RU2197992C2 - Генная терапия заболеваний сосудов с помощью специфического к клетке и зависимого от клеточного цикла активного вещества - Google Patents
Генная терапия заболеваний сосудов с помощью специфического к клетке и зависимого от клеточного цикла активного вещества Download PDFInfo
- Publication number
- RU2197992C2 RU2197992C2 RU97104482/14A RU97104482A RU2197992C2 RU 2197992 C2 RU2197992 C2 RU 2197992C2 RU 97104482/14 A RU97104482/14 A RU 97104482/14A RU 97104482 A RU97104482 A RU 97104482A RU 2197992 C2 RU2197992 C2 RU 2197992C2
- Authority
- RU
- Russia
- Prior art keywords
- interleukin
- sequence
- dna construct
- dna
- construct according
- Prior art date
Links
- 239000013543 active substance Substances 0.000 title claims abstract description 51
- 238000000034 method Methods 0.000 title abstract description 8
- 238000002560 therapeutic procedure Methods 0.000 title description 5
- 208000019553 vascular disease Diseases 0.000 title description 5
- 230000001413 cellular effect Effects 0.000 title 1
- 230000002068 genetic effect Effects 0.000 title 1
- 210000004027 cell Anatomy 0.000 claims abstract description 82
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 79
- 108020004414 DNA Proteins 0.000 claims abstract description 67
- 210000000329 smooth muscle myocyte Anatomy 0.000 claims abstract description 55
- 108091028043 Nucleic acid sequence Proteins 0.000 claims abstract description 46
- 239000003446 ligand Substances 0.000 claims abstract description 27
- 239000003130 blood coagulation factor inhibitor Substances 0.000 claims abstract description 19
- 210000002540 macrophage Anatomy 0.000 claims abstract description 17
- 239000013598 vector Substances 0.000 claims abstract description 16
- 210000004698 lymphocyte Anatomy 0.000 claims abstract description 15
- 239000003814 drug Substances 0.000 claims abstract description 11
- 241000700605 Viruses Species 0.000 claims abstract description 8
- 102000004169 proteins and genes Human genes 0.000 claims description 29
- 239000012190 activator Substances 0.000 claims description 27
- 210000002889 endothelial cell Anatomy 0.000 claims description 25
- 102000005962 receptors Human genes 0.000 claims description 24
- 108020003175 receptors Proteins 0.000 claims description 24
- 230000027455 binding Effects 0.000 claims description 22
- 230000022131 cell cycle Effects 0.000 claims description 19
- 108010056785 Myogenin Proteins 0.000 claims description 18
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 claims description 18
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 claims description 18
- 230000001105 regulatory effect Effects 0.000 claims description 18
- 229960000187 tissue plasminogen activator Drugs 0.000 claims description 18
- 239000012528 membrane Substances 0.000 claims description 16
- 102000004190 Enzymes Human genes 0.000 claims description 15
- 108090000790 Enzymes Proteins 0.000 claims description 15
- 108010076504 Protein Sorting Signals Proteins 0.000 claims description 15
- 229940088598 enzyme Drugs 0.000 claims description 15
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 claims description 14
- 239000003112 inhibitor Substances 0.000 claims description 14
- 229940123587 Cell cycle inhibitor Drugs 0.000 claims description 12
- 108010060309 Glucuronidase Proteins 0.000 claims description 11
- 238000001246 colloidal dispersion Methods 0.000 claims description 11
- 230000002062 proliferating effect Effects 0.000 claims description 11
- 102000053187 Glucuronidase Human genes 0.000 claims description 10
- 102000040945 Transcription factor Human genes 0.000 claims description 10
- 108091023040 Transcription factor Proteins 0.000 claims description 10
- 108090000435 Urokinase-type plasminogen activator Proteins 0.000 claims description 10
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 claims description 10
- 239000000824 cytostatic agent Substances 0.000 claims description 10
- 102000008070 Interferon-gamma Human genes 0.000 claims description 9
- 108010074328 Interferon-gamma Proteins 0.000 claims description 9
- 102000000589 Interleukin-1 Human genes 0.000 claims description 9
- 108010002352 Interleukin-1 Proteins 0.000 claims description 9
- 150000001413 amino acids Chemical group 0.000 claims description 9
- 239000002773 nucleotide Substances 0.000 claims description 9
- 125000003729 nucleotide group Chemical group 0.000 claims description 9
- 210000004204 blood vessel Anatomy 0.000 claims description 8
- 229940079593 drug Drugs 0.000 claims description 8
- 239000012634 fragment Substances 0.000 claims description 8
- 229940044627 gamma-interferon Drugs 0.000 claims description 8
- 239000003102 growth factor Substances 0.000 claims description 8
- 239000013612 plasmid Substances 0.000 claims description 8
- 108700028146 Genetic Enhancer Elements Proteins 0.000 claims description 7
- 102100025390 Integrin beta-2 Human genes 0.000 claims description 7
- 102000004388 Interleukin-4 Human genes 0.000 claims description 7
- 108090000978 Interleukin-4 Proteins 0.000 claims description 7
- 239000003623 enhancer Substances 0.000 claims description 7
- 230000002401 inhibitory effect Effects 0.000 claims description 7
- 229940028885 interleukin-4 Drugs 0.000 claims description 7
- 210000003205 muscle Anatomy 0.000 claims description 7
- 230000035772 mutation Effects 0.000 claims description 7
- 210000003705 ribosome Anatomy 0.000 claims description 7
- 210000001519 tissue Anatomy 0.000 claims description 7
- 102000004127 Cytokines Human genes 0.000 claims description 6
- 108090000695 Cytokines Proteins 0.000 claims description 6
- 102000000588 Interleukin-2 Human genes 0.000 claims description 6
- 108010002350 Interleukin-2 Proteins 0.000 claims description 6
- 102000000646 Interleukin-3 Human genes 0.000 claims description 6
- 108010002386 Interleukin-3 Proteins 0.000 claims description 6
- 108010000499 Thromboplastin Proteins 0.000 claims description 6
- 102000002262 Thromboplastin Human genes 0.000 claims description 6
- 125000000266 alpha-aminoacyl group Chemical group 0.000 claims description 6
- 230000001580 bacterial effect Effects 0.000 claims description 6
- 230000001085 cytostatic effect Effects 0.000 claims description 6
- 229940076264 interleukin-3 Drugs 0.000 claims description 6
- 230000037353 metabolic pathway Effects 0.000 claims description 6
- 102000004411 Antithrombin III Human genes 0.000 claims description 5
- 108090000935 Antithrombin III Proteins 0.000 claims description 5
- 102100025064 Cellular tumor antigen p53 Human genes 0.000 claims description 5
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 claims description 5
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 claims description 5
- 102000007625 Hirudins Human genes 0.000 claims description 5
- 108010007267 Hirudins Proteins 0.000 claims description 5
- 102000004889 Interleukin-6 Human genes 0.000 claims description 5
- 108090001005 Interleukin-6 Proteins 0.000 claims description 5
- 101800004937 Protein C Proteins 0.000 claims description 5
- 102000017975 Protein C Human genes 0.000 claims description 5
- 108050002653 Retinoblastoma protein Proteins 0.000 claims description 5
- 101800001700 Saposin-D Proteins 0.000 claims description 5
- 102000006601 Thymidine Kinase Human genes 0.000 claims description 5
- 108020004440 Thymidine kinase Proteins 0.000 claims description 5
- 108060008682 Tumor Necrosis Factor Proteins 0.000 claims description 5
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 claims description 5
- 108010000134 Vascular Cell Adhesion Molecule-1 Proteins 0.000 claims description 5
- 230000004913 activation Effects 0.000 claims description 5
- 229960005348 antithrombin iii Drugs 0.000 claims description 5
- 230000033228 biological regulation Effects 0.000 claims description 5
- 230000001419 dependent effect Effects 0.000 claims description 5
- 230000003511 endothelial effect Effects 0.000 claims description 5
- 229940006607 hirudin Drugs 0.000 claims description 5
- 229940100601 interleukin-6 Drugs 0.000 claims description 5
- 230000002265 prevention Effects 0.000 claims description 5
- 229960000856 protein c Drugs 0.000 claims description 5
- 230000028327 secretion Effects 0.000 claims description 5
- 102000007469 Actins Human genes 0.000 claims description 4
- 108010085238 Actins Proteins 0.000 claims description 4
- 102000005367 Carboxypeptidases Human genes 0.000 claims description 4
- 108010006303 Carboxypeptidases Proteins 0.000 claims description 4
- 101150021185 FGF gene Proteins 0.000 claims description 4
- 108091008794 FGF receptors Proteins 0.000 claims description 4
- 102000044168 Fibroblast Growth Factor Receptor Human genes 0.000 claims description 4
- 102000003814 Interleukin-10 Human genes 0.000 claims description 4
- 108090000174 Interleukin-10 Proteins 0.000 claims description 4
- 108010064548 Lymphocyte Function-Associated Antigen-1 Proteins 0.000 claims description 4
- 101100013973 Mus musculus Gata4 gene Proteins 0.000 claims description 4
- 108091008606 PDGF receptors Proteins 0.000 claims description 4
- 102000003992 Peroxidases Human genes 0.000 claims description 4
- 102000011653 Platelet-Derived Growth Factor Receptors Human genes 0.000 claims description 4
- 241000700584 Simplexvirus Species 0.000 claims description 4
- 108010051583 Ventricular Myosins Proteins 0.000 claims description 4
- WQPDUTSPKFMPDP-OUMQNGNKSA-N hirudin Chemical compound C([C@@H](C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC=1C=CC(OS(O)(=O)=O)=CC=1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CCCCN)NC(=O)[C@H]1N(CCC1)C(=O)[C@@H](NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)CNC(=O)[C@@H](NC(=O)[C@@H](NC(=O)[C@H]1NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(O)=O)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CO)NC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H]2CSSC[C@@H](C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(=O)N[C@H](C(NCC(=O)N[C@@H](CCC(N)=O)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N2)=O)CSSC1)C(C)C)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]1NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)CNC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=2C=CC(O)=CC=2)NC(=O)[C@@H](NC(=O)[C@@H](N)C(C)C)C(C)C)[C@@H](C)O)CSSC1)C(C)C)[C@@H](C)O)[C@@H](C)O)C1=CC=CC=C1 WQPDUTSPKFMPDP-OUMQNGNKSA-N 0.000 claims description 4
- 229940076144 interleukin-10 Drugs 0.000 claims description 4
- 108040007629 peroxidase activity proteins Proteins 0.000 claims description 4
- 239000002243 precursor Substances 0.000 claims description 4
- LOGFVTREOLYCPF-KXNHARMFSA-N (2s,3r)-2-[[(2r)-1-[(2s)-2,6-diaminohexanoyl]pyrrolidine-2-carbonyl]amino]-3-hydroxybutanoic acid Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H]1CCCN1C(=O)[C@@H](N)CCCCN LOGFVTREOLYCPF-KXNHARMFSA-N 0.000 claims description 3
- 101710157736 ATP-dependent 6-phosphofructokinase Proteins 0.000 claims description 3
- 102100036912 Desmin Human genes 0.000 claims description 3
- 108010044052 Desmin Proteins 0.000 claims description 3
- 102100037241 Endoglin Human genes 0.000 claims description 3
- 108010036395 Endoglin Proteins 0.000 claims description 3
- 102000002045 Endothelin Human genes 0.000 claims description 3
- 108050009340 Endothelin Proteins 0.000 claims description 3
- 108010092372 Granulocyte-Macrophage Colony-Stimulating Factor Receptors Proteins 0.000 claims description 3
- 102000016355 Granulocyte-Macrophage Colony-Stimulating Factor Receptors Human genes 0.000 claims description 3
- 101001046686 Homo sapiens Integrin alpha-M Proteins 0.000 claims description 3
- 101000935040 Homo sapiens Integrin beta-2 Proteins 0.000 claims description 3
- 102000003777 Interleukin-1 beta Human genes 0.000 claims description 3
- 108090000193 Interleukin-1 beta Proteins 0.000 claims description 3
- 102000003815 Interleukin-11 Human genes 0.000 claims description 3
- 108090000177 Interleukin-11 Proteins 0.000 claims description 3
- 102000003816 Interleukin-13 Human genes 0.000 claims description 3
- 108090000176 Interleukin-13 Proteins 0.000 claims description 3
- 108010002616 Interleukin-5 Proteins 0.000 claims description 3
- 102100021592 Interleukin-7 Human genes 0.000 claims description 3
- 108010002586 Interleukin-7 Proteins 0.000 claims description 3
- 102000004890 Interleukin-8 Human genes 0.000 claims description 3
- 108090001007 Interleukin-8 Proteins 0.000 claims description 3
- 108090000362 Lymphotoxin-beta Proteins 0.000 claims description 3
- 108010058398 Macrophage Colony-Stimulating Factor Receptor Proteins 0.000 claims description 3
- 101000962498 Macropis fulvipes Macropin Proteins 0.000 claims description 3
- 102000004316 Oxidoreductases Human genes 0.000 claims description 3
- 108090000854 Oxidoreductases Proteins 0.000 claims description 3
- 102000004887 Transforming Growth Factor beta Human genes 0.000 claims description 3
- 108090001012 Transforming Growth Factor beta Proteins 0.000 claims description 3
- 102000005937 Tropomyosin Human genes 0.000 claims description 3
- 108010030743 Tropomyosin Proteins 0.000 claims description 3
- 102000013534 Troponin C Human genes 0.000 claims description 3
- 102000003990 Urokinase-type plasminogen activator Human genes 0.000 claims description 3
- 102000015395 alpha 1-Antitrypsin Human genes 0.000 claims description 3
- 108010050122 alpha 1-Antitrypsin Proteins 0.000 claims description 3
- 229940024142 alpha 1-antitrypsin Drugs 0.000 claims description 3
- 210000005045 desmin Anatomy 0.000 claims description 3
- 229940074383 interleukin-11 Drugs 0.000 claims description 3
- 229940100602 interleukin-5 Drugs 0.000 claims description 3
- 229940100994 interleukin-7 Drugs 0.000 claims description 3
- 229940096397 interleukin-8 Drugs 0.000 claims description 3
- 238000001361 intraarterial administration Methods 0.000 claims description 3
- 238000002955 isolation Methods 0.000 claims description 3
- 208000032839 leukemia Diseases 0.000 claims description 3
- KUMKLUDNETVLDS-UHFFFAOYSA-N ser-601 Chemical compound O=C1C2=CC(C(C)C)=CC=C2N(CCCCC)C=C1C(=O)NC1(C2)CC(C3)CC2CC3C1 KUMKLUDNETVLDS-UHFFFAOYSA-N 0.000 claims description 3
- 241000701161 unidentified adenovirus Species 0.000 claims description 3
- 201000006082 Chickenpox Diseases 0.000 claims description 2
- 108010009685 Cholinergic Receptors Proteins 0.000 claims description 2
- 102000000311 Cytosine Deaminase Human genes 0.000 claims description 2
- 108010080611 Cytosine Deaminase Proteins 0.000 claims description 2
- 108060006698 EGF receptor Proteins 0.000 claims description 2
- 102000001301 EGF receptor Human genes 0.000 claims description 2
- 241000196324 Embryophyta Species 0.000 claims description 2
- 102000010180 Endothelin receptor Human genes 0.000 claims description 2
- 108050001739 Endothelin receptor Proteins 0.000 claims description 2
- 102000054184 GADD45 Human genes 0.000 claims description 2
- 108700007032 GADD45 Proteins 0.000 claims description 2
- 102400001301 Gasdermin-B, C-terminal Human genes 0.000 claims description 2
- 102000001398 Granzyme Human genes 0.000 claims description 2
- 108060005986 Granzyme Proteins 0.000 claims description 2
- 108010008212 Integrin alpha4beta1 Proteins 0.000 claims description 2
- 108090000467 Interferon-beta Proteins 0.000 claims description 2
- 102000003996 Interferon-beta Human genes 0.000 claims description 2
- 102000013462 Interleukin-12 Human genes 0.000 claims description 2
- 108010065805 Interleukin-12 Proteins 0.000 claims description 2
- 108010092694 L-Selectin Proteins 0.000 claims description 2
- 102000016551 L-selectin Human genes 0.000 claims description 2
- 101710128836 Large T antigen Proteins 0.000 claims description 2
- -1 MRF-4 Proteins 0.000 claims description 2
- 102000007651 Macrophage Colony-Stimulating Factor Human genes 0.000 claims description 2
- 102000019218 Mannose-6-phosphate receptors Human genes 0.000 claims description 2
- 102000004459 Nitroreductase Human genes 0.000 claims description 2
- KHGNFPUMBJSZSM-UHFFFAOYSA-N Perforine Natural products COC1=C2CCC(O)C(CCC(C)(C)O)(OC)C2=NC2=C1C=CO2 KHGNFPUMBJSZSM-UHFFFAOYSA-N 0.000 claims description 2
- 108700019535 Phosphoprotein Phosphatases Proteins 0.000 claims description 2
- 102000045595 Phosphoprotein Phosphatases Human genes 0.000 claims description 2
- 108060006706 SRC Proteins 0.000 claims description 2
- 206010046980 Varicella Diseases 0.000 claims description 2
- 102000034337 acetylcholine receptors Human genes 0.000 claims description 2
- 239000000853 adhesive Substances 0.000 claims description 2
- 230000001070 adhesive effect Effects 0.000 claims description 2
- 102000055574 bcl-2 Homologous Antagonist-Killer Human genes 0.000 claims description 2
- 108700039689 bcl-2 Homologous Antagonist-Killer Proteins 0.000 claims description 2
- 108700021031 cdc Genes Proteins 0.000 claims description 2
- 239000003145 cytotoxic factor Substances 0.000 claims description 2
- 201000010099 disease Diseases 0.000 claims description 2
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 2
- 230000000977 initiatory effect Effects 0.000 claims description 2
- 229940117681 interleukin-12 Drugs 0.000 claims description 2
- 239000002502 liposome Substances 0.000 claims description 2
- 210000003712 lysosome Anatomy 0.000 claims description 2
- 230000001868 lysosomic effect Effects 0.000 claims description 2
- 125000000311 mannosyl group Chemical group C1([C@@H](O)[C@@H](O)[C@H](O)[C@H](O1)CO)* 0.000 claims description 2
- 108020001162 nitroreductase Proteins 0.000 claims description 2
- 229930192851 perforin Natural products 0.000 claims description 2
- 108091008598 receptor tyrosine kinases Proteins 0.000 claims description 2
- 102000027426 receptor tyrosine kinases Human genes 0.000 claims description 2
- ZRKFYGHZFMAOKI-QMGMOQQFSA-N tgfbeta Chemical compound C([C@H](NC(=O)[C@H](C(C)C)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC(C)C)NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O)C1=CC=C(O)C=C1 ZRKFYGHZFMAOKI-QMGMOQQFSA-N 0.000 claims description 2
- 229960005356 urokinase Drugs 0.000 claims description 2
- 108010047303 von Willebrand Factor Proteins 0.000 claims description 2
- 102100036537 von Willebrand factor Human genes 0.000 claims description 2
- 229960001134 von willebrand factor Drugs 0.000 claims description 2
- 210000004379 membrane Anatomy 0.000 claims 5
- 102100031358 Urokinase-type plasminogen activator Human genes 0.000 claims 4
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 claims 3
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 claims 3
- 238000002347 injection Methods 0.000 claims 3
- 239000007924 injection Substances 0.000 claims 3
- 108090000266 Cyclin-dependent kinases Proteins 0.000 claims 2
- 102000003903 Cyclin-dependent kinases Human genes 0.000 claims 2
- 102000014150 Interferons Human genes 0.000 claims 2
- 108010050904 Interferons Proteins 0.000 claims 2
- 102000019223 Interleukin-1 receptor Human genes 0.000 claims 2
- 108050006617 Interleukin-1 receptor Proteins 0.000 claims 2
- 102000004125 Interleukin-1alpha Human genes 0.000 claims 2
- 108010082786 Interleukin-1alpha Proteins 0.000 claims 2
- ZUBDGKVDJUIMQQ-UBFCDGJISA-N endothelin-1 Chemical compound C([C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(O)=O)NC(=O)[C@H]1NC(=O)[C@H](CC=2C=CC=CC=2)NC(=O)[C@@H](CC=2C=CC(O)=CC=2)NC(=O)[C@H](C(C)C)NC(=O)[C@H]2CSSC[C@@H](C(N[C@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@H](CC(C)C)C(=O)N[C@@H](CCSC)C(=O)N[C@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N2)=O)NC(=O)[C@@H](CO)NC(=O)[C@H](N)CSSC1)C1=CNC=N1 ZUBDGKVDJUIMQQ-UBFCDGJISA-N 0.000 claims 2
- XKTZWUACRZHVAN-VADRZIEHSA-N interleukin-8 Chemical compound C([C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@@H](NC(C)=O)CCSC)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CCSC)C(=O)N1[C@H](CCC1)C(=O)N1[C@H](CCC1)C(=O)N[C@@H](C)C(=O)N[C@H](CC(O)=O)C(=O)N[C@H](CCC(O)=O)C(=O)N[C@H](CC(O)=O)C(=O)N[C@H](CC=1C=CC(O)=CC=1)C(=O)N[C@H](CO)C(=O)N1[C@H](CCC1)C(N)=O)C1=CC=CC=C1 XKTZWUACRZHVAN-VADRZIEHSA-N 0.000 claims 2
- 210000003491 skin Anatomy 0.000 claims 2
- 108010064733 Angiotensins Proteins 0.000 claims 1
- 102000015427 Angiotensins Human genes 0.000 claims 1
- 241000193755 Bacillus cereus Species 0.000 claims 1
- 241000894006 Bacteria Species 0.000 claims 1
- 102100026189 Beta-galactosidase Human genes 0.000 claims 1
- 101100289995 Caenorhabditis elegans mac-1 gene Proteins 0.000 claims 1
- 102000019034 Chemokines Human genes 0.000 claims 1
- 108010012236 Chemokines Proteins 0.000 claims 1
- 101710154385 D-aminopeptidase Proteins 0.000 claims 1
- 241000702421 Dependoparvovirus Species 0.000 claims 1
- 108090000371 Esterases Proteins 0.000 claims 1
- 101710082961 GATA-binding factor 2 Proteins 0.000 claims 1
- 108010031186 Glycoside Hydrolases Proteins 0.000 claims 1
- 102000005744 Glycoside Hydrolases Human genes 0.000 claims 1
- 108010031792 IGF Type 2 Receptor Proteins 0.000 claims 1
- 102000006992 Interferon-alpha Human genes 0.000 claims 1
- 108010047761 Interferon-alpha Proteins 0.000 claims 1
- 102000010789 Interleukin-2 Receptors Human genes 0.000 claims 1
- 108010038453 Interleukin-2 Receptors Proteins 0.000 claims 1
- 108010038452 Interleukin-3 Receptors Proteins 0.000 claims 1
- 102000010790 Interleukin-3 Receptors Human genes 0.000 claims 1
- 102000010787 Interleukin-4 Receptors Human genes 0.000 claims 1
- 108010038486 Interleukin-4 Receptors Proteins 0.000 claims 1
- 102000000743 Interleukin-5 Human genes 0.000 claims 1
- 102000000585 Interleukin-9 Human genes 0.000 claims 1
- 108010002335 Interleukin-9 Proteins 0.000 claims 1
- 108010059881 Lactase Proteins 0.000 claims 1
- 102000004083 Lymphotoxin-alpha Human genes 0.000 claims 1
- 108090000542 Lymphotoxin-alpha Proteins 0.000 claims 1
- 108010046938 Macrophage Colony-Stimulating Factor Proteins 0.000 claims 1
- 102000004364 Myogenin Human genes 0.000 claims 1
- 108010058765 Oncogene Protein pp60(v-src) Proteins 0.000 claims 1
- 108010056995 Perforin Proteins 0.000 claims 1
- 102000004503 Perforin Human genes 0.000 claims 1
- 241000589516 Pseudomonas Species 0.000 claims 1
- 108090000919 Pyroglutamyl-Peptidase I Proteins 0.000 claims 1
- 102000001332 SRC Human genes 0.000 claims 1
- 102100030951 Tissue factor pathway inhibitor Human genes 0.000 claims 1
- 101710098624 Tyrosine-protein kinase ABL1 Proteins 0.000 claims 1
- 102100023543 Vascular cell adhesion protein 1 Human genes 0.000 claims 1
- 238000009825 accumulation Methods 0.000 claims 1
- 108010005774 beta-Galactosidase Proteins 0.000 claims 1
- 210000002808 connective tissue Anatomy 0.000 claims 1
- 229940079322 interferon Drugs 0.000 claims 1
- 229940047124 interferons Drugs 0.000 claims 1
- 102000002467 interleukin receptors Human genes 0.000 claims 1
- 108010093036 interleukin receptors Proteins 0.000 claims 1
- 229940118526 interleukin-9 Drugs 0.000 claims 1
- 238000010253 intravenous injection Methods 0.000 claims 1
- 210000001503 joint Anatomy 0.000 claims 1
- 210000003734 kidney Anatomy 0.000 claims 1
- 229940116108 lactase Drugs 0.000 claims 1
