BRPI0307791B1 - Métodos para modificar característica de planta selecionada - Google Patents
Métodos para modificar característica de planta selecionada Download PDFInfo
- Publication number
- BRPI0307791B1 BRPI0307791B1 BRPI0307791-8A BRPI0307791A BRPI0307791B1 BR PI0307791 B1 BRPI0307791 B1 BR PI0307791B1 BR PI0307791 A BRPI0307791 A BR PI0307791A BR PI0307791 B1 BRPI0307791 B1 BR PI0307791B1
- Authority
- BR
- Brazil
- Prior art keywords
- plant
- gene
- dna
- sequence
- plants
- Prior art date
Links
- 238000000034 method Methods 0.000 title claims abstract description 145
- 241000196324 Embryophyta Species 0.000 claims abstract description 645
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 393
- 230000014509 gene expression Effects 0.000 claims abstract description 181
- 244000061456 Solanum tuberosum Species 0.000 claims abstract description 165
- 102000040430 polynucleotide Human genes 0.000 claims abstract description 112
- 108091033319 polynucleotide Proteins 0.000 claims abstract description 112
- 239000002157 polynucleotide Substances 0.000 claims abstract description 112
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 71
- 102000039446 nucleic acids Human genes 0.000 claims abstract description 63
- 108020004707 nucleic acids Proteins 0.000 claims abstract description 63
- 241000894007 species Species 0.000 claims abstract description 44
- 230000001976 improved effect Effects 0.000 claims abstract description 33
- 230000036541 health Effects 0.000 claims abstract description 15
- 238000003860 storage Methods 0.000 claims abstract description 14
- 235000002595 Solanum tuberosum Nutrition 0.000 claims description 148
- 239000003550 marker Substances 0.000 claims description 144
- 108020004414 DNA Proteins 0.000 claims description 107
- 241000589158 Agrobacterium Species 0.000 claims description 98
- 239000002773 nucleotide Substances 0.000 claims description 65
- 125000003729 nucleotide group Chemical group 0.000 claims description 65
- 230000001965 increasing effect Effects 0.000 claims description 61
- 108010031396 Catechol oxidase Proteins 0.000 claims description 59
- 101150054546 ppo gene Proteins 0.000 claims description 51
- 230000001105 regulatory effect Effects 0.000 claims description 36
- 108700026220 vif Genes Proteins 0.000 claims description 30
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 claims description 29
- 235000021307 Triticum Nutrition 0.000 claims description 26
- 108010073135 Phosphorylases Proteins 0.000 claims description 25
- 235000002637 Nicotiana tabacum Nutrition 0.000 claims description 20
- 101150106690 R1 gene Proteins 0.000 claims description 17
- 235000016709 nutrition Nutrition 0.000 claims description 16
- 229910019142 PO4 Inorganic materials 0.000 claims description 15
- 229920002472 Starch Polymers 0.000 claims description 15
- 235000019698 starch Nutrition 0.000 claims description 15
- 239000008107 starch Substances 0.000 claims description 15
- 240000004808 Saccharomyces cerevisiae Species 0.000 claims description 14
- 235000002017 Zea mays subsp mays Nutrition 0.000 claims description 14
- 240000008042 Zea mays Species 0.000 claims description 13
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 claims description 13
- 239000010452 phosphate Substances 0.000 claims description 13
- 108091032973 (ribonucleotides)n+m Proteins 0.000 claims description 12
- 240000003829 Sorghum propinquum Species 0.000 claims description 12
- 235000011684 Sorghum saccharatum Nutrition 0.000 claims description 12
- 235000005824 Zea mays ssp. parviglumis Nutrition 0.000 claims description 12
- 235000005822 corn Nutrition 0.000 claims description 12
- 230000001131 transforming effect Effects 0.000 claims description 11
- 208000034656 Contusions Diseases 0.000 claims description 10
- RYYVLZVUVIJVGH-UHFFFAOYSA-N caffeine Chemical compound CN1C(=O)N(C)C(=O)C2=C1N=CN2C RYYVLZVUVIJVGH-UHFFFAOYSA-N 0.000 claims description 10
- 239000000203 mixture Substances 0.000 claims description 10
- 235000002678 Ipomoea batatas Nutrition 0.000 claims description 9
- 244000017020 Ipomoea batatas Species 0.000 claims description 9
- 240000007594 Oryza sativa Species 0.000 claims description 9
- 235000007164 Oryza sativa Nutrition 0.000 claims description 9
- 241001233957 eudicotyledons Species 0.000 claims description 9
- 230000010474 transient expression Effects 0.000 claims description 9
- 108010021809 Alcohol dehydrogenase Proteins 0.000 claims description 8
- 108010025815 Kanamycin Kinase Proteins 0.000 claims description 8
- 235000003228 Lactuca sativa Nutrition 0.000 claims description 8
- 240000008415 Lactuca sativa Species 0.000 claims description 8
- 241000209510 Liliopsida Species 0.000 claims description 8
- 235000007688 Lycopersicon esculentum Nutrition 0.000 claims description 8
- 240000003183 Manihot esculenta Species 0.000 claims description 8
- 235000016735 Manihot esculenta subsp esculenta Nutrition 0.000 claims description 8
- 235000017587 Medicago sativa ssp. sativa Nutrition 0.000 claims description 8
- 240000003768 Solanum lycopersicum Species 0.000 claims description 8
- 108091036066 Three prime untranslated region Proteins 0.000 claims description 8
- 201000010099 disease Diseases 0.000 claims description 8
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 8
- 235000009566 rice Nutrition 0.000 claims description 8
- 238000011144 upstream manufacturing Methods 0.000 claims description 8
- 241001133760 Acoelorraphe Species 0.000 claims description 7
- 235000007319 Avena orientalis Nutrition 0.000 claims description 7
- 244000075850 Avena orientalis Species 0.000 claims description 7
- 244000000626 Daucus carota Species 0.000 claims description 7
- 235000002767 Daucus carota Nutrition 0.000 claims description 7
- 240000009088 Fragaria x ananassa Species 0.000 claims description 7
- 235000010469 Glycine max Nutrition 0.000 claims description 7
- 235000007340 Hordeum vulgare Nutrition 0.000 claims description 7
- 240000005979 Hordeum vulgare Species 0.000 claims description 7
- 244000305267 Quercus macrolepis Species 0.000 claims description 7
- 235000016976 Quercus macrolepis Nutrition 0.000 claims description 7
- 241000208140 Acer Species 0.000 claims description 6
- 235000002732 Allium cepa var. cepa Nutrition 0.000 claims description 6
- 241000219357 Cactaceae Species 0.000 claims description 6
- 244000260524 Chrysanthemum balsamita Species 0.000 claims description 6
- 235000005633 Chrysanthemum balsamita Nutrition 0.000 claims description 6
- 108091026898 Leader sequence (mRNA) Proteins 0.000 claims description 6
- 241000234435 Lilium Species 0.000 claims description 6
- 235000006679 Mentha X verticillata Nutrition 0.000 claims description 6
- 235000002899 Mentha suaveolens Nutrition 0.000 claims description 6
- 235000001636 Mentha x rotundifolia Nutrition 0.000 claims description 6
- 240000005561 Musa balbisiana Species 0.000 claims description 6
- 235000018290 Musa x paradisiaca Nutrition 0.000 claims description 6
- 241000233855 Orchidaceae Species 0.000 claims description 6
- 241000220317 Rosa Species 0.000 claims description 6
- 239000013566 allergen Substances 0.000 claims description 6
- 239000005516 coenzyme A Substances 0.000 claims description 6
- 229940093530 coenzyme a Drugs 0.000 claims description 6
- 150000003839 salts Chemical class 0.000 claims description 6
- 235000013311 vegetables Nutrition 0.000 claims description 6
- 101150096316 5 gene Proteins 0.000 claims description 5
- 235000011299 Brassica oleracea var botrytis Nutrition 0.000 claims description 5
- 235000017647 Brassica oleracea var italica Nutrition 0.000 claims description 5
- 240000003259 Brassica oleracea var. botrytis Species 0.000 claims description 5
- 235000002566 Capsicum Nutrition 0.000 claims description 5
- RGJOEKWQDUBAIZ-IBOSZNHHSA-N CoASH Chemical compound O[C@@H]1[C@H](OP(O)(O)=O)[C@@H](COP(O)(=O)OP(O)(=O)OCC(C)(C)[C@@H](O)C(=O)NCCC(=O)NCCS)O[C@H]1N1C2=NC=NC(N)=C2N=C1 RGJOEKWQDUBAIZ-IBOSZNHHSA-N 0.000 claims description 5
- 229920000742 Cotton Polymers 0.000 claims description 5
- 101710088194 Dehydrogenase Proteins 0.000 claims description 5
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 claims description 5
- 240000002395 Euphorbia pulcherrima Species 0.000 claims description 5
- 235000016623 Fragaria vesca Nutrition 0.000 claims description 5
- 235000011363 Fragaria x ananassa Nutrition 0.000 claims description 5
- 244000299507 Gossypium hirsutum Species 0.000 claims description 5
- 241000238631 Hexapoda Species 0.000 claims description 5
- 108090000604 Hydrolases Proteins 0.000 claims description 5
- LPHGQDQBBGAPDZ-UHFFFAOYSA-N Isocaffeine Natural products CN1C(=O)N(C)C(=O)C2=C1N(C)C=N2 LPHGQDQBBGAPDZ-UHFFFAOYSA-N 0.000 claims description 5
- 239000006002 Pepper Substances 0.000 claims description 5
- 235000016761 Piper aduncum Nutrition 0.000 claims description 5
- 240000003889 Piper guineense Species 0.000 claims description 5
- 235000017804 Piper guineense Nutrition 0.000 claims description 5
- 235000008184 Piper nigrum Nutrition 0.000 claims description 5
- 229960001948 caffeine Drugs 0.000 claims description 5
- VJEONQKOZGKCAK-UHFFFAOYSA-N caffeine Natural products CN1C(=O)N(C)C(=O)C2=C1C=CN2C VJEONQKOZGKCAK-UHFFFAOYSA-N 0.000 claims description 5
- 235000013339 cereals Nutrition 0.000 claims description 5
- WBYWAXJHAXSJNI-UHFFFAOYSA-N cinnamic acid Chemical compound OC(=O)C=CC1=CC=CC=C1 WBYWAXJHAXSJNI-UHFFFAOYSA-N 0.000 claims description 5
- RGJOEKWQDUBAIZ-UHFFFAOYSA-N coenzime A Natural products OC1C(OP(O)(O)=O)C(COP(O)(=O)OP(O)(=O)OCC(C)(C)C(O)C(=O)NCCC(=O)NCCS)OC1N1C2=NC=NC(N)=C2N=C1 RGJOEKWQDUBAIZ-UHFFFAOYSA-N 0.000 claims description 5
- 230000003247 decreasing effect Effects 0.000 claims description 5
- KDTSHFARGAKYJN-UHFFFAOYSA-N dephosphocoenzyme A Natural products OC1C(O)C(COP(O)(=O)OP(O)(=O)OCC(C)(C)C(O)C(=O)NCCC(=O)NCCS)OC1N1C2=NC=NC(N)=C2N=C1 KDTSHFARGAKYJN-UHFFFAOYSA-N 0.000 claims description 5
- 102000040650 (ribonucleotides)n+m Human genes 0.000 claims description 4
- 102000007698 Alcohol dehydrogenase Human genes 0.000 claims description 4
- 102000003668 Destrin Human genes 0.000 claims description 4
- 108090000082 Destrin Proteins 0.000 claims description 4
- 108090001064 Gelsolin Proteins 0.000 claims description 4
- 108700023224 Glucose-1-phosphate adenylyltransferases Proteins 0.000 claims description 4
- 102000004157 Hydrolases Human genes 0.000 claims description 4
- 240000007049 Juglans regia Species 0.000 claims description 4
- 235000009496 Juglans regia Nutrition 0.000 claims description 4
- 244000061176 Nicotiana tabacum Species 0.000 claims description 4
- 235000020234 walnut Nutrition 0.000 claims description 4
- 108020004705 Codon Proteins 0.000 claims description 3
- 208000005156 Dehydration Diseases 0.000 claims description 3
- 101150089323 HOS1 gene Proteins 0.000 claims description 3
- 229940114081 cinnamate Drugs 0.000 claims description 3
- 230000035784 germination Effects 0.000 claims description 3
- 229910001385 heavy metal Inorganic materials 0.000 claims description 3
- 239000011785 micronutrient Substances 0.000 claims description 3
- 235000013369 micronutrients Nutrition 0.000 claims description 3
- WBYWAXJHAXSJNI-VOTSOKGWSA-M trans-cinnamate Chemical compound [O-]C(=O)\C=C\C1=CC=CC=C1 WBYWAXJHAXSJNI-VOTSOKGWSA-M 0.000 claims description 3
- 101710130006 Beta-glucanase Proteins 0.000 claims description 2
- 108091081024 Start codon Proteins 0.000 claims description 2
- 244000098338 Triticum aestivum Species 0.000 claims 2
- 244000291564 Allium cepa Species 0.000 claims 1
- 241000219310 Beta vulgaris subsp. vulgaris Species 0.000 claims 1
- 101710116650 FAD-dependent monooxygenase Proteins 0.000 claims 1
- 240000004658 Medicago sativa Species 0.000 claims 1
- 101710128228 O-methyltransferase Proteins 0.000 claims 1
- 108090000854 Oxidoreductases Proteins 0.000 claims 1
- 102000004316 Oxidoreductases Human genes 0.000 claims 1
- 108090000612 Proline Oxidase Proteins 0.000 claims 1
- 240000000111 Saccharum officinarum Species 0.000 claims 1
- 235000007201 Saccharum officinarum Nutrition 0.000 claims 1
- 235000021536 Sugar beet Nutrition 0.000 claims 1
- 230000003828 downregulation Effects 0.000 claims 1
- 230000024346 drought recovery Effects 0.000 claims 1
- 235000003599 food sweetener Nutrition 0.000 claims 1
- 239000003765 sweetening agent Substances 0.000 claims 1
- 239000013598 vector Substances 0.000 abstract description 188
- 230000009466 transformation Effects 0.000 abstract description 92
- 230000009261 transgenic effect Effects 0.000 abstract description 34
- 102000004169 proteins and genes Human genes 0.000 abstract description 32
- 230000008569 process Effects 0.000 abstract description 21
- 238000009395 breeding Methods 0.000 abstract description 15
- 230000001488 breeding effect Effects 0.000 abstract description 12
- 238000003976 plant breeding Methods 0.000 abstract description 12
- 238000004519 manufacturing process Methods 0.000 abstract description 11
- 230000009418 agronomic effect Effects 0.000 abstract description 7
- 238000000338 in vitro Methods 0.000 abstract description 7
- 206010027146 Melanoderma Diseases 0.000 abstract description 4
- 244000038559 crop plants Species 0.000 abstract description 3
- 210000004027 cell Anatomy 0.000 description 181
- 230000010354 integration Effects 0.000 description 56
- 102000030523 Catechol oxidase Human genes 0.000 description 45
- 239000012634 fragment Substances 0.000 description 45
- 230000002068 genetic effect Effects 0.000 description 45
- 230000000692 anti-sense effect Effects 0.000 description 40
- 238000012546 transfer Methods 0.000 description 38
- 230000001404 mediated effect Effects 0.000 description 29
- 108010039811 Starch synthase Proteins 0.000 description 27
- 230000000694 effects Effects 0.000 description 27
- 235000018102 proteins Nutrition 0.000 description 25
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 24
- 241000209140 Triticum Species 0.000 description 24
- 239000008103 glucose Substances 0.000 description 24
- 239000013615 primer Substances 0.000 description 24
- 239000003112 inhibitor Substances 0.000 description 23
- 238000003752 polymerase chain reaction Methods 0.000 description 23
- 108091028043 Nucleic acid sequence Proteins 0.000 description 22
- 108010051210 beta-Fructofuranosidase Proteins 0.000 description 22
- 239000001573 invertase Substances 0.000 description 21
- 235000011073 invertase Nutrition 0.000 description 21
- 235000013305 food Nutrition 0.000 description 20
- 235000012015 potatoes Nutrition 0.000 description 20
- 239000000047 product Substances 0.000 description 19
- 101150079413 virD2 gene Proteins 0.000 description 19
- 241000208125 Nicotiana Species 0.000 description 18
- 101150093170 codA gene Proteins 0.000 description 18
- 102000053602 DNA Human genes 0.000 description 16
- 150000001413 amino acids Chemical class 0.000 description 15
- 238000006243 chemical reaction Methods 0.000 description 15
- 230000004927 fusion Effects 0.000 description 15
- 230000006798 recombination Effects 0.000 description 15
- 230000009467 reduction Effects 0.000 description 15
- 238000010353 genetic engineering Methods 0.000 description 14
- 238000005215 recombination Methods 0.000 description 14
- 125000006850 spacer group Chemical group 0.000 description 14
- 206010020649 Hyperkeratosis Diseases 0.000 description 13
- 229930006000 Sucrose Natural products 0.000 description 13
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 13
- 238000009825 accumulation Methods 0.000 description 13
- 238000013459 approach Methods 0.000 description 13
- 230000001580 bacterial effect Effects 0.000 description 13
- 230000015784 hyperosmotic salinity response Effects 0.000 description 13
- 239000002609 medium Substances 0.000 description 13
- 230000035772 mutation Effects 0.000 description 13
- 235000021317 phosphate Nutrition 0.000 description 13
- 229960004793 sucrose Drugs 0.000 description 13
- 238000011426 transformation method Methods 0.000 description 13
- 102000009097 Phosphorylases Human genes 0.000 description 12
- 238000002474 experimental method Methods 0.000 description 12
- 229930027917 kanamycin Natural products 0.000 description 12
- 229960000318 kanamycin Drugs 0.000 description 12
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 12
- 229930182823 kanamycin A Natural products 0.000 description 12
- 239000013612 plasmid Substances 0.000 description 12
- 210000001519 tissue Anatomy 0.000 description 12
- 230000001052 transient effect Effects 0.000 description 12
- 101150076274 upp gene Proteins 0.000 description 12
- 101100093450 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) ubi::crp-6 gene Proteins 0.000 description 11
- 108020004511 Recombinant DNA Proteins 0.000 description 11
- 230000003115 biocidal effect Effects 0.000 description 11
- 230000006870 function Effects 0.000 description 11
- 238000012545 processing Methods 0.000 description 11
- 241000589155 Agrobacterium tumefaciens Species 0.000 description 10
- 241000700605 Viruses Species 0.000 description 10
- 235000001014 amino acid Nutrition 0.000 description 10
- 229940024606 amino acid Drugs 0.000 description 10
- UQHKFADEQIVWID-UHFFFAOYSA-N cytokinin Natural products C1=NC=2C(NCC=C(CO)C)=NC=NC=2N1C1CC(O)C(CO)O1 UQHKFADEQIVWID-UHFFFAOYSA-N 0.000 description 10
- 239000004062 cytokinin Substances 0.000 description 10
- 239000004009 herbicide Substances 0.000 description 10
- 101150012864 ipt gene Proteins 0.000 description 10
- 230000008929 regeneration Effects 0.000 description 10
- 238000011069 regeneration method Methods 0.000 description 10
- 230000035897 transcription Effects 0.000 description 10
- 238000013518 transcription Methods 0.000 description 10
- RYGMFSIKBFXOCR-UHFFFAOYSA-N Copper Chemical compound [Cu] RYGMFSIKBFXOCR-UHFFFAOYSA-N 0.000 description 9
- 108010060309 Glucuronidase Proteins 0.000 description 9
- 102000053187 Glucuronidase Human genes 0.000 description 9
- 108700019146 Transgenes Proteins 0.000 description 9
- 229910052802 copper Inorganic materials 0.000 description 9
- 239000010949 copper Substances 0.000 description 9
- 230000030279 gene silencing Effects 0.000 description 9
- 238000003780 insertion Methods 0.000 description 9
- 230000037431 insertion Effects 0.000 description 9
- 239000005720 sucrose Substances 0.000 description 9
- 150000008163 sugars Chemical class 0.000 description 9
- 238000012360 testing method Methods 0.000 description 9
- JLIDBLDQVAYHNE-YKALOCIXSA-N (+)-Abscisic acid Chemical compound OC(=O)/C=C(/C)\C=C\[C@@]1(O)C(C)=CC(=O)CC1(C)C JLIDBLDQVAYHNE-YKALOCIXSA-N 0.000 description 8
- 108091034117 Oligonucleotide Proteins 0.000 description 8
- 108700026244 Open Reading Frames Proteins 0.000 description 8
- -1 atractilosides Chemical compound 0.000 description 8
- 101150054900 gus gene Proteins 0.000 description 8
- 230000002363 herbicidal effect Effects 0.000 description 8
- 230000001939 inductive effect Effects 0.000 description 8
- 210000004940 nucleus Anatomy 0.000 description 8
- 108090000765 processed proteins & peptides Proteins 0.000 description 8
- 230000005030 transcription termination Effects 0.000 description 8
- 244000025254 Cannabis sativa Species 0.000 description 7
- 241000219823 Medicago Species 0.000 description 7
- 108020005120 Plant DNA Proteins 0.000 description 7
- 238000012300 Sequence Analysis Methods 0.000 description 7
- 150000001875 compounds Chemical class 0.000 description 7
- 238000011161 development Methods 0.000 description 7
- 230000018109 developmental process Effects 0.000 description 7
- 230000012010 growth Effects 0.000 description 7
- 208000015181 infectious disease Diseases 0.000 description 7
- 108020004999 messenger RNA Proteins 0.000 description 7
- 235000000346 sugar Nutrition 0.000 description 7
- 229920001817 Agar Polymers 0.000 description 6
- 241000589156 Agrobacterium rhizogenes Species 0.000 description 6
- 108050004944 Alpha-glucan phosphorylases Proteins 0.000 description 6
- 108091026890 Coding region Proteins 0.000 description 6
- 108700007698 Genetic Terminator Regions Proteins 0.000 description 6
- 108090000848 Ubiquitin Proteins 0.000 description 6
- 239000008272 agar Substances 0.000 description 6
- 239000003242 anti bacterial agent Substances 0.000 description 6
- 230000015572 biosynthetic process Effects 0.000 description 6
- CWJSHJJYOPWUGX-UHFFFAOYSA-N chlorpropham Chemical compound CC(C)OC(=O)NC1=CC=CC(Cl)=C1 CWJSHJJYOPWUGX-UHFFFAOYSA-N 0.000 description 6
- 238000010367 cloning Methods 0.000 description 6
- 238000005520 cutting process Methods 0.000 description 6
- 235000012020 french fries Nutrition 0.000 description 6
- 230000006698 induction Effects 0.000 description 6
- 230000002018 overexpression Effects 0.000 description 6
- 244000052769 pathogen Species 0.000 description 6
- 102000004196 processed proteins & peptides Human genes 0.000 description 6
- 230000010076 replication Effects 0.000 description 6
- 239000002689 soil Substances 0.000 description 6
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 6
- 239000000126 substance Substances 0.000 description 6
- 108090000344 1,4-alpha-Glucan Branching Enzyme Proteins 0.000 description 5
- 102000003925 1,4-alpha-Glucan Branching Enzyme Human genes 0.000 description 5
- 241000234282 Allium Species 0.000 description 5
- 235000016068 Berberis vulgaris Nutrition 0.000 description 5
- 241000335053 Beta vulgaris Species 0.000 description 5
- 241000588724 Escherichia coli Species 0.000 description 5
- 229920000294 Resistant starch Polymers 0.000 description 5
- 230000003321 amplification Effects 0.000 description 5
- 230000008859 change Effects 0.000 description 5
- 101150036876 cre gene Proteins 0.000 description 5
- 238000005516 engineering process Methods 0.000 description 5
- 210000002615 epidermis Anatomy 0.000 description 5
- 239000007788 liquid Substances 0.000 description 5
- 239000000463 material Substances 0.000 description 5
- 230000000877 morphologic effect Effects 0.000 description 5
- 239000006870 ms-medium Substances 0.000 description 5
- 238000003199 nucleic acid amplification method Methods 0.000 description 5
- 235000021254 resistant starch Nutrition 0.000 description 5
- 229940027257 timentin Drugs 0.000 description 5
- 231100000331 toxic Toxicity 0.000 description 5
- 230000002588 toxic effect Effects 0.000 description 5
- 229920000856 Amylose Polymers 0.000 description 4
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 4
- 101100520142 Caenorhabditis elegans pin-2 gene Proteins 0.000 description 4
- 241000701489 Cauliflower mosaic virus Species 0.000 description 4
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 4
- 108091093105 Nuclear DNA Proteins 0.000 description 4
- 108010076504 Protein Sorting Signals Proteins 0.000 description 4
- 108010091086 Recombinases Proteins 0.000 description 4
- 102000018120 Recombinases Human genes 0.000 description 4
- 239000002253 acid Substances 0.000 description 4
- 238000004458 analytical method Methods 0.000 description 4
- 230000000433 anti-nutritional effect Effects 0.000 description 4
- 230000008901 benefit Effects 0.000 description 4
- YCIMNLLNPGFGHC-UHFFFAOYSA-N catechol Chemical compound OC1=CC=CC=C1O YCIMNLLNPGFGHC-UHFFFAOYSA-N 0.000 description 4
- 210000000349 chromosome Anatomy 0.000 description 4
- 230000009519 contusion Effects 0.000 description 4
- 230000007423 decrease Effects 0.000 description 4
- FCRACOPGPMPSHN-UHFFFAOYSA-N desoxyabscisic acid Natural products OC(=O)C=C(C)C=CC1C(C)=CC(=O)CC1(C)C FCRACOPGPMPSHN-UHFFFAOYSA-N 0.000 description 4
- IXORZMNAPKEEDV-UHFFFAOYSA-N gibberellic acid GA3 Natural products OC(=O)C1C2(C3)CC(=C)C3(O)CCC2C2(C=CC3O)C1C3(C)C(=O)O2 IXORZMNAPKEEDV-UHFFFAOYSA-N 0.000 description 4
- IXORZMNAPKEEDV-OBDJNFEBSA-N gibberellin A3 Chemical compound C([C@@]1(O)C(=C)C[C@@]2(C1)[C@H]1C(O)=O)C[C@H]2[C@]2(C=C[C@@H]3O)[C@H]1[C@]3(C)C(=O)O2 IXORZMNAPKEEDV-OBDJNFEBSA-N 0.000 description 4
- 230000006872 improvement Effects 0.000 description 4
- 101150111412 npt gene Proteins 0.000 description 4
- 230000008488 polyadenylation Effects 0.000 description 4
- 230000000644 propagated effect Effects 0.000 description 4
- 238000012216 screening Methods 0.000 description 4
- GEWDNTWNSAZUDX-WQMVXFAESA-N (-)-methyl jasmonate Chemical compound CC\C=C/C[C@@H]1[C@@H](CC(=O)OC)CCC1=O GEWDNTWNSAZUDX-WQMVXFAESA-N 0.000 description 3
- 108010030526 1-aminocyclopropanecarboxylate synthase Proteins 0.000 description 3
- 241000219194 Arabidopsis Species 0.000 description 3
- 241000894006 Bacteria Species 0.000 description 3
- 102100028637 CLOCK-interacting pacemaker Human genes 0.000 description 3
- 108010080611 Cytosine Deaminase Proteins 0.000 description 3
- GHASVSINZRGABV-UHFFFAOYSA-N Fluorouracil Chemical compound FC1=CNC(=O)NC1=O GHASVSINZRGABV-UHFFFAOYSA-N 0.000 description 3
- 241000233866 Fungi Species 0.000 description 3
- 239000005980 Gibberellic acid Substances 0.000 description 3
- 239000004471 Glycine Substances 0.000 description 3
- 241000282414 Homo sapiens Species 0.000 description 3
- 101000766839 Homo sapiens CLOCK-interacting pacemaker Proteins 0.000 description 3
- 241000218922 Magnoliophyta Species 0.000 description 3
- 108091023045 Untranslated Region Proteins 0.000 description 3
- 230000002159 abnormal effect Effects 0.000 description 3
- 230000009471 action Effects 0.000 description 3
- 238000007792 addition Methods 0.000 description 3
- 230000009286 beneficial effect Effects 0.000 description 3
- 238000004113 cell culture Methods 0.000 description 3
- 239000003795 chemical substances by application Substances 0.000 description 3
- 239000002299 complementary DNA Substances 0.000 description 3
- 238000012217 deletion Methods 0.000 description 3
- 230000037430 deletion Effects 0.000 description 3
- 238000010790 dilution Methods 0.000 description 3
- 239000012895 dilution Substances 0.000 description 3
- 238000004520 electroporation Methods 0.000 description 3
- 210000002257 embryonic structure Anatomy 0.000 description 3
- 230000002255 enzymatic effect Effects 0.000 description 3
- 229960002949 fluorouracil Drugs 0.000 description 3
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 3
- 125000000404 glutamine group Chemical group N[C@@H](CCC(N)=O)C(=O)* 0.000 description 3
- 238000010438 heat treatment Methods 0.000 description 3
- 238000001727 in vivo Methods 0.000 description 3
- 238000011534 incubation Methods 0.000 description 3
- GEWDNTWNSAZUDX-UHFFFAOYSA-N methyl 7-epi-jasmonate Natural products CCC=CCC1C(CC(=O)OC)CCC1=O GEWDNTWNSAZUDX-UHFFFAOYSA-N 0.000 description 3
- 230000003505 mutagenic effect Effects 0.000 description 3
- 108010058731 nopaline synthase Proteins 0.000 description 3
- 235000014571 nuts Nutrition 0.000 description 3
- 210000000056 organ Anatomy 0.000 description 3
- 230000035755 proliferation Effects 0.000 description 3
- 230000001568 sexual effect Effects 0.000 description 3
- 239000000243 solution Substances 0.000 description 3
- 229960005322 streptomycin Drugs 0.000 description 3
- 238000003786 synthesis reaction Methods 0.000 description 3
- 230000002123 temporal effect Effects 0.000 description 3
- 238000000844 transformation Methods 0.000 description 3
- 238000013519 translation Methods 0.000 description 3
- JXYWFNAQESKDNC-BTJKTKAUSA-N (z)-4-hydroxy-4-oxobut-2-enoate;2-[(4-methoxyphenyl)methyl-pyridin-2-ylamino]ethyl-dimethylazanium Chemical compound OC(=O)\C=C/C(O)=O.C1=CC(OC)=CC=C1CN(CCN(C)C)C1=CC=CC=N1 JXYWFNAQESKDNC-BTJKTKAUSA-N 0.000 description 2
- NOEGNKMFWQHSLB-UHFFFAOYSA-N 5-hydroxymethylfurfural Chemical compound OCC1=CC=C(C=O)O1 NOEGNKMFWQHSLB-UHFFFAOYSA-N 0.000 description 2
- 101710145411 Acid beta-fructofuranosidase Proteins 0.000 description 2
- 108010085238 Actins Proteins 0.000 description 2
- 239000004382 Amylase Substances 0.000 description 2
- 102000013142 Amylases Human genes 0.000 description 2
- 108010065511 Amylases Proteins 0.000 description 2
- 102000003669 Antiporters Human genes 0.000 description 2
- 108090000084 Antiporters Proteins 0.000 description 2
- 241000219195 Arabidopsis thaliana Species 0.000 description 2
- 235000017060 Arachis glabrata Nutrition 0.000 description 2
- 244000105624 Arachis hypogaea Species 0.000 description 2
- 235000010777 Arachis hypogaea Nutrition 0.000 description 2
- 235000018262 Arachis monticola Nutrition 0.000 description 2
- 108020000946 Bacterial DNA Proteins 0.000 description 2
- 108010059892 Cellulase Proteins 0.000 description 2
- 240000006670 Chlorogalum pomeridianum Species 0.000 description 2
- 108010074879 Cinnamoyl-CoA reductase Proteins 0.000 description 2
- 102000000311 Cytosine Deaminase Human genes 0.000 description 2
- 108010066133 D-octopine dehydrogenase Proteins 0.000 description 2
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 2
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 2
- 241000206602 Eukaryota Species 0.000 description 2
- 229930091371 Fructose Natural products 0.000 description 2
- 239000005715 Fructose Substances 0.000 description 2
- RFSUNEUAIZKAJO-ARQDHWQXSA-N Fructose Chemical compound OC[C@H]1O[C@](O)(CO)[C@@H](O)[C@@H]1O RFSUNEUAIZKAJO-ARQDHWQXSA-N 0.000 description 2
- 102000000340 Glucosyltransferases Human genes 0.000 description 2
- 108010055629 Glucosyltransferases Proteins 0.000 description 2
- IMQLKJBTEOYOSI-GPIVLXJGSA-N Inositol-hexakisphosphate Chemical class OP(O)(=O)O[C@H]1[C@H](OP(O)(O)=O)[C@@H](OP(O)(O)=O)[C@H](OP(O)(O)=O)[C@H](OP(O)(O)=O)[C@@H]1OP(O)(O)=O IMQLKJBTEOYOSI-GPIVLXJGSA-N 0.000 description 2
- 108091092195 Intron Proteins 0.000 description 2
- 101150062031 L gene Proteins 0.000 description 2
- 108090001090 Lectins Proteins 0.000 description 2
- 102000004856 Lectins Human genes 0.000 description 2
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 241001465754 Metazoa Species 0.000 description 2
- 101100369993 Mus musculus Tnfsf10 gene Proteins 0.000 description 2
- 238000010222 PCR analysis Methods 0.000 description 2
- 101710091688 Patatin Proteins 0.000 description 2
- 244000025272 Persea americana Species 0.000 description 2
- 235000008673 Persea americana Nutrition 0.000 description 2
- IAJOBQBIJHVGMQ-UHFFFAOYSA-N Phosphinothricin Natural products CP(O)(=O)CCC(N)C(O)=O IAJOBQBIJHVGMQ-UHFFFAOYSA-N 0.000 description 2
- 108700001094 Plant Genes Proteins 0.000 description 2
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- 102000044159 Ubiquitin Human genes 0.000 description 2
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical compound O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 2
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 2
- 229930013930 alkaloid Natural products 0.000 description 2
- 108090000637 alpha-Amylases Proteins 0.000 description 2
- 235000019418 amylase Nutrition 0.000 description 2
- 230000003466 anti-cipated effect Effects 0.000 description 2
- 229940088710 antibiotic agent Drugs 0.000 description 2
- 238000003556 assay Methods 0.000 description 2
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 2
- 230000033228 biological regulation Effects 0.000 description 2
- 208000034526 bruise Diseases 0.000 description 2
- 150000001720 carbohydrates Chemical class 0.000 description 2
- 235000014633 carbohydrates Nutrition 0.000 description 2
- 230000015556 catabolic process Effects 0.000 description 2
- 210000003855 cell nucleus Anatomy 0.000 description 2
- 230000002759 chromosomal effect Effects 0.000 description 2
- 239000012881 co-culture medium Substances 0.000 description 2
- 230000000295 complement effect Effects 0.000 description 2
- 230000000875 corresponding effect Effects 0.000 description 2
- 238000009402 cross-breeding Methods 0.000 description 2
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 2
- 235000018417 cysteine Nutrition 0.000 description 2
- 238000006731 degradation reaction Methods 0.000 description 2
- 235000019820 disodium diphosphate Nutrition 0.000 description 2
- 238000001035 drying Methods 0.000 description 2
- 238000002635 electroconvulsive therapy Methods 0.000 description 2
- 229940088598 enzyme Drugs 0.000 description 2
- 239000013604 expression vector Substances 0.000 description 2
- 238000012226 gene silencing method Methods 0.000 description 2
- BRZYSWJRSDMWLG-CAXSIQPQSA-N geneticin Chemical compound O1C[C@@](O)(C)[C@H](NC)[C@@H](O)[C@H]1O[C@@H]1[C@@H](O)[C@H](O[C@@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](C(C)O)O2)N)[C@@H](N)C[C@H]1N BRZYSWJRSDMWLG-CAXSIQPQSA-N 0.000 description 2
- IAJOBQBIJHVGMQ-BYPYZUCNSA-N glufosinate-P Chemical compound CP(O)(=O)CC[C@H](N)C(O)=O IAJOBQBIJHVGMQ-BYPYZUCNSA-N 0.000 description 2
- 230000036252 glycation Effects 0.000 description 2
- 229930008677 glyco alkaloid Natural products 0.000 description 2
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 2
- XDDAORKBJWWYJS-UHFFFAOYSA-N glyphosate Chemical compound OC(=O)CNCP(O)(O)=O XDDAORKBJWWYJS-UHFFFAOYSA-N 0.000 description 2
- 238000003306 harvesting Methods 0.000 description 2
- 125000000487 histidyl group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C([H])=N1 0.000 description 2
- 238000010348 incorporation Methods 0.000 description 2
- AWJUIBRHMBBTKR-UHFFFAOYSA-N isoquinoline Chemical compound C1=NC=CC2=CC=CC=C21 AWJUIBRHMBBTKR-UHFFFAOYSA-N 0.000 description 2
- 239000002523 lectin Substances 0.000 description 2
- 238000012423 maintenance Methods 0.000 description 2
- 230000013011 mating Effects 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- 239000003471 mutagenic agent Substances 0.000 description 2
- 229910052757 nitrogen Inorganic materials 0.000 description 2
- 238000012261 overproduction Methods 0.000 description 2
- 235000020232 peanut Nutrition 0.000 description 2
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 2
- 150000002989 phenols Chemical class 0.000 description 2
- 150000003013 phosphoric acid derivatives Chemical class 0.000 description 2
- 229910052698 phosphorus Inorganic materials 0.000 description 2
- 239000011574 phosphorus Substances 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 235000002949 phytic acid Nutrition 0.000 description 2
- 229920001184 polypeptide Polymers 0.000 description 2
- 230000008707 rearrangement Effects 0.000 description 2
- 230000001172 regenerating effect Effects 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 238000003757 reverse transcription PCR Methods 0.000 description 2
- 230000035939 shock Effects 0.000 description 2
- 210000003491 skin Anatomy 0.000 description 2
- 239000011734 sodium Substances 0.000 description 2
- 239000007787 solid Substances 0.000 description 2
- 235000021012 strawberries Nutrition 0.000 description 2
- 238000006467 substitution reaction Methods 0.000 description 2
- 230000005026 transcription initiation Effects 0.000 description 2
- 238000010361 transduction Methods 0.000 description 2
- 230000026683 transduction Effects 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 238000012250 transgenic expression Methods 0.000 description 2
- 239000002753 trypsin inhibitor Substances 0.000 description 2
- 210000003462 vein Anatomy 0.000 description 2
- 101150102050 virE2 gene Proteins 0.000 description 2
- NEEQFPMRODQIKX-UHFFFAOYSA-N (S)-Dencichine Natural products OC(=O)C(N)CNC(=O)C(O)=O NEEQFPMRODQIKX-UHFFFAOYSA-N 0.000 description 1
- 101150084750 1 gene Proteins 0.000 description 1
- PRPINYUDVPFIRX-UHFFFAOYSA-N 1-naphthaleneacetic acid Chemical compound C1=CC=C2C(CC(=O)O)=CC=CC2=C1 PRPINYUDVPFIRX-UHFFFAOYSA-N 0.000 description 1
- SNBCLPGEMZEWLU-QXFUBDJGSA-N 2-chloro-n-[[(2r,3s,5r)-3-hydroxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methyl]acetamide Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CNC(=O)CCl)[C@@H](O)C1 SNBCLPGEMZEWLU-QXFUBDJGSA-N 0.000 description 1
- SEYLPRWNVFCVRQ-UHFFFAOYSA-N 3,4-dihydroxybutan-2-one Chemical compound CC(=O)C(O)CO SEYLPRWNVFCVRQ-UHFFFAOYSA-N 0.000 description 1
- QVYAWBLDJPTXHS-UHFFFAOYSA-N 5-Hydroxymethyl-2-furfural Natural products OC1=CC=C(C=O)O1 QVYAWBLDJPTXHS-UHFFFAOYSA-N 0.000 description 1
- GOSWTRUMMSCNCW-HNNGNKQASA-N 9-ribosyl-trans-zeatin Chemical compound C1=NC=2C(NC\C=C(CO)/C)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O GOSWTRUMMSCNCW-HNNGNKQASA-N 0.000 description 1
- 108010000700 Acetolactate synthase Proteins 0.000 description 1
- 241000251468 Actinopterygii Species 0.000 description 1
- 102000007469 Actins Human genes 0.000 description 1
- 229920000945 Amylopectin Polymers 0.000 description 1
- 102000044503 Antimicrobial Peptides Human genes 0.000 description 1
- 108700042778 Antimicrobial Peptides Proteins 0.000 description 1
- 102000003823 Aromatic-L-amino-acid decarboxylases Human genes 0.000 description 1
- 108090000121 Aromatic-L-amino-acid decarboxylases Proteins 0.000 description 1
- 241000271566 Aves Species 0.000 description 1
- 108010077805 Bacterial Proteins Proteins 0.000 description 1
- LSNNMFCWUKXFEE-UHFFFAOYSA-M Bisulfite Chemical compound OS([O-])=O LSNNMFCWUKXFEE-UHFFFAOYSA-M 0.000 description 1
- 241000253373 Caldanaerobacter subterraneus subsp. tengcongensis Species 0.000 description 1
- 108010022172 Chitinases Proteins 0.000 description 1
- 102000012286 Chitinases Human genes 0.000 description 1
- 241000701248 Chlorella virus Species 0.000 description 1
- 108010077544 Chromatin Proteins 0.000 description 1
- 208000003322 Coinfection Diseases 0.000 description 1
- 206010009944 Colon cancer Diseases 0.000 description 1
- 208000001333 Colorectal Neoplasms Diseases 0.000 description 1
- 108020004635 Complementary DNA Proteins 0.000 description 1
- 241000218631 Coniferophyta Species 0.000 description 1
- 108091035707 Consensus sequence Proteins 0.000 description 1
- 108010051219 Cre recombinase Proteins 0.000 description 1
- GSXOAOHZAIYLCY-UHFFFAOYSA-N D-F6P Natural products OCC(=O)C(O)C(O)C(O)COP(O)(O)=O GSXOAOHZAIYLCY-UHFFFAOYSA-N 0.000 description 1
- IMXSCCDUAFEIOE-UHFFFAOYSA-N D-Octopin Natural products OC(=O)C(C)NC(C(O)=O)CCCN=C(N)N IMXSCCDUAFEIOE-UHFFFAOYSA-N 0.000 description 1
- LMKYZBGVKHTLTN-NKWVEPMBSA-N D-nopaline Chemical compound NC(=N)NCCC[C@@H](C(O)=O)N[C@@H](C(O)=O)CCC(O)=O LMKYZBGVKHTLTN-NKWVEPMBSA-N 0.000 description 1
- IMXSCCDUAFEIOE-RITPCOANSA-N D-octopine Chemical compound [O-]C(=O)[C@@H](C)[NH2+][C@H](C([O-])=O)CCCNC(N)=[NH2+] IMXSCCDUAFEIOE-RITPCOANSA-N 0.000 description 1
- YAHZABJORDUQGO-NQXXGFSBSA-N D-ribulose 1,5-bisphosphate Chemical compound OP(=O)(O)OC[C@@H](O)[C@@H](O)C(=O)COP(O)(O)=O YAHZABJORDUQGO-NQXXGFSBSA-N 0.000 description 1
- 239000003155 DNA primer Substances 0.000 description 1
- 101150092880 DREB1A gene Proteins 0.000 description 1
- 102100024746 Dihydrofolate reductase Human genes 0.000 description 1
- 208000035240 Disease Resistance Diseases 0.000 description 1
- 101100218970 Drosophila melanogaster borr gene Proteins 0.000 description 1
