WO2016010161A1 - 多量体IgA型遺伝子組換え抗体及びその利用 - Google Patents
多量体IgA型遺伝子組換え抗体及びその利用 Download PDFInfo
- Publication number
- WO2016010161A1 WO2016010161A1 PCT/JP2015/070742 JP2015070742W WO2016010161A1 WO 2016010161 A1 WO2016010161 A1 WO 2016010161A1 JP 2015070742 W JP2015070742 W JP 2015070742W WO 2016010161 A1 WO2016010161 A1 WO 2016010161A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- antibody
- iga
- protein
- type
- multimeric
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images




















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/01—Preparation of mutants without inserting foreign genetic material therein; Screening processes therefor
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/10—Antimycotics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
- A61P31/16—Antivirals for RNA viruses for influenza or rhinoviruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P33/00—Antiparasitic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/06—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies from serum
- C07K16/065—Purification, fragmentation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from animals or humans
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1086—Preparation or screening of expression libraries, e.g. reporter assays
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/66—General methods for inserting a gene into a vector to form a recombinant vector using cleavage and ligation; Use of non-functional linkers or adaptors, e.g. linkers containing the sequence for a restriction endonuclease
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
- A61K2039/507—Comprising a combination of two or more separate antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IG], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—RNA viruses
- C07K16/102—Coronaviridae (F)
Definitions
- the present invention relates to a multimeric IgA type recombinant antibody and use thereof.
- This application claims priority on July 18, 2014 based on Japanese Patent Application No. 2014-148328 for which it applied to Japan, and uses the content here.
- Antibodies have functional activities such as neutralization activity due to the sum of antigen binding activity and specificity, and are widely used in the biotechnology industry as antibody drugs, diagnostic agents, biological research tools, and the like.
- IgM immunoglobulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like globulin-like ⁇ , ⁇ A, ⁇ -A, and the secretory fluid.
- IgA is the main antibody in the blood in the body
- IgA secretory fluid
- antibodies such as IgA and IgM are known to function in vivo because antibodies having the same variable region form multimers such as dimers and pentamers.
- multimeric IgA exists in secretions such as colostrum and saliva, and in the serum of patients with multiple myeloma.
- an object of the present invention is to provide a multimeric IgA type recombinant antibody.
- Another object of the present invention is to provide a medicament containing a multimeric IgA type recombinant antibody as an active ingredient.
- Another object of the present invention is to provide a method for producing a multimeric IgA antibody. Moreover, it aims at providing the method of improving the antigen binding activity of an antibody.
- the present invention is as follows. (1) Multimeric IgA type recombinant antibody. (2) The multimeric IgA type recombinant antibody according to (1), wherein the amino acid residue at position 458 of the heavy chain constant region is an amino acid residue derived from a hydrophobic amino acid. (3) The multimeric IgA-type recombinant antibody according to (1) or (2), wherein the tetramer content is 20 mol% or more of the total IgA. (4) A medicament comprising the multimeric IgA gene recombinant antibody according to any one of (1) to (3) as an active ingredient. (5) The medicament according to (4), which is used for treatment or prevention of infectious diseases.
- a method for producing a multimeric IgA type antibody comprising co-expressing an IgA type antibody heavy chain protein, an antibody light chain protein, an antibody J chain protein and a secretory component protein in one cell.
- the production method according to any one of (6) to (8), wherein the cell is a CHO YA7 cell line (Accession No. NITE BP-01535).
- the step is performed by introducing an expression vector for expressing an IgA type antibody heavy chain protein, antibody light chain protein, antibody J chain protein and secretory component protein into the cell,
- An RNA binding protein recognizes, binds or interacts downstream of the promoter and upstream of the start codon of a nucleic acid encoding IgA type antibody heavy chain protein, antibody light chain protein, antibody J chain protein or secretory component protein
- the production method according to any one of (6) to (9), comprising a cis-element.
- the cis-element is a sequence motif GAN 1- (X) n -ACN 2 (n is an integer of 3 to 6, and N 1 and N 2 are each independently A, T, G, The production method according to (10), comprising 1 to several base sequences comprising any one of C.).
- the cis-element is The base sequence shown in any of SEQ ID NOs: 21 to 23, A nucleotide sequence consisting of a nucleotide sequence in which one to several bases are deleted, substituted or added in the nucleotide sequence shown in any of SEQ ID NOs: 21 to 23, and an RNA binding protein recognizes, binds or interacts , Consisting of a base sequence that is 80% or more identical to the base sequence shown in any of SEQ ID NOs: 21 to 23, and a base sequence that an RNA binding protein recognizes, binds or interacts, or Consisting of a base sequence capable of hybridizing under stringent conditions with a nucleic acid consisting of a base sequence complementary to the nucleic acid consisting of the base sequence shown in any of SEQ ID NOs: 21 to 23, and recognizing an RNA binding protein;
- a method for improving the antigen-binding activity or neutralizing activity of the antibody comprising the step of converting the antibody into multimeric IgA.
- the step includes the step of co-expressing the IgA-type antibody heavy chain protein having the heavy chain variable region of the antibody, the light chain protein of the antibody, the antibody J chain protein and the secretory component protein in one cell. , (13) or (14).
- 14 is a photograph showing the result of Experimental Example 18.
- 14 is a photograph showing the result of Experimental Example 19.
- 14 is a photograph showing the result of Experimental Example 19.
- 14 is a photograph showing the result of Experimental Example 19. It is a graph which shows the result of example 20 of an experiment. It is a graph which shows the result of example 20 of an experiment. It is a graph which shows the result of example 20 of an experiment. It is a graph which shows the result of example 20 of an experiment. It is a graph which shows the result of example 20 of an experiment. It is a graph which shows the result of example 20 of an experiment. It is a graph which shows the result of example 20 of an experiment. It is a graph which shows the result of example 20 of an experiment. It is a graph which shows the result of example 20 of an experiment.
- IgA (Monomer IgA, mIgA)
- IgA exists mainly in the serum as monomeric IgA with a molecular weight of around 170,000, and IgA1 is the main component.
- multimeric IgA secreted by plasma cells a complex having a heavy chain: light chain: J chain composition ratio of 4: 4: 1 and referred to as “multimeric IgA” and secreted from mucosal epithelial cells.
- S-IgA dashed chain: light chain: J chain: SC complex of 4: 4: 1: 1 is referred to as “multimeric IgA” and sometimes refers to both. Used strictly without distinction.
- a component having a molecular weight higher than that of dimer IgA is detected from the mobility in electrophoresis and the behavior in gel filtration chromatography, and it is expected to be higher than the dimer, but it may be called multimeric IgA. Since the differentiation from the aggregate has not been made, the details of the molecular structure are unknown.
- the J chain is a polypeptide chain having a molecular weight of 15 kDa and has an N-linked sugar chain and is folded so as to have an immunoglobulin structure.
- the J chain is essential for the dimer IgA to interact with pIgR. Dimer IgA incorporating the J chain causes an interaction between the D1 of pIgR and the Fc region of IgA or pIgR and the J chain, followed by the 311 th cys residue in the IgAC ⁇ 2 domain and the D5 of SC.
- a secreted trimer antibody and a recombinant secreted trimer antibody are referred to as “trimer” without distinction.
- the secretory tetramer antibody and the recombinant secretory tetramer antibody may be referred to as “tetramer” without particular distinction.
- a tetrameric or higher secretory antibody and a tetrameric or higher recombinant secreted multimeric antibody may be referred to as “polymer” without any particular distinction.
- the present invention provides multimeric IgA type recombinant antibodies.
- the multimeric IgA gene recombinant antibody of the present embodiment may be an antibody that was originally non-IgA type converted to IgA type by genetic recombination and further multimerized.
- the non-IgA type antibody is not particularly limited, and examples thereof include non-IgA type human antibodies, non-human mammal-derived antibodies, rodent-derived antibodies, and avian-derived antibodies.
- the class of the antibody is not particularly limited, and examples thereof include IgG type, IgM type, IgE type, IgD type, IgY type antibody and the like.
- immunoglobulin molecules have allotypes having genetically different antigenicity found among individuals within the same species. Allotypes often result from one to several amino acid variations in the constant region of an immunoglobulin molecule.
- IgA2m (1) and IgA2m (2) are generally recognized, and the presence of a third allotype called IgA2 (n) has also been reported.
- IgA2 may be any of the above allotypes.
- the multimeric IgA type recombinant antibody of the present embodiment preferably contains a secretory component protein (SC) in the molecule. Further, it is preferably a dimer or more, more preferably a trimer or more, and further preferably a tetramer or more.
- SC secretory component protein
- the antibody of this embodiment makes it possible to industrially apply an IgA gene recombinant antibody.
- the 458th amino acid residue of the heavy chain constant region is preferably an amino acid residue derived from a hydrophobic amino acid.
- the inventors determined that the ratio of the trimer / tetramer antibody when the amino acid residue at position 458 of the heavy chain constant region is an amino acid residue derived from a hydrophobic amino acid. It was found that it can be greatly enhanced.
- hydrophobic amino acid examples include isoleucine (I), leucine (L), methionine (M), tryptophan (W), and glycine (G).
- isoleucine is preferable because of its high activity of increasing the ratio of trimer / tetramer antibody.
- the multimeric IgA type recombinant antibody of the present embodiment may be a mixture of a dimer, a trimer and a tetramer antibody. Moreover, the monomer may mix.
- the multimeric IgA type recombinant antibody of this embodiment has a tetramer content of 20 mol% or more of the total IgA.
- the content of the tetramer is preferably 30 mol% or more of the total IgA, more preferably 40 mol% or more, further preferably 50 mol% or more, and preferably 60 mol% or more. Particularly preferred.
- the ratio of the monomer, dimer, trimer / tetramer antibody in the multimeric IgA type recombinant antibody can be measured by size exclusion chromatography, for example, as described later in Examples. . As will be described later in the Examples, the trimer and tetramer peaks may not be separated in the measurement by size exclusion chromatography.
- the multimeric IgA type recombinant antibody of the present embodiment has a trimer / tetramer (trimer or tetramer) content of 20 mol% or more of the total IgA. Is preferably 30 mol% or more, more preferably 40 mol% or more, particularly preferably 50 mol% or more, and most preferably 60 mol% or more.
- the dimer IgA has a molecular weight of 300 to 400 kDa.
- Trimer IgA has a molecular weight of 500 to 600 kDa.
- Tetrameric IgA has a molecular weight of 700 to 800 kDa.
- the molecular weight of IgA can be measured, for example, by mass spectrometry under mild ion introduction conditions achieved by lowering the degree of vacuum in the vicinity of the ion intake portion.
- the antibody may be a chimera of an IgA type antibody and a non-IgA type antibody.
- the IgA type antibody means an antibody having an amino acid sequence derived from at least a part of the IgA type antibody.
- an IgA type antibody can be said to be a protein that reacts with a general anti-IgA polyclonal antibody.
- multimeric antibodies have higher antigen-binding or neutralizing activity against antigens than monomeric antibodies.
- the present invention provides a method for quantifying a component of a multimeric IgA antibody comprising a step of performing mass spectrometry using a peptide having an amino acid sequence set forth in SEQ ID NOs: 70 to 97 as an internal standard.
- a stable isotope-labeled amino acid is produced by adding a stable isotope-labeled amino acid to IgA antibody-secreting cells by the method and culturing, this can also be used as an internal standard.
- the multimeric IgA type antibody is preferably human type.
- the above-mentioned components may be IgA1 antibody heavy chain, IgA2 antibody heavy chain, ⁇ -type antibody light chain, ⁇ -type antibody light chain, J chain, and SC.
- the present invention provides a standard for quantifying a component of a multimeric IgA antibody, comprising a peptide set having the amino acid sequences set forth in SEQ ID NOs: 70 to 97.
- the present invention provides a medicament containing a multimeric IgA type recombinant antibody as an active ingredient.
- the medicament of this embodiment is preferably used for treatment or prevention of infectious diseases. Infectious diseases include those caused by pathogens such as parasites, bacteria, fungi, viruses, and abnormal prions.
- IgA The main antibody in mucus and secretions covering the mucosal epithelium is IgA, and although it functions as the forefront of biological defense mechanisms in mucosal infections, IgA has not been put into practical use as an antibody drug It is.
- the inventors have conducted research on the development of a nasal inactivated whole particle influenza vaccine characterized by a safe and simple inoculation using an influenza virus inactivated whole particle antigen as a next generation influenza vaccine. So far, we have obtained good results in basic research using animals and clinical research that recruited healthy adult volunteers, and we are entering the stage of clinical development with an eye toward practical application.
- the inventors found that there are multimeric antibodies larger than dimers among IgA antibodies that play an important role in protecting against viral infection on the respiratory mucosa of humans vaccinated with nasal inactivated influenza vaccine And found that influenza virus neutralizing activity is higher than that of monomeric and dimeric antibodies.
- the inventors succeeded for the first time in efficiently producing a multimeric IgA type recombinant antibody, and the multimeric IgA type recombinant antibody is a monomer, a dimeric antibody. It was revealed that it has higher influenza virus neutralization activity and HA protein binding activity.
- the medicament of this embodiment is used for mucosal infections such as influenza virus infection, RS virus infection, severe acute respiratory syndrome (SARS), Middle East respiratory syndrome (MERS), acquired immune deficiency syndrome (AIDS), and the like. It is useful as a therapeutic or prophylactic agent.
- mucosal infections such as influenza virus infection, RS virus infection, severe acute respiratory syndrome (SARS), Middle East respiratory syndrome (MERS), acquired immune deficiency syndrome (AIDS), and the like. It is useful as a therapeutic or prophylactic agent.
- the multimeric IgA type recombinant antibody can bind to and neutralize influenza virus, RS virus and the like in a small amount.
- the medicament of the present embodiment can be applied as a respiratory administration type antibody medicament as a prophylactic or therapeutic agent for infectious diseases, an in vitro diagnostic agent, an antibody for research use, and the like.
- the medicament of this embodiment may be administered for the purpose of prevention to subjects with a high risk of infection due to the above-mentioned infectious disease-causing virus.
- it may be administered for the purpose of treatment and prevention of virus spread to patients who have suffered from the above infection.
- the medicament of this embodiment can be formulated and administered in dosage forms such as powders and liquids.
- dosage forms such as powders and liquids.
- a thickener that is usually contained in nasal drops for allergic rhinitis that is already on the market may be added to the medicament of this embodiment.
- the medicament of this embodiment can be administered by spraying on the nasal mucosa, by inhalation into the lower respiratory tract using a nebulizer, or the like.
- Intranasal vaccination with an inactivated whole influenza virus vaccine induces strong antibody responses in serum and nasal mucus of healthy adults., Hummmunacc. , 1962-1970, 2013., and can be carried out in the same manner as the nasal whole particle inactivated influenza vaccine.
- the therapeutic or prophylactic agent of the present embodiment is administered on the nasal mucosa by spraying, for example, 250 ⁇ L of one nose may be sprayed on both nostrils.
- the amount of antibody administered may be several hundred ⁇ g to several mg per inoculation (500 ⁇ L).
- a spray device used for nasal inactivated whole-particle influenza vaccine may be used. Further, it may be sprayed from 2 (morning and evening) to 4 times (once every 6 hours) per day. Examples of the administration period include one week.
- the medicine of this embodiment when the medicine of this embodiment is administered by inhalation into the lower respiratory tract, for example, a commonly used aerosol type inhaler may be used.
- the amount of inhaled antibody may be, for example, several mg to several tens mg per inhalation. Further, it may be inhaled about twice a day (morning and evening). Examples of the administration period include one week.
- the medicine of this embodiment may be for humans, for example, domestic animals such as horses, cows, goats, sheep, pigs; pets such as dogs and cats; primates such as chimpanzees, gorillas, cynomolgus monkeys, etc.
- domestic animals such as horses, cows, goats, sheep, pigs
- pets such as dogs and cats
- primates such as chimpanzees, gorillas, cynomolgus monkeys, etc.
- rodents such as mice, rats and guinea pigs may be used.
- the IgA heavy chain, IgA light chain, J chain, and secretory component protein have an amino acid sequence derived from the target animal (subject) It is preferable that the animal type
- the target animal type means that the constant regions of the IgA heavy chain and IgA light chain encoding the multimeric antibody have the amino acid sequences of the constant regions of the target animal IgA heavy chain and IgA light chain.
- the J chain and the secretory component protein are the target animal type means that the J chain and the secretory component protein have the amino acid sequence of the target animal's J chain and the secretory component protein.
- the amino acid sequences of the IgA heavy chain, IgA light chain, J chain, and secretory component protein may contain mutations as long as they have the desired antigen-binding activity.
- the present invention provides a method for producing a multimeric IgA type antibody, comprising co-expressing IgA type antibody heavy chain protein, antibody light chain protein, antibody J chain protein and secretory component protein in one cell.
- a method for producing a multimeric IgA type antibody comprising co-expressing IgA type antibody heavy chain protein, antibody light chain protein, antibody J chain protein and secretory component protein in one cell.
- the multimeric IgA type antibody may be a recombinant antibody.