- 108010013555 lipoprotein-associated coagulation inhibitor Proteins 0.000 claims 1
- 210000004185 liver Anatomy 0.000 claims 1
- 210000004072 lung Anatomy 0.000 claims 1
- 108091005446 macrophage receptors Proteins 0.000 claims 1
- 210000004400 mucous membrane Anatomy 0.000 claims 1
- 210000003281 pleural cavity Anatomy 0.000 claims 1
- 229940083538 smallpox vaccine Drugs 0.000 claims 1
- 210000000952 spleen Anatomy 0.000 claims 1
- 210000000130 stem cell Anatomy 0.000 claims 1
- 210000002330 subarachnoid space Anatomy 0.000 claims 1
- 241001430294 unidentified retrovirus Species 0.000 claims 1
- 239000000126 substance Substances 0.000 abstract description 16
- 230000000694 effects Effects 0.000 abstract description 8
- 230000003213 activating effect Effects 0.000 abstract description 5
- 210000003038 endothelium Anatomy 0.000 abstract description 5
- 230000023555 blood coagulation Effects 0.000 abstract description 2
- 239000003795 chemical substances by application Substances 0.000 abstract description 2
- 238000001994 activation Methods 0.000 abstract 1
- 239000003966 growth inhibitor Substances 0.000 abstract 1
- 206010028980 Neoplasm Diseases 0.000 description 21
- 201000011510 cancer Diseases 0.000 description 20
- 208000007536 Thrombosis Diseases 0.000 description 19
- 102100032970 Myogenin Human genes 0.000 description 18
- 210000004369 blood Anatomy 0.000 description 16
- 239000008280 blood Substances 0.000 description 16
- 239000002253 acid Substances 0.000 description 15
- 238000005345 coagulation Methods 0.000 description 13
- 150000007513 acids Chemical class 0.000 description 12
- 239000003146 anticoagulant agent Substances 0.000 description 12
- 230000015271 coagulation Effects 0.000 description 12
- 230000035755 proliferation Effects 0.000 description 12
- 239000002299 complementary DNA Substances 0.000 description 11
- 239000013603 viral vector Substances 0.000 description 10
- 101000589002 Homo sapiens Myogenin Proteins 0.000 description 8
- 102000001938 Plasminogen Activators Human genes 0.000 description 8
- 108010001014 Plasminogen Activators Proteins 0.000 description 8
- 238000001727 in vivo Methods 0.000 description 8
- 229940127126 plasminogen activator Drugs 0.000 description 8
- 238000013518 transcription Methods 0.000 description 8
- 230000035897 transcription Effects 0.000 description 8
- 208000031481 Pathologic Constriction Diseases 0.000 description 7
- 230000036262 stenosis Effects 0.000 description 7
- 208000037804 stenosis Diseases 0.000 description 7
- 108060003951 Immunoglobulin Proteins 0.000 description 6
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 6
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 6
- 230000002785 anti-thrombosis Effects 0.000 description 6
- 238000010276 construction Methods 0.000 description 6
- 102000018358 immunoglobulin Human genes 0.000 description 6
- 230000005764 inhibitory process Effects 0.000 description 6
- 238000002360 preparation method Methods 0.000 description 6
- 230000003966 vascular damage Effects 0.000 description 6
- 102000019063 CCAAT-Binding Factor Human genes 0.000 description 5
- 108010026988 CCAAT-Binding Factor Proteins 0.000 description 5
- 101150012716 CDK1 gene Proteins 0.000 description 5
- 108091026890 Coding region Proteins 0.000 description 5
- 101100059559 Emericella nidulans (strain FGSC A4 / ATCC 38163 / CBS 112.46 / NRRL 194 / M139) nimX gene Proteins 0.000 description 5
- 101000933465 Homo sapiens Beta-glucuronidase Proteins 0.000 description 5
- 101100273808 Xenopus laevis cdk1-b gene Proteins 0.000 description 5
- 210000004556 brain Anatomy 0.000 description 5
- 230000006378 damage Effects 0.000 description 5
- 238000002474 experimental method Methods 0.000 description 5
- 230000012010 growth Effects 0.000 description 5
- 239000013600 plasmid vector Substances 0.000 description 5
- 238000012552 review Methods 0.000 description 5
- 230000009885 systemic effect Effects 0.000 description 5
- 101800004490 Endothelin-1 Proteins 0.000 description 4
- 241000991587 Enterovirus C Species 0.000 description 4
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 4
- 102100026019 Interleukin-6 Human genes 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 210000001772 blood platelet Anatomy 0.000 description 4
- 230000010261 cell growth Effects 0.000 description 4
- VAYGXNSJCAHWJZ-UHFFFAOYSA-N dimethyl sulfate Chemical compound COS(=O)(=O)OC VAYGXNSJCAHWJZ-UHFFFAOYSA-N 0.000 description 4
- 239000012636 effector Substances 0.000 description 4
- 229960002897 heparin Drugs 0.000 description 4
- 229920000669 heparin Polymers 0.000 description 4
- 210000002865 immune cell Anatomy 0.000 description 4
- 230000002103 transcriptional effect Effects 0.000 description 4
- 108020003589 5' Untranslated Regions Proteins 0.000 description 3
- 108020000948 Antisense Oligonucleotides Proteins 0.000 description 3
- 206010003210 Arteriosclerosis Diseases 0.000 description 3
- 108050006400 Cyclin Proteins 0.000 description 3
- 108010068192 Cyclin A Proteins 0.000 description 3
- 102100025191 Cyclin-A2 Human genes 0.000 description 3
- 102000030168 Endothelin A Receptor Human genes 0.000 description 3
- 108010090549 Endothelin A Receptor Proteins 0.000 description 3
- 102400000686 Endothelin-1 Human genes 0.000 description 3
- 102000009123 Fibrin Human genes 0.000 description 3
- 108010073385 Fibrin Proteins 0.000 description 3
- BWGVNKXGVNDBDI-UHFFFAOYSA-N Fibrin monomer Chemical compound CNC(=O)CNC(=O)CN BWGVNKXGVNDBDI-UHFFFAOYSA-N 0.000 description 3
- 101100457913 Homo sapiens CDC25C gene Proteins 0.000 description 3
- 108091000080 Phosphotransferase Proteins 0.000 description 3
- 108010039918 Polylysine Proteins 0.000 description 3
- 102100036691 Proliferating cell nuclear antigen Human genes 0.000 description 3
- 241000700159 Rattus Species 0.000 description 3
- 108700025701 Retinoblastoma Genes Proteins 0.000 description 3
- 239000000074 antisense oligonucleotide Substances 0.000 description 3
- 238000012230 antisense oligonucleotides Methods 0.000 description 3
- 229960004676 antithrombotic agent Drugs 0.000 description 3
- 208000011775 arteriosclerosis disease Diseases 0.000 description 3
- 238000013461 design Methods 0.000 description 3
- 229950003499 fibrin Drugs 0.000 description 3
- 229960002963 ganciclovir Drugs 0.000 description 3
- IRSCQMHQWWYFCW-UHFFFAOYSA-N ganciclovir Chemical compound O=C1NC(N)=NC2=C1N=CN2COC(CO)CO IRSCQMHQWWYFCW-UHFFFAOYSA-N 0.000 description 3
- 238000001415 gene therapy Methods 0.000 description 3
- 230000002779 inactivation Effects 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- 238000001990 intravenous administration Methods 0.000 description 3
- 102000006240 membrane receptors Human genes 0.000 description 3
- 108020004084 membrane receptors Proteins 0.000 description 3
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 3
- 208000010125 myocardial infarction Diseases 0.000 description 3
- 102000039446 nucleic acids Human genes 0.000 description 3
- 108020004707 nucleic acids Proteins 0.000 description 3
- 150000007523 nucleic acids Chemical class 0.000 description 3
- 230000026731 phosphorylation Effects 0.000 description 3
- 238000006366 phosphorylation reaction Methods 0.000 description 3
- 229920000656 polylysine Polymers 0.000 description 3
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 230000003612 virological effect Effects 0.000 description 3
- BSYNRYMUTXBXSQ-UHFFFAOYSA-N Aspirin Chemical compound CC(=O)OC1=CC=CC=C1C(O)=O BSYNRYMUTXBXSQ-UHFFFAOYSA-N 0.000 description 2
- ZUBDGKVDJUIMQQ-VVTNISDDSA-N CC[C@H](C)[C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](Cc1c[nH]cn1)NC(=O)[C@@H]1CSSC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H]2CSSC[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC2=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](Cc2ccc(O)cc2)C(=O)N[C@H](Cc2ccccc2)C(=O)N1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](Cc1c[nH]c2ccccc12)C(O)=O Chemical compound CC[C@H](C)[C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](Cc1c[nH]cn1)NC(=O)[C@@H]1CSSC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H]2CSSC[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC2=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](Cc2ccc(O)cc2)C(=O)N[C@H](Cc2ccccc2)C(=O)N1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](Cc1c[nH]c2ccccc12)C(O)=O ZUBDGKVDJUIMQQ-VVTNISDDSA-N 0.000 description 2
- 108020004635 Complementary DNA Proteins 0.000 description 2
- 102000009024 Epidermal Growth Factor Human genes 0.000 description 2
- 102000009465 Growth Factor Receptors Human genes 0.000 description 2
- 108010009202 Growth Factor Receptors Proteins 0.000 description 2
- 101000925493 Homo sapiens Endothelin-1 Proteins 0.000 description 2
- 102100039897 Interleukin-5 Human genes 0.000 description 2
- 108700020796 Oncogene Proteins 0.000 description 2
- 101710098940 Pro-epidermal growth factor Proteins 0.000 description 2
- 108010087776 Proto-Oncogene Proteins c-myb Proteins 0.000 description 2
- 102000009096 Proto-Oncogene Proteins c-myb Human genes 0.000 description 2
- 101001023030 Toxoplasma gondii Myosin-D Proteins 0.000 description 2
- 108091008605 VEGF receptors Proteins 0.000 description 2
- 108010053099 Vascular Endothelial Growth Factor Receptor-2 Proteins 0.000 description 2
- 102000009484 Vascular Endothelial Growth Factor Receptors Human genes 0.000 description 2
- OMOVVBIIQSXZSZ-UHFFFAOYSA-N [6-(4-acetyloxy-5,9a-dimethyl-2,7-dioxo-4,5a,6,9-tetrahydro-3h-pyrano[3,4-b]oxepin-5-yl)-5-formyloxy-3-(furan-3-yl)-3a-methyl-7-methylidene-1a,2,3,4,5,6-hexahydroindeno[1,7a-b]oxiren-4-yl] 2-hydroxy-3-methylpentanoate Chemical compound CC12C(OC(=O)C(O)C(C)CC)C(OC=O)C(C3(C)C(CC(=O)OC4(C)COC(=O)CC43)OC(C)=O)C(=C)C32OC3CC1C=1C=COC=1 OMOVVBIIQSXZSZ-UHFFFAOYSA-N 0.000 description 2
- 229960001138 acetylsalicylic acid Drugs 0.000 description 2
- 230000000996 additive effect Effects 0.000 description 2
- 229940127219 anticoagulant drug Drugs 0.000 description 2
- 229940127218 antiplatelet drug Drugs 0.000 description 2
- 230000032823 cell division Effects 0.000 description 2
- ZYGHJZDHTFUPRJ-UHFFFAOYSA-N coumarin Chemical compound C1=CC=C2OC(=O)C=CC2=C1 ZYGHJZDHTFUPRJ-UHFFFAOYSA-N 0.000 description 2
- 238000002337 electrophoretic mobility shift assay Methods 0.000 description 2
- 208000014674 injury Diseases 0.000 description 2
- 210000002751 lymph Anatomy 0.000 description 2
- 230000007246 mechanism Effects 0.000 description 2
- 230000014508 negative regulation of coagulation Effects 0.000 description 2
- 102000020233 phosphotransferase Human genes 0.000 description 2
- 239000000106 platelet aggregation inhibitor Substances 0.000 description 2
- 230000000069 prophylactic effect Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 239000003001 serine protease inhibitor Substances 0.000 description 2
- 208000011580 syndromic disease Diseases 0.000 description 2
- 230000002195 synergetic effect Effects 0.000 description 2
- 230000001225 therapeutic effect Effects 0.000 description 2
- 230000002792 vascular Effects 0.000 description 2
- AFWRJOYNLMVZQO-GMFATLNBSA-N (1r,2r,4as,8as)-1-[(1e,3e)-5-hydroxy-3-methylpenta-1,3-dienyl]-2,5,5,8a-tetramethyl-3,4,4a,6,7,8-hexahydro-1h-naphthalen-2-ol Chemical compound CC1(C)CCC[C@]2(C)[C@@H](/C=C/C(=C/CO)/C)[C@](C)(O)CC[C@H]21 AFWRJOYNLMVZQO-GMFATLNBSA-N 0.000 description 1
- LAQPKDLYOBZWBT-NYLDSJSYSA-N (2s,4s,5r,6r)-5-acetamido-2-{[(2s,3r,4s,5s,6r)-2-{[(2r,3r,4r,5r)-5-acetamido-1,2-dihydroxy-6-oxo-4-{[(2s,3s,4r,5s,6s)-3,4,5-trihydroxy-6-methyloxan-2-yl]oxy}hexan-3-yl]oxy}-3,5-dihydroxy-6-(hydroxymethyl)oxan-4-yl]oxy}-4-hydroxy-6-[(1r,2r)-1,2,3-trihydrox Chemical compound O[C@H]1[C@H](O)[C@H](O)[C@H](C)O[C@H]1O[C@H]([C@@H](NC(C)=O)C=O)[C@@H]([C@H](O)CO)O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)[C@@H](O)[C@@H](CO)O1 LAQPKDLYOBZWBT-NYLDSJSYSA-N 0.000 description 1
- 108020005065 3' Flanking Region Proteins 0.000 description 1
- 108020005029 5' Flanking Region Proteins 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- 102000008873 Angiotensin II receptor Human genes 0.000 description 1
- 108050000824 Angiotensin II receptor Proteins 0.000 description 1
- 101000574464 Arabidopsis thaliana Purple acid phosphatase 17 Proteins 0.000 description 1
- 102000015081 Blood Coagulation Factors Human genes 0.000 description 1
- 108010039209 Blood Coagulation Factors Proteins 0.000 description 1
- 108010059108 CD18 Antigens Proteins 0.000 description 1
- 101100381481 Caenorhabditis elegans baz-2 gene Proteins 0.000 description 1
- 108010060273 Cyclin A2 Proteins 0.000 description 1
- 230000006820 DNA synthesis Effects 0.000 description 1
- 230000004568 DNA-binding Effects 0.000 description 1
- 108090000204 Dipeptidase 1 Proteins 0.000 description 1
- 101100457919 Drosophila melanogaster stg gene Proteins 0.000 description 1
- 108050002772 E3 ubiquitin-protein ligase Mdm2 Proteins 0.000 description 1
- 102000012199 E3 ubiquitin-protein ligase Mdm2 Human genes 0.000 description 1
- 206010014523 Embolism and thrombosis Diseases 0.000 description 1
- 102000013128 Endothelin B Receptor Human genes 0.000 description 1
- 108010090557 Endothelin B Receptor Proteins 0.000 description 1
- 102000010834 Extracellular Matrix Proteins Human genes 0.000 description 1
- 108010037362 Extracellular Matrix Proteins Proteins 0.000 description 1
- 102000009109 Fc receptors Human genes 0.000 description 1
- 108010087819 Fc receptors Proteins 0.000 description 1
- 102100024785 Fibroblast growth factor 2 Human genes 0.000 description 1
- 108090000379 Fibroblast growth factor 2 Proteins 0.000 description 1
- 230000010337 G2 phase Effects 0.000 description 1
- 102000011852 GATA2 Transcription Factor Human genes 0.000 description 1
- 108010075641 GATA2 Transcription Factor Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 1
- 102000006587 Glutathione peroxidase Human genes 0.000 description 1
- 108700016172 Glutathione peroxidases Proteins 0.000 description 1
- 101000715499 Homo sapiens Catalase Proteins 0.000 description 1
- 101001054867 Homo sapiens Protein-lysine 6-oxidase Proteins 0.000 description 1
- 101000799461 Homo sapiens Thrombopoietin Proteins 0.000 description 1
- 101000694103 Homo sapiens Thyroid peroxidase Proteins 0.000 description 1
- 206010021143 Hypoxia Diseases 0.000 description 1
- 102000004289 Interferon regulatory factor 1 Human genes 0.000 description 1
- 108090000890 Interferon regulatory factor 1 Proteins 0.000 description 1
- 206010025323 Lymphomas Diseases 0.000 description 1
- 108050006616 Mannose-6-phosphate receptors Proteins 0.000 description 1
- 102100031790 Myelin expression factor 2 Human genes 0.000 description 1
- 101710107751 Myelin expression factor 2 Proteins 0.000 description 1
- 102100038380 Myogenic factor 5 Human genes 0.000 description 1
- 101710099061 Myogenic factor 5 Proteins 0.000 description 1
- 102100038379 Myogenic factor 6 Human genes 0.000 description 1
- BKAYIFDRRZZKNF-VIFPVBQESA-N N-acetylcarnosine Chemical compound CC(=O)NCCC(=O)N[C@H](C(O)=O)CC1=CN=CN1 BKAYIFDRRZZKNF-VIFPVBQESA-N 0.000 description 1
- 125000001429 N-terminal alpha-amino-acid group Chemical group 0.000 description 1
- 206010028851 Necrosis Diseases 0.000 description 1
- 108700019961 Neoplasm Genes Proteins 0.000 description 1
- 102000048850 Neoplasm Genes Human genes 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 108700026244 Open Reading Frames Proteins 0.000 description 1
- 241000028382 Paulia Species 0.000 description 1
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 description 1
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 description 1
- 235000008331 Pinus X rigitaeda Nutrition 0.000 description 1
- 235000011613 Pinus brutia Nutrition 0.000 description 1
- 241000018646 Pinus brutia Species 0.000 description 1
- 102000004022 Protein-Tyrosine Kinases Human genes 0.000 description 1
- 108090000412 Protein-Tyrosine Kinases Proteins 0.000 description 1
- 101100372762 Rattus norvegicus Flt1 gene Proteins 0.000 description 1
- 201000000582 Retinoblastoma Diseases 0.000 description 1
- 230000018199 S phase Effects 0.000 description 1
- 102000008847 Serpin Human genes 0.000 description 1
- 108050000761 Serpin Proteins 0.000 description 1
- 108010023197 Streptokinase Proteins 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- 108091005735 TGF-beta receptors Proteins 0.000 description 1
- 102100031142 Transcriptional repressor protein YY1 Human genes 0.000 description 1
- 102000016715 Transforming Growth Factor beta Receptors Human genes 0.000 description 1
- 108010009583 Transforming Growth Factors Proteins 0.000 description 1
- 102000009618 Transforming Growth Factors Human genes 0.000 description 1
- 102400001320 Transforming growth factor alpha Human genes 0.000 description 1
- 101800004564 Transforming growth factor alpha Proteins 0.000 description 1
- 206010054094 Tumour necrosis Diseases 0.000 description 1
- 102000016548 Vascular Endothelial Growth Factor Receptor-1 Human genes 0.000 description 1
- 108010053096 Vascular Endothelial Growth Factor Receptor-1 Proteins 0.000 description 1
- 102000016549 Vascular Endothelial Growth Factor Receptor-2 Human genes 0.000 description 1
- 206010057469 Vascular stenosis Diseases 0.000 description 1
- 208000027418 Wounds and injury Diseases 0.000 description 1
- 108010042669 YY1 Transcription Factor Proteins 0.000 description 1
- 101710185494 Zinc finger protein Proteins 0.000 description 1
- 102100023597 Zinc finger protein 816 Human genes 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 108091006088 activator proteins Proteins 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 239000000654 additive Substances 0.000 description 1
- 230000002776 aggregation Effects 0.000 description 1
- 238000004220 aggregation Methods 0.000 description 1
- 238000002399 angioplasty Methods 0.000 description 1
- 230000000702 anti-platelet effect Effects 0.000 description 1
- 230000006907 apoptotic process Effects 0.000 description 1
- 102000006635 beta-lactamase Human genes 0.000 description 1
- 230000000740 bleeding effect Effects 0.000 description 1
- 239000003114 blood coagulation factor Substances 0.000 description 1
- 229940019700 blood coagulation factors Drugs 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 101150069072 cdc25 gene Proteins 0.000 description 1
- 230000009087 cell motility Effects 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 230000009134 cell regulation Effects 0.000 description 1
- 230000010307 cell transformation Effects 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 230000000295 complement effect Effects 0.000 description 1
- 229960000956 coumarin Drugs 0.000 description 1
- 235000001671 coumarin Nutrition 0.000 description 1
- 229940043378 cyclin-dependent kinase inhibitor Drugs 0.000 description 1
- 230000002559 cytogenic effect Effects 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 238000007405 data analysis Methods 0.000 description 1
- 230000002074 deregulated effect Effects 0.000 description 1
- 230000003831 deregulation Effects 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 239000002158 endotoxin Substances 0.000 description 1
- 230000002327 eosinophilic effect Effects 0.000 description 1
- 210000002744 extracellular matrix Anatomy 0.000 description 1
- 230000020764 fibrinolysis Effects 0.000 description 1
- 239000003527 fibrinolytic agent Substances 0.000 description 1
- 238000010230 functional analysis Methods 0.000 description 1
- 230000004927 fusion Effects 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 150000002308 glutamine derivatives Chemical class 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 239000002565 heparin fraction Substances 0.000 description 1
- 210000003630 histaminocyte Anatomy 0.000 description 1
- 102000045501 human CAT Human genes 0.000 description 1
- 102000051318 human LOX Human genes 0.000 description 1
- 102000053400 human TPO Human genes 0.000 description 1
- 230000001146 hypoxic effect Effects 0.000 description 1
- 230000028993 immune response Effects 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 230000003834 intracellular effect Effects 0.000 description 1
- 229920006008 lipopolysaccharide Polymers 0.000 description 1
- 230000033001 locomotion Effects 0.000 description 1
- 208000030159 metabolic disease Diseases 0.000 description 1
- 102000051367 mu Opioid Receptors Human genes 0.000 description 1
- 108010084677 myogenic factor 6 Proteins 0.000 description 1
- 230000017074 necrotic cell death Effects 0.000 description 1
- 230000006715 negative regulation of smooth muscle cell proliferation Effects 0.000 description 1
- 208000018360 neuromuscular disease Diseases 0.000 description 1
- 239000002547 new drug Substances 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 150000003833 nucleoside derivatives Chemical class 0.000 description 1
- 238000006384 oligomerization reaction Methods 0.000 description 1
- 231100000590 oncogenic Toxicity 0.000 description 1
- 230000002246 oncogenic effect Effects 0.000 description 1
- 210000000496 pancreas Anatomy 0.000 description 1
- 235000020004 porter Nutrition 0.000 description 1
- 230000008569 process Effects 0.000 description 1
- 210000002307 prostate Anatomy 0.000 description 1
- 230000029865 regulation of blood pressure Effects 0.000 description 1
- 210000002460 smooth muscle Anatomy 0.000 description 1
- 208000010110 spontaneous platelet aggregation Diseases 0.000 description 1
- 229960005202 streptokinase Drugs 0.000 description 1
- 229940124597 therapeutic agent Drugs 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000032258 transport Effects 0.000 description 1
- 230000008733 trauma Effects 0.000 description 1
- 239000002753 trypsin inhibitor Substances 0.000 description 1
- 210000005167 vascular cell Anatomy 0.000 description 1
- 231100000216 vascular lesion Toxicity 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- 108020001612 μ-opioid receptors Proteins 0.000 description 1
Images








Landscapes
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
Изобретение относится к генной терапии и касается генной терапии заболеваний сосудов с помощью специфического к клетке и зависимого от клеточного цикла активного вещества. Сущность изобретения включает ДНК-конструкцию, существенными элементами которой являются активаторная последовательность, промоторный модуль и ген активного вещества. Активаторная последовательность активируется специфически к клетке в гладкомышечных клетках, активированных эндотелиальных клетках, активированных макрофагах или активированных лимфоцитах. Это активирование регулируется специфически к клеточному циклу промоторным модулем. Активное вещество представляет собой ингибитор роста гладкомышечных клеток и/или свертывания (крови). Описанную ДНК-последовательность вставляют в вирусный или невирусный вектор, дополненный лигандом со сродством к клетке-мишени. Преимущество изобретения в том, что активное вещество действует только на находящиеся в стадии деления гладкомышечные клетки, подавляя их пролиферацию после повреждения сосудов. 33 з.п. ф-лы, 11 ил., 1 табл.