- 206010052806 Drug tolerance increased Diseases 0.000 description 1
- 241000620209 Escherichia coli DH5[alpha] Species 0.000 description 1
- 244000004281 Eucalyptus maculata Species 0.000 description 1
- 108091029865 Exogenous DNA Proteins 0.000 description 1
- 239000004606 Fillers/Extenders Substances 0.000 description 1
- 239000004366 Glucose oxidase Substances 0.000 description 1
- 108010015776 Glucose oxidase Proteins 0.000 description 1
- 241000428456 Gluma Species 0.000 description 1
- 244000068988 Glycine max Species 0.000 description 1
- 239000005562 Glyphosate Substances 0.000 description 1
- 206010020751 Hypersensitivity Diseases 0.000 description 1
- 101100484788 Hypocrea virens (strain Gv29-8 / FGSC 10586) virD gene Proteins 0.000 description 1
- 101150062179 II gene Proteins 0.000 description 1
- 102000004195 Isomerases Human genes 0.000 description 1
- 108090000769 Isomerases Proteins 0.000 description 1
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- 241000270322 Lepidosauria Species 0.000 description 1
- 108700005092 MHC Class II Genes Proteins 0.000 description 1
- 239000007993 MOPS buffer Substances 0.000 description 1
- 235000011430 Malus pumila Nutrition 0.000 description 1
- 244000070406 Malus silvestris Species 0.000 description 1
- 235000015103 Malus silvestris Nutrition 0.000 description 1
- 241000219470 Mirabilis Species 0.000 description 1
- NEEQFPMRODQIKX-REOHCLBHSA-N N(3)-oxalyl-L-2,3-diaminopropionic acid Chemical compound OC(=O)[C@@H](N)CNC(=O)C(O)=O NEEQFPMRODQIKX-REOHCLBHSA-N 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 101710138657 Neurotoxin Proteins 0.000 description 1
- 101710163270 Nuclease Proteins 0.000 description 1
- JLPDBLFIVFSOCC-UHFFFAOYSA-N Oleandrin Natural products O1C(C)C(O)C(OC)CC1OC1CC(CCC2C3(CC(C(C3(C)CCC32)C=2COC(=O)C=2)OC(C)=O)O)C3(C)CC1 JLPDBLFIVFSOCC-UHFFFAOYSA-N 0.000 description 1
- 102000015636 Oligopeptides Human genes 0.000 description 1
- 108010038807 Oligopeptides Proteins 0.000 description 1
- 229930012538 Paclitaxel Natural products 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 102000003992 Peroxidases Human genes 0.000 description 1
- 244000264897 Persea americana var. americana Species 0.000 description 1
- OAICVXFJPJFONN-UHFFFAOYSA-N Phosphorus Chemical compound [P] OAICVXFJPJFONN-UHFFFAOYSA-N 0.000 description 1
- 108091000080 Phosphotransferase Proteins 0.000 description 1
- 241000209049 Poa pratensis Species 0.000 description 1
- 241000209504 Poaceae Species 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 239000004365 Protease Substances 0.000 description 1
- 108010029485 Protein Isoforms Proteins 0.000 description 1
- 102000001708 Protein Isoforms Human genes 0.000 description 1
- 101710167959 Putative UTP-glucose-1-phosphate uridylyltransferase Proteins 0.000 description 1
- 241000220324 Pyrus Species 0.000 description 1
- 238000012181 QIAquick gel extraction kit Methods 0.000 description 1
- 108020004412 RNA 3' Polyadenylation Signals Proteins 0.000 description 1
- 235000002226 Ranunculus ficaria Nutrition 0.000 description 1
- 244000081426 Ranunculus ficaria Species 0.000 description 1
- 108700005079 Recessive Genes Proteins 0.000 description 1
- 102000052708 Recessive Genes Human genes 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 108700005075 Regulator Genes Proteins 0.000 description 1
- 241000701507 Rice tungro bacilliform virus Species 0.000 description 1
- 101150113575 SBEI gene Proteins 0.000 description 1
- 101150110449 SBEII gene Proteins 0.000 description 1
- 101100166255 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) CEP3 gene Proteins 0.000 description 1
- 108010016634 Seed Storage Proteins Proteins 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- 244000079002 Solanum demissum Species 0.000 description 1
- 101000979697 Streptomyces carzinostaticus 2-hydroxy-5-methyl-1-naphthoate 7-hydroxylase Proteins 0.000 description 1
- 229940100389 Sulfonylurea Drugs 0.000 description 1
- 108010006785 Taq Polymerase Proteins 0.000 description 1
- 101710203193 Thaumatin-like protein Proteins 0.000 description 1
- 108010036937 Trans-cinnamate 4-monooxygenase Proteins 0.000 description 1
- HSCJRCZFDFQWRP-JZMIEXBBSA-N UDP-alpha-D-glucose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1OP(O)(=O)OP(O)(=O)OC[C@@H]1[C@@H](O)[C@@H](O)[C@H](N2C(NC(=O)C=C2)=O)O1 HSCJRCZFDFQWRP-JZMIEXBBSA-N 0.000 description 1
- 208000036142 Viral infection Diseases 0.000 description 1
- 208000027418 Wounds and injury Diseases 0.000 description 1
- 235000016383 Zea mays subsp huehuetenangensis Nutrition 0.000 description 1
- 241000482268 Zea mays subsp. mays Species 0.000 description 1
- 238000000184 acid digestion Methods 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 108010050516 adenylate isopentenyltransferase Proteins 0.000 description 1
- 239000011543 agarose gel Substances 0.000 description 1
- 208000026935 allergic disease Diseases 0.000 description 1
- 230000007815 allergy Effects 0.000 description 1
- 102000004139 alpha-Amylases Human genes 0.000 description 1
- 229940024171 alpha-amylase Drugs 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 125000000539 amino acid group Chemical group 0.000 description 1
- 229940126575 aminoglycoside Drugs 0.000 description 1
- 229940025131 amylases Drugs 0.000 description 1
- 230000010165 autogamy Effects 0.000 description 1
- 101150103518 bar gene Proteins 0.000 description 1
- ZYGHJZDHTFUPRJ-UHFFFAOYSA-N benzo-alpha-pyrone Natural products C1=CC=C2OC(=O)C=CC2=C1 ZYGHJZDHTFUPRJ-UHFFFAOYSA-N 0.000 description 1
- BGWGXPAPYGQALX-ARQDHWQXSA-N beta-D-fructofuranose 6-phosphate Chemical compound OC[C@@]1(O)O[C@H](COP(O)(O)=O)[C@@H](O)[C@@H]1O BGWGXPAPYGQALX-ARQDHWQXSA-N 0.000 description 1
- 230000000035 biogenic effect Effects 0.000 description 1
- 239000000872 buffer Substances 0.000 description 1
- 238000010804 cDNA synthesis Methods 0.000 description 1
- 229940027138 cambia Drugs 0.000 description 1
- 230000034303 cell budding Effects 0.000 description 1
- 210000002421 cell wall Anatomy 0.000 description 1
- 230000001413 cellular effect Effects 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 239000003153 chemical reaction reagent Substances 0.000 description 1
- 210000003483 chromatin Anatomy 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 101150043616 cna gene Proteins 0.000 description 1
- 238000004737 colorimetric analysis Methods 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 230000002596 correlated effect Effects 0.000 description 1
- 235000001671 coumarin Nutrition 0.000 description 1
- 150000004775 coumarins Chemical class 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 1
- 210000000805 cytoplasm Anatomy 0.000 description 1
- 231100000599 cytotoxic agent Toxicity 0.000 description 1
- 229940127089 cytotoxic agent Drugs 0.000 description 1
- 239000002254 cytotoxic agent Substances 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000004665 defense response Effects 0.000 description 1
- 230000007850 degeneration Effects 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- KXZOIWWTXOCYKR-UHFFFAOYSA-M diclofenac potassium Chemical compound [K+].[O-]C(=O)CC1=CC=CC=C1NC1=C(Cl)C=CC=C1Cl KXZOIWWTXOCYKR-UHFFFAOYSA-M 0.000 description 1
- 235000005911 diet Nutrition 0.000 description 1
- 230000000378 dietary effect Effects 0.000 description 1
- 108020001096 dihydrofolate reductase Proteins 0.000 description 1
- 238000002845 discoloration Methods 0.000 description 1
- GYQBBRRVRKFJRG-UHFFFAOYSA-L disodium pyrophosphate Chemical compound [Na+].[Na+].OP([O-])(=O)OP(O)([O-])=O GYQBBRRVRKFJRG-UHFFFAOYSA-L 0.000 description 1
- 210000005069 ears Anatomy 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 239000003623 enhancer Substances 0.000 description 1
- 230000007613 environmental effect Effects 0.000 description 1
- 238000006911 enzymatic reaction Methods 0.000 description 1
- 239000002532 enzyme inhibitor Substances 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 230000035558 fertility Effects 0.000 description 1
- 230000004720 fertilization Effects 0.000 description 1
- 235000012631 food intake Nutrition 0.000 description 1
- 230000008014 freezing Effects 0.000 description 1
- 238000007710 freezing Methods 0.000 description 1
- 239000000499 gel Substances 0.000 description 1
- 238000012239 gene modification Methods 0.000 description 1
- 230000005017 genetic modification Effects 0.000 description 1
- 244000037671 genetically modified crops Species 0.000 description 1
- 235000013617 genetically modified food Nutrition 0.000 description 1
- 229940116332 glucose oxidase Drugs 0.000 description 1
- 235000019420 glucose oxidase Nutrition 0.000 description 1
- 108010046301 glucose peroxidase Proteins 0.000 description 1
- 125000004383 glucosinolate group Chemical group 0.000 description 1
- 229940097068 glyphosate Drugs 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 230000007407 health benefit Effects 0.000 description 1
- 239000000185 hemagglutinin Substances 0.000 description 1
- 230000006801 homologous recombination Effects 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 238000009396 hybridization Methods 0.000 description 1
- 150000002429 hydrazines Chemical class 0.000 description 1
- 108010002685 hygromycin-B kinase Proteins 0.000 description 1
- 230000028993 immune response Effects 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 230000000415 inactivating effect Effects 0.000 description 1
- 230000002779 inactivation Effects 0.000 description 1
- 238000009399 inbreeding Methods 0.000 description 1
- 108091006086 inhibitor proteins Proteins 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 208000014674 injury Diseases 0.000 description 1
- 230000003834 intracellular effect Effects 0.000 description 1
- 238000007852 inverse PCR Methods 0.000 description 1
- 150000002500 ions Chemical class 0.000 description 1
- JEIPFZHSYJVQDO-UHFFFAOYSA-N iron(III) oxide Inorganic materials O=[Fe]O[Fe]=O JEIPFZHSYJVQDO-UHFFFAOYSA-N 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- 238000005304 joining Methods 0.000 description 1
- 238000002386 leaching Methods 0.000 description 1
- 125000001909 leucine group Chemical group [H]N(*)C(C(*)=O)C([H])([H])C(C([H])([H])[H])C([H])([H])[H] 0.000 description 1
- 108010053156 lipid transfer protein Proteins 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 238000009630 liquid culture Methods 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 235000009973 maize Nutrition 0.000 description 1
- 230000000442 meristematic effect Effects 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 229960000485 methotrexate Drugs 0.000 description 1
- 238000009629 microbiological culture Methods 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 238000000520 microinjection Methods 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 238000007479 molecular analysis Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- MEFBJEMVZONFCJ-UHFFFAOYSA-N molybdate Chemical compound [O-][Mo]([O-])(=O)=O MEFBJEMVZONFCJ-UHFFFAOYSA-N 0.000 description 1
- 231100000219 mutagenic Toxicity 0.000 description 1
- 231100000707 mutagenic chemical Toxicity 0.000 description 1
- 231100000243 mutagenic effect Toxicity 0.000 description 1
- 239000013642 negative control Substances 0.000 description 1
- 238000007857 nested PCR Methods 0.000 description 1
- 239000002581 neurotoxin Substances 0.000 description 1
- 231100000618 neurotoxin Toxicity 0.000 description 1
- 231100000252 nontoxic Toxicity 0.000 description 1
- 230000003000 nontoxic effect Effects 0.000 description 1
- 238000001216 nucleic acid method Methods 0.000 description 1
- 230000035764 nutrition Effects 0.000 description 1
- JLPDBLFIVFSOCC-XYXFTTADSA-N oleandrin Chemical compound O1[C@@H](C)[C@H](O)[C@@H](OC)C[C@@H]1O[C@@H]1C[C@@H](CC[C@H]2[C@]3(C[C@@H]([C@@H]([C@@]3(C)CC[C@H]32)C=2COC(=O)C=2)OC(C)=O)O)[C@]3(C)CC1 JLPDBLFIVFSOCC-XYXFTTADSA-N 0.000 description 1
- 229950010050 oleandrin Drugs 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 238000009400 out breeding Methods 0.000 description 1
- 150000003891 oxalate salts Chemical class 0.000 description 1
- 230000003647 oxidation Effects 0.000 description 1
- 238000007254 oxidation reaction Methods 0.000 description 1
- 229960001592 paclitaxel Drugs 0.000 description 1
- 239000002245 particle Substances 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 101150072645 pea gene Proteins 0.000 description 1
- 235000021017 pears Nutrition 0.000 description 1
- 239000008194 pharmaceutical composition Substances 0.000 description 1
- 150000003017 phosphorus Chemical class 0.000 description 1
- 102000020233 phosphotransferase Human genes 0.000 description 1
- 210000000745 plant chromosome Anatomy 0.000 description 1
- 230000008121 plant development Effects 0.000 description 1
- 239000013600 plasmid vector Substances 0.000 description 1
- 210000002706 plastid Anatomy 0.000 description 1
- 231100000614 poison Toxicity 0.000 description 1
- 239000002574 poison Substances 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 235000013606 potato chips Nutrition 0.000 description 1
- 235000013573 potato product Nutrition 0.000 description 1
- 230000003389 potentiating effect Effects 0.000 description 1
- 239000000843 powder Substances 0.000 description 1
- 230000001376 precipitating effect Effects 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 210000001938 protoplast Anatomy 0.000 description 1
- 238000000746 purification Methods 0.000 description 1
- 239000000376 reactant Substances 0.000 description 1
- 239000011541 reaction mixture Substances 0.000 description 1
- 230000005849 recognition of pollen Effects 0.000 description 1
- 230000033458 reproduction Effects 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 238000010839 reverse transcription Methods 0.000 description 1
- 150000008223 ribosides Chemical class 0.000 description 1
- 101150040247 rl gene Proteins 0.000 description 1
- 229930182490 saponin Natural products 0.000 description 1
- 235000017709 saponins Nutrition 0.000 description 1
- 150000007949 saponins Chemical class 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 238000010187 selection method Methods 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 239000011780 sodium chloride Substances 0.000 description 1
- 239000002904 solvent Substances 0.000 description 1
- 229960000268 spectinomycin Drugs 0.000 description 1
- UNFWWIHTNXNPBV-WXKVUWSESA-N spectinomycin Chemical compound O([C@@H]1[C@@H](NC)[C@@H](O)[C@H]([C@@H]([C@H]1O1)O)NC)[C@]2(O)[C@H]1O[C@H](C)CC2=O UNFWWIHTNXNPBV-WXKVUWSESA-N 0.000 description 1
- 230000006641 stabilisation Effects 0.000 description 1
- 238000011105 stabilization Methods 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 108010008664 streptomycin 3''-kinase Proteins 0.000 description 1
- 239000000758 substrate Substances 0.000 description 1
- YROXIXLRRCOBKF-UHFFFAOYSA-N sulfonylurea Chemical class OC(=N)N=S(=O)=O YROXIXLRRCOBKF-UHFFFAOYSA-N 0.000 description 1
- 239000006228 supernatant Substances 0.000 description 1
- 230000008093 supporting effect Effects 0.000 description 1
- 230000001629 suppression Effects 0.000 description 1
- 239000000725 suspension Substances 0.000 description 1
- 238000004114 suspension culture Methods 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- RCINICONZNJXQF-MZXODVADSA-N taxol Chemical compound O([C@@H]1[C@@]2(C[C@@H](C(C)=C(C2(C)C)[C@H](C([C@]2(C)[C@@H](O)C[C@H]3OC[C@]3([C@H]21)OC(C)=O)=O)OC(=O)C)OC(=O)[C@H](O)[C@@H](NC(=O)C=1C=CC=CC=1)C=1C=CC=CC=1)O)C(=O)C1=CC=CC=C1 RCINICONZNJXQF-MZXODVADSA-N 0.000 description 1
- 210000003411 telomere Anatomy 0.000 description 1
- 102000055501 telomere Human genes 0.000 description 1
- 108091035539 telomere Proteins 0.000 description 1
- 231100000167 toxic agent Toxicity 0.000 description 1
- 239000003440 toxic substance Substances 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 108700012359 toxins Proteins 0.000 description 1
- UZKQTCBAMSWPJD-UQCOIBPSSA-N trans-Zeatin Natural products OCC(/C)=C\CNC1=NC=NC2=C1N=CN2 UZKQTCBAMSWPJD-UQCOIBPSSA-N 0.000 description 1
- UZKQTCBAMSWPJD-FARCUNLSSA-N trans-zeatin Chemical compound OCC(/C)=C/CNC1=NC=NC2=C1N=CN2 UZKQTCBAMSWPJD-FARCUNLSSA-N 0.000 description 1
- GOSWTRUMMSCNCW-UHFFFAOYSA-N trans-zeatin riboside Natural products C1=NC=2C(NCC=C(CO)C)=NC=NC=2N1C1OC(CO)C(O)C1O GOSWTRUMMSCNCW-UHFFFAOYSA-N 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 230000032258 transport Effects 0.000 description 1
- 125000000430 tryptophan group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C2=C([H])C([H])=C([H])C([H])=C12 0.000 description 1
- 101150016042 udp gene Proteins 0.000 description 1
- 241001515965 unidentified phage Species 0.000 description 1
- 229940035893 uracil Drugs 0.000 description 1
- 108091000036 uracil phosphoribosyltransferase Proteins 0.000 description 1
- 210000003934 vacuole Anatomy 0.000 description 1
- 230000009385 viral infection Effects 0.000 description 1
- 230000003612 virological effect Effects 0.000 description 1
- 230000001018 virulence Effects 0.000 description 1
- 230000000007 visual effect Effects 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 1
- 239000003643 water by type Substances 0.000 description 1
- 230000004580 weight loss Effects 0.000 description 1
- 229940023877 zeatin Drugs 0.000 description 1
Images





Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8202—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation by biological means, e.g. cell mediated or natural vector
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/415—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from plants
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8202—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation by biological means, e.g. cell mediated or natural vector
- C12N15/8205—Agrobacterium mediated transformation
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8209—Selection, visualisation of transformants, reporter constructs, e.g. antibiotic resistance markers
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8209—Selection, visualisation of transformants, reporter constructs, e.g. antibiotic resistance markers
- C12N15/821—Non-antibiotic resistance markers, e.g. morphogenetic, metabolic markers
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8216—Methods for controlling, regulating or enhancing expression of transgenes in plant cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8216—Methods for controlling, regulating or enhancing expression of transgenes in plant cells
- C12N15/8218—Antisense, co-suppression, viral induced gene silencing [VIGS], post-transcriptional induced gene silencing [PTGS]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8216—Methods for controlling, regulating or enhancing expression of transgenes in plant cells
- C12N15/8222—Developmentally regulated expression systems, tissue, organ specific, temporal or spatial regulation
- C12N15/8223—Vegetative tissue-specific promoters
- C12N15/8226—Stem-specific, e.g. including tubers, beets
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
- C12N15/8243—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
- C12N15/8243—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine
- C12N15/8245—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine involving modified carbohydrate or sugar alcohol metabolism, e.g. starch biosynthesis
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
- C12N15/8243—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine
- C12N15/825—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine involving pigment biosynthesis
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8271—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance
- C12N15/8273—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance for drought, cold, salt resistance
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0055—Oxidoreductases (1.) acting on diphenols and related substances as donors (1.10)
- C12N9/0057—Oxidoreductases (1.) acting on diphenols and related substances as donors (1.10) with oxygen as acceptor (1.10.3)
- C12N9/0059—Catechol oxidase (1.10.3.1), i.e. tyrosinase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
- C12N9/1051—Hexosyltransferases (2.4.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1294—Phosphotransferases with paired acceptors (2.7.9)
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L19/00—Products from fruits or vegetables; Preparation or treatment thereof
- A23L19/10—Products from fruits or vegetables; Preparation or treatment thereof of tuberous or like starch containing root crops
- A23L19/12—Products from fruits or vegetables; Preparation or treatment thereof of tuberous or like starch containing root crops of potatoes
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L19/00—Products from fruits or vegetables; Preparation or treatment thereof
- A23L19/10—Products from fruits or vegetables; Preparation or treatment thereof of tuberous or like starch containing root crops
- A23L19/12—Products from fruits or vegetables; Preparation or treatment thereof of tuberous or like starch containing root crops of potatoes
- A23L19/18—Roasted or fried products, e.g. snacks or chips
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L5/00—Preparation or treatment of foods or foodstuffs, in general; Food or foodstuffs obtained thereby; Materials therefor
- A23L5/10—General methods of cooking foods, e.g. by roasting or frying
- A23L5/11—General methods of cooking foods, e.g. by roasting or frying using oil
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- Organic Chemistry (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Zoology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Biophysics (AREA)
- Cell Biology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Nutrition Science (AREA)
- Medicinal Chemistry (AREA)
- Virology (AREA)
- Botany (AREA)
- Gastroenterology & Hepatology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
métodos de modificação de uma característica ou uma célula de um.a planta selecionada e da expressão de um. gene funcional, para fazer um.a planta transformada de modo estável com um polinucleotídeo pretendido, de identi:ficação de uma célula alvo de planta, de redução da mancha preta e do adocicamento induzido por frio, plantas, caracteristicas, polinucleotídeos e seqüências, vetor, p-dna, promotor específico de tubérculo e método de fazer células transgênicas de planta a presente invenção relaciona-se a novos processos de procriação de plantas. o processo melhora o desempenho agronômico de plantas de colheitas pelo uso de material genético que é também usado na procriação clássica. em vez de recombinar sexualmente genomas inteiros de modo aleatôrio, como é feito na procr1açao clássica, são rearrumados in vitro elementos específicos e inseridos de volta em células individuais de plantas. as plantas obtidas através deste novo processo de procriação de plantas não contêm ácidos nucleicos estranhos, mas, das espécies contêm apenas ácidos nucleicos de plantas selcionadas para transformação ou plantas que sejam sexualmente compatíveis com a espécie de planta selecionada. são proporcionadas plantas desenvolvidas através deste novo processo de procriação de plantas. em particular. são providas plantas de batatas que exibem características de armazenagem e de saúde melhoradas.
Description
[001] Este Pedido reivindica prioridade dos Pedidos Provisórios de números de série U.S. 60/357.661 e 60/377.602, que são aqui incorporados por referência. A presente invenção relacionase a métodos para melhorar as características nutricionais, de saúde e agronômicas de uma planta por modificação de DNA específico, bem caracterizado no genoma da planta. Em oposição à reprodução clássica de plantas, o processo inventivo não introduz genes desconhecidos ou potencialmente tóxicos na maquiagem genética da planta. Além disso, o método inventivo, diferentemente das estratégias de engenharia genética convencional, não incorpora ácidos nucleicos de espécie estranhas, isto é espécies que não sejam férteis entre si com a planta a ser modificada por engenharia genética, no genoma da planta. As plantas desenvolvidas através do processo inventivo de procriação de plantas exibem características agronômicas aperfeiçoadas. As plantas particularmente preferidas da presente invenção incluem batatas que apresentam características melhoradas de saúde e de armazenamento em tubérculos e gramas que exibem tolerância incrementada à seca e à doença.
[002] O desempenho agronômico de plantas tem sido melhorado caracteristicamente por procriação clássica de plantas ou engenharia genética. A procriação clássica resulta tipicamente na transferência de ácidos nucleicos desconhecidos de uma planta para outra. As técnicas de engenharia genética introduzem ácidos nucleicos estranhos no genoma da planta, isto é, DNA que não é de uma planta ou que não é de uma planta que seja naturalmente fértil com a planta a ser modificada por engenharia genética. Por exemplo, a engenharia genética introduz ácidos nucleicos que não são de plantas num genoma da planta. Tanto a procriação clássica como as estratégias de engenharia genética criam genomas da plantas que contêm material genético indesejável e não pretendido e as plantas de resultantes de procriação cruzada ou transgênicas podem exibir características desfavoráveis. As insuficiências de ambas as estratégias podem provarse prejudiciais para as plantas transgênicas, assim como também para os animais e pessoas humanas que consomem tais produtos.
[003] A procriação de plantas conta tipicamente com a recombinação fortuita de cromossomos de plantas para criar variedades que tenham características novas e melhoradas. Deste modo, por triagem de grandes populações de progénie que resultam de cruzamentos de plantas, os criadores conseguem identificar aquelas plantas que exibem uma característica desejada, como aumento no rendimento, vigor melhorado, resistência incrementada contra doenças e insetos ou maior capacidade de sobrevivência sob condições de seca. Contudo, os métodos de procriação clássicos são laboriosos e demorados e as novas variedades exibem tipicamente melhorias apenas relativamente modestas.
[004] Além disso, a criação clássica de plantas resulta tipicamente na transferência de centenas de genes desconhecidos para o genoma da planta. É provável que alguns desses genes transferidos codifiquem alégenos potencialmente prejudiciais, tais como patatina, lectinas, quitinases, proteases, proteínas semelhantes a taumatinas, proteínas de transferência de lípidos, amilases, inibidores de tripsina e proteínas de armazenamento em sementes (Breiteneder e colaboradores, J. Allergy Clin. Immunol. 106: 27—36).
[005] De modo semelhante, os genes introduzidos podem ser envolvidos na biossíntese de toxinas inclusive latirógenos, hidrazinas, glucosinolatos e bociógenos, cumarinas, saponinas, alcalóides, glicoalcalóides, aminas biogênicas, inibidores de enzimas, como lectinas (hemaglutininas), inibidores de tripsina, substâncias quelantes como fitatos e oxalatos, ribotoxinas, peptídeos antimicrobianos, aminoácidos como betaNoxalilaminoLalanina, atractilosidas, oleandrina, taxol e isoquinolina (Pokorny, Cas Lek Cesk 136: 267- 70,1997). O risco de introduzir inadvertidamente tais venenos no material de humano e alimentos animais é ainda aumentado através de esforços para “liberar” a diversidade genética de parentes de colheitas selvagens que não foram usados antes no consumo de alimentos (Hoisington e colaboradores, ProcNatlAcad Sci USA 96: 5937-43, 1999).
[006] Embora a procriação clássica de plantas possa introduzir facilmente genes envolvidos em compostos antinutricionais indesejáveis em colheitas de alimentos e plantas, não consegue removêlos facilmente. Por exemplo, levou mais ou menos 15 anos para reduzir níveis prejudiciais de fitatos no milho e no arroz por genes Lpa de inativação (Raboy, J. Nutr 132: 503S-505S, 2002). O longo período de tempo para perceber resultados positivos não é prático, especialmente porque existe a necessidade urgente de métodos que mais efetiva e eficazmente melhorem a qualidade de colheitas de alimentos. Um exemplo de um gene que apenas recentemente foi achado estar associado à síntese de compostos antinutricionais é o gene da polifenol oxidase (PPO), que oxida certos compostos fenólicos para produzir agentes mutagênicos, carcinológicos e citotóxicos como radicais fenoxil e derivados quinóides (Kagan e colaboradores, Biochemistry 33: 9651- 60, 1994). A presença de cópias múltiplas deste gene no genoma de plantas como a batata torna particularmente difícil reduzir a atividade de PPO através da procriação.
[007] Até mais tempo é necessário para a remoção de compostos antinutricionais se pouco ou nada for conhecido sobre a sua base genética. Por exemplo, nenhum gene foi ligado à acumulação de concentrações altas de acrilamida, uma potente neurotoxina e mutágeno em algumas batatas que são aquecidas a 160°C ou mais alto (Tareke e colaboradores, J. Agric. Food Chem. 50: 4998-5006, 2002). É, portanto, muito difícil desenvolver eficazmente novas variedades de batata que produzam menos acrilamida durante o processamento usando a procriação convencional. Deste modo existe uma necessidade de fazer crescer batatas e outros alimentos ricos em carboidrato, como trigo, com níveis reduzidos desses compostos perigosos, mas, sem o uso de ácidos nucleicos desconhecidos ou estranhos.
[008] Outros compostos antinutricionais que podem acumularse durante o processamento e são difíceis de minimizar ou eliminar através da procriação incluem os produtos de reação de Maillard NNitrosoN(3-ceto-l,2 butanediol) — 3'nitrotiramina (Wang e colaboradores, Arch. Toxicol. 70: 10-5, 1995) e 5-hidroximetil-2 furfural (Janzowski e colaboradores, Food Chem Toxicol 38:801-9, 2000). Ê também sabido que produtos adicionais da reação de Maillard que não foram bem caracterizados exibem propriedades mutagênicas (Shibamoto, Prog Clin Biol Res 304: 359-76, 1989).
[009] Pode ser igualmente difícil aumentar rapidamente os níveis de compostos nutricionais positivos em colheitas de alimentos devido à imprecisão inerente à procriação convencional de plantas. Por exemplo, seria desejável aumentar os níveis de “amido resistente” (Topping e colaboradores, Physiol Rev 81: 1031-64, 2001) numa variedade de colheitas. Esse amido, em última instância, é responsável por promover respostas imuno, suprimindo patógenos potenciais e reduzindo a incidência de doenças, inclusive câncer coloretal (Bird e colaboradores, Curr Issues Intest Microbiol 1: 25-37, 2000). Contudo, as únicas plantas disponíveis com níveis aumentados de amido resistente são variedades de baixo rendimento como mutantes de milho “amilose extender”, “dull” e “sugary-2”. A criação de novas fontes de amido de alta resistência, tais como a batata, possibilitaria a modalidade dietética mais ampla deste componente promotor de saúde.
[0010] A incapacidade de manipular seguramente os genótipos de plantas conduz frequentemente ao uso de substâncias químicas externas para induzir um fenótipo desejado. Apesar de numerosos programas de procriação para demorar a brotação de tubérculos, por exemplo, nenhumas variedades de batata estão comercialmente disponíveis que possam ser armazenadas por meses sem tratamento com inibidores de brotação. Os últimos, tais como isopropilNclorofenilcarbamato (CIPC), estão ligados a toxicidade e desenvolvimento agudos de tumores e podem estar presentes em alimentos de batata processada em concentrações entre 1 mg/kg e 5 mg/kg.
[0011] A Engenharia Genética pode ser usada para modificar, produzir ou remover certas características das plantas. Se bem que tenha havido progresso limitado na melhoria das características de valor nutritivo e de saúde das plantas, a maior parte das melhorias visa características de plantas que promovem a facilidade do cultivo das colheitas. Deste modo, certas plantas são resistentes ao herbicida glifosato porque elas contêm o gene bacteriano 5-enolpiruvilshiquimato- 3-fosfatase sintase (Padgette e colaboradores, Arch Biochem Biophys. 258: 564-73, 1987). Semelhantemente, a engenharia genética tem produzido variedades de plantas resistentes a insetos, vírus e fungos (Shah e colaboradores, Trends in Biotechnology 13: 362-368, 1995; Gao e colaboradores, Nat. Biotechnol. 18: 1307-10, 2000; Osusky e colaboradores, Nat. Biotechnol. 18: 1162-6, 2000), mas, poucas com benefícios incrementados de nutrição ou saúde.
[0012] De acordo com as técnicas padrão, bem conhecidas, “cassetes de expressão” genética, incluindo genes e elementos reguladores, são inseridos dentro das bordas de DNAs de transferência isolados em Agrobacterium (“T-DNAs”) e integrados no genoma de plantas. Deste modo, o material de T-DNA de transferência mediado por Agrobacterium compreende tipicamente os seguintes procedimentos normais: (1) recombinação in vitro de elementos genéticos, um dos quais, pelo menos, é de origem estranha, para produzir um cassete de expressão para seleção da transformação, (2) inserção deste cassete de expressão, frequentemente junto com, pelo menos, outro cassete de expressão contendo DNA estranhos, numa região de T-DNA de um vetor binário, que normalmente consiste em várias centenas de pares de base de DNA de Agrobacterium flanqueado por sequências de borda de T- DNA, (3) transferência das sequências localizadas entre as bordas de T- DNA, frequentemente acompanhadas de algumas ou todas as sequências de vetor binário adicionais de Agrobacterium para a célula da planta e (4) seleção de células de planta transformadas de modo estável. Ver, por exemplo, Patentes U.S. 4.658.082; 6.051.757; 6.258.999; 5.453.367; 5.767.368; 6.403.865; 5.629.183; 5.464.763; 6.201.169; 5.990.387; 4.693.976; 5.886.244; 5.221.623; 5.736.369; 4.940.838; 6.153.812; 6.100.447; 6.140.553; 6.051.757; 5.731.179; 5.149.645 e EP 0 120.516; EP 0 257.472; EP 0 561.082; 1.009.842A1, 0 853.675A1; 0 486.233B1; 0 554.273A1; 0 270.822A1; 0 174.166A1 e WO 01/25459.
[0013] Assim, os métodos de engenharia genética contam com a introdução de ácidos nucleicos estranhos nos suprimentos de alimentos. Aquelas técnicas transferem fusões complexas desde alguns até mais de 20 elementos genéticos isolados de vírus, bactérias e plantas, que não são nativos para a espécie de planta transformada. Esses elementos estranhos incluem elementos reguladores tais como promotores e terminadores e genes que são envolvidos na expressão de uma nova característica ou função como marcadores para identificar ou selecionar eventos de transformação. Apesar dos testes de alimentos contendo DNA estranhos quanto à segurança antes da aprovação regulatória, muitos consumidores estão preocupados com os efeitos a longo prazo de comerem alimentos que expressam proteínas estranhas, que são produzidas por genes obtidos de outras espécies de não plantas.
[0014] Um elemento regulador comumente usado é o “super” promotor 35S do vírus do mosaico da couveflor (CaMV), que é tipicamente usado em engenharia de plantas para induzir níveis altos de expressão de transgenes aos quais está diretamente ligado. Contudo, o promotor 35S pode aumentar também a expressão de genes nativos em sua vizinhança (Weigel e colaboradores, Plant Physiol., 122: 1003-13, 2000). Esses promotores podem, assim, induzir alterações impossíveis de prever na expressão de genes endógenos, resultando possivelmente em efeitos indesejáveis, como a produção aumentada de alcalóides. Os promotores “fortes” preferidos são geralmente aqueles isolados de vírus, como o vírus baciliforme tungro do arroz, o vírus de estria do milho, o vírus vein da cassava, o vírus de Mirabilis, o caulimovírus da veia clorótica do amendoim, o vírus do mosaico do figwort e o vírus chlorella. Outros promotores frequentemente usados são clonados de espécies bacterianas e incluem os promotores da sintase da nopalina e o gene da sintase da octopina.
[0015] Para obter a terminação apropriada da tradução de genes, são fundidas sequências terminadoras na extremidade 3' de transgenes e incluem elementos genéticos da sintase da nopalina e genes da sintase da octopina de Agrobacterium. Podem ser usados outros elementos genéticos para intensificar a expressão genética ou visar a proteína expressa para certos compartimentos das células. Estes elementos incluem introns, para impulsionar as sequências da expressão transgênica, e peptídeo de sinal, para visar o gene estranho para certos compartimentos celulares, frequentemente derivados de espécies de plantas estranhas.
[0016] Certos genes envolvidos na expressão de uma nova característica são mais frequentemente derivados de fontes estranhas. Se forem usados genes nativos, eles estão frequentemente invertidos, para silenciar a expressão daqueles genes em plantas transgénicas e cotransformados com DNA estranhos como marcador selecionável. A desvantagem principal desta tecnologia “de antisentido” é que o DNA invertido normalmente contém molduras novas e não caracterizadas de leitura aberta inseridas entre um promotor e um terminador. Deste modo, plantas de batata que foram geneticamente modificadas com construtos antisentido derivados do gene do amido relacionado Rl (Kossmann e colaboradores, Patente US 6.207.880), os genes glucan fosforilase do tipo L e H (Kawchuk e colaboradores, Patente US 5.998.701, 1999), o gene da polifenol oxidase (Steffens, Patente US 6.160.204, 2000) e os genes das enzimas I e II da ramificação do amido (Schwall e colaboradores, Nature Biotechnology 18: 551-554, 2000) todos expressam potencialmente novos peptídeos que consistem em, pelo menos, 50 aminoácidos (Tabela 1). Estes novos peptídeos podem interferir no desenvolvimento da planta e/ou reduzir o valor nutricional da batata e são, portanto, indesejáveis.
[0017] Os genes marcadores convencionais são incorporados em construtos genéticos e usados para seleção de eventos de transformação. Eles conferem resistência a antibiótico ou herbicida (Patente US 6.174.724), uma vantagem metabólica (Patente US 5.767.378) ou um fenótipo morfologicamente anormal (Patente US 5.965.791) para a planta transformada. Tais marcadores são tipicamente derivados de fontes bacterianas.
[0018] Além disso, por causa da infidelidade da transferência de T- DNA, mais ou menos 75% dos eventos de transformação em plantas como tomate, tabaco e batata contêm sequências de “esqueleto” de plasmideos, além do T-DNA (Kononov e colaboradores, Plant J 11: 945- 57, 1997). A presença dessas sequências de esqueleto é indesejável, porque são estranhas e contêm tipicamente origens de replicação e marcadores de gene de resistência a antibióticos.
