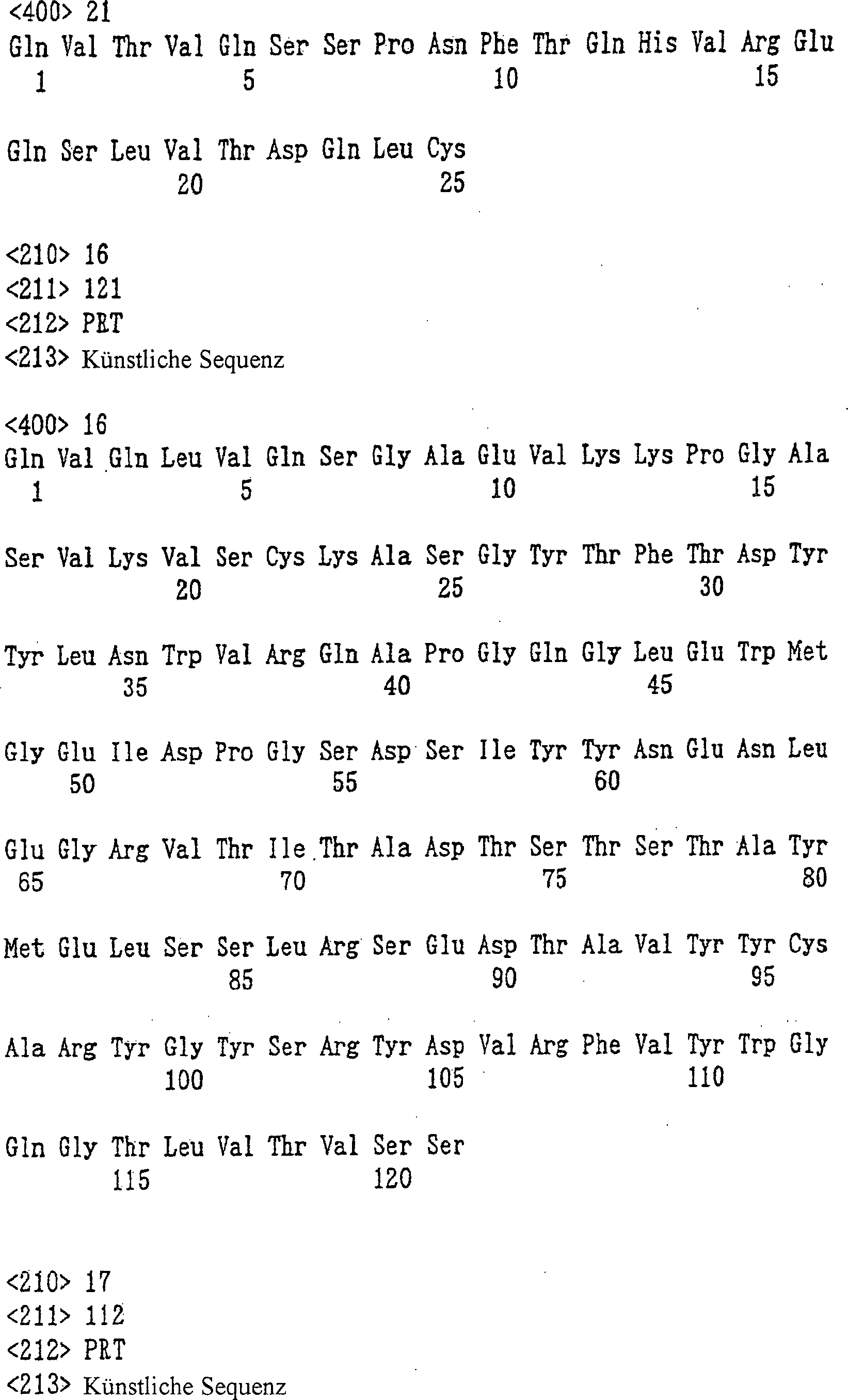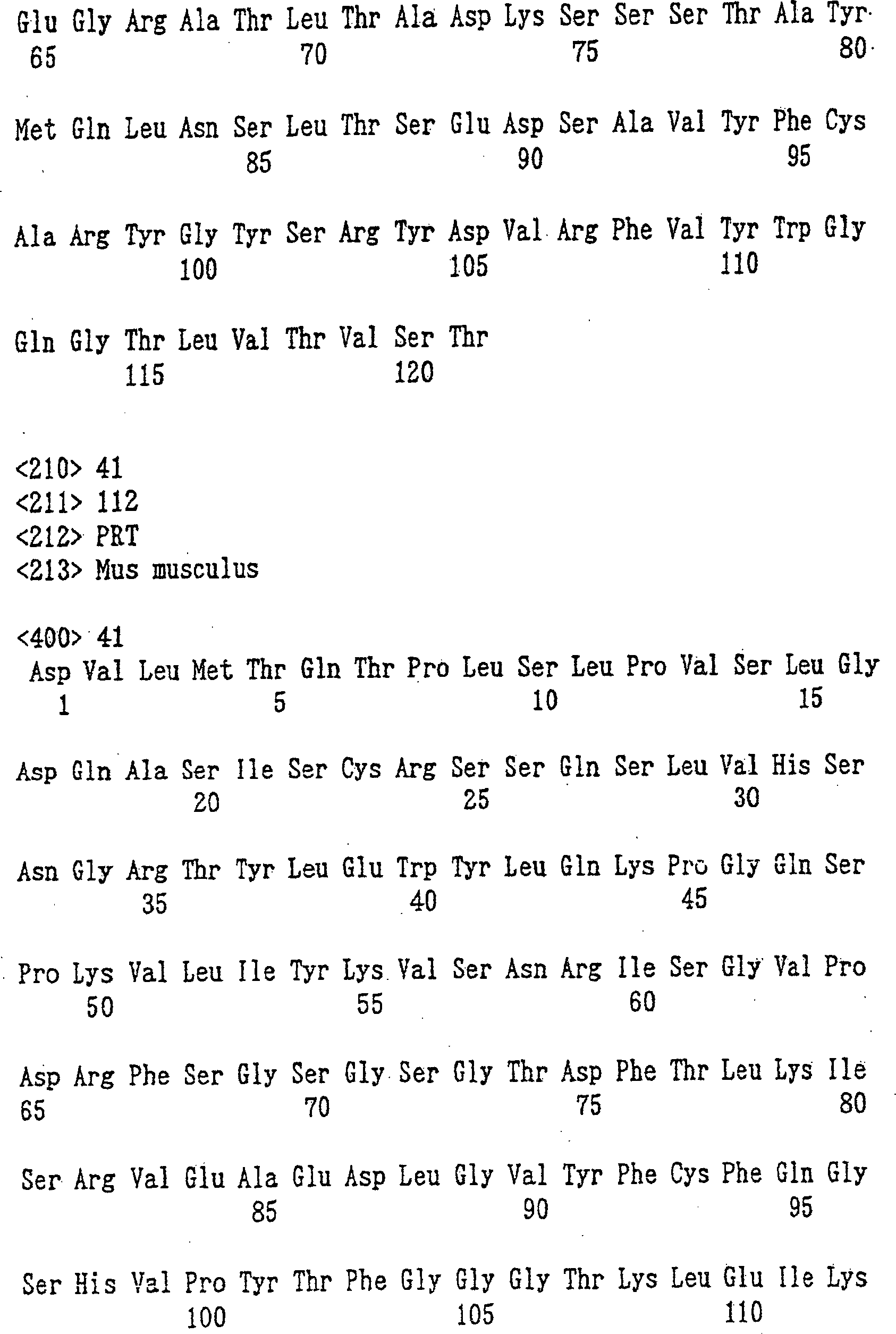-
Technischer Hintergrund
-
Die
vorliegende Erfindung betrifft humanisierte Antikörper gegen
FGF-8 und Antikörperfragmente
davon. Außerdem
betrifft die vorliegende Erfindung eine DNA-Sequenz, welche diese Antikörper oder
das Antikörperfragment
codiert. Die vorliegende Erfindung betrifft einen Vektor, umfassend
die DNA-Sequenz, sowie eine Wirtszelle, welche mit dem Vektor transformiert
ist. Ferner betrifft die vorliegende Erfindung ein Verfahren zur
Herstellung der vorstehenden Antikörper oder der Antikörperfragmente
unter Verwendung der Wirtszelle sowie ein therapeutisches Mittel
und ein diagnostisches Mittel für
Krebs unter Verwendung der Antikörper
oder der Antikörperfragmente.
-
Hintergrund des Fachgebiets
-
Der
durch ein Androgen induzierte Wachstumsfaktor (AIGF) ist ein Faktor,
welcher in 1992 aus einem Kulturüberstand
der Maus-Brusttumorzelllinie SC-3 [J. Steroid Biochem. 27 (1987),
459], die ein Geschlechtshormon-abhängiges Wachstum zeigt, isoliert
wurde. Der AIGF ist ein Wachstumsfaktor, welcher unter Androgenstimulation
induziert und hergestellt wird und das Wachstum von SC-3-Zellen
in einer autokrinen Weise aktiviert [Proc. Natl. Acad. Sci. 89 (1992),
8928–8932].
Als Ergebnis einer Genclonierung wurde festgestellt, dass er in
Bezug auf die Aminosäuresequenz
