한다. 다양한 종은 특정 아미노산의 특정 코돈에 대해 특정한 편향을 나타낸다. 전형적으로, 코돈 최적화는 원래 번역된 단백질의 아미노산 서열을 변경시키지 않는다. 최적화된 코돈은 예를 들어, Aptagen's Gene Forge® 코돈 최적화 및 맞춤형 유전자 합성 플랫폼 (Aptagen, Inc., 2190 Fox Mill Rd. Suite 300, Herndon, Va. 20171) 또는 또 다른 공개적으로 이용 가능한 데이터베이스를 사용하여 결정될 수 있다.
많은 유기체는 성장하는 펩티드 사슬에 특정 아미노산의 삽입을 코딩하기 위해 특정 코돈을 사용하기 위한 편향을 나타낸다. 유기체 간의 코돈 사용의 차이인 코돈 선호도 또는 코돈 편향은 유전자 코드의 축퇴에 의해 제공되며, 많은 유기체 사이에서 잘 기록되어 있다. 코돈 편향은 종종 메신저 RNA (mRNA)의 번역 효율과 상관 관계가 있으며, 이는 특히 번역되는 코돈의 특성 및 특정 전이 RNA (tRNA) 분자의 유용성에 의존하는 것으로 여겨진다. 세포에서 선택된 tRNA의 우세는 일반적으로 펩티드 합성에 가장 빈번하게 사용되는 코돈의 반영이다. 따라서, 유전자는 코돈 최적화에 기초하여 주어진 유기체에서 최적의 유전자 발현을 위해 조정될 수 있다.
광범위한 동물, 식물 및 미생물 종에 이용 가능한 많은 유전자 서열을 고려할 때, 코돈 사용의 상대 빈도를 계산할 수 있다 (Nakamura, Y., et al. "Codon usage tabulated from the international DNA sequence databases: status for the year 2000" Nucl. Acids Res. 28:292 (2000)).
일부 구현예에서, 유전자 편집 유전자 (예를 들어, 공여체 서열) 또는 가이드 RNA는 치료 유전자를 표적으로 한다. 일부 구현예에서, 가이드 RNA는 항체 또는 항체 단편 또는 이의 항원-결합 단편, 예를 들어, 중화 항체 또는 항체 단편 등을 표적으로 한다.
특히, 유전자 편집 유전자 (예를 들어, 공여체 서열) 또는 가이드 RNA는 질환, 기능 장애, 손상 및/또는 장애의 하나 이상의 증상의 치료, 예방 및/또는 개선에 사용하기 위한, 예를 들어, 단백질(들), 폴리펩티드(들), 펩티드(들), 효소(들), 항체, 항원 결합 단편, 및 이의 변이체 및/또는 이의 활성 단편을 포함하지만, 이에 제한되지 않는, 하나 이상의 치료제(들)를 표적으로 한다. 가이드 RNA로 표적화하기 위한 예시적인 유전자는 본원에서 "치료 방법"이라는 제목의 섹션에 기재되어 있다.
플라스미드 기반 발현 벡터와 다른 ceDNA 벡터의 많은 구조적 특징이 있다. ceDNA 벡터는 다음의 특징들 중 하나 이상을 가질 수 있다: 원래 (즉, 삽입되지 않은) 박테리아 DNA의 결핍, 원핵생물 복제 기원의 결핍, 자가-포함(being self-containing), 즉, 이들은 Rep 결합 및 말단 분해 부위 (RBS 및 TRS)를 포함하는 2개의 ITR, 및 ITR들 사이의 외인성 서열 이외의 임의의 서열을 요구하지 않음, 진핵생물 기원의, 헤어핀을 형성하는 ITR 서열의 존재 (즉, 이들은 진핵생물 세포에서 생성됨), 및 박테리아 유형 DNA 메틸화 또는 실제로 포유동물 숙주에 의해 비정상적인 것으로 간주되는 임의의 다른 메틸화의 부재. 일반적으로, 본 벡터는 임의의 원핵생물 DNA를 함유하지 않는 것이 바람직하지만, 일부 원핵생물 DNA는 프로모터 또는 인핸서 영역에서 비제한적인 예로서 외인성 서열로서 삽입될 수 있는 것으로 고려된다. 플라스미드 발현 벡터로부터 ceDNA 벡터를 구별하는 또 다른 중요한 특징은 ceDNA 벡터가 폐쇄 말단을 갖는 단일 가닥 선형 DNA인 반면, 플라스미드는 항상 이중 가닥 DNA라는 것이다.
본원에 제공된 방법에 의해 생성된 유전자 편집용 ceDNA 벡터는 바람직하게는 제한 효소 소화 분석에 의해 결정된 바와 같이, 비-연속 구조보다는 선형 및 연속 구조를 갖는다 (도 4d). 선형 및 연속 구조는 세포 엔도뉴클레아제에 의한 공격으로부터 보다 안정할뿐만 아니라 재조합되고 돌연변이를 유발할 가능성이 적은 것으로 여겨진다. 따라서, 선형 및 연속 구조에서 ceDNA 벡터를 편집하는 유전자가 바람직일 구현예이다. 연속적인, 선형의 단일 가닥 분자내 듀플렉스 ceDNA 벡터는 AAV 캡시드 단백질을 인코딩하는 서열 없이 공유 결합된 말단을 가질 수 있다. 이들 유전자 편집 ceDNA 벡터는 박테리아 기원의 원형 듀플렉스 핵산 분자인 플라스미드 (본원에 기재된 ceDNA 플라스미드 포함)와 구조적으로 구별된다. 상보적인 플라스미드 가닥은 변성 후에 분리되어 2개의 핵산 분자를 생성할 수 있는 반면, 대조적으로, 상보적인 가닥을 가지면서 ceDNA 벡터는 단일 DNA 분자이고, 따라서 변성되더라도 단일 분자로 남아있다. 일부 구현예에서, 본원에 기재된 바와 같은 ceDNA 벡터는 플라스미드와 달리, 원핵생물 유형의 DNA 염기 메틸화 없이 생성될 수 있다. 따라서, ceDNA 벡터 및 ceDNA-플라스미드는 둘 모두 구조 측면에서 (특히 선형 대 원형) 및 또한 이들 상이한 대상을 생성 및 정제하는데 사용된 방법의 관점에서 (아래 참조), 및 ceDNA-플라스미드에 대해 원핵생물 유형이고 ceDNA 벡터에 대해 진핵생물 유형인 DNA 메틸화의 관점에서 상이하다.
플라스미드-기반 발현 벡터에 비해 유전자 편집을 위해 본원에 기재된 ceDNA 벡터를 사용하는 것의 몇 가지 이점이 있으며, 이러한 이점은 다음을 포함하지만, 이에 제한되지는 않는다: 1) 플라스미드는 박테리아 DNA 서열을 함유하고 원핵생물-특이적 메틸화, 예를 들어, 6-메틸 아데노신 및 5-메틸 시토신 메틸화를 거친 박테리아 DNA 서열을 함유하고, 반면 캡시드-비함유 AAV 벡터 서열은 진핵생물 기원이고 원핵생물-특이적 메틸화를 겪지 않으며; 결과적으로, 캡시드-비함유 AAV 벡터는 플라스미드와 비교하여 염증 및 면역 반응을 유발할 가능성이 적고; 2) 플라스미드는 생산 공정 동안 저항성 유전자의 존재를 요구하지만, ceDNA 벡터는 그렇지 않고; 3) 원형 플라스미드가 세포 내로 도입될 때 핵으로 전달되지 않고, 세포 뉴클레아제에 의한 분해를 우회하기 위해 과부하가 필요한 반면, ceDNA 벡터는 바이러스성 시스-요소, 즉, 클레아제에 대한 저항성을 부여하고 핵으로 표적화되고 전달되도록 설계될 수 있는 ITR을 함유한다. ITR 기능에 없어서는 안 될 최소 정의 요소는 Rep-결합 부위 (RBS; AAV2의 경우 5'-GCGCGCTCGCTCGCTC-3' (서열 번호: 531)) 및 말단 분해 부위 (TRS; AAV2의 경우 5'-AGTTGG-3' (서열 번호: 48)) 및 헤어핀 형성을 허용하는 가변 팔린드롬 서열인 것으로 가정되고; 4) ceDNA 벡터는 T 세포-매개 면역 반응을 유발하는, 톨-유사 수용체 패밀리의 일원에 결합하는 것으로 보고된 원핵생물-유래 플라스미드에서 종종 발견되는 CpG 디뉴클레오티드의 과다-표현(over-representation)을 갖지 않는다. 대조적으로, 본원에 개시된 캡시드-비함유 AAV 벡터를 이용한 형질도입은 다양한 전달 시약을 사용하여 통상적인 AAV 비리온으로 형질도입하기 어려운 세포 및 조직 유형을 효율적으로 표적화할 수 있다.
III. 원하는 핵산 서열의 녹인
본원에 기재된 유전자 편집 ceDNA 벡터, 방법 및 조성물은 새로운 핵산 서열을 도입하거나, 게놈 서열의 돌연변이를 교정하거나, 숙주 세포에서 표적 유전자 서열로 돌연변이를 도입하는데 사용될 수 있다. 이러한 방법은 "DNA 녹인 시스템"이라고 할 수 있다. 본원에 기재된 바와 같은 DNA 녹인 시스템은 공여체 서열이 임의의 원하는 표적 부위에 고효율로 삽입될 수 있도록 하여, 외인성 유전자를 발현하는 형질전환 동물의 생성, 질환의 세포 배양 모델의 제조, 스크리닝 분석 시스템의 제조, 조작된 조직 작제물의 유전자 발현의 변형, 게놈 유전자좌의 변형 (예를 들어, 돌연변이), 및 예를 들어 외인성 비-코딩 서열 (예컨대 서열 태그 또는 조절 요소)을 게놈에 첨가함에 의한 유전자 편집과 같은 많은 용도를 위해 실행 가능하도록 한다. 본원에 제공된 방법을 사용하여 생산된 세포 및 동물은, 예를 들어 세포 치료제, 질환 모델, 연구 도구, 및 다양한 목적에 유용한 인간화 동물로서 다양한 용도를 발견할 수 있다.
본 개시내용의 DNA 녹인 시스템은 또한 큰 공여체 서열 (<5kb)을 사용하는 유전자 편집 기술이 게놈 내 임의의 원하는 표적 부위에 삽입될 수 있도록 하여, 현재 기술보다 더 큰 유전자의 유전자 편집을 제공한다. 일부 구현예에서, 큰 상동성 아암, 예를 들어 50개의 염기쌍 내지 2천 개의 염기쌍이 포함되어 탁월한 효율 (높은 온-표적(on-target)) 및 탁월한 특이성 (낮은 오프-표적)으로 유전자 편집을 제공하고, 일부 구현예에서는, 뉴클레아제 사용 없이 HDR을 제공한다.
본 개시내용의 DNA 녹인 시스템은 또한 유전자 편집을 위한 공여체 서열의 투여와 관련하여 몇 가지 이점을 제공한다. 먼저, 본 개시내용의 전달 입자 내에 본원에 기재된 바와 같은 ceDNA 벡터의 투여는 기준선 면역에 의해 배제되지 않으므로 특정 장애를 갖는 임의의 그리고 잠재적으로 모든 환자에게 투여될 수 있다. 둘째로, 본 개시내용의 입자를 투여하는 것은 바이러스성 벡터-기반 전달 시스템에 대해 전형적으로 발생된 것과 같은 전달된 치료제에 대한 적응성 면역 반응을 생성하지 않으므로, 구현예는 임상 효과를 위해 필요에 따라 재투여될 수 있다. 생체내 전달과 같은 본 개시내용에 따른 하나 이상의 ceDNA 벡터의 투여는 반복 가능하고 강력하다.
특정 구현예에서, 본 개시내용의 ceDNA 벡터를 사용한 유전자 편집은 유전자 교정의 효율을 평가하기 위해 치료된 환자로부터의 적절한 바이오마커로 모니터링될 수 있고, 적절한 수준의 유전자 편집이 달성될 때까지 치료 제품의 반복 투여가 이루어질 수 있다.
또 다른 측면에서, 본 개시내용에 따른 ceDNA 벡터와 함께 본원에 기재된 유전자 녹인 시스템을 사용하여 유전자 변형된 동물을 생성하는 방법이 제공된다. 이러한 방법은 아래에 더 자세히 설명되어 있다.
특정 구현예에서, 본 개시내용은 진핵 세포 또는 원핵 세포와 같은 숙주 세포의 염색체 상의 미리 결정된 삽입 부위에 공여체 서열을 삽입하기 위해 ceDNA 벡터를 사용하는 방법에 관한 것이다.
IV. 유전자 편집 시스템 성분- 일반
RNA 유도된 뉴클레아제를 포함하는 것들과 같은 추가의 구현예에서, 유전자 편집에 필요한 성분은 뉴클레아제, 가이드 RNA (Cas9 등이 이용되는 경우), 공여체 서열 및 본 개시의 단일 ceDNA 벡터 내에 포함된 하나 이상의 상동성 아암을 포함할 수 있다. 이러한 구현예는 유전자 편집 성분을 전달하기 위해 별개의 또는 다양한 입자를 필요로 하는 접근법과 비교하여 유전자 편집의 효율을 증가시킨다.
추가의 구현예에서, 뉴클레아제는 유전자 편집 후에 비활성화/감소될 수 있고, 존재한다면, 세포 내에 첨가된 뉴클레아제의 지속성으로 발생할 수 있는 오프-표적 편집을 감소시키거나 제거할 수 있다.
또 다른 측면에서, 본 개시는 본원에 기재된 방법 중 임의의 하나에 사용하기 위한 하나 이상의 ceDNA 벡터를 포함하는 키트에 관한 것이다.
본원에 기재된 방법 및 조성물은 또한, 예를 들어, 하기에 기재된 바와 같이 세포 스위치를 포함하는 유전자 편집 시스템을 제공한다: Oakes et al. Nat. Biotechnol. 34:646-651 (2016) (이의 내용은 그 전문이 본원에 참고로 포함됨).
본원에 기재된 방법 및 조성물은 당 업계에 공지된 방법을 사용하여 고처리량 방식으로 수행될 수 있는 것으로 또한 구체적으로 고려된다 (예를 들어, Shalem et al. Nat Rev Genet 16:299-311 (2015); Shalem et al. Science 343:84-88 (2014); 이들 각각의 내용은 그 전문이 본원에 참고로 포함됨).
V. ITR
본원에 개시된 바와 같이, ceDNA 벡터는 2개의 역 말단 반복 (ITR) 서열 사이에 위치된 유전자 편집 핵산 서열을 함유하며, 여기서 ITR 서열은 이 용어가 본원에 정의된 바와 같이, 비대칭 ITR 쌍 또는 대칭 또는 실질적으로 대칭 ITR 쌍일 수 있다. 본원에 개시된 유전자 편집용 ceDNA 벡터는 (i) 적어도 하나의 WT ITR 및 적어도 하나의 변형된 AAV 역 말단 반복부 (모드-ITR) (예를 들어, 비대칭인 변형된 ITR); (ii) 모드-ITR 쌍이 서로 상이한 3차원 공간 구성을 갖는 2개의 변형된 ITR (예를 들어, 비대칭인 변형된 ITR), 또는 (iii) 대칭 또는 실질적으로 대칭인 WT-WT ITR 쌍 (여기서 각각의 WT-ITR은 동일한 3차원 공간 구성을 가짐), 또는 (iv) 대칭 또는 실질적으로 대칭인 변형된 ITR 쌍 (여기서 각각의 모드-ITR은 동일한 3차원 공간 구성을 가짐) 중 어느 것으로부터 선택된 ITR 서열을 포함할 수 있으며, 여기서 본 개시의 방법은 비제한적으로 리포좀 나노입자 전달 시스템과 같은 전달 시스템을 추가로 포함할 수 있다.
A. 대칭 ITR 쌍
일부 구현예에서, ITR 서열은 2개의 서브패밀리: 척추동물을 감염시키는 파보비리나에, 및 곤충을 감염시키는 덴소비리나에를 포함하는 파보비리다에 패밀리의 바이러스로부터 유래될 수 있다. 파보비리나에 서브패밀리 (파보바이러스라고 함)에는 데펜도바이러스 속이 포함되는데, 대부분의 조건에서, 이의 구성원은 생산적인 감염을 위해 아데노바이러스 또는 헤르페스 바이러스와 같은 헬퍼 바이러스로 동시 감염이 필요하다. 데펜도바이러스 속에는 일반적으로 사람 (예를 들어, 혈청형 2, 3A, 3B, 5 및 6) 또는 영장류 (예를 들어, 혈청형 1 및 4)를 감염시키는 아데노 관련 바이러스 (AAV) 및 다른 온혈 동물 (예를 들어, 소, 개, 말 및 양 아데노-관련 바이러스)을 감염시키는 관련 바이러스가 포함한다. 파보바이러스 및 파보비리다에 패밀리의 다른 구성원은 일반적으로 하기에 기재되어 있다: Kenneth I. Berns, "Parvoviridae: The Viruses and Their Replication," Chapter 69 in FIELDS VIROLOGY (3d Ed. 1996).
본원의 명세서 및 실시예에서 예시된 ITR은 AAV2 WT-ITR이지만, 당업자는, 상기 언급된 바와 같이, 임의의 공지된 파보바이러스, 예를 들어 데펜도바이러스 예컨대 AAV (예를 들어, AAV1, AAV2, AAV3, AAV4, AAV5, AAV 5, AAV7, AAV8, AAV9, AAV10, AAV 11, AAV12, AAVrh8, AAVrh10, AAV-DJ 및 AAV-DJ8 게놈. 예를 들어, NCBI:NC 002077; NC 001401; NC001729; NC001829; NC006152; NC 006260; NC 006261)로부터의 ITR, 키메라 ITR 또는 임의의 합성 AAV로부터의 ITR을 사용할 수 있다는 것을 알고 있다. 일부 구현예에서, AAV는 온혈 동물, 예를 들어, 조류 (AAAV), 소 (BAAV), 개, 말 및 양 아데노-관련 바이러스를 감염시킬 수 있다. 일부 구현예에서, ITR은 B19 파보바이러스 (GenBank 수탁 번호: NC 000883), 마우스로부터의 미세 바이러스 (Minute Virus from Mouse; MVM) (GenBank 수탁 번호 NC 001510); 거위 파보바이러스 (GenBank 수탁 번호 NC 001701); 뱀 파보바이러스 1 (GenBank 수탁 번호 NC 006148)에서 유래된다. 일부 구현예에서, 5' WT-ITR은 본원에 논의된 바와 같이, 하나의 혈청형으로부터 유래될 수 있고, 3' WT-ITR은 상이한 혈청형으로부터 유래될 수 있다.
통상의 숙련가는 ITR 서열이 이중 가닥 홀리데이 접합의 공통 구조를 가지며, 이는 전형적으로 T형 또는 Y형 헤어핀 구조 (예를 들어, 도 2a 및 도 3a 참조)임을 알고 있으며, 여기서 각각의 WT-ITR은 더 큰 팔린드롬 아암 (A-A')에 포매된 2개의 팔린드롬 아암 또는 루프 (B-B' 및 C-C')와 단일 가닥 D 서열 (여기서 이러한 팔린드롬 서열의 순서는 ITR의 플립 또는 플롭 배향을 한정함)에 의해 형성된다. 예를 들어, 하기에 기재된, 상이한 AAV 혈청형 (AAV1-AAV6)으로부터의 ITR의 구조 분석 및 서열 비교를 참조한다: Grimm et al., J. Virology, 2006; 80(1); 426-439; Yan et al., J. Virology, 2005; 364-379; Duan et al., Virology 1999; 261; 8-14. 당업자는 본원에 제공된 예시적인 AAV2 ITR 서열에 기초하여 ceDNA 벡터 또는 ceDNA-플라스미드에 사용하기 위한 임의의 AAV 혈청형으로부터 WT-ITR 서열을 쉽게 결정할 수 있다. 예를 들어, AAV2의 좌측 ITR과 다른 혈청형으로부터의 좌측 ITR의% 동일성: AAV-1 (84%), AAV-3 (86%), AAV-4 (79%), AAV-5 (58%), AAV-6 (좌측 ITR) (100%) 및 AAV-6 (우측 ITR) (82%)을 보여주는, 문헌(Grimm et al., J. Virology, 2006; 80(1); 426-439)에 기재된 상이한 AAV 혈청형 (AAV1-AAV6, 및 조류 AAV (AAAV) 및 소 AAV (BAAV))으로부터의 ITR의 서열 비교를 참조한다.
본원에서 논의된 바와 같이, 일부 구현예에서 유전자 편집용 ceDNA 벡터는 대칭 ITR 서열 (예를 들어, 대칭 ITR 쌍)을 포함할 수 있으며, 여기서 5' ITR 및 3' ITR은 서로 동일한 대칭 3차원 구성을 가질 수 있다 (즉, 대칭 또는 실질적으로 대칭). 즉, 유전자 편집용 ceDNA 벡터는 그 구조가 기하학적 공간에서 동일한 형상이거나 3D 공간에서 동일한 A, C-C' 및 B-B' 루프를 갖도록 (즉, 이들은 동일하거나 서로에 대한 거울상임) 대칭인 3차원 공간 구성을 갖는 ITR 서열을 포함한다. 이러한 구현예에서, 대칭 ITR 쌍, 또는 실질적으로 대칭 ITR 쌍은 야생형 ITR이 아닌 변형된 ITR (예를 들어, 모드-ITR)일 수 있다. 모드-ITR 쌍은 야생형 ITR로부터 하나 이상의 변형을 갖고 서로의 역 보체 (역상)인 동일한 서열을 가질 수 있다. 대안적인 구현예에서, 변형된 ITR 쌍은 본 명세서에 정의된 바와 같이 실질적으로 대칭이고, 즉, 변형된 ITR 쌍은 상이한 서열을 가질 수 있지만 상응하는 또는 동일한 대칭 3차원 형상을 가질 수 있다.
(i) 야생형 ITR
일부 구현예에서, 대칭 ITR 또는 실질적으로 대칭 ITR은 본원에 기재된 야생형 (WT-ITR)이다. 즉, 두 ITR 모두 야생형 서열을 갖지만 반드시 동일한 AAV 혈청형으로부터의 WT-ITR일 필요는 없다. 즉, 일부 구현예에서, 하나의 WT-ITR은 하나의 AAV 혈청형으로부터 유래될 수 있고, 다른 WT-ITR은 상이한 AAV 혈청형으로부터 유래될 수 있다. 이러일 구현예에서, WT-ITR 쌍은 본원에 정의된 바와 같이 실질적으로 대칭이며, 즉, 대칭인 3차원 공간 구성을 여전히 유지하면서 하나 이상의 보존적 뉴클레오티드 변형을 가질 수 있다.
따라서, 본원에 개시된 바와 같이, 유전자 편집용 ceDNA 벡터는 2개의 플랭킹 야생형 역 말단 반복 (WT-ITR) 서열 사이에 위치된 유전자 편집 서열을 함유하며, 이는 서로의 역 보체 (역상)이거나 또는 대안적으로는 서로 실질적으로 대칭이고, 즉, WT-ITR 쌍이 대칭인 3차원 공간 구성을 갖는다. 일부 구현예에서, 야생형 ITR 서열 (예를 들어, AAV WT-ITR)은 기능적 Rep 결합 부위 (RBS; 예를 들어, AAV2에 대한 5'-GCGCGCTCGCTCGCTC-3', 서열 번호: 531) 및 기능성 말단 분해 부위 (TRS; 예를 들어 5'-AGTT-3', 서열 번호: 46)를 포함한다.
일 측면에서, 유전자 편집용 ceDNA 벡터는 2개의 WT 역 말단 반복 서열 (WT-ITR) (예를 들어 AAV WT-ITR) 사이에 작동적으로 위치된 이종 핵산을 인코딩하는 벡터 폴리뉴클레오티드로부터 얻을 수 있다. 즉, 두 ITR 모두 야생형 서열을 갖지만, 반드시 동일한 AAV 혈청형으로부터의 WT-ITR일 필요는 없다. 즉, 일부 구현예에서, 하나의 WT-ITR은 하나의 AAV 혈청형으로부터 유래될 수 있고, 다른 WT-ITR은 상이한 AAV 혈청형으로부터 유래될 수 있다. 이러일 구현예에서, WT-ITR 쌍은 본원에 정의된 바와 같이 실질적으로 대칭이며, 즉, 대칭인 3차원 공간 구성을 여전히 유지하면서 하나 이상의 보존적 뉴클레오티드 변형을 가질 수 있다. 일부 구현예에서, 5' WT-ITR은 하나의 AAV 혈청형으로부터 유래되고, 3' WT-ITR은 동일하거나 상이한 AAV 혈청형으로부터 유래된다. 일부 구현예에서, 5' WT-ITR 및 3'WT-ITR은 서로의 거울상이고, 즉 대칭이다. 일부 구현예에서, 5' WT-ITR 및 3' WT-ITR은 동일한 AAV 혈청형으로부터 유래된다.
WT ITR은 잘 알려져 있다. 일 구현예에서, 2개의 ITR은 동일한 AAV2 혈청형으로부터 유래된다. 특정 구현예에서, 다른 혈청형으로부터의 WT를 사용할 수 있다. 상동성인 다수의 혈청형, 예를 들어 AAV2, AAV4, AAV6, AAV8이 있다. 일 구현예에서, 밀접한 상동성 ITR (예를 들어, 유사한 루프 구조를 갖는 ITR)이 사용될 수 있다. 또 다른 구현예에서, 보다 다양한 AAV WT ITR, 예를 들어, AAV2 및 AAV5를 사용할 수 있고, 또 다른 구현예에서, 실질적으로 WT인 ITR을 사용할 수 있으며, 즉, 그것은 WT의 기본 루프 구조를 갖지만 특성을 변경하거나 특성에 영향을 미치지 않는 일부 보존적 뉴클레오티드 변화를 갖는다. 동일한 바이러스 혈청형으로부터 WT-ITR을 사용할 때, 하나 이상의 조절 서열이 추가로 사용될 수 있다. 특정 구현예에서, 조절 서열은 ceDNA의 활성의 조절을 허용하는 조절 스위치이다.
일부 구현예에서, 본원에 기재된 기술의 한 측면은 공유적으로 폐쇄된 말단을 갖는 비-바이러스 캡시드 비함유 DNA 벡터 (ceDNA 벡터)에 관한 것이며, 여기서 ceDNA 벡터는 2개의 야생형 역 말단 반복 서열 (WT-ITR) 사이에 작동 가능하게 위치된 적어도 하나의 이종 뉴클레오티드 서열을 포함하며, WT-ITR은 동일한 혈청형, 상이한 혈청형으로부터 유래될 수 있거나 서로 실질적으로 대칭일 수 있다 (즉, 이들의 구조가 기하학적 공간에서 동일한 형상이거나 3D 공간에서 동일한 A, C-C' 및 B-B' 루프를 가짐). 일부 구현예에서, 대칭 WT-ITR은 기능성 말단 분해 부위 및 Rep 결합 부위를 포함한다. 일부 구현예에서, 이종 핵산 서열은 이식유전자를 인코딩하고, 벡터는 바이러스 캡시드에 있지 않다.
일부 구현예에서, WT-ITR은 동일하지만 서로의 역 보체이다. 예를 들어, 5' ITR에서 서열 AACG는 상응하는 부위에서 3' ITR에서 CGTT (즉, 역 보체)일 수 있다. 일례에서, 5' WT-ITR 센스 가닥은 ATCGATCG의 서열을 포함하고 상응하는 3' WT-ITR 센스 가닥은 CGATCGAT (즉, ATCGATCG의 역 보체)를 포함한다. 일부 구현예에서, WT-ITR ceDNA는 말단 분해 부위 및 복제 단백질 결합 부위 (RPS) (때로는 복제 단백질 결합 부위로 지칭됨), 예를 들어 Rep 결합 부위를 추가로 포함한다.
WT-ITR을 포함하는 ceDNA 벡터에 사용하기 위한 예시적인 WT-ITR 서열은 본원의 표 2에 도시되어 있으며, 이는 WT-ITR의 쌍 (5' WT-ITR 및 3' WT-ITR)을 나타낸다.
예시적인 예로서, 본 개시내용은 조절 스위치의 존재 또는 부재 하에 이식유전자 (예를 들어, 유전자 편집 서열)에 작동 가능하게 연결된 프로모터를 포함하는 폐쇄 말단 DNA 벡터를 제공하며, 여기서 ceDNA는 캡시드 단백질이 없고, (a) ceDNA는 WT-ITR을 인코딩하는 ceDNA-플라스미드 (예를 들어, 도 1f-1g 참조)로부터 생성되며, 여기서 각각의 WT-ITR은 이의 헤어핀 이차 구성에서 동일한 수의 분자내 이중화 염기쌍을 갖고 (바람직하게는 이러한 참조 서열과 비교하여 이 구성에서 임의의 AAA 또는 TTT 말단 루프의 결실을 배제함), (b) ceDNA는 미변성 겔 및 실시예 1의 변성 조건 하에서 아가로스 겔 전기영동에 의해 ceDNA 식별을 위한 분석을 사용하여 ceDNA로서 식별된다.
일부 구현예에서, 플랭킹 WT-ITR은 서로 실질적으로 대칭이다. 이 구현예에서, 5' WT-ITR은 AAV의 하나의 혈청형으로부터 유래될 수 있고, 3' WT-ITR은 AAV의 상이한 혈청형으로부터 유래될 수 있으므로 WT-ITR은 동일한 역 보체가 아니다. 예를 들어, 5' WT-ITR은 AAV2로부터 유래될 수 있고, 3' WT-ITR은 상이한 혈청형 (예를 들어, AAV1, 3, 4, 5, 6, 7, 8, 9, 10, 11 및 12)으로부터 유래될 수 있다. 일부 구현예에서, WT-ITR은 AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12, AAV13, 뱀 파보바이러스 (예를 들어, 로얄 피톤(royal python) 파보바이러스), 소 파보바이러스, 염소 파보바이러스, 조류 파보바이러스, 개 파보바이러스, 말 파보바이러스, 새우 파보바이러스, 돼지 파보바이러스 또는 곤충 AAV 중 어느 것으로부터 선택된 2개의 상이한 파보바이러스로부터 선택될 수 있다. 일부 구현예에서, 이러한 WT ITR의 조합은 AAV2 및 AAV6으로부터의 WT-ITR의 조합이다. 일 구현예에서, 실질적으로 대칭인 WT-ITR은 하나가 다른 ITR에 대해 반전될 때, 적어도 90% 동일하고, 적어도 95% 동일하고, 적어도 96% ... 97% ... 98% ... 99% .... 99.5% 및 그 사이의 모든 포인트 동일하고, 동일한 대칭 3차원 공간 구성을 갖는다. 일부 구현예에서, WT-ITR 쌍은 대칭인 3차원 공간 구성, 예를 들어, A, C-C'. B-B' 및 D 아암의 동일한 3D 구성을 갖기 때문에 실질적으로 대칭이다. 일 구현예에서, 실질적으로 대칭인 WT-ITR 쌍은 다른 것에 대해 반전되고, 서로 적어도 95% 동일하고, 적어도 96% ... 97% ... 98% ... 99% .... 99.5% 및 그 사이의 모든 포인트 동일하고, 하나의 WT-ITR은 5'-GCGCGCTCGCTCGCTC-3'(서열 번호: 531)의 Rep-결합 부위 (RBS) 및 말단 분해 부위 (trs)를 보유한다. 일부 구현예에서, 실질적으로 대칭인 WT-ITR 쌍은 서로 반전되고, 서로 적어도 95% 동일하고, 적어도 96% ... 97% ... 98% ... 99% .... 99.5% 및 그 사이의 모든 포인트 동일하고, 하나의 WT-ITR은 헤어핀 이차 구조 형성을 허용하는 가변 팔린드롬 서열에 추가하여 5'-GCGCGCTCGCTCGCTC-3' (서열 번호: 531)의 Rep-결합 부위 (RBS) 및 말단 분해 부위 (trs)를 보유한다. 상동성은 디폴트 설정에서 BLAST (기본 지역 정렬 검색 도구), BLASTN과 같은 당 업계에 잘 알려진 표준 수단에 의해 결정될 수 있다.
일부 구현예에서, ITR의 구조적 요소는 큰 Rep 단백질 (예를 들어, Rep 78 또는 Rep 68)과 ITR의 기능적 상호 작용에 관여하는 임의의 구조적 요소일 수 있다. 특정 구현예에서, 구조적 요소는 큰 Rep 단백질과 ITR의 상호 작용에 대한 선택성을 제공하고, 즉, Rep 단백질이 ITR과 기능적으로 상호 작용하는 것을 적어도 부분적으로 결정한다. 다른 구현예에서, Rep 단백질이 ITR에 결합될 때 구조적 요소는 큰 Rep 단백질과 물리적으로 상호 작용한다. 각각의 구조적 요소는, 예를 들어, ITR의 2차 구조, ITR의 뉴클레오티드 서열, 2개 이상의 요소 사이의 간격, 또는 임의의 상기의 조합일 수 있다. 일 구현예에서, 구조적 요소는 A 및 A' 아암, B 및 B' 아암, C 및 C' 아암, D 아암, Rep 결합 부위 (RBE) 및 RBE' (즉, 상보적인 RBE 서열) 및 말단 분해 부위 (trs)로 구성된 군으로부터 선택된다.
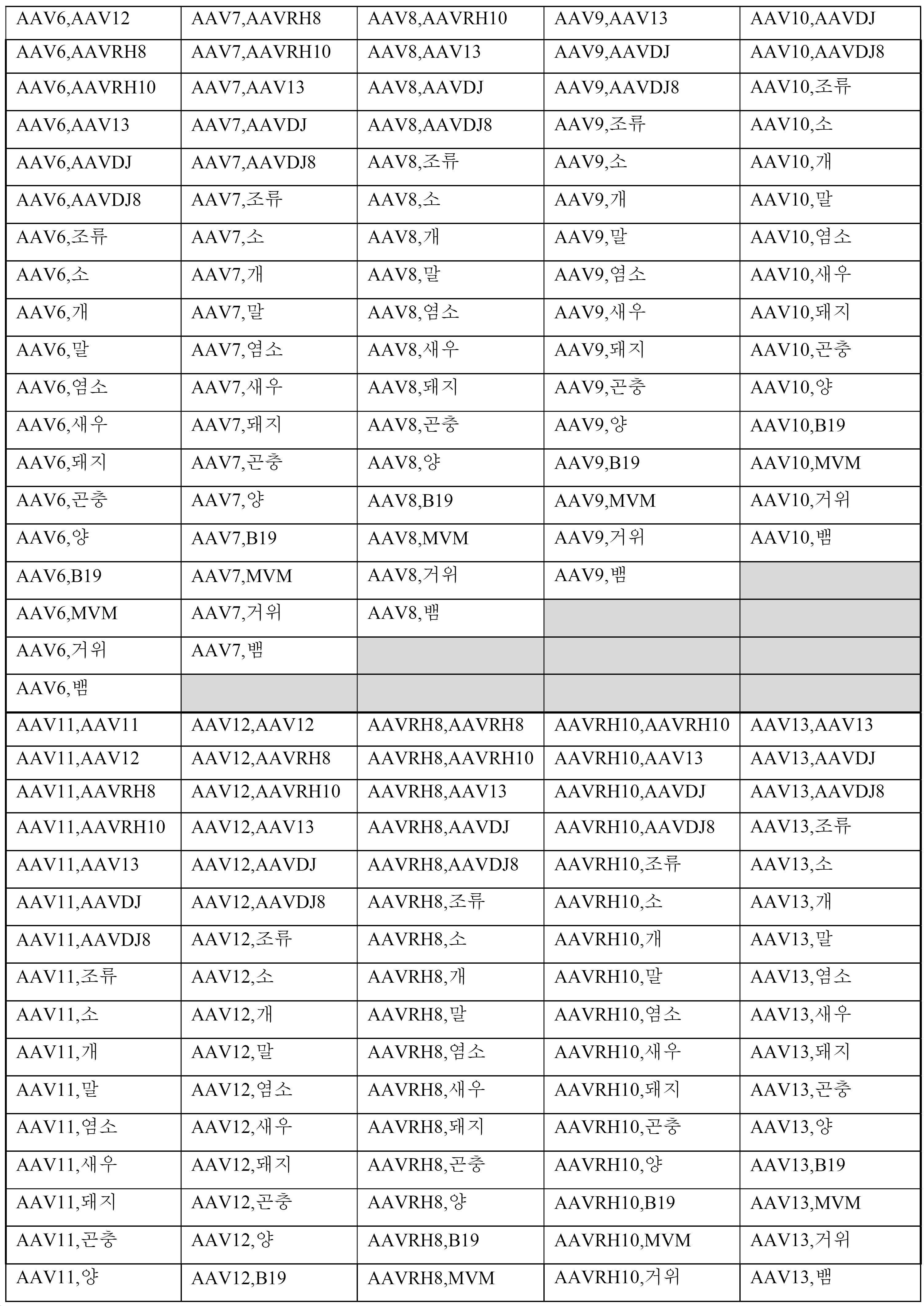
단지 예로서, 표 1은 WT-ITR의 예시적인 조합을 나타낸다.
표 1: 동일한 혈청형 또는 상이한 혈청형 또는 상이한 파로바이러스로부터의 WT-ITR의 예시적인 조합. 표시된 순서는 ITR 위치를 나타내지 않으며, 예를 들어, "AAV1, AAV2"는 ceDNA가 5' 위치에 WT-AAV1 ITR을, 3' 위치에 WT-AAV2 ITR을 포함할 수 있고, 반대로, 5' 위치에 WT-AAV2 ITR 및 3' 위치에 WT-AAV1 ITR을 포함할 수 있음을 나타낸다. 약어: AAV 혈청형 1 (AAV1), AAV 혈청형 2 (AAV2), AAV 혈청형 3 (AAV3), AAV 혈청형 4 (AAV4), AAV 혈청형 5 (AAV5), AAV 혈청형 6 (AAV6), AAV 혈청형 7 (AAV7) AAV 혈청형 8 (AAV8), AAV 혈청형 9 (AAV9), AAV 혈청형 10 (AAV10), AAV 혈청형 11 (AAV11) 또는 AAV 혈청형 12 (AAV12); AAVrh8, AAVrh10, AAV-DJ 및 AAV-DJ8 게놈 (예를 들어, NCBI: NC 002077; NC 001401; NC001729; NC001829; NC006152; NC 006260; NC 006261), 온혈 동물로부터의 ITR (조류 AAV (AAAV), 소 AAV (BAAV), 개, 말 및 양 AAV), B19 파보비리스로부터의 ITR (GenBank 수탁 번호: NC 000883), 마우스로부터의 미세 바이러스 (MVM) (GenBank 수탁 번호 NC 001510); 거위: 거위 파보바이러스 (GenBank 수탁 번호 NC 001701); 뱀: 뱀 파보바이러스 1 (GenBank 수탁 번호 NC 006148).
[표 1]
단지 예로서, 표 2는 일부 상이한 AAV 혈청형으로부터의 예시적인 WT-ITR의 서열을 보여준다.
[표 2]
일부 구현예에서, WT-ITR 서열의 뉴클레오티드 서열은 (예를 들어, 1, 2, 3, 4 또는 5개 이상의 뉴클레오티드 또는 그 안의 임의의 범위의 뉴클레오티드를 변형시킴으로써) 변형될 수 있는데, 변형은 상보적 뉴클레오티드에 대한 치환이고, 예를 들어, C 대신 G, 및 그 반대, 및 A 대신 T, 및 그 반대이다.
본 발명의 특정 구현예에서, ceDNA 벡터는 서열 번호: 550-557 중 임의의 것으로부터 선택된 뉴클레오티드 서열로 구성된 WT-ITR을 갖지 않는다.
본 발명의 대안의 구현예에서, ceDNA 벡터가 서열 번호: 550-557 중 임의의 것으로부터 선택된 뉴클레오티드 서열을 포함하는 WT-ITR을 갖는 경우, 플랭킹 ITR은 또한 WT이고, cDNA는, 예를 들어, 본원 및 PCT/US18/49996 (예를 들어, PCT/US18/49996의 표 11 참조)에 개시된 바와 같은 조절 스위치를 포함한다. 일부 구현예에서, ceDNA 벡터는 본원에 개시된 바와 같은 조절 스위치 및 서열 번호: 550-557로 구성된 군 중 어느 것으로부터 선택된 뉴클레오티드 서열을 갖는 선택된 WT-ITR을 포함한다.
본원에 기재된 ceDNA 벡터는 작동 가능한 RBE, trs 및 RBE' 부분을 보유하는 WT-ITR 구조를 포함할 수 있다. 도 2a 및 도 2b는 예시적인 목적으로 야생형 ITR을 사용하여 ceDNA 벡터의 야생형 ITR 구조 부분 내에서 trs 부위의 작동을 위한 하나의 가능한 메커니즘을 보여준다. 일부 구현예에서, ceDNA 벡터는 Rep-결합 부위 (RBS; AAV2에 대해 5'-GCGCGCTCGCTCGCTC-3'(서열 번호: 531)) 및 말단 분해 부위 (TRS; 5'-AGTT (서열 번호: 46))를 포함하는 하나 이상의 기능성 WT-ITR 폴리뉴클레오티드 서열을 함유한다. 일부 구현예에서, 적어도 하나의 WT-ITR은 기능적이다. 대안적인 구현예에서, ceDNA 벡터가 서로 실질적으로 대칭인 2개의 WT-ITR을 포함하는 경우, 적어도 하나의 WT-ITR은 기능적이고 적어도 하나의 WT-ITR은 비-기능적이다.
B. 비대칭 ITR 쌍 또는 대칭 ITR 쌍을 포함하는 ceDNA 벡터에 대한 일반적으로 변형된 ITR (모드-ITR)
본원에서 논의된 바와 같이, ceDNA 벡터는 대칭 ITR 쌍 또는 비대칭 ITR 쌍을 포함할 수 있다. 두 경우 모두에서, ITR은 변형된 ITR일 수 있으며, 차이점은 첫 번째 경우 (즉, 대칭 모드-ITR), 모드-ITR은 동일한 3차원 공간 구성을 가지며 (즉, 동일한 A-A', C-C' 및 B-B' 아암 구성를 가짐), 반면 두 번째 경우 (즉, 비대칭 모드-ITR), 모드-ITR은 다른 3차원 공간 구성을 갖는다 (즉, A-A', C-C' 및 B-B' 아암의 다른 구성을 가짐).
일부 구현예에서, 변형된 ITR은 야생형 ITR 서열 (예를 들어 AAV ITR)과 비교하여 결실, 삽입 및/또는 치환에 의해 변형된 ITR이다. 일부 구현예에서, ceDNA 벡터의 ITR 중 적어도 하나는 기능적 Rep 결합 부위 (RBS; 예를 들어, AAV2에 대해 5'-GCGCGCTCGCTCGCTC-3', 서열 번호: 531) 및 기능성 말단 분해 부위 (TRS; 5'-AGTT-3', 서열 번호: 46)를 포함한다. 일 구현예에서, ITR 중 적어도 하나는 비-기능적 ITR이다. 일 구현예에서, 상이한 또는 변형된 ITR은 각각 상이한 혈청형으로부터의 야생형 ITR이 아니다.
ITR에서의 특정 변경 및 돌연변이는 본원에 상세하게 기술되어 있지만, ITR의 맥락에서, "변경된" 또는 "돌연변이된" 또는 "변형된"은, 뉴클레오티드가 야생형, 참조 또는 원래 ITR 서열에 대해 삽입, 결실 및/또는 치환된 것을 나타낸다. 변경된 또는 돌연변이된 ITR은 조작된 ITR일 수 있다. 본원에 사용된 "조작된"은 인간의 손에 의해 조작된 측면을 지칭한다. 예를 들어, 폴리펩티드의 적어도 하나의 측면, 예를 들어, 이의 서열이 사람의 손에 의해 조작되어 자연에 존재하는 측면과 상이할 때 폴리펩티드가 "조작된" 것으로 간주된다.
일부 구현예에서, 모드-ITR은 합성일 수 있다. 일 구현예에서, 합성 ITR은 하나 초과의 AAV 혈청형으로부터의 ITR 서열에 기초한다. 또 다른 구현예에서, 합성 ITR은 AAV 기반 서열을 포함하지 않는다. 또 다른 구현예에서, 합성 ITR은 AAV에서 얻어진 서열의 일부만을 갖거나 갖지 않더라도 상기 기술된 ITR 구조를 보존한다. 일부 측면에서, 합성 ITR은 야생형 Rep 또는 특정 혈청형의 Rep와 우선적으로 상호 작용할 수 있거나, 일부 경우 야생형 Rep에 의해 인식되지 않고 돌연변이된 Rep에 의해서만 인식될 것이다.
당업자는 공지된 수단에 의해 다른 혈청형에서 상응하는 서열을 결정할 수 있다. 예를 들어, 변화가 A, A', B, B', C, C' 또는 D 영역에 있는지 확인하고 또 다른 혈청형에서 해당 영역을 결정한다. BLAST® (기본 지역 정렬 검색 도구) 또는 다른 상동성 정렬 프로그램을 디폴트 상태로 사용하여 상응하는 서열을 결정할 수 있다. 본 발명은 또한 상이한 AAV 혈청형의 조합으로부터의 모드-ITR을 포함하는 ceDNA 벡터의 집단 및 복수의 ceDNA 벡터를 제공하고, 즉, 하나의 모드-ITR은 하나의 AAV 혈청형으로부터 유래될 수 있고, 다른 모드-ITR은 상이한 혈청형으로부터 유래될 수 있다. 이론에 구속되지 않고, 일 구현예에서 하나의 ITR은 AAV2 ITR 서열로부터 유래하거나 이를 기초로 할 수 있고, ceDNA 벡터의 다른 ITR은 AAV 혈청형 1 (AAV1), AAV 혈청형 4 (AAV4), AAV 혈청형 5 (AAV5), AAV 혈청형 6 (AAV6), AAV 혈청형 7 (AAV7), AAV 혈청형 8 (AAV8), AAV 혈청형 9 (AAV9), AAV 혈청형 10 (AAV10) , AAV 혈청형 11 (AAV11), 또는 AAV 혈청형 12 (AAV12)의 임의의 하나 이상의 ITR 서열로부터 유래하거나 이를 기초로 할 수 있다.
임의의 파보바이러스 ITR은 변형을 위해 ITR 또는 기본 ITR로 사용될 수 있다. 바람직하게는, 파보바이러스는 데펜도바이러스이다. 보다 바람직하게는 AAV. 선택된 혈청형은 혈청형의 조직 향성(tropism)에 기초할 수 있다. AAV2는 광범위한 조직 향성을 가지며, AAV1은 뉴런 및 골격근을 우선적으로 표적화하고, AAV5는 뉴런, 망막 색소 상피 및 광수용체를 우선적으로 표적화한다. AAV6은 골격근 및 폐를 우선적으로 표적화한다. AAV8은 간, 골격근, 심장 및 췌장 조직을 우선적으로 표적화한다. AAV9는 간, 골격 및 폐 조직을 우선적으로 표적화한다. 일 구현예에서, 변형된 ITR은 AAV2 ITR에 기초한다.
보다 구체적으로, 구조적 요소가 특정한 큰 Rep 단백질과 기능적으로 상호 작용하는 능력은 구조적 요소를 변형시킴으로써 변경될 수 있다. 예를 들어, 구조적 요소의 뉴클레오티드 서열은 ITR의 야생형 서열과 비교하여 변형될 수 있다. 일 구현예에서, ITR의 구조적 요소 (예를 들어, A 아암, A' 아암, B 아암, B' 아암, C 아암, C' 아암, D 아암, RBE, RBE' 및 trs)는 제거되고 다른 파보바이러스로부터의 야생형 구조적 요소로 대체될 수 있다. 예를 들어, 대체 구조는 AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12, AAV13, 뱀 파보바이러스 (예를 들어, 로얄 피톤 파보바이러스), 소 파보바이러스, 염소 파보바이러스, 조류 파보바이러스, 개 파보바이러스, 말 파보바이러스, 새우 파보바이러스, 돼지 파보바이러스 또는 곤충 AAV로부터 유래될 수 있다. 예를 들어, ITR은 AAV2 ITR일 수 있고, A 또는 A' 아암 또는 RBE는 AAV5로부터의 구조적 요소로 대체될 수 있다. 또 다른 예에서, ITR은 AAV5 ITR일 수 있고, C 또는 C' 아암, RBE, 및 trs는 AAV2로부터의 구조적 요소로 대체될 수 있다. 또 다른 예에서, AAV ITR은 AAV2 ITR B 및 B' 아암으로 대체된 B 및 B' 아암을 갖는 AAV5 ITR일 수 있다.
단지 예로서, 표 3은 변형된 ITR의 영역에서 적어도 하나의 뉴클레오티드의 예시적인 변형 (예를 들어, 결실, 삽입 및/또는 치환)을 나타내며, 여기서 X는 상응하는 야생형 ITR에 대한 해당 섹션에서의 적어도 하나의 핵산의 변형 (예를 들어, 결실, 삽입 및/또는 치환)을 나타낸다. 일부 구현예에서, C 및/또는 C' 및/또는 B 및/또는 B'의 임의의 영역에서 적어도 하나의 뉴클레오티드의 임의의 변형 (예를 들어, 결실, 삽입 및/또는 치환)은 적어도 하나의 말단 루프에서 3개의 순차적인 T 뉴클레오티드 (즉, TTT)를 보유한다. 예를 들어, 변형 결과, 단일 아암 ITR (예를 들어, 단일 C-C' 아암 또는 단일 B-B' 아암) 또는 변형된 C-B' 아암 또는 C'-B 아암, 또는 적어도 하나의 절단된 아암 (예를 들어, 절단된 C-C' 아암 및/또는 절단된 B-B' 아암)을 갖는 2개의 아암 ITR 중 어느 것이 된다면, 적어도 하나의 단일 아암 또는 2개의 아암 ITR 중 적어도 하나의 아암 (하나의 아암이 절단될 수 있는 경우)은 적어도 하나의 말단 루프에서 순차적인 T 뉴클레오티드 (즉, TTT)를 보유한다. 일부 구현예에서, 절단된 C-C' 아암 및/또는 절단된 B-B' 아암은 말단 루프에서 3개의 순차적인 T 뉴클레오티드 (즉, TTT)를 갖는다.
[표 3]
ITR의 상이한 B-B' 및 C-C' 영역 또는 아암에 대한 적어도 하나의 뉴클레오티드 변형 (예를 들어, 결실, 삽입 및/또는 치환)의 예시적인 조합 (X는 뉴클레오티드 변형, 예를 들어, 상기 영역에서 적어도 하나의 뉴클레오티드의 첨가, 결실 또는 치환을 나타냄).
일부 구현예에서, 비대칭 ITR 쌍, 또는 본원에 개시된 바와 같은 대칭 모드-ITR 쌍을 포함하는 유전자 편집 ceDNA 벡터에 사용하기 위한 모드-ITR은 표 3에 나타낸 변형, 및 또한 A'와 C 사이, C와 C' 사이, C'와 B 사이, B와 B' 사이, 및 B'와 A 사이에서 선택된 영역 중 어느 하나 이상에서 적어도 하나의 뉴클레오티드의 변형의 조합 중 어느 하나를 포함할 수 있다. 일부 구현예에서, C 또는 C' 또는 B 또는 B' 영역에서 적어도 하나의 뉴클레오티드의 임의의 변형 (예를 들어, 결실, 삽입 및/또는 치환)은 여전히 스템-루프의 말단 루프를 보존한다. 일부 구현예에서, C 및 C' 및/또는 B 및 B' 사이의 적어도 하나의 뉴클레오티드의 임의의 변형 (예를 들어, 결실, 삽입 및/또는 치환)은 적어도 하나의 말단 루프에 3개의 순차적인 T 뉴클레오티드 (즉, TTT)를 보유한다. 대안적인 구현예에서, C 및 C' 및/또는 B 및 B' 사이의 적어도 하나의 뉴클레오티드의 임의의 변형 (예를 들어, 결실, 삽입 및/또는 치환)은 적어도 하나의 말단 루프에 3개의 순차적인 A 뉴클레오티드 (즉, AAA)를 보유한다 일부 구현예에서, 본원에 사용하기 위한 변형된 ITR은 표 3에 나타낸 변형, 및 또한 A', A 및/또는 D로부터 선택된 영역 중 임의의 하나에서 적어도 하나의 뉴클레오티드의 변형 (예를 들어, 결실, 삽입 및/또는 치환)의 조합 중 임의의 하나를 포함할 수 있다. 예를 들어, 일부 구현예에서, 본원에 사용하기 위한 변형된 ITR은 표 3에 나타낸 변형, 및 또한 A 영역에서 적어도 하나의 뉴클레오티드 변형 (예를 들어, 결실, 삽입 및/또는 치환)의 조합 중 임의의 하나를 포함할 수 있다. 일부 구현예에서, 본원에 사용하기 위한 변형된 ITR은 표 3에 나타낸 변형, 및 또한 A' 영역에서 적어도 하나의 뉴클레오티드 변형 (예를 들어, 결실, 삽입 및/또는 치환)의 조합 중 임의의 하나를 포함할 수 있다. 일부 구현예에서, 본원에 사용하기 위한 변형된 ITR은 표 3에 나타낸 변형, 및 또한 A 및/또는 A' 영역에서 적어도 하나의 뉴클레오티드 변형 (예를 들어, 결실, 삽입 및/또는 치환)의 조합 중 임의의 하나를 포함할 수 있다. 일부 구현예에서, 본원에 사용하기 위한 변형된 ITR은 표 3에 나타낸 변형, 및 또한 D 영역에서 적어도 하나의 뉴클레오티드 변형 (예를 들어, 결실, 삽입 및/또는 치환)의 조합 중 임의의 하나를 포함할 수 있다.
일 구현예에서, 구조적 요소의 뉴클레오티드 서열을 (예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 또는 20개 이상의 뉴클레오티드 또는 그 안의 임의의 범위의 뉴클레오티드를 변형함으로써) 변형시켜 변형된 구조적 요소를 생성할 수 있다. 일 구현예에서, ITR에 대한 특정 변형은 본원에 예시되어 있거나 (예를 들어, 서열 번호: 2, 52, 63, 64, 99-100, 469-499, 또는 본원의 도 7a-7b (예를 들어, 97-98, 101-103, 105-108, 111-112, 117-134, 545-54)에 도시되어 있다. 일부 구현예에서, ITR은 (예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 또는 20개 이상의 뉴클레오티드 또는 그 안의 임의의 범위의 뉴클레오티드를 변형시킴으로써) 변형될 수 있다. 다른 구현예에서, ITR은 서열 번호: 469-499 또는 545-547의 변형된 ITR, 또는 서열 번호: 97-98, 101-103, 105-108, 111-112, 117-134, 545-547의 A-A' 아암 및 C-C' 및 B-B' 아암의 RBE-함유 섹션, 또는 PCT/US18/49996 (이는 그 전문이 본원에 참고로 포함됨)의 표 2-9 (즉, 서열 번호: 110-112, 115-190, 200-468)에 나타낸 것 중 하나와 적어도 80%, 적어도 85%, 적어도 90%, 적어도 95%, 적어도 96%, 적어도 97%, 적어도 98%, 적어도 99% 또는 그 이상의 서열 동일성을 가질 수 있다.
일부 구현예에서, 변형된 ITR은 예를 들어 특정 아암의 전부, 예를 들어, A-A' 아암의 전부 또는 일부, 또는 B-B' 아암의 전부 또는 일부 또는 C-C' 아암의 전부 또는 일부의 제거 또는 결실, 또는 대안적으로, 스템 (예를 들어, 단일 아암)을 캡핑하는 최종 루프가 존재하는 한 루프의 스템을 형성하는 1, 2, 3, 4, 5, 6, 7, 8, 9개 이상의 염기쌍의 제거를 포함할 수 있다 (예를 들어, 도 7a의 ITR-21 참조). 일부 구현예에서, 변형된 ITR은 B-B' 아암으로부터 1, 2, 3, 4, 5, 6, 7, 8, 9개 이상의 염기쌍의 제거를 포함할 수 있다. 일부 구현예에서, 변형된 ITR은 C-C' 아암으로부터 1, 2, 3, 4, 5, 6, 7, 8, 9개 이상의 염기쌍의 제거를 포함할 수 있다 (예를 들어, 도 3b의 ITR-1 또는 도 7a의 ITR-45 참조). 일부 구현예에서, 변형된 ITR은 C-C' 아암으로부터 1, 2, 3, 4, 5, 6, 7, 8, 9개 이상의 염기쌍의 제거 및 B-B' 아암으로부터 1, 2, 3, 4, 5, 6, 7, 8, 9개 이상의 염기쌍의 제거를 포함할 수 있다. 염기쌍의 제거의 임의의 조합이 구상되며, 예를 들어, C-C' 아암에서 6개의 염기쌍이 제거될 수 있고, B-B' 아암에서 2개의 염기쌍이 제거될 수 있다. 예시적인 예로서, 도 3b는 각각의 C 부분 및 C' 부분으로부터 결실된 적어도 7개의 염기쌍, C 및 C' 영역 사이의 루프에서 뉴클레오티드의 치환, 및 변형된 ITR이 적어도 하나의 아암 (예를 들어, C-C')이 절단된 2개의 아암을 포함하도록 B 영역 및 B' 영역 각각으로부터 적어도 하나의 염기쌍 결실을 갖는 예시적인 변형된 ITR을 보여준다. 일부 구현예에서, 변형된 ITR은 또한, B-B' 아암이 WT ITR에 대해 또한 절단되도록 B 영역 및 B' 영역 각각으로부터 적어도 하나의 염기쌍 결실을 포함한다.
일부 구현예에서, 변형된 ITR은 전장 야생형 ITR 서열에 대해 1 내지 50개 (예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 또는 50개)의 뉴클레오티드 결실을 가질 수 있다. 일부 구현예에서, 변형된 ITR은 전장 WT ITR 서열에 대해 1 내지 30개의 뉴클레오티드 결실을 가질 수 있다. 일부 구현예에서, 변형된 ITR은 전장 야생형 ITR 서열에 대해 2 내지 20개의 뉴클레오티드 결실을 갖는다.
일부 구현예에서, 변형된 ITR은 A 또는 A' 영역의 RBE-함유 부분에서 임의의 뉴클레오티드 결실을 함유하지 않아서 DNA 복제 (예를 들어, Rep 단백질에 의한 RBE에의 결합 또는 말단 분해 부위에서의 닉킹)를 방해하지 않는다. 일부 구현예에서, 본원에 사용하기 위해 포함된 변형된 ITR은 본원에 기재된 바와 같이 B, B', C 및/또는 C 영역에서 하나 이상의 결실을 갖는다.
일부 구현예에서, 대칭 ITR 쌍 또는 비대칭 ITR 쌍을 포함하는 유전자 편집 ceDNA 벡터는 본원에 개시된 바와 같은 조절 스위치 및 서열 번호: 550 -557로 구성된 군 중 어느 것으로부터 선택된 뉴클레오티드 서열을 갖는 선택된 적어도 하나의 변형된 ITR을 포함한다.
또 다른 구현예에서, 구조적 요소의 구조는 변형될 수 있다. 예를 들어, 구조적 요소는 스템의 높이 및/또는 루프에서 뉴클레오티드 수의 변화이다. 예를 들어, 스템의 높이는 약 2, 3, 4, 5, 6, 7, 8 또는 9개 이상의 뉴클레오티드 또는 그 안의 임의의 범위일 수 있다. 일 구현예에서, 스템 높이는 약 5개의 뉴클레오티드 내지 약 9개의 뉴클레오티드일 수 있고, Rep와 기능적으로 상호 작용할 수 있다. 또 다른 구현예에서, 스템 높이는 약 7개의 뉴클레오티드일 수 있고, Rep와 기능적으로 상호 작용할 수 있다. 또 다른 예에서, 루프는 3, 4, 5, 6, 7, 8, 9 또는 10개 이상의 뉴클레오티드 또는 그 안의 임의의 범위를 가질 수 있다.
또 다른 구현예에서, RBE 또는 확장된 RBE 내의 GAGY 결합 부위 또는 GAGY 관련 결합 부위의 수는 증가 또는 감소될 수 있다. 일 예에서, RBE 또는 확장된 RBE는 1, 2, 3, 4, 5 또는 6개 이상의 GAGY 결합 부위 또는 그 안의 임의의 범위를 포함할 수 있다. 서열이 Rep 단백질에 결합하기에 충분하다면, 각각의 GAGY 결합 부위는 독립적으로 정확한 GAGY 서열 또는 GAGY와 유사한 서열일 수 있다.
또 다른 구현예에서, 두 요소 (예컨대 비제한적으로 RBE 및 헤어핀) 사이의 간격은 큰 Rep 단백질과의 기능적 상호 작용을 변경하기 위해 변경 (예를 들어, 증가 또는 감소)될 수 있다. 예를 들어, 간격은 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 또는 21개 이상의 뉴클레오티드 또는 그 안의 임의의 범위일 수 있다.
본원에 기재된 ceDNA 벡터는 본원에 개시된 야생형 AAV2 ITR 구조와 관련하여 변형되지만, 여전히 작동 가능한 RBE, trs 및 RBE' 부분을 보유하는 ITR 구조를 포함할 수 있다. 도 2a 및 도 2b는 ceDNA 벡터의 야생형 ITR 구조 부분 내에서 trs 부위의 작동을 위한 하나의 가능한 메커니즘을 도시한다. 일부 구현예에서, ceDNA 벡터는 Rep-결합 부위 (RBS; AAV2에 대해 5'-GCGCGCTCGCTCGCTC-3' (서열 번호: 531)) 및 말단 분해 부위 (TRS; 5'-AGTT (서열 번호: 46))를 포함하는 하나 이상의 기능적 ITR 폴리뉴클레오티드 서열을 함유한다. 일부 구현예에서, 적어도 하나의 ITR (wt 또는 변형된 ITR)은 기능적이다. 대안적인 구현예에서, ceDNA 벡터가 서로 상이하거나 비대칭인 2개의 변형된 ITR을 포함하는 경우, 적어도 하나의 변형된 ITR은 기능적이고 적어도 하나의 변형된 ITR은 비-기능적이다.
일부 구현예에서, ceDNA 벡터는 본원에 제공된 바와 같이, 서열 번호: 500-529로 구성되거나 본질적으로 구성된 임의의 서열로부터 선택된 변형된 ITR을 갖지 않는다. 일부 구현예에서, ceDNA 벡터는 서열 번호: 500-529로부터 선택된 임의의 서열로부터 선택된 ITR을 갖지 않는다.
일부 구현예에서, 본원에 기재된 ceDNA 벡터의 변형된 ITR (예를 들어, 좌측 또는 우측 ITR)은 루프 아암, 절단된 아암 또는 스페이서 내의 변형을 갖는다. 루프 아암, 절단된 아암 또는 스페이서 내의 변형을 갖는 ITR의 예시적인 서열은 PCT 출원 PCT/US18/49996 (이는 그 전문이 본원에 참고로 포함됨)의 표 2 (즉, 서열 번호: 135-190, 200-233); 표 3 (예를 들어, 서열 번호: 234-263); 표 4 (예를 들어, 서열 번호: 264-293); 표 5 (예를 들어, 본원의 서열 번호: 294-318); 표 6 (예를 들어, 서열 번호: 319-468; 및 표 7-9 (예를 들어, 서열 번호: 101-110, 111-112, 115-134) 또는 표 10a 또는 10b (예를 들어, 서열 번호: 9, 100, 469-483, 484-499)에 열거되어 있다.
일부 구현예에서, 비대칭 ITR 쌍 또는 대칭 모드-ITR 쌍을 포함하는 ceDNA 벡터에 사용하기 위한 변형된 ITR은 PCT 출원 PCT/US18/49996 (이는 그 전문이 본원에 참고로 포함됨)의 표 2, 3, 4, 5, 6, 7, 8, 9 및 10a-10b에 도시된 것들 중 임의의 것 또는 조합으로부터 선택된다.
상기 클래스 각각에서 비대칭 ITR 쌍, 또는 대칭 모드-ITR 쌍을 포함하는 ceDNA 벡터에 사용하기 위한 추가의 예시적인 변형된 ITR이 표 4a 및 4b에 제공된다. 표 4a에서 우측 변형된 ITR의 예측된 2차 구조는 도 7a에 도시되어 있고, 표 4b에서 좌측 변형된 ITR의 예측된 2차 구조는 도 7b에 도시되어 있다.
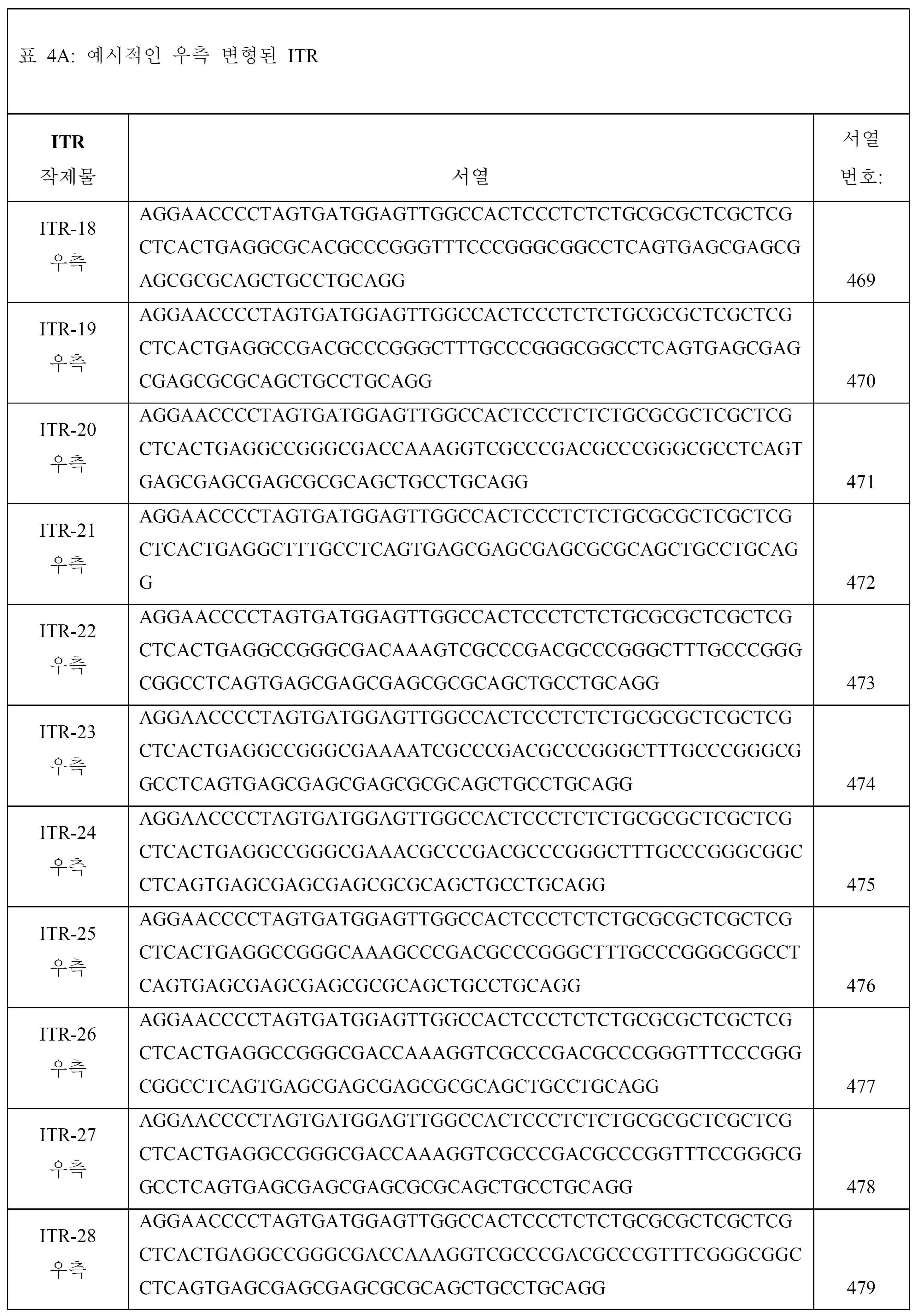
표 4a 및 표 4b는 예시적인 우측 및 좌측 변형된 ITR을 나타낸다.
표 4a: 예시적인 변형된 우측 ITR. 이러한 예시적인 변형된 우측 ITR은 GCGCGCTCGCTCGCTC-3'의 RBE (서열 번호: 531), ACTGAGGC의 스페이서 (서열 번호: 532), 스페이서 보체 GCCTCAGT (서열 번호: 535) 및 GAGCGAGCGAGCGCGC (서열 번호: 536)의 RBE' (즉, RBE에 대한 보체)를 포함할 수 있다.
[표 4a]
표 4b: 예시적인 변형된 좌측 ITR. 이러한 예시적인 변형된 좌측 ITR은 GCGCGCTCGCTCGCTC-3'의 RBE (서열 번호: 531), ACTGAGGC의 스페이서 (서열 번호: 532), 스페이서 보체 GCCTCAGT (서열 번호: 535) 및 GAGCGAGCGAGCGCGC의 RBE 보체 (RBE') (서열 번호: 536)를 포함할 수 있다.
[표 4b]
일 구현예에서, 유전자 편집 ceDNA 벡터는 2개의 대칭인 모드-ITR을 포함하며, 즉, 두 ITR은 동일한 서열을 갖지만, 서로의 역 보체 (역상)이다. 일부 구현예에서, 대칭 모드-ITR 쌍은 동일한 AAV 혈청형으로부터의 야생형 ITR 서열에 대한 결실, 삽입 또는 치환 중 적어도 하나 또는 임의의 조합을 포함한다. 대칭 ITR에서의 첨가, 결실 또는 치환은 동일하지만 서로의 역 보체이다. 예를 들어, 5' ITR의 C 영역에서 3개의 뉴클레오티드의 삽입은 3' ITR의 C' 영역에서의 상응하는 섹션에서 3개의 역 보체 뉴클레오티드의 삽입에 반영될 것이다. 단지 설명의 목적으로만, 5' ITR에서 첨가가 AACG인 경우, 상응하는 부위에서의 3' ITR에서 첨가는 CGTT이다. 예를 들어, 5' ITR 센스 가닥이 ATCGATCG인 경우, G와 A 사이에 AACG가 첨가되어 ATCG AACG ATCG 서열이 생성된다. 상응하는 3' ITR 센스 가닥은 CGATCGAT (ATCGATCG의 역 보체)이고, T와 C 사이에 CGTT (즉, AACG의 역 보체)를 첨가하여 CGAT CGTT CGAT (ATCG AACG ATCG의 역 보체) 서열이 생성된다.
