BRPI0820483B1 - Variantes de glicoamilase com propriedades alteradas e composição compreendendo as mesmas - Google Patents
Variantes de glicoamilase com propriedades alteradas e composição compreendendo as mesmas Download PDFInfo
- Publication number
- BRPI0820483B1 BRPI0820483B1 BRPI0820483-7A BRPI0820483A BRPI0820483B1 BR PI0820483 B1 BRPI0820483 B1 BR PI0820483B1 BR PI0820483 A BRPI0820483 A BR PI0820483A BR PI0820483 B1 BRPI0820483 B1 BR PI0820483B1
- Authority
- BR
- Brazil
- Prior art keywords
- glucoamylase
- seq
- trga
- variants
- sequence
- Prior art date
Links
Images

















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/24—Hydrolases (3) acting on glycosyl compounds (3.2)
- C12N9/2402—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1)
- C12N9/2405—Glucanases
- C12N9/2408—Glucanases acting on alpha -1,4-glucosidic bonds
- C12N9/2411—Amylases
- C12N9/2428—Glucan 1,4-alpha-glucosidase (3.2.1.3), i.e. glucoamylase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/02—Monosaccharides
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/14—Preparation of compounds containing saccharide radicals produced by the action of a carbohydrase (EC 3.2.x), e.g. by alpha-amylase, e.g. by cellulase, hemicellulase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/02—Preparation of oxygen-containing organic compounds containing a hydroxy group
- C12P7/04—Preparation of oxygen-containing organic compounds containing a hydroxy group acyclic
- C12P7/06—Ethanol, i.e. non-beverage
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02E—REDUCTION OF GREENHOUSE GAS [GHG] EMISSIONS, RELATED TO ENERGY GENERATION, TRANSMISSION OR DISTRIBUTION
- Y02E50/00—Technologies for the production of fuel of non-fossil origin
- Y02E50/10—Biofuels, e.g. bio-diesel
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Zoology (AREA)
- Engineering & Computer Science (AREA)
- Wood Science & Technology (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
VARIANTES DE GLICOAMILASE COM PROPRIEDADES ALTERADAS. A presente invenção refere-se a variantes de uma glicoamilase parental tendo propriedades alteradas (por exemplo, termoestabilidade melhorada e/ou atividade específica). Em particular, a presente descrição fornece composições compreendendo as glicoamilases variantes, incluindo composições de hidrólise de amido e composições de limpeza. A descrição também 0se relaciona a construtos de DNA que codificam as variantes e métodos de produção das variantes de glicoamilase em células hospedeiras.
Description
Este pedido de patente reivindica benefício sobre o Pedido de Patente Provisória norte-americana No 60/989.426 depositado em 20 de novembro de 2007.
Também está anexada uma listagem de sequência compreendendo as SEQ ID NOs: 1 a 167, a qual está neste pedido incorporada por referência em sua totalidade.
Variantes de glicoamilase vantajosamente têm propriedades alteradas (por exemplo, termoestabilidade e/ou atividade específica melhorada). Composições compreendendo as glicoamilases variantes, construtos de DNA que codificam as variantes, e métodos de produção das variantes de glicoa- milase em células hospedeiras são fornecidos.
Enzimas glicoamilase (glican 1,4-α-glico-hidrolases, EC 3.2.1.3) são carbohidrases de exoatuação que hidrolisam amido, as quais catalisam a remoção de sucessivas unidades de glicose de extremidades de amido não reduzidas ou moléculas de oligo e polissacarídeos relacionadas. Glicoamilases podem hi- drolisar ligações glicosídicas tanto lineares quanto ramificadas de amido (por exemplo, amilose e amilopectina).
Glicoamilases são produzidas por numerosas cepas de bactérias, fungos, levedura e plantas. Particularmente interessante, e comercialmente im-portante, glicoamilases são enzimas fúngicas que são produzidas extracelular- mente, por exemplo, em cepas de Aspergillus (Svensson et al. (1983) Carlsberg Res. Commun. 48:529-544; Boel et al., (1984) EMBO J. 3:1097-1102; Hayashida et al., (1989) Agric. Biol. Chem. 53:923-929; Patente Norte-americana No 5.024.941; Patente Norte-americana No 4.794.175 e WO 88/09795); Tala- romyces (Patente Norte-americana No 4.247.637; Patente Norte-americana No 6.255.084 e Patente Norte-americana No 6.620.924); Rhizopus (Ashikari et al., (1986) Agric. Biol. Chem. 50:957-964; Ashikari et al., (1989) App. Mi-crobiol. Biotech. 32:129-133 e Patente Norte-americana Ne 4.863.864); Hu-micola (\NO 05/052148 e Patente Norte-americana NQ 4.618.579) e Mucor (Houghton-Larsen et al., (2003) Appl. Microbiol. Biotechnol. 62:210-217). Muitos dos genes que codificam para estas enzimas foram clonados e ex-pressos em células de levedura, fúngicas e/ou bacterianas.
Comercialmente, glicoamilases são enzimas muito importantes e têm sido usadas em uma larga variedade de aplicações que necessitam de hidrólise de amido (por exemplo, para produzir glicose e outros monossaca- rídeos a partir de amido). Glicoamilases são usadas para produzir adoçantes de milho de alta frutose, que compreendem mais de 50% do mercado de adoçante nos Estados Unidos. Em geral, as glicoamilases podem ser, e co- mumente são usadas com alfa amilases em processos de hidrólise de amido para hidrolisar o amido a dextrinas e então glicose. A glicose então pode ser convertida em frutose por outras enzimas (por exemplo, glicose isomerases); cristalizada; ou usada em fermentações para produzir numerosos produtos finais (por exemplo, etanol, ácido cítrico, ácido láctico, succinato, intermediá-rios de ácido ascórbico, ácido glutâmico, glicerol e 1,3-propanodiol). Etanol produzido usando glicoamilases na fermentação de amido e/ou material con-tendo celulose pode ser usado como uma fonte de combustível ou para con-sumo alcoólico.
Embora glicoamilases tenham sido usadas com sucesso em a- plicações comerciais por muitos anos, existe ainda uma necessidade de no-vas glicoamilases com propriedades alteradas, tais como atividade específica melhorada e termoestabilidade aumentada.
Diferentes mutações foram feitas em glicoamilases de Aspergillus que aumentam estabilidade térmica e atividade específica. Referência é feita à Patente Norte-americana Ns 6.537.792; Patente Norte-americana Ns 6.352.851; Chen et al. (1996) Prot. Eng. 9:499-505, Chen et al., (1995) Prot Eng. 8:575-582; Fierobe et al. (1996) Biochem. 35:8698-8704; e Li et al., (1997) Prot. Eng. 10:1199-1204. Existe ainda necessidade de fornecer vari-antes de glicoamilase com propriedades alteradas em relação ao seu paren- tal.
A presente descrição relaciona-se a variantes de glicoamilase de uma glicoamilase parental. As variantes de glicoamilase contêm substituições de aminoácidos no domínio catalítico e/ou no domínio de ligação ao amido. As variantes exibem propriedades alteradas, tais como termoestabili- dade e/ou atividade específica melhoradas.
Em um aspecto, a presente descrição relaciona-se a uma variante de glicoamilase compreendendo duas ou mais substituições de aminoácidos correspondendo à posição 61, 73, 417, 430, 431, 503, 511, 535, 539, ou 563 da SEQ ID NO: 2, ou posição equivalente em uma glicoamilase parental. Em outro aspecto, a presente descrição relaciona-se a uma variante de glicoamilase tendo pelo menos identidade de sequência de 80%, 85%, 90%, 95%, ou 99,5% com uma glicoamilase parental das SEQ ID NO: 1, 2, 3, 5, 6, 7, 8, ou 9. Em uma modalidade, a glicoamilase parental tem um domínio catalítico com a identidade de sequência de pelo menos 80% com SEQ ID NO: 1, 2, 3, 5, 6, 7, 8, ou 9, ou um domínio de ligação a amido com a identidade de sequência de pelo menos 80% com SEQ ID NO: 1 ou 2. Em outro aspecto, a glicoamilase parental é SEQ ID NO: 1 ou 2. Um aspecto adicional da presente descrição relaciona-se à variante de glicoamilase compreendendo ainda uma ou mais substituições de aminoácidos correspondendo à posição: 4, 5, 12, 24, 43, 44, 45, 46, 47, 49, 51, 70, 75, 6, 94, 100, 108, 114, 116, 119, 122, 124, 125, 137, 141, 143, 146, 148, 169, 171, 172, 175, 178, 180, 181, 208, 211, 228, 242, 243, 245, 292, 294.197, 309, 310, 313, 314, 315, 316, 317, 321, 340, 341, 350, 353, 356, 363, 368, 369, 375, 376, 395, 398, 401, 408, 409, 412, 415, 418, 421, 433, 436 ou 451 da SEQ ID NO: 2, ou uma posição equivalente na glicoamilase parental. Em alguns aspectos, a variante de glicoamilase compreende ainda uma ou mais substituições de aminoácidos correspondendo à posição: 4, 5, 24, 29, 43, 44, 49, 70, 75, 76, 100, 108, 119, 124, 137, 146, 148, 169, 171, 172, 175, 178, 181, 208, 211, 243, 292, 294, 297, 314, 316, 317, 340, 341, 350, 356, 363, 368, 369, 376, 395, 401, 412, 433, 436 ou 451 da SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental. Em alguns aspectos, a glicoamilase compre-ende ainda uma ou mais substituições de aminoácidos correspondendo à posição: 5, 24, 43, 44, 49, 70, 75, 76, 94, 119, 141, 146, 148, 172, 175, 178, 180, 181, 208, 211, 243, 294, 309, 314, 353, 369, 375, ou 409 da SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental. Em alguns aspectos, a glicoamilase compreende ainda uma ou mais substituições de aminoácidos correspondendo à posição: 43, 44, ou 294 da SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental.
Em aspectos adicionais da invenção, a variante de glicoamilase compreende duas ou mais substituições de aminoácidos correspondendo à posição: N61I, G73F, L417R/V, T430A/M, A431L/Q, E503A/V, Q511H, A535R, A539R, ou N563I/K da SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental. Em alguns aspectos, a variante de glicoamilase compreende ainda uma ou mais das seguintes substituições: D4L/E/R/S/C/A/Q/W, F5C/M/N/R/SfT/V/W, I12L/R, D24E/L/Y/T,F29L/I/D/C/S/V/W, I43F/R/D/Y/S/Q, D44E/H/K/S/N/Y/F/R/C, Y47W, Y49N, Q70R/K/M/P/G/L/F, Q75R/K/A, R76L/M/K/T/P, P94L, D100W/I/Q/M/P/A/N, N119P/T/Y/D/E, N146S/G/C/H/E/D/T/W/L/F/M, Q148V/Y/H/A/C/D/G/M/R/S/T, Y169D/F, Q172C/A/D/R/E/F/H/V/L/M/N/S/T/V, F175H/A/G/R/S/T/C/W/Y, W178A/C/D/E/F/G/H/K/N/R/S/T/V/Y, E180A/C/G/H/I/L/N/P/Q/R/S/T/V/Y/, V181 E/C/D/G/H/l/P/T/Y/S/L/K/F/A, Q208L/A/C/E/N/F/H/T,S211C/R/E/A/Y/W/M/H/L/I/R/Q/T, E243S/R/N/M/Y/A/L, R245A/E/M/I/P/V, I292D/H/P/R/T/N/V/F/L, G294C/D/E/T/Q/1/A,K297F/L/P/T/M/D/N/Q/A/Y/H/S/R/W, R309A/C/G/H/I/N/P/Q/S/T/W/Y/L,Y310E/G/L/P/S/W/R/Q, D313Q, V314A/R/N/D/C/E/Q/G/H/I/L/K/M/F/P/S/T/W/Y, Y315F, Y316Q/R, N317T/H, K340D/T, K341F/D/P/V/G/S, T350S/E/A/N, Q356H/D/E, T363L/R/C/H/W, S368W/D/F/L, S369F, N376Q/T/H/S/V, Y395Q/R/S, A398S/I/T, S401C/V, R408S, N409W/T/K, T412A/H/K/G, R433H/Q, I436A/T, ou S451M/T/H da SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental. Em alguns aspectos, a variante de glicoamilase compreende ainda uma ou mais das seguintes substituições: I43F/R/D/Y/S/Q, D44E/H/K/S/N/Y/F/R/C, ou G294C/D/E/T/Q/I/A da SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental.
Em um aspecto, a presente descrição relaciona-se a uma glico-amilase variante compreendendo substituições de aminoácidos correspondendo a posições: I43Q/D44C, D44C/G294C, I43Q/G294C, ou I43Q/D44C/G294 da SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental. A variante de glicoamilase tem identidade de sequência de pelo menos 80%, 85%, 90%, 95%, ou 99,5% com a SEQ ID NO: 1, 2, 3, 5, 6, 7, 8, ou 9. Em uma modalidade, a glicoamilase parental tem um domínio catalítico com identidade de sequência de pelo menos 80% com a SEQ ID NO: 1, 2, 3, 5, 6, 7, 8, ou 9, ou um domínio de ligação a amido com identidade de sequência de pelo menos 80% com a SEQ ID NO: 1 ou 2.
A glicoamilase parental pode ser a enzima obtida a partir de: um Trichoderma spp., um Aspergillus spp., um Humicola spp., um Penicillium spp., um Talaromycese spp., ou um Schizosaccharmyces spp. Em alguns aspectos, a glicoamilase parental pode ser de um Trichoderma spp. ou um Aspergillus spp.
Em um aspecto, a glicoamilase variante exibe termoestabilidade alterada quando comparada com a glicoamilase parental. A termoestabilidade alterada pode ser termoestabilidade aumentada. Altemativamente, ou, além disso, a variante exibe atividade específica alterada comparada com a glicoamilase parental. A atividade específica alterada pode ser atividade es-pecífica aumentada. Um aspecto adicional da descrição é um polinucleotídeo que codifica a variante descrita. Um aspecto adicional é um vetor compreendendo o polinucleotídeo. Um aspecto adicional é uma célula hos-pedeira contendo o vetor.
Um aspecto adicional da descrição é uma composição enzimáti- ca incluindo a variante de glicoamilase. Em um aspecto, a composição en- zimática é usada em um processo de conversão de amido ou um processo de fermentação alcoólica.
Um aspecto adicional da invenção é um método de produção de uma glicoamilase variante pelo cultivo de célula hospedeira contendo polinu- cleotídeo sob condições adequadas para expressão e produção da variante de glicoamilase e produção da variante. O método também pode incluir a etapa de recuperação da variante de glicoamilase da cultura.
A FIG. 1A representa a glicoamilase parental glicoamilase de Trichoderma reesei (TrGA), tendo 632 aminoácidos (SEQ ID NO: 1). O pep-tídeo sinal é sublinhado, a região catalítica (SEQ ID NO: 3) começando com resíduos de aminoácidos SVDDFI (SEQ ID NO: 160) e tendo 453 resíduos de aminoácidos está em negrito; a região ligantea está em itálico; e o domínio de ligação a amido (SEQ ID NO: 161) está tanto em itálico como sublinhado. A proteína madura, que inclui o domínio catalítico (SEQ ID NO: 3), região ligantea, e domínio de ligação a amido (SEQ ID NO: 161), é representada pela SEQ ID NO: 2. A FIG. 1B representa o cDNA (SEQ ID NO: 4) que codifica para a TrGA.
A FIG. 2 representa o plasmídeo pDONR-TrGA que inclui o cDNA (SEQ ID NO: 4) da TrGA parental.
A FIG. 3 representa os plasmídeos pREP3Y-DEST (A) e pREP3Y-TrGA (B).
As FIGURAS 4A-4B representam uma comparação de alinhamento dos domínios catalíticos de glicoamilases parentais incluindo a glicoamilase derivada de Aspergillus awamori (AaGA) (SEQ ID NO: 5); Aspergillus niger (AnGA) (SEQ ID NO: 6); Aspergillus orzyae (AoGA) (SEQ ID NO: 7); Trichoderma reesei (TrGA) (SEQ ID NO: 3); Humicola grisea (HgGA) (SEQ ID NO: 8); e Hypocrea vinosa (HvGA) (SEQ ID NO: 9). Aminoácidos idênticos são indicados por um asterisco (*).
As FIGURAS 4C-4D representam um alinhamento que compara o Domínio de Ligação a Amido (SBD) de glicoamilases parentais incluindo Trichoderma reesei (TrGA) (SEQ ID NO: 161), Humicola grisea (HgGA) (SEQ ID NO: 162), Thielavia terrestris (TtGA) (SEQ ID NO: 163), Thermomy- ces lanuginosus (ThGA) (SEQ ID NO: 164), Talaromyces emersonii (TeGA) (SEQ ID NO: 165), Aspergillus niger (AnGA) (SEQ ID NO: 166), e Aspergillus awamori (AaGA) (SEQ ID NO: 167). Aminoácidos idênticos são indicados por um ponto (.).
A FIG. 5A representa o plasmídeo pTrex3g-DEST. A FIG. 5B representa o plasmídeo pTrex3g-TrGA. Os plasmídeos foram usados como vetores de expressão para expressão e produção de glicoamilases variantes em um hospedeiro Trichoderma reesei.
A FIG. 6 representa a comparação de Vmax (μM de glicose/s) entre a TrGA parental (tipo selvagem) e variantes, V314, S211R, Q172F, e Q208N a 60°C e 32°C como discutido ainda no Exemplo 8.
A FIG. 7 representa a atividade de variantes combinatórias sobre um substrato de amido. Variantes combinatórias descritas neste pedido in-cluem ET7-1 (D24Y/V181L/Q208C/G294A/T353R/N375N/N409W), LR8 (Q172F/Q208N), LR12 (Q172F/S211R), LR6 (Q172F/Q208N/V314H),ET8-1 (D24E/V181K/E243Y/I292V/G294Q/N409K) e ET7-2 (Q24L/V181L/Q208C/G294A/T353R/N375Q/N409W). Atividade é representada em unidades da absorção em 340 nm como uma função de ng de variantes de glicoamilase indicadas.
A FIG. 8 representa a atividade de variantes de sítio único sobre um substrato de amido de milho. Variantes de sítio único descritas neste pe-dido incluem V314H, G294Q, S211R, Q208N, Q172F, G294I e P94N. Atividade é representada em unidades de absorção em 340 nm como uma função de ng de variantes de glicoamilase indicadas.
A FIG. 9 representa a atividade de glicoamilase de TrGA e da TrGA variante LR8 (Q172F/Q208N) sobre uma amostra triturada liquefeita de milho (NE).
A FIG. 10 representa o perfil de atividade de TrGA e da TrGA variante LR8 (Q172F/Q208N) sobre uma amostra triturada liquefeita de milho (BSE).
A FIG. 11 representa o perfil de atividade de TrGA e da TrGA variante LR8 (Q172F/Q208N) sobre um substrato de amostra de amido de milho solúvel.
A FIG. 12 representa uma comparação de estruturas tridimensi-onais de glicoamilase de Trichoderma reesei (preto) (SEQ ID NO: 2) e glico- amilase de Aspergillus awamorii (cinza) vistas ao lado.
A FIG. 13 representa uma comparação de estruturas tridimensi-onais de glicoamilase de Trichoderma reesei (preto) e glicoamilase de As- pergillus awamorii (cinza) vistas no topo.
A FIG. 14A representa o plasmídeo pTTT-Dest. A FIG. 14B re-presenta o plasmídeo pTTT-TrGA (B).
Glicoamilases são enzimas comercialmente importantes em uma ampla variedade de aplicações que requerem a hidrólise de amido. Variantes de glicoamilases descritas neste pedido contêm substituições de aminoácidos dentro do domínio catalítico ou no domínio de ligação a amido. As variantes podem exibir propriedades alteradas, tais como termoestabilidade melhorada e/ou atividade específica. As variantes com termoestabilidade melhorada e/ou atividade específica podem melhorar significativamente a eficiência e produção de glicose e etanol combustível de amido de milho, por exemplo.
A menos que definido de outra maneira, todos os termos técnicos e científicos usados neste pedido têm o mesmo significado que comu- mente entendido por versado ordinário na técnica à qual esta descrição pertence. Singleton, et al., DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY, 2D ED., John Wiley and Sons, New York (1994), e Hale & Markham, THE HARPER COLLINS DICTIONARY OF BIOLOGY, Harper Perennial, N.Y. (1991) fornecem a um versado o significado geral de muitos dos termos usados neste pedido. Certos termos são definidos abaixo por causa de clareza e facilidade de referência.
Como usado neste pedido, o termo "glicoamilase (EC 3.2.1.3)" refere-se a uma enzima que catalisa a liberação de D-glicose das extremi-dades não reduzidas do amido e oligo- e polissacarídeos relacionados.
O termo "parental" ou "sequência parental" refere-se a uma se-quência que é nativa ou de ocorrência natural em uma célula hospedeira. As sequências parentais incluem, mas não são limitadas às sequências de gli- coamilase apresentadas nas SEQ ID NOs: 1, 2, 3, 5, 6, 7, 8, e 9.
Como usado neste pedido, uma "posição equivalente" significa uma posição que é comum a duas sequências parentais que é baseada em um alinhamento da sequência de aminoácido da glicoamilase parental em questão bem como alinhamento da estrutura tridimensional da glicoamilase parental em questão com a sequência de aminoácido da glicoamilase de referência TrGA (SEQ ID NO: 2) e sequência tridimensional.
O termo "TrGA" refere-se à sequência de glicoamilase parental de Trichoderma reesei tendo a sequência proteica madura ilustrada na SEQ ID NO: 2 que inclui o domínio catalítico que ilustra a sequência na SEQ ID NO: 3. O isolamento, clonagem e expressão da TrGA são descritos na Patente Norte-americana N9 7.413.887, que são incorporados neste pedido por referência. Em algumas modalidades, a sequência parental refere-se a uma glicoamilase que é o ponto de partida para engenharia de proteínas. A numeração dos aminoácidos de glicoamilase neste pedido é baseada no alinhamento de sequência de uma glicoamilase com TrGA (SEQ ID NO: 2 e SEQ ID NO: 3).
A frase "forma madura de uma proteína ou polipeptídeo" refere- se à forma funcional final da proteína ou polipeptídeo. Para exemplificar, uma forma madura da TrGA inclui o domínio catalítico, região ligantea, e domínio de ligação a amido tendo a sequência de aminoácidos da SEQ ID NO: 2.
Como usado neste pedido, os termos "variante de glicoamilase" e "variante" são usados em referência a glicoamilases que têm algum grau de identidade de sequência de aminoácido a uma sequência de glicoamilase parental e que pode conservar as características funcionais de uma glicoa-milase. Uma variante é similar a uma sequência parental, mas tem pelo menos uma substituição, deleção ou inserção em sua sequência de aminoácidos que as torna diferentes em sequência de uma glicoamilase parental. Em alguns casos, as variantes foram manipuladas e/ou engendradas para incluir pelo menos uma substituição, deleção ou inserção em sua sequência de aminoácidos que as toma diferentes em sequência de um parental.
"Variantes" podem ter pelo menos identidade de sequência de pelo menos 99,5%, pelo menos 99%, pelo menos 98%, pelo menos 97%, pelo menos 96%, pelo menos 95%, pelo menos 94%, pelo menos 93%, pelo menos 92%, pelo menos 91%, pelo menos 90%, pelo menos 88%, pelo menos 85%, pelo menos 80%, pelo menos 75%, pelo menos 70%, pelo menos 65%, pelo menos 60%, pelo menos 55%, pelo menos 50%, ou 45% a uma sequência polipeptídica quando alinhadas otimamente por comparação. Em algumas modalidades, a variante de glicoamilase pode ter identidade de sequência de pelo menos 99,5%, pelo menos 99%, pelo menos 98%, pelo menos 97%, pelo menos 96%, pelo menos 95%, pelo menos 94%, pelo menos menos 92%, menos 85%, menos 65%, pelo menos pelo menos pelo menosmenos 90%,menos 75%,menos 55%, pelo menos pelo menos pelo menos 50%, ou pelo menos 45% ao domínio catalítico de uma glicoamilase parental. Em algumas modalidades, a variante de glicoamilase pode ter identidade de sequência de pelo menos 99,5%, pelo menos 99%, pelo menos 98%, pelo menos 97%, pelo menos 96%, pelo menos 95%, pelo menos 94%, pelo menos 93%, pelo menos 92%, pelo menos 91%, pelo menos 90%, pelo menos 88%, pelo menos 85%, pelo menos 80%, pelo menos 75%, pelo menos 70%, pelo menos 65%, pelo menos 60%, pelo menos 55%, pelo menos 50%, ou pelo menos 45% ao domínio de ligação a amido de uma glicoamilase parental. A identidade de sequência pode ser medida ao longo do comprimento total da sequência parental ou da variante.
Identidade de sequência é determinada usando técnicas padrão conhecidas na técnica (ver, por exemplo, Smith and Waterman, Adv. Appl. Math. 2: 482 (1981); Needleman and Wunsch, J. Mol. Biol. 48: 443 (1970); Pearson and Lipman, Proc. Natl. Acad. Sci. USA 85: 2444 (1988); programas, tais como GAP, BESTHT, FASTA, e TFASTA no Pacote de Programas de Genética de Wisconsin (Genetics Computer Group, Madison, Wl); e Devereux et al., Nucleic Acid Res., 12: 387-395 (1984)).
O "percentual (%) de identidade de sequência de ácido nucleico" ou "percentual (%) de identidade de sequência de aminoácidos" é definido como a porcentagem de resíduos de nucleotídeo ou resíduos de aminoácidos em uma sequência candidata que é idêntica aos resíduos de nucleotídeo ou resíduos de aminoácidos da sequência inicial (por exemplo, TrGA). A identidade de sequência pode ser medida ao longo do comprimento total da sequência inicial (por exemplo, SEQ ID NO: 2).
Identidade de sequência é determinada por métodos conhecidos de alinhamento de sequência. Um método de alinhamento comumente usado é BLAST descrito por Altschul et al., (Altschul et al., J. Mol. Biol. 215: 403- 410 (1990); e Karlin et al, Proc. Natl. Acad. Sci. USA 90: 5873-5787 (1993)). Um programa BLAST particularmente útil é o programa WU-BLAST-2 (ver Altschul et al, Meth. Enzymol. 266: 460-480 (1996)). WU-BLAST-2 usa vários parâmetros de busca, a maioria dos quais são estabelecidos nos valores pré-configurados. Os parâmetros ajustáveis são definidos com os seguintes valores: amplitude de sobreposição =1, fração de sobreposição = 0,125, limiar de termo (T) = 11. Os parâmetros HSP S e HSP S2 são valores dinâmicos e são estabelecidos pelo próprio programa dependendo da composição da sequência particular e da composição do banco de dados particular contra o qual a sequência de interesse está sendo buscada. Entretanto, os valores podem ser ajustados para aumentar a sensibilidade. Um valor de % de identidade de sequência de aminoácidos é determinado pelo número de combinações com resíduos idênticos dividido pelo número total de resíduos da sequência "mais longa" na região alinhada. A sequência "mais longa" é aquela que tem os resíduos mais genuínos na região alinhada (as lacunas introduzidas por WU-Blast-2 para maximizar o escore de alinhamento são ignoradas).
Outros métodos encontram uso em sequências de alinhamento. Um exemplo de um algoritmo útil é PILEUP. PILEUP cria múltiplo alinhamento de sequência a partir de um grupo de sequências relacionadas usando alinhamentos progressivos, aos pares. Também pode traçar uma árvore mostrando as relações de agrupamento usadas para criar o alinhamento. PILEUP usa uma simplificação do método de alinhamento progressivo de Feng e Doolittle (Feng and Doolittle, J. Mol. Evol. 35: 351-360 (1987)). O mé todo é similar àquele descrito por Higgins e Sharp (Higgins and Sharp, CA- BIOS 5: 151-153 (1989)). Parâmetros de PILEUP úteis incluindo um peso de lacuna pré-configurado de 3,00, um peso de comprimento de lacuna pré- configurado de 0,10, e lacunas de extremidade ponderada.
O termo "alinhamento ótimo" refere-se ao alinhamento que fornece o escore de porcentagem de identidade mais alto.
Como usado neste pedido o termo "domínio catalítico" refere-se a uma região estrutural de um polipeptídeo, que contém o sítio ativo para hidrólise de substrato.
O termo "ligante" refere-se a uma sequência de aminoácidos curta geralmente tendo entre 3 e 40 resíduos de aminoácidos que se ligam covalentemente a uma sequência de aminoácidos compreendendo um domínio de ligação a amido com uma sequência de aminoácidos compreendendo um domínio catalítico.
O termo "domínio de ligação a amido" refere-se a uma sequência de aminoácidos que se liga preferencialmente a um substrato de amido.
Como usado neste pedido, os termos "sequência mutante" e "gene mutante" são usados intercambiavelmente e referem-se a uma se-quência polinucleotídica que tem uma alteração em pelo menos um códon que ocorre na sequência parental de uma célula hospedeira. O produto de expressão da sequência mutante é uma proteína variante com uma sequência de aminoácidos alterada em relação ao parental. O produto de expressão pode ter uma capacidade funcional alterada (por exemplo, atividade enzimá- tica aumentada).
O termo "propriedade" ou equivalentes gramaticais do mesmo no contexto de um polipeptídeo, como usado neste pedido, refere-se a qualquer característica ou atributo de um polipeptídeo que pode ser selecionado ou detectado. Estas propriedades incluem, mas não são limitadas à estabilidade oxidativa, especificidade de substrato, atividade catalítica, estabilidade térmica, perfil de atividade de pH, resistência à degradação proteolítica, KM, KCAT, razão de KCAT/KM, enovelamento proteico, capacidade de ligação a um substrato e capacidade de ser secretado.
O termo "propriedade" de equivalente gramatical do mesmo no contexto de um ácido nucleico, como usado neste pedido, refere-se a qualquer característica ou atributo de um ácido nucleico que podem ser selecionados ou detectados. Estas propriedades incluem, mas não são limitadas a uma propriedade que afeta transcrição gênica (por exemplo, força de promotor ou reconhecimento de promotor), uma propriedade que afeta processamento de RNA (por exemplo, junção de RNA e estabilidade de RNA), uma propriedade que afeta tradução (por exemplo, regulação, ligação de mRNA a proteínas ribossomais).
Os termos "termicamente estável" e "termoestável" referem-se a variantes de glicoamilase da presente descrição que conservam uma quanti-dade especificada de atividade enzimática após exposição a uma temperatura ao longo de um dado período de tempo sob condições que prevalecem durante a hidrólise de substratos de amido, por exemplo, enquanto expostos a temperaturas alteradas.
O termo "estabilidade aumentada" no contexto de uma propriedade, tal como termoestabilidade refere-se à atividade hidrolítica superior conservada de um amido ao longo do tempo comparada com outras glicoa-milases de referência (isto é, parentais).
O termo "estabilidade diminuída" no contexto de uma propriedade, tal como termoestabilidade refere-se a uma atividade hidrolítica inferior de amido conservada ao longo do tempo comparada com outras glicoamilases de referência.
O termo "atividade específica" é definido como a atividade por mg de proteína glicoamilase. Em algumas modalidades, a atividade de glico-amilase é determinada pelo ensaio de etanol descrito neste pedido e expressa como a quantidade de glicose que é produzida a partir do substrato de amido. Em algumas modalidades, a concentração proteica pode ser determinada usando o ensaio de Compasso de calibre descrito neste pedido.
Os termos "ativo" e "biologicamente ativo" referem-se a uma ati-vidade biológica associada a uma proteína particular. Segue-se que a ativi-dade biológica de uma dada proteína refere-se a qualquer atividade biológi ca tipicamente atribuída àquela proteína pelos versados na técnica. Por e- xemplo, uma atividade enzimática associada com uma glicoamilase é hidrolí- tica e, dessa forma uma glicoamilase ativa tem atividade hidrolítica.
Os termos "polinucleotídeo" e "ácido nucleico", usados intercam- biavelmente neste pedido, referem-se a uma forma polimérica de nucleotídeos de qualquer comprimento, ou ribonucleotídeos ou desoxirribonucleotí- deos. Estes termos incluem, mas não são limitados a um DNA de fita única, dupla ou tripla, DNA genômico, cDNA, RNA, híbrido de DNA-RNA, ou um polímero compreendendo bases de purina e pirimidina, ou outras bases nu- cleotídicas naturais, quimicamente, bioquimicamente modificadas, não naturais ou derivatizadas.
Como usado neste pedido, os termos "construto de DNA", "DNA de transformação" e "vetor de expressão" são usados intercambiavelmente para referir-se a DNA usado para introduzir sequências em uma célula ou organismo hospedeiro. O DNA pode ser gerado in vitro por PCR ou qualquer outra técnica(s) adequada conhecida por aqueles na técnica. O construto de DNA, DNA de transformação ou cassete de expressão recombinante pode ser incorporado em um plasmídeo, cromossomo, DNA mitocondrial, DNA plastidial, vírus, ou fragmento de ácido nucleico. Tipicamente, a porção de cassete de expressão recombinante de um vetor de expressão, construto de DNA ou DNA de transformação inclui, entre outras sequências, uma se-quência de ácido nucleico a ser transcrita e um promotor. Em algumas mo-dalidades, os vetores de expressão têm a capacidade de incorporar e ex-pressar fragmentos de DNA heterólogos em uma célula hospedeira.
Como usado neste pedido, o termo "vetor" refere-se a um cons-truto polinucleotídico desenhado para introduzir ácidos nucleicos em um ou mais tipos celulares. Os vetores incluem vetores de clonagem, vetores de expressão, vetores de transporte, plasmídeos, cassetes e similares.
Como usado neste pedido, no contexto de introdução de uma sequência de ácido nucleico em uma célula, o termo "introduzido" refere-se a qualquer método adequado para transferir a sequência de ácido nucleico para a célula. Tais métodos de introdução incluem, mas não são limitados à fusão, transfecção, transformação, conjugação e transdução de protoplasto.
Como usado neste pedido, os termos "transformado" e "trans-formado estavelmente" referem-se a uma célula que tem uma sequência polinucleotídica não nativa (heteróloga) integrada em seu genoma ou como um plasmídeo epissomal que é mantido por pelo menos duas gerações.
Como usado neste pedido, os termos "marcador selecionável" e "marcador seletivo" referem-se a um ácido nucleico (por exemplo, um gene) capaz de expressão em células hospedeiras que permitem a facilidade de seleção daqueles hospedeiros contendo o vetor. Tipicamente, os marcadores selecionáveis são genes que conferem resistência antimicrobiana ou uma vantagem metabólica na célula hospedeira para permitir a células contendo o DNA exógeno serem distinguidas de células que não receberam nenhuma sequência exógena durante a transformação.
Como usado neste pedido, o termo "promotor" refere-se a uma sequência de ácido nucleico que funciona para dirigir a transcrição de um gene a jusante. O promotor, em conjunto com outras sequências de ácidos nucleicos regulatórias transcricionais e traducionais (também denominadas "sequências controle") é necessário para expressar um dado gene. Em geral, as sequências regulatórias transcricionais e traducionais incluem, mas não são limitadas a sequências promotoras, sítios de ligação ribnssomal, sequências de partida e parada transcricional, sequências de partida e parada traducional, e sequências ativadoras ou potencializadoras.
Um ácido nucleico é "operacionalmente ligado" quando é colocado em uma relação funcional com outra sequência de ácido nucleico. Por exemplo, DNA que codifica um líder secretório (isto é, um peptídeo sinal), é operacionalmente ligado a DNA de um polipeptídeo se for expresso como uma pré-proteína que participa na secreção do polipeptídeo. Geralmente, "operacionalmente ligado" significa que as sequências de DNA que são liga-das são contíguas, e, no caso de um líder secretório, contíguas e na fase de leitura.
Como usado neste pedido o termo "gene" refere-se a um polinu-cleotídeo (por exemplo, um segmento de DNA), que codifica um polipeptídeo e inclui regiões precedentes e seguintes às regiões de codificação, bem como sequências intervenientes (introns) entre os segmentos de codificação individuais (éxons).
Como usado neste pedido, "ortólogo" e "genes ortólogos" referem-se a genes em espécies diferentes que se desenvolveram a partir de um gene ancestral comum (isto é, um gene homólogo) por especiação. Tipicamente, os ortólogos conservam a mesma função durante o curso da evolução. Identificação de ortólogos encontra uso na predição confiável da função gênica em genomas recentemente sequenciados.
Como usado neste pedido, "parálogo" e "genes parálogos" refe-rem-se a genes que estão relacionados pela duplicação dentro de um genoma. Enquanto os ortólogos conservam a mesma função através do curso da evolução, os parálogos desenvolvem novas funções, embora algumas funções muitas vezes estejam relacionadas à original. Exemplos de genes parálogos incluem, mas não são limitados a genes que codificam tripsina, quimiotripsina, elastase e trombina, que são todas as serina proteinases e que ocorrem em conjunto dentro da mesma espécie.
Como usado neste pedido, o termo "hibridização" refere-se ao processo pelo qual uma fita de ácido nucleico se junta com uma fita com-plementar através de pareamento de bases, como conhecido na técnica.
Uma sequência de ácido nucleico é considerada ser "seletiva-mente hibridizável" a uma sequência de ácido nucleico de referência se as duas sequências especificamente hibridizam entre si sob condições de mo-derada à alta estringência de hibridização e lavagem. As condições de hibri-dização são baseadas na temperatura de fusão (Tm) do complexo de ligação de ácido nucleico ou sonda. Por exemplo, "estringência máxima" tipicamente ocorre em aproximadamente Tm - 5°C (5o abaixo da Tm da sonda); "alta es-tringência" a aproximadamente 5 a 10°C abaixo da Tm; "estringência inter-mediária" a aproximadamente 10 a 20°C abaixo da Tm da sonda; e "baixa estringência" a aproximadamente 20 a 25°C abaixo da Tm. Funcionalmente, condições de estringência máxima podem ser usadas para identificar se-quências tendo identidade estrita ou identidade mais estrita com a sonda de hibridização; enquanto uma estringência de hibridização intermediária ou baixa pode ser usada para identificar ou detectar homólogos de sequência polinucleotidica.
Condições de hibridização de estringência moderada e alta são bem conhecidas na técnica. Um exemplo de condições de alta estringência inclui hibridização a aproximadamente 42°C em formamida 50%, SSC 5 x, solução de Denhardt 5 x, SDS 0,5% e DNA carreador desnaturado 100 μ- g/ml seguido por lavagem duas vezes em SSC 2 x e SDS 0,5% à temperatura ambiente e duas vezes adicionais em SSC 0,1 x e SDS 0,5% a 42°C. Um exemplo de condições estringentes moderadas inclui uma incubação noturna a 37°C em uma solução compreendendo formamida 20%, SSC 5 x (NaCI 150 mM, citrato trissódico 15 mM), fosfato de sódio 50 mM (pH 7,6), solução de Denhardt 5 x, sulfato de dextrana 10% e DNA de esperma de salmão desnaturado cortado 20 mg/ml, seguido por lavagem dos filtros em SSC 1 x a aproximadamente 37 a 50°C. Aqueles versados na técnica sabem como ajustar a temperatura, força iônica, etc. como necessário para acomodar fa-tores, tais como comprimento de sonda e similares.
Como usado neste pedido, "recombinante" inclui referência a uma célula ou vetor, que tenham sido modificados pela introdução de uma sequência de ácido nucleico heteróloga ou homóloga ou que a célula seja derivada de uma célula assim modificada. Dessa forma, por exemplo, células recombinantes expressam genes que não são encontrados em forma idêntica dentro da forma nativa (não recombinante) dos genes nativos celulares ou expressos que são de outra maneira anormalmente expressos, sub- expressos ou não expressos em absoluto em consequência da intervenção humana deliberada.
Em uma modalidade da descrição, as sequências de DNA muta- das são geradas por mutagênese de saturação de sítio em pelo menos um códon. Em outra modalidade, a mutagênese de saturação de sítio é realizada para dois ou mais códons. Em uma modalidade adicional, as sequências de DNA mutante têm homologia de mais de 50%, mais de 55%, mais de 60%, mais de 65%, mais de 70%, mais de 75%, mais de 80%, mais de 85%, mais de 90%, mais de 95%, ou mais de 98% à sequência parental. Em mo-dalidades alternativas, DNA mutante é gerado in vivo usando algum proce-dimento mutagênico conhecido tal como, por exemplo, radiação, nitrosogua- nidina e similares. A sequência de DNA desejada então é isolada e usada nos métodos fornecidos neste pedido.
Como usado neste pedido, "proteína heteróloga" refere-se a uma proteína ou polipeptídeo que não ocorre naturalmente na célula hospedeira.
Uma enzima é "superexpressa" em uma célula hospedeira se a enzima for expressa na célula em um nível mais alto do que o nível em que é expressa em uma célula de tipo selvagem correspondente.
Os termos "proteína" e "polipeptídeo" são usados intercambia- velmente neste pedido. Na presente descrição e reivindicações, os códigos convencionais de uma letra e de três letras de resíduos de aminoácidos são usados. O código de 3 letras para aminoácidos como definido conforme IU- PAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Também é entendido que um polipeptídeo pode ser codificado por mais de uma sequência nucleotídica devido à degeneração do código genético.
Variantes da descrição são descritas pela seguinte nomenclatura: [resíduo de aminoácido original/posição/resíduo de aminoácido substituído]. Por exemplo, a substituição da leucina por arginina na posição 76 é re-presentada como R76L. Quando mais de um aminoácido é substituído em uma dada posição, a substituição é representada como 1) Q172C, Q172D ou Q172R; 2) Q172C, D, ou R, ou 3) Q172C/D/R. Quando uma posição adequada para a substituição é identificada neste pedido sem um aminoácido específico sugerido, deve ser entendido que qualquer resíduo de aminoácido pode ser substituído pelo presente de resíduo de aminoácido na posição. Onde uma glicoamilase variante contém uma deleção comparada com outras glicoamilases, a deleção é indicada com Por exemplo, uma deleção na posição R76 é representada como R76*. Uma deleção de dois ou mais aminoácidos consecutivos é indicada, por exemplo, como (76 a 78)*.
Uma "pró-sequência" é uma sequência de aminoácidos entre a sequência de sinal e proteína madura que é necessária para a secreção da proteína. A clivagem da pró-sequência resultará em uma proteína ativa madura.
O termo "sequência sinal" ou "peptídeo sinal" refere-se a qualquer sequência de nucleotídeos e/ou aminoácidos que possa participar na secreção das formas madura ou precursora da proteína. Esta definição da sequência de sinal é uma funcional daquela, destinada a incluir todas aquelas sequências de aminoácidos codificadas pela porção N-terminal do gene proteico, que participam na efetuação da secreção da proteína. São muitas vezes, mas não universalmente, ligadas à porção N-terminal de uma proteína ou à porção N-terminal de uma proteína precursora. A sequência de sinal pode ser endógena ou exógena. A sequência de sinal pode ser aquela normalmente associada à proteína {por exemplo, glicoamilase), ou pode ser de um gene que codifica outra proteína secretada.
O termo forma "precursora" de uma proteína ou peptídeo refere- se a uma forma madura da proteína tendo uma pró-sequência operacionalmente ligada ao terminal amino ou carbonila da proteína. O precursor também pode ter uma sequência "sinal" operacionalmente ligada, ao terminal amino da pró-sequência. O precursor também pode ter polinucleotídeos adicionais que estão envolvidos na atividade pós-traducional {por exemplo, polinucleotídeos clivados do mesmo para deixar a forma madura de uma proteína ou peptídeo).
"Cepa hospedeira" ou "célula hospedeira" refere-se a um hospe-deiro adequado de um vetor de expressão compreendendo DNA de acordo com a presente descrição.
Os termos "derivado de" e "obtido de" referem-se a não somente uma glicoamilase produzida ou produtível por uma cepa do organismo em questão, mas também uma glicoamilase codificada por uma sequência de DNA isolada de tal cepa e produzido em um organismo hospedeiro contendo tal sequência de DNA. Adicionalmente, o termo refere-se a uma glicoamilase que é codificada por uma sequência de DNA de origem sintética e/ou cDNA e que tem características de identificação da glicoamilase em questão.
Um "derivado" dentro do escopo desta definição geralmente conserva a atividade característica de hidrólise observada na forma de tipo selvagem, nativa ou parental até o ponto que o derivado é útil para fins simi-lares como a forma de tipo selvagem, nativa ou parental. Os derivados fun-cionais de glicoamilases englobam peptídeos de ocorrência natural, sintética ou recombinantemente produzidos ou fragmentos peptídicos que têm as ca-racterísticas gerais das glicoamilases da presente descrição.
O termo "isolado" refere-se a um material que é removido do ambiente natural se estiver ocorrendo naturalmente.
Uma proteína "purificada" refere-se a uma proteína que pelo menos é parcialmente purificada à homogeneidade. Em algumas modalidades, uma proteína purificada é mais de 10% pura, opcionalmente mais de 20% pura, e opcionalmente mais de 30% pura, como determinado por SDS- PAGE. Aspectos adicionais da descrição englobam a proteína em uma forma altamente purificada (isto é, mais de 40% pura, mais de 60% pura, mais de 80% pura, mais de 90% pura, mais de 95% pura, mais de 97% pura, e até mais de 99% pura), como determinado por SDS-PAGE.
Como usado neste pedido, o termo "mutagênese combinatória" refere-se a métodos nos quais as bibliotecas variantes de uma sequência inicial são geradas. Nestas bibliotecas, as variantes contêm uma ou mais mutações escolhidas de um conjunto predefinido de mutações. Além disso, os métodos fornecem meios para introduzir mutações randômicas que não foram membros do conjunto predefinido de mutações. Em algumas modalidades, os métodos incluem aqueles apresentados na Patente Norte- americana N2 6.582.914, por meio deste incorporado por referência. Em modalidades alternativas, os métodos de mutagênese combinatória englobam conjuntos comercialmente disponíveis (por exemplo, QuikChange® Multisite, Stratagene, San Diego, CA).
Como usado neste pedido, o termo "biblioteca de mutantes" re- fere-se a uma população de células que são idênticas na maioria do seu ge- noma, mas incluem homólogos diferentes de um ou mais genes. Tais biblio-tecas podem ser usadas, por exemplo, para identificar genes ou opérons com traços melhorados.
Como usado neste pedido, o termo "conteúdo de sólidos secos (DS ou ds)" refere-se aos sólidos totais de uma pasta fluida em % em uma base de peso seco.
Como usado neste pedido, o termo "acerto inicial" refere-se a uma variante que foi identificada pelo rastreamento de uma biblioteca de mutagênese consenso combinatória. Em algumas modalidades, os acertos iniciais melhoraram características de desempenho, quando comparados com o gene inicial.
Como usado neste pedido, o termo "acerto melhorado" refere-se a uma variante que foi identificada pelo rastreamento de uma biblioteca de mutagênese consenso combinatória aumentada.
Como usado neste pedido, o termo "propriedade-alvo" refere-se à propriedade do gene inicial que deve ser alterada. Não é pretendido que a presente descrição seja limitada a qualquer propriedade-alvo particular. En-tretanto, em algumas modalidades, a propriedade-alvo é a estabilidade de um produto gênico (por exemplo, resistência à desnaturação, proteólise ou outros fatores degradativos), enquanto em outras modalidades, o nível de produção em um hospedeiro de produção é alterado. De fato, é contemplado que qualquer propriedade de um gene inicial encontrará uso na presente descrição. Outras definições de termos podem aparecer em todas as partes do relatório descritivo.
