CN100342014C - 重组d-氨基酸氧化酶 - Google Patents
重组d-氨基酸氧化酶 Download PDFInfo
- Publication number
- CN100342014C CN100342014C CNB2004100308428A CN200410030842A CN100342014C CN 100342014 C CN100342014 C CN 100342014C CN B2004100308428 A CNB2004100308428 A CN B2004100308428A CN 200410030842 A CN200410030842 A CN 200410030842A CN 100342014 C CN100342014 C CN 100342014C
- Authority
- CN
- China
- Prior art keywords
- sequence
- leu
- gly
- ser
- amino acid
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
Images

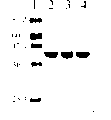
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0012—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7)
- C12N9/0014—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7) acting on the CH-NH2 group of donors (1.4)
- C12N9/0022—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7) acting on the CH-NH2 group of donors (1.4) with oxygen as acceptor (1.4.3)
- C12N9/0024—D-Amino acid oxidase (1.4.3.3)
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Molecular Biology (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Medicinal Chemistry (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Enzymes And Modification Thereof (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Immobilizing And Processing Of Enzymes And Microorganisms (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Cephalosporin Compounds (AREA)
Abstract
本发明提供一种其催化头胞菌素C的活力比亲本D-氨基酸氧化酶所具有的催化活力至少高出25%的重组D-氨基酸氧化酶及其编码DNA序列。
Description
技术领域
本发明涉及生物工程技术领域,特别是,涉及新的D-氨基酸氧化酶的制备和应用。本发明提供的二枚高酶活的重组D-氨基酸氧化酶,可用于将头孢菌素C(cephalosporin C)转化成戊二酰-7-氨基头孢霉烷酸(glutaryl-7-aminocephalosporanic acid)。
背景技术
半合成头孢菌素的母核7-氨基头孢霉烷酸(7-aminocephalosporanic acid)可由化学法裂解头孢菌素C而成。但化学法使用大量有毒化学试剂,污染环境,且反应步骤繁复,得率较低。近年来,酶法成为制备7-氨基头孢霉烷酸的另一主要工艺。酶法转化头孢菌素C包括两个步骤:(1)D-氨基酸氧化酶氧化头孢菌素C(cephalosporin C)生成戊二酰-7-氨基头孢霉烷酸(glutaryl-7-aminocephalosporanic acid);(2)戊二酰-7-氨基头孢霉烷酸酰基酶水解戊二酰-7-氨基头孢霉烷酸产生7-氨基头孢霉烷酸。目前,工业上采用的D-氨基酸氧化酶主要来自红冬孢酵母(Rhodotorula gracilis)或三角酵母(Trigonopsis variabilis)。但是该两种酶催化头孢菌素C的比活均较低(Simonetta,et al.,Biochimica et Biophysica Acta,914:136-142(1987);U.S.Pat.No.5,453,374;U.S.Pat.No.5,208,155)。因而,在现有技术中,仍然存在对高催化活性的D-氨基酸氧化酶的需求,可从而降低酶法制备7-氨基头孢霉烷酸的成本。
发明内容
本发明的目的在于提供具高催化头孢菌素C活力的重组D-氨基酸氧化酶。本发明的目的还在于提供使用本发明的高催化活性重组D-氨基酸氧化酶生产戊二酰-7-氨基头孢霉烷酸。
本发明将三角酵母(Trigonopsis variabilis)FA10的D-氨基酸氧化酶基因(Li,W.et al.,Acta Microbiologica Sinica 31:251-253,1991)克隆至适当的载体上,采用定点突变方法进行基因改进。具体而言,对第53位氨基酸的编码核苷酸进行定点突变,从而获得高催化活性的重组D-氨基酸氧化酶。
一方面,本发明提供一种编码重组D-氨基酸氧化酶的DNA序列,其特征在于所述重组D-氨基酸氧化酶的基因与序列表中的序列1相比,在编码第53位氨基酸的核苷酸密码子中存在导致其编码的苏氨酸被其他氨基酸取代的至少一个核苷酸的差异,且其编码蛋白质所具有的D-氨基酸氧化酶催化头孢菌素C的活力比具有序列2所示氨基酸序列的亲本D-氨基酸氧化酶所具有的催化头孢菌素C的活力至少高出25%,优选高出35%,更优选高出50%,最优选高出100%。
优选的是,本发明的DNA序列含有编码序列表中序列4或6所示氨基酸序列的核苷酸序列,更优选该DNA序列包含序列3或5所示的核苷酸序列。
另一方面,本发明提供一种多肽,其特征在于以序列表中的序列2为参考序列,对应于序列2中第53位苏氨酸的氨基酸被其他氨基酸替代,且其所具有的D-氨基酸氧化酶催化头孢菌素C的活力比具有序列2所示氨基酸序列的亲本D-氨基酸氧化酶所具有的D-氨基酸氧化酶催化活力至少高出25%,优选高出35%,更优选高出50%,最优选高出100%。
优选的是,本发明的多肽,其特征在于,以序列表中的序列2为参考序列,对应于序列2中第53位苏氨酸的氨基酸为丝氨酸或脯氨酸。在本发明的实施例中,具体举出了两个新的重组D-氨基酸氧化酶,重组D-氨基酸氧化酶GHA和重组D-氨基酸氧化酶GHB。重组D-氨基酸氧化酶GHA具有序列4所示的氨基酸序列,其催化头孢菌素C的比活较亲本高105%;重组D-氨基酸氧化酶GHB具有序列6所示的氨基酸序列,其催化头孢菌素C的比活较亲本高35%。本发明中亲本基因系指來自三角酵母(Trigonopsis variabilis)FA10的D-氨基酸氧化酶基因(Li,W.et al.,Acta Microbiologica Sinica 31:251-253,1991),其核苷酸序列如序列1所示,氨基酸序列如序列2所示。
本发明的重组D-氨基酸氧化酶还包括,对GHA或GHB的氨基酸序列进行一个或多个氨基酸的保守替代的形式,增加或减少一个或多个氨基酸的衍生物的形式。
本发明制备重组D-氨基酸氧化酶GHA或重组D-氨基酸氧化酶GHB的方法中,适用的载体包括但不限于原核表达载体pRSET-A和pET;包括但不限于真核表达载体pYD1和pYES2;包括但不限于克隆载体pGEM-T Easy、pUC18、pUC19和pBluescript-SK(+/-)。
本发明制备重组D-氨基酸氧化酶的方法中,宿主细胞可以是原核细胞也可以是真核细胞。适用的原核微生物包括但不限于大肠杆菌(E.coli)、枯草芽孢杆菌(Bacillus subtilis)、短芽孢杆菌(Bacillus brevis)和链霉菌(Streptomyces);适用的真核微生物包括但不限于酿酒酵母(Saccharomyces cerevisiae)、红冬孢酵母、三角酵母、黑曲霉(Aspergillusniger)、乳酸克鲁维酵母(Kluyveromyces lactis)和毕赤巴斯德酵母(Pichiapastoris)。
本发明制备重组D-氨基酸氧化酶GHA和重组D-氨基酸氧化酶GHB的方法中,采用本领域已知的适当方法,所述重组D-氨基酸氧化酶可在原核细胞或真核细胞胞内或胞外表达。可通过将上述本发明的DNA序列通过本领域已知的任何方法导入适当的微生物宿主细胞例如大肠杆菌或酵母细胞中,进行多肽的表达。
如果表达的重组D-氨基酸氧化酶是胞外酶,可以从细胞产物中通过常规技术如硫酸铵分离和丙酮沉淀等,获得部分纯化的重组D-氨基酸氧化酶,或通过常规纯化技术如离子交换亲和层析等从细胞产物中完全纯化。因此,在使用时,本发明的重组D-氨基酸氧化酶可以是未经提纯的粗酶;也可以是部分纯化的酶;也可以是纯化的酶。
如果重组D-氨基酸氧化酶是胞内酶,则可先将宿主细胞破碎,然后例如通过离心去除细胞碎片,接着再通过分级分离,分离纯化重组的D-氨基酸氧化酶。
此外,为了方便在工业上的各种应用,可根据本领域中已知的任何适当的方法将本发明的重组D-氨基酸氧化酶制成固相细胞。为制备所述的固相细胞,可以将含有本发明重组D-氨基酸氧化酶的转化细胞按照本领域已知的方法与固相载体连接起来,制成包含所述D-氨基酸氧化酶的固相细胞。在本发明中,还可以用常规方法将未经纯化的粗酶、部分纯化的酶或纯化的重组D-氨基酸氧化酶本身固定在载体上。可通过将本发明的重组D-氨基酸氧化酶吸附到离子交换树脂以制备固定化的固相酶。
