WO2021100643A1 - 電子伝達体、電子伝達体の製造方法、及び、電子伝達方法 - Google Patents
電子伝達体、電子伝達体の製造方法、及び、電子伝達方法 Download PDFInfo
- Publication number
- WO2021100643A1 WO2021100643A1 PCT/JP2020/042553 JP2020042553W WO2021100643A1 WO 2021100643 A1 WO2021100643 A1 WO 2021100643A1 JP 2020042553 W JP2020042553 W JP 2020042553W WO 2021100643 A1 WO2021100643 A1 WO 2021100643A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- protein
- outer membrane
- cyanobacteria
- cell wall
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images














Classifications
-
- H—ELECTRICITY
- H01—ELECTRIC ELEMENTS
- H01M—PROCESSES OR MEANS, e.g. BATTERIES, FOR THE DIRECT CONVERSION OF CHEMICAL ENERGY INTO ELECTRICAL ENERGY
- H01M8/00—Fuel cells; Manufacture thereof
- H01M8/16—Biochemical fuel cells, i.e. cells in which microorganisms function as catalysts
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPR]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12R—INDEXING SCHEME ASSOCIATED WITH SUBCLASSES C12C - C12Q, RELATING TO MICROORGANISMS
- C12R2001/00—Microorganisms ; Processes using microorganisms
- C12R2001/01—Bacteria or Actinomycetales ; using bacteria or Actinomycetales
-
- H—ELECTRICITY
- H01—ELECTRIC ELEMENTS
- H01M—PROCESSES OR MEANS, e.g. BATTERIES, FOR THE DIRECT CONVERSION OF CHEMICAL ENERGY INTO ELECTRICAL ENERGY
- H01M14/00—Electrochemical current or voltage generators not provided for in groups H01M6/00 - H01M12/00; Manufacture thereof
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02E—REDUCTION OF GREENHOUSE GAS [GHG] EMISSIONS, RELATED TO ENERGY GENERATION, TRANSMISSION OR DISTRIBUTION
- Y02E60/00—Enabling technologies; Technologies with a potential or indirect contribution to GHG emissions mitigation
- Y02E60/30—Hydrogen technology
- Y02E60/50—Fuel cells
Definitions
- the present disclosure relates to an electron carrier containing modified cyanobacteria, a method for producing an electron carrier, and a method for electron transfer.
- Non-Patent Document 1 A bioelectrochemical system (for example, a power generation device) capable of introducing such a phenomenon into, for example, sewage treatment technology and recovering resources from organic wastewater is expected (Patent Document 1).
- the electrons generated in the cells by photosynthesis can be used as a microbial solar cell (Non-Patent Document 3).
- Non-Patent Documents 5 and 6 modification of the electrode surface (Non-Patent Documents 5 and 6) or a mediator compound (Non-Patent Document 7) improves the electron transfer efficiency between the cyanobacteria and the electrode.
- cyanobacteria are also known to produce various useful substances (for example, alcohols, alkanes, fatty acids, etc.) by photosynthesis (Patent Documents 2 and 2).
- Non-Patent Document 4 Non-Patent Document 4).
- Non-Patent Document 2 On the other hand, technological development is also underway to send electrons from outside the cell to the microbial cell to activate the metabolism of the microbial cell and improve the efficiency of substance production (for example, Non-Patent Document 2).
- the present disclosure provides an electron carrier having improved electron transfer efficiency to the outside, a method for producing the electron carrier, and an electron transfer method by including a modified cyanobacteria having improved extracellular electron transfer efficiency. ..
- the electron transmitter according to one aspect of the present disclosure is: (i) the function of a protein involved in the binding between the outer membrane and the cell wall is suppressed or lost in cyanobacteria, and (ii) the protein permeation of the outer membrane. It comprises at least one modified cyanobacteria in which a sex-enhancing channel protein is expressed, said modified cyanobacteria at least one of supplying electrons to the outside and taking in electrons from the outside. I do.
- the electron carrier of the present disclosure and the method for manufacturing the electron carrier, it is possible to provide an electron carrier having improved electron transfer efficiency with the outside. Further, according to the electron transfer method of the present disclosure, the efficiency of at least one of the donation of electrons from the electron carrier to the outside and the reception of electrons by the electron carrier from the outside is improved.
- FIG. 1 is a diagram schematically showing the cell surface layer of cyanobacteria.
- FIG. 2 is a schematic view showing an example of an electron carrier according to the present embodiment.
- FIG. 3 is a transmission electron microscope observation image of an ultrathin section of the modified cyanobacteria of Example 1.
- FIG. 4 is an enlarged image of the broken line region A of FIG.
- FIG. 5 is a transmission electron microscope image of an ultrathin section of the modified cyanobacteria of Example 2.
- FIG. 6 is an enlarged image of the broken line region B of FIG.
- FIG. 7 is a transmission electron microscope image of an ultrathin section of the modified cyanobacteria of Comparative Example 1.
- FIG. 8 is an enlarged view of the broken line region C of FIG. 7.
- FIG. 10 is an exploded perspective view schematically showing an example of the configuration of the electrochemical measuring device.
- FIG. 11 is a schematic cross-sectional view taken along the XI-XI cross-sectional line of FIG.
- FIG. 12 is a diagram showing the results of measuring the current flowing when the culture solution of the modified cyanobacteria of Comparative Example 1 was irradiated with light.
- FIG. 13 is a diagram showing the results of measuring the current flowing when the culture solution of the modified cyanobacteria of Example 2 was irradiated with light.
- FIG. 14 is a diagram showing the results of measuring the current flowing when the culture solution of the modified cyanobacteria of Example 3 was irradiated with light.
- Non-Patent Document 1 if the electrons emitted from the microorganism are received by the external electrode, it can be used as a microbial fuel cell using organic matter as fuel. For example, it can be introduced into a sewage treatment process containing organic matter and used as a power generation device. It is expected (Patent Document 1).
- Non-Patent Document 2 nanoparticles of indium phosphate are adhered to yeast cells, and electrons generated by the photoelectric conversion reaction of the nanoparticles are taken up into the cells to obtain the reducing power used for intracellular metabolism. It has been reported that supplementation can improve the efficiency of shikimic acid production from glucose raw materials.
- microorganisms does not depend on chemical fuels, and can realize energy production and substance production with low environmental load.
- photosynthetic microorganisms such as cyanobacteria and algae are expected to be carbon-neutral next-generation material production systems because they can use light as an energy source and carbon dioxide (CO 2) in the air as a raw material. ..
- Cyanobacteria also called blue-green bacteria or blue-green algae
- Cyanobacteria are a group of eubacteria that decompose water by photosynthesis to produce oxygen and fix CO 2 in the air with the obtained energy. Cyanobacteria can also fix nitrogen (N 2 ) in the air depending on the species.
- N 2 nitrogen
- cyanobacteria grow quickly and have high light utilization efficiency, and in addition, genetic manipulation is easier than other algae species. Therefore, regarding the utilization of cyanobacteria among photosynthetic microorganisms. Active research and development is taking place.
- cyanobacteria accept the electrons generated in the cells of cyanobacteria during electrolysis of water by photosynthesis at an external electrode by utilizing the property of cyanobacteria to transfer electrons outside the cell, water, cyanobacteria, and a pair of electrodes It can be used as a microbial solar cell with a simple configuration including (Non-Patent Document 3).
- cyanobacteria have high photosynthetic ability, they cannot obtain sufficient light energy in low light such as at night or in the evening, in rainy weather or in cloudy weather. Therefore, if electrons can be sent into the cell from the outside of the cyanobacteria, the intracellular reducing power that is insufficient at night or in low light can be replenished from the outside, which is expected to lead to improvement in substance production efficiency.
- Non-Patent Document 5 As means for improving the electron transfer efficiency between cyanobacteria and the external electrode, modification of the electrode surface (Non-Patent Document 5 and Non-Patent Document 6) and a mediator compound (Non-Patent Document 7) have been reported. There is.
- modified cyanobacteria by partially detaching the outer membrane of cyanobacteria from the cell wall or improving the substance permeability of the outer membrane.
- the efficiency of electron transfer between cells and external electrodes is improved. More specifically, by removing the outer membrane of cyanobacteria or improving the permeability of substances, the modified cyanobacteria secrete intracellular electron carriers extracellularly, and extracellular electron carriers are secreted into cells.
- the extracellular electron transfer efficiency of the modified cyanobacteria is improved, so that the electron carrier containing the modified cyanobacteria can efficiently transfer electrons to the outside.
- the present disclosure provides an electron carrier having improved electron transfer efficiency with the outside, and a method for manufacturing the electron carrier.
- the present disclosure also provides an electron transfer method that improves the efficiency of at least one of the donation of electrons from the electron carrier to the outside and the reception of electrons from the outside of the electron carrier.
- the electron transmitter according to one aspect of the present disclosure is: (i) the function of a protein involved in the binding between the outer membrane and the cell wall is suppressed or lost in cyanobacteria, and (ii) the protein permeation of the outer membrane. It comprises at least one modified cyanobacteria in which a sex-enhancing channel protein is expressed, said modified cyanobacteria at least one of supplying electrons to the outside and taking in electrons from the outside. I do.
- the binding between the cell wall and the outer membrane (for example, the binding amount and the binding force) is partially reduced, so that the outer membrane is easily partially detached from the cell wall. Therefore, the electron generated inside the cell or the substance or molecule having the electron easily leaks to the outside of the outer membrane, that is, to the outside of the cell.
- the protein permeability of the outer membrane is improved, so that the substance permeability of the outer membrane is improved.
- the modified cyanobacteria secrete extracellularly generated electrons or substances or molecules having electrons into the cells, and take up extracellular electrons or substances or molecules having electrons into the cells. At least one can be done. Therefore, in the modified cyanobacteria, the extracellular electron transfer efficiency is improved. Therefore, the electron transport chain according to one aspect of the present disclosure has improved electron transfer efficiency with the outside.
- the outside is a substance or molecule that exists as an individual different from the electron carrier, and is, for example, a redox substance involved in the movement of electrons between substances, or a molecule having a redox reactive group.
- the modified cyanobacteria may receive light to generate electrons and release the generated electrons to the outside of the outer membrane.
- the modified cyanobacteria release electrons or substances or molecules with electrons to the outside of the cell when they receive light. Therefore, the electron carrier according to one aspect of the present disclosure can generate electrons inside when irradiated with light, and can supply electrons or substances or molecules having electrons to the outside.
- the modified cyanobacteria may take in electrons existing outside the outer membrane inside the cell wall and utilize the electrons inside the cell wall. ..
- the modified cyanobacteria take in electrons or substances or molecules having electrons existing outside the cell into the cell (in the cytoplasm), and for example, energy (ATP: adenosine triphosphate) in the process of flowing through the photosynthetic electron transport chain. ) Is generated. Then, the modified cyanobacteria use this energy to produce organic matter based on carbon dioxide. Therefore, the electron carrier according to one aspect of the present disclosure can take in electrons from the outside to the inside to generate energy and produce an organic substance such as a protein.
- energy ATP: adenosine triphosphate
- the proteins involved in the binding between the outer membrane and the cell wall are SLH (Surface Layer Homology) domain-retaining outer membrane protein and It may be at least one of the cell wall-pyruvate modifying enzymes.
- the modified cyanobacteria for example, (a) an enzyme that catalyzes the reaction of modifying the SLH domain-retaining outer membrane protein that binds to the cell wall and the bound sugar chain on the surface of the cell wall with pyruvate (that is, cell wall-pyruvate modification) At least one function of (enzyme) is suppressed or lost, or (b) expression of at least one of SLH domain-retaining outer membrane protein and cell wall-pyruvate modifying enzyme is suppressed. Therefore, the binding (that is, the binding amount and binding force) between the SLH domain of the SLH domain-retaining outer membrane protein in the outer membrane and the covalently bound sugar chain on the surface of the cell wall is reduced.
- pyruvate that is, cell wall-pyruvate modification
- the outer membrane is likely to be detached from the cell wall at the portion where the bond between the outer membrane and the cell wall is weakened.
- the modified cyanobacteria secretes intracellular electrons or substances or molecules having electrons to the outside of the cell, and takes up extracellular electrons or substances or molecules having electrons into the cells at least one of them. Therefore, the efficiency of extracellular electron transfer is improved. Therefore, the electron transport chain according to one aspect of the present disclosure has improved electron transfer efficiency with the outside.
- the SLH domain-retaining outer membrane protein is Slr1841 consisting of the amino acid sequence shown in SEQ ID NO: 1, NIES970_09470 consisting of the amino acid sequence shown in SEQ ID NO: 2, and SEQ ID NO: It may be Anacy_3458 consisting of the amino acid sequence represented by 3 or a protein having an amino acid sequence 50% or more identical to that of any of these SLH domain-retaining outer membrane proteins.
- any SLH domain-retaining outer membrane protein shown in SEQ ID NOs: 1 to 3 above, or any of these SLH domain-retaining outer membrane proteins and an amino acid sequence The function of proteins that are 50% or more identical is suppressed or lost, or (b) any SLH domain-retaining outer membrane protein set forth in SEQ ID NOs: 1-3 above, or any of these SLH domains.
- the expression of a protein having an amino acid sequence that is 50% or more identical to that of a retained outer membrane protein is suppressed.
- the modified cyanobacteria (a) the function of the SLH domain-retaining outer membrane protein in the outer membrane or the protein having the same function as the SLH domain-retaining outer membrane protein is suppressed or lost, or (b) The expression level of the SLH domain-retaining outer membrane protein or the protein having the same function as the SLH domain-retaining outer membrane protein in the outer membrane is reduced.
- the binding amount and binding force of the binding domain (for example, SLH domain) of the outer membrane to bind to the cell wall are reduced, so that the outer membrane is easily partially detached from the cell wall. Therefore, the electron transport chain according to one aspect of the present disclosure has improved electron transfer efficiency with the outside.
- the cell wall-pyruvate modifying enzyme is Slr0688 consisting of the amino acid sequence shown in SEQ ID NO: 4, Synpcc7942_1529 consisting of the amino acid sequence shown in SEQ ID NO: 5, and SEQ ID NO: 6. It may be Anacy_1623 consisting of the amino acid sequence shown by, or a protein having an amino acid sequence of 50% or more identical to that of any of these cell wall-pyruvate modifying enzymes.
- the modified cyanobacteria for example, (a) one of the cell wall-pyruvate modifying enzymes shown in SEQ ID NOs: 4 to 6 above or any of these cell wall-pyruvate modifying enzymes and the amino acid sequence are 50%.
- the functions of the proteins that are the same as described above are suppressed or lost, or (b) any cell wall-pyruvate modifying enzyme shown in SEQ ID NOs: 4 to 6 above or any of these cell walls-pyruvate modification.
- the expression of proteins having the same amino acid sequence as the enzyme by 50% or more is suppressed.
- the function of the cell wall-pyruvic acid modifying enzyme or a protein having the same function as the enzyme is suppressed or lost, or (b) the cell wall-pyruvic acid modifying enzyme or the enzyme is used.
- the expression level of proteins with equivalent functions is reduced. This makes it difficult for the covalently bound sugar chains on the surface of the cell wall to be modified with pyruvic acid, so that the amount and binding force of the sugar chains on the cell wall to bind to the SLH domain of the SLH domain-retaining outer membrane protein in the outer membrane. Is reduced.
- the covalent sugar chains on the surface of the cell wall are less likely to be modified with pyruvic acid, so that the binding force between the cell wall and the outer membrane is weakened, and the outer membrane is easily partially detached from the cell wall. Become. Therefore, the electron transport chain according to one aspect of the present disclosure has improved electron transfer efficiency with the outside.
- the gene expressing the protein involved in the binding between the outer membrane and the cell wall may be deleted or inactivated.
- the modified cyanobacteria of (i) above the expression of the protein involved in the binding between the cell wall and the outer membrane is suppressed, or the function of the protein is suppressed or lost, so that the cell wall and the outer membrane are suppressed. Bonding with (so-called binding amount and binding force) is partially reduced. As a result, the outer membrane is likely to be detached from the cell wall at the portion where the bond between the outer membrane and the cell wall is weakened.
- intracellular electrons or substances or molecules having electrons are secreted extracellularly, and extracellular electrons or substances or molecules having electrons are taken up into the cells. Since at least one of these can be done, the efficiency of extracellular electron transfer is improved. Therefore, the electron transport chain according to one aspect of the present disclosure has improved electron transfer efficiency with the outside.
- the genes expressing the protein involved in the binding between the outer membrane and the cell wall are the gene encoding the SLH domain-retaining outer membrane protein and the cell wall-pyruvate. It may be at least one of the genes encoding the modifying enzyme.