Onde uma faixa de valores é fornecida, é entendido que cada valor interveniente, ao décimo de unidade do limite mais baixo a menos que o contexto claramente dite de outra maneira, entre os limites superiores e inferiores daquela faixa também é especificamente revelado. Cada faixa menor entre qualquer valor determinado ou valor interveniente em uma faixa determinada e qualquer outro valor determinado ou interveniente naquela faixa determinada é englobado dentro da descrição. Os limites superiores e inferiores destas faixas menores podem ser independentemente incluídos ou excluídos na faixa, e cada faixa onde, nenhum ou ambos os limites estão incluídos nas faixas menores também são englobados dentro da descrição, sujeitos a qualquer limite especificamente excluído na faixa determinada. Onde a faixa determinada inclui um ou ambos os limites, as faixas excluindo ou ambos daqueles limites incluídos também estão incluídas na descrição.
Antes que as modalidades exemplares sejam descritas em mais detalhes, deve ser entendido que esta descrição não é limitada a modalidades particulares descritas, como tal pode, naturalmente, variar. Embora quaisquer métodos e materiais similares ou equivalentes aos descritos neste pedido possam ser usados na prática ou no teste da presente descrição, métodos e materiais exemplares são descritos agora.
Como usado neste pedido e nas reivindicações adicionadas, as formas singulares "um", "uma", e "o", "a", incluem os referentes plurais a menos que o contexto claramente dite de outra maneira. Dessa forma, por exemplo, a referência a "um gene" inclui uma pluralidade de tais agentes candidatos e a referência "à célula" inclui referência a uma ou mais células e equivalentes da mesma conhecidos pelos versados na técnica, e similares.
As publicações discutidas neste pedido são fornecidas sozinhas para sua descrição antes da data de depósito do presente pedido de patente. Nada neste pedido deve ser interpretado como uma admissão que a presente descrição não tem direito a antedatar tal publicação em virtude de invenção prévia.
Em algumas modalidades, a presente descrição fornece uma variante de glicoamilase. A variante de glicoamilase é uma variante de uma glicoamilase parental, que pode compreender tanto um domínio catalítico como um domínio de ligação a amido. Em algumas modalidades, a glicoami-lase parental compreende um domínio catalítico tendo uma sequência de aminoácidos como ilustrado nas SEQ ID NO: 1,2,3, 5, 6, 7, 8 ou 9 ou tem uma sequência de aminoácidos que exibe identidade de sequência de pelo menos 80% com uma ou mais das sequências de aminoácidos ilustradas nas SEQ ID NO: 1, 2, 3, 5, 6, 7, 8, ou 9. Ainda em outras modalidades, a glicoamilase parental compreende um domínio catalítico codificado por uma sequência de DNA que hibridiza sob condições estringentes médias, altas, ou com DNA que codifica o domínio catalítico de uma glicoamilase que tern uma das sequências de aminoácido das SEQ ID NO: 1,2 ou 3.
Em algumas modalidades, a glicoamilase parental compreende um domínio de ligação a amido tendo uma sequência de aminoácidos como ilustrado nas SEQ ID N. 1, 2, 161, 162, 163, 164, 165, 166, ou 167, ou tem uma sequência de aminoácidos que exibe identidade de sequência de pelo menos 80% com uma ou mais da sequência de aminoácidos ilustrada nas SEQ ID NO: 1, 2, 161, 162, 163, 164, 165, 166, ou 167. Ainda em outras modalidades, a glicoamilase parental compreende um domínio de ligação a amido codificado por uma sequência de DNA que hibridiza sob condições estringentes médias, altas, ou com DNA que codifica domínio de ligação a amido de uma glicoamilase que tem uma das sequências de aminoácido da SEQ ID NO: 1 ou 2.
Estrutura predita e sequências conhecidas de glicoamilases são conservadas entre espécies fúngicas (Coutinho et al., 1994, Protein Eng., 7:393-400 e Coutinho et al., 1994, Protein Eng., 7: 749-760). Em algumas modalidades, a glicoamilase parental é uma glicoamilase de fungo filamen-toso. Em algumas modalidades, a glicoamilase parental é obtida de uma cepa de Trichoderma (por exemplo, T. reesei, T. longibrachiatum, T. strictipilis, T. asperellum, T. konilangbra e T. hazianum), uma cepa de Aspergillus (por exemplo, A. niger, A. nidulans, A. kawachi, A. awamori e A. orzyae), uma cepa de Talaromyces (por exemplo, T. emersonii, T. thermophilus, e T. du- ponti), uma cepa de Hypocrea (por exemplo, H. gelatinosa, H. orientalis, H. vinosa, e H. citrina), uma cepa de Fusarium (por exemplo, F. oxysporum, F. roseum, e F. venenatum), uma cepa de Neurospora (por exemplo, N. crassa) e uma cepa de Humicola (por exemplo, H. grisea, H. insolens e H. lanugino- se), uma cepa de Penicillium (por exemplo, P. notatum ou P. chrysogenum), ou cepa de Saccharomycopsis (por exemplo, S. fibuligera).
Em algumas modalidades, a glicoamilase parental pode ser uma glicoamilase bacteriana. Por exemplo, o polipeptídeo pode ser obtido de uma cepa bacteriana gram-positiva, tal como Bacillus (por exemplo, B. alkalophi- lus, B. amyloliquefaciens, B. lentus, B. licheniformis, B. stearothermophilus, B. subtilis e B. thuringiensis) ou uma cepa de Streptomyces (por exemplo, S. lividans).
Em algumas modalidades, a glicoamilase parental compreenderá um domínio catalítico tendo identidade de sequência de pelo menos 80%, identidade de sequência de pelo menos 85%, identidade de sequência de pelo menos 90%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 97% e também identidade de sequência de pelo menos 98% com o domínio catalítico da sequência de aminoácidos da TrGA de SEQ ID NO: 3.
Em outras modalidades, a glicoamilase parental compreenderá um domínio catalítico tendo identidade de sequência de pelo menos 90%, identidade de sequência de pelo menos 93%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 96%, identidade de sequência de pelo menos 97%, identidade de sequência de pelo menos 98% e também identidade de sequência de pelo menos 99% com o domínio catalítico da glicoamilase parental de Aspergillus da SEQ ID NO: 5 ou SEQ ID NO: 6.
Ainda em outras modalidades, a glicoamilase parental compre-enderá um domínio catalítico tendo identidade de sequência de pelo menos 90%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 97% e também identidade de sequência de pelo menos 99% com o domínio catalítico de glicoamilase parental de Humicola grisea (HgGA) da SEQ ID NO: 8.
Em algumas modalidades, a glicoamilase parental compreenderá um domínio de ligação a amido tendo identidade de sequência de pelo menos 80%, identidade de sequência de pelo menos 85%, identidade de sequência de pelo menos 90%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 97% e também identidade de sequência de pelo menos 98% com o domínio de ligação a amido da sequência de aminoácidos de TrGA das SEQ ID NO: 1, 2, ou 161.
Em outras modalidades, a glicoamilase parental compreenderá um domínio de ligação a amido tendo identidade de sequência de pelo me- nos 90%, identidade de sequência de pelo menos 95%, identidade de se-quência de pelo menos 97% e também identidade de sequência de pelo menos 99% com o domínio catalítico da glicoamilase de Humicola grísea (Hg- GA) da SEQ ID NO: 162.
Em outras modalidades, a glicoamilase parental compreenderá um domínio de ligação a amido tendo identidade de sequência de pelo menos 90%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 97% e também identidade de sequência de pelo menos 99% com o domínio catalítico da glicoamilase de Thielavia terrestris (Tt- GA) da SEQ ID NO: 163.
Em outras modalidades, a glicoamilase parental compreenderá um domínio de ligação a amido tendo identidade de sequência de pelo menos 90%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 97% e também identidade de sequência de pelo menos 99% com o domínio catalítico da glicoamilase de Thermomyces lanugi- nosus (ThGA) da SEQ ID NO; 164.
Em outras modalidades, a glicoamilase parental compreenderá um domínio de ligação a amido tendo identidade de sequência de pelo menos 90%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 97% e também identidade de sequência de pelo menos 99% com o domínio catalítico da glicoamilase de Talaromyces emersoni- ft(TeGA) da SEQ ID NO: 165.
Ainda em outras modalidades, a glicoamilase parental compre-enderá um domínio de ligação a amido tendo identidade de sequência de pelo menos 90%, identidade de sequência de pelo menos 93%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 96%, identidade de sequência de pelo menos 97%, identidade de sequência de pelo menos 98% e também identidade de sequência de pelo menos 99% com o domínio de ligação a amido da glicoamilase parental de Aspergillus das SEQ ID NO: 166 ou 167.
Em algumas modalidades, a glicoamilase parental terá identidade de sequência de pelo menos 80%, identidade de sequência de pelo me- nos 85%, identidade de sequência de pelo menos 88%, identidade de se-quência de pelo menos 90%, identidade de sequência de pelo menos 93%, identidade de sequência de pelo menos 95%, identidade de sequência de pelo menos 96%, identidade de sequência de pelo menos 97%, identidade de sequência de pelo menos 98% e também identidade de sequência de pelo menos 99% com a sequência de aminoácidos de TrGA da SEQ ID NO: 2.
Em modalidades adicionais, um homólogo de glicoamilase de Trichoderma será obtido de uma linhagem de Hypocrea ou Trichoderma. Alguns homólogos de glicoamilase de Trichoderma típicos são descritos na Patente Norte-americana Ns 7.413.887 e a referência é feita especificamente a sequências de aminoácidos apresentadas nas SEQ ID NOs: 17 a 22 e 43 a 47 da referência.
Em algumas modalidades, a glicoamilase parental é TrGA com-preendendo sequência de aminoácidos da SEQ ID NO: 2, ou um homólogo da glicoamilase de Trichoderma tendo pelo menos identidade de sequência de 80%, pelo menos 85%, pelo menos 88%, pelo menos 90%, pelo menos 93%, pelo menos 95%, pelo menos 96%, pelo menos 97%, pelo menos 98%, pelo menos 99% à sequência de TrGA (SEQ ID NO: 2).
Uma glicoamilase parental pode ser isolada e/ou identificada usando técnicas padrão de DNA recombinante. Quaisquer técnicas padrão que podem ser usadas que são conhecidas pelo versado. Por exemplo, sondas e/ou iniciadores específicos para regiões conservadas da glicoamilase podem ser usados para identificar homólogos em células bacterianas ou fúngicas (o domínio catalítico, o sítio ativo, etc.). Altemativamente, PCR degenerada pode ser usada para identificar homólogos em células bacterianas ou fúngicas. Em alguns casos, as sequências conhecidas, tal como em um banco de dados, podem ser analisadas para sequência e/ou identidade estrutural a uma das glicoamilases conhecidas, incluindo SEQ ID NO: 2, ou domínios de ligação a amido conhecidos, incluindo SEQ ID NO: 161. Ensaios funcionais também podem ser usados para identificar a atividade de glicoamilase em uma célula bacteriana ou fúngica. Proteínas tendo atividade de glicoamilase podem ser isoladas e sequenciadas reversamente para isolar a sequência de DNA correspondente. Tais métodos são conhecidos pelo versado.
O dogma central da biologia molecular é que a sequência de DNA que codifica um gene de uma enzima particular, determina a sequência de aminoácidos da proteína, esta sequência por sua vez determina o enove-lamento tridimensional da enzima. Este enovelamento induz resíduos desi-guais que criam um centro catalítico e substrato que se liga à superfície e isto resulta na alta especificidade e atividade das enzimas em questão.
Glicoamilases consistem de não menos que três domínios estru-turais distintos, um domínio catalítico de aproximadamente 450 resíduos que é estruturalmente conservado em todas as glicoamilases, geralmente seguidas por uma região ligantea consistindo de entre 30 e 80 resíduos que são unidos a um domínio de ligação a amido de aproximadamente 100 resíduos. A estrutura da glicoamilase de Trichoderma reesei com as três regiões intactas foi determinada em resolução de 1,8 Angstrõns neste pedido (ver Tabela 20 e Exemplo 13). Usando as coordenadas (ver Tabela 20), a estrutura foi alinhada com as coordenadas do domínio catalítico da cepa X100 de Aspergillus awamorii que foi determinada anteriormente (Aleshin, A.E., Hoffman, C., Firsov, L.M., and Honzatko, R.B. 1994 Refined crystal structures of glu-coamylase from Aspergillus awamori var. X100. J Mol Biol 238: 575-591.). A estrutura cristalina de Aspergillus awamori somente incluiu o domínio catalí-tico. Como visto nas Figuras 12 e 13, a estrutura dos domínios catalíticos sobrepõe-se muito estreitamente e é possível identificar resíduos equivalentes com base nesta sobreposição estrutural. Acredita-se que todas as glico-amilases compartilham a estrutura básica representada nas Figuras 12 e 13.
A figura 12 é uma comparação das estruturas tridimensionais da glicoamilase de Trichoderma (preto) da SEQ ID NO: 1 (ver Figura 1 para se-quência de aminoácidos) e de Aspergillus awamorii (cinza) vista ao lado. Nesta visão, a relação entre o domínio catalítico e a região ligantea e o do-mínio de ligação a amido pode ser vista.
A figura 13 é uma comparação de estruturas tridimensionais de glicoamilase de Trichoderma (preto) e de Aspergillus awamorii (cinza) vista do topo. As glicoamilases mostradas aqui e de fato, todas glicoamilases co-nhecidas até a data compartilham esta homologia estrutural. A conservação da estrutura correlaciona-se com a conservação de atividade e um mecanismo conservado de ação de todas as glicoamilases. Considerando esta alta homologia, modificações que resultam a partir de variantes específicas de sítio de glicoamilase de Trichoderma resultando em função alterada também teriam estrutural similar e por isso consequências funcionais em outras glicoamilases. Por isso, nos ensinamentos de que as variantes resultam em benefícios desejáveis podem ser aplicados a outras glicoamilases.
Uma estrutura cristalina adicional foi produzida usando as coor-denadas na Tabela 20 para o Domínio de Ligação a Amido (SBD). O SBD de TrGA foi alinhado com o SBD de A. niger. Como mostrado na Figura 13, a estrutura de SBDs de A. niger e TrGA sobrepõem-se muito estreitamente. Acredita-se que, enquanto todos os domínios de ligação a amido compartilham pelo menos a estrutura básica representada na Figura 13, alguns SBDs são mais estruturalmente similares do que outros. Por exemplo, o SBD de TrGA pode ser classificado como dentro da família 20 do módulo de ligação a carboidrato dentro do banco de dados CAZY (cazy.org). O banco de dados CAZY descreve as famílias de módulos catalíticos e de ligação a carboidrato estruturalmente relacionados (ou domínios funcionais) de enzimas que de-gradam, modificam, ou criam ligações glicosídicas. Considerando uma alta homologia estrutural, variantes específicas de sítio do SBD de TrGA resul-tando em função alterada também teriam consequências estruturais similares e por isso funcionais em outras glicoamilases que têm SBDs com estrutura similar àquela do SBD de TrGA, particularmente aqueles classificados dentro da família 20 do módulo de ligação a carboidrato. Dessa forma, nos ensinamentos dos quais as variantes resultam em benefícios desejáveis po-dem ser aplicados a outros SBDs tendo similaridade estrutural.
Dessa forma, os números de posição de aminoácido discutidos neste pedido referem-se aos destinados à sequência de glicoamilase madu- ra de Trichoderma reesei apresentada na Figura 1. A presente descrição, entretanto, não é limitada às variantes da glicoamilase de Trichoderma, mas estende-se a glicoamilases contendo resíduos de aminoácidos em posições que são "equivalentes" aos resíduos particulares identificados em glicoami-lase de Trichoderma reesei (SEQ ID NO: 2). Em uma modalidade da presente descrição, a glicoamilase parental é a glicoamilase de Taleromyces e as substituições são feitas nas posições de resíduo de aminoácido equivalentes na glicoamilase de Taleromyces como aquelas descritas neste pedido. Em outras modalidades, a glicoamilase parental é uma das listadas na Tabela 1. Em modalidades adicionais, a glicoamilase parental é uma glicoamilase de Penicillium, tal como Penicillium chrysogenum.
Identidade estrutural determina se os resíduos de aminoácidos são equivalentes. Identidade estrutural é um equivalente topológico individual quando as duas estruturas (estruturas tridimensionais e de aminoácido) são alinhadas. Uma posição de resíduo (aminoácido) de uma glicoamilase é equivalente a um resíduo de glicoamilase de T. reesei se for homólogo (isto é, correspondente na posição na estrutura primária ou terciária) ou análogo a um resíduo ou porção específicos daquele resíduo em glicoamilase de T. reesei (tendo a mesma capacidade funcional ou similar de combinar-se, reagir, ou interagir quimicamente).
A fim de estabelecer a identidade à estrutura primária, a se-quência de aminoácidos de uma glicoamilase pode ser diretamente comparada à sequência primária de glicoamilase de Trichoderma reesei e particularmente ao grupo de resíduos conhecidos sendo invariáveis em glicoamilases pelas quais a sequência é conhecida. Por exemplo, as Figuras 4A e 4B neste pedido mostram os resíduos conservados entre glicoamilases. As figuras 4C e 4D mostram um alinhamento dos domínios de ligação a amido de várias glicoamilases. Após alinhar os resíduos conservados, permitir inserções necessárias e deleções a fim de manter o alinhamento (isto é, evitar eliminação de resíduos conservados através de deleção e inserção arbitrá-rias), os resíduos equivalentes a aminoácidos particulares na sequência pri-mária de glicoamilase de Trichoderma reesei são definidos. O alinhamento de resíduos conservados tipicamente deve conservar 100% de tais resíduos. Entretanto, o alinhamento de maiores do que 75% ou tão pouco quanto 50% de resíduos conservados é também adequado para definir resíduos equivalentes. Além disso, a identidade estrutural pode ser usada em combinação com a identidade de sequência para identificar resíduos equivalentes.
Por exemplo, nas Figuras 4A e 4B, os domínios catalíticos de glicoamilases de seis organismos são alinhados para fornecer a quantidade máxima de homologia entre as sequências de aminoácido. Uma comparação destas sequências mostra que há diversos resíduos conservados contidos em cada sequência como indicado por um asterisco. Estes resíduos conservados, dessa forma, podem ser usados para definir os resíduos de aminoácidos equivalentes correspondentes de glicoamilase de Trichoderma reesei em outras glicoamilases, tais como glicoamilase de Aspergillus niger. Simi-larmente, as Figuras 4C e 4D mostram os domínios de ligação a amido de glicoamilases de sete organismos alinhados para identificar resíduos equiva-lentes.
Identidade estrutural envolve a identificação de resíduos equiva-lentes entre as duas estruturas. "Resíduos equivalentes" podem ser definidos pela determinação de homologia no nível de estrutura terciária (identidade estrutural) para uma enzima cuja estrutura terciária tenha sido determinada por cristalografia de raio x. Os resíduos equivalentes são definidos como aqueles para os quais as coordenadas atômicas de dois ou mais dos átomos da cadeia principal de um resíduo particular de aminoácido de glicoamilase de Trichoderma reesei (N em N, CA em CA, C em C e O em O) estão em 0,13 nm e opcionalmente 0,1 nm após alinhamento. Alinhamento é alcançado após o melhor modelo ter sido orientado e posicionado para dar sobreposição máxima de coordenadas atômicas de átomos proteicos não hidrogênio da glicoamilase em questão à glicoamilase de Trichoderma reesei. O melhor modelo é o modelo cristalográfico que fornece o fator R mais baixo para dados experimentais de difração na resolução mais alta disponível.
Resíduos equivalentes que são funcionalmente análogos a um resíduo específico da glicoamilase de Trichoderma reesei são definidos como aqueles aminoácidos da enzima que podem adotar uma conformação tal que alterem, modifiquem ou contribuam para estrutura proteica, ligação de substrato ou catálise em uma maneira definida e atribuída a um resíduo es-pecífico da glicoamilase de Trichoderma reesei. Além disso, são aqueles resíduos da enzima (para a qual uma estrutura terciária tenha sido obtida pela cristalografia de raio x) que ocupam uma posição análoga até o ponto que, embora os átomos da cadeia principal do dado resíduo possam não satisfazer os critérios de equivalência com base na ocupação de uma posição homóloga, as coordenadas atômicas de pelo menos dois dos átomos da cadeia lateral do resíduo têm 0,13 nm dos átomos da cadeia lateral correspondentes de glicoamilase de Trichoderma reesei. As coordenadas de estrutura tridimensional da glicoamilase de Trichoderma reesei são apresentadas na Tabela 15 e podem ser usadas como delineado acima para determinar resíduos equivalentes no nível de estrutura terciária.
Alguns dos resíduos identificados para substituição são resíduos conservados ao passo que outros não são. No caso de resíduos que não são conservados, a substituição de um ou mais aminoácidos é limitada a substituições que produzem uma variante que tem uma sequência de ami-noácidos que não corresponde a uma encontrada na natureza. No caso de resíduos conservados, tais substituições não devem resultar em uma se-quência que ocorre naturalmente.
As variantes de acordo com a descrição incluem pelo menos uma substituição, deleção ou inserção na sequência de aminoácidos de uma glicoamilase parental que toma a variante diferente na sequência da glicoa-milase parental. Em algumas modalidades, as variantes da descrição terão pelo menos 20%, pelo menos 40%, pelo menos 60%, pelo menos 80%, pelo menos 85%, pelo menos 90%, pelo menos 95%, ou pelo menos 100% da atividade de glicoamilase da atividade de TrGA da SEQ ID NO: 2.
Em algumas modalidades, as variantes de acordo com a descri- ção compreenderão uma substituição, deleção ou inserção em pelo menos uma posição de aminoácido da TrGA parental (SEQ ID NO: 2), ou em uma posição equivalente na sequência de outra glicoamilase parental tendo identidade de sequência de pelo menos 80% à sequência de TrGA, incluindo, mas não limitadas a identidade de sequência de pelo menos 90%, pelo menos 93%, pelo menos 95%, pelo menos 97%, e pelo menos 99%. Em outras modalidades, a variante de acordo com a descrição compreenderá uma substituição, deleção ou inserção em pelo menos uma posição de aminoácido de um fragmento da TrGA parental, em que o fragmento compreende o domínio catalítico da sequência de TrGA (SEQ ID NO: 3) ou em uma posição equivalente em um fragmento compreendendo o domínio catalítico de uma glicoamilase parental tendo identidade de sequência de pelo menos 80% ao fragmento da sequência de TrGA, incluindo mas não limitado a, pelo menos 90%, pelo menos 95%, pelo menos 97%, e pelo menos 99%. Em algumas modalidades, o fragmento compreenderá pelo menos 400, 425, 450 e/ou 500 resíduos de aminoácido.
Em outras modalidades, a variante de acordo com a descrição compreenderá uma substituição, deleção ou inserção em pelo menos uma posição de aminoácido de um fragmento da TrGA parental, em que o frag-mento compreende o domínio de ligação a amido da sequência de TrGA (SEQ ID NO: 161) ou em uma posição equivalente em um fragmento com-preendendo o domínio de ligação a amido de uma glicoamilase parental tendo identidade de sequência de pelo menos 80% ao fragmento da sequência de TrGA, incluindo, mas não limitado a pelo menos 90%, pelo menos 95%, pelo menos 97% e pelo menos 99%. Em algumas modalidades, o fragmento compreenderá pelo menos 40, 50, 60, 70, 80, 90, 100, e/ou 109 resíduos de aminoácidos do domínio de ligação a amido de TrGA (SEQ ID NO: 161).
Em algumas modalidades, quando a glicoamilase parental inclui um domínio catalítico, uma região ligantea, e um domínio de ligação a amido, a variante compreenderá uma substituição, deleção ou inserção em pelo menos uma posição de aminoácido de um fragmento compreendendo parte da região ligantea. Em algumas modalidades, a variante compreenderá uma substituição, deleção ou inserção na sequência de aminoácidos de um frag-mento da sequência de TrGA (SEQ ID NO: 2).
Identidade estrutural com referência a uma substituição de ami-noácido significa que a substituição ocorre na posição de aminoácido equi-valente na glicoamilase homóloga ou glicoamilase parental. O termo posição equivalente significa uma posição que é comum a duas sequências parentais que é baseada em um alinhamento da sequência de aminoácidos da glicoamilase parental em questão bem como alinhamento da estrutura tridi-mensional da glicoamilase parental em questão com a sequência de amino-ácidos de glicoamilase de referência TrGA e sequência tridimensional. Por exemplo, com referência à Figura 5, a posição 24 em TrGA (SEQ ID NO: 2 ou 3) é D24 e a posição equivalente de Aspergillus niger (SEQ ID NO: 6) é posição D25, e a posição equivalente de Aspergillus oryzea (SEQ ID NO: 7) é posição D26. Ver Figuras 12 e 13 para um alinhamento exemplar da sequência tridimensional.
Em algumas modalidades, a variante de glicoamilase incluirá pelo menos uma substituição na sequência de aminoácidos de um parental. Em modalidades adicionais, a variante pode ter mais de uma substituição (por exemplo, dois, três, ou quatro substituições).
Em algumas modalidades, uma variante de glicoamilase com-preende uma substituição, deleção ou inserção, e tipicamente uma substitui-ção em pelo menos uma posição de aminoácido em uma posição corres-pondente às regiões de aminoácidos não conservados como ilustrado na Figura 5 (por exemplo, posições de aminoácido correspondente àquelas po-sições que não são indicadas por na Figura 5).
Enquanto as variantes podem estar em qualquer posição na se-quência proteica madura (SEQ ID NO: 2), em uma modalidade, uma variante de glicoamilase compreende uma ou mais substituições nas seguintes posi-ções na sequência de aminoácidos apresentada nas SEQ ID NO: 2: 4, 5, 12, 24, 29, 43, 44, 45, 46, 47, 49, 51, 61, 70, 73, 75, 76, 94, 100, 108, 114, 116, 119, 122, 124, 125, 137, 143, 146, 148, 169, 171, 172, 175, 178, 180, 181, 208, 211, 228, 242, 243, 245, 292, 294, 297, 309, 310, 313, 314, 315, 316, 317, 321, 340, 341, 350, 353, 356, 363, 368, 369, 375, 376, 395, 398, 401, 408, 409, 412, 415, 417, 418, 421, 430, 431, 433, 436, 451, 503, 511, 535, 539, ou 563; ou em uma posição equivalente em uma glicoamilase parental. Em algumas modalidades, a glicoamilase parental terá identidade de pelo menos 90%, pelo menos 95%, pelo menos 96%, pelo menos 97% pelo menos 98%, e pelo menos 99% com SEQ ID NO: 2. Em outras modalidades, a glicoamilase parental será um homólogo de glicoamilase de Trichoderma.
Em algumas modalidades, a variante de glicoamilase compreende uma ou mais substituições nas seguintes posições na sequência de ami-noácidos apresentada em SEQ ID NO: 2: D4, F5, 112, D24, F29, I43, D44, P45, D46, Y47, Y49, W51, N61, Y70, G73, Q75, R76, P94, D100, K108, K114, F116, N119, R122, Q124, R125, G137, N146, Q148, Y169, N171, Q172, F175, W178, E180, V181, Q208, S211, W228, N242, E243, R245, I292, G294, K297, R309, Y310, D313, V314, Y315, Y316, N317, W321, K340, K341, T350, Q356, T363, S368, S369, N376, Y395, A398, S401, R408, N409, T412, L417, H418, W421, T430, A431, R433, I436, S451, E503, Q511, A535, A539, ou N563; ou uma posição equivalente em glicoamilase parental (por exemplo, um homólogo de glicoamilase de Trichoderma).
Em outras modalidades, a variante de uma glicoamilase parental compreende uma ou mais substituições nas seguintes posições na sequência de aminoácidos apresentada nas SEQ ID NO:2: 4, 5, 24, 29, 43, 44, 49, 61, 70, 73, 75, 76, 100, 108, 119, 124, 137, 146, 148, 169, 171, 172, 175, 178, 181, 208, 211, 243, 292, 294, 297, 314, 316, 317, 340, 341, 350, 356, 363, 368, 369, 376, 395, 401, 409, 412, 417, 430, 431, 433, 436, 451, 503, 511, 535, 539, ou 563; ou uma posição equivalente em uma glicoamilase parental (por exemplo, um homólogo de glicoamilase de Trichoderma).
Em modalidades adicionais, a variante de uma glicoamilase pa-rental compreende pelo menos uma das seguintes substituições nas seguintes posições em uma sequência de aminoácidos apresentada na SEQ ID NO: 2: D4L/E/R/S/C/A/Q/W, F5C/M/N/R/S/T/V/W, I12L/R, D24E/L/Y/T, F29L/I/D/C/S/V/W, I43F/R/D/Y/S/Q, D44E/H/K/S/N/Y/F/R/C, Y47W, Y49N, N61 D/l/L/Q/V/W, Q70R/K/M/P/G/L/F, G73F/C/L/W, Q75R/K/A, R76L/M/K/T/P, P94L, D100W/I/Q/M/P/A/N, N119P/T/Y/D/E, N146S/G/C/H/E/D/T/W/L/F/M, Q148V/Y/H/A/C/D/G/M/R/S/T, Y169D/F, Q172C/A/D/R/E/F/H/V/L/M/N/S/T/V, F175H/A/G/R/S/T/C/W/Y, W178A/C/D/E/F/G/H/K/N/R/S/T/V/Y, E180A/C/G/H/I/L/N/P/Q/R/S/T/V/Y/, V181 E/C/D/G/H/l/P/T/Y/S/L/K/F/A, Q208L/A/C/E/N/F/H/T, S211C/R/E/A/Y/W/M/H/L/I/R/Q/T, E243S/R/N/M/Y/A/L, R245A/E/M/I/P/V, I292D/H/P/R/T/N/V/F/L, G294C/D/E/17Q/I/A, K297F/L/P/T/M/D/N/Q/A/Y/H/S/R/W, R309A/C/G/H/I/N/P/Q/S/T/W/Y/L, Y310E/G/L/P/S/W/R/Q, D313Q, V314A/R/N/D/C/E/Q/G/H/I/L/K/M/F/P/S/T/W/Y, Y315F, Y316Q/R, N317T/H, K340D/T, K341F/D/P/V/G/S, T350S/E/A/N, Q356H/D/E, T363L/R/C/H/W, S368W/D/F/L, S369F, N376Q/T/H/S/V, Y395Q/R/S, A398S/I/T, S401C/V, R408S, N409W/T/K, T412A/H/K/G, L417A/D/E/F/G/I/K/Q/R/S/T/V/W/Y, T430A/E/F/G/H/I/K/M/N/Q/R/V, A431 C/E/H/l/L/M/Q/R/S/W/Y, R433H/Q, I436A/T, S451M/T/H, E503A/C/D/H/S/V/W, Q511C/G/H/I/K/T7V, A535E/F/G/K/L/N/P/R/S/T/V/W/Y, A539E/H/M/R/S/W, ou N563/A/C/E/I/K/L/Q/T/V; ou uma substituição em uma posição equivalente em um homólogo de glicoamilase parental.
Em algumas modalidades, a variante de glicoamilase compreende pelo menos uma substituição em uma posição correspondente à posição de resíduo de aminoácido apresentada nas SEQ ID NO: 2: 5, 24, 43, 44, 49, 61, 70, 73, 75, 76, 94, 119, 146, 148, 172, 175, 178, 180, 181, 208, 211, 245, 294, 353, 315, 375, 409, 309, 314, 369, 412, 417, 430, 431, 503, 511, 535, 539, ou 563; ou uma posição equivalente em uma glicoamilase parental homóloga.
Em algumas modalidades representativas, a variante de glicoa-milase compreende pelo menos uma substituição selecionada a partir do grupo consistindo de F5W, D24E, I43R, I43Y, I43Q, I43S, I43F, D44C, D44R, Y47W, Y49N, N61I, Q70K, G73F, Q75R, R76L, P94L, N119P/17Y/D, N146S/D/T/E/W/L, Q148V N171D, Q172C/D/R/E/F/V/L/T, F175R/W/Y, W178K/N/Y, E180H/N/V/R, V181 E/F/G/l/H, Q208A/T/N, S211H/M/L/R, R245E, R245M, G294C, R309W, V314F/G/H/K/P/R/Y, Y315F, S369F, T412K, L417R, L417V, T430A, T430M, A431L, A431Q, E503A, E503V, Q511H, A535R, A539R, N563I, e N563K correspondente à posição apresentada na SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental homóloga.
Em modalidades adicionais particulares, a variante de glicoami-lase compreende pelo menos uma substituição de um resíduo de aminoácido selecionado a partir das posições correspondentes à posição 5, 43, 44, 61, 73, 75, 76, 94, 108, 119, 124, 146, 148, 171, 172, 175, 178, 180, 181, 208, 211, 294, 297, 314, 316, 412, 417, 430, 431,503, 511, 535, 539, ou 563 da SEQ ID NO: 2, ou uma posição equivalente em um homólogo de glicoamilase de Trichoderma. Em algumas modalidades, a substituição está em uma posição correspondente à posição número 43, 44, 61, 73, 148, 172, 175, 178, 180, 208, 211, 294, 297, 314, 412, 417, 430, 431, 503, 511, 535, 539, ou 563 da SEQ ID NO: 2, ou uma posição equivalente em um homólogo de glicoamilase de Trichoderma.
Em algumas modalidades representativas, a substituição está em uma posição correspondente à posição número 43, 44, 61, 73,108, 124, 171, 172, 208, 211,294, 314, 316, 417, 430, 431,503, 511, 535, 539, ou 563 da SEQ ID NO: 2, ou uma glicoamilase parental homóloga (por exemplo, homólogo de glicoamilase de Trichoderma).
Em algumas modalidades, as variantes de glicoamilase compre-endem múltiplas substituições. Algumas das múltiplas substituições incluirão uma substituição em uma ou mais das posições equivalentes e incluindo as posições 24, 43, 44, 108, 124, 171, 175, 181, 208, 243, 292, 294, 297, 310, 314, 363, 417, 430, 431, 503, 511, 535, 539, ou 563 da SEQ ID NO: 2. Al-gumas substituições múltiplas típicas incluirão uma ou mais das posições equivalentes e correspondentes a posições 43, 44, 61, 73, 08,124,171,208, 211,294, 314, 417, 430, 431, 503, 511, 535, 539, ou 563 da SEQ ID NO: 2.
Alguns exemplos de variantes com múltiplas substituições inclu-em substituições nas posições: D24/I43/D44/F175/V181 /V314fT363; D24/Q208/I292/G294/K297/Y310; V181/E243/l292/k297/N317/Y395; D24/V181 /Q208/G294/T363/N376/N409; D24/V181/I292/G294/E243/N409; I43R/E243/I292/G294/K297; I43/D44/N61/L417/E503/Q511/A539; I43/D44/L417/E503/Q511/A539; I43/N61ZL417/T430/Q511 /A539; I43/N61 /L417/E503/Q511 /A539; I43/N61 /T430/A431 /Q511 /A539; I43/N61/T430/Q511; I43/N61/T430/Q511/A539; I43/N61/Q511; I43/N61/Q511/A539; I43/G73/T430; I43/L417/E503/Q511 /A539; I43/L417/Q511; I43/L417/T430/A431 /Q511 /A539; I43/L417/T430/Q511; I43/L417/T430/Q511/A539; I43/L417/E503/A539; I43/L417/E503/Q511/A539; I43/T430; I43/T430/A431 /E503/Q511; I43/T430/A431/Q511; I43/T430/A431/Q511/A539; I43/T430/E503/Q511; I43/T430/Q511; I43Q/T430/Q511/A539; I43/A431/Q511; I43/T430/E503/Q511/N563; I43/T430/E503/A535/N563; I43/E503/Q511/A539; I43/Q511/A539; D44/G73/L417/N563; D44/G73/E503/Q511; D44/G73/N563; D44/L417/N563; D44/T430/Q511/A535; D44/E503/Q511/N563; G73/T430/E503/Q511; G73/T430/Q511 ; G294/L417/A431; G294/L417/A431; G294/L417/A431/Q511; L417/T430/A431/Q511/A535/A539/N563; L417/A431/Q511; L417/T430/Q511/A535/N563; L417/T430/Q511/A539/N563; e E503/N563; da SEQ ID NO: 2, ou posições equivalentes em glicoamilases parentais e particularmente homólogos de glicoamilase de Trichoderma.
Algumas variantes com múltiplas substituições podem incluir as substituições nas posições: Y47F/W.Y315F/W; D24E,L/I43F,R/D44H,N/F175H/V181K,L/V314D,H,K/T363R; D24L,W,Y/Q208F/l292F,N,V/G294A,l,Q/K297A/Y310F,Q,R; V181 F,K,L/E243A,N,M,R,Y/I292F,L,N,V/K297A,D,H,M,N,Q/N317H/Y395Q.R; D24E,L,Y/V181 F,K,L/Q208C,F/G294A,l,Q/T363R/N376Q/N409K,W; D24E,L,Y/V181 F,K,L/l292F,L,N,V/G294AJ,Q/E243A,M,N,RrY/N409K,W; I43R/E243A,M,N,R,Y/I292F,L>N,V/G294A/K297A,D,H,M,N,Q,S,R,W,Y; I43Q/D44C/N611/L417V/E503A/Q511H/A539R; I43Q/D44C/L417V/E503A/Q511H/A539R; I43Q/N611/L417V/E503A/Q511H/A539R; I43Q/N611/L417WT430M/Q511H/A539R; I43R/N611/L417R.V/E503A/Q511H/A539R; I43Q/N611/T430A/A431L/Q511H/A539R; I43Q/N61I/T430A/Q511H; I43Q/N611/T430A/Q511H/A539R; I43Q/N611/T430M/Q511H/A539R I43Q/N611/Q511H; I43Q/N61I/Q511H/A539R; I43R/G73F/T430A; I43Q/L417V/T430A/A431L/Q511H/A539R; I43Q/L417V/T430A/Q511H; I43Q/L417V/T430A/Q511H/A539R; I43R/L417R/E503A/A539R; I43R.Q/L417V/E503A/Q511H/A539R; I43Q/L417V/Q511H; I43R.Q/T430A; I43Q/T430A/A431L/E503A/Q511H; I43Q/T430A/A431L/Q511H; I43Q/T430A/A431L/Q511H/A539R; I43Q/T430A/E503A/Q511H; I43Q/T430A/Q511H; I43Q/T430A.M/Q511H/A539R; I43R/T430A/E503A.V/Q511H/N563K; I43Q/A431L/Q511H; I43Q/E503A/Q511H/A539R; I43Q/Q511H/A539R; D44C/G73F/E503V/Q511H; D44C/G73F/L417R/N563K; D44C/G73F/N563K; D44C/L417R/N563K; D44R/E503A/Q511H/N563I; D44R/T430A/Q511H/A535R; G73F/T430A/E503V/Q511H; G73F/T430A/Q511H; G294C/L417R/A431L; G294C/L417R/A431 L.Q/Q511H; G294C/L417V/A431Q; L417R.V/A431 L.Q/Q511H; L417V/T430A/A431 L.Q/Q511H/A535R/A539R/N563I; L417V/T430A/Q511H/A535R/N563I; L417V/T430A/Q511H/A539R/N563I; e E503A/N563I da SEQ ID NO: 2, ou posições equivalentes em glicoamilases parentais e particularmente homólogos de glicoamilase de Trichoderma.
Diversas glicoamilases parentais têm sido alinhadas com a se-quência de aminoácidos de TrGA. As figuras 4A e 4B incluem o domínio ca-talítico das seguintes glicoamilases parentais: Aspergillus awamori (AaGA) (SEQ ID NO: 5); Aspergillus niger (AnGA) (SEQ ID NO: 6); Aspergillus orz- yae (AoGA) (SEQ ID NO: 7), Humicola grisea (HgGA) (SEQ ID NO: 8) e Hy- pocrea vinosa (HvGA) (SEQ ID NO: 9). A identidade percentual dos domínios catalíticos é representada na Tabela 1 abaixo. As figuras 4C e 4D incluem o domínio de ligação a amido das seguintes glicoamilases parentais: Tri-choderma reesei (TrGA) (SEQ ID NO: 161), Humicola grisea (HgGA) (SEQ ID NO: 162), Thielavia terrestris (TtGA) (SEQ ID NO: 163), Thermomyces lanuginosus (ThGA) (SEQ ID NO: 164), Talaromyces emersonii (TeGA) (SEQ ID NO: 165), Aspergillus niger (AnGA) (SEQ ID NO: 166), e Aspergillus awamori (AaGA) (SEQ ID NO: 167). Em algumas modalidades, por exemplo, a glicoamilase variante será derivada de uma glicoamilase parental que é uma glicoamilase de Aspergillus e a variante incluirá pelo menos uma substituição em uma posição equivalente a uma posição apresentada na SEQ ID NO: 2, e particularmente em uma posição correspondente a D4, F5, 112, D24, F29, I43, D44, P45, D46, Y47, Y49, W51, N61, Y70, G73, Q75, R76, P94, D100, K108, K114, F116, N119, R122, Q124, R125, G137, N146, Q148, Y169, N171, Q172, F175, W178, E180, V181, Q208, S211, W228, N242, E243, R245, I292, G294, K297, R309, Y310, D313, V314, Y315, Y316, N317, W321, K340, K341, T350, Q356, T363, S368, S369, N376, Y395, A398, S401, R408, N409, T412, L417, H418, W421, T430, A431, R433, I436, S451, E503, Q511, A535, A539 ou N563.
Remoção de H endógeno de açúcares N-ligados em glicoamilase de Trichoderma reesei teve um efeito de estabilização (ao olhar a Tm). Dessa forma, variantes tendo uma substituição N171D podem ter termoes- tabilidade aumentada quando comparadas com o parental de tipo selvagem. Em algumas modalidades, as variantes que têm uma ou mais substituições em sítios tendo açúcares N-ligados são fornecidas, incluindo N171D em Trichoderma reesei (SEQ ID NO: 2).Tabela 1
A presente descrição também fornece variantes de glicoamilase tendo pelo menos uma propriedade alterada (por exemplo, propriedade melhorada) quando comparadas com uma glicoamilase parental e particularmente à TrGA. Em algumas modalidades, pelo menos uma propriedade alterada (por exemplo, propriedade melhorada) é selecionada a partir do grupo consistindo em estabilidade ao ácido, estabilidade térmica e atividade específica. Tipicamente, a propriedade alterada é estabilidade ao ácido aumentada, estabilidade térmica aumentada e/ou atividade específica aumentada. A estabilidade térmica aumentada tipicamente é a temperaturas mais altas. Em uma modalidade, a estabilidade de pH aumentada é em alto pH. Em uma modalidade adicional, a estabilidade de pH aumentada é em pH baixo.
As variantes de glicoamilase da descrição também podem fornecer taxas mais altas de hidrólise de amido em baixas concentrações de substrato quando comparadas à glicoamilase parental. A variante pode ter um Vmax superior ou Km inferior do que uma glicoamilase parental quando testada sob as mesmas condições. Por exemplo, a glicoamilase variante pode ter um Vmax superior em uma faixa de temperatura de 25°C a 70°C (por exemplo, em 25°C a 35°C; 30°C a 35°C; 40°C a 50°C; em 50°C a 55°C e em 55°C a 62°C). Os valores da constante de Michaelis-Menten, Km e Vmax po- 5 dem ser facilmente determinados usando procedimentos padrão conhecidos.
Em um aspecto, a descrição relaciona-se a uma glicoamilase variante tendo estabilidade térmica alterada em temperaturas alteradas quando comparada a uma parental ou tipo selvagem. Temperaturas altera- 10 das incluem temperaturas aumentadas ou reduzidas. Em algumas modalidades, a variante de glicoamilase terá termoestabilidade melhorada, tal como retenção de pelo menos 50%, 60%, 70%, 75%, 80%, 85%, 90%, 92%, 95%, 96%, 97%, 98% ou de 99% de atividade enzimática após exposição a temperaturas alteradas ao longo de um dado período de tempo, por exem- 15 pio, pelo menos 60 minutos, 120 minutos, 180 minutos, 240 minutos, 300 minutos, etc. Em algumas modalidades, a variante tem estabilidade térmica aumentada quando comparada com a glicoamilase parental em temperaturas selecionadas na faixa de 40 a 80°C, também na faixa de 50 a 75°C e na faixa de 60 a 70°C, e tipicamente em uma faixa de pH de 4,0 a 6,0. Em al- 20 gumas modalidades, a termoestabilidade é determinada como descrito nos Exemplos.
Em algumas modalidades, variantes particularmente interessantes com relação a uma melhora na termoestabilidade incluem uma ou mais deleções, substituições ou inserções e particularmente substituições nas 25 seguintes posições na sequência de aminoácidos apresentada na SEQ ID NO: 2: D4, F5, 112, D24, F29, I43, D44, P45, D46, Y47, Y49, W51, N61, Y70, G73, Q75, R76, P94, D100, K108, K114, F116, N119, R122, Q124, R125, G137, N146, Q148, Y169, N171, Q172, F175, W178, E180, V181, Q208, S211, W228, N242, E243, R245, I292, G294, K297, R309, Y310, D313, V314, Y315, Y316, N317, W321, K340, K341, T350, Q356, T363, S368, S369, N376, Y395, A398, S401, R408, N409, T412, L417, H418, W421, T430, A431, R433, I436, S451, E503, Q511, A535, A539, ou N563; ou uma posição equivalente em uma glicoamilase parental. Em algumas modalidades, a glicoamilase parental será um homólogo de glicoamilase de Trichoderma, e em modalidades típicas, a glicoamilase parental terá identidade de sequência de pelo menos 90%, pelo menos 95%, ou pelo menos 98% à SEQ ID NO: 2.
Variantes de glicoamilase da descrição também podem incluir glicoamilases quiméricas ou híbridas com, por exemplo, um domínio de ligação a amido (SBD) de uma glicoamilase e um domínio catalítico e ligante de outra. Por exemplo, uma glicoamilase híbrida pode ser feita pela troca do SBD de AnGA pelo SBD de TrGA, produzindo um híbrido com o SBD de AnGA e o domínio catalítico e ligante de TrGA. Alternativamente, o SBD e ligante de AnGA podem ser trocados pelo SBD e ligante de TrGA.
Em outro aspecto, a descrição relaciona-se a uma glicoamilase variante tendo atividade específica alterada quando comparada com uma glicoamilase parental ou de tipo selvagem.
Em algumas modalidades, variantes particularmente interessantes com relação a uma melhora na atividade específica incluem uma ou mais deleções, substituições ou inserções e particularmente substituições nas seguintes posições na sequência de aminoácidos apresentada na SEQ ID NO: 2: D4, F5, 112, D24, F29, I43, D44, P45, D46, Y47, Y49, W51, N61, Y70, G73, Q75, R76, P94, D100, K108, K114, F116, N119, R122, Q124, R125, G137, N146, Q148, Y169, N171, Q172, F175, W178, E180, V181, Q208, S211, W228, N242, E243, R245, I292, G294, K297, R309, Y310, D313, V314, Y315, Y316, N317, W321, K340, K341, T350, Q356, T363, S368, S369, N376, Y395, A398, S401, R408, N409, T412, L417, T430, A431, H418, W421, R433, I436, S451, E503, Q511, A535, A539, ou N563; ou uma posição equivalente em uma glicoamilase parental. Em algumas modalidades, as variantes da descrição tendo atividade específica melhorada incluem uma substituição nas seguintes posições na sequência de aminoácidos a- presentada na SEQ ID NO: 2: D4, D24, I43, D44, N61, Y70, G73, Q75, R76, D100, K108, N119, Q124, N146, Q148, N171, Q172, F175, V181, Q208, S211, E243, R245, I292, G294, K297, V314, Y316, N317, K340, K341, T350, Q356, T363, S368, N376, Y395, A398, S401, N409, T412, L417, T430, A431, I436, S451, E503, Q511, A535, A539, ou N563; ou uma posição equivalente em uma glicoamilase parental. Em algumas modalidades, a glicoamilase parental compreenderá uma sequência tendo identidade de sequência de pelo menos 90% ou de 95% à sequência da SEQ ID NO: 2.
A presente descrição também se relaciona a polinucleotídeos isolados que codificam uma glicoamilase variante da descrição. Os polinucleotídeos que codificam uma glicoamilase variante podem ser preparados por técnicas estabelecidas conhecidas na técnica. Os polinucleotídeos podem ser preparados sinteticamente, tal como por um sintetizador automático de DNA. A sequência de DNA pode ser de origem genômica variada (ou cDNA) e sintética preparada por ligação de fragmentos em conjunto. Os polinucleotídeos também podem ser preparados pela reação de polimerase em cadeia (PCR) usando iniciadores específicos. Em geral, referência é feita a Minshull J., et al., (2004), Engineered protein function by selective amino acid diversification, Methods 32 (4):416-427. Além disso, diversas companhias agora sintetizam DNA, tal como Geneart AG, Regensburg, Germany.
A presente descrição também fornece polinucleotídeos isolados compreendendo uma sequência nucleotídica (i) tendo identidade de pelo menos 70% a SEQ ID NO: 4, ou (ii) sendo capaz de hibridizar a uma sonda derivada da sequência nucleotídica apresentada na SEQ ID NO: 4, sob condições de estringência intermediária à alta, ou (iii) sendo complementar a uma sequência nucleotídica tendo identidade de sequência de pelo menos 90% à sequência apresentada na SEQ ID NO: 4. Sondas úteis de acordo com a descrição podem incluir pelo menos 50, 100, 150, 200, 250, 300 ou mais nucleotídeos contíguos da SEQ ID NO: 4.
A presente descrição fornece ainda polinucleotídeos isolados que codificam glicoamilases variantes que compreendem uma sequência de aminoácidos compreendendo identidade de sequência de aminoácidos de pelo menos 80% a SEQ ID NO: 2. Em algumas modalidades, as glicoamilases variantes têm identidade de sequência de aminoácidos de pelo menos 80% a SEQ ID NO: 2. Em algumas modalidades, as glicoamilases variantes têm identidade de sequência de aminoácidos de pelo menos 90% a SEQ ID NO: 2. Em algumas modalidades, as glicoamilases variantes têm identidade de sequência de aminoácidos de pelo menos 93% a SEQ ID NO: 2. Em algumas modalidades, as glicoamilases variantes têm identidade de sequência de aminoácidos de pelo menos 95% a SEQ ID NO: 2. Em algumas modalidades, as glicoamilases variantes têm identidade de sequência de aminoácidos de pelo menos 97% a SEQ ID NO: 2. Em algumas modalidades, as gli-coamilases variantes têm identidade de sequência de aminoácidos de pelo menos 98% a SEQ ID NO: 2. Em algumas modalidades, as glicoamilases variantes têm identidade de sequência de aminoácidos de pelo menos 99% a SEQ ID NO: 2. A presente descrição também fornece vetores de expressão compreendendo qualquer um dos polinucleotídeos fornecidos acima.