eine Homologie von 30% bis 40% mit der Fibroblastenwachstumsfaktor
(FGF)-Familie aufweist, und er wurde als FGF-8 bezeichnet. Danach
wurde das menschliche FGF-8-Gen unter Verwendung eines Maus-FGF-8-Gens
als Sonde aus einer menschlichen Plazenta-Genombank cloniert, und
es wurde gezeigt, dass es mit dem Maus-FGF-8 zu 85% hinsichtlich
der Nucleotidsequenz und zu 100% hinsichtlich der Aminosäuresequenz übereinstimmte
[FEBS Letters 363 (1995), 226]. Es ist angenommen worden, dass Geschlechtshormon-induzierte
Wachstumsfaktoren bei Krebserkrankungen wie Prostatakrebs und Brustkrebs,
die ein Geschlechtshormon- abhängiges Wachstum
zeigen, eine autokrine Funktion haben, und die Entdeckung von FGF-8,
obgleich in einem Maussystem, war der erste Beweis für einen
solchen Mechanismus. Es ist vermutet worden, dass FGF-8 durch einen ähnlichen
Mechanismus auch mit der Entstehung und dem Wachstum von Krebs beim
Menschen in Beziehung steht, aber dies ist nicht bewiesen worden.
Jedoch wurde berichtet, dass das Wachstum einer bestimmten menschlichen
Prostatakrebszelllinie durch die Zugabe von FGF-8 begünstigt wurde
[FEBS Letters 363 (1995), 226]. Die Expression der mRNA wurde auch
in verschiedenen menschlichen Krebszelllinien wie Prostatakrebs,
Brustkrebs und Eierstockkrebs bestätigt [Cell Growth & Differ. 7 (1996),
1425; Oncogene 18 (1999), 1053; Int. J. Cancer 88 (2000), 718].
