Isoliertes Photoprotein Bolinopsin, sowie dessen Verwendung
Die Erfindung betrifft das Photoprotein Bolinopsin, dessen Nukleotid- und Aminosäuresequenz, sowie die Aktivität und Verwendung des Photoproteins Bolinopsin.
Photoproteine
Als Biolumineszenz bezeichnet man das Phänomen der Lichterzeugung durch Lebewesen. Sie ist das Ergebnis von biochemischen Reaktionen in Zellen, bei denen die chemische Energie in Form von Lichtquanten abgegeben wird (sog. kalte Emission durch Chemolumineszenz). Derartig erzeugtes Licht ist monochromatisch, denn es wird bei einem diskreten Elektronen-Übergang abgestrahlt, kann aber durch sekundäre Leuchtfarbstoffe (z.B. fluoreszierende Proteine bei Leucht- quallen der Gattung Aequora) in längerwellige Spektralbereiche verschoben werden.
Die biologische Funktion ist vielfältig: In der Meerestiefe zwischen 200 und 1000 m (Meso- pelagial) leuchten rund 90% aller Lebewesen. Die Leuchtsignale werden hier zur Partnerwerbung, Täuschung und als Köder eingesetzt. Auch Glühwürmchen und Leuchtkäfer nutzen die Lichtsignale zur Partnersuche. Die Bedeutung des Leuchtens von Bakterien, Pilzen und einzelligen Algen ist dagegen unklar. Es wird vermutet, dass es zur Koordination von vielen Einzel-Individuen einer großen Population eingesetzt wird oder eine Art biologische Uhr darstellt.
Eine Vielzahl an Coelenteraten ist biolumineszent (JMorin et al., 1974). Diese Organismen emittieren blaues oder grünes Licht. Das 1962 als erstes licht produzierendes Protein identifizierte Aequorin aus Aequoria victoria (Shimomura et al., 1969) emittierte als isoliertes Protein ein blaues Licht und nicht grünes Licht wie phänotypisch beobachtet bei Aequoria victoria. Später konnte das grün fluoreszierende Protein (GFP) aus Aequoria victoria isoliert werden, das aufgrund der Anregung durch das Aequorin die Meduse phänotypisch grün erscheinen lässt (Johnson et al, 1962; Hastings et al., 1969; Inouye et al, 1994). Als weitere Photoproteine konnten noch Clytin (Inouye et al., 1993), Mitrocomin (Fagan et al., 1993) und Obelin (Ularionov et al., 1995) identifiziert und beschrieben werden.
Tabelle 1: Übersicht über einige Photoproteine. Angegeben ist der Name, der Organismus aus dem das Protein isoliert worden ist und die Identifikationsnummer (Acc. No.) des Datenbankeintrages .
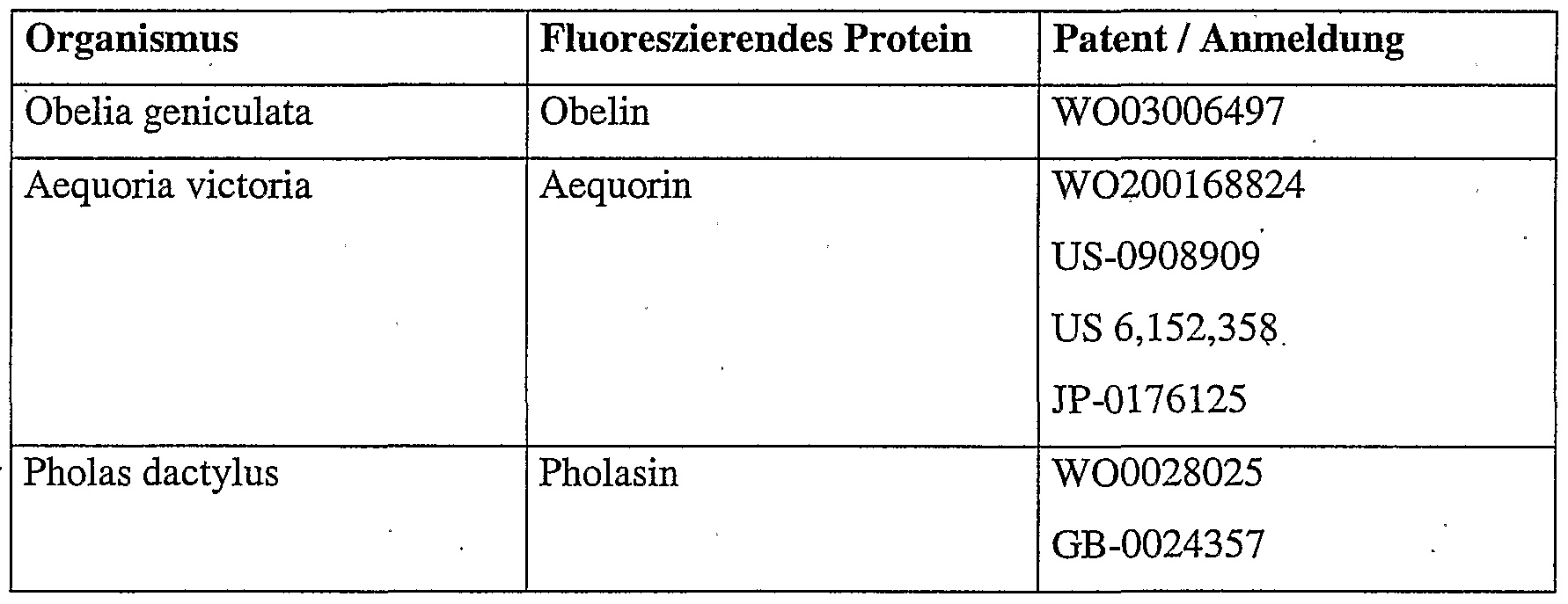
Tabelle 2; Übersicht über einige Photoproteine. Angegeben ist der Organismus aus dem das Protein isoliert worden ist, der Name des Photoproteins und eine Auswahl an Patenten bzw. Anmeldungen.