A presente descrição também fornece fragmentos (isto é, por-ções) de DNA que codificam as glicoamilases variantes fornecidas neste pedido. Estes fragmentos encontram uso na obtenção de fragmentos de DNA de comprimento parcial capazes de serem usados para isolar ou identificar polinucleotídeos que codificam enzimas glicoamilase maduras descritas neste pedido a partir de células de fungos filamentosos (por exemplo, Trichoderma, Aspergillus, Fusarium, Penicillium, Schizosaccharomyces e Humicola), ou um segmento dos mesmos tendo atividade de glicoamilase. Em algumas modalidades, os fragmentos de DNA podem compreender pelo menos 50, 100, 150, 200, 250, 300 ou mais nucleotídeos contíguos. Em algumas modalidades, as porções de DNA fornecidas na SEQ ID NO: 4 podem ser usadas na obtenção de glicoamilase parental e particularmente homólogos de glicoamilase de Trichoderma de outras espécies, tais como fungos filamentosos que codificam uma glicoamilase.
De acordo com uma modalidade da descrição, um construto de DNA compreendendo um polinucleotídeo como descrito acima que codifica uma glicoamilase variante englobada pela descrição e operacionalmente ligado a uma sequência promotora é montado para transferência em uma célula hospedeira.
O construto de DNA pode ser introduzido em uma célula hospedeira usando um vetor. O vetor pode ser qualquer vetor que quando introduzido em uma célula hospedeira é tipicamente integrado no genoma de célula hospedeira e é replicado. Os vetores incluem vetores de clonagem, vetores de expressão, vetores de transporte, plasmídeos, partículas de fago, cassetes e similares. Em algumas modalidades, o vetor é um vetor de expressão que compreende sequências regulatórias operacionalmente ligadas à sequência de codificação de glicoamilase.
Exemplos de vetores de expressão e/ou integração adequados são fornecidos em Sambrook et al., (1989) supra, e Ausubel (1987) supra, e van den Hondel et al. (1991) em Bennett e Lasure (Eds.) More Gene Manipulations in Fungi, Academic Press pp. 396-428 e Patente Norte-americana N- 5.874.276. Fungal Genetics Stock Center Catalogue of Strains (FGSC, <www.fgsc.net>) também revela vetores úteis. Vetores particularmente úteis incluem vetores obtidos de, por exemplo, Invitrogen e Promega.
Vetores específicos adequados para uso em células hospedeiras fúngicas incluem vetores, tais como pFB6, pBR322, pUC18, pUC100, pDONR®201, pDONR®221, pENTR®, pGEM®3Z e pGEM®4Z. Um vetor de expressão para fins gerais úteis em Aspergillus inclui pRAX com um promotor g/aA, e em Hypocrea/Trichoderma inclui pTrex3g com um promotor cbtrt.
Plasmídeos adequados para uso em células bacterianas incluem pBR322 e de pUC19 que permitem replicação em E. coll e pE194 que, por exemplo, permite replicação em Bacillus. Em algumas modalidades, o promotor mostra atividade transcricional em uma célula hospedeira bacteriana ou fúngica e pode ser derivado de genes que codificam proteínas homólogas ou heterólogas à célula hospedeira. O promotor pode ser um promotor mutante, truncado e/ou híbrido. Os promotores supracitados são conhecidos na técnica.
Exemplos de promotores adequados úteis em células fúngicas e células fúngicas particularmente filamentosas, tais como células de Trichoderma ou Aspergillus incluem tais promotores exemplares como promotores de T. reesei cbh1, cbh2, egft, eg!2, eg5, xln1 e xln2. Outros exemplos de promotores úteis incluem promotores de genes de glicoamilase de A. awamori e A. niger (glaA) (ver, Nunberg et al., (1984) Mol. Cell Biol. 4:2306-2315 e Boel et al., (1984) EMBO J. 3:1581-1585), promotor de amilase de TACA de A. oryzae, o promotor de TPI (triose fosfato isomerase) de S. cerevisiae, os genes promotores de acetamidase de Aspergillus nidulans e genes de lípase de Rhizomucor miehei.
Exemplos de promotores adequados úteis em células bacteria- nas incluem aqueles obtidos de operon lac de E. coli; gene de alfa amilase de Bacillus licheniformis (amyL), gene de amilase de B. stearothermophilus (amyM); genes xy/A e xy/B de Bacillus subtilis, o gene de beta-lactamase, e o promotor tac.
Em uma modalidade, o promotor é aquele que é nativo à célula hospedeira. Por exemplo, quando T. reesei é o hospedeiro, o promotor é o promotor natural T. reesei. Em outra modalidade, o promotor é aquele que é heterólogo à célula hospedeira fúngica. Em algumas modalidades, o promotor será o promotor de glicoamilase parental, tal como o promotor de TrGA.
Em algumas modalidades, o construto de DNA inclui ácidos nu- cleicos que codificam para uma sequência de sinal que é uma sequência de aminoácidos ligada ao terminal amino do polipeptídeo que dirige o polipeptídeo codificado na via secretória da célula. A extremidade 5' da sequência de codificação da sequência de ácido nucleico pode incluir naturalmente uma região de codificação de peptídeo de sinal que é naturalmente ligada na região de leitura de tradução com o segmento da sequência de codificação de glicoamilase que codifica a glicoamilase secretada ou a extremidade 5' da sequência de codificação da sequência de ácido nucleico pode incluir um peptídeo sinal que é estranho à sequência de codificação. Em algumas modalidades, o construto de DNA inclui uma sequência de sinal está naturalmente associada a um gene de glicoamilase parental a partir de uma glicoamilase variante foi obtida. Em algumas modalidades, a sequência de sinal será a sequência representada na SEQ ID NO: 1 ou uma sequência tendo identidade de sequência de pelo menos 90%, pelo menos 94% e de pelo menos 98% a esta. Sequências de sinal eficazes podem incluir as sequências de sinal obtidas de glicoamilases de outras enzimas de fungos filamentosos, tais como de Trichoderma (glicoamilase de T. reesei), Humicola (celu- lase de H. insolens ou glicoamilase de H. grisea), Aspergillus (glicoamilase de A. niger e amilase TACA de A. oryzae) e Rhizopus.
Em modalidades adicionais, um construto de DNA ou vetor compreendendo uma sequência de sinal e uma sequência promotora a ser introduzida em uma célula hospedeira é derivado da mesma fonte. Em algumas modalidades, a sequência de sinal de glicoamilase nativa de um homólogo de glicoamilase de Trichoderma, tal como uma sequência de sinal de uma cepa de Hypocrea pode ser usada.
Em algumas modalidades, o vetor de expressão também inclui uma sequência de terminação. Qualquer sequência terminadora funcional na célula hospedeira pode ser usada na presente descrição. Em uma modalidade, a sequência de terminação e a sequência promotora são derivadas da mesma fonte. Em outra modalidade, a sequência de terminação é homóloga à célula hospedeira. Sequência terminadora útil inclui sequências terminado- ras obtidas dos genes oW?1 de Trichoderma reesei-, glicoamilase de A. niger ou A. awamori (Nunberg et al. (1984) supra, e Boel et al., (1984) supra), an- tranilato sintase de Aspergillus nidulans, amilase TACA de Aspergillus oryzae, ou trpC de A. nidulans (Punt et al., (1987) Gene 56:117-124).
Em algumas modalidades, um vetor de expressão inclui um marcador selecionável. Exemplos de marcadores selecionáveis típicos incluem aqueles que conferem resistência antimicrobiana (por exemplo, higromi- cina e fleomicina). Marcadores seletivos nutricionais também encontram uso na presente descrição incluindo aqueles marcadores conhecidos na técnica como amdS (acetamidase), argB (omitina carbamoiltransferase) e pyrG (oro- tidina-5'fosfato decarboxilase). Marcadores úteis em sistemas de vetor para transformação de Trichoderma são conhecidos na técnica (ver, por exemplo, Finkelstein, capítulo 6 em BIOTECHNOLOGY OF FILAMENTOUS FUNGI, Finkelstein et al. Eds. Butterworth-Heinemann, Boston, MA (1992); Kinghom et al. (1992) APPLIED MOLECULAR GENETICS OF FILAMENTOUS FUNGI, Blackie Academic and Professional, Chapman and Hall, London; Berges and Barreau (1991) Curr. Genet. 19:359-365; e van Hartingsveldt et al., (1987) Mol. Gen. Genet. 206:71-75). Em uma modalidade típica, o marcador seletivo é o gene amdS, que codifica a enzima acetamidase, permitindo células transformadas crescerem em acetamida como uma fonte de nitrogênio. O uso do gene amdS de A. nidulans como um marcador seletivo é descrito em Kelley et al., (1985) EMBO J. 4:475-479 e Penttilã et al., (1987) Gene 61:155-164.
Os métodos usados para ligar o construto de DNA compreen-dendo uma sequência de ácido nucleico que codifica uma glicoamilase variante, um promotor, sequências terminadoras e outras e inseri-los em um vetor adequado são bem conhecidos na técnica. A ligação é geralmente realizada pela ligação em sítios de restrição adequados. Se tais sítios não existirem, ligantes oligonucleotídicos sintéticos são usados conforme a prática convencional. (Ver, Sambrook (1989) supra, e Bennett and Lasure, MORE GENE MANIPULATIONS IN FUNGI, Academic Press, San Diego (1991) pp 70-76.). Adicionalmente, vetores podem ser construídos usando técnicas de recombinação conhecidas (por exemplo, Invitrogen Life Technologies, Gateway Technology).
A presente descrição também se relaciona a células hospedeiras compreendendo um polinucleotídeo que codifica uma glicoamilase variante da descrição, que são usadas para produzir as glicoamilases da descrição. Em algumas modalidades, as células hospedeiras são selecionadas a partir de células bacterianas, fúngicas, vegetais e de levedura. O termo célula hospedeira inclui tanto as células, progénie das células como protoplastos criados a partir das células que são usadas para produzir uma glicoamilase variante de acordo com a descrição.
Em algumas modalidades, as células hospedeiras são células fúngicas e células hospedeiras de fungos tipicamente filamentosos. O termo "fungos filamentosos" refere-se a todas as formas filamentosas da subdivisão Eumycotina (Ver, Alexopoulos, C. J. (1962), INTRODUCTORY MYCOLOGY, Wiley, New York). Estes fungos são caracterizados por um micélio vegetativo com uma parede celular composta de quitina, celulose e outros polissacarídeos complexos. Os fungos filamentosos da presente descrição são morfológica, fisiológica e geneticamente distintos de leveduras. Crescimento vegetativo por fungos filamentosos é pelo alongamento de hifas e o catabolismo de carbono é aeróbico obrigatório. Na presente descrição, a célula parental fúngica filamentosa pode ser uma célula de uma espécie de, mas não limitada a Trichoderma, (por exemplo, Trichoderma reesei, a forma assexuada de Hypocrea jecorina, anteriormente classificada como T. longi- brachiatum, Trichoderma viride, Trichoderma koningii, Trichoderma harzia- num) (Sheir-Neirs et al., (1984) Appl. Microbiol. Biotechnol 20:46-53; ATCC N. 56765 e ATCC N. 26921); Penicillium sp., Humicola sp. (por exemplo, H. insolens, H. lanuginosa e H. grisea); Chrysosporium sp. (por exemplo, C. lucknowense), Gliocladium sp., Aspergillus sp. (por exemplo, A. oryzae, A. niger, A. sojae, A. japonicus, A. nidulans e A. awamori) (Ala et al., (1993) Appl. Microbiol. Biotechnol. 39:738-743 e Goedegebuur et al., (2002) Genet 41:89-98), Fusarium sp., (por exemplo, F. roseum, F. graminum F. cerealis, F. oxysporuim e F. venenatum), Neurospora sp., (N. crassa), Hypocrea sp., Mucor sp., (M. miehei), Rhizopus sp. e Emericella sp. (ver também, Innis et al., (1985) Sci. 228:21-26). O termo "Trichoderma" ou "Trichoderma sp." ou "Trichoderma spp.” refere-se a qualquer gênero fúngico anteriormente ou atualmente classificado como Trichoderma.
Em algumas modalidades, as células hospedeiras serão células bacterianas gram-positivas. Exemplos não limitantes incluem cepas de S- treptomyces, (por exemplo, S. lividans, S. coelicolor e S. griseus) e Bacillus. Como usado neste pedido, "o gênero Bacillus” inclui todas as espécies dentro do gênero "Bacillus”, como conhecido por aqueles versados na técnica incluindo, mas não limitado a B. subtilis, B. licheniformis, B. lentus, B. brevis, B. stearothermophilus, B. alkalophilus, B. amyloliquefaciens, B. clausii, B. halodurans, B. megaterium, B. coagulans, B. circulans, B. lautus, e B. thurin- giensis. É reconhecido que o gênero Bacillus continua a sofrer reorganização taxonômica. Dessa forma, é pretendido que o gênero inclua espécies que foram reclassificadas incluindo, mas não limitadas a tais organismos como B. stearothermophilus, que é denominado agora "Geobacillus stearo- thermophilus".
Em algumas modalidades, a célula hospedeira é uma cepa bacteriana gram-negativa, tal como E. coli ou Pseudomonas sp. Em outras modalidades, as células hospedeiras podem ser células de levedura, tais como Saccharomyces sp., Schizosaccharomyces sp., Pichia sp. ou Candida sp.
Em outras modalidades, a célula hospedeira será uma célula hospedeira geneticamente engendrada em que os genes nativos foram inati- vados, por exemplo, pela deleção em células bacterianas ou fúngicas. Onde é desejado obter uma célula hospedeira fúngica tendo um ou mais genes inativados, métodos conhecidos podem ser usados (por exemplo, métodos revelados na Patente Norte-americana Ns 5.246.853, Patente Norte- americana Ns 5.475.101 e WO 92/06209). A inativação gênica pode ser realizada pela deleção completa ou parcial, pela inativação insercional ou por qualquer outro meio que forneça um gene não funcional para seu alvo desejado (tal que o gene seja impedido de expressão de uma proteína funcional). Em algumas modalidades, quando a célula hospedeira é uma célula Trichoderma e particularmente uma célula hospedeira T. reesei, os genes cbtrt, cbh2, egll e egl2 serão inativados e/ou tipicamente deletados. Tipicamente, células hospedeiras de Trichoderma reesei tendo proteínas quad-deletadas são apresentadas e descritas na Patente Norte-americana N- 5.847.276 e WO 05/001036. Em outras modalidades, a célula hospedeira é uma cepa deficiente em protease ou protease menos.
A introdução de um construto ou vetor de DNA em uma célula hospedeira inclui técnicas, tais como transformação; eletroporação; microin- jeção nuclear; transdução; transfecção, (por exemplo, transfecção mediada por lipofecção e mediada por DEAE-dextrina); incubação com DNA precipitado por fosfato de cálcio; bombardeio de alta velocidade com microprojéteis recobertos com DNA; e fusão de protoplasto. Técnicas de transformação gerais são conhecidas na técnica (Ver, por exemplo, Ausubel et al., (1987), supra, capítulo 9; e Sambrook (1989) supra, e Campbell et al., (1989) Curr. Genet. 16:53-56).
Métodos de transformação de Bacillus são revelados em numerosas referências incluindo Anagnostopoulos C e J. Spizizen (1961) J. Bacte- riol. 81:741-746 e WO 02/14490.
Métodos de transformação de Aspergillus são descritos em Yel- ton et al (1984) Proc. Natl. Acad. Sci. USA 81:1470-1474; Berka et al., (1991) em Applications of Enzyme Biotechnology, Eds. Kelly and Baldwin, Plenum Press (NY); Cao et al., (2000) Science 9:991-1001; Campbell et al., (1989) Curr. Genet. 16:53-56 e EP 238023. A expressão da proteína heteró- loga em Trichoderma é descrita na Patente Norte-americana Ns 6.022.725; Patente Norte-americana N2 6.268.328; Harkki et al. (1991); Enzyme Microb. Technol. 13:227-233; Harkki et al., (1989) Technol Bio. 7:596-603; EP 244.234; EP 215.594; e Nevalainen et al., "The Molecular Biology of Trichoderma and its Application to the Expression of Both Homologous and Heterologous Genes", em MOLECULAR INDUSTRIAL MYCOLOGY, Eds. Leong and Berka, Marcel Dekker Inc., NY (1992) pp. 129-148). Referência também é feita a W096/00787 e Bajar et al., (1991) Proc. Natl. Acad. Sci. USA 88:8202-8212 para transformação de cepas de Fusarium.
Em uma modalidade específica, a preparação de Trichoderma sp. para transformação envolve preparação de protoplastos do micélios fúngicos (Ver, Campbell et al., (1989) Curr. Genet. 16:53-56; Pentilla et al., (1987) Gene 61:155-164). Transformação mediada por Agrobacterium tume- faciens de fungos filamentosos é conhecida (Ver, de Groot et al., (1998) Nat. Biotechnol. 16:839-842). Referência também é feita a Patente Norte- americana N2 6.022.725 e Patente Norte-americana N2 6.268.328 para procedimentos de transformação usados com hospedeiros fúngicos filamentosos.
Tipicamente, transformantes geneticamente estáveis são construídos com sistemas de vetor pelo qual o ácido nucleico que codifica a gli- coamilase variante está integrado estavelmente em um cromossomo da cepa hospedeira. Transformantes então são purificados por técnicas conhecidas.
Em algumas modalidades adicionais, as células hospedeiras são células vegetais, tais como células de uma planta monocotiledônea (por e- xemplo, milho, trigo e sorgo) ou células de uma planta dicotiledônea (por exemplo, soja). Métodos para produzir construtos de DNA úteis na transformação de plantas e métodos para transformação vegetal são conhecidos. Alguns destes métodos incluem transferência gênica mediada por Agrobac- terium tumefaciens-, bombardeio de microprojétil, transformação de proto- plastos mediada por PEG, eletroporação e similares. Referência é feita a (Patente Norte-americana Ns 6.803.499, Patente Norte-americana Ns 6.777.589; Fromm et al (1990) Biotechnol. 8:833-839; Potrykus et al (1985) Mol. General. Genet. 199:169-177.
A presente descrição relaciona-se ainda a métodos de produção de glicoamilases variantes compreendendo transformação de uma célula hospedeira com um vetor de expressão compreendendo um polinucleotídeo que codifica uma glicoamilase variante de acordo com a descrição, opcionalmente cultivando a célula hospedeira sob condições adequadas para a produção da glicoamilase variante e opcionalmente recuperando a glicoamilase.
Nos métodos de expressão e produção da presente descrição, as células hospedeiras são cultivadas sob condições adequadas em cultivo em frasco de agitação, fermentações em pequena escala ou larga escala (incluindo fermentações contínuas, de batelada e de batelada alimentada) em fermentadores de laboratório ou industriais, com meio adequado contendo sais e nutrientes fisiológicos (ver, por exemplo, Pourquie, J. et al., BIOCHEMISTRY AND GENETICS OF CELLULOSE DEGRADATION, eds. Aubert, J. P. et al., Academic Press, pp. 71-86, 1988 e Ilmen, M. et al., (1997) Appl. Environ. Microbiol. 63:1298-1306). Meios comuns comercialmente preparados (por exemplo, caldo de Extrato de Malte de Levedura (YM), caldo de Luria Bertani (LB) e caldo de Dextrose Sabouraud (SD)) encontram uso na presente descrição. Condições de cultura representativas de células bacterianas e fúngicas filamentosas são conhecidas na técnica e podem ser encontradas na literatura científica e/ou a partir da fonte dos fungos, tais como a American Type Culture Collection e Fungal Genetics Stock Center. Em casos onde uma sequência de codificação de glicoamilase está sob o controle de um promotor induzível, agente de indução (por exemplo, um açúcar, sal metálico ou antimicrobiano), é adicionado ao meio em uma concentração eficaz para induzir a expressão de glicoamilase.
Em algumas modalidades, a presente descrição relaciona-se a métodos de produção de glicoamilase variante compreendendo cultivo de uma célula vegetal ou vegetal compreendendo um polinucleotídeo que codifica uma glicoamilase variante de acordo com a descrição sob condições adequadas para a produção da variante e opcionalmente recuperação da glicoamilase.
Em algumas modalidades, a fim de avaliar a expressão de uma glicoamilase variante por uma linhagem celular que foi transformada com um polinucleotídeo que codifica uma glicoamilase variante englobada pela descrição, ensaios são realizados em nível proteico, em nível de RNA e/ou pelo uso de bioensaios funcionais particulares para atividade e/ou produção de glicoamilase. Alguns destes ensaios incluem Northern blotting, dot blotting (análise de DNA ou RNA), RT-PCR (reação de polimerase em cadeia de transcriptase reversa), ou hibridização in situ, usando uma sonda apropriadamente marcada (baseada na sequência de codificação de ácido nucleico) e Southern blotting convencional e autorradiografia.
Além disso, a produção e/ou expressão de uma glicoamilase variante podem ser medida em uma amostra diretamente, por exemplo, por ensaios que diretamente medem a redução de açúcares, tais como glicose em meio de cultura e por ensaios para medir a atividade, expressão e/ou produção de glicoamilase. Em particular, a atividade de glicoamilase pode ser analisada pelo método de ácido 3,5-dinitrossalicílico (DNS) (ver, Goto et al., (1994) Biosci. Biotechnol. Biochem. 58:49-54). Em modalidades adicio nais, a expressão proteica é avaliada por métodos imunológicos, tais como marcação imunohistoquímica de células, seções de tecido ou imunoensaio de meio de cultura de tecido, (por exemplo, por Western blotting ou ELISA). Tais imunoensaios podem ser usados qualitativa e quantitativamente para avaliar a expressão de uma glicoamilase. Os detalhes de tais métodos são conhecidos por aqueles versados na técnica e muitos reagentes para praticar tais métodos estão comercialmente disponíveis.
As glicoamilases da presente descrição podem ser recuperadas ou purificadas de meios de cultura por uma variedade de procedimentos conhecidos na técnica incluindo centrifugação, filtração, extração, precipitação e similares.
As glicoamilases variantes podem ser usadas em composições enzimáticas incluindo, mas não limitadas a composições de hidrólise e sacarificação de amido, composições de limpeza e detergentes (por exemplo, detergentes de lavanderia, detergentes de lavagem de prato, e composições de limpeza pesada), composições de fermentação alcoólica, e em composições de ração. Além disso, as glicoamilases variantes podem ser usadas em aplicações de padaria, tais como produção de pão e bolo, preparação de cerveja, serviço de saúde, têxtil, processos de conversão de dejetos ambientais, processamento de biopolpas e aplicações de conversão de biomassa.
Em algumas modalidades, uma composição enzimática incluindo uma glicoamilase variante englobada pela descrição obtida em meios de culturas ou recuperada e purificada do meio de cultura será opcionalmente usada em combinação com qualquer uma ou combinação das seguintes enzimas - alfa amilases, proteases, pululanases, isoamilases, celulases, hemi- celulases, xilanases, ciclodextrina glicotransferases, lipases, fitases, lacases, oxidases, esterases, cutinases, xilanases, enzima de hidrólise de amido granular e outras glicoamilases. Em uma modalidade, as proteases são proteases fúngicas ácidas (AFP). Em uma modalidade adicional, as proteases fúngicas ácidas são de Trichoderma (por exemplo, NSP-24, ver também US 2006/0154353, publicado em 13 de julho de 2006, incorporado neste pedido por referência). Em uma modalidade adicional, a fitase é de Buttiauxiella. spp. (por exemplo, BP-17, ver também variantes reveladas na publicação de patente PCT WO 2006/043178).
Em algumas composições representativas, as glicoamilases variantes da descrição serão combinadas com uma alfa amilase, tal como alfa amilases fúngicas (por exemplo, Aspergillus sp.) ou alfa amilases bacteria- nas (por exemplo, Bacillus sp., tais como B. stearothermophilus, B. amyloli- quefaciens e B. licheniformis) e variantes e híbridos dos mesmos. Em uma modalidade, a alfa amilase é uma alfa amilase ácida estável. Em uma modalidade, a alfa amilase é uma enzima de hidrólise de amido granular (GSHE). Em uma modalidade, a alfa amilase é alfa amilase de Aspergillus kawachi (AKAA), ver US 7.037.704. As alfa amilases comercialmente disponíveis contempladas para uso nas composições da descrição são conhecidas e incluem GZYME G997, SPEZYME FRED, SPEZYME XTRA, STARGEN (Danisco US, Inc, Genencor Division), TERMAMYL 120-L e SUPRA (No- vozymes, Biotech.) e VIRIDIUM (Diversa).
Em outras modalidades, as glicoamilases variantes da descrição podem ser combinadas com outras glicoamilases. Em algumas modalidades, as glicoamilases da descrição serão combinadas com uma ou mais glicoamilases derivadas de cepas de Aspergillus ou variantes das mesmas, tais como A. oryzae, A. niger, A. kawachi, e A. awamorf, glicoamilases derivadas de cepas de Humicola ou variantes da mesma, particularmente, H. grisea, tal como a glicoamilase tendo identidade de sequência de pelo menos 90%, 93%, 95%, 96%, 97%, 98% e de 99% a SEQ ID NO: 3 revelada em WO 05/052148; glicoamilases derivadas de cepas de Talaromyces ou variantes das mesmas, particularmente T. emersonii; glicoamilases derivadas de cepas de Athelia e particularmente A. rolfsif, glicoamilases derivadas de cepas de Penicillium, particularmente P. chrysogenum.
Em particular, as glicoamilases variantes podem ser usadas para processos de conversão de amido, e particularmente na produção de dextrose para xaropes de frutose, açúcares especiais e em álcool e outro produ- to final (por exemplo, ácido orgânico, ácido ascórbico, e aminoácidos) produção da fermentação do amido contendo substratos (G.M.A van Beynum et al., Eds. (1985) STARCH CONVERSION TECHNOLOGY, Marcel Dekker Inc. NY. Dextrinas produzidas usando as composições de glicoamilase variante da descrição podem resultar em rendimentos de glicose de pelo menos 80%, pelo menos 85%, pelo menos 90% e pelo menos 95%. A produção de álcool pela fermentação de substratos de amido usando glicoamilases englobadas pela descrição pode incluir a produção de álcool combustível ou álcool potável. Em algumas modalidades, a produção de álcool será maior quando a glicoamilase variante for usada sob as mesmas condições que a glicoamilase parental. Em algumas modalidades, a produção de álcool será entre aproximadamente 0,5% e 2,5% melhor, incluindo mas não limitada a 0,6%, 0,7%, 0,8%, 0,9%, 1,0%, 1,1%, 1,2%, 1,3%, 1,4%, 1,5%, 1,6%, 1,7%, 1,8%, 1,9%, 2,0%, 2,1%, 2,2%, 2,3%, e 2,4% mais álcool do que a glicoamilase parental.
Em uma modalidade representativa, as glicoamilases variantes da descrição encontrarão uso na hidrólise do amido de vários substratos baseados em vegetais, que são usados para a produção de álcool. Em algumas modalidades, os substratos baseados em vegetais incluirão milho, trigo, cevada, centeio, milo, arroz, cana de açúcar, batatas e combinações dos mesmos. Em algumas modalidades, o substrato baseado em vegetal será material vegetal fracionado, por exemplo, um grão de cereal, tal como milho, que é fracionado em componentes, tais como fibra, germe, proteína e amido (endosperma) (Patente Norte-americana Na 6.254.914 e Patente Norte- americana N- 6.899.910). Métodos de fermentações alcoólicas são descritos em THE ALCOHOL TEXTBOOK, A REFERENCE FOR THE BEVERAGE, FUEL AND INDUSTRIAL ALCOHOL INDUSTRIES, 3rd Ed., Eds K.A. Jacques et al., 1999, Nottingham University Press, UK. Em certas modalidades, o álcool será etanol. Em particular, os processos de produção de fermenta-ção alcoólica são caracterizados como processos de moagem úmida ou moagem seca. Em algumas modalidades, a glicoamilase variante será usada em um processo de fermentação de moagem úmida e em outras modalida- des, a glicoamilase variante encontrará uso em um processo de moagem seca.
Moagem seca de grão envolve diversas etapas básicas, que geralmente incluem: moagem, cozimento, liquefação, sacarificação, fermentação e separação de líquidos e sólidos para produzir álcool e outros coprodu- tos. Material vegetal e particularmente grãos de cereais inteiros, tais como milho, trigo ou centeio são triturados. Em alguns casos, o grão pode ser primeiro fracionado em partes componentes. O material vegetal triturado pode ser moído para obter uma partícula grossa ou fina. O material vegetal triturado é misturado com líquido (por exemplo, água e/ou vinhaça fina) em um tanque de pasta fluida. A pasta fluida é submetida a altas temperaturas (por exemplo, 90°C a 105°C ou maior) em um fogareiro junto com enzimas lique- fatoras (por exemplo, alfa amilases) para solubilizar e hidrolisar o amido no grão a dextrinas. A mistura é resfriada e tratada ainda com enzimas de sacarificação, tais como glicoamilases englobadas pela presente descrição, para produzir a glicose. O mosto contendo glicose então pode ser fermentado por aproximadamente 24 a 120 horas na presença de micro-organismos de fermentação, tais como microrganismo de produção de etanol e particularmente levedura (Saccharomyces spp.). Os sólidos no mosto são separados da fase líquida e álcool, tal como etanol e os coprodutos úteis, tais como grãos de destilarias são obtidos.
Em algumas modalidades, a etapa de sacarificação e a etapa de fermentação são combinadas e o processo é referido como sacarificação e fermentação simultâneas ou sacarificação, propagação de levedura e fermentação simultâneas.
Em outras modalidades, a glicoamilase variante é usada em um processo para hidrólise de amido em que a temperatura do processo é realizada a uma temperatura entre 30°C e 75°C e também em uma temperatura entre 40°C e 65°C em uma faixa de pH entre pH 3,0 e pH 6,5. Os processos de fermentação em algumas modalidades incluem moagem de um grão de cereal ou grão fracionado e combinação do grão triturado de cereal com líquido para formar uma pasta fluida que então é misturada em um vaso único com uma glicoamilase variante de acordo com a descrição e opcionalmente outras enzimas tais como mas não limitadas a alfa amilases, outras glicoamilases, fitases, proteases, pululanases, isoamilases ou outras enzimas tendo atividade de hidrólise de amido granular e levedura para produzir etanol e outros coprodutos (Patente Norte-americana N— 4.514.496, WO 04/081193 e WO 04/080923).
Em algumas modalidades, a descrição pertence a um método de sacarificação de uma solução líquida de amido, que compreende uma etapa de sacarificação enzimática usando uma glicoamilase variante da descrição.
A presente descrição também fornece uma ração animal com-preendendo pelo menos uma glicoamilase variante englobada pela descri-ção. Métodos de uso de uma enzima glicoamilase na produção de rações compreendendo amido são fornecidos em WO 03/049550, depositado em 13 de dezembro de 2002 (neste pedido incorporado por referência em sua totalidade). Resumidamente, a variante de glicoamilase é misturada com uma ração compreendendo amido. A glicoamilase é capaz de degradar o amido resistente para uso pelo animal.
Outros objetos e vantagens da presente descrição são evidentes da presente Relatório Descritivo.
Os seguintes exemplos são fornecidos a fim de demonstrar e ilustrar ainda certas modalidades e aspecto representativos da presente descrição e não devem ser interpretados como limitação do escopo do mesmo.
Na descrição e seção experimental que se segue, as seguintes abreviaturas aplicam-se: GA (glicoamilase); GAU (unidade de glicoamilase); % por peso (porcentagem por peso); °C (graus Centígrados); rpm (rotações por minuto); H2O (água); dHaO (água deionizada); dlHaO (água deionizada, filtração por Milli-Q); aa ou AA (aminoácido); pb (par de bases); Kb (par de quilobases); kD (quilodáltons); g ou grama (gramas); μg (microgramas); mg (miligramas); μL (microlitros); ml e mL (mililitros); mm (milímetros); μm (mi- crômetro); M (molar); mM (milimolar); μM (micromolar); U (unidades); V (volts); PM (peso molecular); seg(s) ou s(s) (segundo/segundos); min(s) ou m(s) (minuto/minutos); hora(s) ou h(s) (hora/horas); DO (oxigênio dissolvido); ABS (Absorvância); EtOH (etanol); PSS (solução salina fisiológica; m/v (massa/volume); e MTP (placa de microtítulo); N (Normal); DP1 (monossaca- rideos); DP2 (dissacarideos); DP> 3 (oligossacarídeos, açúcares que têm um grau de polimerização maior do que 3); ppm (partes por milhão).
Os métodos usados para fornecer variantes são descritos abaixo. Entretanto, deve ser observado que métodos diferentes podem ser usados para fornecer variantes de uma molécula parental e a descrição não é limitada aos métodos usados nos exemplos. É pretendido que quaisquer meios adequados para produção de variantes e seleção de variantes possam ser usados.
As soluções de reagente foram: tampão de NaAc; tampão de acetato de sódio 200 mM pH 4,5; Substrato: p-nitrofenil-a-D-glicopiranosídeo 50 mM (Sigma N-1377) em tampão de NaAc (0,3 g/20ml) e solução de parada: tampão de glicina-NaOH 800 mM pH 10. 30 μl de sobrenadante filtrado fresco foram colocados em MTP de fundo chato de 96 poços. A cada um dos poços de 50 μl de tampão de NaAc e 120 μl de substrato foram adicionados e incubados por 30 minutos a 50°C (Thermolab systems iEMS Incuba- tor/shaker HT). A reação foi terminada pela adição de 100 μl de solução de parada. A absorvância foi medida em 405 nm em um leitor de MTP (Molecular Devices Spectramax 384 plus) e a atividade foi calculada usando um coeficiente de extinção molar de 0,011 μM/cm.
Sobrenadante bruto (100 μl) foi adicionado a 100 μl de tampão de NaAc 50 mM pH 4,5. A amostra foi igualmente dividida em mais 2 MTP. Uma MTP (placa inicial) foi incubada por 1 hora a 4°C e outra MTP (placa residual) foi incubada a 60°C (Thermolab systems iEMS Incubator/shaker HT) por 1 hora. A placa residual foi resfriada por 15 minutos em gelo. A atividade é medida em ambas as placas usando o ensaio de aplicação de etanol descrito abaixo, com a seguinte modificação: a quantidade da amostra tomada para o ensaio de termoestabilidade é 25 μl e a quantidade de tampão de NaAc 30 mM pH 4,0 é 35 μl.
A termoestabilidade é calculada como % de atividade residual como se segue:
O material sobrenadante bruto é testado para a glicose restante no meio de cultura após período de crescimento. Se glicose remanescente for encontrada, o valor de absorvância é subtraído dos valores de absorvân- cia medidos tanto da atividade inicial como da atividade residual.
A solução de reagente foi a solução de trabalho Bradford Quickstart (BioRad cat#500-0205). 100 μl de sobrenadante filtrado de 10 kD foram colocados em uma nova placa de fundo chato de 96 poços. A cada um dos poços, 200 μl de reagente foram adicionados e incubados por 5 minutos à temperatura ambiente. A absorvância foi medida em 595 nm em um leitor de MTP (Molecular Devices Spectramax 384 plus). Concentrações proteicas foram calculadas de acordo com uma curva padrão de Albumina Séri- ca Bovina (BSA) (0 a 50 μg/ml).
Coquetel de hexoquinase: 10 a 15 minutos antes do uso, 90 ml de água foram adicionados a um recipiente BoatIL glucose HK R1 (conjunto IL test glucose (HK), Instruments Laboratory # 182507-40) e suavemente misturados. 85 μl do coquetel de hexoquinase foram adicionados a 100 μl de dH2O. 15 μl da amostra foram adicionados às misturas e incubados por 5 min no escuro à temperatura ambiente. Absorvância foi lida em 340 nm em um leitor de MTP. Concentrações de glicose foram calculadas de acordo com uma curva padrão de glicose (0 a 1 mg/ml).
Para preparar a solução estoque 8%, 8 g de amido de milho solúvel (Sigma #S4180) foram suspensos em 40 ml de dH2O à temperatura ambiente. Cinquenta mililitros de dH2O fervente foram adicionados à pasta fluida em um frasco de 250 ml e cozidas por 5 minutos. A solução de amido foi resfriada a 25°C e o volume ajustado a 100 ml com dH2O. A solução de trabalho solúvel de amido 4% (m/v) foi preparada pela diluição (1:1) da solução estoque com tampão de acetato de sódio 100 mM pH 3,7.
Para o ensaio de rastreamento, 5 μl de sobrenadante bruto foram diluídos com 175 μl de tampão de NaAc 50 mM pH 4,5 em uma MTP de fundo chato de 96 poços. Sessenta microlitros desta diluição foram adicionados a 120 μl de amido de milho solúvel 4% e incubados em 900 rpm por 2 h a 32°C (Thermolab systems iEMS Incubator/shaker HT). A reação foi parada pela adição de 90 μl de Solução de Parada gelada a 4°C (tampão de Glicina-NaOH 800 mM, pH 10). A amostra foi colocada no gelo. O amido foi centrifugado sob 1118 x g a 15°C por 5 minutos (SIGMA 6K15) e 15 μl de sobrenadante foram usados no ensaio de atividade de hexoquinase descrito acima para determinar o conteúdo de glicose.
O material sobrenadante bruto foi testado para glicose remanescente no meio de cultura após o período de crescimento. Se glicose remanescente for encontrada, a quantidade de glicose produzida pela glicoamilase não foi calculada.
Para preparar a solução estoque 8%, 8 g de amido solúvel (Sigma #S4180) foram suspensos em 40 ml de água à temperatura ambiente. Então, 50 ml de dH2O fervente foram adicionados à pasta fluida em um frasco de 250ml e cozidas por 5 min. A solução de amido foi resfriada a 25°C e o volume ajustado a 100 ml com dH2O. A solução de trabalho de amido solúvel 4% (m/v) foi preparada pela diluição da solução estoque 1:1 com tampão de acetato de sódio 100 mM pH 4,5.
Cinquenta microlitros de tampão de NaAc 80 mM pH 4,5 foram colocados em uma nova placa de fundo chato de 96 poços. A cada poço, 120 μl de amido de milho solúvel 4% e 5 μl de sobrenadante filtrado de 10 kD foram adicionados e incubados por 1 h a 60°C. A reação foi parada pela adição de 90 μl de Solução de Parada gelada a 4°C (tampão de Glicina- NaOH 800 mM, pH 10,0). A amostra foi colocada no gelo por 30 min. O amido foi centrifugado abaixo de 716 rpm a 15°C por 5 minutos (Sigma 6K15, centrífuga) e 15 μl do sobrenadante foram usados no ensaio de atividade de hexoquinase descrito neste pedido para determinar o conteúdo de glicose.
O cassete de expressão de TrGA composto da sequência de DNA (SEQ ID NO: 4) codificando peptídeo sinal de TrGA, a pró-sequência, e a proteína madura, incluindo o domínio catalítico, região ligante e domínio de ligação a amido, foi clonado em pDONR®201, um vetor Gateway® Entry (ln- vitrogen, Carlsbad, CA, USA). O cassete de expressão de TrGA foi clonado no vetor Gateway de destino compatível pREP3Y-DEST (FIG. 3) pelo reação de recombinação do Gateway® LR.
O vetor de expressão pRep3Y-TrGA (Figura 3B) permitiu a ex-pressão da proteína TrGA (SEQ ID NO: 2) em Schizosaccharomyces pomba.
Sessenta e cinco bibliotecas de mutagênese saturada de sítio de TrGA (SSM) foram construídas usando o vetor de entrada pDONR-TrGA (FIG. 2) como um molde e os iniciadores listados na Tabela 2. Os iniciadores de mutagênese usados nos experimentos, todos contêm o código de sequência de DNA triplo NNS (N = A, C, T, G e S=C ou G) na posição que corresponde ao códon da sequência de TrGA a ser mutada (SEQ ID NO: 2) e iniciou incorporação randômica de nucleotídeos naquela posição. A construção de cada biblioteca SSM começou com duas amplificações por PCR u- sando o iniciador de mutagênese Gateway de sentido direto (pDONR201- FW) e um de sentido reverso específico (Tabela 3), e o iniciador Gateway de sentido reverso (pDONR201-RV) e um iniciador de mutagênese de sentido direto específico (Tabela 2) (posições iguais nos iniciadores de mutagênese). Fusão de DNA polimerase de alta fidelidade (Finnzymes OY, Espoo, Finland) foi usado para amplificação por PCR (iniciadores 0,2 μM, 25 ciclos) de acordo com o protocolo fornecido por Finnzymes. Resumidamente, 1 μl (SEQ ID NO: 1), de fragmento de DNA de ambas as misturas de PCR espe cífica, ambas direcionadas no mesmo códon foi adicionado a 48 μl da solução de reação de PCR fresca em conjunto com iniciadores Gateway FW e Gateway RV (Invitrogen) e misturados. Esta fusão de amplificação por PCR (22 ciclos) resultou em um fragmento de DNA de cassete de expressão line- ar com um códon de TrGA específico randomicamente mutado e sítios de recombinação de Gateway únicos em ambas as extremidades. A purificação deste fragmento de DNA (ChargeSwitch® PCR clean-up, Invitrogen, Carlsbad USA) e uma reação de recombinação BP (Invitrogen, Carlsbad, USA) com pDONR201 (Invitrogen) gerou um DNA multimérico circular (vetor de entrada) que foi posteriormente transformado em E. coli de eficiência Máxi-ma DH5a (Invitrogen) e plaqueado em meio 2xTY [Bacto Triptona (Difco) 16 g/L, Extrato de Levedura de Bacto (Difco) 10 g/L, NaCL 5 g/L] suplementado com canamicina 50 μg/mL. Tabela 2. Iniciadores de sentido direto usados para gerar bibliotecas de SSM de TrGA
 Tabela 3. Iniciadores usados para gerar bibliotecas de SSM de TrGA
Tabela 3. Iniciadores usados para gerar bibliotecas de SSM de TrGA
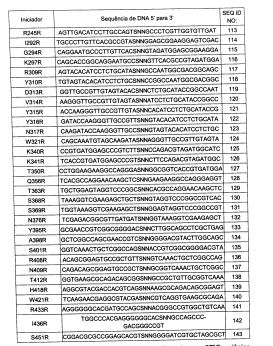
Para cada biblioteca, após incubação noturna a 37°C, colônias foram agrupadas pela ressuspensão dos clones em PSS. Das E. coli trans- formantes agrupadas, plasmídeo total foi isolado (Qiagen) usando técnicas padrão. Resumidamente 1 μl da solução de plasmídeo foi adicionado a 1 μl 5 da solução de vetor de destino pRep3Y (Figura 1A) e adicionado à mistura de enzima LR CLONASE® II de acordo com o protocolo fornecido pela Invi-trogen. Um DNA multimérico circular foi gerado e transformado para E. coli de Eficiência Máxima DH5a como descrito pelo fornecedor.
Após incubação noturna a 37°C, 96 colônias únicas de cada bi-blioteca foram selecionadas de placas de ágar 2xTY com ampicilina 100 μg/ml e cultivadas por 24 horas a 37°C em uma MTP contendo 200 μl de meio 2xTY com ampicilina 100 μg/ml. As culturas foram usadas para análi-ses de sequência (BaseClear B.V., Leiden, Netherlands).
Os números de biblioteca variaram de 1 a 65 com uma adição referindo-se ao códon da sequência de TrGA que é randomicamente muta- do. Após seleção, cada biblioteca incluiu um máximo de 19 variantes de Tr-GA. Estas variantes foram individualmente transferidas para Schizosaccha- romyces pombe de acordo com instrução dos fabricantes. (Zymo Research, Orange CA. USA).
Transformações de S. pombe foram plaqueadas em meio seleti-vo (ágar EMM, Qbiogene, Irvine, USA Cat. N- 4110-232) e incubadas a 28°C por 4 dias. Transformantes foram purificados da placa de transformação pela semeadura das colônias no ágar EMM.
Transformantes de S. pombe foram inoculados em placas de microtítulo de 96 poços (MTP) contendo meio seletivo (2xEMM-C) [Caldo EMM 64,4 g/L (Qbiogene Cat. N2: 4110-032), 0,62 g/L Mistura de Suplemento Completa (CSM-HIS-LEU-TRP, Qbiogene, Cat. N2 4530-122)] e incubados durante a noite a 28°C. Da placa de microtítulo incubada durante a noite, 200 μl de cultura de S. pombe cultivada foram inoculados em 20 ml de meio líquido 2xEMM-C em um frasco de agitação de 100 ml e incubados por 4 dias a 26°C em 280 rpm em uma incubadora de agitação Multitron (Infors AG, Bottmingen, Sweden). Da cultura cultivada, 2 ml de cultura de S. pombe foram amostrados e centrifugados por 5 minutos em 14.000 rpm (Sigma). O sobrenadante foi transferido em uma configuração de filtro de Vivaspin 500 HT de 10 kD (VivaScience AG, Hannover, Germany) e centrifugado por 25 minutos em 1000 g. O retentor foi diluído de volta ao volume de partida original com NaAc 50 mM pH 4,5 suplementado com Tween-80 0,015%. Esta solução foi usada nos diferentes ensaios.
(A) Experimentos foram conduzidos para a construção de variantes de TrGA carregando combinações das seguintes mutações de sítio único: Q172F; Q208N; S211R e V314H. Uma revisão das variantes é mostrada abaixo: a) Q172F; Q208N b) Q172F; S211R c) Q172F; V314H d) Q208N; S211R e) Q208N V314H f)S211R; V314H g) Q172F; Q208N; S211R h) Q172F; Q208N; V314H i) Q172F; S211R; V314H j) Q208N; S211R;V314H k) Q172F; Q208N; S211R; V314H
O conjunto mutagênese sítio-dirigida multi Quikchange® (QCMS) (Stratagene) foi usado para construir a biblioteca. Os iniciadores 5' fosforila- dos usados para criar a biblioteca são mostrados na Tabela 4. Resultados ótimos em termos de incorporação de iniciadores de comprimento total bem como redução significante em erros derivados do iniciador foram obtidos pelo uso de HPLC, PAGE ou qualquer outro tipo de iniciadores purificados (Invitrogen). Tabela 4. Iniciadores usados para construir variantes combinatórias selecionadas
O plasmídeo molde pDONR-TrGA (Figura 2) foi usado para construir a biblioteca combinatória usando o conjunto QCMS de Stratagene. A biblioteca foi construída como descrito pelo fornecedor com concentrações de iniciador modificadas usadas nas reações. Especificamente, 4 μl de pDONR-TrGA (25 a 50 ng) foram misturados com 11 μl de água destilada estéril; 1,5 μl de dNTP; 2,5 μl de tampão QCMS 10*; 1 μl da enzima misturada e 1 μl de cada mistura de iniciador mutante fornecendo um total de 100ng de iniciadores em cada reação. As condições de PCR foram 95°C por 1 minuto, seguido por 30 ciclos de 95° por 1 minuto, 55°C por 1 minuto, e 65°C por 6 minutos, em um termociclador MJ Research usando tubos de PCR de 0,2 ml de parede fina. O produto de reação foi digerido com 1 μl de Dpn1 do conjunto QCMS pela incubação a 37°C durante a noite. Um conjunto de purificação por PCR (Qiagen) foi usado para a purificação da amostra e um segundo ciclo de digestão foi realizado com Dpn1 (Stratagene) por 1 hora a 37°C.
A mistura de reação foi transformada em E. coli de eficiência máxima DH5a (Invitrogen) e plaqueada em ágar seletivo (2xTY suplementa- do com 50 μg de canamicina/ml). Após incubação noturna a 37°C, 96 colônias únicas foram selecionadas para análise de sequência (BaseClear B.V., Leiden, Netherlands). As variantes combinatórias foram clonadas e expressas em uma cepa hospedeira de T. reesei como descrito abaixo e no WO 5 06/060062. (B) seis bibliotecas combinatórias adicionais (Tabela 5) foram sinteticamente construídas por Geneart (Regensburg, Germany) e foram testadas para estabilidade térmica e em ensaios de aplicação de etanol e de adoçante como descritos neste pedido. Tabela 5. Bibliotecas combinatórias
A molécula de TrGA parental sob as condições descritas tinha uma atividade residual entre 15 e 44% (variação diária). O índice de desem- 15 penho foi calculado com base na termoestabilidade da TrGA do mesmo lote.
Os índices de desempenho são os quocientes PI = (Atividade residual de variantes)/(atividade residual de TrGA). Um índice de desempenho > 1 indica uma estabilidade melhorada. Variantes que têm um índice de desempenho de estabilidade térmica de mais de 1,0 são mostradas na Tabela 6 se- 20 guinte. Tabela 6. Classificação de termoestabilidade
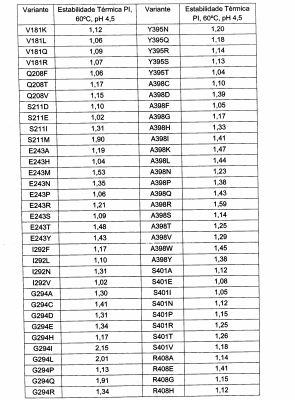