[0019] Existem vários métodos para remover elementos como genes marcadores estranhos, mas, poucos são facilmente aplicáveis à engenharia genética de plantas. De acordo com um desses métodos, o gene marcador e o gene ou sequência de nucleotídeos pretendidos são colocados em vetores diferentes. A infecção de plantas com uma cepa única de Agrobacterium portando dois vetores (Patente US 6.265.638) ou duas cepas de Agrobacterium, cada uma das quais porta um dos vetores, pode resultar ocasionalmente em eventos de integração não ligados, que podem ser geneticamente separados através da outbreeding. A desvantagem principal deste método é que a separação genética de loci pode ser muito laboriosa e delongada especialmente se os eventos de integração de T-DNA estiverem ligados. Além disso, este método não é amplamente aplicável a plantas apomíticas, que se reproduzem assexuadamente, como a Kentucky bluegrass, ou colheitas propagadas vegetativamente como a batata, que não pode ser rapidamente criada, devido à depressão de procriação consaguínea, altos níveis de heterozigosidade e baixos níveis de fertilidade.
[0020] Outro método para remover elementos genéticos estranhos conta com inserção de gene estranhos, como o marcador selecionável, num elemento transponível. O elemento modificado transponível pode então, ser juntado fora do genoma a baixas frequências. Os cruzamentos tradicionais com plantas não transformadas deve, então, ser executado para separar o elemento transposto do hospedeiro (Patente US 5.482.852). Como descrito para o método anterior, este método alternativo não pode ser usado para sistemas propagados vegetativamente nem plantas apomíticas.
[0021] Um terceiro método de remoção de um gene marcador usa o sistema de recombinação de sítio específico Cre/lox do bacteriófago Pl (Dale & Ow, Proc. Natl. Acad. Sci. EUA, 88: 10558-62, 1991). A inserção de um gene marcador junto com o gene da recombinase Cre e um gene quimérico envolvido na indução de Cre (ambos com seus próprios promotores e terminadores) entre dois sítios lox conduz à excisão da região delineada pelos sítios lox durante o processo de regeneração (Zuo e colaboradores, Nat. Biotechnol., 19: 157-61, 2001). Este processo complicado é ineficiente e não confiável e pode causar instabilidade do genoma.
[0022] Estudos recentes reportam que alguns genes de plantas podem ser usados eles mesmos como marcadores de transformação. Os exemplos desses marcadores de plantas incluem Pga22 (Zuo e colaboradores, Curr Opin Biotechnol. 13: 173-80, 2002), Ckil (Kakimoto Science, 274: 982-985, 1996) e Esrl (Banno e colaboradores, Plant Cell, 13: 2609-18, 2001). Todos os genes, porém, acionam respostas de citoquinina, que conferem um fenótipo indesejável para a planta transformada. Além disso esses marcadores de planta ainda precisariam de ser removidos, após a transformação, por qualquer dos métodos descritos acima.
[0023] Os métodos alternativos para transformar plantas são também baseados na recombinação in vitro de elementos genéticos estranhos e contam com sequências de plasmídeos bacterianos para manutenção em E. coli, cujas partes são cointegradas durante o processo de transformação. Os exemplos desses métodos para transformar plantas com DNA estranhos são descritos nas Patentes US 5.591.616; 6.051.757; 4.945.050; 6,143.949; 4.743.548; 5.302.523 e 5.284.253.
[0024] Podem ser também obtidas plantas transgênicas isentas de marcadores omitindo quaisquer procedimentos de seleção anteriores à regeneração. Uma desvantagem deste método é que a maioria dos eventos gerados através deste método representarão plantas não transformadas ou quiméricas, porque normalmente não serão derivadas de células de planta transformada única. Ê extremamente difícil e laborioso usar um procedimento isento de marcador para a identificação de plantas transgênicas que contêm a(s) mesma(s) inserção(ões) de DNA(s) em todas as suas células.
[0025] Assim existe uma necessidade muito importante da melhoria de plantas para além do que pode ser realizado através dos cruzamentos da procriação clássica e das técnicas de engenharia genética convencional e que não conte com a inserção de ácidos nucleicos desconhecidos ou estranhos no genoma da planta. Consequentemente, a presente invenção proporciona métodos e composições para modificar com precisão o próprio material genético da planta. Deste modo, a estratégia inventiva da “procriação precisa” não induz fenótipos indesejáveis e não introduz ácidos nucleicos desconhecidos ou estranhos no genoma da planta.
[0026] A presente invenção proporciona métodos para aumentar geneticamente o valor nutricional e o desempenho agronômico de uma planta sem a incorporação permanente ou estável de DNA desconhecido ou estranho no genoma daquela planta. De acordo com os métodos da presente invenção, ácidos nucleicos específicos, bem caracterizados, elementos de genes e genes são isolados de uma espécie de planta desejada ou de uma espécie de planta que seja sexualmente compatível com a planta desejada, modificados e, depois, reinseridos de volta no genoma da espécie da planta desejada. A modificação pode acarretar a mutação da sequência de ácido nucléico isolada, deletando partes do ácido nucléico isolado ou simplesmente juntando o ácido nucléico isolado a outro polinucleotídeo, tal como subclonar o ácido nucléico isolado num vetor de plasmideo.
[0027] Consequentemente, as plantas transgénicas produzidas pela metodologia inventiva não possuem genomas que compreendam ácidos nucleicos de nenhuma espécie estranha. Deste modo, os métodos da presente invenção produzem uma planta transgênica cujo genoma não compreende um promotor de espécie de nãoplanta, não compreende um terminador de espécie de nãoplanta, não compreende uma região 5' não transladada de espécie de nãoplanta, não compreende uma região 3' não transladada de espécie de nãoplanta, não compreende um gene marcador de espécie de nãoplanta, não compreende um elemento regulador de espécie de nãoplanta, não compreende um gene de espécie de nãoplanta e não compreende nenhum outro polinucleotídeo que seja obtido de um genoma de espécie de nãoplanta.
[0028] Assim, a presente invenção proporciona um método para produzir uma planta transgênica estável que exibe um fenótipo modificado que não é exibido pela planta não transformada, compreendendo (a) transformar células de plantas com um polinucleotídeo pretendido; (b) cultivar plantas a partir das células transformadas; e (c) selecionar uma planta transformada de modo estável com o referido polinucleotídeo pretendido que exibe um novo fenótipo que não é exibido por plantas crescidas a partir das células da planta não transformada correspondente. Preferentemente, o polinucleotídeo pretendido consiste essencialmente em (i) sequências de ácidos nucleicos que são isoladas e/ou nativas para o genoma das células da planta ou para outras plantas da mesma espécie ou são isoladas e/ou nativas para o genoma de uma espécie de planta que seja sexualmente compatível com a planta a partir da qual as células da planta foram isoladas; e (ii), pelo menos, uma sequência de DNA que é uma sequência semelhante a borda que tem uma sequência que é nativa para o genoma das referidas células d planta ou é nativa para o genoma de células de plantas da mesma espécie ou é nativa para uma planta que seja sexualmente compatível com a planta de que as células de planta foram isoladas e em que a sequência semelhante a borda é capaz de integrar de modo estável o polinucleotídeo pretendido no genoma das citadas células de planta.
[0029] Um método preferido da presente invenção acarreta a produção de uma planta transgênica que exibe um fenótipo modificado que não é exibido pela planta não transformada, compreendendo (a) infectar explantes com Agrobacterium portando (i) um vetor “P-DNA”, que contém um polinucleotídeo pretendido, que é nativo para a planta transgênica e (ii) um vetor de “LifeSupport” que contém um cassete de expressão contendo um gene marcador selecionável; (b) selecionar a expressão transiente do gene marcador selecionável, preferentemente para 1-10 dias, para 3-7 dias ou para 4-5 dias; (c) transferir explantes para meios de regeneração para permitir a formação de brotos; (d) fazer a triagem de populações de brotos para determinar quais compreendem, pelo menos, uma cópia do polinucleotídeo pretendido em seus genomas e, desses, que brotos não contêm nenhuns ácidos nucleicos estranhos, tal como o gene marcador selecionável em seus genomas; e (e) permitir brotos que contenham o polinucleotídeo pretendido em seus genomas, mas, nenhum DNA de gene marcador, para desenvolver plantas inteiras em que as plantas inteiras resultantes exibem um fenótipo modificado que não é exibido por plantas cultivadas a partir de células de plantas não transformadas da mesma espécie.
[0030] De acordo com esse método, o polinucleotídeo pretendido (i) consiste essencialmente apenas em elementos que são isolados e/ou nativos para o genoma da espécie da célula da planta ou espécies sexualmente compatíveis com a mesma; (ii) compreende, pelo menos, um elemento de borda que tem uma sequência que é isolada ou nativa para o genoma da espécie de células da planta ou espécie sexualmente compatível com a mesma e é capaz de integrar de modo estável o polinucleotídeo pretendido no genoma de uma célula de planta exposta ao vetor; e (iii) é integrada de modo estável no genoma da planta transformada; em que o método não integra DNA de espécies de nãoplanta ou estranhos no genoma da planta transformada.
[0031] Além disso, qualquer gene marcador selecionável pode ser usado como um indicador de transformação bem sucedida. Por exemplo, pode ser usado um gene marcador de “fosfotransferase de neomicina” ou um “gene marcador de “hpt” para conferir resistência aos antibióticos de aminoglicosídeos, canamicina e higromicina, respectivamente. Outros genes marcadores incluem o gene marcador de “bar”, que confere resistência para o herbicida fosfinotricina; o gene marcador de “DHFR”, que confere resistência ao metotrexato; e o gene marcador de “ESPS”, que confere resistência ao herbicida Roundup. Ê bem conhecido na técnica como seguir a expressão desses genes marcadores para determinar se o gene marcador foi ou não expresso de modo estável no genoma de uma célula da planta transformada. Consequentemente, o especialista qualificado sabe como seguir a expressão do gene marcador para determinar que o gene marcador seja apenas transientemente expresso na célula da planta transformada.
[0032] Noutro aspecto da invenção, provêse um método para fazer uma planta transformada de modo estável compreendendo as etapas de: (1) identificar um gene objetivo; (2) isolar uma sequência de DNA líder ou de arrasto associada ao referido gene objetivo; (3) de modo opcional, modificar o citado DNA isolado líder ou de arrasto; (4) ligar operacionalmente dito DNA líder ou de arrasto a elementos reguladores nativos para formar um cassete de expressão; (5) inserir o referido cassete de expressão num P-DNA que fica localizado num vetor binário em que o vetor binário porta também um gene operável citocinina tal que a inserção inadvertida de sequências de vetores binários adicionais, que são de origem estranha, seja detectada pela expressão do gene citocinina; (6) introduzir o vetor binário modificado no Agrobacterium; (7) integrar de modo estável o DNA nativo reorganizado nos genomas de células de planta usando a transformação mediada por LifeSupport; (8) regenerar células de plantas que contém o DNA nativo reorganizado; (9) descartar plantas que exibam um fenótipo de superprodução de citocinina e não se regenerem completamente; e (10) manter, para análise adicional, as plantas desejáveis que sejam indistinguíveis de plantas não transformadas.
[0033] Noutro aspecto da presente invenção, é provido um método para modificar a expressão de uma característica numa espécie de planta selecionada. Numa modalidade, o método compreende (1) identificar a característica a ser modificada; (2) construir uma molécula de DNA recombinante consistindo essencialmente em elementos genéticos isolados ou nativos para a espécie de planta selecionada em que a molécula de DNA recombinante, quando integrada no genoma da espécie de planta selecionada, modifica a expressão da característica na espécie de planta transformada; (3) integrar de modo estável a molécula de DNA recombinante em células da espécie da planta selecionada usando transformação mediada por LifeSupport; e (4) identificar plantas transformadas que exibam expressão modificada da característica.
[0034] Numa modalidade preferida, o polinucleotídeo que é nativo para uma planta pretendida é inserido no genoma da planta desejada infectando explantes com duas cepas diferentes de Agrobacterium. Uma primeira cepa de Agrobacterium é capaz de transferir o DNA nativo de vetores P-DNA para células da planta; uma segunda cepa pode transferir um T-DNA portando um cassete de expressão para um gene marcador selecionável para células da planta. Os exemplos do último vetor incluem os vetores denominados “LifeSupport” aqui descritos. Selecionando, de preferência, plantas que expressam transientemente o gene marcador durante 1-10 dias, durante 3-7 dias, ou durante 4-5 dias e transferindo subsequentemente os explantes para meios de regeneração, é obtida uma população de eventos, parte da qual representa plantas que contêm, pelo menos, uma cópia do polinucleotídeo, mas, a que faltam quaisquer cópias do T-DNA ou do gene marcador.
[0035] Noutra modalidade, é usada uma cepa única de Agrobacterium que porta tanto um vetor P-DNA, que aloja o gene nativo, pretendido, de interesse, como o polinucleotídeo entre sequências semelhantes à borda P-DNA e um vetor de LifeSupport, que contém um gene marcador. O gene marcador pode ser ou não inserido entre sequências semelhantes a borda P-DNA, sequências semelhantes a borda T-DNA ou outras sequências semelhantes a borda T-DNA.
[0036] Assim, noutra modalidade preferida, o vetor de P-DNA contém, pelo menos, dois cassetes de expressão, um dos quais compreende um gene marcador nativo rastreável ou selecionável acionado por um promotor nativo e seguido de um terminador nativo.
[0037] Selecionando preferentemente durante pelo menos 2 dias e, com maior preferência, pelo menos 5 dias para a expressão do gene marcador nativo e transferindo subsequentemente os explantes para meios de regeneração, é obtida uma população de eventos que representam plantas que contêm, pelo menos, uma cópia do DNA introduzido, integrado de modo estável em seu genomas. Em modalidades preferidas, o gene marcador derivado da planta codifica uma sintase de ácido 5-enolpiruvul-3-fosfoshiquímico ou triptofano decarboxilase mutante. Numa modalidade de maior preferência, o marcador selecionável codifica para tolerância a sal. Numa modalidade de maior preferência, o gene de tolerância a sal tem a sequência de nucleotideos mostrada na SEQ ID NO35 e é usado para selecionar para eventos de transformação em batata.
[0038] Ainda noutra modalidade, a expressão modificada da característica é caracterizada por um aumento na expressão, uma diminuição na expressão ou em expressão indetectável.
[0039] Noutro aspecto da presente invenção, é provida uma planta feita pelo método de (1) identificando a característica a ser modificada; (2) construindo uma molécula de DNA recombinante consistindo essencialmente em elementos genéticos isolados da espécie de planta selecionada em que a molécula de DNA recombinante, quando integrada no genoma da espécie da planta selecionada, modifica a expressão da característica na espécie da planta transformada; (3) integrando de modo estável a molécula de DNA recombinante em células da espécie da planta selecionada através da transformação mediada por LifeSupport; e (4) identificando as plantas transformadas que exibem expressão modificada da característica.
[0040] Num aspecto adicional, é proporcionado um método de modificar a expressão de uma característica numa espécie de planta selecionada. Este método compreende (1) identificar a característica a ser modificada; (2) construir uma molécula de DNA recombinante que consiste essencialmente em (a) elementos genéticos isolados da espécie de planta selecionada em que os elementos genéticos, quando integrados no genoma da espécie da planta selecionada, modificam a expressão da característica na espécie da planta transformada; e (b) um gene marcador selecionável que é isolado da mesma espécie de planta; (3) integrar de modo estável a molécula de DNA recombinante em células da espécie da planta selecionada através da transformação mediada por LifeSupport; (4) detectar o gene marcador selecionável; e (5) identificar as plantas transformadas que exibem expressão modificada da característica.
[0041] Ainda noutro aspecto, é provida uma planta que exibe uma expressão modificada de uma característica. Numa modalidade, a planta tem integrada de modo estável em seu genoma uma molécula de DNA recombinante que consiste essencialmente em elementos genéticos isolados de uma planta da mesma espécie ou de uma planta que seja sexualmente compatível com aquela espécie em que a molécula de DNA recombinante modifica a expressão da característica.
[0042] Noutro aspecto da presente invenção, é proporcionada uma sequência isolada de nucleotideos referida como “plantaDNA” (“P-DNA”). Numa modalidade preferida, ao P-DNA propriamente faltam quaisquer genes ou partes dos mesmos e é delineado por sequências “semelhantes a bordas” T-DNA, terminais, que compartilham pelo menos 50%, pelo menos 75%, pelo menos 90% ou pelo menos 95% de identidade de sequência com a sequência de nucleotideos das bordas de T-DNA de qualquer cepa virulenta de Agrobacterium e que suporte uma transferência eficiente do P-DNA inteiro de Agrobacterium para células da planta
[0043] Numa modalidade preferida uma sequência “semelhante a borda” promove e facilita a integração de um polinucleotídeo a que está ligado. Noutra modalidade preferida, cada sequência de término do P- DNA modificado está entre 5-100 bp de comprimento, 10-80 bp de comprimento, 15-75 bp de comprimento, 15-60 bp de comprimento, 15- 50 bp de comprimento, 15-40 bp de comprimento, 15-30 bp de comprimento, 16-30 bp de comprimento, 20-30 bp de comprimento, 21- 30 bp de comprimento, 22-30 bp de comprimento, 23-30 bp de comprimento, 24-30 bp de comprimento, 25-30 bp de comprimento, ou 26-30 bp de comprimento. Com maior preferência, a sequência semelhante a borda está entre 20 e 28 nucleotídeos de comprimento.
[0044] Numa modalidade preferida, as sequências de borda P-DNA esquerda e direita da presente invenção são isoladas e/ou são nativas para o genoma de uma planta que deve ser modificada e não são idênticas em sequência de nucleotídeos a nenhuma sequência de borda T-DNA derivada de Agrobacterium. Assim, numa modalidade, uma sequência de borda P-DNA pode possuir 1, 2, 3, 4, 5, 6, 7, 8, 9,10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 ou mais nucleotídeos que são diferentes de uma sequência de borda T-DNA de uma espécie de Agrobacterium, tal como Agrobacterium tumefaciens ou Agrobacterium rhizogenes. Alternativamente, noutra modalidade, uma sequência de borda P-DNA ou uma sequência semelhante a borda da presente invenção tem pelo menos 95%, pelo menos 90%, pelo menos 80%, pelo menos 75%, pelo menos 70%, pelo menos 60% ou pelo menos 50% de identidade de sequência com uma sequência de borda de T-DNA de uma espécie de Agrobacterium, tal como Agrobacterium tumefaciens ou Agrobacterium rhizogenes. Com maior preferência, uma sequência de borda P-DNA de planta nativa compartilha mais ou 99%, 98%, 97%, 96%, 95%, 94%, 93%, 92%, 91%, 90%, 89%, 88%, 87%, 86%, 85%, 84%, 83%, 82%, 81%, 80%, 79%, 78%, 77%, 76%, 75%, 74%, 73%, 72%, 71%, 70%, 69%, 68%, 67%, 66%, 65%, 64%, 63%, 62%, 61% ou 60% de identidade de sequência de nucleotideo com uma sequência de borda T-DNA de Agrobacterium.
[0045] Noutra modalidade preferida, uma sequência semelhante a borda pode ser isolada do genoma de uma planta e, depois, modificada ou mutacionada para alterar a eficiência pela qual é capaz de integrar uma sequência de nucleotídeo noutra sequência de nucleotídeo. Em outra modalidade, outras sequências de polinucleotídeos podem ser adicionadas ou incorporadas numa sequência semelhante a borda da presente invenção. Deste modo, ainda noutra modalidade, uma borda esquerda P-DNA ou uma borda direita P-DNA pode ser modificada de modo a possuir sítios de clonagem 5’ e 3’ múltiplos ou sítios de restrição adicionais. Numa modalidade adicional, uma sequência de borda P- DNA pode ser modificada para aumentar a probabilidade de que o DNA de esqueleto do vetor acompanhante não seja integrado no genoma da planta.
[0046] Numa modalidade de maior preferência, os P-DNAs são isolados de qualquer planta pelo uso de primários degenerados numa reação de cadeia de polimerase. Numa modalidade preferida, o P-DNA é derivado da batata, é delineado por terminais de 25 bps com 80 e 88% de identidade com bordas T-DNA convencionais, respectivamente, e tem a sequência de nucleotideos mostrada na SEQ ID NO.: 1. Noutra modalidade de maior preferência, o P-DNA é derivado do trigo, é delineado por terminais de 25 bps com 72% e 92% identidade com bordas T-DNA convencionais, respectivamente, e contém a sequência de nucleotideos mostrada na SEQ ID NO.: 34.
[0047] Esse P-DNA pode ser modificado de modo a compreender outros polinucleotídeos posicionados entre as sequências semelhantes a borda. Numa modalidade preferida, o P-DNA modificado consiste essencialmente, na direção de 5’ para 3', numa primeira sequência semelhante a borda que promove a transferência de DNA, num promotor, num polinucleotídeo pretendido que é ligado operaionalmente ao promotor, num terminador e numa segunda sequência semelhante a borda que promove também a transferência de DNA. Noutra modalidade, o polinucleotídeo pretendido representa uma ou várias cópias de um líder, um arrasto ou um gene em orientações de sentido e/ou antisentido. Numa modalidade de maior preferência, o P-DNA modificado contém cassetes de expressão tanto para um gene PPO quanto um gene inibidor de invertase mutante.
[0048] Deste modo, numa modalidade preferida, o polinucleotídeo pretendido compreende uma sequência de sentido e de antisentido de uma sequência líder. Numa modalidade de maior preferência, a sequência líder está associada a um gene que é endógeno para uma célula da espécie de planta selecionada. Ainda numa modalidade de maior preferência, o líder está associado a um gene que é selecionado do grupo que consiste num gene PPO, um gene Rl, um gene de alfa glucan fosforilase do tipo L ou H, um gene de glicose glucosiltransferase de UDP, um gene HOS1, um gene Sadenosilhomocisteína hidrolase, um gene da classe II de cinamato 4-hidroxilase, um gene cinamoilcoenzima A reductase, um gene de cinamoil álcool dehidrogenase, um gene cafeoil coenzima A Ometiltransferase, um gene do fator de despolimerização da actina, um gene de Nin88, um Lol p 5 gene, um gene alérgeno, um gene P450 hidroxilase, um gene ADPglucose pirofosforilase, um gene prolina dehidrogenase, um gene endo-l,4-betaglucanase, um gene zeaxantin epoxidase e um gene 1-aminociclopropano-l-carboxilato sintase.
[0049] Ainda noutra modalidade preferida, a sequência de polinucleotídeos pretendida compreende uma sequência de sentido e de antisentido de uma sequência de arrasto. Numa modalidade preferida, a sequência de arrasto está associada com um gene selecionado do grupo que consiste num gene PPO, um gene Rl, um gene alfa glucan fosforilase do tipo L ou H, um gene de glicose glucosiltransferase UDP, um gene HOSI, um gene Sadenosilhomocisteína hidrolase, um gene cinamato 4-hidroxilase da classe II, um gene cinamoilcoenzima A reductase, um gene cinamoil álcool dehidrogenase, um gene cafeoil coenzima A Ometiltransferase, um gene do fator despolimerizante da actina, um gene Nin88, um gene Lol p 5, um gene alérgeno, um gene P450 hidroxilase, um gene ADP glicose pirofosforilase, um gene prolina dehidrogenase, um gene endo-l,4-betaglucanase, um gene zeaxantina epoxidase e um gene 1-aminociclopropano-l carboxilato sintase.
[0050] Numa modalidade preferida, o polinucleotídeo pretendido, tal como um gene, é isolado e/ou é nativo para a planta que deve ser transformada. Noutra modalidade preferida, o polinucleotídeo pretendido é modificado ou mutacionado. Numa modalidade, uma mutação para o polinucleotídeo isolado pode tornar o nucleotide pretendido maior ou igual a 99%, 98%, 97%, 96%, 95%, 94%, 93%, 92%, 91%, 90%, 89%, 88%, 87%, 86%, 85%, 84%, 83%, 82%, 81%, 80%, 79%, 78%, 77%, 76%, 75%, 74%, 73%, 72%, 71%, 70%, 69%, 68%, 67%, 66%, 65%, 64%, 63%, 62%, 61%, ou 60% dissemelhante da sua forma não mutacionada.
[0051] Numa modalidade preferida da presente invenção, o promotor de um cassete de expressão localizado dentro de um P-DNA é um promotor constitutivo. Numa modalidade de maior preferência, o promotor constitutivo é o promotor do gene Ubiquitin-3 da batata. Numa modalidade até de maior preferência, o promotor constitutivo é o promotor do gene Ubiquitin-7 da batata.
[0052] Noutra modalidade, o promotor de um cassete de expressão localizado dentro de um P-DNA é um promotor regulável. Numa modalidade de maior preferência, o promotor regulável é sensível à temperatura. Numa modalidade até de maior preferência, o promotor regulável é um promotor ci21A ou um promotor C17, cada um isolado da batata (Schneider e colaboradores, Plant Physiol. 113: 335-45, 1997; Kirch e colaboradores, Plant Mol Biol 33: 897-909, 1997).
[0053] Noutra modalidade, o promotor de um cassete de expressão localizado dentro de um P-DNA pode ser regulado de um modo temporal. Numa modalidade preferida, o promotor é um promotor rbcS (Ueda e colaboradores, Plant Cell 1: 217-27, 1989).
[0054] Ainda noutra modalidade, o promotor de um cassete de expressão localizado dentro de um P-DNA é regulado por qualquer um dentre ácido abscíssico, wounding, metil jasmonato ou ácido giberélico. Numa modalidade adicional, este promotor é um promotor selecionado quer de um promotor de gene Rab 16A, um promotor de gene de <x- amilase ou um promotor de gene pin2.
[0055] Noutra modalidade, o promotor de um cassete de expressão localizado dentro de um P-DNA é um promotor de tecido específico. Numa modalidade particularmente preferida, este promotor é um promotor GBSS isolado de S. tuberosum.
[0056] Numa modalidade, a presente invenção provê um vetor P- DNA que é capaz de replicação tanto na E. coli como no Agrobacterium e contém um P-DNA ou um P-DNA modificado. Numa modalidade preferida, este vetor contém também um cassete de expressão para um gene citocinina no seu esqueleto para possibilitar a seleção contra eventos de integração de esqueleto.
[0057] Noutra modalidade preferida, a sequência de nucleotideos pretendida compreende ainda um elemento espaçador. Numa modalidade de maior preferência, o elemento espaçador é uma sequência do intron Ubi ou uma sequência do espaçador GBSS.
[0058] Noutra modalidade preferida, a sequência de nucleotideos pretendida compreende um gene nativo mutacionado que codifica uma proteína funcionalmente inativa, que reduz a atividade global daquela proteína se expressa em plantas transgênicas. Ainda numa modalidade de maior preferência, este gene mutacionado codifica uma polifenol oxidase funcionalmente inativa carente de um domínio de ligação de cobre.
[0059] Noutra modalidade preferida, a sequência de nucleotideos pretendida inclui um gene nativo que codifica uma proteína funcionalmente ativa. Ainda numa modalidade de maior preferência, este gene codifica para uma proteína com homologia para o inibidor da invertase do tabaco vacuolar.
[0060] Noutra modalidade, o terminador de um cassete de expressão localizado dentro de um P-DNA é uma sequência terminadora Ubi3 ou uma região 3' não transladada de um gene de uma espécie de planta selecionada.
[0061] Noutro aspecto da presente invenção, é proporcionado um método para modificar uma célula de planta alvo. Numa modalidade, o método compreende: (1) inserir um P-DNA modificado no genoma de, pelo menos, uma célula na célula alvo da planta usando a transformação mediada por LifeSupport; e (2) observar se existe uma mudança fenotípica na célula da planta alvo; em que o promotor no P- DNA modificado transcreve as sequências de sentido e/ou de antisentido não transladadas associadas com um gene nativo para reduzir a expressão daquele gene nativo, modificando, assim, a célula de planta alvo. Noutra modalidade preferida, o promotor no P-DNA modificado transcreve um gene para sobrexpressar esse gene na célula de planta alvo.
[0062] Ainda noutro aspecto, é provido um método para produzir uma célula de planta transgênica de uma espécie de planta selecionada que contém um P-DNA modificado. O método compreende cotransfectar uma célula de planta da espécie de planta selecionada com um vetor de P-DNA e um vetor de LifeSupport que compreende um gene marcador flanqueado por um T-DNA de borda esquerda e um T- DNA de borda direita e um gene mutante virD2 inserido no esqueleto do vetor e selecionar uma célula de planta que transientemente expresse o gene marcador e isolar uma célula de planta que contenha o P-DNA modificado integrado em seu genoma, mas, não contenha nenhuns nucleotideos do vetor LifeSupport. Numa modalidade preferida, o gene marcador confere resistência à canamicina. Numa modalidade de maior preferência, a levedura ADH terminadora segue o gene de resistência à canamicina.
[0063] Numa modalidade preferida, a célula de planta da espécie de planta selecionada visada para transformação está em cultura. Noutra modalidade preferida, a célula de planta da espécie de planta selecionada visada para transformação está dentro de uma planta.
[0064] A presente invenção proporciona também uma planta da espécie selecionada que compreende, pelo menos, uma célula com um genoma que contém um P-DNA modificado. Numa modalidade preferida, o P-DNA modificado consiste essencialmente, na direção de 5’ para 3’, num primeiro término que funciona como uma borda de T-DNA seguida de sequências de P-DNA, um promotor, uma sequência de nucleotideos pretendida ligada operavelmente tanto a um promotor, como a uma sequência terminadora e sequências adicionais de P-DNA delineadas por um segundo terminal. Noutra modalidade, o polinucleotídeo pretendido representa uma ou várias cópias de um líder, um arrasto e um gene numa orientação de sentido e/ou de antisentido.
[0065] Noutra modalidade, é encarada uma planta que compreende, pelo menos, uma célula com um genoma que contém um P-DNA modificado.
[0066] Noutro aspecto da invenção, é provido um método para reduzir a expressão de um gene numa espécie de planta selecionada. O método compreende a transformação mediada por LifeSupport de uma célula de planta de uma espécie de planta selecionada com um vetor de P-DNA em que o P-DNA modificado deste vetor é integrado de modo estável no genoma da célula de planta. Noutro aspecto da invenção, o P-DNA modificado compreende um polinucleotídeo desejado que reduz a expressão de um gene endógeno da espécie de planta selecionada.
[0067] Noutro aspecto da presente invenção, um gene nativo para a espécie de planta selecionada pode ser mutacionado e reintroduzido na planta usando os métodos inventivos. Preferentemente, o gene mutacionado, por exemplo, um gene PPO mutacionado, é integrado no genoma da célula da planta usando um vetor de P-DNA.
[0068] A presente invenção também proporciona um método para reduzir a expressão indesejável do gene da polifenol oxidase numa espécie de planta selecionada. Numa modalidade preferida, o método compreende integrar no genoma de uma espécie de planta selecionada um P-DNA modificado compreendido apenas de sequências de nucleotideos isoladas da espécie de planta selecionada ou de uma planta que seja sexualmente compatível com a espécie de planta selecionada, consistindo essencialmente, na direção de 5’ para 3‘, num primeiro término de P-DNA que funciona como uma borda de T-DNA seguida de sequências flanqueadoras de P-DNA; num promotor; num nucleotideo pretendido, que é uma sequência de nucleotideos de arrasto de orientação de sentido associada com um gene PPO específico; numa sequência de orientação de antisentido da sequência de arrasto de nucleotideos do gene PPO específico; numa sequência de terminação e sequências adicionais de P-DNA delineadas por um segundo término que funciona como uma borda T-DNA em que o promotor produz uma molécula de RNA de filamento duplo que reduz a expressão do gene PPO específico, reduzindo, assim, a mancha preta contundida em tecidos específicos da planta. Noutra modalidade, as sequências de nucleotideos de orientação de sentido e de antisentido das sequências líder de nucleotideos são obtidas a partir da região não transladada 5’ que precede o gene PPO específico. Numa modalidade adicional, a sequência líder ou de arrasto de orientação de sentido e de antisentido associada com o gene PPO pode ser separada por outra sequência de polinucleotídeos, aqui referida quer como um intron quer como um “espaçador”. Numa modalidade preferida, a sequência líder ou de arrasto está associada com um gene PPO da batata. Numa modalidade de maior preferência, a sequência líder ou de arrasto está associada com um gene PPO da batata que é expresso em tubérculos da batata. Numa modalidade de maior preferência, a sequência líder ou de arrasto está associada com um gene PPO da batata que é expresso em todas as partes do tubérculo de batata com exceção da epiderme.
[0069] A presente invenção provê também um método para reduzir a produção de acrilamida, a indução de broto durante o armazenamento, a acumulação de fosfato e/ou adocicamento induzido pelo frio em tubérculos de uma espécie de planta selecionada.
[0070] Numa modalidade preferida, o método inclui a transformação mediada por LifeSupport de uma espécie de planta selecionada com um P-DNA modificado compreendido apenas de sequências de nucleotideos isoladas da espécie de planta selecionada ou de plantas que sejam sexualmente compatíveis com a espécie de planta selecionada, consistindo essencialmente, na direção de 5’ para 3’, num primeiro P-DNA com uma sequência esquerda semelhante a borda, num promotor, numa sequência de nucleotídeos pretendida, que é uma sequência de nucleotídeos de orientação de sentido da sequência líder associada com o gene Rl, numa sequência de orientação de antisentido desta sequência líder, numa sequência de terminação e numa sequência direita semelhante a borda. Após a expressão, é produzido um duplex de RNA líder que reduz a expressão do gene Rl, reduzindo, dessa maneira, o adocicamento induzido pelo frio na planta. Noutra modalidade, as sequências de nucleotídeos de orientação de sentido e de antisentido pretendidas representam o arrasto associado com o gene Rl. Numa modalidade adicional, o líder de orientação de sentido e de antisentido ou arrasto associados com Rl podem ser separados por outra sequência de polinucleotideos, aqui referida como um intron ou como um “espaçador”.
[0071] Noutra modalidade preferida, o método compreende a transformação mediada por LifeSupport de uma espécie de planta selecionada com um P-DNA modificado que é semelhante ao descrito acima, mas, contém uma sequência líder ou de arrasto associada com um gene alfa glucan fosforilase.
[0072] Ainda noutra modalidade preferida, o método compreende a transformação mediada por LifeSupport de uma espécie de planta selecionada com um P-DNA modificado que contém um gene inibidor da invertase.
[0073] Noutra modalidade preferida, os P-DNA modificados descritos nos parágrafos precedentes são usados para reduzir a acumulação de produtos indesejáveis adicionais da reação de Maillard, que ocorre durante o aquecimento de alimentos ricos em carboidrato como tubérculos de batata. Estes produtos indesejáveis incluem produtos avançados de terminais de glicação (AGEs) que têm sido associados a várias patologias.
[0074] A presente invenção proporciona também um método para aumentar níveis de amido resistente nos órgãos de armazenamento de plantas e de colheitas de alimentos.
[0075] Numa modalidade preferida, o método compreende a transformação mediada por LifeSupport de uma espécie de planta selecionada com um P-DNA modificado que contém um cassete de expressão para uma fusão das sequências de arrasto associadas com os genes das enzimas I e II de ramificação do amido.
[0076] A presente invenção também proporciona sequências de nucleotideos isoladas que compreendem os promotores do gene GBSS da batata e o gene inibidor da proteinase da batata, que estão predominantemente expressos em tubérculos. Os promotores isolados têm a sequência de nucleotideos mostrada na SEQ ID NO.: 6 e SEQ ID NO.: 40, respectivamente.
[0077] Num aspecto, a presente invenção proporciona um método de modificar uma característica de uma planta selecionada que compreende: a. transformar de modo estável células da planta selecionada com um polinucleotídeo pretendido em que o polinucleotídeo pretendido consiste essencialmente numa sequência de ácidos nucleicos que é nativa para a planta selecionada, nativa para uma planta da mesma espécie ou é nativa para uma planta que seja sexualmente fértil com a planta selecionada, b. obter uma planta transformada de modo estável a partir das células da planta transformada em que a planta transformada contém o polinucleotídeo pretendido integrado de modo estável no genoma e em que o polinucleotídeo pretendido modifica a característica.
[0078] Numa modalidade preferida, o método compreende ainda cotransfectar as células da planta com um gene marcador selecionável que seja transientemente expresso nas células da planta e identificar as células transformadas da planta e plantas transformadas obtidas das células da planta transformada em que o gene marcador selecionável não fica integrado de modo estável e o polinucleotídeo pretendido é integrado de modo estável no genoma.
[0079] Numa modalidade preferida, o polinucleotídeo pretendido inclui um P-DNA, um promotor GBSS, um promotor Ubi7, um promotor Ubi3, um promotor PIP, um gene modificado de PPO, um gene inibidor da invertase, um gene de tolerância a sal, um líder associado a Rl, um líder associado à fosforilase, um arrasto associado a Rl, arrastos associados a SBE, um intron Ubi , um espaçador GBSS, UbiT.
[0080] Noutra modalidade preferida, uma “planta” da presente invenção é uma planta monocotiledônea, selecionada do grupo que consiste em trigo, relva, grama de relva, cereal, milho, arroz, aveia, trigo, cevada, sorgo, orquídea, íris, lírio, cebola, banana, canadeaçúcar, sorgo e palma.
[0081] Ainda noutra modalidade, uma “planta” da presente invenção é uma planta dicotiledônea, selecionada do grupo que consiste em abacate, batata, tabaco, tomate, beterraba, brócolis, cassava, batata doce, pimenta, algodão, poinsétia, legumes, alfafa, soja, cenoura, morango, alface, carvalho, ácer, noz, rosa, hortelã, esprema, margarida e cactos.
[0082] Ainda noutra modalidade, células de plantas e plantas dos métodos inventivos presentes são transformadas via transformação mediada por Agrobacterium. Preferentemente, a transformação mediada por Agrobacterium conta com o uso de, pelo menos, um vetor binário. Ainda noutra modalidade, o método de transformação mediada por Agrobacterium usa um primeiro vetor binário e um segundo vetor binário. Numa modalidade preferida, o primeiro vetor binário contém o polinucleotídeo pretendido e o segundo vetor binário contém um gene marcador selecionável em que o gene marcador selecionável é operavelmente ligado a um promotor e um terminador.
[0083] De acordo com os métodos presentes, a característica que é modificada é selecionada do grupo que consiste em saúde e características nutricionais aumentadas, armazenamento melhorado, rendimento aumentado, tolerância aumentada a sal, tolerância aumentado a metal pesado, tolerância aumentada á seca, tolerância aumentada contra doença, tolerância aumentada contra insetos, tolerância aumentada contra stress de água, tolerância aumentada contra frio e geada, tolerância aumentada contra adocicamento, vigor melhorado, gosto melhorado, textura melhorada, conteúdo de fosfato diminuído, germinação aumentada, captura de micronutrientes aumentada, composição de amido melhorada, longevidade de flor melhorada.
[0084] A presente invenção engloba também uma planta feita pelos métodos presentes.
[0085] Noutro aspecto, a presente invenção compreende um método para modificar uma característica numa planta selecionada que compreende: (a) identificar a característica a ser modificada; (b) construir um primeiro polinucleotídeo que consiste essencialmente em elementos genéticos nativos isolados da planta selecionada, uma planta da mesma espécie ou uma planta que seja sexualmente fértil com a planta selecionada em que os elementos genéticos nativos são capazes de modificar a expressão de um gene que controla a característica; (c) construir um segundo polinucleotídeo que compreende um gene marcador selecionável que está operavelmente ligado a um promotor e um terminador; (d) cotransfectar células da planta selecionada com o primeiro e o segundo polinucleotídeos; (e) selecionar a expressão transiente do gene marcador selecionável; (f) rastrear células de planta transformadas de modo estável com o primeiro polinucleotídeo, mas, que não contenham a segunda molécula de DNA integrada no genoma; e (g) obter uma planta transformada de modo estável a partir das células transformadas da planta que exibam uma expressão modificada da característica.
[0086] Numa modalidade, os elementos genéticos compreendem, pelo menos, um promotor, uma sequência de interesse, um terminador, um intensificador, um intron, um espaçador ou elementos reguladores. Noutra modalidade, o método da reivindicação 4 em que as células da planta são transfectadas com o primeiro polinucleotídeo antes do segundo polinucleotídeo ou viceversa.
[0087] Numa modalidade, a sequência de interesse é um gene. Noutra modalidade, o gene é um gene mutacionado ou do tipo selvagem da polifenol oxidase ou um gene mutacionado ou do tipo selvagem de RI. Noutra modalidade, a sequência de interesse é uma sequência de líder ou arrasto em que a sequência de líder ou arrasto representa uma sequência a montante ou a jusante de um gene que é nativo para a célula da planta. Ainda noutra modalidade, a sequência de interesse compreende uma sequência de orientação de sentido operavelmente ligada a uma sequência líder de antisentido. Noutra modalidade, a sequência de interesse compreende uma sequência de arrasto de orientação de sentido operavelmente ligada a uma sequência arrasto de antisentido. Noutra modalidade, o promotor é um promotor induzível. Noutra modalidade, o terminador é uma sequência terminadora da levedura ADH.