Biolumineszenz wird heute in der Technik vielfältig genutzt, z.B. in Form von Bio-Indikatoren für Umweltverschmutzung oder in der Biochemie zum empfindlichen Nachweis von Proteinen, zur Quantifizierung bestimmter Verbindungen oder als sogenannte "Reporter" bei der Untersuchung zellulärer Gen-Regulation.
Die Photoproteine unterscheiden sich nicht nur aufgrund ihrer Nukleotid- und Aminosäuresequenz, sondern auch aufgrund ihrer biochemischen und physikalischen Eigenschaften. Es konnte gezeigt werden, dass durch die Veränderung der Aminosauresequenz von Photo- proteinen die physikalischen und biochemischen Eigenschaften verändert werden können.
Beispiele von mutagenisierten Photoproteinen sind in der Literatur beschrieben (US 6,495,355; US 5,541,309; US 5,093,240; Shimomura et al., 1986).
Die Lichterzeugung durch die oben genannten Photoproteine erfolgt durch die Oxidation von Coelenterazin (Haddock et al., 2001; Jones et al., 1999).
Reportersysteme
Als Reporter- oder Indikatorgen bezeichnet man generell Gene, deren Genprodukte sich mit Hilfe einfacher biochemischer oder histochemischer Methoden leicht nachweisen lassen. Man unterscheidet mindestens 2 Typen von Reportergenen.
1. Resistenzgene. Als Resistenzgene werden Gene bezeichnet, deren Expression einer Zelle die Resistenz gegen Antibiotika oder andere Substanzen verleiht, deren Anwesenheit im Wachstumsmedium zum Zelltod führt, wenn das Resistenzgen fehlt.
2. Reportergene. Die Produkte von Reportergenen werden in der Gentechnologie als fusionierte oder unfusionierte Indikatoren verwendet. Zu den gebräuchlichsten Reportergenen gehört die beta-Galaktosidase (Alam et al., 1990), alkalische Phosphatase (Yang et al., 1997; Cullen et al., 1992), Luciferasen und andere Photoproteine (Shinomura, 1985; Phillips GN, 1997; Snowdowne et.al., 1984).
Als Lumineszenz bezeichnet man die Abstrahlung von Photonen im sichtbaren Spektralbereich, wobei diese durch angeregte Emittermoleküle erfolgt. Im Unterschied zur Fluoreszenz wird hierbei die Energie nicht von Außen in Form von Strahlung kürzerer Wellenlänge zugeführt.
Man unterscheidet Chemilumineszenz und Biolumineszenz. Als Chemolumineszenz bezeichnet man eine chemische Reaktion die zu einem angeregten Molekül führt, das selbst leuchtet, wenn die angeregten Elektronen in den Grundzustand zurückkehren. Wird diese Reaktion durch ein Enzym katalysiert, spricht man von Biolümineszenz. Die an der Reaktion beteiligten Enzyme werden generell als Luziferasen bezeichnet.
Einordung der Spezies Bolinopsis infundibulum
Eumetazoa → Radiata → Ctenophora → Tentaculata → Lobata → Bolinopsidae → Bolinopsis infundibulum
Isolierung der cDNA
Zur Untersuchung der Biolumineszenz-Aktivität der Spezies Bolinopsis infundibulum wurden Exemplare im Weißen Meer (Biologische Station Kartesh, Russland) gefangen und in flüssigem Stickstoff gelagert. Zur Erstellung der cDNA-Bibliotheken von Bolinopsis infundibulum, wurde die poly(a)+ RNA mit Hilfe des „Straight A" Isolationsmethode von Novagen (USA) isoliert.
Zur Herstellung der cDNA wurde eine RT-PCR durchgeführt. Hierzu wurden 1 μg RNA mit Reverser Transkriptase (Superscribt Gold II) nach folgendem Schema inkubiert:
PCR 1. 30 Sekunden 95°C 2. 6 Minuten 68°C 3. 10 Sekunden 95°C 4. 6 Minuten 68°C 17 Zyklen von Schritt 4 nach Schritt 3
Die Reaktionsprodukte wurden zur Inaktivierung der Polymerase für 30 Minuten bei 37°C mit Proteinase K inkubiert und die cDNA mit Ethanol präzipitiert. Die Expression-cDNA Bank wurde mit Hilfe des „SMART cDNA Library Construction Kits" der Firma Clontech (USA) nach Herstellerangaben durchgeführt. Die Klonierung erfolgte in den Expressionsvektor pTriplEx2 (Clontech; USA). Die Expressionsvektoren wurden durch Elektroporation in Bakterien des Stammes E. coli XLl-Blue transformiert.
Die Bakterien wurden auf LB-Nährböden plattiert und für 24 Stunden bei 37°C inkubiert. An- schließend wurde eine Replikaplattierung durchgeführt, indem die Bakterien mit Hilfe eines Nitro- cellulosefilters auf eine weitere Nährbodenplatte übertragen wurde. Die Replikaplatte wurde wiederum für 24 Stunden bei 37°C inkubiert und die gewachsenen Bakterienkolonien in LB- Flüssigmedium übertragen. Nach der Zugabe von JPTG (Endkonzentration 0,1 mM) wurden die Bakterien für 4 Stunden bei 37°C auf einem Schüttler inkubiert. Die Bakterien wurden durch Zentrifugation geerntet und die Bakterienmasse in 0,5 ml Aufschlusspuffer (5 mM EDTA, 20 mM Tris-HCL pH 9,0) bei 0°C resuspendiert. Anschließend erfolgte der Aufschluss der Bakterien durch Ultraschall.