- the modified cyanobacteria As a result, in the modified cyanobacteria, at least one gene encoding the SLH domain-retaining outer membrane protein and the gene encoding the cell wall-pyruvic acid modifying enzyme are deleted or inactivated. Therefore, in the modified cyanobacteria, for example, the expression of at least one of (a) SLH domain-retaining outer membrane protein and cell wall-pyruvic acid modifying enzyme is suppressed, or (b) SLH domain-retaining outer membrane protein and cell wall. -At least one function of pyruvate modifying enzyme is suppressed or lost.
- the binding that is, the binding amount and binding force
- the outer membrane is easily detached from the cell wall at the portion where the bond between the outer membrane and the cell wall is weakened, so that the intracellular electron or the substance or molecule having an electron is easily leaked to the outside of the cell. .. Therefore, in the electron carrier according to one aspect of the present disclosure, intracellular electrons or substances or molecules having electrons easily leak out of the cell in the modified cyanobacteria, so that the efficiency of electron transfer to the outside is improved.
- the gene encoding the SLH domain-retaining outer membrane protein is slr1841 consisting of the base sequence shown by SEQ ID NO: 7, and nies970_09470 consisting of the base sequence shown by SEQ ID NO: 8. It may be anacy_3458 consisting of the base sequence shown by SEQ ID NO: 9, or a gene whose base sequence is 50% or more identical to any of these genes.
- the gene encoding any of the SLH domain-bearing outer membrane proteins shown in SEQ ID NOs: 7 to 9 above, or a gene that is 50% or more identical to the base sequence of any of these genes. Is deleted or inactivated. Therefore, in the modified cyanobacteria, the expression of (a) any of the above SLH domain-retaining outer membrane proteins or a protein having a function equivalent to that of any of these proteins is suppressed, or (b) the above. The function of any SLH domain-retaining outer membrane protein or a protein having a function equivalent to that of any of these proteins is suppressed or lost.
- the binding amount and binding force of the binding domain (for example, SLH domain) for the outer membrane to bind to the cell wall are reduced.
- the outer membrane is likely to be partially detached from the cell wall, so that the intracellular electron or the substance or molecule having the electron is easily leaked to the outside of the cell.
- the modified cyanobacteria improve the extracellular electron transfer efficiency, so that the electron carrier according to one aspect of the present disclosure improves the electron transfer efficiency with the outside.
- the gene encoding the cell wall-pyruvate modifying enzyme is slr0688 consisting of the base sequence shown by SEQ ID NO: 10 and synpcc7942_1529 consisting of the base sequence shown by SEQ ID NO: 11. , Anacy_1623 consisting of the base sequence shown by SEQ ID NO: 12, or a gene whose base sequence is 50% or more identical to any of these genes.
- the nucleotide sequence of the gene encoding any cell wall-pyruvic acid modifying enzyme shown in SEQ ID NOs: 10 to 12 above or the gene encoding any of these enzymes is 50% or more identical.
- the gene is deleted or inactivated. Therefore, in the modified cyanobacteria, (a) expression of any of the above cell wall-pyruvic acid modifying enzymes or a protein having a function equivalent to that of any of these enzymes is suppressed, or (b) any of the above.
- the function of the cell wall-pyruvic acid modifying enzyme or a protein having a function equivalent to that of any of these enzymes is suppressed or lost.
- the electron transfer efficiency with the outside is improved.
- the channel protein for improving the protein permeability of the outer membrane is CppS consisting of the amino acid sequence shown in SEQ ID NO: 13, SEQ ID NO: 14. It may be CppF consisting of the indicated amino acid sequence, or a protein whose amino acid sequence is 50% or more identical to that of any of these channel proteins.
- the modified cyanobacteria of (ii) above with CppS (SEQ ID NO: 13) or CppF (SEQ ID NO: 14), which is a channel protein that improves the protein permeability of the outer membrane, or any of these channel proteins. Proteins with equivalent function are expressed. Therefore, in the modified cyanobacteria of (ii) above, the protein permeability of the outer membrane is improved, so that the substance permeability of the outer membrane is improved.
- the modified cyanobacteria of (ii) described above secrete intracellular electrons or substances or molecules having electrons to the outside of the cell, and take up extracellular electrons or substances or molecules having electrons into the cells. Since at least one of these can be done, the efficiency of extracellular electron transfer is improved. Therefore, the electron transport chain according to one aspect of the present disclosure has improved electron transfer efficiency with the outside.
- a gene encoding a channel protein that improves protein permeability of the outer membrane may be introduced into the (ii).
- the modified cyanobacteria of (ii) above a channel protein that improves protein permeability of the outer membrane is expressed. Therefore, in the modified cyanobacteria of (ii) above, the protein permeability of the outer membrane is improved, so that the substance permeability of the outer membrane is improved.
- the modified cyanobacteria of (ii) described above secrete intracellular electrons or substances or molecules having electrons to the outside of the cell, and take up extracellular electrons or substances or molecules having electrons into the cells. Since at least one of these can be done, the efficiency of extracellular electron transfer is improved. Therefore, the electron transport chain according to one aspect of the present disclosure has improved electron transfer efficiency with the outside.
- the gene encoding the channel protein that improves the protein permeability of the outer membrane may be a gene derived from chloroplast.
- the gene encoding the channel protein that improves the protein permeability of the outer membrane is cppS consisting of the base sequence shown by SEQ ID NO: 15, cppF consisting of the base acid sequence shown by SEQ ID NO: 16, or any of these. It may be a gene whose base sequence is 50% or more identical to that gene.
- the gene encoding any of the channel proteins shown in SEQ ID NO: 15 and SEQ ID NO: 16 or the base sequence of any of these genes is 50% or more identical.
- a gene is introduced. Therefore, in the modified cyanobacteria of (ii) above, a protein having a function of improving protein permeability in the outer membrane or a protein having a function equivalent to the protein is expressed. As a result, in the modified cyanobacteria of (ii) above, the protein permeability of the outer membrane is improved, so that the substance permeability of the outer membrane is improved.
- the modified cyanobacteria of (ii) described above secrete intracellular electrons or substances or molecules having electrons to the outside of the cell, and take up extracellular electrons or substances or molecules having electrons into the cells. Since at least one of these can be done, the efficiency of extracellular electron transfer is improved. Therefore, the electron transport chain according to one aspect of the present disclosure has improved electron transfer efficiency with the outside.
- the function of a protein involved in the binding between the outer membrane and the cell wall is suppressed or lost in cyanobacteria, and (ii) the above. It comprises the step of producing modified cyanobacteria, which is at least one of which channel proteins are expressed that improve the protein permeability of the outer membrane.
- the modified cyanobacteria secrete extracellular electrons or substances or molecules having electrons into cells, and take up extracellular electrons or substances or molecules having electrons into cells at least one of them. It can be carried out. As a result, the produced modified cyanobacteria have improved extracellular electron transfer efficiency. Therefore, according to the method for manufacturing an electron carrier according to one aspect of the present disclosure, it is possible to provide an electron carrier with improved electron transfer efficiency with the outside.
- any of the above electron carriers is used.
- the electron carrier contains modified cyanobacteria with improved extracellular electron transfer efficiency, it is possible to efficiently donate electrons to the outside (for example, an external electrode) and efficiently receive electrons from the outside. .. Therefore, if an electron carrier is used, the modified cyanobacteria contained in the electron carrier releases the intracellular electron transport chain to the outside of the cell, thereby efficiently supplying electrons to an external electrode, for example, to generate an electric current. Can be done. Further, for example, the modified cyanobacteria contained in the electron carrier can receive electrons instead of light energy from the outside to perform photosynthesis or respiration. As a result, the modified cyanobacteria contained in the electron carrier can also produce useful substances such as proteins inside the cells and secrete them extracellularly.
- each figure is not necessarily exactly illustrated.
- substantially the same configuration is designated by the same reference numerals, and duplicate description may be omitted or simplified.
- the numerical range does not only represent a strict meaning, but also includes measuring a substantially equivalent range, for example, the amount of protein (for example, number or concentration, etc.) or the range thereof.
- both the bacterial cell and the cell represent one cyanobacterial individual.
- the identity of the base sequence and the amino acid sequence is calculated by the BLAST (Basic Local Alignment Search Tool) algorithm. Specifically, it is calculated by performing pairwise analysis with the BLAST program available on the website of NCBI (National Center for Biotechnology Information) (https://blast.ncbi.nlm.nih.gov/Blast.cgi). To. Information on cyanobacterial and plant genes and the proteins encoded by these genes is published, for example, in the NCBI database described above and Cyanobase (http://genome.microbedb.jp/cyanobase/). From these databases, the amino acid sequence of the protein of interest and the base sequence of the gene encoding those proteins can be obtained.
- NCBI National Center for Biotechnology Information
- Cyanobacteria also called cyanobacteria or cyanobacteria, collect light energy with chlorophyll and use the obtained energy to cause charge separation of the reaction center chlorophyll, thereby electrolyzing water and generating oxygen while photosynthesizing. It is a group of prokaryotes. Cyanobacteria are rich in diversity, for example, in cell shape, there are unicellular species such as Synechocysis sp. PCC 6803 and multicellular filamentous species such as Anabaena sp. PCC 7120.
- thermophilic species such as Thermosynechococcus elongatus
- marine species such as Synechococcus elongatus
- freshwater species such as Synechocystis.
- Microcystis aeruginosa a species that has gas vesicles and produces toxins
- Gloeobacter violaceus which has a protein called phycobilisome that is a focusing antenna on the plasma membrane without thylakoids.
- phycobilisome a protein called phycobilisome that is a focusing antenna on the plasma membrane without thylakoids.
- cyanobacteria electrons are generated inside the cell when water is decomposed by photosynthesis and when organic compounds such as sugar synthesized by photosynthesis are catabolized as their own nutrient source.
- the electrons generated by the decomposition of water flow through the photosynthetic electron transport chain existing on the thylakoid membrane, which is the membrane structure in the cytoplasm, and in the process generate a proton driving force used as a bioenergy source, and finally. It is used in the reaction to reduce NADP + and produce NADPH.
- FIG. 1 is a diagram schematically showing the cell surface layer of cyanobacteria.
- the cell surface layer of cyanobacteria is composed of a plasma membrane (also referred to as inner membrane 1), peptidoglycan 2, and an outer membrane 5, which is a lipid membrane forming the outermost layer of cells, in this order from the inside.
- a plasma membrane also referred to as inner membrane 1
- peptidoglycan 2 and an outer membrane 5, which is a lipid membrane forming the outermost layer of cells, in this order from the inside.
- a sugar chain 3 composed of glucosamine, mannosamine, etc. is covalently bound to peptidoglycan 2, and pyruvate is covalently bound to these covalently bound sugar chains 3 (Non-Patent Document 9: Jurgens and). Weckesser, 1986, J. Bacteriol., 168: 568-573).
- the peptidoglycan 2 and the covalently bound sugar chain 3 are collectively referred to as a cell wall 4.
- the gap between the plasma membrane (that is, the inner membrane 1) and the outer membrane 5 is called periplasm, and is called periplasm, which decomposes proteins or forms a three-dimensional structure, decomposes lipids or nucleic acids, or takes up extracellular nutrients.
- periplasm which decomposes proteins or forms a three-dimensional structure, decomposes lipids or nucleic acids, or takes up extracellular nutrients.
- periplasm which decomposes proteins or forms a three-dimensional structure, decomposes lipids or nucleic acids, or takes up extracellular nutrients.
- enzymes involved in There are various enzymes involved in.
- the enzyme that catalyzes the pyruvate modification reaction of the covalent sugar chain 3 in peptidoglycan 2 (hereinafter referred to as cell wall-pyruvate modifying enzyme 9) has been identified in the Gram-positive bacterium Bacillus anthracis and is named CsaB.
- Non-Patent Document 10 Mesnage et al., 2000, EMBO J., 19: 4473-4484.
- cyanobacteria whose genomic nucleotide sequence is open to the public, many species carry a gene encoding a homologous protein having an amino acid sequence identity of 30% or more with CsaB. Examples include slr0688 held by Synechococcus sp. PCC 6803 or syn7502_03092 held by Synechococcus sp. 7502.
- Non-Patent Document 11 Kowata et al., 2017).
- J. Bacteriol., 199: e00371-17 It is known that peptidoglycan-bound sugar chains are pyruvic acid-modified for the binding between the outer membrane protein and the cell wall.
- Non-Patent Document 12 Kojima et al., 2016, Biosci. Biotech. Biochem., 10 : 1954-1959).
- a channel protein is a membrane protein that forms a pathway (that is, a channel) for selectively permeating a predetermined substance from the inside to the outside or from the outside to the inside of a lipid membrane (for example, outer membrane 5).
- the outer membrane of common heterotrophic Gram-negative bacteria such as Escherichia coli and Salmonella, selectively permeates relatively low-molecular-weight nutrients such as sugars and amino acids from the outside to the inside of the outer membrane into cells.
- Porin for uptake Non-Patent Document 13: Nikaido, 2003, Microbiol. Mol. Biol. Rev., 67 (4): 593-656).
- Porin does not exist in the outer membrane 5 of cyanobacteria, and instead, a large amount of ion channel proteins (for example, SLH domain-retaining outer membrane protein 6) that selectively permeate only inorganic ions are present in the outer membrane 5.
- ion channel protein accounts for about 80% of the total protein of the outer membrane 5 (Non-Patent Document 11). Therefore, in cyanobacteria, unless the properties of the outer membrane 5 are significantly modified by using a technique such as gene transfer, a high molecular weight substance such as a protein permeates the outer membrane 5 and is extracellular (that is, the outer membrane 5). It is difficult to spread to (outside).
- the chloroplasts of plants originated from cyanobacteria that coexisted in the cells of primitive eukaryotic cells about 1.5 to 2 billion years ago, and were transformed into chloroplasts by subsequent evolution (Non-Patent Documents). 14: Ponce-Toledo et al., 2017, Curr. Biol., 27 (3): 386-391).
- the chloroplasts carried by gray algae, which are the most primitive plants of unicellular algae, have peptidoglycan and retain a surface structure similar to that of cyanobacteria.
- peptidoglycan does not exist in the chloroplasts of seed plants that have evolved more than unicellular algae.
- the above-mentioned chloroplast outer membrane protein of Glaucophyte is significantly different from the composition of the outer membrane protein of cyanobacteria.
- the outer membrane 5 of cyanobacteria contains a large amount of ion channel proteins that allow inorganic substances such as Slr1841 (SLH domain-retaining outer membrane protein 6) to permeate.
- the ion channel protein accounts for about 80% of the total protein in the outer membrane 5.
- the outer membrane of the chloroplast of Glaucophyte contains a large amount of channel proteins (hereinafter, also referred to as organic channel proteins 18) that allow organic substances named CppS and CppF to permeate.
- the organic channel protein 18 accounts for 80% or more of the total protein in the outer membrane of the chloroplast of gray algae (Non-Patent Document 15: Kojima et al., 2016, J. Biol. Chem., 291: 20198-20209. ).
- CppS and CppF are channel proteins having a channel function that selectively permeates relatively high-molecular-weight organic substances (for example, biomolecules such as proteins), and are used to control the inside of chlorophyll in plant cells and the cytoplasm of plant cells. It is thought to function as a connecting substance transport route.
- CppS and CppF are widely distributed in gray algae.
- CppS and CppF-like proteins are present only in bacteria belonging to the phylum Planctomycetes. Cyanobacteria do not carry CppS and CppF and their similar proteins (see Non-Patent Document 15).
- FIG. 2 is a schematic view showing an example of the electron carrier 30 according to the present embodiment.
- the electron carrier 30 has a function of supplying electrons to the outside and taking in electrons from the outside.
- the outside is a substance or molecule that exists as an individual different from the electron carrier 30, and is, for example, a redox substance involved in the movement of electrons between substances, or a molecule having a redox reactive group.
- supplying electrons means not only supplying electrons but also supplying any substance or molecule having electrons.
- taking in an electron means not only taking in an electron but also taking in any substance or molecule having an electron.
- the function of the protein involved in the binding between the outer membrane 5 and the cell wall 4 in cyanobacteria (the parent cyanobacteria described later) is suppressed or lost, and (Ii) Includes modified cyanobacteria 31, which is at least one of which a channel protein (so-called organic channel protein 18) that improves protein permeability of the outer membrane 5 is expressed. Further, the modified cyanobacteria 31 performs at least one of supplying electrons to the outside of the electron carrier 30 and taking in electrons from the outside.
- the electron carrier 30 may be the modified cyanobacteria 31 of the above (i), the modified cyanobacteria 31 of the above (ii), or the modified cyanobacteria of the above (i) and (ii). It may be 31.