此外,可根据例如Miyake Y.,Aki K.,Hashimoto S.,and Yamano T.(1965)Crystallization and some properties of D-Amino Acid OxidaseApoenzyme,Biochemica et Biophysica Acta,105:86-99来制备结晶酶。很显然,这些固相酶或结晶酶及其应用也在本发明的范围之内。
附图说明
图1质粒pRSET-kan图谱。
图2质粒pRSET-kan序列。
图3重组D-氨基酸氧化酶GHA和重组D-氨基酸氧化酶GHB的SDS-PAGE电泳图。1,蛋白质分子量标记BenchMarkTM Pre-StainedProtein Ladder(Invitrogen),单位是KDa;2,3和4分别是纯化的亲本三角酵母D-氨基酸氧化酶、重组D-氨基酸氧化酶GHA,以及重组D-氨基酸氧化酶GHB。
具体实施方式
下列实施例仅用于说明而不应视为限定本发明的范围。实施实例中未注明具体条件者,按常规条件或制造商建议的条件进行。
实例1 D-氨基酸氧化酶基因重组质粒pRSET-A-DAO的构建
根据已知三角酵母D-氨基酸氧化酶基因5’和3’端序列(Gonzalez,F.J.,Montes,J.,Martin,F.,Lopez,M.C.,Ferminan,E.,Catalan,J.,Galan,M.A.Dominguez,A.Molecular cloning of TvDAO1,a gene encoding aD-amino acid oxidase from Trigonopsis variabilis and its expression inSaccharomyces cerevisiae and Kluyveromyces lactis.Yeast 13:1399-1408;1997)设计引物如下:
5’-NdeI(引入NdeI酶切位点):
5’-TAGGGCTGA
CATATGGCTAAAATCGTTGTTATTGGTGC-3’
(序列7)
3’-BglII(引入BglII酶切位点):
5’-TAGGGCTGA
AGATCTCTAAAGGTTTGGACGAGTAAGAGC-3’(序列8)
以质粒pJL(杨藴刘等,中国专利申请公开号:CN 1371999A)为模板,用以上两个引物,在Pfu DNA聚合酶(Promega)的作用下,合成三角酵母D-氨基酸氧化酶基因。pJL携带有三角酵母FA10 D-氨基酸氧化酶基因(Li,W.et al.,Acta Microbiologica Sinica 31:251-253,1991)。PCR反应条件为:40ng pJL,0.4μM 5’-NdeI,0.4μM 3’-BglII,50μM dATP,50μM dTTP,50μM dCTP,50μM dGTP,20mMTris-HCl(pH 8.8),10mM KCl,10mM(NH4)2SO4,2mM MgSO4,0.1%Triton X-100,2.5U Pfu DNA聚合酶,用无菌水调反应体积至50μl。
PCR扩增反应程序为:
94℃,60sec 循环10次 94℃,60sec 循环25次
94℃,5min→50℃,60sec→→→→→60℃,60sec→→→→→72℃,10min
72℃,120sec 72℃,120sec
得1,098bp长PCR产物,其5’和3’端分别具有NdeI和BglII限制性酶切位点。PCR产物经1%琼脂糖电泳提纯,NdeI及BglII酶切后,与经NdeI及BglII酶切质粒pRSET-A(Invitrogen)得到的2.9Kb的片段连接,得连接产物pRSET-A-DAO。将pRSET-A-DAO转化感受态大肠杆菌BL21(DE3)pLysS(Novagen),在氨苄青霉素LB平板上37℃培养,按照Molecular Cloning-A Laboratory Mannua,ed.By J.Sambrook,et.al.,1989,CSHL Press描述的方法提取质粒,经DNA测序,确定三角酵母D-氨基酸氧化酶基因核苷酸序列如序列1,其推测出的氨基酸序列如序列2所示。
实例2 pRSET-kan载体的构建
为从pRSET-A去除氨苄青霉素抗性基因,根据载体pRSET-A的序列设计引物如下:
VET-F:5’-CTGTCAGACCAAGTTTACTCATATATACTTTAG-3’
(序列9)
VET-R:5’-ACTCTTCCTTTTTCAATATTATTGAAGC-3’(序列10)
为从载体pET-28b(Novogen)扩增出卡那霉素抗性基因,根据载体pET-28b(Novogen)的序列设计引物如下:
KAN-F:5’-ATGAGTCATATTCAACGGGAAAC-3’(序列11)
KAN-R:5’-TTAGAAAAACTCATCGAGCATCAAATG-3’(序列12)
扩增去除氨苄青霉素抗性基因的pRSET-A片段的PCR条件为:50ng pRSET-A,0.4μM VET-F,0.4μM VET-R,50μM dATP,50μMdTTP,50μM dCTP,50μM dGTP,20mM Tris-HCl(pH 8.8),10mM KCl,10mM(NH4)2SO4,2mM MgSO4,0.1%Triton X-100,2.5U Pfu DNA聚合酶,用无菌水调反应体积至50μl。扩增pET-28b卡那霉素抗性基因的PCR条件为:50ng pET-28b,0.4μM KAN-F,0.4μM KAN-R,50μMdATP,50μM dTTP,50μM dCTP,50μM dGTP,20mM Tris-HCl(pH8.8),10mM KCl,10mM(NH4)2SO4,2mM MgSO4,0.1%Triton X-100,2.5U Pfu DNA聚合酶,用无菌水调反应体积至50μl。PCR扩增反应程序均为:
94℃,60sec 循环35次
94℃,5min→→50℃,60sec→→→→→72℃,10min 72℃,4min
72℃,4min
1%琼脂糖电泳并提纯PCR产物(去除氨苄青霉素抗性基因的pRSET-A片段长度为2036bp,卡那霉素抗性基因为816bp),连接该两个片断得连接产物pRSET-kan(图1)。将pRSET-kan转化感受态大肠杆菌BL21(DE3)pLysS,在卡那霉素LB平板上37℃培养,提取质粒,经DNA测序确定核苷酸序列如图2和序列13所示。
实例3 重组D-氨基酸氧化酶GHA的构建
重组D-氨基酸氧化酶GHA由定点突变构建。定点突变技术主要参考PCR Protocols(编者:John M.S.Bartlett and David Stirling.Totowa,N.J.:Humana Press,2003.)一书之描述。
根据实例1克隆的三角酵母D-氨基酸氧化酶基因之序列(序列1),设计引物如下:
引物A:5’TAGGGCTGA
CATATGGCTAAAATCGTTGTTATTG-3’(序列14)
引物B:5’TAGGGCTGA
AGATCTCTAAAGGTTTGGACGAG-3’(序列15)
引物C 1:5’-GCAGGTGCCAACTGGCTC
CCGTTTTACGATGGAGGCAAG-3’(序列16)
引物D:5’-GAGCCAGTTGGCACCTGCCCAAGG-3’(序列17)
引物A和B是一对外引物。引物A包含NdeI酶切位点,并有部份碱基与D-氨基酸氧化酶基因5’端序列交迭;引物B包含BglII酶切位点,并有部份碱基与D-氨基酸氧化酶基因3’端序列交迭。引物C1和D是内引物。引物C1将亲本D-氨基酸氧化酶基因53位之苏氨酸转变成脯氨酸。引物D部份碱基与引物C1的序列交迭。
利用聚合酶链式反应,以pRSET-A-DAO为模板,用引物对A和D扩增模板片段1;用引物对B和C1扩增模板片段2。扩增反应条件为:20ng pRSET-A-DAO,20mM Tris-HCl(pH 8.8),10mM KCl,10mM(NH4)2SO4,2mM MgSO4,0.1%Triton X-100,0.4μM引物A和0.4μM引物D(扩增模板片段1)或0.4μM引物B和0.4μM引物C1(扩增模板片段2),50μM dATP,50μM dTTP,50μM dCTP,50μMdGTP,1.5U Pfu DNA聚合酶,用无菌水调反应体积至50μl。聚合酶链式反应扩增程序为:
94℃,60sec 循环30次
94℃,2min→53℃,60sec→→→→→72℃,10min
72℃,60sec
扩增得到的模板片段1和模板片段2,经1%琼脂糖凝胶电泳分离纯化后,用以扩增全长基因。扩增全长基因的反应条件为:20ng模板片段1,20ng模板片段2,20mM Tris-HCl(pH 8.8),10mM KCl,10mM(NH4)2SO4,2mM MgSO4,0.1%Triton X-100,0.4μM引物A和0.4μM引物B,50μM dATP,50μM dTTP,50μM dCTP,50μMdGTP,1.5U Pfu DNA聚合酶,用无菌水调反应体积至50μl。聚合酶链式反应扩增程序为:
94℃,60sec 循环35次
94℃,2min→53℃,60sec→→→→→ →72℃,10min
72℃,120sec
全长扩增反应得到重组D-氨基酸氧化酶GHA基因,经NdeI及BglII酶切后连接到pRSET-kan得连接产物pRSET-kan-DAOGHA,将pRSET-kan-DAOGHA转化感受态大肠杆菌BL21(DE3)pLysS,卡那霉素LB平板37℃培养,提取质粒,经DNA测序确定引入的突变无误,确定重组D-氨基酸氧化酶GHA核苷酸序列如序列3所示,其推测的氨基酸序列如序列4所示。
实例4 重组D-氨基酸氧化酶GHB的构建