Die Lysate wurden nach der Zugabe von Cöelenterazine (Endkonzentration 10E-07 M) bei 4°C für 3 Stunden inkubiert. Anschließend erfolgte die Messung der Biolumineszenz nach der Zugabe von Calziumchlorid (Endkonzentration 20 mM) im Luminometer.
Es wurde ein Photoprotein identifiziert. Das Photoprotein wurde als Bolinopsin bezeichnet. Im Folgenden wird das Photoprotein Bolinopsin im einzelnen dargestellt.
Bolinopsin
Das Photoprotein Bolinopsin zeigt die höchste Homologie auf Aminosäureebene zu Aequorin aus Aequoria victoria mit einer Identität von 44 % (gezeigt in Beispiel 8; Fig. 7). Auf Nukleinsäure- ebene liegt die Identität unter 30 % (gezeigt in Beispiel 9; Fig. 6). Zum Sequenzvergleich wurde das BLAST-Verfahren verwendet (Altschul et al., 1997).
Die Erfindung betrifft auch funktioneile Äquivalente von Bolinopsin. Fun tionelle Äquivalente sind solche Proteine, die vergleichbare physikochemische Eigenschaften haben und mindestens 70 % homolog sind zu SEQ LD NO: 2. Bevorzugt ist eine Homologie von mindestens 80 % oder 90 %. Besonders bevorzugt ist eine Homologie von mindestens 95 %.
Das Photoprotein Bolinopsin eignet sich als Reportergen für zelluläre Systeme speziell für Rezeptoren, für Ionenkanäle, für Transporter, für Transkriptionsfaktoren oder für induzierbare Systeme.
Das Photoprotein Bolinopsin eignet sich als Reportergen in bakteriellen und eukaryotischen Systemen speziell in Säugerzellen, in Bakterien, in Hefen, in Bakulo, in Pflanzen.
Das Photoprotein Bolinopsin eignet sich als Reportergene für zelluläre Systeme in Kombination mit biolumineszenten oder chemolumineszenten Systemen speziell Systemen mit Luziferasen, mit Oxygenasen, mit Phosphatasen.
Das Photoprotein Bolinopsin eignet sich als Fusionsprotein speziell für Rezeptoren, für Ionenkanäle, für Transporter, für Transkriptionsfaktoren, für Proteinasen, für Kinasen, für Phospho- diesterasen, für Hydrolasen, für Peptidasen, für Transferasen, für Membranproteine, für Glykoproteine.
Das Photoprotein Bolinopsin eignet sich zur Immobilisierung speziell durch Antikörper, durch Biotin, durch magnetische oder magnetisierbare Träger.
Das Photoprotein Bolinopsin eignet sich als Protein für Systeme des Energietransfers speziell der FRET- (Fluorescence Resonance Energy Transfer), BRET- (Bioluminescence Resonance Energy Transfer), FET (field effect transistors), FP (fluorescence polarization), FJLTRF (Homogeneous time-resolved fluorescence) Systemen.
Das Photoprotein Bolinopsin eignet sich als Markierung von Substraten oder JLiganden speziell für Proteasen, für Kinasen, für Transferasen.
Das Photoprotein Bolinopsin eignet sich zur Expression in bakteriellen Sytemen speziell zur Titer- bestimmung, als Substrat für biochemische Systeme speziell für Proteinasen und Kinasen.
Das Photoprotein Bolinopsin eignet sich als Marker speziell gekoppelt an Antikörper, gekoppelt an Enzyme, gekoppelt an Rezeptoren, gekoppelt an Ionenkanäle und andere Proteine.
Das Photoprotein Bolinopsin eignet sich als Reportergen bei der pharmakologischen Wirkstoffsuche speziell im HTS (High Throughput Screening).
Das Photoprotein Bolinopsin eignet sich als Komponente von Detektionssystemen speziell für ELISA (enzyme-linked immunosorbent assay), für Irnmunohistochemie, für Western-Blot, für die konfokale Mikroskopie.
Das Photoprotein Bolinopsin eignet sich als Marker für die Analyse von Wechselwirkungen speziell für Protein-Protein- Wechselwirkungen, für DNA-Protein-Wechselwirkungen, für DNA- RNA-Wechselwirkungen, für RNA-RNA- Wechselwirkungen, für RNA-Protein-Wechsel- wirkungen (DNA : deoxyribonucleic acid; RNA : ribonucleic acid; ).
Das Photoprotein Bolinopsin eignet sich als Marker oder Fusion. sprotein für die Expression in transgenen Organismen speziell in Mäusen, in Ratten, in Hamstern und anderen Säugetieren, in Primaten, in Fischen, in Würmern, in Pflanzen.
Das Photoprotein Bolinopsin eignet sich als Marker oder Fusionsprotein zur Analyse der Embryonalentwicklung.
Das Photoprotein Bolinopsin eignet sich als Marker über einen Kopplungs Vermittler speziell über Biotin, über NHS (N-hydroxysulfosuccimide), über CN-Br.
Das Photoprotein Bolinopsin eignet sich als Reporter gekoppelt an Nukleinsäuren speziell an DNA, an RNA.
Das Photoprotein Bolinopsin eignet sich als Reporter gekoppelt an Proteine oder Peptide.
Das Photoprotein Bolinopsin eignet sich als Reporter zur Messung von intra- oder extrazellulären Calziumkonzentrationen.
Das Photoprotein Bolinopsin eignet sich zur Charakterisierung von Signalkaskaden in zellulären Systemen.