- the bond between the cell wall 4 and the outer membrane 5 (for example, the amount of binding and the binding force) is partially reduced, so that the outer membrane 5 is partially detached from the cell wall 4. It will be easier. Therefore, the electrons generated inside the cells of the modified cyanobacteria 31 or the substances or molecules having electrons easily leak out of the outer membrane 5, that is, outside the cells. Further, according to the above (ii), in the modified cyanobacteria 31, the protein permeability of the outer membrane 5 is improved, so that the substance permeability of the outer membrane 5 is improved.
- the modified cyanobacteria 31 secretes intracellularly generated electrons or substances or molecules having electrons to the outside of the cell, and takes up extracellular electrons or substances or molecules having electrons to the inside of the cell. At least one of can be done. As a result, the modified cyanobacteria 31 has improved extracellular electron transfer efficiency. Therefore, the electron transfer body 30 according to the present embodiment improves the electron transfer efficiency with the outside.
- the electron carrier 30 may contain at least one of the electron transfer substance 33, the electron mediator 35, and the conductive substance 37 in addition to the at least one modified cyanobacteria 31 of the above (i) and (ii). Good.
- the electron carrier 30 may include a modified cyanobacteria 31, an electron carrier 33, an electron mediator 35, and a conductive substance 37.
- the extracellular electron transfer efficiency of the modified cyanobacteria 31 can be further improved.
- the electron transfer substance 33 is a substance responsible for an electron transfer reaction, and is a so-called redox substance including an oxidation type substance that receives electrons and a reduction type substance that gives electrons.
- the electron transport chain 33 is not particularly limited as long as it is a redox substance involved in the electron transport chain in cells, and for example, non-heme iron such as peptides, proteins, flavins, quinones, heme iron, and iron-sulfur clusters. Alternatively, it may be a copper ion or the like.
- the electron mediator 35 is a substance that assists or promotes the electron transfer function of the electron transfer substance 33, and is a so-called redox active species.
- the electron mediator 35 may be, for example, a quinone, a phenothen, a ferricianide, a cytochrome, a viologen, a phenazine, a phenoxazine, a phenothiazine, a ferredoxin, or a derivative thereof.
- a substance may be appropriately selected depending on the type.
- the conductive substance 37 is a substance having a property that electrons easily move in the substance, and for example, even if it is selected from one or more materials selected from the group consisting of carbon-based substances, conductive polymers, semiconductors and metals. Good.
- the carbon-based substance means a substance containing carbon as a constituent component.
- the carbon-based substance may be carbon powder such as graphite, activated carbon or carbon black, carbon fiber such as graphite felt, carbon wool or carbon woven fabric, carbon nanotubes, carbon plate, carbon paper or carbon disc.
- the conductive polymer is a general term for polymer compounds having conductivity.
- the conductive polymer is, for example, a polymer of a single monomer or two or more kinds of monomers whose constituent units are aniline, aminophenol, diaminophenol, pyrrole, thiophene, paraphenylene, fluorene, furan, acetylene or derivatives thereof. There may be. More specifically, the conductive polymer may be, for example, polyaniline, polyaminophenol, polydiaminophenol, polypyrrole, polythiophene, polyfuran, polyacetylene or the like.
- the conductive substance 37 may be a metal or a metal oxide, and from the viewpoint of further increasing the current production amount, tungsten, tungsten oxide, copper, silver, platinum, gold, niobium, iron, cobalt, titanium and molybdenum. , Molybdenum oxide, tin, tin oxide, nickel, nickel oxide, alloys containing these, or oxides thereof.
- modified cyanobacteria 31 [3. Modified cyanobacteria] Subsequently, the modified cyanobacteria 31 will be described. In this embodiment, the modified cyanobacteria 31 is included in the electron carrier 30.
- the modified cyanobacteria 31 for example, receives light to generate electrons and emits the generated electrons to the outside of the outer membrane 5.
- the modified cyanobacteria 31 releases an electron or a substance or molecule having an electron to the outside of the cell (that is, outside the outer membrane 5). Therefore, when the electron carrier 30 receives light, it generates electrons inside and can supply electrons or substances or molecules having electrons to the outside.
- the modified cyanobacteria 31 takes in the electrons existing on the outside of the outer membrane 5 into the inside of the cell wall 4 (that is, in the cytoplasm), and utilizes the electrons inside the cell wall 4.
- the modified cyanobacteria 31 takes in electrons or substances or molecules having electrons existing outside the cell into the cell (in the cytoplasm), and for example, energy (ATP: adenosine) in the process of flowing through the photosynthetic electron transport chain. triphosphate) is generated.
- the modified cyanobacteria 31 uses this energy to produce an organic substance based on carbon dioxide. Therefore, the electron carrier 30 can take in electrons from the outside to the inside to generate energy and produce an organic substance such as a protein.
- the function of a protein involved in the binding between the outer membrane 5 and the cell wall 4 (hereinafter, also referred to as a binding-related protein) in the cyanobacteria is suppressed or lost, and
- At least one of the channel proteins (so-called organic channel proteins 18) that improve the protein permeability of the outer membrane 5 are expressed.
- the protein involved in the binding between the outer membrane 5 and the cell wall 4 may be, for example, at least one of the SLH domain-retaining outer membrane protein 6 and the cell wall-pyruvic acid modifying enzyme 9.
- the modified cyanobacteria 31 have, for example, suppressed or lost the function of at least one protein of SLH domain-retaining outer membrane protein 6 and cell wall-pyruvic acid modifying enzyme 9.
- at least one function of (a) SLH domain-retaining outer membrane protein 6 and cell wall-pyruvate modifying enzyme 9 may be suppressed or lost, and (b) SLH binding to cell wall 4 may be suppressed.
- the modified cyanobacteria 31 for example, (a) an enzyme that catalyzes the reaction of modifying the SLH domain-retaining outer membrane protein 6 that binds to the cell wall and the bound sugar chain on the surface of the cell wall with pyruvate (that is, the cell wall-pilvin). At least one function of the acid modifying enzyme 9) is suppressed or lost, or (b) the expression of at least one of the SLH domain-retaining outer membrane protein 6 and the cell wall-pyruvate modifying enzyme 9 is suppressed. There is.
- the binding that is, the binding amount and binding force
- the outer membrane 5 is likely to be detached from the cell wall 4 at the portion where the bond between the outer membrane 5 and the cell wall 4 is weakened.
- the modified cyanobacteria 31 can secrete the intracellular electron transmitter 33 out of the cell and take up the extracellular electron transmitter 33 into the cell at least one of them.
- the efficiency of extracellular electron transfer is improved. Therefore, the electron transfer body 30 according to the present embodiment improves the electron transfer efficiency with the outside.
- the type of cyanobacteria (hereinafter referred to as parent cyanobacteria) before the expression of the channel protein (that is, the organic channel protein 18) that improves the protein permeability of the membrane 5 is not particularly limited, and any type of cyanobacteria can be used. There may be.
- the parent cyanobacteria may be of the genus Synechococcus, Synechococcus, Anabaena, or Thermosynechococcus, among which Synechococcus sp. PCC 6803, Synechococcus sp. PCC 7942, or Thermosynechococcus elongatus BP-1. May be good.
- the nucleotide sequences of the genes encoding these binding-related proteins and the positions of the genes on the chromosomal DNA or plasmid can be confirmed in the NCBI database and Cyanobase described above.
- the protein involved in the binding between the outer membrane and the cell wall whose function is suppressed or lost in the modified cyanobacteria 31 in the present embodiment is that of any parent cyanobacteria as long as it is possessed by the parent cyanobacteria. They may be, and are not limited by the location of the genes encoding them (eg, on chromosomal DNA or plasmids).
- the SLH domain-retaining outer membrane protein 6 may be Slr1841, Slr1908, Slr0042, etc. when the parent cyanobacteria belongs to the genus Synchocystis, or NIES970_09470 or the like when the parent cyanobacteria belongs to the genus Synechococcus.
- the parent cyanobacteria belongs to the genus Anabaena, it may be Anacy_5815 or Anacy_3458, etc.
- the parent cyanobacteria belongs to the genus Microcystis it may be A0A0F6U6F8_MICAE, etc.
- the parent cyanobacteria belongs to the genus Leptolyngbya, it may be A0A1Q8ZE23_9CYAN, etc., if the parent cyanobacteria belongs to the genus Calothrix, it may be A0A1Z4R6U0_9CYAN, and if the parent cyanobacteria belongs to the genus Nostoc, it may be A0A1C0VG86_9NOSO.
- the parent cyanobacteria belongs to the genus Crocosphaera, it may be B1WRN6_CROS5 or the like, and if the parent cyanobacteria belongs to the genus Pleurocapsa, it may be K9TAE4_9CYAN or the like.
- the SLH domain-retaining outer membrane protein 6 is, for example, Slr1841 (SEQ ID NO: 1) of Synechococcus sp. PCC 6803, NIES970_09470 (SEQ ID NO: 2) of Synechococcus sp. NIES-970, or Anabaena cylindrica PCC. It may be Anacy_3458 (SEQ ID NO: 3) of 7122 or the like. Further, it may be a protein having an amino acid sequence of 50% or more identical to these SLH domain-retaining outer membrane proteins 6.
- any SLH domain-retaining outer membrane protein 6 shown in SEQ ID NOs: 1 to 3 above, or any of these SLH domain-retaining outer membrane proteins 6 and The function of the protein having the same amino acid sequence of 50% or more may be suppressed or lost, and (b) any SLH domain-retaining outer membrane protein 6 shown in SEQ ID NOs: 1 to 3 above, or any of these.
- the expression of a protein having an amino acid sequence that is 50% or more identical to that of the SLH domain-retaining outer membrane protein 6 may be suppressed.
- the modified cyanobacteria 31 (a) the function of the SLH domain-retaining outer membrane protein 6 or the protein having the same function as the SLH domain-retaining outer membrane protein 6 in the outer membrane 5 is suppressed or lost, or , (B) The expression level of the SLH domain-retaining outer membrane protein 6 or the protein having the same function as the SLH domain-retaining outer membrane protein 6 in the outer membrane 5 is reduced.
- the binding domain for example, SLH domain 7
- the electron transfer body 30 improves the electron transfer efficiency with the outside.
- the SLH domain-retaining outer membrane protein 6 whose function is suppressed or lost includes, for example, 40% of the amino acid sequence of any of the SLH domain-retaining outer membrane proteins shown in SEQ ID NOs: 1 to 3 above.
- the cell wall 4 comprises an amino acid sequence having an identity of preferably 50% or more, more preferably 60% or more, further preferably 70% or more, still more preferably 80% or more, still more preferably 90% or more. It may be a protein or polypeptide having a function of binding to the covalent bond type sugar chain 3.
- the cell wall-pyruvate modifying enzyme 9 may be Slr0688 or the like when the parent cyanobacteria belongs to the genus Synchocystis, or may be Syn7502_03092 or Synpcc7942_1529 or the like when the parent cyanobacteria belongs to the genus Synechococcus.
- the cyanobacteria belongs to the genus Anabaena, it may be ANA_C20348 or Anacy_1623, etc.
- the parent cyanobacteria belongs to the genus Microcystis, it may be CsaB (NCBI access ID: TRU80220), etc.
- the parent cyanobacteria belongs to the genus Cyanothese.
- it may be CsaB (NCBI access ID: WP_107667006.1) or the like
- the parent cyanobacteria belongs to the genus Spirulina
- it may be CsaB (NCBI access ID: WP_026079530.1) or the like.
- CsaB NCBI access ID: WP_096658142.1
- the parent cyanobacteria is of the genus Nostoc
- it may be CsaB (NCBI access ID: WP_099068528.1) or the like.
- the parent cyanobacteria belongs to the genus Crocosphaera, it may be CsaB (NCBI access ID: WP_012361697.1), etc.
- the parent cyanobacteria belongs to the genus Pleurocapsa it may be CsaB (NCBI access ID: WP_036798735), etc. May be good.
- the cell wall-pyruvic acid modifying enzyme 9 is, for example, Slr0688 (SEQ ID NO: 4) of Synechococcus sp. PCC 6803, Synpcc 7942_1529 (SEQ ID NO: 5) of Synechococcus sp. PCC 7942, or Anabaena cylindrica PCC 7122. It may be Anacy_1623 (SEQ ID NO: 6) or the like. Further, it may be a protein having an amino acid sequence of 50% or more identical to these cell wall-pyruvic acid modifying enzymes 9.
- any cell wall-pyruvate modifying enzyme 9 shown in SEQ ID NOs: 4 to 6 above, or any of these cell walls-pyruvate modifying enzyme 9 and an amino acid sequence The function of the protein is suppressed or lost by 50% or more, or (b) any cell wall shown in SEQ ID NOs: 4 to 6 above-the cell wall of pyruvate modifying enzyme 9 or any of these. -Expression of a protein having an amino acid sequence 50% or more identical to that of pyruvate modifying enzyme 9 is suppressed.
- the function of (a) cell wall-pyruvic acid modifying enzyme 9 or a protein having a function equivalent to that enzyme is suppressed or lost, or (b) cell wall-pyruvic acid modifying enzyme 9 or The expression level of a protein having the same function as that of the enzyme is reduced.
- the covalently bound sugar chain 3 on the surface of the cell wall 4 is less likely to be modified with pyruvic acid, so that the sugar chain 3 of the cell wall 4 becomes the SLH domain 7 of the SLH domain-retaining outer membrane protein 6 in the outer membrane 5.
- the amount of binding and the binding force are reduced.
- the electron transfer body 30 improves the electron transfer efficiency with the outside.
- the cell wall-pyruvate modifying enzyme 9 whose function is suppressed or lost includes, for example, 40% or more of the amino acid sequence of any of the cell wall-pyruvate modifying enzymes 9 shown in SEQ ID NOs: 4 to 6 above.
- Peptidoglycan consisting of an amino acid sequence having an amino acid sequence of preferably 50% or more, more preferably 60% or more, still more preferably 70% or more, even more preferably 80% or more, still more preferably 90% or more, and having cell wall 4 peptide glycan. It may be a protein or polypeptide having a function of catalyzing a reaction of modifying the covalent sugar chain 3 of 2 with pyruvate.
- suppressing or losing the function of SLH domain-retaining outer membrane protein 6 means suppressing or losing the ability of the protein to bind to the cell wall 4, and to the outer membrane 5 of the protein.
- the transport of the protein is suppressed or lost, or the ability of the protein to implant and function in the outer membrane 5 is suppressed or lost.
- suppressing or losing the function of the cell wall-pyruvic acid modifying enzyme 9 means that the protein suppresses or loses the function of modifying the covalent sugar chain 3 of the cell wall 4 with pyruvic acid.
- the means for suppressing or losing the function of these proteins is not particularly limited as long as it is a means usually used for suppressing or losing the function of the protein.
- the means include, for example, deleting or inactivating the gene encoding the SLH domain-retaining outer membrane protein 6 and the gene encoding the cell wall-pyruvate modifying enzyme 9, and inhibiting the transcription of these genes. It may be to inhibit the translation of transcripts of these genes, or to administer an inhibitor that specifically inhibits these proteins.
- the gene expressing the protein involved in the binding between the outer membrane 5 and the cell wall 4 may be deleted or inactivated.
- the modified cyanobacteria 31 of (i) above the expression of the protein involved in the binding between the cell wall 4 and the outer membrane 5 is suppressed, or the function of the protein is suppressed or lost, so that the cell wall is suppressed.
- the bond between 4 and the outer membrane 5 (so-called bond amount and bond force) is partially reduced.
- the outer membrane 5 is likely to be detached from the cell wall 4 at the portion where the bond between the outer membrane 5 and the cell wall 4 is weakened.
- intracellular electrons or substances or molecules having electrons are secreted extracellularly, and extracellular electrons or substances or molecules having electrons are intracellularly secreted. Since at least one of uptake can be performed, the efficiency of extracellular electron transfer is improved. Therefore, the electron transfer body 30 according to the present embodiment improves the electron transfer efficiency with the outside.
- the gene that expresses the protein involved in the binding between the outer membrane 5 and the cell wall 4 is at least one of the gene encoding the SLH domain-retaining outer membrane protein 6 and the gene encoding the cell wall-pyruvate modifying enzyme 9. There may be.
- the modified cyanobacteria 31 at least one gene encoding the SLH domain-retaining outer membrane protein 6 and the gene encoding the cell wall-pyruvic acid modifying enzyme 9 are deleted or inactivated. Therefore, in the modified cyanobacteria 31, for example, the expression of at least one of (a) SLH domain-retaining outer membrane protein 6 and cell wall-pyruvic acid modifying enzyme 9 is suppressed, or (b) SLH domain-retaining outer membrane.