重组D-氨基酸氧化酶GHB由定点突变构建。定点突变技术主要参考PCR Protocols(编者:John M.S.Bartlett and Dayid Stirling.Totowa,N.J.:Humana Press,2003.)一书之描述。
根据实例1克隆的三角酵母D-氨基酸氧化酶基因之序列(序列1),设计引物如下:
引物A:5’-TAGGGCTGA
CATATGGCTAAAATCGTTGTTATTG-3’(序列14)
引物B:5’-TAGGGCTGA
AGATCTCTAAAGGTTTGGACGAG-3’(序列15)
引物C2:5’GCAGGTGCCAACTGGCTC
AGCTTTTACGATGGAGGCAAG-3’(序列18)
引物D:5’-GAGCCAGTTGGCACCTGCCCAAGG-3’(序列17)
以上引物A、B和D与实例3所述相同。引物C2是内引物,将亲本D-氨基酸氧化酶基因53位之苏氨酸转变成丝氨酸。引物D部份碱基与引物C2的序列交迭。模板片段1之扩增如实例3。利用聚合酶链式反应,以pRSET-A-DAO DNA为模板,用引物对B和C2扩增模板片段3。扩增反应条件为:20ng pRSET-A-DAO,20mM Tris-HCl(pH 8.8),10mM KCl,10mM(NH4)2SO4,2mM MgSO4,0.1%Triton X-100),0.4μM引物B和0.4μM引物C2,50μM dATP,50μM dTTP,50μM dCTP,50μM dGTP,1.5U Pfu DNA聚合酶,用无菌水调反应体积至50μl。聚合酶链式反应扩增程序为:
94℃,60sec 循环30次
94℃,2min→53℃,60sec→→→→→→72℃,10min
72℃,60sec
扩增得到模板片段3,经1%琼脂糖凝胶电泳分离纯化后,用以扩增全长基因。扩增全长基因的反应条件为:20ng模板片段1,20ng模板片段3,20mM Tris-HCl(pH 8.8),10mM KCl,10mM(NH4)2SO4,2mM MgSO4,0.1%Triton X-100,0.4μM引物A和0.4μM引物B,50μM dATP,50μM dTTP,50μM dCTP,50μM dGTP,1.5U Pfu DNA聚合酶,用无菌水调反应体积至50μl。聚合酶链式反应扩增程序为:
94℃,60sec 循环35次
94℃,2min→53℃,60sec→→→→→ →72℃,10min
72℃,120sec
全长扩增反应得到重组D-氨基酸氧化酶GHB基因,经NdeI及BglII酶切后连接到pRSET-kan上,转化感受态大肠杆菌BL21(DE3)pLysS,卡那霉素LB平板37℃培养,提取质粒,经DNA测序确定引入的突变无误,确定重组D-氨基酸氧化酶GHB核苷酸序列如序列5所示,其推测的氨基酸序列如序列6所示。
实例5 D-氨基酸氧化酶的纯化
D-氨基酸氧化酶的纯化基本按Alonso,J.,Barredo,J.L.,Diez,B.,Mellado,E.,Salto,F.,Garcia,J.L.,Cortes,E.(D-amino-acid oxidase genefrom Rhodotorula gracilis[Rhodosporidium toruloides]ATCC 26217.Microbiology 144:1095-1101;1998)所述。取单菌落pRSET-kan-DAOGHA转化之大肠杆菌BL21(DE3)pLysS细胞(实例3)在200ml卡那霉素LB培养液37℃,250rpm培养12小时,然后加入1mM IPTG诱导细胞6小时。离心收集细胞,洗涤并溶于20ml缓冲液A[20mM磷酸钠缓冲液(pH 8.0)含20%甘油,5mM 2-巯基乙醇,1mM PMSF和2mM EDTA],用超声波裂解细胞。离心(4℃,13,000g,20分钟)去除细胞碎片,取上清液上柱DEAE-纤维素柱(Sigma公司,6×2.5cm),用缓冲液A洗脱活性成份,得部分纯化的重组D-氨基酸氧化酶GHA。部分纯化的重组D-氨基酸氧化酶GHA可继续由下列方法纯化:上缓冲液A平衡之Cibacron Blue 3GA-琼脂糖柱(Pharmacia LKBBiotechnology,4×1cm),用30ml 1M磷酸钠(pH 8.0)冲洗,然后用10ml缓冲液A+50μM FAD洗脱活性部分。用SDS-PAGE检测蛋白质的纯度(图3)。三角酵母D-氨基酸氧化酶和重组D-氨基酸氧化酶GHB的纯化也依照上述步骤实行(图3)。
实例6 D-氨基酸氧化酶活性的测定
参照Isogai,T.,Ono,H.,Ishitani,Y.,Kojo,H.,Ueda,Y.,Kohsaka,M.(Structure and expression of cDNA for D-amino acid oxidase activeagainst cephalosporin C from Fusarium solani.J Biochem[Tokyo].108:1063-1069,1990)所述,具体步骤有变更。取三毫升反应液含磷酸钠缓冲液(50mM,pH 7.5)、75mM头孢菌素C钠盐以及三毫升如实例5所述、部分纯化的三角酵母D-氨基酸氧化酶,加氧在22℃振荡反应60分钟。反应开始后在不同时间(1、5、10、30、60分钟)抽取100μl反应液与10μl 3%过氧化氢混匀,再加入50μl 10%三氯醋酸,混匀以终止反应。将终止反应液离心(10,000g,3分钟),取10μl上清液和990μl HPLC流动相混合,上HPLC柱检测。HPLC色谱柱:DiamonsilTM C18,250×4.6mm(迪马公司,北京);流动相:50mMK2HPO4/KH2PO4(pH 7.0),5%乙腈;柱温度:30℃;流速:1ml/min;检测:260nm UV。一单位酶活性定义为在上述条件每分钟转化一微摩尔头孢菌素C为戊二酰-7-氨基头孢霉烷酸的酶量。重组D-氨基酸氧化酶GHA和重组D-氨基酸氧化酶GHB活性的测定也如上述。测得重组D-氨基酸氧化酶GHA的比活是亲本三角酵母D-氨基酸氧化酶比活的205%;重组D-氨基酸氧化酶GHB的比活是亲本D-氨基酸氧化酶比活的135%。
实例7 D-氨基酸氧化酶 固相酶的制备
酶的提取和纯化按实例5所述。D-氨基酸氧化酶固定化酶的制备参照德国Rhm公司(Darmstadt,Germany)的说明进行。量取50mL200mg总蛋白、部分纯化的重组D-氨基酸氧化酶GHA酶溶液,加入K2HPO4与KH2PO4使溶液的磷酸盐浓度至0.5M,pH值为7.5。加入Eupergit CTM(Rhm GmbH,Darmstadt,Germany)干载体5g,室温(17℃-23℃)75rpm搅拌72小时,过滤除去上清液,用蒸馏水反复冲洗,抽滤后,得固定化酶19.4g。酶比活的测定方法基本与实例6所述相同,惟所用反应液体积为1,000ml,加入重组D-氨基酸氧化酶GHA固相酶19.4g。测得重组D-氨基酸氧化酶GHA的比活为65单位/克湿载体。
本发明不受上述具体文字描述的限制。本发明可在权利要求书所概括的范围内做各种改变。这些改变均在本发明的范围之内。
<110>Bioright
<120>工程D-氨基酸氧化酶
<130>SPI020563-09
<160>18
<170>PatentIn version 3.1
<210>1
<211>1071
<212>DNA
<213>Trigonopsis variabilis
<400>1
atggctaaaa tcgttgttat tggtgccggt gttgccggtt taactacagc tcttcaactt 60
cttcgtaaag gacatgaggt tacaattgtg tccgagttta cgcccggtga tcttagtatc 120
ggatatacct cgccttgggc aggtgccaac tggctcacat tttacgatgg aggcaagtta 180
gccgactacg atgccgtctc ttatcctatc ttgcgagagc tggctcgaag cagccccgag 240
gctggaattc gactcatcaa ccaacgctcc catgttctca agcgtgatct tcctaaactg 300
gaaggtgcca tgtcggccat ctgtcaacgc aacccctggt tcaaaaacac agtcgattct 360
ttcgagatta tcgaggacag gtccaggatt gtccacgatg atgtggctta tctagtcgaa 420
tttgcttccg tttgtatcca caccggagtc tacttgaact ggctgatgtc ccaatgctta 480
tcgctcggcg ccacggtggt taaacgtcga gtgaaccata tcaaggatgc caattttcta 540
cactcctcag gatcacgccc cgacgtgatt gtcaactgta gtggtctctt tgcccggttc 600
ttgggaggcg tcgaggacaa gaagatgtac cctattcgag gacaagtcgt ccttgttcga 660
aactctcttc cttttatggc ctccttttcc agcactcctg aaaaagaaaa tgaagacgaa 720
gctctatata tcatgacccg attcgatggt acttctatca ttggcggttg tttccaatcc 780
aacaactggt catccgaacc cgatccttct ctcacccatc gaatcctgtc tagagccctc 840
gaccgattcc cggaactgac caaagatggc cctcttgaca ttgtgcgcga atgcgttggc 900
caccgtcctg gtagagaggg cggtccccga gtagaattag agaagatccc cggcgttggc 960
