WO1997010703A1 - Conditionally male-fertile plants and methods and compositions for restoring the fertility thereof - Google Patents
Conditionally male-fertile plants and methods and compositions for restoring the fertility thereof Download PDFInfo
- Publication number
- WO1997010703A1 WO1997010703A1 PCT/US1996/015131 US9615131W WO9710703A1 WO 1997010703 A1 WO1997010703 A1 WO 1997010703A1 US 9615131 W US9615131 W US 9615131W WO 9710703 A1 WO9710703 A1 WO 9710703A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- plant
- male
- plants
- fertile
- acid
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Classifications
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01N—PRESERVATION OF BODIES OF HUMANS OR ANIMALS OR PLANTS OR PARTS THEREOF; BIOCIDES, e.g. AS DISINFECTANTS, AS PESTICIDES OR AS HERBICIDES; PEST REPELLANTS OR ATTRACTANTS; PLANT GROWTH REGULATORS
- A01N37/00—Biocides, pest repellants or attractants, or plant growth regulators containing organic compounds containing a carbon atom having three bonds to hetero atoms with at the most two bonds to halogen, e.g. carboxylic acids
- A01N37/42—Biocides, pest repellants or attractants, or plant growth regulators containing organic compounds containing a carbon atom having three bonds to hetero atoms with at the most two bonds to halogen, e.g. carboxylic acids containing within the same carbon skeleton a carboxylic group or a thio analogue, or a derivative thereof, and a carbon atom having only two bonds to hetero atoms with at the most one bond to halogen, e.g. keto-carboxylic acids
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01H—NEW PLANTS OR NON-TRANSGENIC PROCESSES FOR OBTAINING THEM; PLANT REPRODUCTION BY TISSUE CULTURE TECHNIQUES
- A01H1/00—Processes for modifying genotypes ; Plants characterised by associated natural traits
- A01H1/02—Methods or apparatus for hybridisation; Artificial pollination ; Fertility
- A01H1/026—Methods or apparatus for hybridisation; Artificial pollination ; Fertility by treatment with chemicals
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01N—PRESERVATION OF BODIES OF HUMANS OR ANIMALS OR PLANTS OR PARTS THEREOF; BIOCIDES, e.g. AS DISINFECTANTS, AS PESTICIDES OR AS HERBICIDES; PEST REPELLANTS OR ATTRACTANTS; PLANT GROWTH REGULATORS
- A01N37/00—Biocides, pest repellants or attractants, or plant growth regulators containing organic compounds containing a carbon atom having three bonds to hetero atoms with at the most two bonds to halogen, e.g. carboxylic acids
- A01N37/06—Unsaturated carboxylic acids or thio analogues thereof; Derivatives thereof
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01N—PRESERVATION OF BODIES OF HUMANS OR ANIMALS OR PLANTS OR PARTS THEREOF; BIOCIDES, e.g. AS DISINFECTANTS, AS PESTICIDES OR AS HERBICIDES; PEST REPELLANTS OR ATTRACTANTS; PLANT GROWTH REGULATORS
- A01N61/00—Biocides, pest repellants or attractants, or plant growth regulators containing substances of unknown or undetermined composition, e.g. substances characterised only by the mode of action
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
- C12N15/8243—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
- C12N15/8243—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine
- C12N15/8247—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine involving modified lipid metabolism, e.g. seed oil composition
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8287—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for fertility modification, e.g. apomixis
- C12N15/8289—Male sterility
Definitions
- the present invention is related to male sterility in plants and more particularly to compositions and methods for producing and/or identifying conditionally male-fertile plants and for restoring fertility to such conditionally male-fertile plants.
- male fertility in higher plants has tremendous practical importance to the breeding of crop and horticultural plants. Dominant, recessive, nuclear, and cytoplasmic male-sterile genetic traits are used in various breeding schemes. The most economically-important use of male sterility is in hybrid plant breeding. In most higher plant species, hybrid cultivars are generally superior to open-pollinated cultivars in yield or in other production-related characteristics. Hybrid plant breeding requires a functionally male-sterile plant as the female parent, but the hybrid must be fully fertile if seeds or fruit are the harvested crop. Furthermore, to take full advantage of heterosis, the female parent must come from an inbred line.
- a cytoplasmic male-sterile line and a normal-cytoplasm maintainer are used during in-breeding of the female parent.
- the male parental line in the breeding program contains a fertility-restoring allele (e.g., of a nuclear gene, Rf) that overcomes the cytoplasmic male-sterility trait to ensure that the hybrid is fertile.
- Rf a fertility-restoring allele
- a breeding scheme employing a cytoplasmic male-sterile parental line is complex, requiring identification of both a suitable cytoplasmic male-sterile trait and a suitable dominant fertility-restoring allele.
- the cytoplasmic male-sterile and maintainer lines must be carried together during the inbreeding phase.
- Nuclear-encoded male-sterile traits have not been widely used in hybrid breeding systems. In principle, recessive male-sterile traits can be maintained as heterozygotes during inbreeding with homozygous plants being chosen for crosses with the male parental line.
- Several strategies have been developed to assist this approach, but the difficulty of obtaining homozygous male-sterile plants for the production phase has limited the use of this approach in practice. These strategies are described in greater detail, e.g. , in Kaul, Male Sterility in Higher Plants, Springer-Verlag, Berlin, 1988, and U.S. Patents 4,654,465 and 4,727,219, which are incorporated herein by reference.
- the emasculation treatment or genotype must have no significant effect on female fertility, since this would compromise hybrid seed production.
- the female parent used must be free of any barriers to cross- pollination by the male parent.
- a cytoplasmic male-sterile trait must be completely recessive in the presence of the restorer and preferably does not confer an undesirable phenotype such as susceptibility to a plant pathogen.
- a nuclear male-sterile trait must also be completely recessive and free of undesirable pleiotropic effects.
- a complex system for the establishment of conditional male fertility involves incorporating into the genome of a plant a transgene the expression of which can be induced by administration of a chemical inducer.
- the transgenic plant is normally male sterile but is rendered male-fertile upon expression of the transgene.
- Such a system is described in U.S. Patent No. 5,432,068.