[0088] De acordo com a presente invenção, um construto líder compreende, na direção de 5’ para 3’, um promotor, uma sequência líder de orientação de sentido, a sequência de antisentido da líder e um terminador, em que a expressão do construto líder produz uma molécula de RNA de filamento duplo que facilita a subregulação da expressão do gene a que está associado. Noutra modalidade, a sequência líder está associada e localizada a montante da região de codificação do gene PPO, do gene Rl, de um gene da fosforilase do tipo L ou um gene alfa glucan fosforilase.
[0089] Noutra modalidade, o construto de arrasto compreende, na direção de 5’ para 3’, um promotor, uma sequência de arrasto de orientação de sentido, a sequência de arrasto de antisentido e um terminador em que a expressão do construto de arrasto produz uma molécula de filamento duplo que facilita a subregulação da expressão do gene a que está associado. Numa modalidade preferida, a sequência de arrasto está associada e localizada a jusante da região de codificação do gene PPO, do gene Rl, de um gene do tipo L da fosforilase ou um gene alfa glucan fosforilase.
[0090] O método ainda compreende expor a célula da planta a um segundo vetor que compreende um elemento marcador em que o marcador é transientemente expresso na planta transformada e não é integrado de modo estável no genoma da planta transformada. Numa modalidade, o marcador é um gene resistente a herbicida, um gene de resistência a antibiótico ou NPTII.
[0091] Preferentemente, as células de planta são transformadas via transformação mediada por Agrobacterium. Numa modalidade, a transformação mediada por Agrobacterium conta com o uso de, pelo menos, um vetor binário. Ainda noutra modalidade, o método de transformação mediado por Agrobacterium usa um primeiro vetor binário e um segundo vetor binário. Noutra modalidade, o primeiro vetor binário porta o primeiro polinucleotídeo e o segundo vetor binário porta o segundo polinucleotídeo.
[0092] A presente invenção proporciona outro método de modificar a expressão de um gene numa planta selecionada, compreendendo: (a) identificar o gene funcional; (b) construir um primeiro polinucleotídeo consistindo essencialmente em elementos genéticos nativos isolados da planta selecionada, uma planta da mesma espécie que a planta selecionada ou uma planta que seja seO-xualmente fértil com a planta selecionada em que os elementos genéticos nativos são capazes de modificar a expressão do gene; (c) construir um segundo polinucleotídeo que compreende um gene marcador selecionável funcional; (d) cotransfectar células de planta da planta selecionada com o primeiro e o segundo poylnucleotídeos; (e) selecionar a expressão transiente do gene marcador selecionável; (f) rastrear células da planta transformadas de modo estável com o primeiro polinucleotídeo, mas, que não contenham o segundo polinucleotídeo integrado no genoma; e (g) obter uma planta transformada a partir das células da planta transformada que exibam expressão modificada do gene.
[0093] De preferência, as células da planta são transformadas via transformação mediada por Agrobacterium. Numa modalidade, a transformação mediada por Agrobacterium conta com o uso de, pelo menos, um vetor binário. Ainda noutra modalidade, o método de transformação mediado por Agrobacterium usa um primeiro vetor binário e um segundo vetor binário. Noutra modalidade, o primeiro vetor binário porta o primeiro polinucleotídeo e o segundo vetor binário porta o segundo polinucleotídeo.
[0094] Noutra modalidade, o primeiro polinucleotídeo compreende, pelo menos, um P-DNA, um promotor GBSS, um promotor Ubi7, um promotor Ubi3, um promotor PIP, um gene modificado de PPO, um gene inibidor da invertase, um gene de tolerância a sal, um líder associado a Rl, um líder associado a fosforilase, um arrasto associado a Rl, arrastos associados a SBE, intron Ubi, espaçador GBSS, UbiT.
[0095] Noutra modalidade, o segundo polinucleotídeo compreende, pelo menos, um gene marcador selecionável, um polinucleotídeo ómegamutacionado virD2, um polinucleotídeo codA e um polinucleotídeo de fusão codA::upp.
[0096] A presente invenção também engloba uma planta feita por esse método.
[0097] Noutra modalidade, é proporcionada uma planta transgênica que exibe uma expressão modificada de uma característica comparada com a planta não transgênica de que foi derivada em que a planta transgênica é transformada de modo estável com o polinucleotídeo pretendido consistindo essencialmente em elementos genéticos nativos isolados da planta, uma planta aa mesma espécie ou uma planta que seja sexualmente fértil com a planta e em que o polinucleotídeo modifica a expressão da característica.
[0098] Noutra modalidade preferida, a “planta” da presente invenção é uma planta monocotiledônea, selecionada do grupo que consiste em trigo, relva, grama de relva, cereal, milho, arroz, aveia, trigo, cevada, sorgo, orquídea, íris, lírio, cebola, banana, canadeaçúcar, sorgo e palma.
[0099] Ainda noutra modalidade, a “planta” da presente invenção é uma planta dicotiledônea, selecionada do grupo que consiste em avacado, batata, tabaco, tomate, beterraba, brócolis, cassava, batata doce, pimenta, algodão, poinsétia, legumes, alfafa, soja, cenoura, morango, alface, carvalho, ácer, noz, rosa, hortelã, esprema, margarida e cactos.
[OOIOO] Noutra modalidade, a característica é selecionada do grupo que consiste em saúde e características nutricionais aumentadas, armazenamento melhorado, rendimento aumentado, tolerância aumentada contra sal, tolerância aumentada contra metais pesados, tolerância aumentada contra a seca, tolerância aumentada contra doenças, tolerância aumentada contra insetos, tolerância aumentada contra stress de água, tolerância aumentada contra frio e geada, cor aumentada, doçura aumentada, vigor melhorado, gosto melhorado, textura melhorada, conteúdo de fosfato diminuído, germinação aumentada, captura de micronutrientes aumentada, composição de amido melhorada, longevidade de flor melhorada.
[00101] Noutra modalidade, o polinucleotídeo pretendido compreende, pelo menos, um P-DNA, um promotor GBSS, um promotor Ubi7, um promotor Ubi3, um promotor PIP, gene PPO modificado, um gene inibidor da invertase, um gene de tolerância a sal, um líder associado a Rl, um líder associado a fosforilase, arrasto associado a Rl, arrastos associado a SBE, intron Ubi, espaçador GBSS, UbiT.
[00102] A presente invenção também engloba uma sequência de nucleotideos isolada, semelhante a borda situandose em tamanho de 20 até 100 bp, que compartilha entre 52% e 96% de identidade de sequência com uma sequência de borda de T-DNA de Agrobacterium tumefaciens. Numa modalidade preferida, a sequência de nucleotideos isolada é isolada de uma planta monocotiledônea, selecionada do grupo que consiste em trigo, relva, grama de relva, cereal, milho, arroz, aveia, trigo, cevada, sorgo, orquídea, íris, lírio, cebola, banana, canadeaçúcar, sorgo e palma. Noutra modalidade, a sequência de nucleotideos é isolada de uma planta dicotiledônea selecionada do grupo que consiste em batata, tabaco, tomate, beterraba, brócolis, cassava, batata doce, pimenta, algodão, poinsétia, legumes, alfafa, soja, cenoura, morango, alface, carvalho, ácer, noz, rosa, hortelã, esprema, margarida e cactos.
[00103] Ainda noutra modalidade, a sequência de nucleotídeos isolada é isolada a partir da batata e tem uma sequência de nucleotídeos mostrada em SEQ ID NO.: 94 ou 95. Numa modalidade preferida, a sequência de nucleotídeos isolada compartilha 52% de identidade de sequência com uma sequência de borda T-DNA de Agrobacterium tumafaciens. A presente invenção engloba um vetor que inclui essas sequência de nucleotídeos.
[00104] A presente invenção proporciona também um método de fazer uma planta transformada de modo estável com um polinucleotídeo pretendido compreendendo: (a) isolar um P-DNA que é flanqueado por sequências semelhantes a bordas a partir da planta em que as sequências semelhantes a bordas compartilham entre 52% e 96% de identidade de sequência com uma sequência de borda T-DNA de Agrobacterium tumafaciens; (b) inserir o polinucleotídeo pretendido entre as sequências P-DNA semelhantes a borda para formar um construto P- DNA; e (c) transformar uma célula da planta a partir da planta com o construto P-DNA; e (d) recuperar uma planta de célula da planta transformada de modo estável com o construto P-DNA.
[00105] Numa modalidade, o construto P-DNA é portado num vetor compreendido de um gene marcador de integração de esqueleto e são selecionadas células da planta transformada que não contêm o gene backone do marcador de integração. Noutra modalidade, o gene marcador de integração do esqueleto é um gene citocinina. Noutra modalidade, não são selecionados brotos da planta que exibam um fenótipo de superprodução de citocinina. Ainda noutra modalidade, o gene marcador de integração backnone é o gene IPT e não são selecionados brotos da planta que exibam um fenótipo anormal ou não consigam desenvolver raízes.
[00106] Noutra modalidade, as células da planta são de uma planta monocotiledônea selecionada do grupo que consiste em trigo, relva, grama de relva, cereal, milho, arroz, aveia, trigo, cevada, sorgo, orquídea, íris, lírio, cebola, banana, canadeaçúcar, sorgo e palma.
[00107] Noutra modalidade, as células da planta são de uma planta dicotiledônea selecionada do grupo que consiste em batata, tabaco, tomate, beterraba, brócolos, cassava, batata doce, pimenta, algodão, poinsétia, legumes, alfafa, soja, cenoura, morango, alface, carvalho, ácer, noz, rosa, hortelã, esprema, margarida e cactos.
[00108] Preferentemente, as células da planta são transformadas via transformação mediada por Agrobacterium. Numa modalidade, a transformação mediada por Agrobacterium conta com o uso de, pelo menos, um vetor binário. Ainda noutra modalidade, o método de transformação mediado por Agrobacterium usa um primeiro vetor binário e um segundo vetor binário. Noutra modalidade, o primeiro vetor bináio porta o primeiro polinucleotídeo e o segundo vetor binário porta o segundo polinucleotídeo. Numa modalidade adicional, o segundo vetor binário compreende, pelo menos, um gene marcador selecionável negativo e um gene ómegamutacionado virD2 em que o gene marcador selecionável negativo fica posicionado dentro da borda direita de T-DNA e a borda esquerda de T-DNA e em que o gene ómegamutacionado virD2 fica posicionado dentro do esqueleto do segundo vetor binário. Numa modalidade preferida, o segundo vetor binário compreende tanto um gene marcador selecionável negativo posicionado dentro da borda direita de T-DNA como da borda esquerda de T-DNA e um gene ómegamutacionado virD2 posicionado dentro do esqueleto do segundo vetor binário.
[00109] A presente invenção proporciona também um P-DNA consistindo essencialmente, na direção de 5’ para 3’, numa primeira sequência T-DNA semelhante a borda, num promotor, numa sequência de polinucleotídeos pretendida, operavelmente ligada ao promotor, num terminador e num segunda sequência T-DNA semelhante a borda, em que as sequências semelhantes a borda têm menos de 100% de identidade de sequência com sequências de borda T-DNA.
[00110] Numa modalidade preferida, a sequência T-DNA semelhante a bordas, o promotor, o polinucleotídeo pretendido e o terminador são todos isolados a partir da mesma planta, da mesma espécie de planta ou de plantas que sejam sexualmente férteis entre si.
[00111] Ainda noutra modalidade, o P-DNA consiste essencialmente num gene marcador selecionável.
[00112] Ainda noutra modalidade, a sequência T-DNA semelhante a bordas, o promotor, o polinucleotídeo pretendido, o terminador e o gene marcador selecionável são todos isolados da mesma planta, da mesma espécie de planta ou de plantas que sejam sexualmente férteis entre si.
[00113] Ainda noutra modalidade, a sequência de polinucleotídeos pretendida no P-DNA é uma sequência a montante ou a jusante da região de codificação de um gene em que a sequência a montante é uma sequência líder e em que a sequência a jusante é uma sequência de arrasto. Nesta modalidade, a sequência T-DNA semelhante a bordas, o promotor, a sequência líder, a sequência de arrasto, o terminador e o gene marcador selecionável são todos isolados da mesma planta, da mesma espécie de planta ou de plantas que sejam sexualmente férteis entre si.
[00114] Noutra modalidade, são proporcionados pela presente invenção vetores que compreendem esses construtos P-DNA.
[00115] Noutra modalidade, o promotor é um promotor regulável. Ainda noutra modalidade, o promotor regulável é sensível à temperatura. Numa modalidade preferida, o promotor regulável é um promotor trigo wcsl20. Noutra modalidade, o promotor está sob regulação temporal. Ainda noutra modalidade, o promotor é um promotor da carboxilase. Numa modalidade adicional, o promotor da carboxilase é um promotor da carboxilase do milho.
[00116] O promotor pode ser regulado por qualquer um de ácido abscíssico, wounding, metil jasmonate ou ácido giberélico. Noutra modalidade, o promotor é um promotor selecionado de qualquer um dentre o promotor do gene Rab 16A, um promotor do gene da a-amilase ou um promotor do gene pin2. Ainda noutra modalidade, o promotor é um promotor específico de tecido.
[00117] Noutra modalidade, a sequência líder é uma parte de uma região não transladada 5’ de um gene que é endógeno para uma célula da espécie de planta selecionada. Noutra modalidade, a região 5’ não transladada está a montante de um códon de partida de um gene que é selecionado do grupo que consiste num gene PPO, um gene Rl, um gene HOS 1, um gene Sadenosilhomocisteína hidrolase, um gene da classe II do cinamato 4-hidroxilase, um gene da reductase da cinamoilcoenzima A, um gene da cinamoil álcool dehidrogenase, um gene da cafeoil coenzima A Ometiltransferase, um fator despolimerizante da actina, um gene Nin88, um gene Lol p 5, um gene alérgeno, um gene P450 hidroxilase, um gene da ADPglicose pirofosforilase, um gene da prolina dehidrogenase, um gene da endo-l,4-betaglucanase, um gene da zeaxantina epoxidase e um gene da sintase do 1-aminociclopropano-l- carboxilato.
[00118] Noutra modalidade, a sequência de arrasto é uma parte da região não transladada 3’ de um gene que está a jusante de um códon de terminação de um gene selecionado do grupo que consiste num gene PPO, um gene Rl, um gene HOS1, um gene da Sadenosilhomocisteína hidrolase, um gene da classe II do cinamato de 4-hidroxilase, um gene da cinamoilcoenzima A reductase, um gene da cinamoil álcool dehidrogenase, um gene da cafeoil coenzima A Ometiltransferase, um fator despolimerizante da actina, um gene de Nin88, um gene Lol p 5, um gene alérgeno, um gene P450 hidroxilase, um gene da glicose de ADP pirofosforilase, um gene da prolina dehidrogenase, um gene da endo-l,4-betaglucanase, um gene da zeaxantina epoxidase e um gene da 1-aminociclopropano-l-carboxilato sintase.
[00119] O vetor presente pode ainda compreender um elemento espaçador que é uma sequência do intron Ubi ou uma sequência do espaçador GBSS. Noutra modalidade, o vetor compreende um terminador que é uma sequência terminadora Ubi3 ou uma região não transladada 3’ de um gene endógeno da planta.
[00120] Noutra modalidade, o vetor compreende um gene marcador selecionável operavelmente ligado a um promotor constitutivo e um gene de Cre operavelmente ligado a um promotor induzível em que o gene marcador selecionável e o gene de Cre são flanqueados por um primeiro sítio de reconhecimento da recombinase e um segundo sítio de reconhecimento da recombinase. Noutra modalidade, o sítio de reconhecimento da recombinase e o segundo sítio de reconhecimento da recombinase são sítios lox.
[00121] Noutra modalidade, o promotor induzível é um promotor sensível à temperatura, um promotor quimicamente induzido ou um promotor temporal. Ainda noutra modalidade, o promotor induzível é um promotor Ha hspl7.7 G4, um promotor do trigo wcsl20, um promotor do gene Rab 16A, um promotor do gene da amilase A, um promotor do gene pin2, um promotor da carboxilase. Ainda noutra modalidade preferida, compreende também um gene marcador derivado da planta. Noutra modalidade preferida, o gene marcador derivado da planta é um gene da sintase do ácido enolpiruvul-3-fosfoshiquímico.
[00122] Noutro aspecto da presente invenção, é proporcionado um método para modificar uma célula da planta, que compreende integrar uma sequência de P-DNA no genoma de uma célula da planta em que o P-DNA consiste essencialmente, na direção de 5’ para 3’, numa primeira sequência T-DNA semelhante a borda, num promotor, numa sequência pretendida de polinucleotídeo de P-DNA operavelmente ligada a promotor, num terminador e numa segunda sequência T-DNA semelhante a borda em que as sequências semelhantes a borda têm menos de 100% de identidade de sequência com sequências de borda de T-DNA e em que a sequência T-DNA semelhante a borda, o promotor, o polinucleotídeo pretendido e o terminador são todos isolados ou nativos do genoma da célula da planta em que o polinucleotídeo pretendido compreende sequências de sentido e de antisentido de uma sequência líder ou sequência de arrasto que estão associadas a montante ou a jusante com as regiões de não codificação de um gene na planta e em que a expressão do polinucleotídeo pretendido produz um transcrito de RNA de filamento duplo que visa o gene associado com o polinucleotídeo pretendido, modificando, assim, a célula da planta.
[00123] A presente invenção engloba também um método para modificar uma planta, compreendendo: (i) transfectar, pelo menos, uma célula na planta com o vetor da presente invenção; (ii) selecionar uma célula que expresse o marcador selecionável funcional; (iii) isolar a célula que expressa o marcador selecionável funcional; (iv) induzir a expressão do gene de Cre funcional na célula isolada; (v) cultivar a célula isolada; e (vi) observar o fenótipo das células cultivadas; em que um fenótipo que é diferente para uma célula da planta não transfectada indica que a célula da planta alvo foi modificada.
[00124] Numa modalidade preferida, a etapa de seleção deste e doutros métodos da presente invenção é executada identificando que células são resistentes a um antibiótico.
[00125] Noutro aspecto, um método para identificar uma célula de planta alvo, cujo genoma contém um P-DNA, inclui cotransfectar uma célula alvo da planta com o vetor da presente invenção e um segundo vetor derivado de Agrobacterium que inclui um gene marcador flanqueado por uma borda esquerda de T-DNA e uma borda direita de T-DNA e um gene ómegamutacionado vir D2 em que o P-DNA é integrado no genoma da célula alvo da planta e em que nenhuma parte do segundo vetor derivado de Agrobacterium fica integrado no genoma da célula alvo da planta. Numa modalidade preferida, o marcador no segundo vetor derivado de Agrobacterium é um gene da neomicina fosfotransferase.
[00126] Noutro aspecto, é proporcionado o método para identificar uma célula alvo da planta cujo genoma contenha, pelo menos, uma parte de um cassete de integração, ainda compreende selecionar células que sobrevivem a crescimento temporário num meio que contém canamicina em que o genoma das células selecionadas contém só o cassete de integração. Numa modalidade, a célula alvo da planta fica dentro da planta. Uma planta que compreende, pelo menos, uma célula cujo genoma compreende esse P-DNA está também englobada pela presente invenção.
[00127] A presente invenção inclui também uma planta que compreende, pelo menos, uma célula cujo genoma é artificialmente manipulado de modo a conter só ácidos nucleicos derivados da planta em que nenhuma célula da planta contém ácidos nucleicos estranhos integrados no genoma da célula.
[00128] A presente invenção engloba também um polinucleotídeo que compreende a sequência de polinucleotídeos de SEQ ID NO.: 93, em que o polinucleotídeo está entre 20 e 80 nucleotideos de comprimento. Numa modalidade, o polinucleotídeo está entre 21 e 70 nucleotideos de comprimento, entre 22 e 50 nucleotideos de comprimento, entre 23 e 40 nucleotideos de comprimento ou entre 24 e 30 nucleotideos de comprimento.
[00129] Noutro aspecto, a invenção engloba um promotor específico de tubérculo como mostrado na SEQ ID NO.: 40.
[00130] A presente invenção também engloba um método baseado em Agrobacterium para fazer células de planta transgênica que não contêm um gene marcador selecionável integrado de modo estável no DNA nuclear, que compreende: construir um primeiro vetor binário compreendido de um polinucleotídeo que consiste essencialmente num gene funcional pretendido operavelmente ligado a bordas T-DNA ou sequências T-DNA semelhantes a bordas nos terminais 5’ e 3’ do gene funcional desejado; construir um segundo vetor binário compreendido de um gene marcador selecionável funcional operavelmente ligado a bordas T- DNA ou sequências semelhantes a bordas de T-DNA nos terminais 5’ e 3’ do gene marcador selecionável funcional; incubar células de plantas com: uma cepa Agrobacterium que porta o primeiro e o segundo vetores binários; ou ii. uma primeira cepa Agrobacterium que porta o primeiro vetor binário e uma segunda cepa Agrobacterium que porta o segundo vetor binário; selecionar células de planta em que o gene funcional pretendido é integrado no DNA nuclear da planta sem integração do gene marcador selecionável no DNA nuclear da planta a seguir à incubação por um período de tempo apropriado num meio que contém um agente de seleção apropriada.
[00131] Numa modalidade preferida, o gene marcador selecionável é um gene resistente a herbicida ou um gene de resistência a antibiótico. Noutra modalidade preferida, o gene de resistência a antibiótico é o gene NNPTII. Noutra modalidade, o gene de resistência a antibiótico é o gene estrutural npt II operavelmente ligado ao promotor do gene Ubiquitin-7 e o terminador do gene do álcool da dehidrogenase 1 (ADH1) da levedura. De acordo com este método, as células de planta são primeiro incubadas com a primeira cepa Agrobacterium e, então, subsequentemente incubadas com a segunda cepa Agrobacterium ou viceversa.
[00132] Numa modalidade preferida, o primeiro vetor binário ainda compreende um gene marcador de integração binária que pode ser usado para detectar células de planta transformadas de modo estável com sequências de esqueleto de vetor binário. Noutra modalidade, o gene marcador de integração do vetor binário é selecionado do grupo que consiste no gene da resistência a herbicida, no gene da resistência a antibiótico ou NPTII. Ainda noutra modalidade, o segundo vetor binário compreende também uma fusão de gene entre os genes da citosina deaminase bacteriana (codA) e a uracil fosforibsiltransferase (upp), que é inserido entre as sequências semelhantes a bordas T-DNA ou T-DNA e as células da planta são expostas a 5 fluorocitosinas a seguir à incubação com a primeira e a segundo cepas de Agrobacterium, a fim de selecionar aquelas células da planta transformadas com o segundo vetor binário.
[00133] Ainda noutra modalidade, o vetor binário secundário compreende também um gene que reduz a probabilidade de integração de esqueleto. Numa modalidade, esse gene é o gene ómegamutacionado virD2 em que o gene ómegamutacionado virD2 reduz a frequência de integração do gene marcador selecionável no DNA nuclear da planta.
[00134] A presente invenção também engloba uma sequência de nucleotideos isolada que compreende o promotor GBSS isolado de S. tuberosum. Numa modalidade preferida, esta sequência de nucleotideos isolada tem a sequência de nucleotideos que é SEQ ID. NO. 6 ou 13.
[00135] A FIG. 1 - Ilustrações esquemáticas de alguns vetores P- DNA usados na presente invenção. A região P-DNA é indicada como caixa cinzenta. “zpt”=cassete de expressão para o gene ipt; “npt”=cassete de expressão para o gene nptii;” mPPO”=ca.ssete de expressão para um gene PPO modificado; “I7VH”=cassete de expressão para um gene inibidor da invertase; “GUS’= cassete de expressão para o gene GUS; “LPPO”=cassete de expressão para uma cópia de sentido e de antisentido do líder associado com um gene PPO-, “LPH”=cassete de expressão para uma cópia de sentido e de antisentido do líder associado com um gene da fosforilase; “Alj”=cassete de expressão para um homólogo Alfin da batata. Ver texto para detalhes.
[00136] A FIG. 2 - Cassetes de expressão livre de genes.
[00137] A FIG. 3 - Alinhamento de proteínas do inibidor da invertase da batata e do tabaco. ”St”=Solarium tuberosum (batata); “Nt”= Nicotiana tabacum (tabaco).
[00138] A FIG. 4 - Alinhamento de arrastos associados com vários genes PPO.
[00139] A FIG. 5 - Ilustrações esquemáticas de alguns vetores de LifeSupport usados na presente invenção. “codA” é um cassete de expressão para o gene codA; “codA::upp” é um cassete de expressão para o gene codA fundido a upp; “ΩQvirD2” é um cassete de expressão para o gene 'ΩQvirD2.
[00140] A estratégia da “procriação precisa” da presente invenção melhora o desempenho agronômico, o valor nutricional e as características de saúde de plantas e colheitas sem introduzir ácidos nucleicos desconhecidos ou ácidos nucleicos de uma espécie estranha no genoma de uma espécie da planta e sem produzir fenótipos indesejáveis ou efeitos colaterais prejudiciais.
[00141] Deste modo, a presente invenção proporciona uma planta transgênica e métodos para produzir essa planta que não integram ácidos nucleicos de espécie de não planta naquele genoma da planta. Os ácidos nucleicos, os promotores, os elementos reguladores, outras sequências de genes de não codificação, marcadores, polinucleotideos e genes que são integrados no genoma da planta selecionada são todos preferencialmente isolados da planta que deve ser transformada, de plantas da mesma espécie a ser transformada ou de plantas que sejam sexualmente férteis entre si com a planta a ser transformada. Esse ácidos nucleicos “nativos” podem ser mutacionados, modificados ou cojuntados com outros ácidos nucleicos nativos num cassete de expressão e reintegrados no genoma da planta selecionada, de acordo com os métodos aqui descritos. Consequentemente, o genótipo e o fenótipo da planta transgênica é alterado usando apenas os próprios ácidos nucleicos daquela planta selecionada ou usando ácido nucleico de uma planta que seja sexualmente compatível com a planta selecionada.
[00142] Para facilitar a produção dessas plantas transgênicas, a presente invenção faz uso do fato de que nem todos os vetores de T-DNA usados na transformação mediada por Agrobacterium são realmente integrados no genoma da planta; isto é, enquanto um vetor pode ser capturado pela célula da planta, pode não acontecer um evento de integração real. De acordo com a presente invenção, pode usarse esse vetor para portar um gene marcador selecionável para uma célula da planta. As células da planta podem, então, ser rastreadas para determinar se o marcador foi integrado de modo estável no genoma da planta, determinando durante quanto tempo o gene marcador é expresso. Consequentemente, são desejadas células de planta que só transientemente expressem o gene marcador selecionável, porque representam células que capturaram, mas, não integraram em seus genomas o gene marcador selecionável.
[00143] Deste modo, cotransformando uma planta com esse “vetor marcador” e também com outro vetor que contenha o gene nativo ou polinucleotídeo pretendido, podese selecionar células de planta que capturaram ambos os vetores e, destes, determinar que células possuem genomas que contém apenas o gene ou polinucleotídeo pretendido. O “vetor marcador” pode ser modificado para reduzir mais a possibilidade de que o marcador seja integrado no genoma da planta. A presente invenção proporciona esses “vetores marcadores” na forma de vetores de “LifeSupport”.
[00144] A menos que de outra forma definidos, todos os termos técnicos e científicos aqui usados têm o mesmo significado conforme comumente entendido por uma pessoa de conhecimentos ordinários na técnica a que esta invenção pertence. Geralmente, a nomenclatura aqui usada e os procedimentos laboratoriais em culturas de células, na genética molecular e na química e hibridização de ácidos nucleicos aqui descritos são aqueles bem conhecidos e comumente empregados na técnica. As técnicas padrão são usadas para métodos de ácidos nucleicos recombinantes, síntese de polinucleotideos, culturas microbianas, culturas de células, culturas de tecidos, transformação, transfecção, transdução, química analítica, química orgânica sintética, sínteses químicas, análise química e formulação e suprimento farmacêuticos. Geralmente, as reações e purificação enzimáticos e/ou etapas de isolamento são realizados de acordo com a descrição dos fabricantes. As técnicas e procedimentos são geralmente praticados de acordo com a metodologia convencional descrita, por exemplo em “Molecular cloning: a laboratory manual”, 2a ed., Cold Spring Harbor Laboratory Press, NY (1989) e “Current protocols in molecular biology”, John Wiley & Sons, Baltimore, MD (1989).
[00145] Sequência de aminoácido: conforme aqui usado, inclui um oligopeptídeo, um peptídeo, um polipeptideo ou proteína e fragmentos dos mesmos, que são isolados, nativos ou de ocorrência natural numa planta ou são feitos sinteticamente, mas, compreendem a sequência de ácido nucléico da contrapartida endógena.
[00146] Manipulado artificialmente: conforme aqui usado, “manipulado artificialmente” significa mover, organizar, operar ou controlar com as mãos ou por meios mecânicos ou meios recombinantes, tais como por técnicas de engenharia genética, uma planta ou célula de planta, para produzir uma célula de planta ou planta que tenha um fenótipo e/ou genótipo biológico, bioquímico, morfológico ou fisiológico diferente em comparação com a contrapartida não manipulada, de ocorrência natural.
[00147] Propagação assexual: produção de progénie por geração de uma planta inteira a partir de recortes de folha, recortes de caule, recortes de raiz, olhos de tubérculo, estolhos, protoplastos únicos de células de planta, calo e similar, que não envolva fusão de gametas.
[00148] Esqueleto: a sequência de ácido nucléico de um vetor binário que exclui a sequência de T-DNA ou P-DNA pretendida para a transferência.
[00149] Borda e sequências semelhantes a borda: “sequências de borda” são sequências específicas derivadas de Agrobacterium. Tipicamente, uma sequência de borda esquerda e uma sequência de borda direita flanqueiam um T-DNA e ambas funcionam como sítios de reconhecimento para reações de incisão catalisadas por virD2. Essa atividade libera ácido nucléico que fica posicionado entre essas bordas. Ver Tabela 2 abaixo para exemplos de sequências de borda. O ácido nucléico liberado, complexado com virD2 e virE2, é direcionado para núcleos de células da planta onde o ácido nucléico é frequentemente integrado no genoma da célula da planta. Normalmente, duas sequências de borda, uma borda esquerda e uma borda direita, são usadas para integrar uma sequência de nucleotideos que fica localizada entre elas noutra sequência de nucleotideos. Ê também possível usar apenas uma borda ou mais de duas bordas, para realizar a integração de um ácido nucléico pretendido dessa maneira.
[00150] De acordo com a presente invenção, uma sequência “semelhante a borda” é isolada da espécie de planta selecionada que deve ser modificada ou de uma planta que seja sexualmente compatível com a espécie de planta a ser modificada e funciona como as sequências de borda de Agrobacterium. Isto é, uma sequência semelhante a borda da presente invenção promove e facilita a integração de um polinucleotídeo ao qual está ligado. Um DNA de planta, isto é, P-DNA da presente invenção contém preferentemente sequências semelhantes a borda.
[00151] Uma sequência semelhante a borda de um P-DNA está entre 5-100 bp de comprimento, 10-80 bp de comprimento, 15-75 bp de comprimento, 15-60 bp de comprimento, 15-50 bp de comprimento, 15- 40 bp de comprimento, 15-30 bp de comprimento, 16-30 bp de comprimento, 20-30 bp de comprimento, 21-30 bp de comprimento, 22- 30 bp de comprimento, 23-30 bp de comprimento, 24-30 bp de comprimento, 25-30 bp de comprimento ou 26-30 bp de comprimento.
[00152] As sequências semelhantes a borda da presente invenção podem ser isoladas a partir de qualquer planta, tal como de batata e trigo. Ver SEQ ID NO.: 1 e SEQ ID NO.: 34, para sequências que contêm, em qualquer término, as sequências semelhantes a borda isoladas da batata e do trigo, respectivamente. Deste modo, as sequências P-DNA de borda esquerda e direita de uso para a presente invenção são isoladas e/ou nativas para o genoma de uma planta que deva ser modificada. Uma sequência P-DNA semelhante a borda não é idêntica em sequência de nucleotideos a nenhuma sequência de borda de T-DNA derivada de Agrobacterium conhecida. Deste modo, uma sequência P-DNA semelhante a borda pode possuir 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 ou mais nucleotideos que são diferentes de uma sequência de borda de T-DNA de uma espécie de Agrobacterium, como Agrobacterium tumefaciens ou Agrobacterium rhizogenes. Isto é, uma borda de P-DNA ou uma sequência semelhante a borda da presente invenção tem, pelo menos, 95%, pelo menos, 90%, pelo menos 80%, pelo menos 75%, pelo menos 70%, pelo menos 60% ou pelo menos 50% de identidade de sequência com uma sequência de borda de T-DNA de uma espécie de Agrobacterium, tal como Agrobacterium tumefaciens ou Agrobacterium rhizogenes, mas, não 100% de identidade de sequência. Conforme aqui usados, os termos descritivos “borda de P-DNA” e “semelhante a borda de P-DNA” são permutáveis.
[00153] Uma sequência nativa de borda de P-DNA tem semelhança maior ou igual a 99%, 98%, 97%, 96%, 95%, 94%, 93%, 92%, 91%, 90%, 89%, 88%, 87%, 86%, 85%, 84%, 83%, 82%, 81%, 80%, 79%, 78%, 77%, 76%, 75%, 74%, 73%, 72%, 71%, 70%, 69%, 68%, 67%, 66%, 65%, 64%, 63%, 62%, 61%, 60%, 59%, 58%, 57%, 56%, 55%, 54%, 53%, 52%, 51% ou 50% em sequência de nucleotídeos com uma sequência de borda de T-DNA de Agrobacterium. Uma sequência semelhante a borda pode, então, ser isolada do genoma de uma planta e ser modificada ou mutacionada de modo a alterar a eficiência pela qual são capazes de integrar uma sequência de nucleotídeos noutra sequência de nucleotídeos. Outras sequências de polinucleotideos podem ser adicionadas ou incorporadas numa sequência semelhante a borda da presente invenção. Deste modo, uma borda esquerda de P- DNA ou uma borda direita de P-DNA pode ser modificada para possuir sítios múltiplos 5’ e 3’ de clonagem ou sítios de restrição adicionais. Uma sequência de borda de P-DNA pode ser modificada para aumentar a probabilidade de que o DNA de esqueleto do vetor acompanhante não seja integrado no genoma da planta.
[00154] A Tabela 2 abaixo representa as sequências de sequências de borda de T-DNA conhecidas e sequências aqui identificadas como “sequências semelhantes a borda”. Nenhuma das sequências identificadas como “semelhantes a borda” da Tabela 2 foram identificadas previamente como tendo uma estrutura de T-DNA semelhante a borda. As sequências semelhantes a borda da batata foram isoladas pelos métodos inventivos presentes usando primários degenerados em reações em cadeia de polimerase do DNA genômico da batata. A presente invenção engloba o uso de qualquer sequência semelhante a borda de P-DNA por transferência de um polinucleotídeo cojuntado no genoma de uma célula de planta.
[00155] Realmente, a presente invenção engloba qualquer sequência semelhante a borda que tenha a estrutura de sequência de ácido nucléico de SEQ ID NO.: 93: ANGATNTATN6GT (SEQ ID NO.: 93), onde “N” é qualquer nucleotideo, como aqueles representados por “A”,“G”, “C” ou “T. Esta sequência representa a sequência de consenso de ácidos nucleicos semelhantes a borda identificados pela presente invenção.Tabela 2 - Sequências e Sequências semelhantes a bordas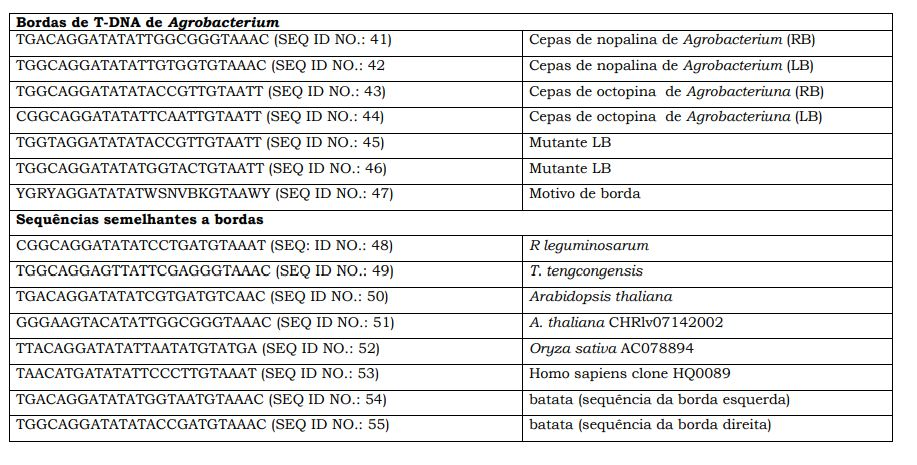 Y=C ou T; R=A ou G; K=G ou T; M=A ou C; W=A ou T; S=C ou G; V=A, C ou G; B=C, G ou T. Os números de acesso para as sequências semelhantes a borda são: sequência genômica do cromossomo 10 BAC OSJNBA0096G08 (AC078894. 11) da Oryza sativa; cromossomo 3 (NM_114337. 1) da Arabidopsis thaliana; cromossomo 1 da Arabidopsis thaliana 1 (NM_105664.1); cepa MB4T da T. tengcongensis, seção 118 de 244 do genoma completo (AE013091.1); clone HQ0089 (AF090888. do Homo sapiens 1); Clone rhiz98el2.qlk de Rhizobiunz:. *a batata partiu e foram obtidas e isoladas as sequências de borda esquerda e direita da batata de acordo com os métodos inventivos presentemente descritos.
Y=C ou T; R=A ou G; K=G ou T; M=A ou C; W=A ou T; S=C ou G; V=A, C ou G; B=C, G ou T. Os números de acesso para as sequências semelhantes a borda são: sequência genômica do cromossomo 10 BAC OSJNBA0096G08 (AC078894. 11) da Oryza sativa; cromossomo 3 (NM_114337. 1) da Arabidopsis thaliana; cromossomo 1 da Arabidopsis thaliana 1 (NM_105664.1); cepa MB4T da T. tengcongensis, seção 118 de 244 do genoma completo (AE013091.1); clone HQ0089 (AF090888. do Homo sapiens 1); Clone rhiz98el2.qlk de Rhizobiunz:. *a batata partiu e foram obtidas e isoladas as sequências de borda esquerda e direita da batata de acordo com os métodos inventivos presentemente descritos.
[00156] Portador DNA: um “portador de DNA” é um segmento de DNA que é usado para portar certos elementos genéticos e suprilos a uma célula da planta. Em transferência convencional de DNA estranho, este portador de DNA é frequentemente o T-DNA da Agrobacterium, delineado pelas sequências de borda. O portador de DNA aqui descrito é obtido da espécie de planta selecionada para ser modificada e contém extremidades que podem ser, estrutural e funcionalmente, diferentes das bordas T-DNA, mas, compartilha com esses T-DNAs a capacidade de sustentar ambas as transferência de DNA de Agrobacterium para os núcleos de células de planta ou certos outros eucariotas e a integração subsequente deste DNA nos genomas desses eucariotas.
[00157] Consistindo essencialmente em: uma composição que “consiste essencialmente em” certos elementos é limitada à inclusão daqueles elementos, como também aqueles elementos que não afetam materialmente as características básicas e novas da composição inventiva. Deste modo, enquanto a composição não afetar as características básicas e novas da presente invenção, isto é, não contiver DNA estranho que não seja da espécie de planta selecionada ou de uma planta que seja sexualmente compatível com a espécie de planta selecionada, então, aquela composição pode ser considerada um componente de uma composição inventiva que seja caracterizada pela linguagem de “consistindo essencialmente em”.
[00158] Primário degenerado: um “primário degenerado” é um oligonucleotídeo que contém tantas variações de nucleotideo quantas consegue acomodar nos desencontros de base, quando hibridizado em sequências de homologia semelhante, mas, não exata.
[00159] Dicotiledôneas: planta de florescência cujos embriões têm duas folhas de semente ou cotilédones. Os exemplos de dicotiledôneas incluem, mas, sem limitação, tabaco, tomate, batata, batata doce, cassava, legumes inclusive alfafa e soja, cenoura, morango, alface, carvalho, ácer, noz, rosa, hortelã, esprema, margarida e cactos.
[00160] Sequências reguladoras: referese àquelas sequências que são padrão e conhecidas dos especialistas na técnica, que podem ser incluídas nos vetores de expressão para aumentar e/ou maximizar a transcrição de um gene de interesse ou translação do RNA resultante num sistema de planta. Estes incluem, mas, sem limitação, promotores, sequências de sinal de exportação de peptídeo, introns, poliadenilação e sítios de terminação de transcrição. Os métodos para modificar construtos de ácido nucleico para aumentar os níveis de expressão em plantas são também geralmente conhecidos na técnica (ver, por exemplo, Rogers e colaboradores, 260 J. Biol. Chem. 3731-38, 1985; Cornejo e colaboradores, 23 Plant Mol. Biol. 567: 81, 1993). Ao engenhar um sistema de planta para afetar a taxa de transcrição de uma proteína, vários fatores conhecidos na técnica, inclusive sequências reguladoras tais como sequências de ação positiva ou negativa, intensificadores e silenciadores, como também a estrutura de cromatina, podem ter impacto. A presente invenção proporciona que, pelo menos, um destes fatores possa ser utilizado na engenharia de plantas para expressar uma proteína de interesse. As sequências reguladoras da presente invenção são elementos genéticos nativos, isto é, são isoladas da espécie de planta selecionada para ser modificada.