Das an Nukleinsäuren oder Peptiden gekoppelte Photoprotein Bolinopsin eignet sich als Sonde speziell für Northern-Blots, für Southern-Blots, für Western-Blots, für ELISA, für Nuklein- säuresequenzierungen, für Proteinanalysen, Chip-Analysen.
Das Photoprotein Bolinopsin eignet sich Markierung von pharmakologischen Formulierungen speziell von infektiösen Agentien, von Antikörpern, von „small molecules".
Das Photoprotein 33olinopsin eignet sich für geologische Untersuchungen speziell für Meeres-, Grundwasser- und Flussströmungen.
Das Photoprotein Bolinopsin eignet sich zur Expression in Expressionssystemen speziell in in- vitro Translationssystemen, in bakteriellen Systemen, in Hefe Systemen, in Bakulo Systemen, in viralen Systemen, in eukaryotischen Systemen.
Das Photoprotein Bolinopsin eignet sich zur Visualisierung von Geweben oder Zellen bei chirurgischen Eingriffen speziell bei invasiven, bei nicht-invasiven, bei minimal-invasiven.
Das Photoprotein Bolinopsin eignet sich auch zur Markierung von Tumorgeweben und anderen phänotypisch veränderten Geweben speziell bei der histologischen Untersuchung, bei operativen Eingriffen.
Die Erfindung betrifft auch die Reinigung des Photoprotein Bolinopsin speziell als wildtyp Protein, als Fusionsprotein, als mutagenisiertes Protein.
Die Erfindung betrifft auch die Verwendung des Photoprotein Bolinopsin auf dem Gebiet der Kosmetik speziell von Badezusätzen, von Lotionen, von Seifen, von Körperfarben, von Zahncreme, von Körperpudern. (
Die Erfindung betrifft auch die Verwendung des Photoprotein Bolinopsin zur Färbung speziell von Nahrungsmitteln, von Badezusätzen, von Tinte, von Textilien, von Kunststoffen.
Die Erfindung betrifft auch die Verwendung des Photoprotein Bolinopsin zur Färbung von Papier speziell von Grußkarten, von Papierprodukten, von Tapeten, von Bastelartikehi.
Die Erfindung betrifft auch die Verwendung des Photoprotein Bolinopsin zur Färbung von Flüssigkeiten speziell für Wasserpistolen, für Springbrunnen, für Getränke, für Eis.
Die Erfindung betrifft auch die Verwendung des Photoprotein Bolinopsin zur Herstellung von Spielwaren speziell von Fingerfarbe, von Schminke.
Die Erfindung betrifft Nukleinsäuremoleküle, die das Polypeptid offenbart durch SEQ LD NO: 2 kodieren.
Die Erfindung betrifft das Polypeptid mit der Aminosauresequenz, die in SEQ LD NO: 2 offenbart ist.
Die Erfindung bezieht sich des weiteren auf Nukleinsäuremoleküle, ausgewählt aus der Gruppe bestehend aus
a) Nukleinsäuremolekülen, die ein Polypeptid kodieren, welches die Aminosäuresequenz offenbart durch SEQ ID NO: 2 umfasst;
b) Nukleinsäuremolekülen, welche die durch SEQ ID NO: 1 dargestellte Sequenz enthalten;
c) Nukleinsäuremolekülen, deren komplementärer Strang mit einem Nukleinsäuremolekül aus a) oder b) unter stringenten Bedingungen hybridisiert und welche die biologische Funktion eines Photoproteins aufweisen;
d) Nukleinsäuremolekülen, welche sich auf Grund der Degenerierung des genetischen Kodes von den unter c) genannten unterscheiden;
e) Nukleinsäuremolekülen, welche eine Sequenzhomologie von mindestens 95 % zu SEQ LD NO: 1 zeigen, und deren Proteinprodukt die biologische Funktion eines Photoproteins aufweist; und
f) Nukleinsäuremolekülen, welche eine Sequenzhomologie von mindestens 65 % zu SEQ ID NO: 1 zeigen, und deren Proteinprodukt die biologische Funktion eines Photoproteins aufweist.
Die Erfindung betrifft auch Nukleinsäuremoleküle, die eine Sequenzhoπiologie von mindestens 95 %, 90 %, 85 %, 80 %, 75 %, 70 %, 65 % oder 60 % zu SEQ ID NO: 1 aufweisen und für ein Polypeptid kodieren, welches die Eigenschaften eines Photoproteins besitzt.
Die Erfindung betrifft die oben genannten Nukleinsäuremoleküle, bei denen die Sequenz einen funktionalen Promotor 5" zu der das Photoprotein kodierenden Sequenz enthält.
Die Erfindung betrifft auch Nukleinsäuremoleküle wie vorhergehend beschrieben, die Bestandteil von rekombinanten DNA oder RNA Vektoren sind.
Die Erfindung betrifft Organismen, die einen solchen Vektor enthalten.
Die Erfindung bezieht sich auf Oligonukleotide mit mehr als 1O aufeinanderfolgenden Nukleotiden, die identisch oder komplementär zur DNA oder RNA Sequenz der Bolinopsin Moleküle oder der weiteren erfindungsgemäßen Molekülen sind.
Die Erfindung betrifft Photoproteine, die durch die vorhergehend beschriebenen Nukleotid- sequenzen kodiert sind.
Die Erfindung bezieht sich auf Verfahren zur Expression der erfindungsgemäßen Photoprotein Polypeptide in Bakterien, eukaryontischen Zellen oder in in vitro Expressionssystemen.
Die Erfindung betrifft auch Verfahren zur Aufreinigung/Isolierung eines erfindungsgemäßen Photoprotein Polypeptides.