tttgttgtcc ataactatgg tgccgccggt gctggttacc agtcctctta cggcatggct 1020
gatgaagctg tttcttacgt cgaaagagct cttactcgtc caaaccttta g 1071
<210>2
<211>356
<212>PRT
<213>Trigonopsis variabilis
<400>2
Met Ala Lys Ile Val Val Ile Gly Ala Gly Val Ala Gly Leu Thr Thr
1 5 10 15
Ala Leu Gln Leu Leu Arg Lys Gly His Glu Val Thr Ile Val Ser Glu
20 25 30
Phe Thr Pro Gly Asp Leu Ser Ile Gly Tyr Thr Ser Pro Trp Ala Gly
35 40 45
Ala Asn Trp Leu Thr Phe Tyr Asp Gly Gly Lys Leu Ala Asp Tyr Asp
50 55 60
Ala Val Ser Tyr Pro Ile Leu Arg Glu Leu Ala Arg Ser Ser Pro Glu
65 70 75 80
Ala Gly Ile Arg Leu Ile Asn Gln Arg Ser His Val Leu Lys Arg Asp
85 90 95
Leu Pro Lys Leu Glu Gly Ala Met Ser Ala Ile Cys Gln Arg Asn Pro
100 105 110
Trp Phe Lys Asn Thr Val Asp Ser Phe Glu Ile Ile Glu Asp Arg Ser
115 120 125
Arg Ile Val His Asp Asp Val Ala Tyr Leu Val Glu Phe Ala Ser Val
130 135 140
Cys Ile His Thr Gly Val Tyr Leu Asn Trp Leu Met Ser Gln Cys Leu
145 150 155 160
Ser Leu Gly Ala Thr Val Val Lys Arg Arg Val Asn His Ile Lys Asp
165 170 175
Ala Asn Phe Leu His Ser Ser Gly Ser Arg Pro Asp Val Ile Val Asn
180 185 190
Cys Ser Gly Leu Phe Ala Arg Phe Leu Gly Gly Val Glu Asp Lys Lys
195 200 205
Met Tyr Pro Ile Arg Gly Gln Val Val Leu Val Arg Asn Ser Leu Pro
210 215 220
Phe Met Ala Ser Phe Ser Ser Thr Pro Glu Lys Glu Asn Glu Asp Glu
225 230 235 240
Ala Leu Tyr Ile Met Thr Arg Phe Asp Gly Thr Ser Ile Ile Gly Gly
245 250 255
Cys Phe Gln Ser Asn Asn Trp Ser Ser Glu Pro Asp Pro Ser Leu Thr
260 265 270
His Arg Ile Leu Ser Arg Ala Leu Asp Arg Phe Pro Glu Leu Thr Lys
275 280 285
Asp Gly Pro Leu Asp Ile Val Arg Glu Cys Val Gly His Arg Pro Gly
290 295 300
Arg Glu Gly Gly Pro Arg Val Glu Leu Glu Lys Ile Pro Gly Val Gly
305 310 315 320
Phe Val Val His Asn Tyr Gly Ala Ala Gly Ala Gly Tyr Gln Ser Ser
325 330 335
Tyr Gly Met Ala Asp Glu Ala Val Ser Tyr Val Glu Arg Ala Leu Thr
340 345 350
Arg Pro Asn Leu
355
<210>3
<211>1071
<212>DNA
<213>Trigonopsis variabilis
<400>3
atggctaaaa tcgttgttat tggtgccggt gttgccggtt taactacagc tcttcaactt 60
cttcgtaaag gacatgaggt tacaattgtg tccgagttta cgcccggtga tcttagtatc 120
ggatatacct cgccttgggc aggtgccaac tggctcccgt tttacgatgg aggcaagtta 180
gccgactacg atgccgtctc ttatcctatc ttgcgagagc tggctcgaag cagccccgag 240
gctggaattc gactcatcaa ccaacgctcc catgttctca agcgtgatct tcctaaactg 300
gaaggtgcca tgtcggccat ctgtcaacgc aacccctggt tcaaaaacac agtcgattct 360
ttcgagatta tcgaggacag gtccaggatt gtccacgatg atgtggctta tctagtcgaa 420
tttgcttccg tttgtatcca caccggagtc tacttgaact ggctgatgtc ccaatgctta 480
tcgctcggcg ccacggtggt taaacgtcga gtgaaccata tcaaggatgc caattttcta 540
cactcctcag gatcacgccc cgacgtgatt gtcaactgta gtggtctctt tgcccggttc 600
ttgggaggcg tcgaggacaa gaagatgtac cctattcgag gacaagtcgt ccttgttcga 660
aactctcttc cttttatggc ctccttttcc agcactcctg aaaaagaaaa tgaagacgaa 720
gctctatata tcatgacccg attcgatggt acttctatca ttggcggttg tttccaatcc 780
aacaactggt catccgaacc cgatccttct ctcacccatc gaatcctgtc tagagccctc 840
gaccgattcc cggaactgac caaagatggc cctcttgaca ttgtgcgcga atgcgttggc 900
caccgtcctg gtagagaggg cggtccccga gtagaattag agaagatccc cggcgttggc 960
tttgttgtcc ataactatgg tgccgccggt gctggttacc agtcctctta cggcatggct 1020
gatgaagctg tttcttacgt cgaaagagct cttactcgtc caaaccttta g 1071
<210>4
<211>356
<212>PRT
<213>Trigonopsis variabilis
<400>4
Met Ala Lys Ile Val Val Ile Gly Ala Gly Val Ala Gly Leu Thr Thr
1 5 10 15
Ala Leu Gln Leu Leu Arg Lys Gly His Glu Val Thr Ile Val Ser Glu
20 25 30
Phe Thr Pro Gly Asp Leu Ser Ile Gly Tyr Thr Ser Pro Trp Ala Gly
35 40 45
Ala Asn Trp Leu Pro Phe Tyr Asp Gly Gly Lys Leu Ala Asp Tyr Asp
50 55 60
Ala Val Ser Tyr Pro Ile Leu Arg Glu Leu Ala Arg Ser Ser Pro Glu
65 70 75 80
Ala Gly Ile Arg Leu Ile Asn Gln Arg Ser His Val Leu Lys Arg Asp
85 90 95
Leu Pro Lys Leu Glu Gly Ala Met Ser Ala Ile Cys Gln Arg Asn Pro
100 105 110
Trp Phe Lys Asn Thr Val Asp Ser Phe Glu Ile Ile Glu Asp Arg Ser
115 120 125
Arg Ile Val His Asp Asp Val Ala Tyr Leu Val Glu Phe Ala Ser Val
130 135 140
Cys Ile His Thr Gly Val Tyr Leu Asn Trp Leu Met Ser Gln Cys Leu
145 150 155 160
Ser Leu Gly Ala Thr Val Val Lys Arg Arg Val Asn His Ile Lys Asp
165 170 175
Ala Asn Phe Leu His Ser Ser Gly Ser Arg Pro Asp Val Ile Val Asn
180 185 190
Cys Ser Gly Leu Phe Ala Arg Phe Leu Gly Gly Val Glu Asp Lys Lys