대안적인 구현예에서, 변형된 ITR 쌍은 본원에 정의된 바와 같이 실질적으로 대칭이고, 즉, 변형된 ITR 쌍은 상이한 서열을 가질 수 있지만 대응하는 또는 동일한 대칭 3차원 형상을 가질 수 있다. 예를 들어, 하나의 변형된 ITR은 하나의 혈청형으로부터 유래될 수 있고, 다른 변형된 ITR은 상이한 혈청형으로부터 유래될 수 있지만, 이들은 동일한 영역에서 동일한 돌연변이 (예를 들어, 뉴클레오티드 삽입, 결실 또는 치환)를 갖는다. 달리 언급하면, 단지 설명의 목적으로, 5' 모드-ITR은 AAV2로부터 유래될 수 있고, C 영역에서 결실을 가질 수 있으며, 3' 모드-ITR은 AAV5로부터 유래될 수 있고, C' 영역에서 상응하는 결실을 가질 수 있으며, 5' 모드-ITR 및 3' 모드-ITR이 동일하거나 대칭인 3차원 공간 구성을 갖는 경우, 이들은 본 명세서에서 변형된 ITR 쌍으로 사용하기 위해 포함된다.
일부 구현예에서, 실질적으로 대칭인 모드-ITR 쌍은 3D 공간에서 동일한 A, C-C' 및 B-B' 루프를 가지며, 예를 들어, 실질적으로 대칭인 모드-ITR 쌍에서 변형된 ITR이 C-C' 아암의 결실을 갖는 경우, 동족 모드-ITR은 C-C' 루프의 상응하는 결실을 가지며 또한 이의 동족 모드-ITR의 기하학적 공간에서 동일한 형태로 나머지 A 및 B-B' 루프의 유사한 3D 구조를 갖는다. 단지 예로서, 실질적으로 대칭인 ITR은 그 구조가 기하학적 공간에서 동일한 형상이 되도록 대칭 공간 구성을 가질 수 있다. 이것은, 예를 들어, G-C 쌍이, 예를 들어, C-G 쌍으로 또는 그 반대로 변형될 때, 또는 A-T 쌍이 T-A 쌍으로 또는 그 반대로 변형될 때 발생할 수 있다. 따라서, ATCG AACG ATCG (서열 번호: 570)로서 변형된 5' ITR 및 CGAT CGTT CGAT (서열 번호: 571) (즉, ATCG AACG ATCG (서열 번호: 570)의 역 보체)로서 변형된 3' ITR의 상기 예시적인 예를 사용하면, 예를 들어, 5' ITR이 ATCG AAC C ATCG (서열 번호: 572)의 서열을 갖는 경우, 이러한 변형된 ITR은 여전히 대칭일 것이며, 여기서 첨가에서 G는 C로 변형되고, 실질적으로 대칭인 3' ITR은 A로의 첨가에서 T의 상응하는 변형 없이 CGATCGTT CGAT (서열 번호: 571)의 서열을 갖는다. 일부 구현예에서, 이러한 변형된 ITR 쌍은 변형된 ITR 쌍이 대칭 입체화학을 갖기 때문에 실질적으로 대칭이다.
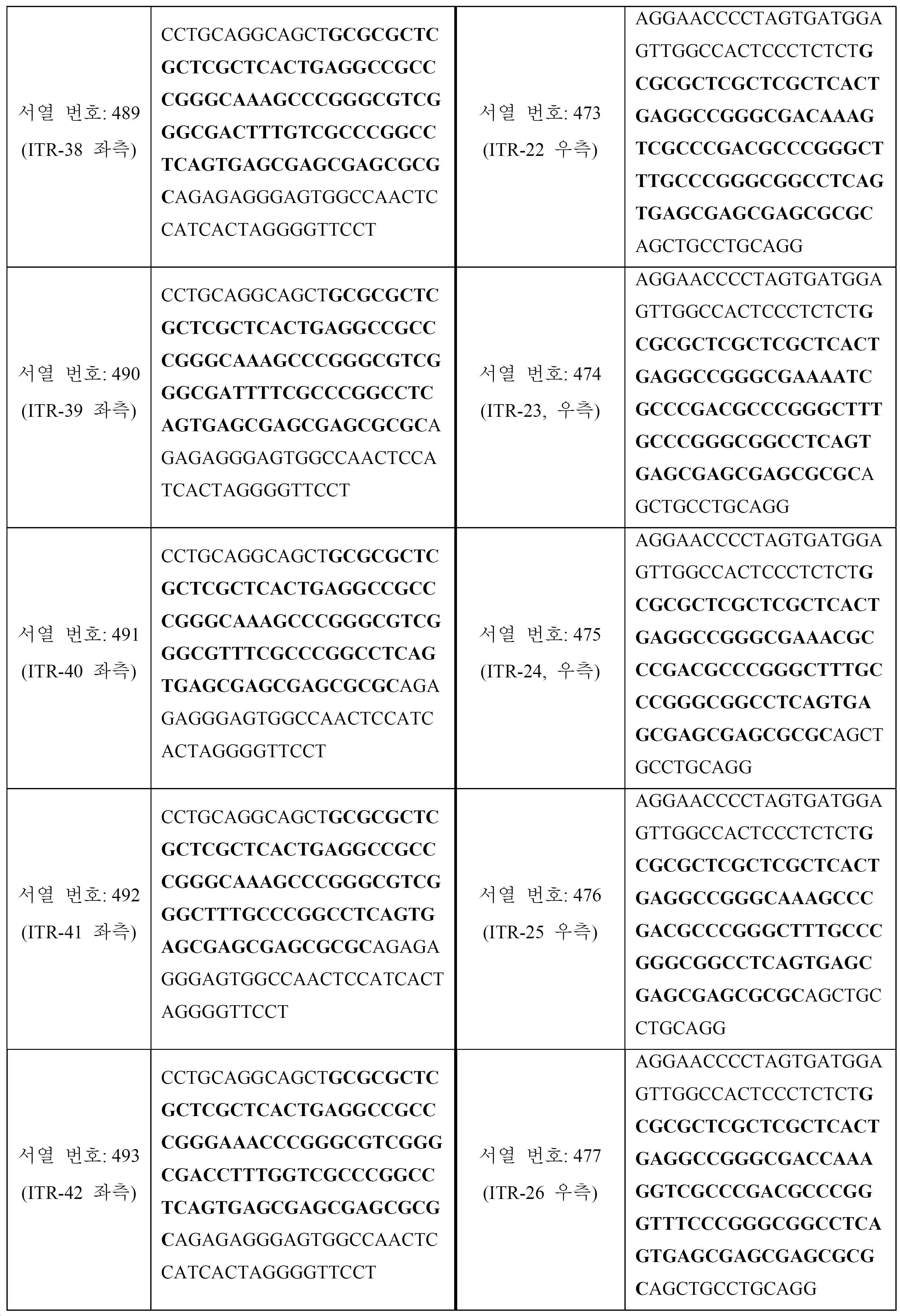
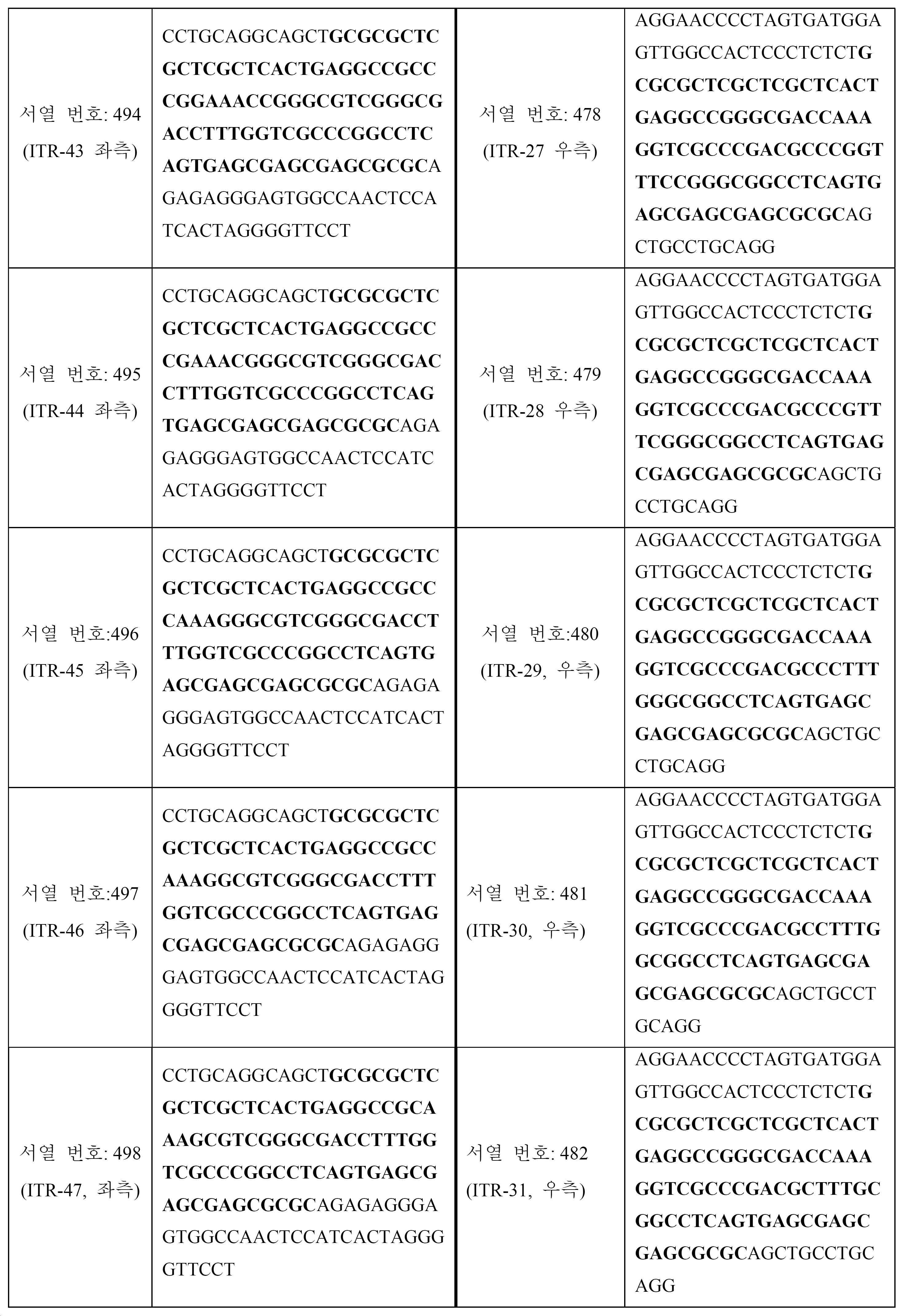
표 5는 예시적인 대칭 변형된 ITR 쌍 (즉, 좌측 변형된 ITR 및 대칭 우측 변형된 ITR)을 보여준다. 서열의 굵은 체 (빨간색) 부분은 도 31a-46b에 또한 도시된 부분 ITR 서열 (즉, A-A', C-C' 및 B-B' 루프의 서열)을 식별한다. 이러한 예시적인 변형된 ITR은 GCGCGCTCGCTCGCTC-3' (서열 번호: 531)의 RBE, ACTGAGGC (서열 번호: 532)의 스페이서, 스페이서 보체 GCCTCAGT (서열 번호: 535) 및 GAGCGAGCGAGCGCGC (서열 번호: 536)의 RBE' (즉, RBE에 대한 보체)를 포함할 수 있다.
[표 5]
일부 구현예에서, 비대칭 ITR 쌍을 포함하는 유전자 편집용 ceDNA 벡터는 본원의 표 4a-4b 중 임의의 하나 이상에 나타낸 ITR 서열 또는 ITR 부분 서열 또는 도 7a 또는 도 7b에 도시되거나, 또는 2018년 9월 7일에 출원된 PCT/US18/49996 (이는 그 전문이 본원에 참고로 포함됨)의 표 2, 3, 4, 5, 6, 7, 8, 9 또는 10a-10b에 개시된 서열에서의 임의의 변형에 상응하는 변형을 갖는 ITR을 포함할 수 있다.
VI. 예시적인 유전자 편집 ceDNA 벡터
상술한 바와 같이, 본 개시내용은 비대칭 ITR 쌍, 대칭 ITR 쌍, 또는 전술한 바와 같이 실질적으로 대칭인 ITR 쌍 중 임의의 하나를 포함하는 재조합 ceDNA 발현 벡터 (예를 들면, 공여체 벡터 (프로모터에 작동 가능하게 연결되거나 연결되지 않을 수 있음) 및 유전자 편집 분자를 인코딩하는 ceDNA 벡터)에 관한 것이다. 특정 구현예에서, 본 개시내용은 플랭킹 ITR 서열 및 유전자 편집 능력을 갖는 재조합 ceDNA 벡터에 관한 것으로, 여기서 ITR 서열은 본원에 정의된 바와 같이 서로 비대칭, 대칭 또는 실질적으로 대칭이며, ceDNA는 플랭킹 ITR 사이에 위치한 관심 뉴클레오티드 서열 (예를 들어 유전자 편집 서열의 발현 카세트, 또는 가이드 RNA)을 추가로 포함하며, 여기서 상기 핵산 분자는 바이러스 캡시드 단백질 코딩 서열이 없다.
일부 구현예에서, ceDNA 벡터는 뉴클레아제, 하나 이상의 상동성 아암, 가이드 RNA, 활성화제 RNA 및 제어 요소 중 적어도 하나를 포함한다. 일부 구현예에서, 5' 상동성 아암, 공여체 서열 및 3' 상동성 아암을 포함하는 폴리뉴클레오티드. 본 개시내용에 따른 적합한 ceDNA 벡터는 하기 실시예에 따라 수득될 수 있다. 특정 구현예에서, 본 개시내용은 유전자 편집 시스템의 적어도 2개의 성분, 예를 들어 CAS 및 적어도 하나의 gRNA, 또는 2개의 ZNF 등을 포함하는 재조합 ceDNA 발현 벡터에 관한 것이다. 따라서, 일부 구현예에서, ceDNA 벡터는 유전자 편집 시스템의 다수의 성분을 포함한다.
재조합 ceDNA 발현 벡터는 적어도 하나의 ITR이 변경된 경우 본원에 기재된 바와 같은 뉴클레오티드 서열(들)을 포함하는 재조합 DNA 절차에 편리하게 적용될 수 있는 임의의 ceDNA 벡터일 수 있다. 본 개시내용의 ceDNA 벡터는 ceDNA 벡터가 도입될 숙주 세포와 양립 가능하다. 특정 구현예에서, ceDNA 벡터는 선형일 수 있다. 특정 구현예에서, ceDNA 벡터는 염색체외 엔티티로서 존재할 수 있다. 특정 구현예에서, 본 개시내용의 ceDNA 벡터는 공여체 서열을 숙주 세포의 게놈으로 통합시킬 수 있는 요소(들)를 함유할 수 있다. 본원에 사용된 "공여체 서열" 및 "이식유전자" 및 "이종 뉴클레오티드 서열"은 동의어이다.
이제 도 1a-1g를 참조하면, 본 개시내용의 ceDNA 벡터를 제조하는데 유용한 2개의 비제한적인 플라스미드의 기능성 성분의 개략도가 도시되어 있다. 도 1a, 1b, 1d, 1f는 유전자 편집용 ceDNA 벡터의 작제물 또는 상응하는 ceDNA 플라스미드 서열을 보여준다. ceDNA 벡터는 캡시드가 없고, 제1 ITR, 발현 가능한 이식유전자 카세트 및 제2 ITR의 순서로 인코딩된 플라스미드로부터 수득될 수 있으며, 여기서 제1 및 제2 ITR 서열은 본원에 정의된 바와 같이 서로 비대칭이거나, 대칭이거나 실질적으로 대칭이다. ceDNA 벡터는 캡시드가 없고, 제1 ITR, 발현 가능한 이식유전자 (단백질 또는 핵산) 또는 공여체 카세트 (예를 들어 HDR 공여체) 및 제2 ITR의 순서로 인코딩된 플라스미드로부터 수득될 수 있으며, 여기서 제1 및 제2 ITR 서열은 본원에 정의된 바와 같이 서로 비대칭이거나, 대칭이거나 실질적으로 대칭이다. 일부 구현예에서, 발현 가능한 이식유전자 카세트는, 필요에 따라, 인핸서/프로모터, 하나 이상의 상동성 아암, 공여체 서열, 전사 후 조절 요소 (예를 들어, WPRE, 예를 들어, 서열 번호: 8)), 및 폴리아데닐화 및 종결 신호 (예를 들어, BGH 폴리A, 예를 들어, 서열 번호: 7)를 포함한다.
도 5는 실시예에 기재된 방법을 사용하여 다중 플라스미드 작제물로부터 ceDNA의 생성을 확인하는 겔이다. ceDNA는 상기 도 4a와 관련하여 그리고 실시예에서 논의된 바와 같이, 겔에서의 특징적인 밴드 패턴에 의해 확인된다.
이제 도 8을 참조하면, 제1 및 제2 ITR (여기서 ITR 서열은 본원에 정의된 바와 같이 서로 비대칭, 대칭 또는 실질적으로 대칭임), 5' 상동성 아암, 공여체 서열, 및 3' 상동성 아암을 포함하는 제1 뉴클레오티드 서열 (여기서, 공여체 서열은 유전자 편집 기능성을 가짐)을 포함하는 본 개시내용에 따른 비제한적인 예시적인 ceDNA 벡터가 도시된다. 일부 구현예에서, 상기 기재된 TR (예를 들어, ITR)은 관심 유전자 편집 분자 (예를 들어, 뉴클레아제 (예를 들어, 서열 특이적 뉴클레아제), 하나 이상의 가이드 RNA, Cas 또는 다른 리보핵단백질 (RNP), 또는 이들의 임의의 조합을 인코딩하는 핵산 서열의 플랭킹 말단에 포함된다. 본 개시내용의 핵산 작제물의 비제한적 예는 서열 번호: 1, 또는 서열 번호: 51의 뉴클레오티드 서열을 갖는 AAV2의 야생형 기능성 ITR을 포함하는 핵산 작제물, 및 추가로 서열 번호: 2 또는 서열 번호: 52의 뉴클레오티드 서열과 적어도 60%, 보다 바람직하게는 적어도 65%, 보다 바람직하게는 적어도 70%, 보다 바람직하게는 적어도 75%, 보다 바람직하게는 적어도 80%, 보다 바람직하게는 적어도 85%, 더욱 더 바람직하게는 적어도 90%, 가장 바람직하게는 적어도 95%의 서열 동일성을 갖는 AAV2의 변형된 ITR을 포함한다. 추가의 ITR은 WO 2017/152149 및 PCT 출원 PCT/US18/49996에 기재되어 있으며, 그 전문이 본원에 참고로 포함된다.
도 8을 참조하면, ceDNA는 도시된 바와 같이 제1 뉴클레오티드 서열의 상류에 있는 제2 뉴클레오티드 서열을 포함할 수 있다. 본원에 기재된 임의의 ceDNA 벡터의 특정 구현예에서, ceDNA 벡터는 공여체 서열 및, 임의로, 상동성 아암을 포함하는 제1 뉴클레오티드 서열의 이러한 제2 뉴클레오티드 서열 5' 또는 3'를 추가로 포함할 수 있다. 일부 구현예에서, 도 8을 참조하면, ceDNA 벡터는 가이드 서열 및/또는 활성화제 RNA 서열을 인코딩하는 하나 이상의 뉴클레오티드에 작동 가능하게 연결된 제2 프로모터를 포함하는 제3 뉴클레오티드 서열을 포함할 수 있다. 특정 구현예에서, 프로모터는 Pol III (U6 (서열 번호: 18) 또는 H1 (서열 번호: 19))이다.
또 다른 구현예에서, ceDNA 벡터는 비틀림 해제(torsional release) 및 보다 효율적인 상동성 지정 수리 (HDR)를 위해 각각의 ceDNA ITR로 향하거나 상동성 도메인 영역 외부로 향하는 하나 이상의 가이드 RNA 및 뉴클레아제를 인코딩한다. 뉴클레아제는 돌연변이 뉴클레아제일 필요는 없으며, 예를 들어 공여체 HDR 주형은 이러한 절단에 의해 ceDNA로부터 방출될 수 있다.
일부 구현예에서, 하나의 비제한적인 예에서, 유전자 편집용 ceDNA 벡터는 특정 유전자에 대한 5' 및 3' 상동성 아암, 또는 5' 상동성 및 3' 상동성 아암의 어느 말단에 본원에 기재된 엔도뉴클레아제에 특이적인 제한 부위를 갖는 표적 통합 부위를 포함할 수 있다. ceDNA 벡터가 제한 부위(들)에 특이적인 하나 이상의 제한 엔도뉴클레아제로 절단될 때, 생성된 카세트는 5' 상동성 아암-공여체 서열-3' 상동성 아암을 포함하고, 원하는 게놈 유전자좌와 보다 쉽게 재조합될 수 있다. 특정 측면에서, ceDNA 벡터 자체는 제한 엔도뉴클레아제를 인코딩하여 ceDNA 벡터를 핵으로 전달할 때 제한 엔도뉴클레아제가 발현되고 벡터를 절단할 수 있도록 한다. 특정 측면에서, 제한 엔도뉴클레아제는 별도로 전달되는 제2 ceDNA 벡터에 인코딩된다. 특정 측면에서, 제한 엔도뉴클레아제는 비-ceDNA-기반 전달 수단에 의해 핵에 도입된다. 따라서, 일부 구현예에서, 본원에 기재된 기술은 하나 초과의 유전자 편집 ceDNA가 대상체에게 전달되도록 한다. 본원에 논의된 바와 같이, 일 구현예에서, ceDNA는 특정 표적 유전자 또는 유전자좌를 표적화하는 공여체 서열의 측면에 상동성 아암을 가질 수 있고, 일부 구현예에서, 본원에 기재된 바와 같이, 게놈 DNA의 절단을 표적화하기 위한 하나 이상의 가이드 RNA (예를 들어, sgRNA)를 또한 포함할 수 있으며, 또 다른 ceDNA는 실제 유전자 편집 단계에 대해 본원에 기재된 바와 같이 뉴클레아제 효소 및 활성화제 RNA를 포함할 수 있다.
A. DNA 엔도뉴클레아제
본 개시내용의 ceDNA 벡터는 서열-특이적 뉴클레아제와 같은 뉴클레아제를 인코딩하는 뉴클레오티드 서열을 함유할 수 있다. 서열-특이적 또는 부위-특이적 뉴클레아제는 표적화된 게놈 유전자좌에서 부위-특이적 이중 가닥 파괴 또는 닉을 도입하는데 사용될 수 있다. 이 뉴클레오티드 절단, 예를 들어, DNA 또는 RNA 절단은 자연 복구 기구, 예를 들어, DNA 복구 기구를 자극하여 두 가지 가능한 복구 경로 중 하나를 유도한다. 공여체 주형이 없는 경우, DNA의 작은 삽입 또는 결실을 초래하는 오류가 발생하기 쉬운 복구 경로인 비-동종 말단 결합 (NHEJ)에 의해 파괴가 복구될 것이다 (예를 들어, Suzuki et al. Nature 540:144-149 (2016), 이의 내용은 그 전문이 참고로 포함됨). 이 방법은 표적화된 유전자 서열의 판독 프레임을 의도적으로 파괴, 결실 또는 변경하는데 사용될 수 있다. 그러나, 뉴클레아제 이외에 공여체 주형이 제공된다면, 세포 기구는 상동성 재조합 (HDR)에 의해 파괴를 복구할 것이며, 이는 DNA 절단의 존재하에 또는 NHEJ를 통한 공여체 주형의 삽입에 의해 10의 몇 승 정도 향상된다.
상기 방법은 표적 부위에서 DNA 서열의 특정 변화를 도입하는데 사용될 수 있다. 본원에 사용된 용어 "부위-특이적 뉴클레아제"는 특정 DNA 서열을 특이적으로 인식하고 절단할 수 있는 효소를 지칭한다. 부위-특이적 뉴클레아제는 조작될 수 있다. 조작된 부위-특이적 뉴클레아제의 예는 아연 핑거 뉴클레아제 (ZFN), TAL 이펙터 뉴클레아제 (TALEN), 메가뉴클레아제 및 CRISPR/Cas9-효소 및 조작된 유도체를 포함한다. 당업자라면 이해할 수 있는 바와 같이, 유전자 편집이 완료되면 엔도뉴클레아제가 일반적으로 더 이상 필요하지 않기 때문에 유전자 편집에 필요한 엔도뉴클레아제가 일시적으로 발현될 수 있다. 이러한 일시적 발현은 오프-표적 효과 및 면역원성에 대한 가능성을 감소시킬 수 있다. 일시적 발현은 당 업계에 공지된 임의의 수단에 의해 달성될 수 있으며, 본원에 기재된 바와 같은 조절 스위치를 사용하여 편리하게 수행될 수 있다.
일부 구현예에서, 뉴클레아제를 인코딩하는 뉴클레오티드 서열은 cDNA이다. 서열-특이적 뉴클레아제의 비제한적 예는 RNA-유도 뉴클레아제, 아연 핑거 뉴클레아제 (ZFN), 전사 활성화제-유사 이펙터 뉴클레아제 (TALEN) 또는 메가뉴클레아제를 포함한다. 적합한 RNA-유도 뉴클레아제의 비제한적인 예는 본원에 기재된 바와 같은 CRISPR 효소를 포함한다.
본원에 기재된 뉴클레아제는, 예를 들어, 서열 특이적 뉴클레아제를 설계하도록 조작될 수 있다 (예를 들어, 미국 특허 8,021,867 참조). 뉴클레아제는 예를 들어, 하기에 기재된 방법을 사용하여 설계될 수 있다: Certo, MT et al. Nature Methods (2012) 9:073-975; 미국 특허 번호 8,304,222; 8,021,867; 8,119,381; 8,124,369; 8,129,134; 8,133,697; 8,143,015; 8,143,016; 8,148,098; 또는 8,163,514 (각각의 내용은 그 전문이 본원에 참고로 포함됨). 대안적으로, 부위 특이적 절단 특성을 갖는 뉴클레아제는 상업적으로 이용 가능한 기술, 예를 들어, Precision BioSciences의 Directed Nuclease Editor™ 게놈 편집 기술을 사용하여 얻을 수 있다.
특정 구현예에서, 예를 들어 상동성 지정 복구 주형, 가이드 RNA 및/또는 Cas 효소, 또는 임의의 다른 뉴클레아제를 포함하는 프로모터가 없는 ceDNA 작제물을 사용하는 경우, 예를 들어 i) 가이드 RNA를 인코딩하는 핵산 ii) 또는 원하는 뉴클레아제, 예를 들어 Cas 효소 또는 다른 뉴클레아제를 인코딩하는 mRNA를 투여함으로써 iii) 또는 Cas 효소 및 가이드 RNA를 포함하는 리보뉴클레오티드 단백질 (RNP) 복합체를 투여함으로써, 또는 iv) 예를 들어, 벡터, 예를 들어 바이러스, 플라스미드 또는 또 다른 ceDNA 벡터에 의한 재조합 뉴클레아제 단백질의 전달에 의해 트랜스로 전달된다. 특정 측면에서, 트랜스로 전달된 분자는 제1 ceDNA 벡터에 순차적으로 공동 투여되거나 투여될 수 있는 하나 이상의 추가의 ceDNA 벡터에 의해 전달된다.
따라서, 일 구현예에서, ceDNA 벡터는 유도성 프로모터에 의해 전사적으로 조절되는 엔도뉴클레아제 (예를 들어, Cas9)를 포함할 수 있다. 일부 구현예에서, 엔도뉴클레아제는 별도의 ceDNA 벡터 상에 있으며, 이는 상동성 아암 및 공여체 서열을 포함하는 ceDNA로 대상체에게 투여될 수 있으며, 이는 임의로 가이드 RNA (sgRNA)를 또한 포함할 수 있다. 대안적인 구현예에서, 엔도뉴클레아제는 본원에 기재된 바와 같은 올인원(all-in-one) ceDNA 벡터 상에 존재할 수 있다.
일부 구현예에서, ceDNA는 프로모터의 제어하에 본원에 기재된 바와 같은 엔도뉴클레아제를 인코딩한다. 유도성 프로모터의 비제한적인 예는, 예를 들어, 알코올, 테트라사이클린, 스테로이드, 금속 및 발병-관련 단백질 (예를 들어, 살리실산, 에틸렌 및 벤조티아디아졸)의 존재 또는 부재에 의해 전사 활성을 조절하는 화학적으로 조절된 프로모터, 및 예를 들어, 빛의 존재 또는 부재, 및 저온 또는 고온에 의해 전사 활성을 조절하는 물리적으로 조절되는 프로모터를 포함한다. 유도성 프로모터의 조절은 ceDNA 벡터의 유전자 편집 활성을 끄거나 켤 수 있게 한다. 유도성 Cas9 프로모터는, 예를 들어, 하기에서 추가로 검토된다: Cao J., et al. Nucleic Acids Research. 44(19)2016, 및 Liu KI, et al. Nature Chemical Biol. 12: 90-987 (2016) (이들은 그 전문이 본원에 포함됨).
일 구현예에서, 본원에 기재된 ceDNA 벡터는 제1 엔도뉴클레아제 (예를 들어, Cas9)의 활성을 일시적으로 표적화하고 억제하는 제2 엔도뉴클레아제를 추가로 포함한다. 다른 엔도뉴클레아제의 활성을 표적화하고 억제하는 엔도뉴클레아제는 당업자에 의해 결정될 수 있다. 또 다른 구현예에서, 본원에 기재된 ceDNA 벡터는 "항-CRISPR 유전자" (예를 들어, L. 모노사이토제네스(L. monocytogenes) ArcIIa)의 일시적 발현을 추가로 포함한다. 본원에 사용된 "항-CRISPR 유전자"는 일반적으로 사용되는 S. 파이오게네스 Cas9를 억제하는 것으로 나타난 유전자를 지칭한다. 또 다른 구현예에서, 제1 엔도뉴클레아제 활성, 또는 항-CRISPR 유전자의 활성을 표적화하고 억제하는 제2 엔도뉴클레아제는 원하는 유전자 편집이 완료된 후 투여되는 제2 ceDNA 벡터에 포함된다. 대안적으로, 제2 엔도뉴클레아제는 관심 유전자, 예를 들어, 본원에 기재된 ceDNA 벡터에 의해 전사적으로 증강된 유전자를 표적화하고 억제한다.
본원에 기재된 ceDNA 벡터 또는 이의 조성물은 표적 유전자를 활성화시키는 전사 활성화제를 인코딩하는 뉴클레오티드 서열을 포함할 수 있다. 예를 들어, 전사 활성화제는 조작될 수 있다. 예를 들어, 조작된 전사 활성화제는 CRISPR/Cas9-기반 시스템, 아연 핑거 융합 단백질 또는 TALE 융합 단백질일 수 있다. 상기 기재된 바와 같이 CRISPR/Cas9-기반 시스템은 RNA를 이용한 표적 유전자의 전사를 활성화시키는 데 사용될 수 있다. CRISPR/Cas9-기반 시스템은 전술한 바와 같이 융합 단백질을 포함할 수 있으며, 여기서 제2 폴리펩티드 도메인은 전사 활성화 활성 또는 히스톤 변형 활성을 갖는다. 예를 들어, 제2 폴리펩티드 도메인은 VP64 또는 p300을 포함할 수 있다. 대안적으로, 전사 활성화제는 아연 핑거 융합 단백질일 수 있다. 상기 기재된 바와 같이, 아연 핑거 표적화된 DNA-결합 도메인은 전사 활성화 활성 또는 히스톤 변형 활성을 갖는 도메인과 조합될 수 있다. 예를 들어, 도메인은 VP64 또는 p300을 포함할 수 있다. TALE 융합 단백질은 표적 유전자의 전사를 활성화시키는 데 사용될 수 있다. TALE 융합 단백질은 TALE DNA-결합 도메인 및 전사 활성화 활성 또는 히스톤 변형 활성을 갖는 도메인을 포함할 수 있다. 예를 들어, 도메인은 VP64 또는 p300을 포함할 수 있다.
전사 수준에서 유전자 발현을 조절하는 또 다른 방법은 본원에 기술된 변형된 DNA 엔도뉴클레아제를 사용하여 후성적 변형을 표적화하는 것이다. 후성적 수준에서 유전자 발현의 조절은 CRISPRa 또는 CRISPRi를 사용하여 달성된 활성화/억제보다 더 높은 속도로 딸 세포에 의해 유전될 수 있다는 이점을 갖는다. 일 구현예에서, p300 아세틸트랜스퍼라제의 촉매 도메인에 융합된 dCas9는 게놈의 원하는 영역에 후성적 변형 (예를 들어, 히스톤 변형 증가)을 만들기 위해 본원에 기재된 방법 및 조성물과 함께 사용될 수 있다. 후생적 변형은 Tet1 데메틸라제 촉매 도메인에 대한 TALEN의 융합 (예를 들어, Maeder et al. Nature Biotechnology 31(12):1137-42 (2013) 참조) 또는 LSD1 히스톤 데메틸라제에 융합된 TAL 이펙터 (Mendenhall et al. Nature Biotechnology 31(12):1133-6 (2013))와 같이, 변형된 TALEN 작제물을 사용하여 달성될 수도 있다.
(i) 변형된 DNA 엔도뉴클레아제, 뉴클레아제-죽은(dead) Cas9 및 이의 용도
바이러스 벡터와 달리, 본원에 기재된 ceDNA 벡터는 세포로 전달될 수 있는 핵산 서열, 이펙터 서열, 조절 서열 등의 크기 또는 수를 제한하는 캡시드를 갖지 않는다. 따라서, 본원에 기재된 ceDNA 벡터는 원하는 벡터의 생성 및/또는 세포로의 벡터의 전달 향상을 위해 변형된 기능 (예를 들어, 독특한 PAM 결합 서열)을 갖는 뉴클레아제-죽은 DNA 엔도뉴클레아제, 닉카아제 또는 다른 DNA 엔도뉴클레아제를 인코딩하는 핵산을 포함할 수 있다. 이러한 ceDNA 벡터는 또한 프로모터 서열 및 원하는 경우 다른 조절 또는 이펙터 서열을 포함할 수 있다. 크기 제약이 없으면, 당업자는, 예를 들어, 변형된 기능을 갖는 원하는 뉴클레아제의 발현, 및 임의로, 적어도 하나의 가이드 RNA가 동일한 벡터 상의 핵산 서열로부터 유래될 수 있으며, 동일한 또는 상이한 프로모터의 제어하에 있을 수 있다는 것을 쉽게 이해할 것이다. 또한, 적어도 2개의 상이한 변형된 엔도뉴클레아제가, 예를 들어, 다중화된 유전자 발현 조절을 위해 (본원의 "다중화된 유전자 발현 조절" 섹션 참조) 및 동일하거나 상이한 프로모터의 제어하에 동일한 벡터로 인코딩될 수 있는 것으로 고려된다. 따라서, 당업자는 원하는 경우, 예를 들어, 하나 이상의 유도성 프로모터에 의한 적어도 2개의 상이한 변형된 엔도뉴클레아제의 일시적으로 조절된 발현을 포함하는, 적어도 2개의 상이한 Cas9 엔도뉴클레아제 (예를 들어, 적어도 3개, 적어도 4개, 적어도 5개, 적어도 6개, 적어도 7개, 적어도 8개, 적어도 9개, 적어도 10개 또는 그 이상)의 원하는 기능성을 조합할 수 있다.
일부 구현예에서, 본원에 기재된 방법 및 조성물과 함께 사용하기 위한 DNA 엔도뉴클레아제는 DNA 엔도뉴클레아제가 예를 들어, 가이드 RNA 서열에 의해 결정된 게놈의 표적 부위에서 DNA 결합 활성을 유지하지만, 절단 활성 (예를 들어, 뉴클레아제 죽은 Cas9 (dCas9))을 보유하지 않거나 또는 비변형된 DNA 엔도뉴클레아제 (예를 들어, Cas9 닉카아제)와 비교하여 절단 활성이 (예를 들어, 적어도 10%, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60%, 적어도 70%, 적어도 80%, 적어도 90%, 적어도 95%, 적어도 99%까지) 감소되도록 변형될 수 있다. 일부 구현예에서, 변형된 DNA 엔도뉴클레아제는 본원에서 표적 유전자의 발현을 억제하기 위해 사용된다. 예를 들어, 변형된 DNA 엔도뉴클레아제가 DNA 결합 활성을 보유하기 때문에, RNA 폴리머라제의 결합을 방지하고/하거나 RNA 폴리머라제를 대체하여 표적 유전자의 전사를 방지할 수 있다. 따라서, 원하는 유전자로부터 유전자 산물 (예를 들어, mRNA, 단백질)의 발현이 방지된다.
예를 들어, "불활성화된 Cas9 (dCas9)", "뉴클레아제 죽은 Cas9" 또는 달리 불활성화된 형태의 Cas9는 특정 유전자에 대한 결합을 지시하는 가이드 RNA와 함께 도입될 수 있다. 이러한 결합은, 원하는 경우, 표적 유전자의 발현 억제를 감소시킬 수 있다. 일부 구현예에서, 그러한 유전자 발현 억제를 역전시키는 능력을 원할 수 있다. 이는, 예를 들어, 죽은 Cas9 단백질에 상이한 가이드 RNA를 제공하여 게놈 부위에 대한 Cas9의 결합을 약화시킴으로써 달성될 수 있다. 이러한 역전은 죽은 Cas9 결합의 안정성을 감소시키도록 설계된 적어도 2개 또는 일련의 가이드 RNA가 연속적으로 투여되는 반복적인 방식으로 발생할 수 있다. 예를 들어, 각각의 연속적인 가이드 RNA는 이전 반복에서 가이드 RNA에 의해 생성된 죽은 Cas9 결합의 불안정성/안정성 정도로부터 불안정성을 증가시킬 수 있다. 따라서, 일부 구현예에서, 게놈 서열의 절단 없이 "원하는 유전자를 불활성화"시키기 위해 가이드 RNA와 함께 표적 유전자 서열로 향하는 dCas9를 사용하여 관심 유전자가 기능성 단백질 형태로 발현되지 않도록 할 수 있다. 대안적인 구현예에서, 가이드 RNA는 dCas9 결합의 안정성이 감소되지만 제거되지 않도록 설계될 수 있다. 즉, RNA 폴리머라제의 변위가 완료되지 않아서 원하는 유전자의 "유전자 발현 감소"를 허용한다.
특정 구현예에서, 하이브리드 재조합 효소는 표적 DNA 상에 통합 인용을 생성하도록 본 개시내용의 ceDNA 벡터에 사용하기에 적합할 수 있다. 예를 들어, Cys2-His2 아연-핑거 또는 TAL 이펙터 DNA-결합 도메인에 융합된 세린 재조합 효소의 레솔바제(resolvase)/인버타제 패밀리로부터 유래된 활성화된 촉매 도메인에 기초한 하이브리드 재조합 효소는 포유동물 세포에서 표적화 특이성을 개선하고 탁월한 부위 특이적 통합 속도를 달성할 수 있는 시약 클래스이다. 본 개시내용에 따라 ceDNA 벡터에서 뉴클레오티드에 의해 인코딩된 적합한 하이브리드 재조합 효소는 하기에 기재된 것을 포함한다: Gaj et al., Enhancing the Specificity of Recombinase-Mediated Genome Engineering through Dimer Interface Redesign, Journal of the American Chemical Society, March 10, 2014 (그 전문이 본원에 참고로 포함됨).
(ii) 아연 핑거 엔도뉴클레아제 및 TALEN
ZFN 및 TALEN-기반 제한 엔도뉴클레아제 기술은 아연 핑거 및 전사 활성화제-유사 이펙터 (TALE)와 같은 펩티드(들)를 인식하는 특정 DNA 서열에 연결된 비특이적 DNA 절단 효소를 이용한다. 전형적으로, DNA 인식 부위 및 절단 부위가 서로 분리된 엔도뉴클레아제가 선택되고 이의 절단 부분이 분리된 다음 펩티드를 인식하는 서열에 연결되어, 원하는 서열에 대해 매우 높은 특이성을 갖는 엔도뉴클레아제가 생성된다. 이러한 특성을 갖는 예시적인 제한 효소는 FokI이다. 또한, FokI는 뉴클레아제 활성을 갖기 위해 이량체화를 요구하는 이점을 가지며, 이는 각각의 뉴클레아제 파트너가 고유한 DNA 서열을 인식하므로 특이성이 급격히 증가한다는 것을 의미한다. 이 효과를 향상시키기 위해, FokI 뉴클레아제는 이종이량체로서만 기능하고 촉매 활성이 증가될 수 있도록 조작되었다. 이종이량체 기능성 뉴클레아제는 원치 않는 동종이량체 활성의 가능성을 피하여 이중 가닥 파괴의 특이성을 증가시킨다.
ZFN 및 TALEN 둘 다의 뉴클레아제 부분은 유사한 특성을 갖지만, 이들 조작된 뉴클레아제 사이의 차이는 이들의 DNA 인식 펩티드에 있다. ZFN은 TALE 상의 Cys2-His2 아연 핑거 및 TALEN에 의존한다. 이들 DNA 인식 펩티드 도메인 둘 다는 이들의 단백질에서 자연적으로 조합되어 발견되는 특징을 갖는다. Cys2-His2 아연 핑거는 전형적으로 3 bp 간격으로 반복적으로 발생하며 전사 인자와 같은 다양한 핵산 상호 작용 단백질에서 다양한 조합으로 발견된다. 한편, TALE는 아미노산과 인식된 뉴클레오티드 쌍 사이의 일대일 인식 비율로 반복적으로 발견된다. 아연 핑거와 TALE 모두 반복된 패턴으로 발생하기 때문에, 다양한 서열 특이성을 만들기 위해 다른 조합을 시도할 수 있다. 부위-특이적 아연 핑거 엔도뉴클레아제를 제조하기 위한 접근법은, 예를 들어, 모듈형 어셈블리 (삼중 서열과 상관된 아연 핑거가 요구되는 서열을 커버하기 위해 일렬로 부착되는 경우), OPEN (펩티드 도메인의 낮은 엄중 선택(stringency selection) 대 삼중 뉴클레오티드, 이어서 펩티드 조합의 높은 엄중 선택 대 박테리아 시스템에서의 최종 표적), 및 아연 핑거 라이브러리의 박테리아 원-하이브리드 스크리닝 등을 포함한다. 본원에 기재된 방법 및 조성물과 함께 사용하기 위한 ZFN은 예를 들어, Sangamo Biosciences™ (리치몬드, 캘리포니아)로부터 상업적으로 입수할 수 있다.
본원에서 상호 교환적으로 사용되는 용어 "전사 활성화제-유사 이펙터 뉴클레아제" 또는 "TALEN"은 엔도뉴클레아제 FokI과 같은, 뉴클레아제의 촉매 도메인의 조작된 융합 단백질, 및 맞춤형 DNA 서열로 표적화될 수 있는 설계된 TALE DNA-결합 도메인을 지칭한다. "TALEN 단량체"는 촉매 뉴클레아제 도메인 및 설계된 TALE DNA-결합 도메인을 갖는 조작된 융합 단백질을 지칭한다. 2개의 TALEN 단량체는 TALEN 표적 영역을 표적화하고 절단하도록 설계될 수 있다.
본원에 사용된 용어 "전사 활성화제-유사 이펙터" 또는 "TALE"는 특정 DNA 서열을 인식하고 이에 결합하는 단백질 구조를 지칭한다. "TALE DNA-결합 도메인"은 RVD 모듈로도 알려진 탠덤 33-35 아미노산 반복 배열을 포함하는 DNA-결합 도메인을 지칭하며, 이들 각각은 단일 염기쌍의 DNA를 특이적으로 인식한다. RVD 모듈은 정의된 서열을 인식하는 어레이를 조립하기 위해 임의의 순서로 배열될 수 있다. TALE DNA-결합 도메인의 결합 특이성은 RVD 어레이에 의해 이어서 20개의 아미노산의 단일 절단된 반복에 의해 결정된다. TALE DNA-결합 도메인은 12 내지 27개의 RVD 모듈을 가질 수 있으며, 이들 각각은 RVD를 함유하고 단일 염기쌍의 DNA를 인식한다. 4개의 가능한 DNA 뉴클레오티드 (A, T, C 및 G) 각각을 인식하는 특정 RVD가 확인되었다. TALE DNA-결합 도메인은 모듈식이기 때문에, 4개의 상이한 DNA 뉴클레오티드를 인식하는 반복부는 함께 연결되어 임의의 특정 DNA 서열을 인식할 수 있다. 이어서, 이들 표적화된 DNA-결합 도메인을 촉매 도메인과 조합하여 인공 전사 인자, 메틸트랜스퍼라제, 인테그라제, 뉴클레아제, 및 재조합 효소를 비롯한 기능성 효소를 생성할 수 있다.
TALEN은 TALEN 표적 영역에서 표적 서열 또는 유전자에 결합하는 뉴클레아제 및 TALE DNA-결합 도메인을 포함할 수 있다. "TALEN 표적 영역"은 2개의 TALEN에 대한 결합 영역, 및 결합 영역 사이에서 발생하는 스페이서 영역을 포함한다. 2개의 TALEN은 TALEN 표적 영역 내의 상이한 결합 영역에 결합한 후, TALEN 표적 영역이 절단된다. TALEN의 예는 국제 특허 출원 WO2013163628에 기재되어 있으며, 이는 그 전문이 참고로 포함된다.
본원에서 상호 교환적으로 사용되는 용어 "아연 핑거 뉴클레아제" 또는 "ZFN"은 완전히 조립될 때 DNA를 절단할 수 있는 적어도 하나의 뉴클레아제 또는 뉴클레아제의 일부에 효과적으로 연결된 적어도 하나의 아연 핑거 DNA 결합 도메인을 포함하는 키메라 단백질 분자를 지칭한다. 본원에 사용된 "아연 핑거"는 DNA 서열을 인식하고 이에 결합하는 단백질 구조를 지칭한다. 아연 핑거 도메인은 인간 프로테옴에서 가장 흔한 DNA 결합 모티프이다. 단일 아연 핑거는 대략 30개의 아미노산을 함유하고, 도메인은 전형적으로 염기쌍당 단일 아미노산 측쇄의 상호 작용을 통해 연속 3개의 염기쌍 DNA를 결합시킴으로써 기능한다.
특정 구현예에서, 본 개시내용에 따른 ceDNA 벡터는 포유동물 세포와 같은 세포에 표적화된 변형을 도입하기에 적합한 아연-핑거 재조합 효소 (ZFR) 또는 키메라 단백질을 인코딩하는 뉴클레오티드 서열을 포함한다. 표적화된 뉴클레아제 및 통상적인 SSR 시스템과는 달리, ZFR 특이성은 모듈형 부위-특이적 DNA 인식 및 서열-의존적 촉매 작용의 협동 산물이다. 플러그 앤 플레이(plug-and-play) 방식으로 다양한 표적화 능력을 갖춘 ZFR을 생성할 수 있다. 향상된 촉매 도메인을 포함하는 ZFR은 개선된 표적화 특이성 및 효율을 나타내고, 낮은 독성으로 인간 게놈으로 치료 유전자의 부위-특이적 전달을 가능하게 한다. Cre 재조합 효소 이량체 인터페이스의 돌연변이 유발은 재조합 특이성을 개선시킨다.
구현예에서, 본 개시내용에 따른 ceDNA 벡터는 하기에 기재된 것과 같은 뉴클레아제 비함유 HDR 시스템에서 사용하기에 적합하다: Porro et al., Promoterless gene targeting without nucleases rescues lethality of a Crigler-Najjar syndrome mouse model, EMBO Molecular Medicine, July 27, 2017 (그 전문이 본원에 참고로 포함됨). 이러한 구현예에서, 생체내 유전자 표적화 접근법은 뉴클레아제를 사용하지 않고 공여체 서열의 삽입에 기초한 ceDNA 적용에 적합하다. 일부 구현예에서, 공여체 서열은 프로모터가 없을 수 있다.
TALEN 및 ZFN은 DNA 편집 (예를 들어, 게놈 DNA 편집)을 위한 ceDNA 벡터의 사용을 위해 예시되지만, 또한, mtZFN 및 mitoTALEN 기능, 또는 하기에 기재된 바와 같이, 미토콘드리아 DNA (mtDNA)의 편집을 위한 ceDNA 벡터의 사용을 위한 미토콘드리아-적응 CRISPR/Cas9 플랫폼의 사용이 본 명세서에 포함된다: Maeder, et al. "Genome-editing technologies for gene and cell therapy." Molecular Therapy 24.3 (2016): 430-446 및 Gammage PA, et al. Mitochondrial Genome Engineering: The Revolution May Not Be CRISPR-Ized. Trends Genet. 2018;34(2):101-110.
(iii) 핵산 유도 엔도뉴클레아제
ceDNA-매개 유전자 편집을 용이하게 하기 위해 상이한 유형의 핵산 유도 엔도뉴클레아제가 본 발명의 조성물 및 방법에 사용될 수 있다. 본 발명의 조성물 및 방법에 적합한 예시적인, 비제한적 유형의 핵산-유도 엔도뉴클레아제는 RNA-유도 엔도뉴클레아제, DNA-유도 엔도뉴클레아제 및 단일-염기 편집기를 포함한다.
일부 구현예에서, 뉴클레아제는 RNA-유도 엔도뉴클레아제일 수 있다. 본원에 사용된 용어 "RNA-유도 엔도뉴클레아제"는 선택된 표적 DNA 서열에 상보적인 영역을 포함하는 RNA 분자와 복합체를 형성하는 엔도뉴클레아제를 지칭하며, 따라서 RNA 분자는 선택된 서열에 결합하여 엔도뉴클레아제 활성을 선택된 표적 DNA 서열로 지시한다.
일 구현예에서, RNA-유도 엔도뉴클레아제는 본원에 논의된 바와 같은 CRISPR 효소이다. 일부 구현예에서, RNA-유도 엔도뉴클레아제는 닉카아제 활성을 포함한다. 일부 구현예에서, RNA-유도 엔도뉴클레아제는 표적 서열의 위치에서, 예컨대 표적 서열 내에서 및/또는 표적 서열의 보체 내에서 하나 또는 양쪽 가닥의 절단을 지시한다. 일부 구현예에서, RNA-유도 엔도뉴클레아제는 표적 서열의 첫 번째 또는 마지막 뉴클레오티드로부터 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 50, 100, 200, 500개 이상의 염기쌍 이내에서 하나 또는 양쪽 가닥의 절단을 지시한다. 다른 구현예에서, 닉카아제 활성은, 예를 들어, HDR 주형이 표적 유전자의 게놈 서열과 HDR 상호 작용에 노출되도록 서열 제한을 완화시키기 위한 ceDNA 벡터 자체 상의 하나 이상의 서열에 관한 것이다.
특정 구현예에서, 닉카아제는 원하는 핵산 서열 (예를 들어, ceDNA 벡터의 하나 이상의 영역) 상의 적어도 1개의 부위, 적어도 2개의 부위, 적어도 3개의 부위, 적어도 4개의 부위, 적어도 5개의 부위, 적어도 6개의 부위, 적어도 7개의 부위, 적어도 8개의 부위, 적어도 9개의 부위, 적어도 10개의 부위 또는 그 이상을 절단하는 것으로 고려된다. 또 다른 구현예에서, 닉카아제가 트랜스-닉킹(trans-nicking)을 통해 1개 및/또는 2개의 부위에서 절단되는 것으로 고려된다. 트랜스-닉킹은 HDR에 의한 게놈 편집을 향상시킬 수 있으며, 이는 정확도(fidelity)가 높고, 오류가 적으므로 원치 않는 오프-표적 효과를 감소시킨다.
일부 구현예에서, 발현 작제물 또는 벡터는 돌연변이된 엔도뉴클레아제가 표적 서열을 함유하는 표적 폴리뉴클레오티드의 하나의 가닥을 절단하는 능력이 결핍되도록 상응하는 야생형 효소에 대해 돌연변이된 RNA-유도 엔도뉴클레아제를 인코딩한다.
일부 구현예에서, RNA-유도 엔도뉴클레아제를 인코딩하는 핵산 서열은 진핵 세포와 같은 특정 세포에서의 발현에 대해 코돈 최적화된다. 진핵 세포는 포유동물과 같은 특정 유기체로부터 유래될 수 있다. 포유동물의 비제한적인 예는 인간, 마우스, 랫트, 토끼, 개 또는 비인간 영장류를 포함할 수 있다. 일반적으로, 코돈 최적화는 천연 서열의 적어도 하나의 코돈 (예를 들어, 약 1, 2, 3, 4, 5, 10, 15, 20, 25, 50 또는 그 이상의 코돈)을, 천연 아미노산 서열을 유지하면서 숙주 세포의 유전자에서 보다 빈번하게 또는 가장 빈번하게 사용되는 코돈으로 대체함으로써 관심 숙주 세포에서 발현을 향상시키기 위한 핵산 서열을 변형시키는 과정을 지칭한다.
일부 구현예에서, RNA-유도 엔도뉴클레아제는 하나 이상의 이종 단백질 도메인 (예를 들어, 엔도뉴클레아제 이외에 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 또는 그 이상의 도메인)을 포함하는 융합 단백질의 일부이다. RNA-유도 엔도뉴클레아제 융합 단백질은 임의의 추가 단백질 서열, 및 임의로 임의의 두 도메인 사이의 링커 서열을 포함할 수 있다. RNA-유도 엔도뉴클레아제에 융합될 수 있는 단백질 도메인의 예로는, 비제한적으로, 에피토프 태그, 리포터 유전자 서열, 정제 태그, 형광 단백질 및 메틸라제 활성, 탈메틸라제 활성, 전사 활성화 활성, 전사 억제 활성, 전사 방출 인자 활성, 히스톤 변형 활성, RNA 절단 활성 및 핵산 결합 활성 중 하나 이상을 갖는 단백질 도메인이 포함된다. 에피토프 태그의 비제한적인 예는 히스티딘 (His) 태그, V5 태그, FLAG 태그, 인플루엔자 헤마글루티닌 (HA) 태그, Myc 태그, VSV-G 태그, 글루타티온-S-트랜스퍼라제 (GST), 키틴 결합 단백질 (CBP), 말토오스 결합 단백질 (MBP), 폴리(NANP), 탠덤 친화성 정제 (TAP) 태그, myc, AcV5, AU1, AU5, E, ECS, E2, nus, Softag 1, Softag 3, Strep, SBP, Glu-Glu, HSV, KT3, S, SI, T7, 비오틴 카복실 캐리어 단백질 (BCCP), 칼모둘린 및 티오레독신 (Trx) 태그를 포함한다. 리포터 유전자의 예는 글루타티온-S-트랜스퍼라제 (GST), 호스래디쉬 퍼옥시다제 (HRP), 클로람페니콜 아세틸트랜스퍼라제 (CAT) 베타-갈락토시다제, 베타-글루쿠로니다제, 루시퍼라제, 녹색 형광 단백질 (예를 들어, GFP, GFP-2, tagGFP, turboGFP, sfGFP, EGFP, 에메랄드, 아자미 그린, 모노머 아자미 그린, CopGFP, AceGFP, ZsGreen1), HcRed, DsRed, 시안 형광 단백질 (CFP), 노란색 형광 단백질 (예를 들어, YFP, EYFP, 시트린, 비너스 YPet, PhiYFP, ZsYellow1), 시안 형광 단백질 (예를 들어, ECFP, 세룰리언, CyPet AmCyan1, 미도리이시-시안(Midoriishi-Cyan)) 적색 형광 단백질 (예를 들어, mKate, mKate2, mPlum, DsRed 모노머, mCherry, mRFP1, DsRed-Express, DsRed2 , HcRed-Tandem, HcRed1, AsRed2, eqFP611, mRaspberry, mStrawberry, Jred), 주황색 형광 단백질 (예를 들어, mOrange, mKO, 쿠사비라-오렌지(Kusabira-Orange), 모노머 쿠사비라-오렌지, mTangerine, tdTomato) 및 청색 형광 단백질 (BFP)을 포함한 자가형광(autofluorescent) 단백질을 포함한다. RNA-유도 엔도뉴클레아제는 DNA 분자에 결합하거나, 말토오스 결합 단백질 (MBP), S-태그, Lex A DNA 결합 도메인 (DBD) 융합물, GAL4 DNA 결합 도메인 융합물, 및 단순 포진 바이러스 (HSV) BP16 단백질 융합물을 포함하지만, 이에 제한되지 않는 다른 세포 분자에 결합하는 단백질 또는 단백질의 단편을 인코딩하는 유전자 서열에 융합될 수 있다. 일부 구현예에서, 태그된 엔도뉴클레아제는 표적 서열의 위치를 식별하기 위해 사용된다.
본원에서는, 적어도 2개 (예를 들어, 적어도 3개, 적어도 4개, 적어도 5개, 적어도 6개, 적어도 7개, 적어도 8개, 적어도 9개, 적어도 10개, 적어도 12개, 적어도 15개 이상)의 상이한 Cas 효소가 실질적으로 동시에 투여되거나 세포와 접촉하고 있음이 고려된다. 이중 가닥 파괴 유도 Cas 효소, Cas 닉카아제, 촉매적으로 비활성인 Cas 효소 (예를 들어, dCas9), 변형된 Cas 효소, 절단된 Cas9 등의 임의의 조합이 본원에 기재된 방법 및 조성물과 조합하여 사용하기 위해 고려된다.
일부 구현예에서, 핵산 유도 엔도뉴클레아제는 DNA 유도 엔도뉴클레아제이다. 예를 들어, Varshney and Burgess Genome Biol. 17:187 (2016)을 참조한다. 일 구현예에서, DNA 복구 및/또는 복제에 관여하는 효소는 엔도뉴클레아제에 융합되어 DNA-유도 뉴클레아제를 형성할 수 있다. 하나의 비제한적인 예는 flap 엔도뉴클레아제 1 (FEN-1)의 Fok1 엔도뉴클레아제로의 융합이다 (Xu et al., Genome Biol. 17:186 (2016). 또 다른 구현예에서, 천연 발생 DNA-유도된 뉴클레아제가 사용될 수 있다. 이러한 천연 발생 뉴클레아제의 비제한적인 예는 아르고너트(Argonaute) 단백질 패밀리로부터의 원핵 생물 엔도뉴클레아제이다 (Kropocheva et al., FEBS Open Bio. 8(S1): P01-074 (2018). 일부 구현예에서, 핵산-유도 엔도뉴클레아제는 "단일 염기 편집기"이며, 이는 DNA 표적화 모듈 및 단일 유형의 뉴클레오티드 염기를 변형시킬 수 있는 촉매 도메인으로 구성된 키메라 단백질이다(Rusk, N, Nature Methods 15:763 (2018); Eid et al., Biochem J. 475(11): 1955-64 (2018)). 이러한 단일 염기 편집기는 DNA 염기의 편집을 수행하기 위해 표적 DNA에서 이중 가닥 파괴를 초래하지 않기 때문에, 삽입 및 결실 (예를 들어, 인델)의 생성이 제한되며, 따라서 편집 과정의 정확도를 향상시킨다. 다른 유형의 단일 염기 편집기가 알려져 있다. 예를 들어, 시티딘 데미아미나제 (시토신의 우라실로의 전환을 촉매하는 효소)는 APOBEC-dCas9와 같은 뉴클레아제에 결합될 수 있으며, 여기서 APOBEC는 시티딘 데미아미나제 기능에 기여하고, 특정 시티딘을 우라실로 탈아미노화하기 위해 dCas9에 의해 유도된다. 결과적인 U-G 불일치는 복구 메커니즘을 통해 해결되고, U-A 염기쌍을 형성하며, 이는 C-대-T 점 돌연변이로 번역된다 (Komor et al., Nature 533: 420-424 (2016); Shimatani et al., Nat. Biotechnol. 35: 441-443 (2017)). 아데닌 데아미나제-기반 DNA 단일 염기 편집기가 조작되었다. 이들은 아데노신을 탈아미노화하여 이노신을 형성하는데, 이는 시티딘과 염기쌍을 이루고 구아닌으로 교정되어 A-T 쌍이 G-C 쌍으로 전환될 수 있다. 이러한 편집기의 예에는 TadA, ABE5.3, ABE7.8, ABE7.9 및 ABE7.10이 포함된다 (Gaudelli et al., Nature 551: 464-471 (2017).
(iv) CRISPR/Cas 시스템
당 업계에 알려진 바와 같이, CRISPR-CAS9 시스템은 유기체의 유전자 서열을 변경할 수 있는 단백질 및 리보핵산 ("RNA")의 조합을 포함하는 특정 핵산 유도-뉴클레아제-기반 시스템 세트이다. CRISPR-CAS9 시스템은 미생물, 진균, 식물 및 동물과 같은 많은 유기체의 게놈에서 특정 데옥시리보핵산 ("DNA")을 변형시키는 강력한 도구로 계속 개발되고 있다. 예를 들어, 인간 질환의 마우스 모델은 개별 유전자를 훨씬 더 빠르게 연구하고, 세포에서 여러 유전자를 한 번에 쉽게 변경하여 이들의 상호 작용을 연구하기 위해 신속하게 개발될 수 있다. 당업자는 유형 I, 유형 II 및 유형 III과 같은 다수의 공지된 CRISPR 시스템 중에서 선택할 수 있다. 유형 II CRISPR-CAS 시스템은 3가지 성분: (1) "가이드 서열" 또는 "타겟터(targeter)-RNA"로 불리는 crDNA 분자; (2) "tracr RNA" 또는 "활성화제-RNA"; 및 (3) Cas9라는 단백질을 포함하는 잘 알려진 메커니즘을 갖는다.
DNA 분자를 변경하기 위해, 시스템에서 다음을 포함하는 많은 상호 작용이 발생한다: (1) 특정 염기 짝짓기에 의해 관심 DNA ("표적 DNA")의 특정 서열에 결합하는 가이드 서열, (2) 가이드 서열은 또 다른 서열에서 특정 염기 짝짓기에 의해 활성화제-RNA에 결합하고, (3) 활성화제-RNA는 Cas 단백질 (예를 들어, Cas9 단백질)과 상호 작용하여, 특정 부위에서 표적 DNA를 절단하는 뉴클레아제로서 작용한다. 본 개시내용에 따른 ceDNA 벡터에 따라 사용하기에 적합한 시스템이 하기에 추가로 기재되어 있다: Van Nierop, et al. Stimulation of homology-directed gene targeting at an endogenous human locus by a nicking endonuclease, Nucleic Acid Research, August 2009 및 Ran et al., Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity.
본 개시내용에 따른 ceDNA 벡터는 가이드 서열, tracr RNA, 또는 Cas (예를 들어, Cas9)와 같은 이들 시스템의 하나 이상의 성분을 인코딩하는 뉴클레오티드를 포함하도록 설계될 수 있다. 특정 구현예에서, 단일 프로모터는 가이드 서열 및 tracr RNA의 발현을 구동하고, 별도의 프로모터는 Cas (예를 들어, Cas9) 발현을 구동한다. 당업자는 특정 Cas 뉴클레아제가 표적 핵산 서열에 인접한 프로토스페이서 인접 모티프 (PAM)의 존재를 필요로 한다는 것을 이해할 것이다. 일부 구현예에서, PAM은 표적 서열의 3 '말단의 1, 2, 3 또는 4개의 뉴클레오티드에 인접하거나 그 내에 있을 수 있다. PAM의 길이 및 서열은 특정 Cas 단백질에 따라 달라질 수 있다. 예시적인 PAM 서열은 NGG, NGGNG, NG, NAAAAN, NNAAAAAW, NNNNACA, GNNNCNNA, TTN 및 NNNNGATT (여기서 N은 임의의 뉴클레오티드로 정의되고, W는 A 또는 T로 정의됨)를 포함한다. 일부 구현예에서, PAM 서열은, 예를 들어, RNA를 편집할 때 가이드 RNA 상에 존재할 수 있다.
일부 구현예에서, Cas 및 Cas9를 포함하는 RNA-유도 뉴클레아제는 CRISPR-Cas9 시스템을 사용한 게놈 조작을 위한 하나 이상의 성분을 제공하도록 설계된 ceDNA 벡터에 사용하기에 적합하다. 예를 들어, 그 전문이 본원에 참고로 포함된 미국 공보 2014/0170753을 참조한다. CRISPR-Cas 9는 포유동물 세포에서 비-동종 말단 결합 (NHEJ) 또는 상동성 지정 복구 (HDR)를 통한 Cas9 매개 게놈 편집을 위한 일련의 도구, 및 하류 기능 연구를 위한 변형된 세포주 생성을 제공한다. 오프-표적 절단을 최소화하기 위해, CRISPR-Cas9 시스템은 쌍을 이룬 가이드 RNA와 함께 Cas9 닉카아제 돌연변이체를 사용하는 이중-닉킹 전략을 포함할 수 있다. 이 시스템은 당 업계에 공지되어 있으며, 예를 들어, 하기에 기재되어 있다: Ran et al., Genome engineering using the CRISPR-Cas9 system, Nature Protocols, 24 October 2013, 및 Zhang, et al., Efficient precise knockin with a double cut HDR donor after CRISPR/Cas9-mediated double-stranded DNA cleavage, Genome Biology, 2017 (두 참조 문헌은 그 전문이 본원에 참고로 포함됨).
특정 구현예에서, ceDNA 시스템은 ceDNA 서열로 향하는 가이드 RNA 및 뉴클레아제를 포함한다. 예를 들어, 유전자 편집 효율을 높이기 위해 nCAS9 D10A와 같은 닉킹 CAS를 사용할 수 있다. 가이드 RNA는 ceDNA의 nCAS 닉킹을 지시함으로써 보다 효율적인 유전자 복구 및/또는 발현을 위해 ceDNA의 비틀림 제약을 해제할 수 있다. 닉킹 뉴클레아제를 사용하여 닉킹된 DNA가 핵 내에서 지속될 수 있게 하는 서열 및 구조적 완전성을 유지하면서 비틀림 제약을 완화시킨다. 가이드 RNA는 동일한 가닥의 DNA 또는 상보적 가닥으로 향할 수 있다. 가이드 RNA는 예를 들어, ITRS, 또는 프로모터 선행 서열(sequences proceeding promoters), 또는 상동성 도메인 등으로 향할 수 있다.
일 구현예에서, RNA-유도 엔도뉴클레아제는 CRISPR 효소, 예컨대 Cas 단백질이다. Cas 단백질의 비제한적인 예는 Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8, Cas8a1, Cas8a2, Cas8b, Cas8c, Cas9 (Csn1 및 Csx12로도 알려짐), Cas10, Cas10d, Cas13, Cas13a, Cas13c, CasF, CasH, Csy1, Csy2, Csy3, Cse1, Cse2, Cse3, Cse4, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx11, Csx16, CsaX, Csz1, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, Cul966, Cpf1, C2c1, C2c3, 이의 상동체 또는 이의 변형된 버전을 포함한다. 일 구현예에서, Cas 단백질은 Cas9이다. 또 다른 구현예에서, Cas 단백질은 뉴클레아제-죽은 Cas9 (dCas9) 또는 Cas9 닉카아제이다. 일 구현예에서, Cas 단백질은 닉킹 Cas 효소 (nCas)이다.
전형적으로, RNA-유도 엔도뉴클레아제는 Cas9에 의해 개시된 이중 가닥 파괴와 같은 DNA 절단 활성을 포함한다. 일부 구현예에서, RNA-유도 엔도뉴클레아제는 Cas9, 예를 들어 S. 피오게네스(S. pyogenes) 또는 S. 뉴모니아에(S. pneumoniae) 유래 Cas9이다. 기타 비제한적인 Cas9의 박테리아 공급원은 스트렙토코커스 피오게네스(Streptococcus pyogenes), 스트렙토코커스 파스퇴리아누스(Streptococcus pasteurianus) 스트렙토코커스 써모필루스(Streptococcus thermophilus), 스트렙토코커스 종, 노카르디옵시스 다손빌레이(Nocardiopsis dassonvillei), 스트렙토마이세스 프리스티나에스피랄리스(Streptomyces pristinaespiralis), 스트렙토마이세스 비리도크로모게네스(Streptomyces viridochromogenes), 스트렙토스포랑지움 로세움(Streptosporangium roseum), 스트렙토스포랑지움 로세움(Streptosporangium roseum), 스타필로코커스 아우레우스(Staphylococcus aureus), 알리사이클로바실러스 애시도칼다리우스(Alicyclobaccillus acidocaldarius), 바실러스 슈도마이코이데스(Bacillus pseudomycoides), 바실러스 셀레니티레두센스(Bacillus selenitireducens), 엑시구오박테리움 시비리쿰(Exiguobacterium sibiricum), 프란시셀라 노빅 이다(Francisella novic ida), 울리넬라 석시노게네스(Wolinella succinogenes), 락토바실러스 델브루키(Lactobacillus delbrueckii), 락토바실러스 살리바리우스(Lactobacillus salivarius), 리스테리아 이노쿠아(Listeria innocua), 락토바실러스 가세리(Lactobacillus gasseri), 마이크로실라 마리나(Microscilla marina), 버콜더리알레스 박테리움(Burkholderiales bacterium), 폴라로모나스 나프탈레니보란스(Polaromonas naphthalenivorans), 폴라로모나스 종, 크로코스패라 왓소니(Crocosphaera watsonii), 시아노테세 종(Cyanothece sp.), 마이크로시스티스 아에루기노사(Microcystis aeruginosa), 시네초코커스 종(Synechococcus sp.), 아세토할로븀 아라바티쿰(Acetohalobium arabaticum), 암모니펙스 데겐시(Ammonifex degensii), 칼디셀룰로시럽터 벡시(Caldicelulosiruptor becscii), 칸디다투스 데설포루디스(Candidatus Desulforudis), 클로스트리듐 보툴리눔(Clostridium botulinum), 클로스트리듐 디피실레(Clostridium difficile), 피네골디아 마그나(Finegoldia magna), 피브로박터 석시노겐(Fibrobacter succinogene), 나트라내로비우스 써모필루스(Natranaerobius thermophilus), 펠로토마쿨룸써모프로피오니쿰(Pelotomaculumthermopropionicum), 애시디티오바실러스 칼두스(Acidithiobacillus caldus), 애시디티오바실러스 페로옥시단스(Acidithiobacillus ferrooxidans), 알로크로마튬 비노숨(Allochromatium vinosum), 마리노박터 종(Marinobacter sp.), 니트로소코커스 할로필루스(Nitrosococcus halophilus), 니트로소코커스 왓소니(Nitrosococcus watsoni), 슈도알테로모나스 할로플랑크티스(Pseudoalteromonas haloplanktis), 크테도노박터 라세미퍼(Ktedonobacter racemifer), 메타노할로븀 에베스티가툼(Methanohalobium evestigatum), 아나배나 바리아빌리스(Anabaena variabilis), 노둘라리아 스푸미게나(Nodularia spumigena), 노스톡 종(Nostoc sp.), 아트로스피라 맥시마(Arthrospira maxima), 아트로스피라 플라텐시스(Arthrospira platensis), 아트로스피라 종, 링비아 종(Lyngbya sp.), 마이크로콜레우스 크토노플라스테스(Microcoleus chthonoplastes), 오실라토리아 종(Oscillatoria sp.), 페트로토가 모빌리스(Petrotoga mobilis), 써모시포 아프리카누스(Thermosipho africanus), 수테렐라 워즈워쎈시스(Sutterella wadsworthensis), 감마 프로테오박테리움(Gamma proteobacterium), 나이세리아 시네레아(Neisseria cinerea), 나이세리아 메닝기티디스(Neisseria meningitidis), 캄필로박터 제주니(Campylobacter jejuni), 캄필로박터 라리(Campylobacter lari), 파르비바쿨룸 라바멘티보란스(Parvibaculum lavamentivorans), 코르니박테리움 디프테리아(Corneybacterium diphtheria), 파스퇴렐라 물토시다(Pasteurella multocida), 로도스피릴룸 루브룸(Rhodospirillum rubrum), 노카르디옵시스 다손빌레이(Nocardiopsis dassonvillei), 또는 아카리오클로리스 마리나(Acaryochloris marina)를 포함한다.