Es wurde auch gezeigt, dass der in menschlichen Krebsgeweben wie
Prostatakrebs, Brustkrebs und Eierstockkrebs vorkommende FGF-8 im
Vergleich zu dem FGF-8 in normalen Geweben überexprimiert wurde [Cancer
Res. 58, 2053; Oncogene 18 (1999), 2755; Oncogene 18 (1999), 1053;
Int. J. Cancer 88 (2000), 718]. Daher wurde von der Möglichkeit
ausgegangen, dass FGF-8 in Bezug auf das Geschlechtshormon-abhängige Wachstum
von Krebszellen beim Menschen auch eine autokrine und eine parakrine
Funktion hat. Andererseits, da auch bei verschiedenen hormonunabhängigen menschlichen
Prostatakrebszellen, Prostatakrebsgeweben und Brustkrebszellen eine
Expression von FGF-8 mit einer hohen Häufigkeit beobachtet worden
ist [Cell Growth & Differ. 7
(1996), 1425; Oncogene 18 (1999), 2755; Oncogene 18 (1999), 1053],
besteht eine hohe Wahrscheinlichkeit, dass die Expression von FGF-8
auf eine Geschlechtshormon-unabhängige
Weise kontrolliert wird. Außerdem, da
es einen Bericht gibt, dass eine Antisense-DNA gegen FGF-8 das in-vitro-
und in-vivo-Wachstum einer hormonunabhängigen Prostatakrebszelllinie
hemmte [Oncogene 16 (1998), 1487], wird auch das Vorhandensein eines
FGF-8-abhängigen Wachstumsmechanismus
für Krebsarten
angenommen, welche ihre Hormonabhängigkeit verloren haben.
-
Basierend
auf diesen Fakten sind Antikörper
gegen FGF-8 bei der Untersuchung der biologischen Funktion von FGF-8
für Krebszellen
und auch bei der Diagnose von Krebszellen wie Prostatakrebs und
Brustkrebs durch ein immunologisches Nachweisverfahren nützlich.
Außerdem
wird bei neutralisierenden Antikörpern,
welche die Funktionen von FGF-8 inhibieren, erwartet, dass sie bei
der Untersuchung der biologischen Funktion von FGF-8, der Diagnose
von Krebserkrankungen wie Prostatakrebs, Brustkrebs und Eierstockkrebs sowie
bei der Behandlung von Geschlechtshormon-abhängigen Krebserkrankungen und
Geschlechtshormon-unabhängigen
Krebserkrankungen nützlich
sind. Daher betrieben die Erfinder der vorliegenden Erfindung die
Herstellung eines Antikörpers
gegen FGF-8, wobei ihnen als Ergebnis die Herstellung des monoclonalen Maus-Antikörpers KM1334
gelang, der mit FGF-8 spezifisch reagiert und die Funktion von FGF-8
inhibiert (ungeprüfte
veröffentlichte
japanische Patentanmeldung Nr.
271391/97 ;
EP-A2 799 835 ).
-
Es
ist allgemein bekannt, dass, wenn ein Antikörper, der von einem nicht-menschlichen Tier
stammt, z. B. ein Maus-Antikörper,
an einen Menschen verabreicht wird, dieser Antikörper als eine fremde Substanz erkannt
wird und einen menschlichen Antikörper gegen den Maus-Antikörper (menschlichen
Anti-Maus-Antikörper;
nachstehend bezeichnet als "HAMA") in dem menschlichen
Körper
induziert. Es ist bekannt, dass der HAMA mit dem verabreichten Mausantikörper reagiert,
wodurch Nebenwirkungen hervorgerufen werden [J. Clin. Oncol. 2 (1984),
881; Blood 65 (1985), 1349; J. Natl. Cancer Inst. 80 (1988), 932;
Proc. Natl. Acad. Sci. USA 82 (1985), 1242], die Eliminierung des
verabreichten Mausantikörpers
aus dem Körper
beschleunigt wird [J. Nucl. Med. 26 (1985), 1011; Blood 65 (1985),
1349; J. Natl. Cancer Inst. 80 (1988), 937] und die therapeutische
Wirkungen des Mausantikörpers
vermindert werden [J. Immunol. 135 (1985), 1530; Cancer Res. 46 (1986),
6489].
-
Um
diese Probleme zu lösen,
sind Versuche unternommen worden, um einen Antikörper, der von einem nicht-menschlichen
Tier stammt, mit Hilfe der Gentechnik in einen humanisierten Antikörper wie
einen menschlichen chimären
Antikörper
oder einen menschlichen CDR-transplantierten Antikörper („CDR-grafted antibody”) umzuwandeln.
-
Der
menschliche chimäre
Antikörper
besteht aus einem Antikörper,
wobei die V-Region von einem Antikörper eines nicht-menschlichen
Tieres abgeleitet ist und die C-Region von einem menschlichen Antikörper stammt
[Proc. Natl. Acad. Sci. USA 81 (1984), 6851]. Der menschliche CDR-transplantierte
Antikörper
umfasst die Aminosäuresequenz
einer CDR-Region in der V-Region, welche von einem Antikörper eines
nicht-menschlichen Tieres stammt, transplantiert in eine geeignete
Position eines menschlichen Antikörpers [Nature 321 (1986), 522].
Im Vergleich zu Antikörpern,
welche von nicht-menschlichen Tieren stammen, wie Mausantikörper und
dergleichen, weisen diese humanisierten Antikörper verschiedene Vorteile
für die
klinische Anwendung auf. Zum Beispiel ist im Hinblick auf die Immunogenität und die
Stabilität
im Blut berichtet worden, dass sich die Halbwertszeit im Blut eines
menschlichen chimären
Antikörpers
bei der Verabreichung an einen Menschen um etwa das sechsfache der
Halbwertszeit eines Mausantikörpers
verlängert
[Proc. Natl. Acad. Sci. USA 86 (1989), 4220]. Im Falle eines menschlichen
CDR-transplantierten Antikörpers
ist berichtet worden, dass dessen Immunogenität und dessen Halbwertszeit
im Serum in einem Experiment unter Verwendung eines Affens im Vergleich
zu einem Mausantikörper
geringer war bzw. verlängert
wurde [J. Immunol. 147 (1991), 1352]. Es wird daher erwartet, dass
die humanisierten Antikörper
weniger Nebenwirkungen haben und dass ihre therapeutischen Wirkungen über einen
längeren
Zeitraum anhalten als bei Antikörpern,
die von nicht-menschlichen Tieren stammen.