[00161] Estranhos: “estranho”, com respeito a um ácido nucleico, significa que esse ácido nucleico é derivado de organismos de nãoplanta ou derivado de uma planta que não é da mesma espécie que a planta a ser transformada ou não é derivada de uma planta que seja fértil com a planta a ser transformada, não pertence à espécie da planta visada.
[00162] De acordo com a presente invenção, o DNA ou RNA estranho representa ácidos nucleicos que estão ocorrendo naturalmente na maquilagem genética de fungos, bactéria, vírus, mamíferos, peixe ou pássaros, mas, não estão ocorrendo naturalmente na planta que deve ser transformada. Deste modo, um ácido nucleico estranho é um que codifica, por exemplo, um polipeptideo que não é naturalmente produzido pela planta transformada. Um ácido nucleico estranho não tem que codificar um produto de proteína. De acordo com a presente invenção, uma planta transgênica pretendida é uma que não contém nenhuns ácidos nucleicos estranhos integrados em seu genoma.
[00163] , Por outro lado, podem ser incorporados e integrados elementos genéticos nativos numa espécie de genoma da planta selecionada de acordo com a presente invenção. Os elementos genéticos nativos são isolados de plantas que pertencem à espécie de planta selecionada ou a partir de plantas que são sexualmente compatíveis com a espécie de planta selecionada. Por exemplo, o DNA nativo incorporado em batatas cultivadas (Solanum tuberosum) pode ser derivado de qualquer genótipo de S. tuberosum ou qualquer genótipo de uma espécie de batata selvagem que seja sexualmente compatível com S. tuberosum (por exemplo, S. demissum).
[00164] Gene: “gene” referese à região de codificação e não inclui sequência de nucleotídeos que sejam 5’ ou 3’ para aquela região. Um gene funcional é a região de codificação operavelmente ligada a um promotor ou terminador.
[00165] Reestruturação genética: referese à reassociação de elementos genéticos que pode ocorrer espontaneamente in vivo como também in vitro, que introduz uma nova organização do material genético. Por exemplo, a junção de polinucleotídeos em diferentes locais cromossômicos pode acontecer espontaneamente in vivo durante tanto o desenvolvimento da planta quanto por recombinação sexual. Consequentemente, a recommbinação in vitro de elementos genéticos por técnicas de modificação genética nãonaturais é similar para eventos de recombinação que podem ocorrer também através da recombinação sexual in vivo.
[00166] Em moldura: tripletes de nucleotideos (códons) são traduzidos para uma sequência de aminoácidos nascente da proteína recombinante pretendida numa célula de planta. Especificamente, a presente invenção contempla um primeiro ácido nucleico ligado em moldura de leitura a um segundo ácido nucleico em que a primeira sequência de nucleotideos é um gene e o segundo nucleotídeo é um promotor ou elemento regulador semelhante.
[00167] Integrar: referese à inserção de uma sequência de ácido nucleico de uma espécie da planta selecionada ou de uma planta que seja da mesma espécie que a planta selecionada ou de uma planta que seja sexualmente compatível com a espécie de planta selecionada, no genoma de uma célula de uma espécie da planta selecionada. A “integração” referese à incorporação apenas de elementos nativos genéticos num genoma da célula da planta. A fim de integrar um elemento genético nativo, tal como por recombinação homóloga, a presente invenção pode “usar” DNA nãonativo como uma etapa nesse processo. Deste modo, a presente invenção distingue entre o “uso de” uma molécula particular de DNA e a “integração” de uma molécula particular de DNA num genoma da célula da planta.
[00168] Introdução: conforme aqui usada, referese à inserção de uma sequência de ácido nucleico numa célula, por métodos inclusive infecção, transfecção, transformação ou transdução.
[00169] Isolado: “isolado” referese a qualquer ácido nucleico ou composto que esteja fisicamente separado de seu ambiente normal, nativo. O material isolado pode ser mantido numa solução adequada contendo, por exemplo, um solvente, um tampão, um íon ou outro componente e pode estar na forma purificada ou não purificada.
[00170] Líder: sequência transcrita, mas, não traduzida precedendo (ou 5’ para) um gene.
[00171] Vetor de LifeSupport: um vetor de LifeSupport é um construto que contém um gene marcador selecionável expressável como um marcador de neomicina fosfotransferase, que fica posicionado entre bordas T-DNA ou semelhantes a T-DNA. O vetor de LifeSupport pode ser modificado para limitar a integração desse marcador, como também outros polinucleotídeos, que ficam situados entre a borda ou sequências semelhantes a borda num genoma da planta. Por exemplo, um vetor de LifeSupport pode incluir um virD2 mutante, uma fusão codA::upp ou qualquer combinação desses elementos genéticos mutacionados. Deste modo, uma proteína modificada virD2 suportará a transferência de T- DNA para núcleos da planta, mas, limitará a eficiência de uma integração genômica subsequente de T-DNAs (Shurvinton e colaboradores, Proc. Natl. Acad. Sei. USA, 89: 11837-11841, 1992; Mysore e colaboradores, Mol. Plant Microbe Interact., 11: 668-683, 1998). Alternativamente, a fusão de genes codA::upp pode ser usada como marcador selecionável negativo anterior à regeneração. Num construto preferido, o vetor de LifeSupport inclui o marcador npt operavelmente ligado ao elemento terminador ADH de levedura.
[00172] Monocotiledônea: planta de florescência cujos embriões têm um cotilédone ou folha de semente. Os exemplos de monocotiledôneas incluem, mas, sem limitação, a grama de relva, milho, arroz, aveia, trigo, cevada, sorgo, orquídea, íris, lírio, cebola e palma.
[00173] Nativo: um elemento genético “nativo” referese a um ácido nucléico que existe naturalmente, originase ou pertence ao genoma de uma planta que deve ser transformada. Deste modo, qualquer ácido nucléico, gene, polinucleotídeo, DNA, RNA, mRNA ou molécula de cNA que é isolada do genoma de uma planta ou espécie de planta que deve ser transformada ou é isolada de uma planta ou espécie que seja sexualmente compatível ou fértil com a espécie de planta que deve ser transformada, é “nativa”, isto é, indígena para a espécie de planta. Noutras palavras, um elemento genético nativo representa todo o material genético que está acessível para criadores de plantas para a melhoria de plantas através da procriação clássica de plantas. Quaisquer variantes de um ácido nucleico nativo são também consideradas “nativas”, conforme a presente invenção. A este respeito, um ácido nucleico “nativo” pode ser também isolado de uma planta ou espécie sexualmente compatível com a mesma e modificado ou mutacionado de forma que a variante resultante seja maior ou igual a 99%, 98%, 97%, 96%, 95%, 94%, 93%, 92%, 91%, 90%, 89%, 88%, 87%, 86%, 85%, 84%, 83%, 82%, 81%, 80%, 79%, 78%, 77%, 76%, 75%, 74%, 73%, 72%, 71%, 70%, 69%, 68%, 67%, 66%, 65%, 64%, 63%, 62%, 61%, ou 60% de semelhança com a sequência de nucleotideos para o ácido nucleico inalterado, nativo, isolado de uma planta. Uma variante nativa de ácido nucleico pode ser também menos do que 60%, menos do que cerca de 55% ou menos do que cerca de 50% semelhante em sequência de nucleotideos.
[00174] Um ácido nucleico “nativo” isolado de uma planta pode codificar também uma variante do produto de proteína de ocorrência natural transcrito e transladado daquele ácido nucleico. Deste modo, um ácido nucleico nativo pode codificar uma proteína que seja maior ou igual a 99%, 98%, 97%, 96%, 95%, 94%, 93%, 92%, 91%, 90%, 89%, 88%, 87%, 86%, 85%, 84%, 83%, 82%, 81%, 80%, 79%, 78%, 77%, 76%, 75%, 74%, 73%, 72%, 71%, 70%, 69%, 68%, 67%, 66%, 65%, 64%, 63%, 62%, 61%, ou 60% de semelhança em sequência de aminoácidos com a proteína inalterada, nativa, expressa na planta de que o ácido nucleico foi isolado.
[00175] Ácido nucleico de ocorrência natural: esta frase significa que o ácido nucleico é achado dentro do genoma de uma espécie de planta selecionada e pode ser uma molécula de DNA ou uma molécula de RNA. A sequência de um sítio de restrição que está normalmente presente no genoma de uma espécie de planta pode ser engenhada numa molécula de DNA exógena, tal como um vetor ou oligonucleotídeo, embora aquele sítio de restrição não estivesse fisicamente isolado daquele genoma. Deste modo, a presente invenção permite a criação sintética de uma sequência de nucleotídeos, tal como uma sequência de reconhecimento de enzima de restrição, enquanto aquela sequência estiver ocorrendo naturalmente no genoma da espécie de planta selecionada ou numa planta que seja sexualmente compatível com a espécie de planta selecionada que deve ser transformada.
[00176] Operavelmente ligada: combinando duas ou mais moléculas de tal modo que, em combinação, elas funcionam corretamente numa célula de planta. Por exemplo, um promotor está operavelmente ligado a um gene estrutural, quando o promotor controlar a transcrição do gene estrutural.
[00177] P-DNA: de acordo com a presente invenção, P-DNA (“DNA de planta”) é isolado do genoma da planta e compreende, em cada extremidade ou apenas numa extremidade, uma sequência T-DNA semelhante a borda. A sequência semelhante a borda compartilha preferentemente pelo menos 50%, pelo menos 60%, pelo menos 70%, pelo menos 75%, pelo menos 80%, pelo menos 90% ou pelo menos 95%, mas, menos de 100% de identidade de sequência, com uma sequência de borda de T-DNA de uma espécie de Agrobacterium, como Agrobacterium tumefaciens ou Agrobacterium rhizogenes. Deste modo, podem ser usados P-DNAs em vez de T-DNAs para transferir uma sequência de nucleotídeos de Agrobacterium para outra sequência de polinucleotideos. O P-DNA pode ser modificado para facilitar a clonagem e deve, de preferência, não codificar naturalmente proteínas ou partes de proteínas. O P-DNA é caracterizado por que contém, em cada extremidade, pelo menos, uma sequência de borda, referida quer como uma “sequência de borda de P-DNA” quer como “uma sequência semelhante a borda de P-DNA”, que são termos intercambiáveis. Ver a definição, acima, de uma “sequência de borda” e “semelhante a borda”. Um P-DNA pode ser também considerado como uma sequência ’’semelhante a T-DNA”; ver definição abaixo.
[00178] Planta: inclui angiospermas e gimnospermas como batata, tomate, tabaco, alfafa, alface, cenoura, morango, beterraba, cassava, batata doce, soja, milho, grama de relva, trigo, arroz, cevada, sorgo, aveia, carvalho eucalipto, noz e palma. Deste modo, uma planta pode ser uma monocotiledônea ou uma dicotiledônea. A palavra “planta”, conforme aqui usada, engloba também células de planta, semente, progénie de planta, propagula quer gerada sexualmente quer assexualmente e descendentes de qualquer destes, tais como recortes ou semente. As células de plantas incluem culturas de suspensão, calos, embriões, regiões meristemáticas, tecido de calos, folhas, raízes, brotos, gametófitos, esporófitos, pólen, sementes e microsporos. As plantas podem estar em vários estágios de maturidade e podem ser cultivadas em cultura líquida ou sólida ou no solo ou em meios adequados em panelas, estufas ou campos. A expressão de um líder, arrasto ou sequências de genes introduzidos em plantas pode ser transiente ou permanente. Uma “espécie de planta selecionada” pode ser, mas sem limitação, uma espécie de qualquer uma destas “plantas.”
[00179] Procriação precisa: referese ao melhoramento de plantas por introdução estável de ácidos nucleicos, como genes nativos e elementos reguladores isolados da espécie de planta selecionada ou de outra planta na mesma espécie da planta selecionada ou de espécies que são sexualmente compatíveis com a espécie de planta selecionada em células de planta individuais e subsequente regeneração destas células de planta geneticamente modificadas em plantas inteiras. Visto que nenhum ácido nucléico desconhecido ou estranho fica permanentemente incorporado no genoma da planta, a tecnologia inventiva faz uso do mesmo material genético que está também acessível através da procriação de planta convencional.
[00180] Espécie de planta: o grupo de plantas que pertence a várias espécies de planta oficialmente designadas que exibem, pelo menos, alguma compatibilidade sexual.
[00181] Transformação de planta e cultura de célula: referese no geral ao processo pelo qual as células da planta são geneticamente modificadas e transferidas para um meio de cultura de plantas apropriado para a manutenção, crescimento adicional e/ou desenvolvimento adicional.
[00182] Recombinante: conforme aqui usado, descreve em geral várias tecnologias por meio das quais os genes podem ser clonados, o DNA pode ser sequenciado e podem ser produzidos produtos de proteína. Conforme aqui usado, o termo também descreve proteínas que têm sido produzidas a seguir à transferência de genes para as células de sistemas hospedeiros de plantas.
[00183] Marcador selecionável: um “marcador selecionável” é tipicamente um gene que codifica uma proteína que confere algum tipo de resistência a um antibiótico, herbicida ou composto tóxico e é usado para identificar eventos de transformação. Os exemplos de marcadores selecionáveis incluem o gene da estreptomicina fosfotransferase (spt) que codifica a resistência à estreptomicina, o gene da fosfomanose isomerase (pmi) que converte fosfatos de manose-6 em fosfato de frutose-6; o gene da neomicina fosfotransferase (nptZZ) que codifica a resistência à canamicina e geneticina, o gene da higromicina fosfotransferase (hpt ou aphiv) que codifica a resistência à higromicina, genes da acetolactato sintase (ais) que codificam a resistência a herbicidas do tipo sulfonilurea, genes que codificam a resistência a herbicidas que atuam de forma a inibir a ação da glutamina sintase como fosfinotricina ou basta (por exemplo, o gene bar) ou outros genes semelhantes conhecidos na técnica.
[00184] Supressão de sentido: redução na expressão de um gene endógeno por expressão de uma ou mais umas cópias adicionais de todo ou parte daquele gene em plantas transgênicas.
[00185] Semelhante a T-DNA: uma sequência “semelhante a T- DNA” é um ácido nucleico que é isolado de uma espécie de planta selecionada ou de uma planta que seja sexualmente compatível com a espécie de planta selecionada e que compartilha pelo menos 75%, 80%, 85%, 90%, ou 95%, mas, não 100% de identidade de sequência com o T-DNA da espécie de Agrobacterium. A sequência semelhante a T-DNA pode conter uma ou mais bordas ou sequências semelhantes a borda que são, cada uma, capazes de integrar uma sequência de nucleotídeos noutro polinucleotídeo. Um “P-DNA”, conforme aqui usado, é um exemplo de uma sequência semelhante a T-DNA.
[00186] Arrasto: sequência transcrita, mas, não transladada seguinte (ou 3’ para) a um gene.
[00187] DNA transcrito: DNA que compreende tanto um gene como o líder não transladado e a sequência de arrasto que estão associados com aquele gene, que é transcrito como um mRNA único pela ação do promotor precedente.
[00188] Terminadores de transcrição e translação: os vetores de expressão da presente invenção têm tipicamente uma região de terminação de transcrição na extremidade oposta da região reguladora da iniciação de transcrição. A região de terminação de transcrição pode ser selecionada, para estabilidade do mRNA para intensificar a expressão e/ou para a adição de caudas de poliadenilação adicionadas ao produto de transcrição de gene (Alber & Kawasaki, Mol. Appl. Genetics 4: 19-34, 1982). Regiões de terminação de transcrição ilustrativas incluem a sequência E9 do gene da ervilha RBCS (Mogen e colaboradores, Mol. Cell. Biol., 12: 5406-14, 1992) e os sinais de terminação de vários genes ubiquitin.
[00189] Transformação de células de planta: um processo pelo qual o DNA é integrado de modo estável no genoma de uma célula de planta. “De modo estável” referese à retenção e/ou expressão permanente ou não transiente de um polinucleotídeo e por um genoma de célula. Deste modo, um polinucleotídeo integrado de modo estável é um que é um acessório dentro de um genoma de célula transformada e pode ser replicado e propagado através da progénie sucessiva da célula ou planta transformada resultante. A transformação pode acontecer sob condições naturais ou artificiais usando vários métodos bem conhecidos na técnica. A transformação pode contar com qualquer método conhecido para a inserção de sequências de ácidos nucleicos numa célula hospedeira procariota ou eucariota, inclusive protocolos de transformação mediados por Agrobacterium, infecção viral, whiskers, eletroporação, choque de calor, lipofecção, tratamento com polietileno glicol, microinjeção e bombardeio de partículas.
[00190] Transgene: um gene que será inserido num genoma hospedeiro, incluindo uma região de codificação de proteína. No contexto da presente invenção, os elementos que compreendem o transgene são isolados do genoma hospedeiro.
[00191] Planta transgênica: uma planta geneticamente modificada que contém, pelo menos, um transgene.
[00192] Usando/De uso: a presente invenção encara o uso de ácido nucléico de espécie diferente daquela da espécie de planta selecionada para ser transformada para facilitar a integração de elementos genéticos nativos no genoma da planta selecionada, enquanto esse ácido nucléico estranho não é integrado de modo estável no mesmo genoma da planta hospedeira. Por exemplo, o plasmideo, vetor ou construto de clonagem em que são clonados, posicionados ou manipulados elementos genéticos nativos pode ser derivado de uma espécie diferente daquela de que os elementos genéticos nativos foram derivados.
[00193] Variante: uma “variante”, conforme aqui usada, é entendida significar um nucleotideo ou sequência de aminoácidos que se desvia da sequência padrão ou dada, nucleotideo ou aminoácido de um gene ou proteína particulares. Os termos “isoforme”, “isótipo” e “análogo” referemse também a “formas variantes de um nucleotideo ou uma sequência de aminoácidos. Uma sequência de aminoácidos que é alterada pela adição, remoção ou substituição de um ou mais aminoácidos ou uma mudança na sequência de nucleotideos pode ser considerada uma sequência “variante”. A variante pode ter mudanças “conservadoras”, em que um aminoácido substituído tem propriedades estruturais ou químicas semelhantes, por exemplo, substituição de leucina por isoleucina. Uma variante pode ter mudanças “não conservativas”, por exemplo, substituição de uma glicina por um triptofano. As variações secundárias análogas podem incluir também deleções de aminoácido ou inserções ou ambas. A orientação na determinação de quais resíduos de aminoácidos podem ser substituídos, inseridos ou deletados pode ser achada usando programas de computador bem conhecidos na técnica como o software Vetor NTI Suite (InforMax, MD).
[00194] Fica entendido que a presente invenção não está limitada à metodologia, protocolos, vetores e reativos etc. particulares aqui descritos, conforme possam variar. Fica também entendido que a terminologia aqui usada é usada com a finalidade de descrever modalidades particulares somente e não se pretende que limite o âmbito da presente invenção. Deve ser notado que, conforme aqui usado e nas reivindicações anexadas, as formas singulares “um”, “uma”, e “o” incluem a referência plural, a menos que o contexto dite claramente de outra forma. Deste modo, por exemplo, uma referência a “um gene” é uma referência a um ou mais genes e inclui equivalentes dos mesmos conhecidos dos especialistas na técnica e assim por diante. Realmente, um especialista na técnica pode usar os métodos aqui descritos para expressar qualquer gene nativo (conhecido presentemente ou subsequentemente) em sistemas hospedeiros de plantas.
[00195] Métodos de transformação mediados por Agrobacterium são os meios preferidos para incorporar o DNA recombinado em células de plantas De acordo com a presente invenção, foi desenvolvido um vetor binário para produzir plantas de batata modificadas geneticamente que contém apenas ácidos nucleicos de batata nativa. Esse vetor é diferente dos vetores convencionais de transformação mediada por Agrobacterium em três formas: (1) em vez de uma sequência de T-DNA derivada de Agrobacterium delineada por bordas de T-DNA, o vetor presente contém um fragmento nativo (P-DNA) de DNA da planta que é flanqueado por sequências semelhantes a borda, que suportam a transferência de P- DNA de Agrobacterium para células de plantas, embora sejam estrutural e funcionalmente diferentes de bordas de T-DNA, (2) o esqueleto do vetor presente pode conter um marcador que, se integrado no genoma da célula de planta, impede estas células de se desenvolverem em plantas maduras e (3) o vetor presente não contém um gene marcador selecionável estranho entre terminais P-DNA.
[00196] A presente invenção demonstra, surpreendentemente, que os fragmentos de P-DNA flanqueados por sequências semelhantes a borda suportam a transferência de DNA de Agrobacterium para células da planta. O P-DNA pode ser isolado do genoma de qualquer planta usando primários que são projetados em base de homologia entre os terminais de um P-DNA de batata e bordas convencionais de T-DNA. Esses fragmentos podem, então, ser testados e, se eficazes, usados para transformar aquela planta com DNA exclusivamente nativo. Ê também possível procurar bases de dados genômicos de plantas para fragmentos de DNA com regiões que mostram homologia com bordas de T-DNA usando programas tais como ‘blastn’; (Altschul e colaboradores, J. Mol. Biol., 215: 403-10, 1990). Os P-DNAs identificados podem, então, ser modificados para aumentar a sua utilidade. Por exemplo, fragmentos internos dos P-DNAs isolados podem ser deletados e podem ser adicionados sítios de restrição para facilitar a clonagem. Podem ser também eficazes para introduzir mutações pontuais nas sequências terminais para tornar o P-DNA mais efetivo na transferência de DNA.
[00197] Qualquer cassete de expressão de gene pode ser inserido entre sequências P-DNA semelhantes a bordas. Para transformações de batata, esse cassete de expressão poderia consistir num promotor de batata, operavelmente ligado a um gene de batata e/ou uma sequência líder ou de arrasto associada àquele gene e seguida de um terminador de batata. O cassete de expressão pode conter elementos genéticos adicionais da batata tal como uma sequência peptídea de sinal fundida em moldura para o terminal 5’ do gene e um intron da batata que podería, por exemplo, ser colocado entre o promotor e o gene de interesse para aumentar a expressão. Para a transformação de trigo com um P-DNA modificado, todos os elementos genéticos que são inseridos no P-DNA do trigo, inclusive o P-DNA propriamente, seriam derivados de espécies de trigo ou planta que sejam sexualmente compatíveis com o trigo.
[00198] Outro caminho para isolar P-DNAs é gerar uma biblioteca de cepas de Agrobacterium que contenham fragmentos aleatórios de DNA da planta em vez de um T-DNA flanqueando um gene marcador selecionável. Explantes infectados com esta biblioteca podem ser colocados em meio de proliferação que contém um agente selecionável apropriado para identificar P-DNAs que suportem a transferência do gene marcador do vetor em Agrobacterium para a célula da planta.
[00199] Ê possível que não apenas o P-DNA modificado nativo, mas, também sequências de plasmídeos adicionais sejam cotransferidas de Agrobacterium para a célula da planta durante o processo de transformação. Para os propósitos da presente invenção, isto é um processo indesejável porque essas sequências de “esqueleto” de plasmídeos representam DNA estranho de nãoplanta, tal como DNA bacteriano. A presente invenção impede que células de plantas transformadas que contêm sequências de esqueleto se desenvolvam em plantas maduras. Deste modo, a presente invenção torna possível distinguir eventos de transformação contendo esqueleto e isentos de esqueleto durante a fase de broto regenerado.
[00200] O método para selecionar ou rastrear eventos de integração de esqueleto conta com a presença de um cassete de expressão para um marcador, tal como o gene da isopentenil fosfotransferase (IPT), no esqueleto do vetor, fora do P-DNA. Após integração do esqueleto, a acumulação de citocinina induzida por IPT alterará a forma dos brotos transformados e impedirá que estes brotos desenvolvam raízes. Em vez do gene de IPT, pode ser usado qualquer outro gene que altere a forma, a textura ou a cor das folhas, raízes, talo, altura ou alguma outra característica morfológica da planta transformada, para rastrear e/ou selecionar eventos de integração de esqueleto. Esse gene é aqui referido como “marcador de integração de esqueleto”. Deste modo, a planta transformada que exiba uma característica morfológica alterada atribuível à expressão do gene marcador de integração de esqueleto é sabida conter em seu genoma DNA estranho além do desejado P-DNA. Consequentemente, não são desejadas plantas que exibem um fenótipo associado com o marcador de integração de esqueleto.
[00201] A presente invenção não está limitada ao uso de apenas um gene de IPT como marcador de integração de esqueleto; outros genes podem ser usados dessa maneira. Por exemplo, um marcador de integração de esqueleto pode ser um gene da sintase da transzeatine (TZS) do Agrobacterium (Krall e colaboradores, FEBS Lett 527: 315-8, 2002) ou um gene recessivo de Arabidopsis hocl (Catterou e colaboradores, Plant J, 30: 273-87, 2002). Este método pode ser aplicado mais facilmente para uso na presente invenção do que alguns métodos que inserem genes tóxicos em sequências de esqueleto de vetor. Ver, por exemplo, EP 1.009.842.
[00202] Posicionando um gene marcador de integração de esqueleto, tal como um gene funcional da citocinina a montante ou a jusante do P- DNA, é imediato distinguir entre eventos de transformação. As plantas transformadas que exibem uma característica morfológica alterada são descartadas, porque contêm sequências de DNA estranhas integradas no genoma.
[00203] Outra estratégia para identificar plantas que são transformadas de modo estável apenas com DNA nativo é empregar a reação em cadeia da polimerase. Usando primários que são especificamente projetados para detectar sequências de esqueleto, podem ser identificadas e descartadas as plantas que contenham sequências de esqueleto estranhas, além do P-DNA. Outros conjuntos primários podem ser subsequentemente usados para confirmar a transferência intacta do P-DNA. Deste modo, quer usando a expressão de um gene para mudar uma característica morfológica de uma planta quer por rastreamento quanto a DNA estranho integrado de modo estável numa planta transformada, podem ser identificadas e selecionadas plantas transformadas de modo estável apenas com sequências de DNA nativas.
[00204] Os elementos genéticos de uma planta hospedeira particular podem ser inseridos na sequência de P-DNA de um vetor binário capaz de replicação tanto em E. coli como Agrobacterium. A introdução dos vetores resultantes em cepas desarmadas de Agrobacterium tais como LBA4404 pode ser realizada através de eletroporação, acasalamento triparental ou tratamento de choque térmico de células quimicamente competentes. As novas cepas podem, então, ser usadas para transformar células de planta individuais através da infecção de plantas inteiras ou explantes.
[00205] Os elementos genéticos de uma planta hospedeira particular podem ser inseridos na sequência de P-DNA de um vetor binário capaz de replicação tanto em E. coli como em Agrobacterium. A introdução dos vetores resultantes em cepas de Agrobacterium, tais como LBA4404, pode ser realizada através de eletroporação, acasalamento triparental ou tratamento de choque térmico de células quimicamente competentes. As novas cepas podem, então, ser usadas para transformar células de plantas individuais através da infecção de plantas inteiras ou explantes. LBA4404 contém o plasmideo Ti desarmado pAL4404, que porta as funções de virulência e um gene de resistência à estreptomicina.
[00206] Embora a integração estável de genes de marcadores bacterianos no genoma de células de planta facilite a identificação de eventos de transformação, essas modificações do genoma das plantas são indesejáveis, porque os genes de marcadores representam DNA estranhos. O uso de um gene marcador estranho pode ser evitado por desenvolvimento de novos métodos de transformação baseados em Agrobacterium.
[00207] Uma modalidade preferida é um método novo que conta com o uso de duas cepas de Agrobacterium: uma cepa contendo um vetor binário com um gene marcador selecionável pretendido para expressão transiente em núcleos de planta e outra cepa que porta o P- DNA com as sequências reais de interesse pretendidas para integração estável no genoma da planta (ver Exemplo 7).
[00208] Após coinfecção com as cepas de Agrobacterium, algumas células de planta receberão tanto um T-DNA com o gene marcador como um P-DNA com as sequências de interesse. Em vez de selecionar subsequentemente para a integração estável do gene marcador sujeitando os explantes infectados ao antibiótico apropriado por um longo período de tempo, os explantes são apenas brevemente expostos ao antibiótico. Deste modo, todas as células da planta que expressam transientemente o gene marcador sobreviverão. Como os T-DNAs na maioria dos casos degradamse devido a atividades de nuclease endógena em lugar de integrarse de modo estável ao genoma do seu hospedeiro, a maioria das células da planta que sobreviveram à seleção transiente são aqui mostradas desenvolveremse em brotos carentes de gene marcador. A presente invenção, além disso, demonstra que uma proporção significativa destes brotos isentos de marcador contém P- DNAs integrados de modo estável.
[00209] Existem várias ferramentas para aumentar a eficiência de transformação isenta de marcador. Primeira, a presente invenção demonstra que esta frequência pode ser aumentada infectando consecutivamente explantes com duas cepas de Agrobacterium que portam T-DNA/marcador e P-DNA/sequências de interesse, respectivamente. Os explantes são, primeiramente, infectados com a cepa de P-DNA e, depois de mais ou menos 4 a 6 horas, com a cepa de T-DNA.
[00210] Segunda, a cepa de T-DNA pode ser modificada para expressar um gene ómegamutacionado virD2. A proteína modificada virD2 suportará a transferência de T-DNA para núcleos de plantas, mas, limita a eficiência de uma integração genômica subsequente de T- DNAs (Shurvinton e colaboradores, Proc. Natl. Acad. Sci. USA, 89: 11837-11841, 1992; Mysore e colaboradores, Mol. Plant Microbe Interact, 11: 668-683, 1998). O método de maior preferência para expressar um gene modificado virD2 é inserir um gene ómegamutacionado virD2 dirigido pelo promotor virD no esqueleto do vetor de T-DNA.
[00211] Terceira, a integração estável de T-DNA pode ser ainda prejudicada inserindo sequências telomere perto das sequências da borda esquerda e direita do T-DNA (Chiurazzi & Signer, Plant Mol. Biol., 26: 923-934, 1994).
[00212] Quarta, o tamanho da região de T-DNA que porta o gene marcador pode ser aumentado para aumentar a frequência de T-DNAs e P DNAs que se movem juntos para o núcleo da célula de planta e para reduzir a frequência da integração genômica do T-DNA.
[00213] Quinta, a frequência de T-DNAs e P DNAs que se deslocam em conjunto para o núcleo da célula da planta pode ser também aumentada usando uma cepa única de Agrobacterium que porta dois vetores binários compatíveis com o T-DNA e P-DNA, respectivamente. Um exemplo de dois vetores binários compatíveis são um vetor derivado pSIM 1301 e vetor derivado pBI121.
[00214] Como o gene marcador transientemente expresso não se integrará normalmente no genoma da planta, não é necessário que tanto este gene quanto as suas sequências reguladoras representem DNA nativo. De fato, pode ser vantajoso usar sequências reguladoras estranhas para promover níveis altos de expressão transiente de gene nas células infectadas da planta. Uma constatação surpreendente da presente invenção é que um cassete de expressão contendo o gene de GUS seguido do terminador do álcool da dehidrogenase 1 da levedura (ADHL) foi transientemente expresso em níveis altos em células da batata. Um construto semelhante com o terminador CYCI da levedura, porém, não funcionou adequadamente. Pode ser também possível aumentar níveis de expressão transiente ligando operavelmente um gene marcador a um promotor não nativo. Os exemplos desses promotores são, entre outros, promotores sintéticos, tais como os promotores induzíveis por glucocorticóide (Mori e colaboradores, Plant J., 27:79-86, 2001; Bohner e colaboradores, Mol. Gen. Genet., 264: 860- 70, 2001) e promotores não nativos tais como os promotores 35S do vírus do mosaico da couveflor e vírus do mosaico da escrofulária e promotores de fungos.
[00215] Como alternativa para a abordagem de transformação mediada pelas duas cepas de Agrobacterium acima descritas, as plantas podem ser também transformadas com uma cepa única que contém um P-DNA com tanto um gene marcador nativo como as sequências reais de interesse. A presente invenção demonstra que é possível usar genes de tolerância ao sal como marcadores nativos para transformação. Esses genes de tolerância a sal incluem homólogos de colheita dos genes S0S1 da Arabidopsis (Shi e colaboradores, Nat. Biotechnol. 2002), ATNHXI (Apse e colaboradores, Science, 285: 1256-8, 1999), Avpl (Gaxiola e colaboradores, Proc Natl Acad Sei USA 98:11444-9, 2001) e CBF3 (Kasuga e colaboradores, Nat. Biotechnol. 17: 287-91, 1999).
[00216] Os rearranjos de elementos genéticos realizados através da metodologia inventiva da Procriação Precisa poderiam ocorrer também espontaneamente através do processo da recombinação genética. Por exemplo, todas as plantas contêm elementos que podem transpor de uma localização cromossômica para outra. Pela inserção em promotores ou genes, esses elementos transponíveis podem aumentar, alterar e/ou reduzir a expressão do gene. Por exemplo, a inserção do AMu4 do elemento Mutator do milho no promotor do gene regulador transcricional Pwr ocasiona pericarpos vermelhos em tiras. A inserção do mesmo elemento no promotor do gene MADSbox específico da folha ZMM19 resultou na expressão deste gene nas inflorescências do milho, causando um alongamento foliáceo das glumas e outras mudanças nas inflorescências macho e fêmea, resultando no famoso fenótipo do milho pod (vagem). Por causa de suas borlas e orelhas estranhas, o milho pod era de significado religioso para certas tribos nativas americanas. Muitos genes são também reorganizados através de outras modificações induzidas por transposon como inversões, deleções, adições e recombinações ectópicas (Bennetzen, Plant Mol Biol 42: 251-69, 2000). Além disso, os rearranjos do DNA da planta ocorrem frequentemente através do processo da recombinação intragênica. Por exemplo, recombinando genes envolvidos na resistência contra patógenos específicos, as plantas são capazes de desenvolver genes de resistência com novas especificidades e, deste modo, coevoluir com seus patógenos (Ellis e colaboradores, Trends Plant Sei 5: 373-9, 2000). Outro exemplo de recombinação intragênica diz respeito a como as plantas se reproduzem: a transição de plantas de fertilização cruzada para autofertilização por recombinação de genes envolvidos em autoincompatibilidade (Kusaba e colaboradores, Plant Cell 13: 627-43, 2001). Outros processos que promovem a evolução do genoma incluem, por exemplo, a clivagem do cromossomo e a recombinação intercromossômica.
[00217] Para modificar características negativas tais como a acumulação de acrilamida durante o processamento, a acumulação de glicoalcalóides, a acumulação de produtos de glicação avançada indesejáveis, a acumulação de CIPC, os baixos níveis de amido resistente, a suscetibilidade à contusão, o adocicamento induzido pelo frio, a suscetibilidade à doença, o baixo rendimento e a baixa qualidade em plantas de colheita através da procriação precisa, é incorporado pelo menos um cassete de expressão específico num genoma do hospedeiro. São usados três métodos diferentes para eliminar as características negativas: (1) a sobrexpressão de genes que impedem a ocorrência de características negativas, (2) a sobrexpressão de versões mutacionadas de genes associados com características negativas, a fim de neutralizar os produtos do gene do tipo selvagem com proteínas não funcionais e (3) o silenciamento de genes específicos que estão associados com uma característica negativa expressando pelo menos uma cópia de um fragmento líder ou de arrasto associado com aquele gene na orientação de sentido e/ou de antisentido.
[00218] Um exemplo de um gene endógeno que está associado com uma característica negativa na batata e pode ser modificado in intro de forma que codifique uma proteína não funcional é o gene da polifenol oxidase (PPO). Após lesão de impacto, o produto do gene de PPO é liberado do plastídeo no citoplasma (Koussevitzky e colaboradores, J. Biol. Chem., 273: 27064-9, 1998), onde ele mediará a oxidação de fenóis de modo a criar uma variedade de radicais fenoxil e derivados quinóides, que são tóxicos e/ ou, em última instância, formam polímeros indesejáveis que deixam descolorações escuras ou “manchas pretas” na colheita.
[00219] A sobrexpressão de um gene mutante PPO que contém um domínio de ligação a cobre não funcional consegue abaixar a atividade de todos os genes PPO que são principalmente expressos em tubérculos e órgãos associados, tais como brotos. As mutações tornam a proteina da polifenol oxidase inativa, porque fica impossibilitada de se ligar ao cobre. O especialista sabería onde fazer mutações pontuais que comprometeríam, neste caso, a função de um produto de gene. Os requerentes identificaram o que domínio de ligação ao cobre no PPO da batata alinhando a sequência da proteina PPO da batata com uma sequência de proteina PPO da batata doce (Klabunde e colaboradores, Nat. Struct. Biol., 5:1084-90, 1998). Áreas de conservação, particularmente aquelas que contêm resíduos conservados de histidina em sítios de ligação a cobre, foram alvo de inativação do produto transgene. Dado que a quase completa ausência de atividade PPO nesses órgãos pode impactar negativamente a capacidade da planta de resistir a patógenos, a presente invenção descreve também um método aperfeiçoado de baixar apenas um gene PPO específico que é predominantemente expresso em todas as partes do tubérculo maduro, com exceção da epiderme. O silenciamento deste gene PPO específico pelo uso de uma sequência de arrasto associada com esse gene não reduz a expressão de PPO na epiderme do tubérculo, a parte do tubérculo que é mais diretamente exposta a patógenos que tentam infectar.
[00220] O escurecimento enzimático induzido pelo gene PPO não só reduz a qualidade dos tubérculos da batata; também afeta negativamente alimentos de colheitas tais como o trigo, o abacate, a banana, a alface, a maçã e as peras.
[00221] Outros genes que estão associados a características negativas e podem ser silenciados usando as sequências líder ou de arrasto associadas com aqueles genes incluem o gene Rl da batata e genes de fosforilases do tipo L. Ambos os genes são envolvidos na degradação do amido para açúcares de redução, como glicose e frutose, que, por aquecimento, participam na reação de Maillard de modo a produzir produtos tóxicos como a acrilamida. A presente invenção demonstra que uma redução do adocicamento induzido pelo frio baixando Rl ou a atividade da fosforilase conduz a uma redução tanto no escurecimento não enzimático como na acumulação da acrilamida durante o processo de fritura das batatas.
[00222] A invenção demonstra também a utilidade da sobrexpressão de certos genes nativos em colheitas geneticamente modificadas. Os níveis de produtos da reação de Maillard como a acrilamida foram significativamente reduzidos baixando a conversão de sucrose para açúcares de redução através da sobrexpressão de um gene inibidor da invertase vacuolar recentemente isolado na batata.
[00223] A presente invenção também prediz que os tubérculos da batata que exibem um nível aumentado da expressão do inibidor da invertase ou um nível reduzido da expressão de Rl ou da fosforilase não exigirão tratamento intensivo com inibidores químicos de brotos, tais como CIPC antes do armazenamento, porque os seus níveis abaixados de açúcares de redução (1) farão demorar a brotação e (2) permitirão o armazenamento a temperaturas mais baixas, delongando, deste modo, a brotação. Os níveis altamente reduzidos de resíduos CIPC ou a ausência dos mesmos aumenta mais o valor nutricional de alimentos processados derivados de plantas que contêm certos P-DNAs modificados aqui descritos.
[00224] Deste modo, batatas fritas derivadas de tubérculos que contêm o P-DNA modificado conterão níveis de resíduos CIPC fortemente reduzidos, elevando o seu valor nutricional.
[00225] O efeito de simultaneamente subregular a expressão do PPO e o gene Rl ou a fosforilase em tubérculos de batata é sinergético, porque os açúcares redutores não são só exigidos para o escurecimento não enzimático através da reação de Maillard, mas, também para escurecimento mediado pela enzima PPO. Os níveis diminuídos de açúcares redutores em tubérculos de batata transgênica limitarão, portanto, também a atividade de PPO e a suscetibilidade da mancha preta à contusão. Deste modo, os genes PPO, Rl e fosforilase e/ou as sequências líder ou de arrasto que estão associadas a estes genes, representam segmentos de DNA de interesse que podem ser isolados, modificados e reintroduzidos de volta na planta para subregular a expressão destes genes.
[00226] Além de desenvolver a resistência à contusão e o adocicamento reduzido induzido pelo frio existem muitas outras características que podem ser introduzidas através da Procriação Precisa sem usar DNA estranhos. Por exemplo, podem ser isolados genes da resistência a doenças a partir de espécies da batata selvagem e inseridos no genoma de variedades suscetíveis à doença.