- At least one function of protein 6 and cell wall-pyruvate modifying enzyme 9 is suppressed or lost. Therefore, the binding (that is, the binding amount and binding force) between the SLH domain 7 of the SLH domain-retaining outer membrane protein 6 in the outer membrane 5 and the covalently bound sugar chain 3 on the surface of the cell wall 4 is reduced.
- the outer membrane 5 is more likely to be detached from the cell wall 4 at the portion where the bond between the outer membrane 5 and the cell wall 4 is weakened, so that the intracellular electron or the substance or molecule having an electron is extracellular. It becomes easy to leak to. Therefore, in the electron carrier 30 according to the present embodiment, the intracellular electron or the substance or molecule having an electron in the modified cyanobacteria 31 is likely to leak out of the cell, so that the electron transfer efficiency with the outside is improved.
- SLH domain-retaining outer membrane protein 6 is used to suppress or eliminate at least one function of SLH domain-retaining outer membrane protein 6 and cell wall-pyruvate modifying enzyme 9 in cyanobacteria.
- the encoding gene and the cell wall-the transcription of at least one of the genes encoding the pyruvate modifying enzyme 9 may be suppressed.
- the gene encoding SLH domain-bearing outer membrane protein 6 may be slr1841, slr1908, slr0042, etc. when the parent cyanobacteria belongs to the genus Syenchocystis, or may be nies970_09470, etc. in the case of the genus Synechococcus.
- the parent cyanobacteria belongs to the genus Anabaena, it may be anacy_5815 or anacy_3458, etc.
- the parent cyanobacteria belongs to the genus Microcystis it may be A0A0F6U6F8_MICAE, etc.
- the parent cyanobacteria belongs to the genus Leptolyngbya, it may be A0A1Q8ZE23_9CYAN, etc., if the parent cyanobacteria belongs to the genus Calothrix, it may be A0A1Z4R6U0_9CYAN, etc., if the parent cyanobacteria belongs to the genus Nostoc, A0A1C0VG86_9NOSO, etc.
- the parent cyanobacteria belongs to the genus Crocosphaera, it may be B1WRN6_CROS5 or the like, and if the parent cyanobacteria belongs to the genus Pleurocapsa, it may be K9TAE4_9CYAN or the like.
- the nucleotide sequences of these genes can be obtained from the NCBI database described above or Cyanobase.
- genes encoding SLH domain-retaining outer membrane protein 6 are slr1841 (SEQ ID NO: 7) of Synechococcus sp. PCC 6803, nies970_09470 (SEQ ID NO: 8) of Synechococcus sp. NIES-970, and Anabaena cylindrica PCC. It may be anacy_3458 (SEQ ID NO: 9) of 7122, or a gene whose amino acid sequence is 50% or more identical to these genes.
- the modified cyanobacteria 31 is 50% or more identical to the gene encoding any of the SLH domain-bearing outer membrane proteins 6 shown in SEQ ID NOs: 7 to 9 above or the base sequence of any of these genes.
- a gene is deleted or inactivated. Therefore, in the modified cyanobacteria 31, (a) expression of any of the above SLH domain-retaining outer membrane proteins 6 or a protein having a function equivalent to that of any of these proteins is suppressed, or (b) The function of any of the above SLH domain-retaining outer membrane proteins 6 or a protein having a function equivalent to that of any of these proteins is suppressed or lost.
- the modified cyanobacteria 31 improves the extracellular electron transfer efficiency, so that the electron carrier 30 according to the present embodiment improves the electron transfer efficiency with the outside.
- the amino acid sequence of a protein is 30% or more the same, it is said that there is a high possibility that it has the same function as the protein. Therefore, if the base sequence of the gene encoding the protein is 30% or more the same, it is considered that there is a high possibility that a protein having the same function as the protein will be expressed. Therefore, the gene encoding the SLH domain-retaining outer membrane protein 6 whose function is suppressed or lost is, for example, any of the genes encoding the SLH domain-retaining outer membrane protein 6 shown in SEQ ID NOs: 7 to 9 above.
- the gene encoding the cell wall-pyruvate modifying enzyme 9 may be slr0688 or the like when the parent cyanobacteria belongs to the genus Synchocystis, or syn7502_03092 or synpcc7942_1529 or the like when the parent cyanobacteria belongs to the genus Synechococcus. If the parent cyanobacteria belongs to the genus Anabaena, it may be ana_C20348 or anacy_1623, etc., and if the parent cyanobacteria belongs to the genus Microcystis, it may be csaB (NCBI access ID: TRU80220), etc.
- Cynahothese it may be csaB (NCBI access ID: WP_107667006.1), etc., and if the parent cyanobacteria is Spirulina, it may be csaB (NCBI access ID: WP_026079530.1), etc.
- the parent cyanobacteria belongs to the genus Calothrix, it may be csaB (NCBI access ID: WP_096658142.1), etc.
- the parent cyanobacteria belongs to the genus Nostoc it may be csaB (NCBI access ID: WP_099068528.1), etc.
- the parent cyanobacteria belongs to the genus Crocosphaera, it may be csaB (NCBI access ID: WP_012361697.1), etc., and if the parent cyanobacteria belongs to the genus Pleurocapsa, csaB (NCBI access ID: WP_036798735). And so on.
- the nucleotide sequences of these genes can be obtained from the NCBI database described above or Cyanobase.
- the gene encoding the cell wall-pyruvic acid modifying enzyme 9 is slr0688 (SEQ ID NO: 10) of Synechococcus sp. PCC 6803, synpcc7942_1529 (SEQ ID NO: 11) of Synechococcus sp. PCC 7942, or Anabaena cylindrica PCC. It may be anacy_1623 (SEQ ID NO: 12) of 7122. Moreover, it may be a gene whose base sequence is 50% or more identical to these genes.
- the modified cyanobacteria 31 50% with the base sequence of the gene encoding any cell wall-pyruvic acid modifying enzyme 9 shown in SEQ ID NOs: 10 to 12 above or the gene encoding any of these enzymes. The same gene is deleted or inactivated. Therefore, in the modified cyanobacteria 31, (a) expression of a protein having a function equivalent to that of any of the above cell wall-pyruvic acid modifying enzymes 9 or any of these enzymes is suppressed, or (b) the above. The function of any of the cell walls-pyruvic acid modifying enzyme 9 or a protein having a function equivalent to that of any of these enzymes is suppressed or lost.
- the covalently bound sugar chain 3 on the surface of the cell wall 4 is less likely to be modified with pyruvic acid, so that the sugar chain 3 of the cell wall 4 becomes the SLH domain 7 of the SLH domain-retaining outer membrane protein 6 in the outer membrane 5.
- the amount of binding and the binding force are reduced.
- the amount of the covalently bound sugar chain 3 on the surface of the cell wall 4 modified with pyruvic acid is reduced, so that the binding force between the cell wall 4 and the outer membrane 5 is weakened, and the outer membrane 5 is weakened. Is more likely to partially detach from the cell wall 4.
- the electron transfer body 30 improves the electron transfer efficiency with the outside.
- the gene encoding the cell wall-pyruvate modifying enzyme 9 whose function is suppressed or lost is, for example, any of the genes encoding the cell wall-pyruvate modifying enzyme 9 shown in SEQ ID NOs: 10 to 12 above. From a base sequence having 40% or more, preferably 50% or more, more preferably 60% or more, still more preferably 70% or more, still more preferably 80% or more, still more preferably 90% or more identity with the base sequence. It may also be a gene encoding a protein or polypeptide having a function of catalyzing a reaction of modifying the covalent sugar chain 3 of the peptidoglycan 2 of the cell wall 4 with pyruvate.
- expressing the organic channel protein 18 on the outer membrane 5 of the cyanobacteria means that the gene encoding the organic channel protein 18 is inserted into the chromosomal DNA or plasmid of the cyanobacteria and synthesized through transcription and translation of the gene.
- the organic channel protein 18 is transported to the outer membrane 5 and expresses a channel function for selectively permeating the gene in the outer membrane 5 of cyanobacteria.
- the means for inserting and expressing the gene is not particularly limited as long as it is a commonly used means, the base sequence of the promoter for transcription activation, the ribosome binding sequence for translation, and the transport to the outer membrane 5. It is not limited by the type of signal sequence for.
- the organic channel protein 18 expressed on the outer membrane 5 of cyanobacteria may be an outer membrane channel protein derived from chloroplasts.
- the organic channel protein 18 may be, for example, CppS (SEQ ID NO: 13) or CppF (SEQ ID NO: 14) of the gray alga Cyanophora paradoxa (hereinafter, also referred to as C. paradoxa).
- the organic channel protein 18 may be a protein having an amino acid sequence 50% or more identical to that of CppS or CppF.
- the protein having an amino acid sequence of 50% or more identical to CppS or CppF is not limited to a protein derived from chloroplasts, and may be, for example, a protein similar to CppS or CppF derived from a microorganism such as a bacterium.
- CppS SEQ ID NO: 13
- CppF SEQ ID NO: 14
- a protein having a function equivalent to that of the organic channel protein 18 of the above is expressed. Therefore, in the modified cyanobacteria 31 of the above (ii), the protein permeability of the outer membrane 5 is improved, so that the substance permeability of the outer membrane 5 is improved.
- the modified cyanobacteria 31 of the above (ii) secretes intracellular electrons or substances or molecules having electrons to the outside of the cell, and extracellular electrons or substances or molecules having electrons to the inside of the cell. Since at least one of uptake can be performed, the efficiency of extracellular electron transfer is improved. Therefore, the electron transfer body 30 according to the present embodiment improves the electron transfer efficiency with the outside.
- the organic channel protein 18 includes, for example, 40% or more, preferably 50% or more, more preferably 60% or more, and further, with the amino acid sequence of any of the proteins shown in SEQ ID NO: 13 and SEQ ID NO: 14 above.
- the modified cyanobacteria 31 may be introduced with a gene encoding an organic channel protein 18 that improves the protein permeability of the outer membrane 5.
- the organic channel protein 18 that improves the protein permeability of the outer membrane 5 is expressed. Therefore, in the modified cyanobacteria 31 of the above (ii), the protein permeability of the outer membrane 5 is improved, so that the substance permeability of the outer membrane 5 is improved.
- the modified cyanobacteria 31 of the above (ii) secretes intracellular electrons or substances or molecules having electrons to the outside of the cell, and extracellular electrons or substances or molecules having electrons to the inside of the cell. Since at least one of uptake can be performed, the efficiency of extracellular electron transfer is improved. Therefore, the electron transfer body 30 according to the present embodiment improves the electron transfer efficiency with the outside.
- the above gene may be, for example, a gene derived from a chloroplast.
- the gene encoding the organic channel protein 18 derived from chloroplast may be, for example, cppS (SEQ ID NO: 15) or cppF (SEQ ID NO: 16) of the gray alga Cyanophora paradoxa.
- the organic channel protein 18 may be a gene having a base sequence of 50% or more identical to any of these genes.
- the modified cyanobacteria 31 of (ii) above a protein having a function of improving protein permeability in the outer membrane 5 or a protein having a function equivalent to the protein is expressed.
- the protein permeability of the outer membrane 5 is improved, so that the substance permeability of the outer membrane 5 is improved.
- the modified cyanobacteria 31 of the above (ii) secretes intracellular electrons or substances or molecules having electrons to the outside of the cell, and extracellular electrons or substances or molecules having electrons to the inside of the cell. Since at least one of uptake can be performed, the efficiency of extracellular electron transfer is improved. Therefore, the electron transfer body 30 according to the present embodiment improves the electron transfer efficiency with the outside.
- the gene encoding the organic channel protein 18 is not limited to the chloroplast-derived gene.
- Examples of the gene encoding the organic channel protein 18 include the base sequence of any of the above genes cppS (SEQ ID NO: 15) and cppF (SEQ ID NO: 16) and 40% or more, preferably 50% or more, more preferably 60. % Or more, more preferably 70% or more, even more preferably 80% or more, still more preferably 90% or more, and a protein having a function of improving the protein permeability of the outer membrane 5.
- it may be a gene encoding a polypeptide.
- the electron transmitter 30 (i) suppresses or loses the function of the protein involved in the binding between the outer membrane 5 and the cell wall 4 in cyanobacteria, and (ii) improves the protein permeability of the outer membrane 5. It comprises the step of producing the modified cyanobacteria 31 which is at least one of the organic channel proteins 18 expressed (hereinafter referred to as the production step of the modified cyanobacteria 31).
- the steps for producing the modified cyanobacteria 31 are (i) a step of suppressing or losing the function of a protein involved in the binding of the outer membrane 5 to the cell wall 4 in cyanobacteria, and (ii) the protein permeability of the outer membrane 5. Includes at least one step of expressing the improving organic channel protein 18.
- the protein involved in the binding between the outer membrane 5 and the cell wall 4 may be, for example, at least one of the SLH domain-retaining outer membrane protein 6 and the cell wall-pyruvic acid modifying enzyme 9.
- the means for suppressing or losing the function of the protein is not particularly limited, but for example, the gene encoding the SLH domain-retaining outer membrane protein 6 and the gene encoding the cell wall-pyruvate modifying enzyme 9 are deleted or deleted. Even by inactivating, inhibiting the transcription of these genes, inhibiting the translation of transcripts of these genes, or administering inhibitors that specifically inhibit these proteins, etc. Good.
- the means for deleting or inactivating the above gene is, for example, introduction of a mutation for one or more bases on the base sequence of the relevant gene, substitution of the relevant base sequence with another base sequence, or replacement of another base sequence. It may be inserted, or part or all of the base sequence of the relevant gene may be deleted.
- the means for inhibiting transcription of the gene is, for example, introduction of a mutation into the promoter region of the gene, inactivation of the promoter by substitution with another base sequence or insertion of another base sequence, or CRISPR interferometry (non-).
- Patent Document 16 Yao et al., ACS Synth. Biol., 2016, 5: 207-212) and the like may be used.
- the specific method for introducing the mutation or substituting or inserting the base sequence may be, for example, ultraviolet irradiation, site-specific mutation introduction, or homologous recombination method.
- the means for inhibiting the translation of the transcript of the above gene may be, for example, RNA (ribonucleic acid) interferometry or the like.
- the modified cyanobacteria 31 may be produced by suppressing or losing the function of the protein involved in the binding between the outer membrane 5 and the cell wall 4 in the cyanobacteria.
- the binding between the cell wall 4 and the outer membrane 5 that is, the binding amount and the binding force
- the outer membrane 5 is the cell wall. It becomes easy to partially separate from 4. Therefore, in the modified cyanobacteria 31, intracellular electrons or substances or molecules having electrons are likely to leak out of the cell, so that the extracellular electron transfer efficiency is improved.
- the organic channel protein 18 that improves the protein permeability of the outer membrane 5 is, for example, a chlorophyll-derived channel protein, and specifically, CppS consisting of the amino acid sequence shown in SEQ ID NO: 13. , CppF consisting of the amino acid sequence shown in SEQ ID NO: 14. Further, the organic channel protein 18 may be a protein having an amino acid sequence of 50% or more identical to any of these channel proteins.
- a gene encoding the organic channel protein 18 that improves the protein permeability of the outer membrane 5 is inserted into the chromosomal DNA or plasmid of cyanobacteria. Then, the organic channel protein 18 synthesized through transcription and translation of the gene is transported to the outer membrane 5 and expresses the channel function in the outer membrane 5 of cyanobacteria.
- the means for inserting and expressing the gene is not particularly limited as long as it is a commonly used means, and the base sequence of the promoter for transcriptional activation, the ribosome binding sequence for translation, and the outer membrane 5 are provided. It is not limited by the type of signal sequence for transport.
- the modified cyanobacteria 31 may be produced by expressing the organic channel protein 18 that improves the protein permeability of the outer membrane 5.
- the protein permeability of the outer membrane 5 is improved, so that the substance permeability of the outer membrane 5 is improved.
- the modified cyanobacteria 31 secretes intracellular electrons or substances or molecules having electrons to the outside of the cell, and takes up extracellular electrons or substances or molecules having electrons into the cells at least one of them. Therefore, the efficiency of extracellular electron transfer is improved.
- the modified cyanobacteria 31 produced by the above method secretes intracellular electrons or substances or molecules having electrons to the outside of the cell, and intracellular electrons or substances or molecules having electrons to the cells. Since at least one of the uptake into the cell can be performed, the efficiency of extracellular electron transfer is improved. Therefore, according to the method for manufacturing the electron carrier 30 according to the present embodiment, it is possible to provide the electron carrier 30 with improved electron transfer efficiency to the outside.