일 구현예에서, Cas9 닉카아제는 nCas9 D10A를 포함한다. 예를 들어, S. 파이오게네스로부터의 Cas9의 RuvC I 촉매 도메인에서 아스파테이트-대-알라닌 치환 (D10A)은 Cas9를 두 가닥을 절단하는 뉴클레아제로부터 닉카아제 (단일 가닥을 절단함)로 전환한다. Cas9를 닉카아제로 만드는 돌연변이의 다른 예에는, 비제한적으로, H840A, N854A 및 N863A가 포함된다. 일부 구현예에서, Cas9 닉카아제는 각각 DNA 표적의 센스 및 안티센스 가닥을 표적화하는 가이드 서열(들), 예를 들어, 2개의 가이드 서열과 조합하여 사용될 수 있다. 이 조합은 두 가닥이 닉킹되고 비-동종 말단 결합 (NHEJ) 복구를 유도하는 데 사용되도록 한다.
일부 구현예에서, RNA-유도 엔도뉴클레아제는 Cas13이다. 촉매적으로 비활성인 Cas13 (dCas13)은 예를 들어, 하기에 기재된 바와 같은 mRNA 서열을 편집하는 데 사용될 수 있다: Cox, D et al. RNA editing with CRISPR-Cas13 Science (2017) DOI: 10.1126/science.aaq0180 (이는 그 전문이 본원에 참조로 포함됨).
일부 구현예에서, 엔도뉴클레아제를 인코딩하는 본원에 기재된 바와 같은 ceDNA 벡터는 Cas9 (예를 들어, 서열 번호: 829), 또는 서열 번호: 829 (Cas9)와 적어도 60%, 보다 바람직하게는 적어도 65%, 보다 바람직하게는 적어도 70%, 보다 바람직하게는 적어도 75%, 보다 바람직하게는 적어도 80%, 보다 바람직하게는 적어도 85%, 더욱 더 바람직하게는 적어도 90%, 가장 바람직하게는 적어도 95% 서열 동일성을 갖거나 서열 번호: 829로 구성된 뉴클레아제의 아미노산 또는 기능성 단편이다. 특정 구현예에서, Cas 9는 촉매 도메인에서 하나 이상의 돌연변이를 포함하여 Cas 9를 단일 DNA 가닥을 절단하는 닉카아제, 예컨대 미국 특허 공보 번호 2017-0191078-A9 (그 전문이 참조로 포함됨)에 기재된 것으로 만든다.
일부 구현예에서, 본 개시내용의 ceDNA 벡터는 유전자-표적화 및 게놈 편집 기능을 갖는 RNA-프로그래밍된 Cas9에 기초한 시스템 및 방법에 사용하기에 적합하다. 예를 들어, 본 개시내용의 ceDNA 벡터는 유전자 표적화 및 유전자 편집을 위한 클러스터된 규칙적으로 간격을 둔 짧은 회문 구조 반복 또는 CRISPR 관련 (Cas) 시스템과 함께 사용하기에 적합하다. CRISPR cas9 시스템은 당 업계에 공지되어 있고, 예를 들어, 2013년 3월에 출원된 미국 특허 출원 번호 13/842,859, 및 미국 특허 번호 8,697,359, 8771,945, 8795,965, 8,865,406, 8,871,445 (이들 모두는 그 전문이 본원에 참조로 포함됨)에 기재되어 있다.
Cas9, Cas9 닉카아제, 또는 불활성화된 Cas9 (dCas9, 또는 뉴클레아제 죽은 Cas9 또는 "촉매적으로 비활성"으로도 지칭됨)는 또한 FokI과의 융합 단백질로서 제조되어 FokI 이종이량체의 형성시에 유전자 편집 또는 유전자 발현 조절이 발생한다.
또한, dCas9는 표적 유전자 서열의 상류에 있는 조절 서열 수준에서 원하는 유전자의 발현을 활성화 (CRISPRa) 또는 억제 (CRISPRi)하는 데 사용될 수 있다. CRISPRa 및 CRISPRi는, 예를 들어, dCas9를 이펙터 영역과 융합시키고 (예를 들어, dCas9/이펙터 융합) dCas9/이펙터 융합 단백질이 (예를 들어, 프로모터 영역에서) 원하는 또는 표적 유전자의 상류에 있는 서열에 결합하도록 지시하는 가이드 RNA를 공급함으로써 수행될 수 있다. dCas9는 뉴클레아제 활성이 없기 때문에, 프로모터 영역의 표적 부위에 결합된 상태로 유지되고, dCas9/이펙터 융합 단백질의 이펙터 부분은 전사 활성화제 또는 억제자를 프로모터 부위로 동원할 수 있다. 이와 같이, 원하는 대로 표적 유전자의 유전자 발현을 활성화 또는 감소시킬 수 있다. 문헌에서의 이전 연구는 dCas9와 공동 발현된 복수의 가이드 RNA의 사용이 원하는 유전자의 발현을 증가시킬 수 있음을 나타낸다 (예를 들어, Maeder et al. CRISPR RNA-guided activation of endogenous human genes Nat Methods 10(10):977-979 (2013) 참조. 일부 구현예에서, 원하는 유전자의 유도성 억제를 허용하는 것이 바람직하다. 이는, 예를 들어, 전사 개시 부위의 상류에 있는 프로모터 영역에서 가이드 RNA 결합 부위를 사용함으로써 달성될 수 있다 (예를 들어, Gao et al. Complex transcriptional modulation with orthogonal and inducible dCas9 regulators. Nature Methods (2016) 참조). 일부 구현예에서, DNA 엔도뉴클레아제의 뉴클레아제 죽은 버전 (예를 들어, dCas9)은, 예를 들어, dCas9/이펙터 융합 단백질의 이펙터 도메인과 상호 작용하는 제제 (예를 들어, 소분자 또는 적어도 하나의 가이드 RNA)의 도입에 의해 원하는 유전자의 발현을 유도적으로 활성화 또는 증가시키기 위해 사용될 수 있다. 다른 구현예에서, 유전자 발현이 외인성 신호를 사용하여 조절될 수 있도록, dCas9가 화학물질-유도성 또는 광-유도성 도메인에 융합될 수 있는 것으로 본원에서 또한 고려된다. 일 구현예에서, 표적 유전자 발현의 억제는 KRAB 리프레서 도메인에 융합된 dCas9를 사용하여 수행되며, 이는 포유동물 시스템에서 유전자 발현의 억제 개선에 유리할 수 있고, 오프-표적 효과가 거의 없다. 대안적으로, 유전자의 전사-기반 활성화는 RNA 폴리머라제의 오메가 서브유닛, 또는 전사 활성화제 VP64 또는 p65에 융합된 dCas9를 사용하여 수행될 수 있다.
따라서, 일부 구현예에서, 본원에 기재된 방법 및 조성물, 예를 들어, ceDNA 벡터는 CRISPRi (CRISPR 간섭) 및/또는 CRISPRa (CRISPR 활성화) 시스템을 숙주 세포로 전달하기 위해 사용되고/되거나 포함할 수 있다. CRISPRi 및 CRISPRa 시스템은 이중 가닥 파괴 (DSB)를 생성할 수 없는 불활성화된 RNA-유도 엔도뉴클레아제 (예를 들어, Cas9)를 포함한다. 이는 가이드 RNA와 조합하여 엔도뉴클레아제가 게놈 내의 표적 서열에 특이적으로 결합하고 RNA-유도된 가역적 전사 조절을 제공할 수 있게 한다. 일 구현예에서, ceDNA 벡터는 뉴클레아제 및/또는 가이드 RNA를 인코딩하는 핵산을 포함하지만 상동성 지정 복구 주형 또는 상응하는 상동성 아암을 포함하지 않는다.
CRISPRi의 일부 구현예에서, 엔도뉴클레아제는 KRAB 이펙터 도메인을 포함할 수 있다. KRAB 이펙터 도메인의 유무에 관계없이, 게놈 서열에 불활성화된 뉴클레아제의 결합은, 예를 들어, 전사 개시 또는 진행을 차단하고/하거나 전사 기구 또는 전사 인자의 결합을 방해할 수 있다.
CRISPRa에서, 불활성화된 엔도뉴클레아제는 하나 이상의 전사 활성화 도메인과 융합될 수 있으며, 이에 의해 엔도뉴클레아제에 의해 표적화된 부위에서 또는 그 근처에서 전사를 증가시킨다. 일부 구현예에서, CRISPRa는 추가의 전사 활성화 도메인을 동원하는 gRNA를 추가로 포함할 수 있다. CRISPRi 및 CRISPRa에 대한 sgRNA 설계는 당 업계에 공지되어 있다 (예를 들어, Horlbeck et al. eLife. 5, e19760 (2016); Gilbert et al., Cell. 159, 647-661 (2014); 및 Zalatan et al., Cell. 160, 339-350 (2015) 참조; 이들 각각은 그 전문이 본원에 참고로 포함됨). CRISPRi 및 CRISPRa-호환성 sgRNA는 또한 주어진 표적에 대해 상업적으로 얻어질 수 있다 (예를 들어, Dharmacon; Lafayette, CO 참조). CRISPRi 및 CRISPRa에 대한 추가 설명은, 예를 들어, 하기에서 발견될 수 있다: Qi et al., Cell. 152, 1173-1183 (2013); Gilbert et al., Cell. 154, 442-451 (2013); Cheng et al., Cell Res. 23, 1163-1171 (2013); Tanenbaum et al. Cell. 159, 635-646 (2014); Konermann et al., Nature. 517, 583-588 (2015); Chavez et al., Nat. Methods. 12, 326-328 (2015); Liu et al., Science. 355 (2017); 및 Goyal et al., Nucleic Acids Res. (2016) (이들 각각은 그 전문이 본원에 참고로 포함됨).
따라서, 일부 구현예에서, 불활성화된 엔도뉴클레아제, 예를 들어, RNA-유도 엔도뉴클레아제 및/또는 Cas9를 포함하는 ceDNA 벡터가 본원에 기재되어 있으며, 여기서 불활성화된 엔도뉴클레아제는 엔도뉴클레아제 활성이 결여되지만, 예를 들어, 하나 이상의 가이드 RNA 및/또는 sgRNA와 함께 부위-특이적 방식으로 DNA에 결합하는 능력을 보유한다. 일부 구현예에서, 벡터는 하나 이상의 tracrRNA, 가이드 RNA 또는 sgRNA를 추가로 포함할 수 있다. 일부 구현예에서, 불활성화된 엔도뉴클레아제는 전사 활성화 도메인을 추가로 포함할 수 있다. 일부 구현예에서, 본 개시내용의 ceDNA 벡터는 또한, 모두 당 업계에 잘 알려진, CRISPRi 또는 CRISPRa dCas 시스템, nCas 또는 Cas13 시스템과 같은 불활성화된 뉴클레아제 시스템에 유용하다.
본원에 기재된 벡터는 살아있는 세포에서 게놈 유전자좌를 가시화하기 위해 dCas9와 조합하여 사용될 수 있는 것으로 고려된다 (예를 들어, Ma et al. Multicolor CRISPR labeling of chromosomal loci in human cells PNAS 112(10):3002-3007 (2015) 참조). 핵 내에서 게놈과 그 조직의 CRISPR 매개 시각화는 4-D 뉴클레옴(nucleome)으로도 불린다. 일 구현예에서, dCas9는 형광 태그를 포함하도록 변형된다. 다수의 유전자좌는, 예를 들어, 각각 다른 형광 라벨에 융합된 오쏘로그를 사용하여 별개의 색상으로 라벨링될 수 있다. 이 기술은, 예를 들어, Alu 서열에 결합하는 가이드 RNA를 사용하여 가이드 RNA-지정된 반복의 위치를 맵핑하는데 도움을 줌으로써 게놈 구조를 연구하도록 확장될 수 있다 (예를 들어, McCaffrey et al. Nucleic Acids Res 44(2):e11 (2016) 참조). 따라서, 일부 구현예에서, 예를 들어, 헌팅턴병 등의 확인 및/또는 진단을 위해 임상적으로 유의미한 유전자좌의 맵핑이 본원에서 고려된다. 유전자 편집 시스템을 인코딩하는 ceDNA 벡터(들)로 게놈 시각화 또는 유전자 스크린을 수행하는 방법은 당 업계에 공지되어 있고/있거나, 예를 들어, 하기에 기재되어 있다: Chen et al. Cell 155:1479-1491 (2013); Singh et al. Nat Commun 7:1-8 (2016); Korkmaz et al. Nat Biotechnol 34:1-10 (2016); Hart et al. Cell 163:1515-1526 (2015) (이들 각각의 내용은 그 전문이 본원에 참고로 포함됨).
일부 구현예에서, 게놈에서 단일 염기를 편집하는 것, 예를 들어, 특정 질환과 관련된 단일 뉴클레오티드 다형성을 변형시키는 것이 바람직할 수 있다 (예를 들어, Komor, AC et al. Nature 533:420-424 (2016); Nishida, K et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science (2016) 참조). 단일 뉴클레오티드 염기 편집은 표적 유전자 유전자좌를 절단하지 않는 촉매적으로 비활성인 엔도뉴클레아제 (예를 들어, 뉴클레아제 죽은 Cas9)에 테더링된 염기 전환 효소를 사용한다. 염기 편집 효소에 의한 염기 전환 후, 시스템은 반대의 편집되지 않은 가닥에 닉을 만들고, 이는 세포 자체의 DNA 복구 메커니즘에 의해 복구된다. 이는 이제 "미스매치된 뉴클레오티드"인 원래 뉴클레오티드를 대체하여 단일 뉴클레오티드 염기쌍의 전환을 완료시킨다. 내인성 효소, 예를 들어, 시티딘 데아미나제는 G/C 뉴클레오티드 쌍의 A/T 뉴클레오티드 쌍으로의 전환을 수행하는데 이용 가능하지만, A/T 뉴클레오티드 쌍의 G/C 뉴클레오티드 쌍으로의 역 전환을 촉매하는 내인성 효소는 없다. 일반적으로 아데닌을 이노신으로 전환시키기 위해 RNA에만 작용하는 아데닌 데아미나제 (예를 들어, TadA)는 박테리아 시스템에서 아데노신을 이노신으로 전환시키기 위해 DNA에 작용하는 아데닌 데아미나제 돌연변이체를 확인하기 위해 점진적으로 선택되어 왔다 (예를 들어, Gaudelli et al Nature (2017), in press doi:10.1038/nature24644 참조, 이의 내용은 그 전문이 참고로 포함됨).
일부 구현예에서, dCas9 또는 닉카아제 기능을 갖는 변형된 Cas9는 염기 편집 기능을 갖는 효소 (예를 들어, 시티딘 데아미나제 APOBEC1 또는 돌연변이체 TadA)에 융합될 수 있다. 게놈 DNA로부터 우라실을 제거하는 내인성 염기 절제 복구 시스템의 억제제를 포함시킴으로써 염기 편집 효율을 더욱 향상시킬 수 있다. 2017년 10월 25일에 온라인으로 공개된, Gaudelli et al. (2017) programmable base editing of A-T to G-C in genomic DNA without DNA cleavage, Nature (그 전문이 본원에 참고로 포함됨).
본원에서는, 원하는 엔도뉴클레아제가 유비퀴틴 또는 폴리유비퀴틴 사슬의 첨가에 의해 변형되는 것으로 고려된다. 일부 구현예에서, 유비퀴틴은 유비퀴틴-유사 단백질 (UBL)일 수 있다. 유비퀴틴-유사 단백질의 비제한적인 예는 소형 유비퀴틴-유사 개질제 (SUMO), 유비퀴틴 교차-반응성 단백질 (UCRP, 인터페론-자극 유전자 15 (ISG-15)로도 알려져 있음), 유비퀴틴-관련 개질제-1 (URM1), 뉴런-전구-세포-발현된 발달상 하향 조절된 단백질-8 (NEDD8, S. 세레비시아에(S. cerevisiae)에서 Rubl이라고도 함), 인간 백혈구 항원 F-관련 (FAT 10), 자가포식-8 (ATG8) 및 자가포식-12 (ATG12), Fau 유비퀴틴 유사 단백질 (FUB1), 막 고정 UBL (MUB), 유비퀴틴 폴드-개질제-1 (UFM1) 및 유비퀴틴 유사 단백질-5 (UBL5)를 포함한다.
CeDNA 벡터 또는 이의 조성물은 예를 들어, 하기에 기재된 바와 같이 변형된 DNA 엔도뉴클레아제를 인코딩할 수 있다: Fu et al. Nat Biotechnol 32:279-284 (2013); Ran et al. Cell 154:1380-1389 (2013); Mali et al. Nat Biotechnol 31:833-838 (2013); Guilinger et al. Nat Biotechnol 32:577-582 (2014); Slaymaker et al. Science 351:84-88 (2015); Klenstiver et al. Nature 523:481-485 (2015); Bolukbasi et al. Nat Methods 12:1-9 (2015); Gilbert et al. Cell 154;442-451 (2012); Anders et al. Mol Cell 61:895-902 (2016); Wright et al. Proc Natl Acad Sci USA 112:2984-2989 (2015); Truong et al. Nucleic Acids Res 43:6450-6458 (2015) (이들 각각의 내용은 그 전문이 본원에 참고로 포함됨).
(v) megaTALS
일부 구현예에서, 본원에 기재된 엔도뉴클레아제는 megaTAL일 수 있다. megaTAL은 전사 활성화제-유사 (TAL) 이펙터 도메인 및 메가뉴클레아제 도메인을 포함하는 조작된 융합 단백질이다. megaTAL은 오프-표적 효과와 전체 효소 크기를 줄이고 활성을 증가시키면서 TAL의 표적 특이성 엔지니어링의 용이성을 유지한다. megaTAL 작제 및 사용은, 예를 들어, 하기에 보다 상세하게 기재되어 있다: Boissel et al. 2014 Nucleic Acids Research 42(4):2591-601 and Boissel 2015 Methods Mol Biol 1239:171-196 (이들 각각은 그 전문이 본원에 참고로 포함됨). megaTAL-매개 유전자 녹아웃 및 유전자 편집에 대한 프로토콜은 당 업계에 공지되어 있으며, 예를 들어, 하기를 참조한다: Sather et al. Science Translational Medicine 2015 7(307):ra156 and Boissel et al. 2014 Nucleic Acids Research 42(4):2591-601 (이들 각각은 그 전문이 본원에 참고로 포함됨). megaTAL은 본원에 기재된 임의의 방법 및 조성물에서 대안적인 엔도뉴클레아제로서 사용될 수 있다.
(vi) 유전자 발현 및 복합 시스템의 다중 조절
본원에 기재된 바와 같은 ceDNA 벡터의 크기 제한의 결여는 다중 가이드 RNA가, 원한다면, 동일한 ceDNA 벡터로부터 발현될 수 있기 때문에 다중화된 편집, CRISPRa 또는 CRISPRi에 특히 유용하다. CRISPR은 강력한 시스템이며 다중 가이드 RNA의 첨가는 유전자 편집 효율, CRISPRa, CRISPRi 또는 핵산의 CRISPR 매개 라벨링을 실질적으로 변경하지 않는다. 다른 곳에 기술된 바와 같이, 복수의 가이드 RNA는 단일 프로모터 (예를 들어, 폴리시스트로닉 전사체)의 제어하에 또는 복수의 프로모터 (예를 들어, 적어도 2개, 적어도 3개, 적어도 4개, 적어도 5개, 적어도 6개 등, 최대 가이드 RNA:프로모터 서열의 1:1 비의 한계까지)의 제어하에 있을 수 있다.
다중 CRISPR/Cas9 기반 시스템은 sgRNA 설계의 단순성과 저렴한 비용을 이용하며, CRISPR/Cas9 기술을 사용하여 처리량이 많은 게놈 연구에서 진보를 활용하는 데 도움이 될 수 있다. 예를 들어, 본원에 기재된 ceDNA 벡터는 어려운 세포주에서 Cas9 및 수많은 단일 가이드 RNA (sgRNA)를 발현시키는 데 유용하다. 다중 CRISPR/Cas9-기반 시스템은 전술한 CRISPR/Cas9-기반 시스템과 동일한 방식으로 사용될 수 있다. 다중 CRISPR/Cas는 하기에 기재된 바와 같이 수행될 수 있다: Cong, L et al. Science 819 (2013); Wang et al. Cell 153:910-918 (2013); Ma et al. Nat Biotechnol 34:528-530 (2016) (이들 각각의 내용은 그 전문이 본원에 참고로 포함됨).
기재된 전사 활성화 및 뉴클레아제 기능성에 더하여, 이 시스템은 게놈 아키텍처의 조사 및 내인성 유전자 조절 경로를 포함하여, 다양한 목적으로 후성적 변형을 제어하는 다른 신규한 Cas9-기반 이펙터를 발현하는 데 유용할 것이다. 내인성 유전자 조절이 다수의 효소 사이의 섬세한 균형이기 때문에, 상이한 기능성을 갖는 다중화 Cas9 시스템은 상이한 조절 신호 사이의 복잡한 상호 작용을 조사할 수 있게 한다. 본원에 기재된 벡터는 단일 세트의 gRNA를 사용하여 독립적인 유전자 조작을 가능하게 하기 위해 압타머-변형된 gRNA 및 직교 Cas9와 호환될 수 있어야 한다.
다중 CRISPR/Cas9-기반 시스템은 세포에서 적어도 하나의 내인성 유전자를 활성화시키는 데 사용될 수 있다. 상기 방법은 세포를 변형된 렌티바이러스 벡터와 접촉시키는 단계를 포함한다. 내인성 유전자는 일시적으로 활성화되거나 안정적으로 활성화될 수 있다. 내인성 유전자는 일시적으로 억제되거나 안정적으로 억제될 수 있다. 융합 단백질은 sgRNA와 유사한 수준으로 발현될 수 있다. 융합 단백질은 sgRNA와 비교하여 상이한 수준으로 발현될 수 있다. 세포는 1차 인간 세포일 수 있다.
다중 CRISPR/Cas9-기반 시스템은 세포에서 다중 유전자 편집 방법에 사용될 수 있다. 상기 방법은 세포를 ceDNA 벡터와 접촉시키는 단계를 포함한다. 다중 유전자 편집은 돌연변이 유전자를 교정하거나 이식유전자를 삽입하는 것을 포함할 수 있다. 돌연변이 유전자를 교정하는 것은 돌연변이 유전자를 결실, 재배열 또는 대체하는 것을 포함할 수 있다. 돌연변이 유전자를 교정하는 것은 뉴클레아제-매개 비-동종 말단 결합 또는 상동성-지정 복구를 포함할 수 있다. 다중 유전자 편집은 적어도 하나의 유전자를 결실 또는 교정하는 것을 포함할 수 있으며, 여기서 유전자는 내인성 정상 유전자 또는 돌연변이 유전자이다.
다중 유전자 편집은 적어도 2개의 유전자를 결실 또는 교정하는 것을 포함할 수 있다. 예를 들어, 적어도 2개의 유전자, 적어도 3개의 유전자, 적어도 4개의 유전자, 적어도 5개의 유전자, 적어도 6개의 유전자, 적어도 7개의 유전자, 적어도 8개의 유전자, 적어도 9개의 유전자, 또는 적어도 10개의 유전자는 결실 또는 교정될 수 있다.
다중 CRISPR/Cas9-기반 시스템은 세포에서 유전자 발현의 다중 조절 방법에 사용될 수 있다. 상기 방법은 세포를 변형된 렌티바이러스 벡터와 접촉시키는 단계를 포함한다. 상기 방법은 적어도 하나의 유전자의 유전자 발현 수준을 조절하는 단계를 포함할 수 있다. 적어도 하나의 표적 유전자의 유전자 발현은 적어도 하나의 표적 유전자의 유전자 발현 수준이 적어도 하나의 표적 유전자에 대한 정상 유전자 발현 수준과 비교하여 증가 또는 감소될 때 조절된다. 유전자 발현 수준은 RNA 또는 단백질 수준일 수 있다.
일부 구현예에서, 다수의 유전자의 발현은 다수의 가이드 RNA (예를 들어, 유전자 발현의 다중 조절 또는 "직교 dCas9 시스템")와 함께 다수의 직교 Cas를 도입함으로써 조절되는 것으로 또한 고려된다. 예를 들어, 다른 Cas 단백질 또는 Cas9 단백질. 당업자는 복수의 가이드 RNA가 RNA의 오프-표적 효과 또는 서로에 대한 상호 작용을 최소화하도록 설계되어야 한다는 것을 이해할 것이다. 직교 dCas9 시스템은 다른 원하는 유전자의 억제와 함께 원하는 특정 유전자의 동시 활성화를 허용한다. 예를 들어, 박테리아 종 예를 들면, S. 파이오게네스, N. 메니니기티디스(N. meninigitidis), S. 써모필루스 및 T. 덴티콜라(T. denticola)의 조합으로부터 유래된 복수의 직교 Cas 단백질 (예를 들어, Cas9 단백질)은 예를 들어, 하기에 기재된 바와 같이 조합으로 사용될 수 있다: Esvelt, K et al. Nature Methods 10(11):1116-1121 (2013) (이는 그 전문이 본원에 참고로 포함됨). 일부 구현예에서, 복수의 가이드 RNA를 인코딩하는 복수의 핵산 서열은 동일한 벡터 상에 존재한다. 또한, 각각의 dCas9는 별개의 유도성 시스템과 쌍을 이룰 수 있으며, 이는 원하는 유전자의 활성화 및/또는 억제를 독립적으로 제어할 수 있게 한다. 또한, 이 유도성 직교 dCas9 시스템은 또한 유전자 발현의 조절을 일시적으로 허용할 수 있다 (예를 들어, Gao et al. Nature Methods Complex transcriptional modulation with orthogonal and inducible dCas9 regulators (2016) 참조).
B. 상동성 지정 복구 주형
일부 구현예에서, 상동성-지정 재조합 주형 또는 "복구" 주형은 또한, 예를 들어, 공여체 서열 및/또는 공여체 서열의 일부로서 ceDNA 벡터에 제공된다. 상동성 지정 복구 주형은 유전자 서열을 복구하거나 새로운 서열을 삽입하기 위해, 예를 들어, 치료 단백질을 제조하는데 사용될 수 있는 것으로 고려된다. 일부 구현예에서, 복구 주형은 본원에 기재된 뉴클레아제, 예를 들어, RNA 유도된 엔도뉴클레아제, 예컨대 CRISPR 복합체의 일부로서 CRISPR 효소, 또는 ZFN 또는 TALE에 의해 닉킹 또는 절단된 표적 서열 내에서 또는 그 근처에서 상동 재조합의 주형으로서 기능하도록 설계된다. 주형 폴리뉴클레오티드는 임의의 적합한 길이, 예컨대 약 10, 15, 20, 25, 50, 75, 100, 150, 200, 500, 1000개 이상의 뉴클레오티드 길이일 수 있다. 일부 구현예에서, 주형 폴리뉴클레오티드는 숙주 세포 게놈에서 표적 서열을 포함하는 폴리뉴클레오티드의 일부에 상보적이다. 최적으로 정렬될 때, 주형 폴리뉴클레오티드는 표적 서열의 하나 이상의 뉴클레오티드 (예를 들어, 약 1, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100개 이상의 뉴클레오티드)와 중첩될 수 있다. 일부 구현예에서, 주형 서열 및 표적 서열을 포함하는 폴리뉴클레오티드가 최적으로 정렬될 때, 주형 폴리뉴클레오티드의 가장 가까운 뉴클레오티드는 표적 서열로부터 약 1, 5, 10, 15, 20, 25, 50, 75, 100, 200, 300, 400, 500, 1000, 5000, 10000개 이상의 뉴클레오티드 내에 있다. 일 구현예에서, 복구 주형의 상동성 아암은 지향성(directional)이다 (즉, 동일하지 않으므로 특정 배향으로 서열에 결합한다). 일부 구현예에서, 세포에서 단일 유전자, 또는 세포에서 2개의 상이한 유전자를 복구하기 위해 2개 이상의 HDR 주형이 제공된다. 일부 구현예에서, 적어도 하나의 주형의 다수의 카피가 세포에 제공된다.
일부 구현예에서, 주형 서열은 내인성 표적 유전자 서열의 일부와 실질적으로 동일할 수 있지만 적어도 하나의 뉴클레오티드 변화를 포함한다. 일부 구현예에서, 절단된 표적 핵산 분자의 복구는, 예를 들어, (i) 표적 유전자로부터 발현된 RNA의 하나 이상의 뉴클레오티드 변화, (ii) 표적 유전자의 발현 수준의 변화, (iii) 유전자 녹다운, (iv) 유전자 녹아웃, (v) 유전자 기능 복원, 또는 (vi) 유전자 녹아웃 및 유전자의 동시 삽입을 초래할 수 있다. 당업자에 의해 쉽게 이해될 수 있는 바와 같이, 주형으로 절단된 표적 핵산 분자의 복구는 표적 유전자의 엑손 서열, 인트론 서열, 조절 서열, 전사 조절 서열, 번역 조절 서열, 스플라이싱 부위, 또는 비-코딩 서열의 변화를 초래할 수 있다. 다른 구현예에서, 주형 서열은 유전자 녹인을 초래할 수 있는 외인성 서열을 포함할 수 있다. 외인성 서열의 통합은 유전자 녹아웃을 초래할 수 있다.
특정 구현예에서, 공여체 서열은 5' 상동성 아암 및/또는 3' 상동성 아암과 같은 하나 이상의 통합 요소를 또한 포함하는 캡시드-비함유 ceDNA 벡터에 있다. 최소한 이러한 특정 구현예에서, ceDNA는 5' 내지 3', 5' HDR 아암, 공여체 서열, 3' HDR 아암 및 적어도 하나의 ITR을 포함하고, 여기서 적어도 하나의 ITR은 5' HDR 아암의 상류 또는 3' HDR 아암의 하류에 있다. 특정 구현예에서, 공여체 서열 (예컨대, 비제한적으로, 인자 IX 또는 인자 VIII (또는 예를 들면, 기타 임의의 관심 치료 단백질)은 숙주 세포의 염색체에 삽입되는 뉴클레오티드 서열이다. 특정 구현예에서, 공여체 서열은 원래 숙주 세포에 존재하지 않거나 숙주 세포와 이질적일 수 있다. 특정 구현예에서, 공여체 서열은 미리 결정된 표적 부위 이외의 부위에 존재하는 내인성 서열이다. 특정 구현예에서, 공여체 서열은 미리 결정된 표적 부위의 것과 유사한 내인성 서열이다 (예를 들어, 기존의 잘못된 서열을 대체함). 특정 구현예에서, 공여체 서열은 숙주 세포에 내인성이지만, 미리 결정된 표적 부위 이외의 부위에 존재하는 서열이다. 일부 구현예에서, 공여체 서열은 코딩 서열 또는 비-코딩 서열이다. 일부 구현예에서, 공여체 서열은 유전자의 돌연변이 유전자좌이다. 특정 구현예에서, 공여체 서열은 염색체에 삽입되는 외인성 유전자, 표적 부위에서의 내인성 서열을 대체하는 변형된 서열, 조절 요소, 리포터 단백질 및/또는 RNA를 인코딩하는 태그 또는 코딩 서열일 수 있다. 일부 구현예에서, 공여체 서열은 융합 단백질의 발현을 위해 표적 유전자의 코딩 서열 내로 프레임으로 삽입될 수 있다. 특정 구현예에서, 공여체 서열은 전체 ORF (코딩/공여체 서열)가 아니라 원하는 표적을 대체하기 위한 DNA의 교정 부분일 뿐이다. 특정 구현예에서, 공여체 서열은 천연 발생 서열과 유사하게 공여체 서열이 조절되도록 내인성 프로모터 뒤에 프레임으로(in-frame) 삽입된다.
특정 구현예에서, 공여체 서열은 코딩 서열을 구동하기 위해 상기 기재된 바와 같은 본원의 프로모터를 임의로 포함할 수 있다. 이러일 구현예는 발현을 용이하게 하기 위해 공여체 서열 내에 폴리-A 테일을 추가로 포함할 수 있다.
특정 구현예에서, 공여체 서열은 미리 결정된 크기이거나, 당업자에 의해 크기가 정해질 수 있다. 특정 구현예에서, 공여체 서열은 적어도 또는 약 10개의 염기쌍, 15개의 염기쌍, 20개의 염기쌍, 25개의 염기쌍, 50개의 염기쌍, 60개의 염기쌍, 75개의 염기쌍, 100개의 염기쌍, 적어도 150개의 염기쌍, 200개의 염기쌍, 300개의 염기쌍, 500개의 염기쌍, 800개의 염기쌍, 1000개의 염기쌍, 1,500개의 염기쌍, 2,000개의 염기쌍, 2500개의 염기쌍, 3000개의 염기쌍, 4000개의 염기쌍, 4500개의 염기쌍 및 5,000개의 염기쌍 길이 또는 약 1개의 염기쌍 내지 약 10개의 염기쌍, 또는 약 10개의 염기쌍 내지 약 50개의 염기쌍, 또는 약 50개의 염기쌍 내지 약 100개의 염기쌍, 또는 약 100개의 염기쌍 내지 약 500개의 염기쌍, 또는 약 500개의 염기쌍 내지 약 5,000개의 염기쌍 길이 중 어느 것일 수 있다. 특정 구현예에서, 공여체 서열은 게놈에서 단일 돌연변이된 뉴클레오티드를 복구하기 위해 단 1개의 염기쌍만을 포함한다.
본 개시내용에 따라 사용하기에 적합한 공여체 서열(들)의 비제한적 예는 본원에 기재된 질환-관련 분자 중 하나와 적어도 60%, 보다 바람직하게는 적어도 65%, 보다 바람직하게는 70%, 보다 바람직하게는 적어도 75%, 보다 바람직하게는 적어도 80%, 보다 바람직하게는 적어도 85%, 더욱 더 바람직하게는 적어도 90%, 가장 바람직하게는 적어도 95%의 서열 동일성을 갖는 하나 이상의 질환-관련 서열에 상응하는 프로모터가 없는 코딩 서열을 포함한다. 일 구현예에서, 코딩 서열은 서열 번호: 825 또는 서열 번호: 825로 구성된 공여체 서열과 적어도 60%, 보다 바람직하게는 적어도 65%, 보다 바람직하게는 적어도 70%, 보다 바람직하게는 적어도 75%, 보다 바람직하게는 적어도 80%, 보다 바람직하게는 적어도 85%, 더욱 더 바람직하게는 적어도 90%, 가장 바람직게는 적어도 95%의 서열 동일성을 갖는다. 서열이 대체되기보다는 첨가되는 것과 같은 특정 구현예에서, 프로모터가 제공될 수 있다.
공여체 서열을 숙주 세포 게놈 내로 통합하기 위해, ceDNA 벡터는 본원에 나타낸 5' 및 3' 상동성 아암과 같은 상동성 재조합에 의해 게놈 내로 통합시키기 위해 공여체 서열 또는 벡터의 임의의 다른 요소를 인코딩하는 폴리뉴클레오티드 서열에 의존할 수 있다. (예를 들어, 도 8 참조). 예를 들어, ceDNA 벡터는 염색체(들) 내의 정확한 위치(들)에서 숙주 세포의 게놈 내로 상동성 재조합에 의해 통합을 지시하기 위해 5' 및 3' 상동성 아암을 인코딩하는 뉴클레오티드를 함유할 수 있다. 정확한 위치에서의 통합 가능성을 증가시키기 위해, 5' 및 3' 상동성 아암은 충분한 수의 핵산, 예컨대 50 내지 5,000개의 염기쌍, 또는 100 내지 5,000개의 염기쌍, 또는 500 내지 5,000개의 염기쌍을 포함할 수 있고, 이는 상동성 재조합의 가능성을 향상시키기 위해 상응하는 표적 서열에 대해 높은 정도의 서열 동일성 또는 상동성을 갖는다. 5' 및 3' 상동성 아암은 숙주 세포의 게놈에서 표적 서열과 상동성인 임의의 서열일 수 있다. 또한, 5' 및 3' 상동성 아암은 뉴클레오티드 서열을 인코딩하지 않거나 인코딩할 수 있다. 특정 구현예에서, 5' 상동성 아암과 염색체 상의 상응하는 서열 사이의 상동성은 적어도 80%, 85%, 90%, 95%, 97%, 98%, 99% 또는 100% 중 어느 것이다. 특정 구현예에서, 3' 상동성 아암과 염색체 상의 상응하는 서열 사이의 상동성은 적어도 80%, 85%, 90%, 95%, 97%, 98%, 99% 또는 100% 중 어느 것이다. 특정 구현예에서, 5' 및/또는 3' 상동성 아암은 염색체 상의 통합 또는 DNA 절단 부위의 바로 상류 및/또는 하류에 있는 서열과 상동성일 수 있다. 대안적으로, 5' 및/또는 3' 상동성 아암은 통합 또는 DNA 절단 부위로부터 먼 서열, 예컨대 통합 또는 DNA 절단 부위로부터 적어도 1, 2, 5, 10, 15, 20, 25, 30, 50, 100, 200, 300, 400 또는 500 bp 떨어져 있거나, 또는 DNA 절단 부위와 부분적으로 또는 완전히 중첩되는 서열과 상동성일 수 있다. 특정 구현예에서, 뉴클레오티드 서열의 3' 상동성 아암은 변경된 ITR에 근접하다.
특정 구현예에서, 공여체 서열의 통합 효율은 통합 전에 ceDNA 벡터로부터 공여체 서열을 포함하는 카세트의 추출에 의해 개선된다. 하나의 비제한적인 예에서, 특정 제한 부위는 5' 상동성 아암으로 5'에서, 3' 상동성 아암으로 3'에서 또는 둘 다로 조작될 수 있다. 그러한 제한 부위가 두 상동성 아암 모두에 대해 존재한다면, 제한 부위는 두 상동성 아암 사이에서 동일하거나 상이할 수 있다. ceDNA 벡터가 조작된 제한 부위(들)에 특이적인 하나 이상의 제한 엔도뉴클레아제로 절단될 때, 생성된 카세트는 5' 상동성 아암-공여체 서열 -3' 상동성 아암을 포함하고, 원하는 게놈 유전자좌와 보다 용이하게 재조합될 수 있다. 당업자는 이 절단된 카세트가, 비제한적으로, 조절 영역, 뉴클레아제 및 추가 공여체 서열 중 하나 이상과 같은 다른 요소를 추가로 포함할 수 있다는 것을 이해할 것이다. 특정 측면에서, ceDNA 벡터 자체는 제한 엔도뉴클레아제를 인코딩하여 ceDNA 벡터를 핵으로 전달할 때 제한 엔도뉴클레아제가 발현되고 벡터를 절단할 수 있도록 할 수 있다. 특정 측면에서, 제한 엔도뉴클레아제는 별도로 전달되는 제2 ceDNA 벡터에 인코딩된다. 특정 측면에서, 제한 엔도뉴클레아제는 비-ceDNA-기반 전달 수단에 의해 핵에 도입된다. 특정 구현예에서, 제한 엔도뉴클레아제는 ceDNA 벡터가 핵으로 전달된 후에 도입된다. 특정 구현예에서, 제한 엔도뉴클레아제 및 ceDNA 벡터는 동시에 핵으로 운반된다. 특정 구현예에서, 제한 엔도뉴클레아제는 ceDNA 벡터의 도입시 이미 존재한다.
특정 구현예에서, 공여체 서열은 5' 상동성 아암 또는 3' 상동성 아암과 이질적이다. 특정 구현예에서, 공여체 서열은 5' 상동성 아암 및 3' 상동성 아암을 포함하는 서열 사이에서 내생적으로 발견되지 않는다. 특정 구현예에서, 공여체 서열은 5' 상동성 아암 또는 3' 상동성 아암을 포함하는 고유 서열에 대해 내생적이지 않다. 특정 구현예에서, 5' 상동성 아암은 염색체 상의 뉴클레아제 절단 부위의 상류에 있는 뉴클레오티드 서열과 상동성이다. 특정 구현예에서, 3' 상동성 아암은 염색체 상의 뉴클레아제 절단 부위의 하류에 있는 뉴클레오티드 서열과 상동성이다. 특정 구현예에서, 5' 상동성 아암 또는 3' 상동성 아암은 적어도 하나의 변경된 ITR에 근접한다. 특정 구현예에서, 5' 상동성 아암 또는 3' 상동성 아암은 약 250 내지 2000 bp이다.
본 개시내용에 따라 사용하기 위한, 특히 간 세포 또는 조직의 유전자 편집에 사용하에 적합한 5' 상동성 아암의 비제한적 예는 서열 번호: 823 또는 서열 번호: 826 내의 적합한 분절, 또는 서열 번호: 823 내의 적합한 분절 또는 서열 번호: 826 내의 적합한 분절로 구성된 5' 상동성 아암과 적어도 60%, 보다 바람직하게는 적어도 65%, 보다 바람직하게는 적어도 70%, 보다 바람직하게는 적어도 75%, 보다 바람직하게는 적어도 80%, 보다 바람직하게는 적어도 85%, 더욱 더 바람직하게는 적어도 90%, 가장 바람직하게는 적어도 95%의 서열 동일성을 갖는 5' 알부민 상동성 아암을 포함한다. 이러한 분절은 모든 각각의 서열일 수 있다.
본 개시내용에 따라 사용하기에 적합한 3' 상동성 아암의 비제한적 예는 서열 번호: 824 또는 서열 번호: 14 827 내의 적합한 분절, 또는 서열 번호: 824 또는 서열 번호: 827 내의 적합한 분절로 구성된 3' 상동성 아암과 적어도 60%, 보다 바람직하게는 적어도 65%, 보다 바람직하게는 적어도 70%, 보다 바람직하게는 적어도 75%, 보다 바람직하게는 적어도 80%, 보다 바람직하게는 적어도 85%, 더욱 더 바람직하게는 적어도 90%, 가장 바람직하게는 적어도 95%의 서열 동일성을 갖는 3' 알부민 상동성 아암을 포함한다. 이러한 분절은 모든 각각의 서열일 수 있다.
일 구현예에서, 본원에 기재된 바와 같이 공여체 서열의 측면에 5'- 및 3' 상동성 아암을 포함하는 유전자 편집 ceDNA 벡터는 Cas 닉카아제 (nCas; 예를 들어, Cas9 닉카아제)를 인코딩하는 또 다른 벡터 (예를 들어, 추가 ceDNA 벡터, 렌티바이러스 벡터, 바이러스 벡터 또는 플라스미드)와 함께 투여될 수 있다. 본원에서 이러한 nCas 효소는 본원에 기재된 바와 같은 ceDNA 벡터에 대한 상동성을 포함하는 가이드 RNA와 함께 사용되며, 예를 들어, 물리적으로 제한된 서열을 방출하거나 비틀림 해제를 제공하는 데 사용될 수 있는 것으로 고려된다. 물리적으로 제한된 서열의 방출은, 예를 들어, ceDNA 벡터 내의 상동성 지정 복구 (HDR) 주형 상동성 아암 (들)이 게놈 서열과의 상호 작용을 위해 노출되도록 ceDNA 벡터를 "언와인드(unwind)"할 수 있다. 또한, 필요한 경우, 이러한 시스템을 사용하여 ceDNA 벡터를 불활성화시킬 수 있는 것으로 고려된다. ceDNA 벡터에서 이중 가닥 파괴를 유도하는 Cas 효소는 이러한 ceDNA 벡터의 더 강한 불활성화제일 것이라는 것이 당업자에 의해 이해될 것이다. 일 구현예에서, 가이드 RNA는 뉴클레아제를 인코딩하는 서열 또는 공여체 서열 또는 주형과 같은 ceDNA 벡터에 삽입된 서열에 대한 상동성을 포함한다. 또 다른 구현예에서, 가이드 RNA는 역 말단 반복부 (ITR)에 대한 상동성 또는 ceDNA 벡터의 상동성/삽입 요소를 포함한다. 일부 구현예에서, 본원에 기재된 바와 같은 ceDNA 벡터는 5' 및 3' 말단 각각에 ITR을 포함하므로, ITR과 상동성을 갖는 가이드 RNA는 하나 이상의 ITR의 닉킹을 실질적으로 동일하게 생성할 것이다. 일부 구현예에서, 가이드 RNA는 ceDNA 벡터의 일부 및 공여체 서열 또는 주형에 대해 상동성을 갖는다 (예를 들어, ceDNA 벡터의 언와인딩을 보조하기 위해). 또한, 닉킹될 때 ceDNA 벡터가 핵에 유지될 수 없게 하는 ceDNA 벡터 상에 특정 부위가 존재하는 것으로 고려된다. 당업자는 이러한 서열을 쉽게 식별할 수 있고, 따라서 이러한 서열 영역에 대한 엔지니어링 가이드 RNA를 피할 수 있다. 대안적으로, 핵 국재화를 담당하는 서열을 닉킹하는 가이드 RNA를 사용하여 뉴클레아제 외부 영역으로 ceDNA 벡터의 세포하 국재화를 변형시키는 것은, 필요하거나 원하는 경우, ceDNA 벡터를 불활성화시키는 방법으로서 사용될 수 있다.
특정 구현예에서, 다른 통합 전략 및 성분은 본 개시내용의 ceDNA 벡터에 따라 사용하기에 적합하다. 예를 들어, 도 1a-1g 또는 도 8 또는 도 9에는 도시되지 않았지만, 일 구현예에서, 본 개시내용에 따른 ceDNA 벡터는 게놈 rDNA 내로 상동성 재조합을 할 수 있는 리보솜 DNA (rDNA) 서열 측면에 있는 발현 카세트를 포함할 수 있다. 예를 들어, rAAV-rDNA 벡터가 입증된, 하기와 같은 유사한 전략이 수행되었다: Lisowski, et al., Ribosomal DNA Integrating rAAV-rDNA Vectors Allow for Stable Transgene Expression, The American Society of Gene and Cell Therapy, 18 September 2012 (그 전문이 본원에 참고로 포함됨). 특정 구현예에서, ceDNA-rDNA 벡터의 전달은 증가된 빈도로 게놈 rDNA 유전자좌에 통합될 수 있으며, 여기서 통합은 rDNA 유전자좌에 특이적이다. 또한, 인간 인자 IX (hFIX) 또는 인간 인자 VIII 발현 카세트를 함유하는 ceDNA-rDNA 벡터는 혈청 hFIX 또는 인간 인자 VIII의 치료 수준을 증가시킨다. rDNA 유전자좌에서의 통합의 상대적 안전성으로 인해, ceDNA-rDNA 벡터는 분열 세포로의 장기 유전자 전이를 필요로 하는 치료제에 대한 ceDNA의 사용을 확장시킨다.
일 구현예에서, 프로모터가 없는 ceDNA 벡터는 상동성 복구 주형 (예를 들어, 2개의 플랭킹 상동성 아암을 갖는 복구 서열)의 전달을 위해 고려되지만, 뉴클레아제 또는 가이드 RNA를 인코딩하는 핵산 서열을 포함하지는 않는다.
본원에 기재된 방법 및 조성물은, 예를 들어, 하기에 기재된 바와 같이 상동성 재조합을 포함하는 방법에 사용될 수 있다: Rouet et al. Proc Natl Acad Sci 91:6064-6068 (1994); Chu et al. Nat Biotechnol 33:543-548 (2015); Richardson et al. Nat Biotechnol 33:339-344 (2016); Komor et al. Nature 533:420-424 (2016) (이들 각각의 내용은 그 전문이 본원에 참고로 포함됨).
본원에 기재된 방법 및 조성물은, 예를 들어, 하기에 기재된 바와 같이 상동성 재조합을 포함하는 방법에 사용될 수 있다: Rouet et al. Proc Natl Acad Sci 91:6064-6068 (1994); Chu et al. Nat Biotechnol 33:543-548 (2015); Richardson et al. Nat Biotechnol 33:339-344 (2016); Komor et al. Nature 533:420-424 (2016) (이들 각각의 내용은 그 전문이 본원에 참고로 포함됨).
C. 가이드 RNA (gRNA)
일반적으로, 가이드 서열은 표적 서열과 하이브리드화되고 선택된 게놈 표적 서열에 대한 RNA-유도 엔도뉴클레아제 복합체의 서열-특이적 표적화를 지시하기 위해 표적 폴리뉴클레오티드 서열과 충분한 상보성을 갖는 임의의 폴리뉴클레오티드 서열이다. 일부 구현예에서, 가이드 RNA는 결합하고, 예를 들어, Cas 단백질은 리보핵단백질 (RNP), 예를 들어, CRISPR/Cas 복합체를 형성할 수 있다.
일부 구현예에서, 가이드 RNA (gRNA) 서열은 가이드 서열과 RNA-유도 엔도뉴클레아제의 회합을 허용하는 crRNA 및/또는 tracrRNA 서열에 융합된, gRNA 서열을 게놈의 원하는 부위로 유도하는 표적화 서열을 포함한다. 일부 구현예에서, 적합한 정렬 알고리즘을 사용하여 최적으로 정렬될 때, 가이드 서열 및 이의 상응하는 표적 서열 사이의 상보성의 정도는 적어도 50%, 60%, 75%, 80%, 85%, 90%, 95%, 97.5%, 99% 이상이다. Smith-Waterman 알고리즘, Needleman-Wunsch 알고리즘, Burrows-Wheeler Transform (예를 들어, Burrows Wheeler Aligner), ClustalW, Clustal X, BLAT, Novoalign (Novocraft Technologies, ELAND (Illumina, San Diego, CA), SOAP 및 Maq에 기반하는 알고리즘과 같이, 서열 정렬에 적합한 임의의 알고리즘을 사용하여 최적의 정렬을 결정할 수 있다. 일부 구현예에서, 가이드 서열은 5, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50, 75개 이상의 뉴클레오티드 길이이다. 본원에서 가이드 RNA의 표적화 서열 및 표적 핵산 분자 상의 표적 서열은 1, 2, 3, 4, 5, 6, 7, 8, 9 또는 10개의 미스매치를 포함할 수 있는 것으로 고려된다. 일부 구현예에서, 가이드 RNA 서열은 팔린드롬 서열을 포함하고, 예를 들어, 자기-표적 서열은 팔린드롬을 포함한다. 가이드 RNA의 표적화 서열은 전형적으로 19-21개의 염기쌍 길이이고, 전체 가이드 RNA (표적화 서열 + 헤어핀)를 Cas9와 같은 Cas에 결합시키는 헤어핀 바로 앞에 있다. 팔린드롬 서열이 가이드 RNA의 자기-표적화 서열로서 사용되는 경우, 역 반복 요소는 예를 들어, 9, 10, 11, 12개 이상의 뉴클레오티드 길이일 수 있다. 가이드 RNA의 표적화 서열이 가장 흔히 19-21 bp인 경우, 9개 또는 10개 뉴클레오티드의 팔린드롬 역 반복 요소는 바람직한 길이의 표적화 서열을 제공한다. 이후 Cas9- 가이드 RNA 헤어핀 복합체는 임의의 뉴클레오티드 서열 (DNA 또는 RNA), 예를 들어, 19-21개의 염기쌍 서열과 일치하는 DNA 서열을 인식하고 절단할 수 있고, 이어서 "PAM" 서열, 예를 들어, NGG 또는 NGA, 또는 다른 PAM이 뒤따른다.
RNA 유도된 엔도뉴클레아제 복합체의 서열-특이적 결합을 표적 서열에 유도하는 가이드 서열의 능력은 임의의 적합한 분석에 의해 평가될 수 있다. 예를 들어, RNA-유도 엔도뉴클레아제 복합체를 형성하기에 충분한 RNA-유도 엔도뉴클레아제 시스템의 성분은 상응하는 표적 서열을 갖는 숙주 세포에, 예컨대 RNA-유도 엔도뉴클레아제 서열의 성분을 인코딩하는 벡터를 이용한 형질감염에 의해 제공될 수 있으며, 이어서 Surveyor 분석법 (Transgenomic™, New Haven, CT)과 같은, 표적 서열 내의 우선적 절단 평가가 뒤따른다. 유사하게, 표적 폴리뉴클레오티드 서열의 절단은 표적 서열, 시험될 가이드 서열 및 시험 가이드 서열과 상이한 대조 가이드 서열을 포함하는, RNA-유도 엔도뉴클레아제 복합체의 성분을 제공하고, 시험 및 대조 가이드 서열 반응 사이에 표적 서열에서의 결합 또는 절단 속도를 비교함으로써 시험관에서 평가될 수 있다. 당업자는 다른 분석법이 gRNA 서열을 시험하는데 또한 사용될 수 있음을 이해할 것이다.
임의의 표적 서열을 표적화하기 위해 가이드 서열을 선택할 수 있다. 일부 구현예에서, 표적 서열은 세포의 게놈 내의 서열이다. 일부 구현예에서, 표적 서열은 본원에 기재된 바와 같이, 자가-클로닝 플라스미드에서 제1 가이드 RNA를 인코딩하는 서열이다. 전형적으로, 게놈 내 표적 서열은 RNA-유도 엔도뉴클레아제의 결합을 위한 프로토스페이서 인접 (PAM) 서열을 포함할 것이다. 당업자는 PAM 서열 및 RNA-유도 엔도뉴클레아제가 엔도뉴클레아제와 표적화 서열의 적절한 회합을 허용하기 위해 동일한 (박테리아) 종으로부터 선택되어야 한다는 것을 이해할 것이다. 예를 들어, CAS9의 PAM 서열은 cpF1의 PAM 서열과 다르다. 설계는 적절한 PAM 서열을 기반으로 한다. 가이드 RNA의 분해를 방지하기 위해, 가이드 RNA의 서열은 PAM 서열을 함유해서는 안된다. 일부 구현예에서, 가이드 RNA에서 표적화 서열의 길이는 12개 뉴클레오티드이고; 다른 구현예에서, 가이드 RNA에서 표적화 서열의 길이는 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35 또는 40개의 뉴클레오티드이다. 가이드 RNA는 표적화된 DNA 서열의 양 가닥에 상보적일 수 있다. 일부 구현예에서, 삽입 또는 결실을 포함하도록 게놈을 변형시키는 경우, gRNA는 단백질 코딩 영역의 N-말단에 더 가깝게 표적화될 수 있다.
RNA-유도 엔도뉴클레아제에 의한 표적화된 절단의 목적상, 게놈에서 고유한 표적 서열이 게놈에서 1회 이상 발생하는 표적 서열보다 바람직하다는 것이 당업자에 의해 이해될 것이다. 생물 정보학 소프트웨어는 가이드 RNA의 오프-표적 효과를 예측하고 최소화하기 위해 사용될 수 있다 (예를 들어, NNaito et al. "CRISPRdirect: software for designing CRISPR/Cas guide RNA with reduced off-target sites" Bioinformatics (2014), epub; Heigwer, F., et al. "E-CRISP: fast CRISPR target site identification" Nat. Methods 11, 122-123 (2014); Bae et al. "Cas-OFFinder: a fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases" Bioinformatics 30(10):1473-1475 (2014); Aach et al. "CasFinder: Flexible algorithm for identifying specific Cas9 targets in genomes" BioRxiv (2014), 등 참조).
S. 파이오게네스 Cas9의 경우, 게놈 내 고유한 표적 서열은 MMMMMMMMNNNNNNNNNNNNXGG (서열 번호: 590) 형태의 Cas9 표적 부위를 포함할 수 있으며, 여기서 NNNNNNNNNNNNXGG (서열 번호: 591) (N은 A, G, T 또는 C이고; X는 임의의 뉴클레오티드일 수 있음)는 게놈에서 단일 발생을 갖는다. 게놈 내 고유한 표적 서열은 MMMMMMMMMNNNNNNNNNNNXGG (서열 번호: 592) 형태의 S. 파이오게네스 Cas9 표적 부위를 포함할 수 있으며, 여기서 NNNNNNNNNNNXGG (서열 번호: 593) (N은 A, G, T 또는 C이고; X는 임의의 뉴클레오티드일 수 있음)는 게놈에서 단일 발생을 갖는다. S. 써모필루스 CRISPR1 Cas9의 경우, 게놈 내 고유한 표적 서열은 MMMMMMMMNNNNNNNNNNNNXXAGAAW (서열 번호: 594) 형태의 Cas9 표적 부위를 포함할 수 있으며, 여기서 NNNNNNNNNNNNXXAGAAW (서열 번호: 595) (N은 A, G, T 또는 C이고; X는 임의의 뉴클레오티드일 수 있고; W는 A 또는 T임)는 게놈에서 단일 발생을 갖는다. 게놈 내 고유한 표적 서열은 MMMMMMMMMNNNNNNNNNNNXXAGAAW (서열 번호: 596) 형태의 S. 써모필루스 CRISPR 1 Cas9 표적 부위를 포함할 수 있으며, 여기서 NNNNNNNNNNNXXAGAAW (서열 번호: 597) (N은 A, G, T 또는 C이고; X는 임의의 뉴클레오티드일 수 있고; W는 A 또는 T임)는 게놈에서 단일 발생을 갖는다. S. 파이오게네스 Cas9의 경우, 게놈 내 고유한 표적 서열은 MMMMMMMMNNNNNNNNNNNNXGGXG (서열 번호: 598) 형태의 Cas9 표적 부위를 포함할 수 있으며, 여기서 NNNNNNNNNNNNXGGXG (서열 번호: 599) (N은 A, G, T 또는 C이고; X는 임의의 뉴클레오티드일 수 있음)는 게놈에서 단일 발생을 갖는다. 게놈 내 고유한 표적 서열은 MMMMMMMMMNNNNNNNNNNNXGGXG (서열 번호: 600) 형태의 S. 파이오게네스 Cas9 표적 부위를 포함할 수 있으며, 여기서 NNNNNNNNNNNXGGXG (서열 번호: 601) (N은 A, G, T 또는 C이고; X는 임의의 뉴클레오티드일 수 있음)는 게놈에서 단일 발생을 갖는다. 이들 각각의 서열에서 "M"은 A, G, T 또는 C일 수 있으며, 서열을 고유한 것으로 식별하는 데 고려될 필요는 없다.
일반적으로, 본원에 사용된 용어 "crRNA/tracrRNA 융합 서열"은 고유한 표적화 서열에 융합되고, 가이드 RNA 및 RNA-유도 엔도뉴클레아제를 포함하는 복합체의 형성을 허용하는 기능을 하는 핵산 서열을 지칭한다. 이러한 서열은 (i) 본원에 기재된 바와 같은 표적 서열에 상응하는 "프로토스페이서"로 명명된 가변 서열, 및 (ii) CRISPR 반복부를 포함하는, 원핵생물에서 CRISPR RNA (crRNA) 서열 후에 모델링될 수 있다. 유사하게, 융합물의 tracrRNA ("전사 활성화 CRISPR RNA") 부분은 엔도뉴클레아제 복합체의 형성을 허용하기 위해 원핵생물의 tracrRNA 서열과 유사한 이차 구조 (예를 들어, 헤어핀)를 포함하도록 설계될 수 있다. 일부 구현예에서, 융합은 tracrRNA 서열과의 충분한 상보성을 가지므로, (1) 상응하는 tracr 서열을 함유하는 세포에서 tracrRNA 서열의 측면에 있는 가이드 서열의 절제; 및 (2) 표적 서열에서 엔도뉴클레아제 복합체의 형성 (여기서 상기 복합체는 tracrRNA 서열에 하이브리드화된 crRNA 서열을 포함함) 중 하나 이상을 촉진하기 위해 tracrRNA 서열과 충분한 상보성을 갖는다. 일반적으로, 상보성의 정도는 두 서열 중 더 짧은 서열의 길이를 따라, crRNA 서열 및 tracrRNA 서열의 최적 정렬과 관련된다. 최적의 정렬은 임의의 적합한 정렬 알고리즘에 의해 결정될 수 있고, tracrRNA 서열 또는 crRNA 서열 내의 자기 상보성과 같이, 2차 구조를 추가로 설명할 수 있다. 일부 구현예에서, 최적으로 정렬된 경우 둘 중 더 짧은 길이를 따라 tracrRNA 서열과 crRNA 서열 사이의 상보성의 정도는 약 25%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97.5%, 99% 이상이다. 일부 구현예에서, tracrRNA 서열은 적어도 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 40, 50, 60, 70, 80, 90, 100 또는 그 이상의 뉴클레오티드 길이 (예를 들어, 70-80, 70-75, 75-80 뉴클레오티드 길이)이다. 일 구현예에서, crRNA는 60개 미만, 50개 미만, 40개 미만, 30개 미만, 또는 20개 미만의 뉴클레오티드 길이이다. 다른 구현예에서, crRNA는 30-50개의 뉴클레오티드 길이이고; 다른 구현예에서 crRNA는 30-50, 35-50, 40-50, 40-45, 45-50 또는 50-55개의 뉴클레오티드 길이이다. 일부 구현예에서, crRNA 서열 및 tracrRNA 서열은 단일 전사체 내에 함유되어 있어서, 둘 사이의 하이브리드화는 헤어핀과 같은 2차 구조를 갖는 전사체를 생성한다. 일부 구현예에서, 헤어핀 구조에 사용하기 위한 루프 형성 서열은 길이가 4개의 뉴클레오티드, 예를 들어 서열 GAAA이다. 그러나, 대안적 서열처럼, 더 길거나 더 짧은 루프 서열이 사용될 수 있다. 서열은 바람직하게는 뉴클레오티드 삼중항 (예를 들어, AAA) 및 추가의 뉴클레오티드 (예를 들어 C 또는 G)를 포함한다. 루프 형성 서열의 예는 CAAA 및 AAAG를 포함한다. 일 구현예에서, 전사체 또는 전사된 gRNA 서열은 적어도 하나의 헤어핀을 포함한다. 일 구현예에서, 전사체 또는 전사된 폴리뉴클레오티드 서열은 적어도 2개 또는 그 이상의 헤어핀을 갖는다. 다른 구현예에서, 전사체는 2, 3, 4 또는 5개의 헤어핀을 갖는다. 추가의 구현예에서, 전사체는 최대 5개의 헤어핀을 갖는다. 일부 구현예에서, 단일 전사체는 전사 종결 서열, 예컨대 폴리 T 서열, 예를 들어, 6개의 T 뉴클레오티드를 추가로 포함한다. 가이드 서열, crRNA 서열 및 tracr 서열을 포함하는 단일 폴리뉴클레오티드의 비제한적인 예는 다음과 같으며 (5'에서 3'로 열거됨), 여기서 "N"은 가이드 서열의 염기를 나타내고, 소문자의 첫 번째 블록은 crRNA 서열을 나타내고, 소문자의 두 번째 블록은 tracr 서열을 나타내며, 최종 폴리-T 서열은 전사 종결자를 나타낸다:
(i) NNNNNNNNNNNNNNNNNNNNgtttttgtactctcaagatttaGAAAtaaatcttgcagaagctacaaagataaggctt catgccgaaatcaacaccctgtcattttatggcagggtgttttcgttatttaaTTTTTT (서열 번호: 602); (ii) NNNNNNNNNNNNNNNNNNNNgtttttgtactctcaGAAAthcagaagctacaaagataaggcttcatgccgaaatca acaccctgtcattttatggcagggtgttttcgttatttaaTTTTTT (서열 번호: 603); (iii) NNNNNNNNNNNNNNNNNNNNgtttttgtactctcaGAAAtgcagaagctacaaagataaggcttcatgccgaaatca acaccctgtcattttatggcagggtgtTTTTTT (서열 번호: 604); (iv) NNNNNNNNNNNNNNNNNNNNgttttagagctaGAAAtagcaagttaaaataaggctagtccgttatcaacttgaaaa agtggcaccgagtcggtgcTTTTTT (서열 번호: 605); (v) NNNNNNNNNNNNNNNNNNNNgttttagagctaGAAATAGcaagttaaaataaggctagtccgttatcaacttgaa aaagtTTTTTTT (서열 번호: 606); 및 (vi) NNNNNNNNNNNNNNNNNNNNgttttagagctagAAATAGcaagttaaaataaggctagtccgttatcaTTTTT TTT (서열 번호: 607). 일부 구현예에서, 서열 (i) 내지 (iii)은 S. 써모필루스 CRISPR1로부터의 Cas9와 조합하여 사용된다. 일부 구현예에서, 서열 (iv) 내지 (vi)은 S. 파이오게네스로부터의 Cas9와 조합하여 사용된다. 일부 구현예에서, tracrRNA 서열은 crRNA 서열을 포함하는 전사체와 별개의 전사체이다.
일부 구현예에서, 가이드 RNA는 2개의 RNA 분자를 포함할 수 있고, 본원에서 "이중 가이드 RNA" 또는 "dgRNA"로 지칭된다. 일부 구현예에서, dgRNA는 crRNA를 포함하는 제1 RNA 분자, 및 tracrRNA를 포함하는 제2 RNA 분자를 포함할 수 있다. 제1 및 제2 RNA 분자는 crRNA 상의 플래그폴(flagpole)과 tracrRNA 사이의 염기 짝짓기를 통해 RNA 듀플렉스를 형성할 수 있다. dgRNA를 사용할 때, 플래그폴은 길이에 대해 상한을 가질 필요는 없다.
다른 구현예에서, 가이드 RNA는 단일 RNA 분자를 포함할 수 있고, 본원에서 "단일 가이드 RNA" 또는 "sgRNA"로 지칭된다. 일부 구현예에서, sgRNA는 tracrRNA에 공유적으로 연결된 crRNA를 포함할 수 있다. 일부 구현예에서, crRNA 및 tracrRNA는 링커를 통해 공유적으로 연결될 수 있다. 일부 구현예에서, sgRNA는 crRNA 상의 플래그폴과 tracrRNA 사이의 염기 짝짓기를 통해 스템-루프 구조를 포함할 수 있다. 일부 구현예에서, 단일-가이드 RNA는 적어도 50, 적어도 60, 적어도 70, 적어도 80, 적어도 90, 적어도 100, 적어도 110, 적어도 120개 또는 그 이상의 뉴클레오티드 길이 (예를 들어, 75-120, 75-110, 75-100, 75-90, 75-80, 80-120, 80-110, 80-100, 80-90, 85-120, 85-110, 85-100, 85-90, 90-120, 90-110, 90-100, 100-120, 100-120개의 뉴클레오티드 길이)이다. 일부 구현예에서, ceDNA 벡터 또는 이의 조성물은 적어도 1개의 gRNA를 인코딩하는 핵산을 포함한다. 예를 들어, 제2 폴리뉴클레오티드 서열은 적어도 1개 gRNA, 적어도 2개 gRNA, 적어도 3개 gRNA, 적어도 4개 gRNA, 적어도 5개 gRNA, 적어도 6개 gRNA, 적어도 7개 gRNA, 적어도 8개 gRNA, 적어도 9개 gRNA, 적어도 10개 gRNA, 적어도 11개 gRNA, 적어도 12개 gRNA, 적어도 13개 gRNA, 적어도 14개 gRNA, 적어도 15개 gRNA, 적어도 16개 gRNA, 적어도 17개 gRNA, 적어도 18개 gRNA, 적어도 19개 gRNA, 적어도 20개 gRNA, 적어도 25개 gRNA, 적어도 30개 gRNA, 적어도 35개 gRNA, 적어도 40개 gRNA, 적어도 45개 gRNA, 또는 적어도 50개 gRNA를 인코딩할 수 있다. 제2 폴리 뉴클레오티드 서열은 1개 gRNA 내지 50개 gRNA, 1개 gRNA 내지 45개 gRNA, 1개 gRNA 내지 40개 gRNA, 1개 gRNA 내지 35개 gRNA, 1개 gRNA 내지 30개 gRNA, 1개 gRNA 내지 25개의 상이한 gRNA, 1개 gRNA 내지 20개 gRNA, 1개 gRNA 내지 16개 gRNA, 1개 gRNA 내지 8개의 상이한 gRNA, 4개의 상이한 gRNA 내지 50개의 상이한 gRNA, 4개의 상이한 gRNA 내지 45개의 상이한 gRNA, 4개의 상이한 gRNA 내지 40개의 상이한 gRNA, 4개의 상이한 gRNA 내지 35개의 상이한 gRNA, 4개의 상이한 gRNA 내지 30개의 상이한 gRNA, 4개의 상이한 gRNA 내지 25개의 상이한 gRNA, 4개의 상이한 gRNA 내지 20개의 상이한 gRNA, 4개의 상이한 gRNA 내지 16개의 상이한 gRNA, 4개의 상이한 gRNA 내지 8개의 상이한 gRNA, 8개의 상이한 gRNA 내지 50개의 상이한 gRNA, 8개의 상이한 gRNA 내지 45개의 상이한 gRNA, 8개의 상이한 gRNA 내지 40개의 상이한 gRNA, 8개의 상이한 gRNA 내지 35개의 상이한 gRNA, 8개의 상이한 gRNA 내지 30개의 상이한 gRNA, 8개의 상이한 gRNA 내지 25개의 상이한 gRNA, 8개의 상이한 gRNA 내지 20개의 상이한 gRNA, 8개의 상이한 gRNA 내지 16개의 상이한 gRNA, 16개의 상이한 gRNA 내지 50개의 상이한 gRNA, 16개의 상이한 gRNA 내지 45개의 상이한 gRNA, 16개의 상이한 gRNA 내지 40개의 상이한 gRNA, 16개의 상이한 gRNA 내지 35개의 상이한 gRNA, 16개의 상이한 gRNA 내지 30개의 상이한 gRNA, 16개의 상이한 gRNA 내지 25개의 상이한 gRNA, 또는 16개의 상이한 gRNA 내지 20개의 상이한 gRNA를 인코딩할 수 있다. 상이한 gRNA를 인코딩하는 각각의 폴리뉴클레오티드 서열은 프로모터에 작동 가능하게 연결될 수 있다. 상이한 gRNA에 작동 가능하게 연결된 프로모터는 동일한 프로모터일 수 있다. 상이한 gRNA에 작동 가능하게 연결된 프로모터는 상이한 프로모터일 수 있다. 프로모터는 구성적 프로모터, 유도성 프로모터, 억제성 프로모터, 또는 조절 가능한 프로모터일 수 있다.