Die Erfindung bezieht sich auf Peptide mit mehr als 5 aufeinanderfolgenden Aminosäuren, die immunologisch durch Antikörper gegen die erfindungsgemäßen Photoproteme erkannt werden.
Die Erfindung betrifft die Verwendung der erfindungsgemäßen, für Photoproteine kodierende Nukleinsäuren als Marker- oder Reportergene, insbesondere für die pharmakologische Wirkstoffsuche und Diagnostik.
Die Erfindung betrifft die Verwendung der cifmdungsgemäßen Photoproteme bzw. eine erfindungsgemäße, für ein Photoprotein kodierende Nukleinsaüre als Marker oder Reporter bzw. als Marker- oder Reportergen.
Die Erfindung betrifft die Verwendung des Photoproteins Bolinopsin. (SEQ JJD NO: 2) bzw. die Verwendung einer für das Photoprotein Bolinopsin kodierenden Nvxideinsäure als Marker oder Reporter bzw. als Marker oder Reportergen insbesondere für die pharmakologische Wirkstoffsuche und Diagnostik.
Die Erfindung betrifft die Verwendung der in SEQ ID NO: 1 dargestellten Nukleinsaüre als Marker- oder Reportergen, insbesondere für die pharmakologische Wirikstoffsuche und Diagnostik.
Gegenstand der Erfindung sind auch polyklonale oder monoklonale Avntikörper, welche ein erfin- dungsgemäßes Polypeptid erkennen.
Die Erfindung betrifft auch monoklonale oder polyklonale Antikörper, die das Photoprotein Bolinopsin (SEQ ID NO: 2) erkennen.
Expression der erfindungsgemäßen Photoproteine
Als Expression bezeichnet man die Produktion eines Moleküls, das nach dem Einbringen des Gens in eine geeignete Wirtszelle die Transcription und Translation des in einen Expressionsvektor Monierte Fremdgen erlaubt. Expressionsvektoren enthalten die für die Expression von Genen in Zellen von Prokaryoten oder Eukaryonten erforderlichen Kontrollsignale.
Expressionsvektoren können prinzipiell auf zwei verschiedene Weisen konstruiert werden. Bei den sogenannten Transcriptionsfusionen wird das vom einklonierten Fremdgen codierte Protein als
authentisches, biologisch aktives Protein synthetisiert. Der Expressionsvektor trägt hierzu alle zur Expression benötigten 5 - und 3 - Kontrollsignale.
Bei den sogenannten Translationsfusionen wird das vom einklonierten Fremdgen codierte Protein als Hybridprotein zusammen mit einem anderen Protein exprimiert, das sich leicht nachweisen lässt. Die zur Expression benötigten 5 - und 3V- Kontrollsignale inklusive des Startcodons und eventuell ein Teil der für die N-terminalen Bereiche des zu bildenden Fϊybridproteins codierenden Sequenzen stammen vom Vektor. Der zusätzliche eingeführte Proteinteil stabilisiert nicht nur in vielen Fällen das vom einklonierten Fremdgen codierte Protein vor dem Abbau durch zelluläre Proteasen, sondern lässt sich auch zum Nachweis und zur Isolierung des gebildeten Hybridproteins einsetzen. Die Expression kann sowohl transient, als auch stabil erfolgen. Als Wirtsorganismen eignen sich sowohl Bakterien, Hefen, Viren als auch eukaryotische Systeme.
Reinigung der erfindungsgemäßen Photoproteine
Die Isolierung von Proteinen (auch nach Überexpression) wird häufig als Proteinreinigung bezeichnet. Zur Proteinreinigung steht eine Vielzahl an etablierten Methoden und Verfahren zur Verfügung.
Die Fest-Flüssig-Trennung ist eine Grundoperation bei Proteinisolierungen. Sowohl bei der Abtrennung der Zellen vom Kulturmedium als auch bei der Klärung des Rohextraktes nach Zellaufschluss und Entfernung der Zelltrümmer, bei der Abtrennung von Niederschlägen nach Fällungen usw. ist der Verfahrensschritt erforderlich. Er erfolgt durch Zentrifugation und Filtration.
Durch Gewinnung intrazellulärer Proteine muss die Zellwand zerstört bzw. durchlässig gemacht werden. Je nach Maßstab und Organismus werden dazu Hochdruckhomogenisatoren oder Rührwerkskugel- bzw. Glasperlenmühlen eingesetzt. Im Labormaßstab kommen u.a. mechanische Zellintegrationen und Ultraschallbehandlung zum Einsatz.
Sowohl für extrazelluläre als auch intrazelluläre Proteine (nach Zellaufschluss) sind verschiedene Fällungsverfahren mit Salzen (insbesondere Ammoniumsulfat) oder organischen Lösungsmitteln (Alkohole, Aceton) eine schnelle und effiziente Methode zur Konzentration von Proteinen. Bei der Reinigung intrazellulärer Proteine ist die Entfernung der löslichen Nukleinsäuren erstrebenswert (Fällung z.B. mit Streptomycin- oder Protaminsulfat). Bei der Gewinnung extrazellulärer Proteine werden häufig Träger (z.B. Stärke, Kieselgur) vor Zugabe der Fällungsmittel zugesetzt, um besser handhabbare Niederschläge zu erhalten.
Für die Feinreinigung stehen zahlreiche chromatographische und Verteilungsverfahren zur Verfügung (Absorptions- und Ionenaustauschchromatographie, Gelfiltration, Affinitätschromatographie, Elektrophoresen). Eine Säulenchromatographie wird auch im technischen Maßstab angewandt. Für den Labormaßstab ist vor allem die Affinitätschromatographie von Bedeutung, die Reinigungsfaktoren bis zu mehreren 100 pro Schritt ermöglicht.