- the electron transfer method according to the present embodiment uses an electron carrier 30 containing any of the above-mentioned modified cyanobacteria 31.
- the electron transmitter 30 suppresses or loses the function of the protein involved in the binding between the outer membrane 5 and the cell wall 4 in the above-mentioned (i) cyanobacteria, and (ii) improves the protein permeability of the outer membrane 5.
- the modified cyanobacteria 31, which is at least one of the organic channel proteins 18 to be expressed, are contained, and the modified cyanobacteria 31 (I) supply electrons to the outside and (II) receive electrons from the outside. Do at least one of the uptakes.
- the electron is an electron or a substance or molecule having an electron.
- supplying electrons means not only supplying electrons but also supplying any substance or molecule having electrons.
- taking in an electron means not only taking in an electron but also taking in any substance or molecule having an electron.
- the electron carrier 30 may be a modified cyanobacteria 31 which is at least one of the above (i) and (ii), and as shown in FIG. 2, in addition to the modified cyanobacteria 31 It may contain at least one of an electron transfer substance 33, an electron mediator 35, and a conductive substance 37.
- the modified cyanobacteria 31 supplies electrons to the outside, for example, the modified cyanobacteria 31 receives light to generate an electron or a substance or molecule having an electron.
- the substance or molecule having electrons may be, for example, an electron transport chain 33.
- the modified cyanobacteria 31 may release the generated electrons or substances or molecules having electrons to the outside of the outer membrane.
- the modified cyanobacteria 31 may release a part of the electron transport chain 33 involved in the photosynthetic electron transport chain to the outside of the cell (that is, outside the outer membrane 5).
- the electron transport chain 33 released extracellularly may be incorporated into the cells of another modified cyanobacteria 31 and participate in the generation of a bioenergy source, for example, via a plurality of other modified cyanobacteria 31.
- the movement of electrons may occur.
- the released electron transfer material 33 may supply electrons to the electrode by a redox reaction with the outside (for example, an external electrode).
- the movement of electrons can be confirmed by measuring the current value.
- an electrode is placed in a cell suspension (so-called culture solution), and an electric potential is applied from the outside, electron transfer between the cell and the electrode occurs with high efficiency. An electric current is generated.
- the modified cyanobacteria 31 takes in electrons from the outside, for example, the modified cyanobacteria 31 takes in an electron existing outside the outer membrane or a substance or molecule having an electron inside the cell wall 4. Further, the modified cyanobacteria 31 may utilize an electron or a substance or molecule having an electron inside the cell wall 4 (in the cytoplasm). As described in the prior art, cyanobacteria generally have high photosynthetic ability and produce various organic substances in cells. Like cyanobacteria, modified cyanobacteria 31 produce various organic substances intracellularly (intracellular and thylakoids).
- the modified cyanobacteria 31 takes in an electron or a substance or molecule having an electron (which may be a part of the electron transport chain 33) existing outside the outer membrane 5 into the cell wall 4 and takes in the cell wall. Electrons may be received from the electron transport chain 33 inside the 4 (that is, in the cytoplasm) and used for substance production. Further, the modified cyanobacteria 31 may utilize the electrons taken in from the outside for respiration (catabolism of organic matter). As described above, since the modified cyanobacteria 31 can utilize electric energy instead of light energy, the intracellular reducing power is less likely to be insufficient even in an environment where sunlight irradiation is insufficient. Therefore, the modified cyanobacteria 31 can utilize both light energy and electric energy, so that the efficiency of substance production in the cell is improved.
- the modified cyanobacteria 31 contained in the electron carrier 30 uses electric energy instead of light energy to stably generate the energy and reducing power required in the cell, catabolize the substance, and the like. , Can produce substances.
- the electron carrier 30 since the electron carrier 30 according to the present embodiment contains the modified cyanobacteria 31 having improved extracellular electron transfer efficiency, it efficiently donates electrons to the outside (for example, an external electrode) and electrons from the outside. Can be efficiently received. Therefore, for example, when the electron carrier 30 is used, the modified cyanobacteria 31 contained in the electron carrier 30 releases the intracellular electron transfer substance to the outside of the cell, thereby efficiently supplying electrons to, for example, an external electrode. It can generate an electric current.
- modified cyanobacteria in which the outer membrane of the cyanobacteria is partially detached from the cell wall by (i) suppressing or abolishing the function of the protein related to the binding between the outer membrane and the cell wall in the cyanobacteria. It was prepared (Examples 1 and 2), and (ii) modified cyanobacteria having improved material permeability of the outer membrane were prepared by expressing a channel protein that improves protein permeability in the outer membrane (Example). 3). Whether or not these three types of modified cyanobacteria secrete the intracellular electron transmitter 33 to supply electrons to the outside and to take up electrons from the outside of the cell into the cell at least one of them.
- the evaluation was performed by quantifying and identifying the proteins secreted extracellularly by these modified cyanobacteria, and measuring the intracellular current value generated by photosynthesis.
- cyanobacteria Synechocystis sp. PCC 6803 (hereinafter, simply referred to as "cyanobacteria").
- Example 1 In Example 1, a modified cyanobacteria in which the expression of the slr1841 gene (SEQ ID NO: 7) encoding the SLH domain-retaining outer membrane protein was suppressed was produced.
- a cyanobacterial modified strain in which the expression of the slr1841 gene was suppressed As a gene suppression method, a CRISPR (Clustered Regularly Interspaced Short Palindromic Repeat) interferometry was used. In this method, the gene encoding the dCas9 protein (hereinafter referred to as the dCas9 gene) and the slr1841_sgRNA (single-guide Ribonucleic Acid) gene are introduced into the chromosomal DNA of cyanobacteria to introduce the slr1841 gene (SEQ ID NO: 7). Expression can be suppressed.
- CRISPR Clustered Regularly Interspaced Short Palindromic Repeat
- the mechanism of gene expression suppression by this method is as follows.
- a Cas9 protein lacking nuclease activity and an sgRNA (slr1841__sgRNA) that complementarily binds to the base sequence of the slr1841 gene (SEQ ID NO: 7) form a complex.
- this complex recognizes the slr1841 gene on the chromosomal DNA of cyanobacteria and specifically binds to the slr1841 gene.
- this binding becomes steric hindrance, transcription of the slr1841 gene is inhibited.
- the expression of the cyanobacterial slr1841 gene is suppressed.
- the above three genes are inserted into the psbA1 gene on the chromosomal DNA in a linked state, they can be amplified as one DNA fragment by the PCR method.
- the obtained DNA fragment is referred to as "psbA1 :: dCas9 cassette".
- the psbA1 :: dCas9 cassette was inserted into the pUC19 plasmid to obtain the pUC19-dCas9 plasmid.
- Transformed cells were selected by growing on BG-11 agar medium containing 20 ⁇ g / mL spectinomycin. In the selected cells, homologous recombination occurs between the psbA1 gene on the chromosomal DNA and the psbA1 upstream fragment region and the psbA1 downstream fragment region on the pUC19-dCas9 plasmid.
- a Synechocystis dCas9 strain in which a dCas9 cassette was inserted into the psbA1 gene region was obtained.
- the composition of the BG-11 medium used is as shown in Table 2.
- sgRNA specifically binds to the target gene by introducing a sequence of about 20 bases complementary to the target sequence into the region called protospacer on the sgRNA gene. To do.
- the protospacer sequences used in this example are shown in Table 3.
- the sgRNA gene (excluding the protospacer region) and the kanamycin resistance marker gene are linked and inserted into the slr2030-slr2031 gene on the chromosomal DNA. Therefore, by adding a protospacer sequence (SEQ ID NO: 33) complementary to the slr1841 gene (SEQ ID NO: 7) to the primer used when amplifying the sgRNA gene by the PCR method, an sgRNA that specifically recognizes slr1841 (SEQ ID NO: 33) slr1841_sgRNA) can be easily obtained.
- amplification by the PCR method using the primers slr2030-Fw (SEQ ID NO: 19) and slr2031-Rv (SEQ ID NO: 22) shown in Table 1 was carried out to obtain ((SEQ ID NO: 19).
- a DNA fragment (slr2030-2031 :: slr1841_sgRNA) was obtained in which i) slr2030 gene fragment, (ii) slr1841_sgRNA, (iii) kanamycin resistance marker gene, and (iv) slr2031 gene fragment were linked in this order.
- slr2030-2031 Using the In-Fusion PCR cloning method (registered trademark), slr2030-2031 :: slr1841_sgRNA was inserted into the pUC19 plasmid to obtain the pUC19-slr1841_sgRNA plasmid.
- the pUC19-slr1841_sgRNA plasmid was introduced into Synechocystis dCas9 strain by the same method as in (1-1) above, and transformed cells were selected on BG-11 agar medium containing 30 ⁇ g / mL kanamycin.
- a transformant Synechocystis dCas9 slr1841_sgRNA strain (hereinafter, also referred to as slr1841 inhibitory strain) in which slr1841_sgRNA was inserted into the slr2030-slr2031 gene on the chromosomal DNA was obtained.
- Example 2 a modified cyanobacteria in which the expression of the slr0688 gene encoding the cell wall-pyruvic acid modifying enzyme was suppressed was obtained by the following procedure.
- In-Fusion PCR was used and DNA fragments (slr2030-2031 :: slr0688_sgRNA) in which (i) slr2030 gene fragment, (ii) slr0688_sgRNA, (iii) canamycin resistance marker gene, and (iv) slr2031 gene fragment were linked in order.
- Comparative Example 1 a Synechocystis dCas9 strain was obtained by the same procedure as in (1-1) of Example 1.
- Example 1 Subsequently, the state of the cell surface was observed for each of the strains obtained in Example 1, Example 2, and Comparative Example 1. The details will be described below.
- Example 3 (3-1) Culturing of strains
- the cells were shake-cultured for 5 days under the conditions of s and 30 ° C.
- the slr0688-suppressed strain of Example 2 and the Control strain of Comparative Example 1 were also cultured under the same conditions as in Example 1.
- the cells in the resin were sliced to a thickness of 70 nm using an ultramicrotome (Ultracut) to prepare ultrathin sections.
- This ultrathin section was stained with 2% uranium acetate and 1% lead citrate solution to prepare a transmission electron microscope sample of the slr1841 inhibitor strain of Example 1.
- the same operation was performed for the slr0688-suppressed strain of Example 2 and the Control strain of Comparative Example 1, and samples for a transmission electron microscope were prepared.
- FIG. 3 is a TEM (Transmission Electron Microscope) image of the slr1841 inhibitory strain of Example 1.
- FIG. 4 is an enlarged image of the broken line region A of FIG.
- FIG. 4A is an enlarged TEM image of the broken line region A of FIG. 3
- FIG. 4B is a diagram depicting an enlarged TEM image of FIG. 4A.
- the adventitia was partially exfoliated from the cell wall (that is, the adventitia was partially exfoliated) and the adventitia was partially flexed. There was.
- the outer membrane was partially peeled off as shown in FIGS. 4 (a) and 4 (b).
- the parts one-dot dashed line areas a1 and a2 in the figure
- a portion where the outer membrane was greatly bent could be confirmed near the one-dot dashed line region a1.
- This part is a part where the bond between the outer membrane and the cell wall is weakened, and it is considered that the outer membrane is expanded outward and bent because the culture solution permeated into the periplasm from the outer membrane.
- FIG. 5 is a TEM image of the slr0688 inhibitory strain of Example 2.
- FIG. 6 is an enlarged image of the broken line region B of FIG.
- FIG. 6A is an enlarged TEM image of the broken line region B of FIG. 5
- FIG. 6B is a diagram depicting an enlarged TEM image of FIG. 6A.
- FIG. 7 is a TEM image of the Control strain of Comparative Example 1.
- FIG. 8 is an enlarged image of the broken line region C of FIG. 7.
- FIG. 8A is an enlarged TEM image of the broken line region C of FIG. 7, and
- FIG. 8B is a diagram depicting an enlarged TEM image of FIG. 8A.
- the cell surface layer of the Control strain of Comparative Example 1 was in order, and the inner membrane, the cell wall, the outer membrane, and the S layer were kept laminated in this order. That is, in the Control strain, the portion where the outer membrane was detached from the cell wall, the portion where the outer membrane was detached from the cell wall (that is, peeled off), and the portion where the outer membrane was bent as in Examples 1 and 2 were I could't see it.
- Example 3 the Synechocystis cppS tetR strain (hereinafter, cppS introduced) in which the chloroplast outer membrane channel protein CppS (SEQ ID NO: 13) retained by the gray alga Cyanophora paradoxa was introduced into the outer membrane of cyanobacteria by the following procedure. Also called a stock).
- slr2031-Rv SEQ ID NO: 22
- amplification was performed by PCR to obtain PL22 and KmR.
- these are inserted into the slr2030-slr2031 gene on the chromosome and are present. Therefore, when amplified by the PCR method using the above four primers, the slr2030 gene fragment is located on the 5'end side of PL22. Is amplified in a linked form, and the slr2031 gene fragment is amplified in a linked form on the 3'terminal side of KmR.
- amplification is performed by the PCR method using the four primers (SEQ ID NOs: 19, 22, 27, 28) shown in Table 1.
- a gene cassette (slr2030-2031 :: slr0042-KmR cassette) in which the slr2030 gene fragment, PL22, slr0042 gene, KmR, and slr2031 gene fragment were sequentially linked was obtained from the 5'terminal side.
- the slr2030-2031 :: slr0042-KmR cassette was inserted into the pUC19 plasmid to obtain the pUC19-slr0042 plasmid.
- total cDNA was prepared from the gray alga C. paradoxa NIES-547 using the SMART cDNA library synthesis kit (Clontech).
- amplification was performed by PCR using the primers cppS-Fw (SEQ ID NO: 29) and cppS-Rv (SEQ ID NO: 30) shown in Table 1 to obtain the cppS gene (SEQ ID NO: 13).
- the cppS gene SEQ ID NO: 13
- the cppS gene was inserted into the pUC19-slr0042 plasmid to obtain the pUC19-CppS plasmid.
- the cppS gene is inserted in a form linked to the 3'end side of the outer membrane transfer signal sequence of the slr0042 gene, and the region other than the outer membrane transfer signal sequence of the slr0042 gene is replaced with the coding region of cppS. Is removed with.
- amplification was performed by the PCR method to obtain a tetR gene and a spectinomycin resistance marker gene (SpcR) as a marker for gene transfer.
- SpcR spectinomycin resistance marker gene
- the upstream fragment of the psbA1 gene is linked to the 5'end side of the tetR gene. It is amplified in the form that the downstream fragment of the psbA1 gene is linked to the 3'terminal side of SpcR.
- amplification by PCR using the primers psbA1-Fw (SEQ ID NO: 17) and psbA1-Rv (SEQ ID NO: 18) shown in Table 1 is used to 5'.
- a gene cassette (psbA1 :: tetR cassette) in which the upstream fragment of the psbA1 gene, tetR, SpcR, and the downstream fragment of the psbA1 gene were linked in this order was obtained.
- the psbA1 :: tetR cassette cassette was inserted into the pUC19 plasmid to obtain the pUC19-tetR plasmid.
- the amount (hereinafter, also referred to as the amount of secreted protein) was measured.
- the secretory productivity of each of the above strains was evaluated based on the amount of protein in the culture medium.
- the protein secretory productivity refers to the ability to produce a protein by secreting the protein produced inside the cell extracellularly.
- Example 1 Culturing of strain The slr1841 inhibitory strain of Example 1 was cultured in the same manner as in (3-1) above. The culture was independently performed 3 times. The strains of Example 2, Example 3 and Comparative Example 1 were also cultured under the same conditions as those of Example 1.
- the slr1841 inhibitory strain of Example 1 As shown in FIG. 9, the slr1841 inhibitory strain of Example 1, the slr0688 inhibitory strain of Example 2, and the cppS-introduced strain of Example 3 were all culture supernatants as compared with the Control strain of Comparative Example 1.
- the amount of protein secreted inside (mg / L) was improved about 25 times.
- Example 1 when the absorbance (730 nm) of the culture solution was measured and the amount of secreted protein (mg protein / g cell dry weight) per 1 g of dry cell weight was calculated, the slr1841 inhibitory strain of Example 1 was calculated. In both the slr0688-suppressed strain of Example 2 and the cppS-introduced strain of Example 3, the amount of secreted protein (mg protein / g cell dry weight) per 1 g of dry cell weight was compared with that of the Control strain of Comparative Example 1. It was improved about 36 times.