- pIgR expressed on the cell membrane on the basement membrane side of mucosal epithelial cells is specific to the J chain protein in the dimer / multimeric Ig molecule produced by plasma cells existing in the lamina intestinal. It is recognized and bound to be taken up into the cell, transported to the apical side after being taken into the epithelial cell, then cleaved between the extracellular domain part of pIgR and the transmembrane part, It is released from the cell to the mucosal surface.
- IgA type antibody heavy chain protein, antibody light chain protein, antibody J chain protein and secretory component protein are not co-expressed in one cell.
- the inventors unexpectedly co-expressed the secretory component protein together with IgA type antibody heavy chain protein, antibody light chain protein, and antibody J chain protein in one cell, thereby surprisingly producing multimeric IgA type.
- the first successful production of antibodies The production method of this embodiment makes it possible to industrially apply multimeric IgA-type antibodies.
- the antibody J chain protein and the secretory component protein preferably have an amino acid sequence derived from the subject animal species.
- the antibody J chain protein and the secretory component protein may be a chimera having amino acid sequences derived from a plurality of animal species.
- the IgA-type antibody heavy chain protein in the present embodiment may be an antibody that was originally non-IgA type converted to IgA type by gene recombination.
- the non-IgA type antibody is not particularly limited, and examples thereof include non-IgA type human antibodies, non-human mammal-derived antibodies, rodent-derived antibodies, and avian-derived antibodies.
- the class of the antibody is not particularly limited, and examples thereof include IgG type, IgM type, IgE type, IgD type, IgY type antibody and the like.
- variable region of an IgG type antibody can be converted to an IgA type by transplanting it to an IgA type antibody backbone framework.
- IgA type antibody backbone framework Alternatively, by converting only the CDR region of an IgG type antibody into the CDR region of an IgA type antibody, it can be converted to an IgA type.
- the subclass of IgA may be IgA1 type or IgA2 type.
- Examples of host cells include mammalian cells and insect cells.
- Mammalian cells include 293F cells, CHO cells, CHO YA7 cells and the like, and CHO YA7 cells are particularly preferable.
- Insect cells include the Sf9 cell line and the Sf21 cell line.
- the present invention includes a step of co-expressing IgA-type antibody heavy chain protein, antibody light chain protein, antibody J chain protein, secretory component protein, p180 protein and SF3b4 protein in one cell.
- a method for producing an IgA-type antibody is provided.
- the multimeric IgA type antibody may be a recombinant antibody.
- the amino acid sequence of the p180 protein is shown in SEQ ID NO: 98, and the base sequence encoding the p180 protein is shown in SEQ ID NO: 99.
- the amino acid sequence of the SF3b4 protein is shown in SEQ ID NO: 100, and the base sequence encoding the SF3b4 protein is shown in SEQ ID NO: 101.
- polysome formation can be promoted on the endoplasmic reticulum membrane in the cell.
- the polysome is one in which one molecule of mRNA is bound to a plurality of ribosomes present on the intracellular endoplasmic reticulum membrane.
- Suitable host cells are the same as those described above.
- Human p180 protein is a type I transmembrane protein present in the endoplasmic reticulum, and consists of a short cytoplasmic region, a transmembrane region, and a cytoplasmic domain portion.
- p180 protein having any number of repeats can be used, but p180 protein having 54 repeats is preferred.
- the C-terminal side of the p180 protein forms a coiled-coil domain, and the 945th to 1540th parts of SEQ ID NO: 98 are important for promoting polysome formation by interaction with the SF3b4 protein (UenoUT. et al., Regulation of polysome assembly on the endoplasmic reticulum by a coiled-coil protein, p180, Nucleic Acids Res., 40 (7), 3006-3017, 2012.).
- the production method of this embodiment makes it possible to efficiently produce multimeric IgA antibodies.
- the mRNA precursor transcribed from intracellular DNA is converted to mature mRNA by removing the intron portion by splicing.
- This process is carried out by a large complex consisting of spliceosome, a small nuclear RNA (snRNA) -protein.
- snRNA small nuclear RNA
- nucleic acid encoding a target protein when expressed in a cell in which the expression of p180 protein and SF3b4 protein is enhanced, the result is that mRNA transcribed from the nucleic acid interacts with SF3b4 protein or p180 protein, or Interaction with SF3b4 protein, followed by interaction between the coiled-coil domain of p180 protein and SF3b4 protein promotes the localization of mRNA to the endoplasmic reticulum, and the ability to synthesize or secrete the target protein in this cell Is enhanced.
- the p180 protein may be a protein consisting of the amino acid sequence represented by SEQ ID NO: 98, and the sequence identity with the amino acid sequence represented by SEQ ID NO: 98 is 80% or more, preferably 85% or more, More preferably 90% or more, particularly preferably 95% or more of the amino acid sequence, which may be a protein having a function of promoting polysome formation on the endoplasmic reticulum membrane in the cell, and is represented by SEQ ID NO: 98 It comprises an amino acid sequence having a sequence similarity with an amino acid sequence of 80% or more, preferably 85% or more, more preferably 90% or more, and particularly preferably 95% or more, and is capable of forming a polysome on the endoplasmic reticulum membrane in a cell.
- a base sequence that may be a protein having a function to promote and can hybridize with a nucleic acid comprising a base sequence complementary to the base sequence shown in SEQ ID NO: 99 under stringent conditions And a protein having a function of promoting polysome formation on the endoplasmic reticulum membrane in the cell.
- the p180 protein may be derived from mammals other than humans.
- the SF3b4 protein may be a protein consisting of the amino acid sequence represented by SEQ ID NO: 100, and the sequence identity with the amino acid sequence represented by SEQ ID NO: 100 is 80% or more, preferably 85% or more, more preferably 90% or more, particularly preferably 95% or more of an amino acid sequence equivalent to the protein consisting of the amino acid sequence shown in SEQ ID NO: 100, or the ability to synthesize or secrete a protein as a target product when expressed in a cell May be a protein having a function of enhancing the sequence, and the sequence similarity with the amino acid sequence shown in SEQ ID NO: 100 is 80% or more, preferably 85% or more, more preferably 90% or more, particularly preferably 95% or more Is the same as the protein consisting of the amino acid sequence shown in SEQ ID NO: 100.
- a protein having a function of enhancing the ability to synthesize or secrete a target protein when expressed in a cell In the amino acid sequence shown in SEQ ID NO: 100, one or several amino acids are A function consisting of a deleted, substituted or added amino acid sequence, which is equivalent to the protein consisting of the amino acid sequence shown in SEQ ID NO: 100, and enhances the ability to synthesize or secrete the target protein when expressed in cells.
- the present invention relates to IgA-type antibody heavy chain protein, antibody light chain protein, antibody J chain protein, and secretory component protein, CHO YA7 cell line (name of depositary institution: National Institute of Technology and Evaluation Technology Patent) Microorganism Deposit Center (NPMD), name of depositary institution: Japan Room 2-5-8, Kazusa Kamashichi, Kisarazu City, Chiba Prefecture, 292-0818, Japan, Date of deposit: February 13, 2013, Deposit number: NITE BP-01535
- the multimeric IgA type antibody may be a recombinant antibody.
- the CHO YA7 cell line is a cell line established by the inventors and constitutively expresses p180 protein and SF3b4 protein in the cell. Therefore, the production method of this embodiment makes it possible to efficiently produce multimeric IgA antibodies.
- the step of co-expressing IgA type antibody heavy chain protein, antibody light chain protein, antibody J chain protein and secretory component protein in one cell comprises IgA type antibody heavy chain protein, antibody light chain
- An expression vector for expressing a chain protein, antibody J chain protein and secretory component protein is introduced into the cell.
- the expression vector is downstream of the promoter, and is an IgA-type antibody heavy chain protein, antibody light chain
- Provided is a method for producing a multimeric IgA type antibody having a cis-element that an RNA binding protein recognizes, binds to, or interacts with, upstream of a start codon of a nucleic acid encoding a chain protein, antibody J chain protein or secretory component protein .
- the multimeric IgA type antibody may be a recombinant antibody.
- the cis-element is a sequence motif GAN 1- (X) n -ACN 2 (n is an integer of 3 to 6, and N 1 and N 2 are each independently any one of A, T, G, and C It is preferable that the base sequence comprises 1 to several nucleotide sequences.
- the inventors When the cis-element is present in the 5 ′ untranslated region of the mature mRNA, the inventors bind the cis-element to the RRM protein that recognizes the cis-element, and is a small site that synthesizes secreted proteins. It has been found that there is a function of enhancing the transport / localization of mRNA on the membrane of the endoplasmic reticulum and increasing the translation efficiency.
- the production method of this embodiment makes it possible to produce a multimeric IgA type recombinant antibody more efficiently.
- Suitable host cells are the same as those described above.
- the cis-element may be the base sequence shown in any of SEQ ID NOs: 21 to 23, and 1 to several bases are deleted in the base sequence shown in any of SEQ ID NOs: 21 to 23 May be a base sequence that is substituted or added and that is recognized, bound, or interacts with an RNA-binding protein, and has an identity of 80 to the base sequence shown in any of SEQ ID NOs: 21 to 23 % Or more, preferably 85% or more, more preferably 90% or more, particularly preferably 95% or more, and the RNA binding protein may be a base sequence that recognizes, binds or interacts, Hybridizes under stringent conditions with a nucleic acid consisting of a base sequence complementary to the nucleic acid consisting of the base sequence shown in any of SEQ ID NOs: 21 to 23
- the base sequence capable Rukoto and recognition RNA binding protein or may be a nucleic acid fragment consisting of binding or interacting nucleotide sequence.
- the cis-element may be a
- the number of bases that may be deleted, substituted or added is, for example, 1 to 30, for example 1 to 15, for example 1 to 10, for example 1 to 5.
- the number of amino acids that may be deleted, substituted or added is, for example, 2 to 40, for example 2 to 30, for example 2 to 20, for example 2 to 10, for example 2 to 7, for example 2 ⁇ 5, such as 5, such as 4, such as 3, such as 2, for example.
- amino acid sequences refers to the identity of two target amino acid sequences, and matches in the optimal alignment of amino acid sequences created using mathematical algorithms known in the art. Expressed by the percentage of amino acid residues.
- Amino acid sequence identity can be determined by visual inspection and mathematical calculations, homology search programs well known to those skilled in the art (eg BLAST, FASTA), sequence alignment programs (eg ClustalW), genetic information processing software (For example, GENETYX (registered trademark)) or the like.
- amino acid sequence similarity refers to the similarity between two target amino acid sequences, and matches in the optimal alignment of amino acid sequences created using mathematical algorithms known in the art. Expressed by the percentage of amino acid residues and amino acid residues showing similarity. The similarity of amino acid sequences is indicated by the relationship between amino acid residues whose physicochemical properties are similar to each other.
- aromatic amino acids Phe, Tyr, Trp
- hydrophobic amino acids Al, Leu, Ile, Val
- Gly, Pro, Met, Phe, Trp aliphatic amino acids
- Al amino acids
- polar amino acids Al, Arg, His
- acidic amino acids Asp, Glu
- Amino acids containing hydroxyl groups Ser, Thr
- amino acids with small side chains Gly, Ala, Ser, Thr, Met
- Similarity of amino acid sequences can be determined by visual inspection and mathematical calculation as well as identity, homology search programs well known to those skilled in the art (for example, BLAST, PSI-BLAST, HMMER), genetic information processing software (For example, GENETYX (registered trademark)) or the like.
- the identity of base sequences refers to the identity of two target base sequences, and matches in the optimal alignment of base sequences created using a mathematical algorithm known in the art. Expressed by the percentage of nucleic acid residues.
- a typical computer program for calculating the identity of a base sequence is the Wisconsin package of the Genetics Computer Group (GCG; Madison, Wis.), Version 10.0 program “GAP” (Devereux, et al., 1984) , Nucl. Acids Res., 12: 387), BLASTN program, version available from the National Library of Medicine website: http://www.ncbi.nlm.nih.gov/blast/bl2seq/bls.html 2.2.7 or UW-BLAST 2.0 algorithm.
- “under stringent conditions” includes, for example, the method described in Molecular Cloning-A LABORATORY MANUUAL SECOND EDITION (Sambrook et al., Cold Spring Harbor Laboratory Press).
- 5 ⁇ SSC composition of 20 ⁇ SSC: 3M sodium chloride, 0.3M citric acid solution, pH 7.0
- 0.1 wt% N-lauroyl sarcosine 0.02 wt% SDS
- the conditions for hybridization can be exemplified by incubation for several hours to overnight at 55-70 ° C. in a hybridization buffer comprising a nucleic acid hybridization probe and 50% formamide.
- the washing buffer used for washing after incubation is preferably a 0.1 ⁇ SSC solution containing 0.1 wt% SDS, more preferably a 0.1 ⁇ SSC solution containing 0.1 wt% SDS.
- the cis-element described above is a multimeric IgA-type antibody as long as it is contained in any one or more of IgA-type antibody heavy chain protein, antibody light chain protein, antibody J-chain protein, and secretory component protein expression vectors. The effect of improving the expression level or secretion level is obtained.
- the method of this embodiment makes it possible to produce a multimeric antibody more efficiently 26 to 35 times or more than the conventional method. Furthermore, the monoclonal tetramer antibody (rS-qIgA) against influenza virus produced by this method has an increased antigen binding activity compared to the monomer antibody, and showed a neutralization activity of 100 times or more at maximum. .
- the present invention provides a method for improving the antigen-binding activity or neutralizing activity of the antibody, comprising the step of converting the antibody into multimeric IgA.
- the step of converting the antibody into multimeric IgA comprises co-expressing IgA type antibody heavy chain protein having the heavy chain variable region of the antibody, light chain protein of the antibody, antibody J chain protein and secretory component protein in one cell. It is preferable to include the process to make.
- the antigen-binding activity of an antibody is the ability of the antibody to bind to the target molecule itself, but the specificity is the ability to bind only to the target molecule. In order to ensure high specificity, It is necessary to set a site that exists only as a recognition site (epitope). Although these antigen-binding activities and specificities are generated by the diversity of the variable region sequences of antibody molecules, it is difficult to predict the antigen-binding activity and specificity from the variable region sequences to the target molecules, and antigen binding activities In addition, it is necessary to clone a highly specific antibody from a living body such as a mouse, a randomly produced antibody-producing cell or antibody gene, or an artificially generated random variable region library by some method. In addition, since both specificity and antigen binding activity are independently dependent on the sequence of the variable region, without changing the specificity of the obtained antibody, that is, without changing the recognized epitope, It is very difficult to artificially improve the antigen binding activity.
- the inventors co-expressed the IgA-type antibody heavy chain protein, the light chain protein of the antibody, the antibody J chain protein, and the secretory component protein in one cell, thereby producing a multimer. It was found that an IgA type antibody can be produced. Suitable host cells are the same as those described above.
- the antigen-binding activity or neutralizing activity of the antibody can be improved by multimerizing the monomer antibody. For example, it is possible to improve the virus neutralization activity per mole of the antibody by 100 times or more compared to the monomer antibody by multimerizing the monomer antibody as described later.
- the antibody to be used for improving antigen-binding activity or neutralizing activity examples include human antibodies, antibodies derived from non-human mammals, rodent-derived antibodies, and avian-derived antibodies.
- the class of the antibody is not particularly limited, and examples thereof include IgG type, IgM type, IgE type, IgA type, IgD type, IgY type antibody and the like.
- the method of this embodiment can be applied by binding the variable region of the antibody to the constant region of an IgA type antibody and converting it to an IgA type by gene recombination.
- an IgG type monoclonal antibody variable region can be converted to an IgA type monoclonal antibody by transplanting it to an IgA type antibody backbone framework.
- the antibody may be converted to the IgA type by transplanting only the CDR region of the antibody to be improved in antigen binding activity into the CDR region of the IgA type antibody.
- the subclass of IgA may be IgA1 type or IgA2 type.
- the method of the present embodiment is very versatile as a technique capable of converting an IgG type monoclonal antibody having a variable region that recognizes the same epitope into a high binding type and a high activity type. Therefore, it can be widely applied to products using monoclonal antibodies such as pharmaceutical antibodies, diagnostic antibodies used in immunochromatography, immunohistochemistry, ELISA, and other research-use antibodies.
- the position of the amino acid residue in the amino acid sequence of the constant region of the IgA1 antibody is referred to as Liu YS et al. Complete covalent structure of a human IgA1 immunoglobulin. Science. 1976; 193: 1017-20. Number was used.
- the position of the amino acid residue in the constant region of the IgA2 antibody allotype (IgA2m1, IgA2m2, IgA2 (n))
- the constant region of each IgA2 antibody allotype and the constant region of IgA1 antibody are aligned, and the corresponding IgA1
- the number of amino acid residues of the antibody was used.
- the amino acid sequence 224-236 (STPPTPSPSTPPT) of the IgA1 antibody was omitted because there was no corresponding amino acid in each IgA2 antibody allotype.
- Example 1 Isolation of antibody variable region gene induced in human by nasal inactivated whole particle influenza vaccine and production of monoclonal IgG1 antibody using the same] (Vaccination and collection of peripheral blood lymphocytes) A highly pathogenic avian influenza virus A / H5N1 inactivated whole particle vaccine was intranasally inoculated into healthy adults twice at intervals of 3 weeks (one nose 250 ⁇ L, total 500 ⁇ L). As the vaccine, an inactivated whole particle vaccine containing 45 ⁇ g of hemagglutinin (HA) was used. Seven days after the second vaccination, peripheral blood was collected, and peripheral blood lymphocytes were collected using a blood cell separation solution Lymphoprep TM (AXIS-SHIELD).