일부 실험에서, 가이드 RNA는 녹인, 녹아웃 결실 또는 결함 유전자의 교정에 성공한 공지된 ZFN 서열 표적화된 영역을 표적으로 할 것이다. 공지된 ZFN 표적 영역에 결합하는 다수의 sgRNA 서열이 설계되었고, 미국 특허 공보 2015/0056705 (그 전문이 본원에 참고로 포함됨)의 표 1-2에 기재되어 있으며, 예를 들어 인간 베타-글로빈, 인간, BCLIIA, 인간 KLF1, 인간 CCR5, 인간 CXCR4, PPP1R12C, 마우스 및 인간 HPRT, 인간 알부민, 인간 인자 IX, 인간 인자 VIII, 인간 LRRK2, 인간 Htt, 인간 RH, CFTR, TRAC, TRBC, 인간 PD1, 인간 CTLA-4, HLA c11, HLA A2, HLA A3, HLA B, HLA C, HLA c1. II DBp2. DRA, Tap 1 및 2. 타파신, DMD, RFX5, 등,)에 대한 gRNA 서열을 포함한다.
변형된 뉴클레오시드 또는 뉴클레오티드는 본원에 기재된 가이드 RNA 또는 mRNA에 존재할 수 있다. 가이드 RNA 또는 DNA 엔도뉴클레아제 (예를 들어, RNA-유도 뉴클레아제)를 인코딩하는 mRNA는 하나 이상의 변형된 뉴클레오시드 또는 뉴클레오티드를 포함할 수 있으며; 이러한 mRNA는 정규 A, G, C 및 U 잔기 대신에 또는 이에 추가로 사용되는 하나 이상의 비-천연 및/또는 천연 발생 성분 또는 구성의 존재를 설명하기 위해 "변형된"으로 지칭된다. 일부 구현예에서, 변형된 RNA는 비-정규 뉴클레오시드 또는 뉴클레오티드로 합성되며, 여기서 "변형"으로 불린다. 변형된 뉴클레오시드 및 뉴클레오티드는 다음 중 하나 이상을 포함할 수 있다: (i) 포스포디에스테르 골격 연결에서 비-연결 포스페이트 산소 중 하나 또는 둘 모두 및/또는 연결 포스페이트 산소 중 하나 이상의 변경, 예를 들어, 교체 (예시적인 골격 변형); (ii) 리보스 당의 구성 요소, 예를 들어, 리보스 당 상의 2' 하이드록실의 변경, 예를 들어, 대체 (예시적인 당 변형); (iii) 포스페이트 모이어티를 "데포스포" 링커로의 대규모 대체 (예시적인 골격 변형); (iv) 천연 발생 핵염기의 비정규 핵염기로의 변형 또는 대체 (예시적인 염기 변형); (v) 리보스-포스페이트 골격의 대체 또는 변형 (예시적인 골격 변형); (vi) 올리고뉴클레오티드의 3' 말단 또는 5' 말단의 변형, 예를 들어, 말단 포스페이트 기의 제거, 변형 또는 대체 또는 모이어티, 캡 또는 링커의 컨쥬게이션 (이러한 3' 또는 5' 캡 변형은 당 및/또는 골격 변형을 포함할 수 있음); 및 (vii) 당의 변형 또는 대체 (예시적인 당 변형). 변형되지 않은 핵산은, 예를 들어, 세포 뉴클레아제에 의해 분해되기 쉬울 수 있다. 예를 들어, 뉴클레아제는 핵산 포스포디에스테르 결합을 가수분해할 수 있다. 따라서, 한 측면에서, 본원에 기재된 가이드 RNA는, 예를 들어, 뉴클레아제에 대한 안정성을 도입하기 위해 하나 이상의 변형된 뉴클레오시드 또는 뉴클레오티드를 함유할 수 있다. 특정 구현예에서, 본원에 기재된 mRNA는, 예를 들어, 뉴클레아제에 대한 안정성을 도입하기 위해 하나 이상의 변형된 뉴클레오시드 또는 뉴클레오티드를 함유할 수 있다. 일 구현예에서, 변형은 2'-O-메틸 뉴클레오티드를 포함한다. 다른 구현예에서, 변형은 포스포로티오에이트 (PS) 연결을 포함한다.
변형된 포스페이트 기의 예에는 포스포로티오에이트, 포스포로셀레네이트, 보라노 포스페이트, 보라노 포스페이트 에스테르, 수소 포스포네이트, 포스포로아미데이트, 알킬 또는 아릴 포스포네이트 및 포스포트리에스테르가 포함된다. 비변형된 포스페이트 기의 인 원자는 비키랄이다. 그러나, 비-브리징 산소 중 하나를 상기 원자 또는 원자 그룹 중 하나로 대체하면 인 원자가 키랄이 될 수 있다. 입체 발생(stereogenic) 인 원자는 "R" 배열 (여기서는 Rp) 또는 "S" 배열 (여기서는 Sp)을 가질 수 있다. 골격은 또한 브리징 산소 (즉, 포스페이트를 뉴클레오시드에 연결하는 산소)를 질소 (브릿징된 포스포로아미데이트), 황 (브릿징된 포스포로티오에이트) 및 탄소 (브릿징된 메틸렌포스포네이트)로 대체함으로써 변형될 수 있다. 대체는 연결 산소 중 어느 것 또는 연결 산소 둘 다에서 발생할 수 있다. 포스페이트 기는 특정 골격 변형에서 비-인 함유 커넥터로 대체될 수 있다. 일부 구현예에서, 하전된 포스페이트 기는 중성 모이어티로 대체될 수 있다. 포스페이트 기를 대체할 수 있는 모이어티의 예는, 비제한적으로, 예를 들어, 메틸 포스포네이트, 하이드록실아미노, 실록산, 카보네이트, 카복시 메틸, 카바메이트, 아미드, 티오에테르, 에틸렌 옥사이드 링커, 설포네이트, 설폰아미드, 티오포름아세탈, 포름아세탈, 옥심, 메틸렌이미노, 메틸렌메틸이미노, 메틸렌하이드라조, 메틸렌디메틸하이드라조 및 메틸렌옥시메틸이미노를 포함할 수 있다.
변형된 뉴클레오시드 및 뉴클레오티드는 당 그룹에 대한 하나 이상의 변형, 즉 당 변형을 포함할 수 있다. 예를 들어, 2' 하이드록실 기 (OH)는, 예를 들어, 다수의 상이한 "옥시" 또는 "데옥시" 치환체로 대체될 수 있다. 일부 구현예에서, 2' 하이드록실 기에 대한 변형은 하이드록실이 더 이상 탈양성자화되어 2'-알콕시드 이온을 형성할 수 없기 때문에 핵산의 안정성을 향상시킬 수 있다. 2' 하이드록실 기 변형의 예는 알콕시 또는 아릴옥시 (OR, 여기서 "R"은, 예를 들어, 알킬, 사이클로알킬, 아릴, 아르알킬, 헤테로아릴 또는 당일 수 있음); 폴리 에틸렌 글리콜 (PEG), 0(CH2CH20)nCH2CH2OR (여기서, R은, 예를 들어, H 또는 임의로 치환된 알킬일 수 있고, n은 0 내지 20의 정수 (예를 들어, 0 내지 4, 0 내지 8, 0 내지 10, 0 내지 16, 1 내지 4, 1 내지 8, 1 내지 10, 1 내지 16, 1 내지 20, 2 내지 4, 2 내지 8, 2 내지 10, 2 내지 16, 2 내지 20, 4 내지 8, 4 내지 10, 4 내지 16, 4 내지 20)일 수 있음)을 포함할 수 있다. 일부 구현예에서, 2' 하이드록실 기 변형은 2'-0-Me일 수 있다. 일부 구현예에서, 2' 하이드록실 기 변형은 2' 하이드록실 기를 플루오라이드로 대체하는, 2'-플루오로 변형일 수 있다. 일부 구현예에서, 2' 하이드록실 기 변형은 2' 하이드록실이, 예를 들어, Ci-6 알킬렌 또는 Ci-6 헤테로알킬렌 브릿지에 의해 동일한 리보스 당의 4' 탄소에 연결될 수 있는 "고정된(locked)" 핵산 (LNA)을 포함할 수 있으며, 여기서 예시적인 브릿지는 메틸렌, 프로필렌, 에테르 또는 아미노 브릿지; O-아미노 (여기서, 아미노는, 예를 들어, NH2; 알킬아미노, 디알킬아미노, 헤테로사이클릴, 아릴아미노, 디아릴아미노, 헤테로아릴아미노 또는 디헤테로아릴아미노, 에틸렌디아민 또는 폴리아미노일 수 있음) 및 아미노알콕시, 0(CH2)n-아미노 (여기서 아미노는, 예를 들어, NH2; 알킬아미노, 디알킬아미노, 헤테로사이클릴, 아릴아미노, 디아릴아미노, 헤테로아릴아미노 또는 디헤테로아릴아미노, 에틸렌디아민 또는 폴리아미노일 수 있음)를 포함할 수 있다. 일부 구현예에서, 2' 하이드록실 기 변형은 리보스 고리가 C2'-C3' 결합이 결여된 "고정되지 않은(unlocked)" 핵산 (UNA)을 포함할 수 있다. 일부 구현예에서, 2' 하이드록실 기 변형은 메톡시에틸 기 (MOE), (OCH2CH2OCH3, 예를 들어, PEG 유도체)를 포함할 수 있다.
용어 "데옥시" 2' 변형은 수소 (즉, 예를 들어, 부분 dsRNA의 오버행 부분에서의 데옥시리보스 당); 할로 (예를 들어, 브로모, 클로로, 플루오로 또는 요오도); 아미노 (여기서, 아미노는, 예를 들어, -NH2, 알킬아미노, 디알킬아미노, 헤테로사이클릴, 아릴아미노, 디아릴아미노, 헤테로아릴아미노, 디헤테로아릴아미노 또는 아미노산일 수 있음); NH(CH2CH2NH)nCH2CH2-아미노 (여기서 아미노는, 예를 들어, 본원에 기재된 바와 같을 수 있음), -NHC(O)R (여기서, R은, 예를 들어, 알킬, 사이클로알킬, 아릴, 아르알킬, 헤테로아릴 또는 당일 수 있음), 시아노; 머캅토; 알킬-티오-알킬; 티오알콕시; 및 알킬, 사이클로알킬, 아릴, 알케닐 및 알키닐 (이는 예를 들어, 본원에 기재된 바와 같은 아미노로 임의로 치환될 수 있음)을 포함할 수 있다. 당 변형은 리보스에서 상응하는 탄소의 입체화학 구성과 반대의 입체화학 구성을 갖는 하나 이상의 탄소를 또한 함유할 수 있는 당 기를 포함할 수 있다. 따라서, 변형된 핵산은 당으로서 예를 들어, 아라비노스를 함유하는 뉴클레오티드를 포함할 수 있다. 변형된 핵산은 또한 비염기성 당을 포함할 수 있다. 이들 비염기성 당은 또한 구성 당 원자 중 하나 이상에서 추가로 변형될 수 있다. 변형된 핵산은 또한 L 형태인 하나 이상의 당, 예를 들어 L-뉴클레오시드를 포함할 수 있다.
변형된 핵산에 통합될 수 있는, 본원에 기재된 변형된 뉴클레오시드 및 변형된 뉴클레오티드는 핵염기라고도 불리는 변형된 염기를 포함할 수 있다. 핵염기의 예는 아데닌 (A), 구아닌 (G), 시토신 (C) 및 우라실 (U)을 포함하지만, 이에 제한되지는 않는다. 이들 핵염기는 변형된 핵산 내로 통합될 수 있는 변형된 잔기를 제공하도록 변형되거나 완전히 대체될 수 있다. 뉴클레오티드의 핵염기는 퓨린, 피리미딘, 퓨린 유사체 또는 피리미딘 유사체로부터 독립적으로 선택될 수 있다. 일부 구현예에서, 핵염기는, 예를 들어, 염기의 천연 발생 및 합성 유도체를 포함할 수 있다.
이중 가이드 RNA를 사용하는 구현예에서, crRNA 및 tracr RNA 각각은 변형을 함유할 수 있다. 이러한 변형은 crRNA 및/또는 tracr RNA의 한쪽 또는 양쪽 말단에 있을 수 있다. sgRNA를 포함하는 특정 구현예에서, sgRNA의 한쪽 또는 양쪽 말단에 있는 하나 이상의 잔기가 화학적으로 변형될 수 있거나, 전체 sgRNA가 화학적으로 변형될 수 있다. 특정 구현예는 5' 말단 변형을 포함한다. 특정 구현예는 3' 말단 변형을 포함한다. 특정 구현예에서, 가이드 RNA 분자의 단일 가닥 오버행에서 뉴클레오티드는 중 하나 이상 또는 모두는 데옥시뉴클레오티드이다. 변형된 mRNA는 5' 말단 및/또는 3' 말단 변형을 함유할 수 있다.
D. 조절 요소.
본원에 정의된 비대칭 ITR 쌍 또는 대칭 ITR 쌍을 포함하는 유전자 편집용 ceDNA 벡터는 시스-조절 요소의 특정 조합을 추가로 포함하는 발현 작제물로부터 생성될 수 있다. 시스-조절 요소는 프로모터, 리보스위치, 절연체, 미르-조절 가능 요소, 전사 후 조절 요소, 조직- 및 세포 유형-특이적 프로모터 및 인핸서를 포함하지만, 이에 제한되지는 않는다. 일부 구현예에서, ITR은 이식유전자의 프로모터로서 작용할 수 있다. 일부 구현예에서, ceDNA 벡터는 이식유전자의 발현을 조절하기 위한 추가 성분, 예를 들어, 이식유전자의 발현을 조절하기 위한 본원에 기재된 조절 스위치, 또는 ceDNA 벡터를 포함하는 세포를 사멸시킬 수 있는 사멸 스위치를 포함한다. 본 발명에서 사용될 수 있는 조절 스위치를 포함하는 조절 요소는 PCT/US18/49996에 보다 상세하게 논의되어 있으며, 이의 전문이 본원에 참고로 포함된다.
구현예에서, 제2 뉴클레오티드 서열은 조절 서열, 및 뉴클레아제를 인코딩하는 뉴클레오티드 서열을 포함한다. 특정 구현예에서, 유전자 조절 서열은 뉴클레아제를 인코딩하는 뉴클레오티드 서열에 작동 가능하게 연결된다. 특정 구현예에서, 조절 서열은 숙주 세포에서 뉴클레아제의 발현을 제어하기에 적합하다. 특정 구현예에서, 조절 서열은 프로모터 서열에 작동 가능하게 연결된 유전자, 예컨대 본 개시내용의 뉴클레아제(들)를 인코딩하는 뉴클레오티드 서열의 전사를 지시할 수 있는, 적합한 프로모터 서열을 포함한다. 특정 구현예에서, 제2 뉴클레오티드 서열은 뉴클레아제를 인코딩하는 뉴클레오티드 서열의 5' 말단에 연결된 인트론 서열을 포함한다. 특정 구현예에서, 프로모터의 효능을 증가시키기 위해 인핸서 서열이 프로모터의 상류에 제공된다. 특정 구현예에서, 조절 서열은 인핸서 및 프로모터를 포함하고, 여기서 제2 뉴클레오티드 서열은 뉴클레아제를 인코딩하는 뉴클레오티드 서열의 상류에 있는 인트론 서열을 포함하고, 상기 인트론은 하나 이상의 뉴클레아제 절단 부위(들)를 포함하고, 상기 프로모터는 뉴클레아제를 인코딩하는 뉴클레오티드 서열에 작동 가능하게 연결된다.
ceDNA 벡터는 시스-조절 요소, 예컨대 WHP 전사 후 조절 요소 (WPRE) (예를 들어, 서열 번호: 8) 및 BGH 폴리A (서열 번호: 9)의 특정 조합을 추가로 포함하는 발현 작제물로부터 생성될 수 있다. 발현 작제물에 사용하기에 적합한 발현 카세트는 바이러스 캡시드에 의해 부과된 패키징 제약에 의해 제한되지 않는다.
(i). 프로모터:
본 발명의 유전자-편집 ceDNA 벡터에 사용된 프로모터는 이들이 촉진하는 특정 서열에 적합하게 맞추어져야 한다는 것이 당업자에게 이해될 것이다. 예를 들어, 가이드 RNA는 프로모터가 전혀 필요하지 않을 수 있는데, 그 이유는 이의 기능이 재조합 사건을 수행하기 위해 천연 DNA 상에 특정 표적 서열을 갖는 듀플렉스를 형성하는 것이기 때문이다. 대조적으로, ceDNA 벡터에 의해 인코딩된 뉴클레아제는 프로모터로부터 이익을 얻을 수 있으므로 이는 벡터로부터 효율적으로 그리고, 선택적으로, 조절 가능한 방식으로 발현될 수 있다.
본 발명의 발현 카세트는 세포-특이성뿐만 아니라 전체 발현 수준에 영향을 줄 수 있는 프로모터를 포함한다. 이식유전자 발현을 위해, 이들은 매우 활성인 바이러스-유래 즉각적인 조기 프로모터를 포함할 수 있다. 발현 카세트는 조직 특이적 진핵생물 프로모터를 함유하여 이식유전자 발현을 특정 세포 유형으로 제한하고 조절되지 않은 이소성 발현으로 인한 독성 효과 및 면역 반응을 감소시킬 수 있다. 바람직한 구현예에서, 발현 카세트는 합성 조절 요소, 예컨대 CAG 프로모터 (서열 번호: 3)를 함유할 수 있다. CAG 프로모터는 (i) 사이토메갈로바이러스 (CMV) 조기 인핸서 요소, (ii) 치킨 베타-액틴 유전자의 프로모터, 제1 엑손 및 제1 인트론, 및 (iii) 토끼 베타-글로빈 유전자의 스플라이스 수용체를 포함한다. 대안적으로, 발현 카세트는 알파-1-항트립신 (AAT) 프로모터 (서열 번호: 4 또는 서열 번호: 74), 간 특이적 (LP1) 프로모터 (서열 번호: 5 또는 서열 번호: 16), 또는 인간 신장 인자-1 알파 (EF1a) 프로모터 (예를 들어, 서열 번호: 6 또는 서열 번호: 15)를 함유할 수 있다. 일부 구현예에서, 발현 카세트는 하나 이상의 구성적 프로모터, 예를 들어, 레트로바이러스 라우스 육종 바이러스 (RSV) LTR 프로모터 (선택적으로 RSV 인핸서를 가짐) 또는 사이토메갈로바이러스 (CMV) 즉각적인 조기 프로모터 (선택적으로 CMV 인핸서, 예를 들어, 서열 번호: 22를 가짐)를 포함한다. 대안적으로, 유도성 프로모터, 이식유전자를 위한 천연 프로모터, 조직-특이적 프로모터, 또는 당 업계에 공지된 다양한 프로모터가 사용될 수 있다.
상기 기재된 것들을 포함하여, 적합한 프로모터는 바이러스로부터 유래될 수 있으며 따라서 바이러스 프로모터로 지칭될 수 있거나, 원핵생물 또는 진핵생물 유기체를 포함하는 임의의 유기체로부터 유래될 수 있다. 적합한 프로모터를 사용하여 임의의 RNA 폴리머라제 (예를 들어, pol I, pol II, pol III)에 의한 발현을 구동시킬 수 있다. 예시적인 프로모터는 SV40 조기 프로모터, 마우스 유방 종양 바이러스 긴 말단 반복 (LTR) 프로모터; 아데노바이러스 주요 후기 프로모터 (Ad MLP); 단순 포진 바이러스 (HSV) 프로모터, CMV 즉각적인 조기 프로모터 영역 (CMVIE)과 같은 사이토메갈로바이러스 (CMV) 프로모터, 라우스 육종 바이러스 (RSV) 프로모터, 인간 U6 소형 핵 프로모터 (U6, 예를 들어, 서열 번호: 18) (Miyagishi et al., Nature Biotechnology 20, 497-500 (2002)), 향상된 U6 프로모터 (예를 들어, Xia et al., Nucleic Acids Res. 2003 Sep. 1; 31(17)), 인간 H1 프로모터 (H1) (예를 들어, 서열 번호: 19), CAG 프로모터, 인간 알파 1-안티팁신(antitypsin) (HAAT) 프로모터 (예를 들어, 서열 번호: 21) 등을 포함하지만, 이에 제한되지 않는다. 특정 구현예에서, 이들 프로모터는 하나 이상의 뉴클레아제 절단 부위를 포함하도록 하류 인트론 함유 말단에서 변경된다. 특정 구현예에서, 뉴클레아제 절단 부위(들)를 함유하는 DNA는 프로모터 DNA에 이질적이다.
일 구현예에서, 사용된 프로모터는 치료 단백질을 인코딩하는 유전자의 천연 프로모터이다. 치료 단백질을 인코딩하는 각각의 유전자에 대한 프로모터 및 다른 조절 서열은 공지되어 있고 특성화되어 있다. 사용된 프로모터 영역은 하나 이상의 추가 조절 서열 (예를 들어, 천연), 예를 들어 인핸서 (예를 들어, 서열 번호: 22 및 서열 번호: 23)를 추가로 포함할 수 있다.
본 발명에 따라 사용하기에 적합한 프로모터의 비제한적인 예는, 예를 들어, (서열 번호: 3)의 CAG 프로모터, HAAT 프로모터 (서열 번호: 21), 인간 EF1-α 프로모터 (서열 번호: 6) 또는 EF1a 프로모터의 단편 (서열 번호: 15), IE2 프로모터 (예를 들어, 서열 번호: 20) 및 랫트 EF1-α 프로모터 (서열 번호: 24)를 포함한다.
(ii). 폴리아데닐화 서열:
ceDNA 벡터로부터 발현된 mRNA를 안정화시키고 핵 수출 및 번역을 돕기 위해 폴리아데닐화 서열을 인코딩하는 서열이 ceDNA 벡터에 포함될 수 있다. 일 구현예에서, ceDNA 벡터는 폴리아데닐화 서열을 포함하지 않는다. 다른 구현예에서, 벡터는 적어도 1, 적어도 2, 적어도 3, 적어도 4, 적어도 5, 적어도 10, 적어도 15, 적어도 20, 적어도 25, 적어도 30, 적어도 40, 적어도 45, 적어도 50개 또는 그 이상의 아데닌 디뉴클레오티드를 포함한다. 일부 구현예에서, 폴리아데닐화 서열은 약 43개 뉴클레오티드, 약 40-50개 뉴클레오티드, 약 40-55개 뉴클레오티드, 약 45-50개 뉴클레오티드, 약 35-50개 뉴클레오티드, 또는 이들 사이의 임의의 범위를 포함한다.
발현 카세트는 당 업계에 공지된 폴리아데닐화 서열 또는 이의 변형, 예컨대 소 BGHpA (예를 들어, 서열 번호: 74) 또는 바이러스 SV40pA (예를 들어, 서열 번호: 10), 또는 합성 서열 (예를 들어, 서열 번호: 27)로부터 단리된 천연 발생 서열을 포함할 수 있다. 일부 발현 카세트는 또한 SV40 후기 폴리 A 신호 상류 인핸서 (USE) 서열을 포함할 수 있다. 일부 구현예에서, USE는 SV40pA 또는 이종 폴리-A 신호와 조합하여 사용될 수 있다.
발현 카세트는 또한 이식유전자의 발현을 증가시키기 위해 전사 후 요소를 포함할 수 있다. 일부 구현예에서, 우드척 간염 바이러스 (WHP) 전사 후 조절 요소 (WPRE) (예를 들어, 서열 번호: 8)는 이식유전자의 발현을 증가시키는 데 사용된다. 단순 포진 바이러스 또는 B형 간염 바이러스 (HBV)의 티미딘 키나제 유전자로부터의 전사 후 요소와 같은 다른 전사 후 처리 요소가 사용될 수 있다. 분비 서열은 이식유전자, 예를 들어, VH-02 및 VK-A26 서열, 예를 들어, 서열 번호: 25 및 서열 번호: 26에 연결될 수 있다.
(iii). 핵 국재화 서열
일부 구현예에서, RNA 유도 엔도뉴클레아제를 인코딩하는 벡터는 하나 이상의 핵 국재화 서열 (NLS), 예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10개 또는 그 이상의 NLS를 포함한다. 일부 구현예에서, 하나 이상의 NLS는 아미노-말단에 또는 그 근처, 카복시-말단에 또는 그 근처에 위치하거나, 또는 이들의 조합 (예를 들어, 아미노-말단에서 하나 이상의 NLS 및/또는 카복시 말단에서 하나 이상의 NLS)에 위치한다. 하나 초과의 NLS가 존재하는 경우, 단일 NLS가 하나 초과의 카피에 존재하고/하거나 하나 이상의 다른 NLS와 조합하여 하나 이상의 카피에 존재하도록 다른 것과 독립적으로 선택될 수 있다. NLS의 비제한적인 예는 표 6에 제시되어 있다.
[표 6]
E.
유전자 편집 시스템의 추가 성분
본 개시내용의 ceDNA 벡터는 유전자 편집을 위해 다른 성분을 인코딩하는 뉴클레오티드를 함유할 수 있다. 예를 들어, 특정 유전자 표적화 사건을 선택하기 위해, 보호 shRNA가 마이크로RNA에 포매되어 알부민 유전자좌와 같은 고도로 활성인 유전자좌에 부위 특이적으로 통합되도록 설계된 재조합 ceDNA 벡터에 삽입될 수 있다. 이러일 구현예는 하기에 기재된 바와 같은 임의의 유전자 배경에서 유전자 변형 간세포의 생체내 선택 및 확장을 위한 시스템을 제공할 수 있다: Nygaard et al., A universal system to select gene-modified hepatocytes in vivo, Gene Therapy, June 8, 2016. 본 개시내용의 ceDNA 벡터는 형질전환, 형질감염, 형질도입 또는 이와 유사한 세포의 선택을 허용하는 하나 이상의 선택 가능한 마커를 함유할 수 있다. 선택 가능한 마커는 그 산물이 살생물제 또는 바이러스 내성, 중금속에 대한 내성, 영양요구성에 대한 프로토트로피(prototrophy), NeoR 등을 제공하는 유전자이다. 특정 구현예에서, 양성 선별 마커는 NeoR과 같은 공여체 서열에 포함된다. 음성 선별 마커는 공여체 서열의 하류에 통합될 수 있으며, 예를 들어 음성 선별 마커를 인코딩하는 핵산 서열 HSV-tk는 공여체 서열의 하류에 있는 핵산 작제물에 통합될 수 있다. 도 8을 참조하면, 이식유전자는 0.05 내지 6kb 신장 상동성 아암이 측면에 있는 바이러스 2A 펩티드 절단 부위 (2A)를 통해 선별 마커 (NeoR)에 임의로 융합된다. 특정 구현예에서, 성공적인 올바른 부위 사용을 위해 제어하고 선택될 수 있는 HSV TK)와 같은 음성 선별 마커 및 발현 유닛은 선택적으로 상동성 아암 외부에 위치될 수 있다.
구현예에서, 본 개시내용의 ceDNA 벡터는 5' 상동성 아암의 상류에 있고 이에 근접한 폴리아데닐화 부위를 포함할 수 있다.
도 9를 참조하면, ceDNA 특이적 ITR을 포함하는 본 개시내용에 따른 ceDNA 벡터가 도시되어 있다. ceDNA 벡터는 전사 방향에 대해 임의의 배향을 갖는 Pol III 프로모터 구동 (예컨대 U6 및 H1) sgRNA 발현 유닛을 포함한다. "이중 돌연변이 닉카아제"에 대한 sgRNA 표적 서열은 돌연변이 ITR에 근접한 3' 상동성 아암의 하류에 있는 비틀림을 해제하도록 임의로 제공된다. 이러한 구현예는 어닐링을 증가시키고 HDR 빈도를 촉진시킨다.
일부 구현예에서, 본원에 기재된 ceDNA 벡터에 의해 포함된 뉴클레아제는 유전자 편집 후에 불활성화/감소될 수 있다. 예를 들어, 본원의 실시예 6 (또한 도 8, 9 및 13 참조)을 참조한다.
F. 조절 스위치
분자 조절 스위치는 신호에 응답하여 측정 가능한 상태 변화를 생성하는 스위치이다. 이러한 조절 스위치는 ceDNA 벡터의 출력을 제어하기 위해 본원에 기재된 ceDNA 벡터와 유용하게 결합될 수 있다. 일부 구현예에서, ceDNA 벡터는 이식유전자의 발현을 미세 조정하는 역할을 하는 조절 스위치를 포함한다. 예를 들어, 이는 ceDNA 벡터의 생화학적 봉쇄(biocontainment) 기능으로 작용할 수 있다. 일부 구현예에서, 스위치는 제어 가능하고 조절 가능한 방식으로 ceDNA에서 관심 유전자의 발현을 시작 또는 중지 (즉, 셧다운)하도록 설계된 "ON/OFF" 스위치이다. 일부 구현예에서, 스위치는 스위치가 활성화되면 ceDNA 벡터를 포함하는 세포가 세포 프로그래밍된 사멸을 겪도록 지시할 수 있는 "사멸 스위치"를 포함할 수 있다. 유전자 편집 분자 (예를 들어, 엔도뉴클레아제를 인코딩하는 이식유전자, 가이드 RNA, gDNA, RNA 활성화제, 또는 공여체 서열의 발현을 조절하기 위해 유전자 편집 ceDNA에 사용하기 위해 포함된 예시적인 조절 스위치는 PCT/US18/49996 (이는 그 전문이 본원에 참고로 포함됨)에서 보다 상세히 논의된다.
(i) 이진 조절 스위치
일부 구현예에서, ceDNA 벡터는 이식유전자의 발현을 제어 가능하게 조절하는 역할을 할 수 있는 조절 스위치를 포함한다. 예를 들어, ceDNA 벡터의 ITR 사이에 위치한 발현 카세트는 관심 유전자에 작동적으로 연결된, 조절 영역, 예를 들어, 프로모터, 시스-요소, 리프레서, 인핸서 등을 추가로 포함할 수 있으며, 여기서 조절 영역은 하나 이상의 보조 인자 또는 외인성 제제에 의해 조절된다. 단지 예로서, 조절 영역은 소분자 스위치 또는 유도성 또는 억제성 프로모터에 의해 조절될 수 있다. 유도성 프로모터의 비제한적인 예는 호르몬-유도성 또는 금속-유도성 프로모터이다. 다른 예시적인 유도성 프로모터/인핸서 요소는 RU486-유도성 프로모터, 엑디손-유도성 프로모터, 라파마이신-유도성 프로모터 및 메탈로티오네인 프로모터를 포함하지만, 이에 제한되지는 않는다.
(ii) 소분자 조절 스위치
다양한 당 업계에 공지된 소분자 기반 조절 스위치가 당 업계에 공지되어 있고, 조절 스위치 제어 ceDNA 벡터를 형성하기 위해 본원에 개시된 ceDNA 벡터와 조합될 수 있다. 일부 구현예에서, 조절 스위치는 문헌(Taylor, et al. BMC Biotechnology 10 (2010): 15)에 기재된 것과 같이, 작동적으로 연결된 이식유전자의 발현을 제어하는 인공 프로모터와 함께, 직교 리간드/핵 수용체 쌍, 예를 들어 레티노이드 수용체 변이체/LG335 및 GRQCIMFI; 조작된 스테로이드 수용체, 예를 들어, 프로게스테론에 결합할 수 없지만 RU486 (미페프리스톤)에 결합하는 C-말단 절단을 갖는 변형된 프로게스테론 수용체 (미국 특허 번호 5,364,791); 드로소필라(Drosophila) 유래 엑디손 수용체 및 이의 엑디스테로이드 리간드 (Saez, et al., PNAS, 97(26)(2000), 14512-14517; 또는 문헌(Sando R 3rd; Nat Methods. 2013, 10(11):1085-8)에 개시된 바와 같은 항생제 트리메토프림 (TMP)에 의해 제어되는 스위치 중 임의의 하나 또는 조합으로부터 선택될 수 있다. 일부 구현예에서, 이식유전자를 제어하거나 ceDNA 벡터에 의해 발현되는 조절 스위치는 미국 특허 8,771,679 및 6,339,070에 개시된 것과 같은 전구 약물 활성화 스위치이다.
(iii) "패스코드(Passcode)" 조절 스위치
일부 구현예에서, 조절 스위치는 "패스코드 스위치" 또는 "패스코드 회로"일 수 있다. 패스코드 스위치는 특정 조건이 발생할 때 ceDNA 벡터에서 이식유전자의 발현 제어를 미세 조정할 수 있도록 하며, 즉, 이식유전자 발현 및/또는 억제가 발생하려면 조건의 조합이 필요한다. 예를 들어, 이식유전자의 발현이 발생하기 위해서는 적어도 조건 A 및 B가 발생해야 한다. 패스코드 조절 스위치는 이식유전자 발현이 발생하기 위해 임의의 수의 조건, 예를 들어, 적어도 2, 적어도 3, 적어도 4, 적어도 5, 적어도 6 또는 적어도 7개 또는 그 이상의 조건이 존재할 수 있다. 일부 구현예에서, 적어도 2개의 조건 (예를 들어, A, B 조건)이 발생해야 하고, 일부 구현예에서, 적어도 3개의 조건 (예를 들어, A, B 및 C, 또는 A, B 및 D)이 발생해야 한다. 단지 예로서, 패스코드 "ABC" 조절 스위치를 갖는 ceDNA로부터의 유전자 발현이 발생하기 위해서는 조건 A, B 및 C가 존재해야 한다. 조건 A, B 및 C는 다음과 같을 수 있다; 조건 A는 병태 또는 질환의 존재이고, 조건 B는 호르몬 반응이고, 조건 C는 이식유전자 발현에 대한 반응이다. 예를 들어, 이식유전자이 결함 있는 EPO 유전자를 편집하는 경우, 조건 A는 만성 신장 질환 (CKD)의 존재이고, 조건 B는 대상체가 신장에 저산소 상태에 있는 경우 발생하고, 조건 C는 신장에서 에리스로포이에틴 생성 세포 (EPC) 동원이 손상된 것이거나; 또는 대안적으로, HIF-2 활성화가 손상된 것이다. 산소 수준이 증가하거나 원하는 수준의 EPO에 도달하면, 3가지 조건이 발생할 때까지 이식유전자이 다시 꺼지고, 다시 켜진다.
일부 구현예에서, ceDNA 벡터에 사용하기 위해 포함된 패스코드 조절 스위치 또는 "패스코드 회로"는 생화학적 봉쇄 조건을 정의하는데 사용되는 환경 신호의 범위 및 복잡성을 확장시키기 위해 하이브리드 전사 인자 (TF)를 포함한다. 미리 결정된 조건의 존재하에 세포 사멸을 유발하는 데드맨 스위치(deadman switch)와는 달리, "패스코드 회로"는 특정 "패스코드"의 존재하에서 세포 생존 또는 이식유전자 발현을 허용하고, 미리 결정된 환경 조건 또는 패스코드가 존재하는 경우에만 이식유전자 발현 및/또는 세포 생존을 허용하도록 쉽게 재프로그래밍될 수 있다.
본원에 개시된 조절 스위치의 임의의 및 모든 조합, 예를 들어, 소분자 스위치, 핵산-기반 스위치, 소분자-핵산 하이브리드 스위치, 전사 후 이식유전자 조절 스위치, 번역 후 조절, 방사선-제어 스위치, 저산소증-매개 스위치 및 본원에 개시된 바와 같은 당업자에게 공지된 다른 조절 스위치는 본원에 개시된 바와 같은 패스코드 조절 스위치에 사용될 수 있다. 사용을 위해 포함된 조절 스위치는 리뷰 논문 (Kis et al., J R Soc Interface. 12: 20141000 (2015))에서 또한 논의되며, Kis의 표 1에 요약되어 있다. 일부 구현예에서, 패스코드 시스템에 사용하기 위한 조절 스위치는 표 11의 스위치 중 임의의 것 또는 조합으로부터 선택될 수 있다.
(iv). 이식유전자 발현을 제어하기 위한 핵산 기반 조절 스위치
일부 구현예에서, ceDNA에 의해 발현된 이식유전자를 제어하기 위한 조절 스위치는 핵산 기반 제어 메커니즘에 기초한다. 예시적인 핵산 제어 메카니즘이 당 업계에 공지되어 있으며 사용이 구상된다. 예를 들어, 이러한 메커니즘은, 예를 들어, US2009/0305253, US2008/0269258, US2017/0204477, WO2018026762A1, 미국 특허 9,222,093 및 EP 출원 EP288071에 개시되고, 또한 문헌(Villa JK et al., Microbiol Spectr. 2018 May;6(3))에 의한 검토에 개시된 것과 같은 리보스위치를 포함한다. 또한, WO2018/075486 및 WO2017/147585에 개시된 것과 같은 대사 물질-반응성 전사 바이오센서가 포함된다. 사용이 구상된 당 업계에 공지된 다른 메커니즘은 siRNA 또는 RNAi 분자 (예를 들어, miR, shRNA)로 이식유전자의 침묵화를 포함한다. 예를 들어, ceDNA 벡터는 ceDNA 벡터에 의해 발현된 이식유전자에 상보적인 RNAi 분자를 인코딩하는 조절 스위치를 포함할 수 있다. 이식유전자이 ceDNA 벡터에 의해 발현되더라도 이러한 RNAi가 발현될 때, 상보적 RNAi 분자에 의해 침묵화될 것이고, 이식유전자이 ceDNA 벡터에 의해 발현될 때 RNAi가 발현되지 않을 때 이식유전자는 RNAi에 의해 침묵화되지 않는다.
일부 구현예에서, 조절 스위치는, 예를 들어 US2002/0022018에 개시된 바와 같은 조직-특이적 자가-활성화 조절 스위치이며, 따라서 조절 스위치는 이식유전자 발현이 그렇지 않으면 불리할 수 있는 부위에서 이식유전자 발현을 의도적으로 차단한다. 일부 구현예에서, 조절 스위치는, 예를 들어 US2014/0127162 및 미국 특허 8,324,436에 개시된 바와 같은 재조합 효소 가역 유전자 발현 시스템이다.
(V). 전사 후 및 번역 후 조절 스위치.
일부 구현예에서, ceDNA 벡터에 의해 발현된 관심 이식유전자 또는 유전자를 제어하기 위한 조절 스위치는 전사 후 변형 시스템이다. 예를 들어, 이러한 조절 스위치는 US2018/0119156, GB201107768, WO2001/064956A3, EP 특허 2707487 및 문헌(Beilstein et al., ACS Synth. Biol., 2015, 4 (5), pp 526-534; Zhong et al., Elife. 2016 Nov 2;5. pii: e18858)에 개시된 바와 같이 테트라사이클린 또는 테오필린에 민감한 압타자임(aptazyme) 리보스위치일 수 있다. 일부 구현예에서, 당업자는 리간드 감수성 (OFF-스위치) 압타머를 함유하는 억제성 siRNA 및 이식유전자 둘 다를 인코딩할 수 있으며, 최종 결과는 리간드 감수성 ON-스위치인 것으로 예상된다.
(vi). 다른 예시적인 조절 스위치
환경 변화에 의해 유발된 것들을 포함하여, ceDNA 벡터에 의해 발현된 이식유전자의 유전자 발현을 제어하기 위해 임의의 공지된 조절 스위치가 ceDNA 벡터에서 사용될 수 있다. 추가의 예는 다음을 포함하지만, 이에 제한되지 않는다; 문헌(Suzuki et al., Scientific Reports 8; 10051 (2018))의 BOC 방법; 유전자 코드 확장 및 비-생리적 아미노산; 방사선 제어 또는 초음파 제어 온/오프 스위치 (예를 들어, Scott S et al., Gene Ther. 2000 Jul;7(13):1121-5; 미국 특허 5,612,318; 5,571,797; 5,770,581; 5,817,636; 및 WO1999/025385A1 참조. 일부 구현예에서, 조절 스위치는, 예를 들어, 미국 특허 7,840,263; US2007/0190028A1 (여기서 유전자 발현은 ceDNA 벡터의 이식유전자에 작동적으로 연결된 프로모터를 활성화시키는 전자기 에너지를 포함하는 하나 이상의 형태의 에너지에 의해 제어됨)에 개시된 바와 같은 이식 가능한 시스템에 의해 제어된다.
일부 구현예에서, ceDNA 벡터에 사용하기 위해 구상된 조절 스위치는 저산소증-매개 또는 스트레스-활성화 스위치, 예를 들어, WO1999060142A2, 미국 특허 5,834,306; 6,218,179; 6,709,858; US2015/0322410; 문헌(Greco et al., (2004) Targeted Cancer Therapies 9, S368)에 기재된 것, 뿐만 아니라, 예를 들어, 미국 특허 9,394,526에 개시된 바와 같은, FROG, TOAD 및 NRSE 요소 및 저산소증 반응 요소 (HRE), 염증 반응 요소 (IRE) 및 전단 응력 활성화 요소 (SSAE)를 포함한 조건부로 유도 가능한 침묵 요소이다. 이러한 구현예는 허혈 후 또는 허혈성 조직 및/또는 종양에서 ceDNA 벡터로부터 이식유전자의 발현을 켜는데 유용하다.
(iv). 사멸 스위치
본 발명의 다른 구현예는 사멸 스위치를 포함하는 ceDNA 벡터에 관한 것이다. 본원에 개시된 사멸 스위치는 ceDNA 벡터를 포함하는 세포가 대상체 시스템으로부터 도입된 ceDNA 벡터를 영구적으로 제거하기 위한 수단으로 사멸되거나 프로그래밍된 세포 사멸을 겪도록 할 수 있다. 당업자는 본 발명의 ceDNA 벡터에서 사멸 스위치의 사용이 전형적으로 대상체가 수용 가능하게 상실할 수 있는 제한된 수의 세포 또는 아폽토시스가 바람직한 세포 유형 (예를 들어, 암 세포)에 대한 ceDNA 벡터의 표적화와 결합될 수 있음을 이해할 것이다. 모든 측면에서, 본원에 개시된 바와 같은 "사멸 스위치"는 입력 생존 신호 또는 다른 특정 조건이 없는 경우 ceDNA 벡터를 포함하는 세포의 빠르고 강력한 세포 사멸을 제공하도록 설계된다. 달리 말하면, 본 명세서에서 ceDNA 벡터에 의해 인코딩된 사멸 스위치는 ceDNA 벡터를 포함하는 세포의 세포 생존을 특정 입력 신호에 의해 정의된 환경으로 제한할 수 있다. 이러한 사멸 스위치는 대상체로부터 ceDNA 벡터를 제거하거나 그것이 인코딩된 이식유전자를 발현하지 않도록 보장하는 것이 바람직한 경우 생물학적 봉쇄 기능을 한다.
VII. ceDNA 벡터의 상세한 생산 방법
A. 일반적인 생산
본원에 정의된 비대칭 ITR 쌍 또는 대칭 ITR 쌍을 포함하는 유전자 편집용 ceDNA 벡터의 생산을 위한 특정 방법은 2018년 9월 7일에 출원된 PCT/US18/49996의 섹션 IV에 기재되어 있으며, 이는 그 전문이 본원에 참고로 포함된다. 본원에 기재된 바와 같이, ceDNA 벡터는, 예를 들어, a) 숙주 세포 내에서 ceDNA 벡터의 생성을 유도하기에 효과적인 조건 하에서 및 충분한 시간 동안 Rep 단백질의 존재하에, 바이러스 캡시드 코딩 서열이 없는, 폴리뉴클레오티드 발현 작제물 주형 (예를 들어, ceDNA-플라스미드, ceDNA-박미드 및/또는 ceDNA-바쿨로바이러스)을 보유하는 숙주 세포 (예를 들어, 곤충 세포) 집단을 인큐베이션하는 단계로서, 상기 숙주 세포는 바이러스 캡시드 코딩 서열을 포함하지 않는, 상기 단계; 및 b) 숙주 세포로부터 ceDNA 벡터를 수거하고 분리하는 단계를 포함하는 과정에 의해 얻어질 수 있다. Rep 단백질의 존재는 변형된 ITR로 벡터 폴리뉴클레오티드의 복제를 유도하여 숙주 세포에서 ceDNA 벡터를 생성한다. 그러나, 바이러스 입자 (예를 들어 AAV 비리온)는 발현되지 않는다. 따라서, AAV 또는 다른 바이러스 기반 벡터에 자연적으로 부과되는 것과 같은 크기 제한은 없다.
숙주 세포로부터 단리된 ceDNA 벡터의 존재는 ceDNA 벡터 상에 단일 인식 부위를 갖는 제한 효소로 숙주 세포로부터 단리된 DNA를 소화하고 비-변성 겔 상에서 소화된 DNA 물질을 분석하여 선형 및 비연속 DNA와 비교하여 선형 및 연속 DNA의 특징적인 밴드의 존재를 확인함으로써 확인될 수 있다.
또 다른 측면에서, 본 발명은, 예를 들어, 하기에 기재된 바와 같이, 비-바이러스성 DNA 벡터의 생산에서 DNA 벡터 폴리뉴클레오티드 발현 주형 (ceDNA 주형)을 그들 자신의 게놈에 안정적으로 통합시킨 숙주 세포주의 사용을 제공한다: Lee, L. et al. (2013) Plos One 8(8): e69879. 바람직하게는, Rep는 약 3의 MOI에서 숙주 세포에 첨가된다. 숙주 세포주가 포유동물 세포주, 예를 들어, HEK293 세포인 경우, 세포주는 안정적으로 통합된 폴리뉴클레오티드 벡터 주형을 가질 수 있고, 헤르페스 바이러스와 같은 제2 벡터를 사용하여 Rep 단백질을 세포 내로 도입할 수 있으며, 이는 Rep 및 헬퍼 바이러스의 존재하에 ceDNA의 절제 및 증폭을 허용한다.
일 구현예에서, 본원에 기재된 ceDNA 벡터를 제조하는데 사용된 숙주 세포는 곤충 세포이고, 바쿨로바이러스는 Rep 단백질을 인코딩하는 폴리뉴클레오티드 및, 예를 들어, 도 4a-4c 및 실시예 1에 기재된 바와 같이, ceDNA에 대한 비-바이러스성 DNA 벡터 폴리뉴클레오티드 발현 작제물 주형 둘 다를 전달하는데 사용된다. 일부 구현예에서, 숙주 세포는 Rep 단백질을 발현하도록 조작된다.
이어서 ceDNA 벡터를 수거하고 숙주 세포로부터 단리한다. 세포로부터 본원에 기재된 ceDNA 벡터를 수거하고 수집하는 시간은 ceDNA 벡터의 고-수율 생산을 달성하도록 선택되고 최적화될 수 있다. 예를 들어, 수거 시간은 세포 생존력, 세포 형태, 세포 성장 등을 고려하여 선택될 수 있다. 일 구현예에서, 세포는 충분한 조건 하에서 성장하고, ceDNA 벡터를 생성하기 위해 바쿨로바이러스 감염 후 그러나 바쿨로바이러스 독성으로 인해 대부분의 세포가 죽기 시작하기 전에 충분한 시간 수거한다. Qiagen Endo-Free Plasmid 키트와 같은 플라스미드 정제 키트를 사용하여 DNA 벡터를 분리할 수 있다. 플라스미드 분리를 위해 개발된 다른 방법도 DNA 벡터에 적용될 수 있다. 일반적으로, 임의의 핵산 정제 방법이 채택될 수 있다.
DNA 벡터는 DNA의 정제를 위해 당업자에게 공지된 임의의 수단에 의해 정제될 수 있다. 일 구현예에서, ceDNA 벡터는 DNA 분자로서 정제된다. 또 다른 구현예에서, ceDNA 벡터는 엑소좀 또는 미세입자로서 정제된다.
ceDNA 벡터의 존재는 DNA 벡터 상에 단일 인식 부위를 갖는 제한 효소로 세포로부터 단리된 벡터 DNA를 소화시키고, 겔 전기영동을 사용하여 소화된 및 소화되지 않은 DNA 물질 모두를 분석하여, 선형 및 비-연속 DNA와 비교하여 선형 및 연속 DNA의 특징적인 밴드의 존재를 확인함으로써 확인할 수 있다. 도 4c 및 도 4d는 본원의 공정에 의해 생성된 폐쇄 말단 ceDNA 벡터의 존재를 확인하기 위한 일 구현예를 도시한다.
B. ceDNA 플라스미드
ceDNA-플라스미드는 ceDNA 벡터의 추후 생산에 사용되는 플라스미드이다. 일부 구현예에서, ceDNA-플라스미드는 적어도 다음을 전사 방향으로 작동적으로 연결된 성분으로서 제공하기 위해 공지된 기술을 사용하여 구축될 수 있다: (1) 변형된 5' ITR 서열; (2) 시스-조절 요소, 예를 들어, 프로모터, 유도성 프로모터, 조절 스위치, 인핸서 등을 함유하는 발현 카세트; 및 (3) 3' ITR 서열이 5' ITR 서열에 대해 대칭인 변형된 3' ITR 서열. 일부 구현예에서, ITR이 측면에 있는 발현 카세트는 외인성 서열을 도입하기 위한 클로닝 부위를 포함한다. 발현 카세트는 AAV 게놈의 rep 및 cap 코딩 영역을 대체한다.
일 측면에서, ceDNA 벡터는 본원에서, 제1 아데노-관련 바이러스 (AAV) 역 말단 반복부 (ITR), 이식유전자를 포함하는 발현 카세트, 및 돌연변이 또는 변형된 AAV ITR 순서로 인코딩하는 "ceDNA-플라스미드"로 지칭되는 플라스미드로부터 수득되며, 여기서 상기 ceDNA-플라스미드는 AAV 캡시드 단백질 코딩 서열이 없다. 대안적인 구현예에서, ceDNA-플라스미드는 제1 (또는 5') 변형 또는 돌연변이된 AAV ITR, 이식유전자를 포함하는 발현 카세트, 및 제2 (또는 3') 변형된 AAV ITR 순서로 인코딩하며, 여기서 상기 ceDNA-플라스미드 AAV 캡시드 단백질 코딩 서열이 없고, 5' 및 3' ITR은 서로 대칭이다. 대안적인 구현예에서, ceDNA-플라스미드는 제1 (또는 5') 변형 또는 돌연변이된 AAV ITR, 이식유전자를 포함하는 발현 카세트, 및 제2 (또는 3') 돌연변이 또는 변형된 AAV ITR 순서로 인코딩하며, 여기서 상기 ceDNA-플라스미드는 AAV 캡시드 단백질 코딩 서열이 없고, 5' 및 3' 변형된 ITR은 동일한 변형을 갖는다 (즉, 이들은 서로 역 보체이거나 대칭이다).
추가의 구현예에서, ceDNA-플라스미드 시스템은 바이러스 캡시드 단백질 코딩 서열이 없다 (즉, AAV 캡시드 유전자가 없고 다른 바이러스의 캡시드 유전자도 없다). 또한, 특정 구현예에서, ceDNA-플라스미드는 또한 AAV Rep 단백질 코딩 서열이 없다. 따라서, 바람직한 구현예에서, ceDNA-플라스미드는 AAV2)에 대한 기능성 AAV 캡 및 AAV rep 유전자 GG-3', 및 헤어핀 형성을 허용하는 가변 팔린드롬 서열이 없다.
본 발명의 ceDNA-플라스미드는 당 업계에 널리 공지된 임의의 AAV 혈청형의 게놈의 천연 뉴클레오티드 서열을 사용하여 생성될 수 있다. 일 구현예에서, ceDNA-플라스미드 골격은 AAV1, AAV2, AAV3, AAV4, AAV5, AAV 5, AAV7, AAV8, AAV9, AAV10, AAV 11, AAV12, AAVrh8, AAVrh10, AAV-DJ 및 AAV-DJ8 게놈으로부터 유도된다. 예를 들어, NCBI: NC 002077; NC 001401; NC001729; NC001829; NC006152; NC 006260; NC 006261; Kotin and Smith, The Springer Index of Viruses, Springer가 유지 관리하는 URL에서 이용 가능 (www 웹 주소: oesys.springer.de/viruses/database/mkchapter.asp?virID=42.04.)(참고 -URL 또는 데이터베이스에 대한 참조는 본 출원의 유효 출원일자의 URL 또는 데이터베이스의 내용을 지칭함). 특정 구현예에서, ceDNA-플라스미드 골격은 AAV2 게놈으로부터 유래된다. 또 다른 특정 구현예에서, ceDNA-플라스미드 골격은 이들 AAV 게놈 중 하나로부터 유래된 5' 및 3' ITR을 포함하도록 유전자 조작된 합성 골격이다.
ceDNA-플라스미드는 선택적으로 ceDNA 벡터-생성 세포주의 확립에 사용하기 위한 선택 가능한 또는 선별 마커를 포함할 수 있다. 일 구현예에서, 선별 마커는 3' ITR 서열의 하류 (즉 , 3')에 삽입될 수 있다. 또 다른 구현예에서, 선별 마커는 5' ITR 서열의 상류 (즉 , 5')에 삽입될 수 있다. 적절한 선별 마커는, 예를 들어, 약물 내성을 부여하는 마커를 포함한다. 선별 마커는, 예를 들어, 블라스티시딘 S-저항성 유전자, 카나마이신, 제네티신 등일 수 있다. 바람직일 구현예에서, 약물 선별 마커는 블라스티시딘 S-저항성 유전자이다.
예시적인 ceDNA (예를 들어, rAAV0)는 rAAV 플라스미드로부터 생성된다. rAAV 벡터의 생산 방법은 (a) 상기 기재된 바와 같은 rAAV 플라스미드를 숙주 세포에 제공하는 단계로서, 상기 숙주 세포 및 상기 플라스미드 둘 다는 캡시드 단백질 코딩 유전자가 없는, 상기 단계, (b) 숙주 세포를 ceDNA 게놈의 생성을 허용하는 조건 하에 배양하는 단계, 및 (c) 세포를 수거하고 상기 세포로부터 생성된 AAV 게놈을 분리하는 단계를 포함한다.
C. ceDNA 플라스미드로부터 ceDNA 벡터를 제조하는 예시적인 방법
캡시드가 없는 ceDNA 벡터를 제조하는 방법, 특히 생체내 실험에 충분한 벡터를 제공하기 위해 충분히 높은 수율을 갖는 방법이 또한 본원에 제공된다.
일부 구현예에서, ceDNA 벡터의 생산 방법은 (1) 발현 카세트 및 2개의 대칭 ITR 서열을 포함하는 핵산 작제물을 숙주 세포 (예를 들어, Sf9 세포)에 도입하는 단계, (2) 선택적으로, 예를 들어, 플라스미드 상에 존재하는 선별 마커를 사용하여 클론 세포주를 확립하는 단계, (3) Rep 코딩 유전자를 상기 곤충 세포 내로 (상기 유전자를 운반하는 바쿨로바이러스에 의한 형질감염 또는 감염에 의해) 도입하는 단계; 및 (4) 세포를 수거하고 ceDNA 벡터를 정제하는 단계를 포함한다. ceDNA 벡터의 생성을 위해 상기 기술된 발현 카세트 및 2개의 ITR 서열을 포함하는 핵산 작제물은 ceDNA 플라스미드, 또는 하기 기재된 바와 같이 ceDNA 플라스미드로 생성된 박미드 또는 바쿨로바이러스의 형태일 수 있다. 핵산 작제물은 형질감염, 바이러스 형질도입, 안정적인 통합 또는 당 업계에 공지된 다른 방법에 의해 숙주 세포로 도입될 수 있다.
D. 세포주
:
ceDNA 벡터의 생산에 사용되는 숙주 세포주는 Sf9 Sf21과 같은 스포도프테라 프루기페르다(Spodoptera frugiperda), 또는 트리코플루시아 니(Trichoplusia ni) 세포, 또는 기타 무척추동물, 척추동물 또는 포유동물 세포를 포함하는 다른 진핵 세포주로부터 유래된 곤충 세포주를 포함할 수 있다. HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1 180, 단핵구 및 성숙 및 미성숙 수지상 세포과 같은, 당업자에게 공지된 다른 세포주가 또한 사용될 수 있다. 높은 수율의 ceDNA 벡터 생산을 위해 ceDNA-플라스미드의 안정적인 발현을 위해 숙주 세포주를 형질감염시킬 수 있다.
CeDNA-플라스미드는 당 업계에 공지된 시약 (예를 들어, 리포좀, 인산칼슘) 또는 물리적 수단 (예를 들어, 전기천공)을 사용하여 일시적 형질감염에 의해 Sf9 세포 내로 도입될 수 있다. 대안적으로, ceDNA-플라스미드를 게놈에 안정적으로 통합시킨 안정적인 Sf9 세포주가 확립될 수 있다. 이러한 안정적인 세포주는 상기 기술된 바와 같이 선별 마커를 ceDNA-플라스미드에 통합시킴으로써 확립될 수 있다. 세포주를 형질감염시키기 위해 사용된 ceDNA-플라스미드가 항생제와 같은 선별 마커를 포함하는 경우, ceDNA-플라스미드로 형질감염되고 ceDNA-플라스미드 DNA를 게놈에 통합시킨 세포는 세포 성장 배지에 항생제의 첨가에 의해 선별될 수 있다. 이어서, 세포의 내성 클론을 단일 세포 희석 또는 콜로니 전달 기술에 의해 단리하고 증식시킬 수 있다.
E. ceDNA 벡터 분리 및 정제
:
유전자 편집용 ceDNA 벡터를 수득 및 분리하는 과정의 예는 도 4a-4e 및 아래의 특정 예에 기재되어 있다. 본원에 개시된 ceDNA-벡터는 ceDNA-플라스미드, ceDNA-박미드 또는 ceDNA-바쿨로바이러스로 추가로 형질전환된, AAV Rep 단백질(들)을 발현하는 생산 세포로부터 얻을 수 있다. ceDNA 벡터의 생산에 유용한 플라스미드는 도 6a (REP BIIC 생산에 유용), 도 6b (ceDNA 벡터를 얻기 위해 사용된 플라스미드)에 도시된 플라스미드를 포함한다.
일 측면에서, 폴리뉴클레오티드는 플라스미드 (Rep-플라스미드), 박미드 (Rep-박미드) 또는 바쿨로바이러스 (Rep-바쿨로바이러스)에서 생산 세포로 전달된 AAV Rep 단백질 (Rep 78 또는 68)을 인코딩한다. Rep-플라스미드, Rep-박미드 및 Rep-바쿨로바이러스는 위에서 설명한 방법으로 생성될 수 있다.
예시적인 ceDNA 벡터인, ceDNA-벡터를 생성하는 방법이 본원에 기재되어 있다. 본 발명의 ceDNA 벡터를 생성하기 위해 사용되는 발현 작제물은 플라스미드 (예를 들어, ceDNA-플라스미드), 박미드 (예를 들어, ceDNA-박미드) 및/또는 바쿨로바이러스 (예를 들어, ceDNA-바쿨로바이러스)일 수 있다. 단지 예로서, ceDNA-벡터는 ceDNA-바쿨로바이러스 및 Rep-바쿨로바이러스로 공동 감염된 세포로부터 생성될 수 있다. Rep-바쿨로바이러스로부터 생산된 Rep 단백질은 ceDNA-바쿨로바이러스를 복제하여 ceDNA-벡터를 생성할 수 있다. 대안적으로, ceDNA 벡터는 Rep-플라스미드, Rep-박미드 또는 Rep-바쿨로바이러스로 전달된 AAV Rep 단백질 (Rep78/52)을 인코딩하는 서열을 포함하는 작제물로 안정적으로 형질감염된 세포로부터 생성될 수 있다. ceDNA-바쿨로바이러스는 세포로 일시적으로 형질감염될 수 있고, Rep 단백질에 의해 복제되고 ceDNA 벡터를 생성할 수 있다.
박미드 (예를 들어, ceDNA- 박미드)는 Sf9, Sf21, Tni (트리코플루시아 니) 세포, 하이 파이브(High Five) 세포와 같은 허용 곤충 세포로 형질감염될 수 있고, ceDNA-바쿨로바이러스를 생성할 수 있으며, 이는 대칭 ITR 및 발현 카세트를 포함하는 서열을 포함하는 재조합 바쿨로바이러스이다. ceDNA-바쿨로바이러스는 곤충 세포 내로 다시 감염되어 차세대 재조합 바쿨로바이러스를 수득할 수 있다. 선택적으로, 단계는 재조합 바쿨로바이러스를 대량으로 생성하기 위해 한 번 또는 여러 번 반복될 수 있다.
세포로부터 본원에 기재된 ceDNA 벡터를 수거하고 수집하는 시간은 ceDNA 벡터의 고수율 생산을 달성하도록 선택되고 최적화될 수 있다. 예를 들어, 수거 시간은 세포 생존력, 세포 형태, 세포 성장 등을 고려하여 선택될 수 있다. 일반적으로, 세포는 ceDNA 벡터 (예를 들어, ceDNA 벡터)를 생성하기 위해 바쿨로바이러스 감염 후 충분한 시간 후에 그러나 바이러스 독성으로 인해 대부분의 세포가 죽기 시작하기 전에 수거될 수 있다. ceDNA-벡터는 Qiagen ENDO-FREE PLASMID® 키트와 같은 플라스미드 정제 키트를 사용하여 Sf9 세포로부터 분리될 수 있다. 플라스미드 단리를 위해 개발된 다른 방법들도 ceDNA 벡터에 적용될 수 있다. 일반적으로, 상업적으로 입수 가능한 DNA 추출 키트뿐만 아니라 임의의 당 업계에 공지된 핵산 정제 방법이 채택될 수 있다.
대안적으로, 정제는 세포 펠릿을 알칼리 용해 공정에 적용하고, 생성된 용해물을 원심분리하고 크로마토그래피 분리를 수행함으로써 구현될 수 있다. 하나의 비제한적인 예로서, 상기 공정은 핵산을 보유하는 이온 교환 컬럼 (예를 들어, SARTOBIND Q®) 상에 상청액을 로딩한 후 (예를 들어 1.2 M NaCl 용액으로) 용리하고 겔 여과 칼럼 (예를 들어, 6 빠른 흐름 GE) 상에서 추가 크로마토그래피 정제를 수행함으로써 수행될 수 있다. 이어서, 캡시드-비함유 AAV 벡터는, 예를 들어, 침전에 의해 회수된다.
일부 구현예에서, ceDNA 벡터는 또한 엑소좀 또는 미세입자 형태로 정제될 수 있다. 많은 세포 유형이 막 미세소포 쉐딩(membrane microvesicle shedding)을 통해 가용성 단백질뿐만 아니라 복잡한 단백질/핵산 카고도 방출한다는 것이 당 업계에 공지되어 있다 (Cocucci et al, 2009; EP 10306226.1) 이러한 소포는 미세소포 (미세입자로도 지칭됨) 및 엑소좀 (나노소포로도 지칭됨)을 포함하는데, 이 둘은 모두 단백질 및 RNA를 카고로서 포함한다. 미세소포는 원형질막의 직접 버딩(budding)으로부터 생성되고, 엑소좀은 다소포성 엔도좀과 원형질막의 융합시 세포외 환경으로 방출된다. 따라서, ceDNA 벡터-함유 미세소포 및/또는 엑소좀은 ceDNA-플라스미드 또는 ceDNA-플라스미드로 생성된 박미드 또는 바쿨로바이러스로 형질도입된 세포로부터 단리될 수 있다.
미세소포는 배양 배지를 여과하거나 20,000 xg에서, 엑소좀은 100,000 x g에서 초원심분리함으로써 단리될 수 있다. 초원심분리의 최적 지속 시간은 실험적으로 결정될 수 있으며, 소포가 분리되는 특정 세포 유형에 의존할 것이다. 바람직하게는, 배양 배지는 먼저 저속 원심분리 (예를 들어, 2000 x g에서 5-20분 동안)로 제거되고, 예를 들어, AMICON® 스핀 컬럼 (Millipore, Watford, UK)을 사용하여 스핀 농축된다. 미세소포 및 엑소좀은 미세소포 및 엑소좀 상에 존재하는 특정 표면 항원을 인식하는 특정 항체를 사용함으로써 FACS 또는 MACS를 통해 추가로 정제될 수 있다. 다른 미세소포 및 엑소좀 정제 방법은 면역침전, 친화성 크로마토그래피, 여과 및 특정 항체 또는 압타머로 코팅된 자성 비드를 포함하지만, 이에 제한되지는 않는다. 정제시, 소포는, 예를 들어, 포스페이트-완충 염수로 세척된다. ceDNA-함유 소포를 전달하기 위해 미세소포 또는 엑소좀을 사용하는 것의 하나의 이점은 이들 소포가 각각의 세포 유형 상의 특정 수용체에 의해 인식되는 막 단백질을 포함하여 다양한 세포 유형에 표적화될 수 있다는 것이다. (또한 EP 10306226 참조)
본원의 또 다른 측면은 ceDNA 작제물을 자신의 게놈에 안정적으로 통합한 숙주 세포주로부터 ceDNA 벡터를 정제하는 방법에 관한 것이다. 일 구현예에서, ceDNA 벡터는 DNA 분자로서 정제된다. 또 다른 구현예에서, ceDNA 벡터는 엑소좀 또는 미세입자로서 정제된다.
PCT/US18/49996의 도 5는 실시예에 기재된 방법을 사용하여 다수의 ceDNA-플라스미드 작제물로부터 ceDNA의 생성을 확인하는 겔을 보여준다. ceDNA는 실시예에서 도 4d와 관련하여 논의된 바와 같이, 겔에서 특징적인 밴드 패턴에 의해 확인된다.
VIII. 약제학적 조성물
또 다른 측면에서, 약제학적 조성물이 제공된다. 약제학적 조성물은 본원에 개시된 바와 같은 유전자 편집용 ceDNA 벡터 및 약제학적으로 허용되는 담체 또는 희석제를 포함한다.
본원에 개시된 유전자 편집 DNA-벡터는 대상체의 세포, 조직 또는 기관으로의 생체내 전달을 위해 대상체에게 투여하기에 적합한 약제학적 조성물에 포함될 수 있다. 전형적으로, 약제학적 조성물은 본원에 개시된 바와 같은 ceDNA-벡터 및 약제학적으로 허용되는 담체를 포함한다. 예를 들어, 본원에 기재된 ceDNA 벡터는 원하는 치료적 투여 경로 (예를 들어, 비경구 투여)에 적합한 약제학적 조성물에 포함될 수 있다. 고압 정맥내 또는 동맥내 주입, 및 세포내 주사, 예컨대 핵내 미세주입 또는 세포질내 주사를 통한 수동적인 조직 형질도입이 또한 고려된다. 치료 목적을 위한 약제학적 조성물은 용액, 마이크로에멀젼, 분산액, 리포좀 또는 높은 ceDNA 벡터 농도에 적합한 다른 정렬된 구조로서 제형화될 수 있다. 멸균 주사용 용액은 ceDNA 벡터 화합물을 필요한 양으로, 필요에 따라, 상기 열거된 성분 중 하나 또는 조합과 함께 적절한 완충제에 혼입한 후 멸균 여과함으로써 제조될 수 있으며, ceDNA 벡터를 포함하여 제형화하여 핵 내의 이식유전자를 수용자의 유전자에 전달하여, 그 안의 이식유전자 또는 공여체 서열의 치료적 발현을 초래한다. 상기 조성물은 또한 약제학적으로 허용되는 담체를 포함할 수 있다.
ceDNA 벡터를 포함하는 약제학적 활성 조성물은 다양한 목적을 위해 이식유전자 또는 공여체 서열을 세포, 예를 들어, 대상체의 세포로 전달하도록 제형화될 수 있다.
치료 목적을 위한 약제학적 조성물은 전형적으로 제조 및 저장 조건 하에서 멸균되고 안정적이어야 한다. 조성물은 용액, 마이크로에멀젼, 분산액, 리포좀 또는 높은 ceDNA 벡터 농도에 적합한 다른 정렬 구조로 제형화될 수 있다. 멸균 주사용 용액은 ceDNA 벡터 화합물을 필요한 양으로, 필요에 따라, 상기 열거된 성분 중 하나 또는 조합과 함께 적절한 완충제에 혼입한 후 멸균 여과함으로써 제조될 수 있다.
본원에 개시된 바와 같은 ceDNA 벡터는 국소, 전신, 양막내, 경막내, 두개내, 동맥내, 정맥내, 림프내, 복강내, 피하, 기관, 조직내 (예를 들어, 근육내, 심장내, 간내, 신장내, 뇌내), 경막내, 방광내, 결막 (예를 들어, 안와외(extra-orbital), 안와내, 안와후(retroorbital), 망막내, 망막하, 맥락막, 맥락막하, 간질내, 전방내 및 유리체내), 달팽이관내 및 점막 (예를 들어, 구강, 직장, 코) 투여에 적합한 약제학적 조성물에 포함될 수 있다. 고압 정맥내 또는 동맥내 주입, 및 세포내 주사, 예컨대 핵내 미세주입 또는 세포질내 주사를 통한 수동 조직 형질도입이 또한 고려된다.
치료 목적을 위한 약제학적 조성물은 전형적으로 제조 및 저장 조건 하에서 멸균되고 안정적이어야 한다. 조성물은 용액, 마이크로에멀젼, 분산액, 리포좀 또는 높은 ceDNA 벡터 농도에 적합한 다른 정렬 구조로 제형화될 수 있다. 멸균 주사용 용액은 ceDNA 벡터 화합물을 필요한 양으로, 필요에 따라, 상기 열거된 성분 중 하나 또는 조합과 함께 적절한 완충제에 혼입한 후 멸균 여과함으로써 제조될 수 있다.