- the amount of secreted protein (mg / L) of the slr0688 inhibitory strain of Example 2 and the gene (cppS) encoding the organic channel protein CppS were expressed on the outer membrane of cyanobacteria. Comparing with the amount of secreted protein (mg / L) of the introduced strain, the slr0688-suppressed strain of Example 2 was slightly higher (about 20 mg / L) than the cppS-introduced strain of Example 3.
- IAA iodoacetamide
- cysteine with a final concentration of 60 mM was added and allowed to stand at room temperature for 10 minutes.
- 400 ng of trypsin was added and allowed to stand overnight at 37 ° C. to fragment the protein into peptide fragments.
- TFA Trifluoroacetic Acid
- the sample was dried by a centrifugal evaporator. Then, 3% acetonitrile and 0.1% formic acid were added, and the sample was dissolved using a closed ultrasonic crusher. The peptide concentration was adjusted to 200 ng / ⁇ L.
- Table 4 shows 10 types of proteins in order from the one with the largest relative quantitative value among the identified proteins.
- the expression of the cppS gene improved the protein permeability of the outer membrane, and the protein in the periplasm permeated the channel protein CppS to the outside of the outer membrane (that is, outside the cells). It was confirmed that it was easily secreted. Therefore, it was shown that the modified strains of Examples 1 to 3 had improved substance permeability of the outer membrane.
- FIG. 10 is an exploded perspective view schematically showing an example of the configuration of the electrochemical measuring device 100.
- FIG. 11 is a schematic cross-sectional view taken along the XI-XI cross-sectional line of FIG.
- the electrochemical measuring device 100 includes a measuring unit 10 and a light irradiation unit 20.
- the measuring unit 10 has a reaction tank 12 having a storage unit 11 for accommodating the culture solution 40 of cyanobacteria, and a first electrode installed inside the reaction tank 12 so as to be in contact with the culture solution 40 in the storage unit 11. 13, a second electrode 14 installed inside the reaction vessel 12 so as to be in contact with the culture solution 40 in the accommodating portion 11, a potentiostat 15 for controlling the potential of the first electrode 13, and the inside of the accommodating portion 11.
- a reference electrode 16 is provided inside the reaction vessel 12 so as to be in contact with the culture solution 40 of the above.
- the reaction tank 12 has electrical insulation and does not permeate the culture solution 40. Further, the reaction tank 12 is a material that is not corroded or damaged by the culture solution 40, and may be made of, for example, plastic or ceramic.
- the measuring unit 10 applies, for example, a voltage between the first electrode 13 and the second electrode 14, or causes a current to flow, and measures the current or potential corresponding to the voltage or current.
- the measuring unit 10 applies a voltage between the first electrode 13 and the second electrode 14 to measure the current.
- the first electrode 13 is a so-called working electrode, and is an electrode that sensitively undergoes an electrochemical response to a trace amount of a substance in the culture solution 40 on the surface of the electrode.
- the second electrode 14 is a so-called counter electrode, and is an electrode for setting a potential difference with the working electrode (first electrode 13) or for passing an electric current.
- the first electrode 13 and the second electrode 14 are made of a conductive substance.
- the conductive substance may be, for example, a carbon material, a conductive polymer material, a semiconductor, a metal, or the like.
- the carbon material may be carbon nanotube, Ketjen black, glassy carbon, graphene, fullerene, carbon fiber, carbon fabric, carbon aerogel, or the like.
- the conductive polymer material polyaniline, polyacetylene, polypyrrole, poly (3,4-ethylenedioxythiophene), poly (p-phenylene vinylene), polythiophene, poly (p-phenylene sulfide) and the like can be used. There may be.
- the semiconductor may be silicone, germanium, indium tin oxide (ITO: Indium Tin Oxide), titanium oxide, copper oxide, silver oxide or the like.
- the metal may be gold, platinum, silver, titanium, aluminum, tungsten, copper, iron, palladium or the like.
- the first electrode 13 is an indium tin oxide (ITO) electrode
- the second electrode 14 is a platinum electrode.
- the conductive substance is not particularly limited as long as the conductive substance is not decomposed by its own oxidation reaction.
- the reference electrode 16 is an electrode that does not react with the substance in the culture solution 40 and maintains a constant potential, and is used by the potentiostat 15 to control the potential difference between the first electrode 13 and the reference electrode 16 to be constant. Will be done.
- the reference electrode 16 is a silver / silver chloride electrode.
- the potentiostat 15 applies a voltage between the first electrode 13 and the second electrode 14 to control the potential between the first electrode 13 and the reference electrode 16 to a predetermined value.
- the light irradiation unit 20 includes a light source 21 and a housing 22 that holds the light source 21.
- the light source 21 includes, for example, one or more light emitting bodies (for example, an LED (light emission diode)) and a reflecting surface surrounding the light emitting body.
- the light irradiation unit 20 may be arranged at a predetermined distance from the measurement unit 10 in the Z-axis plus direction.
- FIG. 11 illustrates a configuration in which the first electrode 13, the second electrode 14, and the reference electrode 16 each have a lead-out electrode below the reaction vessel 12 (that is, in the negative direction of the Z-axis). As long as it can be electrically connected to the stat 15, it may be in a form that does not pop out from the reaction tank 12. Further, the first electrode 13, the second electrode 14 and the reference electrode 16 may each be drawn out to the side surface of the reaction vessel 12.
- the configuration of the electrochemical measuring device 100 used is as follows.
- First electrode 13 Indium tin oxide electrode (surface area: 3.14 cm 2 )
- Second electrode 14 Platinum electrode Reference electrode 16: Silver / silver chloride electrode
- Light source 21 Light source that emits white light of about 120 ⁇ mol / m 2 / s
- FIG. 12 is a diagram showing the results of measuring the current flowing when the culture solution 40 of the Control strain of Comparative Example 1 was irradiated with light.
- the potentials of the first electrode 13 with respect to the reference electrode 16 are + 0.0V, + 0.1V, + 0.2V, + 0.25V, and +.
- the measured current values are about + 0.0nA, about + 0.0nA, about + 0.0nA, about + 0.0nA, and about + 1.0nA. Met.
- FIGS. 13 and 14 are diagram showing the results of measuring the current flowing when the culture solution 40 of the slr0688 inhibitory strain of Example 2 was irradiated with light.
- FIG. 14 is a diagram showing the results of measuring the current flowing when the culture solution 40 of the cppS-introduced strain of Example 3 was irradiated with light.
- the potentials of the first electrode 13 with respect to the reference electrode 16 are + 0.0V, + 0.1V, + 0.2V, + 0.25V, and In each case of + 0.3V, the measured current values (maximum after baseline correction) were about + 90nA, about + 250nA, about + 560nA, about + 750nA, and about + 1100nA.
- the potential of the first electrode 13 with respect to the reference electrode 16 was + 0.0V, + 0.1V, + 0.2V, + 0.25V. And, in each case of + 0.3V, the measured current value (maximum value after baseline correction) is about + 10nA, about + 20nA, about + 70nA, about + 150nA, and about + 260nA. It was.
- the current flowing in the culture solution 40 (hereinafter referred to as photocurrent) when the culture solution 40 is irradiated with light increases the potential of the first electrode 13 with respect to the reference electrode 16 to + 0.3V.
- the culture solution 40 of the slr0688-suppressed strain of Example 2 was improved 1000 times as much as the culture solution 40 of the control strain of Comparative Example 1.
- the photocurrent when the potential of the first electrode 13 with respect to the reference electrode 16 was controlled to be + 0.3 V was the culture solution of the control strain of Comparative Example 1. It improved about 300 times that of 40.
- the modified cyanobacteria in the present embodiment are extracellular by partially detaching the outer membrane of the cyanobacteria from the cell wall or improving the substance permeability of the outer membrane of the cyanobacteria. It was shown that the electron transfer efficiency was improved about 300 to 1000 times.
- the modified cyanobacteria modified to facilitate the leakage of intracellular proteins by weakening the bond between the outer membrane of the cyanobacteria and the cell wall, as in Examples 1 and 2. It is considered that the outer membrane is torn and peeled off, which makes it easier for organic substances other than proteins (for example, quinone) to leak out of the cell. Therefore, it is considered that the photocurrent was about 5 times higher than that of the modified cyanobacteria in which the protein permeability of the outer membrane of the cyanobacteria was improved as in Example 3.
- the protein production efficiency of the modified cyanobacteria is also improved, and the modified cyanobacteria can be used repeatedly even after the protein is recovered, so that the outer membrane can be appropriately detached and peeled off. It is very beneficial in that it is.
- the useful substance can be produced stably and efficiently by giving electric energy other than sunlight.
- the electron carrier, the method for manufacturing the electron carrier, and the electron transfer method according to the present disclosure have been described above based on the embodiments, but the present disclosure is not limited to these embodiments. .. As long as the gist of the present disclosure is not deviated, various modifications that can be conceived by those skilled in the art are applied to the embodiment, and other forms constructed by combining some components in the embodiment are also included in the scope of the present disclosure. included.
- an example in which the binding between the outer membrane and the cell membrane is weakened and the extracellular electron transfer efficiency is improved by suppressing or losing the function of the protein involved in the binding between the outer membrane and the cell wall in cyanobacteria is not limited to this.
- the bond between the outer membrane and the cell wall may be weakened, or the outer membrane may be weakened.
- the outer membrane may be weakened by adding the enzyme or the drug to the culture solution of cyanobacteria.
- the electron carrier the method for manufacturing the electron carrier, and the electron transfer method of the present disclosure
- electrons can be efficiently donated to the outside of the electron carrier, so that sludge or sewage can be treated and power generation can be performed. Can be done at the same time.