- FIG. 1 shows the relative growth rate of wild-type Arabidopsis and the fad3 fad7-2 fad ⁇ triple mutant. At intervals between 8 and 24 days after sowing, samples of plants were harvested and the fresh weight of the above- ground parts was measured. The relative growth rate ( ⁇ 1 ) for the wild-type (•) was 0.388 ⁇ 0.025 and that for the mutant (o) was 0.388 ⁇ 0.033.
- the conditionally male-fertile phenotype is caused by mutations that interfere with normal jasmonic acid metabolism.
- mutations can include, but are not limited to, mutations that: interfere with biosynthesis of jasmonic acid from ⁇ -linolenate (e.g., mutations in structural genes for enzymes in the jasmonate biosynthetic pathway) or tr ⁇ /w-acting regulators of such structural genes or their binding sites; receptors for jasmonate or its precursors; etc.
- the present invention is useful for identifying conditionally male- fertile plants arising naturally or as a result of various conventional mutagenesis techniques, including genetic engineering. This discovery also facilitates the production of additional conditionally male-fertile plant varieties for use, for example, in hybrid breeding programs.
- the ability to produce and/or identify conditionally male-fertile plants and to rescue the male-fertility of these plants by simply applying a chemical compound forms the basis for a conditional male-fertility system that is broadly applicable to hybrid breeding of crop and horticultural plants.
- compositions that include a conditionally male-fertile plant and an effective amount of jasmonate or a related compound, i.e., an amount of a composition comprising jasmonate or a related compound that is effective to restore male fertility to the plant when applied to the plant, preferably jasmonic acid, methyl jasmonate, or mixtures thereof.
- conditionally male-sterile plants may be jasmonate-deficient (including, for example, plants having at least one mutation at a FAD locus, but are not necessarily so (e.g., Arabidopsis line CS2338).
- Conditionally male-fertile plants may, for example, have one or more mutations of a FAD gene and/or one or more mutations in the locus that causes male- sterility in CS2338.
- Another aspect of the present invention encompasses methods of identifying a conditionally male-fertile plant by applying to a male-sterile plant an effective amount of a composition comprising jasmonic acid or a related compound and detecting whether male fertility is thereby restored to the plant.
- Such methods can be used to screen a population of male-sterile plants produced by mutagenizing fertile plants by conventional methods (e.g., by chemical mutagenesis, irradiation, gene disruption by mobile genetic elements, antisense expression, cosuppression, gene replacement, etc.).
- Another aspect of the present invention encompasses methods is the use of conditionally male-fertile plants for the production of hybrid plants in plant breeding programs.
- a conditionally male-fertile plant is made fertile by application of a composition that includes an effective amount of jasmonic acid or a related compound and the plant is self-fertilized to produce an inbred plant. Then, during the production phase, the inbred plant (which is allowed to exhibit the male-sterile phenotype) is crossed with a second plant to produce a hybrid plant.
- conditionally male-fertile refers to a plant having a dominant or recessive male-sterile trait (which is inherited in a Mendelian manner), whereby the fertility of the plant can be recovered by the administration of exogenous jasmonate or a related compound(s).
- an "effective amount" of a composition comprising jasmonate or a related compound(s) is an amount that, when applied to a conditionally male fertile plant as described herein, restores the fertility of the plant to a level acceptable for plant breeding purposes.
- conditionally male-fertile plants that are useful, for example, for hybrid breeding programs.
- conditionally male-fertile plants may be found in any plant species in which jasmonate or related compounds are required for pollen fertility including, but not limited to, rapeseed, canola and other Brassica species, soybean, wheat, barley, corn, sunflower, tomato, tobacco, cotton, and rice.
- male-sterile lines are, in fact, conditionally male fertile by determining whether the lines recover male fertility upon administration of jasmonate or a related compound.
- line CS23308 it is practical to grow progeny from a plant that is heterozygous for a particular male-sterile trait. Once homozygous (male-sterile) segregants have been identified among the progeny by their inability to set seed (or by other criteria), then these sterile individuals are treated with jasmonate or a related compound. If the plants produce seed after jasmonate treatment, but not after control treatments lacking jasmonate or a related compound, then it can be concluded that these plants come from a line that contains a conditionally male- fertile trait.
- conditionally male-fertile lines according to the invention are substantially deficient in jasmonic acid in their vegetative tissues as the result of a mutation affecting the biosynthesis of jasmonic acid from or-linolenate.
- Such mutants can be identified by conventional techniques for identifying a deficiency in jasmonic acid or the buildup of a precursor compound, e.g., by enzyme assays, radiotracer studies, gas chromatography (Creelman et al. , Proc. Natl. Acad. Sci. USA 89:4938-4941, 1992), high performance liquid chromatography, or other techniques.
- jasmonic acid and some related compounds are involved in the defense of plants from insect attack, it is possible to visually screen male-sterile plants for conditional male fertility on the basis of their greater susceptibility to insect attack and damage.
- a conditionally male-fertile fad3-2 fad.7-2 fad8 triple mutant of Arabidopsis thaliana and of the Arabidopsis line CS2338 displayed normal anther and pollen development up until the very last stages of pollen maturation and/or dehiscence of the anther locules.
- Visual inspection and other experimental procedures e.g., as described in Regan and Moffatt, Plant Cell 2:877-889, 1990
- conditionally male-fertile plants by conventional mutagenesis techniques.
- conditionally male-fertile plants are produced by mutations that eliminate or substantially reduce the expression of genes encoding, for example: enzymes for the biosynthesis of jasmonate from ⁇ -linolenate; polypeptides that are responsible for the regulation of such genes; receptors for precursors of jasmonate; polypeptides necessary for intra- or intercellular transport of jasmonate or related compounds or the intermediates or products of its biosynthesis or metabolism.
- genes include, but are not limited, to: FAD (fatty acid desaturation) genes (Somerville and Browse, Trends in Cell. Biol.
- Mutagenesis techniques useful for producing conditionally male-sterile plants include, but are not limited to, genetic approaches such as the use of ionizing radiation (e.g., irradiation with X-rays or gamma rays) or chemical mutagens, gene disruption by mobile genetic elements such as transposons, or genetic engineering techniques, include targeted gene disruption or gene replacement (mediated by homologous recombination or other means), antisense expression, or cosuppression of an appropriate gene, or other conventional techniques.
- genetic approaches such as the use of ionizing radiation (e.g., irradiation with X-rays or gamma rays) or chemical mutagens, gene disruption by mobile genetic elements such as transposons, or genetic engineering techniques, include targeted gene disruption or gene replacement (mediated by homologous recombination or other means), antisense expression, or cosuppression of an appropriate gene, or other conventional techniques.