일부 측면에서, 본원에 제공된 방법은 본원에 개시된 바와 같은 유전자 편집을 위한 하나 이상의 ceDNA 벡터를 숙주 세포로 전달하는 것을 포함한다. 또한, 이러한 방법에 의해 생성된 세포, 및 이러한 세포를 포함하거나 이로부터 생성된 유기체 (예컨대 동물, 식물 또는 진균)가 본원에 제공된다. 핵산의 전달 방법은 리포펙션, 뉴클레오펙션, 마이크로인젝션, 유전자총법(biolistics), 리포좀, 면역리포좀, 다가 양이온 또는 지질:핵산 접합체, 네이키드 DNA, 및 작용제-강화된 DNA 흡수를 포함할 수 있다. 리포펙션은 예를 들어, 미국 특허 번호 5,049,386, 4,946,787; 및 4,897,355)에 기재되어 있으며, 리포펙션 시약은 상업적으로 판매된다 (예를 들어, Transfectam™ 및 Lipofectin™). 전달은 세포 (예를 들어, 시험관내 또는 생체외 투여) 또는 표적 조직 (예를 들어, 생체내 투여)으로의 전달일 수 있다.
핵산을 세포로 전달하기 위한 다양한 기술 및 방법이 당 업계에 공지되어 있다. 예를 들어, ceDNA와 같은 핵산은 지질 나노입자 (LNP), 리피도이드, 리포좀, 지질 나노입자, 리포플렉스 또는 코어-쉘 나노입자로 제형화될 수 있다. 전형적으로, LNP는 핵산 (예를 들어, ceDNA) 분자, 하나 이상의 이온화 가능한 또는 양이온성 지질 (또는 이의 염), 하나 이상의 비이온성 또는 중성 지질 (예를 들어, 인지질), 응집을 방지하는 분자 (예를 들어, PEG 또는 PEG-지질 접합체), 및 임의로 스테롤 (예를 들어, 콜레스테롤)로 구성된다.
ceDNA와 같은 핵산을 세포로 전달하는 또 다른 방법은 세포에 의해 내재화된 리간드와 핵산을 접합시키는 것이다. 예를 들어, 리간드는 세포 표면상의 수용체에 결합하고 세포내이입을 통해 내재화될 수 있다. 리간드는 핵산에서 뉴클레오티드에 공유적으로 연결될 수 있다. 핵산을 세포 내로 전달하기 위한 예시적인 접합체는, 예를 들어, WO2015/006740, WO2014/025805, WO2012/037254, WO2009/082606, WO2009/073809, WO2009/018332, WO2006/112872, WO2004/090108, WO2004/091515 및 WO2017/177326에 기재되어 있다.
ceDNA와 같은 핵산은 또한 형질감염에 의해 세포로 전달될 수 있다. 유용한 형질감염 방법은 지질-매개 형질감염, 양이온성 폴리머-매개 형질감염 또는 인산칼슘 침전을 포함하지만, 이에 제한되지는 않는다. 형질감염 시약은 당 업계에 잘 알려져 있으며, TurboFect 형질감염 시약 (Thermo Fisher Scientific), Pro-Ject 시약 (Thermo Fisher Scientific), TRANSPASS™ P 단백질 형질감염 시약 (New England Biolabs), CHARIOT™ 단백질 전달 시약 (Active Motif), PROTEOJUICE™ 단백질 형질감염 시약 (EMD Millipore), 293fectin, LIPOFECTAMINE™ 2000, LIPOFECTAMINE™ 3000 (Thermo Fisher Scientific), LIPOFECTAMINE™ (Thermo Fisher Scientific), LIPOFECTIN™ (Thermo Fisher Scientific), DMRIE-C, CELLFECTIN™ (Thermo Fisher Scientific), OLIGOFECTAMINE™ (Thermo Fisher Scientific), LIPOFECTACE™, FUGENE™ (Roche, Basel, Switzerland), FUGENE™ HD (Roche), TRANSFECTAM™ (Transfectam, Pr오메가, Madison, Wis.), TFX-10™ (Pr오메가), TFX-20™ (Pr오메가), TFX-50™ (Pr오메가), TRANSFECTIN™ (BioRad, Hercules, Calif.), SILENTFECT™ (Bio-Rad), Effectene™ (Qiagen, Valencia, Calif.), DC-chol (Avanti Polar Lipids), GENEPORTER™ (Gene Therapy Systems, San Diego, Calif.), DHARMAFECT 1™ (Dharmacon, Lafayette, Colo.), DHARMAFECT 2™ (Dharmacon), DHARMAFECT 3™ (Dharmacon), DHARMAFECT 4™ (Dharmacon), ESCORT™ III (Sigma, St. Louis, Mo.) 및 ESCORT™ IV (Sigma Chemical Co.)를 포함하지만, 이에 제한되지 않는다. ceDNA와 같은 핵산은 또한 당업자에게 공지된 미세유체 방법을 통해 세포로 전달될 수 있다.
생체내 또는 생체외에서 핵산의 비-바이러스성 전달 방법은 전기천공, 리포펙션 (미국 특허 번호 5,049,386; 4,946,787 및 Transfectam™ 및 Lipofectin™과 같은 상업적으로 입수 가능한 시약 참조), 미세주입, 유전자총법, 비로좀(virosome), 리포좀 (예를 들어, Crystal, Science 270:404-410 (1995); Blaese et al., Cancer Gene Ther. 2:291-297 (1995); Behr et al., Bioconjugate Chem. 5:382-389 (1994); Remy et al., Bioconjugate Chem. 5:647-654 (1994); Gao et al., Gene Therapy 2:710-722 (1995); Ahmad et al., Cancer Res. 52:4817-4820 (1992); 미국 특허 번호 4,186,183, 4,217,344, 4,235,871, 4,261,975, 4,485,054, 4,501,728, 4,774,085, 4,837,028, 및 4,946,787), 면역리포좀, 다가 양이온 또는 지질:핵산 접합체, 네이키드 DNA, 및 작용제-강화된 DNA 흡수를 포함한다. 예를 들어, Sonitron 2000 시스템 (Rich-Mar)을 사용한 소노포레이션(sonoporation)도 핵산의 전달에 사용될 수 있다.
본원에 기재된 바와 같은 ceDNA 벡터는 또한 생체내에서 세포의 형질도입을 위해 유기체에 직접 투여될 수 있다. 투여는 주사, 주입, 국소 적용 및 전기천공을 포함하지만, 이에 제한되지 않는, 분자를 혈액 또는 조직 세포와 궁극적으로 접촉되게 도입하기 위해 통상적으로 사용되는 임의의 경로에 의해 이루어진다. 이러한 핵산을 투여하는 적합한 방법은 이용 가능하고 당업자에게 잘 알려져 있으며, 하나 초과의 경로가 특정 조성물을 투여하는데 사용될 수 있지만, 특정 경로는 종종 또 다른 경로보다 더 즉각적이고 보다 효과적인 반응을 제공할 수 있다.
본원에 개시된 바와 같은 핵산 벡터 ceDNA 벡터의 도입 방법은, 예를 들어, 미국 특허 번호 5,928,638에 기재된 바와 같은 방법에 의해 조혈 줄기세포로 전달될 수 있다.
본 발명에 따른 ceDNA 벡터는 대상체에서 세포 또는 표적 기관으로 전달하기 위해 리포좀에 첨가될 수 있다. 리포좀은 적어도 하나의 지질 이중층을 갖는 소포이다. 리포좀은 제약 개발의 맥락에서 약물/치료적 전달을 위한 담체로서 전형적으로 사용된다. 이들은 세포막과 융합하고 지질 구조를 재배치하여 약물 또는 활성 제약 성분 (API)을 전달함으로써 작동한다. 이러한 전달을 위한 리포좀 조성물은 인지질, 특히 포스파티딜콜린 기를 갖는 화합물로 구성되지만, 이들 조성물은 또한 다른 지질을 포함할 수 있다.
일부 측면에서, 본 개시내용은 화합물(들)의 면역원성/항원성을 감소시키고 화합물(들)에 친수성 및 소수성을 제공하고, 투여 빈도를 감소시킬 수 있는 폴리에틸렌 글리콜 (PEG) 작용기를 갖는 하나 이상의 화합물 (소위 "PEG화 화합물")을 포함하는 리포좀 제형을 제공한다. 또는 리포좀 제형은 단순히 추가 성분으로서 폴리에틸렌 글리콜 (PEG) 폴리머를 포함한다. 이러한 측면에서, PEG 또는 PEG 작용기의 분자량은 62 Da 내지 약 5,000 Da일 수 있다.
일부 측면에서, 본 개시내용은 수 시간 내지 수 주에 걸쳐 연장 방출 또는 제어 방출 프로파일을 갖는 API를 전달할 리포좀 제형을 제공한다. 일부 관련 측면에서, 리포좀 제형은 지질 이중층에 의해 결합된 수성 챔버를 포함할 수 있다. 다른 관련 측면에서, 리포좀 제형은 승온에서 물리적 전이를 겪는 성분으로 API를 캡슐화하여 수 시간 내지 수 주에 걸쳐 API를 방출한다.
일부 측면에서, 리포좀 제형은 스핑고미엘린 및 본원에 개시된 하나 이상의 지질을 포함한다. 일부 측면에서, 리포좀 제형은 옵티좀을 포함한다.
일부 측면에서, 본 개시내용은 N-(카보닐-메톡시폴리에틸렌 글리콜 2000)-1,2-디스테아로일-sn-글리세로-3-포스포에탄올아민 나트륨 염 (디스테아로일-sn-글리세로-포스포에탄올아민), MPEG (메톡시 폴리에틸렌 글리콜)-공액 지질, HSPC (수소화 대두 포스파티딜콜린); PEG (폴리에틸렌 글리콜); DSPE (디스테아로일-sn-글리세로-포스포에탄올아민); DSPC (디스테아로일포스파티딜콜린); DOPC (디오레오일포스파티딜콜린); DPPG (디팔미토일포스파티딜글리세롤); EPC (계란 포스파티딜콜린); DOPS (디올레오일포스파티딜세린); POPC (팔미토일로레오일포스파티딜콜린); SM (스핑고미엘린); MPEG (메톡시 폴리에틸렌 글리콜); DMPC (디미리스토일 포스파티딜콜린); DMPG (디미리스토일 포스파티딜글리세롤); DSPG (디스테아로일포스파티딜글리세롤); DEPC (디에루코일포스파티딜콜린); DOPE (디올레올리-sn-글리세로-포스포에탄올아민). 콜레스테릴 설페이트 (CS), 디팔미토일포스파티딜글리세롤 (DPPG), DOPC (디올레올리-sn-글리세로-포스파티딜콜린) 또는 이들의 임의의 조합으로부터 선택된 하나 이상의 지질을 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 인지질, 콜레스테롤 및 PEG화 지질을 56:38:5의 몰비로 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형의 전체 지질 함량은 2-16 mg/mL이다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질 및 PEG화 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질 및 PEG화 지질을 각각 3:0.015:2의 몰비로 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기, 콜레스테롤 및 PEG화 지질을 함유하는 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기 및 콜레스테롤을 함유하는 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, PEG화 지질은 PEG-2000-DSPE이다. 일부 측면에서, 본 개시내용은 DPPG, 대두 PC, MPEG-DSPE 지질 접합체 및 콜레스테롤을 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 하나 이상의 지질 및 에탄올아민 작용기를 함유하는 하나 이상의 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질, 및 스테롤, 예를 들어, 콜레스테롤을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형은 DOPC/DEPC; 및 DOPE를 포함한다.
일부 측면에서, 본 개시내용은 하나 이상의 약제학적 부형제, 예를 들어, 수 크로스 및/또는 글리신을 추가로 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 구조상 단일 라멜라 또는 다중 라멜라가 있는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 다중-소포성 입자 및/또는 발포체-기반 입자를 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 일반적인 나노입자에 비해 크기가 크고, 크기가 약 150 내지 250 nm인 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형은 동결 건조된 분말이다.
일부 측면에서, 본 개시내용은 리포좀 외부에 단리된 ceDNA를 갖는 혼합물에 약한 염기를 첨가함으로써 본원에 개시되거나 기재된 ceDNA 벡터로 제조되고 로딩되는 리포좀 제형을 제공한다. 이 첨가는 리포좀 외부의 pH를 약 7.3으로 증가시키고 API를 리포좀으로 유도한다. 일부 측면에서, 본 개시내용은 리포좀 내부에서 산성인 pH를 갖는 리포좀 제형을 제공한다. 이러한 경우, 리포좀 내부는 pH 4-6.9, 보다 바람직하게는 pH 6.5일 수 있다. 다른 측면에서, 본 개시내용은 리포좀내 약물 안정화 기술을 사용하여 제조된 리포좀 제형을 제공한다. 이러한 경우에, 폴리머 또는 비-폴리머 고도로 하전된 음이온 및 리포좀내 포획제, 예를 들어 폴리포스페이트 또는 수크로스 옥타설페이트가 사용된다.
다른 측면에서, 본 개시내용은 인지질, 레시틴, 포스파티딜콜린 및 포스파티딜에탄올아민을 포함하는 리포좀 제형을 제공한다.
리포좀, 나노캡슐, 미세입자, 미세구, 지질 입자, 소포 등과 같은 전달 시약은 본 개시내용의 조성물을 적합한 숙주 세포로 도입하기 위해 사용될 수 있다. 특히, 핵산은 지질 입자, 리포좀, 소포, 나노구, 나노입자, 금 입자 등으로 캡슐화되어 전달되도록 제형화될 수 있다. 이러한 제형은 본원에 개시된 핵산의 약제학적으로 허용되는 제형의 도입에 바람직할 수 있다.
당 업계에 공지된 다양한 전달 방법 또는 이의 변형을 사용하여 시험관내 또는 생체내에서 ceDNA 벡터를 전달할 수 있다. 예를 들어, 일부 구현예에서, ceDNA 벡터는 표적화된 세포로의 DNA 유입이 촉진되도록 기계적, 전기, 초음파, 유체 역학적 또는 레이저-기반 에너지에 의해 세포막에 일시적 침투를 함으로써 전달된다. 예를 들어, ceDNA 벡터는 크기 제한 채널을 통해 또는 당 업계에 공지된 다른 수단을 통해 세포를 압착함으로써 일시적으로 세포막을 파괴함으로써 전달될 수 있다. 일부 경우에, ceDNA 벡터는 단독으로 피부, 흉선, 심장 근육, 골격근 또는 간 세포에 네이키드 DNA로서 직접 주사된다.
일부 경우에, ceDNA 벡터는 유전자 총에 의해 전달된다. 캡시드가 없는 AAV 벡터로 코팅된 금 또는 텅스텐 구형 입자 (1-3 μm 직경)는 가압된 가스를 통해 표적 조직 세포로 침투하기 위해 고속으로 가속화될 수 있다.
ceDNA 벡터 및 약제학적으로 허용되는 담체를 포함하는 조성물이 본원에서 구체적으로 고려된다. 일부 구현예에서, ceDNA 벡터는 지질 전달 시스템, 예를 들어, 본원에 기재된 리포좀으로 제형화된다. 일부 구현예에서, 이러한 조성물은 숙련된 전문가가 원하는 임의의 경로로 투여된다. 상기 조성물은 경구, 비경구, 설하, 경피, 직장, 경점막, 국소, 흡입, 협측 투여, 흉막내, 정맥내, 동맥내, 복강내, 피하, 근육내, 비강내 경막내, 및 관절내 또는 이들의 조합을 포함하여 상이한 경로로 대상체에게 투여될 수 있다. 수의용으로, 조성물은 정상적인 수의학적 실무에 따라 적합하게 허용되는 제형으로 투여될 수 있다. 수의사는 특정 동물에 가장 적합한 투여 요법 및 투여 경로를 쉽게 결정할 수 있다. 조성물은 전통적인 주사기, 바늘이 없는 주입 장치, "미세발사체 충격 건(microprojectile bombardment gone gun)" 또는 전기천공 ("EP"), "유체 역학적 방법" 또는 초음파와 같은 다른 물리적 방법에 의해 투여될 수 있다.
조성물은 본원에 기재된 바와 같이, 리포좀 매개된 또는 나노입자 촉진된 생체내 전기천공과 함께 또는 없이 DNA 주사 (DNA 백신 접종으로도 지칭됨)를 포함하는 여러 기술에 의해 대상체에게 전달될 수 있다.
일부 구현예에서, 전기천공은 ceDNA 벡터를 전달하는데 사용된다. 전기 천공은 한 쌍의 전극을 조직에 삽입함으로써 세포막 표적 세포 조직의 일시적인 불안정화를 야기하여 불안정화 막의 주변 매질의 DNA 분자가 세포의 세포질 및 핵원형질 내로 침투할 수 있도록 한다. 전기천공은 피부, 폐 및 근육과 같은 많은 유형의 조직에 대해 생체내에서 사용되어 왔다.
일부 경우에, ceDNA 벡터는 유체 역학적 주사에 의해 전달되는데, 이는 임의의 수용성 화합물 및 입자를 사지 전체의 내부 기관 및 골격근으로 직접 세포내로 전달하기 위한 간단하고 매우 효율적인 방법이다.
일부 경우에, ceDNA 벡터는 내부 장기 또는 종양의 세포 내로 DNA 입자의 세포내 전달을 용이하게 하기 위해 막에 나노스코픽 공극을 만들어 초음파에 의해 전달되므로, 플라스미드 DNA의 크기 및 농도는 시스템의 효율에 큰 역할을 한다. 일부 경우에, ceDNA 벡터는 핵산을 함유하는 입자를 표적 세포 내로 농축시키기 위해 자기장을 사용하여 자기감염에 의해 전달된다.
일부 경우에, 화학적 전달 시스템은, 예를 들어, 나노머 복합체를 사용함으로써 사용될 수 있으며, 이는 양이온성 리포좀/미쉘 또는 양이온성 폴리머에 속하는, 다가 양이온 나노머 입자에 의한 음전하 핵산의 압축을 포함한다. 전달 방법에 사용되는 양이온성 지질은 1가 양이온성 지질, 다가 양이온성 지질, 구아니딘 함유 화합물, 콜레스테롤 유도체 화합물, 양이온성 폴리머 (예를 들어, 폴리(에틸렌이민), 폴리-L-리신, 프로타민, 기타 양이온성 폴리머), 및 지질-폴리머 하이브리드를 포함하지만, 이에 제한되지 않는다.
A. 엑소좀:
일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 엑소좀에 패키징되어 전달된다. 엑소좀은 다소포체와 원형질막의 융합 후 세포외 환경으로 방출되는 식작용 기원의 작은 막 소포이다. 이들의 표면은 공여체 세포의 세포막으로부터의 지질 이중층으로 구성되며, 이들은 엑소좀을 생성하는 세포로부터의 시토졸을 함유하고, 표면상의 모 세포로부터 막 단백질을 나타낸다. 엑소좀은 상피 세포, B 및 T 림프구, 비만 세포 (MC) 및 수지상 세포 (DC)를 포함하는 다양한 세포 유형에 의해 생성된다. 일부 구현예, 직경이 10nm 내지 1μm, 20nm 내지 500nm, 30nm 내지 250nm, 50nm 내지 100nm인 엑소좀이 사용되는 것으로 예상된다. 공여체 세포를 사용하거나 특정 핵산을 이들에 도입함으로써 표적 세포로의 전달을 위한 엑소좀을 단리할 수 있다. 당 업계에 공지된 다양한 접근법을 사용하여 본 발명의 캡시드-비함유 AAV 벡터를 함유하는 엑소좀을 생성할 수 있다.
B. 미세입자/나노입자:
일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 지질 나노입자에 의해 전달된다. 일반적으로, 지질 나노입자는 예를 들어 하기에 개시된 바와 같이, 이온화 가능한 아미노 지질 (예를 들어, 헵타트리아콘타-6,9,28,31-테트라엔-19-일 4-(디메틸아미노)부타노에이트, DLin-MC3-DMA, 포스파티딜콜린 (1,2-디스테아로일-sn-글리세로-3-포스포콜린, DSPC), 콜레스테롤 및 코트 지질 (폴리에틸렌 글리콜-디미리스토글리세롤, PEG-DMG)을 포함한다: Tam et al. (2013). Advances in Lipid Nanoparticles for siRNA delivery. Pharmaceuticals 5(3): 498-507.
일부 구현예에서, 지질 나노입자는 약 10 내지 약 1000 nm의 평균 직경을 갖는다. 일부 구현예에서, 지질 나노입자는 300 nm 미만의 직경을 갖는다. 일부 구현예에서, 지질 나노입자는 약 10 내지 약 300 nm의 직경을 갖는다. 일부 구현예에서, 지질 나노입자는 200 nm 미만의 직경을 갖는다. 일부 구현예에서, 지질 나노입자는 약 25 내지 약 200 nm의 직경을 갖는다. 일부 구현예에서, 지질 나노입자 제제 (예를 들어, 복수의 지질 나노입자를 포함하는 조성물)는 평균 크기 (예를 들어, 직경)가 약 70 nm 내지 약 200 nm이고, 보다 전형적으로 평균 크기가 약 100nm 이하인 크기 분포를 갖는다.
당 업계에 공지된 다양한 지질 나노입자가 본원에 개시된 ceDNA 벡터를 전달하는데 사용될 수 있다. 예를 들어, 지질 나노입자를 사용한 다양한 전달 방법은 미국 특허 번호 9,404,127, 9,006,417 및 9,518,272에 기재되어 있다.
일부 구현예에서, 본원에 개시된 ceDNA 벡터는 금 나노입자에 의해 전달된다. 일반적으로, 핵산은, 예를 들어, 하기에 기재된 바와 같이, 금 나노입자에 공유 결합될 수 있거나 금 나노입자에 비공유 결합 (예를 들어, 전하-전하 상호작용에 의해 결합)될 수 있다: Ding et al. (2014). Gold Nanoparticles for Nucleic Acid Delivery. Mol. Ther. 22(6); 1075-1083. 일부 구현예에서, 금 나노입자-핵산 접합체는, 예를 들어, 미국 특허 번호 6,812,334에 기재된 방법을 사용하여 제조된다.
C. 접합체
일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 접합된다 (예를 들어, 세포 흡수를 증가시키는 작용제에 공유 결합됨). "세포 흡수를 증가시키는 작용제"는 지질 막을 통한 핵산의 수송을 용이하게 하는 분자이다. 예를 들어, 핵산은 친유성 화합물 (예를 들어, 콜레스테롤, 토코페롤 등), 세포 침투 펩티드 (CPP) (예를 들어, 페네트라틴, TAT, Syn1B 등) 및 폴리아민 (예를 들어, 스퍼민)에 접합될 수 있다. 세포 흡수를 증가시키는 작용제의 추가 예는, 예를 들어, 하기에 개시되어 있다: Winkler (2013). Oligonucleotide conjugates for therapeutic applications. Ther. Deliv. 4(7); 791-809.
일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 폴리머 (예를 들어, 폴리머 분자) 또는 폴레이트 분자 (예를 들어, 엽산 분자)에 접합된다. 일반적으로, 폴리머에 접합된 핵산의 전달은, 예를 들어 WO2000/34343 및 WO2008/022309에 기재된 바와 같이, 당 업계에 공지되어 있다. 일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는, 예를 들어 미국 특허 번호 8,987,377에 기재된 바와 같은 폴리(아미드) 폴리머에 접합된다. 일부 구현예에서, 본 개시내용에 기재된 핵산은 미국 특허 번호 8,507,455에 기재된 바와 같은 엽산 분자에 접합된다.
일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는, 예를 들어, 미국 특허 번호 8,450,467에 기재된 바와 같이 탄수화물에 접합된다.
D. 나노캡슐
대안적으로, 본원에 개시된 바와 같은 ceDNA 벡터의 나노캡슐 제형이 사용될 수 있다. 나노캡슐은 일반적으로 물질을 안정적이고 재현 가능한 방식으로 포획할 수 있다. 세포내 폴리머 과부하로 인한 부작용을 피하기 위해, 이러한 초미립자 (약 0.1 μm 크기)는 생체내에서 분해될 수 있는 폴리머를 사용하여 설계되어야 한다. 이들 요건을 충족시키는 생분해성 폴리알킬-시아노아크릴레이트 나노입자가 사용될 것으로 고려된다.
E. 리포솜
본 개시내용에 따른 ceDNA 벡터는 대상체에서 세포 또는 표적 기관으로 전달하기 위해 리포좀에 첨가될 수 있다. 리포좀은 적어도 하나의 지질 이중층을 갖는 소포이다. 리포좀은 제약 개발의 맥락에서 약물/치료적 전달을 위한 담체로서 전형적으로 사용된다. 이들은 세포막과 융합하고 이의 지질 구조를 재배치하여 약물 또는 활성 제약 성분 (API)을 전달함으로써 작동한다. 이러한 전달을 위한 리포좀 조성물은 인지질, 특히 포스파티딜콜린 기를 갖는 화합물로 구성되지만, 이들 조성물은 또한 다른 지질을 포함할 수 있다.
리포좀의 형성 및 사용은 일반적으로 당업자에게 공지되어 있다. 리포좀은 개선된 혈청 안정성 및 순환 하프-타임(half-time)으로 개발되었다 (미국 특허 번호 5,741,516). 또한, 잠재적 약물 담체로서의 리포좀 및 리포좀 유사 제제의 다양한 방법이 기술되어 있다 (미국 특허 번호 5,567,434; 5,552,157; 5,565,213; 5,738,868 및 5,795,587).
리포좀은 다른 절차에 의해 일반적으로 형질감염에 저항성인 다수의 세포 유형과 함께 성공적으로 사용되었다. 또한, 리포좀은 바이러스-기반 전달 시스템에 전형적인 DNA 길이 제한이 없다. 리포좀은 유전자, 약물, 방사선 치료제, 바이러스, 전사 인자 및 알로스테릭 이펙터를 다양한 배양 세포주 및 동물에 도입하는데 효과적으로 사용되어 왔다. 또한, 리포좀-매개 약물 전달의 효과를 검사하는 몇몇 성공적인 임상 시험이 완료되었다.
리포좀은 수성 매질에 분산되어 자발적으로 다중 라멜라 동심 이중층 소포 (다중 라멜라 소포 (multilamellar vesicle; MLV)라고도 함)를 형성하는 인지질로부터 형성된다. MLV는 일반적으로 25 nm 내지 4 μm의 직경을 갖는다. MLV의 초음파 처리에 의해 코어에 수용액을 함유하는, 200 내지 500 ANG 범위의 직경을 갖는 작은 단일 라멜라 소포 (SUV)가 형성된다.
일부 구현예에서, 리포좀은 양이온성 지질을 포함한다. 용어 "양이온성 지질"은 극성 및 비극성 도메인 둘 다를 갖고, 생리학적 pH에서 또는 그 근처에서 양으로 하전될 수 있고, 핵산과 같이 다가 음이온에 결합하고, 세포로 핵산의 전달을 용이하게 하는 지질 및 합성 지질을 포함한다. 일부 구현예에서, 양이온성 지질은 포화 및 불포화 알킬 및 지환족 에테르 및 아민, 아미드 또는 이의 유도체의 에스테르를 포함한다. 일부 구현예에서, 양이온성 지질은 직쇄, 분지형 알킬, 알케닐 기 또는 상기의 임의의 조합을 포함한다. 일부 구현예에서, 양이온성 지질은 1 내지 약 25개의 탄소 원자 (예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 또는 25개의 탄소 원자를 함유한다. 일부 구현예에서, 양이온성 지질은 25개 초과의 탄소 원자를 함유한다. 일부 구현예에서, 직쇄 또는 분지형 알킬 또는 알켄 기는 6개 이상의 탄소 원자를 갖는다. 양이온성 지질은 또한, 일부 구현예에서, 하나 이상의 지환족 기를 포함할 수 있다. 지환족 기의 비제한적인 예는 콜레스테롤 및 다른 스테로이드 기를 포함한다. 일부 구현예에서, 양이온성 지질은 하나 이상의 반대 이온으로 제조된다. 반대 이온 (음이온)의 예로는 Cl-, Br-, I-, F-, 아세트산염, 트리플루오로아세트산염, 황산염, 아질산염, 질산염을 포함하지만, 이에 제한되지 않는다.
일부 측면에서, 본 개시내용은 화합물(들)의 면역원성/항원성을 감소시키고 화합물(들)에 친수성 및 소수성을 제공하고 투여 빈도를 감소시킬 수 있는 폴리에틸렌 글리콜 (PEG) 작용기를 갖는 하나 이상의 화합물 (소위 "PEG화 화합물")을 포함하는 리포좀 제형을 제공한다. 또는 리포좀 제형은 단순히 추가 성분으로서 폴리에틸렌 글리콜 (PEG) 폴리머를 포함한다. 이러한 측면에서, PEG 또는 PEG 작용기의 분자량은 62 Da 내지 약 5,000 Da일 수 있다.
일부 측면에서, 본 개시내용은 수 시간 내지 수 주에 걸쳐 연장 방출 또는 제어 방출 프로파일을 갖는 API를 전달할 리포좀 제형을 제공한다. 일부 관련 측면에서, 리포좀 제형은 지질 이중층에 의해 결합된 수성 챔버를 포함할 수 있다. 다른 관련 측면에서, 리포좀 제형은 승온에서 물리적 전이를 겪는 성분으로 API를 캡슐화하여 수 시간 내지 수 주에 걸쳐 API를 방출한다.
일부 측면에서, 리포좀 제형은 스핑고미엘린 및 본원에 개시된 하나 이상의 지질을 포함한다. 일부 측면에서, 리포좀 제형은 옵티좀을 포함한다.
일부 측면에서, 본 개시내용은 N-(카보닐-메톡시폴리에틸렌 글리콜 2000)-1,2-디스테아로일-sn-글리세로-3-포스포에탄올아민 나트륨 염, (디스테아로일-sn-글리세로-포스포에탄올아민), MPEG (메톡시폴리에틸렌 글리콜)-공액 지질, HSPC (수소화 대두 포스파티딜콜린); PEG (폴리에틸렌 글리콜); DSPE (디스테아로일-sn-글리세로-포스포에탄올아민); DSPC (디스테아로일포스파티딜콜린); DOPC (디올레오일포스파티딜콜린); DPPG (디팔미토일포스파티딜글리세롤); EPC (계란 포스파티딜콜린); DOPS (디올레오일포스파티딜세린); POPC (팔미토일로레오일포스파티딜콜린); SM (스핑고미엘린); MPEG (메톡시 폴리에틸렌 글리콜); DMPC (디미리스토일 포스파티딜콜린); DMPG (디미리스토일 포스파티딜글리세롤); DSPG (디스테아로일포스파티딜글리세롤); DEPC (디에루코일포스파티딜콜린); DOPE (디올레올리-sn-글리세로-포스포에탄올아민). 콜레스테릴 설페이트 (CS), 디팔미토일포스파티딜글리세롤 (DPPG), DOPC (디올레올리-sn-글리세로-포스파티딜콜린) 또는 이들의 임의의 조합으로부터 선택된 하나 이상의 지질을 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 인지질, 콜레스테롤 및 PEG화 지질을 56:38:5의 몰비로 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형의 전체 지질 함량은 2-16 mg/mL이다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질 및 PEG화 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질 및 PEG화 지질을 각각 3:0.015:2의 몰비로 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 콜레스테롤 및 PEG화 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기 및 콜레스테롤을 함유하는 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, PEG화 지질은 PEG-2000-DSPE이다. 일부 측면에서, 본 개시내용은 DPPG, 대두 PC, MPEG-DSPE 지질 접합체 및 콜레스테롤을 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 하나 이상의 지질 및 에탄올아민 작용기를 함유하는 하나 이상의 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질 및 스테롤, 예를 들어 콜레스테롤 중 하나 이상을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형은 DOPC/DEPC; 및 DOPE를 포함한다.
일부 측면에서, 본 개시내용은 하나 이상의 약제학적 부형제, 예를 들어 수 크로스 및/또는 글리신을 추가로 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 구조상 단일 라멜라 또는 다중 라멜라가 있는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 다중-소포성 입자 및/또는 발포체-기반 입자를 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 일반적인 나노입자에 비해 크기가 크고 크기가 약 150 내지 250 nm인 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형은 동결 건조된 분말이다.
일부 측면에서, 본 개시내용은 리포좀 외부에 단리된 ceDNA를 갖는 혼합물에 약한 염기를 첨가함으로써 본원에 개시되거나 기재된 ceDNA 벡터로 제조되고 로딩된 리포좀 제형을 제공한다. 이 첨가는 리포좀 외부의 pH를 약 7.3으로 증가시키고 API를 리포좀으로 유도한다. 일부 측면에서, 본 개시내용은 리포좀 내부에서 산성인 pH를 갖는 리포좀 제형을 제공한다. 이러한 경우, 리포좀 내부는 pH 4-6.9, 보다 바람직하게는 pH 6.5일 수 있다. 다른 측면에서, 본 개시내용은 리포좀내 약물 안정화 기술을 사용하여 제조된 리포좀 제형을 제공한다. 이러한 경우에, 폴리머 또는 비-폴리머 고도로 하전된 음이온 및 리포좀내 포획제, 예를 들어 폴리포스페이트 또는 수크로스 옥타설페이트가 사용된다.
다른 측면에서, 본 개시내용은 인지질, 레시틴, 포스파티딜콜린 및 포스파티딜에탄올아민을 포함하는 리포좀 제형을 제공한다.
양이온성 지질의 비제한적인 예는 폴리에틸렌이민, 폴리아미도아민 (PAMAM) 스타버스트 덴드리머, 리포펙틴 (DOTMA 및 DOPE의 조합), 리포펙타제, LIPOFECTAMINE™ (예를 들어, LIPOFECTAMINE™ 2000), DOPE, 시토펙틴(Cytofectin) (Gilead Sciences, Foster City, Calif.) 및 유펙틴(Eufectins) (JBL, San Luis Obispo, CA)를 포함한다. 예시적인 양이온성 리포좀은 N-[1-(2,3-디올레올옥시)-프로필]-N,N,N-트리메틸암모늄 클로라이드 (DOTMA), N-[1-(2,3-디올레올옥시)-프로필]-N,N,N-트리메틸암모늄 메틸설페이트 (DOTAP), 3β-[N-(N',N'-디메틸아미노에탄)카바모일] 콜레스테롤 (DC-Chol), 2,3,-디올레일옥시-N-[2(스퍼민카복사미도)에틸]-N,N-디메틸-1-프로판아미늄 트리플루오로아세테이트 (DOSPA), 1,2-디미리스틸옥시프로필-3-디메틸-하이드록시에틸 암모늄 브로마이드; 및 디메틸디옥타데실암모늄 브로마이드 (DDAB)로부터 제조될 수 있다. 핵산 (예를 들어, CELiD)은 또한, 예를 들어, 폴리 (L-라이신) 또는 아비딘과 복합체화될 수 있고, 지질은 이 혼합물, 예를 들어, 스테릴-폴리 (L-라이신)에 포함될 수 있거나 포함되지 않을 수 있다.
일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 미국 특허 번호 8,158,601에 기재된 양이온성 지질, 또는 미국 특허 번호 8,034,376에 기재된 바와 같은 폴리아민 화합물 또는 지질을 사용하여 전달된다.
F. 예시적인 리포좀 및 지질 나노입자 (LNP) 조성물
본 발명에 따른 ceDNA 벡터는 유전자 편집이 필요한, 예를 들어, 공여체 서열이 필요한 세포로 전달하기 위해 리포좀에 첨가될 수 있다. 리포좀은 적어도 하나의 지질 이중층을 갖는 소포이다. 리포좀은 제약 개발의 맥락에서 약물/치료적 전달을 위한 담체로서 전형적으로 사용된다. 이들은 세포막과 융합하고 이의 지질 구조를 재배치하여 약물 또는 활성 제약 성분 (API)을 전달함으로써 작동한다. 이러한 전달을 위한 리포좀 조성물은 인지질, 특히 포스파티딜콜린 기를 갖는 화합물로 구성되지만, 이들 조성물은 또한 다른 지질을 포함할 수 있다.
일부 측면에서, 본 개시내용은 화합물(들)의 면역원성/항원성을 감소시키고, 친수성 및 소수성을 제공하고, 투여 빈도를 감소시킬 수 있는 폴리에틸렌 글리콜 (PEG) 작용기를 갖는 하나 이상의 화합물 (소위 "PEG화 화합물")을 포함하는 리포좀 제형을 제공한다. 또는 리포좀 제형은 단순히 추가 성분으로서 폴리에틸렌 글리콜 (PEG) 폴리머를 포함한다. 이러한 측면에서, PEG 또는 PEG 작용기의 분자량은 62 Da 내지 약 5,000 Da일 수 있다.
일부 측면에서, 본 개시내용은 수 시간 내지 수 주에 걸쳐 연장 방출 또는 제어 방출 프로파일을 갖는 API를 전달할 리포좀 제형을 제공한다. 일부 관련 측면에서, 리포좀 제형은 지질 이중층에 의해 결합된 수성 챔버를 포함할 수 있다. 다른 관련 측면에서, 리포좀 제형은 승온에서 물리적 전이를 겪는 성분으로 API를 캡슐화하여 수 시간 내지 수 주에 걸쳐 API를 방출한다.
일부 측면에서, 리포좀 제형은 스핑고미엘린 및 본원에 개시된 하나 이상의 지질을 포함한다. 일부 측면에서, 리포좀 제형은 옵티좀을 포함한다.
일부 측면에서, 본 개시내용은 N-(카보닐-메톡시폴리에틸렌 글리콜 2000)-1,2-디스테아로일-sn-글리세로-3-포스포에탄올아민 나트륨염 (디스테아로일-sn-글리세로-포스포에탄올아민), MPEG (메톡시 폴리에틸렌 글리콜)-공액 지질, HSPC (수소화 대두 포스파티딜콜린); PEG (폴리에틸렌 글리콜); DSPE (디스테아로일-sn-글리세로-포스포에탄올아민); DSPC (디스테아로일포스파티딜콜린); DOPC (디올레오일포스파티딜콜린); DPPG (디팔미토일포스파티딜글리세롤); EPC (계란 포스파티딜콜린); DOPS (디올레오일포스파티딜세린); POPC (팔미토일로레오일포스파티딜콜린); SM (스핑고미엘린); MPEG (메톡시 폴리에틸렌 글리콜); DMPC (디미리스토일 포스파티딜콜린); DMPG (디미리스토일 포스파티딜글리세롤); DSPG (디스테아로일포스파티딜글리세롤); DEPC (디에루코일포스파티딜콜린); DOPE (디올레올리-sn-글리세로-포스포에탄올아민). 콜레스테릴 설페이트 (CS), 디팔미토일포스파티딜글리세롤 (DPPG), DOPC (디오롤리-sn-글리세로-포스파티딜콜린) 또는 이들의 임의의 조합으로부터 선택된 하나 이상의 지질을 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 인지질, 콜레스테롤 및 PEG화 지질을 56:38:5의 몰비로 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형의 전체 지질 함량은 2-16 mg/mL이다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질 및 PEG화 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질 및 PEG화 지질을 각각 3:0.015:2의 몰비로 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 콜레스테롤 및 PEG화 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질 및 콜레스테롤을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, PEG화 지질은 PEG-2000-DSPE이다. 일부 측면에서, 본 개시내용은 DPPG, 대두 PC, MPEG-DSPE 지질 접합체 및 콜레스테롤을 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 하나 이상의 지질 및 에탄올아민 작용기를 함유하는 하나 이상의 지질을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 포스파티딜콜린 작용기를 함유하는 지질, 에탄올아민 작용기를 함유하는 지질 및 스테롤, 예를 들어 콜레스테롤 중 하나 이상을 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형은 DOPC/DEPC; 및 DOPE를 포함한다.
일부 측면에서, 본 개시내용은 하나 이상의 약제학적 부형제, 예를 들어, 수 크로스 및/또는 글리신을 추가로 포함하는 리포좀 제형을 제공한다.
일부 측면에서, 본 개시내용은 구조상 단일 라멜라 또는 다중 라멜라인 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 다중 소포성 입자 및/또는 발포체 기반 입자를 포함하는 리포좀 제형을 제공한다. 일부 측면에서, 본 개시내용은 일반적인 나노입자에 비해 크기가 크고 크기가 약 150 내지 250 nm인 리포좀 제형을 제공한다. 일부 측면에서, 리포좀 제형은 동결 건조된 분말이다.
일부 측면에서, 본 개시내용은 리포좀 외부에 단리된 ceDNA를 갖는 혼합물에 약한 염기를 첨가함으로써 본원에 개시되거나 기재된 ceDNA 벡터로 제조되고 로딩된 리포좀 제형을 제공한다. 이 첨가는 리포좀 외부의 pH를 약 7.3으로 증가시키고 API를 리포좀으로 유도한다. 일부 측면에서, 본 개시내용은 리포좀 내부에서 산성인 pH를 갖는 리포좀 제형을 제공한다. 이러한 경우, 리포좀 내부는 pH 4-6.9, 보다 바람직하게는 pH 6.5일 수 있다. 다른 측면에서, 본 개시내용은 리포좀내 약물 안정화 기술을 사용하여 제조된 리포좀 제형을 제공한다. 이러한 경우에, 폴리머 또는 비-폴리머 고도로 하전된 음이온 및 리포좀내 포획제, 예를 들어 폴리포스페이트 또는 수크로스 옥타설페이트가 사용된다.
다른 측면에서, 본 개시내용은 인지질, 레시틴, 포스파티딜콜린 및 포스파티딜에탄올아민을 포함하는 리포좀 제형을 제공한다. 일부 구현예에서, 리포좀 제형은 하기 표 7에 기재된 제형이다.
[표 7]
일부 측면에서, 본 개시내용은 ceDNA 및 이온화 가능한 지질을 포함하는 지질 나노입자를 제공한다. 예를 들어, 본원에 포함된, 2018년 9월 7일자로 출원된 국제 출원 PCT/US2018/050042에 개시된 바와 같은 방법에 의해 수득된 ceDNA로 제조되고 로딩된 지질 나노입자 제형. 이는 이온화 가능한 지질을 양성화하고 ceDNA/지질 회합 및 입자의 핵 형성에 유리한 에너지학을 제공하는 낮은 pH에서 에탄올성 지질과 수성 ceDNA의 고에너지 혼합에 의해 달성될 수 있다. 입자는 유기 용매의 수성 희석 및 제거를 통해 추가로 안정화될 수 있다. 입자는 원하는 수준으로 농축될 수 있다.
일반적으로, 지질 입자는 약 10:1 내지 30:1의 총 지질 대 ceDNA (질량 또는 중량) 비로 제조된다. 일부 구현예에서, 지질 대 ceDNA 비 (질량/질량 비; w/w 비)는 약 1:1 내지 약 25:1, 약 10:1 내지 약 14:1, 약 3:1 내지 약 15:1, 약 4:1 내지 약 10:1, 약 5:1 내지 약 9:1, 또는 약 6:1 내지 약 9:1의 범위일 수 있다. 지질 및 ceDNA의 양은 원하는 N/P 비, 예를 들어, 3, 4, 5, 6, 7, 8, 9, 10 이상의 N/P 비를 제공하도록 조정될 수 있다. 일반적으로, 지질 입자 제형의 전체 지질 함량은 약 5 mg/ml 내지 약 30 mg/mL의 범위일 수 있다.
이온화 가능한 지질은 전형적으로 핵산 카고, 예를 들어, ceDNA를 낮은 pH에서 응축시키고 막 회합 및 융합유도성(fusogenicity)을 유도하기 위해 사용된다. 일반적으로, 이온화 가능한 지질은 산성 조건, 예를 들어 pH 6.5 이하에서 양으로 하전되거나 양성화되는 적어도 하나의 아미노기를 포함하는 지질이다. 이온화 가능한 지질은 또한 본원에서 양이온성 지질로 지칭된다.
예시적인 이온화 가능한 지질은 PCT 특허 공보 WO2015/095340, WO2015/199952, WO2018/011633, WO2017/049245, WO2015/061467, WO2012/040184, WO2012/000104, WO2015/074085, WO2016/081029, WO2017/004143, WO2017/075531, WO2017/117528, WO2011/022460, WO2013/148541, WO2013/116126, WO2011/153120, WO2012/044638, WO2012/054365, WO2011/090965, WO2013/016058, WO2012/162210, WO2008/042973, WO2010/129709, WO2010/144740 , WO2012/099755, WO2013/049328, WO2013/086322, WO2013/086373, WO2011/071860, WO2009/132131, WO2010/048536, WO2010/088537, WO2010/054401, WO2010/054406 , WO2010/054405, WO2010/054384, WO2012/016184, WO2009/086558, WO2010/042877, WO2011/000106, WO2011/000107, WO2005/120152, WO2011/141705, WO2013/126803, WO2006/007712, WO2011/038160, WO2005/121348, WO2011/066651, WO2009/127060, WO2011/141704, WO2006/069782, WO2012/031043, WO2013/006825, WO2013/033563, WO2013/089151, WO2017/099823, WO2015/095346, 및 WO2013/086354, 및 미국 특허 공보 US2016/0311759, US2015/0376115, US2016/0151284, US2017/0210697, US2015/0140070, US2013/0178541, US2013/0303587, US2015/0141678, US2015/0239926, US2016/0376224, US2017/0119904, US2012/0149894, US2015/0057373, US2013/0090372, US2013/0274523, US2013/0274504, US2013/0274504, US2009/0023673, US2012/0128760, US2010/0324120, US2014/0200257, US2015/0203446, US2018/0005363, US2014/0308304, US2013/0338210, US2012/0101148, US2012/0027796, US2012/0058144, US2013/0323269, US2011/0117125, US2011/0256175, US2012/0202871, US2011/0076335, US2006/0083780, US2013/0123338, US2015/0064242, US2006/0051405, US2013/0065939, US2006/0008910, US2003/0022649, US2010/0130588, US2013/0116307, US2010/0062967, US2013/0202684, US2014/0141070, US2014/0255472, US2014/0039032, US2018/0028664, US2016/0317458, 및 US2013/0195920 (이들 내용은 모두 그 전문이 본원에 참고로 포함됨)에 기재되어 있다.
일부 구현예에서, 이온화 가능한 지질은 다음 구조를 갖는 MC3 (6Z,9Z,28Z,31Z)-헵타트리아콘타-6,9,28,31-테트라엔-19-일-4-(디메틸아미노) 부타노에이트 (DLin-MC3-DMA 또는 MC3)이다.
지질 DLin-MC3-DMA는 Jayaraman et al., Angew. Chem. Int. Ed Engl. (2012), 51(34): 8529-8533에 기재되어 있으며, 이의 내용은 그 전문이 본원에 참고로 포함된다.
일부 구현예에서, 이온화 가능한 지질은 다음 구조를 갖는 지질 ATX-002이다:
지질 ATX-002는 WO2015/074085에 기재되어 있으며, 이의 내용은 그 전문이 본원에 참고로 포함된다.
일부 구현예에서, 이온화 가능한 지질은 다음 구조를 갖는 (13Z,16Z)-N,N-- 디메틸-3-노닐도코사-13,16-디엔-1-아민 (화합물 32)이다:
화합물 32는 WO2012/040184에 기재되어 있으며, 이의 내용은 그 전문이 본원에 참고로 포함된다.
일부 구현예에서, 이온 가능한 지질은 다음 구조를 갖는 화합물 6 또는 화합물 22이다:
화합물 6 및 22는 WO2015/199952에 기재되어 있으며, 이의 내용은 그 전문이 본원에 참고로 포함된다.
제한 없이, 이온화 가능한 지질은 지질 나노입자에 존재하는 총 지질의 20-90% (mol)를 포함할 수 있다. 예를 들어, 이온화 가능한 지질 몰 함량은 지질 나노입자에 존재하는 총 지질의 20-70% (mol), 30-60% (mol) 또는 40-50% (mol)일 수 있다. 일부 구현예에서, 이온화 가능한 지질은 지질 나노입자에 존재하는 총 지질의 약 50 mol% 내지 약 90 mol%를 포함한다.
일부 측면에서, 지질 나노입자는 비-양이온성 지질을 추가로 포함할 수 있다. 비이온성 지질은 양친매성 지질, 중성 지질 및 음이온성 지질을 포함한다. 따라서, 비-양이온성 지질은 중성, 하전된, 양쪽이온성 또는 음이온성 지질일 수 있다. 비양이온성 지질은 전형적으로 융합유도성을 향상시키기 위해 사용된다.
예시적인 비-양이온성 지질은 디스테아로일-sn-글리세로-포스포에탄올아민, 디스테아로일포스파티딜콜린 (DSPC), 디올레오일포스파티딜콜린 (DOPC), 디팔미토일포스파티딜콜린 (DPPC), 디올레오일포스파티딜글리세롤 (DOPG), 디팔미토일포스파티딜글리세롤 (DPPG), 디올레오일-포스파티딜에탄올아민 (DOPE), 팔미토일로레오일포스파티딜콜린 (POPC), 팔미토일로레오일포스파티딜에탄올아민 (POPE), 디올레오일-포스파티딜에탄올아민 4-(N-말레이미도메틸)-사이클로헥산-1-카복실레이트 (DOPE-mal), 디팔미토일 포스파티딜 에탄올아민 (DPPE), 디미리스토일포스포에탄올아민 (DMPE), 디스테아로일-포스파티딜-에탄올아민 (DSPE), 모노메틸-포스파티딜에탄올아민 (예컨대 16-O-모노메틸 PE), 디메틸-포스파티딜에탄올아민 (예컨대 16-O-디메틸 PE), 18-1-트랜스 PE, 1-스테아로일-2-올레오일-포스파티딜에탄올아민 (SOPE), 수소화된 대두 포스파티딜콜린 (HSPC), 계란 포스파티딜콜린 (EPC), 디올레오일포스파티딜세린 (DOPS), 스핑고미엘린 (SM), 디미리스토일 포스파티딜콜린 (DMPC), 디미리스토일 포스파티딜글리세롤 (DMPG), 디스테아로일포스파티딜글리세롤 (DSPG), 디에루코일포스파티딜콜린 (DEPC), 팔미토일로레이올포스파티딜글리세롤 (POPG), 디엘라이도일-포스파티딜에탄올아민 (DEPE), 레시틴, 포스파티딜에탄올아민, 리소레시틴, 리소포스파티딜에탄올아민, 포스파티딜세린, 포스파티딜이노시톨, 스핑고미엘린, 계란 스핑고미엘린 (ESM), 세팔린, 카디오리핀, 포스파티드산, 세레브로사이드, 디세틸포스페이트, 리소포스파티딜콜린, 딜리놀레오일포스파티딜콜린, 또는 이들의 혼합물을 포함하지만, 이에 제한되지 않는다. 또한, 다른 디아실포스파티딜콜린 및 디아실포스파티딜에탄올아민 인지질도 사용될 수 있음이 이해된다. 이들 지질 중의 아실 기는 바람직하게는 C10-C24 탄소 사슬을 갖는 지방산, 예를 들어, 라우로일, 미리스토일, 팔미토일, 스테아로일, 또는 올레오일로부터 유도된 아실기이다.
지질 나노입자에 사용하기에 적합한 비-양이온성 지질의 다른 예는 비인성(nonphosphorous) 지질, 예컨대 스테아릴아민, 도데실아민, 헥사데실아민, 아세틸 팔미테이트, 글리세롤리시놀레에이트, 헥사데실 스테레에이트, 이소프로필 미리스테이트, 양쪽성 아크릴 폴리머, 트리에탄올아민-라우릴 설페이트, 알킬-아릴 설페이트 폴리에틸옥실화 지방산 아미드, 디옥타데실디메틸 암모늄 브로마이드, 세라마이드, 스핑고미엘린 등을 포함한다.
일부 구현예에서, 비-양이온성 지질은 인지질이다. 일부 구현예에서, 비-양이온성 지질은 DSPC, DPPC, DMPC, DOPC, POPC, DOPE 및 SM으로부터 선택된다. 일부 바람직일 구현예에서, 비-양이온성 지질은 DPSC이다.
예시적인 비-양이온성 지질은 PCT 공보 WO2017/099823 및 미국 특허 공보 US2018/0028664에 기재되어 있으며, 이들의 내용은 모두 그 전문이 본원에 참고로 포함된다. 일부 예에서, 비-양이온성 지질은 US2018/0028664 (그 내용은 그 전문이 본원에 참고로 포함됨)에 정의된 바와 같이, 올레산 또는 식 (I),
, 식 (II)
또는 식 (IV),
의 화합물이다.
비-양이온성 지질은 지질 나노입자에 존재하는 총 지질의 0-30% (mol)를 포함할 수 있다. 예를 들어, 비-양이온성 지질 함량은 지질 나노입자에 존재하는 총 지질의 5-20% (mol) 또는 10-15% (mol)이다. 다양일 구현예에서, 이온화 가능한 지질 대 중성 지질의 몰비는 약 2:1 내지 약 8:1의 범위이다.
일부 구현예에서, 지질 나노입자는 임의의 인지질을 포함하지 않는다.
일부 측면에서, 지질 나노입자는 막 완전성을 제공하기 위해 스테롤과 같은 성분을 추가로 포함할 수 있다.
지질 나노입자에 사용될 수 있는 하나의 예시적인 스테롤은 콜레스테롤 및 이의 유도체이다. 콜레스테롤 유도체의 비제한적인 예는 극성 유사체 예컨대 5α-콜레스탄올, 5β-코프로스탄올, 콜레스테릴-(2'-하이드록시)-에틸 에테르, 콜레스테릴-(4'-하이드록시)-부틸 에테르, 및 6-케토콜레스탄올; 비극성 유사체 예컨대 5α-콜레스탄, 콜레스테논, 5α-콜레스타논, 5β-콜레스타논, 및 콜레스테릴 데카노에이트; 및 이들의 혼합물을 포함한다. 일부 구현예에서, 콜레스테롤 유도체는 콜레스테릴-(4'-하이드록시)-부틸 에테르와 같은 극성 유사체이다.
예시적인 콜레스테롤 유도체는 PCT 공보 WO2009/127060 및 미국 특허 공보 US2010/0130588에 기재되어 있으며, 이들의 내용은 모두 그 전문이 본원에 참고로 포함된다.
스테롤과 같은 막 완전성을 제공하는 성분은 지질 나노입자에 존재하는 총 지질의 0-50% (mol)를 포함할 수 있다. 일부 구현예에서, 이러한 성분은 지질 나노입자의 총 지질 함량의 20-50% (mol) 30-40% (mol)이다.
일부 측면에서, 지질 나노입자는 폴리에틸렌 글리콜 (PEG) 또는 공액 지질 분자를 추가로 포함할 수 있다. 일반적으로, 이들은 지질 나노입자의 응집을 억제하고/하거나 입체 안정화를 제공하는데 사용된다. 예시적인 공액 지질은 PEG-지질 접합체, 폴리옥사졸린 (POZ)-지질 접합체, 폴리아미드-지질 접합체 (예컨대 ATTA-지질 접합체) , 양이온-폴리머 지질 (CPL) 접합체, 및 이들의 혼합물을 포함하지만, 이에 제한되지 않는다. 일부 구현예에서, 접합된 지질 분자는 PEG-지질 접합체, 예를 들어 (메톡시 폴리에틸렌 글리콜)-접합된 지질이다.
예시적인 PEG-지질 접합체는 PEG-디아실글리세롤 (DAG) (예컨대 1-(모노메톡시-폴리에틸렌글리콜)-2,3-디미리스토일글리세롤 (PEG-DMG)), PEG-디알킬옥시프로필 (DAA), PEG-인지질, PEG-세라마이드 (Cer), 페길화된 포스파티딜에탄올로아민 (PEG-PE), PEG 석시네이트 디아실글리세롤 (PEGS-DAG) (예컨대 4-O-(2',3'-디(테트라데카노일옥시)프로필-1-O-(w-메톡시(폴리에톡시)에틸) 부탄디오에이트 (PEG-S-DMG)), PEG 디알콕시프로필카밤, N-(카보닐-메톡시폴리에틸렌 글리콜 2000)-1,2-디스테아로일-sn-글리세로-3-포스포에탄올아민 나트륨염, 또는 이들의 혼합물을 포함하지만, 이에 제한되지는 않는다. 추가의 예시적인 PEG-지질 접합체는 US5,885,613, US6,287,591, US2003/0077829, US2003/0077829, US2005/0175682, US2008/0020058, US2011/0117125, US2010/0130588, US2016/0376224, 및 US2017/0119904에 기재되어 있으며, 이들의 내용은 그 전문이 본원에 참고로 포함된다.
일부 구현예에서, PEG- 지질은 US2018/0028664(이의 내용은 그 전문이 본원에 참고로 포함됨)에 정의된 바와 같은, 식 (III),
, 식 (III-a-I),
, 식 (III-a-2),
, 식 (III-b-1),
, 식 (III-b-2),
, 또는 식 (V),
의 화합물이다.
일부 구현예에서, US20150376115 또는 US2016/0376224 (이들 내용은 모두 그 전문이 본원에 참고로 포함됨)에 정의된 바와 같은, 식 (II),
의 PEG-지질이다.
PEG-DAA 접합체는, 예를 들어, PEG-디라우릴옥시프로필, PEG-디미리스틸옥시프로필, PEG-디팔미틸옥시프로필 또는 PEG-디스테아릴옥시프로필일 수 있다. PEG-지질은 PEG-DMG, PEG-디라우릴글리세롤, PEG-디팔미토일글리세롤, PEG-디스테릴글리세롤, PEG-디라우릴글리카미드, PEG-디미리스틸글리카미드, PEG-디팔미토일글리카미드, PEG-디스테릴글리카미드, PEG-콜레스테롤 (1-[8'-(콜레스트-5-엔-3[베타]-옥시)카복사미도-3',6'-디옥사옥타닐] 카바모일-[오메가]-메틸-폴리(에틸렌 글리콜), PEG-DMB (3,4-디테트라데콕실벤질- [오메가]-메틸-폴리(에틸렌 글리콜) 에테르), 및 1,2-디미리스토일-sn-글리세로-3-포스포에탄올아민-N-[메톡시(폴리에틸렌 글리콜)-2000],
,
,
및
중 하나 이상일 수 있다.
PEG 이외의 분자와 접합된 지질도 PEG-지질 대신 사용될 수 있다. 예를 들어, 폴리옥사졸린 (POZ)-지질 접합체, 폴리아미드-지질 접합체 (예컨대 ATTA-지질 접합체), 및 양이온성-폴리머 지질 (CPL) 접합체가 PEG-지질 대신에 또는 이에 더하여 사용될 수 있다.
예시적인 접합된 지질, 즉, PEG-지질, (POZ)-지질 접합체, ATTA-지질 접합체 및 양이온성 폴리머-지질은 PCT 특허 출원 공보 WO1996/010392, WO1998/051278, WO2002/087541, WO2005/026372, WO2008/147438, WO2009/086558, WO2012/000104, WO2017/117528, WO2017/099823, WO2015/199952, WO2017/004143, WO2015/095346, WO2012/000104, WO2012/000104, 및 WO2010/006282, 미국 특허 출원 공보 US2003/0077829, US2005/0175682, US2008/0020058, US2011/0117125, US2013/0303587, US2018/0028664, US2015/0376115, US2016/0376224, US2016/0317458, US2013/0303587, US2013/0303587, 및 US20110123453 및 미국 특허 US5,885,613, US6,287,591, US6,320,017, 및 US6,586,559 (이들 내용은 모두 그 전문이 본원에 참고로 포함됨)에 기재되어 있다.
PEG 또는 접합된 지질은 지질 나노입자에 존재하는 총 지질의 0-20% (mol)를 포함할 수 있다. 일부 구현예에서, PEG 또는 접합된 지질 함량은 지질 나노입자에 존재하는 총 지질의 0.5-10% 또는 2-5% (mol)이다.
이온화 가능한 지질, 비-양이온성 지질, 스테롤 및 PEG/공액 지질의 몰비는 필요에 따라 변할 수 있다. 예를 들어, 지질 입자는 조성물의 몰 또는 총 중량 기준으로 30-70% 이온화 가능한 지질, 조성물의 몰 또는 총 중량 기준으로 0-60% 콜레스테롤, 조성물의 몰 또는 총 중량 기준으로 0-30%의 비-양이온성-지질 및 조성물의 몰 또는 총 중량 기준으로 1-10% 공액 지질을 포함할 수 있다. 바람직하게는, 조성물은 조성물의 몰 또는 총 중량 기준으로 30-40% 이온화 가능한 지질, 조성물의 몰 또는 총 중량 기준으로 40-50% 콜레스테롤, 및 조성물의 몰 또는 총 중량 기준으로 10-20% 비-양이온성 지질을 포함한다. 일부 다른 구현예에서, 조성물은 조성물의 몰 또는 총 중량 기준으로 50-75% 이온화 가능한 지질, 조성물의 몰 또는 총 중량 기준으로 20-40%의 콜레스테롤, 및 조성물의 몰 또는 총 중량 기준으로 5 내지 10% 비-양이온성 지질, 및 조성물의 몰 또는 총 중량 기준으로 1-10% 공액 지질이다. 조성물은 조성물의 몰 또는 총 중량 기준으로 60-70% 이온화 가능한 지질, 조성물의 몰 또는 총 중량 기준으로 25-35% 콜레스테롤, 및 조성물의 몰 또는 총 중량 기준으로 5-10% 비-양이온성-지질을 함유할 수 있다. 조성물은 또한 조성물의 몰 또는 총 중량 기준으로 최대 90%의 이온화 가능한 지질 및 조성물의 몰 또는 총 중량 기준으로 2 내지 15%의 비-양이온성 지질을 함유할 수 있다. 제형은 또한, 예를 들어, 조성물의 몰 또는 총 중량 기준으로 8-30% 이온화 가능한 지질, 조성물의 몰 또는 총 중량 기준으로 5-30% 비-양이온성 지질, 및 조성물의 몰 또는 총 중량 기준으로 0-20% 콜레스테롤; 몰 또는 조성물의 총 중량 기준으로 4-25% 이온화 가능한 지질, 몰 또는 조성물의 총 중량 기준으로 4-25% 비-양이온성 지질, 몰 또는 조성물의 총 중량 기준으로 2 내지 25% 콜레스테롤, 몰 또는 조성물의 총 중량 기준으로 10 내지 35% 공액 지질, 및 몰 또는 조성물의 총 중량 기준으로 5%의 콜레스테롤; 또는 몰 또는 조성물의 총 중량 기준으로 2-30% 이온화 가능한 지질, 몰 또는 조성물의 총 중량 기준으로 2-30%의 비-양이온성 지질, 몰 또는 조성물의 총 중량 기준으로 1 내지 15%의 콜레스테롤, 몰 또는 조성물의 총 중량 기준으로 2 내지 35%의 공액 지질, 및 몰 또는 조성물의 총 중량 기준으로 1-20%의 콜레스테롤; 또는 심지어 몰 또는 조성물의 총 중량 기준으로 최대 90%의 이온화 가능한 지질 및 몰 또는 조성물의 총 중량 기준으로 2-10%의 비-양이온성 지질, 또는 심지어 몰 또는 조성물의 총 중량 기준으로 100% 양이온성 지질일 수 있다. 일부 구현예에서, 지질 입자 제형은 이온화 가능한 지질, 인지질, 콜레스테롤 및 PEG화 지질을 50:10:38.5:1.5의 몰비로 포함한다. 일부 다른 구현예에서, 지질 입자 제형은 이온화 가능한 지질, 콜레스테롤 및 PEG화 지질을 60:38.5:1.5의 몰비로 포함한다.
일부 구현예에서, 지질 입자는 이온화 가능한 지질, 비-양이온성 지질 (예를 들어 인지질) 스테롤 (예를 들면, 콜레스테롤) 및 PEG화 지질을 포함하며, 여기서 지질의 몰비는 이온화 가능한 지질에 대해 20 내지 70 몰 퍼센트이고, 목표는 40-60이고, 비-양이온성 지질은 0 내지 30의 범위이고, 목표는 0 내지 15이며, 스테롤의 몰%는 20 내지 70의 범위이고, 목표는 30 내지 50이고, PEG화 지질의 몰 퍼센트는 1 내지 6의 범위이고, 목표는 2 내지 5이다.
ceDNA를 포함하는 지질 나노입자 (LNP)는 2018년 9월 7일자로 출원된 국제 출원 PCT/US2018/050042에 개시되어 있으며, 이는 그 전문이 본원에 포함되고, 본원에 개시된 바와 같은 방법 및 조성물에서 사용되도록 구상된다.
지질 나노입자 입자 크기는 Malvern Zetasizer Nano ZS (Malvern, UK)를 사용하여 준탄성 광 산란에 의해 결정될 수 있으며, 대략 50-150 nm 직경, 대략 55-95 nm 직경, 또는 대략 70-90 nm 직경이다.
제형화된 양이온성 지질의 pKa는 핵산의 전달을 위한 LNP의 효과와 상관될 수 있다 (Jayaraman et al, Angewandte Chemie, International Edition (2012), 51(34), 8529-8533; Semple et al, Nature Biotechnology 28, 172-176 (2010) 참조, 둘 다 그 전문이 참고로 포함됨). pKa의 바람직한 범위는 약 5 내지 약 7이다. 각각의 양이온성 지질의 pKa는 2-(p-톨루이디노)-6-나프탈렌 설폰산 (TNS)의 형광에 기초한 분석을 사용하여 지질 나노입자에서 결정된다. 0.4 mM의 총 지질 농도의 PBS 중의 양이온성 지질/DSPC/콜레스테롤/PEG-지질 (50/10/38.5/1.5 mol%)을 포함하는 지질 나노입자는 본원 및 다른 곳에 기재된 인라인 공정을 사용하여 제조될 수 있다. TNS는 증류수에서 100 μM 스톡 용액으로 준비될 수 있다. 소포는 10 mM HEPES, 10 mM MES, 10 mM 암모늄 아세테이트, 130 mM NaCl을 함유하는 2 mL의 완충 용액에서 24 μM 지질로 희석될 수 있으며, 여기서 pH는 2.5 내지 11의 범위이다. TNS 용액의 분취액을 첨가하여 1 μM의 최종 농도를 제공할 수 있으며, 와동 혼합 후 형광 강도는 321 nm 및 445 nm의 여기 및 방출 파장을 사용하여 SLM Aminco 시리즈 2 발광 분광광도계에서 실온에서 측정된다. 시그모이달 최적합(sigmoidal best fit) 분석을 형광 데이터에 적용할 수 있으며, pKa는 반-최대 형광 강도를 발생시키는 pH로 측정된다.
꼬리 정맥 주사를 통한 투여 4시간 후 간에서의 루시퍼라제 발현을 측정함으로써 상대 활성을 결정할 수 있다. 활성은 0.3 및 1.0 mg ceDNA/kg의 용량에서 비교되고 투여 4 시간 후에 측정된 루시퍼라제/g 간으로 표현된다.
제한 없이, 본 발명의 지질 나노입자는 캡시드가 없는 비-바이러스성 DNA 벡터를 관심 표적 부위 (예를 들어, 세포, 조직, 기관 등)로 전달하는데 사용될 수 있는 지질 제형을 포함한다. 일반적으로, 지질 나노입자는 캡시드가 없는, 비-바이러스성 DNA 벡터 및 이온화 가능한 지질 또는 이의 염을 포함한다.
일부 구현예에서, 지질 입자는 이온화 가능한 지질/비-양이온성-지질/스테롤/공액 지질을 50:10:38.5:1.5의 몰비로 포함한다.
다른 측면에서, 본 개시내용은 인지질, 레시틴, 포스파티딜콜린 및 포스파티딜에탄올아민을 포함하는 지질 나노입자 제형을 제공한다.
일부 구현예에서, 하나 이상의 추가 화합물이 또한 포함될 수 있다. 이들 화합물은 별도로 투여될 수 있거나 추가의 화합물이 본 발명의 지질 나노입자에 포함될 수 있다. 다시 말해서, 지질 나노입자는 ceDNA 이외의 다른 화합물 또는 제1 ceDNA와는 다른 적어도 제2 ceDNA를 함유할 수 있다. 제한 없이, 다른 추가 화합물은 소형 또는 대형 유기 또는 무기 분자, 단당류, 이당류, 삼당류, 올리고당류, 다당류, 펩티드, 단백질, 펩티드 유사체 및 이의 유도체, 펩티도미메틱, 핵산, 핵산 유사체 및 유도체, 생물학적 물질로 만들어진 추출물, 또는 이들의 임의의 조합으로 이루어진 군으로부터 선택될 수 있다.
일부 구현예에서, 하나 이상의 추가 화합물은 치료제일 수 있다. 치료제는 치료 목적에 적합한 임의의 부류로부터 선택될 수 있다. 다시 말해, 치료제는 치료 목적에 적합한 임의의 부류로부터 선택될 수 있다. 즉, 치료제는 원하는 치료 목적 및 생물학적 작용에 따라 선택될 수 있다. 예를 들어, LNP 내의 ceDNA가 암 치료에 유용한 경우, 추가의 화합물은 항암제 (예를 들어, 화학요법제, 표적화된 암 요법 (소분자, 항체 또는 항체-약물 접합체를 포함하지만, 이에 제한되지 않음)일 수 있다. 또 다른 예에서, ceDNA를 함유하는 LNP가 감염 치료에 유용한 경우, 추가의 화합물은 항균제 (예를 들어, 항생제 또는 항바이러스 화합물)일 수 있다. 또 다른 예에서, ceDNA를 함유하는 LNP가 면역 질환 또는 장애의 치료에 유용한 경우, 추가 화합물은 면역 반응을 조절하는 화합물 (예를 들어, 면역억제제, 면역자극 화합물, 또는 하나 이상의 특정 면역 경로를 조절하는 화합물일 수 있다. 일부 구현예에서, 상이한 단백질을 인코딩하는 ceDNA와 같은 상이한 화합물, 또는 치료제와 같은 상이한 화합물을 함유하는 상이한 지질 나노입자의 상이한 칵테일이 본 발명의 조성물 및 방법에 사용될 수 있다.
일부 구현예에서, 추가의 화합물은 면역 조절제이다. 예를 들어, 추가 화합물은 면역억제제이다. 일부 구현예에서, 추가의 화합물은 면역자극성이다.
지질 나노입자 및 약제학적으로 허용되는 담체 또는 부형제를 포함하는 약제학적 조성물이 또한 본원에 제공된다.
일부 측면에서, 본 개시내용은 하나 이상의 약제학적 부형제를 추가로 포함하는 지질 나노입자 제형을 제공한다. 일부 구현예에서, 지질 나노입자 제형은 수크로스, 트리스, 트레할로스 및/또는 글리신을 추가로 포함한다.
일반적으로, 본 발명의 지질 나노입자는 의도된 치료 효과를 제공하기 위해 선택된 평균 직경을 갖는다. 따라서, 일부 측면에서, 지질 나노입자는 약 30 nm 내지 약 150 nm, 보다 전형적으로 약 50 nm 내지 약 150 nm, 보다 전형적으로 약 60 nm 내지 약 130 nm, 보다 전형적으로 약 70 nm 내지 약 110 nm, 가장 일반적으로 약 85 nm 내지 약 105nm, 및 바람직하게는 약 100 nm의 평균 직경을 갖는다. 일부 측면에서, 본 개시내용은 일반적인 나노입자에 비해 크기가 크고 크기가 약 150 내지 250 nm인 지질 입자를 제공한다. 지질 나노입자 입자 크기는, 예를 들어, Malvern Zetasizer Nano ZS (Malvern, UK) 시스템을 사용하는 준탄성 광 산란에 의해 결정될 수 있다.