- the electron carrier since the electron carrier can efficiently transfer and receive electrons from the outside, the electron carrier can utilize electric energy when the amount of sunlight is insufficient. Therefore, according to the present disclosure, it is possible to efficiently produce useful substances in the fields of food, medicine, or chemistry, soil improvement, wastewater treatment, power generation, and the like.
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biochemistry (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Biotechnology (AREA)
- Molecular Biology (AREA)
- General Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Biophysics (AREA)
- Medicinal Chemistry (AREA)
- Gastroenterology & Hepatology (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Manufacturing & Machinery (AREA)
- Sustainable Development (AREA)
- Sustainable Energy (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Electrochemistry (AREA)
- General Chemical & Material Sciences (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description
微生物細胞が細胞外環境と電子伝達を行う現象を利用する技術開発が近年注目されている。例えば、グラム陰性細菌のShewanelle属又はGeobacter属の細菌は、有機物を細胞内で異化するときに生じる電子を、シトクロムなどの電子伝達機能を持つ生体分子を介して細胞外へ放出することが知られている(非特許文献1)。このように、微生物から放出された電子を外部電極で受容すれば、有機物を燃料とした微生物燃料電池として利用でき、例えば有機物を含む汚水処理過程に導入して発電装置として利用するなどの応用が期待されている(特許文献1)。
本開示の一態様の概要は、以下の通りである。
[1.定義]
本明細書において、塩基配列及びアミノ酸配列の同一性は、BLAST(Basic Local Alignment Search Tool)アルゴリズムによって計算される。具体的には、NCBI(National Center for Biotechnology Information)(https://blast.ncbi.nlm.nih.gov/Blast.cgi)のウェブサイトで利用できるBLASTプログラムにてペアワイズ解析を行うことにより算出される。シアノバクテリア及び植物の遺伝子、並びに、これらの遺伝子がコードするタンパク質に関する情報は、例えば上述のNCBIデータベース及びCyanobase(http://genome.microbedb.jp/cyanobase/)において公開されている。これらのデータベースから、目的のタンパク質のアミノ酸配列及びそれらのタンパク質をコードする遺伝子の塩基配列を取得することができる。
続いて、本実施の形態に係る電子伝達体について説明する。図2は、本実施の形態に係る電子伝達体30の一例を示す模式図である。
続いて、改変シアノバクテリア31について説明する。本実施の形態では、改変シアノバクテリア31は、電子伝達体30に含まれる。
続いて、本実施の形態に係る電子伝達体30の製造方法について説明する。電子伝達体30は、(i)シアノバクテリアにおいて外膜5と細胞壁4との結合に関与するタンパク質の機能が抑制又は喪失されている、及び、(ii)外膜5のタンパク質透過性を向上させる有機物チャネルタンパク質18が発現されている、の少なくとも1つである改変シアノバクテリア31を製造するステップ(以下、改変シアノバクテリア31の製造ステップという)を含む。
本実施の形態に係る電子伝達方法は、上記のいずれかの改変シアノバクテリア31を含む電子伝達体30を用いる。
実施例1では、SLHドメイン保持型外膜タンパク質をコードするslr1841遺伝子(配列番号7)の発現が抑制された改変シアノバクテリアを製造した。
遺伝子抑制法として、CRISPR(Clustered Regularly Interspaced Short Palindromic Repeat)干渉法を用いた。本方法では、dCas9タンパク質をコードする遺伝子(以下、dCas9遺伝子という)と、slr1841_sgRNA(single-guide Ribonucleic Acid)遺伝子とを、シアノバクテリアの染色体DNAに導入することにより、slr1841遺伝子(配列番号7)の発現を抑制することができる。
Synechocystis LY07株(以下、LY07株ともいう)(非特許文献16参照)の染色体DNAを鋳型として、dCas9遺伝子及びdCas9遺伝子の発現制御のためのオペレーター遺伝子、並びに、遺伝子導入の目印となるスペクチノマイシン耐性マーカー遺伝子を、表1に記載のプライマーpsbA1-Fw(配列番号17)及びpsbA1-Rv(配列番号18)を用いてPCR(Polymerase chain reaction)法により増幅した。なお、LY07株では、上記の3つの遺伝子が連結した状態で染色体DNA上のpsbA1遺伝子に挿入されているため、1つのDNA断片としてPCR法により増幅することができる。ここでは、得られたDNA断片を「psbA1::dCas9カセット」と表記する。In-Fusion PCRクローニング法(登録商標)を用いて、psbA1::dCas9カセットをpUC19プラスミドに挿入し、pUC19-dCas9プラスミドを得た。
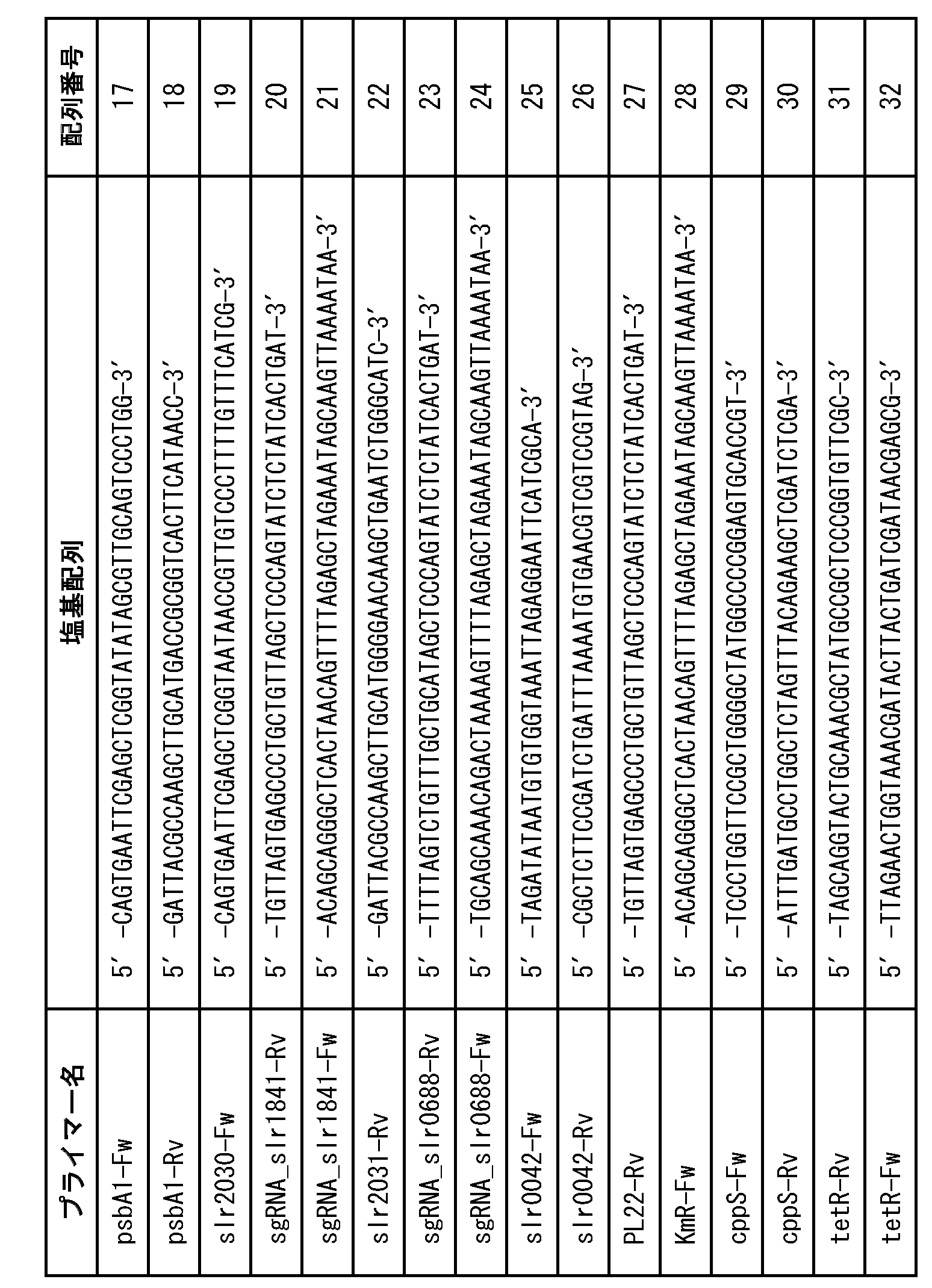
CRISPR干渉法では、sgRNA遺伝子上のprotospacerと呼ばれる領域に、標的配列と相補的な約20塩基の配列を導入することにより、sgRNAが標的遺伝子に特異的に結合する。本実施例で用いたprotospacer配列は表3に示される。

上記dCas9遺伝子及びslr1841_sgRNA遺伝子は、アンヒドロテトラサイクリン(aTc)の存在下で発現誘導されるようにプロモーター配列が設計されている。本実施例では、培地中に終濃度1 μg/mL aTc を添加することによりslr1841遺伝子の発現を抑制した。
実施例2では、下記の手順により、細胞壁-ピルビン酸修飾酵素をコードするslr0688遺伝子の発現が抑制された改変シアノバクテリアを得た。
上記(1-2)と同様の手順により、slr0688遺伝子(配列番号10)と相補的なprotospacer配列(配列番号34)を含むsgRNA遺伝子をSynechocystis dCas9株に導入し、Synechocystis dCas9 slr0688_sgRNA株を得た。なお、表1に記載のプライマーslr2030-Fw(配列番号19)及びsgRNA_slr0688-Rv(配列番号23)のセット、並びに、sgRNA_slr0688-Fw(配列番号24)及びslr2031-Rv(配列番号22)のセットを用いたことと、(i)slr2030遺伝子断片、(ii)slr0688_sgRNA、(iii)カナマイシン耐性マーカー遺伝子、(iv)slr2031遺伝子断片が順に連結したDNA断片(slr2030-2031::slr0688_sgRNA)をIn-Fusion PCRクローニング法(登録商標)を用いて、pUC19プラスミドに挿入し、pUC19-slr0688_sgRNAプラスミドを得たこと以外は、上記(1-2)と同様の条件で行った。
比較例1では、実施例1の(1-1)と同様の手順により、Synechocystis dCas9株を得た。
実施例1で得られた改変シアノバクテリアSynechocystis dCas9 slr1841_sgRNA株(いわゆる、slr1841抑制株)、実施例2で得られた改変シアノバクテリアSynechocystis dCas9 slr0688_sgRNA株(以下、slr0688抑制株ともいう)、及び、比較例1で得られた改変シアノバクテリアSynechocystis dCas9株(以下、Control株という)のそれぞれの超薄切片を作製し、電子顕微鏡を用いて細胞表層の状態(言い換えると、外膜構造)を観察した。
初発菌体濃度OD730 = 0.05となるように、実施例1のslr1841抑制株を、1 μg/mL aTcを含むBG-11培地に接種し、光量100 μmol/m2/s、30°Cの条件下で5日間振盪培養した。なお、実施例2のslr0688抑制株及び比較例1のControl株も実施例1と同様の条件で培養した。
上記(3-1)で得られた培養液を、室温にて2,500 g で10分間遠心分離し、実施例1のslr1841抑制株の細胞を回収した。次いで、細胞を-175℃の液体プロパンで急速凍結した後、2%グルタルアルデヒド及び1%タンニン酸を含むエタノール溶液を用いて-80℃で2日間固定した。固定後の細胞をエタノールにより脱水処理し、脱水した細胞を酸化プロピレンに浸透させたあと、樹脂(Quetol-651)溶液中に沈めた。その後60℃で48時間静置し、樹脂を硬化させて、細胞を樹脂で包埋した。樹脂中の細胞を、ウルトラミクロトーム(Ultracut)を用いて70 nmの厚さに薄切し、超薄切片を作成した。この超薄切片を、2%酢酸ウラン及び1%クエン酸鉛溶液を用いて染色して、実施例1のslr1841抑制株の透過型電子顕微鏡の試料を準備した。なお、実施例2のslr0688抑制株及び比較例1のControl株についてもそれぞれ同様の操作を行い、透過型電子顕微鏡の試料を準備した。
透過型電子顕微鏡(JEOL JEM-1400Plus)を用いて、加速電圧100 kV下で、上記(3-2)で得られた超薄切片の観察を行った。観察結果を図3~図8に示す。
実施例3では、下記の手順により、灰色藻Cyanophora paradoxaが保持する葉緑体の外膜チャネルタンパク質CppS(配列番号13)を、シアノバクテリアの外膜に導入したSynechocystis cppS tetR株(以下、cppS導入株ともいう)を得た。
(4-1)cppS遺伝子発現カセットの構築
cppS遺伝子と、cppS遺伝子の発現制御のためのプロモーター領域(PL22)と、シアノバクテリアにおける外膜移行シグナル配列(slr0042-signal)と、遺伝子導入の目印となるカナマイシン耐性マーカー遺伝子(KmR)と、が連結した遺伝子カセットを、以下の手順で作製した。
上記PL22のプロモーター活性は、TetRリプレッサーを介した制御により、アンヒドロテトラサイクリン(aTc)の存在下でのみ誘導される。従ってPL22の活性制御のためのtetR遺伝子を改変シアノバクテリアに導入する必要がある。
上記手順により得られたpUC19-cppSプラスミド1μgと、シアノバクテリア培養液(菌体濃度OD730 = 0.5程度)とを混合し、自然形質転換によりプラスミドを細胞内に導入した。形質転換された細胞を30 μg/mLのカナマイシンを含むBG-11寒天培地上で生育させることにより、選抜した。選抜された細胞では、染色体上のslr2030-2031遺伝子と、pUC19-cppSプラスミド上のslr2030遺伝子断片領域およびslr2031遺伝子断片領域との間で相同組み換えが起こる。これにより、slr2030-2031遺伝子領域にcppS遺伝子発現カセットが挿入されたSynechocystis cppS株を得た。なお、用いたBG-11培地の組成は表2の通りである。
上記Synechocystis cppS tetR 株のcppS遺伝子は、アンヒドロテトラサイクリン(aTc)の存在下で発現誘導される。本実施例では、BG-11培地中に終濃度1 μg/mL aTc を添加して培養することによりcppS遺伝子を発現誘導した。
以下の方法により、ペリプラズム(外膜と内膜との間隙)に存在するタンパク質が細胞外に分泌しているか否かを確認した。
実施例1のslr1841抑制株を上記(3-1)と同様の方法で培養した。培養は、独立して3回行った。なお、実施例2、実施例3及び比較例1の菌株についても実施例1の菌株と同様の条件で培養した。
上記(5-1)で得られた培養液を、室温にて2,500 gで10分間遠心分離し、培養上清を得た。得られた培養上清を、ポアサイズ0.22 μmのメンブレンフィルターを用いてろ過し、実施例1のslr1841抑制株の細胞を完全に除去した。ろ過後の培養上清に含まれる総タンパク質量をBCA(Bicinchoninic acid)法により定量した。この一連の操作を、独立して培養した3つの培養液のそれぞれについて行い、実施例1のslr1841抑制株の細胞外に分泌されたタンパク質量の平均値及び標準偏差を求めた。なお、実施例2、実施例3及び比較例1の菌株についても、それぞれ、同様の条件で、上記(5-1)で得られた3つの培養液のタンパク質量の定量を行い、3つの培養液中のタンパク質量の平均値及び標準偏差を求めた。
続いて、上記(5-2)で得られた培養上清中に含まれるタンパク質を、LC-MS/MSにより同定した。方法を以下に説明する。
培養上清の液量に対して8倍量の冷アセトンを加え、20℃で2時間静置後、20,000 gで15分間遠心分離し、タンパク質の沈殿物を得た。この沈殿物に100 mM Tris pH 8.5、0.5%ドデカン酸ナトリウム(SDoD)を加え、密閉式超音波破砕機によってタンパク質を溶解した。タンパク質濃度1 μg/mLに調整後、終濃度10 mMのジチオスレイトール(DTT)を添加して 50℃で30分間静置した。続いて、終濃度30 mMのヨードアセトアミド(IAA)を添加し、室温(遮光)で30分間静置した。IAA の反応を止めるために、終濃度60 mMのシステインを添加して室温で 10 分間静置した。トリプシン400 ngを添加して37℃で一晩静置し、タンパク質をペプチド断片化した。5% TFA(Trifluoroacetic Acid)を加えた後、室温にて15,000 gで10分間遠心分離し、上清を得た。この作業によりSDoDが除去された。C18スピンカラムを用いて脱塩後、遠心エバポレーターにより試料を乾固した。その後、3%アセトニトリル、0.1% formic acidを加え、密閉式超音波破砕機を用いて試料を溶解した。ペプチド濃度 200 ng/μL になるように調製した。
上記(5-3-1)で得られた試料をLC-MS/MS装置(UltiMate 3000 RSLCnano LC System) を用いて以下の条件で解析を実施した。
カラム:CAPCELL CORE MP 75 μm × 250 mm
溶媒:A溶媒は0.1%ギ酸水溶液、B溶媒は0.1%ギ酸+80%アセトニトリル
グラジエントプログラム:試料注入4分後にB溶媒8%、27分後にB溶媒44%、28分後にB溶媒80%、34分後に測定終了
得られたデータは以下の条件で解析し、タンパク質及びペプチドの同定ならびに定量値の算出を行った。
データベース:UniProtKB/Swiss Prot database ( Synechocystis sp. PCC 6803)
Fragmentation:HCD
Precursor Tolerance:8 ppm
Fragment Tolerance:1 0 ppm
Data Acquisition Type:Overlapping DIA
Peptide Length:8-70
Peptide Charge:2-8
Max Missed Cleavages:1
Fixed Modification:Carbamidomethylation
Peptide FDR: 1%以下
実施例2のslr0688抑制株、実施例3のcppS導入株及び比較例1のControl株について、細胞外電子伝達効率の評価を行った。より具体的には、これらの改変株の培養液にそれぞれ光を照射し、光合成による水の分解により発生した細胞内電流を外部電極で検出することにより、細胞外電子伝達効率を評価した。使用した装置及び電流の測定手順を以下に説明する。
まず、電気化学測定装置100の構成について図面を参照しながら説明する。図10は、電気化学測定装置100の構成の一例を概略的に示す分解斜視図である。図11は、図10のXI-XI断面線における概略断面図である。
比較例1のControl株、実施例2のslr0688抑制株、及び、実施例3のcppS導入株について、以下の手順で、上記の3つの菌株それぞれの電流生成量を測定した。
第二電極14:白金電極
参照電極16:銀/塩化銀電極
光源21:約120 μmol/m2/sの白色光を発する光源
まず、比較例1のControl株を、初発菌体濃度OD730 = 0.1となるように1 μg/mL aTcを含むBG-11培地に接種し、光量100 μmol/m2/s、30°Cの条件下で3日間振盪培養した。なお、実施例2のslr0688抑制株及び実施例3のcppS導入株についても、それぞれ、同様の条件で培養した。
上記(6-2-1)で得られた比較例1のControl株の培養液40を、反応槽12に4mL注入した後、参照電極16に対する第一電極13の電位が+0.0Vとなるようにポテンショスタット15により制御した。そして、光照射部20から収容部11内の培養液40に約120 μmol/m2/sの白色光を一定時間照射し、第一電極13と第二電極14との間に流れる電流を測定した。続いて、参照電極16に対する第一電極13の電位を+0.1V、+0.2V、+0.25V、及び、+0.3Vの順に変えて、それぞれの電位に制御した場合において、同様に、第一電極13と第二電極14との間に流れる電流を測定した。結果を図12に示す。図12は、比較例1のControl株の培養液40に光を照射した際に流れる電流を測定した結果を示す図である。

以上の結果から、実施例1及び実施例2のように、シアノバクテリアの外膜と細胞壁との結合を弱めて細胞内のタンパク質が漏出しやすくなるように改変した改変シアノバクテリアでは、外膜が破れて剥がれ落ちることにより、タンパク質以外の有機物(例えば、キノンなど)も細胞外に漏出しやすくなったと考えられる。そのため、実施例3のように、シアノバクテリアの外膜のタンパク質透過性を向上した改変シアノバクテリアよりも光電流が約5倍高かったと考えられる。
2 ペプチドグリカン
3 糖鎖
4 細胞壁
5 外膜
6 SLHドメイン保持型外膜タンパク質
7 SLHドメイン
8、18 有機物チャネルタンパク質
9 細胞壁-ピルビン酸修飾酵素
10 測定部
11 収容部
12 反応槽
13 第一電極
14 第二電極
15 ポテンショスタット
16 参照電極
20 光照射部
21 光源
22 筐体
30 電子伝達体
31 改変シアノバクテリア
33 電子伝達物質
35 電子メディエータ
37 導電性物質
40 培養液
100 電気化学測定装置
Claims (16)
- (i)シアノバクテリアにおいて外膜と細胞壁との結合に関与するタンパク質の機能が抑制又は喪失されている、及び、(ii)前記外膜のタンパク質透過性を向上させるチャネルタンパク質が発現されている、の少なくとも1つである改変シアノバクテリアを含み、
前記改変シアノバクテリアは、外部に電子を供給すること、及び、外部から電子を取り込むことの少なくとも1つを行う、
電子伝達体。 - 前記改変シアノバクテリアは、
光を受けて電子を生成し、
生成した前記電子を前記外膜の外に放出する、
請求項1に記載の電子伝達体。 - 前記改変シアノバクテリアは、
前記外膜の外側に存在する電子を前記細胞壁の内側に取り込み、
前記細胞壁の内側で前記電子を利用する、
請求項1又は2に記載の電子伝達体。 - 前記(i)において、前記外膜と前記細胞壁との結合に関与するタンパク質は、SLH(Surface Layer Homology)ドメイン保持型外膜タンパク質、及び、細胞壁-ピルビン酸修飾酵素の少なくとも1つである、
請求項1に記載の電子伝達体。 - 前記SLHドメイン保持型外膜タンパク質は、
配列番号1で示されるアミノ酸配列からなるSlr1841、
配列番号2で示されるアミノ酸配列からなるNIES970_09470、
配列番号3で示されるアミノ酸配列からなるAnacy_3458、又は、
これらのいずれかのSLHドメイン保持型外膜タンパク質とアミノ酸配列が50%以上同一であるタンパク質である、
請求項4に記載の電子伝達体。 - 前記細胞壁-ピルビン酸修飾酵素は、
配列番号4で示されるアミノ酸配列からなるSlr0688、
配列番号5で示されるアミノ酸配列からなるSynpcc7942_1529、
配列番号6で示されるアミノ酸配列からなるAnacy_1623、又は、
これらのいずれかの細胞壁-ピルビン酸修飾酵素とアミノ酸配列が50%以上同一であるタンパク質である、
請求項4に記載の電子伝達体。 - 前記(i)において、前記外膜と前記細胞壁との結合に関与するタンパク質を発現させる遺伝子が欠失又は不活性化されている、
請求項1に記載の電子伝達体。 - 前記外膜と細胞壁との結合に関与するタンパク質を発現させる遺伝子は、SLHドメイン保持型外膜タンパク質をコードする遺伝子、及び、細胞壁-ピルビン酸修飾酵素をコードする遺伝子の少なくとも1つである、
請求項7に記載の電子伝達体。 - 前記SLHドメイン保持型外膜タンパク質をコードする遺伝子は、
配列番号7で示される塩基配列からなるslr1841、
配列番号8で示される塩基配列からなるnies970_09470、
配列番号9で示される塩基配列からなるanacy_3458、又は、
これらのいずれかの遺伝子と塩基配列が50%以上同一である遺伝子である、
請求項8に記載の電子伝達体。 - 前記細胞壁-ピルビン酸修飾酵素をコードする遺伝子は、
配列番号10で示される塩基配列からなるslr0688、
配列番号11で示される塩基配列からなるsynpcc7942_1529、
配列番号12で示される塩基配列からなるanacy_1623、又は、
これらのいずれかの遺伝子と塩基配列が50%以上同一である遺伝子である、
請求項8に記載の電子伝達体。 - 前記(ii)において、前記外膜のタンパク質透過性を向上させるチャネルタンパク質は、
配列番号13で示されるアミノ酸配列からなるCppS、
配列番号14で示されるアミノ酸配列からなるCppF、又は、
これらのいずれかのチャネルタンパク質とアミノ酸配列が50%以上同一であるタンパク質である、
請求項1に記載の電子伝達体。 - 前記(ii)において、前記外膜のタンパク質透過性を向上させるチャネルタンパク質をコードする遺伝子が導入されている、
請求項1に記載の電子伝達体。 - 前記外膜のタンパク質透過性を向上させるチャネルタンパク質をコードする遺伝子は、葉緑体由来の遺伝子である、
請求項12に記載の電子伝達体。 - 前記外膜のタンパク質透過性を向上させるチャネルタンパク質をコードする遺伝子は、
配列番号15で示される塩基配列からなるcppS、
配列番号16で示される塩基酸配列からなるcppF、又は、
これらのいずれかの遺伝子と塩基配列が50%以上同一である遺伝子である、
請求項12又は13に記載の電子伝達体。 - (i)シアノバクテリアにおいて外膜と細胞壁との結合に関与するタンパク質の機能が抑制又は喪失されている、及び、(ii)前記外膜のタンパク質透過性を向上させるチャネルタンパク質が発現されている、の少なくとも1つである改変シアノバクテリアを製造するステップを含む、
電子伝達体の製造方法。 - 請求項1から14のいずれか1項に記載の電子伝達体を用いる、
電子伝達方法。
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2021558359A JP7668465B2 (ja) | 2019-11-21 | 2020-11-16 | 改変シアノバクテリア |
| CN202080078392.7A CN114729303B (zh) | 2019-11-21 | 2020-11-16 | 电子传递体、电子传递体的制造方法以及电子传递方法 |
| EP20890054.8A EP4063485A4 (en) | 2019-11-21 | 2020-11-16 | ELECTRON CARRIER, METHOD OF MAKING AN ELECTRON CARRIER AND METHOD OF ELECTRON TRANSFER |
| US17/748,678 US20220293990A1 (en) | 2019-11-21 | 2022-05-19 | Electron carrier, electron carrier production method, and electron transfer method |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2019210255 | 2019-11-21 | ||
| JP2019-210255 | 2019-11-21 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| US17/748,678 Continuation US20220293990A1 (en) | 2019-11-21 | 2022-05-19 | Electron carrier, electron carrier production method, and electron transfer method |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2021100643A1 true WO2021100643A1 (ja) | 2021-05-27 |