Description
Область техники, к которой относится изобретение
Описывается ДНК-последовательность для генной терапии заболеваний сосудов. Существенными элементами ДНК-последовательности являются активаторная последовательность, промоторный модуль и ген активного вещества. Активаторная последовательность специфически к клетке активируется в гладкомышечных клетках, активированных эндотелиальных клетках, активированных макрофагах или активированных лимфоцитах. Это активирование специфически к клеточному циклу регулируется промоторным модулем. Активное вещество представляет собой ингибитор роста гладкомышечных клеток и/или свертывания. Описанную ДНК-последовательность включают в вирусный или невирусный вектор, дополненный лигандом со сродством к летке-мишени.
Описывается ДНК-последовательность для генной терапии заболеваний сосудов. Существенными элементами ДНК-последовательности являются активаторная последовательность, промоторный модуль и ген активного вещества. Активаторная последовательность специфически к клетке активируется в гладкомышечных клетках, активированных эндотелиальных клетках, активированных макрофагах или активированных лимфоцитах. Это активирование специфически к клеточному циклу регулируется промоторным модулем. Активное вещество представляет собой ингибитор роста гладкомышечных клеток и/или свертывания. Описанную ДНК-последовательность включают в вирусный или невирусный вектор, дополненный лигандом со сродством к летке-мишени.
1). Заболевания сосудов за счет гладкомышечных клеток
Гладкомышечные клетки сосудов преобладающе локализованы в средней оболочке артериальных кровеносных сосудов и участвуют в локальной и системной регуляции кровяного давления. В случае неповрежденного, здорового сосуда эти гладкомышечные клетки находятся в состоянии покоя при делении клетки (R. Ross, Nature, 362, 801 (1993)). Повреждения сосудов приводят к перемещению гладкомышечных клеток во внутреннюю оболочку стенки сосуда, где они пролиферируют (образование новой внутренней оболочки) и образуют внеклеточные матричные компоненты.
Гладкомышечные клетки сосудов преобладающе локализованы в средней оболочке артериальных кровеносных сосудов и участвуют в локальной и системной регуляции кровяного давления. В случае неповрежденного, здорового сосуда эти гладкомышечные клетки находятся в состоянии покоя при делении клетки (R. Ross, Nature, 362, 801 (1993)). Повреждения сосудов приводят к перемещению гладкомышечных клеток во внутреннюю оболочку стенки сосуда, где они пролиферируют (образование новой внутренней оболочки) и образуют внеклеточные матричные компоненты.
Относящуюся к внутренней оболочке пролиферацию гладкомышечных клеток рассматривают как существенный компонент при возникновении артериосклероза (J. S. Forrester и др., Am. Coll. Cardiol., 17, 758 (1991)). Далее, эта пролиферация гладкомышечных клеток приводит к повторному стенозу (сужению) сосудов после ангиопластических операций, как также после увеличения размеров полости суженных сосудов (R.S. Schwartz и др., Am. Coll. Cardiol., 20, 1284 (1992), M.W. Liu и др., Circulation, 79, 1374 (1989)).
Как известно, артериосклероз, так же как стеноз и повторный стеноз, сосудов приводит в конечном итоге к тромбозу сосуда и тем самым зачастую к угрожающему жизни инфаркту.
До сих пор, однако, еще нет никакой терапии, которая путем ингибирования роста гладкомышечных клеток предотвращала бы стенозы сосудов. Правда, известно, что гепарин может ингибировать пролиферацию гладкомышечных клеток (Cochran и др., J. Cell. Physiol., 124, 29 (1995)), однако с помощью гепарина нельзя в достаточной мере предотвращать возникновение стенозов. Таким образом, существует потребность в новых способах для предотвращения роста гладкомышечных клеток в поврежденных сосудах и тем самым устранения опасности инфаркта. При этом используют знание о генах и молекулах, которые регулирующе вмешиваются в рост гладкомышечных клеток.
Так, известно, что протоонкоген c-myb, так же как cdc2-киназа и "ядерный антиген пролиферируюших клеток" (PCNA) принимает участие в пролиферации гладкомышечных клеток. Путем введения антисмысловых к c-myb олигонуклеотидов (Simons и др., Nature, 359, 67 (1992)), так же, как антисмысловых к сdс2-киназе олигонуклеотидов в комбинации с антисмысловыми к PCNA олигонуклеотидами (Morishita и др., Proc. Natl. Acad. Sci., 90, 8474 (1993)) непосредственно после и локально в место повреждения сосуда можно предотвращать пролиферацию гладкомышечных клеток у крысы.
Подобных результатов достигают в случае крысы и свиньи путем введения известного в качестве онкогенного супрессора гена ретинобластомы (Rb), также и здесь непосредственно после и локально в место повреждения сосуда. Для того чтобы предотвратить инактивирование вырабатываемого геном ретинобластомы продукта путем фосфорилирования применяют точечно мутированный Rb-ген (обмен Thr-246, Ser-601, Ser-605, Ser-780, Ser-786, Ser-787, Ser-800 на Ala; Thr-350 на Аrg и Ser-804 на Glu (Hamel и др., Mol. Cell. Biol., 12, 3431 (1992)), который кодирует нефосфорилируемую конститутивно активную форму Rb-протеина. Этот мутированный Rb-ген встраивают в рекомбинантный аденовирус с неполной репликацией и этот вектор вводят локально (Chang и др., Science, 267, 518 (1995)).
В другом эксперименте применяют рекомбинантный аденовирус с неполной репликацией, в котором ген вводят в тимидинкиназу вируса простого герпеса (AV-HS-TK). Продукт, вырабатываемый геном киназы, способен фосфорилировать предшественник активного вещества ("пропрепарат") ганцикловир и таким образом превращать в неуклеозидный аналог, который ингибирует синтез ДНК.
Генный вектор AV-HS-TK вводят спустя 7 дней после повреждения сосуда, однако здесь также локально в место повреждения, и затем ежедневно в течение 14 дней интраперитонеально инъецируют ганцикловир. Путем этой обработки в случае крысы достигают отчетливого ингибирования роста гладкомышечных клеток (Guzman и др. , Proc. Natl. Acad. Sci., 91, 10732 (1994)). Путем такого же рода обработки достигают подобных результатов также в случае свиньи (Ohno и др. , Science, 265, 781 (1994)). Однако здесь вектор вводят непосредственно после повреждения сосуда, а введение ганцикловира осуществляют ежедневно в течение 6 дней.
В целом эти опыты показывают возможность с помощью генотерапевтических мер, которые представляют собой вмешательство в процессы деления гладкомышечных клеток, предотвращать стеноз после повреждения сосудов.
Недостатком известных из литературы способов, однако, является то, что эффективные вещества (векторы) нужно вводить локально в место повреждений сосудов, причем, возможно, даже время от времени нужно подвергать обтурации соответствующий участок сосуда, чтобы предотвращать "смывание" векторов. Такого рода инвазивные вмешательства проводят, правда, обычным образом в рамках увеличения размеров полости суженных сосудов, однако они требуют значительных расходов и со своей стороны представляют собой значительную угрозу для пациента вследствие опасности тромбозов и эмболии.
Между прочим, несмотря на локальное введение векторов, нет гарантии в том, что они трансдуцируют только пролиферирующие гладкомышечные клетки. Трансдукция находящихся вблизи или удаленных клеток может приводить к побочным воздействиям (в том числе к трансформации клеток и индукциям опухолей), как это в настоящее время обсуждается широким кругом специалистов (Friedmann, Science, 244, 1275 (1989); Plummer, Scrip Magazine, III/29 (1995)).
В качестве альтернативы введению вышеописанных векторов системное (например, внутривенное или оральное) введение цитостатических препаратов для ингибирования пролиферации гладкомышечных клеток в место заболевания сосуда оказывает только незначительное и временное действие, с другой стороны, вызывает опасность повреждения эндотелия и приводит к значительным острым, а также хроническим побочным воздействиям.
2). Тромбозы
Тромбозы представляют собой все еще трудно излечиваемое, отчасти угрожающее жизни осложнение метаболических заболеваний, как артериосклероз, заболевания артериальных и венозных сосудов и локальные, так же как системные иммуно-реактивные синдромы (см. обзоры Philipps и др. , Blood, 71, 831 (1988); Harker, Biomed. Progr., 8, 17 (1995)).
Тромбозы представляют собой все еще трудно излечиваемое, отчасти угрожающее жизни осложнение метаболических заболеваний, как артериосклероз, заболевания артериальных и венозных сосудов и локальные, так же как системные иммуно-реактивные синдромы (см. обзоры Philipps и др. , Blood, 71, 831 (1988); Harker, Biomed. Progr., 8, 17 (1995)).
Хотя ряд антикоагулянтов, антитромботических средств, фибринолитических препаратов и ингибиторов агрегации тромбоцитов уже давно находит применение в клинической практике и новые вещества проходят клинические испытания, до сих пор угрожающие жизни осложнения тромбоза невозможно в достаточной мере ни предотвращать, ни сдерживать (White, Scrip Magazine, 4, 6 (1994); Antiplatelet Trialists' Collaboration BMJ, 308, 81 (1994)).
Таким образом, существует потребность в новых лекарственных средствах для предотвращения и терапии тромбозов (BMJ, 305, 567 (1992); Vinazzer, Biomedical Progress, 6, 17 (1993)). Причиной значительной части тромбозов являются активированные или поврежденные эндотелиальные клетки. Они сами или стимулированные для пролиферации за счет факторов роста в крови гладкомышечные клетки сами по себе или в совокупности с активированными макрофагами, лимфоцитами, тромбоцитами и гранулоцитами вызывают активирование системы свертывания (Nemerson, Blood, 71, 1 (1988)).
Это активирование приводит в конечном счете к образованию фибрина, активированию и агрегации тромбоцитов и к образованию обогащенных фибрином или обогащенных тромбоцитами сужающих или закупоривающих кровеносные сосуды сгустков крови, тромбозов. Такого рода тромбозы в области артериальной сосудистой системы приводят к инфарктам, которые, например, в случае сердца или мозга, являются угрозой для жизни.
Применяемая до сих пор терапия с помощью антитромботических средств (как гепарин или фракции гепарина), антикоагулянтов (как кумарин), ингибиторов агрегации тромбоцитов (как аспирин) и фибринолотических препаратов (как стреп-токиназа, урокиназа или тканевые плазминогенные активаторы (tPA)), правда, оказывает подтвержденное многочисленными клиническими испытаниями профилактическое действие на угрожающие тромбозы и терапевтическое действие на существующие тромбозы, это действие, однако, является недостаточным. Причина этого в значительной мере заключается в том факте, что действие применяемых терапевтических средств не ограничивается местом заболевания, то есть тромбозом, а они действуют системно. Обусловленные тем самым кровотечения, таким образом, ограничивают как увеличение дозы, так и продолжительность применения.
3). Общее описание изобретения
Предметом изобретения является активное вещество (то есть лекарственное средство), которое можно вводить пациенту как локально, так и также системно и которое:
- влияет только на находящиеся в стадии деления гладкомышечные клетки, подавляет пролиферацию гладкомышечных клеток после повреждений сосудов или поражений сосудов и, таким образом, препятствует стенозу или повторному стенозу сосудов;
- ингибирует свертывание крови только в месте возникшего тромбоза, то есть в месте активированных и пролиферирующих эндотелиальных клеток, гладкомышечных клеток внутренней оболочки кровеносных сосудов, макрофагов и/или лимфоцитов; или
- ингибирует как пролиферацию гладкомышечных клеток, так и локально ингибирует там тромбоз.
Предметом изобретения является активное вещество (то есть лекарственное средство), которое можно вводить пациенту как локально, так и также системно и которое:
- влияет только на находящиеся в стадии деления гладкомышечные клетки, подавляет пролиферацию гладкомышечных клеток после повреждений сосудов или поражений сосудов и, таким образом, препятствует стенозу или повторному стенозу сосудов;
- ингибирует свертывание крови только в месте возникшего тромбоза, то есть в месте активированных и пролиферирующих эндотелиальных клеток, гладкомышечных клеток внутренней оболочки кровеносных сосудов, макрофагов и/или лимфоцитов; или
- ингибирует как пролиферацию гладкомышечных клеток, так и локально ингибирует там тромбоз.
Основной составной частью этого активного вещества является конструкция ДНК, которая состоит из следующих элементов:
(ДНК во всем тексте настоящей заявки используют в виде общего понятия как для комплементарной (кДНК), так и для геномной ДНК-последовательности)

Основной элемент этого активного вещества представляет собой регулируемый клеточным циклом промоторный модуль.
(ДНК во всем тексте настоящей заявки используют в виде общего понятия как для комплементарной (кДНК), так и для геномной ДНК-последовательности)
Основной элемент этого активного вещества представляет собой регулируемый клеточным циклом промоторный модуль.
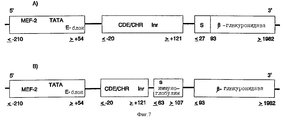
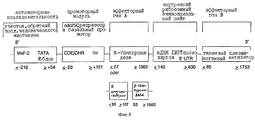
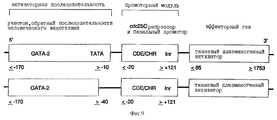
Под регулируемым клеточным циклом промоторным модулем нужно понимать, например, нуклеотидную последовательность CDE-CHR-Inr- (см. ниже). Существенной функцией промоторного модуля является ингибирование функции активаторной последовательности в GO/Gl-фазе клеточного цикла и специфической к клеточному циклу экспрессии в S/G2-фазе и, таким образом, в пролиферирующих клетках.
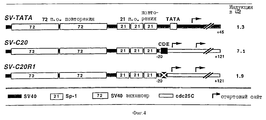
Промоторный модуль CDE-CHR-Inr обнаружен при детальном исследовании G2-специфической экспрессии человеческого промотора cdc25C. Исходным моментом являлось нахождение репрессорного элемента (зависимый от клеточного цикла элемент CDE), который является ответственным за "отключение" промотора в Gl-фазе клеточного цикла (Lucibello и др., EMBO J., 14, 132 (1995)). Путем геномного диметилсульфатного (ДМС) футпринтинга и функционального анализа (фиг. 1, 2) можно показать, что CDE Gl-специфически связывает репрессор ("CDE-связывающий фактор", CDF) и тем самым приводит к ингибированию транскрипции в непролиферирующих (GO) клетках. Локализованный в области базального промотора CDE в своей репримирующей функции зависит от "обратно активирующей (против хода транскрипции) последовательности" (UAS). Это приводит к выводу, что CDE-связывающий фактор подавляет активирующее транскрипцию действие связанных на 5'-конце активаторных протеинов зависимым от клеточного цикла образом, то есть в непролиферирующих клетках, а также в Gl-фазе клеточного цикла (фиг.3).
Этот вывод можно подтвердить следующим экспериментом: слияние вирус-специфического, не регулирующего клеточный цикл раннего энхансера SV40 с минимальным промотором cdc25 (состоящим из CDE и расположенной на 3'-конце стартовой области) приводит к отчетливой регуляции клеточным циклом химерного промотора (фиг.4). Последующие исследования энхансера cdc25C показывают, что в случае регулируемых зависимых от клеточного цикла CDF факторов транскрипции речь идет о NF-Y (CBF) (Dom и др., Cell, 50, 863 (1987); Van Hujisduijnen и др., ЕМВО J., 11, 3119 (1990); Coustry и др., J. Biol. Chem., 270, 468 (1995)), Sp1 (Kadonaga и др., TIBS, 11, 10 (1986)) и, возможно, новом связывающем CBS 7 факторе транскрипции (CIF). Другим представляющим интерес заключением этого исследования является обнаружение того, что NF-Y внутри энхансера cdc25C эффективно активирует транскрипцию только в сочетании по меньшей мере с одним другим комплексом NF-Y или с CIF. Как NF-Y, так и Sp1 относятся к классу обогащенных глутамином активаторов, что является важным указанием на механизм репрессии (например, взаимодействие соответственно интерференция с определенными базальными факторами транскрипции или TAFS).
Сравнение промоторных последовательностей cdc25C, циклина А и cdc2 показывает гомологию в нескольких областях (фиг.5). Во всех трех промоторах (имеющиеся отклонения функционально безотносительны) сохраняется не только CDE, но и также соседние Ус-блоки. Все эти области, как и следовало ожидать, показывают связывание протеина ин виво в случае CDE зависимым от клеточного цикла образом. Кроме того, можно видеть, что все три промотора дерегулируются за счет мутации CDE (таблица 1). Аналогичное подобие наблюдают при сравнении cdc25C, циклина А и cdc2.
Последовательности также отчетливы (фиг.5) в области непосредственно у 3'-конца CDE ("область, гомологичная гену клеточного цикла"; CHR). Эта область функционально так же важна, как и CDE (таблица 1), однако не выявляется в экспериментах ин виво по диметилсульфатному футпринтингу. Возможным объяснением этого является взаимодействие фактора с маленьким участком ДНК. Результаты экспериментов по "анализу на резкое изменение электрофоретической подвижности" (EMSA) указывают на то, что CDE и CHR вместе связываются в протеиновый комплекс, CDF. Эти наблюдения указывают на то, что репрессия через посредство CDF обогащенных глутамином активаторов протекает по зачастую встречающемуся механизму регулируемой клеточным циклом транскрипции.
Для регуляции промотора cdc25C имеет значение, однако, не только CDE-CHR-область, но и также один из сайтов инициации (положение + 1) внутри нуклеотидной последовательности базального промотора (положения ≤ -20 до ≥ +30, см. фиг. 1). Мутации в этой области, которая включает ин витро сайт связывания фактора транскрипции YY-1 (Seto и Shenk, Nature, 354, 241 (1991); Usheva и Shenk, Cell, 76, 1115 (1994)), приводят к полной дерегуляции. Принимая во внимание близость CDE-CHR к базальному промотору, таким образом, очень вероятно взаимодействие CDF с базальным комплексом транскрипции.
В качестве активаторной последовательности (обратно активирующая (против хода транскрипции) последовательность = UAS) нужно понимать нуклеотидную последовательность (промоторную или энхансерную последовательность), с которой взаимодействуют образовавшиеся или активные в клетке-мишени факторы транскрипции. В качестве активаторной последовательности можно использовать CMV-энхансер, CMV-промотор (европейский патент 073177 В1), промотор SV40 или любую другую известную специалисту промоторную или энхансерную последовательность. Согласно настоящему изобретению, однако, к предпочтительным активаторным последовательностям причисляют такие ген-регуляторные последовательности, соответственно элементы генов, которые особенно кодируют образовавшиеся в гладкомышечных клетках, активированных эндотелиальных клетках или в активированных макрофагах или лимфоцитах протеины.
Под активным веществом нужно понимать ДНК-последовательность протеина, который может вызывать в месте образования терапевтический эффект, то есть ингибирование пролиферации гладкомышечных клеток, свертывания или (в случае двух активных веществ) ингибирование пролиферации и одновременно свертывания. При выборе нуклеотидной последовательности для активаторной последовательности и активного вещества руководствуются клеткой-мишенью и желательным активным веществом.
Предлагаемую согласно изобретению конструкцию ДНК известным специалисту образом дополняют до вектора; так, например, вводят в вирусный вектор (см. D. Jolly, Cancer Gene Therapy, 1, 51 (1994)), или, однако, дополняют до плазмиды. Вирусные векторы или плазмиды можно комплексовать с коллоидными дисперсиями, так, например, с липосомами (Farhood и др., Annal. of the New York Academy of Sciences, 716, 23 (1994)) или, однако, с конъюгатами полилизин-лиганд (Curiel и др., Annals of the New York Academy of Sciences, 716, 36 (1994)). Точно также можно получать лекарственные средства с помощью обычных вспомогательных для получения лекарственных средств веществ.
Такого рода вирусные или невирусные векторы можно дополнять лигандом, который обладает сродством к связыванию мембранной структуры с выбранной клеткой-мишенью. При выборе лиганда, таким образом, руководствуются выбором клетки-мишени (см. п.4.4-и последующие и п.5.4 и последующие). Предлагаемое согласно изобретению активное вещество подробнее поясняется, руководствуясь следующими примерами.