[00227] Conforme descrito acima, os níveis reduzidos de Rl ou fosforilase resultam numa fosforilação reduzida do amido. Esta redução na fosforilação do amido resulta numa diminuição de 90% no conteúdo de fosfato de tubérculos da batata (ViksoNielsen, Biomacromolecules, 2: 836-43, 2001). Isto resultará numa redução nos níveis de fosfato em águas de rejeito a partir do processamento de plantas da batata, que são atualmente mais ou menos 25-40 mg/L. Deste modo, o uso de tubérculos de baixo fosfato reduzirá a liberação de fosfatos no ambiente e ajudará a proteger ecossistemas importantes. Além disso, batatas de baixo fosfato podem exigir menos fertilização de fosfato para crescimento e rendimento ótimos, o que apoiaria uma agricultura mais sustentável, postergando a exaustão dos recursos disponíveis de fosfato.
[00228] Além da suscetibilidade reduzida à contusão e adocicamento reduzido ao frio, que são duas características de processamento importantes, a presente invenção também proporciona tolerância ao sal, uma característica de entrada crescentemente importante. Alguns dos construtos de P-DNA modificados descritos na presente invenção contém um gene de tolerância ao sal como marcador nativo para transformação. De modo relevante, a utilidade deste gene não fica limitada a uma etapa de rastreamento no procedimento de transformação. A sobrexpressão do gene de tolerância a sal em plantas da batata reduz sintomas de stress induzidos por níveis de solos de alta salinidade e tornará possível cultivar novas variedades que contêm um P-DNA modificado em porcentagem crescente de terras agrícolas que contêm níveis de salinidade que excedem o máximo de 2 millimhos/cm de níveis de condutividade elétrica que são ótimos para a cultura das variedades convencionais.
[00229] Uma vez o líder, o gene ou o arrasto tenham sido isolados da espécie de planta de interesse e opcionalmente modificados, podem ser operavelmente ligados a um promotor de planta ou elemento regulador semelhante, para expressão apropriada em plantas. Os elementos reguladores como estes servem para expressar sequências não transladadas associadas a um gene de interesse em tecidos específicos ou em certos níveis ou em momentos particulares.
[00230] Dependendo da estratégia envolvida na modificação da característica, pode ser necessário limitar o silenciamento a uma região particular da planta. O promotor que normalmente dirige a expressão do gene endógeno pode não ser adequado para a expressão específica do tecido. Conforme descrito na seção acima, a integração estável de componentes reguladores bacterianos ou virais, como o “super” promotor 35S do vírus do mosaico da couveflor, pode resultar em eventos impossíveis de predizer e indesejáveis. Deste modo, um aspecto da presente invenção usa promotores que são isolados da espécie de planta hospedeira selecionada.
[00231] Numa modalidade preferida da presente invenção, para uso em S. Tuberosum, as sequências líder ou de arrasto associadas com genes Rl, fosforilase e PPO são operavelmente ligadas ao promotor do gene da sintase do amido ligado a grânulo (Rohde e colaboradores, J. Gen. & Breed, 44, 311-315, 1990). Este promotor tem sido frequentemente usado por outros para dirigir a expressão do gene e é particularmente ativo em tubérculos da batata (van der Steege e colaboradores, Plant Mol Biol, 20: 19-30, 1992; Beaujean e colaboradores, Biotechnol. Bioeng., 70: 9-16, 2000; Oxenboll e colaboradores, Proc. Natl. Acad. Sei. USA, 9: 7639-44, 2000). Este promotor pode ser também usado, numa modalidade preferida, para expressão das sequências líder ou de arrasto modificadas de genes de Rl, fosforilase e PPO.
[00232] Alternativamente, outros promotores da batata podem ser operavelmente ligados a sequências de interesse a partir da batata. Esses promotores incluem o promotor do gene patatina (Bevan e colaboradores, Nucleic Acids Res, 14: 4625-38, 1986) ou um fragmento do mesmo, o qual promove a expressão em tubérculos da batata, o promotor do gene da UDPglicose pirofosforilase da batata (Patente US 5.932.783) e o promotor do gene ubiquitin (Garbarino e colaboradores, Plant Physiol., 109: 1371-8, 1995).
[00233] A transcrição de líderes e/ou arrastos pode ser também regulada usando promotores induzíveis e regiões reguladoras que são operavelmente ligados num construto a um polinucleotídeo de interesse. Os exemplos de promotores induzíveis incluem aqueles que são sensíveis á temperatura, como promotores de choques de calor ou frio. Por exemplo, os promotores ci21A e C17- da batata são induzíveis pelo frio (Kirch e colaboradores, Plant Mol. Biol, 33: 897-909, 1997; schneider e colaboradores, Plant Physiol., 113: 335-45, 1997).
[00234] Podem ser usados outros promotores induzíveis que respondem a certos substratos como antibióticos, outras substâncias químicas ou pH. Por exemplo, é sabido que o ácido abscíssico e o ácido giberélico afetam o pH intracelular de células da planta e, ao fazêlo, regulam o gene Rab 16A e o promotor alfaamilase 1/6-4 (Heimovaara Dijkstra e colaboradores, Plant Mol. Biol., 4: 815-20, 1995). O ácido abscíssico, wounding e metil jasmonato são também conhecidos induzirem o promotor pin2 da batata (Lorberth e colaboradores, Plant J., 2:477-86, 1992).
[00235] Noutro exemplo, algumas sequências de nucleotideos estão sob regulação temporal e são ativadas para expressar uma sequência a jusante apenas durante um certo estágio de desenvolvimento da planta ou durante certas horas do dia. Por exemplo, o promotor da batata da pequena subunidade do gene da ribulose-1,5 bisfosfato carboxilase (rbcS) pode dirigir a expressão específica da célula regulada pela luz (Fritz e colaboradores, Proc. Natl. Acad. Sei. USA, 88: 4458-62, 1991). O especialista qualificado está bem versado nestas formas exemplificativas de promotores induzíveis e sequências reguladoras.
[00236] O uso de certos sinais de poliadenilação pode ser também útil para regular a expressão, fazendo variar a estabilidade do transcrito de mRNA. Em particular, alguns sinais de poliadenilação, quando operavelmente ligados à extremidade 3’ de um polinucleotídeo, fazem que o transcrito de mRNA se torne acessível à degradação.
[00237] Deste modo, é possível regular a expressão de um gene ligandoo operavelmente a um ou mais desses promotores, sequências reguladoras, sinais de poliadenilação 3’, regiões 3’ não transladadas, peptídeos de sinal e similares. De acordo com a presente invenção, as sequências de DNA e os elementos reguladores como aqueles aqui descritos e que, em última instância, serão integrados no genoma de uma planta, são obtidos a partir do DNA da espécie de planta selecionada a ser modificada pelo processo da Procriação Precisa da presente invenção. Isto é, as sequências de DNA e os elementos reguladores que são derivados, isolados e clonados de outra espécies, tais como bactérias, vírus, microorganismos, mamíferos, pássaros répteis e espécies de plantas sexualmente incompatíveis não são integrados no genoma da planta transformada. O DNA estranho ao genoma da espécie de planta selecionada pode ser usado na presente invenção para criar um construto de transformação, na medida em que esse DNA estranho não fica integrado no genoma da planta.
[00238] A presente invenção proporciona não apenas um método para transformar uma espécie de planta por integração do DNA obtido da espécie de planta selecionada ou de uma planta que seja sexualmente compatível com a espécie de planta selecionada, também proporciona um meio pelo qual pode ser regulada a expressão desse DNA. Consequentemente, é possível aperfeiçoar a expressão de uma certa sequência, quer por estratégia específica do tecido quer por alguma outra, como previamente descrito.
[00239] Além de elementos reguladores que iniciam a transcrição, o cassete de expressão nativa exige também elementos que terminem a transcrição na extremidade 3’ da região reguladora da iniciação da transcrição. A região de terminação da transcrição e a região de iniciação da transcrição podem ser obtidas do mesmo gene ou de genes diferentes. A região de terminação da transcrição pode ser selecionada, particularmente para estabilidade do mRNA para intensificar a expressão.
[00240] Este elemento particular, a denominada “região não transladada 3”’ é importante no transporte, estabilização, localização e término da transcrição de gene. A este respeito, é bem conhecido dos versados na técnica, que a região não transladada 3’ pode formar certo hairpin loop (laço de grampo de cabelo). Consequentemente, a presente invenção encara a possibilidade de ligar operavelmente uma região não transladada 3’ à extremidade 3’ de um polinucleotídeo clonado de tal forma que o transcrito de mRNA resultante possa ser exposto a fatores que atuam em sequências e estruturas conferidas pela região não transladada 3’.
[00241] Uma sequência 3’ do gene ubiquitin pode ser subclonada da espécie de planta de que o promotor e o transgene foram isolados e inserida a jusante de um transgene, para assegurar a terminação apropriada da transcrição. Ambos os transgenes exemplificativos podem ser fundidos no gene Ubiquitin (Ubi3) da sequência terminadora da batata, não importando se o promotor é usado para dirigir a sua expressão.
[00242] Este exemplo demonstra que as bordas de T-DNA são específicas para Agrobacterium. Mostra também que as plantas contêm sequências T-DNA semelhantes a bordas e provê a sequência de fragmentos de DNA isolados da batata e do trigo que são delineados por essas sequências semelhantes a borda.
[00243] Os sistemas de transformação convencional usam T-DNAs derivados de Agrobacterium como veículos para a transferência de DNA estranho de Agrobacterium para células de plantas (Schilperoort e colaboradores, Patente US 4.940.838, 1990). Embora T-DNAs normalmente incluam várias centenas de pares de base, delineados por uma repetição da borda esquerda (LB) e da borda direita (RB), também podem consistir somente nessas bordas. As bordas de T-DNA desempenham um papel essencial no processo de transferênciaa de DNA, porque funcionam como sítios de reconhecimento específico para a reação de corte catalisada por virD2. O DNA de filamento único liberado, complexado com virD2 e virE2 Agrobacterial, é transferido para núcleos de células de plantas onde frequentemente se integra com sucesso no genoma da planta. Todas as bordas de T-DNA que foram usadas para transferência de DNA estranho são derivadas de cepas de nopalina e octopina de Agrobacterium tumefaciens e A. rhizogenes (Tabela 2). Estas bordas e frequentemente algum DNA flanqueador de Agrobacterium estão presentes em milhares de vetores binários incluindo, por exemplo, pPAM (AY027531), pJawohl (AF408413), pYL156 (AF406991), pINDEX (AF294982), pC1300 (AF294978), pBI121 (AF485783), pLH9000 (AF458478), pAClõl (AJ315956), BinHygTOp (Z37515), pHELLSGATE (AJ311874), pBAR-35S (AJ251014), pGreen (AJ007829), pBIN19 (X77672), pCAMBIA (AF354046), pX6-GFP (AF330636), pER8 (AF309825), PBI101 (U12639), pSKI074 (AF218466), pAJl (AC138659), pAClôl (AJ315956), pSLJ8313 (Y18556) e pGV4939(AY 147202). Recentemente, foram identificados dois homólogos de bordas de T-DNA no plasmideo Ti do tipo crisopina pTiChry5 (Palanichelvam e colaboradores, Mol Planta Microbe Interact 13: 1081-91, 2000). O homólogo da borda esquerda é idêntico ao homólogo da borda inativa localizada no meio do T-DNA de pTil5955. O homólogo da borda direita é raramente divergente da sequência de bordas funcionais de T-DNA. Ê, portanto, improvável que estes homólogos sejam funcionalmente ativos em sustentar a transferência de DNA de pTiChry5 para células de plantas.
[00244] O desenvolvimento de um novo método que torna possível transformar plantas apenas com DNA nativo exige, em primeiro lugar, uma substituição do T-DNA inclusive LB e RB. Infelizmente, buscas avançadas BLAST de bases de dados públicas incluindo aquelas mantidas pelo The National Center For Biotechnology Information, The Institute for Genomic Research e SANGER falharam em identificar quaisquer sequências de borda em plantas. Foi, portanto, necessário considerar sequências de DNA de plantas que são semelhantes, mas, não idênticas a bordas de T-DNA, designadas aqui como “semelhantes a borda” (como borda). Exemplos de sequências de plantas semelhantes a borda que foram identificadas em bases de dados públicas são mostrados na Tabela 2. O desafio de tentar substituir bordas de T-DNA por sequências semelhantes a borda é que as sequências de borda são altamente conservadas (ver Tabela 2). Uma grande parte destas sequências é também altamente conservada nas regiões de de corte de outros sistemas de transferência de DNA bacteriano como aquele de IncP, PC 194 e 0X174, indicando que estas sequências são essenciais para a transferência de DNA do tipo conjugado (Waters e colaboradores, Proc Natl Acad Sci 88: 1456-60, 1991). Porque não existem nenhuns dados confiáveis em requisitos de sequência de borda, a borda inteira parece, então, importante no processo de corte. Um estudo único que tentou resolver este assunto testando a eficácia de mutantes de borda no suporte da transferência de DNA é não confiável, porque os controles negativos não pareceram funcionar apropriadamente (van Haaren e colaboradores Plant Mol. Biol. 13: 523-531, 1989). Além disso, nenhum dos resultados deste estudo foi confirmado molecularmente. Apesar destas preocupações, dois mutantes de borda possivelmente efetivos são também mostrados na Tabela 2.
[00245] Baseado na homologia entre sequências de borda, foi identificado um motivo de borda de T-DNA (Tabela 2). Embora este motivo inclua 13.824 variantes, muitas das quais podem não funcionar - ou podem ser inadequadas - na transferência do DNA, representa a definição mais ampla possível do que uma sequência de borda de T- DNA é ou pode ser. Este motivo de borda foi, então, usado para pesquisar bases de dados de DNA publicamente disponíveis para homólogos usando o “Motif Alignment and Search Toor (Bailey e Gribskov, Bioinformatics 14: 48-54, 1998) e “advanced BLASTN” (“penalidade para não casamento de nucleotideo”=—1; “espera”=105; Altschul e colaboradores, Nucleic Acids Res 25: 3389-3402, 1997). Novamente estas pesquisas não identificaram nenhuns casamentos idênticos em organismos outros que não Agrobacterium.
[00246] Para tentar e aumentar a chance de isolar um fragmento de DNA da batata que contenha sequências semelhantes a borda que correspondam ao motivo de borda, o DNA foi isolado a partir de 100 acessos geneticamente diversos (a denominada “coleção de núcleo”, fornecida pela US Potato Genebank, WI). Este DNA foi agrupado e usado como modelo para reações em cadeia de polimerase usando uma variedade de oligonucleotídeos projetada para ligarse às bordas ou sequências semelhantes a borda. Os fragmentos ampliados foram analisados quanto às sequências e a sequência foi, então, confirmada usando PCR inverso com primários aninhados. Um dos fragmentos de DNA da batata que era de interesse particular contém uma sequência nova sem nenhumas molduras de leitura abertas principais que sejam delineadas por sequências semelhantes a borda (Tabela 2). Uma das sequências semelhantes a borda deste fragmento contém pelo menos 5 desacertos (mismatches) com bordas de T-DNA; a outra sequência semelhante a borda contém, pelo menos, 2 desacertos. Embora ambas as sequências contenham um desacerto com o motivo de borda, elas foram testadas quanto à capacidade de sustentar a transferência de DNA. Com esse propósito, o fragmento foi, primeiro, reduzido em tamanho a 0,4 quilopares de base realizando uma deleção interna (SEQ ID NO.: 1). O fragmento resultante foi designado “P-DNA” (planta DNA) para distinguilo do T-DNA derivado de Agrobacterium. Um fragmento semelhante foi isolado do genoma da variedade de batata Russet Ranger, mas, não foi usado para nenhumas experiências adicionais.
[00247] Com base na divergência entre P-DNA e bordas de T-DNA, foi usado o sistema de amplificação elongase (Life Technologies) com os seguintes primários de degeneração para isolar um P-DNA do trigo: 5’ GTTTACANHNBNATATATCCTGYCA-3’ (BorF) (SEQ ID NO.: 56) e 5’ TGRCAGGATATATNVNDNTGTAAAC-3’ (BorR) (SEQ ID NO.: 57). O fragmento resultante de 825 bps é mostrado na SEQ ID NO.: 2 e foi usado para substituir o T-DNA de um vetor binário convencional. A eficácia deste construto pode ser testada inserindo um cassete de expressão para o gene GUS entre os terminais de P-DNA e infectando o trigo com uma cepa de Agrobacterium portando o vetor resultante.
[00248] Este Exemplo demonstra que, apesar das diferenças estruturais (divergência de sequência) e funcionais (frequências de transformação) entre os términos de P-DNA e as bordas de T-DNA, pode ser usado um P-DNA de um modo semelhante a um T-DNA para transferir DNA de Agrobacterium para células de tabaco.
[00249] Foi obtido um vetor isento de T-DNA que pode ser mantido tanto em E. coli como A. tumefaciens removendo a região inteira de T- DNA do vetor binário convencional pCAMBIA1301 (Cambia, AU). Isto foi realizado ligando simultaneamente um fragmento de 5.9 kb SacIISphl de pSIM1301 com 2 fragmentos ampliados de pCAMBIA1301 usando os pares de oligonucleotídeos: 5’ CCGCGGTGATCACAG- GCAGCAAC-3’ (SEQ ID NO.: 58) e 5’AAGCTTCCAGCCAGCCAACAGCTCCCCGAC-3’(SEQ ID NO.: 59) e 5’ AAGCTTGGCTACTAGTGCGAGAT-CTCTAAGAGAAAAGAGCGTTTA-3’ (SEQ ID NO.:60) e 5’ GCATGCTCGAGATAGGTGACCAC- ATACAAATGGACGAACGG-3’ (SEQ ID NO.: 61), respectivamente.
[00250] Para tornar possível rastrear contra eventos de integração de esqueleto, um cassete de expressão incluindo o gene isopentenil transferase do Agrobacterium {IPT) dirigido pelo promotor Ubi3 e seguido pelo terminador Ubi3 (SEQ ID NO.: 3) foi inserido como fragmento SacII de 2.6 kbp no esqueleto do vetor isento de T-DNA descrito acima, produzindo pSIMlOO-OD-IPT. As células da planta transformada expressando o gene IPT são esperadas acumular citocininas e crescer em brotos anormais que não podem desenvolver raízes.
[00251] O fragmento de 0,4 kb de P-DNA descrito no Exemplo 1 foi inserido em pSIM100-OD IPT para gerar pSIMlll (Figura 1; SEQ ID NO.: 4).
[00252] Para testar se pSIMlll pode ser usado para obter plantas transformadas portando P-DNAs (inclusive quaisquer sequências localizadas entre os terminais de P-DNA) sem o esqueleto de vetor adicional, foi inserido um cassete de expressão do gene neomicina fosfotransferase (NPTIl) no P-DNA de pSIMlll para criar pSIM108 (Figura 1).
[00253] A eficácia dos terminais de P-DNA em sustentar a transferência de DNA foi testada comparando frequências de transformação entre pSIM108 e um vetor de controle que continha um P-DNA modificado com bordas T-DNA convencionais. Este vetor de controle, designado pSIM109, foi gerado por amplificação do P-DNA inteiro que continha o cassete de expressão de gene NPTII com os pares de oligonucleotídeos: 5’ ACTAGTGTTTACCCGCCAATATATCCTGT CAGAG-3’ (SEQ ID NO.: 62) e 5’AAGCTTTGGCAGGATATATTGTGG TGTAAACGAAG-3’ (SEQ ID NO.: 63). Um segundo vetor de controle que foi usado nestas experiências é o vetor binário convencional pBI121 (número de acesso de Genbank AF485783), que contém o mesmo cassete de expressão de NPTII inserido num T-DNA regular. Os vetores binários foram introduzidos em células LBA4404 de Agrobacterium tumefaciens como se segue. Células competentes de LB4404 (50 pL) foram incubadas durante 5 minutos a 37°C na presença de 1 pg de DNA de vetor, congeladas durante mais ou menos 15 segundos em nitrogênio líquido (cerca de — 196°C) e incubadas novamente a 37°C durante 5 minutos. Depois de adicionar 1 mL de caldo de cultura líquido (LB), as células tratadas foram cultivadas durante 3 horas a 28°C e colocadas em placa de LB/agar contendo estreptomicina (100 mg/L) e canamicina (100 mg/L). Os DNAs de vetor foram, então, isolados de culturas durante a noite a partir de colônias individuais de LBA4404 e examinadas por análise de restrição, para confirmar a presença de DNA de plasmideo intacto.
[00254] As transformações de teste do tabaco de planta modelo foram realizadas cultivando uma diluição de 10 vezes de células LBA4404::pSIM108 cultivadas durante a noite em 5-6 horas, precipitando as células durante 15 minutos a 2.800 rpm, lavandoas com meio de MS líquido (Phytotechnology) suplementado com sucrose (3%, pH 5,7) e resuspendendo as células no mesmo meio a um ODeoonm de 0,2. A suspensão foi, então, usada para infectar in vitro explantes de folhas de plantas de Nicotiana tabacum de 4 semanas de idade. Os explantes infectados de tabaco foram incubados durante 2 dias em meio de cocultura (sais 1/10 de MS, 3% sacarose, pH 5,7) contendo 6 g/L de agar a 25°C numa câmara de cultura de Percival (16 hrs de luz) e, subsequentemente, transferidos para meio de M401/agar contendo timentina (150 mg/L) e canamicina (100 mg/L). O número de calos por explante que se desenvolveu dentro das 4 semanas seguintes é mostrado na Tabela 3. Os nossos dados demonstram que os P-DNAs delineados por terminais nativos ou bordas de T-DNA convencionais são cerca de 50% mais efetivos na transformação do tabaco do que T-DNAS. A eficiência aumentada de transferência de P-DNA pode ser devida ao seu conteúdo de CG diferente ou outras características estruturais desconhecidas do P-DNA.
[00255] Este Exemplo demonstra que pode ser usado um P-DNA de um modo semelhante a um T-DNA para transferir DNA de Agrobacterium para células de batata.
[00256] As transformações de batata foram realizadas infectando explantes de caule de 4 semanas de idade de plântulas Russet Ranger cultivadas in vitro com cepas de Agrobacterium de acordo com o procedimento seguinte. As diluições de dez vezes de culturas crescidas durante a noite foram cultivadas durante 5-6 horas, precipitadas durante 15 minutos a 2.800 rpm, lavadas com meio de MS líquido (Phytotechnology) suplementado com sacarose (3%, pH 5,7) e ressuspendidas no mesmo meio para um ODõoonm de 0,2. As células ressuspendidas foram, então, usadas para infectar 0,4-0,6 mm de segmentos internodais de batata. Os caules infectados foram incubados durante 2 dias em meio de cocultura (sais 1/10 de MS, 3% sacarose, pH 5,7) contendo 6 g/L agar a 22°C numa câmara de crescimento de Percival (16 hrs de luz) e, subsequentemente, transferidos para meio de indução de calos (CIM, meio MS suplementado com 3% sacarose 3, 2,5 mg/L de zeatin ribosideo, 0,1 mg/L de ácido naftaleno acético e 6g/L de agar) contendo timentina (150 mg/L) e canamicina (100 mg/L). Depois de 1 mês de cultura em CIM, os explantes foram transferidos para meio de indução de brotos (SIM, meio MS suplementado com 3% de sacarose, 2,5 mg/L de zeatin ribosideo, 0,3 mg/L de ácido giberélico GA3 e 6g/L de agar) contendo timentina e canamicina (150 e 100 mg/L respectivamente). Depois de 3-4 semanas, foi contado o número de explantes que desenvolviam calos transgênicos e/ou brotos. Como mostrado no tobaco, o número de explantes de caules infectados com pSIM108 que mostrou calos era mais alto do que aqueles em experiências de controle com o vetor binário convencional pBI121 (Tabela 3). Os brotos que surgiram subsequentemente destes calos podiam ser agrupados em duas classes diferentes. A primeira classe de brotos era fenotipicamente indistinguível dos brotos de controle transformados com LBA::pBI121. A segunda classe de brotos exibia um fenótipo IPT. Os brotos da última classe eram raquíticos, continham apenas folhas muito pequenas, exibiam uma cor verde claro para amarelo e eram incapazes de emitir raiz por transferência para meios isentos de hormônio. Para confirmar que os brotos com um fenótipo IPT continham o gene IPT integrado de modo estável em seus genomas, todos os brotos foram transferidos para caixas de Magenta contendo meio MS suplementado com 3% de sacarose e timentine 150 mg/L, deixados crescer durante 3 a 4 semanas mais e usados para isolar DNA. Este DNA de planta serviu como modelo em reações de PCR com um par de oligonucleotídeos projetado para ligarse ao gene de IPT: 5’GTC CAA CTT GCA CAG GAA AGA C-3’; e 5’CAT GGA TGA AAT ACT CCT GAG C-3'. Como mostrado na Tabela 4, a experiência de PCR confirmou uma correlação rígida entre o fenótipo IPT e a presença do gene de IPT. A presença do esqueleto de DNA foi também examinada em plantas obtidas a partir de uma transformação com pBI121. Isto foi feito efetuando reações de PCR em DNA isolado dos eventos de transformação com os ‘primários de esqueleto pBI121’: 5’CGGTGTAAGTGAACTGCAGTTGCCATG-3’ (SEQ ID NO.: 64) e 5’ CATCGGCCTCACTCATG-AGCAGATTG-3’ (SEQ ID NO.: 65). A amplificação de uma faixa de 0,7 kbp é indicativa da integração do esqueleto. Comparando os dados apresentado na Tabela 4, pode ser concluído que as frequências de integração de esqueleto são semelhantes para vetores de P-DNA e vetores de T-DNA.
[00257] Foi realizada uma segunda experiência de PCR para testar se as plantas isentas de IPT não continham nenhumas outras sequências de esqueleto. Como o cassete de expressão de IPT fica posicionado perto das sequências semelhantes a borda esquerdas, o par oligonucleotídeo para esta experiência foi projetado para ligarse a sequências de esqueleto perto da sequência semelhante a borda direita: 5’CACGC-TAAGTGCCGGCCGTCCGAG-3’ (SEQ ID NO.: 66) e 5TCCTAATCGACGGCGCACCGGCTG-3’ (SEQ ID NO.: 67). Os dados desta experiência confirmam que as plantas que eram positivas para o gene IPT são também positivas para esta outra parte do esqueleto.
[00258] Foram realizadas experiências semelhantes com a variedade de batata Russet Burbank. Com base numa avaliação de fenótipos de IPT, as frequências de integração de esqueleto para pSIM108 e pSIM109 foram mostradas serem comparáveis àquelas em Russet Ranger (ver Tabelas 4 e 5).
[00259] Usando métodos de transformação convencional, este Exemplo demonstra que a sobrexpressão do novo gene inibidor da invertase da batata aumenta as características de processamento e a saúde dos tubérculos da batata.
[00260] Foram projetados os seguintes primários para ampliar um novo homólogo da batata do inibidor da invertase vacuolar do tabaco Ntinhhl (Greiner e colaboradores, Nature Biotechnology, 17, 708-711, 1999): 5’AAAGTTGAATT-CAAATGAGAAATTTATTC-3’ (SEQ ID NO.: 68) e 5TTTTAAGCTTTCATAATAACATTCTAAT-3’ (SEQ ID NO.: 69). A reação de amplificação foi realizada misturando os seguintes componentes: 4 pL de DNA de planta, 2 pL de primário dianteiro (10 pM/ml), 2 pL de primário invertido, 25 pL de Hot Start Master Mix (Catálogo de Qiagen Nr. 203443) e 17 pL de água. Esta mistura de reação foi sujeita às seguintes condições de reação em cadeia da polimerase (PCR) usando um termociclador PTC-100 (MJ Research): (1) 5 minutos a 95°C (1 ciclo), (2) 1 minuto a 94°C, 1 minuto a 45°C e 4 minutos a 72°C (35 ciclos) e (3) 10 minutos a 720°C (1 ciclo). O produto total foi carregado em gel de agarose 0,8% e uma faixa de pares de base 540 foi purificada de gel usando QIAquick Gel Extraction Kit (Qiagen, CA). Este fragmento purificado foi, então, ligado em pGEMT Easy (Promega, WI) e transformado em E. coli DH5-alfa usando Max Efficiency Competent Cells (GibcoBRL, MD). A análise de sequências do DNA dos plasmideos recombinantes isolados do alfa DH5 transformado revelou a presença de uma única moldura de leitura aberta consistindo em 543 pares de base que codificam uma proteína putativa de 181 aminoácidos (SEQ ID NO.: 5); o alinhamento de núcleos (cluster alignment) revelou 70% de homologia para NTinhh (Figura 2). Este alto nível de homologia estendese para o domínio Nterminal de 15 aminoácidos, indicando que o homólogo da batata é direcionado para o vacúolo. De maneira interessante, o homólogo do inibidor da invertase da batata, designado Stinhl, compartilha só 43% de homologia com o inibidor patenteado da invertase da parede da célula do tabaco, designado NTinhl (Patente W098/04722; Figura 2).
[00261] Embora o gene Stinhl esteja presente em tubérculos de batata inalterada, o seu nível de expressão é inadequado para a inibição completa da invertase e do adocicamento induzido pelo frio reduzido. Para melhorar as características de armazenamento da batata, o gene Stinhl foi fundido com um novo promotor intensificado de tubérculo do gene da sintase do amido limitado a grânulo (GBSS), que é sabido promover níveis altos de expressão do gene em tubérculos. O promotor GBSS foi isolado do cultivar da batata Russet Ranger efetivando uma reação de PCR usando o primário dianteiro 5’GAACCATGCATCTCAATC- 3’ (SEQ ID NO.: 70) e o primário inverso 5’GTCAGGATCCCTACCAAGCTACAGATGAAC-3’ (SEQ ID NO.: 71). A análise de sequências do produto ampliado clonado em pGEMT demonstrou que este novo promotor contém 658 pares de base (SEQ ID NO.:6). O promotor/gene de fusão resultante foi, então, ligado à sequência reguladora 3’ do gene ubiquitin da batata (UbiT; SEQ ID NO.: 7), assegurando, deste modo, a terminação apropriada da transcrição do gene inibidor da invertase.
[00262] Este cassete de expressão foi inserido entre as bordas T- DNA de um vetor binário e foi usado o vetor resultante pSIM320 para transformar Russet Ranger como descrito acima. Três recortes de nove linhas transgênicas independentes foram plantados no solo e cultivadas durante quatro semanas numa câmara de crescimento (11 hrs de luz; 20°C). Foram, então, colhidos pelo menos 3 minitubérculos de cada linha e transferidos para uma geladeira ajustada aos 4°C para induzir o adocicamento por frio. Depois de 4 semanas, foram determinados os níveis de glicose nestes minitubérculos armazenados a frio usando um medidor AccuChek e tiras de teste (de Roche Diagnostics, IN) ou um reativo de glicose oxidase/ peroxidase (Megazyme, Irlanda). Estes níveis foram comparados com os níveis de glicose média tanto em linhas não transformadas quanto em 6 fileiras de “controle de vetor” transformadas com um vetor derivado de pSIMllO a que faltava o gene inibidor da invertase. Conforme mostrado na Tabela 6, três linhas transgênicas acumularam menos de 40% da glicose nas linhas de “controle de vetor”, demonstrando que o homólogo do inibidor da invertase da batata é funcionalmente ativo.
[00263] O experimento seguinte mostrou que a quantidade de açúcares redutores presente em tubérculos se correlaciona com a produção de acrilamida, durante o processamento do tubérculo. Os tubérculos de batata Russet Ranger foram colhidos de fresco do campo e armazenados a 4°C para induzir o adocicamento pelo frio; os tubérculos de controle foram armazenados a 18°C. Depois de 4 semanas, foram determinados os níveis de glicose em ambos os grupos de tubérculos. Subsequentemente, os tubérculos foram lavados, lexiviados durante 8 minutos ou 12 minutos a 74°C, cortados em tiras de 0,290 x 0,290 de cadarço de sapato, imersos numa solução de pirofosfato ácido de sódio a 1% a 71°C, secos a 71°C até que fosse alcançada perda de peso de 14±2% no secador, fritos a 199°C durante 40 segundos para atingir 64±2% de primeira umidade de fritura e congelados por 20 minutos a —26°C, agitando a bandeja 2-3 vezes nos primeiros 6 minutos. As batatas fritas resultantes foram, então, analisadas quanto aos níveis de acrilamida pelo Laboratório de Covance (WI). Conforme mostrado na Tabela 7, os níveis de glicose em tubérculos armazenados a 18°C estavam abaixo do nível de detecção de 0,1 mg/g, enquanto os tubérculos armazenados a frio continham em média 3,4 mg/g glicose. Esta Tabela mostra também que as batatas fritas produzidas a partir das últimas batatas contêm níveis cerca de 10 vezes mais altos de acrilamida do que as batatas fritas produzidas a partir de batatas armazenadas a 18°C. Até mesmo usando um tempo menor de lexiviação para as batatas armazenadas a 18°C do que para as batatas armazenadas a 4°C para produzir batatas fritas com uma cor semelhante (cor ids de 78 e 71, respectivamente), foi obtida uma diferença de 5 vezes na acumulação de acrilamida (Tabela 7). Deste modo, parece haver uma correlação direta entre a quantidade de açúcares reduzidos, tais como glicose, em tubérculos e a acumulação de acrilamida nas batatas fritas derivadas destes tubérculos.
[00264] Para determinar se os níveis de glicose reduzida em linhas pSIM320 limitariam a acumulação de acrilamida induzida pelo processamento, processaramse minitubérculos armazenados a frio pSIM320 cortandoos em cunhas, lexiviando durante 8 minutos, imergindo em SAPP a 0,5% durante 30 segundos, secando durante 4,5 minutos a 71 °C, fritando por 40 segundos a 193°C, congelando durante 15 minutos a —26°C e, finalmente, secando por 3 minutos e 10 segundos a 71°C. O material processado foi, então, embarcado para o Laboratório de Covance para determinações de acrilamida. Conforme mostrado na Tabela 6, as batatas fritas obtidas a partir de minitubérculos com as quantidades mais baixas de glicose acumularam os níveis mais baixos de acrilamida. Uma redução de 40% nos níveis de glicose nas linhas “320-2” e “320-4” está associada a uma redução de 5 vezes nos níveis de acrilamida.
[00265] Usando métodos de transformação convencional, este Exemplo demonstra que uma nova sequência líder associada com o gene Rl da batata pode ser usada eficazmente para melhorar as características de processamento e saúde dos tubérculos da batata. Prediz também que um novo arrasto associado com aquele mesmo gene pode ser explorado da mesma maneira.
[00266] Como alternativa para sobrexpressar o gene inibidor da invertase, foram desenvolvidos métodos para limitar a produção de acrilamida sem usar nenhumas sequências de gene reais. Esse método é baseado no silenciamento do gene Rl expresso no tubérculo. Anteriormente, mostrouse que este gene relacionado a amido pode ser silenciado através da expressão antisentido de um fragmento de gene de 1,9 kbs derivado daquele gene (Kossmann e colaboradores, Patente US 6.207.880). Contudo, a expressão antisentido de fragmentos de DNA grandes é indesejável, porque esses fragmentos contêm novas molduras de leitura abertas (Tabela 1). Como abordagem mais segura para o descrito acima, foi isolada uma pequena sequência líder associada com o gene Rl da batata. Este líder foi obtido realizando uma amplificação rápida de extremidades cDNA com o kit 5’ RACE fornecido por GIBCO BRL sobre o RNA total dos tubérculos de plantas de batata de Russet Ranger. A análise de sequências demonstrou que o líder associado a Rl consiste em 179 pares de base (SEQ ID NO.: 8). Tanto uma cópia de sentido como de antisentido desta sequência líder, separada pelo intron Ubiquitin da batata (SEQ ID NO.: 9) foram colocadas entre o promotor GBSS e UbiT. O cassete de expressão resultante para a sequência líder associada com Rl é mostrado na Figura 3 (SEQ ID NO.: 10). Um cassete semelhante contendo um espaçador derivado do promotor GBSS (SEQ ID NO.: 11) - em vez do intron Ubi -separando as cópias de sentido e de antisentido do arrasto Rl é mostrado (Figura 3; SEQ ID NO.: 12). Variantes adicionais com uma versão mais longa do promotor GBSS (SEQ ID NO.: 13) são mostradas na Figura 3 (SEQ ID NOs.: 14- 15).
[00267] Para testar a eficácia do líder associado a Rl em limitar a produção de acrilamida, o cassete de expressão mostrado na Figura 3 foi inserido como fragmento KpnIXbal entre as bordas de T-DNA de um vetor binário. Foi usada uma cepa LBA4404 de Agrobacterium portando o vetor resultante pSIM332 para transformar a batata Russet Ranger. Para induzir a formação de tubérculos, 25 brotos representando eventos de transformação independentes foram transferidos para o solo e colocados numa câmara de crescimento (11 horas de luz, 25°C). Depois de três semanas, pelo menos 3 minitubérculos/linha foram armazenados durante 4 semanas a 4°C para induzir a mobilização do amido. Os níveis de glicose nestes minitubérculos armazenados a frio foram subsequentemente determinados como descrito no Exemplo 4 e comparados com os níveis de glicose média em plantas e controles de vetor não transformados. Como mostrado na Tabela 8, os minitubérculos derivados de todas as 25 linhas exibiam níveis reduzidos de glicose depois do armazenamento a frio. Uma redução aproximada de 2 vezes nos níveis de acrilamida em batatas fritas derivadas de minitubérculos exibiam níveis reduzidos de expressão de Rl comparados com os controles. São antecipados efeitos muito mais fortes de subregular a expressão do gene Rl em tubérculos maduros.
[00268] Como alternativa para a abordagem baseada em líder, foram gerados cassetes de expressão que continham tanto uma cópia de sentido quanto de antisentido da sequência de arrasto associada com Rl. Este arrasto foi obtido realizando uma reação em cadeia da polimerase de transcrição inversa (RT-PCR) no RNA total isolado de microtubérculos do cultivar da batata Russet Ranger. Foi gerado DNA complementar usando o kit de Omniscript RT (Qiagen, CA) e, então, usado como modelo para uma reação de PCR com polimerase DNA de partida quente (Qiagen, CA) com o primário inverso do gene específico Rl-1 (5’GTTCAGACAA-GACCACAGATGTGA-3’). A análise de sequências do fragmento de DNA ampliado, clonado em pGEMT, demonstrou que o arrasto associado com Rl consiste em 333 pares de base (SEQ ID NO.: 16). As cópias de sentido e de antisentido do arrasto foram separadas pelo intron Ubi ou o espaçador GBSS e comprimidas entre o promotor GBSS e o terminador Ubi3 (Figura 3; SEQ ID NOs.: 17-18). As versões semelhantes com o promotor GBSS maior são mostradas na Figura 3 (SEQ ID NOs.: 19-20).
[00269] Os níveis de glicose e acrilamida podem ser determinados conforme descrito acima. Os tubérculos que exibem reduções de cerca de 50% ou maiores nas concentrações de glicose são também esperados acumularem cerca de 50% menos de acrilamida durante o processo de fritura. As características de saúde e armazenamento melhoradas de plantas modificadas podem ser confirmadas em tubérculos maduros crescidos no campo.
[00270] Podem ser determinados os níveis de fosfato em tubérculos de batata usando o Método Fósforo AOAC 995.11 (Total) em Alimentos (45.1. 33 Official Methods of Analysis of AOAC Internacional, 17a Edição). As amostras são preparadas pelas cinzas secas em forno seguido de digestão ácida. As amostras dissolvidas são, então, neutralizadas e tratadas com uma solução de molibdatoácido ascórbico e comparadas com uma série de padrões de fósforo (tratados de modo semelhante). Um espetrofotômetro de feixe duplo seria usado para a análise colorimétrica a 823 nanômetros. Ê esperada uma diminuição significativa no conteúdo de fosfato, que é benéfica para o ambiente.
[00271] Usando métodos de transformação convencional, este Exemplo demonstra que uma nova sequência líder associada com o gene Lalfa glucan fosforilase da batata pode ser eficazmente usada para melhorar as características de processamento e saúde de tubérculos da batata.
[00272] Anteriormente, foi mostrado que o adocicamento induzido pelo frio pode ser reduzido através da expressão de antisentido de fragmentos 0,9 kbs derivados de genes alfa glucan fosforilase (Kawchuk e colaboradores, Patente US 5.998.701, 1999). Contudo, a expressão antisentido destes fragmentos relativamente grandes de DNA é indesejável, porque contêm novas molduras de leitura aberta não caracterizadas que podem impactar a qualidade nutricional dos alimentos, se expressas em plantas transgênicas (Tabela 1).