-
Außerdem,
da der menschliche CDR-transplantierte Antikörper mit Hilfe der Gentechnik
erzeugt wird, können
Moleküle
in verschiedenen Formen hergestellt werden. Wenn zum Beispiel die
Subklasse 1 als eine H-Ketten-C-Region (CH) eines menschlichen Antikörpers verwendet
wird, kann ein humanisierter Antikörper mit einer starken Effektorfunktion
wie zum Beispiel einer Antikörper-abhängigen zellvermittelten
cytotoxischen Aktivität
hergestellt werden [Cancer Res. 56 (1996), 1118]. Ein humanisierter
Antikörper
mit einer starken Effektorfunktion ist besonders wirksam, wenn ein
Antigen auf der Oberfläche
von Zellen wie Krebszellen vorhanden ist und die Zerstörung der
Zielzelle gewünscht
wird. Andererseits, falls nur eine Funktion erforderlich ist, wodurch
das Zielmolekül
neutralisiert wird, oder falls Nebenwirkungen durch die Zerstörung der
Zielzelle hervorgerufen werden könnten,
wird erwartet, dass diese Nebenwirkungen vermieden werden können und
eine längere
Halbwertszeit im Blutserum im Vergleich zu einem Mausantikörper erhalten
werden kann, wenn die Subklasse 4 als die CH-Region des menschlichen
Antikörpers
verwendet wird [Immunol. 85 (1995), 668], da die Subklasse 4 üblicherweise
eine geringe Effektorfunktion hat [J. Exp. Med. 166 (1987), 1351;
J. Exp. Med. 168 (1998), 1351]. Außerdem können im Einklang mit den jüngsten Fortschritten
im Hinblick auf die Proteinkonstruktion und die Genkonstruktion
Antikörperfragmente
mit einem geringeren Molekulargewicht wie Fab, Fab', F(ab')2,
scFv [Science 242 (1988), 423], ein Diabody [Nature Biotechnol.
15 (1997), 629], dsFv [Molecular Immunol. 32 (1995), 249] und ein
Peptid, umfassend eine CDR-Region [J. Biol. Chem. 271 (1996), 2966],
als humanisierte Antikörper
hergestellt werden. Die Antikörperfragmente
zeigen eine überragende
vorübergehende
Aktivität
in dem Zielgewebe, verglichen mit den vollständigen Antikörpermolekülen [Cancer
Res. 52 (1992), 3402].
-
Es
wird davon ausgegangen, dass die humanisierten Antikörper im
Hinblick auf die klinische Anwendung beim Menschen wünschenswerter
sind als Antikörper,
welche von nicht-menschlichen Tieren stammen, wie Maus-Antikörper.
-
Wie
vorstehend besprochen, wird erwartet, dass, wenn humanisierte Antikörper, welche
mit FGF-8 spezifisch reagieren und die Funktion von FGF-8 inhibieren,
oder Antikörperfragmente
davon hergestellt werden, diese Antikörper und Antikörperfragmente
als wirksame diagnostische Mittel oder therapeutische Mittel für Erkrankungen,
welche mit FGF-8 in Beziehung stehen, wie Krebs, verwendet werden
können.
Jedoch sind solche Antikörper
oder Antikörperfragmente
bis jetzt noch nicht hergestellt worden.
-
Offenbarung der Erfindung
-
Eine
Aufgabe der vorliegenden Erfindung ist die Bereitstellung der humanisierten
Antikörper
oder deren Fragmente, wie in den Ansprüchen gekennzeichnet, welche
mit FGF-8, von dem angenommen wird, dass er ein Wachstumsfaktor
von Prostatakrebs, Brustkrebs, Eierstockkrebs und dergleichen ist,
spezifisch reagieren und welche weiterhin die biologische Funktion
von FGF-8 inhibieren.
-
Die
Erfinder der vorliegenden Erfindung, welche die cDNA der H-Kette
und die cDNA der L-Kette des Antikörpers aus dem Hybridom KM1334
(FERM BP-5451), das einen Mausantikörper gegen FGF-8, welcher der
IgG1-Subklasse angehört,
produziert, gewonnen haben, stellten fest, dass die V-Regionen und
die CDR-Regionen neue Aminosäuresequenzen
sind, und exprimierten und reinigten einen menschlichen chimären Anti-FGF-8-Antikörper und
einen menschlichen CDR-transplantierten Anti-FGF-8-Antikörper durch
das Clonieren der cDNAs, welche die neuen V-Regionen oder die VH-Region
und die VL-Region, umfassend die CDRs, codieren, in einen Expressionsvektor
für eine
tierische Zelle, der die cDNAs umfasst, welche die CH-Region eines menschlichen
Antikörpers
und die L-Ketten-C-Region (CL) eines menschlichen Antikörpers codieren,
um auf diese Weise einen Vektor für die Expression eines humanisierten
Antikörpers
zu konstruieren, und dann durch das Einbringen des Vektors für die Expression
des Antikörpers
in eine tierische Zelle. Auf diese Weise ist die vorliegende Erfindung
durch die Bereitstellung der in den Ansprüchen gekennzeichneten Ausführungsformen
vollendet worden.
-
Folglich
betrifft die vorliegende Erfindung humanisierte Antikörper, die
mit FGF-8 spezifisch reagieren und welche die biologische Funktion
von FGF-8 inhibieren, oder Antikörperfragmente
davon.
-
Die
biologische Funktion von FGF-8 beinhaltet eine Geschlechtshormonabhängige und
-unabhängige Aktivität bei der
Wachstumsbeschleunigung von verschiedenen Krebszellen wie Prostatakrebs,
Brustkrebs und Eierstockkrebs [Proc. Natl. Acad. Sci. USA 89 (1992),
8928], eine Funktion bei der Vaskularisierung [Oncogene 20 (2001),
2791] und eine Funktion bei der Entstehung von Arthrose [Development
127 (2000), 2471; Nature Genet. 26 (2000), 460].