Variantes foram testadas em um ensaio de rastreamento de etanol usando os ensaios descritos acima. A tabela 7 mostra os resultados do 5 ensaio de rastreamento de variantes com um índice de Desempenho (PI) > 1,0 comparado ao PI da TrGA parental. O PI é uma medida da atividade específica (atividade/mg de enzima). O PI da atividade específica é o quociente "atividade variante-específica/atividade tipo selvagem-específica". O PI de atividade específica é 1,0 e uma variante com um Pl> 1,0 tem uma atividade específica que é maior do que a TrGA parental. A atividade específica é a atividade medida pelo ensaio de rastreamento de etanol dividida pelos resultados obtidos no ensaio de Bradford descrito acima.Tabela 7, Rastreamento de Etanol



As variantes foram testadas em um ensaio de rastreamento de 5 adoçante como descrito mais acima. A tabela 8 mostra os resultados do en- saio de rastreamento em que as variantes com um índice de Desempenho (Pl)> 1,00 comparado com o PI da TrGA parental são mostrados. O PI é uma medida da atividade específica (atividade/mg de enzima). O PI da atividade específica é o quociente "atividade variante-específica/atividade tipo selvagem-específica". O PI da atividade específica é 1,0 e uma variante com um Pl> 1,0 tem uma atividade específica que é maior do que a TrGA parental.Tabela 8. Rastreamento de adoçante