- Lymphoprep TM AXIS-SHIELD
- Isolation of antibody-producing plasma cells and cDNA preparation Isolation of antibody-producing plasma cells induced in peripheral blood by nasal vaccination was performed using FACS Aria (BD Bioscience). Cell populations of cell surface markers CD2 ⁇ , CD3 ⁇ , CD4 ⁇ , CD10 ⁇ , CD20 ⁇ , IgD ⁇ , CD19 low , CD27 high and CD38 high were used as antibody-producing plasma cells and separated and collected as single cells. Single antibody-producing plasma cells were collected in a 96-well plate with 9 ⁇ L of sterile water containing 45 ng of carrier RNA in each well. cDNA preparation is described in T.W. This was performed according to the report of Tiller et al.
- the isotype of the antibody heavy chain isolated in each well was determined by Real-time PCR.
- TaqMan probes and primers were prepared for IgG, IgA and IgM constant regions.
- TaqMan probes for IgG, IgA and IgM were labeled with FAM, HEX and Cy5, respectively.
- QuantTect Multiplex PCR NoROX Master Mix Qiagen was used, and analysis was performed using LightCycler 480 (Roche).
- Amplification and sequencing of antibody variable region genes is described in T.W. This was performed according to the report of Tiller et al. (J Immunol Methods, 329, 112-24, 2008). Specifically, 11.5 ⁇ L of HotStarTaq DNA polymerase (Qiagen), dNTP mix and a primer set amplification mixture for each antibody variable region gene were added to 1 ⁇ L of the prepared cDNA, and the first PCR reaction was performed. It was. Further, a second PCR reaction was performed using a primer set designed further inside each gene contained in 1 ⁇ L of the PCR product. In any PCR reaction, amplification was performed under the conditions of 95 ° C. for 15 minutes, (94 ° C. for 30 seconds, 58 ° C. for 20 seconds, 72 ° C. for 60 seconds) ⁇ 43 cycles, 72 ° C. for 2 minutes. Moreover, the base sequence analysis (sequence) of the PCR product was performed by a conventional method.
- PCR of the antibody variable region gene was performed using PrimeSTAR (registered trademark) MAX DNA Polymerase (TaKaRa) according to the instructions.
- the above-mentioned first PCR product was used as a template, and a primer suitable for the locus to be amplified was selected based on the result of the second PCR product sequencing described above.
- PCR conditions were 98 ° C. for 10 seconds, 55 ° C. for 5 seconds, and 72 ° C. for 10 seconds for 25 cycles.
- Purification of the PCR product was performed using MonoFas (registered trademark) DNA purification kit I (GL Sciences Inc.) according to the instructions and eluted in 30 ⁇ L of Buffer C.
- the purified PCR product is 30 ⁇ L in total volume using AgeI-HF (all chains) and SalI-HF (heavy chain), BsiWI ( ⁇ chain) or XhoI ( ⁇ chain) (above, NEB) under appropriate conditions. And treated with a restriction enzyme.
- the expression vectors ⁇ 1, HC (heavy chain), ⁇ LC ( ⁇ chain), and ⁇ LC ( ⁇ chain) corresponding to each chain were treated with the same enzyme combination.
- the restriction enzyme product was purified using MonoFas (registered trademark) DNA purification kit I (GL Sciences Inc.) according to the instructions, and eluted into 20 ⁇ L of Buffer C.
- Ligation of DNA subjected to restriction enzyme treatment was performed in a total volume of 10 ⁇ L using a DNA Ligation Kit ⁇ Mighty Mix> (TaKaRa) according to the instructions.
- the ligation product was transformed into 10 ⁇ L of Competent Quick DH5 ⁇ (TOYOBO) by heating at 42 ° C.
- Plasmid extraction was performed using PureYield TM Plasmid Miniprep System (Promega) according to the instructions.
- Expi293F cells subcultured and maintained was 3.0 ⁇ 10 6 cells / mL or more, the survival rate was 95% or more, and the cells were not aggregated.
- the number of cells was adjusted to 2.9 ⁇ 10 6 cells / mL using Expi293 Expression medium kept at 37 ° C.
- the prepared cell suspension was transferred to a disposable Erlenmeyer flask with a vent filter cap, 25.5 mL, returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2, and cultured with shaking at 125 rpm.
- 30 ⁇ g of plasmid DNA 15 ⁇ g each of IgG heavy chain and light chain was added.
- Expifectamine 293 Reagent 80 ⁇ L of Expifectamine 293 Reagent was added to 1.5 mL of Opti-MEM I medium prepared separately. After allowing to stand at room temperature for 5 minutes, the entire amount of the DNA solution was added to the Expifectamine solution and allowed to stand at room temperature for 20 to 30 minutes. After adding the transfection mix to the cells, the cells were returned to the incubator for cell culture adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. 16-18 hours after transfection, 150 ⁇ L Expifectamine 293 Transfection Enhancer 1 and 1.5 mL Expifectamine 293 Transfection Enhancer 2 were added. The cells were returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. Supernatants were collected 6 days after transfection.
- the column was re-equilibrated with 10 column volumes of PBS.
- the antibody concentration was measured by NanoDrop (Thermo Scientific).
- the antibody was concentrated using Amicon (registered trademark) Ultra Centrifugal Filter Devices (Millipore) according to the instructions.
- the buffer was exchanged into PB (pH 7.4) according to the instruction using Zeba Desert Spin Columns (Thermo Scientific). The concentration after buffer exchange was measured by NanoDrop.
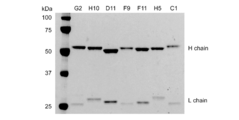
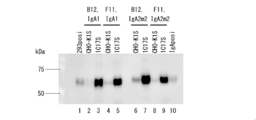
- FIG. 1 is a photograph showing the results of SDS-PAGE. It was confirmed that antibody clones G2, H10, D11, F9, F11, H5, and C1 were IgG1 type. It was confirmed that antibody clones G2, H10, D11, F9, F11, H5, and C1 were IgG1 type. Variations in expression levels were seen from clone to clone.
- IgA type recombinant antibody was prepared by the following method. Not only the IgG1 antibody prepared in Experimental Example 1, but all antibodies with known sequences can be converted to IgA by the following method.
- the IgA1 antibody constant region gene was amplified using pFUSE-CHIg-hA1 (InvivoGen) as a template. PCR conditions were 98 ° C. for 10 seconds, 55 ° C. for 5 seconds, and 72 ° C. for 30 seconds for 30 cycles.
- the PCR product was purified using MonoFas (registered trademark) DNA purification kit I (GL Sciences Inc.) according to the instructions, and eluted in 30 ⁇ L of Buffer C.
- the purified PCR product and ⁇ 1 HC plasmid were treated with a restriction enzyme at 37 ° C. using XhoI and HindIII-HF (NEB) in a total volume of 30 ⁇ L.
- the restriction enzyme-treated product was purified using MonoFas (registered trademark) DNA purification kit I (GL Sciences Inc.) according to the instructions and eluted into 20 ⁇ L of Buffer C. Ligation of the restriction enzyme-treated DNA was performed in a total volume of 10 ⁇ L using a DNA Ligation Kit ⁇ Mighty Mix> (TaKaRa) according to the instructions. The ligation product was transformed to 10 ⁇ L of Competent Quick DH5 ⁇ by heating at 42 ° C.
- Plasmid extraction was performed according to the instructions using PureYield (TM) Plasmid Miniprep System (Promega).
- the sequence reaction of the extracted plasmid was performed according to the instructions using BigDye (registered trademark) Terminator v3.1 Cycle Sequencing Kit (Life Technologies).
- the reaction product was purified using a BigDye XT terminator (trademark) Kit (Life Technologies) according to the instructions, and sequenced with Applied Biosystems 3130 Genetic Analyzer (Life Technologies).
- PCR of the antibody variable region gene was performed using PrimeSTAR (registered trademark) MAX DNA Polymerase (TaKaRa) according to the instructions.
- PrimeSTAR registered trademark
- MAX DNA Polymerase TaKaRa
- the reverse primer was changed to that for the ⁇ 1 HC expression vector, and PCR conditions were 98 ° C. for 10 seconds, 55 ° C. for 5 seconds, and 72 ° C. for 5 seconds for 25 cycles.
- the PCR product was purified using MonoFas (registered trademark) DNA purification kit I (GL Sciences Inc.) according to the instructions, and eluted in 30 ⁇ L of Buffer C.
- the purified PCR product and ⁇ 1 HC expression vector were treated with restriction enzymes under appropriate conditions using AgeI-HF and NheI-HF (NEB) in a total volume of 30 ⁇ L.
- the restriction enzyme product was purified using MonoFas (registered trademark) DNA purification kit I (GL Sciences Inc.) according to the instructions, and eluted into 20 ⁇ L of Buffer C.
- Ligation of the restriction enzyme-treated DNA was performed in a total volume of 10 ⁇ L using a DNA Ligation Kit ⁇ Mighty Mix> (TaKaRa) according to the instructions.
- the ligation product was transformed into 10 ⁇ L of Competent Quick DH5 ⁇ (TOYOBO) by heating at 42 ° C.
- Plasmid extraction was performed according to the instructions using PureYield (TM) Plasmid Miniprep System (Promega).
- the sequence reaction of the extracted plasmid was performed according to the instructions using BigDye (registered trademark) Terminator v3.1 Cycle Sequencing Kit (Life Technologies).
- the reaction product was purified using a BigDye XT terminator (trademark) Kit (Life Technologies) according to the instructions and sequenced with Applied Biosystems 3130 Genetic Analyzer (Life Technologies). As a result of sequencing, it was confirmed to be the same as the antibody gene cloned into the ⁇ 1 HC expression vector.
- Expi293F cells subcultured and maintained was 3.0 ⁇ 10 6 cells / mL or more, the survival rate was 95% or more, and the cells were not aggregated.
- the number of cells was adjusted to 2.9 ⁇ 10 6 cells / mL using Expi293 Expression medium kept at 37 ° C.
- the prepared cell suspension was transferred to a disposable Erlenmeyer flask with a vent filter cap, 25.5 mL, returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2, and cultured with shaking at 125 rpm.
- 30 ⁇ g of plasmid DNA 15 ⁇ g each of IgA1 heavy chain and light chain was added.
- Expifectamine 293 Reagent 80 ⁇ L of Expifectamine 293 Reagent was added to 1.5 mL of Opti-MEM I medium prepared separately. After allowing to stand at room temperature for 5 minutes, the entire amount of the DNA solution was added to the Expifectamine solution and allowed to stand at room temperature for 20 to 30 minutes. After adding the transfection mix to the cells, the cells were returned to the incubator for cell culture adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. 16-18 hours after transfection, 150 ⁇ L Expifectamine 293 Transfection Enhancer 1 and 1.5 mL Expifectamine 293 Transfection Enhancer 2 were added. The cells were returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. Supernatants were collected 6 days after transfection.
- the column was re-equilibrated with 10 column volumes of PBS.
- the antibody concentration was measured by NanoDrop (Thermo Scientific).
- the antibody was concentrated using Amicon (registered trademark) Ultra Centrifugal Filter Devices (Millipore) according to the instructions.
- the buffer was exchanged into PB (pH 7.4) according to the instruction using Zeba Desert Spin Columns (Thermo Scientific). The concentration after buffer exchange was measured by NanoDrop.
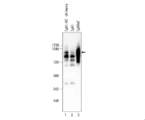
- FIG. 2 is a photograph showing the results of SDS-PAGE. It was confirmed that the antibody clones B12, D11, F9, F11 and H5 were IgA1 type.
- the antibody J chain uses an artificial gene synthesis service (operon biotechnology) to add a XhoI cleavage site and a Kozak sequence to the 5 ′ side of the coding region (CDS) of the J chain (GenBank accession no. NM — 144646).
- An artificial gene SEQ ID NO: 24 with a NotI cleavage site added on the side was synthesized.
- the J chain gene was treated with restriction enzymes under appropriate conditions using XhoI and NotI-HF (NEB).
- pCXSN vector a mammalian cell expression vector composed of CMV promoter and SV40 polyA treated with the same restriction enzyme to obtain an antibody J chain expression plasmid, pCXSN-hJC.
- the density of Expi293F cells subcultured and maintained was 3.0 ⁇ 10 6 cells / mL or more, the survival rate was 95% or more, and the cells were not aggregated.
- the number of cells was adjusted to 2.9 ⁇ 10 6 cells / mL using Expi293 Expression medium kept at 37 ° C.
- the prepared cell suspension was transferred to a disposable Erlenmeyer flask with a vent filter cap, 25.5 mL, returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2, and cultured with shaking at 125 rpm.
- Plasmid DNA (J chain expression group: IgA1 heavy chain and light chain 12 ⁇ g each, J chain 6 ⁇ g; J chain non-expression group: IgA1 heavy chain and light chain 15 ⁇ g each) was added to 1.5 mL of Opti-MEM I medium.
- Expifectamine 293 Reagent 80 ⁇ L of Expifectamine 293 Reagent was added to 1.5 mL of Opti-MEM I medium prepared separately. After allowing to stand at room temperature for 5 minutes, the entire amount of the DNA solution was added to the Expifectamine solution and allowed to stand at room temperature for 20 to 30 minutes. After adding the transfection mix to the cells, the cells were returned to the incubator for cell culture adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. 16-18 hours after transfection, 150 ⁇ L Expifectamine 293 Transfection Enhancer 1 and 1.5 mL Expifectamine 293 Transfection Enhancer 2 were added. The cells were returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. Supernatants were collected 6 days after transfection.
- the column was re-equilibrated with 10 column volumes of PBS.
- the antibody concentration was measured by NanoDrop (Thermo Scientific).
- the antibody was concentrated using Amicon (registered trademark) Ultra Centrifugal Filter Devices (Millipore) according to the instructions.
- the buffer was exchanged into PB (pH 7.4) according to the instruction using Zeba Desert Spin Columns (Thermo Scientific). The concentration after buffer exchange was measured by NanoDrop.
- FIG. 3 is a photograph showing the results of BN-PAGE.
- J chain non-expression group -J
- a band of monomeric IgA recombinant antibody was confirmed, whereas in the J chain expression group (+ J), in addition to the monomer band, A body IgA type recombinant antibody band was confirmed.
- Overlap PCR using PrimeSTAR (registered trademark) MAX DNA Polymerase (TaKaRa) was performed using the two synthesized DNA fragments as templates, and the gene fragment encoding the secretory component (SC) was amplified (SEQ ID NO: 25) , Restriction enzyme treatment with XhoI and NotI, and cloning into pCXSN vector to obtain pCXSN-hSC-HisTag which is an expression plasmid of secretory component.
- inverse PCR was performed using pCXSN-hSC-HisTag as a template to prepare pCXSN-hSC that expresses only the secreted component from which the HindIII cleavage site, thrombin cleavage site, and histidine tag added to the 3 ′ side were removed.
- Multimeric antibodies could be produced using either plasmid as the secretory component.
- Expi293F cells subcultured and maintained was 3.0 ⁇ 10 6 cells / mL or more, the survival rate was 95% or more, and the cells were not aggregated.
- the number of cells was adjusted to 2.9 ⁇ 10 6 cells / mL using Expi293 Expression medium kept at 37 ° C.
- the prepared cell suspension was transferred to a disposable Erlenmeyer flask with a vent filter cap, 25.5 mL, returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2, and cultured with shaking at 125 rpm.
- Plasmid DNA IgA1 heavy chain and light chain 12 ⁇ g each, J chain and secretory component 6 ⁇ g each
- Opti-MEM I medium Plasmid DNA
- Expifectamine 293 Reagent 80 ⁇ L of Expifectamine 293 Reagent was added to 1.5 mL of Opti-MEM I medium prepared separately. After allowing to stand at room temperature for 5 minutes, the entire amount of the DNA solution was added to the Expifectamine solution and allowed to stand at room temperature for 20 to 30 minutes. After adding the transfection mix to the cells, the cells were returned to the incubator for cell culture adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. 16-18 hours after transfection, 150 ⁇ L Expifectamine 293 Transfection Enhancer 1 and 1.5 mL Expifectamine 293 Transfection Enhancer 2 were added. The cells were returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. Supernatants were collected 6 days after transfection.
- the column was re-equilibrated with 10 column volumes of PBS.
- the antibody concentration was measured by NanoDrop (Thermo Scientific).
- the antibody was concentrated using Amicon (registered trademark) Ultra Centrifugal Filter Devices (Millipore) according to the instructions.
- the buffer was exchanged into PB (pH 7.4) according to the instruction using Zeba Desert Spin Columns (Thermo Scientific). The concentration after buffer exchange was measured by NanoDrop.
- the concentrated multimeric IgA1-type antibody was fractionated by gel filtration chromatography using AKTA explorer 10 (GE Healthcare). Superose 6 10/300 GL (GE Healthcare) was used for the column.