4). Активное вещество для ингибирования пролиферации гладкомышечных клеток:
4.1. Выбор активаторной последовательности гладкомышечных клеток
В качестве активаторных последовательностей согласно настоящему изобретению предпочтительно нужно выбирать ген-регуляторные последовательности, соответственно элементы генов, которые кодируют особенно в гладкомышечных клетках образовавшиеся протеины. Этими генами, например, являются следующие:
- тропомиозин (Tsukahara и др., Nucleic Acid Res., 22, 2318 (1994); Novy и др., Cell Motility and Cytosceleton, 25, 267 (1993); Wilton и др., Cytogenetics and Cell Genetics, 68, 122 (1995));
- α-актин (Sartorelli и др., Gens and Developm., 4, 1811 (1990); Miwa и др., Nucleic Acids Res, 18, 4263 (1990));
- α-миозин (Kelly и др., Can. J. Physiol. and Pharm., 72, 1351 (1994); Moussavi и др., Mol. Cell. Biochem., 128, 219 (1993));
- рецепторы факторов роста, как, например, фактор роста, полученный из тромбоцитов (PDGF), FGF (Rubin и др., Int. Congress Ser., 925, 131 (1990); Ross, Ann. Rev. Med., 38, 71 (1987);
- рецепторы ацетилхолина (Dutton и др., PNAS USA, 90, 2040 (1993); Durr и др., Eur. J. Biochem., 224, 353 (1994));
- фосфофруктокиназа-А (Gekakis и др., Biochemistry, 33, 1771, (1994); Tsujino и др., J. Biol. Chem., 264, 15334 (1989); Castella-Escola и др., Gene, 91, 225 (1990));
- фосфоглицератмутаза (Nakatsuji и др. , Mol. Cell. Biol., 12, 4384 (1992));
- тропонин С (Lin и др., Mol. Cell. Biol., 11, 267 (1991));
- десмин (Li и др. , J. Biol. Chem., 266, 6562 (1991); Neuromuscular Disorders, 3, 423 (1993));
- миогенин (Funk и др., PNAS USA, 89, 9484 (1992); Olson, Symp. Soc. Exp. Biol. , 46, 331 (1992); Zhon и др., Mol. Cell. Biol., 14, 6232 (1994); Atchley и др., PNAS USA, 9JL, 11522 (1994));
- рецепторы эндотелина A (Hosoda и др., J. Biol. Chem., 26 7, 18797 (1992); Orelly и др., J. Cardiovasc, Pharm., 22, 18 (1993); Hayzer и др., Am. J. Med. Sci. 304, 231 (1992); Haendler и др., J. Cardiovasc. Pharm., 20, 1 (1992));
- VEGF; VEGF образуется гладкомышечными клетками, особенно в гипоксических условиях (Berse и др., Mol. Biol. Cell, 3, 211 (1992); Finkenzeller и др. , ВВРС, 208, 432 (1995); Tischer и др., ВВРС, 165, 1198 (1989); Leung и др., Science, 246, 1306 (1989); Ferrara и др., Endoc. Fev., 13, 18 (1992)).
4.1. Выбор активаторной последовательности гладкомышечных клеток
В качестве активаторных последовательностей согласно настоящему изобретению предпочтительно нужно выбирать ген-регуляторные последовательности, соответственно элементы генов, которые кодируют особенно в гладкомышечных клетках образовавшиеся протеины. Этими генами, например, являются следующие:
- тропомиозин (Tsukahara и др., Nucleic Acid Res., 22, 2318 (1994); Novy и др., Cell Motility and Cytosceleton, 25, 267 (1993); Wilton и др., Cytogenetics and Cell Genetics, 68, 122 (1995));
- α-актин (Sartorelli и др., Gens and Developm., 4, 1811 (1990); Miwa и др., Nucleic Acids Res, 18, 4263 (1990));
- α-миозин (Kelly и др., Can. J. Physiol. and Pharm., 72, 1351 (1994); Moussavi и др., Mol. Cell. Biochem., 128, 219 (1993));
- рецепторы факторов роста, как, например, фактор роста, полученный из тромбоцитов (PDGF), FGF (Rubin и др., Int. Congress Ser., 925, 131 (1990); Ross, Ann. Rev. Med., 38, 71 (1987);
- рецепторы ацетилхолина (Dutton и др., PNAS USA, 90, 2040 (1993); Durr и др., Eur. J. Biochem., 224, 353 (1994));
- фосфофруктокиназа-А (Gekakis и др., Biochemistry, 33, 1771, (1994); Tsujino и др., J. Biol. Chem., 264, 15334 (1989); Castella-Escola и др., Gene, 91, 225 (1990));
- фосфоглицератмутаза (Nakatsuji и др. , Mol. Cell. Biol., 12, 4384 (1992));
- тропонин С (Lin и др., Mol. Cell. Biol., 11, 267 (1991));
- десмин (Li и др. , J. Biol. Chem., 266, 6562 (1991); Neuromuscular Disorders, 3, 423 (1993));
- миогенин (Funk и др., PNAS USA, 89, 9484 (1992); Olson, Symp. Soc. Exp. Biol. , 46, 331 (1992); Zhon и др., Mol. Cell. Biol., 14, 6232 (1994); Atchley и др., PNAS USA, 9JL, 11522 (1994));
- рецепторы эндотелина A (Hosoda и др., J. Biol. Chem., 26 7, 18797 (1992); Orelly и др., J. Cardiovasc, Pharm., 22, 18 (1993); Hayzer и др., Am. J. Med. Sci. 304, 231 (1992); Haendler и др., J. Cardiovasc. Pharm., 20, 1 (1992));
- VEGF; VEGF образуется гладкомышечными клетками, особенно в гипоксических условиях (Berse и др., Mol. Biol. Cell, 3, 211 (1992); Finkenzeller и др. , ВВРС, 208, 432 (1995); Tischer и др., ВВРС, 165, 1198 (1989); Leung и др., Science, 246, 1306 (1989); Ferrara и др., Endoc. Fev., 13, 18 (1992)).
Промоторные последовательности генов для этих протеинов указываются в следующих работах:
- тропомиозин (Gooding и др., Embo. J., 13, 3861 (1994));
- α-актин (Shimizu и др., J. Biol. Chem., 270, 7631 (1995); Sartorelli и др., Genes Dev., 4, 1811 (1999));
- α-миозин (Kurabayashi и др., J. Biol. Chem., 265, 19271 (1990); Molkentin и др., Mol. Cell. Biol., 14, 4947 (1994));
- рецептор PDGF (Pistritto и др., Antibiot. Chemother., 46, 73 (1994));
- рецептор FGF (Myers и др., J. Biol. Chem., 270, 8257 (1995); Johnson и др. , Adv. Cencer Res., 60, 1 (1993); Chellaiah и др., J. Biol. Chem., 269, 11620 (1994); Yu и др., Hum. Mol. Genetics, 3, 212 (1994); Wang и др., ВВРС, 203, 1781 (1994); Murgue и др., Cancer Res., 54, 5206 (1994); Avraham и др., Genomics, 21, 656 (1994); Burgess и др. , Ann. Rev. Biochem., 58, 575 (1989));
- MRF-4 (Naidu и др., Mol. Cell. Biol., 15, 2707 (1995));
- фосфофруктокиназа A (Gekakis и др., Biochem., 33, 1771 (1994));
- фосфоглицератмутаза (Makatsuji и др. , Mol. Cell. Biol., 12, 4384 (1992)) ;
- тропонин С (Ip и др., Mol. Cell. Biol., 14, 7517 (1994); Parmacek и др., Mol. Cell. Biol., 12, 1967 (1992));
- миогенин (Salminen и др., J. Cell. Biol., 115, 905 (1991); Durr и др., Eur. J. Biochem. , 224, 353 (1994); Edmondson и др., Mol. Cell. Biol., 12, 3665 (1992));
- рецепторы эндотелина A (Hosoda и др., J. Biol. Chem., 267, 18797 (1992); Li и Paulia, J. Biol. Chem., 266, 6562 (1991));
- десмин (Li и др., Neuromisc. Dis., 3, 423 (1993); Li и Capetanaki, Nucl. Acids Res., 21, 335 (1993));
- VEGF.
- тропомиозин (Gooding и др., Embo. J., 13, 3861 (1994));
- α-актин (Shimizu и др., J. Biol. Chem., 270, 7631 (1995); Sartorelli и др., Genes Dev., 4, 1811 (1999));
- α-миозин (Kurabayashi и др., J. Biol. Chem., 265, 19271 (1990); Molkentin и др., Mol. Cell. Biol., 14, 4947 (1994));
- рецептор PDGF (Pistritto и др., Antibiot. Chemother., 46, 73 (1994));
- рецептор FGF (Myers и др., J. Biol. Chem., 270, 8257 (1995); Johnson и др. , Adv. Cencer Res., 60, 1 (1993); Chellaiah и др., J. Biol. Chem., 269, 11620 (1994); Yu и др., Hum. Mol. Genetics, 3, 212 (1994); Wang и др., ВВРС, 203, 1781 (1994); Murgue и др., Cancer Res., 54, 5206 (1994); Avraham и др., Genomics, 21, 656 (1994); Burgess и др. , Ann. Rev. Biochem., 58, 575 (1989));
- MRF-4 (Naidu и др., Mol. Cell. Biol., 15, 2707 (1995));
- фосфофруктокиназа A (Gekakis и др., Biochem., 33, 1771 (1994));
- фосфоглицератмутаза (Makatsuji и др. , Mol. Cell. Biol., 12, 4384 (1992)) ;
- тропонин С (Ip и др., Mol. Cell. Biol., 14, 7517 (1994); Parmacek и др., Mol. Cell. Biol., 12, 1967 (1992));
- миогенин (Salminen и др., J. Cell. Biol., 115, 905 (1991); Durr и др., Eur. J. Biochem. , 224, 353 (1994); Edmondson и др., Mol. Cell. Biol., 12, 3665 (1992));
- рецепторы эндотелина A (Hosoda и др., J. Biol. Chem., 267, 18797 (1992); Li и Paulia, J. Biol. Chem., 266, 6562 (1991));
- десмин (Li и др., Neuromisc. Dis., 3, 423 (1993); Li и Capetanaki, Nucl. Acids Res., 21, 335 (1993));
- VEGF.
Ген-регуляторными последовательностями VEGF-гена являются следующие:
- промоторная последовательность VEGF-гена (5'-фланкирующая область) (Michenko и др. , Cell. Mol. Biol. Res., 40, 35 (1994); Tischer и др., J. Biol. Chem., 266, 11947 (1991)); или
- энхансерная последовательность VEGF-гена (3'-фланкирующая область) (Michenko и др., Cell. Mol. Biol. Res., 40, 35 (1994)); или
- c-Src-ген (Mukhopadhyay и др., Nature, 375, 577 (1995) 7 Bonham и др., Oncogene, 8, 1973 (1993); Parker и др., Mol. Cell. Biol., 5, 831 (1985); Anderson и др., Mol. Cell. Biol., 5., 1122 (1985)); или
- V-Src-ген (Mukhodpadhyay и др., Nature, 375, 577 (1995); Anderson и др. , Mol. Cell. Biol. , 5, 1122 (1985); Gibbs и др., J. Virol., 53, 19 (1985));
- "искусственные" промоторы; факторы семейства спираль-петля-спираль (HLH) (MyoD, Myf-5, миогенин, MRF4 (обзор Olson и Klein, Genes Dev., 8, 1 (1994)) описываются в качестве специфических к мышцам активаторов транскрипции. Далее, к специфическим к мышцам факторам транскрипции относятся протеин "цинковый палец" GATA-4 (Arceci и др., Mol. Cell. Biol., 13, 2235 (1993); Ip и др., Mol. Cell. Biol., 14, 7517 (1994)), а также группы факторов транскрипции MEF-2 (Yu и др., Gene Dev., 6, 1783 (1992)).
- промоторная последовательность VEGF-гена (5'-фланкирующая область) (Michenko и др. , Cell. Mol. Biol. Res., 40, 35 (1994); Tischer и др., J. Biol. Chem., 266, 11947 (1991)); или
- энхансерная последовательность VEGF-гена (3'-фланкирующая область) (Michenko и др., Cell. Mol. Biol. Res., 40, 35 (1994)); или
- c-Src-ген (Mukhopadhyay и др., Nature, 375, 577 (1995) 7 Bonham и др., Oncogene, 8, 1973 (1993); Parker и др., Mol. Cell. Biol., 5, 831 (1985); Anderson и др., Mol. Cell. Biol., 5., 1122 (1985)); или
- V-Src-ген (Mukhodpadhyay и др., Nature, 375, 577 (1995); Anderson и др. , Mol. Cell. Biol. , 5, 1122 (1985); Gibbs и др., J. Virol., 53, 19 (1985));
- "искусственные" промоторы; факторы семейства спираль-петля-спираль (HLH) (MyoD, Myf-5, миогенин, MRF4 (обзор Olson и Klein, Genes Dev., 8, 1 (1994)) описываются в качестве специфических к мышцам активаторов транскрипции. Далее, к специфическим к мышцам факторам транскрипции относятся протеин "цинковый палец" GATA-4 (Arceci и др., Mol. Cell. Biol., 13, 2235 (1993); Ip и др., Mol. Cell. Biol., 14, 7517 (1994)), а также группы факторов транскрипции MEF-2 (Yu и др., Gene Dev., 6, 1783 (1992)).
HLH-Протеины, а также GATA-4 проявляют специфическую к мышце транскрипцию не только с промоторами специфических к мышце генов, но и также в гетерологичном окружении, а также с искусственными промоторами. Такого рода искусственными промоторами, например, являются следующие:
- множественные копии сайтов связывания (ДНК) специфических к мышце HLH-протеинов, как Е-блок (MyoD)
(например, 4хAGCAGGTGTTGGGAGGC) (Weintraub и др. , PNAS, 87, 5623 (1990));
- множественные копии сайта связывания ДНК GATA-4 α-миозин-гена вируса синдрома Франклина
(например, ) (Molkentin и др., Mol. Cell. Biol., 14, 4947 (1994)).
) (Molkentin и др., Mol. Cell. Biol., 14, 4947 (1994)).
- множественные копии сайтов связывания (ДНК) специфических к мышце HLH-протеинов, как Е-блок (MyoD)
(например, 4хAGCAGGTGTTGGGAGGC) (Weintraub и др. , PNAS, 87, 5623 (1990));
- множественные копии сайта связывания ДНК GATA-4 α-миозин-гена вируса синдрома Франклина
(например,
4.2. Выбор активного вещества гладкомышечных клеток
В качестве активного вещества согласно изобретению нужно понимать ДНК-последовательность, экспрессированный протеин которой ингибирует пролиферацию гладкомышечных клеток. К этим ингибиторам клеточного цикла относятся, например, ДНК-последовательности следующих протеинов:
а) Ингибирующие протеины:
- протеин ретинобластомы (pRb=pllO) или родственные р107 и р130 протеины (La Thangue, Curr. Opin. Cell Biol., 6, 443 (1994));
- р53-протеин (Prives и др., Genes Dev., 1, 529 (1993));
- p21 (WAF-1)-протеин (El-Deiry и др., Cell, 75, 817 (1993));
- р16-протеин (Serrano и др. , Nature, 366, 704 (1993); Kamb и др., Science, 264, 436 (1994); Nobori и др., Nature, 368, 753 (1994));
- другие ингибиторы cdK (обзор Pines, TIBS, 19, 143 (1995));
- протеин GADD45 (Papathanasiou и др. , Mol. Cell. Biol., 11, 1009 (1991); Smith и др., Science, 266, 1376 (1994));
- bak-протеин (Farrow и др., Nature, 374, 731 (1995); Chittenden и др., Nature, 374, 733 (1995); Kiefer и др., Nature, 374, 736 (1995)).
В качестве активного вещества согласно изобретению нужно понимать ДНК-последовательность, экспрессированный протеин которой ингибирует пролиферацию гладкомышечных клеток. К этим ингибиторам клеточного цикла относятся, например, ДНК-последовательности следующих протеинов:
а) Ингибирующие протеины:
- протеин ретинобластомы (pRb=pllO) или родственные р107 и р130 протеины (La Thangue, Curr. Opin. Cell Biol., 6, 443 (1994));
- р53-протеин (Prives и др., Genes Dev., 1, 529 (1993));
- p21 (WAF-1)-протеин (El-Deiry и др., Cell, 75, 817 (1993));
- р16-протеин (Serrano и др. , Nature, 366, 704 (1993); Kamb и др., Science, 264, 436 (1994); Nobori и др., Nature, 368, 753 (1994));
- другие ингибиторы cdK (обзор Pines, TIBS, 19, 143 (1995));
- протеин GADD45 (Papathanasiou и др. , Mol. Cell. Biol., 11, 1009 (1991); Smith и др., Science, 266, 1376 (1994));
- bak-протеин (Farrow и др., Nature, 374, 731 (1995); Chittenden и др., Nature, 374, 733 (1995); Kiefer и др., Nature, 374, 736 (1995)).
Для того чтобы предотвратить быструю внутриклеточную инактивацию этих игнибиторов клеточного цикла, нужно предпочтительно применять такие гены, которые обладают мутациями сайтов инактивации экспримированных протеинов, при этом не ухудшая этим их в отношении их функции.
Протеин ретинобластомы (pRb/pllO) и родственные протеины р107 и р130 инактивируются за счет фосфорилирования. Таким образом, предпочтительно используют pRb/p110-, p107- или р130-кДНК-последовательность, которая мутирована таким образом, что участки фосфорилирования кодированного протеина заменены нефосфорилируемыми аминокислотами.
Согласно Hamel и др. (Mol. Cell. Biol., 12, 3431 (1992)) кДНК-последовательность протеина ретинобластомы (р110) за счет обмена аминокислот в положениях 246, 350, 601, 605, 780, 786, 787, 800 и 804 становится более нефосфорилируемой, однако ее активность связывания с большим Т-антигеном не ухудшается. Например, аминокислоты Thr-246, Ser-601, Ser-605, Ser-780, Ser-786, Ser-787 и Ser-800 заменяются на Ala, аминокислота Thr-350 заменяется на Arg и аминокислота Ser-804 заменяется на Glu.
Аналогичным образом мутируется ДНК-последовательность протеина р107 или протеина р130.
Протеин р53 инактивируется в клетке либо за счет связывания со специальными протеинами, как, например, MDM2, или вследствие олигомеризации р53 через дефосфорилированный С-концевой серин-392 (Schikawa и др., Leukemia и Lymphoma, 11, 21 (1993); Brown, Annals of Oncology, 4, 623 (1993)). Таким образом, предпочтительно используют ДНК-последовательность протеина р53, который укорочен на С-конце на серин-392.
б) Цитостатические или цитотоксические протеины
В качестве активного вещества, далее, нужно понимать ДНК-последовательность, которая экспрессирует (выражает) цитостатический или цитотоксический протеин.
В качестве активного вещества, далее, нужно понимать ДНК-последовательность, которая экспрессирует (выражает) цитостатический или цитотоксический протеин.
К такого рода протеинам относятся, например, следующие:
- перфорин (Lin и др., Immunol. Today, 16, 194 (1995));
- гранзим (Smyth и др., Immunol. Today, 16, 202 (1995));
- фактор некроза опухоли (Porter, Tib Tech., 9, 158 (1991); Sidhu и др., Pharmac. Ther., 57, 79 (1993)); в особенности
-α-фактор некроза опухоли (Beutler и др., Nature, 320, 584 (1986); Kriegler и др., Cell, 53, 45 (1988));
-β-фактор некроза опухоли (Gray и др., Nature, 312, 721 (1984); Li и др. , J. Immunol., 138, 4496 (1987); Aggarwal и др., J. Biol. Chem., 260, 2334 (1985)).
- перфорин (Lin и др., Immunol. Today, 16, 194 (1995));
- гранзим (Smyth и др., Immunol. Today, 16, 202 (1995));
- фактор некроза опухоли (Porter, Tib Tech., 9, 158 (1991); Sidhu и др., Pharmac. Ther., 57, 79 (1993)); в особенности
-α-фактор некроза опухоли (Beutler и др., Nature, 320, 584 (1986); Kriegler и др., Cell, 53, 45 (1988));
-β-фактор некроза опухоли (Gray и др., Nature, 312, 721 (1984); Li и др. , J. Immunol., 138, 4496 (1987); Aggarwal и др., J. Biol. Chem., 260, 2334 (1985)).
в) Ферменты
В качестве активного вещества, однако, также нужно понимать ДНК-последовательность фермента, который инактивный предшественник цитостатического препарата превращает в цитостатический препарат.
В качестве активного вещества, однако, также нужно понимать ДНК-последовательность фермента, который инактивный предшественник цитостатического препарата превращает в цитостатический препарат.
Такого рода ферменты, которые расщепляют инактивные предшественники веществ (пропрепараты) до активных цитостатических препаратов (лекарственные средства), и возможные пропрепараты и лекарственные средства уже наглядно описаны Deonarain и др., (Br. J. Cancer, 70, 786 (1994)), Mullen (Pharmac. Ther., 63, 199 (1994)) и Harris и др. (Gene Ther., 1, 170 (1994)).
Например, можно применять ДНК-последовательность одного из следующих ферментов:
- тимидинкиназа вируса простого герпеса
(Garapin и др., PNAS USA, 76, 3755 (1979); Vile и др., Cancer Res., 53, 3860 (1993); Wagner и др., PNAS USA, 78, 1441 (1981); Moelten и др., Cancer Res., 46, 5276 (1986); J. Natl. Cancer Inst., 82, 297 (1990));
- тимидинкиназа вируса ветряной оспы
(Huber и др. , PNAS USA, 88, 8039 (1991); Snoeck, Int. J. Antimicrob. Agents, 4, 211 (1994));
- бактериальная нитроредуктаза
(Michael и др., FEMS Microbiol. Letters, 124, 195 (1994); Bryant и др., J. Biol. Chem. , 266, 4126 (1991); Watanabe и др., Nucleic Acids Res., 18, 1059 (1990));
- бактериальная β-глюкуронидаза
(Jefferson и др., PNAS USA, 83, 8447 (1986));
- растительная β-глюкуронидаза из сухих зерновых культур
(Schulz и др., Phytochemistry, 26, 933 (1987));
- человеческая β-глюкуронидаза
(Bosslet и др., Br. J. Cancer, 65, 234 (1992); Oshima и др., PNAS USA, 84, 685 (1987));
- человеческая карбоксипептидаза (СВ), например:
СВ-А тучной клетки
(Reynolds и др., J. Clin. Invest, 89, 273 (1992));
СВ-В поджелудочной железы
(Yamamoto и др., J. Biol. Chem., 267, 2575 (1992);
Catasus и др., J. Biol. Chem., 270, 6651 (1995));
- бактериальная карбоксипептидаза
(Hamilton и др. , J. Bacteriol., 174, 1626 (1992); Osterman и др., J. Protein Chem., 11, 561 (1992));
- бактериальная β-лактамаза
(Rodriges и др. , Cancer Res., 55, 63 (1995); Hussain и др., J. Bacteriol., 164, 223 (1985); Coqe и др., Embo J., 12, 631 (1993));
- бактериальная цитозиндезаминаза
(Mullen и др., PNAS USA, 89, 33 (1992); Austin и др., Mol. Pharmac., 43, 380 (1993); Danielson и др., Mol. Microbiol., 6, 1335 (1992));
- человеческая каталаза, соответственно пероксидаза
(Ezurum и др., Nucl. Acids Res., 21, 1607 (1993));
- фосфатаза, в особенности
человеческая щелочная фосфатаза
(Gum и др., Cancer Res., 50, 1085 (1990));
человеческая кислая фосфатаза простаты
(Sharieff и др. , Am. J. Hum. Gen., 49, 412 (1991); Song и др., Gene, 129, 291 (1993); Tailor и др., Nucl. Acids Res., 18, 4928 (1990));
- кислая фосфатаза типа 5
(Gene, 130, 201 (1993));
- оксидаза, в особенности
человеческая лизилоксидаза
(Kimi и др., J. Biol. Chem., 270, 7176 (1995));
человеческая кислая D-аминооксидаза
(Fukui и др., J. Biol. Chem., 267, 18631 (1992));
- пероксидаза, в особенности
человеческая глутатион-пероксидаза
(Chada и др., Genomics, 6, 268 (1990); Ishida и др., Nucl. Acids Res., 15, 10051 (1987));
человеческая эозинофильная пероксидаза
(Ten и др. , J. Exp. Med., 169, 1757 (1989); Sahamaki и др., J. Biol. Chem., 264, 16828 (1989));
- человеческая пероксидаза щитовидной железы
(Kimura, PNAS USA, 84, 5555 (1987)).