O cassete de expressão de TrGA compreendendo a sequência de DNA SEQ ID NO: 4 foi clonado em pDQNR®201, um vetor Gateway® Entry (Invitrogen, Carlsbad, CA). O cassete de expressão de TrGA foi clonado no vetor de destino compatível Gateway pTrex3g-DEST (Figura 5), que também é descrito em WO 06/060062, pela reação de recombinação de Gateway® LR. O vetor de expressão pTrex3g-TrGA (Figura 5) permitiu a expressão da proteína TrGA (SEQ ID NO: 2) em um hospedeiro Trichoderma reesei. Vetores foram construídos, os quais incluíram cDNA modificado de TrGA que codifica pelo menos as seguintes variantes (1) V314H; (2) S211R; (3) Q208Ne(4) Q172F.
Um vetor de expressão contendo uma GA variante foi transfor-mado em uma cepa hospedeira de T. reesei derivada de RL-P37 (IA52) e tendo várias deleções gênicas (Δcb/?1, ΔαW?2, Δeg/1, Δeg/2) usando bombardeio de partícula pelo Sistema PDS-1000/Helium (BioRad Cat. N- 165- 02257). O protocolo é delineado abaixo, e referência também é feita nos e- xemplos 6 e 11 do WO 05/001036.
Uma suspensão de esporos (aproximadamente 5x108 espo- ros/ml) da cepa de T. reesei foi preparada. Cem a duzentos microlitros de suspensão de esporos foram espalhados sobre o centro de placas de meio de acetamida Meio Mínimo (MM). O meio de acetamida MM tinha a seguinte composição: acetamida 0,6 g/L; CsCI 1,68 g/L; glicose 20 g/L; KH2PO4 20 g/L; CaCl2*2H2O 0,6 g/L; solução de elementos-traço 1000X 1 ml/L: ágar20 g/L; e pH 5,5; solução salina de elemento-traço 400X 1 ml/L; ácido cítrico 175 g/L, FeSO4.7H2O 200g/L, ZnSO4*7H2O 16 g/L, CuSO4«5H2O 3,2 g/L, MnSO4*H2O 1,4 g/L, H3BO3 0,8 g/L. A suspensão de esporos foi deixada secar na superfície do meio de acetamida MM.
A transformação seguiu a instrução dos fabricantes. Resumida-mente, 60 mg de partículas de tungsténio M10 foram colocados em um tubo de microcentrífuga. 1 mL de etanol foi adicionado e permitido ficar por 15 minutos. As partículas foram centrifugadas em 15.000 rpm por 15 segundos. O etanol foi removido e as partículas foram lavadas três vezes com dHaO estéril antes que 1 mL de glicerol 50% (v/v) estéril fosse adicionado. 25 μl da suspensão de partícula de tungsténio foram colocados em um tubo de mi- crocentrífuga. Enquanto vortexando continuamente, o seguinte foi adicionado: 0,5 a 5 μl (100 a 200 ng/μl) de DNA plasmidial, 25 μl de CaCl2 2,5 M e 10 μl de espermidina 0,1 M. As partículas foram centrifugadas por 3 segundos. O sobrenadante foi removido e as partículas foram lavadas com 200 μl de etanol 70% (v/v) e centrifugadas por 3 segundos. O sobrenadante foi removido e 24 μl de etanol 100% foram adicionados, misturados por pipetagem, e o tubo foi colocado em um banho ultrassónico, alíquotas de 8 μl de partículas foram removidas e colocadas sobre o centro de discos de macroveículo que foram mantidos em um dessecador. Uma vez que a suspensão de tungstênio/DNA tinha secado, o disco de microveículo foi colocado na câmara de bombardeio junto com a placa da acetamida MM com esporos e o processo de bombardeio foi realizado de acordo com instruções dos fabricantes. Após bombardeio dos esporos plaqueados com as partículas de tungstênio/DNA, as placas foram incubadas a 28°C. As colônias transformadas foram transferidas para placas frescas de acetamida MM após 4 dias (Pentillã et al. (1987) Gene 61:155-164).
Após cultivo por 5 dias, placas de acetamida MM transformantes exibindo morfologia estável foram inoculados em frascos de agitação de 250 ml contendo 30 ml de meio Proflo. O meio Proflo continha: a-lactose 30 g/L; (NH4)2SO46,5 g/L; KH2PO42 g/L; MgSO4«7H2O 0,3 g/L; CaCI2’2H2O 0,2 g/L; solução salina de elemento-traço 400X 1 ml/L: ácido cítrico 175 g/L, Fe- SO4«7H2O 200g/L, ZnSO4«7H2O 16 g/L, CuSO4«5H2O 3,2 g/L, MnSO4’H2O 1,4 g/L, H3BO3 0,8 g/L; Tween 80 10% 2 ml/L; farinha de semente de algodão ProFIo 22,5 g/L (Traders protein, Memphis, TN); CaCOs 0,72 g/L. Após dois dias de cultivo a 28°C e 140 rpm, 10% da cultura de Proflo foram transferidos para um frasco de agitação de 250 ml contendo 30 ml de Meios Defi- nidos de Lactose. A composição de Meios definidos de Lactose foi como se segue (NH4)2SO4 5 g/L; tampão 1,4-Piperazinabis(ácido propanossulfônico) 33 g/L; casaminos ácidos 9 g/L; KH2PO4 4,5 g/L; MgSO4*7H2O 1,0 g/L; anti- espumante Mazu DF60-P 5 ml/L (Mazur Chemicals, IL); solução de elemento-traço 1000X; pH 5,5; 40 ml/L de solução de lactose 40% (p/v) foram adicionados ao meio após esterilização. Os frascos de agitação de Meio Definido de Lactose foram incubados a 28°C, 140 rpm por 4 a 5 dias.
Amostras do sobrenadante de cultura foram misturadas com um volume apropriado de tampão de carregamento de amostra 2X com agente redutor. O micélio foi removido por centrifugação e o sobrenadante foi analisado para proteína total (Conjunto de Ensaio de Proteína BCA, Pierce Cat. N- 23225).
A atividade de GA foi medida usando o ensaio de p-nitrofenil- alfa-D-glicopiranosídeo (pNPG) com pNPG como um substrato (Sigma N- 1377). Neste ensaio, a capacidade da glicoamilase de catalisar a hidrólise de p-nitrofenil-alfa-D-glicopiranosídeo (pNPG) à glicose e p-nitrofenol é medida. Em um pH alcalino, o nitrofenol forma uma cor amarela que é proporcional à atividade de glicoamilase e é monitorada em 405 nm e comparada com um padrão enzimático medido como uma GAU (Elder, M. T. and Montgomery R. S., Glucoamylase activity in industrial enzyme preparations using colonmetric enzymatic method, Journal of AOAC International, vol. 78(2), 1995). Uma GAU é definida como a quantidade de enzima que produzirá 1 g de açúcar reduzido calculado como glicose por hora a partir de um substrato de amido solúvel (4% ds) em pH 4,2 e 60°C.
O perfil proteico foi determinado por eletroforese em PAGE em NuPAGE® Novex Bis-Tris Gel 10% com Tampão de Corrida MES SDS (Invitrogen, Carlsbad, CA, USA).
Cepas hospedeiras de Trichoderma reesei expressando as variantes únicas a) V314H, b) S211R, c) Q172F e d) Q208N foram cultivadas em fermentadores batelada-alimentada de 14L a 34°C, pH 3,5 em meios nutritivos incluindo glicose (Cerelose DE99), KH2PO4, MgSθ4-7H2θ, (NH4)2SO4, CaCl2-2H2θ, elementos-traço e antiespumante Mazu (DF6000K). Sob depleção de glicose, a temperatura e o pH de crescimento foram deslocados para 28°C e 4,0, respectivamente. 0 material celular foi removido por filtração e os sobrenadantes de cultura foram coletados e concentrados para conter mais do que 90% de glicoamilase como proteína total.
Várias propriedades cinéticas foram determinadas para produ-ção de glicose no amido solúvel de batata em pH 4,3 a 32°C e a 60°C e comparadas à TrGA de tipo selvagem. Cada uma das quatro variantes de-monstrou valor de Vmax aumentado (μM de glicose/s) quando comparada ao tipo selvagem (TrGA) indicando elevadas taxas catalíticas (kcat (s‘1)). A figura 6 ilustra o Vmax de duas replicatas para cada temperatura testada.
Validação do rastreamento foi realizada nas variantes que foram identificadas como tendo um índice de desempenho superior quando comparadas à TrGA parental (ver Tabela 7/8) usando um novo teste de aplicação de Etanol em pequena escala. Vinte e quatro variantes derivadas das bibliotecas do sítio de avaliação e combinatório (Tabela 9) foram selecionadas e transformadas diretamente em T. reesei para expressão e teste em larga escala. As variantes foram testadas para desdobramento térmico usando Calorimetria Diferencial de Varredura (análise DSC descrita neste pedido abaixo) e desempenho usando um novo ensaio de aplicação de etanol em pequena escala secundário. O método consistiu em duas etapas: 1) injeção de variantes em uma coluna de troca aniônica para determinar exatamente a concentração proteica; e 2) titulação de variantes com três concentrações de TrGA diferentes (0,3-0,15-0,075g/28g ds) a fim de calcular seu desempenho de acordo com a produção de etanol em relação à molécula de tipo selvagem.Tabela 9. Lista de variantes combinatórias
Uma preparação enzimática bruta foi purificada usando um sis-tema AKTA explorer 100 FPLC (Amersham Biosciences, Piscataway, NJ), β- ciclodextrina (Sigma-Aldrich, Zwijndrecht, Netherlands; 85.608-8) foi ligada a contas de Sefarose ativadas por epóxi (GE Healthcare, Diegem, Belgium; 17-0480-01). A coluna foi usada para capturar glicoamilases da preparação enzimática. A enzima foi eluída das contas usando tampão de Tris 25 mM pH 7,5 ou tampão de acetato de sódio 50 mM pH 4,3 contendo a- ciclodextrina 10 mM (Sigma, 28705). Amostras purificadas foram analisadas por SDS-PAGE. Para determinar exatamente a concentração proteica das variantes, um método de determinação de proteína com base em FPLC foi desenvolvido. A concentração proteica da molécula de TrGA marcadora purificada foi primeiro determinada usando um protocolo padrão de Bradford (Bio-Rad cat#500-0205). Posteriormente, as amostras purificadas foram injetadas em uma coluna ResourceQ_1 ml (GE Healthcare) e a enzima foi eluída com tampão de Tris pH 25 mM contendo NaCI 500 mM. A área de pico foi determinada e a concentração proteica foi calculada em relação à área de pico da TrGA padrão com concentração conhecida.
A tabela 10 resume a produção de etanol e açúcares (DP1, DP2, DP> 3) por variantes combinatórias diferentes. Uma amostra de milho triturada liquefeita obtida e diluída a DS 26% usando vinhaça fina. O pH da pas- ta fluida foi ajustado para pH 4,3 usando ácido sulfúrico 4N. Uma alíquota de 100 g de mosto foi colocada em um banho d'água a 32°C e permitido o equilíbrio. Após adição de 100 μl de ureia 400 ppm, 1 ml de amostra de enzima TrGA variante purificada (150 μg/ml) ou TrGA purificada (300, 150, 75 μg/ml) foi adicionado a cada amostra de mosto de milho. Finalmente, 333 μl de 15 g hidratados por 30 minutos em 45 ml solução de água Dl de levedura Red Star Red (Lesaffre. yeast Corp. Milwaukee, Wl) foram adicionados a cada amostra. As amostras foram tomadas em 5, 21, 28, 48 e 52 horas e analisadas por HPLC usando uma coluna 9 Aminex HPX-87H (Bio-Rad).
Um tubo de centrífuga eppendorf de 2 ml foi enchido com fer- mentador de cerveja e resfriado em gelo por 10 minutos. A amostra foi cen-trifugada por 3 minutos em 14.000 x g e 500 μl do sobrenadante foram transferidos para um tubo teste contendo 50 μl de solução de extinção (ácido sulfúrico 1,1 N) e permitidas permanecer por 5 minutos. 5,0 ml de água foram adicionados ao tubo teste e então filtrados em uma placa de filtro de 0,22 μm (multitela, Millipore, Amsterdam, Netherlands) e corridos em HPLC. Temperatura de Coluna: 60°C; fase móvel: ácido sulfúrico 0,01 N; taxa de fluxo 0,6 ml/min; detector: RI; volume de injeção: 20 μl. A coluna separa moléculas com base na carga e peso molecular; DP1 (monossacarídeos); DP2 (dissa- carídeos); DP3 (trissacarídeos); DP> 3 (açúcares oligossacarídicos tendo um grau de polimerização maior do que 3); ácido succínico; ácido lático; glicerol; metanol; etanol.
A temperatura de fusão das amostras enzimáticas purificadas (0,2 a 0,4 mg/ml) foi determinada usando Calorimetria Diferencial de Varre-dura (DSC).Tabela 10. Produção de etanol e sacarídeos
A tabela 11 representa os rendimentos finais de etanol e o de-sempenho das variantes na dosagem de 0,15 mg. O desempenho foi calculado pela interpolação dos valores de 0,3 mg e de 0,15 mg da TrGA pelos valores das variantes. Tabela 11: Rendimentos de Etanol
Todas as variantes combinatórias exceto ET7-2 tiveram melhor desempenho do que a TrGA de tipo selvagem. LR8 teve o melhor desempenho com desempenho melhorado 1,56.
A tabela 12 fornece um resumo de todas as variantes de sítio único e combinatórias testadas usando ensaio de aplicação de etanol em pequena escala. As variantes que estão sombreadas na Tabela 12 tiveram um melhor desempenho do que TrGA e também tiveram uma temperatura de desdobramento térmico mais alta (dTm). Tabela 12. Desempenho e desdobramento térmico de variantes em relação àTrGA