[00273] Como aproximação mais segura para o líder descrito acima, foram isoladas pequenas sequências de arrasto que estão associadas com um gene glucan fosforilase tipo L a partir do RNA de tubérculos maduros. O par primário usado com este propósito é: 5’GGATCCGA- GTGTGGGTAAGTAATTAAG-3’ (SEQ ID NO.: 72) e 5’GAATTCTGTGCTCTCTATGCAAATCTAGC-3’ (SEQ ID NO.: 73). A sequência líder resultante de 273 bp foi ampliada e é mostrada na SEQ ID NO.: 21. Semelhantemente, o primário “direto”, 5’GGAACATTGAAGCTGTGG-3’ (SEQ ID NO.: 74), foi usado com um primário oligodT para ampliar uma “sequência de arrasto” de 158 bp que está associada com o gene da fosforilase do tipo L (SEQ ID NO.: 22).
[00274] Os cassetes de expressão foram, então, projetados usando estas sequências de arrasto ou líder para modificar a expressão do gene da fosforilase do tipo L e, ao fazêlo, diminuir os níveis de acrilamida em produtos fritos, pela limitação da mobilização do amido. Estes cassetes foram construídos de um modo semelhante ao descrito no Exemplo 5 e estão representados na Figura 3 (SEQ ID NOs.: 23-26). Foi usada uma cepa de Agrobacterium contendo um vetor binário com este cassete de expressão, designada pSIM216, para infectar caules de batata e gerar 25 plantas transgênicas. Minitubérculos derivados destas plantas foram armazenados durante 4 semanas a 4°C para induzir o adocicamento pelo frio. Os minitubérculos armazenados a frio foram, então, analisados quanto aos níveis de glicose. Como mostrado na Tabela 9, os minitubérculos de todas as linhas transgênicas exibiram níveis de glicose reduzidos.
[00275] Foram usadas quatro linhas que exibiram pelo menos 50% de concentrações de glicose reduzidas (linhas 216-2, 216-5, 216-10 e 216-21) para avaliar níveis de acrilamida induzidos pelo processamento. Embora os níveis de acrilamida em tubérculos fritos derivados das primeiras três linhas fossem semelhantes aos dos controles, as batatas fritas que foram derivadas da linha 216-21 acumularam só 45% dos níveis de acrilamida do tipo selvagem (136 vs. 305 partes por bilhão). Estes resultados confirmam as experiências descritas no Exemplo 4 para tubérculos que sobrexpressam o gene inibidor da invertase da batata, na medida em que são necessárias reduções relativamente grandes nas concentrações de glicose (e frutose) para limitar a acumulação de acrilamida induzida por aquecimento em minitubérculos armazenados a frio. Como o silenciamento do gene da fosforilase é esperado ser mais efetivo em tubérculos “216” maduros, as reduções nos níveis de acrilamida são também antecipados serem mais pronunciados nas batatas fritas produzidas a partir desses tubérculos. As características de saúde e armazenamento melhoradas de plantas modificadas podem ser confirmadas em tubérculos maduros.
[00276] Usando métodos de transformação convencional, este Exemplo demonstra que pode ser usado eficazmente um gene modificado da polifenol oxidase a que falta um sítio de ligação a cobre funcional para reduzir a suscetibilidade à contusão em tubérculos.
[00277] Anteriormente foi mostrado que a suscetibilidade à contusão da mancha preta pode ser reduzida através da expressão antisentido do gene PPO de 1,8 kbs (Steffens, Patente US 6.160.204, 2000). Contudo, a expressão do complemento inverso deste gene grande é indesejável, porque ele contém novas molduras de leitura aberta não caracterizadas que codificam peptídeos que consistem em mais de 100 aminoácidos, que podem potencialmente impactar a qualidade nutricional dos alimentos (Tabela 1). Como uma aproximação mais segura para o descrito acima, o gene PPO foi modificado para codificar uma proteína não funcional.
[00278] O gene PPO da batata de tipo selvagem foi isolado de Russet Ranger usando um método de reação em cadeia da polimerase (PCR). Primeiro, foi isolado o DNA genômico de brotos de Russet Ranger. O gene PPO da batata foi, então, ampliado a partir do DNA genômico da batata usando a polimerase do DNA e primários de oligonucleotídeos: 5’CGAATTCATGGCA-AGCTTGTGCAATAG-3’ (PPOF) (SEQ ID NO.: 75) e 5’CGAATTCTTAACAATCTGCAAGACTGAT-CG-3’ (PPOR) (SEQ ID NO.: 76). Estes foram projetados para complementar as extremidades 5’e 3’ do gene PPO da batata. Os fragmentos ampliados de 1,6 kb foram clonados num vetor pGEMT EASY (Promega) e foi confirmado re[presentarem um gene PPO funcional por análise de sequências (SEQ ID NO.: 27).
[00279] O domínio de ligação a cobre no PPO da batata foi identificado alinhando esta proteína com uma proteína PPO da batata doce que foi mostrado conter resíduo de Cisteína conservada (Cys) na posição 92, resíduo de Glutamina (Glu) na posição 236 e resíduos de Histidina (His) nas posições 88, 109, 118, 240, 244 e 274 coordenando os dois cobres dos sítios ativos (Klabunde e colaboradores, Nature Structural Biol., 5: 1084-90, 1998). Estes resíduos Cys, Glu e His estão também presentes no PPO da batata.
[00280] O gene de PPO inativo foi criado usando uma abordagem de substituição de mutação de PCR.
[00281] Três fragmentos foram ampliados por Proof Start Taq DNA Polymerase (Qiagen) usando 3 pares de primários e PPO Russet Ranger do tipo selvagem como modelo. As sequências do primeiro par, designado Pl-F e P2-R, respectivamente, são: 5’GAGAGATCTTGATAAGACACAACC-3’ (SEQ ID NO.: 77) e 5’CATTACC1ATAAGCC2CAC3TGTATATTAGCTTGTTGC-3’ (SEQ ID NO.: 78) (1: mutação de “A” para “C”, que resulta na substituição da Cisteína por Glicina na posição 186; 2: mutação “A” para “C”, que resulta na substituição da Cisteína por Triptofano na posição 183; 3: mutação ”A” para “C”, que resulta na substituição da Histina por Glutamina na posição 182). As sequências do segundo par, designado P3-F e P4-R, respectivamente, são 5’GTGCTTATAGAATTGGTGGC-3’ (SEQ ID NO.: 79) e 5TAGTTCCCGGGAGTTCA-GTG-3’ (SEQ ID NO.: 80). As sequências do terceiro par, designadas P5-F e P6-R, respectivamente, são 5’CTCCCGGGAACTATAGG4A-AACATTCCTCT5CGGTCCTGTCCACATCTGGTC -3’ (SEQ ID NO.: 81) e 5’GTGTGATATCTGTTCT-TTTCC-3’ (SEQ ID NO.: 82) (4: mutação “A” para “G”, que resulta na substituição da Glutamina por Glicina na posição 326; 5: mutação “A” para “T”, que resulta na substituição da Histina por Leucina na posição 330).
[00282] Um fragmento de 80 bp foi ampliado usando primário Pl-F e P2-R e digerido com BglII. Este fragmento contém uma extremidade pegajosa (BglII) e uma extremidade rombuda e porta três mutações no sítio de ligação do cobre I. Um fragmento de 0,4 kb ampliado usando o primário P3-F e P4-R e digerido com Xmal contém uma extremidade rombuda e uma extremidade pegajosa (Xmal). Um fragmento de 0,2 kb foi ampliado usando o primário P5-F e P6-R e digerido com Xmal e EcoRV. Este terceiro fragmento com uma extremidade pegajosa (Xmal) e uma extremidade rombuda (EcoRV) tem duas mutações no sítio de ligação de cobre II. O fragmento de BglII e EcoRV a partir do PPO da batata de tipo selvagem clonado foi, então, substituído pelos três fragmentos ampliados PCR ligados acima. A presença de um total de 5 mutações pontuais no gene PPO modificado foi confirmada por análise de sequências (SEQ ID NO.: 28). Para criar um cassete de expressão para PPO modificado (mPPO), foram ligados os seguintes quatro fragmentos simultaneamente em conjunto: (1) um fragmento de BamHIHindlII contendo o promotor GBSS, (2) um fragmento de HindIIISacI contendo PPO mutante, (3) um fragmento de SacIKpnl contendo o terminador Ubi-3 e (4) plasmideo PBluescript, digerido com Kpnl e BamHI. Este cassete de expressão foi, então, inserido entre as bordas de um vetor binário para criar pSIM314.
[00283] A eficácia do cassete de expressão do gene mPPO foi avaliada transformando explantes de caule de Russet Ranger com pSIM314. Os recortes nodais de plantas transgênicas que contêm este cassete de expressão foram colocados em meio MS suplementado com 7% de sacarose. Depois de um período de incubação de 5 semanas às escuras a 18°C, foram isolados e analisados microtubérculos quanto à atividade PPO. Com esta finalidade, foi pulverizado lg de tubérculos de batata em nitrogênio líquido. Este pó foi, então, adicionado a 5 ml de 50 mM de tampão MOPS (pH 6,5) (3-(Nmorfolino) propanoácido sulfônico) contendo 50 mM de catecol e incubados à temperatura ambiente com rotação durante cerca de 1 hora. A fração sólida foi, então, precipitada e o sobrenadante transferido para outro tubo, para determinar a atividade de PPO medindo a mudança de OD-410 com o passar do tempo. Conforme mostrado na Tabela 10, os microtubérculos isolados de algumas das linhas transgênicas exibiram uma atividade significativamente reduzida da polifenol oxidase comparada com os controles não transformados ou os controles transformados com um construto não contendo o gene PPO mutante. A redução mais forte na atividade PPO foi observada em linhas “314-9”, “314-17” e “314-29”. Para testar se a expressão do gene mutante PPO também reduziu a atividade PPO em minitubérculos, foram plantadas no solo plântulas enraizadas de linhas transgênicas e incubadas numa câmara de cultura durante 4 semanas. Um ensaio de PPO em minitubérculos isolados demonstrou que a atividade PPO reduzida em microtubérculos se correlacionava na maioria dos casos com atividades reduzidas em minitubérculos (Tabela 10). Linhas transgênicas que exibem uma atividade reduzida de PPO podem ser propagadas e testadas tanto na estufa quanto no campo para confirmar o fenótipo de “baixa contusão” em tubérculos maduros. Como os micro e minitubérculos expressam uma variedade de polifenol oxidases, algumas das quais compartilham apenas limitada homologia de sequências com a polifenol oxidase visada que está predominantemente expressa em tubérculos maduros, pode ser antecipada uma redução ainda mais profunda da atividade de PPO nos tubérculos maduros de linhas tais como “314-9” e “314-17”. Os dados indicam que a sobrexpressão de um gene PPO funcionalmente inativo pode resultar em reduzida suscetibilidade de contusão. As características de saúde e armazenamento melhoradas de plantas modificadas podem ser também confirmadas em tubérculos maduros cultivados no campo.
[00284] Usando métodos de transformação convencional, este Exemplo demonstra que uma nova sequência de arrasto associada com o gene PPO da batata pode ser eficazmente usada para reduzir a suscetibilidade à contusão em tubérculos.
[00285] A transcrição inversa de PCR foi também utilizada para isolar a sequência de arrasto associada com o gene PPO expresso em tubérculos da batata. Os primários para a primeira reação de PCR foram PPO-1 (5’GAATGA-GCTTGACAAGGCGGAG-3’, (SEQ ID NO.: 83)) e os oligodT; primários para uma segunda reação PCR aninhada foram PPO-2 (5’CTGGCGAT-AACGGAACTGTTG-3’, (SEQ ID NO.: 84)) e oligodT. A análise de sequências dos fragmentos de DNA ampliados clonados em pGEM-T revelou a presença de um arrasto de 154 bp (SEQ ID NO.: 29). Uma cópia de sentido e de antisentido deste arrasto, separado pelo intron Ubi foi, então, fundida ao promotor GBSS e ao terminador Ubi3, conforme descrito acima, para gerar um cassete de expressão, mostrado na Figura 3 (SEQ ID NO.: 30). Um construto alternativo contendo os segmentos de arrasto separados por um espaçador GBSS está mostrado na Figura 3 (SEQ ID NO.: 31). As versões semelhantes com o promotor GBSS maior são mostradas na Figura 3 (SEQ ID NOs.: 32-33). De maneira interessante, o arrasto do gene PPO que está predominantemente expresso em tubérculos maduros (indicado com PPP03 na Figura 4) é diferente do arrasto de genes PPO que estão predominantemente expressos em outros tecidos, incluindo microtubérculos (indicados com PPOM-41 e PPOM-44 na Figura 4). Em razão da baixa homologia entre arrastos associados a genes PPO diferentes, o uso do arrasto P-PP03 resultará num silenciamento do gene de PPO específico do tubérculo maduro somente. Este silenciamento de gene muito específico seria difícil de realizar com sequências derivadas do gene PPO propriamente, demonstrando, deste modo, a vantagem de usar sequências não codificadoras para silenciamento de genes. Para visualizar a estensão da atividade PPO, 0,5 mL de 50 mm catecol foram pipetados nas superfícies de corte de minitubérculos modificados geneticamente, fatiados. Comparado com controles, o escurecimento visual das regiões do tubérculo era reduzido de cerca de 5 a 10 vezes. De maneira interessante, entretanto, nenhum escurecimento reduzido foi observado na pele da batata. Parece que a sequência de arrasto usada silenciou especificamente o gene PPO que está predominantemente expresso no córtex e medula, mas, não na camada epidérmica. Este achado inesperado pode ser benéfico aos tubérculos para protegêlos contra alguns patógenos que tentam infectar através da pele, porque o gene PPO pode desempenhar algum papel em certas respostas de defesa. Para determinar quantitativamente a atividade PPO, foi realizado um ensaio como descrito no Exemplo 7. A Tabela 11 mostra até 80% de redução da atividade PPO em minitubérculos transformados, em comparação com controles não transformados. O nível de redução é esperado ser até maior em tubérculos maduros, porque estes tubérculos expressam o gene PPO visado mais predominantemente do que mini e microtubérculos. As características melhoradas de linhas como “217-7” e “217-26” podem ser confirmadas em tubérculos maduros.
[00286] O aumento das razões amilose/ amilopectina em tubérculos pode intensificar mais o valor nutricional de produtos da batata. Um método que torna possível aumentar o conteúdo de amilose é baseado na expressão de antisentido da codificação de genes para a enzima de ramificação do amido (SBE) I e II (Schwall e colaboradores, Nature Biotechnology 18: 551-554, 2000). As desvantagens deste método são que (1) a eficiência do silenciamento simultâneo de dois genes diferentes através da exploração de tecnologias de antisentido é muito baixa, (2) a expressão de antisentido das sequências do gene do SBEI e SBEII relativamente grandes resulta na expressão indesejável de molduras de leitura aberta (Tabela 1) (3) construtos correspondentes que abrigam os dois cassetes de expressão de antisentido são desnecessariamente grandes e complexos, aumentando, deste modo, as probabilidades de recombinação e diminuindo as frequências de transformação.
[00287] A nossa abordagem para aumentar o conteúdo de amilose na batata é baseada na expressão das sequências de arrasto que estão associadas a ambos os genes. Estes arrastos (SEQ ID NO.: 34 e 35) foram isolados com os pares de primários 5’GTCCATGATGTCTTCAGGGTGGTA-3’ (SEQ ID NO.: 85) e 5’CTAATATT- TGATATATGTGATTGT-3’ (SEQ ID NO.: 86) e 5ACGAACTTGTGATCGCGTTGAAAG-3’ (SEQ ID NO.: 87) e 5ACTAAGCAAAACCTGCTGAAGCC-C-3’ (SEQ ID NO.: 88). Um promotor único dirige a expressão de uma fusão de sentido e de antisentido de ambos os arrastos, separado pelo intron Ubiquitin-7 e seguida pelo terminador Ubiquitin-3. O tamanho do cassete de expressão inteira é apenas de 2,5 kb.
[00288] Este Exemplo demonstra que as plantas podem ser eficazmente transformadas sem necessidade da integração estável de genes marcadores selecionáveis.
[00289] Este método é o primeiro a aproveitarse do fenômeno de que DNAs direcionados para os núcleos de células de planta frequentemente deixam de integrarse subsequentemente no genoma da célula de planta. Os inventores fizeram a constatação surpreendente de que é possível selecionar células que expressam temporariamente um T-DNA que não se integra que contém um gene marcador selecionável colocando explantes infectados durante 5 dias num meio de planta com o agente seletivo apropriado. Um segundo fenômeno que foi aplicado para desenvolver o método atual é que T-DNAs de vetores binários diferentes frequentemente visam o mesmo núcleo de células da planta. Usando dois vetores binários diferentes, um contendo o marcador selecionável num T-DNA e o outro portando um T-DNA ou P-DNA com as sequências de interesse real foi possível aplicar um sistema de seleção transiente e obter populações de calos, brotos ou plantas, uma parte significativa da qual representa eventos de transformação isentos de marcadores.
[00290] Foi usado um vetor binário convencional designado pSIMOll para representar o vetor com a “sequência de interesse”, que é, no caso deste teste, um cassete de expressão para o gene beta glucuronidase (GUS) localizado num T-DNA convencional. O segundo vetor binário que foi usado para estes experimentos contém um cassete de expressão que compreende o gene da neomicina fosfotransferase [NPTII] dirigido pelo promotor forte do gene Ubiquitin-7 e seguido das sequências terminadoras do gene da nopalina sintase (nos) entre as bordas do T-DNA de um derivado pSIMOl 1.
[00291] Surpreendentemente, pôde ser também obtido um nível forte de níveis de expressão do gene de NPTII transiente substituindo o terminador nos pelo gene terminador do álcool da dehidrogenase 1(ADH1) da levedura (Genbank número de acesso VO 1292, SEQ ID NO.: 56). Este achado é interessante, porque o terminador ADH1 da levedura não compartilha a homologia com nenhum terminador de planta. De modo importante, deve ser aqui notado que muitos terminadores de levedura não funcionam adequadamente em plantas. Por exemplo, quase nenhuma expressão do gene GUS foi observada numa experiência semelhante conforme descrito acima com o gene GUS seguido do terminador isolcitocromo c(CYCl) da levedura (número de acesso Genbank SCCYT1). Foi gerado um vetor aperfeiçoado portando o gene marcador selecionável NPTII substituindo o terminador nos pelo terminador ADH1 da levedura. O vetor binário contendo um gene marcador selecionável para transformação transiente é designado “LifeSupport” (Figura 5).
[00292] Explantes do caule da batata foram simultaneamente infectados com duas cepas A. tumefaciens LBA4404 que continham pSIMOll e LifeSupport, respectivamente. Uma diluição 1/10 de culturas de cada cepa crescidas durante a noite foram cultivadas durante 5-6 horas antes fossem precipitadas, lavadas e ressuspendidas a um ODõoonm de 0,4, como descrito no Exemplo 3. As células ressuspendidas foram, então, usadas para infectar segmentos de batata internodais de 0,4-0,6 cm a uma densidade final de cada bactéria de 0,2 (ODeoonm). Os caules infectados foram tratados como no Exemplo 3, com uma diferença principal: a seleção com canamicina foi limitada aos primeiros 5 dias de cultura em meio de indução de calos. Então, os explantes foram deixados desenvolve mais em CIM e SIM fresco contendo apenas timentina 150 mg/L mas, sem nenhum antibiótico seletivo. Dentro de cerca de 3 meses do dia da infecção as folhas dos brotos derivadas de calos desenvolvidos em 40-60% dos caules infectados foram tanto testadas quanto à expressão de GUS como analisadas por PCR para identificar eventos que continham as sequências de interesse, mas, nenhum gene marcador. Como mostrado na Tabela 12, 11% dos brotos representavam eventos de transformação isentos de marcador.
[00293] A abordagem das duas cepas descrita acima foi também usada para transformar tabaco. Os brotos que se desenvolveram dentro de cerca de 2 meses foram analisados quanto a GUS e analisados quanto a PCR. A frequência elevada de eventos de transformação isentos de marcador identificados (18%) implica que o método desenvolvido é aplicável para plantar espécies diferentes da batata (Tabela 12).
[00294] De modo importante, a infecção sequencial em lugar da infecção simultânea com as duas cepas diferentes de Agrobacterium resultou num aumento da eficiência de transformação isenta de marcador. O efeito surpreendente de infecções sequenciais foi descoberto infectando explantes de caules da batata com a cepa Agrobacterium contendo pSIMOl 1, colocando os explantes infectados em placas de cocultivo durante 4 horas e, então, reinfectandoos com o vetor de LifeSupport. Os explantes duplamente infectados foram tratados como previamente descrito neste Exemplo. Conforme mostrado na Tabela 13, o tempo de atraso de 4 horas entre as duas infecções diferentes resultou numa frequência aumentada de 2 vezes os eventos de transformação isentos de marcador na batata.
[00295] Este Exemplo demonstra a eficácia da procriação precisa. A saúde e as características agronômicas de plantas da batata são aumentadas inserindo elementos genéticos da batata (ver Exemplos 1, 4 e 7) na batata, usando a transformação isenta de marcador (Exemplo 10).
[00296] Foi criado um vetor binário contendo dois cassetes de expressão para os genes do inibidor da invertase e mutante da polifenol oxidase inseridos entre os terminais P-DNA, designado pSIM340 (Figura 1) inserindo ambos os cassetes de expressão de PPO mutante e inibidor da invertase num vetor binário pSIM112’. Os explantes do caule da batata foram simultaneamente infectados com pSIM340 e um vetor mais aperfeiçoado LifeSupport. Os explantes infectados foram, então, cocultivados, sujeitos a seleção transiente e induzidos a proliferar e desenvolver brotos, conforme discutido anteriormente. Depois de 3 meses, os brotos pequenos foram transferidos para novos meios e deixados crescer durante 3 semanas adicionais. Os brotos foram, então, fenotipicamente analisados e o material das folhas foi coletado para análises moleculares para determinar a presença de esqueleto, gene marcador e P-DNA com as sequências de interesse, conforme descrito nos Exemplos 2 e 3. Segundo mostrado na Tabela 14, 1,2% de eventos representavam uma planta que continha o P-DNA modificado de pSIM340 sem LifeSupport. Esta frequência de transformação isenta de marcador é mais baixa do que a encontrado para um T-DNA, revelando novamente uma diferença funcional entre P-DNA e T-DNA.
[00297] Este Exemplo demonstra que a eficiência de métodos de procriação precisos pode ser aumentada selecionando contra a integração estável de LifeSupport T-DNAs usando o gene bacteriano da citosina deaminase.
[00298] O exemplo anterior demonstra que a eficiência da transformação isenta de marcador é várias vezes mais baixa com um P- DNA modificado do que com um T-DNA convencional. Para melhorar a eficiência de geração de brotos contendo apenas um P-DNA modificado, foi inserido um cassete de expressão para uma fusão de genes de suicídio incluindo os genes bacterianos da citosina deaminase (codA) e uracil fosforibosiltransferase (upp) (InvivoGen, CA) entre bordas de T- DNA do vetor de LifeSupport, gerando pSIM346 (Figura 5). Explantes de caule da batata foram infectados com uma cepa portando pSIM340 e outra portando pSIM346 e subsequentemente colocados no seguinte meio: (1) meios de cocultivo durante 2 dias, (2) meios CIMTK para selecionar expressão de gene marcador transiente durante 5 dias, (3) meios Cl MT para permitir a proliferação de células de planta que expressassem transientemente o gene marcador durante 30 dias, (4) meios SIMT com 500 mg/L de 5 fluorocitosinas não tóxicas (5-FC), que serão convertidas por células da planta expressando codA::upp no 5 fluorouracil (5-FU) tóxico, para selecionar contra a integração estável do LifeSupport TDNA. O calo ocasionou brotos em SIMT dentro de 4 semanas. Estes brotos foram transferidos para meios MS com timentina e deixados crescer até que estivesse disponível tecido suficiente para análise de PCR. O DNA foi, então, extraído de 100 brotos e usado para determinar a presença de P-DNA, LifeSupport e esqueleto. Conforme mostrado na Tabela 15, nenhum dos brotos analisados continha um LifeSupport T-DNA, indicando, pela primeira vez, que a fusão de genes codA::upp fusão pode ser usada como marcador selecionável negativo anterior à regeneração. De modo mais importante, os nossos resultados demonstram que uma seleção negativa contra a integração LifeSupport T-DNA aumenta a frequência de brotos que só contêm um P-DNA modificado. Juntando uma seleção positiva para expressão transiente do gene marcador com uma seleção negativa contra a integração estável da fusão de gene de codA::upp, a frequência de brotos que só contêm um P-DNA modificado é cerca de 5 vezes mais alta do que empregando apenas a seleção positiva para a expressão transiente do gene marcador (Tabela 15).
[00299] Foi obtido um aumento ainda maior na eficiência de transformação isenta de marcador usando o vetor de LifeSupport pSIM350 (Figura 5), que é semelhante a pSIM346 mas contém o gene codA em vez da fusão de genes codA::upp. Explantes de caule da batata simultaneamente infectados com pSIM340 e pSIM350 foram tratados como descrito acima e foram testados molecularmente 51 brotos resultantes, quanto à ocorrência de eventos contendo apenas a região T-DNA de pSIM340. De maneira interessante, esta análise de PCR revelou que alguns brotos continham o gene codA (Tabela 15). Este achado demonstra que codA não é um marcador selecionável negativo tão apertado quanto codA::upp em plantas. De modo mais importante, um grande número de brotos (29%) mostrou representarem eventos de transformação isentos de marcador.
[00300] As eficiências podem ser ainda aumentadas não infectando os explantes simultaneamente com pSIM340 e pSIM350, mas, consecutivamente. Infectando os explantes com pSIM340 e reinfectandoos com pSIM350, depois de 4 horas, as frequências de transformação isentas de marcador são esperadas serem de aproximadamente 30-40%.
[00301] Este Exemplo demonstra que a eficiência dos métodos de procriação precisa pode ser aumentada prejudicando a integração do LifeSupport T-DNA no genoma da planta usando um gene ómegamutacionado virD2.
[00302] Foi mostrado que o domínio ómega da proteína vird2 do Agrobacterium é importante para a capacidade daquela proteína sustentar a integração de T-DNA no genoma das plantas (Mysore e colaboradores, Mol Plant Microbe Interact 11: 668-83, 1998). Baseado nesta observação, foram criados vetores modificados de LifeSupport que contêm um cassete de expressão para uma proteína ómegamutacionada virD2 inserida no sítio SacII em suas sequências de esqueleto. O cassete de expressão foi obtido ampliando um fragmento de DNA de 2,2 kb a partir do plasmideo pCS45 (cortesia de Dr. Walt Resma - Oregon State Universidade, OR, EUA - SEQ ID NO.: 36). Foi usado um derivado de LifeSupport portando este cassete de expressão, designado pSIM401'Ω (Figura 5), para sustentar a transformação de plantas da batata com o P-DNA modificado de pSIM340. Depois da seleção transiente e indução de brotos, 100 brotos foram testados molecularmente quanto à presença de transgenes. Conforme mostrado na Tabela 15, 4,4% de brotos só continham o P-DNA modificado, indicando que o uso de ómegavirD2 aumenta a eficiência da transformação isenta de marcador cerca de 4 vezes (Tabela 15).
[00303] As eficiências são ainda melhoradas aumentando o tamanho do LifeSupport T-DNA de 3,7 kb (em pSIM401Q) para 8,1 kb (no derivado pSIM401'Ω designado de pSIM341Ω; Figura 5). Regenerando os brotos de explantes de caules da batata simultaneamente infectados com pSIM340 e pSIM341Ω, 7 de 81 eventos analisados (7%) foram mostrados representarem eventos de transformação isentos de marcador (Tabela 15).
[00304] Um aperfeiçoamento adicional pode ser obtido infectando explantes consecutivamente em lugar de simultaneamente com pSIM340 e LifeSupport. De um modo semelhante ao descrito no Exemplo 10, a frequência de plantas que só contém um P-DNA modificado pode ser aproximadamente dobrada infectando os explantes com pSIM340 e reinfectandoos com LifeSupport depois de 4 horas
[00305] Este Exemplo demonstra que podem ser também obtidas altas frequências de transformação isenta de marcador usando uma cepa de Agrobacterium única que contém tanto o vetor de P-DNA como o LifeSupport.
[00306] Foram criados dois vetores binários compatíveis que podem ser simultaneamente mantidos em Agrobacterium. Em vez de usar este sistema para integrar de modo estável dois T-DNAs portando o DNA de interesse e um gene marcador, respectivamente (Komari e colaboradores, Patente US 5.731.179, 1998), pretendese a integração apenas do P-DNA modificado.
[00307] O primeiro vetor, designado pSIM356, contém um cassete de expressão que compreende o gene GUS dirigido pelo promotor Ubi7 e seguido do UbiT inserido entre os terminais P-DNA. A parte de esqueleto deste vetor contém origens bacterianas de replicação de pVSl e pBR322, um gene de resistência à espectinomicina para seleção bacteriana e um cassete de expressão para o gene IPT capacitar a seleção contra integração de esqueleto em plantas (Figura 1). O segundo vetor, designado pSIM363, contém um cassete de expressão que compreende o gene NPTII dirigido pelo promotor Ubi7 e seguido do terminador ADH1 da levedura inserido entre bordas convencionais de T- DNA (Figura 5). A parte de esqueleto deste vetor contém origens bacterianas de replicação a partir de ColEl (número de Genbank V00268) e ori V (número de Genbank M20134) e um gene de resistência à canamicina para seleção bacteriana.
[00308] O conceito de aumentar as frequências de transformação isenta de marcador usando pSIM356 e pSIM363 foi testado em 100 brotos de tabaco. Conforme mostrado na Tabela 16, cerca de 19% dos brotos regenerados continham o DNA de interesse sem gene marcador. Foi também encontrado um aumento na eficiência de transformação isenta de marcador aplicando esta abordagem da cepa-1 à batata. Nove de 60 brotos independentes testados (15%) continham o pSIM340 T- DNA e não tinham o LifeSupport T-DNA (Tabela 16).
[00309] A abordagem da cepa-1 pode ser combinada com o método descrito no Exemplo 12 para casar uma seleção positiva para expressão de gene marcador transiente com uma seleção negativa contra integração estável do gene codA. Com este propósito, foi desenvolvido o vetor de LifeSupport pSIM365 (Figura 5). Uma cepa Agrobacterium portando este vetor junto com um vetor de P-DNA pode ser usada para desenvolver eficazmente plantas que só contém um cassete de expressão de interesse localizado dentro de um P-DNA integrado de modo estável em seu genomas.
[00310] Além de transformar plantas de colheita com P-DNAs que apenas contêm as sequências desejáveis para introduzir características benéficas, a presente invenção proporciona também um método de transformar essas plantas com P-DNAs que contêm um gene marcador nativo adicional. Os nossos novos genes nativos marcadores de escolha são homólogos da batata do gene antiporter Na+/H+ vacuolar Arabidopsis e gene da alfafa alfinl. A expressão destes genes não só permite a identificação de eventos de transformação, mas, também proporciona às plantas transformadas tolerância ao sal. Os níveis de alta salinidade numa área crescente de terra agrícola afetarão menos, então, as plantas de batata que contiverem o marcador de tolerância a sal.
[00311] Duas versões de um homólogo antiporter vacuolar Na+/H+, designado Pst (Potato salt tolerance] foram ampliadas a partir de cDNA de uma variedade resistente à ferrugem recente obtida a partir do US Potato Genbank (WI), designada “LBR4”, usando o par de oligonucleotídeos 5’CCCGGGATGGCTTCTGTGCTG-GCT-3’ (SEQ ID NO.: 89) e 5’GGTACCTCATGG-ACCCTGTTCCGT-3’ (SEQ ID NO.: 90). As suas sequências são mostradas na SEQ ID NO.:37 e 38. Um terceiro gene (SEQ ID NO.: 39) com homologia para alfin-1 foi ampliado a partir do DNA da batata LBR4 usando os primários 5’C- CCGGGTATGGAAAATTCGGTACCCAGGACTG-3’ (SEQ ID NO.: 91) e 5’ACTAGTTAAACTCTAGCT-C-TCTTGC-3’ (SEQ ID NO.: 92). A eficácia dos genes de Pst para funcionarem como marcadores de transformação foi avaliada inserindo uma fusão com o promotor Ubi7 entre bordas convencionais de T-DNA de um vetor modificado pSIM341. Depois de um período de seleção transiente, células resistentes à canamicina são deixadas proliferar e desenvolver brotos. Estes brotos são, então, transferidos para meios que contêm cloreto de sódio 100-150 mM. Os brotos tolerantes a sal representam eventos de transformação que contêm o T-DNA do pSIM341 modificado.
[00312] Um novo promotor específico de tubérculo recentemente isolado pode substituir o promotor GBSS usado para desenvolver os cassetes de expressão descrita em exemplos anteriores. Este promotor foi isolado do genoma de plantas de batata Russet Burbank usando a reação inversa em cadeia da polimerase com primários específicos para um gene do inibidor da proteinase da batata (acesso Genbank D17332) (SEQ ID NO.: 39). Foi testada a eficácia do promotor PIP criando um vetor binário que contém o gene GUS dirigido por este promotor e um cassete de expressão para o gene marcador NPTII. Foi usado um construto semelhante com o promotor PIP substituído pelo promotor GBSS como controle. Os brotos transformados foram obtidos infectando explantes de caule com cepas Agrobacterium portando os vetores binários, cocultivando por 2 dias e selecionando em meio de CIMTK durante 2 meses. Estes brotos foram transferidos para novos meios para induzir formação de raiz e, depois, plantados na terra. Os tubérculos podem ser analisados para expressão de GUS depois de um período de crescimento de 3 meses na estufa.
[00313] Além de pSIM340, podem ser usados muitos outros vetores para melhorar plantas da batata transformandoas com P-DNAs modificados. Dois desses vetores contêm um cassete de expressão para uma cópia de sentido e de antisentido do arrasto associado com um gene PPO que é expresso em todos os tecidos de tubérculo com exceção da epiderme (ver Exemplo 8). O vetor pSIM370 contém um cassete de expressão adicional para uma cópia de sentido e de antisentido do líder associado com o gene da fosforilase (ver Exemplo 6). O vetor pSIM371 contém um terceiro cassete de expressão para o homólogo da batata alfin-1 (Figura 1).
[00314] Um terceiro vetor alternativo, designado pSIM372, contém tanto um cassete de expressão para o homólogo da batata alfin-1 quanto um cassete de expressão para uma cópia de sentido e de antisentido de uma fusão do arrasto associado a PPO, um líder associado a Rl e um líder associado a fosforilase.
[00315] O vetor de LifeSupport preferido para uma abordagem de 1 cepa é pSIM365. Para uma abordagem de 2 cepas, o vetor preferido é pSIM367, que contém cassetes de expressão para ambos os NPTII e codA entre bordas de T-DNA e um cassete de expressão adicional para ómega virD2 no esqueleto do plasmideo (Figura 5).