- DPBS Dynamic Phosphate Buffered Saline
- Flow rate 0.5 mL / min, column equilibration 1.5 column volume, elution 0.5 mL (total 1.5 column volumes).
- the eluted sample was collected for each fraction, and the fraction containing IgA was concentrated using Amicon (registered trademark) Ultra Centrifugal Filter Devices (Millipore).
- Amicon registered trademark
- Ultra Centrifugal Filter Devices Millipore
- the buffer was exchanged with PB (pH 7.4) according to the instruction manual using Zeba Desert Spin Columns (Thermo Scientific). The concentration after buffer exchange was measured by NanoDrop.
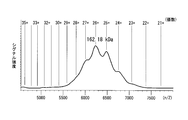
- FIG. 4 is a photograph showing a gel filtration chromatography chart and the results of BN-PAGE of antibodies contained in fractions 19 to 32. It was shown that the expressed IgA1-type antibody contains a tetramer, a dimer, and a monomer. This result is the first result of producing a tetrameric IgA type recombinant antibody.
- Example 5 Examination of influenza virus neutralizing activity
- a human IgG1 antibody against influenza virus was prepared, and in Experimental Example 2, an IgA1 antibody having the same variable region as that of the IgG1 antibody was prepared.
- multimeric IgA1-type antibodies were prepared. Using these antibodies, influenza virus neutralizing activity was examined.
- the neutralizing activity of each prepared antibody was quantified by measuring the minimum neutralization concentration by microneutralization test.
- a 2-fold serial dilution series of the sample was prepared, mixed with 100 TCID 50 (100 times the amount of 50% tissue culture infectious amount) virus solution, and incubated at 37 ° C. for 30 minutes. Then, this mixed solution is added to MDCK cells (canine kidney-derived cell line) and cultured for 4 days, and the value obtained by dividing the sample concentration by the maximum sample dilution factor at which cytopathic effects due to influenza virus cannot be confirmed under a microscope.
- MDCK cells canine kidney-derived cell line
- Influenza virus neutralization activity was measured using monomeric IgG1 type antibodies and monomeric IgA1 type antibodies of antibody clones G2, H10, D11, F9, F11, H5, B12 and C1.
- a / H5N1 strain (clade 2.1) and A / H1N1 strain were used for the virus.
- Table 1 shows the results. The lower the minimum neutralization concentration, the higher the virus neutralization activity. It was revealed that the antibodies of clones F11 and H5 showed good neutralizing activity against both H5N1 and H1N1 strains.
- Influenza virus neutralization activity of dimeric IgA1-type recombinant antibody Influenza virus neutralization activity was measured in the same manner as described above using the monomeric IgA1-type antibody and dimeric IgA1-type antibody of antibody clones D11, F9, F11, H5 and B12. A / H5N1 strain (clade 2.1) and A / H1N1 strain were used for the virus.
- Table 2 shows the results. The lower the minimum neutralization concentration, the higher the virus neutralization activity. Despite the same structure of the variable region of the antibody, the dimer antibody tended to have higher virus neutralizing activity than the monomeric antibody.
- Influenza virus neutralizing activity of tetrameric IgA1-type recombinant antibody Influenza virus neutralizing activity was measured in the same manner as described above using monomeric IgA1-type antibody, dimeric IgA1-type antibody and tetrameric IgA1-type antibody of antibody clones F9, F11 and H5.
- a / H5N1 strain (clade 2.1) was used as the virus. Table 3 shows the results. The lower the minimum neutralization concentration, the higher the virus neutralization activity.
- the dimeric antibody has higher virus neutralizing activity than the monomeric antibody, and the tetrameric antibody is more active in the virus than the dimeric antibody. There was a tendency for high sum activity.
- FIG. 5A and FIG. 5B show the activity of neutralizing ability of monomeric IgG1-type antibody, monomeric IgA1-type antibody, dimeric IgA1-type antibody and tetrameric IgA1-type antibody.
- the neutralization activity ratio of the monomeric IgA1-type antibody, the dimeric IgA1-type antibody and the tetrameric IgA1-type antibody when the virus neutralizing activity of the monomeric IgG1-type antibody is set to 1 is shown.
- a / H5N1 strain (clade 2.1) was used as the virus.
- FIG. 5A shows neutralization per unit protein amount of a monomeric IgA1-type antibody, a dimeric IgA1-type antibody and a tetrameric IgA1-type antibody when the virus neutralizing activity of the monomeric IgG1-type antibody is 1. It is a graph which shows an activity ratio.
- FIG. 5B shows neutralization per unit molecule number of monomeric IgA1-type antibody, dimeric IgA1-type antibody, and tetrameric IgA1-type antibody when the virus neutralizing activity of the monomeric IgG1-type antibody is 1. It is a graph which shows an activity ratio.
- Example 7 Examination of antigen binding activity
- Production of a recombinant viral glycoprotein expression vector and expression and purification of the recombinant viral glycoprotein were carried out in the same manner as in Experimental Example 9 described later.
- the antigen-binding activity against influenza virus HA protein was examined using monomeric IgA1-type antibody, dimeric IgA1-type antibody, and tetrameric IgA1-type antibody.
- recombinant HA protein (derived from A / H5N1 strain, 1 ⁇ g / mL) was added to a 96-well half plate and left standing at 4 ° C. overnight, followed by blocking.
- Antibody clone F11 is a dimer and a tetramer for the virus (A / H5N1 (clade 2.1)) of the same clade (clade 2.1) as the vaccine, and more binding activity than the monomer
- the strongest antigen-binding activity of the tetramer was seen against HA derived from a virus (A / H5N1 (clade 1)) separate from the vaccine. It was shown that the antigen-binding activity of the clones having the antigen-binding activity was improved by multimerization.
- Example 8 Effect of enhancing expression of multimeric antibody by using CHO YA7 cells and cis-element
- the cis-element # 1 shown in SEQ ID NO: 22 is introduced downstream of the promoter of the antibody J chain protein expression plasmid pCXSN-hJC and upstream of the start codon of the J chain protein, and pCXSN-cis # 1-hJC Got. It was confirmed by gene sequence analysis that the orientation of the cis-element was the correct insertion direction.
- cis-element # 2 shown in SEQ ID NO: 23 was introduced downstream of the promoter of pCXSN-hSC, which is the above-described secretory component expression plasmid, and upstream of the start codon of the secretory component protein, and pCXSN-cis # 2-hSC. Got. It was confirmed by gene sequence analysis that the orientation of the cis-element was the correct insertion direction.
- IgA1 antibody heavy chain expression plasmid constructed in Experimental Example 2 the light chain full-length expression for 1 ⁇ 10 5 each of CHO YA7 cells and control CHO cells, which are cell lines that co-express p180 protein and SF3b4 protein 0.5 ⁇ g of each of the four expression plasmids, plasmids pCXSN-cis # 1-hJC and pCXSN-cis # 2-hSC, was transfected by the lipofection method.
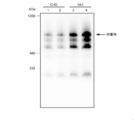
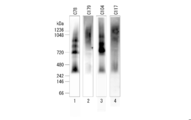
- FIG. 6 is a photograph showing the results of BN-PAGE.
- Lanes 1 and 2 are the results of expressing multimeric IgA type antibodies in CHO cells
- lanes 3 and 4 are the results of expressing multimeric IgA type antibodies in CHO YA7 cells.
- Lanes 1 and 3 are the results of using an antibody J chain protein expression plasmid having no cis-element and a secretory component expression plasmid having no cis-element as controls, and lanes 2 and 4 are cis.
- the arrow indicates the band of tetrameric IgA type antibody.
- Influenza virus HA protein and RS virus F protein ⁇ FP deletion mutant of amino acid sequence 137-146, McLellan JS, et al., Structure of respiratory syncytial virus fusion glycoprotein in the postfusion conformation reveals preservation of neutralizing epitopes., J. Virol. 85 (15), 7788-7796, 2011). Trimer formation sequence and purification tag coding sequence (bacteriophage T4 fibrin trimer) on the C-terminal side of the amino acid sequence encoding the extracellular region.
- Expi293F cells subcultured and maintained was 3.0 ⁇ 10 6 cells / mL or more, the survival rate was 95% or more, and the cells were not aggregated.
- the number of cells was adjusted to 2.9 ⁇ 10 6 cells / mL using Expi293 Expression medium kept at 37 ° C.
- the prepared cell suspension was transferred to a disposable Erlenmeyer flask with a vent filter cap, 25.5 mL, returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2, and cultured with shaking at 125 rpm. 30 ⁇ g of plasmid DNA was added to 1.5 mL of Opti-MEM I medium.
- Expifectamine 293 Reagent 80 ⁇ L of Expifectamine 293 Reagent was added to 1.5 mL of Opti-MEM I medium prepared separately. After allowing to stand at room temperature for 5 minutes, the entire amount of the DNA solution was added to the Expifectamine solution and allowed to stand at room temperature for 20 to 30 minutes. After adding the transfection mix to the cells, the cells were returned to the incubator for cell culture adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. 16-18 hours after transfection, 150 ⁇ L Expifectamine 293 Transfection Enhancer 1 and 1.5 mL Expifectamine 293 Transfection Enhancer 2 were added. The cells were returned to a cell culture incubator adjusted to 37 ° C. and 8% CO 2 and cultured with shaking at 125 rpm. Supernatants were collected 4-6 days after transfection.
- Buffer W 100 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, pH 8.0
- Buffer E 100 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 2.5 mM desthiobiotin, pH 8)
- Buffer R 100 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 1 mM HABA, pH 8.0
- the column was equilibrated with 2 CV Buffer W, the sample was loaded, washed 5 times with 1 CV Buffer W, and the viral glycoprotein was eluted with 3 CV Buffer E.
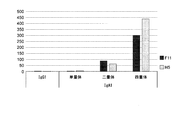
- ELISA against RS virus F protein The reactivity of multimers of anti-RS virus F protein antibody to RS virus F protein was examined by ELISA. First, 50 ⁇ L of recombinant F protein (1 ⁇ g / mL) was immobilized on a 96-well half plate at 4 ° C. overnight, followed by blocking. Subsequently, a 2-fold serial dilution series of the anti-RS virus F protein antibody sample was reacted at room temperature for 2 hours. Subsequently, after washing with PBST, HRP-labeled goat anti-human IgA antibody (30000 times) (BETHYL LABORATORIES) was reacted at room temperature for 1 hour.
- FIG. 7 shows the value of OD 450 nm (average value ⁇ standard deviation) at an anti-RS virus F protein antibody sample concentration of 1 ⁇ g / mL.
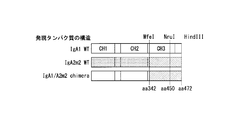
- IgA2m1 a gene encoding the heavy chain constant region of IgA2m1 ( ⁇ 2m1 HC, accession number: J00221)
- a gene encoding the heavy chain constant region of IgA2m2 a gene encoding the heavy chain constant region of IgA2m2 ( ⁇ 2m2 HC, accession numbers: M60192 and AJ012264)
- IgA2 Obtain each nucleotide sequence of the gene ( ⁇ 2 (n) HC, accession number: S71043) encoding the heavy chain constant region of (n) based on sequences registered in IMGT / LIGM-DB, which is a public database Then, codon optimization was performed for humans, and artificial genes were synthesized.
- the NheI cleavage site was created by modifying the codon of the last amino acid alanine in the variable region and the first amino acid serine in the constant region.
- the synthetic sequence treated with NheI and HindIII was cloned into a vector obtained by treating the above ⁇ 1 HC with NheI and HindIII.
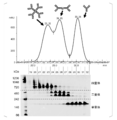
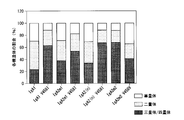
- FIG. 8A shows representative results of IgA1, IgA2m1, IgA2m2, and IgA2 (n) type antibodies in chromatograms obtained by size exclusion chromatography.
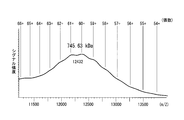
- FIG. 8B is a graph showing the results of calculating the peak area ratio of each of trimer / tetramer, dimer, and monomer antibody based on FIG. 8A. If the valley between peaks in the chromatogram of FIG. 8A or the separation of the peaks is unclear, infer the peak inflection point, draw a line perpendicular to the baseline, and divide each fraction into chromatogram and base The area surrounded by the line was calculated.
- IgA IgA1
- IgA2m1 IgA2m2
- IgA2 (n) IgA2
- the tendency of multimerization of IgA1 and each IgA2 allotype is different.
- the IgA2m2 type antibody was shown to have a high multimer-forming ability.
- ⁇ 1 ⁇ 2m2 HC chimeric expression vector (Preparation of ⁇ 1 ⁇ 2m2 HC chimeric expression vector) Using PrimeSTAR (registered trademark) MAX DNA Polymerase, the above-described ⁇ 1 HC expression vector was used as a template to PCR amplify the sequence from CH1 to CH2 of IgA1 HC ( ⁇ 1 segment: N-terminal to G342 of constant region). Furthermore, using the ⁇ 2m2 HC expression vector as a template, the sequence from the CH3 to the C terminus of the IgA2m2 HC ( ⁇ 2m2 segment: from N343 to the C terminus of the constant region) was PCR amplified.
- FIG. 9 is a graph showing the results. As a result, it was revealed that the formation of multimers was promoted when the sequence from CH3 to C-terminus of the IgA1 antibody heavy chain was replaced with an IgA2m2-derived sequence.
- Example 12 Analysis of region related to multimerization promoting activity of IgA2m2
- various expression vectors were constructed as follows.
- the above-mentioned anti-influenza virus antibody clone B12 IgA1 heavy chain expression plasmid (pIgA1H) and the anti-influenza virus antibody clone B12 IgA2m2 heavy chain expression plasmid (pIgA2m2H) were treated with restriction enzymes NheI and HindIII. Similarly, a fragment for ligation was prepared. Each fragment was ligated to prepare plasmids pIgA1H-NRE and pIgA2m2H-NRE.
- IgA1 / IgA2m2 chimeric heavy chain expression vector An IgA1 / IgA2m2 chimeric heavy chain expression vector was constructed as follows. That is, a fragment obtained by treating pIgA2m2H-NRE with MfeI and HindIII and pIgA1H-NRE treated with MfeI and HindIII were ligated, and the mutant pIgA1H in which the amino acid 342 to 472th region of the IgA1 heavy chain was replaced with IgA2m2 type / PIgA2m2-chimera was constructed.
- FIG. 10A is a schematic diagram showing the structure of each mutant.
- the cis # 1-hJC ORF and cis # 2-hSC ORF were excised from the pCXSN-cis # 1-hJC and pCXSN-cis # 2-hSC of Example 5 to construct a co-expression vector for J chain and SC.
- the cis # 1-hJC ORF and cis # 2-hSC ORF are ligated between the promoter in the expression unit composed of the human EF1 promoter and BGH polyA and polyA, respectively, and the expression vectors pEF-cis-hJC / Zeo and pEF / Each cis-hSC / Zeo was constructed.
- J NQ asparagine (N) to which an N-linked sugar chain binds are replaced with glutamine (Q)
- Q glutamine
- J NQ fragments encoding J NQ and SC NQ were prepared by artificial gene synthesis (Genscript) (SEQ ID NOs: 30, 31).
- J chain replaced N59 with Q.
- SC replaced all N83, N90, N135, N186, N421, N469, and N499 with Q.
- each synthetic fragment was treated with XhoI and NotI, and then ligated with pCXSN-cis # 1-hJC or pCXSN-cis # 2-hSC similarly treated with XhoI and NotI, respectively, and pCXSN-cis # 1-hJC NQ and pCXSN-cis # 2-hSC NQ was constructed.
- spERt technique a technique for enhancing gene expression by introducing an expression vector of expression enhancing two factors (p180 and SF3b4) into cells.
- pEF-cis-hJC / Zeo and pEF / cis-hSC / Zeo are introduced into CHO-K1 (1E26) cells, drug selection is performed with Zeocin (400 ⁇ g / mL), and hJC and hSC are stably expressed.
- CHO-K1 strain C23 was established.
- pCXSN-cis # 1-hJC NQ and pCXSN-cis # 2-hSC NQ were introduced into CHO-K1 (1E26) cells, drug selection was performed with Zeocin (400 ⁇ g / mL), and hJC NQ and hSC NQ CHO-K1 strain C452 was stably expressed.
- C23 cells were transfected with the anti-influenza virus antibody clone B12 light chain expression vector (pIgA-LC) and the anti-influenza virus antibody clone B12 IgA heavy chain expression vector by lipofection.
- the IgA heavy chain expression vector the above-mentioned three types of expression vectors (pIgA1H, pIgA2m2, pIgA1H / pIgA2m2-chimera) were used.
- IgA1 multimers and IgA2m2 multimers were detected in the culture supernatant of C23 cells, respectively (FIG. 10B, lanes 2 and 3), and IgA antibody heavy chain and light chain were detected in C23 cells. It was confirmed that IgA multimeric antibodies can be efficiently produced by gene transfer.
- IgA1 multimers and IgA2m2 multimers were also present in the culture supernatant in the same experiment conducted using the above-described anti-influenza virus antibody clones F11 and C452 cells, respectively. was detected.