지질 입자의 의도된 용도에 따라, 성분의 비율이 변할 수 있고, 특정 제형의 전달 효율은, 예를 들어, 엔도좀 방출 파라미터 (ERP) 분석을 사용하여 측정될 수 있다.
ceDNA는 입자의 지질 부분과 복합체화되거나 지질 나노입자의 지질 부분에서 캡슐화될 수 있다. 일부 구현예에서, ceDNA는 지질 나노입자의 지질 위치에서 완전히 캡슐화될 수 있으며, 이에 의해, 예를 들어, 수용액에서 뉴클레아제에 의한 분해로부터 보호할 수 있다. 일부 구현예에서, 지질 나노입자 중의 ceDNA는 지질 나노입자를 37℃에서 적어도 약 20, 30, 45 또는 60분 동안 뉴클레아제에 노출시킨 후 실질적으로 분해되지 않는다. 일부 구현예에서, 지질 나노입자의 ceDNA는 37℃에서 적어도 약 30, 45 또는 60분 또는 적어도 약 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34 또는 36시간 동안 혈청에서 입자의 인큐베이션 후 실질적으로 분해되지 않는다.
특정 구현예에서, 지질 나노입자는 대상체, 예를 들어, 인간과 같은 포유동물에 실질적으로 비-독성이다.
일부 측면에서, 지질 나노입자 제형은 동결 건조된 분말이다.
일부 구현예에서, 지질 나노입자는 적어도 하나의 지질 이중층을 갖는 고체 코어 입자이다. 다른 구현예에서, 지질 나노입자는 비-이중층 구조, 즉, 비-라멜라 (즉, 비-이중층) 형태를 갖는다. 제한 없이, 비-이중층 형태는, 예를 들어, 3차원 튜브, 막대, 입방체 대칭 등을 포함할 수 있다. 지질 입자의 비-라멜라 형태 (즉, 비-이중층 구조)는 당업자에게 공지되고 사용되는 분석 기술을 사용하여 결정될 수 있다. 이러한 기술은 동결 투과 전자 현미경 ("Cryo-TEM"), 시차 주사 열량 측정 ("DSC"), X-선 회절 등을 포함하지만, 이에 제한되지는 않는다. 예를 들어, 지질 나노입자의 형태 (라멜라 대 비-라멜라)는, 예를 들어, US2010/0130588 (이의 내용은 그 전문이 본원에 참조로 포함됨)에 기재된 바와 같은 Cryo-TEM 분석을 사용하여 쉽게 평가되고 특성화될 수 있다.
일부 추가의 구현예에서, 비-라멜라 형태를 갖는 지질 나노입자는 고전자밀도이다.
일부 측면에서, 본 개시내용은 구조상 단일 라멜라 또는 다중 라멜라인 지질 나노입자를 제공한다. 일부 측면에서, 본 개시내용은 다중-소포성 입자 및/또는 발포체-기반 입자를 포함하는 지질 나노입자 제형을 제공한다.
지질 성분의 조성 및 농도를 제어함으로써, 지질 접합체가 지질 입자로부터 교환되는 속도, 및 이어서 지질 나노입자가 융합유도성이 되는 속도를 제어할 수 있다. 또한, 예를 들어, pH, 온도 또는 이온 강도를 포함하는 다른 변수를 사용하여 지질 나노입자가 융합유도성이 되는 속도를 변화 및/또는 제어할 수 있다. 지질 나노입자가 융합유도성이 되는 속도를 제어하는데 사용될 수 있는 다른 방법은 본 개시내용에 기초하여 당업자에게 명백할 것이다. 지질 접합체의 조성 및 농도를 제어함으로써 지질 입자 크기를 제어할 수 있음이 또한 명백할 것이다.
제형화된 양이온성 지질의 pKa는 핵산의 전달을 위한 LNP의 효과와 상관관계가 있을 수 있다 (Jayaraman et al, Angewandte Chemie, International Edition (2012), 51(34), 8529-8533; Semple et al, Nature Biotechnology 28, 172-176 (20l0) 참조, 둘 다 그 전문이 참고로 포함됨). pKa의 바람직한 범위는 약 5 내지 약 7이다. 양이온성 지질의 pKa는 2-(p-톨루이디노)-6-나프탈렌 설폰산 (TNS)의 형광에 기초한 검정을 사용하여 지질 나노입자에서 측정될 수 있다.
지질 입자에서 ceDNA의 캡슐화는 Oligreen® 분석 또는 PicoGreen® 분석과 같이 핵산과 관련될 때 형광이 강화된 염료를 사용하는, 막-불침투성 형광 염료 배제 분석을 수행하여 확인될 수 있다. 일반적으로, 캡슐화는 염료를 지질 입자 제형에 첨가하고, 생성된 형광을 측정하고, 소량의 비이온성 세제를 첨가할 때 관찰된 형광과 비교함으로써 결정된다. 지질 이중층의 세제-매개 파괴는 캡슐화된 ceDNA를 방출하여 막-불투과성 염료와 상호작용할 수 있게 한다. ceDNA의 캡슐화는 다음과 같이 계산될 수 있으며: E= (I0 - I)/I0, 여기서 I 및 I0는 세제를 첨가하기 전후에 형광 강도를 나타낸다.
IX. ceDNA 벡터를 전달하는 방법
일부 구현예에서, ceDNA 벡터는 다양한 적합한 방법에 의해 시험관내 또는 생체내 표적 세포로 전달될 수 있다. ceDNA 벡터만 적용하거나 주사할 수 있다. CeDNA 벡터는 형질감염 시약 또는 다른 물리적 수단의 도움 없이 세포로 전달될 수 있다. 대안적으로, ceDNA 벡터는 세포로의 DNA의 진입을 용이하게 하는 임의의 당 업계에 공지된 형질감염 시약 또는 다른 당 업계에 공지된 물리적 수단, 예를 들어, 리포좀, 알코올, 폴리리신-풍부 화합물, 아르기닌-풍부 화합물, 인산칼슘, 미세소포, 미세주사, 전기천공 등을 사용하여 전달될 수 있다.
대조적으로, 본원에 개시된 캡시드 비함유 AAV 벡터를 이용한 형질도입은 다양한 전달 시약을 사용하여 통상적인 AAV 비리온으로 형질도입하기 어려운 세포 및 조직 유형을 효율적으로 표적화할 수 있다.
또 다른 구현예에서, ceDNA 벡터는 CNS (예를 들어, 뇌 또는 눈)에 투여된다. ceDNA 벡터는 척수, 뇌간 (숨뇌, 뇌교), 중뇌 (시상하부, 시상, 시상상부, 뇌하수체, 흑질, 송과선), 소뇌, 종뇌(선조체, 후두를 포함한 대뇌, 측두엽, 두정엽, 전두엽, 피질, 기저핵, 해마 및 포르타아미달라(portaamygdala)), 변연계, 신피질, 선조체, 대뇌 및 하구로 도입될 수 있다. ceDNA 벡터는 또한 망막, 각막 및/또는 시신경과 같은 눈의 다른 영역에 투여될 수 있다. ceDNA 벡터는 뇌척수액 내로 전달될 수 있다 (예를 들어, 요추 천자에 의해). ceDNA 벡터는 혈액-뇌 장벽이 교란된 상황 (예를 들어, 뇌종양 또는 뇌경색)에서 CNS에 혈관내로 추가로 투여될 수 있다.
일부 구현예에서, ceDNA 벡터는 경막내, 안구내, 뇌내, 심실내, 정맥내 (예를 들어, 만니톨과 같은 당의 존재하에), 비강내, 귀내, 안구내 (예를 들어, 유리체내, 망막하, 전방) 및 안구주위 (예를 들어, 서브-테논 부위(sub-Tenon's region)) 전달뿐만 아니라 운동 뉴런으로 역행 전달을 통한 근육내 전달을 포함하지만 이에 제한되지 않는, 당 업계에 공지된 임의의 경로에 의해 CNS의 원하는 영역(들)에 투여될 수 있다.
일부 구현예에서, ceDNA 벡터는 CNS의 원하는 영역 또는 구획에 직접 주사 (예를 들어, 정위 주사)에 의해 액체 제형으로 투여된다. 다른 구현예에서, ceDNA 벡터는 원하는 영역에 국소 적용하거나 에어로졸 제형의 비강내 투여에 의해 제공될 수 있다. 눈에의 투여는 액체 액적의 국소 적용에 의한 것일 수 있다. 추가의 대안으로서, ceDNA 벡터는 고체, 서방형 제형으로서 투여될 수 있다 (예를 들어, 미국 특허 번호 7,201,898 참조). 또 다른 추가의 구현예에서, ceDNA 벡터는 운동 뉴런 (예를 들어, 근위축성 측삭 경화증 (ALS); 척수 근육 위축 (SMA) 등)을 포함하는 질환 및 장애를 치료, 개선 및/또는 예방하기 위해 역행 수송에 사용될 수 있다. 예를 들어, ceDNA 벡터는 뉴런으로 이동할 수 있는 근육 조직으로 전달될 수 있다.
X. ceDNA 벡터의 추가 용도
본원에 제공된 조성물 및 ceDNA 벡터는 다양한 목적을 위해 표적 유전자를 유전자 편집하는데 사용될 수 있다. 일부 구현예에서, 생성된 이식유전자는 연구 목적, 예를 들어, 이식유전자 생성물의 기능을 연구하기 위한, 예를 들어, 이식유전자를 보유한 체세포성 형질전환 동물 모델을 생성하기 위해 사용되는 것으로 의도된 단백질 또는 기능성 RNA를 인코딩한다. 또 다른 예에서, 이식유전자는 질환의 동물 모델을 생성하기 위해 사용되는 것으로 의도된 단백질 또는 기능성 RNA를 인코딩한다. 일부 구현예에서, 생성된 이식유전자는 포유동물 대상체에서 질환 상태 또는 장애의 치료, 예방 또는 개선에 유용한 하나 이상의 펩티드, 폴리펩티드 또는 단백질을 인코딩한다. 생성된 이식유전자는 유전자의 발현 감소, 발현 부족 또는 기능 장애와 관련된 질환을 치료하기에 충분한 양으로 대상체에게 전달될 수 있다 (예를 들어, 발현될 수 있다). 일부 구현예에서, 생성된 이식유전자는 증가된 발현, 유전자 산물의 활성, 또는 생성된 이식유전자이 발현을 억제하거나 달리 발현이 감소되도록 하는 유전자의 부적절한 상향 조절과 관련된 질환을 치료하기에 충분한 양으로 대상체에서 발현될 수 있다. 또 다른 구현예에서, 생성된 이식유전자는 천연 유전자의 결함 카피를 대체하거나 보충한다. 이식유전자는 그 자체로 전사될 유전자의 오픈 리딩 프레임이 아닐 수 있고; 대신에, 이는 표적 유전자의 프로모터 영역 또는 억제 영역일 수 있고, ceDNA 유전자 편집 벡터는 관심 유전자의 발현을 조절하는 결과로 이러한 영역을 변형시킬 수 있음을 당업자는 이해할 것이다.
일부 구현예에서, 이식유전자는 질환의 동물 모델을 생성하기 위해 사용되는 것으로 의도된 단백질 또는 기능성 RNA를 인코딩한다. 일부 구현예에서, 이식유전자는 포유동물 대상체에서 질환 상태의 치료 또는 예방에 유용한 하나 이상의 펩티드, 폴리펩티드 또는 단백질을 인코딩한다. 이식유전자 또는 공여체 서열은 유전자의 발현 감소, 발현 부족 또는 기능 장애와 관련된 질환을 치료하기에 충분한 양으로 환자에게 전달 (예를 들어, 발현)될 수 있다. 일부 구현예에서, 이식유전자는 유전자 편집 분자 (예를 들어, 뉴클레아제)이다. 특정 구현예에서, 뉴클레아제는 CRISPR-연관 뉴클레아제 (Cas 뉴클레아제)이다.
XI. 사용 방법
본원에 개시된 바와 같은 유전자 편집용 ceDNA 벡터는 또한 관심 뉴클레오티드 서열 (예를 들어, 유전자 편집 분자, 예를 들어, 뉴클레아제 또는 가이드 서열)을 표적 세포 (예를 들어, 숙주 세포)로 전달하는 방법에 사용될 수 있다. 상기 방법은 특히 유전자 편집 분자를 이를 필요로 하는 대상체의 세포로 전달하고 관심 표적 유전자를 편집하는 방법일 수 있다. 본 발명은 유전자 편집 기구의 치료 효과가 일어나도록 유전자 편집 분자, 예를 들어, 대상체의 세포에서 ceDNA 벡터에 인코딩된 뉴클레아제 또는 가이드 서열의 생체내 발현을 허용한다. 이러한 결과는 ceDNA 벡터 전달의 생체내 및 시험관내 모드 둘 다에서 볼 수 있다.
또한, 본 발명은 상기 관심 핵산을 포함하는 본 발명의 ceDNA 벡터의 다중 투여를 포함하는, 유전자 편집 분자를 이를 필요로 하는 대상체의 세포로 전달하는 방법을 제공한다. 본 발명의 ceDNA 벡터는 캡슐화된 바이러스 벡터에 대해 전형적으로 관찰되는 것과 같은 면역 반응을 유도하지 않기 때문에, 이러한 다중 투여 전략은 ceDNA-기반 시스템에서 더 큰 성공을 거둘 것이다.
ceDNA 벡터 핵산(들)은 원하는 조직의 세포를 형질감염시키고 과도한 부작용 없이 충분한 수준의 유전자 전달 및 발현을 제공하기에 충분한 양으로 투여된다. 통상적이고 약제학적으로 허용되는 투여 경로는 정맥내 (예를 들어, 리포좀 제형으로), 선택된 기관으로의 직접 전달 (예를 들어, 간으로의 문맥내 전달), 근육내 및 기타 모 투여 경로를 포함하지만, 이에 제한되지는 않는다. 원하는 경우, 투여 경로가 조합될 수 있다.
ceDNA 전달은 유전자 편집 성분을 인코딩하는 모든 뉴클레오티드의 ceDNA 벡터 전달로 제한되지 않는다. 예를 들어, 본원에 기재된 ceDNA 벡터는 유전자 편집 성분의 일부를 제공하기 위해 제공된 다른 전달 시스템과 함께 사용될 수 있다. 본 개시냉요에 따른 ceDNA 벡터와 조합될 수 있는 시스템의 하나의 비제한적인 예는 치료 또는 유전자 편집이 필요한 숙주 세포에 Cas9를 개별적으로 전달하는 시스템을 포함한다. 특정 구현예에서, Cas9는 문헌[Lee et al., Nanoparticle delivery of Cas9 ribonucleotideprotein and donor DNA in vivo induces homology-directed DNA repair, Nature Biomedical Engineering, 2017 (그 전문이 본원에 참고로 포함됨)]에 기재된 것과 같은 나노입자로 전달될 수 있는 한편, 다른 성분, 예컨대 공여체 서열은 ceDNA에 의해 제공된다.
본 발명은 또한 선택적으로 약제학적으로 허용되는 담체와 함께, 치료적 유효량의 ceDNA 벡터를 대상체의 이를 필요로 하는 세포 (특히 근육 세포 또는 조직)에 도입하는 것을 포함하는 대상체에서 질환을 치료하는 방법을 제공한다. ceDNA 벡터는 담체의 존재하에 도입될 수 있지만, 이러한 담체는 필요하지 않다. 구현된 ceDNA 벡터는 질환을 치료하는데 유용한 관심 뉴클레오티드 서열을 포함한다. 특히, ceDNA 벡터는 대상체에 도입될 때 외인성 DNA 서열에 의해 인코딩된 원하는 폴리펩티드, 단백질 또는 올리고뉴클레오티드의 전사를 지시할 수 있는 제어 요소에 작동 가능하게 연결된 원하는 외인성 DNA 서열을 포함할 수 있다. ceDNA 벡터는 상기 제공된 임의의 적합한 경로, 및 본원의 다른 곳을 통해 투여될 수 있다.
본원에 제공된 조성물 및 벡터는 다양한 목적으로 이식유전자를 전달하는데 사용될 수 있다. 일부 구현예에서, 이식유전자는 연구 목적으로, 예를 들어, 이식유전자 생성물의 기능을 연구하기 위한, 예를 들어, 이식유전자를 보유한 체세포성 형질전환 동물 모델을 생성하기 위해 사용되는 것으로 의도된 단백질 또는 기능성 RNA를 인코딩한다. 또 다른 예에서, 이식유전자는 질환의 동물 모델을 생성하기 위해 사용되는 것으로 의도된 단백질 또는 기능성 RNA를 인코딩한다. 일부 구현예에서, 이식유전자는 포유동물 대상체에서 질환 상태의 치료 또는 예방에 유용한 하나 이상의 펩티드, 폴리펩티드 또는 단백질을 인코딩한다. 이식유전자는 유전자의 발현 감소, 발현 부족 또는 기능 장애와 관련된 질환을 치료하기에 충분한 양으로 환자에게 전달될 수 있다 (예를 들어, 발현된다). 일부 구현예에서, 이식유전자는 유전자 편집 분자 (예를 들어, 뉴클레아제)이다. 특정 구현예에서, 뉴클레아제는 CRISPR-연관 뉴클레아제 (Cas 뉴클레아제)이다.
원칙적으로, 발현 카세트는 돌연변이로 인해 감소되거나 부재하거나 또는 과발현될 때 치료적 이점을 전달하는 단백질 또는 폴리펩티드를 인코딩하는 임의의 유전자를 표적화하는 핵산 또는 뉴클레아제를 포함할 수 있으며, 본 발명의 범위 내에 있는 것으로 간주된다. ceDNA 벡터는 메가뉴클레아제- 또는 아연 핑거 뉴클레아제에 의해 제공되는 이중 가닥 파괴 후에 삽입되는 교정 DNA 가닥으로서 사용되는 주형 뉴클레오티드 서열을 포함한다. ceDNA 벡터는 메가뉴클레아제- 또는 아연 핑거 뉴클레아제에 의해 제공된 이중 가닥 파괴 후에 삽입되는 교정 DNA 가닥으로서 사용되는 주형 뉴클레오티드 서열을 포함할 수 있다. 바람직하게는, 비삽입된 박테리아 DNA는 존재하지 않으며, 바람직하게는 박테리아 DNA가 본원에 제공된 ceDNA 조성물에 존재하지 않는다.
유전자 편집용 ceDNA 벡터 전달은 한 종의 ceDNA 벡터로 제한되지 않는다. 이와 같이, 또 다른 측면에서, 상이한 공여체 서열 및/또는 유전자 편집 서열을 포함하는 다수의 ceDNA 벡터는 표적 세포, 조직, 기관 또는 대상체에게 동시에 또는 순차적으로 전달될 수 있다. 따라서, 이 전략을 통해 여러 유전자를 동시에 유전자 편집할 수 있다. 유전자 편집 기능의 상이한 부분을, 동시에 또는 상이한 시간에 투여될 수 있고, 개별적으로 조절할 수 있는 별도의 ceDNA 벡터로 분리할 수 있다. 바이러스성 캡시드의 부재로 인한 항-캡시드 숙주 면역 반응의 부족을 고려하면, 전달은 후속적으로 증가하거나 감소하는 용량으로, 중요하게는 임상 환경에서의 유전자 요법에 대해 여러 번 수행될 수 있다. 캡시드가 없으므로 항-캡시드 반응이 일어나지 않을 것으로 예상된다.
본 발명은 또한 선택적으로 약제학적으로 허용되는 담체와 함께, 유전자 편집을 위한 치료적 유효량의 ceDNA 벡터를 대상체의 이를 필요로 하는 표적 세포 (특히 근육 세포 또는 조직)에 도입하는 것을 포함하는 대상체에서 질환을 치료하는 방법을 제공한다. ceDNA 벡터는 담체의 존재하에 도입될 수 있지만, 이러한 담체는 필요하지 않다. 구현된 ceDNA 벡터는 질환을 치료하는데 유용한 관심 뉴클레오티드 서열을 포함한다. 특히, ceDNA 벡터는 대상체에 도입될 때 외인성 DNA 서열에 의해 인코딩된 원하는 폴리펩티드, 단백질 또는 올리고뉴클레오티드의 전사를 지시할 수 있는 제어 요소에 작동 가능하게 연결된 원하는 외인성 DNA 서열을 포함할 수 있다. ceDNA 벡터는 상기 제공된 임의의 적합한 경로, 및 본원의 다른 곳을 통해 투여될 수 있다.
XII. 치료 방법
본원에 기재된 기술은 또한, 예를 들어, 현장외(ex situ), 시험관내 및 생체내 적용, 방법론, 진단 절차 및/또는 유전자 치료 요법을 포함하는 다양한 방식으로 개시된 ceDNA 벡터를 사용하는 방법뿐만 아니라 제조 방법을 입증한다.
선택적으로 약제학적으로 허용되는 담체와 함께, 치료적 유효량의 유전자 편집 ceDNA를 대상체의 이를 필요로 하는 표적 세포 (예를 들어, 근육 세포 또는 조직, 또는 다른 병든 세포 유형)에 도입하는 것을 포함하는 대상체에서 질환 또는 장애를 치료하는 방법이 본원에 제공된다. ceDNA 벡터는 담체의 존재하에 도입될 수 있지만, 이러한 담체는 필요하지 않다. 구현된 ceDNA 벡터는 질환을 치료하는데 유용한 관심 뉴클레오티드 서열을 포함한다. 특히, ceDNA 벡터는 대상체에 도입될 때 외인성 DNA 서열에 의해 인코딩된 원하는 폴리펩티드, 단백질 또는 올리고뉴클레오티드의 전사를 지시할 수 있는 제어 요소에 작동 가능하게 연결된 원하는 외인성 DNA 서열을 포함할 수 있다. ceDNA 벡터는 상기 제공된 임의의 적합한 경로 및 본원의 다른 곳을 통해 투여될 수 있다.
본원에는 본 발명의 ceDNA 벡터 중 하나 이상을, 하나 이상의 약제학적으로 허용되는 완충제, 희석제 또는 부형제와 함께 포함하는 ceDNA 벡터 조성물 및 제형이 개시된다. 이러한 조성물은 질환, 손상, 장애, 외상 또는 기능 장애의 하나 이상의 증상을 진단, 예방, 치료 또는 개선하기 위한 하나 이상의 진단 또는 치료 키트에 포함될 수 있다. 일 측면에서, 질환, 손상, 장애, 외상 또는 기능 장애는 인간 질환, 손상, 장애, 외상 또는 기능 장애이다.
본원에 기재된 기술의 또 다른 측면은 진단적 또는 치료적 유효량의 ceDNA 벡터를 이를 필요로 하는 대상체에게 제공하는 방법을 제공하며, 상기 방법은 이를 필요로 하는 대상체의 세포, 조직 또는 기관에, 본원에 개시된 바와 같은 ceDNA 벡터의 양을; ceDNA 벡터로부터 이식유전자의 발현을 가능하게 하기에 효과적인 시간 동안 제공하여 ceDNA 벡터에 의해 발현된 진단적 또는 치료적 유효량의 단백질, 펩티드, 핵산을 대상체에게 제공하는 단계를 포함한다. 추가 측면에서, 대상체는 인간이다.
본원에 기재된 기술의 또 다른 측면은 대상체에서 질환, 장애, 기능 장애, 손상, 비정상적 상태 또는 외상의 적어도 하나 이상의 증상을 진단, 예방, 치료 또는 개선하는 방법을 제공한다. 전체적이고 일반적인 의미에서, 상기 방법은 적어도, 이를 필요로 하는 대상체에게 하나 이상의 개시된 ceDNA 벡터를, 상기 대상체의 질환, 장애, 기능 장애, 손상, 비정상적 상태 또는 외상의 하나 이상의 증상을 진단, 예방, 치료 또는 개선하기에 충분한 양으로 그리고 시간 동안 투여하는 단계를 포함한다. 추가 측면에서, 대상체는 인간이다.
또 다른 측면은 질환 또는 질환 상태의 하나 이상의 증상을 치료 또는 감소시키기 위한 도구로서 ceDNA 벡터를 사용하는 것이다. 결함이 있는 유전자가 알려져 있고, 전형적으로 다음의 두 가지 등급으로 분류되는 수많은 유전 질환이 있다: 일반적으로 열성 방식으로 유전되는 일반적으로 효소의 결핍 상태, 및 조절 또는 구조 단백질을 포함할 수 있고, 전형적으로 우세 방식으로 유전되지만 항상 그런 것은 아닌 불균형 상태. 결핍 상태 질환의 경우, ceDNA 벡터를 사용하여 이식유전자를 전달하여 대체 요법을 위해 정상 유전자를 병든 조직으로 가져오고, 또한, 일부 구현예에서, 안티센스 돌연변이를 사용하여 질환에 대한 동물 모델을 생성할 수 있다. 불균형 질환 상태의 경우, ceDNA 벡터를 사용하여 모델 시스템에서 질환 상태를 생성한 다음 질병 상태를 저지하기 위한 노력에 사용할 수 있다. 따라서, 본원에 개시된 ceDNA 벡터 및 방법은 유전자 질환의 치료를 허용한다. 본원에 사용된 바와 같이, 질환 상태는 질환을 유발하거나 더 심각하게 만드는 결핍 또는 불균형을 부분적으로 또는 전체적으로 개선함으로써 치료된다.
A. 숙주 세포:
일부 구현예에서, ceDNA 벡터는 이식유전자를 대상체 숙주 세포 내로 전달한다. 일부 구현예에서, 본 발명의 숙주 세포는, 예를 들어 혈액 세포, 줄기세포, 조혈 세포, CD34+ 세포, 간 세포, 암 세포, 혈관 세포, 근육 세포, 췌장 세포, 신경 세포, 눈 또는 망막 세포, 상피 또는 내피 세포, 수지상 세포, 섬유 모세포, 또는, 제한 없이, 간(hepatic) (즉, 간(liver)) 세포, 폐 세포, 심장 세포, 췌장 세포, 장 세포, 횡격막 세포, 신장(renal) (즉, 신장(kidney)) 세포, 신경 세포, 혈액 세포, 골수 세포, 또는 유전자 요법이 고려되는 대상체의 임의의 하나 이상의 선택된 조직을 포함하는 포유동물 기원의 임의의 다른 세포를 포함하는 인간 숙주 세포이다. 일 측면에서, 본 발명의 숙주 세포는 인간 숙주 세포이다.
본 개시내용은 또한, 본원에 기재된 ceDNA 벡터를 포함하여, 상기 언급된 바와 같은 재조합 숙주 세포에 관한 것이다. 따라서, 숙련가에게 명백한 목적에 따라 다수의 숙주 세포를 사용할 수 있다. 공여체 서열을 포함하는 작제물 또는 ceDNA 벡터가 숙주 세포 내로 도입되어 공여체 서열이 전술한 바와 같이 염색체 구성 요소로 유지된다. 용어 숙주 세포는 복제 동안 발생하는 돌연변이로 인해 모 세포와 동일하지 않은 모 세포의 임의의 자손을 포함한다. 숙주 세포의 선택은 공여체 서열 및 그 공급원에 크게 좌우될 것이다. 숙주 세포는 또한 포유동물, 곤충, 식물 또는 진균 세포와 같은 진핵생물일 수 있다. 일 구현예에서, 숙주 세포는 인간 세포 (예를 들어, 1차 세포, 줄기세포 또는 불멸화된 세포주)이다. 일부 구현예에서, 숙주 세포는 결함 유전자의 교정 또는 유전자의 발현을 제거하기 위해 편집된 유전자이다. 예를 들어, CRISPR/CAS는 NHEJ 또는 HDR 복구, 뿐만 아니라 다른 유전자 편집 시스템, 예를 들어 ZFN 또는 TALE에 의해 하나 이상의 gRNA로 게놈을 편집하는데 사용될 수 있다. 숙주 세포는 임의의 세포 유형, 예를 들어, 체세포 또는 줄기세포, 유도된 다능성 줄기세포, 또는 혈액 세포, 예를 들어, T-세포 또는 B-세포, 또는 골수 세포일 수 있다. 특정 구현예에서, 숙주 세포는 동종이계 세포이다. 예를 들어, T-세포 게놈 조작은 암 면역요법, HIV 요법과 같은 질환 조절 (예를 들어, 수용체 녹아웃, 예컨대 CXCR4 및 CCR5) 및 면역결핍 요법에 유용하다. B-세포 상의 MHC 수용체는 면역요법에 대해 표적화될 수 있다. 게놈 편집 골수 줄기세포, 예를 들어, CD34+ 세포, 또는 유도된 다능성 줄기세포는 치료 단백질의 발현을 위해 환자에게 다시 이식될 수 있다.
B. 유전자 편집 ceDNA로 치료될 예시적인 질환
ceDNA 유전자 편집 벡터는 또한 공여체 DNA의 부재하에 결함 유전자를 교정하는데 유용하며, 예를 들어 CRISPR/CAS ceDNA 시스템에서 스플라이스 수용체 또는 스플라이스 공여체를 표적으로 하는 하나의 단일 가이드 RNA는 결함 유전자의 프레임시프트 돌연변이를 교정하고, 기능성 단백질의 발현을 초래한다. 비제한적 예로서, 뒤센느 근위축증(Duchene Muscular Dystrophy)의 DMD 유전자는 단일 가이드 RNA NHEJ를 사용하여 엑손 스킵핑, 및 기능성 디스트로핀의 발현을 위해 다중 가이드 RNA를 사용하여 교정되었고, 예를 들어, US 2016/0201089를 참조하며, 이는 그 전문이 본원에 참고로 포함된다.
ceDNA 유전자 편집 벡터는 또한 유전자 발현을 제거하는데 유용하다. 예를 들어, 일 구현예에서 ceDNA 벡터를 사용하여, 예를 들어, 프레임-시프트 돌연변이를 유발함으로써 넌센스 인델 (예를 들어, 비-코딩 염기쌍의 삽입 또는 결실)이 표적 유전자의 녹다운을 유도할 수 있다. 비제한적인 예로서, HIV 수용체인, CXCR4 및 CCR5의 발현은 CAS9 RNP 및 하나 이상의 가이드 RNA를 사용하여 NHEJ 또는 HDR 경로를 유도함으로써 1차 인간 T-세포에서 성공적으로 제거되었고, Schumann et al. (2015) Generation of knock in primary human cells using Cas9 ribonucleoproteins, PNAS 112(33): 10437-10442 (그 전문이 본원에 참고로 포함됨)를 참조한다. 이 시스템은 단일 가이드 RNA 및 RNP (예를 들어, CAS9)만을 필요로 했다. CeDNA 벡터는 또한 발현을 제거하기 위해 PD-1 유전자좌를 표적화하는데 사용될 수 있다. PD-1은 악성 종양에서 발생하는 만성 활성 T 세포에서 면역 체크포인트 세포 표면 수용체를 발현시킨다. 상기 슈만(Schumann) 등을 참고한다.
일부 구현예에서, ceDNA 유전자 편집 벡터는 병에 걸린 유전자를 표적으로 하는 벡터를 사용하여 결함 유전자를 교정하는데 사용된다. 일 구현예에서, 본원에 기재된 바와 같은 ceDNA 벡터는, 예를 들어, 뒤센느 근위축증을 치료하거나 레베르 선천성 흑암시(Leber Congenital Amaurosis)의 치료에서 LCA10의 돌연변이된 인트론을 제거하기 위해 프레임시프트 돌연변이를 교정하기 위해 원하는 DNA 영역을 여기시키는데 사용될 수 있다. ceDNA 벡터를 사용한 유전자 편집에 의해 치료될 수 있는 질환 또는 장애의 비제한적 예는 미국 특허 공보 2014/0170753의 이들 및 이들의 관련 유전자와 함께 표 A-C에 열거되어 있으며, 이 문헌은 그 전문이 본원에 참고로 포함된다. 대안적인 구현예에서, ceDNA 벡터는 세이프 하버 유전자, 예를 들어, 불활성 인트론에서 치료 단백질 또는 리포터 단백질의 발현을 위한 발현 카세트의 삽입에 사용된다. 특정 구현예에서, 프로모터가 없는 카세트는 세이프 하버 유전자에 삽입된다. 이러일 구현예에서, 프로모터가 없는 카세트는 세이프 하버 유전자 조절 요소 (프로모터, 인핸서 및 시그널링 펩티드)를 이용할 수 있으며, 세이프 하버 유전자좌에서의 삽입의 비제한적인 예는 하기에 기재된 알부민 유전자좌로의 삽입이다: Blood (2015) 126 (15): 1777-1784 (이는 그 전문이 본원에 참고로 포함됨). 알부민으로의 삽입은 이식유전자의 혈액으로의 분비를 가능하게 하는 이점이 있다 (예를 들어, 실시예 22 참조). 또한, 게놈 세이프 하버 부위는 당 업계에 공지되어 있고, 예를 들어, 하기에 기재된 기술을 사용하여 결정될 수 있다: Papapetrou, ER & Schambach, A. Molecular Therapy 24(4):678-684 (2016) or Sadelain et al. Nature Reviews Cancer 12:51-58 (2012) (이들 각각의 내용은 그 전문이 본원에 참고로 포함됨). 본원에서는 아데노 관련 바이러스 (AAV) 게놈의 세이프 하버 부위 (예를 들어, AAVS1 세이프 하버 부위)가 본원에 기재된 방법 및 조성물과 함께 사용될 수 있는 것으로 고려된다 (예를 들어, Oceguera-Yanez et al. Methods 101:43-55 (2016) or Tiyaboonchai, A et al. Stem Cell Res 12(3):630-7 (2014) 참조, 이들 각각의 내용은 그 전문이 참고로 포함됨). 예를 들어, AAVS1 게놈 세이프 하버 부위는 배아 줄기세포 (예를 들어, 인간 배아 줄기세포) 또는 유도된 다능성 줄기세포 (iPS 세포)에서 조혈 특이적 이식유전자 발현 및 유전자 침묵의 목적으로 본원에 기재된 ceDNA 벡터 및 조성물과 함께 사용될 수 있다. 또한, 염색체 19 상의 AASV1 세이프 하버 부위에 삽입하기 위한 합성 또는 상업적으로 이용 가능한 상동성-지정 복구 공여체 주형이 본원에 기재된 바와 같은 ceDNA 벡터 또는 조성물과 함께 사용될 수 있는 것으로 본원에 고려된다. 예를 들어, 상동성-지정 복구 주형, 및 가이드 RNA는, 예를 들어, 캘리포니아주 팔로 알토 소재의 System Biosciences로부터 상업적으로 구입하여 ceDNA 벡터 내로 클로닝될 수 있다.
일부 구현예에서, ceDNA 벡터는 T 세포에서 유전자를 녹아웃시키거나 편집하는데, 예를 들어, 개선된 입양 세포 전달 및/또는 CAR-T 요법을 위해 T 세포를 조작하기 위해 사용된다 (예를 들어, 실시예 24 참조). 일부 구현예에서, ceDNA 벡터는 본원에 기재된 바와 같은 유전자 편집 벡터일 수 있다. 일부 구현예에서, ceDNA 벡터는 본원의 다른 곳에 기재된 바와 같이, 엔도뉴클레아제, 주형 핵산 서열, 또는 엔도뉴클레아제 및 주형 핵산의 조합을 포함할 수 있다. 치료적으로 적절한 녹아웃 및 T 세포의 유전자 편집의 비제한적 예는 하기에 기재되어 있다: PNAS (2015) 112(33):10437-10442 (이는 그 전문이 본원에 참고로 포함됨).
유전자 편집 ceDNA 벡터 또는 이의 조성물은 임의의 유전 질환의 치료에 사용될 수 있다. 비제한적인 예로서, ceDNA 벡터 또는 이의 조성물은 예를 들어 돌연변이 단백질이 신경, 심장, 위장 시스템 등에서 미스폴딩되고 응집되는 희귀 질환(orphan disease)인, 트랜스티레틴 아밀로이드증 (ATTR)의 치료에 사용될 수 있다. 본원에 기재된 유전자 편집 시스템을 사용하여 돌연변이 질환 유전자 (mutTTR)를 결실시킴으로써 질환을 치료할 수 있는 것으로 고려된다. 유전성 질환의 이러한 치료는 질환 진행을 중단시킬 수 있고 확립된 질환의 퇴행 또는 질환의 적어도 하나의 증상의 적어도 10% 감소를 가능하게 할 수 있다.
또 다른 구현예에서, ceDNA 벡터 또는 이의 조성물은 오르니틴 트랜스카바밀라제 결핍증 (OTC 결핍증), 고암모니아혈증 또는 다른 요소 주기 장애의 치료에 사용될 수 있으며, 이는 신생아 또는 영아의 암모니아 해독 능력을 손상시킨다. 모든 선천성 대사 질환에서와 같이, 야생형 대조군과 비교하여 효소 활성의 부분적인 회복 (예를 들어, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60%, 적어도 70%, 적어도 80%, 적어도 90%, 적어도 95% 또는 적어도 99%)조차도 OTC 결핍증을 갖는 대상체에 대해 적어도 하나의 증상 OTC의 감소 및/또는 삶의 질 개선을 위해 충분할 수 있음이 본원에 고려된다. 일 구현예에서, OTC를 인코딩하는 핵산은 생체내 단백질 대체를 위해 알부민 내인성 프로모터 뒤에 삽입될 수 있다.
또 다른 구현예에서, ceDNA 벡터 또는 이의 조성물은 페닐알라닌 하이드록실라제 효소를 인코딩하는 핵산 서열을 전달하여 PKU 환자에게 유독할 수 있는, 식이 페닐알라닌의 축적을 감소시킴으로써 페닐케톤뇨증 (PKU)의 치료에 사용될 수 있다. 모든 선천성 대사 질환에서와 같이, 야생형 대조군과 비교하여 효소 활성의 부분적인 회복 (예를 들어, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60%, 적어도 70%, 적어도 80%, 적어도 90%, 적어도 95% 또는 적어도 99%)조차도 PKU를 갖는 대상체에 대해 PKU의 적어도 하나의 증상의 감소 및/또는 삶의 질 개선을 위해 충분할 수 있음이 본원에 고려된다. 일 구현예에서, 페닐알라닌 하이드록실라제를 인코딩하는 핵산은 생체내 단백질 대체를 위해 알부민 내인성 프로모터 뒤에 삽입될 수 있다.
또 다른 구현예에서, ceDNA 벡터 또는 이의 조성물은 글리코겐 저장병 (GSD)을 갖는 대상체에서 비정상 글리코겐 합성 또는 파괴를 교정하기 위해 효소를 인코딩하는 핵산 서열을 전달함으로써 글리코겐 저장병 (GSD)의 치료에 사용될 수 있다. 본원에 기재된 유전자 편집 방법을 사용하여 보정될 수 있는 효소의 비제한적인 예는 글리코겐 신타제, 글루코스-6-포스파타제, 산-알파 글루코시다제, 글리코겐 탈분지 효소, 글리코겐 분지 효소, 근육 글리코겐 포스포릴라제, 간 글리코겐 포스포릴라제, 근육 포스포프럭토키나제, 포스포릴라제 키나제, 포도당 수송체-2 (GLUT-2), 알돌라제 A, 베타-에놀라제, 포스포글루코뮤타제-1 (PGM-1), 및 글리코게닌-1을 포함한다. 모든 선천성 대사 질환에서와 같이, 야생형 대조군과 비교하여 효소 활성의 부분적인 회복 (예를 들어, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60%, 적어도 70%, 적어도 80%, 적어도 90%, 적어도 95% 또는 적어도 99%)조차도 GSD를 갖는 대상체에 대해 GSD의 적어도 하나의 증상의 감소 및/또는 삶의 질 개선을 위해 충분할 수 있음이 고려된다. 일 구현예에서, 비정상 글리코겐 저장을 교정하기 위한 효소를 인코딩하는 핵산은 생체내 단백질 대체를 위해 알부민 내인성 프로모터 뒤에 삽입될 수 있다.
본원에 기재된 ceDNA 벡터는 또한 레베르 선천성 흑암시 (LCA), 폴리Q 반복부를 포함하는 폴리글루타민 질환, 및 알파-1항트립신 결핍증 (A1AT)의 생체내 복구에 사용하기 위해 고려된다. LCA는 실명을 유발하는 희귀한 선천성 안과 질환으로, 이는 GUCY2D, RPE65, SPATA7, AIPL1, LCA5, RPGRIP1, CRX, CRB1, NMNAT1, CEP290, IMPDH1, RD3, RDH12, LRAT, TULP1, KCNJ13, GDF6 및/또는 PRPH2 중 임의의 하나의 유전자의 돌연변이에 의해 유발될 수 있다. 본원에 기재된 유전자 편집 방법 및 조성물은 LCA의 증상을 담당하는 유전자(들)에서의 오류를 교정하기 위해 LCA와 관련된 하나 이상의 유전자의 전달에 적합할 수 있음이 고려된다. 폴리글루타민 질환은 치상핵적핵담창구시상하핵 위축증 (dentatorubropallidoluysian atrophy), 헌팅턴병, 척수 및 구근 근위축증(spinal and bulbar muscular atrophy), 및 척수소뇌성 실조증(spinocerebellar ataxia) 유형 1, 2, 3 (마카도-조셉 질환(Machado-Joseph disease)으로도 알려져 있음), 6, 7 및 17을 포함하지만, 이에 제한되지는 않는다. ceDNA 벡터를 사용하는 유전자 편집 방법을 사용하여 DNA 돌연변이를 복구하여 폴리글루타민 질환과 관련된 것들과 같은 트리뉴클레오티드 반복 확장부 (예를 들어, 폴리Q 반복부)를 생성할 수 있음이 본원에 구체적으로 고려된다. A1AT 결핍은 알파-1항트립신의 결함 생성을 유발하여 혈액 및 폐에서 효소의 활성을 감소시키고, 결과적으로 병든 대상체에서 폐기종 또는 만성 폐쇄성 폐 질환을 유발할 수 있는 유전 장애이다. A1AT 결핍의 복구는 본원에 개괄된 ceDNA 벡터 또는 이의 조성물을 사용하여 본원에서 구체적으로 고려된다. LCA, 폴리글루타민 질환 또는 A1AT 결핍의 치료를 위해 원하는 단백질을 인코딩하는 핵산은 생체내 단백질 대체를 위해 알부민 내인성 프로모터 뒤에 삽입될 수 있음이 본원에 고려된다.
추가의 구현예에서, 본원에 기재된 바와 같은 ceDNA 벡터를 포함하는 조성물은 바이러스 서열, 병원체 서열, 염색체 서열, 전위 접합(translocation junction) (예를 들어, 암과 관련된 전위), 비-코딩 RNA 유전자 또는 질환 관련 유전자인 RNA 서열 등에서 유전자를 편집하는데 사용될 수 있다.
본원에 개시된 바와 같은 유전자 편집 ceDNA 벡터를 사용하여 관심있는 임의의 핵산 또는 표적 유전자를 편집할 수 있다. 표적 핵산 및 표적 유전자는 폴리펩티드를 인코딩하는 핵산, 또는 비-코딩 핵산 (예를 들어, RNAi, miR 등) 바람직하게는 치료적 (예를 들어, 의학적, 진단적 또는 수의학적 용도를 위한) 또는 면역원성 (예를 들어, 백신을 위한) 폴리펩티드를 포함하지만, 이에 제한되지 않는다. 특정 구현예에서, 본원에 기재된 유전자 편집 ceDNA 벡터에 의해 표적화되는 표적 핵산 또는 표적 유전자는 하나 이상의 폴리펩티드, 펩티드, 리보자임, 펩티드 핵산, siRNA, RNAi, 안티센스 올리고뉴클레오티드, 안티센스 폴리뉴클레오티드, 항체, 항원 결합 단편 또는 이들의 임의의 조합을 인코딩한다.
특히, 본원에 개시된 ceDNA 벡터에 의한 유전자 편집을 위한 유전자 표적은 질환, 기능 장애, 손상 및/또는 장애의 하나 이상의 증상의 치료, 예방 및/또는 개선에 사용하기 위해, 예를 들어, 비제한적으로, 단백질(들), 폴리펩티드(들), 펩티드(들), 효소(들), 항체, 항원 결합 단편, 뿐만 아니라 이의 변이체 및/또는 이의 활성 단편을 인코딩할 수 있다. 일 측면에서, 질환, 기능 장애, 외상, 손상 및/또는 장애는 인간 질환, 기능 장애, 외상, 손상 및/또는 장애이다.
본원에 언급된 바와 같이, 본원에 개시된 ceDNA 벡터를 사용한 유전자 편집을 위한 유전자 표적은 단백질 또는 펩티드, 또는 하나 이상의 작용제, 길항제, 항-아폽토시스 인자, 억제제, 수용체, 사이토카인, 세포독소, 적혈구 생성 촉진제, 당단백질, 성장 인자, 성장 인자 수용체, 호르몬, 호르몬 수용체, 인터페론, 인터류킨, 인터류킨 수용체, 신경 성장 인자, 신경활성 펩티드, 신경활성 펩티드 수용체, 프로테아제, 프로테아제 억제제, 단백질 데카르복실라제, 단백질 키나제, 단백질 키나제 억제제, 효소, 수용체 결합 단백질, 수송 단백질 또는 이의 하나 이상의 억제제, 세로토닌 수용체 또는 이의 하나 이상의 흡수 억제제, 세르핀, 세르핀 수용체, 종양 억제제, 진단 분자, 화학요법제, 세포독소 또는 이들의 임의의 조합을 포함하지만 이에 제한되지 않는, 치료 핵산 서열 또는 치료제를 인코딩할 수 있다.
C. 유전자 편집을 위한 추가 질환:
일반적으로, 본원에 개시된 바와 같은 ceDNA 벡터는 유전자 발현과 관련된 임의의 장애와 관련된 증상을 치료, 예방 또는 개선하기 위해 상기 설명에 따라 임의의 이식유전자를 전달하는데 사용될 수 있다. 예시적인 질환 상태는 낭포성 섬유증 (및 폐의 다른 질환), 혈우병 A, 혈우병 B, 지중해 빈혈, 빈혈 및 기타 혈액 장애, AIDS, 알츠하이머병, 파킨슨병, 헌팅턴병, 근위축성 측삭 경화증, 간질 및 기타 신경계 장애, 암, 당뇨병, 근위축증 (예를 들어, 뒤센느, 베커), 헐러병(Hurler's disease), 아데노신 데아미나제 결핍, 대사 결함, 망막 퇴행성 질환 (및 눈의 다른 질환), 미토콘드리아증 (예를 들어, 레버의 유전성 시신경병증 (LHON), 리 증후군 및 아급성 경화성 뇌병증), 근병증 (예를 들어, 안면견갑상완 근병증 (FSHD) 및 심근병증), 고형 장기 (예를 들어, 뇌, 간, 신장, 심장) 질환 등을 포함하지만, 이에 제한되지는 않는다. 일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 대사 장애 (예를 들어, 오미틴 트랜스카바밀라제 결핍)를 갖는 개체의 치료에 유리하게 사용될 수 있다.
일부 구현예에서, 본원에 기재된 ceDNA 벡터는 유전자 또는 유전자 산물에서의 돌연변이에 의해 야기된 질환 또는 장애를 치료, 개선 및/또는 예방하는데 사용될 수 있다. ceDNA 벡터로 치료될 수 있는 예시적인 질환 또는 장애는 대사 질환 또는 장애 (예를 들어, 파브리병, 고셔병, 페닐케톤뇨증 (PKU), 글리코겐 저장병); 우레아 사이클 질환 또는 장애 (예를 들어, 오르니틴 트랜스카바밀라제 (OTC) 결핍); 리소좀 저장 질환 또는 장애 (예를 들어, 이염성백질디스트로피 (MLD), 점막다당류증 II형 (MPSII; 헌터 증후군)); 간 질환 또는 장애 (예를 들어, 진행 가족성 간내담즙정체 (PFIC); 혈액 질환 또는 장애 (예를 들어, 혈우병 (A 및 B), 지중해 빈혈 및 빈혈); 암 및 종양, 및 유전 질환 또는 장애 (예를 들어, 낭포성 섬유증)를 포함하지만, 이에 제한되지 않는다.
또 다른 측면으로서, 본원에 개시된 바와 같은 ceDNA 벡터는 이식유전자 발현 수준 (예를 들어, 본원에 기재된 바와 같은 호르몬 또는 성장 인자를 인코딩하는 이식유전자)을 조절하는 것이 바람직한 상황에서 이종 뉴클레오티드 서열을 전달하는데 사용될 수 있다.
따라서, 일부 구현예에서, 본원에 기재된 ceDNA 벡터는 질환 또는 장애를 초래하는 유전자 산물의 비정상적인 수준 및/또는 기능 (예를 들어, 단백질의 부재 또는 결함)을 교정하는데 사용될 수 있다. ceDNA 벡터는 기능성 단백질을 생성하고/하거나 단백질의 수준을 변형시켜 단백질에서 부재 또는 결함에 의해 야기되는 특정 질환 또는 장애로 인한 증상을 완화시키거나 감소시키거나 상기 특정 질환 또는 장애에 이점을 제공할 수 있다. 예를 들어, OTC 결핍의 치료는 기능성 OTC 효소를 생성함으로써 달성될 수 있으며; 혈우병 A 및 B의 치료는 인자 VIII, 인자 IX 및 인자 X의 수준을 변형시킴으로써 달성될 수 있으며; PKU의 치료는 페닐알라닌 하이드록실라제 효소의 수준을 변형시킴으로써 달성될 수 있으며; 파브리병 또는 고셔병의 치료는 각각 기능성 알파 갈락토시다제 또는 베타 글루코세레브로시다제를 생성함으로써 달성될 수 있으며; MLD 또는 MPSII의 치료는 각각 기능성 아릴설파타제 A 또는 이듀로네이트-2-설파타제를 생성함으로써 달성될 수 있으며; 낭포성 섬유증의 치료는 기능성 낭포성 섬유증 막횡단 전도 조절제를 생성함으로써 달성될 수 있으며; 글리코겐 저장병의 치료는 기능적 G6Pase 효소 기능을 회복시킴으로써 달성될 수 있으며; PFIC의 치료는 기능적 ATP8B1, ABCB11, ABCB4 또는 TJP2 유전자를 생성함으로써 달성될 수 있다.
대안적인 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 안티센스 핵산 을 시험관내 또는 생체내에서 세포에 제공하는데 사용될 수 있다. 예를 들어, 이식유전자이 RNAi 분자인 경우, 표적 세포에서 안티센스 핵산 또는 RNAi의 발현은 세포에 의한 특정 단백질의 발현을 감소시킨다. 따라서, RNAi 분자 또는 안티센스 핵산인 이식유전자는 이를 필요로 하는 대상체에서 특정 단백질의 발현을 감소시키기 위해 투여될 수 있다. 안티센스 핵산은 또한 세포 생리학을 조절하기 위해, 예를 들어, 세포 또는 조직 배양 시스템을 최적화하기 위해 시험관내에서 세포에 투여될 수 있다.
일부 구현예에서, ceDNA 벡터에 의해 인코딩된 예시적인 이식유전자는 X, 리소좀 효소 (예를 들어, 테이-삭스병과 관련된 헥소사미니다제 A, 또는 헌터 증후군/MPS II와 관련된 이듀로네이트 설파타제), 에리트로포이에틴, 안지오스타틴, 엔도스타틴, 초산화물 디스뮤타제, 글로빈, 렙틴, 카탈라제, 티로신 하이드록실라제, 및 사이토카인 (예를 들어, 인터페론, β-인터페론, 인터페론-γ, 인터류킨-2, 인터류킨-4, 인터류킨 12, 과립구-대식세포 콜로니 자극 인자, 림프독소 등), 펩티드 성장 인자 및 호르몬 (예를 들어, 소마토트로핀, 인슐린, 인슐린-유사 성장 인자 1 및 2, 혈소판 유래 성장 인자 (PDGF), 표피 성장 인자 (EGF), 섬유 모세포 성장 인자 (FGF), 신경 성장 인자 (NGF), 신경 영양 인자-3 및 4, 뇌-유래 신경 영양 인자 (BDNF), 신경교 유래 성장 인자 (GDNF), 형질전환 성장 인자-α 및 -β 등), 수용체 (예를 들어, 종양 괴사 인자 수용체)를 포함하지만, 이에 제한되지 않는다. 일부 예시적인 구현예에서, 이식유전자는 하나 이상의 원하는 표적에 특이적인 모노클로날 항체를 인코딩한다. 일부 예시적인 구현예에서, 하나 초과의 이식유전자는 ceDNA 벡터에 의해 인코딩된다. 일부 예시적인 구현예에서, 이식유전자는 2개의 상이한 관심 폴리펩티드를 포함하는 융합 단백질을 인코딩한다. 일부 구현예에서, 이식유전자는 본원에 정의된 바와 같이, 전장 항체 또는 항체 단편을 포함하는 항체를 인코딩한다. 일부 구현예에서, 항체는 본원에 정의된 바와 같이, 항원-결합 도메인 또는 면역글로불린 가변 도메인 서열이다. 다른 예시적인 이식유전자 서열은 자살 유전자 산물 (팀딘(thymdine) 키나제, 시토신 데아미나제, 디프테리아 독소, 시토크롬 P450, 데옥시시티딘 키나제, 및 종양 괴사 인자), 암 요법에 사용되는 약물에 대한 내성을 부여하는 단백질, 및 종양 억제 유전자 산물을 인코딩한다.
대표적인 구현예에서, ceDNA 벡터에 의해 발현된 이식유전자는 근위축증의 치료를 필요로 하는 대상체에서 근위축증의 치료에 사용될 수 있으며, 상기 방법은 본원에 기재된 치료-, 개선- 또는 예방-유효량의 ceDNA 벡터를 투여하는 단계를 포함하며, 여기서 상기 ceDNA 벡터는 디스트로핀, 미니-디스트로핀, 마이크로-디스트로핀, 미오스타틴 프로펩티드, 폴리스타틴, 액티빈 II형 가용성 수용체, IGF-1, 항염증성 폴리펩티드, 예컨대 Ikappa B 우성 돌연변이체, 사르스코판, 우트로핀, 마이크로-디스트로핀, 라미닌-α2, α-사르코글리칸, β-사르코글리칸, γ-사르코글리칸, δ-사르코글리칸, IGF-1, 미오스타틴 또는 미오스타틴 프로펩티드에 대한 항체 또는 항체 단편, 및/또는 미오스타틴에 대한 RNAi를 인코딩하는 이종 핵산을 포함한다. 특정 구현예에서, ceDNA 벡터는 본원의 다른 곳에 기재된 바와 같이 골격, 횡격막 및/또는 심장 근육에 투여될 수 있다.
일부 구현예에서, ceDNA 벡터는 혈액에서 정상적으로 순환하는 폴리펩티드 (예를 들어, 효소) 또는 기능성 RNA (예를 들어, RNAi, 마이크로RNA, 안티센스 RNA)의 생성, 또는 장애 (예를 들어, 대사 장애, 예컨대 당뇨병 (예를 들어, 인슐린), 혈우병 (예를 들어, VIII), 무코다당류 장애 (예를 들어, Sly 증후군, 헐러 증후군, 샤이 증후군(Scheie Syndrome), 헐러-샤이 증후군, 헌터 증후군, 산필리포 증후군(Sanfilippo Syndrome) A, B, C, D, 모르키오 증후군(Morquio Syndrome), 마로토-라미 증후군(Maroteaux-Lamy Syndrome) 등) 또는 리소좀 저장 장애 (예컨대 고셔병 [글루코세레브로시다제], 폼페병 [리소좀산 .알파-글루코시다제] 또는 파브리병 [.알파-갈락토시다제 A]) 또는 글리코겐 저장 장애 (예컨대, 폼페병 [리소좀산 글루코시다제])를 치료, 개선 및/또는 예방하기 위해 다른 조직으로 전신 전달하기 위해 골격, 심장 또는 횡격막 근육에 이식유전자를 전달하는데 사용될 수 있다. 대사 장애를 치료, 개선 및/또는 예방하기 위한 다른 적합한 단백질이 상기 기재되어 있다.
다른 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 이를 필요로 하는 대상체에서 대사 장애를 치료, 개선 및/또는 예방하는 방법으로 이식유전자를 전달하는데 사용될 수 있다. 예시적인 대사 장애 및 폴리펩티드를 인코딩하는 이식유전자는 본원에 기재되어 있다. 선택적으로, 폴리펩티드는 분비된다 (예를 들어, 천연 상태의 분비된 폴리펩티드이거나, 예를 들어, 당 업계에 공지된 바와 같은 분비 신호 서열과의 작동 가능한 회합에 의해 분비되도록 조작된 폴리펩티드).
본 발명의 또 다른 측면은 이를 필요로 하는 대상체에서 선천성 심부전 또는 PAD를 치료, 개선 및/또는 예방하는 방법에 관한 것으로, 상기 방법은 본원에 기재된 바와 같은 ceDNA 벡터를 포유동물 대상체에게 투여하는 단계를 포함하며, 여기서 ceDNA 벡터는, 예를 들어, 근형질 내부 세망(sarcoplasmic endoreticulum) Ca2+-ATPase (SERCA2a), 혈관 신생 인자, 포스파타제 억제제 I (I-1), 포스포람반에 대한 RNAi; 포스포람반 억제 또는 우세-음성 분자, 예컨대 포스포람반 S16E, 포스포람반 유전자를 조절하는 아연 핑거 단백질, β2-아드레날린 수용체, .베타.2-아드레날린 수용체 키나제 (BARK), PI3 키나제, 칼사르칸, .베타.-아드레날린 수용체 키나제 억제제 (βARKct), 단백질 포스파타제 1의 억제제 1, S100A1, 파발부민, 아데닐릴 사이클라제 6형, 절단된 구성적 활성 βARKct, Pim-1, PGC-1α, SOD-1, SOD-2, EC-SOD, 칼리크레인, HIF, 티모신-β4, mir-1, mir-133, mir-206 및/또는 mir-208과 같은 G-단백질 결합 수용체 키나제 2형 녹다운에 영향을 미치는 분자를 인코딩하는 이식유전자를 포함한다.
본원에 개시된 바와 같은 ceDNA 벡터는 임의의 적합한 수단에 의해, 선택적으로 대상체가 흡입하는, ceDNA 벡터를 포함하는 호흡 가능한 입자의 에어로졸 현탁액을 투여함으로써 대상체의 폐에 투여될 수 있다. 호흡 가능한 입자는 액체 또는 고체일 수 있다. ceDNA 벡터를 포함하는 액체 입자의 에어로졸은 당업자에게 공지된 바와 같이, 임의의 적합한 수단, 예컨대 압력 구동식 에어로졸 분무기 또는 초음파 분무기를 사용하여 제조될 수 있다. 예를 들어, 미국 특허 번호 4,501,729 호를 참조한다. ceDNA 벡터를 포함하는 고체 입자의 에어로졸도 마찬가지로 제약 분야에 공지된 기술에 의해 임의의 고체 미립자 의약 에어로졸 발생기로 제조될 수 있다.
일부 구현예에서, ceDNA 벡터는 CNS의 조직 (예를 들어, 뇌, 눈)에 투여될 수 있다. 특정 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 유전 장애, 신경퇴행성 장애, 정신 장애 및 종양을 포함하는, CNS 질환을 치료, 개선 또는 예방하기 위해 투여될 수 있다. CNS의 예시적인 질환은 알츠하이머병, 파킨슨병, 헌팅턴병, 카나반병(Canavan disease), 라이병(Leigh's disease), 레프숨병(Refsum disease), 투렛 증후군, 1차 측삭 경화증, 근위축성 측삭 경화증, 진행성 근위축증, 피크병(Pick's disease), 근위축증, 다발성 경화증, 중증 근무력증, 빈스완거병(Binswanger's disease), 척수 또는 두부 손상으로 인한 외상, 테이 삭스병, 레쉬-나이안병(Lesch-Nyan disease), 간질, 뇌경색, 기분 장애 (예를 들어, 우울증, 양극성 정동 장애, 지속성 정동 장애, 2차 기분 장애)를 포함한 정신 장애, 조현병, 약물 의존성 (예를 들어, 알코올 중독 및 기타 물질 의존성), 신경증 (예를 들어, 불안, 강박 장애, 신체형 장애, 해리성 장애, 비탄, 산후 우울증), 정신병 (예를 들어, 환각 및 망상), 치매, 편집증, 주의력 결핍 장애, 정신성적 장애, 수면 장애, 통증 장애, 식이 또는 체중 장애 (예를 들어, 비만, 악액질, 신경성 식욕부진 및 폭식증(bulemia)) 및 CNS의 암 및 종양 (예를 들어, 뇌하수체 종양)을 포함하지만, 이에 제한되지는 않는다.
본 발명의 ceDNA 벡터로 치료, 개선 또는 예방될 수 있는 안구 장애에는 망막, 후관(posterior tract) 및 시신경을 포함하는 안과 질환 (예를 들어, 망막색소변성증, 당뇨병성 망막병증 및 기타 망막 퇴행성 질환, 포도막염, 연령-관련 황반 변성, 녹내장)을 포함한다. 많은 안과 질환 및 장애는 3가지 유형의 징후 중 하나 이상과 관련이 있다: (1) 혈관 신생, (2) 염증 및 (3) 변성. 일부 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 항-혈관 신생 인자; 항염증 인자; 세포 변성을 지연시키거나, 세포 스페어링(cell sparing)을 촉진시키거나, 또는 세포 성장를 촉진시키는 인자 및 상기의 조합을 전달하기 위해 사용될 수 있다. 예를 들어, 당뇨병성 망막병증은 혈관 신생을 특징으로 한다. 당뇨병성 망막병증은 안구 내로 (예를 들어, 유리체에서) 또는 안구 주위로 (예를 들어, 서브-테논 부위에서) 하나 이상의 항-혈관 신생 인자를 전달함으로써 치료될 수 있다. 하나 이상의 신경 영양 인자는 또한 안구 내로 (예를 들어, 유리체 내로) 또는 안구 주위로 공동-전달될 수 있다. 본 발명의 ceDNA 벡터로 치료, 개선 또는 예방될 수 있는 추가의 안 질환은 지도모양 위축(geographic atrophy), 혈관 또는 "습성" 황반 변성, 스타가르트, 질환(Stargardt disease), 레베르 선천성 흑암시 (LCA), 어셔 증후군(Usher syndrome), 탄력섬유성 가황색종 (PXE), x-연관 망막색소변성증 (XLRP), x-연관 망막층간분리증 (XLRS), 맥락막결손, 레베르 유전성 시신경병증 (LHON), 아코마톱시아(Archomatopsia), 추체-간체 이영양증, 퓨크스 내피 각막 이영양증(Fuchs endothelial corneal dystrophy), 당뇨병성 황반 부종 및 안암 및 안 종양을 포함한다.
일부 구현예에서, 염증성 안 질환 또는 장애 (예를 들어, 포도막염)는 본 발명의 ceDNA 벡터에 의해 치료, 개선 또는 예방될 수 있다. 하나 이상의 항염증 인자는 본원에 개시된 바와 같은 ceDNA 벡터의 안내 (예를 들어, 유리체 또는 전방) 투여에 의해 발현될 수 있다. 다른 구현예에서, 망막 변성을 특징으로 하는 안 질환 또는 장애 (예를 들어, 망막색소변성증)는 본 발명의 ceDNA 벡터에 의해 치료, 개선 또는 예방될 수 있다. 하나 이상의 신경 영양 인자를 인코딩하는 본원에 개시된 바와 같은 ceDNA 벡터의 안내 (예를 들어, 유리체 투여)는 이러한 망막 변성-기반 질환을 치료하는데 사용될 수 있다. 일부 구현예에서, 혈관 신생 및 망막 변성 (예를 들어, 연령-관련 황반 변성)을 모두 포함하는 질환 또는 장애는 본 발명의 ceDNA 벡터로 치료될 수 있다. 연령-관련 황반 변성은 안구 내로 (예를 들어, 유리체) 하나 이상의 신경 영양 인자 및/또는 안구 내로 또는 안구 주위로 (예를 들어, 서브-테논 부위에서) 하나 이상의 항-혈관 신생 인자를 인코딩하는 본원에 개시된 바와 같은 ceDNA 벡터를 투여함으로써 치료될 수 있다. 녹내장은 안압 증가 및 망막 신경절 세포의 손실을 특징으로 한다. 녹내장의 치료는 본원에 개시된 바와 같은 ceDNA 벡터를 사용하여 세포를 흥분독성 손상으로부터 보호하는 하나 이상의 신경보호제의 투여를 포함한다. 따라서, 이러한 작용제는 N-메틸-D-아스파르테이트 (NMDA) 길항제, 사이토카인 및 신경 영양 인자를 포함하며, 본원에 개시된 바와 같은 ceDNA 벡터를 사용하여 안내, 선택적으로적으로 유리체 내로 전달될 수 있다.
다른 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 발작을 치료하기 위해, 예를 들어 발작의 개시, 발병 또는 중증도를 감소시키기 위해 사용될 수 있다. 발작에 대한 치료적 치료의 효능은 행동 (예를 들어, 눈 또는 입의 전율, 틱(tick)) 및/또는 전기 촬영술(electrographic) 수단 (대부분의 발작은 현저한 전기 촬영술 이상을 갖는다)에 의해 평가될 수 있다. 따라서, 본원에 개시된 바와 같은 ceDNA 벡터는 또한 간질을 치료하는데 사용될 수 있으며, 이는 시간이 지남에 따라 다수의 발작으로 나타난다. 하나의 대표적인 구현예에서, 뇌하수체 종양을 치료하기 위해 소마토스타틴 (또는 이의 활성 단편)이 본원에 개시된 바와 같은 ceDNA 벡터를 사용하여 뇌에 투여된다. 이 구현예에 따르면, 소마토스타틴 (또는 이의 활성 단편)을 인코딩하는 본원에 개시된 바와 같은 ceDNA 벡터는 뇌하수체로의 미세주입에 의해 투여된다. 마찬가지로, 이러한 치료는 말단 비대증 (뇌하수체에서 비정상적인 성장 호르몬 분비)을 치료하는데 사용될 수 있다. 당 업계에 공지된 바와 같이 소마토스타틴의 핵산 (예를 들어, GenBank 수탁 번호 J00306) 및 아미노산 (예를 들어, GenBank 수탁 번호 P01166; 프로세싱된 활성 펩티드 소마토스타틴-28 및 소마토스타틴-14를 함유함) 서열. 특정 구현예에서, ceDNA 벡터는 미국 특허 7,071,172에 기재된 바와 같은 분비 신호를 포함하는 이식유전자를 인코딩할 수 있다.
본 발명의 또 다른 측면은 생체내에서 대상체에게 전신 전달하기 위한 안티센스 RNA, RNAi 또는 다른 기능성 RNA (예를 들어, 리보자임)를 생성하기 위한 본원에 기재된 바와 같은 ceDNA 벡터의 용도에 관한 것이다. 따라서, 일부 구현예에서, ceDNA 벡터는 안티센스 핵산, 리보자임 (예를 들어, 미국 특허 번호 5,877,022에 기재된 바와 같음), 스플라이세오솜(spliceosome)-매개 트랜스-스플라이싱에 영향을 미치는 RNA (Puttaraju et al., (1999) Nature Biotech. 17:246; 미국 특허 번호 6,013,487; 미국 특허 번호 6,083,702 참조), 유전자 침묵을 매개하는 간섭 RNA (RNAi) (Sharp et al., (2000) Science 287:2431 참조) 또는 다른 "비-번역" RNA, 예컨대 "가이드" RNA (Gorman et al., (1998) Proc. Nat. Acad. Sci. USA 95:4929; Yuan et al.의 미국 특허 번호 5,869,248) 등을 인코딩하는 이식유전자를 포함할 수 있다.
일부 구현예에서, ceDNA 벡터는 리포터 폴리펩티드 (예를 들어, 녹색 형광 단백질 또는 알칼리성 포스파타제와 같은 효소)를 인코딩하는 이식유전자를 또한 추가로 포함할 수 있다. 일부 구현예에서, 실험 또는 진단 목적에 유용한 리포터 단백질을 인코딩하는 이식유전자는 β-락타마제, β-갈락토시다제 (LacZ), 알칼리성 포스파타제, 티미딘 키나제, 녹색 형광 단백질 (GFP), 클로람페니콜 아세틸트랜스퍼 라제 (CAT), 루시퍼라제 및 당 업계에 잘 알려진 다른 것들 중 어느 것으로부터 선택된다. 일부 측면에서, 리포터 폴리펩티드를 인코딩하는 이식유전자를 포함하는 ceDNA 벡터는 진단 목적으로 또는 이들이 투여되는 대상체에서 ceDNA 벡터의 활성의 마커로서 사용될 수 있다.
일부 구현예에서, ceDNA 벡터는 숙주 염색체 상의 유전자좌와 상동성을 공유하고 상기 유전자좌와 재조합하는 이식유전자 또는 이종 뉴클레오티드 서열을 포함할 수 있다. 이 접근법은 숙주 세포에서 유전적 결함을 교정하는데 이용될 수 있다.
일부 구현예에서, ceDNA 벡터는, 예를 들어 백신 접종을 위해 대상체에서 면역원성 폴리펩티드를 발현시키는데 사용될 수 있는 이식유전자를 포함할 수 있다. 이식유전자는 인간 면역결핍 바이러스, 인플루엔자 바이러스, gag 단백질, 종양 항원, 암 항원, 박테리아 항원, 바이러스 항원 등으로부터의 면역원을 포함하지만, 이에 제한되지 않는 당 업계에 공지된 임의의 관심 면역원을 인코딩할 수 있다.