195 200 205
Met Tyr Pro Ile Arg Gly Gln Val Val Leu Val Arg Asn Ser Leu Pro
210 215 220
Phe Met Ala Ser Phe Ser Ser Thr Pro Glu Lys Glu Asn Glu Asp Glu
225 230 235 240
Ala Leu Tyr Ile Met Thr Arg Phe Asp Gly Thr Ser Ile Ile Gly Gly
245 250 255
Cys Phe Gln Ser Asn Asn Trp Ser Ser Glu Pro Asp Pro Ser Leu Thr
260 265 270
His Arg Ile Leu Ser Arg Ala Leu Asp Arg Phe Pro Glu Leu Thr Lys
275 280 285
Asp Gly Pro Leu Asp Ile Val Arg Glu Cys Val Gly His Arg Pro Gly
290 295 300
Arg Glu Gly Gly Pro Arg Val Glu Leu Glu Lys Ile Pro Gly Val Gly
305 310 315 320
Phe Val Val His Asn Tyr Gly Ala Ala Gly Ala Gly Tyr Gln Ser Ser
325 330 335
Tyr Gly Met Ala Asp Glu Ala Val Ser Tyr Val Glu Arg Ala Leu Thr
340 345 350
Arg Pro Asn Leu
355
<210>5
<211>1071
<212>DNA
<213>Trigonopsis variabilis
<400>5
atggctaaaa tcgttgttat tggtgccggt gttgccggtt taactacagc tcttcaactt 60
cttcgtaaag gacatgaggt tacaattgtg tccgagttta cgcccggtga tcttagtatc 120
ggatatacct cgccttgggc aggtgccaac tggctcagct tttacgatgg aggcaagtta 180
gccgactacg atgccgtctc ttatcctatc ttgcgagagc tggctcgaag cagccccgag 240
gctggaattc gactcatcaa ccaacgctcc catgttctca agcgtgatct tcctaaactg 300
gaaggtgcca tgtcggccat ctgtcaacgc aacccctggt tcaaaaacac agtcgattct 360
ttcgagatta tcgaggacag gtccaggatt gtccacgatg atgtggctta tctagtcgaa 420
tttgcttccg tttgtatcca caccggagtc tacttgaact ggctgatgtc ccaatgctta 480
tcgctcggcg ccacggtggt taaacgtcga gtgaaccata tcaaggatgc caattttcta 540
cactcctcag gatcacgccc cgacgtgatt gtcaactgta gtggtctctt tgcccggttc 600
ttgggaggcg tcgaggacaa gaagatgtac cctattcgag gacaagtcgt ccttgttcga 660
aactctcttc cttttatggc ctccttttcc agcactcctg aaaaagaaaa tgaagacgaa 720
gctctatata tcatgacccg attcgatggt acttctatca ttggcggttg tttccaatcc 780
aacaactggt catccgaacc cgatccttct ctcacccatc gaatcctgtc tagagccctc 840
gaccgattcc cggaactgac caaagatggc cctcttgaca ttgtgcgcga atgcgttggc 900
caccgtcctg gtagagaggg cggtccccga gtagaattag agaagatccc cggcgttggc 960
tttgttgtcc ataactatgg tgccgccggt gctggttacc agtcctctta cggcatggct 1020
gatgaagctg tttcttacgt cgaaagagct cttactcgtc caaaccttta g 1071
<210>6
<211>356
<212>PRT
<213>Trigonopsis variabilis
<400>6
Met Ala Lys Ile Val Val Ile Gly Ala Gly Val Ala Gly Leu Thr Thr
1 5 10 15
Ala Leu Gln Leu Leu Arg Lys Gly His Glu Val Thr Ile Val Ser Glu
20 25 30
Phe Thr Pro Gly Asp Leu Ser Ile Gly Tyr Thr Ser Pro Trp Ala Gly
35 40 45
Ala Asn Trp Leu Ser Phe Tyr Asp Gly Gly Lys Leu Ala Asp Tyr Asp
50 55 60
Ala Val Ser Tyr Pro Ile Leu Arg Glu Leu Ala Arg Ser Ser Pro Glu
65 70 75 80
Ala Gly Ile Arg Leu Ile Asn Gln Arg Ser His Val Leu Lys Arg Asp
85 90 95
Leu Pro Lys Leu Glu Gly Ala Met Ser Ala Ile Cys Gln Arg Asn Pro
100 105 110
Trp Phe Lys Asn Thr Val Asp Ser Phe Glu Ile Ile Glu Asp Arg Ser
115 120 125
Arg Ile Val His Asp Asp Val Ala Tyr Leu Val Glu Phe Ala Ser Val
130 135 140
Cys Ile His Thr Gly Val Tyr Leu Asn Trp Leu Met Ser Gln Cys Leu
145 150 155 160
Ser Leu Gly Ala Thr Val Val Lys Arg Arg Val Asn His Ile Lys Asp
165 170 175
Ala Asn Phe Leu His Ser Ser Gly Ser Arg Pro Asp Val Ile Val Asn
180 185 190
Cys Ser Gly Leu Phe Ala Arg Phe Leu Gly Gly Val Glu Asp Lys Lys
195 200 205
Met Tyr Pro Ile Arg Gly Gln Val Val Leu Val Arg Asn Ser Leu Pro
210 215 220
Phe Met Ala Ser Phe Ser Ser Thr Pro Glu Lys Glu Asn Glu Asp Glu
225 230 235 240
Ala Leu Tyr Ile Met Thr Arg Phe Asp Gly Thr Ser Ile Ile Gly Gly
245 250 255
Cys Phe Gln Ser Asn Asn Trp Ser Ser Glu Pro Asp Pro Ser Leu Thr
260 265 270
His Arg Ile Leu Ser Arg Ala Leu Asp Arg Phe Pro Glu Leu Thr Lys
275 280 285
Asp Gly Pro Leu Asp Ile Val Arg Glu Cys Val Gly His Arg Pro Gly
290 295 300
Arg Glu Gly Gly Pro Arg Val Glu Leu Glu Lys Ile Pro Gly Val Gly
305 310 315 320
Phe Val Val His Asn Tyr Gly Ala Ala Gly Ala Gly Tyr Gln Ser Ser
325 330 335
Tyr Gly Met Ala Asp Glu Ala Val Ser Tyr Val Glu Arg Ala Leu Thr
340 345 350
Arg Pro Asn Leu
355
<210>7
<211>38
<212>DNA
<213>Artificial Sequence
<220>
<223>引物(primer for introducing NdeI restriction site)
<400>7
tagggctgac atatggctaa aatcgttgtt attggtgc 38
<210>8
<211>39
<212>DNA
<213>Artificial Sequence
<220>
<223>引物(primer for introducing BglII restriction site)
<400>8
tagggctgaa gatctctaaa ggtttggacg agtaagagc 39
<210>9
<211>33
<212>DNA
<213>Artificial Sequence
<220>
<223>正向引物(forward primer for removing Ampr gene from pRSET-A)
<400>9