-
Ein
menschlicher chimärer
Antikörper
ist ein Antikörper,
der die VH-Region und die VL-Region eines Antikörpers, welcher von einem nicht-menschlichen
Tier stammt, und die CH-Region und die CL-Region eines menschlichen
Antikörpers
umfasst. Das nicht-menschliche Tier kann ein beliebiges Tier wie
eine Maus, eine Ratte, ein Hamster, ein Kaninchen und dergleichen
sein, solange wie daraus ein Hybridom erzeugt werden kann.
-
Der
menschliche chimäre
Antikörper
der vorliegenden Erfindung kann durch das Erhalten der cDNAs, welche
die VH-Region und die VL-Region codieren, aus einem Hybridom, das
einen monoclonalen Antikörper herstellt,
welcher mit FGF-8 spezifisch reagiert, das Inserieren der cDNAs
in einen Expressionsvektor für
eine tierische Zelle, enthaltend die Gene, welche die CH-Region
eines menschlichen Antikörpers
und die CL-Region eines menschlichen Antikörpers codieren, um einen Vektor
für die
Expression des menschlichen chimären Antikörpers zu
konstruieren, und das Einbringen des Vektors in eine tierische Zelle,
um den Antikörper
zu exprimieren, hergestellt werden.
-
Es
kann irgendeine CH-Region eines menschlichen chimären Antikörpers verwendet
werden, solange wie sie von einem menschlichen Immunglobulin (hlg)
stammt, wobei solche der hlgG-Klasse bevorzugt werden, und weiterhin
kann irgendeine der Subklassen von hlgG wie γ1, γ2, γ3 und γ4 verwendet werden. Ebenso kann
irgendeine CL-Region eines menschlichen chimären Antikörpers verwendet werden, solange
wie sie von einem hlg stammt, wobei solche der κ-Kiasse oder λ-Klasse verwendet werden können.
-
Der
menschliche chimäre
Antikörper,
welcher mit FGF-8 spezifisch reagiert (chimärer Anti-FGF-8-Antikörper), umfasst
einen menschlichen chimären
Antikörper,
welcher die komplementaritätsbestimmende
Region (nachstehend bezeichnet als "CDR")
1, CDR2 und CDR3 der variablen Region (V-Region) der schweren Kette
(H-Kette) eines Antikörpers,
umfassend die in SEQ ID NO: 5, 6 bzw. 7 dargestellte Aminosäuresequenzen,
umfasst und/oder CDR1, CDR2 und CDR3 der variablen Region (V-Region)
der leichten Kette (L-Kette) eines Antikörpers, umfassend die in SEQ
ID NO: 8, 9 bzw. 10 dargestellte Aminosäuresequenzen, umfasst, und
vorzugsweise einen menschlichen chimären Antikörper, wobei die VH-Region die
in SEQ ID NO: 40 dargestellte Aminosäuresequenz umfasst und/oder
die VL-Region die in SEQ ID NO: 41 dargestellte Aminosäuresequenz
umfasst. Beispiele schließen
den Antikörper
KM3034 ein, wobei die VH-Region des Antikörpers die in SEQ ID NO: 40
dargestellte Aminosäuresequenz
umfasst, die CH-Region des menschlichen Antikörpers eine Aminosäuresequenz
der Subklasse 71 umfasst, die VL-Region des Antikörpers die
in SEQ ID NO: 41 dargestellte Aminosäuresequenz umfasst, und die
CL-Region des menschlichen
Antikörpers
eine Aminosäuresequenz
der κ-Klasse
umfasst.
-
Ein
menschlicher CDR-transplantierter Antikörper ist ein Antikörper, wobei
die CDR-Aminosäuresequenzen
der VH-Region und der VL-Region eines Antikörpers, welcher von einem nicht-menschlichen
Tier stammt, in geeignete Positionen der VH-Region und der VL-Region
eines menschlichen Antikörpers
transplantiert sind.
-
Der
menschliche CDR-transplantierte Antikörper der vorliegenden Erfindung
kann durch das Transplantieren der CDR-Sequenzen der VH-Region und
der VL-Region eines
Antikörpers,
welcher mit FGF-8 spezifisch reagiert, eines nicht-menschlichen
Tiers in FR-Fragmente der VH-Region und der VL-Region eines beliebigen
menschlichen Antikörpers,
um cDNAs zu konstruieren, die die erhaltenen V-Regionen codieren,
das Inserieren der cDNAs in einen Expressionsvektor für eine tierische
Zelle, enthaltend die DNAs, welche die CH-Region und die CL-Region
eines menschlichen Antikörpers
codieren, um einen Vektor für
die Expression des menschlichen CDR-transplantierten Antikörpers zu
konstruieren, und dann das Einbringen davon in eine tierische Zelle,
um den Antikörper
zu exprimieren, hergestellt werden.
-
Es
kann irgendeine CH-Region eines menschlichen CDR-transplantierten
Antikörpers
verwendet werden, solange wie sie von einem hIg stammt, wobei solche
der hIgG-Klasse bevorzugt werden, und irgendeine der Subklassen
von hIgG wie 71, γ2, γ3 und γ4 kann verwendet
werden. Ebenso kann irgendeine CL-Region eines menschlichen CDR-transplantierten
Antikörpers
verwendet werden, solange wie sie von einem hIg stammt, wobei solche
der κ-Klasse
oder λ-Klasse
verwendet werden können.