- amino acid substitution mutant of Fc region An amino acid substitution mutant of the heavy chain constant region (Fc region) was prepared. More specifically, PrimeSTAR (registered trademark) MAX DNA using an ⁇ 2m2 HC expression vector as a template and primers modified in sequence so that five kinds of amino acid substitutions (Y411F, E428D, M451L, I458V, A467V) occur. Inverse PCR was performed using Polymerase.
- FIG. 11 is a graph showing the examination results. As a result, it was clarified that when the 458th I of the IgA2m2 antibody was replaced with V, the multimer decreased and the monomer ratio increased.
- FIG. 12 is a graph showing the examination results. As a result, it was revealed that multimerization was significantly promoted by substituting the 458th V of the IgA1 antibody with I.
- V458 mutant vector In order to construct a mutant in which the V of the 458th IgA1 heavy chain was substituted with various amino acids, pIgA1H-NRE was treated with restriction enzymes MfeI and HindIII to prepare a ligation fragment. Also, to prepare insert linkers, V458I (SEQ ID NOs: 32 and 33), V458A (SEQ ID NOs: 34 and 35), V458W (SEQ ID NOs: 36 and 37), V458C (SEQ ID NOs: 38 and 39), V458D (SEQ ID NO: 40).
- IgAl heavy chain expression vector pIgA1 -HC or IgA2m2 heavy chain expression vector pIgA2m2-HC was introduced by lipofection method, drug selection was performed with Zeocin (400 ⁇ g / mL) and puromycin (10 ⁇ g / mL), IgA1 heavy chain, light chain, hJC NQ , hSC CHO-K1 strain C104 stably expressing NQ, and, IgA2m2 heavy chain, light chain, HJC NQ, were established CHO-K1 strain C117 stably expressing hSCs NQ. The culture supernatant of each cell was analyzed by BN-PAGE and Western blotting.
- IgA crude purified fraction was subjected to solvent substitution with PBS ( ⁇ ) using Vivaspin20 (manufactured by GE) to prepare IgA1 and IgA2m2 multimeric antibody solutions.
- the antibody yield was about 0.404 mg for C78, 0.712 mg for C179, about 0.298 mg for C104, and about 0.179 mg for C117 per 80 mL of cell suspension.
- the antibody yield can be increased by 1 to 2 orders of magnitude by subjecting each cell to serum-free conditioning after suspension culture, studying the culture conditions, and the medium.
- the neutralizing activity of each prepared antibody was quantified by measuring the minimum neutralization concentration by microneutralization test.
- a 2-fold serial dilution series of antibody samples was prepared, mixed with 100 TCID 50 (100 times the amount of 50% tissue culture infectious amount) virus solution, and then incubated at 37 ° C. for 30 minutes. Thereafter, this mixed solution was added to MDCK cells and cultured for 5 days, and the value obtained by dividing the concentration of the sample by the maximum sample dilution ratio at which cytopathic effect by influenza virus could not be confirmed under a microscope was defined as the minimum neutralization concentration.
- vaccine production strain A / X-179A H1N1pdm09
- PR8 laboratory strain A / Puerto Rico / 8/34
- the cell lines C23, CHO-K1 and 1C17 were acclimatized stepwise to the serum-free suspension culture conditions, and cell lines C23S, CHO-K1S and 1C17S were established. Further, expression vectors pIgA1-cis-HC, pIgA2m2-cis-HC in which cis # 2 described in International Publication No.
- the expression vector pIgA-cis-LC was constructed by inserting cis # 1 into the pIgA-LC of anti-influenza virus antibody clone B12 and clone F11.
- FIG. 16B is a photograph showing the results.
- “293posi” represents a positive control of a multimeric IgA fraction prepared in 293expi cells in the same manner as in Example 4, and “IgAposi” represents an IgA standard.
- CHO-K1S cells were treated with the above-described pIgA-LC, pEF-cis-hJC / Zeo, pEF / cis-hSC / Zeo, and pIgA1-HC or pIgA2m2-HC. Transfection was performed by the lipofection method.
- 1C17S cells were transfected with the above-described pIgA-cis-LC, pEF-cis-hJC / Zeo, pEF / cis-hSC / Zeo, and pIgA1-cis-HC or pIgA2m2-cis-HC. .
- C23 cells were transfected with pIgA-cis-LC and pIgA1-cis-HC or pIgA2m2-cis-HC.
- multimeric IgA2m2 was remarkably increased in 1C17S cells and C23 cells.
- the expression of multimeric IgA2m2 was hardly detectable in CHO-K1S cells, which are the parent lines of these cells (FIG. 16C, lanes 4 to 6).
- the solvent of the IgA1 and IgA2m2 multimer fractions was replaced with 12.5 mM ammonium acetate using a desalting column (Thermo Fisher Scientific, “Zeba Spin Desaling Columns”, 7K MWCO, 0.5 mL).
- This sample was diluted 5 or 10 times with 50 mM or 100 mM ammonium acetate, and a quadrupole-time-of-flight mass spectrometer maXis II (manufactured by Bruker Daltonics) using a nano ion source (“TriVersa NanoMate” manufactured by Advion) And measured under the following conditions.
- Ionization ESI positive (High Mass option)
- ion spray voltage 1.4 to 1.8 kV
- ion source temperature 80 ° C.
- 17A to 17D show examples of tetramers measured under mild ion introduction conditions using the High Mass option, and ion peaks corresponding to monomers measured under normal ion introduction conditions. .
- a 154.87 kDa peak corresponding to the monomer of IgA2m2 was detected by ion introduction under conditions close to normal.
- the crosslinker (Bich C., et al., Reactivity and applications of new amine reactive cross-linkers for mass spectrometric detection of protein-protein complexes., Anal. Chem. 82, 172- 179, 2010)), stabilizing the complex confirmed an increase in tetramer and a concomitant decrease in monomer.
- the human nasal mucosa-derived IgA dimer fraction and the recombinant IgA1 multimer fraction were analyzed by LC-MS.
- Human nasal mucosa-derived IgA dimer fraction was described in SuzukiSuT., Et al., Relationship of the quaternary structure of human secretory IgA to neutralization of influenza virus, PNAS 112 (25), 7809-7817, 2015 Prepared in the same manner as the method.
- the recombinant IgA1 multimeric fraction was prepared in the same manner as in Experimental Example 4. For these samples, the subunit ratio of the IgA multimeric fraction was evaluated by trypsin digestion and using LC-MS as follows.
- Tris-HCl pH 7.6 was added to 10 ⁇ g of an IgA sample at a concentration of 100 mM, CaCl 2 at a concentration of 1 mM, and 0.05 nmol of a stable isotope-labeled peptide of each subunit as an internal standard.
- DTT manufactured by Wako
- iodoacetamide manufactured by Wako was added so as to be 25 mM, and an alkylation reaction of free SH groups was performed at room temperature for 30 minutes while shielding light.
- Solution A (0.1% formic acid) 98%, Solution B (100% acetonitrile) 2%; 2.1-6 minutes: Liquid A 98-50%, Liquid B 2-50%; 6.1-8 minutes: Solution A 10%, Solution B 90%; 8.1-10 minutes: Solution A 98%, Solution B 2%.
- the separated peptide component was detected by the mass spectrometer under the following conditions. Ionization: ESI positive, ion spray voltage: 4.5 kV, ion source temperature: 200 ° C.
- FIG. 19B shows the result of the recombinant IgA1 multimer fraction.
- the light chain ⁇ type was detected at a ratio close to 1: 1 with respect to the IgA1 heavy chain.
- the ratios of J chain and SC to IgA1 heavy chain were about 1: 7 and about 1: 8, respectively.
- a multimeric IgA gene recombinant antibody can be provided.
- the pharmaceutical which contains a multimeric IgA gene recombinant antibody as an active ingredient can be provided.
- mold antibody can be provided.
- the method of improving the antigen binding activity of an antibody can be provided.
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Medicinal Chemistry (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Veterinary Medicine (AREA)
- Microbiology (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Immunology (AREA)
- Oncology (AREA)
- Communicable Diseases (AREA)
- Virology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Mycology (AREA)
- Epidemiology (AREA)
- Bioinformatics & Computational Biology (AREA)
- Crystallography & Structural Chemistry (AREA)
- Tropical Medicine & Parasitology (AREA)
- Pulmonology (AREA)
- Peptides Or Proteins (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
Description
(1)多量体IgA型遺伝子組換え抗体。
(2)重鎖定常領域の第458番目のアミノ酸残基が疎水性アミノ酸に由来するアミノ酸残基である、(1)に記載の多量体IgA型遺伝子組換え抗体。
(3)四量体の含有量が全IgAの20モル%以上である、(1)又は(2)に記載の多量体IgA型遺伝子組換え抗体。
(4)(1)~(3)のいずれか一項に記載の多量体IgA型遺伝子組換え抗体を有効成分として含有する医薬。
(5)感染症の治療又は予防用である、(4)に記載の医薬。
(6)IgA型抗体重鎖タンパク質、抗体軽鎖タンパク質、抗体J鎖タンパク質及び分泌成分タンパク質を1つの細胞内で共発現させる工程を含む、多量体IgA型抗体の製造方法。
(7)前記IgA型抗体重鎖タンパク質は、遺伝子組換えによりIgG型からIgA型に変換されたものである、(6)に記載の製造方法。
(8)前記工程において、更にp180タンパク質及びSF3b4タンパク質を前記細胞内で共発現させる、(6)又は(7)に記載の製造方法。
(9)前記細胞がCHO YA7細胞株(受託番号NITE BP-01535)である、(6)~(8)のいずれかに記載の製造方法。
(10)前記工程は、IgA型抗体重鎖タンパク質、抗体軽鎖タンパク質、抗体J鎖タンパク質及び分泌成分タンパク質を発現するための発現ベクターを前記細胞内に導入することにより行われ、前記発現ベクターは、プロモーターの下流で、かつ、IgA型抗体重鎖タンパク質、抗体軽鎖タンパク質、抗体J鎖タンパク質又は分泌成分タンパク質をコードする核酸の開始コドンの上流に、RNA結合タンパク質が認識、結合又は相互作用するcis-エレメントを有する、(6)~(9)のいずれかに記載の製造方法。
(11)前記cis-エレメントが、配列モチーフGAN1-(X)n-ACN2(nは3~6の整数であり、N1及びN2は、それぞれ独立して、A、T、G、Cのいずれかである。)からなる塩基配列を1~数個含む、(10)に記載の製造方法。
(12)前記cis-エレメントが、
配列番号21~23のいずれかに示される塩基配列、