Extrazelluläre Proteine fallen in relativ verdünnten Lösungen an. Sie müssen ebenso wie extrazelluläre Proteine vor ihrer weiteren Verwendung konzentriert werden. Neben den schon erwähnten Verfahren hat sich - auch im industriellen Maßstab - die Ultrafiltration bewährt.
Anorganische Salze als Begleitstoffe von Proteinen sind für spezifische Anwendungen häufig unerwünscht. Sie können u.a. durch Gelfiltration, Dialyse und Diafiltration entfernt werden.
Zahlreiche Proteine kommen als Trockenpräparate zum Einsatz. Als Trocknungsverfahren sind die Vakuum-, Gefrier- und Sprühtrocknung von Bedeutung.
Nukleotid- und Aminosäuresequenzen
Das Photoprotein Bolinopsin wird durch die folgende Nukleotidsequenz kodiert (SEQ ID NO: 1):
5 -
ATGCCTCTAGACGAGACCAACAACGAAAGCTATAGATGGCTGAGAAGTGTGGGTAAC GATTGGCAGTTTGATGTCGAGGACGTTCATCCTAAACAGCTTAGTCGGCTCTACAAGA GATTCGACACCTTCGATCTAGACAGTGACGGTCGTATGGACATGGACGAGATCCTGTA CTGGCCCGACAGAATGAGGCAGCTGGTGAACGCTTCTGACGAACAGGTCGAGAAGAT GAGGGCTGCTTGCTACACCTTCTTCCACAACAAAGGAGTGGATCCAGAAAAGGGACTC CTCAGAGACGACTGGGTTGAGGCTAACAGAGTATTTGCTGAGGCTGAAAGAGAGAGG GAACGACGTGGCATGCCCTCCTTGATTGGTCTTTTGTCAGACGCTTACTACGATGTCCT GGATGATGACGGTGATGGTACTGTTGATGTTGATGAACTCAAAACCATGATGAAGGCT TTTGATGTGCCCCAGGAGGCTGCCTACACCTTCTTTAAGAAA.GCTGACACGGATAATA GTGGAAAACTGGAGAGAAGCGAACTGGTCCATCTCTTCAGAAAGTTCTGGATGGAATC CTACGATCCTCAGTGGGACGGTGTCTACGCTTACAAATATTA-A-3\
Daraus ergibt sich eine Aminosäuresequenz von (SEQ ID NO: 2):
MPLDETNNESYRWLRSVGNDWQFDVEDVHPKQLSP^YΈSFTJΓFDLDSDGRMDMDEΓLY WPDRMRQLVNASDEQVEKMRAACYTFFHNKGVDPEKGLLRDX)WVEANRVFAEAERERE RRGMPSLIGLLSDAYYDVLDDDGDGTVDVDELKTMMKAFDVPQEAAYTFFKKADTDNS GKLERSELVHLFRKFWMESYDPQWDGVYAYKY
Diese Sequenzen finden sich im Sequenzlisting wieder.
Kurze Beschreibung der Figuren
Fig. 1: Die Fig. 1 zeigt die Plasmidkarte des Vektors pTriplEX2-Bolinopsin.
Fig. 2: Die Fig. 2 zeigt die Plasmidkarte des Vektors pcDNT A3 -Bolinopsin
Fig. 3: Die Fig. 3 zeigt die Exitation von Bolinopsin. Y : Intensität; X : Wellenlänge [um].
Fig. 4: Die Fig. 4 zeigt die Fluoreszenz von Bolinopsin. Y : Intensität; X : Wellenlänge [nm] .
Fig. 5: Die Fig. 5 zeigt die Biolumineszenz von Bolinopsin. Y : Intensität; X : Wellenlänge [nm].
Fig. 6: Die Fig. 6 zeigt das Alignment von Bolinopsin und Aequorin (aequoria victoria) auf Nukleinsäureebene.
Fig. 7: Die Fig. 7 zeigt das Alignment von Bolinopsin und Aequorin (aequoria victoria) auf Aminosäureebene.
Fig. 8: Die Fig. 8 zeigt das Ergebnis der Biolumineszenzmessung von Bolinopsin nach bakterieller Expression. Y : Lumineszenz in RLU [relative light units]; X : μl Lysat : 0 = uninduziertes Kontrolllysat.
Fig. 9: Die Fig. 9 zeigt das Ergebnis der Biolumineszenzmessung von Bolinopsin nach bakterieller Expression in Abhängigkeit vom eingesetzten Coelenterazinderivat. Y : Lumineszenz in RLU [relative light units]; X : Coelenterazinderivat : 1 = nativ, 2 = cp, 3 = f , 4 = fcp, 5 = hcp, 6 = h, 7 = i, 8 = ip, 9 = n; Balken : schwarz : uninduziertes Kontrolllysat; hell-grau : 10 μl Lysat; weiss : 20 μl Lysat; dunkel-grau : 40 μl Lysat.
Beispiele
Beispiel 1
Als Vektor zur Herstellung des im folgenden dargestellten Konstruktes wurde das Plasmid pTriplEx2 der Firma Clontech verwendet. Das Derivat des Vektors wurde als pTriplEx2- 5 Bolinopsin bezeichnet. Der Vektor pTriplEx2-Bolinopsin wurde zur Expression von Bolinospin in bakteriellen Systemen verwendet.
Die Fig. 1 zeigt die Plasmidkarte des Vektors pTriplEX2-Bolinopsin .
Beispiel 2
Als Vektor . zur Herstellung des im folgenden dargestellten Konstruktes wurde das PlasmidO pcDNA3.1(+) der Firma Clontech verwendet. Das Derivat des Vektors wurde als pcDNA3- Bolinopsin bezeichnet. Der Vektor pcDNA3-Bolinopsin wurde zur Expression von Bolinospin in eukaryotischen Systemen verwendet.