Family
ID=75980742
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2020/042553 Ceased WO2021100643A1 (ja) | 2019-11-21 | 2020-11-16 | 電子伝達体、電子伝達体の製造方法、及び、電子伝達方法 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US20220293990A1 (ja) |
| EP (1) | EP4063485A4 (ja) |
| JP (1) | JP7668465B2 (ja) |
| CN (1) | CN114729303B (ja) |
| WO (1) | WO2021100643A1 (ja) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2022186218A1 (ja) * | 2021-03-04 | 2022-09-09 | パナソニックIpマネジメント株式会社 | 電子伝達体、電子伝達体の製造方法、及び、電子伝達方法 |
| EP4091453A4 (en) * | 2019-12-23 | 2023-09-20 | Panasonic Intellectual Property Management Co., Ltd. | METHOD FOR PRODUCING A PLANT GROWTH PROMOTER, PLANT GROWTH PROMOTER AND METHOD FOR PROMOTING PLANT GROWTH |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP7766262B2 (ja) * | 2019-11-21 | 2025-11-10 | パナソニックIpマネジメント株式会社 | 分泌されたタンパク質の製造方法 |
| EP4063482A4 (en) * | 2019-11-21 | 2022-12-21 | Panasonic Intellectual Property Management Co., Ltd. | MODIFIED CYANOBACTERIUM, MODIFIED CYANOBACTERIUM MANUFACTURING METHOD AND PROTEIN MANUFACTURING METHOD |
| EP4541190A4 (en) * | 2022-06-20 | 2025-10-15 | Panasonic Ip Man Co Ltd | AGENT FOR INDUCTING PLANT DISEASE RESISTANCE, METHOD FOR INDUCTING PLANT DISEASE RESISTANCE, AND METHOD FOR PRODUCING THE AGENT FOR INDUCTING PLANT DISEASE RESISTANCE |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS6341676B2 (ja) | 1983-02-03 | 1988-08-18 | Mitsubishi Heavy Ind Ltd | |
| JP2018142408A (ja) | 2017-02-27 | 2018-09-13 | パナソニック株式会社 | 微生物燃料電池 |
Family Cites Families (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP2473597A4 (en) * | 2009-09-04 | 2013-05-15 | Harvard College | PREPARATION OF SECRETED BIOPRODUCTS FROM PHOTOSYNTHESIS MICROBES |
| JP2016182063A (ja) * | 2015-03-26 | 2016-10-20 | 国立大学法人 東京大学 | 培地及び強光阻害抑制方法 |
| EP4063482A4 (en) * | 2019-11-21 | 2022-12-21 | Panasonic Intellectual Property Management Co., Ltd. | MODIFIED CYANOBACTERIUM, MODIFIED CYANOBACTERIUM MANUFACTURING METHOD AND PROTEIN MANUFACTURING METHOD |
| CN116004690B (zh) * | 2022-09-09 | 2024-10-15 | 天津大学 | 一种工程改造希瓦氏菌囊泡提高胞外电子传递的方法 |
| CN115725490B (zh) * | 2022-10-19 | 2024-07-12 | 天津大学 | 一种合成与分泌高效电子传递载体吩嗪-1-羧酸的重组希瓦氏菌株构建方法及用途 |
-
2020
- 2020-11-16 CN CN202080078392.7A patent/CN114729303B/zh active Active
- 2020-11-16 JP JP2021558359A patent/JP7668465B2/ja active Active
- 2020-11-16 WO PCT/JP2020/042553 patent/WO2021100643A1/ja not_active Ceased
- 2020-11-16 EP EP20890054.8A patent/EP4063485A4/en active Pending
-
2022
- 2022-05-19 US US17/748,678 patent/US20220293990A1/en active Pending
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS6341676B2 (ja) | 1983-02-03 | 1988-08-18 | Mitsubishi Heavy Ind Ltd | |
| JP2018142408A (ja) | 2017-02-27 | 2018-09-13 | パナソニック株式会社 | 微生物燃料電池 |
Non-Patent Citations (24)
| Title |
|---|
| ALISTAIR J. MCCORMICK ET AL.: "Energy & Environmental Science", vol. 4, 2011, ROYAL SOCIETY OF CHEMISTRY, article "Photosynthetic biofilms in pure culture harness solar energy in a mediatorless bio-photovoltaic cell (BPV) system", pages: 4699 - 4709 |
| DUCAT DC ET AL.: "Trends in Biotechnology", vol. 29, 2011, ELSEVIER BV, article "Engineering cyanobacteria to generate i high-value products", pages: 95 - 103 |
| HASAN K ET AL.: "Physical Chemistry Chemical Physics", vol. 16, 2014, ROYAL SOCIETY OF CHEMISTRY, article "Photo-electrochemical communication between cyanobacteria (Leptolyngbia sp.) and osmium redox polymer modified electrodes", pages: 24676 - 24680 |
| J. A. GRALNICKD. K. NEWMAN: "Molecular Microbiology", vol. 65, 2007, WILEY-BLACKWELL, article "Extracellular respiration", pages: 1 - 11 |
| JENNY Z. ZHANG ET AL.: "Journal of American Chemical Society", vol. 140, 2018, AMERICAN CHEMICAL SOCIETY, article "Photoelectrochemistry of Photosystem II in Vitro vs in Vivo", pages: 6 - 9 |
| JUNLING GUO ET AL.: "Science", vol. 362, 2018, AMERICAN ASSOCIATION FOR THE ADVANCEMENT OF SCIENCE, article "Light-driven fine chemical production in yeast biohybrids", pages: 813 - 816 |
| JURGENSWECKESSER, J. BACTERIOL., vol. 168, 1986, pages 568 - 573 |
| KOJIMA ET AL., BIOSCI. BIOTECH. BIOCHEM., vol. 10, 2016, pages 1954 - 1959 |
| KOJIMA ET AL., J. BIOL. CHEM., vol. 291, 2016, pages 20198 - 20209 |
| KOJIMA SEIJI: "Elucidation and application of bacterial-derived membrane stabilization mechanism and substance permeation mechanism that function in the chloroplast surface membrane", 23 April 2018 (2018-04-23), pages 1 - 2, XP055921776, Retrieved from the Internet <URL:https://kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-18H02117> [retrieved on 20201224] * |
| KOJIMA, S. ET AL.: "Outer membrane proteins derived from non-cyanobacterial lineage cover the peptidoglycan of Cyanophora paradoxa cyanelles and serve as a cyanelle diffusion channel", JOURNAL OF BIOLOGICAL CHEMISTRY, vol. 291, no. 38, 29 June 2010 (2010-06-29), pages 20198 - 20209, XP055825633, ISSN: 0021-9258 * |
| KOJIMA, S. ET AL.: "Outer membrane-deprived cyanobacteria liberate periplasmic and thylakoid luminal components that support the growth of heterotrophs", BIORXIV, 25 March 2020 (2020-03-25), pages 1 - 31, XP055825843, Retrieved from the Internet <URL:https://doi.org/10.1101/2020.03.24.006684> [retrieved on 20201224] * |
| KOWATA, HIKARU: "Studies on molecular basis of cyanobacterial outer membrane function and its evolutionary relationship with primitive chloroplasts", DOCTORAL THESIS, 27 March 2018 (2018-03-27), TOHOKU UNIVERSITY, pages 1 - 11, XP055825637, Retrieved from the Internet <URL:http://hdl.handle.net/10097/00122689> * |
| KOWATA, J. BACTERIOL., vol. 199, 2017, pages e00371 - 17 |
| LEA-SMITH, D. J. ET AL.: "Photosynthetic, respiratory and extracellular electron transport pathways in cyanobacteria", BIOCHIMICA ET BIOPHYSICA ACTA, vol. 1857, 2016, pages 247 - 255, XP029411808, ISSN: 0005-2728, DOI: 10.1016/j.bbabio.2015.10.007 * |
| MESNAGE ET AL., EMBO J., vol. 19, 2000, pages 4473 - 4484 |
| NIKAIDO, MICROBIOL. MOL. BIOL. REV., vol. 67, no. 4, 2003, pages 593 - 656 |
| NISHIO K ET AL.: "Plant and Cell Physiology", vol. 56, 2015, JAPANESE SOCIETY OF PLANT PHYSIOLOGISTS, article "Electrochemical detection of circadian redox i rhythm in cyanobacterial cells via extracellular electron transfer", pages: 1053 - 1058 |
| PONCE-TOLEDO ET AL., CURR. BIOL., vol. 27, no. 3, 2017, pages 386 - 391 |
| See also references of EP4063485A4 |
| SEKAR, N. ET AL.: "Enhanced photo- bioelectrochemical energy conversion by genetically engineered cyanobacteria", BIOTECHNOLOGY AND BIOENGINEERING, vol. 113, no. 3, March 2016 (2016-03-01), pages 675 - 679, XP055825839, ISSN: 0006-3592 * |
| TSCHORTNER, J. ET AL.: "Biophotovoltaics: green power generation from sunlight and water", FRONTIERS IN MICROBIOLOGY, vol. 10, April 2019 (2019-04-01), pages 866, XP055825841, ISSN: 1664-302X * |
| VAN DE MEENE ET AL., ARCH. MICROBIOL., vol. 184, no. 5, 2006, pages 259 - 270 |
| YAO ET AL., ACS SYNTH. BIOL., vol. 5, 2016, pages 207 - 212 |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP4091453A4 (en) * | 2019-12-23 | 2023-09-20 | Panasonic Intellectual Property Management Co., Ltd. | METHOD FOR PRODUCING A PLANT GROWTH PROMOTER, PLANT GROWTH PROMOTER AND METHOD FOR PROMOTING PLANT GROWTH |
| JP2024019520A (ja) * | 2019-12-23 | 2024-02-09 | パナソニックIpマネジメント株式会社 | 植物成長促進剤の製造方法、植物成長促進剤、及び、植物成長促進方法 |
| JP7801672B2 (ja) | 2019-12-23 | 2026-01-19 | パナソニックIpマネジメント株式会社 | 植物成長促進剤の製造方法、植物成長促進剤、及び、植物成長促進方法 |
| WO2022186218A1 (ja) * | 2021-03-04 | 2022-09-09 | パナソニックIpマネジメント株式会社 | 電子伝達体、電子伝達体の製造方法、及び、電子伝達方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| EP4063485A1 (en) | 2022-09-28 |
| CN114729303B (zh) | 2025-04-11 |
| JPWO2021100643A1 (ja) | 2021-05-27 |
| JP7668465B2 (ja) | 2025-04-25 |
| EP4063485A4 (en) | 2023-07-19 |
| CN114729303A (zh) | 2022-07-08 |
| US20220293990A1 (en) | 2022-09-15 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7668465B2 (ja) | 改変シアノバクテリア | |
| Bird et al. | Engineering wired life: synthetic biology for electroactive bacteria | |
| Dong et al. | Engineering cyanobacterium with transmembrane electron transfer ability for bioelectrochemical nitrogen fixation | |
| Tefft et al. | Reversing an extracellular electron transfer pathway for electrode-driven acetoin reduction | |
| Deng et al. | Multi-heme cytochromes provide a pathway for survival in energy-limited environments | |
| Luo et al. | A new electrochemically active bacterium phylogenetically related to Tolumonas osonensis and power performance in MFCs | |
| Kumar et al. | Exoelectrogens in microbial fuel cells toward bioelectricity generation: a review | |
| Liu et al. | Magnetite compensates for the lack of a pilin‐associated c‐type cytochrome in extracellular electron exchange | |
| Lin et al. | Synthetic Saccharomyces cerevisiae‐Shewanella oneidensis consortium enables glucose‐fed high‐performance microbial fuel cell | |
| Commault et al. | Maintenance of Geobacter-dominated biofilms in microbial fuel cells treating synthetic wastewater | |
| TerAvest et al. | Oxidizing electrode potentials decrease current production and coulombic efficiency through cytochrome c inactivation in Shewanella oneidensis MR‐1 | |
| Xu et al. | Enhanced performance and mechanism study of microbial electrolysis cells using Fe nanoparticle-decorated anodes | |
| Suo et al. | Synthetic curli enables efficient microbial electrocatalysis with stainless‐steel electrode | |
| Verma et al. | Moving towards the enhancement of extracellular electron transfer in electrogens | |
| Yu et al. | Engineering outer membrane vesicles to increase extracellular electron transfer of Shewanella oneidensis | |
| Klein et al. | Microbe–Anode Interactions: Comparing the impact of genetic and material engineering approaches to improve the performance of microbial electrochemical systems (MES) | |
| Ding et al. | Modular engineering strategy to redirect electron flux into the electron-transfer chain for enhancing extracellular electron transfer in Shewanella oneidensis | |
| Badalamenti et al. | Coupling dark metabolism to electricity generation using photosynthetic cocultures | |
| Zhang et al. | Construction of an acetate metabolic pathway to enhance electron generation of engineered Shewanella oneidensis | |
| WO2021100640A1 (ja) | 改変シアノバクテリア、改変シアノバクテリアの製造方法、及び、タンパク質の製造方法 | |
| JPWO2021100643A5 (ja) | ||
| Hu et al. | All roads lead to Rome: Cyclic di-GMP differentially regulates extracellular electron transfer in Geobacter biofilms | |
| Gu et al. | Rewiring the respiratory pathway of Lactococcus lactis to enhance extracellular electron transfer | |
| US20100041121A1 (en) | Metabolically engineered organisms for the production of hydrogen and hydrogenase | |
| Samrot et al. | RETRACTED: Electricity generation by Enterobacter cloacae SU-1 in mediator less microbial fuel cell |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 20890054 Country of ref document: EP Kind code of ref document: A1 |
|
| DPE1 | Request for preliminary examination filed after expiration of 19th month from priority date (pct application filed from 20040101) | ||
| ENP | Entry into the national phase |
Ref document number: 2021558359 Country of ref document: JP Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2020890054 Country of ref document: EP Effective date: 20220621 |
|
| WWG | Wipo information: grant in national office |
Ref document number: 202080078392.7 Country of ref document: CN |