- тимидинкиназа вируса простого герпеса
(Garapin и др., PNAS USA, 76, 3755 (1979); Vile и др., Cancer Res., 53, 3860 (1993); Wagner и др., PNAS USA, 78, 1441 (1981); Moelten и др., Cancer Res., 46, 5276 (1986); J. Natl. Cancer Inst., 82, 297 (1990));
- тимидинкиназа вируса ветряной оспы
(Huber и др. , PNAS USA, 88, 8039 (1991); Snoeck, Int. J. Antimicrob. Agents, 4, 211 (1994));
- бактериальная нитроредуктаза
(Michael и др., FEMS Microbiol. Letters, 124, 195 (1994); Bryant и др., J. Biol. Chem. , 266, 4126 (1991); Watanabe и др., Nucleic Acids Res., 18, 1059 (1990));
- бактериальная β-глюкуронидаза
(Jefferson и др., PNAS USA, 83, 8447 (1986));
- растительная β-глюкуронидаза из сухих зерновых культур
(Schulz и др., Phytochemistry, 26, 933 (1987));
- человеческая β-глюкуронидаза
(Bosslet и др., Br. J. Cancer, 65, 234 (1992); Oshima и др., PNAS USA, 84, 685 (1987));
- человеческая карбоксипептидаза (СВ), например:
СВ-А тучной клетки
(Reynolds и др., J. Clin. Invest, 89, 273 (1992));
СВ-В поджелудочной железы
(Yamamoto и др., J. Biol. Chem., 267, 2575 (1992);
Catasus и др., J. Biol. Chem., 270, 6651 (1995));
- бактериальная карбоксипептидаза
(Hamilton и др. , J. Bacteriol., 174, 1626 (1992); Osterman и др., J. Protein Chem., 11, 561 (1992));
- бактериальная β-лактамаза
(Rodriges и др. , Cancer Res., 55, 63 (1995); Hussain и др., J. Bacteriol., 164, 223 (1985); Coqe и др., Embo J., 12, 631 (1993));
- бактериальная цитозиндезаминаза
(Mullen и др., PNAS USA, 89, 33 (1992); Austin и др., Mol. Pharmac., 43, 380 (1993); Danielson и др., Mol. Microbiol., 6, 1335 (1992));
- человеческая каталаза, соответственно пероксидаза
(Ezurum и др., Nucl. Acids Res., 21, 1607 (1993));
- фосфатаза, в особенности
человеческая щелочная фосфатаза
(Gum и др., Cancer Res., 50, 1085 (1990));
человеческая кислая фосфатаза простаты
(Sharieff и др. , Am. J. Hum. Gen., 49, 412 (1991); Song и др., Gene, 129, 291 (1993); Tailor и др., Nucl. Acids Res., 18, 4928 (1990));
- кислая фосфатаза типа 5
(Gene, 130, 201 (1993));
- оксидаза, в особенности
человеческая лизилоксидаза
(Kimi и др., J. Biol. Chem., 270, 7176 (1995));
человеческая кислая D-аминооксидаза
(Fukui и др., J. Biol. Chem., 267, 18631 (1992));
- пероксидаза, в особенности
человеческая глутатион-пероксидаза
(Chada и др., Genomics, 6, 268 (1990); Ishida и др., Nucl. Acids Res., 15, 10051 (1987));
человеческая эозинофильная пероксидаза
(Ten и др. , J. Exp. Med., 169, 1757 (1989); Sahamaki и др., J. Biol. Chem., 264, 16828 (1989));
- человеческая пероксидаза щитовидной железы
(Kimura, PNAS USA, 84, 5555 (1987)).
Для облегчения секреции указанных ферментов, возможно, содержащуюся в ДНК-последовательности гомологичную сигнальную последовательность можно заменять на гетерологичную улучшающую внеклеточное выделение сигнальную последовательность.
Так, например, сигнальную последовательность β-глюкуронидазы (положение в ДНК ≤ 27 до 93; Oshima и др., PNAS, 84, 685 (1987)) можно заменять сигнальной последовательностью человеческого иммуноглобулина (положение в ДНК ≤ 63 до ≥ 107; Riechmann и др., Nature, 332, 323 (1988)).
Далее, предпочтительно нужно выбирать кДНК таких ферментов, которые за счет точечных мутаций в более незначительной степени накапливаются в лизосомах. Такого рода точечные мутации описываются, например, для β-глюкуронидазы (Shipley и др., J. Biol. Chem., 268, 12193 (1993)).
4.3. Комбинация одинаковых или разных активных веществ гладкомышечных клеток
Предметом изобретения, далее, является активное вещество, в котором имеется комбинация ДНК-последовательностей нескольких одинаковых активных веществ (А, А) или разных активных веществ (А, Б). Для экспрессии двух ДНК-последовательностей предпочтительно промежуточно включают кДНК "внутреннего рибосомного аминоацильного сайта" (IRES) в качестве регуляторного элемента. Такого рода IRES описываются Mountford и Smith (TIG, 11, 179 (1995)), Kaufman и др., Nucl. Acids Res., 19, 4485 (1991); Morgan и др., Nucl. Acids Res. , 20, 1293 (1992) и Dirks и др., Gene, 129, 247 (1993); Pelletier и Sonenberg, Nature, 334, 320 (1988); и др., Bio Techn., 12, 694 (1994).
Предметом изобретения, далее, является активное вещество, в котором имеется комбинация ДНК-последовательностей нескольких одинаковых активных веществ (А, А) или разных активных веществ (А, Б). Для экспрессии двух ДНК-последовательностей предпочтительно промежуточно включают кДНК "внутреннего рибосомного аминоацильного сайта" (IRES) в качестве регуляторного элемента. Такого рода IRES описываются Mountford и Smith (TIG, 11, 179 (1995)), Kaufman и др., Nucl. Acids Res., 19, 4485 (1991); Morgan и др., Nucl. Acids Res. , 20, 1293 (1992) и Dirks и др., Gene, 129, 247 (1993); Pelletier и Sonenberg, Nature, 334, 320 (1988); и др., Bio Techn., 12, 694 (1994).
Так, кДНК IRES-последовательности полиовируса (положение ≤ 140 до ≥ 630 5'-UTR (Pelletier и Sonenberg, Nature, 334, 320 (1988)) можно применять для связывания ДНК антитромботического вещества А (по 3'-концу) и ДНК антитромботического вещества Б (по 5'-концу).

Такого рода активное вещество в зависимости от сочетания обладает аддитивным или синергическим действием согласно изобретению.
Вследствие относящегося к внутренней оболочке роста гладкомышечных клеток, так же как за счет их апоптоза или некроза, как результат воздействия ингибитора цикла может активироваться система свертывания и могут возникать тромбозы. Такого рода тромбозы нужно предотвращать путем профилактического введения ингибитора свертывания (аспирин, гепарин или другое антитромботическое средство). Введение ингибитора свертывания осуществляют системно, то есть орально или парентерально.
Однако зачастую побочные действия ингибитора свертывания препятствуют достаточной концентрации в месте относящегося к внутренней оболочке роста гладкомышечных клеток.
Таким образом, профилактика тромбоза благодаря такого рода ингибиторам свертывания является ненадежной (Pukac, Am. J. Pathol., 139, 1501 (1991)).
Следующим предметом изобретения является то, что заявляемое в изобретении активное вещество дополнительно к активному веществу, которое представляет собой ингибитор клеточного цикла, в качестве другого элемента содержит ДНК-последовательность активного вещества, которое представляет собой ингибитор свертывания.

Экспрессия ингибитора свертывания регулируется таким же образом, как и экспрессия ингибитора клеточного цикла, за счет активаторной последовательности и регулируемого клеточным циклом репрессорного модуля. Одновременная экспрессия как ингибитора клеточного цикла, так же как ингибитора свертывания, регулируется предпочтительно элементом гена "внутренним рибосомным аминоацильным сайтом" (IRES).
В качестве ингибитора свертывания можно использовать гены, например, активаторов плазминогена (РА), как тканевый PA (tPA) или урокиназа-подобный PA (uPA), протеин С, антитромбин-III, ингибитор метаболического пути тканевого фактора или гирудин. ДНК-Последовательности этих ингибиторов свертывания описываются ниже:
- активатор тканевого плазминогена (tPA)
(Sasaki и др., Nucl. Acids Res., 16, 5695 (1988); Pennica и др., Nature, 301, 214 (1983); Wei и др., DNA, 4, 76 (1985); Harris и др., Mol. Biol. Med. , 3, 279 (1986));
- плазминогенный активатор урокиназного типа (uРА)
(Miyake и др. , J. Biochem., 104, 643 (1988); Nelles и др., J. Biol. Chem., 262, 5682 (1987));
- гибриды tPA и uPA
(Kalyan и др. , Gene, 68, 205 (1988); Devries и др., Biochem. 27, 2565 (1988));
- протеин С (Foster и др., PNAS, 82, 4673 (1985));
- гирудин
(Maerki и др., Semin. Thromb. Hemostas., 17, 88 (1991); De Taxis du Poet и др., Blood Coag. Fibrin., 2, 113 (1991); Harvey и др., PNAS USA, 83, 1084 (1986); Sachhieri и др., европейский патент 0324712 В1; европейский патент 0142860 В1);
- ингибиторы серин-протеиназы (серпины), как, например, C-lS-ингибитор
(Bock и др., Biochem., 25, 4292 (1986); Davis и др., PNAS USA, 83, 3161 (1986); Que, BBPC, 137, 620 (1986); Rauth и др., Proteine Sequences and Data Analysis, 1, 251 (1988); Carter и др., Eur. J. Biochem., 173, 163 (1988); Tosi и др., Gene, 42, 265 (1986); Carter и др., Eur. J. Biochem., 197, 301 (1991); Eldering и др., J. Biol. Chem., 267, 7013 (1993));
- 1-антитрипсин
(Tosi и др., Gene, 42, 265 (1986); Graham и др., Hum. Genetics, 85, 381 (1990); Hafeez и др. , J. Clin. Invest., 89, 1214 (1992); Тикунова и др., Биоорганическая химия, 17, 1694 (1991); Kay и др., Human Gene Ther., 3, 641 (1992); Lemarchand и др., Mol. Biol., 27, 1014 (1993); Lambach и др., Human Mol. Gen., 7, 1001 (1993));
- антитромбин-III
(Stackhouse и др., J. Biol. Chem., 258, 703 (1983); Olds и др., Bioshem. , 32, 4216 (1993); Laue и др., Nucl. Acids Res., 22, 3556 (1994));
- ингибитор метаболического пути тканевого фактора
(TFPI)
(Enjyoji и др. , Genomics, 17, 423 (1993); Wun и др., J. Biol. Chem., 263, 6001 (1988); Girard и др., Thromb. Res., 55, 37 (1989)).
- активатор тканевого плазминогена (tPA)
(Sasaki и др., Nucl. Acids Res., 16, 5695 (1988); Pennica и др., Nature, 301, 214 (1983); Wei и др., DNA, 4, 76 (1985); Harris и др., Mol. Biol. Med. , 3, 279 (1986));
- плазминогенный активатор урокиназного типа (uРА)
(Miyake и др. , J. Biochem., 104, 643 (1988); Nelles и др., J. Biol. Chem., 262, 5682 (1987));
- гибриды tPA и uPA
(Kalyan и др. , Gene, 68, 205 (1988); Devries и др., Biochem. 27, 2565 (1988));
- протеин С (Foster и др., PNAS, 82, 4673 (1985));
- гирудин
(Maerki и др., Semin. Thromb. Hemostas., 17, 88 (1991); De Taxis du Poet и др., Blood Coag. Fibrin., 2, 113 (1991); Harvey и др., PNAS USA, 83, 1084 (1986); Sachhieri и др., европейский патент 0324712 В1; европейский патент 0142860 В1);
- ингибиторы серин-протеиназы (серпины), как, например, C-lS-ингибитор
(Bock и др., Biochem., 25, 4292 (1986); Davis и др., PNAS USA, 83, 3161 (1986); Que, BBPC, 137, 620 (1986); Rauth и др., Proteine Sequences and Data Analysis, 1, 251 (1988); Carter и др., Eur. J. Biochem., 173, 163 (1988); Tosi и др., Gene, 42, 265 (1986); Carter и др., Eur. J. Biochem., 197, 301 (1991); Eldering и др., J. Biol. Chem., 267, 7013 (1993));
- 1-антитрипсин
(Tosi и др., Gene, 42, 265 (1986); Graham и др., Hum. Genetics, 85, 381 (1990); Hafeez и др. , J. Clin. Invest., 89, 1214 (1992); Тикунова и др., Биоорганическая химия, 17, 1694 (1991); Kay и др., Human Gene Ther., 3, 641 (1992); Lemarchand и др., Mol. Biol., 27, 1014 (1993); Lambach и др., Human Mol. Gen., 7, 1001 (1993));
- антитромбин-III
(Stackhouse и др., J. Biol. Chem., 258, 703 (1983); Olds и др., Bioshem. , 32, 4216 (1993); Laue и др., Nucl. Acids Res., 22, 3556 (1994));
- ингибитор метаболического пути тканевого фактора
(TFPI)
(Enjyoji и др. , Genomics, 17, 423 (1993); Wun и др., J. Biol. Chem., 263, 6001 (1988); Girard и др., Thromb. Res., 55, 37 (1989)).
4.4. Выбор лиганда гладкомышечных клеток
В качестве лиганда в коллоидных дисперсиях, например, полилизин-лиганд-конъюгаты, предпочтительны вещества, которые связываются с поверхностью гладкомышечных клеток. Сюда относятся антитела или фрагменты антител, направленные против мембранных структур гладкомышечных клеток, как, например:
- антитело 10F3
(Printseva и др., Exp. Cell Res., 169, 85 (1987); Am. J. Path., 134, 305 (1989)); или
- антитело против актина
(Desmonliere и др., Comptes Rendus des Seances de la Soc. de Biol. и de ses Filiales, 182, 391 (1988)); или
- антитела против рецепторов ангиотензина-II
(Butcher и др., ВВРА, 196, 1280 (1993)); или
- антитела против рецепторов факторов роста
(обзор Mendelsohn, Prog. All., 45, 147 (1988); Sato и др., J. Nat. Canс. Inst., 81, 1600 (1989); Hynes и др., ВВА, 1198, 165 (1994));
или антитела, направленные, например, против
- EGF-рецепторов
(Fan и др., Cancer Res., 53, 4322 (1993); Bender и др., Cancer Res., 52, 121 (1992); Aboud-Pirak и др., J. Nat. Cancer Inst., 80, 1605 (1988); Sato и др., Mol. Biol. Med., 1, 511 (1983); Kawamoto и др., PNAS, 80, 1337 (1983));
- или против PDGF-рецепторов
(Yu и др. , J. Biol. Chem., 269, 10668 (1994); Kelly и др., J. Biol. Chem. , 266, 8987 (1991); Bowen-Pope и др. , J. Biol. Chem., 257, 5161 (1982));
- или против FGF-рецепторов
(Vanhalteswaran и др., J. Cell. Biol., 115, 418 (1991); Zhan и др., J. Biol. Chem., 269, 20221 (1994));
- или антитела против рецепторов эндотелина-А.
В качестве лиганда в коллоидных дисперсиях, например, полилизин-лиганд-конъюгаты, предпочтительны вещества, которые связываются с поверхностью гладкомышечных клеток. Сюда относятся антитела или фрагменты антител, направленные против мембранных структур гладкомышечных клеток, как, например:
- антитело 10F3
(Printseva и др., Exp. Cell Res., 169, 85 (1987); Am. J. Path., 134, 305 (1989)); или
- антитело против актина
(Desmonliere и др., Comptes Rendus des Seances de la Soc. de Biol. и de ses Filiales, 182, 391 (1988)); или
- антитела против рецепторов ангиотензина-II
(Butcher и др., ВВРА, 196, 1280 (1993)); или
- антитела против рецепторов факторов роста
(обзор Mendelsohn, Prog. All., 45, 147 (1988); Sato и др., J. Nat. Canс. Inst., 81, 1600 (1989); Hynes и др., ВВА, 1198, 165 (1994));
или антитела, направленные, например, против
- EGF-рецепторов
(Fan и др., Cancer Res., 53, 4322 (1993); Bender и др., Cancer Res., 52, 121 (1992); Aboud-Pirak и др., J. Nat. Cancer Inst., 80, 1605 (1988); Sato и др., Mol. Biol. Med., 1, 511 (1983); Kawamoto и др., PNAS, 80, 1337 (1983));
- или против PDGF-рецепторов
(Yu и др. , J. Biol. Chem., 269, 10668 (1994); Kelly и др., J. Biol. Chem. , 266, 8987 (1991); Bowen-Pope и др. , J. Biol. Chem., 257, 5161 (1982));
- или против FGF-рецепторов
(Vanhalteswaran и др., J. Cell. Biol., 115, 418 (1991); Zhan и др., J. Biol. Chem., 269, 20221 (1994));
- или антитела против рецепторов эндотелина-А.
Мышиные моноклональные антитела предпочтительно нужно использовать в гуманизированной форме. Гуманизирование осуществляют способом, описанным Winter и др., Nature, 349, 293 (1991), и Hoogenbooms и др., Rev. Tr. Transfus. Hemobiol., 36, 19 (1993). Фрагменты антител получают согласно уровню техники, например, способом, описанным Winter и др., Nature. 349, 293 (1991); Hoogenboom и др., Rev. Tr. Transfus. Hemobiol., 36, 19 (1993); Girol, Mol. Immunol. , 28, 1379 (1991) или Huston и др., Int. Res. Immunol., 10, 195 (1993).
К лигандам относятся, далее, все активные вещества, которые связываются с мембранными структурами или мембранными рецепторами гладкомышечных клеток (обзор Pusztai и др. , J. Pathol., 169, 191 (1993); Harris, Current Opin. Biotechnol. , 2, 260 (1991)). Например, сюда относятся факторы роста или их фрагменты, соответственно их частичные последовательности, которые связываются с экспрессированными гладкомышечными клетками рецепторами, как, например:
- PDGF
(Westermark и др. , Cancer Res., 51, 5087 (1991); Ponten и др., J. Invest. Dermatol., 102, 304 (1994));
- EGF
(Modjtahedi и др., Int. J. Oncol., 4, 277 (1994);
Carpentar и др., J. Biol. Chem., 265, 7709 (1990);
- TGF-β
(Segarini, BBA, 1155, 269 (1993));
- TGF-α
(Salomon и др., Cancer Cells, 2, 389 (1990));
- FGF
(Burgess и др., Annu. Rev. Biochem., 58, 575 (1989));
- эндотелин-А
(Oreilly и др., J. Cardiovasc. Pharm., 22, 18 (1993)).
- PDGF
(Westermark и др. , Cancer Res., 51, 5087 (1991); Ponten и др., J. Invest. Dermatol., 102, 304 (1994));
- EGF
(Modjtahedi и др., Int. J. Oncol., 4, 277 (1994);
Carpentar и др., J. Biol. Chem., 265, 7709 (1990);
- TGF-β
(Segarini, BBA, 1155, 269 (1993));
- TGF-α
(Salomon и др., Cancer Cells, 2, 389 (1990));
- FGF
(Burgess и др., Annu. Rev. Biochem., 58, 575 (1989));
- эндотелин-А
(Oreilly и др., J. Cardiovasc. Pharm., 22, 18 (1993)).
4.5. Получение биологически активного вещества гладкомышечных клеток
Получение предлагаемого согласно изобретению активного вещества описывается подробнее, руководствуясь следующими примерами:
а). Конструкция химерного промотора миогенин-промотор CDE-CHR-Inr
Человеческий миогениновый промотор (положение ≤ -210 до ≥ +54 ДНК-последовательности, опубликованной Salmin и др. , J. Cell. Biol., 115, 905 (1991)) своим 3'-концом связывается с 5'-концом CDE-CHR-Inr-модуля человеческого cdc25C-гeнa (положение ≤ -20 до ≥ +121 последовательности, опубликованной Lucibello и др., EMBO J., 14, 132 (1995)) (см. фиг.6). Связывание осуществляется с помощью известных специалисту и имеющихся в продаже ферментов. Кроме того, применяют различные фрагменты миогениновой промоторной последовательности (см. фиг. 6). Так, применяют содержащую ТАТА-блок ДНК-последовательность миогенинового промотора. Однако равным образом можно применять также промоторную последовательность в положении ≤ -210 до ≥ -40 (см. фиг.6).
Получение предлагаемого согласно изобретению активного вещества описывается подробнее, руководствуясь следующими примерами:
а). Конструкция химерного промотора миогенин-промотор CDE-CHR-Inr
Человеческий миогениновый промотор (положение ≤ -210 до ≥ +54 ДНК-последовательности, опубликованной Salmin и др. , J. Cell. Biol., 115, 905 (1991)) своим 3'-концом связывается с 5'-концом CDE-CHR-Inr-модуля человеческого cdc25C-гeнa (положение ≤ -20 до ≥ +121 последовательности, опубликованной Lucibello и др., EMBO J., 14, 132 (1995)) (см. фиг.6). Связывание осуществляется с помощью известных специалисту и имеющихся в продаже ферментов. Кроме того, применяют различные фрагменты миогениновой промоторной последовательности (см. фиг. 6). Так, применяют содержащую ТАТА-блок ДНК-последовательность миогенинового промотора. Однако равным образом можно применять также промоторную последовательность в положении ≤ -210 до ≥ -40 (см. фиг.6).
б). Конструкция плазмиды, содержащей основную (центральную) составную часть активного вещества
Таким образом полученную химерную контрольную единицу транскрипции промоторного модуля миогенина ее 3'-концом связывают с 5'-концом ДНК, которая содержит полную кодирующую область человеческой β-глюкуронидазы (положение в ДНК ≤ 27 - ≥ 1982 последовательности, опубликованной Oshima и др., PNAS USA, 84, 684 (1987)). (см. фиг.6).
Таким образом полученную химерную контрольную единицу транскрипции промоторного модуля миогенина ее 3'-концом связывают с 5'-концом ДНК, которая содержит полную кодирующую область человеческой β-глюкуронидазы (положение в ДНК ≤ 27 - ≥ 1982 последовательности, опубликованной Oshima и др., PNAS USA, 84, 684 (1987)). (см. фиг.6).
Эта ДНК также содержит необходимую для секреции сигнальную последовательность (22 N-концевые аминокислоты). Для облегчения клеточного выделения эту сигнальную последовательность предпочтительно обменивают на сигнальную последовательность иммуноглобулина (положение ≤ 63 до ≥ 107; Riechmann и др. , Nature, 332, 323 (1988)) (фиг.7). Таким образом полученные контрольные единицы транскрипции и ДНК человеческой β-глюкуронидазы клонируют в pUC18/19 или производимых от Bluescript плазмидных векторов, которые можно использовать непосредственно или в коллоидных дисперсионных системах для введения ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы и вводить путем инъекции.
в). Конструкция плазмиды, содержащей гены β-глюкуронидазы, как также тканевый плазминогенный активатор
Полученный как в п.2) элемент β-глюкуронидазы миогенинового репрессорного модуля его 3'-концом связывают с 5'-концом кДНК "внутреннего рибосомного аминоациального сайта" полиовируса (положение ≤ 140 до ≥ 630 5'-UTR-элемента Pelletier и Sonenberg, Nature, 334, 320 (1988)). С его 3'-концом снова связывают 5'-конец ДНК тканевого плазминогенного активатора (положение ≤ 85 до ≥ 1753; Pennica и др., Nature, 301, 214 (1983)) (фиг.7). Затем всю конструкцию клонируют в pUC17/19 или производимых от Bluescript плазмидных векторах, которые можно использовать непосредственно или в коллоидальных дисперсионных системах для переноса ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы (фиг. 8).