ctgtcagacc aagtttactc atatatactt tag 33
<210>10
<211>28
<212>DNA
<213>Artificial Sequence
<220>
<223>反向引物(reverse primer for removing Ampr gene from pRSET-A)
<400>10
actcttcctt tttcaatatt attgaagc 28
<210>11
<211>23
<212>DNA
<213>Artificial Sequence
<220>
<223>正向引物(forward primer for amplifying Kanr gene from pET-28b)
<400>11
atgagtcata ttcaacggga aac 23
<210>12
<211>27
<212>DNA
<213>Artificial Sequence
<220>
<223>反向引物(reverse primer for amplifying Kanr gene from pET-28b)
<400>12
ttagaaaaactcatcgagca tcaaatg 27
<210>13
<211>2852
<212>DNA
<213>Trigonopsis variabilis
<400>13
gatctcgatc ccgcgaaatt aatacgactc actataggga gaccacaacg gtttccctct 60
agaaataatt ttgtttaact ttaagaagga gatatacata tgcggggttc tcatcatcat 120
catcatcatg gtatggctag catgactggt ggacagcaaa tgggtcggga tctgtacgac 180
gatgacgata aggatcgatg gggatccgag ctcgagatct gcagctggta ccatggaatt 240
cgaagcttga tccggctgct aacaaagccc gaaaggaagc tgagttggct gctgccaccg 300
ctgagcaata actagcataa ccccttgggg cctctaaacg ggtcttgagg ggttttttgc 360
tgaaaggagg aactatatcc ggatctggcg taatagcgaa gaggcccgca ccgatcgccc 420
ttcccaacag ttgcgcagcc tgaatggcga atgggacgcg ccctgtagcg gcgcattaag 480
cgcggcgggt gtggtggtta cgcgcagcgt gaccgctaca cttgccagcg ccctagcgcc 540
cgctcctttc gctttcttcc cttcctttct cgccacgttc gccggctttc cccgtcaagc 600
tctaaatcgg gggctccctt tagggttccg atttagtgct ttacggcacc tcgaccccaa 660
aaaacttgat tagggtgatg gttcacgtag tgggccatcg ccctgataga cggtttttcg 720
ccctttgacg ttggagtcca cgttctttaa tagtggactc ttgttccaaa ctggaacaac 780
actcaaccct atcgcggtct attcttttga tttataaggg attttgccga tttcggccta 840
ttggttaaaa aatgagctga tttaacaaat atttaacgcg aattttaaca aaatattaac 900
gcttacaatt taggtggcac ttttcgggga aatgtgcgcg gaacccctat ttgtttattt 960
ttctaaatac attcaaatat gtatccgctc atgagacaat aaccctgata aatgcttcaa 1020
taatattgaa aaaggaagag tatgagtcat attcaacggg aaacgtcttg ctctaggccg 1080
cgattaaatt ccaacatgga tgctgattta tatgggtata aatgggctcg cgataatgtc 1140
gggcaatcag gtgcgacaat ctatcgattg tatgggaagc ccgatgcgcc agagttgttt 1200
ctgaaacatg gcaaaggtag cgttgccaat gatgttacag atgagatggt cagactaaac 1260
tggctgacgg aatttatgcc tcttccgacc atcaagcatt ttatccgtac tcctgatgat 1320
gcatggttac tcaccactgc gatccccggg aaaacagcat tccaggtatt agaagaatat 1380
cctgattcag gtgaaaatat tgttgatgcg ctggcagtgt tcctgcgccg gttgcattcg 1440
attcctgttt gtaattgtcc ttttaacagc gatcgcgtat ttcgtctcgc tcaggcgcaa 1500
tcacgaatga ataacggttt ggttgatgcg agtgattttg atgacgagcg taatggctgg 1560
cctgttgaac aagtctggaa agaaatgcat aaacttttgc cattctcacc ggattcagtc 1620
gtcactcatg gtgatttctc acttgataac cttatttttg acgaggggaa attaataggt 1680
tgtattgatg ttggacgagt cggaatcgca gaccgatacc aggatcttgc catcctatgg 1740
aactgcctcg gtgagttttc tccttcatta cagaaacggc tttttcaaaa atatggtatt 1800
gataatcctg atatgaataa attgcagttt catttgatgc tcgatgagtt tttctaactg 1860
tcagaccaag tttactcata tatactttag attgatttaa aacttcattt ttaatttaaa 1920
aggatctagg tgaagatcct ttttgataat ctcatgacca aaatccctta acgtgagttt 1980
tcgttccact gagcgtcaga ccccgtagaa aagatcaaag gatcttcttg agatcctttt 2040
tttctgcgcg taatctgctg cttgcaaaca aaaaaaccac cgctaccagc ggtggtttgt 2100
ttgccggatc aagagctacc aactcttttt ccgaaggtaa ctggcttcag cagagcgcag 2160
ataccaaata ctgtccttct agtgtagccg tagttaggcc accacttcaa gaactctgta 2220
gcaccgccta catacctcgc tctgctaatc ctgttaccag tggctgctgc cagtggcgat 2280
aagtcgtgtc ttaccgggtt ggactcaaga cgatagttac cggataaggc gcagcggtcg 2340
ggctgaacgg ggggttcgtg cacacagccc agcttggagc gaacgaccta caccgaactg 2400
agatacctac agcgtgagct atgagaaagc gccacgcttc ccgaagggag aaaggcggac 2460
aggtatccgg taagcggcag ggtcggaaca ggagagcgca cgagggagct tccaggggga 2520
aacgcctggt atctttatag tcctgtcggg tttcgccacc tctgacttga gcgtcgattt 2580
ttgtgatgct cgtcaggggg gcggagccta tggaaaaacg ccagcaacgc ggccttttta 2640
cggttcctgg gcttttgctg gccttttgct cacatgttct ttcctgcgtt atcccctgat 2700
tctgtggata accgtattac cgcctttgag tgagctgata ccgctcgccg cagccgaacg 2760
accgagcgca gcgagtcagt gagcgaggaa gcggaagagc gcccaatacg caaaccgcct 2820
ctccccgcgc gttggccgattcattaatgc ag 2852
<210>14
<211>34
<212>DNA
<213>Artificial Sequence
<220>
<223>外引物(outer primer for amplifying a gene encoding D-amino acid oxidase)
<400>14
tagggctgac atatggctaa aatcgttgtt attg 34
<210>15
<211>32
<212>DNA
<213>Artificial Sequence
<220>
<223>外引物(outer primer for amplifying a gene encoding D-amino acid oxidase)