Os resultados mostraram que Cromatografia (FPLC) foi um instrumento útil para determinar exatamente a concentração proteica. Os resultados também mostraram que a titulação de variantes com três concentrações de TrGA foi um método valioso para determinar o desempenho de variantes em pequena escala. Sete variantes tiveram melhor desempenho do que a TrGA de tipo selvagem (ver Tabela 12) e também tiveram uma temperatura de desdobramento térmico mais alta e as variantes que não tiveram desempenho tão bom quanto TrGA também tinha uma Tm menor.
A atividade específica de um conjunto das variantes combinatórias e diversas variantes de sítio único que foram usadas para construir variantes combinatórias foi analisada (Tabela 13). LR8 (PI 1,56 determinado com o ensaio de aplicação em pequena escala) foi estudada ainda com respeito à especificidade de substrato. Isto foi feito pela configuração de um ensaio de MTP para determinar as taxas de produção de glicose de variantes de GA e determinar a especificidade por substrato da variante LR8. O ensaio de MTP foi encontrado discriminar entre as variantes e todas as variantes exceto ET7-1 mostraram taxas mais altas do que a glicoamilase de Trichoderma reesei de tipo selvagem (wt). Além disso, diversas variantes (LR8/ET8/Q172F) tiveram desempenho 20 a 30% melhor do que TrGA. LR8 teve melhor desempenho em amido de milho solúvel e duas amostras de milho trituradas liquefeitas diferentes comparadas ao tipo selvagem.
Os substratos usados nos seguintes experimentos foram solução estoque de amido de milho solúvel preparada como se segue: 8g de amido de milho solúvel (Sigma # S4180) foram dissolvidos em 100 ml de água milliQ e aquecidos em um micro-ondas por 1 minuto. A dispersão foi fervida por 5 minutos e após resfriada, o volume foi ajustado a 100 ml. Amido de milho solúvel 4% foi preparado pela diluição da solução estoque 1:1 com tampão de NaAc 100 mM pH 4. Em um experimento, um substrato liquefeito de milho (NE) foi preparado usando um analisador de umidade para medir o ds %, então o substrato foi diluído 7,5x com NaAc 50 mM para obter finalmente ds 4%. O substrato foi centrifugado por 5' em 2000 x g e o sobrenadante foi filtrado com um filtro de 0,22 μm. Em outro experimento, um substrato liquefeito de milho (BSE) foi preparado do mesmo modo, exceto que o substrato foi diluído 10x antes da centrifugação.
A enzima foi diluída usando a solução Estoque de 150 μg enzi- ma/ml (3 μg/180 μl de mistura de reação). As soluções foram ainda diluídas com NaAc 50 mM pH 4,0 como se segue: 300 ng (10 x), 200 ng, 150 ng, 100 ng, 75 ng, 50 ng, 25 ng, 10 ng/180 μl de mistura de reação.
O ensaio foi realizado como se segue: 40 μl de NaAc 50 mM pH 4,0,120 μl de amido de milho solúvel 4%, e 20 μl de enzima foram adicionados a cada poço. As amostras foram incubadas por 2 h a 32°C 900 rpm e terminadas em gelo após adição de 90 μl de tampão de glicina-NaOH 800 mM pH 10 por 5 min. A placa foi centrifugada para 5 minutos em 2000 rpm a 15°C. A uma nova placa, 85 μl de água milliQ e 100 μl de coquetel de Hexoquinase (conjunto II test glucose (HK), Instrumental Laboratory # 182507-40) e 20 μl de sobrenadante foram adicionados. Para uma linha de calibração de glicose (0 a 1 mg/ml), 20 μl de glicose estoque foram adicionados em vez disso. As placas foram incubadas por 10 minutos à temperatura ambiente no escuro seguido pela medida de absorção em 340 nm usando Spectramax.
Os resultados do ensaio para determinar as taxas de produção de glicose por variantes de GA são mostrados na Figura 7 e 8. Nestas figuras, o desempenho relativo à TrGA foi calculado por quantidade de enzima adicionada. As conclusões foram tiradas da região linear do gráfico em 150 ng de enzima. Os resultados nas Figuras 7, 8 e na Tabela 13 mostraram que LR8, ET8, ET7-2, S211R, Q172F e P94N tiveram desempenho melhor do que tipo selvagem ao longo da faixa linear.
Para determinar a especificidade por substrato de LR8, o de-sempenho de LR8 e TrGA de tipo selvagem foi testado em substratos (amido de milho solúvel, e os dois substratos de mosto de milho produzidos no Exemplo 10) usados em rastreamento e aplicação. Quando analisados por HPLC, os substratos mostraram uma diferença no grau de padrão de polime- rização (DP) (ver Figuras 9 a 11). Em NE e BSE dP1-> = DP4 está presente, enquanto o amido de milho solúvel consiste de pelo menos quatro ou mais moléculas de glicose. Em todos os substratos, LR8 teve desempenho melhor do que o tipo selvagem (ver Figuras 9,10 e 11).
Variantes de TrGA adicionais, particularmente variantes com substituições dentro do SBD, foram criadas e rastreadas diretamente em Trichoderma reesei. Similar ao Exemplo 1, outras dez bibliotecas de muta-gênese saturada em sítio de TrGA (SSM) foram construídas usando o vetor de entrada pDONR-TrGA como um molde e iniciadores listados na Tabela 14. Os sítios incluem: N61, G73, L417, T430, A431, E503, Q511, A535, A539, e N563. Entre estes sítios, E503, Q511, A535, A539, e N563 estão localizados dentro do domínio de ligação a amido de TrGA. Posteriormente, a recombinação foi realizada com o vetor pTTT-Dest (Figura 14) usando mistura de enzima LR CLONASE® II de acordo com o protocolo fornecido pela Invitrogen. Os produtos de recombinação foram transformados em E. coli de eficiência Máxima DH5a (Invitrogen) e plaqueados em meio 2xTY [Bacto Triptona (Difco) 16 g/L, Extrato de Levedura Bacto (Difco) 10 g/L, NaCI 5 g/L] suplementado com ampicilina 100 μg/mL. Após incubação noturna a 37°C, 96 colônias únicas de cada biblioteca foram selecionadas a partir de placas de ágar 2xTY com ampicilina 100 μg/ml e cultivadas por 24 h a 37°C em uma MTP contendo 200 μl de meio 2xTY com ampicilina 100 μg/ml. As cultu- ras foram usadas para análises de sequência (analisador de sequência A- BI3100, Applied Biosystems). Cada biblioteca continha de 15 a 19 variantes de TrGA diferentes no vetor de expressão final. Estas variantes foram individualmente transformadas em T. reesei.Tabela 14 Iniciadores usados para gerar bibliotecas de SSM de TrGA adicionais
As SELs foram transformadas em T. reesei usando o método de PEG-protoplasto (ver, por exemplo, Pentillâ et al. (1987) Gene 61: 155-164). O hospedeiro T. reesei é uma cepa derivada de RL-P37 (IA52) e que tem quatro deleções gênicas (Δcb/i1, Δcb/i2, Δeg/1, Δeg/2; isto é, "quad- deletado"; ver Patente Norte-americana Nes-5.847.276, WO 92/06184, e WO 05/001036). Misturas de transformação contendo até 600 ng de DNA e 1 a 5x105 protoplastos em um volume total de 25 μl foram tratadas com 200 ml de solução de PEG 25%, diluídas com 2 volumes de solução de sorbitol 1,2 M, misturadas com cobertura de agarose seletiva MM 3% com acetamida e vertida sobre agarose seletiva 2% com acetamida em placas de microtítulo de 24 poços. As placas foram incubadas a 28°C por 5 a 8 dias. Esporos da população total de transformantes regenerados em cada poço individual foram coletados das placas usando uma solução de NaCI 0,85%, Tween 80 0,015%. Suspensões de esporos foram usadas para inocular fermentações em MTPs de 96 poços. No caso de MTPs de 24 poços, uma etapa de pla- queamento adicional em uma nova MTP de 24 poços com acetamida MM seletiva foi introduzida a fim de enriquecer o número de esporos.
Os transformantes foram fermentados em MTPs e os sobrena- dantes de cultura contendo as variantes proteicas expressas foram usados para os ensaios. Em resumo, MTPs contendo 200 μl do meio LD-GSM foram inoculados em quadruplicata com suspensões de esporos de transformantes de T. reesei expressando variantes de TrGA (mais de 104 esporos por poço). As placas foram incubadas a 28°C com agitação de 230 rpm e umidade de 80% por 6 dias. Os sobrenadantes de cultura foram coletados por filtração a vácuo. Os sobrenadantes foram usados em ensaio diferente para rastreamento de variantes com propriedades melhoradas.
As variantes que exibem um índice de desempenho de mais de 1,0 para estabilidade térmica, atividade específica, e tanto estabilidade térmica como atividade específica são mostradas nas Tabelas 15 a 17. Tabela 15. Rastreamento de estabilidade térmica de variantes de TrGA adi-cionais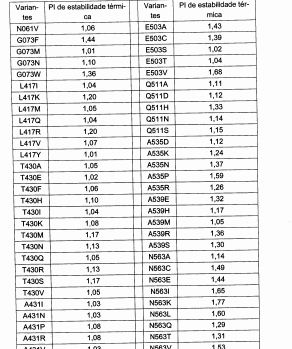 Tabela 16. Rastreamento de atividade específica de variantes de TrGA adicionais.
Tabela 16. Rastreamento de atividade específica de variantes de TrGA adicionais.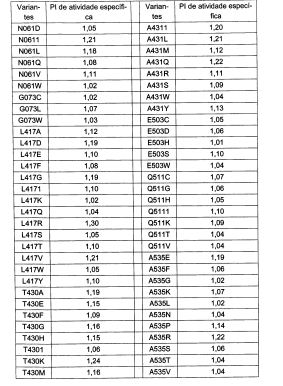
 Tabela 17. Variantes de TrGA adicionais que exibem tanto estabilidade térmica como atividade específica aumentadas
Tabela 17. Variantes de TrGA adicionais que exibem tanto estabilidade térmica como atividade específica aumentadas
Com base nos resultados dos Exemplos 4 a 6 e 11, um conjunto selecionado a partir de variantes combinatórias, e variantes de sítio único foi caracterizado ainda por suas propriedades alteradas. O conjunto seleciona- do inclui variantes combinatórias e de sítio único com substituição(ões) em: I43, D44, N61, G73, G294, L417, T430, A431, E503, Q511, A535, A539 e/ou N563. As variantes foram purificadas em fermentação de larga escala, e o PI de estabilidade térmica e atividades específicas foi determinado. Especifi- camente, as atividades específicas foram determinadas usando diferentes substratos, incluindo DP7, amido de milho e liquefeito. Os resultados são mostrados na Tabela 18 e 19.Tabela 18. Pls de um conjunto selecionado de variantes de sítio único, cada uma das quais é de uma fermentação de 500 ml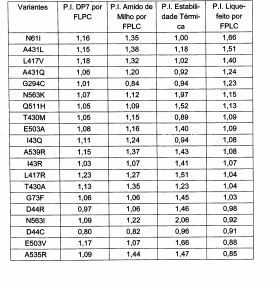 Tabela 19. Pls de um conjunto selecionado de variantes combinatórias
Tabela 19. Pls de um conjunto selecionado de variantes combinatórias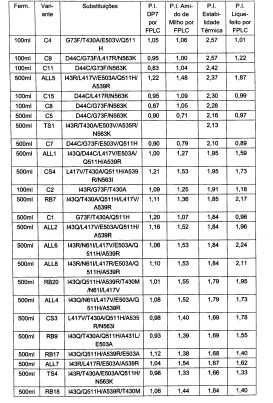


A estrutura tridimensional completa da glicoamilase de Trichoderma reesei (Hypocrea jecorina) (TrGA) foi determinada em resolução de 1,9 A. A tabela 20 mostra as coordenadas da estrutura cristalina da glicoamilase de Trichoderma. TrGA foi cristalizada em uma forma intacta contendo 599 resíduos e todas as modificações pós-traducionais que ocorreriam nor-malmente no hospedeiro natural. A estrutura cristalina foi produzida e analisada como se segue:
Para expressão e purificação proteica, o gene que codifica a GA de H. jecorina foi clonado e expresso de acordo com os protocolos descritos na Patente Norte-americana N2 7.413.887.
O material proteico de TrGA usado para todos os experimentos de cristalização foi inicialmente purificado em uma etapa por cromatografia de troca aniônica como se segue: os sobrenadantes de cultura concentrados da TrGA expressa, consistindo de proteína total 180 mg/ml, foram preparados pela diluição de amostra 1:10 em um tampão Tris-HCI 25 mM, pH 8,0. Uma coluna HiPrep 16/10 Q Sepharose FF (GE Helthcare) foi empregada para a purificação por troca aniônica. A coluna HiPrep foi equilibrada com 4 volumes de coluna (CV) de tampão de partida (Tris-HCI 25 mM, pH 8,0) seguido pela aplicação de 10 ml da amostra proteica diluída. Um gradiente linear de 8 CV de NaCI 0 a 140 mM no tampão de corrida (Tris-HCI 25 mM, pH 8,0) foi aplicado para eluir a proteína ligada. TrGA ligada eluiu da coluna HiPrep Q Sepharose em uma concentração de sal de aproximadamente NaCI 80 mM. As frações contendo proteína TrGA pura foram agrupadas e concentradas a 50 mg/ml usando um tubo de concentração de centrífuga Vivaspin de 25 ml (Viva Science) com um corte de peso molecular (MWCO) de 10 kD. Material de TrGA purificado e concentrado foi trocado em tampão usando uma coluna de dessalinização DG-10 (Bio-Rad) equilibrada com tampão de acetato de sódio 50 mM, pH 4,3. As concentrações proteicas foram determinadas pela medida da absorvância em 280 nm. O estoque proteico de TrGA inicialmente purificado e concentrado foi após isso armazenado a -20°C.
Duas etapas de purificação adicionais, em purificação de troca aniônica adicional, e uma purificação por exclusão de tamanho, foram intro-duzidas para aumentar a cristabilidade do material proteico de TrGA. Estas duas etapas de purificação adicionais foram realizadas como se segue. Na primeira etapa de purificação de troca aniônica, uma coluna MonoQ de 10 ml (GE Helthcare) foi empregada. Uma Amostra de 1 ml do material de TrGA inicialmente purificada e congelada (50 mg de proteína) foi descongelada e o tampão foi modificado para Tris-HCI 20 mM, pH 8,0, pela diluição repetida da amostra a 6 ml no novo tampão, seguido por uma concentração da amostra novamente a 0,5 ml usando um tubo de concentração de 6 ml de MWCO 5 kD. A amostra de TrGA foi diluída após a última etapa de concentração em água destilada até que uma condutividade da amostra proteica fosse alcançada que correspondeu à condutividade do tampão de partida de purificação aniônica, isto é, Tris-HCI 25 mM, pH 8,0. A coluna MonoQ foi primeiro equilibrada com 4 volumes coluna (CV) de tampão de partida, seguido pela aplicação da amostra proteica diluída à coluna A proteína ligada foi eluída da coluna MonoQ por dois gradientes diferentes. No primeiro, um gradiente 4 CV de pH linear foi aplicado onde o pH do tampão inicial foi reduzido de 8,0 para 6,0. No segundo gradiente, um longo gradiente 8 CV de sal foi aplicado no qual a concentração salina foi aumentada de 0 a 350 mM de NaCI no tampão de corrida (Tris-HCI 25 mM, pH 6,0). Foi encontrado que TrGA ligada eluiu da coluna durante o segundo gradiente de sal em uma concentração de NaCI aproximada de 150 mM. As frações contendo TrGA foram a- grupadas e concentradas a 2 ml usando um tubo de concentração Vivaspin de 6 ml de MWCO 5 kD. A amostra de TrGA concentrada foi após isso aplicada a uma coluna de exclusão por tamanho Superdex 200 16/60 (GE Helthcare) equilibrada com 4 CV de Tris-CI 20 mM, pH 8,0, e NaCI 50 mM, que também foi usado como tampão de corrida. As frações do pico de eluição principal após a purificação por exclusão de tamanho foram agrupadas e concentradas a uma concentração proteica aproximada de 7,5 mg/ml usando um tubo de concentração Vivaspin 6 ml de MWCO 5 kD.
Para cristalização proteica, a amostra proteica que foi usada para encontrar as condições de cristalização da TrGA iniciais foi uma amostra do material TrGA que foi purificado uma vez pela purificação por troca aniô- nica e após isso armazenado a -20°C. A amostra proteica de TrGA foi descongelada e diluída com tampão de acetato de sódio 50 mM, pH 4,3, a aproximadamente 12 mg/ml, antes dos experimentos de cristalização iniciais. O conjunto de dados de raio x ortorrômbico, foi usado para resolver a estrutura da TrGA pela substituição molecular (MR.), e o conjunto de dados ortorrôm- bicos de alta resolução, usado para o grupo de modelo espacial final ortorrômbico de estrutura da TrGA. Foi encontrado que cristais ortorrômbicos da TrGA cresciam na solução consistindo de PEG 3350 25%, acetato de amónio 0,20M, Bis-Tris 0,10M pH 5,5 (solução de reservatório), usando o método de difusão de vapor com gotas em suspensão (McPherson 1982), a 20°C. Gotas de cristalização foram preparadas pela mistura de quantidades iguais da solução proteica (12 mg/ml) e solução de reservatório a um volume final de 10 μl. Foi encontrado que os cristais de TrGA pertenciam ao grupo espacial ortorrômbico P212121 com dimensões celulares aproximadas: a = 52,2 A, b = 99,2 A, c = 121,2 A, e tem um Vm calculado de 2,3 (Matthews 1968) com moléculas na unidade assimétrica.
Para coleta de dados por raio x, os dois conjuntos de dados or-torrômbicos de TrGA foram coletados de cristais únicos montados em tubos capilares selados, à temperatura ambiente. O conjunto de dados de raio x de TrGA ortorrômbico de baixa resolução inicial, usado para resolver a estrutura por métodos de substituição molecular (MR.), foi coletada em uma fonte de raio x local, um detector de placa de imagem Raxis IV++ MSC/Rigaku (Molecular Structures Corp., The Woodlands, Texas) com espelhos de focagem usando radiação de Cu Kα de um gerador de anodo rotatório Rigaku RU200. Este conjunto de dados foi processado, escalonado, e calculado a média usando programa d*trek fornecido por MSC/Rigaku. O conjunto de dados C centrado monoclínico foi coletado de um cristal de TrGA congelado único em 100K, equilibrado em um agente crioprotetor compreendido de PEG 3350 25%, Glicerol 15%, CaCfe 50 mM e Bis-Tris 0,1 M pH 5,5 como crioprotetor, montado em alças de fibra de seda artificial, e mergulhado congelado em nitrogênio líquido antes do transporte ao síncroton. O conjunto de dados or- torrômbicos de alta resolução (1,9 A) e conjunto de dados monoclínico cên- trico C (1,8 À) foram ambos coletados em uma fonte de síncroton, linha de raio 911:5 no MAX LAB em Lund, Sweden. Tanto os conjuntos de dados que foram coletados em uma fonte de síncroton foram processados com MOS- FLM, como escalonados com o programa SCALA incluído no pacote de programas CCP4 (Collaborative Computational Project Number 4 1994). Todo o processamento de dados subsequente foi realizado usando o pacote de pro-grama CCP4 (Collaborative Computational Project Number 4 1994), a menos que de outra maneira afirmado. O conjunto de 5% das reflexões de cada conjunto de dados foi deixado de lado e usados para monitorar o R-livre (Brunger, A (1992) Nature, 355:472-475).
A estrutura TrGA foi inicialmente resolvida por MR. com o pro-grama de substituição automática MOLREP (Collaborative Computational Project Number 4 1994), incluído no pacote de programa CCP4, usando o conjunto de dados de baixa resolução inicial ortorrômbica, e usando as co-ordenadas de GA variante de Aspergillus, awamori (AaGA) X100 (entrada pdb 1GLM (Aleshin et al. (1994) J. Mol. Boil. 238: 575-591) como modelo de pesquisa. O modelo de pesquisa de GA de A. awamori foi editado para remover todas as porções de glicosilação ligadas à molécula proteica como N- e O-glicosilações, e todas as moléculas solventes antes de realizar os expe-rimentos de MR. Todas as reflexões entre resolução de 36,8 e 2,8 A, do con-junto de dados de baixa resolução inicial de TrGA, foi usadas para a solução de MR. O programa de MR. encontrou uma solução de função de rotação única, com máximos de 11,1 α acima do fundo, o próximo máximo mais alto foi 3,8a acima do fundo. A solução de função de tradução forneceu um fator R de 48,7% e teve um fator de contraste de 17,4. A solução de MR. foi refinada para 10 ciclos contidos a menos na parte de refinamento de quadrados usando o programa Refmac 5.0 (Murshudov et al (1997) Acta Crystallogr. D53: 240-255). Isto reduziu o fator R cristalográfico a 31,1%, enquanto o valor de R-livre caiu de 42,2% a 41,1%.
O modelo de solução de MR refinado foi usado para calcular um mapa de densidade inicial do conjunto de dados de baixa resolução ortor- rômbica de TrGA. A densidade eletrônica de uma ponte dissulfeto entre re-síduos 19 e 26 de TrGA, uma ponte dissulfeto não presente no modelo de estrutura da variante de A. awamori X100, pode ser prontamente identificada neste mapa de densidade eletrônica. Isto foi tomado como uma indicação que o mapa de densidade eletrônica foi de qualidade suficiente para ser u- sado para construir um modelo de estrutura de TrGA de sua sequência de aminoácidos. O modelo de estrutura de TrGA inicial, baseado no conjunto de dados de baixa resolução, foi refinado com ciclos alternados do modelo de construção usando Coot (Emsley and Cowtan, (2004) Acta Crystallogr. D. boil. Crystallogr. 60: 2126-2132), e refinamento de probabilidade máxima usando Refmac 5.0.
A resolução do modelo de estrutura de TrGA inicial foi estendida para a resolução do conjunto de dados ortorrômbico de alta resolução (1,9 A) pelo refinamento do modelo de estrutura de TrGA inicial contra o conjunto de dados de alta resolução de 10 ciclos de refinamento contido usando o programa Refmac 5.0. A maioria das moléculas de água nos modelos de estrutura foi localizada automaticamente pelo uso dos protocolos de seleção de água nos programas de refinamento, e então manualmente selecionadas ou descartadas por inspeção visual. Todas as comparações estruturais foram feitas com Coot (Emsley and Cowtan (2004) supra) ou O (Jones et al. (1991) Acta Crystallogr. A47: 110-119), e figuras foram preparadas com Py- MOL (Delano W.L. (2002) The PyMOL Molecular Graphics System. Paio Alto, CA, USA; Delano Scientific).
A partir destes resultados, pode ser visto que o segmento de núcleo catalítico de TrGA seguiu a mesma topologia (α/α)6 cilíndrica descrita por Aleshin et al. 1992 para a AaGA, consistindo de um cilindro duplo de alfa hélices com o C-terminal da hélice exterior levando ao N-terminal de uma hélice interna. Foi possível identificar diferenças-chave na densidade eletrô-nica, tais como a ponte dissulfeto entre os resíduos 19 e 26 e uma inserção (resíduos 257 a 260) em relação a AaGA. O segmento compreendendo 80 a 100 também sofreu extensa reconstrução modelar. Um sítio de glicosilação principal foi identificado em Asn 171, que tinha até quatro porções glicosídi- cas ligadas. Um sítio de glicosilação similar foi identificado em AaGA. Adi-cionalmente, o núcleo catalítico contendo três peptídeos cis entre resíduos 22 a 23, 44 a 45 e 122 a 123 foi conservado entre TrGA e AaGA. Em geral, houve uma variação rms de 0,535 Â entre 409 ddos 453 átomos de Ca em comparação às coordenadas dos núcleos catalíticos de TrGA e AaGA.
A estrutura cristalina da TrGA identificada no Exemplo 13, foi superposta na estrutura cristalina anteriormente identificada da GA de As- pergillus awamorí (AaGA). A estrutura cristalina de AaGA foi obtida a partir do banco de dados de proteína (PDB) e a forma de AaGA que foi cristalizada foi a forma contendo somente um domínio catalítico. A estrutura da glicoamilase de Trichoderma reesei com as três regiões intactas foi determinada na resolução de 1,8 Angstrôns neste pedido (ver Tabela 15 e Exemplo 12). U- sando as coordenadas (ver Tabela 20) a estrutura foi alinhada com as coordenadas do domínio catalítico da cepa de Aspergillus awamorii X100 que foi determinada anteriormente (Aleshin, A.E., Hoffman, C., Firsov, L.M., and Honzatko, R.B. 1994 Refined crystal structures of glucoamylase from Aspergillus awamori var. X100. J Mol Biol 238: 575-591 e no PDB). Como visto nas Figuras 12 e 13, a estrutura do domínio catalítico se sobrepôs muito estreitamente e permitiu a identificação de resíduos equivalentes com base nesta sobreposição estrutural.
Com base nesta análise, os sítios foram identificados podendo ser mutados em TrGA e resultar em estabilidade aumentada e/ou atividade específica. Estes sítios incluem 108, 124, 175 e 316 no sítio ativo. Também foram identificadas específicas variantes aos pares Y47W/Y315F e Y47F/Y315W. Outros sítios identificados foram I43, D44, P45, D46, R122, R125, V181, E242, Y310, D313, V314, N317, R408, e N409. Por causa da alta homologia estrutural é esperado que as variantes benéficas encontradas em sítios em TrGA teriam consequências similares em Aspergillus awamori e outras glicoamilases homólogas.
Várias modificações e variações dos métodos descritos e o sis- 5 tema da descrição serão evidentes para os versados na técnica sem se a- fastar do escopo e do espírito da descrição. Embora a descrição tenha sido descrita com relação às modalidades representativas específicas, deve ser entendido que as matérias objeto como reivindicado não devem ser indevi-damente limitadas a tais modalidades específicas. De fato, várias modifica- ções dos modos descritos para realizar a descrição que são óbvios para os versados na técnica são destinados a estar dentro do escopo das seguintes reivindicações.