- Targeted gene disruption will produce a recessive conditionally male- fertile trait similar to those identified from mutant screens.
- antisense expression or cosuppression will produce a dominant trait, causing Fl hybrids to express the conditionally male-fertile phenotype.
- This problem can be overcome in a breeding program by incorporating into the nuclear genome of the male line a second gene that is functionally equivalent to the suppressed gene but which has a DNA sequence that is sufficiently different from the suppressed gene to prevent its own suppression (e.g., homologs of the inactivated gene from the same plant species or from other organisms or genes that have been mutagenized in vitro to introduce silent or conservative mutations that do not substantially interfere with biological function).
- a fad3-2 fad7-2 fadS triple mutant of Arabidopsis thaliana in which ⁇ -linolenic acid was substantially eliminated in all tissues was found to be male-sterile but was otherwise apparently normal with regard to its growth rate and other characteristics, at least under laboratory conditions.
- both ⁇ -linolenic acid and jasmonic acid, a product of ⁇ -linolenic acid metabolism restored fertility to the triple-mutant plants.
- Metabolic and developmental processes are highly conserved among higher plants; many of the genes controlling such processes are also highly conserved in both sequence and function. Mutations in one or more genes encoding an enzyme in the biochemical pathway for jasmonic acid biosynthesis, including mutations analogous to those described herein for Arabidopsis thaliana, produce conditional male fertility in other plant species. Such genetic variants form the basis for successful hybrid breeding of these crops. Mutations in genes encoding regulators of the pathway for the biosynthesis or intracellular or intercellular transport of jasmonate or related compounds or the intermediates or products of its biosynthesis or metabolism also give rise to conditional male fertility.
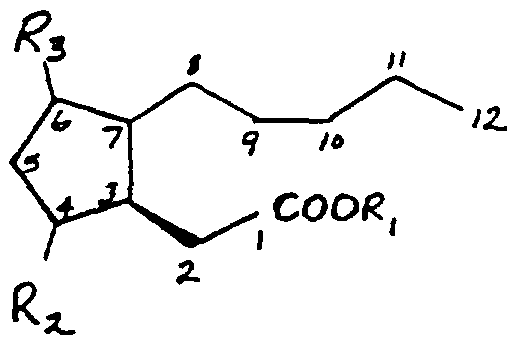
- related compound includes compounds that function like jasmonic acid in restoring the fertility of conditionally male-fertile plants, preferably compounds that are structurally related to jasmonic acid, including, but not limited to, compounds having the formula:
- R is H or a lower alkyl chain of 1 to 6 carbon atoms
- the compound is optionally single or double bonded at one or more of C ⁇ C ⁇ C 3 :C , C 4 :C 5 , C 5 :C 6 , C 6 :C 7 , C 9 :C 10 , or C u :Cj 2 , and where additional -OH groups may be optionally located at one or more of C g , C n , or C ⁇ .
- Representative compounds that are structurally related to jasmonate include, but are not limited to: cucurbic acid 7-wo-jasmonic acid,
- Preferred related compounds include jasmonic acid [(3R,7S)- jasmonic acid], the methyl ester of jasmonic acid ("methyl jasmonate”), and mixtures thereof, including racemic mixtures containing related enantiomers and/ or diastereomers.
- metabolic precursors of jasmonic acid and related compounds including, but not limited to, ⁇ -linolenic acid (9Z, 12Z, 15Z octadecatrienoic acid), 13(S)-hydroperoxy linolenic acid (13-hydroperoxy-9Z, 1 IE, 15Z-octadecatrienoic acid), allene oxide (12,13(S)-epoxy-9Z,ll,15Z-octadecatrienoic acid),
- 12-oxo-phytodienoic acid (8-[2- .s-2'-pentenyl)- 3-oxo-cyclopent-4-enyl]octanoic acid) (Blechert et al. , Proc. Natl. Acad. Sci. USA 92:4099-4105, 1995), (lS,2S)3-oxo-2-(2'-pentenyl)cyclopentane-octanoic acid, (lS,2S)3-oxo-2-(2'- pentenyl)cyclopentanehexanoic acid, (lS,2S)3-oxo-2-(2'- pentenyl)cyclopentanetetranoic acid.
- Structurally-related compounds may act directly as fertility-restoring agents or may be metabolized in the plant to fertility restoring agents.
- Structurally-related compounds also include conjugates of these compounds with other moieties that do not interfere with fertility-restoring activity, including, for example, mono- or polysaccharides, amino acids, or polypeptides.
- “Related compounds” also include other compounds that restore at least partial fertility to conditionally male-fertile plants, e.g., compounds that interact with a jasmonic acid receptor, such as coronatine and coronofacic acid, or that affect other molecules involved in the jasmonate signaling pathway, including proteinase inhibitors such as bestatin (Schaller et al. , Plant Cell 7:1893-1898, 1995).
- a jasmonic acid receptor such as coronatine and coronofacic acid
- proteinase inhibitors such as bestatin (Schaller et al. , Plant Cell 7:1893-1898, 1995).
- Admimstration of Compositions That Include Jasmonate or Related Compound(s) Jasmonate acid and related compounds may be administered to a plant by any conventional means, including direct application to the flowers or other tissues of the plant, e.g. , by painting or spraying on the flowers or other plant tissues a composition that includes jasmonate or related compound(s) or a mixture thereof.
- a composition that includes jasmonate or a related compound may be painted or sprayed onto the flower, flower bud, or other plant tissue, for example.
- the composition may be a solution of jasmonate or a related compound in water (or another suitable non-phytotoxic solvent such as glycerol) and may optionally include a wetting or sticking agent, e.g. , a low concentration of a suitable surfactant.
- the composition may be applied directly to a flower or other plant tissue by any conventional means, such as by painting, misting, spraying, drenching, etc.
- Jasmonic acid or a related compound can also be "applied" to plant tissue via airborne transmission in which volatile source of the inducing agent is placed in the vicinity of the plant tissue and the agent is allowed to diffuse or disperse through the atmosphere to contact the plant tissue.
- the volatile source may be, for example, plant materials which naturally produce the inducing agent or a volatile solution of the agent, such as a solution comprising a volatile solvent and the inducing agent.