<400>15
tagggctgaa gatctctaaa ggtttggacg ag 32
<210>16
<211>39
<212>DNA
<213>Artificial Sequence
<220>
<223>正向内引物(inner forward primer for introducing a mutant of amino acid residue from Thr to Pro
at position 53 of SEQ ID NO:2)
<400>16
gcaggtgcca actggctccc gttttacgat ggaggcaag 39
<210>17
<211>24
<212>DNA
<213>Artificial Sequence
<220>
<223>反向内引物(inner reverse primer)
<400>17
gagccagttg gcacctgccc aagg 24
<210>18
<211>39
<212>DNA
<213>Artificial Sequence
<220>
<223>内引物(inner forward primer for introducing a mutant of amino acid residue from Thr to Ser at
position 53 of SEQ ID NO:2)
<400>18
gcaggtgcca actggctcag cttttacgat ggaggcaag 39
Claims (11)
1.一种编码重组D-氨基酸氧化酶的DNA序列,其特征在于所述重组D-氨基酸氧化酶的基因与序列表中的序列1相比,编码第53位的苏氨酸的核苷酸密码子突变为编码丝氨酸或脯氨酸的密码子。
2.权利要求1所述的DNA序列,其特征在于所述重组D-氨基酸氧化酶的基因编码序列4所述的氨基酸序列。
3.权利要求2所述的DNA序列,其特征在于它包含序列3所示的核苷酸序列。
4.权利要求1所述的DNA序列,其特征在于所述重组D-氨基酸氧化酶的基因编码序列6所述的氨基酸序列。
5.权利要求5所述的DNA序列,其特征在于它包含序列5所示的核苷酸序列。
6.一种多肽,其氨基酸序列与由权利要求1所述的DNA编码的氨基酸序列一致。
7.权利要求6所述的多肽,其特征在于,以序列表中的序列2为参考序列,对应于序列2中第53位苏氨酸的氨基酸为脯氨酸。
8.权利要求7所述的多肽,其氨基酸序列如序列4所示。
9.权利要求6所述的多肽,其特征在于,以序列表中的序列2为参考序列,对应于序列2中第53位苏氨酸的氨基酸为丝氨酸。
10.权利要求9所述的多肽,其氨基酸序列如序列6所示。
11.一种质粒,其含有权利要求1-5中任一项所述的DNA序列。
Priority Applications (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CNB2004100308428A CN100342014C (zh) | 2004-04-08 | 2004-04-08 | 重组d-氨基酸氧化酶 |
| DE602004014824T DE602004014824D1 (de) | 2004-04-08 | 2004-04-12 | Rekombinante d-aminosäure-oxidase |
| US11/547,639 US7517677B2 (en) | 2004-04-08 | 2004-04-12 | Recombinant D-amino acid oxidases |
| EP04726873A EP1748073B1 (en) | 2004-04-08 | 2004-04-12 | Recombinant d-amino acid oxidase |
| AT04726873T ATE399865T1 (de) | 2004-04-08 | 2004-04-12 | Rekombinante d-aminosäure-oxidase |
| PCT/CN2004/000340 WO2005098000A1 (fr) | 2004-04-08 | 2004-04-12 | D-amino acide oxydase recombinante |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CNB2004100308428A CN100342014C (zh) | 2004-04-08 | 2004-04-08 | 重组d-氨基酸氧化酶 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CNB2004101031662A Division CN100386440C (zh) | 2004-04-08 | 2004-04-08 | 重组 d-氨基酸氧化酶的用途 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN1680558A CN1680558A (zh) | 2005-10-12 |
| CN100342014C true CN100342014C (zh) | 2007-10-10 |
Family
ID=35067316
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CNB2004100308428A Expired - Lifetime CN100342014C (zh) | 2004-04-08 | 2004-04-08 | 重组d-氨基酸氧化酶 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US7517677B2 (zh) |
| EP (1) | EP1748073B1 (zh) |
| CN (1) | CN100342014C (zh) |
| AT (1) | ATE399865T1 (zh) |
| DE (1) | DE602004014824D1 (zh) |
| WO (1) | WO2005098000A1 (zh) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN108840448A (zh) * | 2018-06-29 | 2018-11-20 | 博瑞威生物化工(沧州)有限公司 | 一种重组酵母菌处理含d型氨基酸废水的方法 |
Families Citing this family (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN1912127B (zh) * | 2005-08-08 | 2011-05-25 | 百瑞全球有限公司 | 一种大肠杆菌表达载体及其应用 |
| CN1912130B (zh) * | 2005-08-08 | 2011-06-15 | 百瑞全球有限公司 | 一种用于制备7-氨基头孢霉烷酸的两步酶法 |
| WO2007051355A1 (en) * | 2005-11-07 | 2007-05-10 | Bioright Worldwide Company Limited | The use of recombinant d-amino acid oxidase |
| CN1982445B (zh) | 2005-12-16 | 2011-07-27 | 百瑞全球有限公司 | 用于制备固定化酶或固定化细胞的载体以及使用该载体的固定化方法 |
| US20110059503A1 (en) * | 2007-03-08 | 2011-03-10 | Richmond Chemical Corporation | Compositions of variant biocatalysts for preparing enantiopure amino acids |
| RU2362806C2 (ru) * | 2007-07-23 | 2009-07-27 | Общество с ограниченной ответственностью "Инновации и высокие технологии МГУ" | Мутантные оксидазы d-аминокислот |
| CA2817224A1 (en) | 2010-11-19 | 2012-05-24 | Novozymes North America, Inc. | Processes of producing a fermentation product |
| RU2507262C1 (ru) * | 2012-09-25 | 2014-02-20 | Общество с ограниченной ответственностью "Инновации и высокие технологии МГУ" | Мутантная оксидаза d-аминокислот (варианты) |
| CN103710361B (zh) * | 2013-07-12 | 2015-07-15 | 广西大学 | 一种编码D-氨基酸氧化酶的基因daoE及其应用 |
| US10397388B2 (en) * | 2015-11-02 | 2019-08-27 | Hand Held Products, Inc. | Extended features for network communication |
| CN111019916B (zh) | 2018-11-23 | 2020-12-08 | 弈柯莱生物科技(上海)股份有限公司 | 一种d-氨基酸氧化酶突变体及其应用 |