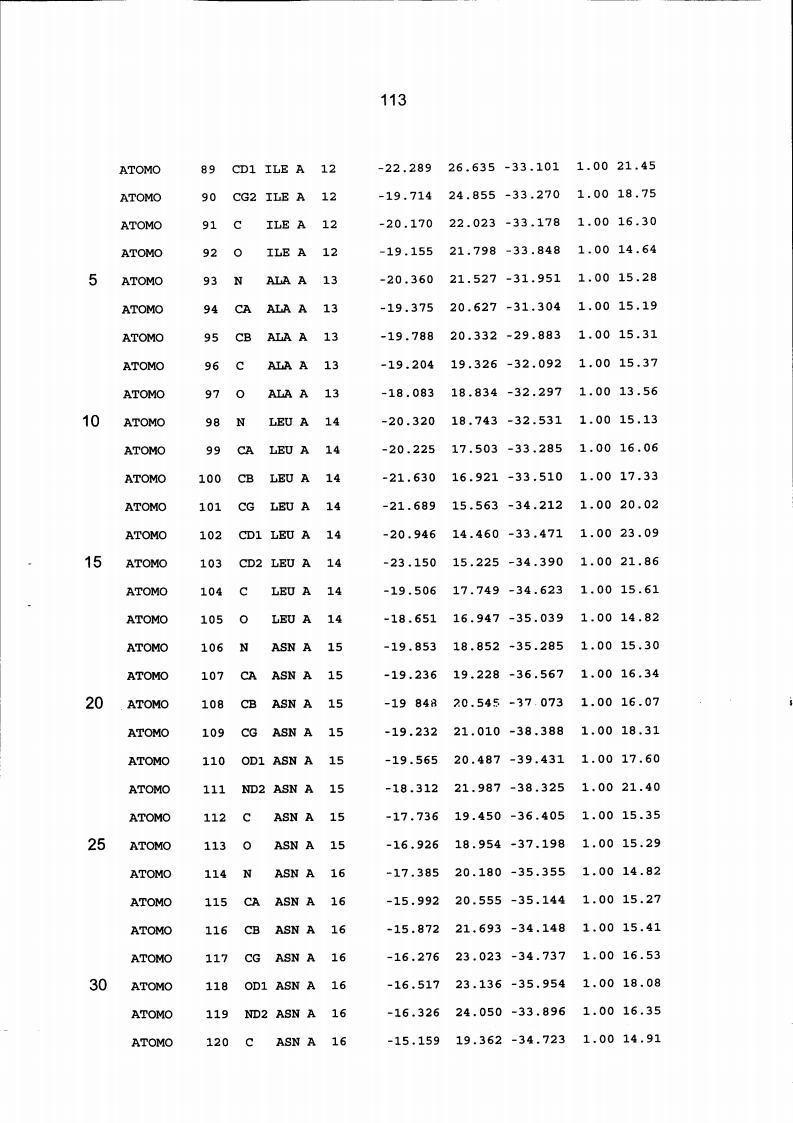

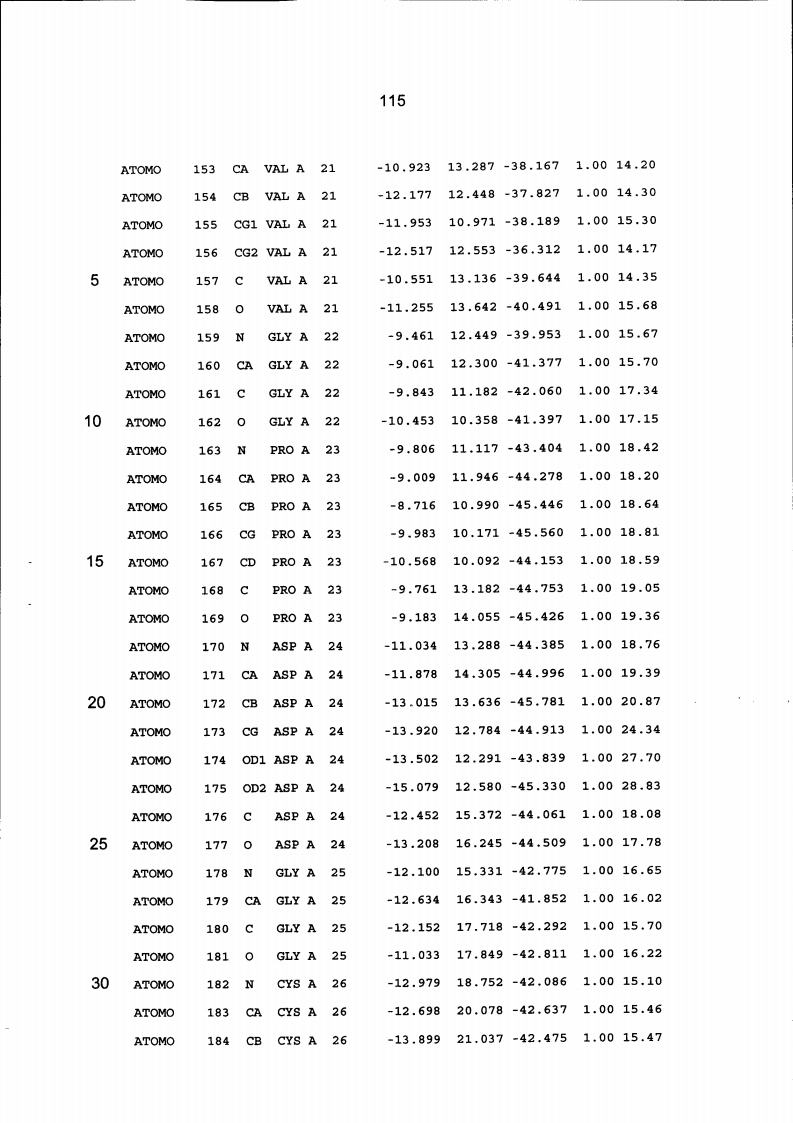























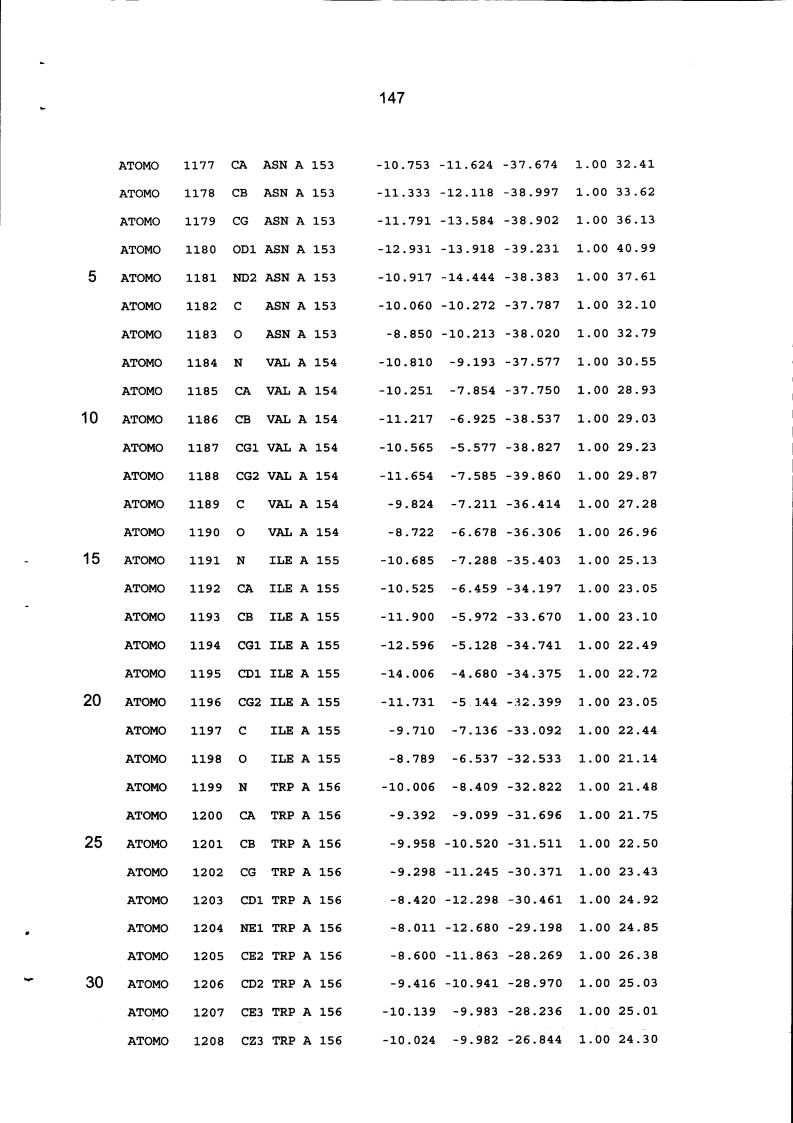




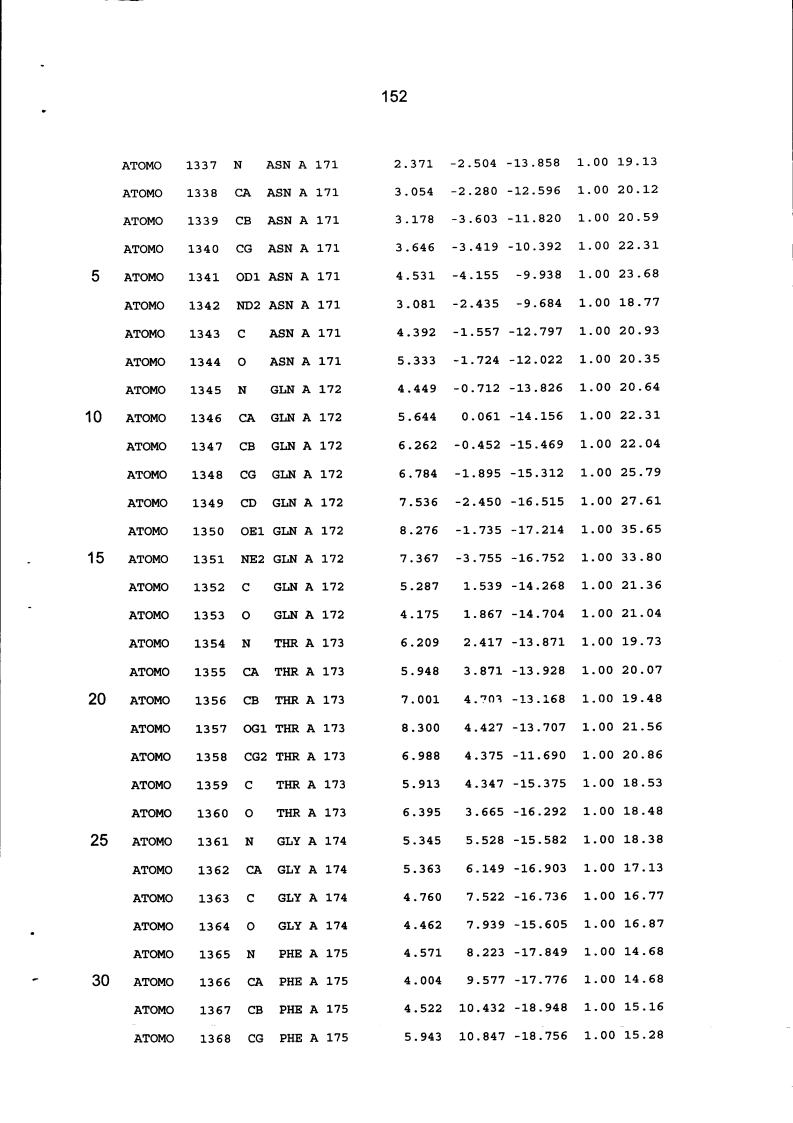
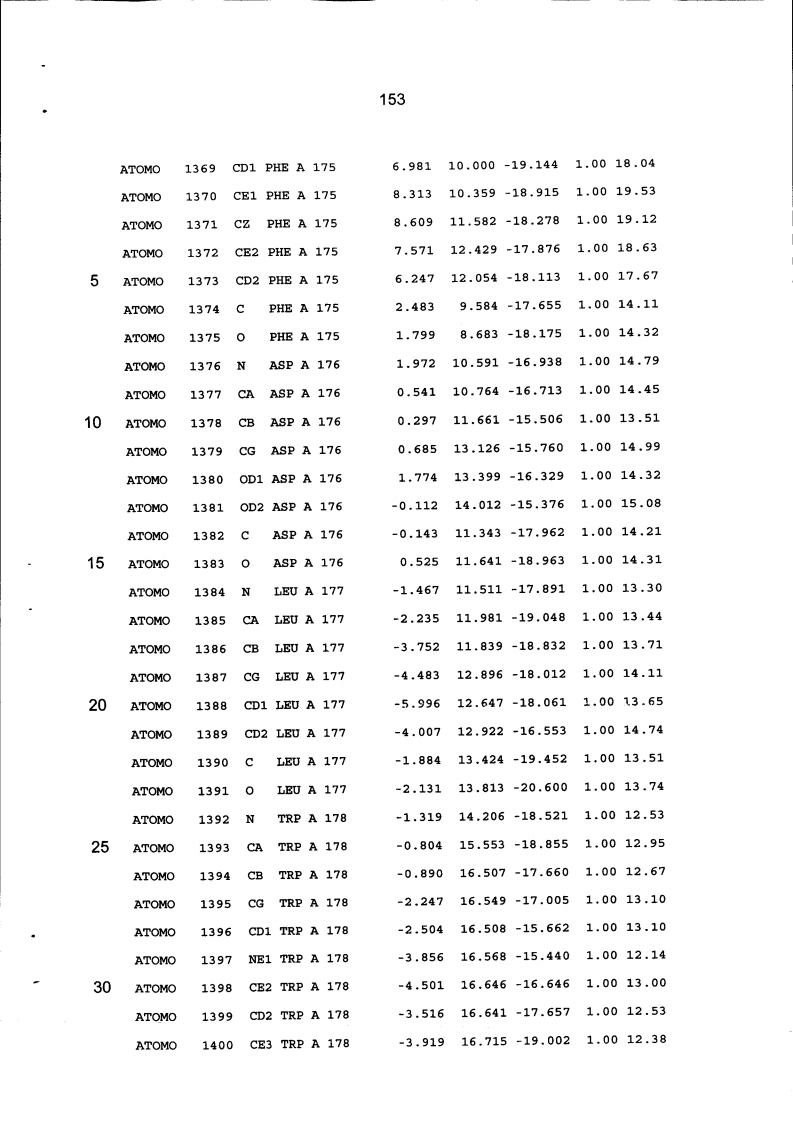
