- Suitable solvents for this purpose include organic solvents, e.g. , alcohols (e.g., methanol and ethanol), or water.
- the manner in which the volatile source of the inducing agent is placed in the vicinity of the plant tissue to be treated is not critical to the practice of the invention.
- the inducing agent may be placed in an open container in the vicinity of the plant tissue, on a matrix support, such as a fiber matrix, in the vicinity of the plant tissue.
- Amounts of jasmonic acid or related compounds effective to induce fertility in treated plants of a given male-sterile line depend on many factors, including the nature, environment and condition of the plants to be treated, the method of contact of the inducing agent with plant tissue to be treated and other factors.
- a solution of jasmonic acid or a related compound for direct application comprises from about 1 pg/ml to about 100 mg/ml of the inducing agent, more preferably from about 1 ng/ml to about 10 mg/ml of the inducing agent, and most preferably from about 1 ⁇ g/ml to about 1 mg/ml of the inducing agent.
- a higher jasmonate concentration will be required to induce fertility if the jasmonate is sprayed onto a plant tissue rather than painted on the tissue.
- Jasmonic acid and related compounds induce compounds that act in the defense of plants from insect attack. It is possible that conditionally male- fertile plants that are rendered fertile by the administration of exogenous jasmonic acid or related compounds will be more susceptible to insect damage (Farmer and Ryan, Proc. Natl. Acad. Sci. USA 87:7713-7716, 1990). Protection against insect damage can be provided through the use of insecticidal treatments. Application of jasmonic acid or related compounds may itself reduce insect damage, as described in WO 91/18512, which is incorporated herein by reference.
- the biophysical reactions of light harvesting and electron transport during photosynthesis take place in a uniquely constructed bilayer membrane, the thylakoid.
- the complement of atypical glycerolipid molecules that form the foundation of this membrane is characterized by sugar headgroups and a very high level of unsaturation in the fatty acid chains that compose the central portion of the thylakoid lamella bilayer.
- monogalactosyldiacyl-glycerol the major thylakoid lipid
- monogalactosyldiacyl-glycerol typically contains more than 90% of ⁇ -linolenic acid (18:3) or a combination of 18:3 and hexadecatrienoic (16:3) acids, depending on the plant species (Jamieson and Reid, Phytochemistry 10: 1837-1843, 1971).
- Both pathways are initiated by the synthesis of 16:0-ACP and 18: 1-ACP by the combined action of a Type II fatty acid synthase and a soluble stearoyl-ACP desaturase located in the chloroplasts or other plastids.
- the "prokaryotic pathway" located in the chloroplast inner envelope uses 18:1-ACP and 16:0-ACP for the sequential acylation of glycerol-3-phosphate and synthesis of glycerolipid components for the chloroplast membranes.
- the "eukaryotic pathway” involves export of 16:0 and 18: 1 fatty acids from the chloroplast to the endoplasmic reticulum and their incorporation into phosphatidylcholine and other phospholipids that are the principal structural lipids of all the membranes of the cell except for the chloroplast.
- the diacylglycerol moiety of phosphatidylcholine can be returned to the chloroplast envelope and used as a second source of precursors for the synthesis of chloroplast glycerolipids.
- the FAD 3 gene product localized predominantly in the endoplasmic reticulum, utilizes 18:2 on phosphatidylcholine as its major substrate although it is possible that it also acts on 18:2 groups of other phospholipids (Browse et al., J. Biol. Chem. 268: 16345-16351, 1993).
- fad7-2fad8 double mutant plants contain no 16:3 but approximately 17% 18:3 in their chloroplast membranes (McConn et al. , Plant Physiol. 106: 1609-1614, 1994).
- the extrachloroplast membranes in leaves of fad3 mutant plants contain considerable 18:3 because 18:2 lipid can be transferred to the chloroplast on the eukaryotic pathway, desaturated by the FAD7 and FAD8 enzymes, and then returned to the endoplasmic reticulum and other extrachloroplast membranes (Browse et al. , J. Biol. Chem. 268:16345-16351, 1993).
- the F 2 progeny from these crosses were screened by gas chromatographic analysis and fad7-2 fad8 and fad.3-2 fad.7-2 double mutant plants were identified.
- To screen the leaf fatty acid compositions of individual F 2 progeny samples of leaf tissue were rapidly immersed in liquid nitrogen, ground to a fine powder, extracted and analyzed for their content of trienoic fatty acids using gas chromatography essentially as described by Miquel and Browse (/. Biol. Chem. 267: 1502-1509, 1992).
- the two double mutant plants were crossed, and an F, plant derived from a cross between the two double mutant plants was allowed to self -pollinate. The resulting seeds were germinated.
- the leaf fatty acid compositions of individual F 2 progeny were examined for their content of trienoic fatty acids. Of 240 F 2 plants analyzed, 17 contained no detectable 16:3 or 18:3 (detection limit approximately 0.1 % of total) while the remaining plants exhibited a range in the proportion of total trienoic fatty acids from 10% to 40% (Table 1).
- triple-mutant plants were identified by fatty acid analysis from among the progeny of plants that contained just one wild-type allele at the fad8 locus, that is, fad3-2(- 1 -) fad7-2(- 1 -) fad8(+ 1 -).
- wild-type plants began to bolt about three weeks after sowing of the seed and flowers are produced continuously for 4-6 weeks thereafter until the plant became senescent.
- the growth rate of linolenate-deficient triple-mutant plants was indistinguishable from the wild-type plant during vegetative development at this temperature (FIG. 1) and triple-mutant and wild-type plants displayed a similar timing for bolting and the start of flowering.
- EXAMPLE 2 The Arabidopsis fa/3-2 faal-2 fadS Triple Mutant is Male Sterile
- anthers from wild-type flowers it was always possible to pollinate the mutant and produce mature seeds. In contrast, anthers from mutant flowers were unable to induce seed set on emasculated wild- type flowers. Closer examination of wild-type and mutant flowers revealed that the locules of the mutant anthers had not dehisced to deposit pollen on the stigmatic surface. Manual disruption of the anther locules to release the enclosed pollen did not result in any seed set.
- the samples were transferred to a Samdri-PVT-3D drying apparatus and critical point dried in CO 2 (Boyde, Scanning Electron Microscopy 2:5945-5951 , 1978; Cohen, Scanning Electron Microscopy 2:303-323, 1979).