Полученный как в п.2) элемент β-глюкуронидазы миогенинового репрессорного модуля его 3'-концом связывают с 5'-концом кДНК "внутреннего рибосомного аминоациального сайта" полиовируса (положение ≤ 140 до ≥ 630 5'-UTR-элемента Pelletier и Sonenberg, Nature, 334, 320 (1988)). С его 3'-концом снова связывают 5'-конец ДНК тканевого плазминогенного активатора (положение ≤ 85 до ≥ 1753; Pennica и др., Nature, 301, 214 (1983)) (фиг.7). Затем всю конструкцию клонируют в pUC17/19 или производимых от Bluescript плазмидных векторах, которые можно использовать непосредственно или в коллоидальных дисперсионных системах для переноса ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы (фиг. 8).
5). Активное вещество для ингибирования свертывания
5.1. Выбор активаторной последовательности для игнибирования или свертывания
В качестве активаторных последовательностей согласно изобретению предпочтительно нужно применять ген-регуляторные последовательности, соответственно элементы из генов, которые кодируют обнаруживаемые в гладкомышечных клетках, в активированных эндотелиальных клетках, в активированных макрофагах или в активированных лимфоцитах протеины.
5.1. Выбор активаторной последовательности для игнибирования или свертывания
В качестве активаторных последовательностей согласно изобретению предпочтительно нужно применять ген-регуляторные последовательности, соответственно элементы из генов, которые кодируют обнаруживаемые в гладкомышечных клетках, в активированных эндотелиальных клетках, в активированных макрофагах или в активированных лимфоцитах протеины.
а) Гладкомышечные клетки
Примеры активаторных последовательностей генов в гладкомышечных клетках уже указаны в п.4.1.
Примеры активаторных последовательностей генов в гладкомышечных клетках уже указаны в п.4.1.
б). Активированные эндотелиальные клетки
Примеры протеинов, которые особенно образуются в активированных эндотелиальных клетках, описываются Burrows и др., (Pharmac. Therp., 64, 155 (1994)) и Plate и сотр., (Brain Pathol., 4, 207 (1994)). В особенности к этим интенсивно образующимся в эндотелиальных клетках протеинам относятся, например, следующие:
- специфический к мозгу эндотелиальный переносчик глюкозы-I; эндотелиальные клетки мозга отличаются очень сильной экспрессией этого переносчика, чтобы осуществить трансэндотелиальную транспортировку D-глюкозы в мозг (Gerhart и др., J. Neurosci. Res., 22, 464 (1989)). Промоторная последовательность описывается Murakami и др., J. Biol. Chem., 267, 9300 (1992);
- эндоглин; эндоглин, по-видимому, является не переносящим сигнала рецептором TGF-β (Gougos и др., J. Biol. Chem., 265, 8361 (1990); Cheifetz, 1. Biol. Chem., 267, 19027 (1992); Moren и др., ВВРС, 189, 356 (1992)). В незначительных количествах он встречается в нормальном эндотелии, однако усиленно экспримируется пролиферирующим эндотелием (Westphal и др., J. Invest. Derm. , 100, 27 (1993); Burrows и др., Pharmac. Ther., 64, 155 (1994)). Промоторная последовательность описывается Bellon и др. (Eur. J. Immunol., 23, 2340 (1993)) и Ge и др. (Gene, 138, 201 (1994)).
Примеры протеинов, которые особенно образуются в активированных эндотелиальных клетках, описываются Burrows и др., (Pharmac. Therp., 64, 155 (1994)) и Plate и сотр., (Brain Pathol., 4, 207 (1994)). В особенности к этим интенсивно образующимся в эндотелиальных клетках протеинам относятся, например, следующие:
- специфический к мозгу эндотелиальный переносчик глюкозы-I; эндотелиальные клетки мозга отличаются очень сильной экспрессией этого переносчика, чтобы осуществить трансэндотелиальную транспортировку D-глюкозы в мозг (Gerhart и др., J. Neurosci. Res., 22, 464 (1989)). Промоторная последовательность описывается Murakami и др., J. Biol. Chem., 267, 9300 (1992);
- эндоглин; эндоглин, по-видимому, является не переносящим сигнала рецептором TGF-β (Gougos и др., J. Biol. Chem., 265, 8361 (1990); Cheifetz, 1. Biol. Chem., 267, 19027 (1992); Moren и др., ВВРС, 189, 356 (1992)). В незначительных количествах он встречается в нормальном эндотелии, однако усиленно экспримируется пролиферирующим эндотелием (Westphal и др., J. Invest. Derm. , 100, 27 (1993); Burrows и др., Pharmac. Ther., 64, 155 (1994)). Промоторная последовательность описывается Bellon и др. (Eur. J. Immunol., 23, 2340 (1993)) и Ge и др. (Gene, 138, 201 (1994)).
- VEGF-рецепторы:
Различают два рецептора (Plate и др., Int. J. Cancer, 59, 520 (1994)):
- VEGF-рецептор-1 (flt-1)
(Vries и др., Science, 255, 989 (1992))
(в цитостатической части содержит fms-подобную тирозинкиназу); и
- VEGF-рецептор-2 (flk-1, KDR)
(Terman и др., ВВРС, 187, 1579 (1992))
(содержит в цитоплазмической части тирозинкиназу).
Различают два рецептора (Plate и др., Int. J. Cancer, 59, 520 (1994)):
- VEGF-рецептор-1 (flt-1)
(Vries и др., Science, 255, 989 (1992))
(в цитостатической части содержит fms-подобную тирозинкиназу); и
- VEGF-рецептор-2 (flk-1, KDR)
(Terman и др., ВВРС, 187, 1579 (1992))
(содержит в цитоплазмической части тирозинкиназу).
Оба рецептора почти исключительно встречаются в эндотелиальных клетках (Senger и др., Cancer Metast. Rev., 12, 303 (1993));
- другие специфические к эндотелию рецепторные тирозинкиназы:
- til-1 или til-2
(Partanen и др. , Mol. Cell Biol., 12, 1698 (1992); и Risau, Development, 119, 957 (1993); Dumont и др., Oncogene, 7, 1471 (1992));
и Risau, Development, 119, 957 (1993); Dumont и др., Oncogene, 7, 1471 (1992));
- В61-рецептор (Eck-рецептор)
(Bartley и др., Nature, 368, 558 (1994); Pandey и др., Science, 268, 567 (1995); van der Geer и др., Ann. Rev. Cell Biol., 10, 251 (1994));
- B61
Молекула B61 представляет собой лиганд В61-рецептора
(Hoizman и др., J. Am. Soc. Nephrol., 4, 466 (1993); Bartley и др., Nature, 368, 558 (1994));
- эндотелии, в особенности:
- эндотелин В
(Oreilly и др., J. Cardiovasc. Pharm., 22, 18 (1993); Benatti и др., J. Clin. Invest., 91, 1149 (1993); O'Reilly и др., ВВРС, 193, 834 (1993)). Промоторная последовательность описывается Benatti и др., J. Clin. Invest., 91, 1149 (1993);
- эндотелин-1
(Yanasigawa и др. , Nature, 332, 411 (1988)). Промоторная последовательность описывается Wilson и др., Mol. Cell. Biol., 10, 4654 (1990);
- рецепторы эндотелина, в особенности рецептор эндотелина-В
(Webb и др. , Mol. Pharmacol., 47, 730 (1995); Haendler и др., J. Cardiovasc. Pharm., 20, 1 (1992));
- манноза-6-фосфат-рецепторы
(Perales и др., Eur. J. Biochem., 226, 225 (1994)).
- другие специфические к эндотелию рецепторные тирозинкиназы:
- til-1 или til-2
(Partanen и др. , Mol. Cell Biol., 12, 1698 (1992);
- В61-рецептор (Eck-рецептор)
(Bartley и др., Nature, 368, 558 (1994); Pandey и др., Science, 268, 567 (1995); van der Geer и др., Ann. Rev. Cell Biol., 10, 251 (1994));
- B61
Молекула B61 представляет собой лиганд В61-рецептора
(Hoizman и др., J. Am. Soc. Nephrol., 4, 466 (1993); Bartley и др., Nature, 368, 558 (1994));
- эндотелии, в особенности:
- эндотелин В
(Oreilly и др., J. Cardiovasc. Pharm., 22, 18 (1993); Benatti и др., J. Clin. Invest., 91, 1149 (1993); O'Reilly и др., ВВРС, 193, 834 (1993)). Промоторная последовательность описывается Benatti и др., J. Clin. Invest., 91, 1149 (1993);
- эндотелин-1
(Yanasigawa и др. , Nature, 332, 411 (1988)). Промоторная последовательность описывается Wilson и др., Mol. Cell. Biol., 10, 4654 (1990);
- рецепторы эндотелина, в особенности рецептор эндотелина-В
(Webb и др. , Mol. Pharmacol., 47, 730 (1995); Haendler и др., J. Cardiovasc. Pharm., 20, 1 (1992));
- манноза-6-фосфат-рецепторы
(Perales и др., Eur. J. Biochem., 226, 225 (1994)).
Промоторные последовательности описываются Ludwig и др.
(Gene, 142, 311 (1994)), Oshima и др. , (J. Biol. Chem., 263, 2553 (1988)) и Pohlmann и др., (PNAS USA, 84, 5575 (1987));
- фактор Виллебранда;
промоторная последовательность описывается Jahroudi и Lynch (Mol. Cell. Biol. , 14, 999 (1994); Ferreira и др., Biochem. J., 293, 641 (1993)) и Aird и др. (PNAS USA, 92, 4567 (1995));
- интерлейкин-lα, интерлейкин-1β.
- фактор Виллебранда;
промоторная последовательность описывается Jahroudi и Lynch (Mol. Cell. Biol. , 14, 999 (1994); Ferreira и др., Biochem. J., 293, 641 (1993)) и Aird и др. (PNAS USA, 92, 4567 (1995));
- интерлейкин-lα, интерлейкин-1β.
Интерлейкин-1 продуцируется активированными эндотелиальными клетками (Wamer и др., J. Immunol., 139, 1911 (1987)). Промоторные последовательности описываются Hangen и др. , Mol. Carcinog., 2, 68 (1986), Turner и др., J. Iimnunol. , 143, 3556 (1989); Fenton и др., J. Immunol., 138, 3972 (1987); Bensi и др., Cell. Growth. Diff., 1, 491 (1990); Hiscott и др., Mol. Cell. Biol., 13, 6231 (1993) и Mori и др., Blood, 84, 1688 (1994);
- рецептор интерлейкина-1;
промоторная последовательность описывается Ye и др., PNAS USA, 90, 2295 (1993);
- адгезионная молекула сосудистой клетки (VCAM-1). Экспрессия VCAM-1 в эндотелиальных клетках активируется липополисахаридами, α-фактором некроза опухоли (Neish и др., Mol. Cell. Biol., 15, 2558 (1995)); интерлейкином-4 (Iademarko и др., J. Clin. Invest., 95, 264 (1995)) и интерлейкином-1 (Marni и др., J. Clin. Invest., 92, 1866 (1993)).
- рецептор интерлейкина-1;
промоторная последовательность описывается Ye и др., PNAS USA, 90, 2295 (1993);
- адгезионная молекула сосудистой клетки (VCAM-1). Экспрессия VCAM-1 в эндотелиальных клетках активируется липополисахаридами, α-фактором некроза опухоли (Neish и др., Mol. Cell. Biol., 15, 2558 (1995)); интерлейкином-4 (Iademarko и др., J. Clin. Invest., 95, 264 (1995)) и интерлейкином-1 (Marni и др., J. Clin. Invest., 92, 1866 (1993)).
Промоторная последовательность VCAM-1 описывается Neish и др., Mol. Cell. Biol., 15, 2558 (1995); Ahmad и др., J. Biol. Chem., 270, 8976 (1995); Neish и др., J. Exp. Med., 176, 1583 (1992); Iademarco и др., J. Biol. Chem. , 267, 16323 (1992) и Cybulsky и др., PNAS USA, 88, 7859 (1991);
- синтетическая активаторная последовательность. В качестве альтернативы природным специфическим к эндотелию промоторам также можно применять синтетические активаторные последовательности, которые состоят из олигомеризованных сайтов связывания факторов транскрипции, предпочтительно или селективно активных в эндотелиальных клетках. Примером их является фактор транскрипции GATA-2, сайтом связывания которого в эндотелине-1 является ген 5'-ТТАТСТ-3' (Lee и др. , Biol. Chem., 266, 16188 (1991); Dorfmann и др., J. Biol. Chem. , 267, 1279 (1992) и Wilson и др., Mol. Cell. Biol., 10, 4854 (1990)).
- синтетическая активаторная последовательность. В качестве альтернативы природным специфическим к эндотелию промоторам также можно применять синтетические активаторные последовательности, которые состоят из олигомеризованных сайтов связывания факторов транскрипции, предпочтительно или селективно активных в эндотелиальных клетках. Примером их является фактор транскрипции GATA-2, сайтом связывания которого в эндотелине-1 является ген 5'-ТТАТСТ-3' (Lee и др. , Biol. Chem., 266, 16188 (1991); Dorfmann и др., J. Biol. Chem. , 267, 1279 (1992) и Wilson и др., Mol. Cell. Biol., 10, 4854 (1990)).
в). Активированные макрофаги и/или активированные лимфоциты
В качестве активаторной последовательности согласно настоящему изобретению, кроме того, нужно понимать промоторные последовательности генов протеинов, которые интенсивно образуются при иммунной реакции в макрофагах и/или лимфоцитах. Сюда относятся, например:
- интерлейкин-1
(Bensi и др., Gene, 52, 95 (1987); Fibbe и др., Blut., 59, 147 (1989));
- рецептор интерлейкина-1
(Colotta и др., Immunol. Today, 15, 562 (1994); Sims и др., Clin. Immunol. Immunopath., 72, 9 (1994); Ye и др., PNAS USA, 90, 2295 (1993));
- интерлейкин-2
(Jansen и др. , Cell, 39, 207 (1994); Ohbo и др., J. Biol. Chem., 270, 7479 (1995));
- рецептор интерлейкина-2 (Semenzato и др., Int. J. Clin. Lab. Res., 22, 133 (1992));
- γ-интерферон
(Kirchner, DMW, 111, 64 (1986); Lehmann и др., J. Immunol., 153, 165 (1994));
- интерлейкин-4
(Paul, Blood, 77, 1859 (1991); te Velde и др., Blood, 76, 1392 (1990));
- рецептор интерлейкина-4
(Vallenga и др., Leukemia, 7, 1131 (1993); Gelizzi и др., Int. Immunol., 2, 669 (1990));
- интерлейкин-3
(Frendl, Int. J. Immunopharm., 14, 421 (1992));
- интерлейкин-5
(Azuma и др., Nucl. Acid Res., 14, 9149 (1986); Yokota и др., PNAS, 84, 7388 (1987));
- интерлейкин-6
(Brack и др., Int. J. Clin. Lab. Res., 22, 143 (1992);
- LIF
(Metcalf, Int. J. Cell Clon. , 9, 95 (1991); Samal, ВВА, 1260, 27 (1995));
- интерлейкин-7
(Joshi и др., 21, 681 (1991));
- интерлейкин-10
(Benjamin и др., Leuk. Lymph., 12, 205 (1994); Fluchiger и др., J. Exp. Med., 179, 91 (1994));
- интерлейкин-11
(Yang и др., Biofactors, 4, 15 (1992));
- интерлейкин-12
(Kiniwa и др., J. Clin. Invest., 90, 262 (1992), Gatelay, Cancer Invest. , 11, 500 (1993));
- интерлейкин-13
(Punnonen и др. , PNAS, 90, 3730 (1993); Muzio и др., Blood, 83., 1738 (1994));
- GM-CSF
(Metcalf, Cancer, 15, 2185 (1990));
- рецептор GM-CSF
Nakagawa и др., J. Biol. Chem., 269, 10905 (1994));
- адгезионные протеины, как интегрин, бета-2-протеин
(Nueda и др., J. Biol. Chem., 268, 19305 (1993)).
В качестве активаторной последовательности согласно настоящему изобретению, кроме того, нужно понимать промоторные последовательности генов протеинов, которые интенсивно образуются при иммунной реакции в макрофагах и/или лимфоцитах. Сюда относятся, например:
- интерлейкин-1
(Bensi и др., Gene, 52, 95 (1987); Fibbe и др., Blut., 59, 147 (1989));
- рецептор интерлейкина-1
(Colotta и др., Immunol. Today, 15, 562 (1994); Sims и др., Clin. Immunol. Immunopath., 72, 9 (1994); Ye и др., PNAS USA, 90, 2295 (1993));
- интерлейкин-2
(Jansen и др. , Cell, 39, 207 (1994); Ohbo и др., J. Biol. Chem., 270, 7479 (1995));
- рецептор интерлейкина-2 (Semenzato и др., Int. J. Clin. Lab. Res., 22, 133 (1992));
- γ-интерферон
(Kirchner, DMW, 111, 64 (1986); Lehmann и др., J. Immunol., 153, 165 (1994));
- интерлейкин-4
(Paul, Blood, 77, 1859 (1991); te Velde и др., Blood, 76, 1392 (1990));
- рецептор интерлейкина-4
(Vallenga и др., Leukemia, 7, 1131 (1993); Gelizzi и др., Int. Immunol., 2, 669 (1990));
- интерлейкин-3
(Frendl, Int. J. Immunopharm., 14, 421 (1992));
- интерлейкин-5
(Azuma и др., Nucl. Acid Res., 14, 9149 (1986); Yokota и др., PNAS, 84, 7388 (1987));
- интерлейкин-6
(Brack и др., Int. J. Clin. Lab. Res., 22, 143 (1992);
- LIF
(Metcalf, Int. J. Cell Clon. , 9, 95 (1991); Samal, ВВА, 1260, 27 (1995));
- интерлейкин-7
(Joshi и др., 21, 681 (1991));
- интерлейкин-10
(Benjamin и др., Leuk. Lymph., 12, 205 (1994); Fluchiger и др., J. Exp. Med., 179, 91 (1994));
- интерлейкин-11
(Yang и др., Biofactors, 4, 15 (1992));
- интерлейкин-12
(Kiniwa и др., J. Clin. Invest., 90, 262 (1992), Gatelay, Cancer Invest. , 11, 500 (1993));
- интерлейкин-13
(Punnonen и др. , PNAS, 90, 3730 (1993); Muzio и др., Blood, 83., 1738 (1994));
- GM-CSF
(Metcalf, Cancer, 15, 2185 (1990));
- рецептор GM-CSF
Nakagawa и др., J. Biol. Chem., 269, 10905 (1994));
- адгезионные протеины, как интегрин, бета-2-протеин
(Nueda и др., J. Biol. Chem., 268, 19305 (1993)).
Промоторные последовательности этих протеинов известны из следующих публикаций:
- рецептор интерлейкина-1
(Ye и др., PNAS USA, 90, 2295 (1993));
- интерлейкин-lα.
- рецептор интерлейкина-1
(Ye и др., PNAS USA, 90, 2295 (1993));
- интерлейкин-lα.
(Hangen и др., Mol. Carcinog., 2, 68 (1986); Turner и др., J. Immunol., 143, 3556 (1989); Mori и др., Blood, 84, 1688 (1994));
- интерлейкин-lβ
(Fenton и др., J. Immunol., 138, 3972 (1987); Bensi и др., Cell Growth. Diff., 1, 491 (1990); Turner и др., J. Immunol., 143, 3556 (1989); Hiscott и др., Mol. Cell. Biol., 13, 6231 (1993));
- интерлейкин-2
(Fujita и др., Cell, 46, 401 (1986); Наmа и др., J. Exp. Med., 181, 1217 (1995); Kant и др. , Lymph. Rec. Interact., 179 (1989); Kamps и др., Mol. Cell. Biol., 10, 5464 (1990); Williams и др., J. Immunol., 141, 662 (1988); Brunvand, FASEB J., 6, А 998 (1992));
- рецептор интерлейкина-2
(Ohbo и др. , J. Biol. Chem., 270, 7479 (1995); Shibuya и др., Nucl. Acids Res. , 18, 3697 (1990); Lin и др., Mol. Cell. Biol., 13, 6201 (1993); Williams и др., J. Immunol., 141, 662 (1988));
- γ-интерферон
(Ye и др., J. Biol. Chem., 269, 25728 (1994));
- интерлейкин-4
(Rooney и др., EMBO J., 13, 625 (1994); Hama и др., J. Exp. Med., 181, 1217 (1995); Li-Weber и др., J. Immunol, 153, 4122 (1994); Min и др., J. Immunol., 148, 1913 (1992); Abe и др., PNAS, 89, 2864 (1992));
- рецептор интерлейкина-4
(Beckmann и др., Chem. Immunol., 51, 107 (1992); Ohara и др., PNAS, 85, 8221 (1988));
- интерлейкин-3
(Mathey-Prevot и др., PNAS USA, 87, 5046 (1990); Cameron и др., Blood, 83, 2851 (1994); Arai и др., Lymphokine Res., 9, 551 (1990));
- рецептор интерлейкина-3 (α-субъединица);
Miyajima и др., Blood, 85, 1246 (1995); Rapaport и др., Gene, 137, 333 (1993); Kosugi и др., ВВРС, 208, 360 (1995));
- рецептор интерлейкина-3 (β-субъединица)
(Gorman и др., J. Biol. Chem., 267, 15842 (1992); Kitamura и др., Cell, 66, 1165 (1991); Hayashida и др., PNAS USA, 87, 9655 (1990));
- интерлейкин-5
(Lee и др., J. Allerg. Clin. Immunol., 94, 594 (1994); Kauhansky и др., J. Immunol., 152, 1812 (1994); Staynov и др., PNAS USA, 92, 3606 (1995));
- интерлейкин-6
(Lu и др. , J. Biol. Chem., 270, 9748 (1995); Gruss и др., Blood, 80, 2563 (1992); Ray и др., PNAS, 85, 6701 (1988); Droogmans и др., DNA-Sequence, 3, 115 (1992); Mori и др., Blood, 84, 2904 (1994); Liberman и др., Mol. Cell. Biol., 10, 2327 (1990); Ishiki и др., Mol. Cell. Biol., 10, 2757 (1990));
- интерлейкин-7
(Pleiman и др. , Mol. Cell. Biol., 11, 3052 (1991); Lapton и др., J. Immunol., 144, 3592 (1990));
- интерлейкин-8
(Chang и др. , J. Biol. Chem., 269, 25277 (1994); Sprenger и др., J. Immunol., 153, 2524 (1994));
- интерлейкин-10
(Kim и др., J. Immunol., 148, 3616 (1992); Platzer и др., DNA-Sequence, 4, 399 (1994); Kube и др., Cytokine, 7, 1 (1995));
- интерлейкин-11
(Yang и др., J. Biol. Chem., 269, 32732 (1994));
- GM-CSF
(Nimer и др. , Mol. Cell. Biol., 10, 6084 (1990); Staynov и др., PNAS USA, 92, 3606 (1995); Koyano-Nakayawa и др., Int. Immunol., 5, 345 (1993); Ye и др., Nucl. Acids Res., 22, 5672 (1994));
- рецептор GM-CSF (α-цепь)
(Nakagawa и др., J. Biol. Chem., 269, 10905 (1994));
- рецептор макрофаг-колониестимулирующего фактора (M-CSF)
(Yue и др., Mol. Cell. Biol., 13, 3191 (1993); Zhang и др., Mol. Cell. Biol., 14, 373 (1994));
- тип I и тип II макрофаговых сапрофитных рецепторов
(Moulton и др., Mol. Cell. Biol., 14, 4408 (1994));
- интерлейкин-13
(Staynov и др., PNAS USA, 92, 3606 (1995));
- LIF
(Gough и др., Ciba Found. Symp., 167, 24 (1992); Stahl и др., Cytokine, 5, 386 (1993));
- интерфероновый регуляторный фактор-1, промотор которого стимулируется интерлейкином-6, также гамма- или бета-интерфероном
(Harrock и др., ЕМВО J., 13, 1942 (1994));
- реактивный промотор γ-интерферона
(Lamb и др., Blood, 83, 2063 (1994));
- γ-интерферон
(Hardy и др., PNAS, 82, 8173 (1985));
- MAC-1
(Dziennis и др., Blood, 85, 319 (1995); Bauer и др., Hum. Gene Ther., 5, 709 (1994); Hickstein и др., PNAS USA, 89, 2105 (1992));
- LFA-1
(Nueda и др., J. Biol. Chem., 268, 19305 (1993); Agura и др., Blood, 79, 602 (1992); Cornwell и др., PNAS USA, 90, 4221 (1993));
- р150, 95
(Noti и др., DNA and Cell. Biol., 11, 123 (1992); Lopezcabrera и др., J. Biol. Chem., 268, 1187 (1993)).