[00316] Os explantes de caule da batata são infectados com 1 cepa portando tanto pSIM365 quanto qualquer dos vetores pSIM370, 371 e 372 ou sequencialmente com 2 cepas portando pSIM366 e quaisquer dos vetores preferidos de interesse, respectivamente. Depois de 2 dias de cocultivo e um período de seleção transiente de 5 dias, os explantes são transferidos para meios para proliferação/ regeneração e eliminação do Agrobacterium. Trinta dias mais tarde, os explantes são novamente transferidos para o mesmo meio, mas, agora contendo também 5-FU para eliminar eventos contendo LifeSupport T-DNAs. Os brotos que surgem subsequentemente em calos são transferidos para meios de regeneração que podem conter sal 100-200 mM para rastrear eventos de tolerância a sal. Os brotos de IPT negativo são deixados enraizar e desenvolver em plantas maduras. Uma proporção grande destas plantas (10%—100%) é predita representar plantas isentas de marcador e isentas de esqueleto contendo um P-DNA com a sequência de nucleotideos de interesse integrada de modo estável em seu genomas. Tabela 1 - Peptídeos não caracterizados potencialmente expressos em linhas de batatas de antisentido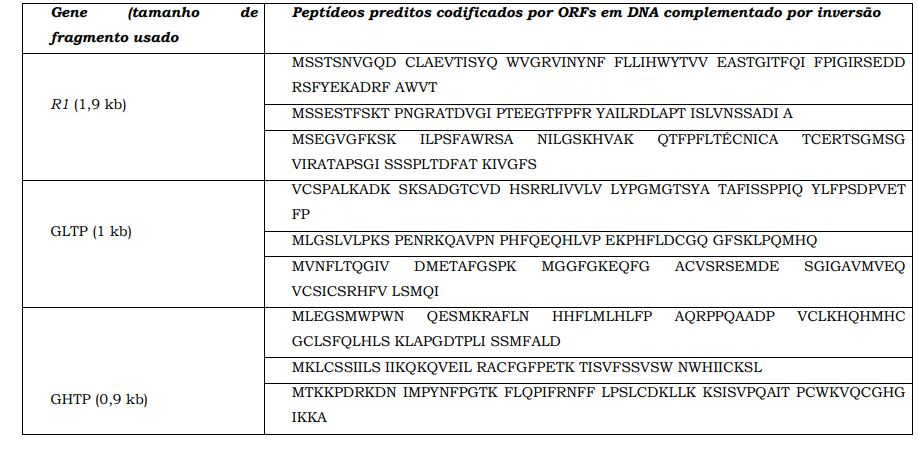
 Tabela de 3 - Eficiência de Transformação
Tabela de 3 - Eficiência de Transformação  Tabela 4 - Integração de Esqueleto resultante de transformação de Guardaflorestal Ruivo
Tabela 4 - Integração de Esqueleto resultante de transformação de Guardaflorestal Ruivo 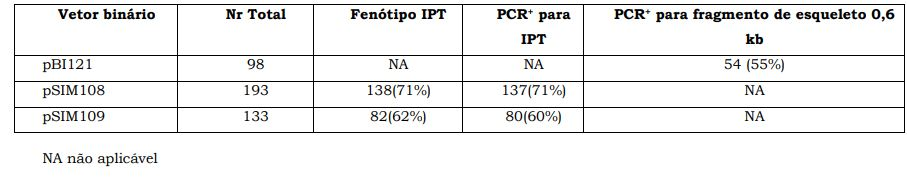 Tabela 5 - Integração de esqueleto resultante de transformação de Russet Burbank
Tabela 5 - Integração de esqueleto resultante de transformação de Russet Burbank 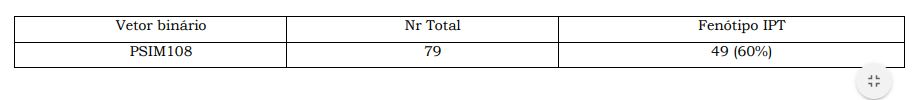
 Tabela 6 - Níveis de acrilamida em batatas fritas derivadas de minitubérculos pSIM320 armazenadas a frio
Tabela 6 - Níveis de acrilamida em batatas fritas derivadas de minitubérculos pSIM320 armazenadas a frio 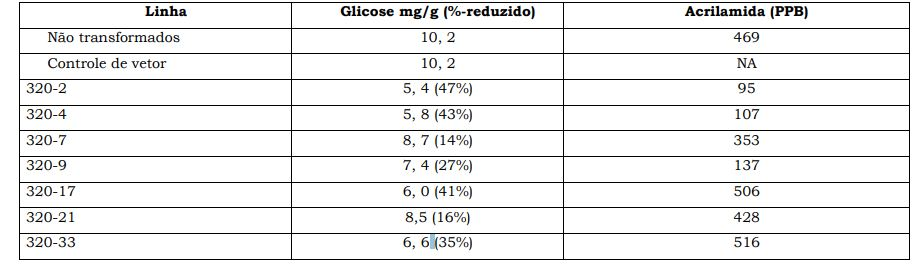 Tabela 7 - Níveis de acrilamida nas batatas fritas derivadas de tubérculos maduros não transformados
Tabela 7 - Níveis de acrilamida nas batatas fritas derivadas de tubérculos maduros não transformados 
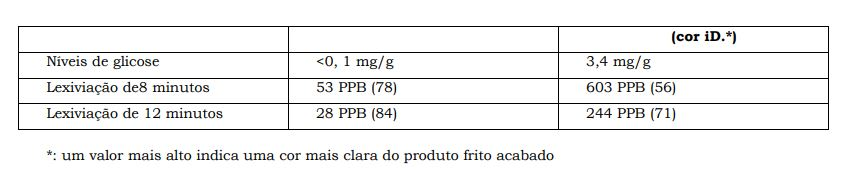 Tabela 8 - Níveis de glicose em minitubérculos pSIM332 armazenados a frio
Tabela 8 - Níveis de glicose em minitubérculos pSIM332 armazenados a frio 
 Tabela 9 - Níveis de glicose em minitubérculos pSIM216 armazenados a frio
Tabela 9 - Níveis de glicose em minitubérculos pSIM216 armazenados a frio 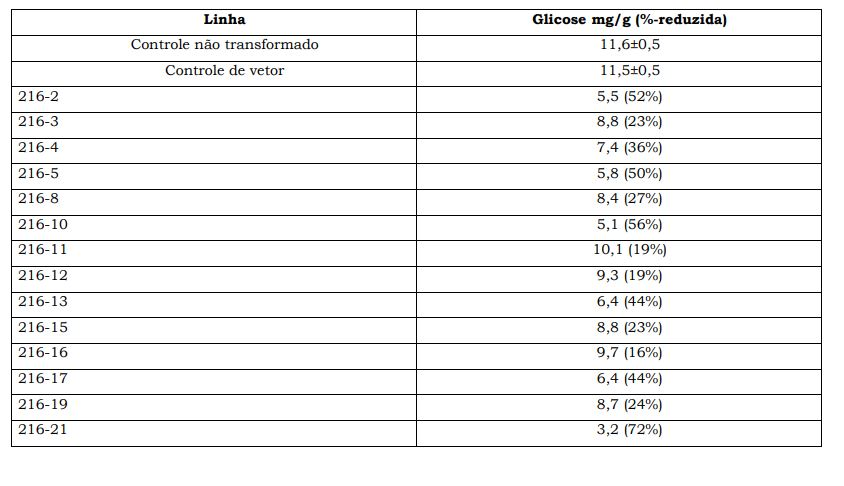
 Tabela 10 - Atividade de PPO em linhas de batata que expressam um gene PPO modificado
Tabela 10 - Atividade de PPO em linhas de batata que expressam um gene PPO modificado 

 Tabela 11 - Atividade PPO em minitubérculos da batata que expressam uma sequência de arrasto associada gene PPO
Tabela 11 - Atividade PPO em minitubérculos da batata que expressam uma sequência de arrasto associada gene PPO 
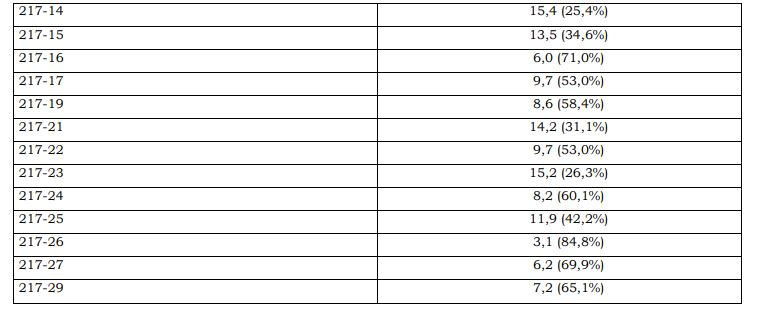 Tabela 12 - Transformação isenta de marcador com o vetor LifeSupport + PSIM011
Tabela 12 - Transformação isenta de marcador com o vetor LifeSupport + PSIM011 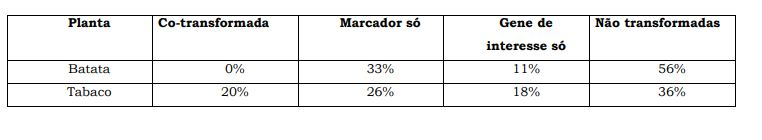 Co-transformado: PCRpositivo para ambos os GUS e NPT Gene de interesse somente: PCRpositivo para GUS Não transformadas: As plantas são PCRnegativo para ambos os GUS e NPT Tabela 13 - Transformação sequencial da batata com o vetor LifeSupport e o Tempo de pSIM011
Co-transformado: PCRpositivo para ambos os GUS e NPT Gene de interesse somente: PCRpositivo para GUS Não transformadas: As plantas são PCRnegativo para ambos os GUS e NPT Tabela 13 - Transformação sequencial da batata com o vetor LifeSupport e o Tempo de pSIM011  Tabela 14 - Transformação isenta de marcador com o vetor P-DNA pSIM340+
Tabela 14 - Transformação isenta de marcador com o vetor P-DNA pSIM340+  Co-transformada: PCRpositivo para ambos o gene PPO de pSIM340 e o gene de NPT de de LifeSupport Não transformados: As plantas são PCRnegativo para de PPO e NPTII Tabela 15 - Transformação de batata isenta de marcador com vetores de LifeSupport pSIM340+ melhorados
Co-transformada: PCRpositivo para ambos o gene PPO de pSIM340 e o gene de NPT de de LifeSupport Não transformados: As plantas são PCRnegativo para de PPO e NPTII Tabela 15 - Transformação de batata isenta de marcador com vetores de LifeSupport pSIM340+ melhorados  Co-transformado: PCR-positivo para ambos o gene PPO de pSIM340 e o gene NPT de LifeSupport Não transformadas: As plantas são PCR-negativo para PPO e NPTII Tabela 16 - A transformação de batata isenta de marcador com uma cepa única de Agrobacterium portando ambos pSIM356 pSIM363
Co-transformado: PCR-positivo para ambos o gene PPO de pSIM340 e o gene NPT de LifeSupport Não transformadas: As plantas são PCR-negativo para PPO e NPTII Tabela 16 - A transformação de batata isenta de marcador com uma cepa única de Agrobacterium portando ambos pSIM356 pSIM363 
 Co-transformado: PCR-positivo para ambos o gene GUS de pSIM356 e o gene NPT de LifeSupport Não transformadas: As plantas são PCR-negativo para PPO e NPTII
Co-transformado: PCR-positivo para ambos o gene GUS de pSIM356 e o gene NPT de LifeSupport Não transformadas: As plantas são PCR-negativo para PPO e NPTII
Claims (17)
1. Método Para Modificar Característica de Planta Selecionada, integrando estavelmente apenas um polinucleotídeo desejado no genoma da referida planta, compreendendo (a) transformar a referida planta com referido polinucleotídeo pretendido, caracterizado por que o referido polinucleotídeo pretendido consiste essencialmente (i) numa sequência de ácido nucleico que é nativa para planta selecionada, nativa para uma planta da mesma espécie como a planta selecionada ou nativa para uma planta que seja sexualmente interfértil com a referida planta selecionada, em que (ii) o polinucleotídeo pretendido não contém DNA estranho que não seja da espécie de planta selecionada ou uma planta que seja sexualmente compatível com as espécies de plantas selecionadas e (iii) o polinucleotídeo pretendido não compreende uma sequência de nucleotídeos que seja idêntica na sequência de nucleotídeos a uma sequência de borda de Agrobacterium T-DNA; (b) selecionar células de plantas compreendendo apenas o referido polinucleotídeo pretendido no genoma; e (c) obter uma planta transformada de modo estável a partir das células de plantas transformadas selecionadas, em que a planta transformada contém apenas o polinucleotídeo pretendido integrado de modo estável em seu genoma e em que a planta transformada de modo estável exibe uma característica que é modificada em comparação com a planta selecionada, em que o polinucleotídeo pretendido compreende pelo menos uma sequência de nucleotídeos de SEQ ID NO: 1.
2. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 1, compreendendo ainda a co-introdução de um gene marcador selecionável nas referidas células de plantas para expressão transitória e (b) identificação de células de plantas transformadas e plantas transformadas obtidas a partir das referidas células de plantas transformadas, caracterizado por que o referido gene marcador selecionável está integrado de modo instável no genoma da referida planta.
3. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 2, caracterizado por que o gene marcador selecionável está ligado a um promotor e terminador a partir de um gene que é expresso de modo não natural numa célula de planta.
4. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 3, caracterizado por que (i) o gene marcador selecionável codifica a neomicina fosfotransferase, (ii) o terminador é um terminador do gene do álcool da dehidrogenase da levedura ou (iii) o gene marcador selecionável codifica a neomicina fosfotransferase e o terminador é um terminador do gene do álcool da dehidrogenase da levedura.
5. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 1, caracterizado por que a planta é uma planta monocotiledônea.
6. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 5, caracterizado por que a referida planta monocotiledônea é selecionada do grupo que consiste em trigo, relva, grama de relva, cereal, milho, arroz, aveia, trigo, cevada, sorgo, orquídea, íris, lírio, cebola, banana, cana-de-açúcar, sorgo e palma.
7. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 1, caracterizado por que a planta é uma planta dicotiledônea.
8. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 7, caracterizado por que a referida planta dicotiledônea é selecionada do grupo que consiste em batata, tabaco, tomate, beterraba sacarina, brócolis, mandioca, batata doce, pimenta, algodão, poinsétia, legumes, alfafa, soja, cenoura, morango, alface, carvalho, ácer, noz, rosa, hortelã, esprema, margarida e cactos.
9. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 1, caracterizado por que a referida característica é selecionada do grupo que consiste em saúde e características nutricionais aumentadas, armazenamento melhorado, rendimento aumentado, tolerância aumentada a sal, tolerância aumentada a metal pesado, tolerância aumentada à seca, tolerância aumentada à doença, tolerância aumentada a inseto, tolerância aumentada a stress de água, tolerância aumentada ao frio e geada, cor intensificada, doçura aumentada, vigor melhorado, gosto melhorado, textura melhorada, conteúdo de fosfato diminuído, germinação aumentada, captura aumentada de micronutrientes, composição de amido melhorada e longevidade de flor melhorada.
10. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 1, caracterizado por que o polinucleotídeo pretendido compreende pelo menos uma cópia de uma parte das sequências de região não traduzidas em 5’ ou 3’ de pelo menos um dos genes Rl, gene da polifenol oxidase e gene da fosforilase e está operacionalmente ligado a elementos reguladores que expressam o polinucleotídeo pretendido na célula de planta.
11. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 10, caracterizado por que (a) o polinucleotídeo pretendido compreende ainda duas cópias de pelo menos parte das sequências de região não traduzidas em 5’ ou 3’, (b) essas duas cópias são posicionadas como repetições invertidas uma da outra entre um promotor e um terminador e (c) a expressão da repetição invertida produz moléculas de RNA de fita dupla que facilitam a regulação negativa da expressão do gene ao qual está associado.
12. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 3, caracterizado por que o polinucleotídeo pretendido é introduzido numa célula de planta antes do gene marcador selecionável.
13. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 10, caracterizado por que a região 5’ está a montante de um códon de início de um gene e a região 3’ está a jusante do códon de parada de um gene, em que o gene é selecionado do grupo que consiste num gene PPO, um gene Rl, um gene HOS1, um gene S- adenosilhomocisteína hidrolase, um gene cinamato 4-hidroxilase da classe II, um gene cinamoil-coenzima A da reductase, um gene de deshidrogenase do álcool cinamoil, um gene cafeoil coenzima A O- metiltransferase, um gene do fator despolimerizante da actina, um gene Nin88, um gene Lol p 5, um gene alérgeno, um gene hidroxilase P450, um gene ADP glicose pirofosforilase, um gene da deshidrogenase da prolina, um gene endo-l,4-beta-glucanase, um gene zeaxantina epoxidase e um gene da sintase do 1-aminociclopropanode-l-carboxilato.
14. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 13, caracterizado por que o polinucleotídeo pretendido está num cassete de expressão, em que o cassete de expressão compreende a sequência de SEQ ID NO: 23.
15. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 1, caracterizado por que a expressão do polinucleotídeo pretendido modifica uma característica numa planta regenerada a partir da célula de planta transformada, em que a característica é pelo menos um dos (i) níveis mais baixos de acrilamida, (ii) contusões reduzidas por manchas pretas, (iii) adoçante reduzido induzido pelo frio e (iv) níveis reduzidos de fosfato de amido em comparação com uma planta que não compreende uma célula que expressa o polinucleotídeo pretendido.
16. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 1, caracterizado por que a expressão do polinucleotídeo pretendido modifica uma característica na planta obtida, em que a característica é um nível mais baixo de acrilamida.
17. Método Para Modificar Característica de Planta Selecionada, de acordo com a Reivindicação 10, caracterizado por que o polinucleotídeo pretendido compreende pelo menos uma cópia de uma parte das sequências da região não traduzidas 5’ ou 3’ de pelo menos dois do gene Rl, gene da polifenol oxidase e gene da fosforilase e está operacionalmente ligado aos elementos reguladores que expressam o polinucleotídeo pretendido na célula de planta.
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US35766102P | 2002-02-20 | 2002-02-20 | |
| US60/357,661 | 2002-02-20 | ||
| US37760202P | 2002-05-06 | 2002-05-06 | |
| US60/377,602 | 2002-05-06 | ||
| PCT/US2003/004947 WO2003069980A2 (en) | 2002-02-20 | 2003-02-20 | Precise breeding |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| BRPI0307791A2 BRPI0307791A2 (pt) | 2019-01-15 |
| BRPI0307791B1 true BRPI0307791B1 (pt) | 2020-08-11 |
Family
ID=27760470
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| BRPI0307791-8A BRPI0307791B1 (pt) | 2002-02-20 | 2003-02-20 | Métodos para modificar característica de planta selecionada |
Country Status (10)
| Country | Link |
|---|---|
| US (11) | US7250554B2 (pt) |
| EP (2) | EP1585384B1 (pt) |
| CN (1) | CN102978236B (pt) |
| AU (3) | AU2003219806B8 (pt) |
| BR (1) | BRPI0307791B1 (pt) |
| CA (2) | CA2477240C (pt) |
| ES (1) | ES2602133T3 (pt) |
| IL (2) | IL163628A0 (pt) |
| NZ (4) | NZ535395A (pt) |
| WO (1) | WO2003069980A2 (pt) |
Families Citing this family (58)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| NZ535395A (en) * | 2002-02-20 | 2009-01-31 | Simplot Co J R | Precise plant breeding using plant DNA border like sequences instead of Agrobacterium borders |
| US7534934B2 (en) * | 2002-02-20 | 2009-05-19 | J.R. Simplot Company | Precise breeding |
| EP1560484B1 (en) * | 2002-03-20 | 2011-05-11 | J.R. Simplot Company | Refined plant transformation |
| JP2004065085A (ja) * | 2002-08-06 | 2004-03-04 | Nippon Suisan Kaisha Ltd | 緑色が変色しにくい枝豆 |
| US7575917B2 (en) | 2003-04-09 | 2009-08-18 | Monsanto Technology Llc | DNA constructs and methods to enhance the production of commercially viable transgenic plants |
| US20050196504A1 (en) * | 2004-03-04 | 2005-09-08 | Finley John W. | Reduction of acrylamide in processed foods |
| WO2005121346A1 (en) * | 2004-06-08 | 2005-12-22 | New Zealand Institute For Crop & Food Research | Transformation vectors |
| WO2006017920A2 (en) * | 2004-08-18 | 2006-02-23 | Alellyx S.A. | Polynucleotides, dna constructs and methods for the alteration of plant lignin content and/or composition |
| US8137961B2 (en) * | 2004-09-08 | 2012-03-20 | J.R. Simplot Company | Plant-specific genetic elements and transfer cassettes for plant transformation |
| WO2006029076A2 (en) * | 2004-09-08 | 2006-03-16 | J.R. Simplot Company | Plant-specific genetic elements and transfer cassettes for plant transformation |
| EP1799028B1 (en) * | 2004-09-24 | 2015-03-04 | J.R. Simplot Company | Gene silencing |
| KR101413122B1 (ko) | 2005-02-28 | 2014-07-18 | 22엔디 센츄리 리미티드, 엘엘씨 | 식물의 니코틴 알칼로이드 수준의 감소 |
| US7611898B2 (en) * | 2005-05-09 | 2009-11-03 | The Samuel Roberts Noble Foundation | Agrobacterium transformation of stolons |
| EP1915453B1 (en) * | 2005-07-27 | 2010-12-01 | J.R. Simplot Company | Marker-free plant transformation |
| US20070074313A1 (en) * | 2005-09-16 | 2007-03-29 | J.R. Simplot Company | Native antibiotic resistance genes |
| WO2007035752A2 (en) | 2005-09-20 | 2007-03-29 | J.R. Simplot Company | Low acrylamide foods |
| US7923600B2 (en) | 2006-02-06 | 2011-04-12 | J.R. Simplot Company | Generation of marker-free and backbone-free transgenic plants using a single binary approach |
| US20070250948A1 (en) * | 2006-03-07 | 2007-10-25 | J.R. Simplot Company | Plant-specific genetic-elements and transfer cassettes for plant transformation |
| PL2038300T3 (pl) | 2006-07-06 | 2014-07-31 | Simplot Co J R | Artykuły żywnościowe zawierające wysoki poziom przeciwutleniaczy |
| DE602007012269D1 (de) * | 2006-08-31 | 2011-03-10 | Monsanto Technology Llc | Verfahren zur herstellung transgener pflanzen |
| WO2007072224A2 (en) | 2006-09-13 | 2007-06-28 | Nara Institute Of Science And Technology | Increasing levels of nicotinic alkaloids |
| US9551003B2 (en) * | 2006-09-13 | 2017-01-24 | 22Nd Century Limited, Llc | Increasing levels of nicotinic alkaloids in plants |
| WO2008046189A1 (en) * | 2006-10-11 | 2008-04-24 | Her Majesty The Queen In Right Of Canada, As Represented By The Minister Of The Province Of Nova Scotia, The Nova Scotia Agricultural College (Nsac) | Proteins involved in after-cooking darkening in potatoes |
| US9102948B2 (en) | 2006-11-17 | 2015-08-11 | 22Nd Century Limited, Llc | Regulating alkaloids |
| US20090123626A1 (en) * | 2007-09-28 | 2009-05-14 | J.R. Simplot Company | Reduced acrylamide plants and foods |
| US20100257632A1 (en) * | 2007-12-05 | 2010-10-07 | Keygene N.V. | Methods for generating marker-free transgenic plants |
| EP2300617B1 (en) | 2008-07-16 | 2016-03-23 | Monsanto Technology LLC | Methods and vectors for producing transgenic plants |
| WO2010091018A1 (en) * | 2009-02-03 | 2010-08-12 | Wisconsin Alumni Research Foundation | Control of cold-induced sweetening and reduction of acrylamide levels in potato or sweet potato |
| US8604279B2 (en) | 2009-10-19 | 2013-12-10 | J.R. Simplot Company | Glyphosate tolerant perennial ryegrass named ‘JS501’ |
| US8604280B2 (en) | 2009-10-19 | 2013-12-10 | J.R. Simplot Company | Glyphosate tolerant perennial ryegrass named ‘replay’ |
| US20130219560A1 (en) * | 2010-09-02 | 2013-08-22 | Richard T. Sayre | Reduction of post-harvest physiological deterioration |
| CA2867139A1 (en) | 2011-04-11 | 2012-10-18 | Targeted Growth, Inc. | Identification and the use of krp mutants in plants |
| US9797004B2 (en) | 2012-06-12 | 2017-10-24 | Syngenta Participations Ag | Methods and compositions for determination of vector backbone in a nucleic acid sample |
| US9253952B2 (en) | 2012-08-17 | 2016-02-09 | University Of Copenhagen | Agrobacterium rhizogenes transformation and expression of rol genes in Kalanchoë |
| US10504132B2 (en) * | 2012-11-27 | 2019-12-10 | American Express Travel Related Services Company, Inc. | Dynamic rewards program |
| WO2014153032A1 (en) | 2013-03-14 | 2014-09-25 | Evolutionary Genomics, Inc. | Identification and use of tomato genes controlling salt/drought tolerance and fruit sweetness |
| PH12015502511B1 (en) * | 2013-05-02 | 2023-09-20 | Simplot Co J R | Potato cultivar e12 |
| HRP20221172T1 (hr) | 2013-10-29 | 2023-02-03 | Biotech Institute, Llc | Uzgoj, proizvodnja, prerada i uporaba specijalnog sinsemilla kanabisa |
| US8889964B1 (en) | 2014-06-17 | 2014-11-18 | J.R. Simplot Company | Potato cultivar W8 |
| CA2951526A1 (en) | 2014-06-17 | 2015-12-23 | J.R. Simplot Company | Potato cultivar w8 |
| EP2977458A1 (en) * | 2014-07-23 | 2016-01-27 | Rheinische Friedrich-Wilhelms-Universität Bonn | Improved strains of Agrobacterium tumefaciens for transferring DNA into plants |
| CN104911180B (zh) * | 2014-12-22 | 2017-11-14 | 华中农业大学 | 马铃薯类t‑dna元件与同源转基因体系及其应用 |
| US9918441B2 (en) | 2015-05-14 | 2018-03-20 | J.R. Simplot Company | Potato cultivar V11 |
| CN108347894B (zh) | 2015-10-08 | 2019-10-29 | 杰.尔.辛普洛公司 | 马铃薯栽培品种y9 |
| KR20180059467A (ko) | 2015-10-08 | 2018-06-04 | 제이.알.심프롯캄패니 | 감자 종자 x17 |
| WO2017185136A1 (en) * | 2016-04-27 | 2017-11-02 | Nexgen Plants Pty Ltd | Construct and vector for intragenic plant transformation |
| CN106258244A (zh) * | 2016-08-15 | 2017-01-04 | 仁怀市助惠蔬菜种植专业合作社 | 一种红薯薯种的越冬贮藏方法 |
| US11332752B2 (en) | 2018-03-12 | 2022-05-17 | Pioneer Hi-Bred International, Inc. | Use of morphogenic factors for the improvement of gene editing |
| US11946057B2 (en) | 2018-12-20 | 2024-04-02 | Benson Hill, Inc. | Pre-conditioning treatments to improve plant transformation |
| CN111139261B (zh) * | 2019-02-28 | 2022-05-31 | 山东省农业科学院作物研究所 | 一种利用基因编辑降低小麦籽粒多酚氧化酶含量的方法 |
| CA3187967A1 (en) | 2020-06-24 | 2021-12-30 | Benson Hill, Inc. | Plant cell treatments to improve plant transformation |
| CN112251460A (zh) * | 2020-10-28 | 2021-01-22 | 中国农业科学院蔬菜花卉研究所 | 一种萝卜功能基因组研究及基因功能验证的方法 |
| US11530419B2 (en) | 2020-10-30 | 2022-12-20 | Fortiphyte, Inc. | Pathogen resistance in plants |
| CN114793792B (zh) * | 2022-04-24 | 2023-06-20 | 河南科技学院 | 一种bns系列温敏核不育小麦不同育性群体花粉育性恢复的方法 |
| US20240093220A1 (en) | 2022-09-09 | 2024-03-21 | Friedrich Alexander Universität Erlangen-Nürnberg | Plant regulatory elements and uses thereof |
| US12225874B2 (en) | 2023-03-27 | 2025-02-18 | Redsea Science And Technology Inc. | Tomato plant designated ‘X22-31’ |
| US20250075226A1 (en) | 2023-08-29 | 2025-03-06 | University Of Freiburg | Proteins for regulation of symbiotic infection and associated regulatory elements |
| US20250382627A1 (en) | 2024-06-13 | 2025-12-18 | J.R. Simplot Company | Engineering remontant flowering in rosaceae |
Family Cites Families (84)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6051757A (en) | 1983-01-14 | 2000-04-18 | Washington University | Regeneration of plants containing genetically engineered T-DNA |
| US6174724B1 (en) | 1983-01-17 | 2001-01-16 | Monsanto Company | Chimeric genes suitable for expression in plant cells |
| NL8300699A (nl) | 1983-02-24 | 1984-09-17 | Univ Leiden | Werkwijze voor het inbouwen van vreemd dna in het genoom van tweezaadlobbige planten; werkwijze voor het produceren van agrobacterium tumefaciens bacterien; stabiele cointegraat plasmiden; planten en plantecellen met gewijzigde genetische eigenschappen; werkwijze voor het bereiden van chemische en/of farmaceutische produkten. |
| NL8300698A (nl) | 1983-02-24 | 1984-09-17 | Univ Leiden | Werkwijze voor het inbouwen van vreemd dna in het genoom van tweezaadlobbige planten; agrobacterium tumefaciens bacterien en werkwijze voor het produceren daarvan; planten en plantecellen met gewijzigde genetische eigenschappen; werkwijze voor het bereiden van chemische en/of farmaceutische produkten. |
| US5231019A (en) * | 1984-05-11 | 1993-07-27 | Ciba-Geigy Corporation | Transformation of hereditary material of plants |
| US5149645A (en) * | 1984-06-04 | 1992-09-22 | Rijksuniversiteit Leiden | Process for introducing foreign DNA into the genome of plants |
| US4658082A (en) * | 1984-07-25 | 1987-04-14 | Atlantic Richfield Company | Method for producing intact plants containing foreign DNA |
| EP0174166A1 (en) | 1984-08-31 | 1986-03-12 | Lubrizol Genetics Inc. | Disarmed T-DNA |
| US4743548A (en) | 1984-09-25 | 1988-05-10 | Calgene, Inc. | Plant cell microinjection technique |
| US4945050A (en) * | 1984-11-13 | 1990-07-31 | Cornell Research Foundation, Inc. | Method for transporting substances into living cells and tissues and apparatus therefor |
| US5182200A (en) * | 1985-04-22 | 1993-01-26 | Lubrizol Genetics, Inc. | T-dna promoters |
| US5221623A (en) * | 1986-07-22 | 1993-06-22 | Boyce Thompson Institute For Plant Research, Inc. | Use of bacterial luciferase structural genes for cloning and monitoring gene expression in microorganisms and for tagging and identification of genetically engineered organisms |
| EP0257472A3 (en) | 1986-08-14 | 1989-10-04 | Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. | Transgenic monocotyledonous plants, seeds thereof and process for the preparation of the plants |
| EP0265556A1 (en) | 1986-10-31 | 1988-05-04 | Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. | Stable binary agrobacterium vectors and their use |
| US5856177A (en) * | 1987-11-18 | 1999-01-05 | Mycogen Corporation | Promoters derived from the maize phosphoenolpyruvate carboxylase gene involved in C4 photosynthesis |
| US5990387A (en) * | 1988-06-10 | 1999-11-23 | Pioneer Hi-Bred International, Inc. | Stable transformation of plant cells |
| US5629183A (en) | 1989-05-08 | 1997-05-13 | The United States Of America As Represented By The Secretary Of Agriculture | Plant transformation by gene transfer into pollen |
| US5302523A (en) | 1989-06-21 | 1994-04-12 | Zeneca Limited | Transformation of plant cells |
| US5225341A (en) | 1990-07-19 | 1993-07-06 | The Regents Of The University Of California | Biologically safe plant transformation system using a ds transposon |
| US6403865B1 (en) * | 1990-08-24 | 2002-06-11 | Syngenta Investment Corp. | Method of producing transgenic maize using direct transformation of commercially important genotypes |
| NL9002116A (nl) | 1990-09-27 | 1992-04-16 | Clovis Matton N V | Werkwijze voor het genetisch manipuleren van plantecellen, recombinante plasmiden, recombinante bacterien, planten. |
| NZ239977A (en) | 1990-11-14 | 1993-08-26 | Pioneer Hi Bred Int | Transforming plants by the use of agrobacterium |
| GB9304200D0 (en) | 1993-03-02 | 1993-04-21 | Sandoz Ltd | Improvements in or relating to organic compounds |
| US5324646A (en) | 1992-01-06 | 1994-06-28 | Pioneer Hi-Bred International, Inc. | Methods of regeneration of Medicago sativa and expressing foreign DNA in same |
| US6160204A (en) | 1992-01-31 | 2000-12-12 | Cornell Research Foundation, Inc. | Polyphenol oxidase cDNA |
| DE4220758A1 (de) * | 1992-06-24 | 1994-01-05 | Inst Genbiologische Forschung | DNA-Sequenz und Plasmide zur Herstellung von Pflanzen mit veränderter Saccharose-Konzentration |
| ATE398679T1 (de) | 1992-07-07 | 2008-07-15 | Japan Tobacco Inc | Verfahren zur transformation einer monokotyledon pflanze |
| WO1994003607A1 (en) | 1992-07-30 | 1994-02-17 | Keygene N.V. | Dna constructs, cells and plants derived therefrom |
| US5284253A (en) * | 1992-11-25 | 1994-02-08 | Atlantic Richfield Company | Stacker for electronic payment system key pads |
| US5646023A (en) * | 1993-04-15 | 1997-07-08 | J.R. Simplot Company | Modulation of sugar content in plants |
| US5648249A (en) * | 1993-05-28 | 1997-07-15 | Monsato Company | Method of improving the quality of stored potatoes |
| EP0628636A1 (en) * | 1993-06-07 | 1994-12-14 | Stichting Agrotechnologisch Onderzoek ( Ato-Dlo) | Transgenic plants exhibiting modified carbohydrate metabolism |
| US6969782B2 (en) | 1993-10-06 | 2005-11-29 | Ajinomoto Co., Inc. | Method of producing transgenic plants having improved amino acid composition |
| US5492852A (en) | 1993-10-07 | 1996-02-20 | Nec Corporation | Method for fabricating a solid imaging device having improved smear and breakdown voltage characteristics |
| US5731179A (en) * | 1993-12-08 | 1998-03-24 | Japan Tobacco Inc. | Method for introducing two T-DNAS into plants and vectors therefor |
| CA2180826A1 (en) * | 1994-01-13 | 1995-07-20 | Richard J. Ii Stoner | Organic disease control system |
| US6103893A (en) | 1994-03-25 | 2000-08-15 | National Starch And Chemical Investment Holding Corporation | High amylose starch from transgenic potato plants |
| US5859351A (en) * | 1994-04-13 | 1999-01-12 | The Regents Of The University Of California | Prf protein and nucleic acid sequences: compositions and methods for plant pathogen resistance |
| US5736369A (en) * | 1994-07-29 | 1998-04-07 | Pioneer Hi-Bred International, Inc. | Method for producing transgenic cereal plants |
| US5631152A (en) * | 1994-10-26 | 1997-05-20 | Monsanto Company | Rapid and efficient regeneration of transgenic plants |
| JP3256952B2 (ja) | 1994-11-09 | 2002-02-18 | 日本製紙株式会社 | 植物への遺伝子導入用ベクター、並びにこれを用いた遺伝子導入植物の作成方法及び植物への遣伝子多重導入方法 |
| US5767368A (en) | 1995-04-03 | 1998-06-16 | Board Of Trustees Operating Michigan State University | Method for producting a cereal plant with foreign DNA |
| PT826061E (pt) | 1995-05-05 | 2007-10-16 | Brunob Ii Bv | ''melhoramentos na composição de amido vegetal ou relacionados com o mesmo'' |
| DE59611362D1 (de) * | 1995-09-19 | 2006-08-17 | Bayer Bioscience Gmbh | Pflanzen, die eine modifizierte stärke synthetisieren, verfahren zu ihrer herstellung sowie modifizierte stärke |
| BR9610738A (pt) | 1995-09-25 | 1999-07-13 | Novartis Ag | Integração aperfeiçoada de dna exógeno transferido para as células eucarióticas |
| DE19608918A1 (de) * | 1996-03-07 | 1997-09-11 | Planttec Biotechnologie Gmbh | Nucleinsäuremoleküle, die neue Debranching-Enzyme aus Mais codieren |
| US5932783A (en) * | 1996-04-01 | 1999-08-03 | J.R. Simplot Co. | Potato UDP-glucose pyrophosphorylase gene promoters and their uses |
| WO1997040707A1 (en) | 1996-04-29 | 1997-11-06 | The Procter & Gamble Company | Process for preparing frozen par-fried potato strips having deep fried texture when oven finished |
| US6096865A (en) * | 1996-05-06 | 2000-08-01 | Amgen Inc. | Mutants of the green fluorescent protein having improved fluorescent properties at 37° |
| AUPO069996A0 (en) | 1996-06-27 | 1996-07-18 | Australian National University, The | Manipulation of plant cellulose |
| ES2251738T3 (es) | 1996-07-30 | 2006-05-01 | Universitat Heidelberg | Inhibidor de la invertasa. |
| US5981840A (en) * | 1997-01-24 | 1999-11-09 | Pioneer Hi-Bred International, Inc. | Methods for agrobacterium-mediated transformation |
| US5959180A (en) | 1997-02-07 | 1999-09-28 | The United States Of America As Represented By The Secretary Of Agriculture | DNA sequences from potato encoding solanidine UDP-glucose glucosyltransferase and use to reduce glycoalkaloids in solanaceous plants |
| US5998701A (en) * | 1997-06-04 | 1999-12-07 | Her Majesty The Queen In Right Of Canada As Represented By The Department Of Agriculture | Potatoes having improved quality characteristics and methods for their production |
| JP2001511007A (ja) * | 1997-02-10 | 2001-08-07 | カナダ国 | 低温誘発性甘味増加の減少を伴うαグルカンL−タイプまたはH−タイプ塊茎ホスホリラーゼのレベルの低下したトランスジェニックポテト |
| US6066781A (en) * | 1997-02-13 | 2000-05-23 | Applied Phytologics, Inc. | Production of mature proteins in plants |
| DE69826596T2 (de) * | 1997-02-20 | 2006-02-09 | Bayer Bioscience N.V. | Verbesserte methode zur transformation von pflanzen |
| JP3151657B2 (ja) | 1997-03-26 | 2001-04-03 | 農林水産省農業生物資源研究所長 | 遺伝子導入法 |
| CN1192110C (zh) | 1997-06-30 | 2005-03-09 | 辛根塔默根有限公司 | 用于植物转化的质粒和使用此质粒的方法 |
| US6096781A (en) * | 1997-11-14 | 2000-08-01 | Eli Lilly And Company | 2-arylbenzo[B]thiophenes useful for the treatment of estrogen deprivation syndrome |
| US6100447A (en) | 1998-02-12 | 2000-08-08 | Applied Phytologics, Inc. | Method of barley transformation |
| CN1202246C (zh) * | 1998-04-08 | 2005-05-18 | 联邦科学和工业研究组织 | 获得修饰表型的方法和措施 |
| US6620987B1 (en) | 1998-04-09 | 2003-09-16 | E. I. Dupont De Nemours & Company | Nucleic acid encoding a starch R1 phosphorylation protein homolog from maize |
| WO1999060106A2 (en) | 1998-05-20 | 1999-11-25 | Universite Du Quebec A Montreal | A universal plant promoter inducible by low- temperatures |
| CA2344700C (en) | 1998-10-01 | 2005-03-29 | Pioneer Hi-Bred International, Inc. | Method of plant transformation |
| DE19846001C2 (de) | 1998-10-06 | 2000-08-24 | Bioplant Biotechnologisches Fo | Transgene Pflanzen mit erhöhter Resistenz gegen den Befall durch Phytopathogene |
| DE19846011C2 (de) | 1998-10-06 | 2001-09-27 | Drk Blutspendedienst Nordrhein | Verfahren zur Aufreinigung von Granulozyten enthaltenden Präparaten |
| JP3571234B2 (ja) * | 1998-10-12 | 2004-09-29 | 本田技研工業株式会社 | 車両用操舵制御装置 |
| TW389394U (en) * | 1998-10-23 | 2000-05-01 | Hon Hai Prec Ind Co Ltd | Electrical connector |
| CA2348366C (en) * | 1998-11-09 | 2012-05-15 | Bayer Cropscience Aktiengesellschaft | Nucleic acid molecules from rice and their use for the production of modified starch |
| JP4394323B2 (ja) | 1999-09-30 | 2010-01-06 | 日本たばこ産業株式会社 | 植物の形質転換用ベクター |
| US6392053B2 (en) | 1999-12-15 | 2002-05-21 | Bristol-Myers Squibb Company | Process for preparing arylacetylaminothiazoles |
| US6750379B2 (en) * | 2000-03-09 | 2004-06-15 | Dekalb Genetics Corporation | Homologous recombination-mediated transgene alterations in plants |
| US20030154518A1 (en) * | 2000-06-12 | 2003-08-14 | Signer Ethan R. | Removal of selectable markers from transformed cells |
| AUPQ815400A0 (en) | 2000-06-14 | 2000-07-06 | Dairy Research And Development Corporation | Modification of lignin biosynthesis |
| EP1356065A2 (en) | 2000-11-03 | 2003-10-29 | Monsanto Technology LLC | Method of imparting disease resistance to plants by reducing polyphenol oxidase activity |
| US6492852B2 (en) * | 2001-03-30 | 2002-12-10 | International Business Machines Corporation | Pre-divider architecture for low power in a digital delay locked loop |
| FR2825579B1 (fr) * | 2001-06-11 | 2004-07-23 | Rhobio | Methode d'obtention de plante monocotyledone contenant un gene d'interet sans sequence auxiliaire etrangere |
| NZ535395A (en) | 2002-02-20 | 2009-01-31 | Simplot Co J R | Precise plant breeding using plant DNA border like sequences instead of Agrobacterium borders |
| US7534934B2 (en) | 2002-02-20 | 2009-05-19 | J.R. Simplot Company | Precise breeding |
| US20060233930A1 (en) * | 2002-11-08 | 2006-10-19 | Bayer Cropscience Gmbh | Process for reducing the acrylamide content of heat-treated foods |
| EP1799028B1 (en) * | 2004-09-24 | 2015-03-04 | J.R. Simplot Company | Gene silencing |
| WO2007035752A2 (en) | 2005-09-20 | 2007-03-29 | J.R. Simplot Company | Low acrylamide foods |
| US20090123626A1 (en) * | 2007-09-28 | 2009-05-14 | J.R. Simplot Company | Reduced acrylamide plants and foods |
-
2003
- 2003-02-20 NZ NZ535395A patent/NZ535395A/en not_active IP Right Cessation
- 2003-02-20 CA CA2477240A patent/CA2477240C/en not_active Expired - Lifetime
- 2003-02-20 WO PCT/US2003/004947 patent/WO2003069980A2/en not_active Ceased
- 2003-02-20 NZ NZ585768A patent/NZ585768A/en not_active IP Right Cessation
- 2003-02-20 US US10/369,324 patent/US7250554B2/en not_active Ceased
- 2003-02-20 EP EP03716081A patent/EP1585384B1/en not_active Expired - Lifetime
- 2003-02-20 IL IL16362803A patent/IL163628A0/xx unknown
- 2003-02-20 ES ES10008266.8T patent/ES2602133T3/es not_active Expired - Lifetime
- 2003-02-20 EP EP10008266.8A patent/EP2248902B1/en not_active Expired - Lifetime
- 2003-02-20 CN CN201210432123.3A patent/CN102978236B/zh not_active Expired - Lifetime
- 2003-02-20 BR BRPI0307791-8A patent/BRPI0307791B1/pt active IP Right Grant
- 2003-02-20 NZ NZ585399A patent/NZ585399A/en not_active IP Right Cessation
- 2003-02-20 AU AU2003219806A patent/AU2003219806B8/en not_active Expired
- 2003-02-20 US US10/505,079 patent/US20050229267A1/en not_active Abandoned
- 2003-02-20 NZ NZ571718A patent/NZ571718A/en not_active IP Right Cessation
- 2003-02-20 CA CA2775650A patent/CA2775650C/en not_active Expired - Lifetime
-
2004
- 2004-08-19 IL IL163628A patent/IL163628A/en active IP Right Grant
-
2006
- 2006-03-15 US US11/375,095 patent/US7619138B2/en not_active Expired - Lifetime
-
2007
- 2007-01-31 US US11/700,169 patent/US7880057B2/en not_active Expired - Lifetime
- 2007-01-31 US US11/700,101 patent/US7947868B2/en not_active Expired - Lifetime
- 2007-01-31 US US11/700,170 patent/US8193412B2/en not_active Expired - Lifetime
-
2009
- 2009-05-06 US US12/436,389 patent/US8252974B2/en not_active Expired - Fee Related
-
2010
- 2010-11-05 AU AU2010241331A patent/AU2010241331A1/en not_active Abandoned
- 2010-12-03 US US12/960,033 patent/US8143477B2/en not_active Expired - Fee Related
- 2010-12-22 AU AU2010257344A patent/AU2010257344A1/en not_active Abandoned
-
2012
- 2012-05-25 US US13/480,970 patent/US8502027B2/en not_active Expired - Fee Related
- 2012-06-04 US US13/488,024 patent/US20130254938A1/en not_active Abandoned
-
2013
- 2013-08-07 US US13/961,434 patent/USRE45722E1/en not_active Expired - Lifetime
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| EP2248902B1 (en) | Precise breeding | |
| US8273949B2 (en) | Precise breeding | |
| AU2013200775B2 (en) | Precise breeding |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| B06H | Technical and formal requirements: requirement cancelled [chapter 6.8 patent gazette] |
Free format text: ANULADA A EXIGENCIA CODIGO 6.7 PUBLICADA NA RPI NO 1871 DE 14/11/2006, POR TER SIDO INDEVIDA. |
|
| B06U | Preliminary requirement: requests with searches performed by other patent offices: procedure suspended [chapter 6.21 patent gazette] | ||
| B16A | Patent or certificate of addition of invention granted [chapter 16.1 patent gazette] |
Free format text: PRAZO DE VALIDADE: 10 (DEZ) ANOS CONTADOS A PARTIR DE 11/08/2020, OBSERVADAS AS CONDICOES LEGAIS. |