- the specimens were then affixed to stubs with paraffin, coated with gold in argon with a Technics Hummer V sputtering device (Echlin, Scanning Electron Microscopy 1:79-80, 1981) and viewed on a Hitachi S-570 scanning electron microscope. Two independent preparations were examined.
- triple-mutant progeny were produced in a Mendelian ratio from eit er fad3-2(+ 1 -) fad7-2(+ 1 -) fad8 (+/-) or fad3-2(-l-) fad7-2(-l-)fad8 (+/-) (where " + " indicates a wild-type allele and "-” indicates a mutant allele at each locus in the diploid genome) parents indicates that the genotype of the maternal tissue (rather than the genotype of the segregating, haploid microspores) mediates the male-sterile phenotype and that very low levels of 18:3 are probably sufficient to ensure fertility.
- flowering triple-mutant plants were grown under different environmental conditions without restoring fertility to them. These conditions included: light intensity ranging from 100-150 and 300 ⁇ mole quanta/m 2 s; photoperiods including continuous light; 12 hr light, 12 hr darkness; and 10 hr light, 14 hr darkness; and a variety of temperatures from 5°C to 22 °C.
- the male sterility of the fad3-2 fad7-2 fad8 triple-mutant plants persisted under a range of environmental conditions. From a total of 15,000 flowers on more than 150 untreated triple- mutant plants, not a single seed of the parental genotype fad3-2 fad7-2 fad8 was ever recovered.
- EXAMPLE 3 Germination and Viability of Pollen from the Triple Mutant Pollen viability was assessed by double staining pollen grains with fluorescein diacetate and propidium iodide essentially as described by Heslop-Harrison and Heslop-Harrison (Stain Technol. 45: 115-120, 1970) and by Regan and Moffatt (Plant Cell 2:877-889, 1990). Equal amounts of fluorescein diacetate and propidium iodide solutions were added to freshly isolated pollen. The pollen was transferred to a glass slide, covered with a coverslip, and viewed under ultraviolet light using filter block 13 with excitation filters BP450-490, dichromatic mirror RKP510 and suppression filter LP520.
- the pollen was transferred to a glass slide, covered with a coverslip, and viewed under ultraviolet light using filter block 13 (Leitz) with excitation filters BP450-490, dichromatic mirror RKP510 and suppression filter LP520 (all from Leitz).
- Fluorescein diacetate a vital stain, is taken up by living cells and converted to impermeant fluorescein, which emits a green fluorescence under ultraviolet light (Heslop-Harrison and Heslop-Harrison, Stain Technology 45:115-120, 1970).
- Propidium iodide is excluded from living cells but labels dead cells with a red-orange fluorescence under ultraviolet light (Regan and Moffatt, Plant Cell 2:877-889, 1990).
- Wild-type and triple-mutant plants were grown side-by-side in four separate experiments. Pollen was isolated from mature flowers by gently releasing them from the anther locules into 17% (w/v) sucrose. The liberated pollen was then placed onto plates of pollen germination medium, consisting of 17% (w/v) sucrose, 2 mM CaCl 2 , 1.65 mM H 3 BO 3 at pH 7 (Preuss et al., Genes and Development 7:974-985, 1993) and solidified with 6% (w/v) agar. Pollen was incubated for 12 hours at room temperature then analyzed for pollen tube formation.
- Pollen development in the mutant followed a course that was very similar to that of wild-type pollen.
- Pollen mother cells undergoing the first division of meiosis formed haploid progeny which, following the second meiotic division, become organized into tetrads of microspores encased in a callose wall.
- the individual microspores were released by the action of a callase enzyme secreted by cells of the parental tissues.
- the layer of specialized tapetal cells that provides many of the nutrients and other factors (including the extracellular callase) required for pollen development persisted until late in development.
- DAPI stain 4',6-diamidino-2- phenylindole which binds specifically to double-stranded DNA, was used to stain pollen from wild ⁇ type and mutant plants that had been removed from anthers at stages immediately before or immediately after flower opening. DAPI staining was performed following the procedure of Coleman and Goff (Stain Technology 60: 145-154, 1985).
- the pollen was double stained with fluorescein diacetate and propidium iodide to show viable (blue-green) and dead (red-orange) pollen grains.
- D API- stained pollen was viewed with ultraviolet light using filter block A, with excitation filters BP340-380 dichromatic mirror RKP400 and suppression filter LP430 (all from Leitz). (Staining for DNA with mithramycin in the same manner provided similar results.)
- the male-sterility of the mutant was evident by the lack of any such enlarged siliques.
- One pot of wild-type and one pot of mutant plants were sprayed with 2 ml of 0.1 % solution of ⁇ -linolenic acid sodium soap each day for 10 consecutive days, during which time the plants were kept on a regime of 12 hours light/ 12 hours dark (at 22°C, 140 ⁇ mole quanta/m 2 /s during the lighted portion of the cycle) and sprayed at the start of the darkened portion of the cycle.
- ⁇ -linolenate content (as a proportion of the total fatty acids) in leaves, roots, and flower tissues (sepals, petals, carpels, and anthers) was determined by gas chromatography as described above after derivatization with 2.5% (v/v) H 2 SO 4 in methanol (Miquel and Browse, J. Biol. Chem. 267: 1502-1509, 1992).
- the results for flowers of fad3-2 fad.7-2 fad8 triple-mutant plants treated in this way and for untreated mutant and wild- type flowers are shown in Table 2. As expected, wild-type plants produced seeds while untreated mutant plants were sterile.
- the octadecanoid signalling compounds jasmonic acid and methyl jasmonate (available from Bedoukian Research Inc. , Danbury, CT), activate wound responses in plants (Farmer and Ryan, Proc. Natl. Acad. Sci. USA 87:7713-7716, 1990) and have been postulated to perform roles in several other developmental and environmental response processes (Sembdner and Parthier, Ann. Rev. Plant Physiol. Plant Mol. Biol. 44:569-589, 1993).
- the structure and biosynthesis of jasmonic acid have encouraged plant biologists because of parallels to eicosanoid second messengers that are central to inflammatory responses and other physiological processes in mammals (Creelman et al., Proc. Natl. Acad. Sci. -75-4 89:4938-4941 , 1992).