Die Fig. 2 zeigt die Plasmidkarte des Vektors pcDl A3-Bolinopsm .
Beispiel 3 5 Bakterielle Expression
Die bakterielle Expression erfolgte im E. coli Stamm BL21(DE3). durch Transformation der Bakterien mit den Expressionsplasmiden pTriplEX2-Bolinopsin und pTriplEX2. Die transformierten Bakterien wurden in LB-Medium bei 37°C für 3 Stunden inkubiert und die Expression für 4 Stunden durch Zugabe von IPTG bis zu einer Endkonzentration von 1 mM induziert. DieO induzierten Bakterien wurden durch Zentrifugation geerntet, in PBS + 5 mM EDTA resuspendiert und durch Ultraschall aufgeschlossen. Das Lysat wurde 3 Stunden mit Coelenterazin im dunkeln inkubiert. Direkt nach der Zugabe von 5 mM Calziumchlόrid wurde die Biolumineszenz im Luminometer gemessen. Die Integrationszeit der Messung betrug 40 Sekunden.
Die Fig. 8 zeigt die Ergebnisse der Biolumineszenzmessung von Bolinopsin. 5 Beispiel 4
Eukaryotische Expression
Die konstitutive eukaryotische Expression erfolgte in CHO-Zellen durch Transfektion der Zellen mit den Expressionsplasmiden pcDNA3-Bolinopsin und pcDNA3.1(+) in transienten Experi-
menten. Hierzu wurden 10000 Zellen pro Loch in DMEM-F12 Medium auf 96 Loch ikrotiter- platten plattiert und über Nacht bei 37°C inkubiert. Die Transfektion erfolgte mit Hilfe des Fugene 6 Kits (Röche) nach Herstellerangaben. Die transfizierten Zellen wurden über Nacht bei 37°C in DMEM-F12 Medium inkubiert.
Beispiel 5
BLAST
Ergebnis einer BLAST-Analyse von Bolinopsin auf der Aminosäureebene.
>pdb|lJF2|A Chain A, Crystal Structure Of W92f Obelin Mutant From Obelia Longissima At 1.72 Angstrom Resolution, Length = 195, Score = 85.1 bits (209), Expect = 8e-16, Identities = 52/177 (29%), Positives = 90/177 (50%), Gaps = 4/177 (2%)
>emb|CAD87698.1| unnamed protein product [synthetic construct], Length = 195, Score =
81.6 bits (200), Expect = 8e-15, Identities = 51/177 (28%), Positives = 89/177 (499b), Gaps = 4/177 (2%)
>pdb|lJF0|A Chain A, The Crystal Structure Of Obelin From Obelia Geniculata At 1.82 A Resolution, gb|AAL86372.1|AF394688_l apoobelin [Obelia geniculata], Length = 195, Score = 80.1 bits (196), Expect = 2e-14, Identities = 51/177 (28%), Positives = 89/177 (49%), Gaps = 4/177 (2%)
>sp|P39047|MYTR_MITCE Mitrocomin precursor, pir||S39022 mitrocomin precursor - rv itrocoma cellularia, gb|AAA29298.1| apomitrocornin, Length = 198, Score = 78.6 bits (192), Expect = 7e-14, Identities = 47/177 (26%), Positives = 91/177 (50%), Gaps = 4/177 (2%)
>sp|Q08121|CLYT_CLYGR Clytin precursor (Phialidin), pir||S28860 clytin - hydromedusa (Clytia gregarium), emb|CAA49754.1| clytin [Clytia gregaria], gb|AAA28293.1| apoclytin, Length = 198, Score = 77.4 bits (189), Expect = 2e-13, Identities = 53/177 (29%), Positives = 89/177 (49%), Gaps = 4/177 (2%)
Beispiel 6
BLAST
Ergebnis einer BLAST-Analyse von Bolinopsin auf Nukleinsäureebene.
>gb|AC073341.10| Homo sapiens BAC clone RP11-549I23 from 7, complete sequence, Length = 185574, Score = 52.6 bits (27), Expect = 4e-04, Identities = 33/36 (91%)
>gb|AC092850.13| Homo sapiens 12 BAC RP11-346B9 (Roswell Park Cancer Institute Human BAC Library) complete sequence, Length = 176733, Score = 46.8 bits (24), Expect = 0.023, Identities = 32/36 (88%)
>gb|AC126564.7| Homo sapiens 12 BAC RP11-638F5 (Roswell Park Cancer Institute Human BAC Library) complete sequence, Length = 121242, Score = 46.8 bits (24), Expect = 0.023, Identities = 32/36 (88%)
>gb|AC093924.3| Genomic sequence for Mus musculus, clone RP23-239M9, from chromosome 17, complete sequence, Length = 166277, Score = 44.9 bits (23), Expect = 0.086, Identities = 31/35 (88%)
>gb|AC060234.11| Homo sapiens chromosome 10 clone RP11-523O18, complete sequence, Length = 170073, Score = 44.9 bits (23), Expect = 0.O86, Identities = 31/35 (88%)
>gb|AC084727.14| Homo sapiens chromosome 10 clone RP11-507P23, complete sequence, Length = 188652, Score = 44.9 bits (23), Expect = 0.086, Identities = 31/35 (88%)
Beispiel 7
Die Fig. 6 zeigt das Alignment von Bolinopsin mit Aequorin (Aequoria victoria) auf Nukleinsäureebene.
Beispiel 8
Die Fig. 7 zeigt das Alignment von Bolinopsin mit Aequorin (Aequoria victoria) auf Aminosäureebene.