配列番号21~23のいずれかに示される塩基配列において、1~数個の塩基が欠失、置換又は付加されている塩基配列からなり、かつRNA結合タンパク質が認識、結合若しくは相互作用する塩基配列、
配列番号21~23のいずれかに示される塩基配列と同一性が80%以上である塩基配列からなり、かつ、RNA結合タンパク質が認識、結合若しくは相互作用する塩基配列、又は、
配列番号21~23のいずれかに示される塩基配列からなる核酸と相補的な塩基配列からなる核酸とストリンジェントな条件下でハイブリダイズすることができる塩基配列からなり、かつRNA結合タンパク質が認識、結合若しくは相互作用する塩基配列からなる、(10)又は(11)に記載の製造方法。
(13)抗体を多量体IgA型化する工程を含む、前記抗体の抗原結合活性又は中和活性を向上させる方法。
(14)前記抗体は、IgG型抗体である、(13)に記載の方法。
(15)前記工程が、前記抗体の重鎖可変領域を有するIgA型抗体重鎖タンパク質、前記抗体の軽鎖タンパク質、抗体J鎖タンパク質及び分泌成分タンパク質を1つの細胞内で共発現させる工程を含む、(13)又は(14)に記載の方法。
IgA型抗体に関連して、従来用いられている用語について以下に説明する。
IgAは、血清中では、主に単量体IgAとして分子量17万前後で存在しており、IgA1が主要構成要素である。
二量体IgAとは、粘膜固有層に存在する形質細胞が産生する二量体IgAであって、重鎖:軽鎖:J鎖の割合が4:4:1である分子を指す。IgA2が約半分程度存在する。
従来、ポリメリックIg受容体により認識される二量体以上のIgAを総称して、多量体IgAという場合が多い。すなわち、重鎖:軽鎖:J鎖の構成比が4:4:1の複合体である二量体IgAが主成分で、抗体J鎖タンパク質(Joining chain)によりIgA二分子が結合し、分泌成分タンパク質(secretory component、SCタンパク質)を含む場合と含まない場合の両方について「多量体IgA」という用語が使用されている。
粘膜上皮細胞の基底膜側の細胞膜上に発現しているpIgRは、免疫グロブリンスーパーファミリーに属するI型の膜貫通型蛋白質で、細胞外ドメイン部分、膜貫通部、細胞質内領域からなる。pIgRは粘膜固有層に存在する形質細胞が産生したJ鎖を含む二量体/多量体型Ig分子を特異的に認識して結合し、上皮細胞中への取り込み後も結合したままアピカル側へ輸送される。上皮細胞内から粘膜面への放出には、pIgRの細胞外ドメイン部分と膜貫通部との間で切断が必要で、上皮細胞のタンパク質分解酵素による切断後に内腔側粘膜層へS-IgAとして分泌される。このpIgRの細胞外ドメイン部分がIgA複合体の構成成分となる事実に由来して、分泌成分タンパク質(secretory component、SCタンパク質)とも呼ばれている。pIgRは五量体IgMの取り込みにも同様にして機能している。
粘膜上皮細胞より分泌され、SCタンパク質を含んだ二量体以上のIgA複合体を指し、Secretory dimeric IgA(S-dIgA)の場合は重鎖:軽鎖:J鎖:SCの構成比が4:4:1:1である。
粘膜固有層に存在する形質細胞が産生する多量体IgAの分子量等の性状解析は技術的に困難なため、分泌型多量体IgA形成の詳細な分子形成過程は不明である。すなわち、生体内で形質細胞が産生する主要な分泌型多量体IgAは、二量体、三量体、四量体等の多量体のうちどれが主であるか、多量体化は上皮細胞内で起こっているかどうか、粘膜部で生体防御に中心的役割を果たしているのがどの型かも不明である。生体内IgA及び組換え体でも二量体(440kDa付近)より大きいIgAも微量検出されているが、凝集体との鑑別はなされていない。
本明細書においては、分泌成分タンパク質(SC)を分子内に有する多量体抗体を分泌型抗体という。また、次のように表記する場合がある。
・単量体抗体=mIgA
・二量体抗体=dIgA
・分泌型二量体抗体=S-dIgA
・分泌型三量体抗体=S-tIgA
・分泌型四量体抗体=S-qIgA
・四量体以上の分泌型多量体抗体=S-pIgA
・組換え単量体抗体=rmIgA
・組換え二量体抗体=rdIgA
・組換え分泌型二量体抗体=recombinant S-dIgA(rS-dIgA)
・組換え分泌型三量体抗体=recombinant S-tIgA(rS-tIgA)
・組換え分泌型四量体抗体=recombinant S-qIgA(rS-qIgA)
・四量体以上の組換え分泌型多量体抗体=recombinant S-pIgA(rS-pIgA)
ここでは、分泌成分タンパク質(SC)を分子内に有する場合に「S-」を分子名の頭に付記する。分子内にSCを有さない抗体は「S-」を付記しない。また、分子の会合状態を、IgAの前に略称を付記することにより表記する。単量体は「m」、二量体は「d」、三量体は「t」、四量体は「q」、四量体以上の多量体は「p」を付記する。また、単量体抗体と組換え単量体抗体を特に区別なく「monomer」と表記する場合がある。また、二量体抗体と分泌型二量体抗体、組換え二量体抗体、組換え分泌型二量体抗体を特に区別なく「dimer」と表記する場合がある。また、分泌型三量体抗体と組換え分泌型三量体抗体を特に区別なく「trimer」と表記する場合がある。また、分泌型四量体抗体と組換え分泌型四量体抗体を特に区別なく「tetramer」と表記する場合がある。また、四量体以上の分泌型抗体と四量体以上の組換え分泌型多量体抗体を特に区別なく「polymer」と表記する場合がある。
1実施形態において、本発明は、多量体IgA型遺伝子組換え抗体を提供する。本実施形態の多量体IgA型遺伝子組換え抗体は、本来非IgA型であった抗体を遺伝子組換えによりIgA型に変換し、更に多量体化したものであってもよい。非IgA型抗体としては、特に制限されず、例えば、非IgA型の、ヒト抗体、非ヒト哺乳動物由来の抗体、げっ歯類由来の抗体、鳥類由来の抗体等が挙げられる。また、抗体のクラスも特に制限されず、例えば、IgG型、IgM型、IgE型、IgD型、IgY型抗体等が挙げられる。
1実施形態において、本発明は、多量体IgA型遺伝子組換え抗体を有効成分として含有する医薬を提供する。本実施形態の医薬は、感染症の治療又は予防用であることが好ましい。感染症としては、寄生虫、細菌、真菌、ウイルス、異常プリオン等の病原体によるものが挙げられる。
1実施形態において、本発明は、IgA型抗体重鎖タンパク質、抗体軽鎖タンパク質、抗体J鎖タンパク質及び分泌成分タンパク質を1つの細胞内で共発現させる工程を含む、多量体IgA型抗体の製造方法を提供する。多量体IgA型抗体は、遺伝子組換え抗体であってもよい。
1実施形態において、本発明は、抗体を多量体IgA型化する工程を含む、前記抗体の抗原結合活性又は中和活性を向上させる方法を提供する。
(ワクチン接種と末梢血リンパ球の回収)
高病原性トリインフルエンザウイルスA/H5N1の不活化全粒子ワクチンを健康成人へ3週間隔で2回経鼻接種(片鼻250μL、計500μL)した。ワクチンとしては、45μgのヘマグルチニン(HA)を含有する不活化全粒子ワクチンを使用した。2回目のワクチン接種から7日後に末梢血を回収し、血球分離溶液Lymphoprep(商標)(AXIS-SHIELD社)を用いて末梢血リンパ球を回収した。
経鼻ワクチン接種により末梢血中に誘導された抗体産生形質細胞の単離は、FACS Aria (BD Bioscience社)を用いて実施した。細胞表面マーカーCD2-、CD3-、CD4-、CD10-、CD20-、IgD-、CD19low、CD27highかつCD38highの細胞集団を抗体産生形質細胞とし、単一細胞として分離・回収した。単一抗体産生形質細胞は、各ウェルに45ngのキャリアRNAを含む滅菌水9μLを分注した96穴プレートに回収した。cDNA調製は、T. Tillerら(J Immunol Methods, 329, 112-24, 2008)の報告に則り実施した。具体的には、細胞を回収した各ウェルにSuperscript III RT(ライフテクノロジーズ社)、Randam Hexamer(ライフテクノロジーズ社)、RNaseOUT(ライフテクノロジーズ社)、dNTP mix(キアゲン社)を含む6μLの混合液を添加し、50℃50分、85℃5分の反応を行うことでcDNAを調製した。
調製したcDNAを2μL用いて、各ウェルに単離された抗体重鎖のアイソタイプをReal-time PCRにより決定した。IgG、IgAおよびIgMの各定常領域に対してTaqManプローブとプライマーを準備した。IgG、IgAおよびIgMに対するTaqManプローブは、それぞれFAM、HEXおよびCy5による標識とした。QuantiTect Multiplex PCR NoROX Master Mix(キアゲン社)を使用し、LightCycler480(ロシュ社)を用いて解析を行った。
抗体可変領域遺伝子の増幅は、T. Tillerら(J Immunol Methods, 329, 112-24, 2008)の報告に則り実施した。具体的には、調製したcDNA1μLに対して11.5μLのHotStarTaq DNA polymerase(キアゲン社)、dNTP mix及び各抗体可変領域遺伝子を増幅するプライマーセットの混合液を添加し、1回目のPCR反応を行った。更にこのPCR産物1μLに含まれる各遺伝子に対して更に内側に設計したプライマーセットを用いて2回目のPCR反応を行った。いずれのPCR反応においても、95℃15分、(94℃30秒、58℃20秒、72℃60秒)×43サイクル、72℃2分の条件で増幅を行った。また、PCR産物の塩基配列解析(シークエンス)を常法により行った。
抗体可変領域遺伝子のPCRはPrimeSTAR(登録商標)MAX DNA Polymerase(TaKaRa社)を使用して、説明書にしたがって実施した。鋳型として上述の1回目のPCR産物を使用し、プライマーとしては、上述の2回目のPCR産物のシークエンスの結果に基づいて、増幅する遺伝子座に適切なペアを選択した。PCR条件は98℃10秒、55℃5秒、72℃10秒で25サイクルとした。PCR産物の精製はMonoFas(登録商標)DNA精製キットI(ジーエルサイエンス社)を使用して、説明書にしたがって実施し、30μLのBuffer Cに溶出した。
遺伝子組換え抗体の作製にはExpi293(商標)Expression System(ライフテクノロジーズ社)を説明書にしたがって用いた。30mLの系を以下に例示する。
細胞のデブリを1000×g、10分間の遠心分離により取り除いた後、Millex-HV Filter Unit (ミリポア社)で上清の濾過を行った。遺伝子組換え抗体の精製はCaptureSelect(商標)human Fc affinity matrix(ライフテクノロジーズ社)を用いて、説明書にしたがって実施した。具体的には、カラムは10カラム容量のリン酸緩衝液(PBS)で平衡化し、サンプルをロードし、10カラム容量のPBSで洗浄し、5カラム容量の0.1M Glycine-HCl(pH3.0)により抗体を溶出し、1M Tris-HCl(pH9.0)で溶出液を中和した。カラムは10カラム容量のPBSで再平衡化した。抗体の濃度はNanoDrop(Thermo Scientific社)で測定した。抗体の濃縮はAmicon(登録商標)Ultra Centrifugal Filter Devices(ミリポア社)を用いて、説明書にしたがって実施した。Zeba Desalt Spin Columns(Thermo Scientific社)で、説明書にしたがってPB(pH7.4)にバッファー交換した。バッファー交換後の濃度はNanoDropで測定した。
精製した抗体の確認はNuPAGE(登録商標)Bis-Tris Gel(ライフテクノロジーズ社)を用いて、説明書にしたがってSDSポリアクリルアミドゲル電気泳動(SDS-PAGE)を実施した。図1はSDS-PAGEの結果を示す写真である。抗体クローンG2、H10、D11、F9、F11、H5及びC1が、IgG1型化されたことが確認された。抗体クローンG2、H10、D11、F9、F11、H5及びC1が、IgG1型化されたことが確認された。クローンによって発現量にばらつきが見られた。40mL培養系において、クローンB12は約40mg、クローンD11は約4mg、クローンF9は約1.5mg、クローンF11は約2.1mg、クローンH5は約1mgの抗体を作製することができた。
IgA型遺伝子組換え抗体を以下の方法により作製した。実験例1で作製したIgG1抗体に限らず、配列が既知の抗体は全て以下の方法でIgA型化することができる。
IgA1抗体定常領域遺伝子を含む発現ベクターα1 HCを作製した。IgA1抗体定常領域遺伝子のPCRは、PrimeSTAR(登録商標)MAX DNA Polymerase(TaKaRa社)を使用して、説明書にしたがって実施した。
抗体可変領域遺伝子のPCRはPrimeSTAR(登録商標)MAX DNA Polymerase(TaKaRa社)を使用して、説明書にしたがって実施した。γ1 HC発現ベクターへクローニングした抗体遺伝子を鋳型として、リバースプライマーをα1 HC発現ベクター用のものに変更し、PCR条件は98℃10秒、55℃5秒、72℃5秒で25サイクルとした。
遺伝子組換え抗体の作製にはExpi293(商標)Expression System(ライフテクノロジーズ社)を説明書にしたがって用いた。30mLの系を以下に例示する。
細胞のデブリを1000×g、10分間の遠心分離により取り除いた後、Millex-HV Filter Unit (ミリポア社)で上清の濾過を行った。遺伝子組換え抗体の精製はCaptureSelect(商標)human IgA affinity matrix(ライフテクノロジーズ社)を用いて、説明書にしたがって実施した。具体的には、カラムは10カラム容量のリン酸緩衝液(PBS)で平衡化し、サンプルをロードし、10カラム容量のPBSで洗浄し、5カラム容量の0.1M Glycine-HCl(pH3.0)により抗体を溶出し、1M Tris-HCl(pH9.0)で溶出液を中和した。カラムは10カラム容量のPBSで再平衡化した。抗体の濃度はNanoDrop(Thermo Scientific社)で測定した。抗体の濃縮はAmicon(登録商標)Ultra Centrifugal Filter Devices(ミリポア社)を用いて、説明書にしたがって実施した。Zeba Desalt Spin Columns(Thermo Scientific社)で、説明書にしたがってPB(pH7.4)にバッファー交換した。バッファー交換後の濃度はNanoDropで測定した。
精製した抗体の確認はNuPAGE(登録商標)Bis-Tris Gel(ライフテクノロジーズ社)を用いて、説明書にしたがってSDSポリアクリルアミドゲル電気泳動(SDS-PAGE)を実施した。図2はSDS-PAGEの結果を示す写真である。抗体クローンB12、D11、F9、F11及びH5が、IgA1型化されたことが確認された。
実験例2において作製したIgA1抗体発現コンストラクトを用いて、二量体IgA1型抗体を作製した。
抗体J鎖は人工遺伝子合成サービス(オペロン バイオテクノロジー)を利用して、J鎖(GenBank accession no.NM_144646)のコード領域(CDS)の5’側にXhoI切断サイト及びKozak配列を付加し、3’側にNotI切断サイトを付加した人工遺伝子(配列番号24)を合成した。J鎖遺伝子をXhoI及びNotI-HF(以上、NEB社)を用いて、適切な条件で制限酵素処理した。同一の制限酵素で処理したpCXSNベクター(CMVプロモーターとSV40 polyAから構成される哺乳類細胞発現用ベクター)にクロ-ニングし、抗体J鎖の発現プラスミドであるpCXSN-hJCを得た。
二量体IgA1型抗体の作製にはExpi293(商標)Expression System(ライフテクノロジーズ社)を説明書にしたがって用いた。30mLの系を以下に例示する。
細胞のデブリを1000×g、10分間の遠心分離により取り除いた後、Millex-HV Filter Unit (ミリポア社)で上清の濾過を行った。遺伝子組換え抗体の精製はCaptureSelect(商標)human IgA affinity matrix(ライフテクノロジーズ社)を用いて、説明書にしたがって実施した。具体的には、カラムは10カラム容量のリン酸緩衝液(PBS)で平衡化し、サンプルをロードし、10カラム容量のPBSで洗浄し、5カラム容量の0.1M Glycine-HCl(pH3.0)により抗体を溶出し、1M Tris-HCl(pH9.0)で溶出液を中和した。カラムは10カラム容量のPBSで再平衡化した。抗体の濃度はNanoDrop(Thermo Scientific社)で測定した。抗体の濃縮はAmicon(登録商標)Ultra Centrifugal Filter Devices(ミリポア社)を用いて、説明書にしたがって実施した。Zeba Desalt Spin Columns(Thermo Scientific社)で、説明書にしたがってPB(pH7.4)にバッファー交換した。バッファー交換後の濃度はNanoDropで測定した。
精製した抗体の確認は、未変性ポリアクリルアミドゲル電気泳動法(BN-PAGE、3-13%、Invitrogen社)により行った。図3はBN-PAGEの結果を示す写真である。J鎖非発現群(-J)では、単量体IgA型遺伝子組換え抗体のバンドが確認されたのに対し、J鎖発現群(+J)では、単量体のバンドに加えて、二量体IgA型遺伝子組換え抗体のバンドが確認された。
実験例2において作製したIgA1抗体発現コンストラクトを用いて、多量体IgA1型抗体を作製した。
GeneArt(登録商標)Strings(商標)DNA Fragments(ライフテクノロジーズ社)を利用して、polymeric immunoglobulin receptor(GenBank accession no. NM_002644)の185~2005残基の5’側にXhoI切断サイト及びKozak配列を付加し、3’側にHindIII切断サイト、thrombin切断サイト、ヒスチジンタグ及びNotI切断サイトを付加した人工DNAフラグメントを、中央付近で重複するように5’側フラグメントと3’側フラグメントの2本合成した。合成した2本のDNAフラグメントを鋳型に用いてPrimeSTAR(登録商標)MAX DNA Polymerase(TaKaRa社)を使用したoverlap PCRを行い、分泌成分(SC)をコードする遺伝子断片を増幅し(配列番号25)、XhoI及びNotIで制限酵素処理し、pCXSNベクターにクローニングし、分泌成分の発現プラスミドであるpCXSN-hSC-HisTagを得た。また、pCXSN-hSC-HisTagを鋳型としてインバースPCRを行い、3’側に付与したHindIII切断サイト、thrombin切断サイト、ヒスチジンタグを除去した分泌成分のみを発現するpCXSN-hSCを作製した。分泌成分としてどちらのプラスミドを用いても多量体抗体を作製することができた。
多量体IgA1型抗体の作製にはExpi293(商標)Expression System(ライフテクノロジーズ社)を説明書にしたがって用いた。30mLの系を以下に例示する。
細胞のデブリを1000×g、10分間の遠心分離により取り除いた後、Millex-HV Filter Unit (ミリポア社)で上清の濾過を行った。遺伝子組換え抗体の精製はCaptureSelect(商標)human IgA affinity matrix(ライフテクノロジーズ社)を用いて、説明書にしたがって実施した。具体的には、カラムは10カラム容量のリン酸緩衝液(PBS)で平衡化し、サンプルをロードし、10カラム容量のPBSで洗浄し、5カラム容量の0.1M Glycine-HCl(pH3.0)により抗体を溶出し、1M Tris-HCl(pH9.0)で溶出液を中和した。カラムは10カラム容量のPBSで再平衡化した。抗体の濃度はNanoDrop(Thermo Scientific社)で測定した。抗体の濃縮はAmicon(登録商標)Ultra Centrifugal Filter Devices(ミリポア社)を用いて、説明書にしたがって実施した。Zeba Desalt Spin Columns(Thermo Scientific社)で、説明書にしたがってPB(pH7.4)にバッファー交換した。バッファー交換後の濃度はNanoDropで測定した。
濃縮した多量体IgA1型抗体を、AKTA explorer10(GEヘルスケア社)を用いて、ゲル濾過クロマトグラフィーにより分画した。カラムには、Superose6 10/300 GL (GEヘルスケア社)を用いた。DPBS(Dulbecco’s Phosphate Buffered Saline)を以下のプロトコールで流した。流速0.5mL/分、カラム平衡化1.5カラム容量、溶出0.5mL(計1.5カラム容量)。