-
Der
menschliche CDR-transplantierte Antikörper, welcher mit FGF-8 spezifisch
reagiert (Anti-FGF-8-CDR-transplantierter Antikörper) schließt einen
menschlichen CDR-transplantierten Antikörper ein, welcher CDR1, CDR2
und CDR3 der VH-Region
eines Antikörpers,
umfassend die in SEQ ID NO: 5, 6 bzw. 7 dargestellten Aminosäuresequenzen,
umfasst, und/oder CDR1, CDR2 und CDR3 der VL-Region eines Antikörpers, umfassend
die in SEQ ID NO: 8, 9 bzw. 10 dargestellten Aminosäuresequenzen,
umfasst; vorzugsweise einen menschlichen CDR-transplantierten Antikörper, wobei
die VH-Region die in SEQ ID NO: 16 dargestellte Aminosäuresequenz
umfasst und/oder die VL-Region die in SEQ ID NO: 17 dargestellte
Aminosäuresequenz
umfasst; und mehr bevorzugt einen menschlichen CDR-transplantierten
Antikörper,
wobei die VH-Region des Antikörpers
eine Aminosäuresequenz
umfasst, worin mindestens ein Aminosäurerest, ausgewählt aus
Lys in Position 12, Lys in Position 13, Ala in Position 40, Pro
in Position 41, Met in Position 48, Val in Position 68, Ile in Position
70, Thr in Position 74, Thr in Position 76, Glu in Position 82,
Ser in Position 84, Arg in Position 87 und Tyr in Position 95, in
der in SEQ ID NO: 16 dargestellten Aminosäuresequenz durch einen anderen
Aminosäurerest
substituiert ist; sowie einen menschlichen CDR-transplantierten
Antikörper,
wobei die VL-Region
des Antikörpers
eine Aminosäuresequenz
umfasst, worin mindestens ein Aminosäurerest, ausgewählt aus
Ile in Position 2, Val in Position 3, Thr in Position 14, Pro in
Position 15, Gln in Position 50, Leu in Position 51 und Tyr in Position
92, in der in SEQ ID NO: 17 dargestellten Aminosäuresequenz durch einen anderen
Aminosäurerest
substituiert ist; und einen menschlichen CDR-transplantierten Antikörper, wobei
die VH-Region des Antikörpers
eine Aminosäuresequenz
umfasst, worin mindestens ein Aminosäurerest, ausgewählt aus
Lys in Position 12, Lys in Position 13, Ala in Position 40, Pro
in Position 41, Met in Position 48, Val in Position 68, Ile in Position
70, Thr in Position 74, Thr in Position 76, Glu in Position 82,
Ser in Position 84, Arg in Position 87 und Tyr in Position 95, in
der in SEQ ID NO: 16 dargestellten Aminosäuresequenz durch einen anderen
Aminosäurerest
substituiert ist, und wobei die VL-Region des Antikörpers eine
Aminosäuresequenz
umfasst, worin mindestens ein Aminosäurerest, ausgewählt aus
Ile in Position 2, Val in Position 3, Thr in Position 14, Pro in
Position 15, Gln in Position 50, Leu in Position 51 und Tyr in Position
92, in der in SEQ ID NO: 17 dargestellten Aminosäuresequenz durch eine andere
Aminosäure
substituiert ist. Beispiele für
die VH-Region schließen
die in SEQ ID NO: 18 dargestellte Aminosäuresequenz ein, wobei sechs
Reste, umfassend Lys in Position 12, Lys in Position 13, Ala in
Position 40, Pro in Position 41, Met in Position 48 und Tyr in Position
95, in der in SEQ ID NO: 16 dargestellten Aminosäuresequenz durch Ala, Arg,
Arg, Ser, Ile bzw. Phe modifiziert sind. Beispiele für die VL-Region
schließen
die in SEQ ID NO: 19 dargestellte Aminosäuresequenz ein, wobei sechs
Reste, umfassend Ile in Position 2, Thr in Position 14, Pro in Position
15, Gln in Position 50, Leu in Position 51 und Tyr in Position 92,
in der in SEQ ID NO: 17 dargestellten Aminosäuresequenz durch Val, Ser,
Leu, Lys, Val bzw. Phe modifiziert sind; sowie die in SEQ ID NO:
38 dargestellte Aminosäuresequenz,
wobei vier Reste, umfassend Thr in Position 14, Pro in Position
15, Leu in Position 51 und Tyr in Position 92, durch Ser, Leu, Val
bzw. Phe modifiziert sind; und die in SEQ ID NO: 39 dargestellte
Aminosäuresequenz,
wobei drei Reste, umfassend Ile in Position 2, Leu in Position 51
und Tyr in Position 92, durch Val, Val bzw. Phe modifiziert sind.
-
Beispiele
umfassen einen menschlichen CDR-transplantierten Antikörper, wobei
die VH-Region des Antikörpers
die in SEQ ID NO: 16 oder 18 dargestellte Aminosäuresequenz umfasst; einen menschlichen CDR-transplantierten
Antikörper,
wobei die VL-Region des Antikörpers
die in SEQ ID NO: 17, 19, 38 oder 39 dargestellte Aminosäuresequenz
umfasst; einen menschlichen CDR-transplantierten Antikörper, wobei
die VH-Region und die VL-Region des Antikörpers die in SEQ ID NO: 16
oder 18 bzw. in SEQ ID NO: 17, 19, 38 oder 39 dargestellten Aminosäuresequenzen
umfassen; und ähnliche
und vorzugsweise einen menschlichen CDR-transplantierten Antikörper, wobei
die VH-Region und die VL-Region des Antikörpers die in SEQ ID NO: 16
bzw. in SEQ ID NO: 19 dargestellte Aminosäuresequenz umfassen; einen
menschlichen CDR-transplantierten Antikörper, wobei die VH-Region und die VL-Region
des Antikörpers
die in SEQ ID NO: 16 bzw. in SEQ ID NO: 38 dargestellte Aminosäuresequenz
umfassen; und einen menschlichen CDR-transpiantierten Antikörper, wobei
die VH-Region und die VL-Region des Antikörpers die in SEQ ID NO: 16
bzw. in SEQ ID NO: 39 dargestellte Aminosäuresequenz umfassen.
-
Das
Antikörperfragment
der vorliegenden Erfindung schließt Fab, Fab', F(ab')2, scFv, einen
Diabody, dsFv, ein Peptid, das eine CDR-Region umfasst, und dergleichen
ein.