- интерлейкин-lβ
(Fenton и др., J. Immunol., 138, 3972 (1987); Bensi и др., Cell Growth. Diff., 1, 491 (1990); Turner и др., J. Immunol., 143, 3556 (1989); Hiscott и др., Mol. Cell. Biol., 13, 6231 (1993));
- интерлейкин-2
(Fujita и др., Cell, 46, 401 (1986); Наmа и др., J. Exp. Med., 181, 1217 (1995); Kant и др. , Lymph. Rec. Interact., 179 (1989); Kamps и др., Mol. Cell. Biol., 10, 5464 (1990); Williams и др., J. Immunol., 141, 662 (1988); Brunvand, FASEB J., 6, А 998 (1992));
- рецептор интерлейкина-2
(Ohbo и др. , J. Biol. Chem., 270, 7479 (1995); Shibuya и др., Nucl. Acids Res. , 18, 3697 (1990); Lin и др., Mol. Cell. Biol., 13, 6201 (1993); Williams и др., J. Immunol., 141, 662 (1988));
- γ-интерферон
(Ye и др., J. Biol. Chem., 269, 25728 (1994));
- интерлейкин-4
(Rooney и др., EMBO J., 13, 625 (1994); Hama и др., J. Exp. Med., 181, 1217 (1995); Li-Weber и др., J. Immunol, 153, 4122 (1994); Min и др., J. Immunol., 148, 1913 (1992); Abe и др., PNAS, 89, 2864 (1992));
- рецептор интерлейкина-4
(Beckmann и др., Chem. Immunol., 51, 107 (1992); Ohara и др., PNAS, 85, 8221 (1988));
- интерлейкин-3
(Mathey-Prevot и др., PNAS USA, 87, 5046 (1990); Cameron и др., Blood, 83, 2851 (1994); Arai и др., Lymphokine Res., 9, 551 (1990));
- рецептор интерлейкина-3 (α-субъединица);
Miyajima и др., Blood, 85, 1246 (1995); Rapaport и др., Gene, 137, 333 (1993); Kosugi и др., ВВРС, 208, 360 (1995));
- рецептор интерлейкина-3 (β-субъединица)
(Gorman и др., J. Biol. Chem., 267, 15842 (1992); Kitamura и др., Cell, 66, 1165 (1991); Hayashida и др., PNAS USA, 87, 9655 (1990));
- интерлейкин-5
(Lee и др., J. Allerg. Clin. Immunol., 94, 594 (1994); Kauhansky и др., J. Immunol., 152, 1812 (1994); Staynov и др., PNAS USA, 92, 3606 (1995));
- интерлейкин-6
(Lu и др. , J. Biol. Chem., 270, 9748 (1995); Gruss и др., Blood, 80, 2563 (1992); Ray и др., PNAS, 85, 6701 (1988); Droogmans и др., DNA-Sequence, 3, 115 (1992); Mori и др., Blood, 84, 2904 (1994); Liberman и др., Mol. Cell. Biol., 10, 2327 (1990); Ishiki и др., Mol. Cell. Biol., 10, 2757 (1990));
- интерлейкин-7
(Pleiman и др. , Mol. Cell. Biol., 11, 3052 (1991); Lapton и др., J. Immunol., 144, 3592 (1990));
- интерлейкин-8
(Chang и др. , J. Biol. Chem., 269, 25277 (1994); Sprenger и др., J. Immunol., 153, 2524 (1994));
- интерлейкин-10
(Kim и др., J. Immunol., 148, 3616 (1992); Platzer и др., DNA-Sequence, 4, 399 (1994); Kube и др., Cytokine, 7, 1 (1995));
- интерлейкин-11
(Yang и др., J. Biol. Chem., 269, 32732 (1994));
- GM-CSF
(Nimer и др. , Mol. Cell. Biol., 10, 6084 (1990); Staynov и др., PNAS USA, 92, 3606 (1995); Koyano-Nakayawa и др., Int. Immunol., 5, 345 (1993); Ye и др., Nucl. Acids Res., 22, 5672 (1994));
- рецептор GM-CSF (α-цепь)
(Nakagawa и др., J. Biol. Chem., 269, 10905 (1994));
- рецептор макрофаг-колониестимулирующего фактора (M-CSF)
(Yue и др., Mol. Cell. Biol., 13, 3191 (1993); Zhang и др., Mol. Cell. Biol., 14, 373 (1994));
- тип I и тип II макрофаговых сапрофитных рецепторов
(Moulton и др., Mol. Cell. Biol., 14, 4408 (1994));
- интерлейкин-13
(Staynov и др., PNAS USA, 92, 3606 (1995));
- LIF
(Gough и др., Ciba Found. Symp., 167, 24 (1992); Stahl и др., Cytokine, 5, 386 (1993));
- интерфероновый регуляторный фактор-1, промотор которого стимулируется интерлейкином-6, также гамма- или бета-интерфероном
(Harrock и др., ЕМВО J., 13, 1942 (1994));
- реактивный промотор γ-интерферона
(Lamb и др., Blood, 83, 2063 (1994));
- γ-интерферон
(Hardy и др., PNAS, 82, 8173 (1985));
- MAC-1
(Dziennis и др., Blood, 85, 319 (1995); Bauer и др., Hum. Gene Ther., 5, 709 (1994); Hickstein и др., PNAS USA, 89, 2105 (1992));
- LFA-1
(Nueda и др., J. Biol. Chem., 268, 19305 (1993); Agura и др., Blood, 79, 602 (1992); Cornwell и др., PNAS USA, 90, 4221 (1993));
- р150, 95
(Noti и др., DNA and Cell. Biol., 11, 123 (1992); Lopezcabrera и др., J. Biol. Chem., 268, 1187 (1993)).
5.2. Выбор активного вещества для ингибирования свертывания
В качестве активного вещества согласно настоящему изобретению нужно понимать ДНК-последовательность, которая кодирует протеин, прямо или косвенно ингибирующий агрегацию тромбоцитов, или фактор свертывания крови, или стимулирующий фибринолиз.
В качестве активного вещества согласно настоящему изобретению нужно понимать ДНК-последовательность, которая кодирует протеин, прямо или косвенно ингибирующий агрегацию тромбоцитов, или фактор свертывания крови, или стимулирующий фибринолиз.
Такого рода активное вещество обозначают как ингибитор свертывания. В качестве ингибиторов свертывания можно использовать гены, например, активаторов плазминогена (РА), так, тканевый PA (tPA), или урокиназоподобный PA (uPA), или протеин С, антитромбин-III, C-IS-ингибитор, α-1-антитрипсин, ингибитор метаболического пути тканевого фактора (TFPI) или гирудин. ДНК-последовательности этих ингибиторов свертывания уже описаны в разделе 4.3.
5.3. Комбинация двух одинаковых или разных активных веществ для ингибирования свертывания
Предметом изобретения, далее, является активное вещество, в котором встречается сочетание ДНК-последовательностей: двух одинаковых, ингибирующих свертывание веществ (А, А), или двух различных, ингибирующих свертывание веществ (А, Б). Для экспрессии обеих ДНК-последовательностей предпочтительно промежуточно включают кДНК "внутреннего рибосомного аминоацильного сайта" (IRES) в качестве регуляторного элемента.
Предметом изобретения, далее, является активное вещество, в котором встречается сочетание ДНК-последовательностей: двух одинаковых, ингибирующих свертывание веществ (А, А), или двух различных, ингибирующих свертывание веществ (А, Б). Для экспрессии обеих ДНК-последовательностей предпочтительно промежуточно включают кДНК "внутреннего рибосомного аминоацильного сайта" (IRES) в качестве регуляторного элемента.
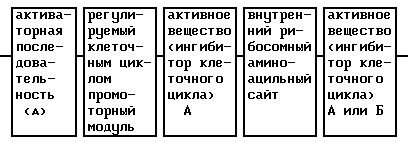
Такого рода IRES описываются, например, Montford и Smith (TIG, 11, 179 (1995); Kaufman и др. , Nucl. Acids Res., 19, 4485 (1991); Morgan и др., Nucl. Acids Res. , 20, 1293 (1992); Dirks и др., Gene, 128, 247 (1993); Pelletier и Sonenberg, Nature, 334, 320 (1988) и Sugitomo и др., Bio-Techn., 12, 694 (1994).
Так, можно использовать кДНК IRES-последовательности полиовируса (положение ≤ 140 до ≥ 630 5'-UTR (Pelletier и Sonenberg, Nature, 334, 320 (1988)) для связывания ДНК антитромботического вещества А (по 3'-концу) и ДНК антитромботического вещества Б (по 5'-концу).
Такого рода сочетание двух одинаковых или разных генов способствует аддитивному действию (в случае одинаковых генов) или синергическому действию выбранных антитромботических веществ.
5.4. Выбор лиганда для ингибирования свертывания
В качестве лиганда вирусных или невирусных векторов, например, в коллоидных дисперсиях, содержащих полилизин-лиганд-конъюгаты, предпочтительны вещества, которые связываются с поверхностью гладкомышечных клеток, или пролиферирующих эндотелиальных клеток, или активированных макрофагов и/или лимфоцитов.
В качестве лиганда вирусных или невирусных векторов, например, в коллоидных дисперсиях, содержащих полилизин-лиганд-конъюгаты, предпочтительны вещества, которые связываются с поверхностью гладкомышечных клеток, или пролиферирующих эндотелиальных клеток, или активированных макрофагов и/или лимфоцитов.
а). Лиганды гладкомышечных клеток
Примеры лигандов, которые связываются с гладкомышечными клетками, уже указаны в разделе 4.4.
Примеры лигандов, которые связываются с гладкомышечными клетками, уже указаны в разделе 4.4.
б). Лиганды активированных эндотелиальных клеток
Сюда относятся согласно изобретению антитела или фрагменты антител, направленные против мембранных структур эндотелиальных клеток, которые описываются, например, Burrows и др. (Pharmac. Ther., 64, 155 (1994)), Hughes и др. (Cancer Res. , 49, 6214 (1989)) и Maruyama и др. (PNAS USA, 87, 5744 (1990)). В особенности сюда относятся антитела против VEGF-рецепторов.
Сюда относятся согласно изобретению антитела или фрагменты антител, направленные против мембранных структур эндотелиальных клеток, которые описываются, например, Burrows и др. (Pharmac. Ther., 64, 155 (1994)), Hughes и др. (Cancer Res. , 49, 6214 (1989)) и Maruyama и др. (PNAS USA, 87, 5744 (1990)). В особенности сюда относятся антитела против VEGF-рецепторов.
Мышиные моноклональные антитела предпочтительно нужно использовать в гуманизированной форме. Гуманизирование осуществляют описанным в разделе 4.4. образом. Фрагменты антител получают согласно уровню техники, например, описанным в разделе 4.4 способом.
К лигандам, далее, относятся все активные вещества, которые связываются с мембранными структурами или мембранными рецепторами эндотелиальных клеток. Например, сюда относятся вещества, которые содержат концевую маннозу, далее, интерлейкин-1 или факторы роста или их фрагменты, соответственно их частичные последовательности, которые связываются с рецепторами, экспрессируемыми эндотелиальными клетками, как, например, PDGF, bFGF, VEGF, TGF-β (Pusztain и др., J. Pathol., 169, 191 (1993)).
Далее, к таким лигандам относятся адгезионные молекулы, которые связываются с активированными и/или пролиферирующими эндотелиальными клетками. Такого рода адгезионные молекулы, как, например, SLex, LFA-1, MAC-1, LECAM-1 или VLA-4, уже описаны (обзоры Augustin-Voss и др., J. Cell. Biol., 119, 483 (1992); Pauli и др., Cancer Metast. Rev., 9, 175 (1990); Honn и др., Cancer Metast. Rev., 11, 353 (1992)).
в). Лиганды активированных макрофагов и/или активированных лимфоцитов
К лигандам согласно изобретению относятся, далее, вещества, которые специфически связываются с поверхностью иммунных клеток. Сюда относятся антитела или фрагменты антител, направленные против мембранных структур иммунных клеток, которые описываются, например, Powelson и др., Biotech. Adv., 11, 725 (1993).
К лигандам согласно изобретению относятся, далее, вещества, которые специфически связываются с поверхностью иммунных клеток. Сюда относятся антитела или фрагменты антител, направленные против мембранных структур иммунных клеток, которые описываются, например, Powelson и др., Biotech. Adv., 11, 725 (1993).
Далее, к лигандам также относятся моноклональные или поликлональные антитела или фрагменты антител, которые с помощью своих постоянных доменов связываются с Fc - или μ-рецепторами иммунных клеток (Rojanasakul и др., Pharm. Res., 11, 1731 (1994)).
Здесь также предпочтительно используют мышиные моноклональные антитела в гуманизированной форме (см. раздел 4.4.) и фрагменты получают, например, с помощью указанной в разделе 4.4 методики.
К лигандам, далее, относятся все вещества, которые связываются с мембранными рецепторами на поверхности иммунных клеток. Например, сюда относятся факторы роста, как цитокины, EGF, TGF, FGF или PDGF или их фрагменты, соответственно их частичные последовательности, которые связываются с рецепторами, экспрессируемыми такого рода клетками.
5.5. Получение активного вещества для ингибирования свертывания
Получение предлагаемого согласно изобретению активного вещества описывается подробнее, руководствуясь следующими примерами.
Получение предлагаемого согласно изобретению активного вещества описывается подробнее, руководствуясь следующими примерами.
а). Конструкция химерного промотора эндотелина-1-CDE-CHR-Inr
Человеческий промотор эндотелин-1 (положение ≤ -170 до ≥ -10; Wilson и др., Mol. Cell. Biol., 10, 4854 (1990)) или укороченный на ТАТА-блок вариант (положение ≤ -170 до ≥ -40) по его 3'-концу связывают с 5'-концом CDE-CHR-Inr-модуля (положение ≤ -20 до ≥ +121) человеческого cdc25C-гена (Lucibello и др. , EMBO J. , 14, 132 (1995)) (фиг.9). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов.
Человеческий промотор эндотелин-1 (положение ≤ -170 до ≥ -10; Wilson и др., Mol. Cell. Biol., 10, 4854 (1990)) или укороченный на ТАТА-блок вариант (положение ≤ -170 до ≥ -40) по его 3'-концу связывают с 5'-концом CDE-CHR-Inr-модуля (положение ≤ -20 до ≥ +121) человеческого cdc25C-гена (Lucibello и др. , EMBO J. , 14, 132 (1995)) (фиг.9). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов.
б). Конструкция плазмиды, содержащей химерный промотор эндотелин-1-CDE-CHR-Inr в центральной составной части активного вещества
Описанную транскрипционную единицу химерного промоторного модуля эндотелина-1 связывают по ее 3'-концу с 5'-концом ДНК, которая содержит полную кодирующую область тканевого плазминогенного активатора (положение ≤ 85 до ≥ 1753; Pennica и др., Nature, 301, 214 (1983)) (фиг.9). Эта ДНК также содержит необходимую для секреции сигнальную последовательность. Транскрипционные контрольные единицы и ДНК тканевого плазминогенного активатора клонируют в pUC19/19 или производимых от Bluescript плазмидных векторах, которые можно использовать непосредственно или в коллоидальных дисперсионных системах для введения ин виво. Альтернативно, химерные гены трансферируют в вирусные векторы или другие пригодные векторы и инъецируют.
Описанную транскрипционную единицу химерного промоторного модуля эндотелина-1 связывают по ее 3'-концу с 5'-концом ДНК, которая содержит полную кодирующую область тканевого плазминогенного активатора (положение ≤ 85 до ≥ 1753; Pennica и др., Nature, 301, 214 (1983)) (фиг.9). Эта ДНК также содержит необходимую для секреции сигнальную последовательность. Транскрипционные контрольные единицы и ДНК тканевого плазминогенного активатора клонируют в pUC19/19 или производимых от Bluescript плазмидных векторах, которые можно использовать непосредственно или в коллоидальных дисперсионных системах для введения ин виво. Альтернативно, химерные гены трансферируют в вирусные векторы или другие пригодные векторы и инъецируют.
в). Конструкция химерного промотора CDE-CHR-Inr миогенина
Человеческий миогениновый промотор (положение ≤ -210 до ≥ +54 ДНК-последовательности, опубликованной Salmin и др. , J. Cell. Biol., 115, 905 (1991)) по его 3'-концу связывают с 5'-концом CDE-CHR-Inr-модуля человеческого cdc25C-гена (положение ≤ -20 до ≥ +121 последовательности, опубликованной Lucibello и др., ЕМВО J., 14, 132 (1995)) (см. фиг.10). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов. Кроме того, используют различные фрагменты промоторной последовательности миогенина (см. фиг.10). Так, применяют содержащую ТАТА-блок ДНК-последовательность промотора миогенина. Однако равным образом также можно использовать промоторную последовательность в положении ≤ -210 до ≥ -40.
Человеческий миогениновый промотор (положение ≤ -210 до ≥ +54 ДНК-последовательности, опубликованной Salmin и др. , J. Cell. Biol., 115, 905 (1991)) по его 3'-концу связывают с 5'-концом CDE-CHR-Inr-модуля человеческого cdc25C-гена (положение ≤ -20 до ≥ +121 последовательности, опубликованной Lucibello и др., ЕМВО J., 14, 132 (1995)) (см. фиг.10). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов. Кроме того, используют различные фрагменты промоторной последовательности миогенина (см. фиг.10). Так, применяют содержащую ТАТА-блок ДНК-последовательность промотора миогенина. Однако равным образом также можно использовать промоторную последовательность в положении ≤ -210 до ≥ -40.
г). Конструкция плазмиды, содержащей химерный промотор CDE-CHR-Inr миогенина в центральной составной части активного вещества
Таким образом полученную контрольную транскрипционную единицу химерного промоторного модуля миогенина по ее 3'-концу связывают с 5'-концом ДНК, которая содержит полную кодирующую область тканевого плазминогенного активатора (см. фиг. 10). Эта ДНК также содержит необходимую для секреции сигнальную последовательность. Транскрипционные контрольные единицы и ДНК тканевого плазминогенного активатора клонируют в pUC18/19 или производимых от Bluescript плазмидных векторах, которые непосредственно или в коллоидальной дисперсионной системе можно использовать для введения ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы и инъецировать.
Таким образом полученную контрольную транскрипционную единицу химерного промоторного модуля миогенина по ее 3'-концу связывают с 5'-концом ДНК, которая содержит полную кодирующую область тканевого плазминогенного активатора (см. фиг. 10). Эта ДНК также содержит необходимую для секреции сигнальную последовательность. Транскрипционные контрольные единицы и ДНК тканевого плазминогенного активатора клонируют в pUC18/19 или производимых от Bluescript плазмидных векторах, которые непосредственно или в коллоидальной дисперсионной системе можно использовать для введения ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы и инъецировать.
д). Конструкция плазмиды, содержащей два гена активных веществ
Описанную в п. в) транскрипционную единицу CDE-CHR-Inr миогенина по ее 3'-концу связывают с 5'-концом ДНК ингибитора метаболического пути тканевого фактора (TFPI; положение ≤ 133 до ≥ 957; Wun и др., J. Biol. Chem., 263, 6001 (1988); или положение ≤ 382 до ≥ 1297; Girard и др., Thromb. Res., 55, 37 (1989)). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов.
Описанную в п. в) транскрипционную единицу CDE-CHR-Inr миогенина по ее 3'-концу связывают с 5'-концом ДНК ингибитора метаболического пути тканевого фактора (TFPI; положение ≤ 133 до ≥ 957; Wun и др., J. Biol. Chem., 263, 6001 (1988); или положение ≤ 382 до ≥ 1297; Girard и др., Thromb. Res., 55, 37 (1989)). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов.
3'-конец ДНК TFPI теперь связывают с 5'-концом кДНК внутреннего рибосомного аминоацильного сайта (положение ≤ 140 до ≥ 630; Pelletier и Sonnenberg, Nature., 334, 320 (1988)) и затем его 3'-конец связывают с 5'-концом ДНК тканевого плазминогенного активатора (см. фиг.11). Это таким образом полученное активное вещество, наконец, клонируют в pUC18/19 или производимых от Bluescript плазмидных векторах, которые непосредственно или в коллоидальной дисперсионной системе можно использовать для введения ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы и инъецировать.
6. Действие активного вещества на гладкомышечные клетки и/или на свертывание
Активное вещество согласно настоящему изобретению после локального или системного, предпочтительно внутривенного или внутриартериального введения оказывает преимущественное, если не исключительное, действие на такие гладкомышечные клетки, которые непосредственно доступны вследствие повреждения или травмирования сосуда (в особенности слой эндотелия) и, возможно, после попадания во внутреннюю оболочку объема кровеносного сосуда.
Активное вещество согласно настоящему изобретению после локального или системного, предпочтительно внутривенного или внутриартериального введения оказывает преимущественное, если не исключительное, действие на такие гладкомышечные клетки, которые непосредственно доступны вследствие повреждения или травмирования сосуда (в особенности слой эндотелия) и, возможно, после попадания во внутреннюю оболочку объема кровеносного сосуда.
Благодаря сочетанию специфической к ткани активаторной последовательности и регулируемого клеточным циклом репрессорного модуля обеспечивается то, что ингибитор клеточного цикла активируется преобладающе или исключительно в делящихся гладкомышечных клетках.
Благодаря предлагаемому согласно изобретению применению мутированных ингибиторов клеточного цикла обеспечивается их более продолжительное по времени ингибирующее пролиферацию действие.
Активное вещество согласно настоящему изобретению, далее, позволяет осуществлять то, что после локального (например, в ткань, полости тела или промежуточные пространства между тканями) или после системного, предпочтительно внутривенного или внутриартериального введения, преобладающе, если не исключительно, в гладкомышечные клетки, активированные пролиферирующие эндотелиальные клетки, активированные лимфоциты или активированные макрофаги экспрессируют антитромботическое вещество, и оно, таким образом, высвобождается в месте возникновения тромбоза.
Так как активное вещество как за счет его специфичности к клетке, так и также за счет его специфичности к клеточному циклу гарантирует высокую степень безопасности, его также можно использовать в высоких дозировках и в случае необходимости многократно с интервалами от дней или недель для профилактики или терапии закупорки сосудов, вызываемой за счет пролиферирующих гладкомышечных клеток, и/или для профилактики и/или терапии тромбозов.
Пояснение к фигурам 1-11
Фиг.1:
Нуклеотидная последовательность cdc25C-промоторного участка с найденными ин виво сайтами связывания протеина (геномный диметилсульфатный футпринтинг; закрашенный кружок - полная конститутивная защита; незакрашенный кружок - частичная конститутивная защита; звездочка - регулируемая клеточным циклом Gl-специфическая защита; CBS - конститутивный связывающий сайт; CDE - зависимый от клеточного цикла элемент. Окрашенные в серый цвет области указывают YC-блоки (NF-Y-сайты связывания). Стартовые участки маркированы закрашенными квадратами.