実験例1において、インフルエンザウイルスに対するヒトIgG1抗体を作製し、実験例2において、当該IgG1抗体と同一の可変領域を有するIgA1抗体を作製した。また、実験例3及び4において多量体IgA1型抗体を作製した。これらの抗体を用いて、インフルエンザウイルス中和活性を検討した。
抗体クローンG2、H10、D11、F9、F11、H5、B12及びC1の単量体IgG1型抗体及び単量体IgA1型抗体を用いて、インフルエンザウイルス中和活性を測定した。ウイルスにはA/H5N1株(clade 2.1)及びA/H1N1株を使用した。
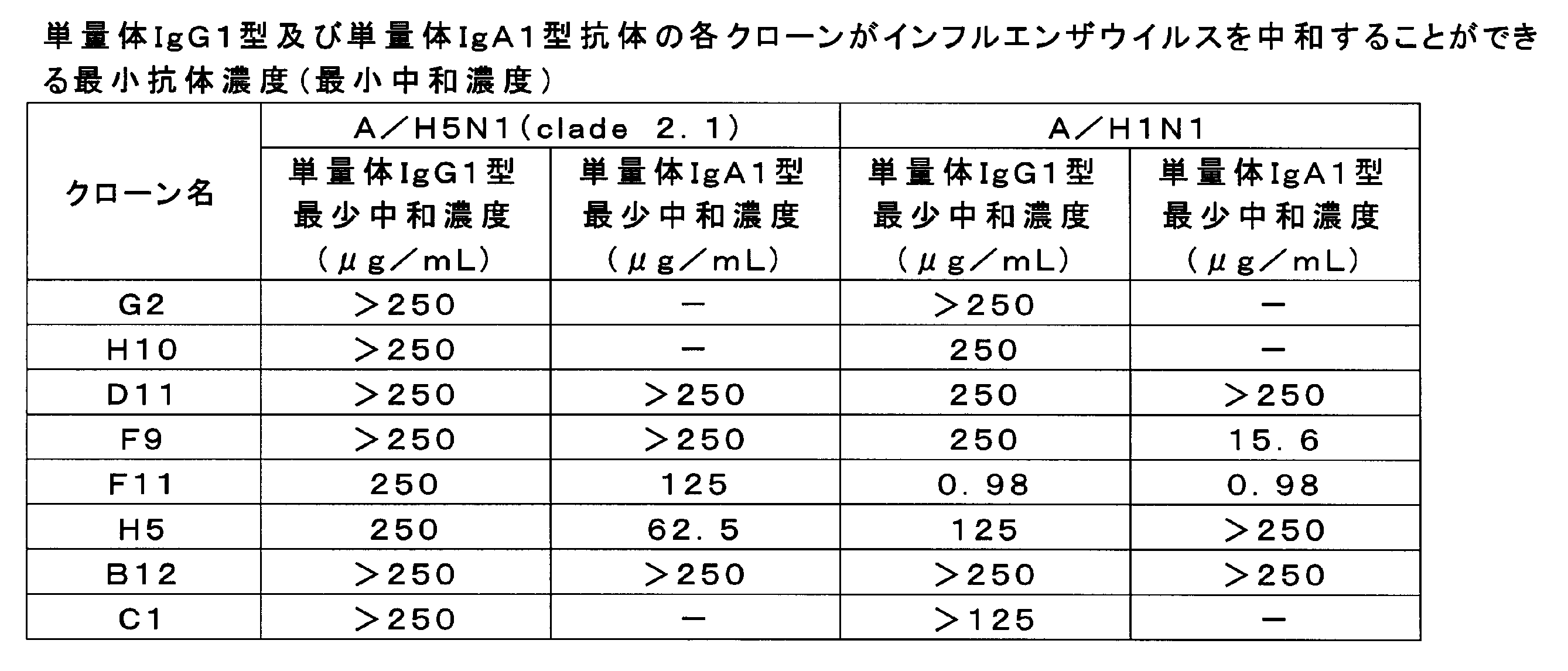
抗体クローンD11、F9、F11、H5及びB12の単量体IgA1型抗体及び二量体IgA1型抗体を用いて、上記と同様にしてインフルエンザウイルス中和活性を測定した。ウイルスにはA/H5N1株(clade 2.1)及びA/H1N1株を使用した。

抗体クローンF9、F11及びH5の単量体IgA1型抗体、二量体IgA1型抗体及び四量体IgA1型抗体を用いて、上記と同様にしてインフルエンザウイルス中和活性を測定した。ウイルスにはA/H5N1株(clade 2.1)を使用した。表3に結果を示す。最少中和濃度が低いほどウイルス中和活性が高いことを示す。

抗体クローンF11及びH5について、単量体IgG1型抗体、単量体IgA1型抗体、二量体IgA1型抗体及び四量体IgA1型抗体の中和能の活性比較のため、図5A及び図5Bに単量体IgG1型抗体のウイルス中和活性を1とした場合における、単量体IgA1型抗体、二量体IgA1型抗体及び四量体IgA1型抗体の中和活性比を示した。ウイルスにはA/H5N1株(clade 2.1)を使用した。
抗体クローンF11及びH5について、重鎖及び軽鎖の塩基配列及びアミノ酸配列をIGBLAST(http://www.ncbi.nlm.nih.gov/igblast/)にて解析した。また、相補性決定領域(complementarity determining region、CDR)をAndrew C.R. Martin博士らの方法(http://www.bioinf.org.uk/abs/)により決定した。配列表の配列番号と、各塩基配列及びアミノ酸配列との対応を表4に示す。クローンF11の軽鎖はκ鎖であった。また、クローンH5の軽鎖はλ鎖であった。

遺伝子組換えウイルス糖タンパク質発現ベクターの作製、遺伝子組換えウイルス糖タンパク質の発現と精製は後述する実験例9と同様の方法により行った。抗体クローンB12及びF11について、単量体IgA1型抗体、二量体IgA1型抗体及び四量体IgA1型抗体を用いて、インフルエンザウイルスHAタンパク質に対する抗原結合活性を検討した。


上述した抗体J鎖タンパク質の発現プラスミドであるpCXSN-hJCのプロモーターの下流かつJ鎖タンパク質の開始コドンの上流に、配列番号22に示すcis-エレメント#1を導入し、pCXSN-cis#1-hJCを得た。遺伝子配列解析によりcis-エレメントの向きが正しい挿入方向であることを確認した。
(抗RSウイルスFタンパク質抗体の作製)
公共データベースであるRCSB PDBに登録されている抗RSウイルスFタンパク質抗体のアミノ酸配列(PDB ID:2HWZ)の重鎖及び軽鎖可変領域のアミノ酸配列をヒト用にコドン最適化したアミノ酸配列をコードするDNA断片をそれぞれ人工合成した。合成した重鎖可変領域をコードするDNA断片を上述したα1 HCに、軽鎖 可変領域をコードするDNA断片を上述したκ LCにクローニングした。続いて、実験例4と同様にして抗RSウイルスFタンパク質抗体の多量体の発現及び精製を行った。
インフルエンザウイルスのHAタンパク質及びRSウイルスのFタンパク質ΔFP(アミノ酸配列137-146番の欠失変異体、McLellan J. S., et al., Structure of respiratory syncytial virus fusion glycoprotein in the postfusion conformation reveals preservation of neutralizing epitopes., J. Virol. 85(15), 7788-7796, 2011 を参照)の細胞外領域をコードするアミノ酸配列のC末端側に三量体形成配列及び精製用タグのコード配列(バクテリオファージT4フィブリチン三量体形成フォールドン配列、トロンビン切断部位(RSRSLVPRGSPGSGYIPEAPRDGQAYVRKDGEWVLLSTFL、配列番号26)、タンパク質精製用のStrep-tag(登録商標)II配列(WSHPQFEK、配列番号27)及び6×His tag配列(Stevens J. et al., Structure of the uncleaved human H1 hemagglutinin from the extinct 1918 influenza virus., Science 303, 1866-1870, 2004 を参照))を融合したアミノ酸配列をコードするDNA断片を合成し、哺乳類細胞発現用のベクターpCXSNにクローニングした。
遺伝子組換えウイルス糖タンパク質の作製にはExpi293(商標)Expression System(ライフテクノロジーズ社)を説明書にしたがって用いた。30mL系を以下に例示する。
まず、回収した上清中の細胞のデブリを1000×g、10分間の遠心分離により取り除いた。続いて、Millex-HV Filter Unit(ミリポア社)を使用して上清をろ過した。続いて、AKTA explorer10(GEヘルスケア社)を用いたアフィニティー精製によりウイルス糖タンパク質を精製した。カラムにはHisTrap excel(GEヘルスケア社)を用いた。より具体的には、平衡化溶液として20mMリン酸ナトリウム、0.5M NaCl、pH7.4を使用した。また、洗浄液として20mMリン酸ナトリウム、0.5M NaCl、10mMイミダゾール、pH7.4を使用した。また、溶出液として、20mMリン酸ナトリウム、0.5M NaCl、500mMイミダゾール、pH7.4を使用した。精製条件は、流速1mL/分、カラム平衡化10CV、カラム洗浄40CV、溶出1mL/フラクション(計50CV)、カラム再平衡化5CVとした。6xHis tagによる精製サンプルを、更に精製する場合にはStrep-tag(登録商標)/Strep-Tactin(登録商標)システムを使用した。Strep-Tactin(登録商標)Superflow(登録商標)(iba社)を用いて、説明書にしたがって精製した。より具体的には、まず、試薬として、Buffer W(100mM Tris-HCl、150mM NaCl、1mM EDTA、pH8.0)、Buffer E(100mM Tris-HCl、150mM NaCl、1mM EDTA、2.5mM desthiobiotin、pH8.0)、Buffer R(100mM Tris-HCl、150mM NaCl、1mM EDTA、1mM HABA、pH8.0)を準備した。続いて、カラムを2CVのBuffer Wで平衡化し、サンプルをロードし、1CVのBuffer Wで5回洗浄し、3CVのBuffer Eでウイルス糖タンパクを溶出した。そして、カラムを5CVのBuffer Rで3回再生洗浄し、4CVのBuffer Wで2回平衡化した。タンパク質を濃縮する場合には、Amicon(登録商標)Ultra Centrifugal Filter Devicesを用いて、説明書にしたがって濃縮した。精製されたタンパク質の濃度はNanoDrop(Thermo SCIENTIFIC社)を用いた吸光測定により測定した。
抗RSウイルスFタンパク質抗体の多量体のRSウイルスFタンパク質に対する反応性をELISAにより検討した。まず、96ウェルハーフプレートに50μLの遺伝子組換えFタンパク質(1μg/mL)を4℃で一晩固相化した後、ブロッキングを行った。続いて、抗RSウイルスFタンパク質抗体サンプルの2倍段階希釈系列を室温で2時間反応させた。続いて、PBSTを用いた洗浄後、HRP標識ヤギ抗ヒトIgA抗体(30000倍)(BETHYL LABORATORIES社)を室温で1時間反応させた。続いて、1-Step(商標)Ultra TMB-ELISA(Thermo SCIENTIFIC社)を用いて発色反応を行い、1M硫酸を加えて反応停止後、吸光度を測定した。抗RSウイルスFタンパク質抗体サンプル濃度1μg/mLにおけるOD 450nmの値(平均値±標準偏差)を図7に示した。その結果、抗RSウイルスFタンパク質抗体の抗原結合活性は、抗体が、二量体、四量体化すると有意に上昇することが確認された。
(α2m1 HC、α2m2 HC、α2(n) HC発現ベクターの作製)
IgA2のアロタイプの重鎖定常領域の遺伝子をクローニングした。具体的には、IgA2m1の重鎖定常領域をコードする遺伝子(α2m1 HC、アクセッション番号:J00221)、IgA2m2の重鎖定常領域をコードする遺伝子(α2m2 HC、アクセッション番号:M60192及びAJ012264)、IgA2(n)の重鎖定常領域をコードする遺伝子(α2(n) HC、アクセッション番号:S71043)の各塩基配列を、公共データベースであるIMGT/LIGM-DBに登録されている配列に基づいて入手し、ヒト用にコドン最適化を行い、人工遺伝子合成した。可変領域の最後のアミノ酸アラニンと定常領域の最初のアミノ酸セリンのコドンを改変してNheI切断サイトを作製した。NheI及びHindIIIで処理した合成配列を、上述のα1 HCをNheI及びHindIIIで処理したベクターにクローニングした。
PrimeSTAR(登録商標)MAX DNA Polymeraseを使用して、上述した抗インフルエンザウイルス抗体クローンB12の抗体可変領域遺伝子のPCRを行った。γ1 HC発現ベクターにクローニングした抗体遺伝子を鋳型とする場合には、α1 HC発現ベクターへの抗体遺伝子のクローニングと同様の方法を用いた。α1 HC発現ベクターにクローニングした抗体遺伝子を利用する場合には、AgeI及びNheIを用いた通常の制限酵素を用いたサブクローニング法を用いた。
コスモスピンフィルターH(ナカライテスク社)を用いてサンプルを前処理し、サイズ排除クロマトグラフィーにより分子サイズに基づいて分離し、抗体の構造を解析した。HPLCシステムとして、Agilent 1260 Infinity(Agilent Technologies社)を用いた。また、カラムにはAgilent Bio SEC-5 500Å (Agilent Technologies社)を用いた。溶離液にはPBS(pH7.4)を用い、流速は1mL/分の条件で行った。1回の解析あたり抗体サンプルを1μg以上使用した。クロマトグラムはOpenLAB CDS ChemStation Edition(Agilent Technologies社)を用いて解析し、三量体/四量体、二量体、単量体各々に相当するピーク面積を算出し、面積比として比較した。図8Aに、サイズ排除クロマトグラフィーにより得られるクロマトグラムのIgA1、IgA2m1、IgA2m2、IgA2(n)型抗体の代表的な結果を示す。図8Bは、図8Aに基づいて、三量体/四量体、二量体、単量体抗体各々のピーク面積比を算出した結果を示すグラフである。図8Aのクロマトグラムのピーク間の谷又はピークの分離が不明瞭な場合はピークの変曲点を推測し、ベースラインへ垂直の線を引き、各々のフラクションに区分けを行い、クロマトグラムとベースラインに囲まれる面積を算出した。
IgA2m2型抗体の多量体化促進活性について解析するため、以下の解析を行った。
PrimeSTAR(登録商標)MAX DNA Polymeraseを使用して、上述したα1 HC発現ベクターを鋳型として、IgA1 HCのCH1からCH2までの配列(α1セグメント:定常領域のN末端からG342まで)をPCR増幅した。また、α2m2 HC発現ベクターを鋳型として、IgA2m2 HCのCH3以降C末までの配列(α2m2セグメント:N343から定常領域のC末端まで)をPCR増幅した。
IgA2m2の多量体化促進活性について解析するため、各種発現ベクターを以下の通りに構築した。
IgA重鎖定常領域の変異体発現ベクターを構築するため、定常領域にアミノ酸置換を伴わないよう制限酵素部位を付加した定常領域フラグメントIgA1H-NRE(配列番号28)、IgA2m2-NRE(配列番号29)を人工遺伝子合成(Genscript社)により合成した。続いて、IgA1H-NRE、IgA2m2-NREを制限酵素NheI及びHindIIIで処理し、ライゲーション用フラグメントを調製した。
IgA1/IgA2m2キメラ重鎖発現ベクターの構築を以下の通りに行った。すなわち、pIgA2m2H-NREをMfeI及びHindIIIで処理し得られたフラグメントとMfeI及びHindIII処理したpIgA1H-NREとを連結し、IgA1重鎖のアミノ酸342~472番目の領域をIgA2m2型に置換した変異体pIgA1H/pIgA2m2-chimeraを構築した。図10Aは各変異体の構造を示す模式図である。
J鎖、SCの共発現ベクター構築のため、実施例5のpCXSN-cis#1-hJC及びpCXSN-cis#2-hSCより、cis#1-hJC ORF、cis#2-hSC ORFを切り出した。cis#1-hJC ORF、cis#2-hSC ORFをそれぞれヒトEF1プロモーターとBGH polyAから構成される発現ユニット内のプロモーターとpolyAの間に連結し、発現ベクターpEF-cis-hJC/Zeo及びpEF/cis-hSC/Zeoをそれぞれ構築した。
IgA1重鎖のCH3以降C末端までのアミノ酸配列と、これに対応するIgA2m2重鎖のアミノ酸配列とで異なるアミノ酸残基は、第411残基、第428残基、第451残基、第458残基、第467残基の5残基であるため、以下の解析を行った。
重鎖定常領域(Fc領域)のアミノ酸置換変異体を作製した。より具体的には、α2m2 HC発現ベクターを鋳型として、5種のアミノ酸置換(Y411F、E428D、M451L、I458V、A467V)が起こるように配列を改編したプライマーを用いて、PrimeSTAR(登録商標)MAX DNA Polymeraseを使用して、インバースPCRを行った。
実験例13と同様にして、Fc領域のアミノ酸置換変異体の作製及び解析を行った。以下、作製した変異体を「IgA1 F411Y」のように表記する。この場合、IgA1の定常領域の第411残基に相当するアミノ酸フェニルアラニン(F)がチロシン(Y)に置換されている。他の変異体についても同様である。
IgAの多量体化促進活性におけるIgA1重鎖内V458の役割について解析するため、V458の変異体発現ベクターを以下の通りに構築した。
IgA1重鎖458番目のVをアミノ酸各種に置換した変異体を構築するため、pIgA1H-NREを制限酵素MfeI及びHindIIIで処理しライゲーション用フラグメントを調製した。また、インサートリンカーを調製するため、V458I(配列番号32及び33)、V458A(配列番号34及び35)、V458W(配列番号36及び37)、V458C(配列番号38及び39)、V458D(配列番号40及び41)、V458E(配列番号42及び43)、V458F(配列番号44及び45)、V458G(配列番号46及び47)、V458H(配列番号48及び49)、V458K(配列番号50及び51)、V458L(配列番号52及び53)、V458M(配列番号54及び55)、V458N(配列番号56及び57)、V458P(配列番号58及び59)、V458Q(配列番号60及び61)、V458R(配列番号62及び63)、V458S(配列番号64及び65)、V458T(配列番号66及び67)、V458Y(配列番号68及び69)の組み合わせで各DNA断片を95℃で10分熱処理後に段階的に25℃まで下げてアニーリングさせた。
続いて、リポフェクション法により、作製したV458の各種変異体発現ベクターと上述した抗インフルエンザウイルス抗体クローンB12の軽鎖発現ベクター(pIgA-LC)とをC23細胞にそれぞれトランスフェクションした。
(Fc領域のアミノ酸置換変異体の作製)
C23細胞の代わりにExpi293F細胞を用いて実験例15と同様の検討を行った。具体的には、実験例15で作製した、pIgA1H-NRE-V458I、pIgA1H-NRE-V458A、pIgA1H-NRE-V458W、pIgA1H-NRE-V458E、pIgA1H-NRE-V458G、pIgA1H-NRE-V458K、pIgA1H-NRE-V458Lの発現プラスミドを使用し、実験例13と同様の実験を行った。
(Fc領域のアミノ酸置換変異体の作製)
IgA1、IgA2m1、IgA2m2、IgA2(n)について、第458残基の変異体を作製し、多量体化における役割を検討した。α2m1 HCのアミノ酸置換変異体を作製する場合、α2m1 HC発現ベクターを鋳型として、目的のアミノ酸置換(V458I)が起こるように配列を改編したプライマーを用いて、PrimeSTAR(登録商標)MAX DNA Polymeraseを使用したインバースPCRを行った。続いて、増幅したPCR産物の5’末端をT4ポリヌクレオチドキナーゼを用いてリン酸化し、T4DNAリガーゼにより環状に連結させた。続いて、反応産物で大腸菌を形質転換し、プラスミドを抽出し、シークエンスにより目的のアミノ酸置換を起こす変異が導入されたことを確認した。α2(n) HCについても同様にして第458残基のアミノ酸変異を有する変異体を作製した。α1 HCについては実験例14、α2m2 HCは実験例13で作製した変異体を用いた。
上述したspERt技術を用いて四量体IgA抗体を製造し、以下の方法で解析した。
J/SC安定発現CHO株C23に抗インフルエンザウイルス抗体クローンF11の軽鎖発現ベクターpIgA-LCと、IgA1重鎖発現ベクターpIgA1-HC又はIgA2m2重鎖発現ベクターpIgA2m2-HCとをリポフェクション法にてトランスフェクションした。ピューロマイシン(10μg/ml)による薬剤選択を経て、IgA1重鎖及び軽鎖を安定に発現するCHO株C78、並びにIgA2m2重鎖及び軽鎖を安定に発現するCHO株C179を樹立した。
上述した細胞株C78、C179、C104及びC117を、それぞれ5%ウシ胎児血清含有DMEMにて7日間培養し、培養液を低速遠心とポアサイズ0.45μmのフィルターを用いたろ過により清澄化後、IgA抗体をCaptureSelect human Fc affinity matrix(ライフテクノロジーズ社製)を用いて実験例1~4と同様にして精製した。
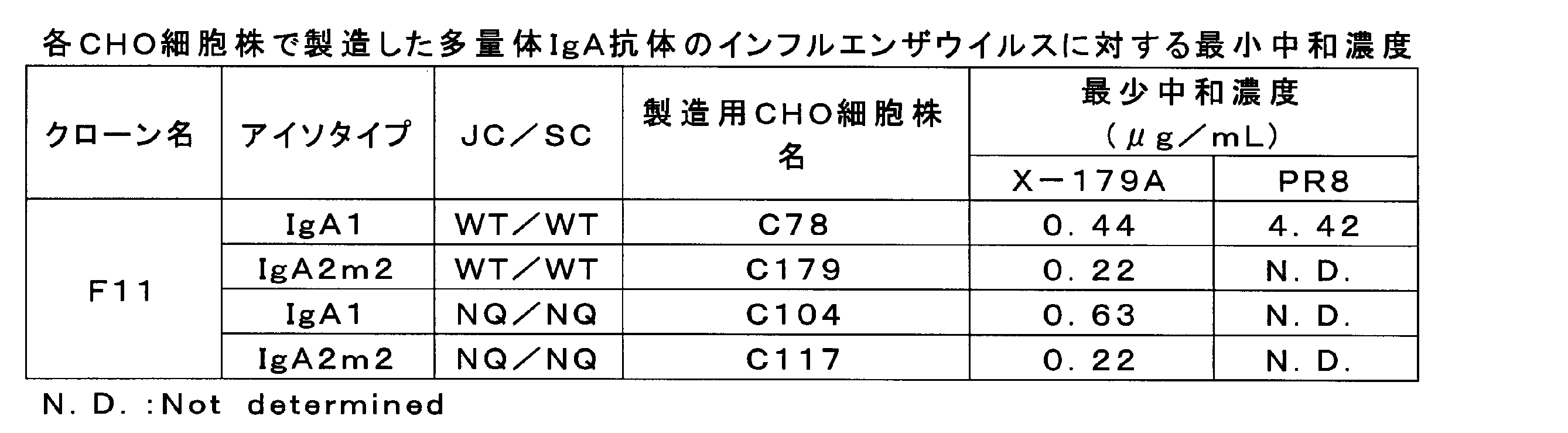
spERt技術によるIgA抗体の生産性増強効果について検討した。
実験例12に記載の方法と同様にして、p180及びSF3b4を安定に発現するCHO-K1細胞株1C17を樹立した。
各クローンの多量体を作製するため、CHO-K1S細胞に、上述したpIgA-LCと、pEF-cis-hJC/Zeoと、pEF/cis-hSC/Zeoと、pIgA1-HC又はpIgA2m2-HCとをリポフェクション法によりトランスフェクションした。
実験例4と同様に調製したIgA1、IgA2m2の多量体(四量体)分画成分の分子量を、質量分析計を用いて測定した。ここで、IgA1にはpCXSN-hSCを用いてタグの無いSCを作製した。また、IgA2m2にはpCXSN-hSC-HisTagを用いてタグ付きSCを発現させて各多量体を作製した。

多量体画分中の各サブユニットの構成比を高精度に定量するため、安定同位体標識ペプチドを内部標準とした質量分析による新規定量方法を確立した。
0~2分:A液(0.1%ギ酸)98%、B液(100%アセトニトリル)2%;
2.1~6分:A液98~50%、B液2~50%;
6.1~8分:A液10%、B液90%;
8.1~10分:A液98%、B液2%。

Claims (15)
- 多量体IgA型遺伝子組換え抗体。
- 重鎖定常領域の第458番目のアミノ酸残基が疎水性アミノ酸に由来するアミノ酸残基である、請求項1に記載の多量体IgA型遺伝子組換え抗体。
- 四量体の含有量が全IgAの20モル%以上である、請求項1又は2に記載の多量体IgA型遺伝子組換え抗体。
- 請求項1~3のいずれか一項に記載の多量体IgA型遺伝子組換え抗体を有効成分として含有する医薬。
- 感染症の治療又は予防用である、請求項4に記載の医薬。
- IgA型抗体重鎖タンパク質、抗体軽鎖タンパク質、抗体J鎖タンパク質及び分泌成分タンパク質を1つの細胞内で共発現させる工程を含む、多量体IgA型抗体の製造方法。