Beispiel 9
Spektrum des Photoproteins Bolinopsin
Zur Messung der spektralen Eigenschaften von Bolinopsin wurden E. coli BL21(DE3) mit den Plasmiden pTriplEX2-CGFP und pTriplEX2 transformiert. Die Induktion erfolgte durch die Zugabe von 1 rnM IPTG und einer Inkubation von 4 Stunden bei 37°C. Anschließend wurden die Bakterien geerntet und in PBS resuspendiert. Die Lyse erfolgte durch Ultraschall. Anschließend erfolgte die Messung der Fluoreszenz bzw. Biolumineszenz. Das Maximum der Exitation lag bei 352 nm, der Fluoreszenz bei 452 nm und das der Biolumineszenz bei 468 nm.
Die Fig. 3 zeigt die Exitation von Bolinopsin Die Fig. 4 zeigt die Fluoreszenz von Bolinopsin
Die Fig. 5 zeigt die Biolumineszenz von Bolinopsin
Beispiel 10
Zur Identifizierung von Substraten für Bolinopsin wurden 5 μl Lösungen verschiedener Coelenterazinderivate (10"4 M) in Methanol mit jeweils 0, 10, 20 und 40 μl Lysat in einem Gesamtvolumen von 75 μl für 3 Stunden bei 4°C inkubiert und die Lumineszenz nach der Zugabe von 25 μl Calziumchlorid (Endkonzentration 5 mM) gemessen. Die Coelenterazine wurden von Sigma (Deutschland) bezogen. Bolinopsin zeigte mit allen eingesezten Coelenterazinderivaten Biolumineszenzaktivität. Die höchste Aktivität könnte mit dem nativen Coelenterazin gemessen werden.
Die Fig. 9 zeigt die Coelenterazin-Derivate als potentielle Substrate für Bolinopsin und eine grafische Darstellung der Lumineszenzmessung für 30 Sekunden bei 8,7 kV im Luminometer (RLU, relative light units).
Literatur / Patente
US 6,495,355 US 5,541,309
US 5,093,240
US-0908909
US 6,152,358
JP-0176125 GB-0024357
WO03006497
WO200168824
Alam J, Cook JL. Reporter genes: application to the study of rnammalian gene transcription. Anal
Biochem. 1990 Aug l;188(2):245-54 Altschul, Stephen F., Thomas L. Madden, Alejandro A. Schäffer, Jinghui Zhang, Zheng
Zhang, Webb Miller, and David J. Lipman (1997); Gapped BLAST and PSI-BLAST: a new generation of protein database search programs; NΗcleic Acids Res. 25:3389-3402
Cullen Bryan R., Malim Michael H., Secreted placental alkaline phosphatase as a eukaryotic reporter gene. Methods in Enzymology. 216:362ff Fagan TF, Ohmiya Y, Blinks JR, Inouye S, Tsutji FI. Cloning, expression and sequence analysis of cDNA for the Ca(2+)-binding photoprotein, mitxocomin. FEBS Lett. 1993 Nov l;333(3):301-5
Hastings, J.W. and Morin, J.G. (1969) Comparative biochemistry of calcium-activated photoproteins from the ctenophore, Mnemiopsis and the coelenterates Aequorea, Obelia, and
Pelagia. Biol. Bull. 137, 402.
Haddock SH, Rivers TJ, Robison BH. Can coelenterates make coelenterazine? Dietary requirement for luciferin in cnidarian bioluminescence. Proc Natl Acad Sei U S A 20O1 Sep
25;98(20): 11148-51
Inouye S, Tsuji FI. (1994) Aequorea green fluorescent protein. Expression of the gene and fluorescence characteristics of the recombinant protein. FEBS Lett 1994 Mar 21;341(2-3):277-80
Inouye S, Tsuji FI. Cloning and sequence analysis of cDNA for the Ca(2+)-activated photoprotein, clytin. FEBS Lett. 1993 Jan 11 ;315(3):343-6.
Illarionov BA, Bondar VS, Illarionova VA, Vysotski ES. Sequence of the cDNA encoding the
Ca(2+)-activated photoprotein obelin from the hydroid polyp Obelia longissima. Gene. 1995 Feb
14;153(2):273-4.
Jones K, Hibbert F, Keenan M. Glowing jellyfish, luminescence and a molecule called coelenterazine. Trends Biotechnol 1999 Dec;17(12):477-81
Johnson, F.H., Shimomura, O., Saiga, Y., Gershman, L.C., Reynolds, G.T., and Waters, J.R.
(1962) Quantum efficiency of Cypridina luminescence, with a note on that of Aequorea. J. Cell.
Comp. Physiol. 60, 85-103.
Morin, J.G. and Hastings, J.YV. (1971) Biochemistry of the bioluminescence of colonial lrydroids and other coelenterates. J. Cell. Physiol. 77, 305-311.
Phillips GN. Structure and dynamics of green fluorescent protein. Curr Opin Struct Biol. 1997
Dec;7(6): 821-7
Shimomura O, Johnson FH. Properties of the bioluminescent protein aequorin.
Biochemistry. 1969 Oct;8(10): 3991-7 Shimomura O., Bioluminescence in the sea: photoprotein Systems. Symp Soc Exp Biol.
1985;39:351-72
Shimomura O. Isolation and properties of various molecular forms of aequorin.
Biochem J. 1986 Mar l;234(2):271-7.
Snowdowne KW, Borle AB. Measurement of cytosolic free calcium in mammalian cells with aequorin. Am J Physiol. 1984 Noy;247(5 Pt l):C396-408.
Yang Te-Tuan, Sinai Parisa, Kitts Paul A. Kain Seven R., Quantification of gene expresssion with a secreted alkaline phosphatase reporter system. Biotechnique. 1997 23(6) lllOff