| CN115896054A (zh) * | 2022-10-11 | 2023-04-04 | 湖南福来格生物技术有限公司 | 一种d-氨基酸氧化酶突变体及其应用 |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6635458B2 (en) * | 1997-09-25 | 2003-10-21 | Antibioticos, S.A.U. | Enzymatic process for the preparation of cephalosporanic 7$B(G)-(4-Carboxybutanamide) acid by means of the modified enzyme D-aminoacid oxidase of trigonopsis variabilis produced in escherichia coli |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS62262994A (ja) * | 1986-05-12 | 1987-11-16 | Asahi Chem Ind Co Ltd | D−アミノ酸オキシダ−ゼ遺伝子 |
| JPH07108225B2 (ja) * | 1986-09-16 | 1995-11-22 | 旭化成工業株式会社 | D−アミノ酸オキシダ−ゼ遺伝子 |
| CA2100987C (en) * | 1992-07-27 | 1999-06-15 | Kaoru Furuya | A transformant capable of producing d-amino acid oxidase |
| EP1344830A1 (en) * | 2002-03-12 | 2003-09-17 | Bioferma Murcia, S.A. | A process for the preparation of cephalosporan acid derivatives from cephalosporin C |
-
2004
- 2004-04-08 CN CNB2004100308428A patent/CN100342014C/zh not_active Expired - Lifetime
- 2004-04-12 AT AT04726873T patent/ATE399865T1/de not_active IP Right Cessation
- 2004-04-12 WO PCT/CN2004/000340 patent/WO2005098000A1/zh not_active Ceased
- 2004-04-12 US US11/547,639 patent/US7517677B2/en not_active Expired - Lifetime
- 2004-04-12 DE DE602004014824T patent/DE602004014824D1/de not_active Expired - Lifetime
- 2004-04-12 EP EP04726873A patent/EP1748073B1/en not_active Expired - Lifetime
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6635458B2 (en) * | 1997-09-25 | 2003-10-21 | Antibioticos, S.A.U. | Enzymatic process for the preparation of cephalosporanic 7$B(G)-(4-Carboxybutanamide) acid by means of the modified enzyme D-aminoacid oxidase of trigonopsis variabilis produced in escherichia coli |
Non-Patent Citations (2)
| Title |
|---|
| Role of Arginine 285 in the Active Site of Rhodotorula gracilisD-Amino Acid Oxidase Gianluca Molla,Davide Porrini,et al,THE JOURNAL OF BIOLOGICAL CHEMISTRY,Vol.275 No.32 2000 * |
| Substitution of the critical methionine residues in trigonopsisvariabilis D-amino acid oxidase with leucine enhances itsresistance to hydrogen peroxide. Ju SS,Lin LL,Chien HR,Hsu WH,FEMS Microbiol Lett,Vol.186 No.2 2000 * |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN108840448A (zh) * | 2018-06-29 | 2018-11-20 | 博瑞威生物化工(沧州)有限公司 | 一种重组酵母菌处理含d型氨基酸废水的方法 |
| CN108840448B (zh) * | 2018-06-29 | 2021-03-23 | 博瑞威生物化工(沧州)有限公司 | 一种重组酵母菌处理含d型氨基酸废水的方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| CN1680558A (zh) | 2005-10-12 |
| US7517677B2 (en) | 2009-04-14 |
| DE602004014824D1 (de) | 2008-08-14 |
| EP1748073B1 (en) | 2008-07-02 |
| WO2005098000A1 (fr) | 2005-10-20 |
| EP1748073A4 (en) | 2007-08-08 |
| ATE399865T1 (de) | 2008-07-15 |
| EP1748073A1 (en) | 2007-01-31 |
| US20080009052A1 (en) | 2008-01-10 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN100342014C (zh) | 重组d-氨基酸氧化酶 | |
| CN1177931C (zh) | Dna的合成方法 | |
| CN1209437A (zh) | 获得具有正确键合的胱氨酸键的胰岛素前体的改进的方法 | |
| CN1656225A (zh) | 新型羰基还原酶及其编码基因、以及利用它们制备光学活性醇的方法 | |
| CN101029311A (zh) | 过氧化氢酶基因及其用途 | |
| CN1294255C (zh) | 一种具有转化黄曲霉毒素活性的解毒酶及编码该酶的基因 | |
| CN1639337A (zh) | 具有催泪成分合成酶活性的蛋白质或多肽、编码该蛋白质或多肽的 DNA、利用该 DNA制造具有催泪成分合成酶活性的蛋白质或多肽的制造方法以及具有抑制该蛋白质或多肽的mRNA翻译的功能的核酸分子 | |
| CN101062954A (zh) | 具有抗血管生成作用的融合蛋白及其编码基因与应用 | |
| CN101074435A (zh) | α-半乳糖苷酶基因、其编码蛋白及其制备方法和应用 | |
| CN101045933A (zh) | 一种适冷蛋白酶的基因mcp01及其制备方法 | |
| CN1650006A (zh) | 葡萄糖脱氢酶β亚基及其编码DNA | |
| CN1680583A (zh) | 重组 d-氨基酸氧化酶的用途 | |
| CN101040047A (zh) | 编码新型的具有d-丝氨酸合成活性的酶的dna、该酶的制备方法、及利用该酶的d-丝氨酸制备方法 | |
| CN1497046A (zh) | α-酮酸还原酶及其制备方法,以及使用此酶制备光学活性的α-羟基酸的方法 | |
| CN1641019A (zh) | 头孢菌素c酰基转移酶 | |
| CN1517436A (zh) | 氧化还原酶 | |
| CN1257269C (zh) | 一种生产7-氨基头孢烷酸的方法及其专用重组酶 | |
| CN1141387C (zh) | ppGpp合成酶及用于改善目的蛋白产生的表达系统 | |
| CN1891820A (zh) | 在表面活性剂存在下稳定的胆固醇氧化酶 | |
| CN1119418C (zh) | 红冬孢属d-氨基酸氧化酶 | |
| CN1163604C (zh) | 减毒水痘病毒冈株的基因62和利用基因62的减毒水痘活疫苗用病毒株的鉴定方法 | |
| CN1613011A (zh) | 信号系统及其使用的元件 | |
| CN1948339A (zh) | 端粒酶活性抑制蛋白的制备和纯化 | |
| CN100344759C (zh) | 小麦脱水应答转录因子DREB基因cDNA序列 | |
| CN1216989C (zh) | 一种新的青霉素g酰化酶及其应用 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| C10 | Entry into substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| C14 | Grant of patent or utility model | ||
| GR01 | Patent grant | ||
| CX01 | Expiry of patent term | ||
| CX01 | Expiry of patent term |
Granted publication date: 20071010 |