- 前記IgA型抗体重鎖タンパク質は、遺伝子組換えによりIgG型からIgA型に変換されたものである、請求項6に記載の製造方法。
- 前記工程において、更にp180タンパク質及びSF3b4タンパク質を前記細胞内で共発現させる、請求項6又は7に記載の製造方法。
- 前記細胞がCHO YA7細胞株(受託番号NITE BP-01535)である、請求項6~8のいずれか一項に記載の製造方法。
- 前記工程は、IgA型抗体重鎖タンパク質、抗体軽鎖タンパク質、抗体J鎖タンパク質及び分泌成分タンパク質を発現するための発現ベクターを前記細胞内に導入することにより行われ、
前記発現ベクターは、プロモーターの下流で、かつ、IgA型抗体重鎖タンパク質、抗体軽鎖タンパク質、抗体J鎖タンパク質又は分泌成分タンパク質をコードする核酸の開始コドンの上流に、RNA結合タンパク質が認識、結合又は相互作用するcis-エレメントを有する、請求項6~9のいずれか一項に記載の製造方法。 - 前記cis-エレメントが、配列モチーフGAN1-(X)n-ACN2(nは3~6の整数であり、N1及びN2は、それぞれ独立して、A、T、G、Cのいずれかである。)からなる塩基配列を1~数個含む、請求項10に記載の製造方法。
- 前記cis-エレメントが、
配列番号21~23のいずれかに示される塩基配列、
配列番号21~23のいずれかに示される塩基配列において、1~数個の塩基が欠失、置換又は付加されている塩基配列からなり、かつRNA結合タンパク質が認識、結合若しくは相互作用する塩基配列、
配列番号21~23のいずれかに示される塩基配列と同一性が80%以上である塩基配列からなり、かつ、RNA結合タンパク質が認識、結合若しくは相互作用する塩基配列、又は、
配列番号21~23のいずれかに示される塩基配列からなる核酸と相補的な塩基配列からなる核酸とストリンジェントな条件下でハイブリダイズすることができる塩基配列からなり、かつRNA結合タンパク質が認識、結合若しくは相互作用する塩基配列からなる、請求項10又は11に記載の製造方法。 - 抗体を多量体IgA型化する工程を含む、前記抗体の抗原結合活性又は中和活性を向上させる方法。
- 前記抗体は、IgG型抗体である、請求項13に記載の方法。
- 前記工程が、前記抗体の重鎖可変領域を有するIgA型抗体重鎖タンパク質、前記抗体の軽鎖タンパク質、抗体J鎖タンパク質及び分泌成分タンパク質を1つの細胞内で共発現させる工程を含む、請求項13又は14に記載の方法。
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2016534515A JP6564777B2 (ja) | 2014-07-18 | 2015-07-21 | 多量体IgA型遺伝子組換え抗体を含む組成物及びその利用 |
| EP15822054.1A EP3170839B1 (en) | 2014-07-18 | 2015-07-21 | POLYMERIC IgA-TYPE RECOMBINANT ANTIBODY AND USE THEREOF |
| EP20196341.0A EP3786181A1 (en) | 2014-07-18 | 2015-07-21 | Polymeric iga-type recombinant antibody and use thereof |
| US15/326,569 US10925962B2 (en) | 2014-07-18 | 2015-07-21 | Polymeric IgA-type recombinant antibody and use thereof |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2014148328 | 2014-07-18 | ||
| JP2014-148328 | 2014-07-18 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2016010161A1 true WO2016010161A1 (ja) | 2016-01-21 |
Family
ID=55078646
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2015/070742 Ceased WO2016010161A1 (ja) | 2014-07-18 | 2015-07-21 | 多量体IgA型遺伝子組換え抗体及びその利用 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US10925962B2 (ja) |
| EP (2) | EP3786181A1 (ja) |
| JP (1) | JP6564777B2 (ja) |
| WO (1) | WO2016010161A1 (ja) |
Cited By (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2671477C2 (ru) * | 2016-12-30 | 2018-10-31 | Федеральное государственное бюджетное образовательное учреждение высшего образования "Московский государственный университет имени М.В. Ломоносова" (МГУ) | РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA2m1F16-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA2m1 |
| WO2021193553A1 (ja) * | 2020-03-23 | 2021-09-30 | 東興薬品工業株式会社 | 多量体IgA抗体の作製法及び多重特異性多量体IgA抗体 |
| JP2022522985A (ja) * | 2019-01-22 | 2022-04-21 | ジェネンテック, インコーポレイテッド | 免疫グロブリンa抗体、並びに産生及び使用の方法 |
| CN115190888A (zh) * | 2020-01-24 | 2022-10-14 | 麦克法兰布奈特医疗研究与公共健康研究所有限公司 | 检测肠道屏障功能障碍和/或肝硬化 |
| JP2023535677A (ja) * | 2020-07-10 | 2023-08-21 | バイオモレキュラー・ホールディングス・エルエルシー | 四面体抗体 |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR20220004979A (ko) | 2019-03-27 | 2022-01-12 | 유엠씨 우트레크트 홀딩 비.브이. | 조작된 iga 항체 및 사용 방법 |
| US20220033851A1 (en) * | 2020-08-03 | 2022-02-03 | Roger B. Swartz | mRNA, episomal and genomic integrated lentiviral and gammaretroviral vector expression of dimeric immunoglobulin A and polymeric immunoglobulin A to Enable Mucosal and Hematological Based Immunity/Protection via Gene Therapy for Allergens, viruses, HIV, bacteria, pneumonia, infections, pathology associated proteins, systemic pathologies, cancer, toxins and unnatural viruses. CAR engineered and non-CAR engineered immune cell expression of dimeric immunoglobulin A and polymeric immunoglobulin A. |
| CA3229487A1 (en) * | 2021-08-20 | 2023-02-23 | Adam WAICKMAN | Iga monoclonal antibodies for treating flavivirus infection |
| US20240425570A1 (en) * | 2021-09-17 | 2024-12-26 | The Board Of Trustees Of The University Of Illinois | Chimeric secretory component polypeptides and uses thereof |
| CN118995809B (zh) * | 2023-05-17 | 2026-03-31 | 清华大学 | 一种用于制备IgA双聚体的方法 |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2013087914A1 (en) * | 2011-12-16 | 2013-06-20 | Synthon Biopharmaceuticals B.V. | EXPRESSION OF SECRETORY IgA ANTIBODIES IN DUCKWEED |
| WO2014157429A1 (ja) * | 2013-03-26 | 2014-10-02 | 株式会社 ニッピ | タンパク質の製造方法 |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU689489B2 (en) * | 1993-07-30 | 1998-04-02 | Oravax, Inc | Monoclonal IgA antibody against respiratory syncytial virus |
| US6063905A (en) * | 1997-01-07 | 2000-05-16 | Board Of Regents, The University Of Texas System | Recombinant human IGA-J. chain dimer |
| JP6086743B2 (ja) | 2013-01-31 | 2017-03-01 | 株式会社吉野工業所 | 詰め替え容器 |
-
2015
- 2015-07-21 JP JP2016534515A patent/JP6564777B2/ja active Active
- 2015-07-21 WO PCT/JP2015/070742 patent/WO2016010161A1/ja not_active Ceased
- 2015-07-21 EP EP20196341.0A patent/EP3786181A1/en not_active Withdrawn
- 2015-07-21 US US15/326,569 patent/US10925962B2/en active Active
- 2015-07-21 EP EP15822054.1A patent/EP3170839B1/en active Active
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2013087914A1 (en) * | 2011-12-16 | 2013-06-20 | Synthon Biopharmaceuticals B.V. | EXPRESSION OF SECRETORY IgA ANTIBODIES IN DUCKWEED |
| WO2014157429A1 (ja) * | 2013-03-26 | 2014-10-02 | 株式会社 ニッピ | タンパク質の製造方法 |
Non-Patent Citations (5)
| Title |
|---|
| JOHANSEN, FE. ET AL.: "Recombinant expression of polymeric IgA: incorporation of J chain and secretory component of human origin", EUROPEAN JOURNAL OF IMMUNOLOGY, vol. 29, no. 5, 1999, pages 1701 - 1708, XP002331960, DOI: doi:10.1002/(SICI)1521-4141(199905)29:05<1701::AID-IMMU1701>3.0.CO;2-Z * |
| LI, C. ET AL.: "Construction of a Chimeric Secretory IgA and Its Neutralization Activity against Avian Influenza Virus H5N1", JOURNAL OF IMMUNOLOGY RESEARCH, vol. 394127, 13 February 2014 (2014-02-13), pages 1 - 10, XP055385659 * |
| LORIN, V. ET AL.: "Efficient generation of human IgA monoclonal antibodies", JOURNAL OF IMMUNOLOGICAL METHODS, vol. 422, 22 April 2015 (2015-04-22), pages 102 - 110, XP029239942, DOI: doi:10.1016/j.jim.2015.04.010 * |
| MURAMATSU, M. ET AL.: "Comparison of Antiviral Activity between IgA and IgG Specific to Influenza Virus Hemagglutinin: Increased Potential of IgA for Heterosubtypic Immunity", PLOS ONE, vol. 9, no. 1, 17 January 2014 (2014-01-17), pages e85582, XP055385668 * |
| See also references of EP3170839A4 * |
Cited By (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2671477C2 (ru) * | 2016-12-30 | 2018-10-31 | Федеральное государственное бюджетное образовательное учреждение высшего образования "Московский государственный университет имени М.В. Ломоносова" (МГУ) | РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA2m1F16-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA2m1 |
| JP2022522985A (ja) * | 2019-01-22 | 2022-04-21 | ジェネンテック, インコーポレイテッド | 免疫グロブリンa抗体、並びに産生及び使用の方法 |
| CN115190888A (zh) * | 2020-01-24 | 2022-10-14 | 麦克法兰布奈特医疗研究与公共健康研究所有限公司 | 检测肠道屏障功能障碍和/或肝硬化 |
| WO2021193553A1 (ja) * | 2020-03-23 | 2021-09-30 | 東興薬品工業株式会社 | 多量体IgA抗体の作製法及び多重特異性多量体IgA抗体 |
| JPWO2021193553A1 (ja) * | 2020-03-23 | 2021-09-30 | ||
| JP7785258B2 (ja) | 2020-03-23 | 2025-12-15 | 東興薬品工業株式会社 | 多量体IgA抗体の作製法及び多重特異性多量体IgA抗体 |
| JP2023535677A (ja) * | 2020-07-10 | 2023-08-21 | バイオモレキュラー・ホールディングス・エルエルシー | 四面体抗体 |
Also Published As
| Publication number | Publication date |
|---|---|
| EP3170839A4 (en) | 2018-01-03 |
| US10925962B2 (en) | 2021-02-23 |
| EP3170839A1 (en) | 2017-05-24 |
| US20170340732A1 (en) | 2017-11-30 |
| EP3170839B1 (en) | 2020-10-28 |
| JPWO2016010161A1 (ja) | 2017-06-01 |
| JP6564777B2 (ja) | 2019-08-21 |
| EP3786181A1 (en) | 2021-03-03 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6564777B2 (ja) | 多量体IgA型遺伝子組換え抗体を含む組成物及びその利用 | |
| CA2956000C (en) | An anti-ctla4 monoclonal antibody or antigen binding fragment thereof, a pharmaceutical composition and use | |
| US11926657B2 (en) | Neutralizing anti-influenza binding molecules and uses thereof | |
| CA3052903A1 (en) | Anti human annexin a1 antibody | |
| WO2023105087A1 (en) | Novel flt3 antibodies and antibody-drug-conjugates based thereon, therapeutic methods and uses thereof in combination with tyrosine kinase inhibitors | |
| CN110325546A (zh) | 靶向流感病毒的dna单克隆抗体 | |
| CN115057939B (zh) | 结合新型冠状病毒的双特异性结合蛋白、药物和方法 | |
| JPWO2016010160A1 (ja) | 抗インフルエンザウイルス抗体及びその利用 | |
| CN110437332B (zh) | 一种抗病毒感染的sftsv蛋白结合分子 | |
| RU2777073C1 (ru) | Однодоменное антитело для нейтрализации вирусов и его модификации, и способ их применения для экстренной профилактики заболеваний, вызываемых вирусом гриппа А | |
| HK40061503B (zh) | 中和抗流感结合分子及其用途 | |
| HK1250725B (zh) | 中和抗流感结合分子及其用途 | |
| HK1253092B (en) | Neutralizing anti-influenza binding molecules and uses thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 15822054 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2016534515 Country of ref document: JP Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| REEP | Request for entry into the european phase |
Ref document number: 2015822054 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2015822054 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 15326569 Country of ref document: US |