- Jasmonic acid in plants is synthesized from ⁇ -linolenic acid (9Z, 12Z, 15Z octadecatrienoic acid), a component of plant cell membranes (which is presumably released from membrane lipids by the action of a phospholipase A 2 ) by a pathway that is initiated by lipoxygenase. Cyclization and 3-oxidation of the lipoxygenase product, 13 (S)-hydroperoxy linolenic acid, result in the formation of jasmonic acid, which has a structure analogous in some respects to the prostaglandin E series of eicosanoids (Vick and Zimmerman, Plant Physiol. 75:458-461, 1984).
- 13(S)-Hydroperoxy linolenic acid may also give rise to other compounds, including products derived from a hydroperoxide lyase reaction sequence: 3Z-hexen-l-ol, 2E-hexen-l-al, 3E-hexen-l-ol, and traumatic acid (Croft et al., Plant Physiol. 101: 13-24, 1993).
- the male-sterile phenotype in the triple mutant is controlled by the genotype of the sporophytic tissue. This is true of the majority of male-sterile mutants and the defects are often localized by direct or circumstantial evidence to processes occurring in the tapetum, the anther cell layer that bounds the locule and is intimately involved in all aspects of pollen microspore development (Chapman, Int'l. Rev. Cytology 107:111-125, 1987; Goldberg et al. , Plant Cell 5:1217-1229, 1993). It would seem reasonable to implicate the tapetum as the physiological source of jasmonic acid, thereby regulating final maturation processes in pollen grains.
- methyl jasmonate which is normally present in the plants
- methyl jasmonate which is normally present in the plants
- specific translocation of jasmonic acid or its derivatives in the vascular system may be involved.
- a second component of male-sterility in the triple mutant is the failure of the anther locules to dehisce correctly. In some male-sterile mutants, the pollen is fully viable and the failure of pollination and fertilization is attributable solely to the fact that anther dehiscence does not occur (Dawson et al. , Can. J. Bot. 71:629-638, 1993).
- jasmonic acid performs (at least) two separate signalling functions during flower development, first, to ensure the maturation of viable pollen, and second, to orchestrate the changes in cell wall structure of the stomium and in the cellular water relations within the endothecium that result in successful dehiscence of the anthers (Stanley and Linskens, Pollen Biology,
- ⁇ -linolenic acid could restore fertility to flowers without penetrating the flower bud to increase the proportion of ⁇ -linolenic acid in the anthers to measurable levels (detection limit approximately 0.1 % of total fatty acids) suggests that jasmonic acid may be produced in other flower organs on mutant plants and then diffuse or be transported to anthers.
- the male-sterile phenotype of line CS2338 included morphological features in common with that of the fad3-2 fad7-2 fad8 triple mutant. Floral organs developed normally and the anther locules were not dehisced in flowers that had developed for two days after the petals appeared at the tip of the bud. When the anther locules were disrupted, the pollen that was released was not capable of fertilizing emasculated wild-type flowers. These results indicated that line CS2338 was indeed segregating for a male-sterile trait. Seed was collected separately from several individual fertile segregants of line CS2338 to form a series of sublines. Seeds from each subline were germinated and the plants grown to maturity.
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Wood Science & Technology (AREA)
- Chemical & Material Sciences (AREA)
- Zoology (AREA)
- General Health & Medical Sciences (AREA)
- Biotechnology (AREA)
- Plant Pathology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Organic Chemistry (AREA)
- Molecular Biology (AREA)
- General Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Environmental Sciences (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- Cell Biology (AREA)
- Dentistry (AREA)
- Microbiology (AREA)
- Pest Control & Pesticides (AREA)
- Biochemistry (AREA)
- Agronomy & Crop Science (AREA)
- Nutrition Science (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Botany (AREA)
- Developmental Biology & Embryology (AREA)
- Oil, Petroleum & Natural Gas (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Agricultural Chemicals And Associated Chemicals (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Nitrogen Condensed Heterocyclic Rings (AREA)
- Polysaccharides And Polysaccharide Derivatives (AREA)
Abstract
Description

Claims



Priority Applications (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| BR9610756A BR9610756A (en) | 1995-09-21 | 1996-09-20 | Conditionally fertile male vegetables and methods and compositions for restoring their fertility |
| JP9512927A JPH11511474A (en) | 1995-09-21 | 1996-09-20 | Conditional male fertile plants and methods and compositions for restoring their fertility |
| AU73670/96A AU706527B2 (en) | 1995-09-21 | 1996-09-20 | Conditionally male-fertile plants and methods and compositions for restoring the fertility thereof |
| NZ320845A NZ320845A (en) | 1995-09-21 | 1996-09-20 | Conditionally male-fertile plants and methods and compositions for restoring the fertility thereof |
| EP96935888A EP0868119A4 (en) | 1995-09-21 | 1996-09-20 | Conditionally male-fertile plants and methods and compositions for restoring the fertility thereof |
| MXPA/A/1998/002247A MXPA98002247A (en) | 1995-09-21 | 1998-03-19 | Conditionally androphertile plants and methods and compositions to restore the fertility of mis |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US421495P | 1995-09-21 | 1995-09-21 | |
| US60/004,214 | 1995-09-21 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO1997010703A1 true WO1997010703A1 (en) | 1997-03-27 |
Family
ID=21709717
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US1996/015131 Ceased WO1997010703A1 (en) | 1995-09-21 | 1996-09-20 | Conditionally male-fertile plants and methods and compositions for restoring the fertility thereof |
Country Status (8)
| Country | Link |
|---|---|
| EP (1) | EP0868119A4 (en) |
| JP (1) | JPH11511474A (en) |