D. 유전자 편집 ceDNA 벡터를 사용한 성공적인 유전자 편집 시험
당 분야에 널리 공지된 분석법을 사용하여 시험관내 및 생체내 모델 모두에서 ceDNA에 의한 유전자 편집의 효율을 시험할 수 있다. ceDNA에 의한 원하는 이식유전자의 녹인 또는 녹아웃은 원하는 이식유전자의 mRNA 및 단백질 수준을 측정함으로써 (예를 들어, 역전사 PCR, 웨스턴 블롯 분석, 및 효소-결합 면역흡착 분석법 (ELISA)) 당업자에 의해 평가될 수 있다. ceDNA에 의한 핵산 변경 (예를 들어, 점 돌연변이 또는 DNA 영역의 결실)은 게놈 표적 DNA의 심층 시퀀싱에 의해 평가될 수 있다. 일 구현예에서, ceDNA는, 예를 들어 형광 현미경 또는 발광 플레이트 판독기에 의해 리포터 단백질의 발현을 조사함으로써, 원하는 이식유전자의 발현을 평가하는데 사용될 수 있는 리포터 단백질을 포함한다. 생체내 적용의 경우, 단백질 기능 분석을 사용하여 유전자 편집이 성공적으로 이루어졌는지 확인하기 위해 주어진 유전자 및/또는 유전자 산물의 기능성을 시험할 수 있다. 예를 들어, 낭포성 섬유증 막횡단 전도 조절 유전자 (CFTR)의 점 돌연변이는 음이온 채널을 통해 음이온 (예를 들어, Cl-)을 이동시키는 CFTR의 능력을 억제하고, ceDNA의 유전자 편집 능력에 의해 교정될 수 있는 것으로 예상된다. ceDNA의 투여 후, 당업자는 CFTR의 점 돌연변이가 교정되었는지를 결정하기 위해 음이온이 음이온 채널을 통해 이동하는 능력을 평가할 수 있다. 당업자는 시험관내 또는 생체내에서 단백질의 기능성을 측정하기 위한 최상의 시험을 결정할 수 있을 것이다.
세포 또는 대상체에서 유전자 편집의 효과는 적어도 1개월, 적어도 2개월, 적어도 3개월, 적어도 4개월, 적어도 5개월, 적어도 6개월, 적어도 10개월, 적어도 12개월, 적어도 18개월, 적어도 2년, 적어도 5년, 적어도 10년, 적어도 20년 동안 지속되거나 영구적일 수 있음이 고려된다.
일부 구현예에서, 본원에 기재된 발현 카세트, 발현 작제물 또는 ceDNA 벡터의 이식유전자는 숙주 세포에 대해 코돈 최적화될 수 있다. 본원에 사용된 용어 "코돈 최적화된" 또는 "코돈 최적화"는 천연 서열 (예를 들어, 원핵생물 서열)의 적어도 하나, 하나 초과 또는 상당수의 코돈을 척추동물의 유전자에서 보다 빈번하게 또는 가장 빈번하게 사용되는 코돈으로 대체함으로써, 관심 척추동물의 세포, 예를 들어 마우스 또는 인간 (예를 들어, 인간화된)의 세포에서 발현을 향상시키기 위해 핵산 서열을 변형시키는 과정을 지칭한다. 다양한 종은 특정 아미노산의 특정 코돈에 대해 특정한 편향을 나타낸다. 전형적으로, 코돈 최적화는 원래 번역된 단백질의 아미노산 서열을 변경시키지 않는다. 최적화된 코돈은 예를 들어, Aptagen's Gene Forge® 코돈 최적화 및 맞춤형 유전자 합성 플랫폼 (Aptagen, Inc.) 또는 또 다른 공개적으로 이용 가능한 데이터베이스를 사용하여 결정될 수 있다.
XIII. 투여
특정 구현예에서, 다양한 간격, 예를 들어, 매일, 매주, 매월, 매년 등에 걸쳐 원하는 수준의 유전자 발현을 달성하기 위해 하나 초과의 투여 (예를 들어, 2, 3, 4회 또는 그 이상의 투여)가 사용될 수 있다.
본원에 개시된 ceDNA 벡터의 예시적인 투여 방식은 경구, 직장, 경점막, 비강내, 흡입 (예를 들어, 에어로졸을 통한), 협측 (예를 들어, 설하), 질, 경막내, 안내, 경피, 내피내, 자궁 내 (또는 난(ovo) 내), 비경구 (예를 들어, 정맥내, 피하, 피내, 두개내, 근육내 (골격, 횡격막 및/또는 심장 근육으로의 투여 포함), 흉막내, 뇌내 및 관절내), 국소 (예를 들어, 기도 표면, 및 경피 투여를 포함하는 피부 및 점막 표면 모두에), 림프내 등, 및 직접 조직 또는 기관 주사 (예를 들어, 간, 눈, 골격근, 심장 근육, 횡격막 근육 또는 뇌에)를 포함한다.
ceDNA 벡터의 투여는 뇌, 골격근, 평활근, 심장, 횡격막, 기도 상피, 간, 신장, 비장, 췌장, 피부 및 눈으로 구성된 군으로부터 선택된 부위를 포함하지만, 이에 제한되지 않는 대상체의 임의의 부위에 투여될 수 있다. ceDNA 벡터의 투여는 또한 종양 (예를 들어, 종양 또는 림프절 내 또는 그 근처)으로의 투여일 수 있다. 임의의 경우에 가장 적합한 경로는 치료, 개선 및/또는 예방되는 상태의 성질 및 중증도 및 사용되는 특정 ceDNA 벡터의 성질에 따라 달라질 것이다. 또한, ceDNA는 하나의 벡터 또는 다수의 ceDNA 벡터 (예를 들어 ceDNA 칵테일)에서 하나 초과의 이식유전자를 투여할 수 있게 한다.
본 발명에 따른 골격근으로 본원에 개시된 ceDNA 벡터의 투여는 사지 (예를 들어, 상완, 하완, 상부 다리 및/또는 하부 다리), 등, 목, 머리 (예를 들어, 혀), 흉부, 복부, 골반/회음 및/또는 손발가락의 골격근으로의 투여를 포함하지만 이에 제한되지 않는다. 본원에 개시된 바와 같은 ceDNA는 정맥내 투여, 동맥내 투여, 복강내 투여, 사지 관류, (선택적으로 다리 및/또는 팔의 고립된 사지 관류; 예를 들어, Arruda et al., (2005) Blood 105: 3458-3464), 및/또는 직접 근육내 주사에 의해 골격근으로 전달될 수 있다. 특정 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 사지 관류, 선택적으로 고립된 사지 관류 (예를 들어, 정맥내 또는 관절내 투여에 의해 대상체 (예를 들어, DMD와 같은 근위축증을 갖는 대상체)의 사지 (팔 및/또는 다리)로 투여된다. 특정 구현예에서, 본원에 개시된 바와 같은 ceDNA 벡터는 "유체 역학" 기술을 사용하지 않고 투여될 수 있다.
본원에 개시된 바와 같은 ceDNA 벡터의 심장 근육으로의 투여는 좌심방, 우심방, 좌심실, 우심실 및/또는 중격으로의 투여를 포함한다. 본원에 기재된 바와 같은 ceDNA 벡터는 정맥내 투여, 대동맥내 투여와 같은 동맥내 투여, 직접 심장 주사 (예를 들어, 좌심방, 우심방, 좌심실, 우심실) 및/또는 관상 동맥 관류에 의해 심장 근육으로 전달될 수 있다. 횡격막 근육으로의 투여는 정맥내 투여, 동맥내 투여 및/또는 복막내 투여를 포함하는 임의의 적합한 방법에 의해 이루어질 수 있다. 평활근으로의 투여는 정맥내 투여, 동맥내 투여 및/또는 복막내 투여를 포함하는 임의의 적합한 방법에 의해 이루어질 수 있다. 일 구현예에서, 투여는 평활근에 존재하고/하거나, 그 근처에 및/또는 평활근 상에 존재하는 내피 세포로 투여될 수 있다.
일부 구현예에서, 본 발명에 따른 ceDNA 벡터는 골격근, 횡격막 근육 및/또는 심장 근육으로 투여된다 (예를 들어, 근이영양증 또는 심장병 (예를 들어, PAD 또는 울혈성 심부전)을 치료, 개선 및/또는 예방하기 위해).
A. 생체외
치료
일부 구현예에서, 대상체로부터 세포를 제거하고, ceDNA 벡터가 그 안에 도입되고, 이어서 세포는 대상체 내로 다시 교체된다. 생체외 치료를 위해 대상체로부터 세포를 제거한 후 대상체로 다시 도입하는 방법은 당 업계에 공지되어 있다 (예를 들어, 미국 특허 번호 5,399,346 참조; 이의 개시내용은 그 전문이 본원에 포함됨). 대안적으로, ceDNA 벡터는 또 다른 대상체 유래 세포, 배양 세포 또는 임의의 다른 적합한 공급원 유래 세포 내로 도입되고, 세포는 이를 필요로 하는 대상체에게 투여된다.
ceDNA 벡터로 형질도입된 세포는 바람직하게는 약제학적 담체와 함께 "치료 적 유효량"으로 대상체에게 투여된다. 당업자는 치료 효과가 대상체에게 약간의 이점이 제공되는 한 완전하거나 치유력이 있을 필요는 없음을 이해할 것이다.
일부 구현예에서, ceDNA 벡터는 바람직하게는, 시험관내, 생체외 또는 생체내 세포에서 생산된 임의의 폴리펩티드인 이식유전자 (때로는 이종 뉴클레오티드 서열이라고도 함)을 인코딩할 수 있다. 예를 들어, 본원에 논의된 바와 같은 치료 방법에서 ceDNA 벡터의 사용과 대조적으로, 일부 구현예에서 ceDNA 벡터는, 예를 들어, 항원 또는 백신의 생산을 위해 배양 세포 및 이로부터 단리된 발현된 유전자 산물 내로 도입될 수 있다.
ceDNA 벡터는 수의학 및 의학적 용도로 사용될 수 있다. 상기 기재된 바와 같은 생체외 유전자 전달 방법에 적합한 대상체는 조류 (예를 들어, 닭, 오리, 거위, 메추라기, 칠면조 및 꿩) 및 포유동물 (예를 들어, 인간, 소, 양, 염소, 말, 고양이, 개 및 토끼목) 둘 다를 포함하며, 포유동물이 바람직하다. 인간 대상체가 가장 바람직하다. 인간 대상체에는 신생아, 유아, 청소년 및 성인이 포함된다.
본원에 기재된 기술의 한 측면은 이식유전자를 세포로 전달하는 방법에 관한 것이다. 전형적으로, 시험관내 방법의 경우, ceDNA 벡터는 본원에 개시된 방법, 뿐만 아니라 당 업계에 공지된 다른 방법을 사용하여 세포 내로 도입될 수 있다. 본원에 개시된 ceDNA 벡터는 바람직하게는 생물학적-유효량으로 세포에 투여된다. ceDNA 벡터가 생체내 세포 (예를 들어, 대상체)로 투여되는 경우, ceDNA 벡터의 생물학적-유효량은 표적 세포에서 이식유전자의 형질도입 및 발현을 야기하기에 충분한 양이다.
B. 투여 범위
생체내 및/또는 시험관내 분석은 선택적으로 사용을 위한 최적의 투약 범위를 식별하는데 도움을 주기 위해 사용될 수 있다. 제형에 사용되는 정확한 용량은 또한 투여 경로 및 상태의 심각성에 따라 달라질 것이며, 당업자의 판단 및 각 대상체의 상황에 따라 결정되어야 한다. 시험관내 또는 동물 모델 시험 시스템으로부터 유도된 용량-반응 곡선으로부터 유효 용량이 추론될 수 있다.
ceDNA 벡터는 원하는 조직의 세포를 형질감염시키고 과도한 부작용 없이 충분한 수준의 유전자 전달 및 발현을 제공하기에 충분한 양으로 투여된다. 통상적이고 약제학적으로 허용되는 투여 경로는 선택된 기관으로의 직접 전달 (예를 들어, 간으로의 문맥내 전달), 경구, 흡입 (비강내 및 기관내 전달 포함), 안내, 정맥내, 근육내, 피하, 피내, 종양내 및 기타 모 투여 경로와 같이 상기 "투여" 섹션에서 상기에 기재된 것을 포함하지만, 이에 제한되지 않는다. 원하는 경우, 투여 경로를 조합할 수 있다.
특정 "치료 효과"를 달성하기 위해 요구되는 ceDNA 벡터의 양의 용량은 핵산 투여 경로, 치료 효과를 달성하기 위해 요구되는 유전자 또는 RNA 발현 수준, 치료되는 특정 질환 또는 장애, 및 유전자(들), RNA 산물(들), 또는 생성된 발현 단백질(들)의 안정성을 포함하지만, 이에 제한되지 않는 몇몇 요인에 기초하여 변할 것이다. 당업자는 상기 언급된 인자, 및 당 업계에 잘 알려진 다른 인자에 기초하여 특정 질환 또는 장애를 갖는 환자를 치료하기 위한 ceDNA 벡터 용량 범위를 쉽게 결정할 수 있다.
투약 요법은 최적의 치료 반응을 제공하도록 조정될 수 있다. 예를 들어, 올리고뉴클레오티드는 반복적으로 투여될 수 있으며, 예를 들어, 수회 용량이 매일 투여될 수 있거나, 치료 상황의 긴급성으로 지시되는 바와 같이 용량을 비례적으로 감소시킬 수 있다. 당업자는 올리고 뉴클레오티드가 세포 또는 대상체에게 투여되는지에 관계없이 대상 올리고뉴클레오티드의 적절한 용량 및 투여 스케줄을 쉽게 결정할 수 있을 것이다.
"치료적 유효 용량"은 임상 시험을 통해 결정될 수 있는 비교적 넓은 범위에 속할 것이고, 특정 적용에 따라 달라질 것이다 (신경 세포는 매우 소량을 필요로 하는 한편, 전신 주사는 다량을 필요로 할 것임). 예를 들어, 인간 대상체의 골격 또는 심장 근육으로의 생체내 직접 주사의 경우, 치료적 유효 용량은 약 1 μg 내지 100 g 정도의 ceDNA 벡터일 것이다. 엑소좀 또는 미세입자가 ceDNA 벡터를 전달하기 위해 사용되는 경우, 치료적 유효 용량은 실험적으로 결정될 수 있지만, 1 μg 내지 약 100 g의 벡터를 전달할 것으로 예상된다. 또한, 치료적 유효 용량은 질환의 하나 이상의 증상을 감소시키지만, 오프-표적 유전자의 유전자 편집을 초래하지 않는 표적 유전자를 편집하는데 영향을 줄 정도로 충분한 양의 유전자 편집 분자를 발현하는 ceDNA 벡터의 양이다.
약제학적으로-허용되는 부형제 및 담체 용액의 제형은 다양한 치료 요법에서 본원에 기재된 특정 조성물을 사용하기 위한 적합한 투여 및 치료 요법의 개발과 같이 당업자에게 잘 알려져 있다.
시험관내 형질감염의 경우, 세포 (1x106 세포)로 전달되는 ceDNA 벡터의 유효량은 0.1 내지 100 μg ceDNA 벡터, 바람직하게는 1 내지 20 μg, 보다 바람직하게는 1 내지 15 μg 또는 8 내지 10 μg 정도일 것이다. 더 큰 ceDNA 벡터는 더 높은 용량을 요구할 것이다. 엑소좀 또는 미세입자가 사용되는 경우, 효과적인 시험관내 용량은 실험적으로 결정될 수 있지만 일반적으로 동일한 양의 ceDNA 벡터를 전달하도록 의도될 것이다.
치료는 단일 용량 또는 다중 용량의 투여를 포함할 수 있다. 일부 구현예에서, 하나 초과의 용량이 대상체에게 투여될 수 있으며; 실제로, ceDNA 벡터가 바이러스 캡시드의 부재로 인해 항-캡시드 숙주 면역 반응을 유발하고 유발하지 않기 때문에, 필요에 따라 다중 용량이 투여될 수 있다. 이와 같이, 당업자는 적절한 용량의 수를 쉽게 결정할 수 있다. 투여되는 용량의 수는, 예를 들어, 1 내지 100, 바람직하게는 2 내지 20개 정도의 용량일 수 있다.
임의의 특정 이론에 구속되지 않고, 본원에 기재된 바와 같은 ceDNA 벡터의 투여에 의해 유발된 전형적인 항-바이러스 면역 반응의 부재 (즉, 캡시드 성분의 부재)는 ceDNA 벡터가 숙주에 여러 번 투여될 수 있게 한다. 일부 구현예에서, 이종 핵산이 대상체에게 전달되는 경우의 수는 2 내지 10배 (예를 들어, 2, 3, 4, 5, 6, 7, 8, 9, 또는 10배) 범위이다. 일부 구현예에서, ceDNA 벡터는 10회 넘게 대상체에게 전달된다.
일부 구현예에서, ceDNA 벡터의 용량은 캘린더 일(calendar day) (예를 들어, 24시간 기간)당 1회 이하로 대상에게 투여된다. 일부 구현예에서, ceDNA 벡터의 용량은 2, 3, 4, 5, 6 또는 7 캘린더 일당 1회 이하로 대상체에게 투여된다. 일부 구현예에서, ceDNA 벡터의 용량은 캘린더 주(calendar week) (예를 들어, 7 캘린더 주)당 1회 이하로 대상체에게 투여된다. 일부 구현예에서, ceDNA 벡터의 용량은 격주 이내에 (예를 들어, 2 캘린더 주 주기 중 1회) 대상체에게 투여된다. 일부 구현예에서, ceDNA 벡터의 용량은 캘린더 개월(calendar month)당 1회 이하로 (예를 들어, 30 캘린더 일 내에 1회) 대상체에게 투여된다. 일부 구현예에서, ceDNA 벡터의 용량은 6 캘린더 개월당 1회 이하로 대상체에게 투여된다. 일부 구현예에서, ceDNA 벡터의 용량은 캘린더 년(calendar year) (예를 들어, 365일 또는 윤년에는 366일)당 1회 이하로 대상체에게 투여된다.
C. 단위 투여 형태
일부 구현예에서, 약제학적 조성물은 단위 투여 형태로 편리하게 제공될 수 있다. 단위 투여 형태는 전형적으로 약제학적 조성물의 하나 이상의 특정 투여 경로에 적합할 것이다. 일부 구현예에서, 단위 투여 형태는 흡입에 의한 투여에 적합하다. 일부 구현예에서, 단위 투여 형태는 기화기에 의한 투여에 적합하다. 일부 구현예에서, 단위 투여 형태는 분무기에 의한 투여에 적합하다. 일부 구현예에서, 단위 투여 형태는 에어로졸에 의한 투여에 적합하다. 일부 구현예에서, 단위 투여 형태는 경구 투여, 협측 투여 또는 설하 투여에 적합하다. 일부 구현예에서, 단위 투여 형태는 정맥내, 근육내 또는 피하 투여에 적합하다. 일부 구현예에서, 단위 투여 형태는 경막내 또는 뇌실내 투여에 적합하다. 일부 구현예에서, 약제학적 조성물은 국소 투여용으로 제형화된다. 단일 투여 형태를 생성하기 위해 담체 물질과 조합될 수 있는 활성 성분의 양은 일반적으로 치료 효과를 생성하는 화합물의 양일 것이다.
XIV. 다양한 응용
본원에 제공된 조성물 및 ceDNA 벡터는 상기 기재된 바와 같은 다양한 목적을 위한 유전자 편집 분자를 전달하는데 사용될 수 있다. 일부 구현예에서, 유전자 편집 분자는 표적 유전자, 예를 들어, 단백질 또는 기능성 RNA를 표적화하고, 이는 연구 목적으로, 예를 들어, 표적 유전자의 기능을 연구하기 위한, 예를 들어, 하나 이상의 돌연변이 또는 교정된 유전자 서열을 보유하는 체세포성 형질전환 동물 모델을 생성하기 위해 편집되어야 한다. 또 다른 예에서, 유전자 편집 분자는 질환의 동물 모델을 생성하기 위해 단백질 또는 기능적 RNA를 인코딩하는 표적 유전자를 유전자 편집하는데 사용된다.
일부 구현예에서, 유전자 편집 분자의 표적 유전자는 포유동물 대상체에서 질환 상태의 치료, 개선 또는 예방에 유용한, 하나 이상의 펩티드, 폴리펩티드 또는 단백질을 인코딩한다. 유전자 편집 분자는 비정상적인 유전자 서열과 관련된 질환을 치료하기에 충분한 양으로 ceDNA 벡터를 통해 환자에게 전달 (예를 들어, 발현)될 수 있으며, 이는 표적 유전자의 발현 결핍 또는 기능 장애 중 하나 이상을 초래할 수 있다.
일부 구현예에서, 진단 및 스크리닝 방법에 사용하기 위해, 유전자 편집 분자가 세포 배양 시스템, 또는 대안적으로 형질전환 동물 모델에서 일시적으로 또는 안정적으로 발현되는 ceDNA 벡터가 구상된다.
본원에 기재된 기술의 또 다른 측면은 포유동물 세포 집단을 형질도입시키는 방법을 제공한다. 전체적이고 일반적인 의미에서, 본 방법은 적어도, 집단의 하나 이상의 세포에, 본원에 개시된 ceDNA 중 하나 이상의 유효량을 포함하는 조성물을 도입하는 단계를 포함한다.
추가로, 본 발명은 하나 이상의 개시된 ceDNA 벡터 또는 ceDNA 조성물을 포함하거나, 하나 이상의 추가 성분으로 제형화되거나, 또는 이들의 사용을 위한 하나 이상의 설명서와 함께 준비된 조성물, 및 치료 및/또는 진단 키트를 제공한다.
본원에 개시된 바와 같은 ceDNA 벡터가 투여되는 세포는 신경 세포 (말초 및 중추 신경계 세포, 특히 뇌 세포 포함), 폐 세포, 망막 세포, 상피 세포 (예를 들어, 장 및 호흡기 상피 세포), 근육 세포, 수지상 세포, 췌장 세포 (섬 세포 포함), 간 세포, 심근 세포, 골 세포 (예를 들어, 골수 줄기세포), 조혈 줄기세포, 비장 세포, 각질 세포, 섬유 모세포, 내피 세포, 전립선 세포, 생식 세포 등을 포함하지만 이에 제한되지 않는 임의의 유형일 수 있다. 대안적으로, 세포는 임의의 전구 세포일 수 있다. 추가의 대안으로서, 세포는 줄기세포 (예를 들어, 신경 줄기세포, 간 줄기세포)일 수 있다. 또 다른 대안으로서, 세포는 암 또는 종양 세포일 수 있다. 더욱이, 세포는, 상기 나타낸 바와 같이, 임의의 기원의 종으로부터 유래될 수 있다.
일부 구현예에서, 본 출원은 하기 단락 중 임의의 단락에서 정의될 수 있다:
1. ceDNA 벡터로서, (i) 적어도 하나의 변경된 AAV 역 말단 반복부 (ITR); 및 (ii) 5' 상동성 아암, 공여체 서열 및 3' 상동성 아암을 포함하는 제1 뉴클레오티드 서열을 포함하고, 여기서 적어도 상기 공여체 서열은 유전자 편집 기능성을 갖는, ceDNA 벡터.
2. 제1 뉴클레오티드 서열은, 제1 뉴클레오티드 서열의 상류에 있는 제2 뉴클레오티드를 추가로 포함하고, 여기서 제2 뉴클레오티드 서열은 유전자의 조절 서열 및 뉴클레아제를 인코딩하는 뉴클레오티드 서열을 포함하고, 상기 유전자 조절 서열은 상기 뉴클레아제를 인코딩하는 뉴클레오티드 서열에 작동 가능하게 연결되는, 단락 1의 ceDNA 벡터.
3. 뉴클레아제가 서열-특이적 뉴클레아제인, 단락 1-2 중 어느 것의 ceDNA 벡터.
4. 서열-특이적 뉴클레아제가 RNA-유도된 뉴클레아제, 아연 핑거 뉴클레아제 (ZFN) 또는 전사 활성화제-유사 이펙터 뉴클레아제 (TALEN)인, 단락 1-3 중 어느 것의 ceDNA 벡터.
5. RNA-유도된 뉴클레아제가 Cas 또는 Cas9인, 단락 1-4 중 어느 것의 ceDNA 벡터.
6. 조절 서열이 인핸서 및 프로모터를 포함하고, 여기서 제2 핵산 서열은 뉴클레아제를 인코딩하는 뉴클레오티드 서열의 상류에 있는 인트론 서열을 포함하고, 인트론은 뉴클레아제 절단 부위를 포함하고, 프로모터는 뉴클레아제를 인코딩하는 뉴클레오티드 서열에 작동 가능하게 연결되는, 단락 1-5 중 어느 것의 ceDNA 벡터.
7. 가이드 서열 및/또는 활성화 RNA 서열을 인코딩하는 뉴클레오티드 서열을 포함하는 제3 뉴클레오티드 서열을 추가로 포함하는, 단락 1-6 중 어느 것의 ceDNA 벡터.
8. 제3 뉴클레오티드 서열이 가이드 서열 및/또는 활성화 RNA 서열을 인코딩하는 뉴클레오티드 서열에 작동 가능하게 연결된 프로모터를 추가로 포함하는, 단락 1-7 중 어느 것의 ceDNA 벡터.
9. 폴리-A 부위가 상기 상동성 아암의 상류에 있고 이에 근접한, 단락 1-8 중 어느 것의 ceDNA 벡터.
10. 공여체 서열이 5' 상동성 아암 또는 3' 상동성 아암과 이질적인, 단락 1-9 중 어느 것의 ceDNA 벡터.
11. 5' 상동성 아암이 염색체 상의 뉴클레아제 절단 부위의 상류에 있는 뉴클레오티드 서열과 상동성인, 단락 1-10 중 어느 것의 ceDNA 벡터.
12. 3' 상동성 아암이 염색체 상의 뉴클레아제 절단 부위의 하류에 있는 뉴클레오티드 서열과 상동성인 단락 1-11 중 어느 것의 ceDNA 벡터.
13. 5' 상동성 아암 또는 3' 상동성 아암이 적어도 하나의 변경된 ITR에 근접한, 단락 1-12 중 어느 것의 ceDNA 벡터.
14. 5' 상동성 아암 및 3' 상동성 아암이 약 250 내지 2000 bp인, 단락 1-13 중 어느 것의 ceDNA 벡터.
15. 뉴클레아제를 인코딩하는 뉴클레오티드 서열이 ceDNA인, 단락 1-14 중 어느 것인, ceDNA 벡터.
16. 프로모터가 CAG 프로모터인, 단락 1-15 중 어느 것의 ceDNA 벡터.
17. 프로모터가 Pol III, U6 또는 H1인, 단락 1-17 중 어느 것의 ceDNA 벡터.
18. 숙주 세포에서 염색체 상의 미리 결정된 삽입 부위에 공여체 서열을 삽입하는 방법으로서, 적어도 하나의 변경된 ITR을 갖는 ceDNA 벡터를 숙주 세포에 도입하는 단계를 포함하고, 여기서 상기 ceDNA 벡터는 5' 상동성 아암, 공여체 서열, 및 3' 상동성 아암을 포함하는 뉴클레오티드 서열을 포함하며, 상기 공여체 서열은 상동성 재조합을 통해 삽입 부위에서 또는 삽입 부위에 인접한 염색체 내로 삽입되는, 방법.
19. 삽입 부위를 인식하는 가이드 RNA (gRNA)를 인코딩하는 뉴클레오티드 서열을 세포에 도입하는 단계를 추가로 포함하는, 단락 18의 방법.
20. 삽입 부위에서 또는 삽입 부위에 인접한 염색체를 절단하는 서열-특이적 뉴클레아제를 인코딩하는 뉴클레오티드 서열을 세포에 도입하는 단계를 추가로 포함하는, 단락 18 또는 19의 방법.
21. 서열-특이적 뉴클레아제가 RNA-유도된 뉴클레아제, 아연 핑거 뉴클레아제 (ZFN) 또는 전사 활성화제-유사 이펙터 뉴클레아제 (TALEN)인, 단락 18-20 중 어느 것의 방법.
22. RNA-유도된 뉴클레아제가 Cas 또는 Cas9인, 단락 18-21 중 어느 것의 방법.
23. 도입 단계는 캡시드가 없는, 단락 18-22 중 어느 것의 방법.
24. 5' 상동성 아암이 염색체 상의 뉴클레아제 절단 부위의 상류에 있는 서열과 상동성인, 단락 18-23 중 어느 것의 방법.
25. 3' 상동성 아암이 염색체 상의 뉴클레아제 절단 부위의 하류에 있는 서열과 상동성인, 단락 18-24 중 어느 것의 방법.
26. 5' 상동성 아암 또는 3' 상동성 아암이 변경된 ITR에 근접한, 단락 18 중 어느 것의 방법.
27. 5' 상동성 아암 및 3' 상동성 아암이 적어도 약 50-2000 염기쌍인, 단락 18-26 중 어느 것의 방법.
28. 상기 뉴클레오티드 서열이 5' 상동성 아암의 상류에 있는 5' 플랭킹 서열 및 3' 상동성 아암의 하류에 있는 3' 플랭킹 서열을 추가로 포함하는, 단락 18-27 중 어느 것의 방법.
29. 동물의 염색체 상의 미리 결정된 삽입 부위에 삽입된 공여체 서열을 포함하는 유전자 변형된 동물의 생성 방법으로서, a) 단락 18에 따라 염색체 상의 미리 결정된 삽입 부위에 공여체 서열이 삽입된 세포를 생성하는 단계; 및 b) a)에 의해 생성된 세포를 운반 동물에 도입하여 유전자 변형된 동물을 생산하는 단계를 포함하는, 방법.
30. 세포가 접합자 또는 다능성 줄기세포인, 단락 29의 방법.
31. 단락 29 또는 30의 방법에 의해 생성된 유전자 변형된 동물.
32. 세포에서 염색체 상의 삽입 부위에 공여체 서열을 삽입하기 위한 키트로서, (a) 다음을 포함하는 제1 ceDNA 벡터: (i) 적어도 하나의 변경된 AAV 역 말단 반복부 (ITR); 및 (ii) 5' 상동성 아암, 공여체 서열 및 3' 상동성 아암을 포함하는 제1 뉴클레오티드 서열로서, 상기 공여체 서열은 유전자 편집 기능성을 갖는, 제1 뉴클레오티드 서열; 및 (b) 다음을 포함하는 제2 ceDNA 벡터: (i) 적어도 하나의 변경된 AAV 역 말단 반복부 (ITR); 및 (ii) 뉴클레아제를 인코딩하는 뉴클레오티드 서열을 포함하고, 여기서 5' 상동성 아암은 염색체 상의 뉴클레아제 절단 부위의 상류에 있는 서열과 상동성이고, 3' 상동성 아암은 염색체 상의 뉴클레아제 절단 부위의 하류에 있는 서열과 상동성이고; 5' 상동성 아암 또는 3' 상동성 아암은 변경된 ITR에 근접한, 키트.
33. 숙주 세포의 염색체 상의 미리 결정된 삽입 부위에 공여체 서열을 삽입하는 방법으로서, (a) 적어도 하나의 변경된 ITR을 갖는 제1 ceDNA 벡터를 숙주 세포에 도입하는 단계로서, ceDNA 벡터는 5' 상동성 아암, 공여체 서열 및 3' 상동성 아암을 포함하는 제1 선형 핵산을 포함하는, 상기 단계; 및 (b) 적어도 하나의 변경된 ITR을 갖는 제2 ceDNA 벡터를 숙주 세포에 도입하는 단계로서, 상기 제2 ceDNA 벡터는 삽입 부위에서 또는 삽입 부위에 인접한 염색체를 절단하는 서열-특이적 뉴클레아제를 인코딩하는 뉴클레오티드 서열을 포함하는 제2 선형 핵산을 포함하고, 상기 공여체 서열은 상동성 재조합을 통해 삽입 부위에서 또는 삽입 부위에 인접한 염색체 내로 삽입되는, 방법.
34. 제2 ceDNA 벡터가 삽입 부위를 인식하는 가이드 서열을 인코딩하는 제3 뉴클레오티드 서열을 추가로 포함하는, 단락 18-33 중 어느 것의 방법.
35. 이식유전자 향상 요소, 및 3' 상동성 아암의 하류에 있고 이에 근접한 폴리-A 사이트(cite) 중 적어도 하나를 추가로 포함하는, 단락 1-17 중 어느 것의 ceDNA 벡터.
36. 변경된 ITR에 근접한 대안적인 뉴클레아제 표적 서열을 추가로 포함하는, 단락 1-17 또는 35 중 어느 것의 ceDNA 벡터.
37. 3' 상동성 아암의 상류에 있고 이에 근접한 2A 및 선별 마커를 추가로 포함하는, 단락 1-17 또는 35-36 중 어느 것의 ceDNA 벡터.
38. ceDNA 핵산 벡터 조성물로서, 플랭킹 말단 반복부 (TR); 및 적어도 하나의 유전자 편집 핵산 서열을 포함하고, 여기서 벡터는 선형 폐쇄-말단 듀플렉스 DNA인, ceDNA 핵산 벡터 조성물.
39. 말단 반복부가 역 TR (ITR)인, 단락 38의 조성물.
40. 말단 반복부 중 적어도 하나가 변형되는, 단락 38 또는 39의 조성물.
41. 벡터가 핵산 변성 조건 하에 단일 가닥 원형 DNA인, 단락 38-40 중 어느 것의 조성물.
42. 유전자 편집 핵산 서열이 서열 특이적 뉴클레아제, 하나 이상의 가이드 RNA, CRISPR/Cas, 리보핵단백질 (RNP), 또는 CRISPRi 또는 CRISPRa 시스템을 위한 불활성화된 CAS, 또는 이들의 임의의 조합으로 구성된 군으로부터 선택된 유전자 편집 분자를 인코딩하는, 단락 38-41 중 어느 것의 조성물.
43. 서열-특이적 뉴클레아제가 TAL-뉴클레아제, 아연-핑거 뉴클레아제 (ZFN), 메가뉴클레아제, megaTAL 또는 RNA 유도된 엔도뉴클레아제 (예를 들어, CAS9, cpf1, dCAS9, nCAS9)를 포함하는, 단락 38-42 중 어느 것의 조성물.
44. 적어도 2개의 변형된 ITR을 추가로 포함하는, 단락 38-43 중 어느 것의 조성물.
45. 관심 핵산을 추가로 포함하는, 단락 38-44 중 어느 것의 조성물.
46. 유전자 편집 핵산 서열이 상동성-지정 복구 주형인, 단락 38-45 중 어느 것의 조성물.
47. 상동성-지정 복구 주형이 5' 상동성 아암, 공여체 서열 및 3' 상동성 아암을 포함하는, 단락 38-46 중 어느 것의 조성물.
48. 엔도뉴클레아제를 인코딩하는 핵산 서열을 추가로 포함하고, 여기서 엔도뉴클레아제는 표적 유전자의 DNA 상의 특정 엔도뉴클레아제 부위 또는 ceDNA 벡터 상의 표적 부위에서 절단되거나 닉킹되는, 단락 38-47 중 어느 것의 조성물.
49. 5' 상동성 아암이 염색체 상의 DNA 엔도뉴클레아제 부위의 상류에 있는 뉴클레오티드 서열과 상동성인, 단락 38-48 중 어느 것의 조성물.
50. 3' 상동성 아암이 DNA 엔도뉴클레아제 부위의 하류에 있는 뉴클레오티드 서열에 상동성인, 단락 38-49 중 어느 것의 조성물.
51. 상동성 아암이 각각 약 250 내지 2000bp인, 단락 38-40 중 어느 것의 조성물.
52. DNA 엔도뉴클레아제가 TAL-뉴클레아제, 아연-핑거 뉴클레아제 (ZFN) 또는 RNA 유도된 엔도뉴클레아제 (예를 들어, Cas9 또는 Cpf1)를 포함하는, 단락 38-52 중 어느 것의 조성물.
53. RNA 유도된 엔도뉴클레아제가 Cas 효소를 포함하는, 단락 38-52 중 어느 것의 조성물.
54. Cas 효소가 Cas9인, 단락 38-53 중 어느 것의 조성물.
55. Cas 효소가 닉킹 Cas9 (nCas9)인, 단락 38-53 중 어느 것의 조성물.
56. nCas9가 Cas의 HNH 또는 RuVc 도메인에서 돌연변이를 포함하는, 단락 38-55 중 어느 것의 조성물.
57. Cas 효소가 표적 유전자의 프로모터 영역을 표적화하는 gRNA와 복합체를 형성하는 불활성화된 Cas 뉴클레아제 (dCas)인, 단락 38-53 중 어느 것의 조성물.
58. KRAB 이펙터 도메인을 추가로 포함하는, 단락 38-57 중 어느 것의 조성물.
59. dCas가 프로모터 영역으로 향할 수 있는 이종 전사 활성화 도메인에 융합된, 단락 38-57 중 어느 것의 조성물.
60. dCas 융합이 표적 유전자의 발현을 상향 조절하기 위해 추가의 전사 활성화 도메인을 동원하는 가이드 RNA에 의해 표적 유전자의 프로모터 영역으로 향하는, 단락 38-59 중 어느 것의 조성물.
61. dCas가 S. 파이오게네스 dCas9인, 단락 38-57 중 어느 것의 조성물.
62. 가이드 RNA 서열이 표적 유전자의 프로모터의 근접부를 표적화하고, CRISPR이 표적 유전자를 침묵시키는 (CRISPRi 시스템), 단락 38-61 중 어느 것의 조성물.
63. 가이드 RNA 서열이 표적 유전자의 전사 개시 부위를 표적화하고 표적 유전자를 활성화시키는 (CRISPRa 시스템), 단락 38-61 중 어느 것의 조성물.
64. RNA-유도된 DNA 엔도뉴클레아제에 대한 적어도 하나의 가이드 RNA (gRNA)를 인코딩하는 핵산을 추가로 포함하는, 단락 38-63 중 어느 것의 조성물.
65. 가이드 RNA (sgRNA)가 결함 유전자의 스플라이스 수용체 또는 스플라이스 공여체 부위를 표적화하여 비-동종 말단 결합 (NHEJ) 및 결함 유전자의 교정을 초래하는, 단락 38-64 중 어느 것의 조성물.
66. 벡터가 하나의 가이드 RNA 서열의 다중 카피를 인코딩하는, 단락 38-65 중 어느 것의 조성물.
67. 뉴클레아제를 인코딩하는 핵산 서열에 작동 가능하게 연결된 조절 서열을 추가로 포함하는, 단락 38-66 중 어느 것의 조성물.
68. 조절 서열은 인핸서 및/또는 프로모터를 포함하는, 단락 38-67 중 어느 것의 조성물.
69. 프로모터가 DNA 엔도뉴클레아제를 인코딩하는 핵산 서열에 작동 가능하게 연결되고, 여기서 DNA 엔도뉴클레아제를 인코딩하는 핵산 서열은 엔도뉴클레아제 서열의 상류에 있는 인트론 서열을 추가로 포함하고, 인트론은 뉴클레아제 절단 부위를 포함하는, 단락 38-68 중 어느 것의 조성물.
70. 폴리-A-부위는 5' 상동성 아암의 상류에 있고 이에 근접한, 단락 38-69 중 어느 것의 조성물.
71. 상기 공여체 서열이 5' 상동성 아암 또는 3' 상동성 아암과 이질적인, 단락 47*** 중 어느 것의 조성물.
72. 5' 상동성 아암 또는 3' 상동성 아암이 적어도 하나의 변형된 ITR에 근접한, 단락 47 중 어느 것의 조성물.
73. 뉴클레아제를 인코딩하는 뉴클레오티드 서열이 cDNA인, 단락 48 중 어느 것의 조성물.
74. 프로모터가 CAG 프로모터인, 단락 68 중 어느 것의 조성물.
75. 프로모터가 Pol III, U6 또는 H1인, 단락 68 중 어느 것의 조성물.
76. 단락 38-75 중 어느 것의 벡터를 포함하는 세포.
77. 단락 38-75 중 어느 것의 벡터 및 지질을 포함하는 조성물.
78. 단락 38-75 중 어느 것의 벡터, 또는 단락 76의 세포를 포함하는 키트.
79. 세포를 유전자 편집 시스템과 접촉시키는 단계를 포함하는 유전자 편집 방법으로서, 상기 유전자 편집 시스템의 하나 이상의 성분은 세포를 폐쇄-말단 DNA (ceDNA) 핵산 벡터 조성물과 접촉시킴으로써 세포로 전달되고, 상기 ceDNA 핵산 벡터 조성물은 플랭킹 말단 반복 (TR) 서열, 및 선택적으로 적어도 하나의 표적 유전자에 상보적인 영역을 갖는 적어도 하나의 유전자 편집 핵산 서열을 포함하는 선형 폐쇄-말단 듀플렉스 DNA인, 방법.
80. 말단 반복부가 역 TR (ITR)인, 단락 79 방법.
81. ITR이 변형된 ITR인, 단락 79 또는 80의 방법.
82. 유전자 편집 시스템이 TALEN 시스템, 아연-핑거 엔도뉴클레아제 (ZFN) 시스템, CRISPR/Cas 시스템 및 메가뉴클레아제 시스템으로 구성된 군으로부터 선택되는, 방법.
83. 적어도 하나의 유전자 편집 핵산 서열이 RNA 유도 뉴클레아제, 가이드 RNA, TALEN 및 아연 핑거 엔도뉴클레아제 (ZFN)로 구성된 군으로부터 선택된 유전자 편집 분자를 인코딩하는, 방법.
84. 단일 ceDNA 벡터가 유전자 편집 시스템의 모든 성분을 포함하는, 단락 79-83 중 어느 것의 방법.
85. 세포를 접촉시키는 단계는 유전자 편집 시스템과 함께 형질감염 시약 또는 지질 시약을 투여하는 단계를 추가로 포함하는, 방법.
86. 유전자 편집 시스템이 형질감염 시약 또는 리포좀 시약을 추가로 포함하는, 단락 79-85 중 어느 것의 방법.
87. ceDNA 핵산 벡터 조성물이 단락 1-77 중 어느 하나인, 단락 79-86 중 어느 것의 방법.
88. 표적 유전자의 발현이 변경되는, 단락 79-87 중 어느 것의 방법.
89. 접촉된 세포가 진핵 세포인, 단락 79-88 중 어느 것의 방법.
90. Cas 단백질이 진핵 세포에서의 발현에 대해 코돈 최적화된, 단락 79-88 중 어느 것의 방법.
91. 표적 유전자를 편집하기에 적합한 조건 하에서 및 충분한 시간 동안, 단락 1-77 중 어느 것의 ceDNA 조성물의 유효량을 세포에 투여하는 단계를 포함하는, 게놈 편집 방법.
92. 표적 유전자가 하나 이상의 가이드 RNA 서열을 사용하여 표적화되고 HDR 공여체 주형의 존재하에 상동성 지정 복구 (HDR)에 의해 편집되는 유전자인, 단락 79-91 중 어느 것의 방법.
93. 표적 유전자가 하나의 가이드 RNA 서열을 사용하여 표적화되고 표적 유전자는 비-동종 말단 결합 (NHEJ)에 의해 편집되는, 단락 79-91 중 어느 것의 방법.
94. 상기 방법이 질환과 관련된 단일 뉴클레오티드 다형성 (SNP)을 교정하기 위해 생체내에서 수행되는, 단락 79-93 중 어느 것의 방법.
95. 질환이 겸상 적혈구 빈혈증, 유전성 혈색소증 또는 암 유전성 실명을 포함하는, 단락 94의 방법.
96. 적어도 2개의 상이한 Cas 단백질이 ceDNA 벡터에 존재하며, Cas 단백질 중 하나는 촉매적으로 비활성이고 (Cas-i), Cas-I와 관련된 가이드 RNA는 표적 세포의 프로모터를 표적화하고, Cas-I를 코딩하는 DNA는 유도성 프로모터의 제어 하에 있어서 원하는 시간에 표적 유전자의 발현을 끌 수 있는, 단락 91의 방법.
97. 세포의 표적 유전자에서 단일 뉴클레오티드 염기쌍을 편집하는 방법으로서, 세포를 CRISPR/Cas 유전자 편집 시스템과 접촉시키는 단계를 포함하고, 여기서 CRISPR/Cas 유전자 편집 시스템의 하나 이상의 성분은 세포를 폐쇄-말단 DNA (ceDNA) 핵산 벡터 조성물과 접촉시킴으로써 세포로 전달되고, 상기 ceDNA 핵산 벡터 조성물은 플랭킹 말단 반복 (TR) 서열 및 적어도 하나의 표적 유전자 또는 표적 유전자에 대한 조절 서열에 상보적인 영역을 갖는 적어도 하나의 유전자 편집 핵산 서열을 포함하는 선형 폐쇄-말단 듀플렉스 DNA이고,
상기 벡터로부터 발현되는 Cas 단백질은 촉매적으로 비활성이고, 염기 편집 모이어티에 융합되고,
상기 방법은 표적 유전자의 발현을 조절하기에 충분한 조건 하에서 및 시간 동안 수행되는, 방법.
98. 말단 반복부는 역 TR (ITR)인, 단락 79-97 중 어느 것의 방법.
99. 플랭킹 말단 반복부 중 적어도 하나가 변형된 말단 반복부인, 단락 79-98 중 어느 것의 방법.
100. 염기 편집 모이어티가 단일-가닥-특이적 시티딘 데아미나제, 우라실 글리코실라제 억제제 또는 tRNA 아데노신 데아미나제를 포함하는, 방법.
101. 벡터로부터 발현된 촉매적으로 비활성 Cas 단백질이 dCas9인, 단락 79-100 중 어느 것의 방법.
102. ceDNA 벡터가 단락 1-77 중 어느 것의 구조를 가지며, 여기서 접촉된 세포는 T 세포 또는 CD34+인, 단락 79-101 중 어느 것의 방법.
103. 표적 유전자가 프로그래밍된 사멸 단백질 (PD1), 세포독성 T-림프구-관련 항원 4 (CTLA4), 또는 종양 괴사 인자-α (TNF-α)를 인코딩하는, 단락 79-102 중 어느 것의 방법.
104. 이를 필요로 하는 대상체에게 단락 102에 의해 생성된 세포를 투여하는 단계를 추가로 포함하는, 단락 79-103 중 어느 것의 방법.
105. 이를 필요로 하는 대상체는 유전 질환, 바이러스 감염, 박테리아 감염, 암 또는 자가면역 질환을 갖는, 단락 104의 방법.
106. 세포 내 2개 이상의 표적 유전자의 발현을 조절하는 방법으로서, 세포 내로 하기를 도입하는 단계를 포함하고:
(i) 플랭킹 말단 반복 (TR) 서열, 및 2개 이상의 표적 유전자에 상보적인 적어도 2개의 가이드 RNA를 인코딩하는 핵산 서열을 포함하는 벡터를 포함하는 조성물로서, 상기 벡터는 선형 폐쇄-말단 듀플렉스 DNA인, 상기 조성물,
(ii) 플랭킹 말단 반복 (TR) 서열, 및 각각 가이드 RNA와 회합하고 2개 이상의 표적 유전자에 결합하는 적어도 2개의 촉매적으로 비활성 DNA 엔도뉴클레아제를 인코딩하는 핵산 서열을 포함하는 벡터를 포함하는 제2 조성물로서, 상기 벡터는 선형 폐쇄-말단 듀플렉스 DNA인, 제2 조성물, 및
(iii) 플랭킹 말단 반복 (TR) 서열, 및 적어도 2개의 전사 조절 단백질 또는 도메인을 인코딩하는 핵산 서열을 포함하는 벡터를 포함하는 제3 조성물로서, 상기 벡터는 선형 폐쇄-말단 듀플렉스 DNA인, 제3 조성물,
여기서 적어도 2개의 가이드 RNA, 적어도 2개의 촉매적으로 비활성 RNA-유도 엔도뉴클레아제 및 적어도 2개의 전사 조절 단백질 또는 도메인은 세포에서 발현되며,
2개 이상의 공동-국재화 복합체는 가이드 RNA, 촉매적으로 비활성 RNA-유도 엔도뉴클레아제, 전사 조절 단백질 또는 도메인 및 표적 유전자 사이에서 형성되고,
상기 전사 조절 단백질 또는 도메인은 적어도 2개의 표적 유전자의 발현을 조절하는, 방법.
107. 말단 반복부가 역 TR (ITR)인, 단락 106의 방법.
108. 플랭킹 TR 서열 중 적어도 하나가 변형된 TR인, 단락 106 또는 107의 방법.
109. 게놈 세이프 하버 유전자에 핵산 서열을 삽입하는 방법으로서, 세포를 (i) 유전자 편집 시스템 및 (ii) 게놈 세이프 하버 유전자와 상동성을 갖고 관심 단백질을 인코딩하는 핵산 서열을 포함하는 상동성 지정 복구 주형과 접촉시키는 단계를 포함하고,
여기서 유전자 편집 시스템의 하나 이상의 성분은 세포를 폐쇄-말단 DNA (ceDNA) 핵산 벡터 조성물과 접촉시킴으로써 세포로 전달되고, 상기 ceDNA 핵산 벡터 조성물은 플랭킹 말단 반복 (TR) 서열 및 적어도 하나의 유전자 편집 핵산 서열을 포함하는 선형 폐쇄-말단 듀플렉스 DNA이고,
상기 방법은 관심 단백질을 인코딩하는 핵산 서열을 게놈 세이프 하버 유전자에 삽입하기에 충분한 조건 하에서 및 시간 동안 수행되는, 방법.
110. 말단 반복부가 역 TR (ITR)인, 단락 109의 방법.
111. 플랭킹 TR 서열 중 적어도 하나가 변형된 TR인, 단락 109 또는 110의 방법.
112. 게놈 세이프 하버 유전자가 높은 발현 수준으로 단백질을 발현시키는 것으로 알려진 적어도 하나의 코딩 서열에 가까운 활성 인트론을 포함하는, 단락 109-111 중 어느 것의 방법.
113. ceDNA 벡터가 단락 1-77 중 어느 하나에서와 같은 구조를 포함하는, 단락 109-112 중 어느 것의 방법.
114. 게놈 세이프 하버 유전자가 알부민 유전자 내 또는 그 근처의 부위를 포함하는, 단락 109-113 중 어느 것의 방법.
115. 관심 단백질이 수용체, 독소, 호르몬, 효소 또는 세포 표면 단백질인, 단락 109-114 중 어느 것의 방법.
116. 관심 단백질이 분비된 단백질인, 단락 109-115 중 어느 것의 방법.
117. 관심 단백질이 인자 VIII (FVIII) 또는 인자 IX (FIX)를 포함하는, 단락 109-116 중 어느 것의 방법.
118. 상기 방법이 혈우병 A 또는 혈우병 B의 치료를 위해 생체내에서 수행되는, 단락 109-117 중 어느 것의 방법.
실시예
하기 실시예는 제한이 아닌 예시로서 제공된다. 당업자는 ceDNA 벡터가 본원에 기재된 임의의 야생형 또는 변형된 ITR로부터 구축될 수 있고, 하기 예시적인 방법이 이러한 ceDNA 벡터의 활성을 구축 및 평가하는데 사용될 수 있음을 이해할 것이다. 상기 방법은 특정 ceDNA 벡터로 예시되지만, 설명에 따라 임의의 ceDNA 벡터에 적용 가능하다.
실시예 1: ceDNA 벡터의 구축
폴리뉴클레오티드 작제 주형을 사용한 ceDNA 벡터의 생성은 PCT/US18/49996의 실시예 1에 기재되어 있다. 예를 들어, 본 발명의 ceDNA 벡터를 생성하는데 사용되는 폴리뉴클레오티드 작제 주형은 ceDNA-플라스미드, ceDNA-박미드 및/또는 ceDNA-바쿨로바이러스일 수 있다. 이론에 제한되지 않고, 허용 숙주 세포에서, 예를 들어, Rep의 존재하에, 2개의 대칭 ITR 및 발현 작제물을 갖는 폴리뉴클레오티드 작제 주형 (여기서 ITR 중 적어도 하나는 야생형 ITR 서열에 대해 변형됨)은 ceDNA 벡터를 생성하기 위해 복제된다. ceDNA 벡터 생산은 두 단계, 즉, 우선, Rep 단백질을 통한 주형 골격 (예를 들어 ceDNA-플라스미드, ceDNA-박미드, ceDNA-바클리오바이러스(bacliovirus) 게놈 등)으로부터 주형의 절제 ("구조") 단계 및 둘째, 절제된 ceDNA 벡터의 Rep 매개 복제 단계를 거친다.
ceDNA 벡터를 생성하는 예시적인 방법은 본원에 기재된 ceDNA-플라스미드로부터의 방법이다. 도 1a 및 1b를 참조하면, 각각의 ceDNA-플라스미드의 폴리뉴클레오티드 작제 주형은 ITR 서열 사이에 다음을 갖는 좌측 변형된 ITR 및 우측 변형된 ITR 둘 모두를 포함한다: (i) 인핸서/프로모터; (ii) 이식유전자의 클로닝 부위; (iii) 전사 후 반응 요소 (예를 들어, 우드척 간염 바이러스 전사 후 조절 요소 (WPRE)); 및 (iv) 폴리아데닐화 신호 (예를 들어, 소 성장 호르몬 유전자 (BGHpA)로부터). 작제물 내의 특정 부위에 새로운 유전자 성분의 도입을 용이하게 하기 위해 고유한 제한 엔도뉴클레아제 인식 부위 (R1-R6) (도 1a 및 도 1b에 도시됨)가 또한 각 성분 사이에 도입되었다. R3 (PmeI) GTTTAAAC (서열 번호: 7) 및 R4 (PacI) TTAATTAA (서열 번호: 542) 효소 부위는 클로닝 부위로 조작되어 이식유전자의 오픈 리딩 프레임을 도입한다. 이들 서열을 ThermoFisher Scientific으로부터 입수한 pFastBac HT B 플라스미드 내로 클로닝하였다.
간략하게, 유전자 편집을 위한 일련의 ceDNA 벡터는 도 4a-4c에 도시된 과정을 사용하여 ceDNA-플라스미드 작제물로부터 수득되었다. 표 8은 본원에 사용하기 위한 유전자 편집 ceDNA 벡터를 생성하기 위한 예시적인 작제물을 나타내고, 이는 또한 서열, 예를 들어, 유전자 편집 분자에 작동 가능하게 연결된 프로모터의 한쪽 말단에 복제 단백질 부위 (RPS) (예를 들어, Rep 결합 부위)를 포함할 수 있다. 표 8의 숫자는 각 성분의 서열에 상응하는 본 문서의 서열 번호를 나타낸다. 표 8의 플라스미드는 서열 번호: 8을 포함하는 WPRE, 이어서 이식유전자 및 우측 ITR 사이의 3' 비번역 영역에서 서열 번호: 9를 포함하는 BGHpA로 구축되었다.
[표 8]
일부 구현예에서, 유전자 편집 ceDNA 벡터를 제조하기 위한 작제물은 본원에 기재된 바와 같은 조절 스위치인 프로모터, 예를 들어 유도성 프로모터를 포함한다.
ceDNA-박미드의 생산:
도 4a를 참조하면, DH10Bac 적격 세포 (MAX EFFICIENCY® DH10Bac™ Competent Cells, Thermo Fisher)를 제조사의 지시에 따른 프로토콜에 따라 시험 또는 대조군 플라스미드로 형질전환시켰다. DH10Bac 세포에서 플라스미드와 바쿨로바이러스 셔틀 벡터 간의 재조합을 유도하여 재조합 ceDNA-박미드를 생성하였다. 박미드 및 트랜스포사제 플라스미드의 유지 및 형질전환체를 위해 선택하는 항생제와 함께 X-gal 및 IPTG를 함유하는 박테리아 한천 플레이트 상에서 이. 콜라이(E. coli)에서의 청색-백색 스크리닝 (Φ80dlacZΔM15 마커는 박미드 벡터로부터 β-갈락토시다제 유전자의 α-상보성을 제공함)에 기초한 양성 선별을 스크리닝함으로써 재조합 박미드를 선별하였다. β-갈락토시드 지표 유전자를 파괴하는 전위에 의해 야기된 백색 콜로니를 선별하여 10 ml의 배지에서 배양하였다.
재조합 ceDNA-박미드를 이. 콜라이로부터 단리하고 FugeneHD를 사용하여 Sf9 또는 Sf21 곤충 세포로 형질감염시켜 감염성 바쿨로바이러스를 생성하였다. 부착성 Sf9 또는 Sf21 곤충 세포를 25℃에서 T25 플라스크 중의 50 ml의 배지에서 배양하였다. 4일 후, 배양 배지 (PO 바이러스 함유)를 세포로부터 제거하고, 0.45 μm 필터를 통해 여과하여, 감염성 바쿨로바이러스 입자를 세포 또는 세포 잔해로부터 분리하였다.
선택적으로, 제1 세대의 바쿨로바이러스 (P0)는 50 내지 500 ml의 배지에서 나이브 Sf9 또는 Sf21 곤충 세포를 감염시킴으로써 증폭되었다. 세포가 직경 18-19 nm (나이브 직경 14-15 nm), 및 밀도 ~4.0E+6 세포/mL에 도달할 때까지, 세포 직경 및 생존력을 모니터링하면서, 25℃에서 130 rpm으로 궤도 진탕기 인큐베이터에서 현탁 배양으로 세포를 유지시켰다. 감염 후 3일에서 8일 사이에, 배지에서 P1 바쿨로바이러스 입자를 원심분리 후 수집하여 세포 및 잔해물을 제거한 다음 0.45 μm 필터를 통해 여과하였다.
시험 작제물을 포함하는 ceDNA-바쿨로바이러스를 수집하고 바쿨로바이러스의 감염성 활성 또는 역가를 측정하였다. 구체적으로, 2.5E+6 세포/ml에서 4 x 20 ml Sf9 세포 배양물을 다음 희석에서 P1 바쿨로바이러스로 처리하고: 1/1000, 1/10,000, 1/50,000, 1/100,000, 25-27℃에서 인큐베이션하였다. 감염력은 4 내지 5일 동안 매일, 세포 직경 증가율 및 세포주기 정지율, 및 세포 생존력의 변화에 의해 결정되었다.
도 4a를 참조하면, 예를 들어, 도 7a에 따른 "Rep-플라스미드"는 Rep 78 (서열 번호: 13) 또는 Rep68 (서열 번호: 12) 및 Rep52 (서열 번호: 14) 또는 Rep40 (서열 번호: 11)을 포함하는 pFASTBAC™-이중 발현 벡터 (ThermoFisher)에서 생성되었다.
Rep-플라스미드를 제조자가 제공한 프로토콜에 따라 DH10Bac 적격 세포 (MAX EFFICIENCY® DH10Bac™ Competent Cells (Thermo Fisher)로 형질전환시켰다. DH10Bac 세포에서 Rep-플라스미드와 바쿨로바이러스 셔틀 벡터 간의 재조합을 유도하여 재조합 박미드 ("Rep-박미드")를 생성하였다. X-gal 및 IPTG를 함유하는 박테리아 한천 플레이트 상의 이. 콜라이에서의 청색-백색 스크리닝 (Φ80dlacZΔM15 마커는 박미드 벡터로부터 β-갈락토시다제 유전자의 α-상보성을 제공함)을 포함하는 양성 선별에 의해 재조합 박미드를 선별하였다. 단리된 백색 콜로니를 선별하고 10 ml의 선별 배지 (LB 브로쓰 중의 카나마이신, 겐타마이신, 테트라사이클린)에 접종하였다. 재조합 박미드 (Rep-박미드)를 이. 콜라이로부터 단리하고, Rep-박미드를 Sf9 또는 Sf21 곤충 세포에 형질감염시켜 감염성 바쿨로바이러스를 생성하였다.
Sf9 또는 Sf21 곤충 세포를 4일 동안 50 ml의 배지에서 배양하고, 감염성 재조합 바쿨로바이러스 ("Rep-바쿨로바이러스")를 배양물로부터 단리하였다. 선택적으로, 제1 세대의 렙-바쿨로바이러스 (P0)는 나이브 Sf9 또는 Sf21 곤충 세포를 감염시켜 증폭되고 50 내지 500 ml의 배지에서 배양되었다. 감염 후 3일 내지 8일 사이에, 배지에서 P1 바쿨로바이러스 입자를 원심분리 또는 여과 또는 또 다른 분별 공정에 의해 세포를 분리함으로써 수집하였다. Rep-바쿨로바이러스를 수집하고 바쿨로바이러스의 감염 활성을 측정하였다. 구체적으로, 2.5x106 세포/mL에서 4 x 20 mL Sf9 세포 배양물을 다음 희석, 1/1000, 1/10,000, 1/50,000, 1/100,000에서 P1 바쿨로바이러스로 처리하고, 인큐베이션하였다. 감염력은 4 내지 5일 동안 매일, 세포 직경 증가율 및 세포주기 정지율, 및 세포 생존력의 변화에 의해 결정되었다.
ceDNA 벡터 생성 및 특성화
도 4b를 참조하면, (1) ceDNA-박미드 또는 ceDNA-바쿨로바이러스를 함유하는 샘플 및 (2) 상기 기재된 Rep-바쿨로바이러스를 함유하는 Sf9 곤충 세포 배양 배지를 각각 1:1000 및 1:10,000의 비율로 Sf9 세포의 새로운 배양액 (2.5E+6 세포/ml, 20ml)에 첨가하였다. 이어서, 세포를 25℃에서 130 rpm으로 배양하였다. 공동 감염 후 4-5일 후에, 세포 직경 및 생존력이 검출된다. 세포 직경이 ~70-80%의 생존력으로 18-20nm에 도달하면, 세포 배양물을 원심분리하고, 배지를 제거하고, 세포 펠릿을 수집하였다. 세포 펠릿은 먼저 물 또는 완충제 중 적절한 부피의 수성 매질에 재현탁된다. ceDNA 벡터는 Qiagen MIDI PLUS™ 정제 프로토콜 (Qiagen, 컬럼당 처리되는 세포 펠릿 덩어리 0.2mg)을 사용하여 세포로부터 단리 및 정제되었다.
Sf9 곤충 세포로부터 생성되고 정제된 ceDNA 벡터의 수율은 260nm에서의 UV 흡광도에 기초하여 초기에 결정되었다.
ceDNA 벡터는 도 4d에 도시된 바와 같이 비변성 또는 변성 조건 하에서 아가로스 겔 전기영동에 의해 동정함으로써 평가될 수 있으며, 여기서 (a) 제한 엔도뉴클레아제 절단 및 겔 전기영동 분석 후 변성 겔 대 비변성 겔 상에서 크기의 2배로 이동하는 특징적인 밴드의 존재 및 (b) 비절단 물질에 대한 변성 겔 상의 단량체 및 이량체 (2x) 밴드의 존재는 ceDNA 벡터의 존재의 특징이다.
단리된 ceDNA 벡터의 구조는 a) ceDNA 벡터 내에 단일 절단 부위만 존재, 및 b) 0.8% 변성 아가로스 겔 상에서 분별될 때 명확하게 보일 정도로 충분히 큰 생성 단편 (>800 bp)에 대해 선택된 제한 엔도뉴클레아제를 사용하여 공동-감염된 Sf9 세포 (본원에 기재된 바와 같음)로부터 수득된 DNA를 소화함으로써 추가로 분석되었다. 도 4d 및 4e에 도시된 바와 같이, 비-연속 구조를 갖는 선형 DNA 벡터 및 선형 및 연속 구조를 갖는 ceDNA 벡터는 반응 생성물의 크기에 의해 구별될 수 있고, 예를 들어, 비-연속 구조를 갖는 DNA 벡터는 1kb 및 2kb 단편을 생성할 것으로 예상되고, 연속 구조를 갖는 비-캡슐화된 벡터는 2kb 및 4kb 단편을 생성할 것으로 예상된다.
따라서, 단리된 ceDNA 벡터가 정의에 의해 요구되는 바와 같이 공유적으로 폐쇄된-말단이라는 것을 정성적인 방식으로 입증하기 위해, 샘플을 단일 제한 부위를 갖는 것으로 특정 DNA 벡터 서열의 맥락에서 확인된 제한 엔도뉴클레아제로 소화하여, 바람직하게는 동일하지 않은 크기 (예를 들어, 1000 bp 및 2000 bp)의 2개의 절단 산물을 생성한다. 소화 및 변성 겔 (2개의 상보적 DNA 가닥을 분리함) 상에서 전기영동 후, 선형의 비공유적으로 폐쇄된 DNA는 1000 bp 및 2000 bp 크기로 분해되며, 공유적으로 폐쇄된 DNA (즉, ceDNA 벡터)는, 2개의 DNA 가닥이 연결되고 현재 펼쳐져(unfolded) 길이가 2배 (단일 가닥을 통해)이므로 2배 크기 (2000 bp 및 4000 bp)로 분해될 것이다. 또한, DNA 벡터의 단량체, 이량체 및 n-머릭(meric) 형태의 소화는 다량체 DNA 벡터의 말단-대-말단 연결로 인해 동일한 크기의 단편으로 모두 분해될 것이다 (도 4d 참조).
본원에 사용된 문구 "비변성 겔 및 변성 조건 하에서 아가로스 겔 전기영동에 의한 DNA 벡터의 동정을 위한 분석"은 제한 엔도뉴클레아제 소화를 수행한 후 소화 생성물의 전기영동 평가를 수행하여 ceDNA의 폐쇄-말단성(close-endedness)을 평가하기 위한 분석을 지칭한다. 하나의 이러한 예시적인 분석이 뒤따르지만, 당업자는 이 예에서 많은 당 업계 공지된 변형이 가능하다는 것을 이해할 것이다. 제한 엔도뉴클레아제는 DNA 벡터 길이의 대략 1/3x 및 2/3x의 생성물을 생성하는 관심 ceDNA 벡터에 대한 단일 절단 효소로 선택된다. 이것은 비변성 겔과 변성 겔 상의 밴드를 분해한다. 변성 전에, 샘플에서 완충제를 제거하는 것이 중요한다. Qiagen PCR 클린-업 키트 또는 탈염 "스핀 컬럼", 예를 들어 GE HEALTHCARE ILUSTRA™ MICROSPIN™ G-25 컬럼이 엔도뉴클레아제 소화를 위해 당 업계에 공지된 일부 옵션이다. 분석은 예를 들어, i) 적절한 제한 엔도뉴클레아제(들)를 사용하여 DNA 소화시키고, 2) 예를 들어, Qiagen PCR 클린-업 키트에 적용하고, 증류수로 용리하고, iii) 10x 변성 용액 (10x = 0.5M NaOH, 10mM EDTA)을 첨가하고, 완충되지 않은 10X 염료를 첨가하고, NaOH 농도가 겔 및 겔 박스에서 균일하도록 보장하기 위해 1mM EDTA 및 200mM NaOH와 함께 사전 인큐베이션된 0.8 - 1.0% 겔 상에서 10X 변성 용액을 4x로 첨가하여 제조된 DNA 레더와 함께 분석하고, 1x 변성 용액 (50 mM NaOH, 1mM EDTA)의 존재하에 겔을 작동시키는 것을 포함한다. 당업자는 크기 및 원하는 결과 타이밍에 기초하여 전기영동을 작동시키기 위해 어떤 전압을 사용할 것인지 이해할 것이다. 전기영동 후, 겔을 배수하고 1x TBE 또는 TAE로 중화시키고 1x SYBR Gold를 사용하여 증류수 또는 1x TBE/TAE로 옮긴다. 그런 다음 밴드를 예를 들어 Thermo Fisher, SYBR® Gold Nucleic Acid Gel Stain (DMSO 중 10,000X 농축물) 및 에피플루오레센트 빛(epifluorescent light) (청색) 또는 UV (312nm)로 시각화할 수 있다.
생성된 ceDNA 벡터의 순도는 임의의 당 업계에 공지된 방법을 사용하여 평가될 수 있다. 하나의 예시적이고 비제한적인 방법으로서, ceDNA 벡터의 형광 강도를 표준과 비교함으로써 샘플의 전체 UV 흡광도에 대한 ceDNA-플라스미드의 기여를 추정할 수 있다. 예를 들어, UV 흡광도를 기준으로 4μg의 ceDNA 벡터가 겔에 로딩되고, ceDNA 벡터 형광 강도가 1μg인 것으로 알려진 2kb 밴드에 해당하는 경우, 1μg의 ceDNA 벡터가 있으며, ceDNA 벡터는 총 UV 흡수재의 25%이다. 그런 다음 겔 상의 밴드 강도는 밴드가 나타내는 계산된 입력에 대해 플로팅되며, 예를 들어 총 ceDNA 벡터가 8kb이고, 절단된 비교 밴드가 2kb인 경우, 밴드 강도는 전체 입력의 25%로 플롯팅될 것이며, 이 경우 1.0μg 입력의 경우 .25μg일 것이다. ceDNA 벡터 플라스미드 적정을 사용하여 표준 곡선을 플롯팅하면, 회귀선 방정식이 ceDNA 벡터 밴드의 양을 계산하는데 사용되고, 그런 다음 ceDNA 벡터로 표시되는 총 입력 백분율 또는 순도 백분율을 결정하는데 사용될 수 있다.
실시예 2: ceDNA 벡터는
시험관내
에서 루시퍼라제 이식유전자를 발현시킨다
작제물은 루시퍼라제 리포터 유전자를 인코딩하는 오픈 리딩 프레임을 루시퍼라제 코딩 서열을 포함하는 ceDNA-플라스미드 작제물: 작제물 15-30, (상기 표 8 참조)의 클로닝 부위에 도입함으로써 생성되었다. HEK293 세포를 배양하고 형질감염제로서 FUGENE® (Promega Corp.)를 사용하여 100 ng, 200 ng 또는 400 ng의 플라스미드 작제물 1-31로 형질감염시켰다. 각각의 플라스미드로부터 루시퍼라제의 발현을 각 세포 배양물에서 루시퍼라제 활성에 기초하여 결정하였고, 이는 루시퍼라제 활성이 플라스미드로부터의 유전자 발현으로부터 초래되었음을 확인하였다.
실시예 3: ceDNA 벡터로부터 루시퍼라제 이식유전자의 생체내 단백질 발현.
작제물로부터 생성된 ceDNA 벡터로부터의 이식유전자의 생체내 단백질 발현은 마우스에서 평가될 수 있다. 예를 들어, ceDNA-플라스미드 작제물 1-31 (표 8에 기재됨)로부터 수득된 ceDNA 벡터를 시험하고, 리포좀, 리도스(redose) (28일에) 없이 ceDNA 작제물의 유체 역학적 주사 후 마우스 모델에서 지속되고 내구성 있는 루시퍼라제 이식유전자 발현, 및 외인성 반딧불이 루시퍼라제 ceDNA의 지속력 (최대 42일)을 입증하였다. 다른 실험에서, 선택된 ceDNA 벡터의 루시퍼라제 발현은 생체내에서 평가되며, 여기서 ceDNA 벡터는 루시퍼라제 이식유전자를 포함하고, 5' ITR 및 3'ITR은 표 2, 표 4a, 표 4b 또는 표 5 중 어느 것에 열거된 임의의 ITR 쌍, 또는 도 7a-7b에 도시된 변형된 ITR 쌍으로부터 선택된다. ceDNA 벡터로부터 생체내 단백질 발현을 평가하기 위해 하기 예시적인 방법이 사용되었다.