Фиг.1:
Нуклеотидная последовательность cdc25C-промоторного участка с найденными ин виво сайтами связывания протеина (геномный диметилсульфатный футпринтинг; закрашенный кружок - полная конститутивная защита; незакрашенный кружок - частичная конститутивная защита; звездочка - регулируемая клеточным циклом Gl-специфическая защита; CBS - конститутивный связывающий сайт; CDE - зависимый от клеточного цикла элемент. Окрашенные в серый цвет области указывают YC-блоки (NF-Y-сайты связывания). Стартовые участки маркированы закрашенными квадратами.
Фиг.2:
Дерепрессия сdс25С-промотора специфически в G0 путем мутации cdc.
Дерепрессия сdс25С-промотора специфически в G0 путем мутации cdc.
Фиг.3:
Схематическое изображение регуляции сdс25С-энхансера за счет CDE.
Схематическое изображение регуляции сdс25С-энхансера за счет CDE.
Фиг.4:
G0/G1 - специфическая репрессия 8У40-энхансера за счет CDE.
G0/G1 - специфическая репрессия 8У40-энхансера за счет CDE.
Фиг.5:
Гомологии в CDE-CHR-области и расположенные у 5'-концов YC-блоки в cdc25C, циклине-А и cdc2-промоторах.
Гомологии в CDE-CHR-области и расположенные у 5'-концов YC-блоки в cdc25C, циклине-А и cdc2-промоторах.
Фиг.6:
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого на 3'-конце промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческой β-глюкуронидазы (полный кодирующий участок, положение ≤ 27 до ≥ 1982) в качестве эффектора (Oshima и др., PNAS USA, 84, 685 (1987)). Данные о расположении относятся к данным Salminen и др., J. Cell. Biol., 115, 905 (1991) для гена миогенина, соответственно к системе cdc25C, применяемой Lucibello и др., ЕМВО J., 14, 132 (1995).
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого на 3'-конце промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческой β-глюкуронидазы (полный кодирующий участок, положение ≤ 27 до ≥ 1982) в качестве эффектора (Oshima и др., PNAS USA, 84, 685 (1987)). Данные о расположении относятся к данным Salminen и др., J. Cell. Biol., 115, 905 (1991) для гена миогенина, соответственно к системе cdc25C, применяемой Lucibello и др., ЕМВО J., 14, 132 (1995).
Фиг.7:
Замена гомологичной сигнальной последовательности β-глюкуронидазы гетерологичной сигнальной последовательностью (человеческого иммуноглобулина). Данные о расположении сигнальной последовательности (MGWSCIILFLVATAT) иммуноглобулина (HuVH-cAMP) относятся к данным Riechmann и др., Nature, 332, 323 (1988).
Замена гомологичной сигнальной последовательности β-глюкуронидазы гетерологичной сигнальной последовательностью (человеческого иммуноглобулина). Данные о расположении сигнальной последовательности (MGWSCIILFLVATAT) иммуноглобулина (HuVH-cAMP) относятся к данным Riechmann и др., Nature, 332, 323 (1988).
Альтернатива: Встраивание сигнального пептида иммуноглобулина для лучшего внеклеточного выделения β-глюкуронидазы; см.(Б).
Фиг.8:
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого на 3'-конце промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческой β-глюкуронидазы, внутреннего рибосомного аминоациального сайта в качестве регулирующей нуклеотидной последовательности и ДНК тканевого плазминогенного активатора. Данные о расположении относятся к таковым Salminen и др., J. Cell. Biol., 115, 905 (1991) для миогенина; Lucibello и др., ЕМВО J., 14, 132 (1995) для элемента CDE/CHR-Inr; Oshima и др. , PNAS USA, 84, 685 (1987) для β-глюкуронидазы; Riechmann и др., Nature, 332? 323 (1988) для сигнальной последовательности иммуноглобулина; Pelletier и Sonenberg, Nature, 334, 320 (1988) для внутреннего рибосомного аминоациального сайта полиовируса и Pennica и др., Nature, 301, 214 (1983) для человеческого тканевого плазминогенного активатора.
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого на 3'-конце промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческой β-глюкуронидазы, внутреннего рибосомного аминоациального сайта в качестве регулирующей нуклеотидной последовательности и ДНК тканевого плазминогенного активатора. Данные о расположении относятся к таковым Salminen и др., J. Cell. Biol., 115, 905 (1991) для миогенина; Lucibello и др., ЕМВО J., 14, 132 (1995) для элемента CDE/CHR-Inr; Oshima и др. , PNAS USA, 84, 685 (1987) для β-глюкуронидазы; Riechmann и др., Nature, 332? 323 (1988) для сигнальной последовательности иммуноглобулина; Pelletier и Sonenberg, Nature, 334, 320 (1988) для внутреннего рибосомного аминоациального сайта полиовируса и Pennica и др., Nature, 301, 214 (1983) для человеческого тканевого плазминогенного активатора.
Фиг.9:
Химерная конструкция, состоящая из различных частей человеческого промотора эндотелина-1, слитого по 3'-концу промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческого тканевого плазминогенного активатора (полная кодирующая область; Pennica и др., Nature, 301, 214 (1983)) в качестве эффектора. Данные о расположении относятся к таковым Wilson и др., Mol. Cell. Biol., 10, 4854 (1990) для гена эндотелина-1, соответственно к используемой Lucibello и др., ЕМВО J., 14, 132 (1995), системе cdc25C.
Химерная конструкция, состоящая из различных частей человеческого промотора эндотелина-1, слитого по 3'-концу промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческого тканевого плазминогенного активатора (полная кодирующая область; Pennica и др., Nature, 301, 214 (1983)) в качестве эффектора. Данные о расположении относятся к таковым Wilson и др., Mol. Cell. Biol., 10, 4854 (1990) для гена эндотелина-1, соответственно к используемой Lucibello и др., ЕМВО J., 14, 132 (1995), системе cdc25C.
Фиг.10:
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого по 3'-концу промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческого тканевого плазминогенного активатора (полная кодирующая область) в качестве эффектора. Данные о расположении относятся к таковым Salminen и др., J. Cell. Biol., 115, 905 (1991) для гена миогенина, соответственно к используемой Lucibello и др., ЕМВО J. , 14, 132 (1995) системе cdc25C и к данным Pennica и др., Nature, 301, 214 (1983) для тканевого плазминогенного активатора.
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого по 3'-концу промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческого тканевого плазминогенного активатора (полная кодирующая область) в качестве эффектора. Данные о расположении относятся к таковым Salminen и др., J. Cell. Biol., 115, 905 (1991) для гена миогенина, соответственно к используемой Lucibello и др., ЕМВО J. , 14, 132 (1995) системе cdc25C и к данным Pennica и др., Nature, 301, 214 (1983) для тканевого плазминогенного активатора.
Фиг.11:
Химерная конструкция с двумя эффекторными генами.
Химерная конструкция с двумя эффекторными генами.
Данные о расположении для ингибитора метаболического пути тканевого фактора относятся к данным
*) Wun и др., J. Biol. Chem., 263, 6001 (1988);
**) Girard и др., Thromb. Res., 55, 37 (1989);
а данные для внутреннего рибосомного аминоациального сайта (IRES) кДНК основываются на таковых Pelletier и Sonenberg, Nature, 334, 320 (1988).
*) Wun и др., J. Biol. Chem., 263, 6001 (1988);
**) Girard и др., Thromb. Res., 55, 37 (1989);
а данные для внутреннего рибосомного аминоациального сайта (IRES) кДНК основываются на таковых Pelletier и Sonenberg, Nature, 334, 320 (1988).
Claims (34)
1. ДНК-конструкция, состоящая из активаторной последовательности, регулируемого клеточным циклом промоторного модуля, включающего элементы CDE-CHR-Inr и содержащего положения ≤ -20 до ≥ +30 cdc25C-промоторного участка (нуклеотидная последовательность GGCTGGCGG-AAGGTTTGAATGGTCAACGCCTGCGG-CTGTTGATATTCTTG), причем CDE представляет собой "зависимый от клеточного цикла элемент" (нуклеотидная последовательность TGGCGG), CHR представляет собой "гомологичную гену клеточного цикла область" (нуклеотидная последовательность GTTTGAA), и Inr означает сайт инициации (положение +1), а также важные для инициации соседние последовательности, и ДНК-последовательности, кодирующей активное вещество, которое представляет собой ингибитор клеточного цикла и/или ингибитор свертывания.
2. ДНК-конструкция по п.1, отличающаяся тем, что активаторная последовательность представлена промоторной или энхансерной последовательностями и регулируется факторами транскрипции, образующимися в гладкомышечных клетках.
3. ДНК-конструкция по п. 2, отличающаяся тем, что содержит в качестве активаторной последовательности CMV-энхансер, CMV-промотор или SV40-промотор; или промоторную последовательность гена тропомиозина, α-актина, α-миозина, PDGF-рецепторов, FGF-рецепторов, MRF-4, ацетилхолиновых рецепторов, фосфофруктокиназы-А, фосфоглицератмутазы, тропонина С, миогенина, рецепторов эндотелина или десмина; или в качестве активаторной последовательности множественные копии сайта связывания специфических к мышцам HLH-протеинов (Е-блок) или GATA-4; или промоторную последовательность VEGF, или энхансерную последовательность VEGF, или ДНК-последовательность с-Src или v-Src, которые регулируют VEGF-ген.
4. ДНК-конструкция по любому из пп.1-3, отличающаяся тем, что ДНК-последовательность кодирует активное вещество ингибитора клеточного цикла, представляющего собой: протеин ретинобластомы pRb/р110, протеин р107 или протеин р130; или протеин р53; или протеин р21, протеин р16 или другой ингибитор "циклин-зависимой киназы (СdК)"; или протеин GADD45; или bak-протеин; или цитостатический или цитотоксический протеин; или фермент для расщепления предшественников цитостатических средств до цитостатических средств.
5. ДНК-конструкция по п.4, отличающаяся тем, что протеин регинобластомы (pRb/р110) за счет замены аминокислот в положениях 246, 350, 601, 605, 780, 786, 787, 800 и 804 становится более нефосфорилируемым, однако, без ухудшения его активности связывания с большим Т-антигеном; например, за счет того, что аминокислоты Thr-246, Ser-601, Ser-605, Ser-780, Ser-786, Ser-787 и Ser-800 заменяются на A1а, аминокислота Thr-350 заменяется на Arg, и аминокислота Ser-804 заменяется на Glu; или протеин р107 мутирован аналогично протеину рRb/110; или протеин р130 мутирован аналогично протеину рRb/110.
6. ДНК-конструкция по п.4, отличающаяся тем, что протеин р53С на С-конце укорочен на серин-392.
7. ДНК-конструкция по любому из пп.1-3, отличающаяся тем, что ингибитором клеточного цикла является перфорин, гранзим, α-фактор некроза опухоли или β-фактор некроза опухоли.
8. ДНК-конструкция по любому из пп.1-3, отличающаяся тем, что ингибитором клеточного цикла является фермент и этот фермент представляет собой тимидинкиназу вируса простого герпеса, цитозиндезаминазу, тимидинкиназу вируса ветряной оспы, нитроредуктазу, β-глюкуронидазу (в особенности человеческую, растительную или бактериальную β-глюкуронидазу), карбоксипептидазу (предпочтительно бактерий рода Pseudomonas), лактомазу (предпочтительно Bacillus cereus), пироглутаматаминопептидазу, D-аминопептидазу, оксидазу, пероксидазу, фосфатазу, гидроксинитриллазу, протеазу, эстеразу или гликозидазу, причем сигнальная последовательность этого фермента может быть гомологична или для лучшего клеточного выделения также гетерологична.
9. ДНК-конструкция по п.8, отличающаяся тем, что путем точечной мутации к ДНК-последовательности фермента снижено накопление лизосом и повышено внеклеточное выделение.
10. ДНК-конструкция по любому из пп.1-9, отличающаяся тем, что содержит ДНК-последовательности нескольких одинаковых или разных ингибиторов клеточного цикла, причем возможно две ДНК-последовательности связаны друг с другом через ДНК-последовательность внутреннего рибосомного аминоацильного сайта.
11. ДНК-конструкция по любому из пп.1-3, отличающаяся тем, что ДНК-последовательность кодирует активное вещество ингибитора свертывания.
12. ДНК-конструкция по любому из пп.1-9, отличающаяся тем, что ДНК-последовательность ингибитора клеточного цикла дополнена ДНК-последовательностью ингибитора свертывания, и эти обе ДНК-последовательности связаны друг с другом через ДНК-последовательность внутреннего рибосомного аминоацильного сайта.
13. ДНК-конструкция по любому из п.11 или 12, отличающаяся тем, что ДНК-последовательность ингибитора кодирует tPA, uPA, гибридные молекулы tPA и uPA, протеин С, антитромбин III, TFPI, С-1-ингибитор, α-1-антитрипсин или гирудин.
14. ДНК-конструкция по п.1, отличающаяся тем, что активаторная последовательность представлена промоторной или энхансерной последовательностями и регулируется факторами транскрипции, образующимися в пролифелирующих эндотелиальных клетках.
15. ДНК-конструкция по п.14, отличающаяся тем, что содержит промоторную последовательность эндотелиального переносчика глюкозы-1, эндоглина, VEGF-рецептора-1 или -2, рецепторной тирозинкиназы til-1 или til-2, рецептора В61, лиганда В61, эндотелина, в особенности эндотелина-В-1, манноза-6-фосфатного рецептора, интерлейкина-1α или интерлейкина-1β, рецептора интерлекина-1, VCAM-1, фактора Виллебранда; или синтетическую активаторную последовательность из олигомеризованных сайтов связывания факторов транскрипции, которые предпочтительно или селективно активны в эндотелиальных клетках, как GATA-2, сайтом связывания которого является 5'-ТТАТСТ-3'.
16. ДНК-конструкция по п.1, отличающаяся тем, что активаторная последовательность представлена промоторной или энхансерной последовательностями и регулируется факторами транскрипции, образующимися при активировании макрофагами и в макрофагах или в лимфоцитах.
17. ДНК-конструкция по п.16, отличающаяся тем, что содержит промоторную последовательность цитокинов или их рецепторов, в особенности интерлейкина-1α или интерлейкина-1β, рецептора интерлейкина-1, интерлейкина-2, рецептора интерлейкина-2, γ-интерферона, интерлейкина-3, рецептора интерлейкина-3, интерлейкина-4, рецептора интерлейкина-4, интерлейкина-5, интерлейкина-6, интерлейкина-7, интерлейкина-8, интерлейкина-9, интерлейкина-10, интерлейкина-11, интерлейкина-12, интерлейкина-13, M-CSF-рецептора, типа I или типа II рецептора сапрофитных макрофагов, фактора 1 регуляции интерферона, γ-интерферона, чувствительного к γ-интерферону промотора, гранулоцит-макрофаг-колониестимулирующего фактора (GM-CSF), GM-CSF-рецептора, гранулоцит-колониестимулирующего фактора (G-CSF), адгезионных протеинов, как LFA-1, Mac-1 или р150/95, или ингибирующего лейкемию фактора (LIF).
18. ДНК-конструкция по любому из пп.14-17, отличающаяся тем, что ДНК-последовательность кодирует активное вещество ингибитора свертывания.
19. ДНК-конструкция по п.18, отличающаяся тем, что ДНК-последовательность ингибитора свертывания кодирует тканевый плазминогенный активатор (tPA), урокиназу (uPA), гибрид tPA и uPA, протеин С, антитромбин-III, ингибитор метаболического пути тканевого фактора, С-1-игибитор, α-1-антитрипсин или гирудин.
20. ДНК-конструкция по любому из пп.1-3 и 14-19, отличающаяся тем, что содержит ДНК-последовательность нескольких одинаковых или разных ингибиторов свертывания, причем возможно две ДНК-последовательности связаны друг с другом через ДНК-последовательность внутреннего рибосомного аминоациального сайта.
21. ДНК-конструкция по любому из пп.1-20, отличающаяся тем, что она вставлена в вектор.
22. ДНК-конструкция по п. 21, отличающаяся тем, что вектором является вирус.
23. ДНК-конструкция по п. 21, отличающаяся тем, что вирусом является ретровирус, аденовирус, адено-ассоциированный вирус, вирус простого герпеса или вирус осповакцины.
24. ДНК-конструкция по любому из пп.1-23, отличающаяся тем, что она вставлена в плазмиду.
25. ДНК-конструкция по любому из пп.21-24, отличающаяся тем, что она получена в коллоидной дисперсионной системе.
26. ДНК-конструкция по п.25, отличающаяся тем, что коллоидной дисперсионной системой являются липосомы.
27. ДНК-конструкция по п.25, отличающаяся тем, что коллоидной дисперсионной системой являются полилизионовые лиганды.
28. ДНК-конструкция по любому из пп.21-27, отличающаяся тем, что она дополнена лигандом, который связывается с мембранными структурами гладкомышечных клеток, активированных эндотелиальных клеток, активированных макрофагов или активированных лимфоцитов.
29. ДНК-конструкция по п. 21, отличающаяся тем, что лигандом является поликлональное или моноклональное антитело или фрагмент этого антитела, которое (который) с помощью своих изменяемых доменов специфически связывается с мембранной структурой гладкомышечных клеток, активированных эндотелиальных клеток, активированных макрофагов или активированных лимфоцитов; или является цитокин или фактор роста или его фрагмент, соответственно, его частичная последовательность, который (которая) связывается с рецептором гладкомышечных клеток, активированных эндотелиальных клеток, активированных макрофагов или активированных лимфоцитов; или лиганд представляет собой адгезионную молекулу, как SLeX, LFA-1, MAC-1, LECAM-1 или VLA-4.
30. ДНК-конструкция по п.29, отличающаяся тем, что мембранная структура рецептора представляет собой маннозу, интерлейкин-1 или фактор роста, как PDGF, FGF, VEGF, TGF-β.
31. ДНК-конструкция по п.29, отличающаяся тем, что мембранными структурами являются актин, ангиотензин, рецепторы интерлейкина, рецепторы EGF, рецепторы PDGF, рецепторы FGF или рецепторы эндотелина.
32. ДНК-конструкция по п.31, отличающаяся тем, что мембранные структуры представляют собой рецепторы цитокинов, факторов роста, интеферонов, хемокинов, в особенности рецепторы фактора стволовой клетки, интерлейкина-1, интерлейкина-2, интерлейкина-3, интерлейкина-4, интерлейкина-6, интерлейкина-8, интерлейкина-10, α-интерферона, β-интерферона, γ-интерферона, GM-CSF, M-CSF, G-CSF или α-фактора некроза опухоли.
33. ДНК-конструкция по любому из пп.1-32, применяемая в составе лекарственного средства для инъекции в ткань, как мышца, соединительная ткань, печень, почки, селезенка, легкие, кожа; для локального нанесения на кожу или слизистые оболочки; для инъекции в полости тела, как суставы, плевральная полость, субарахноидальное пространство; или для инъекции в систему кровеносных сосудов, как внутриартериальная или внутривенная инъекция.
34. ДНК-конструкция по любому из пп.1-32, применяемая для профилактики и лечения заболеваний кровеносных сосудов.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| GB9417366A GB9417366D0 (en) | 1994-08-26 | 1994-08-26 | Cell cycle regulated repressor and DNA element |
| GB9417366.3 | 1994-08-26 | ||
| GB9506466.3 | 1995-03-29 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| RU97104482A RU97104482A (ru) | 1999-05-10 |
| RU2197992C2 true RU2197992C2 (ru) | 2003-02-10 |
Family
ID=10760519
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU97104482/14A RU2197992C2 (ru) | 1994-08-26 | 1995-08-25 | Генная терапия заболеваний сосудов с помощью специфического к клетке и зависимого от клеточного цикла активного вещества |
Country Status (3)
| Country | Link |
|---|---|
| GB (1) | GB9417366D0 (ru) |
| RU (1) | RU2197992C2 (ru) |
| ZA (1) | ZA957074B (ru) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2378375C2 (ru) * | 2003-11-21 | 2010-01-10 | Асубио Фарма Ко., Лтд. | Способ пролиферации кардиомиоцитов |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2395136A1 (en) * | 1999-12-22 | 2001-06-28 | The Scripps Research Institute | Angiogenesis and vascular permeability modulators and inhibitors |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1993008845A1 (en) * | 1991-11-08 | 1993-05-13 | Massachusetts Institute Of Technology | Localized oligonucleotide therapy |
-
1994
- 1994-08-26 GB GB9417366A patent/GB9417366D0/en active Pending
-
1995
- 1995-08-23 ZA ZA9507074A patent/ZA957074B/xx unknown
- 1995-08-25 RU RU97104482/14A patent/RU2197992C2/ru not_active IP Right Cessation
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1993008845A1 (en) * | 1991-11-08 | 1993-05-13 | Massachusetts Institute Of Technology | Localized oligonucleotide therapy |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2378375C2 (ru) * | 2003-11-21 | 2010-01-10 | Асубио Фарма Ко., Лтд. | Способ пролиферации кардиомиоцитов |
Also Published As
| Publication number | Publication date |
|---|---|
| ZA957074B (en) | 1997-02-24 |
| GB9417366D0 (en) | 1994-10-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| PT777739E (pt) | Tratamento de doencas de vasos por meio de terapia genetica atraves de um material activo especifico de celulas e dependente do ciclo celular | |
| US6210939B1 (en) | Recombinant adenoviral vector and methods of use | |
| US20030018005A1 (en) | Gene therapy of diseases associated with the immune system, using a cell-specific active compound which is regulated by the cell cycle | |
| Varley et al. | Inflammation-induced recombinant protein expression in vivo using promoters from acute-phase protein genes. | |
| US6576758B1 (en) | Nucleic acid constructs containing hybrid promoters | |
| US6033856A (en) | Promoter of the cdc25B gene, its preparation and use | |
| AU716178B2 (en) | Nucleic acid constructs containing genes encoding transport signals | |
| Shiratori et al. | Strategy of liver‐directed gene therapy: present status and future prospects | |
| US20040266006A1 (en) | Adenoviral vectors having a protein IX deletion | |
| AU5155799A (en) | Expression system containing chimeric promoters with binding sites for recombinant transcription factors | |
| CZ376898A3 (cs) | Konstrukt nukleové kyseliny pro expresi řízenou onkogenem nebo virem, způsob přípravy takového konstruktu a jeho použití pro přípravu léčiva | |
| Ahmed et al. | In vivo tumor suppression by adenovirus-mediated interferon alpha2b gene delivery | |
| RU2197992C2 (ru) | Генная терапия заболеваний сосудов с помощью специфического к клетке и зависимого от клеточного цикла активного вещества | |
| EP2386319A1 (en) | Promoters exhibiting endothelial cell specificity and methods of using same | |
| WO1996006938A1 (de) | Gentherapeutische behandlung von gefässerkrankungen durch einen zellspezifischen, zellzyklusabhängigen wirkstoff | |
| US7482155B1 (en) | Chimeric promoters for controlling expression in smooth muscle cells | |
| AU2001294793B2 (en) | Viruses targeted to hypoxic cells and tissues | |
| US6743623B2 (en) | Viral recombinant vectors for expression in muscle cells | |
| JP2004509635A5 (ru) | ||
| US6099831A (en) | Viral recombinant vectors for expression in muscle cells | |
| Abaan et al. | Gene therapy in human breast cancer | |
| Ehsan et al. | Antisense and gene therapy to prevent restenosis | |
| Cavenee | Genetics of Human Astrocytic Tumors |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| MM4A | The patent is invalid due to non-payment of fees |
Effective date: 20030826 |