| CN (1) | CN1196655A (en) |
| AU (1) | AU706527B2 (en) |
| BR (1) | BR9610756A (en) |
| CA (1) | CA2232623A1 (en) |
| NZ (1) | NZ320845A (en) |
| WO (1) | WO1997010703A1 (en) |
Cited By (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1999023233A1 (en) * | 1997-10-30 | 1999-05-14 | Mogen International N.V. | Nuclear male sterile plants, method of producing same and methods to restore fertility |
| WO1999045122A1 (en) * | 1998-03-06 | 1999-09-10 | Metabolix, Inc. | Modification of fatty acid metabolism in plants |
| US6586658B1 (en) | 1998-03-06 | 2003-07-01 | Metabolix, Inc. | Modification of fatty acid metabolism in plants |
| WO2004062350A3 (en) * | 2003-01-13 | 2004-12-29 | Florisys Inc | Methods and compositions for producing male sterile plants |
| US6890525B2 (en) | 1999-12-10 | 2005-05-10 | Plant Bioscience Limited | Semiochemical |
| WO2010081005A1 (en) * | 2009-01-08 | 2010-07-15 | Yale University | Method for controlling flower development in plants |
| EP2781151A1 (en) | 2013-03-18 | 2014-09-24 | Bayer CropScience AG | Methods of separating hybrid seed from a mixture of seeds |
Families Citing this family (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US8603502B2 (en) | 2002-02-04 | 2013-12-10 | L'oreal S.A. | Compositions comprising jasmonic acid derivatives and use of these derivatives |
| CN112662684B (en) * | 2020-05-31 | 2022-10-11 | 华中农业大学 | Method for improving male fertility of cotton at high temperature |
| CN113261464A (en) * | 2021-05-25 | 2021-08-17 | 中国农业科学院郑州果树研究所 | Method for inducing fruitage of male flowers of delicious kiwi fruits |
| CN114793792B (en) * | 2022-04-24 | 2023-06-20 | 河南科技学院 | A method for restoring pollen fertility of different fertility populations of BNS series thermosensitive genic male sterile wheat |
Citations (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4654465A (en) | 1985-07-18 | 1987-03-31 | Agracetus | Genic male-sterile maize |
| US4686319A (en) * | 1985-09-25 | 1987-08-11 | Rutgers, The State University Of New Jersey | Synthesis of genetic females and their use in hybrid seed production |
| US4727219A (en) | 1986-11-28 | 1988-02-23 | Agracetus | Genic male-sterile maize using a linked marker gene |
| US4954158A (en) * | 1983-08-16 | 1990-09-04 | University Of Georgia Research Foundation, Inc. | 2,3-methanoproline |
| WO1991018512A1 (en) | 1990-05-25 | 1991-12-12 | Washington State University Research Foundation, Inc. | Method of inducing plant defense mechanisms |
| WO1993018142A1 (en) | 1992-03-09 | 1993-09-16 | Washington State University Research Foundation | Methods for the regulation of plant fertility |
| US5356799A (en) | 1988-02-03 | 1994-10-18 | Pioneer Hi-Bred International, Inc. | Antisense gene systems of pollination control for hybrid seed production |
| US5432068A (en) | 1990-06-12 | 1995-07-11 | Pioneer Hi-Bred International, Inc. | Control of male fertility using externally inducible promoter sequences |
| US5436386A (en) | 1993-10-05 | 1995-07-25 | Seedtec International Inc. | Hybrid safflower production utilizing genetic dwarf male sterility |
-
1996
- 1996-09-20 AU AU73670/96A patent/AU706527B2/en not_active Ceased
- 1996-09-20 JP JP9512927A patent/JPH11511474A/en active Pending
- 1996-09-20 EP EP96935888A patent/EP0868119A4/en not_active Ceased
- 1996-09-20 NZ NZ320845A patent/NZ320845A/en unknown
- 1996-09-20 CA CA002232623A patent/CA2232623A1/en not_active Abandoned
- 1996-09-20 CN CN96197098.7A patent/CN1196655A/en active Pending
- 1996-09-20 BR BR9610756A patent/BR9610756A/en not_active Application Discontinuation
- 1996-09-20 WO PCT/US1996/015131 patent/WO1997010703A1/en not_active Ceased
Patent Citations (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4954158A (en) * | 1983-08-16 | 1990-09-04 | University Of Georgia Research Foundation, Inc. | 2,3-methanoproline |
| US4654465A (en) | 1985-07-18 | 1987-03-31 | Agracetus | Genic male-sterile maize |
| US4686319A (en) * | 1985-09-25 | 1987-08-11 | Rutgers, The State University Of New Jersey | Synthesis of genetic females and their use in hybrid seed production |
| US4727219A (en) | 1986-11-28 | 1988-02-23 | Agracetus | Genic male-sterile maize using a linked marker gene |
| US5356799A (en) | 1988-02-03 | 1994-10-18 | Pioneer Hi-Bred International, Inc. | Antisense gene systems of pollination control for hybrid seed production |
| WO1991018512A1 (en) | 1990-05-25 | 1991-12-12 | Washington State University Research Foundation, Inc. | Method of inducing plant defense mechanisms |
| US5432068A (en) | 1990-06-12 | 1995-07-11 | Pioneer Hi-Bred International, Inc. | Control of male fertility using externally inducible promoter sequences |
| WO1993018142A1 (en) | 1992-03-09 | 1993-09-16 | Washington State University Research Foundation | Methods for the regulation of plant fertility |
| US5436386A (en) | 1993-10-05 | 1995-07-25 | Seedtec International Inc. | Hybrid safflower production utilizing genetic dwarf male sterility |
Non-Patent Citations (4)
| Title |
|---|
| PHYTOCHEMISTRY, February 1995, Vol. 38, No. 3, KNOFEL et al., "Jasmonates From Pine Pollen", pages 569-571. * |
| PLANT BREEDING REVIEWS, 1985, Vol. 3, MICRAE, "Advances in Chemical Hybridization", pages 169-191. * |
| PLANT CELL, May 1994, Vol. 6, FEYS et al., "Arabidopsis Mutants Selected for Resistance to the Phytotoxin Coronatine Are Male Sterile, Insensitive to Methyl Jasmonate and Resistant to a Bacterial Pathogen", pages 751-759. * |
| See also references of EP0868119A4 |
Cited By (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1999023233A1 (en) * | 1997-10-30 | 1999-05-14 | Mogen International N.V. | Nuclear male sterile plants, method of producing same and methods to restore fertility |
| US6603064B1 (en) | 1997-10-30 | 2003-08-05 | Syngenta Mogen B.V. | Nuclear male sterile plants, method of producing same and methods to restore fertility |
| WO1999045122A1 (en) * | 1998-03-06 | 1999-09-10 | Metabolix, Inc. | Modification of fatty acid metabolism in plants |
| US6586658B1 (en) | 1998-03-06 | 2003-07-01 | Metabolix, Inc. | Modification of fatty acid metabolism in plants |
| US6890525B2 (en) | 1999-12-10 | 2005-05-10 | Plant Bioscience Limited | Semiochemical |