생체내 루시퍼라제 발현: 5-7주 수컷 CD-1 IGS 마우스 (Charles River Laboratories)에 0일째 꼬리 정맥에 iv 유체 역학적 투여를 통해 1.2 mL 부피로 루시퍼라제를 발현하는 0.35 mg/kg의 ceDNA 벡터를 투여하였다. 루시퍼라제 발현은 3, 4, 7, 14, 21, 28, 31, 35 및 42일에 IVIS 영상화에 의해 평가된다. 간략하게, 마우스에 150 mg/kg의 루시페린 기질을 복강내 주사 한 다음, IVIS® 영상화를 통해 전신 발광을 평가하였다.
IVIS 영상화는 3일, 4일, 7일, 14일, 21일, 28일, 31일, 35일 및 42일에 수행되고, 수집된 장기는 42일에 희생시킨 후 생체외에서 영상화된다.
연구 과정에서 동물의 체중을 측정하고 일반적인 건강과 웰빙을 매일 모니터링한다. 희생시, 혈액은 말단 심장 스틱에 의해 각각의 동물로부터 수집되고, 두 부분으로 분할되고, 1) 혈장 및 2) 혈청으로 처리되며, 혈장 스냅-냉동 및 혈청은 간 효소 패널에 사용되며 이어서 스냅 냉동된다. 또한, 간, 비장, 신장 및 사타구니 림프절 (LN)은 IVIS에 의해 생체외에서 수집되고 이미지화된다.
루시퍼라제 발현은 MAXDISCOVERY® 루시퍼라제 ELISA 분석 (BIOO Scientific/PerkinElmer), 간 샘플의 루시퍼라제를 위한 qPCR, 간 샘플의 조직병리 및/또는 혈청 간 효소 패널 (VetScanVS2; Abaxis Preventive Care Profile Plus)에 의해 간에서 평가된다.
실시예 4: 변형된 ITR 스크리닝
A. 비대칭 및 대칭 ITR 쌍을 포함하는 ceDNA 벡터에 대한 변형된 ITR 스크리닝.
모드-ITR 구조와 ceDNA 형성의 관계의 분석은 PCT 출원 PCT/US18/49996에 기재된 바와 같이 수행될 수 있으며, 이는 그 전문이 본원에 참고로 포함된다. 본원의 도 7a-7b 및 표 4a 및 4b에 나타낸 바와 같은 일련의 모드-ITR는 ceDNA 형성에 대한 특정 구조적 변화의 영향 및 ceDNA-인코딩된 이식유전자를 발현하는 능력에 대한 의문을 풀기 위해 구축되었다. 인간 세포 배양에서 돌연변이 작제, ceDNA 형성의 분석 및 ceDNA 이식유전자 발현의 평가는 하기에 더욱 상세하게 기술되어 있다. 예상한 대로, 3개의 음성 대조군 (배지만, 공여체 DNA가 없는 모의 형질감염 및 Rep-함유 바쿨로바이러스 세포의 부재 하에서 처리된 샘플)은 현저한 루시퍼라제 발현을 나타내지 않았다. 각각의 돌연변이 샘플에서 강력한 루시퍼라제 발현이 관찰되었으며, 이는 각각의 샘플에 대해 ceDNA-인코딩된 이식유전자이 돌연변이에 관계없이 성공적으로 형질감염되고 발현되었음을 나타낸다. 따라서, 돌연변이 샘플은 비대칭 모드-ITR 쌍을 포함하는 ceDNA를 정확하게 형성하는 것으로 보였다. 모드-ITR은 본 발명의 조성물 및 방법에 사용될 수 있고 하기 예시적인 방법을 사용하여 활성을 스크리닝할 수 있다.
대칭 ITR 쌍을 갖는 ceDNA 벡터가 상기 실시예 1에 기재되고 도 4b에 기재된 바와 같이 생성되고 구축되었다. 대칭 모드-ITR과 대칭 WT-ITR의 관계의 분석은 PCT/US18/49996에 기재된 방법에 따라 평가되었으며, 이는 그 전문이 본원에 참고로 포함된다. ITR 서열에 대한 돌연변이는 우측 및 좌측 ITR 영역 모두에서 대칭적으로 생성되었다. 라이브러리는 표 5에 개시된 바와 같이 16개의 우측 이중 돌연변이체 (예를 들어, 대칭 모드-ITR 쌍)를 함유하였다.
실시예 5: 유전자 편집 ceDNA 벡터의 생성
예시적인 목적으로, 예시적인 유전자 편집 ceDNA 벡터는 인자 VIII을 편집하기 위한 ceDNA 벡터를 생성하는 것과 관련하여 기술되며, 하기에 기술된다. 그러나, 인자 VIII이 본원에 기재된 바와 같은 방법 및 작제물에 유용한 유전자 편집 ceDNA 벡터를 생성하는 방법을 예시하기 위해 본 실시예에서 예시되지만, 당업자는 상기 언급된 바와 같이, 유전자 편집이 바람직한 임의의 유전자를 사용할 수 있음을 인식하고 있다. 편집을 위한 예시적인 유전자는, 예를 들어, "유전자 편집 ceDNA로 치료될 예시적인 질환" 및 "유전자 편집을 위한 추가 질환"이라는 제목의 섹션에서 본원에 기재되어 있다.
인자 VIII 유전자 편집 ceDNA의 생성: 관심 이식유전자 (예를 들어, 비제한적인 일 예로서, 인자 VIII)을 포함하는 오픈 리딩 프레임을 ceDNA 벡터에 삽입하고, 이식유전자의 결함이 있는 천연 카피와 관련된 질환 또는 장애가 있는 환자 (인자 VIII의 경우, 혈우병 A로 고통받는 환자)에 대한 내인성 이식유전자 유전자좌 내의 HDR을 용이하게 하기 위해 오픈 리딩 프레임에 인접한 게놈 DNA 서열의 큰 (각각 최대 2 kb) 상동성 아암에 의해 플랭킹된다. 부위-특이적 뉴클레아제 오픈 리딩 프레임은 선택적으로 sgRNA와 같은 임의의 필요한 보조 성분과 더불어, 천연 이식유전자 유전자좌 (예를 들어, 인자 VIII 유전자좌)에 또는 그 근처의 부위에 특이적이고, 재조합을 증가시키는데 효과적인 뉴클레아제와 함께 벡터에 포함된다. ceDNA 벡터는 또한 뉴클레아제가 ceDNA 벡터로부터 뉴클레아제의 발현을 불가능하게 하는 ceDNA 벡터 자체의 부위에 대해 더 특이적이 되도록 조작될 수 있다. 이러한 추가 특이성은 ceDNA 벡터에 의해 발현된 추가 gRNA에 의해 제공된다. ceDNA는, 예를 들어, 본원에 기재된 바와 같이 지질 나노입자 (LNP)로 전달될 수 있다.
ceDNA-이식유전자 작제물은 뉴클레아제 (예를 들어, Cas9, TALE, MegaTales 또는 ZFN) 및, 필요한 경우 유전자 편집 과정에 DNA 특이성을 제공하는 가이드 RNA를 포함하도록 추가로 조작될 수 있다. 따라서, 이 '올인원' ceDNA 작제물은 핵심 ceDNA 골격 요소 외에 다음과 같은 요소를 갖는다: 이식유전자 코딩 서열 (예를 들어, 이식유전자 인코딩 인자 VIII); 2개의 게놈 상동성 영역 (예를 들어, 내인성 인자 VIII 유전자좌에 특이적인 HR); 뉴클레아제 코딩 영역 및 뉴클레아제의 발현을 유도하기 위한 프로모터; 및 CRISPR 시스템이 이용되는 경우, 가이드 RNA (예를 들어, Cas9의 경우). 이식유전자과 함께 시스로 sgRNA 및 Cas9 발현 카세트를 갖고, sgRNA 및 Cas9가 상동성 아암 외부에 있어 세포 게놈 내로 통합되지 않도록 ceDNA 벡터를 조작할 수 있다. 유전자 편집 사건 후, HDR 후 선형 ceDNA는 DNA 말단을 노출시켜 분해될 것이며, 따라서 이 작제물로부터의 발현을 감소시킬 것이다.
인자 VIII 작제물을 갖는 예시적인 ceDNA 벡터는 그 자체의 불활성화를 유도할 뉴클레아제 서열 (또는 이의 프로모터)로 조작된 DNA 서열을 갖도록 추가로 변형될 수 있다. 예를 들어, Cas9 단백질이 생산될 때, 이는 유전자 편집 (즉, 원하는 효과)을 유발할뿐만 아니라 ceDNA에 결합하여 ceDNA 내에서 이중 가닥 DNA 파괴를 유도하며, 따라서 이는 Cas9의 하향 조절/제거를 보장할 것이다 (지속적인 Cas9에 의해 유도된 오프-표적 DNA 파괴의 가능성을 감소시키기 위해).
인자 VIII을 인코딩하는 유전자 편집 ceDNA 벡터는 알부민 유전자좌 또는 다른 게놈 유전자좌 (삽입된 인자 VIII의 발현을 유도하는 강력한 프로모터 근처)에 대한 게놈 상동성 아암을 이용하여 생성될 수 있다. 실시예 5에 언급된 다양한 실험이 이 프레임워크에서 반복된다.
도 10a는 ceDNA 설계에 포함된 5' 및 3' 상동성 아암에 의해 플랭킹된 본 개시내용에 따른 시험-벡터 발현 유닛을 도시한다. 이 구현예에서, ceDNA는 알부민 게놈 유전자좌에 하이브리드화되고 따라서 내인성 알부민 프로모터 하에서 FIX의 발현을 유도하는 5' 및 3' 상동성 아암으로 플랭킹된 인자 IX (FIX) 오픈 리딩 프레임으로 설계된다. 대조군은 5' 상동성 아암만의 발현 유닛; 및 3' 상동성 아암만을 함유하는 것이다 (각각 도 10b 및 10c). 프로모터, WPRE 요소, pA를 포함하는 발현 유닛 리포터 유전자, 예를 들어, GFP를 사용하여 발현을 실험적으로 확인할 수 있다 (도 10d).
Cas9 mRNA, 아연-핑거 뉴클레아제 (ZFN), 전사 활성화제-유사 이펙터 뉴클레아제 (TALEN), 돌연변이된 "닉카아제" 엔도뉴클레아제, 클래스 II CRISPR/Cas 시스템 (CPF1)과 같이, 뉴클레아제 발현 유닛을 포함하는 ceDNA 벡터는 트랜스로 전달될 수 있다 (도 10e). 본원에 기재된 바와 같은 LNP는 전달 옵션으로서 사용될 수 있다. 5' 또는 3' 효소 펩티드 서열에 융합된 핵 국재화 신호 (NLS) (예를 들어, Cas9, ZFN, TALEN 등과 같은 뉴클레아제 발현 유닛)를 사용하여 뉴클레아제 발현 유닛의 핵으로의 수송을 증가시키거나 개선할 수 있다. ceDNA에 의해 발현된 뉴클레아제에 따라, 원하는 부위에서 이중 가닥 파괴 (DSB)를 유도하기 위해, 하나 이상의 단일 가이드 RNA가 또한 트랜스로 전달될 수 있다. 예를 들어, sgRNA 발현 벡터 또는 화학적으로 합성된 합성 sgRNA로서. (sg = 단일 가이드-RNA 표적 서열) (도 10f). sgRNA 벡터는 ceDNA 벡터 또는 다른 발현 벡터일 수 있다. 단일 가이드 RNA 서열을 자유롭게 이용 가능한 소프트웨어/알고리즘을 사용하여 선택하고 검증할 수 있다. 4개의 잠재적 후보 서열이 선택되고 검증된다. (tools.genome-engineering.org와 같은 공공 리소스를 사용하여 적합한 단일 가이드 RNA 서열을 선택할 수 있다.)
예시적인 5' 및 3' 상동성 아암: 5' 및/또는 3' 상동성 아암은 도 8, 9 및 10a-10f에 도시된 ceDNA 작제물에 사용하기 위해 길이가 약 350bp일 수 있다. 예를 들어, 5' 상동성 아암은 50 내지 2000bp 범위일 수 있다. 유사하게, 3' 상동성 아암은 길이가 약 2000bp일 수 있고, 50 내지 2000bp 범위일 수 있다. 당업자는 하기에 기재된 바와 같이 5' 및/또는 3' 상동성 아암의 길이 및/또는 재조합 빈도를 변형시킬 수 있다: Zhang, Jian-Ping, et al. "Efficient precise knockin with a double cut HDR donor after CRISPR/Cas9-mediated double-stranded DNA cleavage." Genome biology 18.1 (2017): 35.and Wang, Yuanming, et al. "Systematic evaluation of CRISPR-Cas systems reveals design principles for genome editing in human cells." Genome biology 19.1 (2018): 62. 본원의 도 16에 도시된 바와 같이, FIX 또는 FVIII은 임의의 프로모터가 없는 오픈-리딩 프레임 (ORF)으로 대체될 수 있다. WPRE 및 폴리아데닐화 신호, 예컨대 BGHpA를 포함하지만, 이에 제한되지 않는 추가 요소가 유전자 편집 ceDNA 작제물에 첨가될 수 있다. 예를 들어, 삽입될 유전자의 발현 (예를 들어, 예시적인 유전자로서 FIX 또는 FVII)은 내인성 및 매우 강한 알부민 프로모터에 의해 구동된다. WPRE와 같은 전사 향상 요소는 ORF의 3'에 첨가된다. 폴리아데닐화 신호 (예를 들어, BGH-pA)도 첨가될 수 있다. 본원에 개시된 바와 같이, ceDNA 작제물의 용량은 크기 때문에 ITR 사이의 DNA 단편의 길이가 15kb를 넘는 것을 허용한다. 따라서, 큰 ORF를 갖는 ceDNA 벡터 시스템의 사용이 포함된다. 또한, 강한 프로모터 단위를 갖는 다른 발현 유닛이 사용될 수 있다. 예를 들어, 알부민 유전자좌를 표적화하는 대신, 다른 세이프 하버 유전자좌, 예컨대, 비제한적으로, CCR5 또는 AAV-세이프-하버-S1 (AAVS1) 유전자좌를 표적화하는 상동성 아암을 갖는, 다른 세이프 하버 유전자좌를 표적화하는 상동성 아암을 갖는 ceDNA 벡터가 사용될 수 있다. 이는 표적 세포 또는 조직에 어떠한 영향을 미치지 않으면서 유전자 편집 분자 또는 표적 유전자를 인트론 부위에 삽입할 수 있게 한다. 도 11에 도시된 바와 같이, 합성 프로모터 단위, 예를 들어, ceDNA 벡터에 기재된 CAG 프로모터의 인트론에 sgRNA 서열을 도입함으로써 뉴클레아제 활성의 자가 불활성화 특징의 적정을 위한 발현 작제물이 제조될 수 있다. 불활성화 정도는 sgRNA seq 또는 조합 및/또는 돌연변이된 (탈-최적화된(de-optimized)) sgRNA 표적 seq의 수에 의해 조절된다. (Zhang et al, NatPro, 2013 Regulation of Cas9 activity by using de-optimized sgRNA recognition target sequence.) 도 11에서, sgRNA는 단독으로 또는 다중으로 (예를 들어, 4개) 존재하며, 일부 구현예에서, 상이한 흑색 또는 백색으로 표시되는 하나 또는 다수의 고유한 표적 서열로 구성될 수 있다.
도 12는 ceDNA 벡터가 하나 이상의 sgRNA의 발현을 유도하기 위해 다양한 Pol III 프로모터 단위 배열을 포함할 수 있는 예를 보여준다. 이 예에서, 하나 초과의 선택된 프로모터가 ITR 사이에 배치되었다. 전사 방향은 정방향 또는 역방향일 수 있다. sgRNA는 조합 및/또는 복제될 수 있다. 도 14는 ceDNA가 U6 프로모터를 이용하는 것과 같이 다수의 sgRNA (sg1, sg2, sg3 또는 sg4)를 발현시킬 수 있는 또 다른 예를 보여준다.
따라서, 본원에 사용하기 위한 유전자 편집용 ceDNA 벡터는 이들 변형 중 임의의 하나 이상을 포함할 수 있다.
실시예 6: 마스터 ORF를 갖는 올인원 유전자 편집 ceDNA 벡터
유전자 편집 ceDNA 벡터는 도 15에 도시된 바와 같은 피처(feature)를 포함하여 제조될 수 있다. 표지되지 않은 포함된 피처는 뉴클레아제 발현 유닛 (해시된(hashed) 뉴클레아제 요소 포함) 및 예시된 sgRNA 표적화 서열을 갖는 프로모터의 인트론 하류이다. 피처는 ceDNA 특이적 ITR; 전사 방향과 관련하여 임의의 배향을 갖는 Pol III 프로모터 구동 (U6 또는 H1) sgRNA 발현 유닛; 탈-최적화를 갖거나 갖지 않는 sgRNA 표적화 서열을 포함할 수 있는 합성 프로모터 구동 뉴클레아제 (예를 들어, Cas9, 이중 돌연변이 닉카아제, Talen 또는 다른 돌연변이체) 발현 유닛 (지시된 것 이외에 위치한 실험에서); 0.05 내지 6kb 신장 상동성 아암에 의해 플랭킹된 바이러스성 2A 펩티드 절단 부위 (2A)를 통해 선별 마커 (예를 들어, NeoR)에 잠재적으로 융합된 이식유전자 (예를 들어, FIX)을 포함한다. (2A 시스템에 관해서는: Chan et al, Comparison of IRES and F2A-Based Locus-Specific Multicistronic Expression in Stable Mouse LinesHSV-TK suicide, PLOS 2011 HSV-TK suicide gene system; Fesnak et al, Engineered T Cells: The Promise and Challenges of Cancer Immunotherapy, NatRevCan 2016.)
적절하다면, 상동성 아암의 외부에 위치된, 성공적인 올바른 부위 사용을 위해 제어하고 선택할 수 있는 음성 선별 마커 (예를 들어, HSV TK) 및 발현 유닛이 구상된다. 본원에 개시된 바와 같은 다른 조절 요소 또는 조절 스위치는 또한 음성 선별 마커 유전자 대신에 또는 이에 추가하여 포함될 수 있다.
HDR 요소의 삽입을 위한 상동성 아암을 포함하는 예시적인 ceDNA 벡터가 도 13에 도시되어 있다. 이러한 ceDNA 벡터에서, 무작위 통합이 있는 경우, 음성 선별 마커가 있는 전체 벡터가 게놈에 통합된다. 이러한 잘못-형질감염된 세포는 HSV TK 음성 선별 마커에 대한 GVC와 같은 적절한 약물로 사멸될 수 있다. 대안적으로, 음성 선별 가능 마킹은 본원에 기재된 바와 같은 조절 스위치, 예를 들어, 사멸 스위치 유전자 또는 PCT/US18/49996 (이는 그 전문이 본원에 참고로 포함됨)의 표 11에 개시된 임의의 유전자로 대체될 수 있다.
또 다른 예시적인 ceDNA 벡터가 도 9에 도시되어 있으며, 이는 본 실시예의 것과 유사하지만, 음성 선별 마커를 "이중 돌연변이 닉카아제"에 대한 sgRNA 표적 seq로 대체한다 (단단한 아래쪽 화살표로 표시). 단일 가닥 DNA 절단 (닉킹)의 도입은 돌연변이 ITR에 근접한 3' 상동성 아암의 하류의 비틀림을 해제하고 어닐링을 증가시켜 HDR 빈도를 증가시키는데 도움이 될 수 있다. 이러한 ceDNA 벡터에서, 음성 마커는 "이중 돌연변이 닉카아제"에 대한 sgRNA 표적 서열과 함께 사용된다.
본 실시예에서 논의된 ceDNA 벡터는 단지 예시 목적을 위한 것이며, 다른 표적 유전자를 삽입하기 위해 당업자에 의해 변형될 수 있으며, 예를 들어, 사용되는 FIX 대신에, 인자 XIII이 사용되며, 인자 XIII이 사용되는 경우, FIX가 사용된다. 유사하게, 당업자는 유전자 편집이 요구되는 임의의 표적 유전자를 사용할 수 있다는 것을 알고 있다.
실시예 7:
질환의 치료를 위한 유전자 편집 ceDNA 벡터의 생성.
예시적인 목적으로, 실시예 7은 상이한 질환을 치료하기 위한 예시적인 유전자 편집 ceDNA 벡터를 생성하는 것을 기술한다. 그러나, 낭포성 섬유증, 간 장애, 전신 장애, CNS 장애 및 근육 장애에 대한 유전자가 본원에 기재된 바와 같은 방법 및 작제물에 유용한 유전자 편집 ceDNA 벡터를 생성하는 방법을 설명하기 위해 본 실시예에 예시되어 있지만, 당업자는, 유전자 편집이 요구되는 임의의 질환을 치료하기 위해 표적 유전자를 변형시킬 수 있다는 것을 알고 있다. 질환을 본원에 기재된 바와 같은 ceDNA 편집 벡터로 치료하기 위해 유전자 편집이 바람직한 전략인 예시적인 질환 또는 유전 장애는 "유전자 편집 ceDNA로 치료될 예시적인 질환" 및 "유전자 편집을 위한 추가 질환"이라는 제목의 섹션에서 논의된다.
일 예에서, 2-4와 같은 다수의 뉴클레아제 절단 부위를 갖는 sgRNA를 포함하는 ceDNA 벡터가 생성될 수 있으며, 뉴클레아제 및 5' 상동성 아암을 위한 상류 인트론 중 하나 또는 둘 다에 놓인다. 이들은 별개의 또는 공유된 sgRNA에 의해 유도된 특이성을 가질 수 있다. 이들 피처를 모두 갖는 예시적인 "올인원" ceDNA 벡터가 도 15에 도시되어 있다.
ceDNA 벡터를 대체하거나 제공하는 예시적인 이식유전자는 간 장애 (예를 들어, OTC, GSD1a, 크리글러-나자르(Crigler-Najar), PKU 등) 또는 전신 장애 (예를 들어, MPSII, MLD, MPSIIIA, 고셔, 파브리, 폼페 등)를 포함하는, 다른 유전자 장애에 대해 별개의 이식유전자으로 유전자 편집을 유도하도록 구성될 수 있다.
유전 장애 또는 질환을 치료하기 위한 유전자 편집 ceDNA 벡터의 예는 폐에서, 예를 들어 낭포성 섬유증 (CF)에서 유전자 편집을 유도하기 위해 ceDNA 벡터가 변형될 수 있다는 점에서 실시예 6에서 논의된 것과 유사할 수 있다. 이러한 ceDNA 벡터는 CF에서 돌연변이된 유전자인 CFTR을 인코딩하기 위해 생성된다. CFTR은 AAV 내에 포함될 수 없는 큰 유전자이다. 따라서, ceDNA 벡터는 독특한 솔루션을 제공하고, 일부 구현예에서, 폐 상피의 유전자 편집을 유도하기 위해 대상체에게 정맥내로 및/또는 분무 제형으로서 투여될 수 있다. 상기와 같이, ceDNA 유전자 편집 벡터는 CFTR이 내인성 CFTR 유전자좌에 삽입되도록 구성된다. 이러한 예에서, ceDNA 벡터는 또한 뉴클레아제 및 가이드 RNA를 포함할뿐만 아니라 유전자 편집의 효율 및 정확도를 증가시키기 위해 큰 상동성 아암을 이용할 수 있다.
CNS 장애의 유전자 편집을 위한 유전자 편집 ceDNA 벡터의 예는 실시예 6에서 논의된 것과 유사하며, 여기서 ceDNA는 신경퇴행성 장애 (예를 들어, 알츠하이머병, 파킨슨병, 헌팅턴병의 가족성 형태), 리소좀 저장 장애 (예를 들어, MPSII, MLD, MPSIIIA, 카나반(Canavan), 배튼(Batten) 등) 또는 신경발달 장애 (예를 들어, SMA, 레트 증후군(Rett syndrome) 등)를 포함하는 장애를 위한 CNS에서의 유전자 편집을 유도하도록 변형된다.
근육의 유전 장애 또는 질환을 치료하기 위한 유전자 편집 ceDNA 벡터의 예는 ceDNA 벡터가 뒤센느 근위축증, 안면견갑상완 이영양증 등을 포함하지만 이에 제한되지 않는 장애를 위한 근육에서의 유전자 편집을 유도하도록 변형될 수 있다는 점에서 실시예 6에서 논의된 것과 유사할 수 있다.
실시예 6-7에서 논의되고 예시된 유전자 편집 ceDNA (즉, ceDNA 벡터를 대체하거나 제공하는 이식유전자)는 결함이 있는 이식유전자에 대한 동물 모델에서 표적 세포로 전달되어 유전자 편집의 효능을 평가하고, 또한 보다 효과적인 유전자 산물을 생산하는 세포를 제공할 수 있다.
실시예 8:
ceDNA는 메가뉴클레아제가 표적화된 이중 가닥 파괴 (DSB)를 수행하는 유전자 편집에 사용하기에 적합하다.
유전자 편집 ceDNA 벡터는 메가뉴클레아제에 의해 제공되는 이중 가닥 파괴 후 삽입되는 교정 DNA 가닥으로서 주형 뉴클레오티드 서열을 포함할 수 있다. 예시적인 목적으로, 예시적인 유전자 편집 ceDNA 벡터는 Apo A-I 유전자를 편집 및 보정하기 위한 ceDNA 벡터를 생성하는 것과 관련하여 기술되며, 아래에서 설명된다. 그러나, 본원에 기재된 바와 같은 방법 및 작제물에 유용한 유전자 편집 ceDNA 벡터를 생성하는 방법을 예시하기 위해 Apo A-I 유전자의 교정이 본 실시예에 예시되어 있지만, 당업자는, 상기 언급된 바와 같이, 유전자 편집이 요구되는 임의의 다른 유전자의 서열을 교정하기 위해 ceDNA 벡터를 사용할 수 있다는 것을 알고 있다. 편집을 위한 예시적인 유전자는 예를 들어 "유전자 편집 ceDNA로 치료될 예시적인 질환" 및 "유전자 편집을위한 추가 질환"이라는 제목의 섹션에서 본원에 기재되어있다.
생체내에서 돌연변이된 인간 ApoAI 유전자의 메가뉴클레아제-유도된 교정
혈류에서 메가뉴클레아제 발현 카세트 및 ceDNA의 혼합물의 직접 주사에 의해 생체내 포유동물에서 이중 가닥 파괴 (DSB) 유도된 유전자 전환의 사용이 수행된다. 생체내 마우스에서 인간 Apo A-I 이식유전자의 복구에 기초한 시스템이 제공된다. 아포지단백질 A-I (APO A-I)은 고밀도 지단백질 (HDL)의 주요 단백질 요소이며 HDL 대사에서 중요한 역할을 한다. 고밀도 지단백질은 역 콜레스테롤 수송의 주요 매개체로서 주요한 심장보호 역할을 한다. Apo A-I 유전자는 간에서 발현되고 단백질은 혈액으로 분비된다. 또한, 인간의 Apo A-I 결핍은 조기 관상 동맥 심장 질환을 초래한다. 종합하면, 이러한 기준은 Apo A-I 유전자를 ceDNA를 포함한 메가뉴클레아제-유도된 유전자 교정의 연구에 대한 우수한 후보로 만든다.
이식유전자: 인간 Apo A-I 유전자를 코딩하는 게놈 서열이 이식유전자를 구축하는데 사용된다. Apo A-I 유전자의 발현은 간에서의 이식유전자 발현을 촉진시키기에 충분한 것으로 나타난 자체 최소 프로모터 (328 bp)에 의해 구동된다 (Walsh et al., J. Biol. Chem., 1989, 264, 6488-6494). 간략하게, 인간 Apo A-I 유전자는 인간 간 게놈 DNA (Clontech) 상의 PCR에 의해 수득되고 플라스미드 pUC19에서 클로닝된다. 2개의 정지 코돈을 함유하는 I-SceI 부위는 엑손 4와 같은 적합한 엑손의 시작 부분에서 PCR에 의해 삽입된다 (US 20120288943 A9의 도 17). 돌연변이된 유전자 (1-SceI-hApo A-I)는 절단된 형태의 천연 인간 APO A-I (야생형 APO A-I에 대해 80개 잔기 대 267개 아미노산)을 인코딩하도록 만들어진다. 모든 작제물은 인간 Apo A-I 유전자 서열에 대해 서열 분석되고 확인된다.
형질전환 마우스의 생성: 돌연변이된 인간 Apo A-I 유전자를 보유하는 EcoRI/XbaI 게놈 DNA 단편이 형질전환 파운더(founder)의 생성에 사용된다. 마우스 apo a-I 유전자에 대한 녹아웃 수컷 (WT KO 마우스) (The Jackson Laboratory, #002055) 및 B6SJLF1 암컷 (Janvier)의 번식으로부터 수정된 난모세포에 미세주사가 행해진다. 형질전환 파운더 마우스 (F0)는 꼬리로부터 추출된 게놈 DNA에 대한 PCR 및 서던 블롯 분석에 의해 확인된다. 이어서, 내인성 뮤린 apo a-I 유전자에 대한 녹아웃 유전적 배경에서 I-SceI-hApo A-I 형질전환 계통을 유도하기 위해 F0을 WT KO 마우스에 교미시켰다. 총 7개의 독립적인 형질전환 계통이 연구된다. 이식유전자 통합의 분자 특성화는 서던 블롯 실험에 의해 수행된다.
각각의 형질전환 계통에서 이식유전자 발현의 분석은 간으로부터 추출된 전체 RNA에 대해 RT-PCR에 의해 수행된다 (Trizol Reagent, Invitrogen). 뮤린 전사체와의 교차 반응을 피하기 위해, 인간 형질전환 I-SceI-hApo A-I cDNA에 특이적인 프라이머가 사용된다. 액틴 프라이머는 내부 대조군으로 사용된다.
생체내 유체 역학-기반 형질도입 생체내 형질전환 마우스의 간 세포의 형질도입은 유체 역학적 꼬리 정맥 주사에 의해 수행된다. 10 내지 20 g의 동물에게 10초 미만에 PBS에서 체중의 10분의 1의 부피로 원형 플라스미드 DNA를 주사한다. CMV 프로모터의 제어하에 I-SceI를 코딩하는 20 또는 50 마이크로그램의 ceDNA의 혼합물.
유전자 교정 분석: I-SceI 발현 카세트 및 ceDNA 복구 매트릭스의 주사 후 마우스에서의 이식유전자의 교정은 랜덤 6량체를 사용하여 역 전사된 총 간 RNA에 대한 네스티드(nested) PCR에 의해 분석된다. 교정된 유전자를 검출하지만, 교정되지 않은 유전자는 검출하지 않기 위해, 복구된 이식유전자를 특이적으로 증폭시킨 프라이머 세트가 사용된다. 특이성은 I-SceI 부위에 걸친 역방향 올리고뉴클레오티드를 사용함으로써 달성되며, 복구 매트릭스 외부에 정방향으로 위치한 액틴 프라이머를 내부 대조군으로 사용하였다.
결과:
I-SceI-hapo A-I 이식유전자의 하나 또는 여러 카피를 운반하는 다양한 형질전환 계통이 이 실험에 사용된다. 마우스에는 I-SceI-발현 벡터 및 ceDNA의 혼합물 또는 I-SceI-발현 카세트 및 ceDNA 둘 다를 운반하는 벡터가 주사된다. 돌연변이된 인간 Apo A-I 유전자의 복구는 교정된 인간 Apo A-I 유전자와만 쌍을 이루도록 특별히 고안된 프라이머를 사용하여 전체 간 RNA 상에서 RT-PCR에 의해 모니터링된다. PCR 단편은 I-SceI-발현 카세트 및 ceDNA 복구 매트릭스가 주사된 형질전환 마우스에서 구체적으로 시각화된다. 유전자 교정은 하나 또는 여러 개의 이식유전자 카피를 함유하는 시험된 모든 형질전환 계통에서 검출 가능하다.
메가뉴클레아제-유도된 유전자 전환이 생체내 게놈 수술을 수행하는데 사용될 수 있고, 메가뉴클레아제는 이러한 적용을 위한 약물로서 사용될 수 있는 것으로 나타났다. ceDNA 벡터는 메가뉴클레아제에 의해 제공되는 이중 가닥 파괴 후에 삽입되는 교정 DNA 가닥으로서 사용되는 주형 뉴클레오티드 서열을 포함한다.
실시예 9: 1차 인간 간세포의
시험관내
AAV 형질도입
세포 배양 접시 (48웰; CM1048; Lifetech)는 사전 코팅된 상태로 구입할 수 있거나 플레이트 (3548; VWR)를 웰당 150 mL로, 10 mL 간세포 기본 배지 (CC-3199; Lonza) 중 250 mL BDMatrigel (BD Biosciences)의 혼합물로 코팅할 수 있다. 플레이트를 37℃에서 1시간 동안 인큐베이션한다. 해동/플레이팅 배지는 18 mL InVitroGRO CP 배지 (BioreclamationIVT)와 400mL Torpedo 항생제 믹스 (Celsis In vitro Technologies)를 조합하여 제조된다. 플레이트가 준비되면, 플레이팅 가능한 인간 간세포 (lot# AKB; cat# F00995-P)를 액체 질소 증기상에서 37°수조로 바로 옮긴다. 바이알을 세포가 완전히 해동될 때까지 부드럽게 교반한다. 세포를 5mL의 예열된 해동/플레이팅 배지를 함유하는 50mL 원뿔형 튜브로 바로 옮긴다. 세포를 완전히 옮기기 위해, 바이알을 1 mL의 해동/플레이팅 배지로 세척한다. 튜브를 부드럽게 소용돌이쳐서 세포를 재현탁시킨다. 트리판 블루 용액 1:5 (25-900-C1; Cellgro)를 사용하여 세포 수를 세고 세포 생존력을 결정하기 위해 소량의 분취액 (20 mL)을 제거하였다. 이어서 세포를 75g에서 5분 동안 원심분리한다. 상청액을 완전히 따라 내고 세포를 13106 세포/mL로 재현탁시킨다. 마트리겔 혼합물을 웰로부터 흡인하고, 세포를 48웰 접시에서 웰당 23105개의 세포로 시딩한다. 이어서 세포를 37℃에서 5% CO2 인큐베이터에서 인큐베이션한다. 형질도입시, 세포는 유지를 위해 간세포 배양 배지 (HCM) (간세포 기본 배지, CC-3199, Lonza; HCM, CC-4182, SingleQuots)로 전환된다. 본원에 기재된 바와 같은 ceDNA 벡터 및 mAlb ZFN 메신저 RNA (mRNA) [또는 실험에서 각각 ZFN 메신저와 동일한 부위를 표적화하는 Cas9 mRNA 및 mAlb gRNA 또는 TALEN 또는 MN으로 대체; 또는 실험에서 ceDNA 상의 발현 요소를 통합]를 리포펙타민 RNAiMAX™ (Lifetech) (또는 본원에 개시된 다른 적합한 시약)로 형질감염시켰다. 24시간 후, 배지를 새로운 HCM으로 교체하고, 이는 1차 간세포 배양액의 최대 건강을 보장하기 위해 매일 수행된다. ELISA에 의한 hFIX 검출이 필요한 실험의 경우, hFIX가 상청액에 축적될 수 있도록 때때로 배지를 며칠 동안 교환하지 않는다.
실시예 10: ZFN 시스템을 사용한
생체내
혈우병 치료를 위한 ceDNA 벡터
아연 핑거 뉴클레아제-기반 유전자 편집 시스템을 포함하는 ceDNA 벡터가 또한 구축될 수 있다. 예시적인 목적으로, 예시적인 유전자 편집 ceDNA 벡터는 뉴클레아제 이식유전자으로서 아연 핑거 뉴클레아제 (ZFN)를 인코딩하는 ceDNA 벡터를 생성하는 것과 관련하여 기술되며, 아래에서 설명된다. 그러나, 본원에 기재된 바와 같은 방법 및 작제물에 유용한 뉴클레아제를 인코딩하는 유전자 편집 ceDNA를를 생성하는 방법을 예시하기 위해 ZFN이 본 실시예에 예시되어 있지만, 당업자는, 상기 사용에 언급된 바와 같이, 본원에 기재된 임의의 뉴클레아제, 예를 들어 그리고 비제한적으로 아연 핑거 뉴클레아제 (ZFN), TAL 이펙터 뉴클레아제 (TALEN), 메가뉴클레아제 및 CRISPR/Cas9-효소 및 조작된 부위-특이적 유도체 뉴클레아제를 사용할 수 있다는 것을 알고 있다. ceDNA 벡터에 의해 인코딩되는 예시적인 뉴클레아제는, 예를 들어, "DNA 엔도뉴클레아제"라는 제목의 섹션 및 그 안의 하위섹션에서 본원에 기재되어 있다.
전술한 실시예에 기재된 방법을 참고하면, ceDNA 벡터에 이식유전자으로서 포함되는 뉴클레아제는 아연 핑거 뉴클레아제 (ZFN)일 수 있다. 하나의 비제한적인 예로서, 샤마(Sharma) 등 (Blood 126: 1777-1784 (2015)에 의해 기재된 알부민 유전자좌로 치료적 이식유전자의 ZFN 매개 표적화는 본 발명의 ceDNA 벡터를 사용하여 수행될 수 있다. 이러한 ceDNA 벡터는 그 유전자좌를 표적화하는 ceDNA-인코딩된 ZFN의 활성을 통해 표적 대상체 내 알부민 유전자좌에서 인간 인자 VIII 및/또는 인자 IX의 통합을 허용한다. ceDNA 벡터는 본원에 기재된 임의의 전달 방법을 사용하여 환자에게 투여될 수 있다. 치료적 수준으로 혈우병 A 및 B의 마우스 모델에서, 예를 들어, 인간 인자 VIII 및 IX (hFVIII 및 hFIX)의 장기 발현은 이 방법을 사용하여 달성된다.
실시예 11: ZFN 시스템을 사용한
생체내
낭포성 섬유증 치료를 위한 ceDNA 벡터
폐에서, 예를 들어 낭포성 섬유증 (CF)을 갖는 대상체에서 유전자 편집을 유도하기 위해 실시예 10의 실험과 유사한 접근법이 적용된다. 이 실험에서, ceDNA는 CF에서 돌연변이된 유전자인, 야생형 CFTR을 인코딩하기 위해 생성된다. CFTR은 AAV 내에 포함될 수 없는 큰 유전자이다. ceDNA는 AAV보다 상당히 큰 핵산 삽입물을 수용하므로 CFTR의 치료에 독특한 솔루션을 제공한다. ZFN CFTR-특이적 유전자 편집 시스템을 인코딩하는 ceDNA 벡터는 폐 상피의 유전자 편집을 유도하기 위해 정맥내로 또는 분무 제형으로서 투여될 수 있다. 상기와 같이, 실험에서 CFTR은 그 유전자좌에 표적화된 인코딩된 ZFN의 활성을 통해 내인성 CFTR 유전자좌에 삽입되고, 뉴클레아제 및 가이드 RNA의 패키징 및 큰 상동성 아암을 사용하여 유전자 편집 효율 및 정확도를 증가시킬 수 있다.
실시예 12: 생체내 뒤센느 근위축증 치료를 위한 ceDNA 벡터
디스트로핀 유전자에서 돌연변이를 교정함으로써 근육 조직, 예를 들어, 뒤센느 근위축증에서 ZFN-매개 유전자 편집을 유도하기 위해 실시예 10 및 11의 실험과 유사한 접근법이 적용된다.
대안적으로, ceDNA 벡터는 디스트로핀 유전자의 엑손 51 스플라이스 수용체를 플랭킹하는 적어도 2개의 닉 및/또는 DSB를 생성하는 엔도뉴클레아제 (예를 들어, ZNF 또는 TALES)를 인코딩하기 위해 생성된다. 이러한 닉 및/또는 파괴를 복구하면 엑손 51이 결실된다. 엑손 51의 결실은 디스트로핀 전사체로부터 엑손 51을 배제함으로써 특정 DMD-유발 돌연변이, 예를 들어, 엑손 48-50의 결실을 교정한다. 본원에 기재된 ceDNA 벡터의 큰 페이로드 용량은 2개의 엔도뉴클레아제 (예를 들어, Ousterout et al. Molecular Therapy 2015 doi:10.1038/mt.2014.234에 기재된 2개의 ZFN; 그 전문이 본원에 참고로 포함됨) 또는 RNA-유도된 엔도뉴클레아제 및 다수의 sgRNA가 단일 벡터에서 근육 세포로 전달되도록 하여 효율을 증가시킨다. ceDNA 벡터는 근육 조직의 유전자 편집을 유도하기 위해 정맥내 또는 근육내로 투여될 수 있다.
또한, 단 하나의 가이드 RNA 표적 서열 (예를 들어, 하나 또는 다수의 카피 수) 및/또는 CRISPR/Cas 뉴클레아제를 (시스 또는 트랜스로) 발현시키는 ceDNA 유전자 편집 벡터를 사용하여 DMD 유전자의 개별 스플라이스 공여체 또는 스플라이스 수용체 부위를 표적화할 수 있다. 이는 엑손 스키핑 (예를 들어, 엑손 51 스키핑) 및 유전자의 교정이 기능성 단백질을 발현시키도록 하는 NHEJ를 초래한다. DMD 유전자를 표적으로 하는 다중 가이드 RNA는 US. 2016/0201089 (그 전문이 본원에 참고로 포함됨)에서 발견되며, 예를 들어, 그 안의 실시예 5-11을 참조한다. 디스트로핀 발현의 교정은 DMD 근아세포 세포주에서 시험될 수 있다.
실시예 13:
게놈 세이프 하버 유전자로부터의 장기 치료적 발현을 위한 ceDNA 유전자 편집 벡터.
상동성 도메인을 포함하는 ceDNA 유전자 편집 벡터를 사용하여 치료적 이식유전자의 삽입 및 발현을 위한 게놈 세이프 하버 유전자를 표적화할 수 있다. ceDNA 벡터는 실시예 1에 따라 제조된다. 임의의 세이프 하버 유전자좌가 표적화될 수 있으며, 이러한 세이프 하버는, 예를 들어, 공지된 비활성 인트론이거나 대안적으로 높은 발현 수준으로 단백질을 발현시키는 것으로 알려진 코딩 서열에 가까운 활성 인트론이다. 세이프 하버 유전자 내로의 삽입은 삽입 부재와 비교하여 유의미한 부정적인 영향을 미치지 않는다. 예를 들어, 혈청 알부민은 높은 발현 수준 및 간 세포에서의 존재로 인해 관심 있는 원형 표적이다. 알부민의 인트론 서열에 스플라이스 수용체 부위 및 이식유전자를 보유하는 프로모터가 없는 카세트의 통합은 알부민의 첫 번째 엑손이 최종 단백질 생성물로부터 절단된 분비 펩티드를 인코딩하기 때문에 많은 다른 단백질의 발현 및 분비를 뒷받침할 것이다. 적어도 하나의 ceDNA 벡터는 인트론 1을 표적화하는 아연 핑거 쌍을 인코딩한다. 예시적인 아연 핑거 쌍은 하기에 충분히 설명된 바와 같다: Blood (2015) 126 (15): 1777-1784 (예를 들어, 보충 도면 6 쌍 A-B 및 C-D), 이는 그 전체가 본원에 참고로 포함된다. 또한, ceDNA 벡터의 제한이 없기 때문에, ceDNA 벡터는 공여체 DNA를 동일하거나 다른 ceDNA 벡터 상에 제공하도록 조작된다.
예시적인 목적으로, 예시적인 유전자 편집 ceDNA 벡터는 알부민 세이프 하버를 표적화하는 상동성 아암 (상동성 도메인으로도 지칭됨)을 포함하는 ceDNA 벡터를 생성하는 것과 관련하여 기술되며, 아래에서 설명된다. 그러나, 본원에 기재된 바와 같은 방법 및 작제물에 유용한 이식유전자 (또는 공여체 DNA)의 표적화된 삽입을 위한 상동성 아암을 갖는 유전자 편집 ceDNA를 생성하는 방법을 설명하기 위해 이식유전자 (또는 공여체 DNA)을 알부민 알부민의 인트론 1으로의 표적화된 삽입을 위한 상동성 아암을 갖는 유전자 편집 ceDNA가 본 실시예에서 예시되어 있지만, 당업자는, 상기 사용에 언급된 바와 같이, 세이프 하버 유전자를 포함하지만 이에 제한되지 않는 임의의 유전자에 대해 상동성 아암을 사용할 수 있거나 유전자좌가 사용될 수 있고, 예를 들어, CCR5 또는 AAV-세이프-하버-S1 유전자좌가 표적화될 수 있다는 것을 알고 있다.
도 16은 공여체 DNA를 표적 알부민 인트론 서열 내로 통합시키기 위한 몇몇 프로모터가 없는 작제물을 도시한 개략도를 도시하며, 이러한 작제물은 뉴클레아제와 동일하거나 상이한 벡터 상에 있을 것이다. 일 구현예에서, 프로모터가 없는 ceDNA 작제물은 말단 반복부 (예를 들어, ITR)에 의해 플랭킹된 삽입/복구 서열을 포함하고, 뉴클레아제/가이드 RNA는 별도의 작제물 (예를 들어, 제2 ceDNA 벡터, 뉴클레아제를 인코딩하는 mRNA, 재조합 뉴클레아제, RNP 복합체 등)을 사용하여 제공된다. 프로모터 없이 (프로모터가 없는) 임의의 이식유전자, 예를 들어 FVIII 또는 인자 IX, 또는 GFP 또는 GFP 및 neo를 인코딩하는 ceDNA는 게놈 상동성 아암을 사용하여 알부민 유전자좌에 대해 제조된다 (실시예 4 참조). 일부 실험에서, ZFN 대신에, Cas9 또는 cpf1 뉴클레아제가 ceDNA 벡터로 조작되고 가이드 RNA는 ZFN 영역을 표적화하도록 설계된다. 일부 실험에서, 동일한 ceDNA가 가이드 RNA를 발현시키도록 추가로 조작될 것이고 (예를 들어, 도 14, 15 및 16 참조), CAS 또는 cpf1 효소가 사용될 때, 동일하거나 상이한 ceDNA 상에서, 또는 플라스미드 상에서 CRISPR이 제공될 수 있다. 가이드 RNA는, 표적 서열을 정렬시키고 사용되는 CAS 효소 (예를 들어, saCAS9 또는 sp CAS 9, 또는 cpf1 등)와 관련된 PAM 모티프를 식별함으로써, 예를 들어, 하기에서 ZFN 표적 서열에 결합하도록 조작된다: Sharma et al. Blood (2015) 126 (15): 1777-1784 (예를 들어, 보충 도면 6 쌍 A-B 및 C-D). 가이드 RNA에 대한 알부민 내 하나의 ceDNA 표적 센터가 도 17에 도시되어 있다. 유사한 부위가 인간 알부민에 사용된다.
예시된 이식유전자 (예를 들어, 분비된 단백질, 예를 들어, 인자 IX)을 발현시키기 위해 알부민 유전자 내로의 예시적인 삽입을 위한 ceDNA 유전자 편집 시스템은 인간 표적 유전자로 향하는 가이드 RNA를 사용할 때 시험관내 1차 인간 간세포 (예를 들어, Thermo Scientific의 인간 간세포)에서 및 생체내 시험을 위한 마우스 모델에서 시험된다. 예를 들어, 마우스 간은 상기 샤마 등에 기재된 바와 같이, 색성 분석법 및 항원을 사용하여 인자 IX mRNA 수준의 측정 및 인자 IX 활성의 측정을 위해 ceDNA 시스템의 전신 투여 후 단리된다. 인자 IX를 포함하는 시스템에서, 당 업계에 공지되어 있고 상업적으로 이용 가능한 mRNA 수준의 시험, 단백질 활성 분석 및 웨스턴 블롯은 임의의 원하는 이식유전자에 대한 녹인의 평가 및 시험관내 인간 1차 세포 및 생체내 마우스 모델 모두에서의 교정을 시험하는데 적합하다. 알부민 유전자좌 내로의 삽입은 분비 단백질의, 예를 들어, 혈액 내로의 분비를 허용한다. 이식유전자의 혈장 수준을 평가할 것이다. 인간 인자 VIII 및 인자 IX의 분비는 혈우병에 대한 동물 모델에서 생체내에서 시험될 것이다. 당업자는 본원에 기재된 ceDNA를 사용하여 임의의 분비 단백질이 알부민으로 '노킹(knocked)'될 수 있음을 이해할 것이다. 비제한적인 예로는 α-갈락토시다제, 이듀라네이트-2-설파타제, 베타-글루코시다제, α- L-이듀로니다제 등을 포함하고, 적절한 동물 모델에서 시험될 수 있다. 일 구현예에서, 녹인은 Gal1과 같은 유도성 프로모터의 제어를 받는다.
ceDNA 벡터의 비틀림 제약은 ceDNA 벡터 자체를 표적화하는 가이드 RNA와 함께 nCas9 닉카아제를 통해 해제되는 것으로 본원에서 추가로 고려된다. 비틀림 제약의 이러한 해제는 ceDNA 벡터에서 발견되는 HDR 주형에서 하나 이상의 상동성 아암의 능력을 개선시킬 수 있다. 또한, ITR을 표적화하는 가이드 RNA가 염색체에 대한 가이드 RNA와 조합하여 Cas9와 함께 사용될 수 있다는 것이 추가로 고려된다. ceDNA 벡터의 ITR (또는 상동성) 영역뿐만 아니라 염색체 상의 표적 부위를 모두 표적화하는 단일 가이드 RNA가 설계될 수 있다. ceDNA 벡터 내 절단 부위는 DNA 벡터의 한쪽 또는 양쪽 말단에 위치할 수 있다.
실시예 14: ceDNA 벡터에 사용하기 위한 예시적인 표적 유전자 및 sgRNA
예시적인 목적으로, 예시적인 유전자 편집 ceDNA 벡터는 편집을 위한 ZNF 뉴클레아제에 대한 sgDNA를 포함하는 ceDNA 벡터를 생성하는 것과 관련하여 기술되며, 아래에서 설명된다. 그러나, 유전자를 편집하기 위한 ZNF 뉴클레아제에 대한 sgRNA 서열이 본원에 기재된 방법 및 작제물에 유용한 유전자 편집 ceDNA 벡터를 생성하는 방법을 예시하기 위해 본 실시예에 예시되어 있지만, 당업자는, 상기 사용에 언급된 바와 같이, "DNA 엔도뉴클레아제"라는 제목의 본원의 섹션 및 그 안의 하위섹션에서 논의된 바와 같이, 아연 핑거 뉴클레아제 (ZFN), TAL 이펙터 뉴클레아제 (TALEN), 메가뉴클레아제, 및 CRISPR/Cas9 및 조작된 부위-특이적 뉴클레아제에 대한 sgRNA를 포함하는, 본원에 기재된 바와 같은 임의의 뉴클레아제의 sgRNA를 사용할 수 있다는 것을 알고 있다.
ZNF에 대한 비제한적인 예시적인 표적 유전자 및 표적 서열 쌍; 및 ZNF 표적 서열에 기초한 gRNA 서열은 표 9에서 발견된다. ceDNA 벡터는 표적 유전자의 보정 및/또는 조절을 위해 이들 서열을 표적화하는 ZNF를 발현시키도록 조작된다. ceDNA 벡터는 예를 들어 임의의 CRISPR/Cas 시스템을 사용한 표적 유전자의 교정을 위해 이러한 예시적인 gRNA를 발현시키도록 조작된다. 따라서, 특정 구현예에서, ceDNA 벡터는 표 9 또는 표 10에서 선택된 유전자를 표적화한다. 특정 구현예에서, ceDNA 벡터는 표 9로부터 선택된 가이드 RNA를 포함한다. 특정 구현예에서, ceDNA 벡터는하기 표 9로부터 선택된 표적 서열을 표적화하는 ZFN을 인코딩하는 유전자를 포함한다.
[표 9] ZFN 표적 서열을 표적화하는 sgRNA
[표 10]
실시예 15: T 세포의 조작을 위한 ceDNA 유전자 편집 벡터
본원에 개시된 바와 같이, 본원에 기재된 ceDNA 유전자 편집 벡터는 임의의 세포의 게놈, 예를 들어 T 세포에서 유전자를 편집, 복구 및/또는 녹아웃시키는데 사용될 수 있다. 예시적인 목적으로, 예시적인 유전자 편집 ceDNA 벡터는 T-세포에서 CXCR4, CCR5, PD-1 유전자 중 어느 것을 편집하기 위한 ceDNA 벡터를 생성하는 것과 관련하여 기술되고 아래에 설명된다. 그러나, CXCR4, CCR5 또는 PD-1 유전자를 표적화하는 것이 본원에 기재된 바와 같은 방법 및 작제물에 유용한 유전자 편집 벡터 ceDNA를 생성하는 방법을 예시하기 위해 본 실시예에 예시되어 있지만, 당업자는 상기 사용에 언급된 바와 같이, 예를 들어, "유전자 편집 ceDNA로 치료될 예시적인 질환" 및 "유전자 편집을 위한 추가 질환"이라는 제목의 섹션에서 본원에 기재된 바와 같이, 유전자 편집이 요구되는 임의의 유전자를 사용할 수 있다는 것을 알고 있다. 또한, T 세포의 게놈은 이 예시적인 예에서 변형되지만, 당업자는 임의의 세포, 예를 들어 본원에서 "숙주 세포"라는 제목의 섹션 XII.A에 기재된 바와 같은 임의의 세포가 생체외 또는 생체내에서 변형될 수 있다는 것을 알고 있다. 또한, 게놈 DNA가 이 예시적인 예에서 변형되는 것으로 보이지만, ceDNA 벡터는 또한, 미토콘드리아 DNA (mtDNA)를 변형하여, 예를 들어, 하기에 기재된 바와 같이 mtZFN 및 mitoTALEN 기능, 또는 미토콘드리아-적응된 CRISPR/Cas9 플랫폼을 인코딩하도록 변형될 수 있는 것으로 예상된다: Maeder, et al. "Genome-editing technologies for gene and cell therapy." Molecular Therapy 24.3 (2016): 430-446 and Gammage PA, et al. Mitochondrial Genome Engineering: The Revolution May Not Be CRISPR-Ized. Trends Genet. 2018;34(2):101-110.
치료적으로 관련된 임의의 유전자가 표적화될 수 있고 (예를 들어, HIV 진입을 위한 보조수용체(coreceptor)인, CXCR4 또는 CCR5), 제거, 편집, 복구 또는 교체될 수 있다 (CXCR4의 경우, 예를 들어, HIV 진입을 방지하기 위해). 추가의 비제한적인 예에서, T 세포 고갈의 매개체인, PD-1이 제거될 수 있다. 표적 유전자의 제거는 주형 핵산 서열, 예를 들어 공여체 HDR 주형과 함께 또는 없이 수행된다. HDR 주형이 없는 경우 단일 가이드 RNA (sgRNA) 및 상응하는 뉴클레아제를 사용하면 비-동종-말단-결합 (NHEJ)이 초래된다.
치료적으로 관련된 임의의 유전자좌가 표적화될 수 있으며, 이러한 표적화된 유전자좌는, 예를 들어, T 세포 고갈의 공지된 조절제, 바이러스 보조수용체 등이다. ceDNA 유전자 편집 벡터는, 예를 들어, 본원에서 "DNA 엔도뉴클레아제"라는 제목의 섹션 및 그 안의 하위섹션에 기재된 바와 같이 RNA-유도 엔도뉴클레아제, 예를 들어, CRISPR/Cas9, 및 아연 핑거 뉴클레아제 (ZFN), TAL 이펙터 뉴클레아제 (TALEN), 메가뉴클레아제 및 조작된 부위-특이적 유도체 뉴클레아제를 포함하는 다른 엔도뉴클레아제를 포함하는, 본원에 기재된 바와 같은 임의의 엔도뉴클레아제를 포함할 수 있다. CXCR4 및 PD-1의 HDR에 대한 예시적인 적합한 엔도뉴클레아제 및 주형 핵산은 하기에 충분히 기재되어 있다: Schumann et al., PNAS (2015) 112(33):10437-10442, 이는 그 전문이 본원에 참고로 포함된다 (예를 들어 슈만 등의 도 1-4 참조).
또한, ceDNA의 크기 제한이 없기 때문에, ceDNA 벡터는 동일한 ceDNA 벡터 상에 공여체 DNA 및/또는 유전자 편집 분자를 제공하도록 조작될 수 있다. 대안적으로, 공여체 DNA 및/또는 유전자 편집 분자는 하나 이상의 상이한 ceDNA 벡터 상에 제공될 수 있다.
표적 유전자좌의 절제, 예를 들어, 프로모터의 파괴 또는 조기 정지 코돈 또는 미스센스 돌연변이의 삽입에 적합한 ceDNA 인코딩 주형 핵산은 표적 유전자좌에 대한 게놈 상동성 아암과 함께 만들어질 것이다 (예를 들어, 실시예 10 참조). 이와 같이, 변형된 세포에서, 유전자는 절단되거나 유전자 침묵될 것이다. 일부 실험에서, Cas9 또는 cpf1 뉴클레아제는 동일하거나 상이한 ceDNA 벡터로 조작될 것이다. 일부 실험에서, ceDNA는 가이드 RNA를 발현시키도록 추가로 조작될 것이고 (예를 들어, 도 12, 15, 16 참조), Cas 또는 cpf1 효소가 사용될 때, 이는 동일하거나 상이한 ceDNA 상에서, 또는 플라스미드, 또는 mRNA 또는 재조합 단백질에 의해 제공될 수 있다. 가이드 RNA는, 표적 서열을 정렬시키고 사용되는 Cas 효소 (예를 들어, saCas9 또는 spCAS 9, 또는 cpf1 등)와 관련된 PAM 모티프를 식별함으로써, 예를 들어, 하기에서 표적 서열에 결합하도록 조작된다: PNAS (2015) 112(33):10437-10442.
일부 실험에서, 가이드 RNA는 다른 공지된 서열 영역을 표적화할 것이다. 공지된 표적 영역에 결합하는 다수의 sgRNA 서열은 미국 특허 공보 2015/0056705 (이는 그 전문이 본원에 참고로 포함됨)의 표 1-2에 기재되어 있으며, 예를 들어 인간 베타-글로빈, 인간, BCLIIA, 인간 KLF1, 인간 CCR5, 인간 CXCR4, PPP1R12C, 마우스 및 인간 HPRT, 인간 알부민, 인간 인자 IX, 인간 인자 VIII, 인간 LRRK2, 인간 Htt, 인간 RH, CFTR, TRAC, TRBC, 인간 PD1, 인간 CTLA-4, HLA c11, HLA A2, HLA A3, HLA B, HLA C, HLA c1. II DBp2. DRA, Tap 1 및 2. 타파신, DMD, RFX5, 등)에 대한 gRNA 서열을 포함한다.
ceDNA 벡터는 생체외에서 T 세포로 전달될 것이나, 전신 전달이 또한 본원에서 고려된다. 일부 실험에서, Cas9 또는 cpf1 뉴클레아제 대신에, 아연 핑거 뉴클레아제, TALENS 또는 megaTAL이 동일하거나 상이한 ceDNA 벡터로 조작될 것이다.
실시예 16: 위스콧-알드리치 증후군 (Wiscott-Aldrich Syndrome; WAS)을 치료하기 위한
생체외
유전자 편집
AAV를 사용한 생체외 유전자 편집은 AAV가 상동성 복구 주형 또는 DNA 공여체 주형의 유일한 공급원이라는 점에서 도전적이다. AAV 벡터는 전달될 수 있는 상동성 아암의 크기를 제한하는 캡슐화된 DNA를 포함한다. 또한, AAV와 관련된 복잡성 및 고비용은 생체외 유전자 편집에 대한 유용성을 제한한다. 또한, AAV 벡터는 배양시 세포에서 충분한 상동성 지정 복구를 유도하기 위해 매우 높은 역가에서 요구된다.
본원에 기재된 바와 같은 CeDNA 벡터는 DNA 공여체 주형의 AAV 벡터-매개 전달과 관련된 많은 문제를 극복할 수 있다. 예를 들어, ceDNA 벡터는 통상적인 AAV 벡터와 함께 사용되는 것보다 더 긴 상동성 아암을 갖는 공여체 주형의 사용을 허용하며, 이는 더 높은 온-표적 및 더 낮은 오프-표적 효과로 보다 효율적인 유전자 편집을 가능하게 하는 이점을 제공한다.
예시적인 목적으로, 생체외에서 WAS 유전자 CD34+ 줄기세포를 편집하기 위한 ceDNA 벡터를 생성하는 것과 관련하여 예시적인 유전자 편집 ceDNA 벡터가 기술되며, 아래에서 설명된다. 그러나, WAS 유전자의 표적화는 본원에 기재된 바와 같은 방법 및 작제물에 유용한 유전자 편집 벡터 ceDNA를 생성하는 방법을 예시하기 위해 본 실시예에 예시되어 있지만, 당업자는, 상기 사용에 언급된 바와 같이, 유전자 편집이 요구되는 임의의 유전자를 사용될 수 있다는 것을 알고 있다. 편집을 위한 예시적인 유전자는, 예를 들어, "유전자 편집 ceDNA로 치료될 예시적인 질환" 및 "유전자 편집을 위한 추가 질환"이라는 제목의 섹션에서 본원에 기재되어 있다. 또한, 본 예시적인 예에서 조혈 CD34+ 세포의 게놈이 변형되지만, 당업자는 체세포, 배양 세포, 및 줄기세포 및/또는 다능성 세포를 포함하는 임의의 세포, 예를 들어, 본원에서 "숙주 세포"라는 제목의 섹션 XII.A.에 기재된 바와 같은 임의의 세포가 생체외 또는 생체내에서 변형될 수 있음을 알고 있다.
생체내 실험은 배양시 유전자 편집을 수행하기 위해 위스콧-알드리치 증후군 (WAS) 유전자 오픈 리딩 프레임 (ORF)을 인코딩하는 ceDNA 벡터의 능력을 시험하기 위해 인간 CD34+ 조혈 줄기세포를 사용하여 수행될 것이다. 5개의 상이한 처리 아암을 포함하는 예시적인 실험이 이하 본원에 개괄되어 있다.
먼저, ceDNA 벡터는 AAV 벡터를 사용하여 현재 전달되고 WAS ORF (미니유전자; 엑손만) 및 상동성 영역을 인코딩하는 작제물을 전달하는데 사용될 것이다. 유전자 편집 결과의 효율은 AAV (30-40%)를 사용하여 달성된 효율과 비교될 것이고 ceDNA 벡터-기반 전달이 동일한 미니유전자의 AAV-매개 전달 효율을 충족시거나 초과할 수 있는지를 결정할 것이다.
다음으로, ceDNA 벡터를 사용하여 인트론-1을 보유한 WAS 미니유전자 (엑손만)를 전달할 것이다. 인트론-1은 WAS 단백질의 발현에 결정적인 것으로 밝혀졌지만, 인트론-1은 AAV 벡터의 크기 제한을 초과한다. 따라서, WAS 미니유전자 + 인트론-1의 성공적인 전달은 ceDNA 벡터가 유전자 편집, WAS 미니유전자 + 인트론-1의 표적화된 전달 및 WAS 단백질의 성공적인 발현에 대해 AAV 벡터보다 우수함을 보여줄 것이다.
다음으로, WAS ORF 미니유전자 또는 WAS 미니유전자 + 인트론-1을 포함하는 ceDNA 벡터는 더 긴 상동성 아암으로 설계되어, 그러한 상동성 아암의 증가된 길이가 유전자 편집의 온-표적 효율 또는 오프-표적 정확도에 영향을 미치는지를 평가할 것이다.
다음으로, WAS ORF 미니유전자 또는 WAS 미니유전자 + 인트론-1을 포함하는 ceDNA 벡터는 Cas9 부위의 존재가 ceDNA 벡터의 비틀림 긴장을 해제함으로써 공여체 주형으로서 작용하는 ceDNA의 효율을 향상시키는지를 결정하기 위해 ceDNA 상에 Cas9 절단 부위를 포함하도록 설계된다. 마지막으로, ceDNA-GFP/ceDNA-LUC와 같은 리포터 작제물을 인코딩하는 ceDNA 벡터는 생체외 세포에서 ceDNA 벡터의 전기천공 효율을 최적화 및/또는 최대화하도록 설계될 수 있다.
실시예 17 : 배양 세포에서 유전자 편집을 위한 예시적인 작업-흐름 방법(들)
또 다른 예에서, 도 19에 도시된 방법은 배양 세포에서 유전자 편집을 수행하기 위해 본원에서 사용된다. 예를 들어, 본 발명의 임의의 ceDNA 벡터는 본 출원 및 실시예에 기재된 방법을 통해 간 세포 배양물과 같은 배양 세포로 전달될 수 있다. 이어서, 세포를 NHEJ 경로를 사용하여 (ceDNA 벡터가 HDR 주형을 포함하지 않는 경우) 또는 HDR 경로를 통해 (ceDNA 벡터가 HDR 주형을 포함하는 경우) 유전자 편집을 수행하기에 충분한 시간 동안 및 조건 하에서 인큐베이션한다. 성공적인 유전자 편집이 발생했는지를 결정하기 위해, 세포를 공여체 주형 단백질, 예를 들어, 인자 IX (FIX)의 발현에 대해 분석하거나 게놈 표적 DNA의 심층 시퀀싱에 의해 분석하여 공여체 주형의 성공적인 혼입이 발생했는지를 결정할 수 있다. 이 실시예와 관련하여 추가로 고려해야 할 사항은 아래에 개괄된다.
가이드 RNA의 설계: 당 업계에 공지된 임의의 방법을 사용하여 본 발명의 ceDNA 벡터에 혼입시키기 위한 표적 유전자 편집 부위에 상동성을 갖는 맞춤형 가이드 RNA (gRNA)를 설계 및/또는 합성할 수 있다. 본 명세서에서, 맞춤형 gRNA는 다수의 공급업체, 예를 들어, ThermoFisher를 통해 설계되고 합성될 수 있음이 구체적으로 고려된다.
1차 세포 배양: 일부 구현예에서, 간세포와 같은 간 세포에서 유전자 편집 부위를 표적화하는 것이 바람직할 수 있다. 이는, 예를 들어, 알부민 유전자 내의 유전자 편집 부위를 이용함으로써 달성될 수 있다. 당업자는, 간 세포를 포함한 1차 세포의 성공적인 단리 및 성장이 어려울 수 있으며, 해동 및 플레이팅 절차, 플레이트 코팅, Matrigel™ 및/또는 성장 배지 (예를 들어, 간세포 기본 배지)의 최적화를 필요로 할 수 있음을 이해할 것이다. 일 구현예에서, 간 세포 배양 방법은 당 업계, 예를 들어, 하기에 공지된 방법에서 유래된다: Sharma Blood (2017) (상기), 이의 내용은 그 전문이 본원에 참고로 포함된다. 1차 간세포에 대한 성장 조건은 해동 배지, 플레이팅 배지, 인큐베이션 배지 및 매트릭스 시약 (GelTrex™, MatriGel™)과 같이, 예를 들어, ThermoFisher의 시약을 최적화될 수 있다.
HDR 주형 ceDNA: 비제한적인 예에서, HDR 주형을 포함하는 ceDNA 벡터는 도 19에 도시된 바와 같이 설계된다. 예를 들어, ceDNA 벡터는 원하는 길이를 갖는 5' 상동성 아암 및 원하는 길이의 3' 상동성 아암을 포함할 수 있다. 비특이적 효과를 배제하기 위해, 본 명세서에서 ceDNA는 (i) 공여체 주형 서열의 존재 또는 부재하에 5' 상동성 아암 단독, 또는 (ii) 공여체 주형 서열의 존재 또는 부재하에 3' 상동성 아암 단독을 포함하는 ceDNA 대조군이 전체 HDR 주형 (예를 들어, 5' 상동성 아암, 공여체 주형 서열, 및 3' 상동성 아암)을 포함하는 ceDNA 벡터와 실질적으로 유사한 프로토콜로 사용될 수 있음이 구체적으로 고려된다. 이들 대조군은 당업자가, 존재한다면, 상동성 아암에 의해 단독으로 생성될 수 있는 비-특이적 또는 오프-표적 효과를 식별할 수 있게 할 것이다.
실시예 18: 대상체에서
생체내
유전자 편집을 위한 예시적인 작업-흐름 방법(들)
실시예 17에 기재된 방법 및 ceDNA 작제물은 다세포 유기체, 예를 들어, 동물 또는 인간에서 유전자 편집을 수행하도록 적응될 수 있다. ceDNA 벡터는 임의의 편리한 방식으로 유기체 (예를 들어, 마우스)의 배아 줄기세포로 전달될 수 있다. 일부 예에서 유기체는 비-인간 유기체이다. 유기체는 동물 질환 모델의 생성을 위한 설치류 또는 동물 (예를 들어, 비-인간 영장류)일 수 있다. 적절히 재조합된 이식유전자의 존재를 보장하기 위해 생성된 세포를 스크리닝한다. 양성 세포는 야생형 유기체 (예를 들어, 마우스 및 비-인간 설치류)에 이식될 수 있고, 그 결과 자손은 이식유전자의 존재에 대해 스크리닝된다. 표적화된 돌연변이가 유기체의 임의의 균주 (예를 들어, 마우스) 내의 관심 유전자에서 이루어질 수 있기 때문에, 자손의 역교배(backcrossing)는 원하는 유전자 배경에서 형질전환 자손을 얻는데 필요하지 않다.
예시적인 목적으로, 본 실시예는 동물 모델을 생성하기 위해 세포를 편집하기 위한 ceDNA 벡터를 생성하는 것과 관련하여 예시적인 유전자 편집 ceDNA 벡터를 사용하는 것을 논의한다. 그러나, 본원에 기재된 유전자 편집 벡터 ceDNA를 사용하는 방법을 설명하기 위해 동물 (예를 들어, 마우스)의 변형이 본 실시예에 예시되어 있지만, 당업자는, 상기 사용에 언급된 바와 같이, 유전자 편집이 요구되는 임의의 유기체 또는 대상체의 세포 상에 ceDNA 벡터를 사용할 수 있다는 것을 알고 있다. 유전자 편집을 위한 예시적인 대상체는 본원에서 "대상체"의 정의에서 논의된다.
참조
본 출원 및 본원의 실시예에 인용된 특허 및 특허 출원을 포함하지만 이에 제한되지 않는 모든 간행물 및 참고 문헌은, 각각의 개별 간행물 또는 참고 문헌이 본 명세서에 완전히 제시된 것으로 본원에 참고로 포함되는 것으로 구체적이고 개별적으로 표시된 것처럼 그 전문이 참고로 포함된다. 본 출원이 우선권을 주장하는 임의의 특허 출원도 간행물 및 참고 문헌에 대해 전술한 방식으로 본원에 참고로 포함된다.