-
Ein
Fab-Fragment ist ein Antikörperfragment
mit einem Molekulargewicht von etwa 50.000 und einer Antigenbindungsaktivität, wobei
die Fragmente, welche durch die Behandlung von IgG mit der Protease
Papain (die an dem Aminosäurerest
in Position 224 in der H-Kette schneidet) erhalten werden, umfassend
ungefähr die
Hälfte
der N-terminalen Seite der H-Kette und die gesamte L-Kette, über eine
Disulfidbrücke
(S-S-Bindung) miteinander verbunden sind.
-
Das
Fab-Fragment der vorliegenden Erfindung kann durch das Behandeln
eines menschlichen CDR-transplantierten Antikörpers der vorliegenden Erfindung,
welcher mit FGF-8 spezifisch reagiert, mit der Protease Papain erhalten
werden. Das Fab-Fragment kann auch durch die Insertion einer DNA,
welche das Fab-Fragment des Antikörpers codiert, in einen Expressionsvektor
für einen
Prokaryonten oder einen Expressionsvektor für einen Eukaryonten und das
Einbringen des Vektors in einen Prokaryonten oder einen Eukaryonten,
um das Fab-Fragment zu exprimieren, hergestellt werden.
-
Ein
F(ab')2-Fragment
ist ein Antikörperfragment
mit einem Molekulargewicht von etwa 100.000 und einer Antigenbindungsaktivität, welches
etwas größer als
das Fab-Fragment ist, wobei die Fragmente, welche durch die Behandlung
von IgG mit der Protease Pepsin (welche an dem Aminosäurerest
in Position 234 in der H-Kette schneidet) erhalten werden, über eine
S-S-Bindung in der Gelenkregion miteinander verbunden sind.
-
Das
F(ab')2-Fragment
der vorliegenden Erfindung kann durch das Behandeln eines Antikörpers, welcher
mit FGF-8 spezifisch reagiert, mit der Protease Pepsin erhalten
werden. Das F(ab')2-Fragment kann auch durch die Verknüpfung von
Fab'-Fragmenten,
welche nachstehend beschrieben werden, über eine Thioetherbindung oder
eine S-S-Bindung erzeugt werden.
-
Ein
Fab'-Fragment ist
ein Antikörperfragment
mit einem Molekulargewicht von etwa 50.000 und einer Antigenbindungsaktivität, welches
durch die Spaltung einer S-S-Bindung in der Gelenkregion des vorstehenden
F(ab')2-Fragments
erhalten wird.
-
Das
Fab'-Fragment der
vorliegenden Erfindung kann durch die Behandlung des F(ab')2-Fragments, welches
mit FGF-8 spezifisch reagiert, mit dem Reduktionsmittel Dithiothreit
erhalten werden. Das Fab'-Fragment
kann auch durch die Insertion einer DNA, welche das Fab'-Fragment des Antikörpers codiert,
in einen Expressionsvektor für
einen Prokaryonten oder einen Expressionsvektor für einen
Eukaryonten und das Einbringen des Vektors in einen Prokaryonten
oder einen Eukaryonten, um das Fab'-Fragment zu exprimieren, hergestellt
werden.
-
Ein
scFv-Fragment ist ein VH-P-VL- oder VL-P-VH-Polypeptid, worin eine
VH-Kette und eine VL-Kette unter Verwendung eines geeigneten Peptidlinkers
(P) aus 12 oder mehr Resten miteinander verbunden sind, welches
eine Antigenbindungsaktivität
besitzt.
-
Das
scFv-Fragment der vorliegenden Erfindung kann durch Erhalten der
cDNAs, codierend die VH-Region und die VL-Region eines Antikörpers, der
mit FGF-8 spezifisch reagiert, das Konstruieren einer DNA, welche
das scFv-Fragment codiert, die Insertion der DNA in einen Expressionsvektor
für einen
Prokaryonten oder einen Expressionsvektor für einen Eukaryonten und dann
das Einbringen des Expressionsvektors in einen Prokaryonten oder
einen Eukaryonten, um das scFv-Fragment
zu exprimieren, hergestellt werden.
-
Ein
Diabody ist ein Antikörperfragment,
worin scFv-Fragmente, welche die gleiche oder eine unterschiedliche
Antigenbindungsspezifität
haben, ein Dimer bilden, wobei das Fragment eine divalente Antigenbindungsaktivität gegen
das gleiche Antigen oder zwei spezifische Antigenbindungsaktivitäten gegen
verschiedene Antigene besitzt.
-
Der
Diabody der vorliegenden Erfindung, zum Beispiel ein divalenter
Diabody, welcher mit FGF-8 spezifisch reagiert, kann durch Erhalten
der cDNAs, codierend die VH-Region und die VL-Region eines Antikörpers, das
Konstruieren einer DNA, welche ein scFv-Fragment mit einem Polypeptidlinker
aus 3 bis 10 Resten codiert, die Insertion der DNA in einen Expressionsvektor
für einen
Prokaryonten oder einen Expressionsvektor für einen Eukaryonten und dann
das Einbringen des Expressionsvektors in einen Prokaryonten oder
einen Eukaryonten, um den Diabody zu exprimieren, hergestellt werden.
-
Ein
dsFv-Fragment wird durch die Verknüpfung von Polypeptiden, worin
jeweils ein Aminosäurerest
in der VH-Region und der VL-Region durch einen Cysteinrest substituiert
ist, über
eine S-S-Bindung zwischen den Cysteinresten erhalten. Der Aminosäurerest,
welcher durch einen Cysteinrest substituiert wird, kann basierend
auf einer dreidimensionalen Strukturannahme des Antikörpers gemäß der durch
Reiter et al. [Protein Engineering 7 (1994), 697] vorgestellten
Methode ausgewählt
werden.