| US7820153B2 (en) | 1999-12-10 | 2010-10-26 | Plant Bioscience Limited | Semiochemical |
| US8221736B2 (en) | 1999-12-10 | 2012-07-17 | Rothamsted Research Limited | Semiochemical |
| WO2004062350A3 (en) * | 2003-01-13 | 2004-12-29 | Florisys Inc | Methods and compositions for producing male sterile plants |
| WO2010081005A1 (en) * | 2009-01-08 | 2010-07-15 | Yale University | Method for controlling flower development in plants |
| US20120010077A1 (en) * | 2009-01-08 | 2012-01-12 | Yale University | Methods for Controlling Flower Development in Plants |
| US9258998B2 (en) | 2009-01-08 | 2016-02-16 | Yale University | Methods for controlling flower development in plants |
| EP2781151A1 (en) | 2013-03-18 | 2014-09-24 | Bayer CropScience AG | Methods of separating hybrid seed from a mixture of seeds |
Also Published As
| Publication number | Publication date |
|---|---|
| MX9802247A (en) | 1998-08-30 |
| EP0868119A4 (en) | 2000-08-09 |
| CN1196655A (en) | 1998-10-21 |
| NZ320845A (en) | 1998-10-28 |
| AU7367096A (en) | 1997-04-09 |
| CA2232623A1 (en) | 1997-03-27 |
| EP0868119A1 (en) | 1998-10-07 |
| JPH11511474A (en) | 1999-10-05 |
| AU706527B2 (en) | 1999-06-17 |
| BR9610756A (en) | 1999-07-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| McConn et al. | The critical requirement for linolenic acid is pollen development, not photosynthesis, in an Arabidopsis mutant. | |
| US6297426B1 (en) | Methods of mediating female fertility in plants | |
| Ishiguro et al. | The DEFECTIVE IN ANTHER DEHISCENCE1 gene encodes a novel phospholipase A1 catalyzing the initial step of jasmonic acid biosynthesis, which synchronizes pollen maturation, anther dehiscence, and flower opening in Arabidopsis | |
| Footitt et al. | The COMATOSE ATP-binding cassette transporter is required for full fertility in Arabidopsis | |
| US5432068A (en) | Control of male fertility using externally inducible promoter sequences | |
| US5850014A (en) | Nucleotide sequences mediating ferility and method of using same | |
| US5824524A (en) | Nucleotide sequences mediating fertility and method of using same | |
| US5162602A (en) | Corn plants tolerant to sethoxydim and haloxyfop herbicides | |
| Dawson et al. | Characterization and genetic mapping of a mutation (ms35) which prevents anther dehiscence in Arabidopsis thaliana by affecting secondary wall thickening in the endothecium | |
| Browse | The power of mutants for investigating jasmonate biosynthesis and signaling | |
| US5290696A (en) | Method for imparting cyclohexanedione and/or aryloxyphenoxypropanioc acid herbicide tolerance to maize plants | |
| AU706527B2 (en) | Conditionally male-fertile plants and methods and compositions for restoring the fertility thereof | |
| Lung et al. | Deciphering the roles of acyl-CoA-binding proteins in plant cells | |
| EP0630403A4 (en) | METHOD FOR REGULATING THE FERTILITY OF PLANTS. | |
| Dai et al. | Generating novel male sterile tomatoes by editing respiratory burst oxidase homolog genes | |
| US5428001A (en) | Method for combating weeds among maize plants | |
| Damayanti et al. | Functional disruption of the tomato putative ortholog of HAWAIIAN SKIRT results in facultative parthenocarpy, reduced fertility and leaf morphological defects | |
| Cross et al. | Chemical agents that inhibit pollen development: tools for research | |
| MXPA98002247A (en) | Conditionally androphertile plants and methods and compositions to restore the fertility of mis | |
| McConn | Mutants of Arabidopsis deficient in polyunsaturated fatty acids | |
| Koltunow et al. | Paternal transmission of a seed size reduction gene varies with age of a primary transformant and seed set is also influenced by gene expression in maternal tissues | |
| Sanderson | Genetic and phenotypic characterisation of an Arabidopsis thaliana jasmonic acid signalling mutant, jam2 | |
| Ribarits et al. | Dominant-negative inhibition of cytoplasmic glutamine synthetase in developing anthers and pollen leads to reversible male sterility | |
| Tiryaki | Characterization of a new jasmonate signaling mutant in Arabidopsis | |
| Jewell | Jasmonate metabolism and signaling in abiotic stress and male reproductive development |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WWE | Wipo information: entry into national phase |
Ref document number: 96197098.7 Country of ref document: CN |
|
| AK | Designated states |
Kind code of ref document: A1 Designated state(s): AL AM AT AU AZ BB BG BR BY CA CH CN CZ DE DK EE ES FI GB GE HU IL IS JP KE KG KP KR KZ LK LR LS LT LU LV MD MG MK MN MW MX NO NZ PL PT RO RU SD SE SG SI SK TJ TM TR TT UA UG UZ VN AM AZ BY KG KZ MD RU TJ TM |
|
| AL | Designated countries for regional patents |
Kind code of ref document: A1 Designated state(s): KE LS MW SD SZ UG AT BE CH DE DK ES FI FR GB GR IE IT LU MC NL PT SE BF BJ CF CG CI CM GA GN |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application | ||
| DFPE | Request for preliminary examination filed prior to expiration of 19th month from priority date (pct application filed before 20040101) | ||
| ENP | Entry into the national phase |
Ref document number: 2232623 Country of ref document: CA Ref document number: 2232623 Country of ref document: CA Kind code of ref document: A |
|
| WWE | Wipo information: entry into national phase |
Ref document number: PA/a/1998/002247 Country of ref document: MX |
|
| ENP | Entry into the national phase |
Ref document number: 1997 512927 Country of ref document: JP Kind code of ref document: A |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 320845 Country of ref document: NZ |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 1996935888 Country of ref document: EP |
|
| REG | Reference to national code |
Ref country code: DE Ref legal event code: 8642 |
|
| WWP | Wipo information: published in national office |
Ref document number: 1996935888 Country of ref document: EP |
|
| WWR | Wipo information: refused in national office |
Ref document number: 1996935888 Country of ref document: EP |
|
| WWW | Wipo information: withdrawn in national office |
Ref document number: 1996935888 Country of ref document: EP |