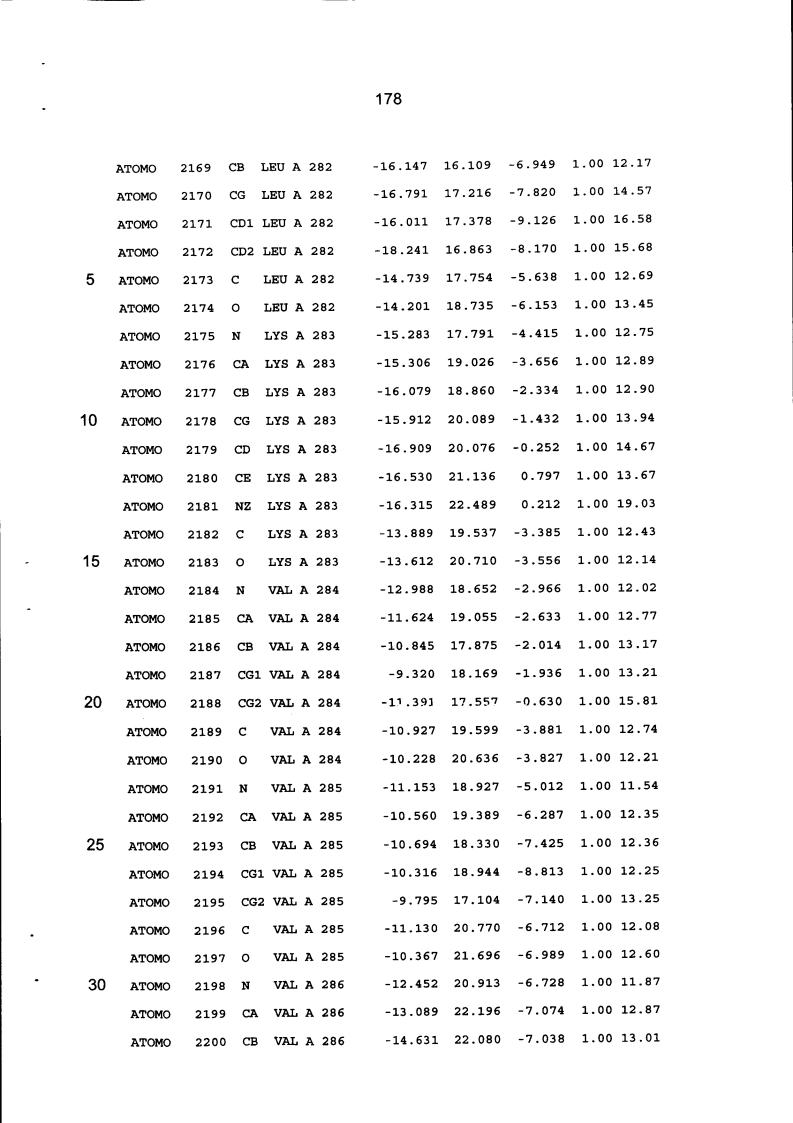













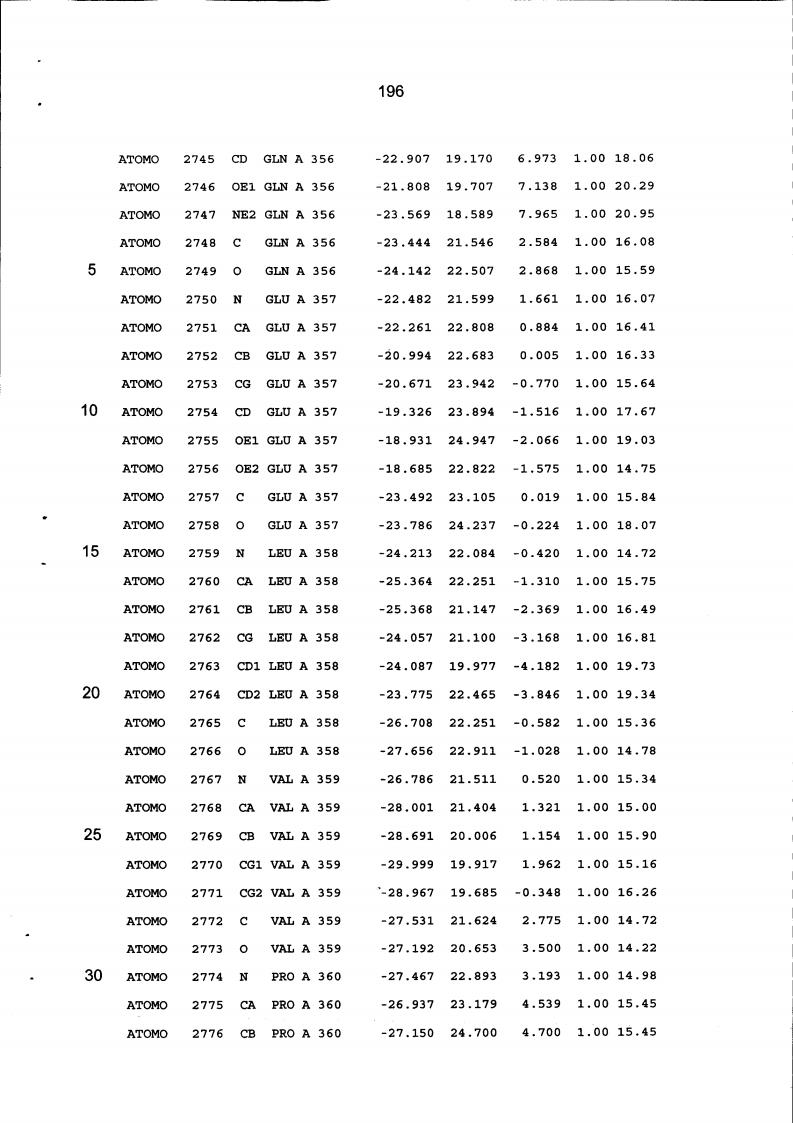


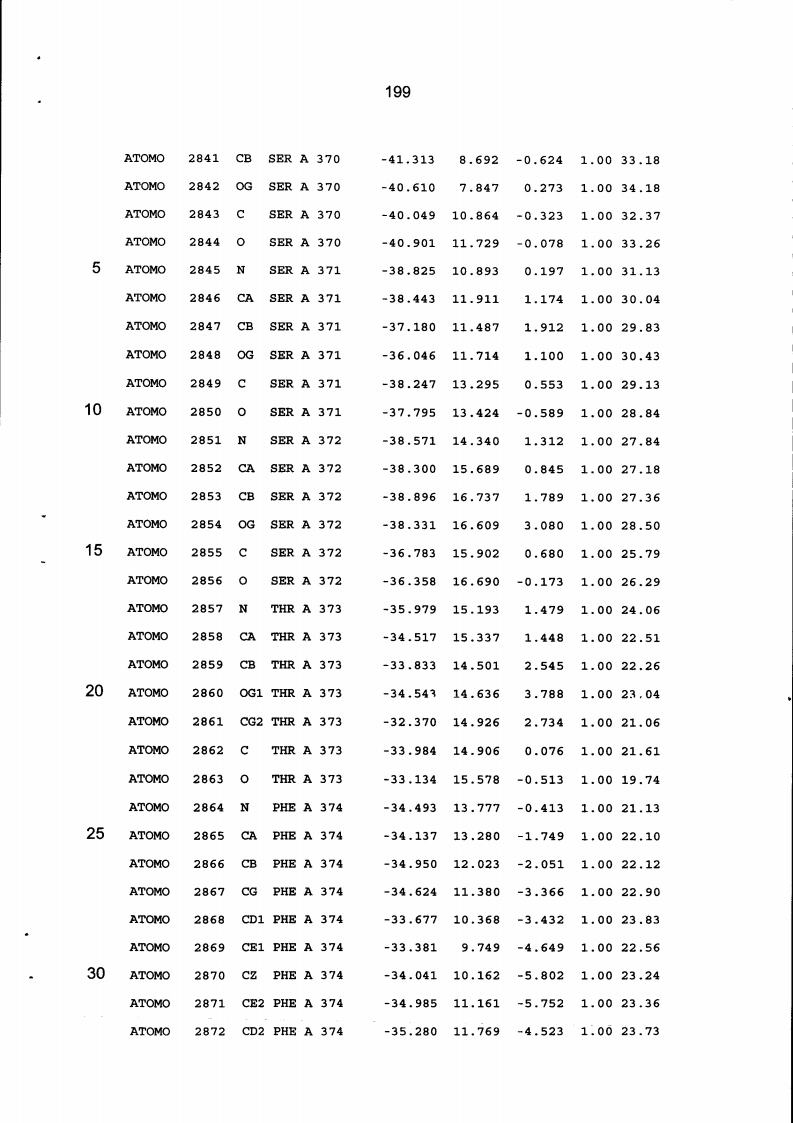







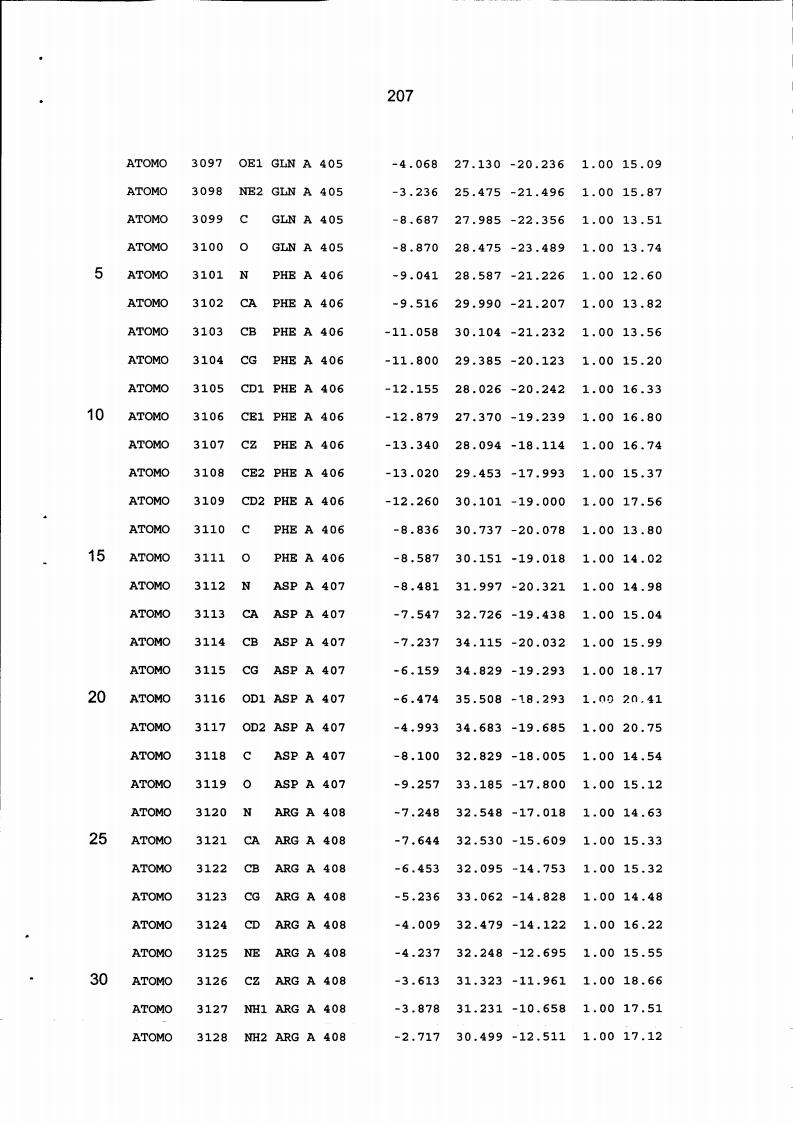














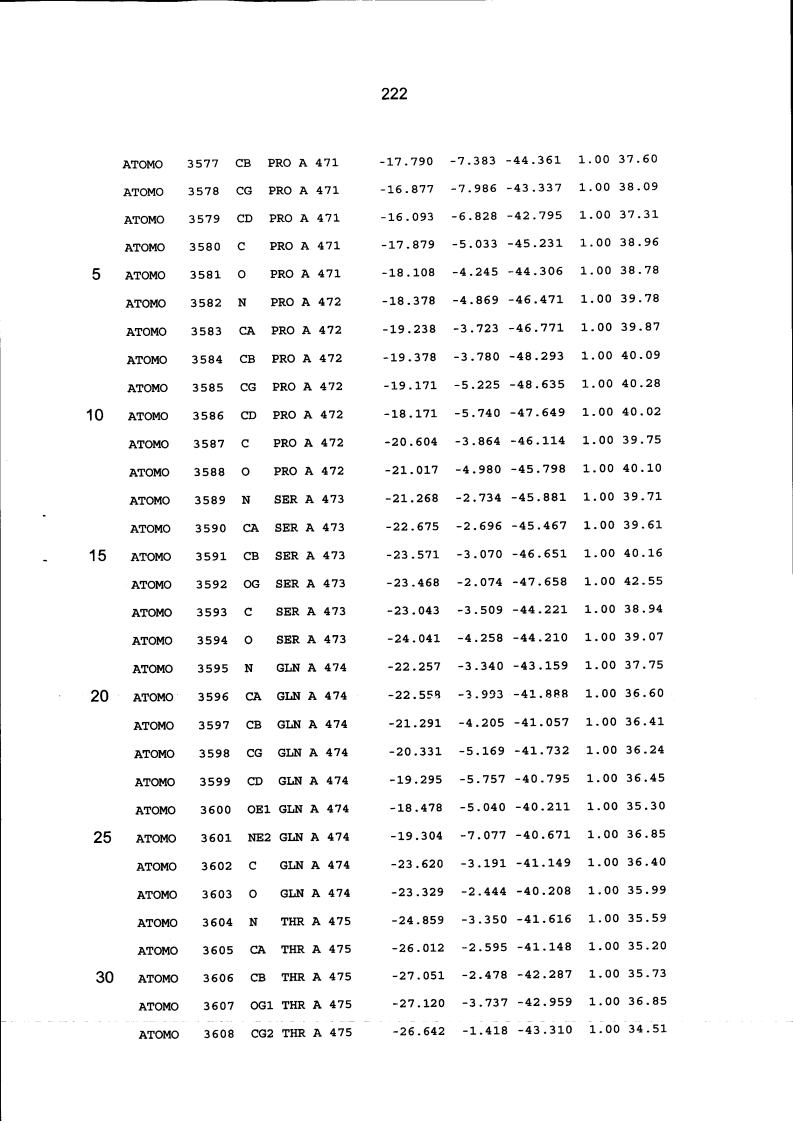









































Claims (14)
1. Variante de glicoamilase, caracterizada pelo fato de que compreendendo substituições de aminoácidos correspondentes à posição 417, 430, 539 e 563 de SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental selecionada do grupo consistindo em uma glicoamilase de Trichoderma reesei compreendendo SEQ ID NO: 1, 2, ou 3, uma glicoamilase de Aspergillus awamori compreendendo SEQ ID NO: 5 e SEQ ID NO: 168, uma glicoamilase de Aspergillus niger compreendendo SEQ ID NO: 6 e SEQ ID NO: 167, uma glicoamilase de Humicola grisea compreendendo SEQ ID NO: 8 e SEQ IDD NO: 163 e uma glicoamilase de Hypocrea vinosa compreendendo SEQ ID NO:9.
2. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que as substituições de aminoácido são: L417V, T430A, Q511H, A539R, e N563I da SEQ ID NO: 2, ou uma posição equivalente em uma glicoamilase parental.
3. Variante glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que a glicoamilase parental possui um domínio catalítico de SEQ ID NO: 1, 2, 3, 5, 6, 8, ou 9.
4. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que a glicoamilase parental possui um domínio de ligação a amido de SEQ ID NO: 1 ou 2.
5. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que a glicoamilase parental compreende a SEQ ID NO: 1 ou 2.
6. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que a glicoamilase parental consiste em SEQ ID NO: 1 ou 2.
7. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que a variante de glicoamilase compreende uma ou mais substituições de aminoácidos adicionais correspondentes às posições: 43, 44, 61, 73, 294, 431, 503 ou 535 de SEQ ID NO: 2, ou uma posição equivalente na glicoamilase parental.
8. Variante de glicoamilase de acordo com a reivindicação 7, caracterizada pelo fato de que as substituições de aminoácidos são: I43Q/R, D44C/R, N61I, G73F, G294C, A431L/Q, E503A/V ou A535R da SEQ ID NO: 2, ou uma posição equivalente na glicoamilase parental.
9. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que apresenta termoestabilidade aumentada em comparação com a glicoamilase parental.
10. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de apresenta atividade específica aumentada em comparação com a glicoamilase parental.
11. Variante de glicoamilase de acordo com a reivindicação 4, caracterizada pelo fato de que o aumento da atividade específica da variante da glicoamilase é medida usando DP7, amido de milho ou liquefato como substrato.
12. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que a variante da glicoamilase exibe tanto termoestabilidade aumentada quanto atividade específica aumentada em comparação com a glicoamilase parental.
13. Variante de glicoamilase de acordo com a reivindicação 1, caracterizada pelo fato de que a glicoamilase parental é obtida de um Trichoderma spp. ou um Aspergillus spp.
14. Composição enzimática, caracterizada pelo fato de que compreende a variante de glicoamilase como definida na reivindicação 1.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US98942607P | 2007-11-20 | 2007-11-20 | |
| US60/989,426 | 2007-11-20 | ||
| PCT/US2008/012934 WO2009067218A2 (en) | 2007-11-20 | 2008-11-20 | Glucoamylase variants with altered properties |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| BRPI0820483A2 BRPI0820483A2 (pt) | 2014-10-14 |
| BRPI0820483B1 true BRPI0820483B1 (pt) | 2022-04-12 |
Family
ID=40352212
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| BRPI0820483-7A BRPI0820483B1 (pt) | 2007-11-20 | 2008-11-20 | Variantes de glicoamilase com propriedades alteradas e composição compreendendo as mesmas |
Country Status (12)
| Country | Link |
|---|---|
| US (3) | US8058033B2 (pt) |
| EP (3) | EP3323889B1 (pt) |
| JP (1) | JP5594899B2 (pt) |
| CN (1) | CN101868537B (pt) |
| BR (1) | BRPI0820483B1 (pt) |
| CA (1) | CA2705941C (pt) |
| DK (3) | DK3323889T3 (pt) |
| ES (2) | ES2795989T3 (pt) |
| HU (1) | HUE050328T2 (pt) |
| MX (1) | MX2010005461A (pt) |
| PL (1) | PL2222842T3 (pt) |
| WO (1) | WO2009067218A2 (pt) |
Families Citing this family (64)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN101522893B (zh) * | 2006-10-10 | 2014-07-09 | 丹尼斯科美国公司 | 具有改变性质的葡糖淀粉酶变体 |
| US8592194B2 (en) | 2007-10-09 | 2013-11-26 | Danisco Us Inc. | Glucoamylase variants with altered properties |
| CN102421911B (zh) * | 2009-05-12 | 2016-05-25 | 丹尼斯科美国公司 | 以改良的液化方法发酵的乙醇产量 |
| US8545907B2 (en) | 2009-08-07 | 2013-10-01 | Danisco Us Inc. | Alpha-amylase blend for starch processing and method of use thereof |
| AR077942A1 (es) * | 2009-08-19 | 2011-10-05 | Danisco | Variantes de glucoamilasa |
| CN102712915B (zh) * | 2009-08-19 | 2014-12-10 | 丹尼斯科美国公司 | 具有改良比活和/或热稳定性的葡糖淀粉酶的组合变体 |
| MX338068B (es) * | 2010-04-14 | 2016-04-01 | Novozymes As | Polipeptidos que tienen actividad de glucoamilasa y polinucleotidos que codifican los mismos. |
| US9617527B2 (en) | 2010-04-14 | 2017-04-11 | Novozymes A/S | Polypeptides having glucoamylase activity and polynucleotides encoding same |
| CA2807558A1 (en) | 2010-08-06 | 2012-02-09 | Danisco Us Inc. | Production of isoprene under neutral ph conditions |
| BR112013001179A2 (pt) | 2010-08-06 | 2016-05-31 | Danisco Us Inc | sacrificação e fermentação em ph neutro. |
| BR112013027170A2 (pt) | 2011-04-29 | 2017-06-20 | Danisco Us Inc | processo em ph único para liquefação e sacarificação de amido para xaropes de glicose de densidade alta |
| US20120276593A1 (en) | 2011-04-29 | 2012-11-01 | Danisco Us Inc. | Use of cellulase and glucoamylase to improve ethanol yields from fermentation |
| WO2013022799A1 (en) | 2011-08-05 | 2013-02-14 | Danisco Us Inc. | PRODUCTION OF ISOPRENOIDS UNDER NEUTRAL pH CONDITIONS |
| RU2014121491A (ru) | 2011-10-28 | 2015-12-10 | ДАНИСКО ЮЭс ИНК. | Варианты вариантной альфа-амилазы, образующей мальтогексаозу |
| WO2013086219A1 (en) | 2011-12-09 | 2013-06-13 | Danisco Us Inc. | Ribosomal promotors from b. subtilis for protein production in microorganisms |
| EP2831256A1 (en) | 2012-03-28 | 2015-02-04 | Danisco US Inc. | Low temperature method for making high glucose syrup |
| CA2870830A1 (en) | 2012-05-11 | 2013-11-14 | Danisco Us Inc. | Use of alpha-amylase from aspergillus clavatus for saccharification |
| WO2013181647A2 (en) | 2012-06-01 | 2013-12-05 | Danisco Us Inc. | Compositions and methods of producing isoprene and/or industrrial bio-products using anaerobic microorganisms |
| CN104379737B (zh) | 2012-06-08 | 2018-10-23 | 丹尼斯科美国公司 | 对淀粉聚合物具有增强的活性的变体α淀粉酶 |
| US20140024064A1 (en) | 2012-07-23 | 2014-01-23 | Butamax(Tm) Advanced Biofuels Llc | Processes and systems for the production of fermentative alcohols |
| AR092099A1 (es) * | 2012-08-14 | 2015-03-25 | Danisco Us Inc | Variantes de glucoamilasa de trichoderma reesei resistentes a la perdida de actividad relacionada con la oxidacion y su uso |
| US20150218606A1 (en) | 2012-08-16 | 2015-08-06 | Danisco Us Inc. | Method of using alpha-amylase from aspergillus clavatus and pullulanase for saccharification |
| CA2878616A1 (en) * | 2012-08-16 | 2014-02-20 | Danisco Us Inc. | Method of using alpha-amylase from aspergillus clavatus and isoamylase for saccharification |
| MX2015002099A (es) * | 2012-08-22 | 2015-05-11 | Dupont Nutrition Biosci Aps | Variantes que tienen actividad glucoamilasa. |
| WO2014039773A1 (en) * | 2012-09-07 | 2014-03-13 | Novozymes A/S | Glucoamylase variants and polynucleotides encoding same and uses thereof |
| WO2014043288A1 (en) | 2012-09-12 | 2014-03-20 | Butamax Advanced Biofuels Llc | Processes and systems for the production of fermentation products |
| CA2887574A1 (en) | 2012-10-11 | 2014-04-17 | Butamax Advanced Biofuels Llc | Processes and systems for the production of fermentation products |
| EP2922951B1 (en) | 2012-11-20 | 2017-08-23 | Danisco US Inc. | Amylase with maltogenic properties |
| US9765374B2 (en) | 2012-12-14 | 2017-09-19 | Danisco Us Inc | Method of using α-amylase from Aspergillus fumigatus and isoamylase for saccharification |
| WO2014099415A1 (en) | 2012-12-20 | 2014-06-26 | Danisco Us Inc. | Method of using alpha-amylase from aspergillus terreus and pullulanase for saccharification |
| WO2014099525A1 (en) | 2012-12-21 | 2014-06-26 | Danisco Us Inc. | Paenibacillus curdlanolyticus amylase, and methods of use, thereof |
| EP3354728B1 (en) | 2012-12-21 | 2020-04-22 | Danisco US Inc. | Alpha-amylase variants |
| WO2014135647A1 (en) * | 2013-03-07 | 2014-09-12 | Novozymes A/S | Glucoamylase variants and polynucleotides encoding same |
| DK2970931T3 (en) | 2013-03-11 | 2018-01-22 | Danisco Us Inc | COMBINATORY ALPHA AMYLASE VARIATIONS |
| WO2014159309A1 (en) | 2013-03-12 | 2014-10-02 | Butamax Advanced Biofuels Llc | Processes and systems for the production of alcohols |
| WO2014200658A1 (en) | 2013-06-13 | 2014-12-18 | Danisco Us Inc. | Alpha-amylase from promicromonospora vindobonensis |
| WO2014200656A1 (en) | 2013-06-13 | 2014-12-18 | Danisco Us Inc. | Alpha-amylase from streptomyces umbrinus |
| WO2014200657A1 (en) | 2013-06-13 | 2014-12-18 | Danisco Us Inc. | Alpha-amylase from streptomyces xiamenensis |
| WO2014204596A1 (en) | 2013-06-17 | 2014-12-24 | Danisco Us Inc. | Alpha-amylase from bacillaceae family member |
| WO2014205355A2 (en) | 2013-06-21 | 2014-12-24 | Danisco Us Inc. | Compositions and methods for clostridial transformation |
| US20160186102A1 (en) | 2013-10-03 | 2016-06-30 | Danisco Us Inc. | Alpha-amylases from exiguobacterium, and methods of use, thereof |
| EP3060659B1 (en) | 2013-10-03 | 2019-05-29 | Danisco US Inc. | Alpha-amylases from exiguobacterium, and methods of use, thereof |
| WO2015057517A1 (en) | 2013-10-17 | 2015-04-23 | Danisco Us Inc. | Use of hemicellulases to improve ethanol production |
| WO2015065978A1 (en) | 2013-10-28 | 2015-05-07 | Danisco Us Inc. | Trehalase in fermentations |
| WO2015066669A1 (en) | 2013-11-04 | 2015-05-07 | Danisco Us Inc. | Proteases in corn processing |
| WO2015066667A1 (en) | 2013-11-04 | 2015-05-07 | Danisco Us Inc. | Proteases in wheat processing |
| US20160272957A1 (en) | 2013-11-20 | 2016-09-22 | Danisco Us Inc. | Variant alpha-amylases having reduced susceptibility to protease cleavage, and methods of use, thereof |
| US10584323B2 (en) | 2013-12-04 | 2020-03-10 | Danisco Us Inc | Glucoamylase variants |
| WO2015094809A1 (en) | 2013-12-19 | 2015-06-25 | Danisco Us Inc. | Chimeric fungal alpha-amylases comprising carbohydrate binding module and the use thereof |
| WO2015094714A1 (en) | 2013-12-19 | 2015-06-25 | Danisco Us Inc. | Proteases in grain processing |
| CN103695386B (zh) * | 2013-12-25 | 2015-09-16 | 南宁邦尔克生物技术有限责任公司 | 一种耐酸性高温β-淀粉酶突变体及其应用 |
| US10612058B2 (en) | 2014-12-19 | 2020-04-07 | Danisco Us Inc | Methods for saccharifying a starch substrate |
| EP3259358B1 (en) | 2015-02-19 | 2024-06-19 | Danisco US Inc. | Enhanced protein expression |
| US20180362946A1 (en) | 2015-12-18 | 2018-12-20 | Danisco Us Inc. | Polypeptides with endoglucanase activity and uses thereof |
| WO2017152169A1 (en) | 2016-03-04 | 2017-09-08 | Danisco Us Inc. | Engineered ribosomal promoters for protein production in microorganisms |
| US9598680B1 (en) * | 2016-08-05 | 2017-03-21 | Fornia Biosolutions, Inc. | G16 glucoamylase compositions and methods |
| RU2763378C2 (ru) * | 2016-09-23 | 2021-12-28 | ДюПон НЬЮТРИШН БАЙОСАЙЕНСИЗ АпС | ПРИМЕНЕНИЕ АКТИВНЫХ ПРИ НИЗКОМ ЗНАЧЕНИИ pH АЛЬФА-1,4/1,6-ГЛИКОЗИДГИДРОЛАЗ В КАЧЕСТВЕ КОРМОВОЙ ДОБАВКИ ДЛЯ ЖВАЧНЫХ ЖИВОТНЫХ ДЛЯ УЛУЧШЕНИЯ ПЕРЕВАРИВАНИЯ КРАХМАЛА |
| EP3703661A1 (en) | 2017-11-02 | 2020-09-09 | Danisco US Inc. | Freezing point depressed solid matrix compositions for melt granulation of enzymes |
| WO2021007379A1 (en) | 2019-07-09 | 2021-01-14 | Dupont Nutrition Biosciences Aps | Fat coated particulate enzyme compositions |
| WO2021046073A1 (en) | 2019-09-05 | 2021-03-11 | Dupont Nutrition Biosciences Aps | Feed composition |
| CA3232987A1 (en) | 2021-09-27 | 2023-03-30 | Marion BERNARDEAU | Feed additive compositions and methods for using the same |
| JP2025501588A (ja) | 2021-12-24 | 2025-01-22 | ダニスコ・ユーエス・インク | ハイブリッドグルコアミラーゼ及びその使用方法 |
| CN120390792A (zh) | 2022-12-19 | 2025-07-29 | 丹尼斯科美国公司 | 重组真菌菌株及其用于产生一致性蛋白质的方法 |
| EP4701430A1 (en) | 2023-04-28 | 2026-03-04 | International N&H Denmark ApS | Ruminant feed additive compositions |
Family Cites Families (42)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS5534046A (en) | 1978-09-01 | 1980-03-10 | Cpc International Inc | Novel glucoamyrase having excellent heat resistance and production |
| GB2089836B (en) | 1980-12-16 | 1984-06-20 | Suntory Ltd | Process for producing alcohol by fermentation without cooking |
| US4794175A (en) | 1983-12-20 | 1988-12-27 | Cetus Corporation | Glucoamylase CDNA |
| US4618579A (en) | 1984-09-28 | 1986-10-21 | Genencor, Inc. | Raw starch saccharification |
| JPH0630586B2 (ja) | 1984-12-15 | 1994-04-27 | サントリー株式会社 | グルコアミラ−ゼ遺伝子 |
| DE3650202T3 (de) | 1985-08-29 | 2004-06-03 | Genencor International, Inc., South San Francisco | Expression von heterologen Polypeptiden in filamentösen Pilzen, Verfahren zu deren Herstellung und Vektoren zu deren Herstellung. |
| US5024941A (en) | 1985-12-18 | 1991-06-18 | Biotechnica International, Inc. | Expression and secretion vector for yeast containing a glucoamylase signal sequence |
| DK122686D0 (da) | 1986-03-17 | 1986-03-17 | Novo Industri As | Fremstilling af proteiner |
| GB8610600D0 (en) | 1986-04-30 | 1986-06-04 | Novo Industri As | Transformation of trichoderma |
| EP0316444B1 (en) | 1987-06-06 | 1995-08-02 | Omnigene Inc | Yeast expression vector |
| US6803499B1 (en) | 1989-08-09 | 2004-10-12 | Dekalb Genetics Corporation | Methods and compositions for the production of stably transformed, fertile monocot plants and cells thereof |
| US6777589B1 (en) | 1990-01-22 | 2004-08-17 | Dekalb Genetics Corporation | Methods and compositions for the production of stably transformed, fertile monocot plants and cells thereof |
| US5475101A (en) | 1990-10-05 | 1995-12-12 | Genencor International, Inc. | DNA sequence encoding endoglucanase III cellulase |
| US5290474A (en) | 1990-10-05 | 1994-03-01 | Genencor International, Inc. | Detergent composition for treating cotton-containing fabrics containing a surfactant and a cellulase composition containing endolucanase III from trichoderma ssp |
| US5246853A (en) | 1990-10-05 | 1993-09-21 | Genencor International, Inc. | Method for treating cotton-containing fabric with a cellulase composition containing endoglucanase components and which composition is free of exo-cellobiohydrolase I |
| DK0553280T4 (da) | 1990-10-05 | 2010-11-15 | Genencor Int | Fremgangsmåde til behandling af bomuldsholdige tekstiler med cellulase |
| WO1992010581A1 (en) | 1990-12-10 | 1992-06-25 | Genencor International, Inc. | IMPROVED SACCHARIFICATION OF CELLULOSE BY CLONING AND AMPLIFICATION OF THE β-GLUCOSIDASE GENE OF TRICHODERMA REESEI |
| US5861271A (en) | 1993-12-17 | 1999-01-19 | Fowler; Timothy | Cellulase enzymes and systems for their expressions |
| ATE294871T1 (de) | 1994-06-30 | 2005-05-15 | Novozymes Biotech Inc | Nicht-toxisches, nicht-toxigenes, nicht- pathogenes fusarium expressionssystem und darin zu verwendende promotoren und terminatoren |
| US5744716A (en) | 1995-06-08 | 1998-04-28 | Scp Global Technologies, A Division Of Preco, Inc. | Fluid displacement level, density and concentration measurement system |
| US6537792B1 (en) | 1996-07-24 | 2003-03-25 | Iowa State University | Protein engineering of glucoamylase to increase pH optimum, substrate specificity and thermostability |
| US6255084B1 (en) | 1997-11-26 | 2001-07-03 | Novozymes A/S | Thermostable glucoamylase |
| CA2331340A1 (en) * | 1998-07-15 | 2000-01-27 | Novozymes A/S | Glucoamylase variants |
| US6352851B1 (en) * | 1998-07-15 | 2002-03-05 | Novozymes A/S | Glucoamylase variants |
| US6268328B1 (en) | 1998-12-18 | 2001-07-31 | Genencor International, Inc. | Variant EGIII-like cellulase compositions |
| US6254914B1 (en) | 1999-07-02 | 2001-07-03 | The Board Of Trustees Of The University Of Illinois | Process for recovery of corn coarse fiber (pericarp) |
| CN100510067C (zh) * | 1999-07-09 | 2009-07-08 | 诺维信公司 | 葡糖淀粉酶变体 |
| CA2418317A1 (en) | 2000-08-11 | 2002-02-21 | Genencor International, Inc. | Bacillus transformation, transformants and mutant libraries |
| US6582914B1 (en) | 2000-10-26 | 2003-06-24 | Genencor International, Inc. | Method for generating a library of oligonucleotides comprising a controlled distribution of mutations |
| AU2002333212A1 (en) * | 2001-10-01 | 2003-04-14 | Novozymes A/S | Glucoamylase variants |
| GB0129864D0 (en) | 2001-12-13 | 2002-02-06 | Danisco | Animal feed |
| AU2004219649A1 (en) | 2003-03-10 | 2004-09-23 | Broin And Associates, Inc. | Method for producing ethanol using raw starch |
| CN102277341B (zh) | 2003-03-10 | 2015-07-22 | 诺维信公司 | 酒精产品生产方法 |
| ATE524491T1 (de) | 2003-05-29 | 2011-09-15 | Genencor Int | Neue trichoderma-gene |
| US6899910B2 (en) | 2003-06-12 | 2005-05-31 | The United States Of America As Represented By The Secretary Of Agriculture | Processes for recovery of corn germ and optionally corn coarse fiber (pericarp) |
| CN102174490B (zh) | 2003-11-21 | 2016-09-28 | 丹尼斯科美国公司 | 颗粒淀粉水解酶在木霉菌中的表达和从颗粒淀粉底物产生葡萄糖的方法 |
| US7413887B2 (en) | 2004-05-27 | 2008-08-19 | Genecor International, Inc. | Trichoderma reesei glucoamylase and homologs thereof |
| CN1981037B (zh) | 2004-05-27 | 2011-08-31 | 金克克国际有限公司 | 白曲霉酸稳定性α淀粉酶的异源表达和在颗粒淀粉水解中的应用 |
| GB0423139D0 (en) | 2004-10-18 | 2004-11-17 | Danisco | Enzymes |
| CA2589725C (en) * | 2004-11-30 | 2014-11-25 | Genencor International, Inc. | Trichoderma reesei glucoamylase and homologs thereof |
| ATE530643T1 (de) | 2004-12-30 | 2011-11-15 | Genencor Int | Säureaktive fungale protease |
| CN101522893B (zh) * | 2006-10-10 | 2014-07-09 | 丹尼斯科美国公司 | 具有改变性质的葡糖淀粉酶变体 |
-
2008
- 2008-11-20 EP EP17205190.6A patent/EP3323889B1/en active Active
- 2008-11-20 WO PCT/US2008/012934 patent/WO2009067218A2/en not_active Ceased
- 2008-11-20 US US12/292,563 patent/US8058033B2/en active Active
- 2008-11-20 JP JP2010534956A patent/JP5594899B2/ja not_active Expired - Fee Related
- 2008-11-20 DK DK17205190.6T patent/DK3323889T3/da active
- 2008-11-20 ES ES17205190T patent/ES2795989T3/es active Active
- 2008-11-20 DK DK14177958.7T patent/DK2816109T3/en active
- 2008-11-20 CN CN2008801166952A patent/CN101868537B/zh active Active
- 2008-11-20 BR BRPI0820483-7A patent/BRPI0820483B1/pt active IP Right Grant
- 2008-11-20 EP EP14177958.7A patent/EP2816109B1/en active Active
- 2008-11-20 DK DK08851977.2T patent/DK2222842T3/en active
- 2008-11-20 PL PL08851977T patent/PL2222842T3/pl unknown
- 2008-11-20 HU HUE17205190A patent/HUE050328T2/hu unknown
- 2008-11-20 CA CA2705941A patent/CA2705941C/en active Active
- 2008-11-20 ES ES08851977.2T patent/ES2527586T3/es active Active
- 2008-11-20 EP EP08851977.2A patent/EP2222842B1/en active Active
- 2008-11-20 MX MX2010005461A patent/MX2010005461A/es active IP Right Grant
-
2011
- 2011-09-30 US US13/251,116 patent/US8679792B2/en active Active
-
2014
- 2014-02-04 US US14/172,790 patent/US9290753B2/en active Active
Also Published As
| Publication number | Publication date |
|---|---|
| DK2222842T3 (en) | 2015-01-19 |
| PL2222842T3 (pl) | 2015-03-31 |
| US9290753B2 (en) | 2016-03-22 |
| US20120149087A1 (en) | 2012-06-14 |
| HUE050328T2 (hu) | 2020-11-30 |
| DK2816109T3 (en) | 2018-04-16 |
| EP2222842A2 (en) | 2010-09-01 |
| CN101868537A (zh) | 2010-10-20 |
| CN101868537B (zh) | 2013-11-06 |
| CA2705941C (en) | 2018-05-22 |
| DK3323889T3 (da) | 2020-06-29 |
| EP3323889A1 (en) | 2018-05-23 |
| EP2816109A1 (en) | 2014-12-24 |
| EP3323889B1 (en) | 2020-03-25 |
| EP2222842B1 (en) | 2014-10-15 |
| ES2795989T3 (es) | 2020-11-25 |
| JP5594899B2 (ja) | 2014-09-24 |
| BRPI0820483A2 (pt) | 2014-10-14 |
| ES2527586T3 (es) | 2015-01-27 |
| MX2010005461A (es) | 2010-06-02 |
| US20140227754A1 (en) | 2014-08-14 |
| WO2009067218A2 (en) | 2009-05-28 |
| WO2009067218A3 (en) | 2009-08-06 |
| JP2011502543A (ja) | 2011-01-27 |
| US20090275080A1 (en) | 2009-11-05 |
| CA2705941A1 (en) | 2009-05-28 |
| US8058033B2 (en) | 2011-11-15 |
| US8679792B2 (en) | 2014-03-25 |
| EP2816109B1 (en) | 2018-01-10 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US11130946B2 (en) | Glucoamylase variants with altered properties | |
| ES2795989T3 (es) | Variantes de glucoamilasa con propiedades modificadas | |
| EP2481796B1 (en) | Glucoamylase variants | |
| US8592194B2 (en) | Glucoamylase variants with altered properties |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| B06F | Objections, documents and/or translations needed after an examination request according [chapter 6.6 patent gazette] | ||
| B06A | Patent application procedure suspended [chapter 6.1 patent gazette] | ||
| B09B | Patent application refused [chapter 9.2 patent gazette] | ||
| B12B | Appeal against refusal [chapter 12.2 patent gazette] | ||
| B16A | Patent or certificate of addition of invention granted [chapter 16.1 patent gazette] |
Free format text: PRAZO DE VALIDADE: 20 (VINTE) ANOS CONTADOS A PARTIR DE 20/11/2008, OBSERVADAS AS CONDICOES LEGAIS. PATENTE CONCEDIDA CONFORME ADI 5.529/DF, QUE DETERMINA A ALTERACAO DO PRAZO DE CONCESSAO. |