-
Das
dsFv-Fragment der vorliegenden Erfindung kann durch Erhalten der
cDNAs, codierend die VH-Region und die VL-Region eines Antikörpers, der
mit FGF-8 spezifisch reagiert, das Konstruieren einer DNA, welche
das dsFv-Fragment codiert, die Insertion der DNA in einen Expressionsvektor
für einen
Prokaryonten oder einen Expressionsvektor für einen Eukaryonten und dann
das Einbringen des Expressionsvektors in einen Prokaryonten oder
einen Eukaryonten, um das dsFv-Fragment
zu exprimieren, hergestellt werden.
-
Ein
Peptid, das eine CDR-Region umfasst, umfasst mindestens eine CDR-Region aus der VH-Region oder
der VL-Region. Ein Peptid, das mehrere CDR-Regionen umfasst, kann hergestellt werden,
indem die CDR-Regionen direkt oder über einen geeigneten Peptidlinker
miteinander verknüpft
werden.
-
Das
Peptid, das eine CDR-Region umfasst, der vorliegenden Erfindung
kann durch die Konstruktion der cDNAs, codierend die CDRs der VH-Region
und der VL-Region eines Antikörpers,
der mit FGF-8 spezifisch reagiert, die Insertion der cDNAs in einen
Expressionsvektor für
einen Prokaryonten oder einen Expressionsvektor für einen
Eukaryonten und dann das Einbringen des Expressionsvektors in einen
Prokaryonten oder einen Eukaryonten, um das Peptid zu exprimieren,
hergestellt werden. Das Peptid, das eine CDR-Region umfasst, kann
auch durch ein chemisches Syntheseverfahren wie die Fmoc-Methode
(Fluorenylmethoxycarbonyl-Methode),
die tBoc-Methode (t-Butyloxycarbonyl-Methode) oder dergleichen hergestellt
werden.
-
Ein
Verfahren zur Herstellung des menschlichen chimären Antikörpers, des menschlichen CDR-transplantierten
Antikörpers
und des Antikörperfragments
davon, welche mit FGF-8 spezifisch reagieren und die Funktion von
FGF-8 inhibieren, sowie eine Methode zur Beurteilung der Aktivität und ein
Verfahren zur Verwendung davon werden nachstehend erläutert.
-
1. Herstellung eines menschlichen
chimären
Antikörpers
und eines menschlichen CDR-transplantierten Antikörpers
-
(1) Konstruktion eines Vektors für die Expression
eines humanisierten Antikörpers
-
Ein
Vektor für
die Expression eines humanisierten Antikörpers ist ein Expressionsvektor
für eine
tierische Zelle, in den die DNAs, welche die CH-Region und die CL-Region
eines menschlichen Antikörpers
codieren, inseriert worden sind, und welcher durch die Clonierung
der jeweiligen DNAs, welche die CH-Region und die CL-Region eines
menschlichen Antikörpers
codieren, in einen Expressionsvektor für eine tierische Zelle konstruiert
wird.
-
Die
C-Region eines menschlichen Antikörpers kann die CH-Region und
die CL-Region eines beliebigen menschlichen Antikörpers umfassen.
Beispiele schließen
die CH-Region der Subklasse γ1
und die CL-Region der κ-Klasse
eines menschlichen Antikörpers
und dergleichen ein. Was die DNAs anbetrifft, welche die CH-Region und die CL-Region
eines menschlichen Antikörpers
codieren, kann eine chromosomale DNA, umfassend ein Exon und ein
Intron, oder eine cDNA verwendet werden. Was den Expressionsvektor
für eine
tierische Zelle anbetrifft, kann ein beliebiger Expressionsvektor
verwendet werden, solange wie die C-Region eines menschlichen Antikörpers darin
inseriert und exprimiert werden kann. Beispiele schließen pAGE107
[Cytotechnology 3 (1990), 133], pAGE103 [J. Biochem. 101 (1987),
1307], pHSG274 [Gene 27 (1984), 223], pKCR [Proc. Natl. Acad. Sci.
USA 78 (1981), 1527], pSGIβd2-4
[Cytotechnology 4 (1990), 173], pSE1 UK1Sed1-3 [Cytotechnology 13
(1993), 79] und dergleichen ein. Beispiele eines Promotors und eines
Enhancers, welche in einem Expressionsvektor für eine tierische Zelle verwendet
werden, schließen
den frühen
SV40-Promotor und -Enhancer [J. Biochem. 101 (1987), 1307], den
Moloney-Maus-Leukämievirus-LTR-Promotor
und -Enhancer [Biochem. Biophys. Res. Commun. 149 (1987), 960],
den Promotor [Cell 41 (1985), 479] und den Enhancer [Cell 33 (1983),
717] der H-Kette eines Immunglobulins und dergleichen ein.
-
Der
Vektor für
die Expression eines humanisierten Antikörpers kann entweder dem Typ
entsprechen, worin die H-Kette und die L-Kette des Antikörpers auf separaten
Vektoren vorliegen, oder dem Typ, worin beide Ketten auf dem gleichen
Vektor vorliegen (Tandem-Typ). Im Hinblick auf die Einfachheit der
Konstruktion eines Vektors für
die Expression eines humanisierten Antikörpers, die Einfachheit der
Einschleusung in tierische Zellen und der Ausgeglichenheit zwischen
den Expressionsmengen der H-Kette und der L-Kette des Antikörpers in
tierischen Zellen wird ein Tandem-Typ des Vektors für die Expression
eines humanisierten Antikörpers
bevorzugt [J. Immunol. Methods 167 (1994), 271]. Beispiele für den Tandem-Typ
des Vektors für
die Expression eines humanisierten Antikörpers schließen pKANTEX93
(
WO 97/10354 ), pEE18
[Hybridoma 17 (1998), 559] und dergleichen ein.
-
Der
konstruierte Vektor für
die Expression eines humanisierten Antikörpers kann zur Expression eines menschlichen
chimären
Antikörpers
und eines menschlichen CDR-transplantierten Antikörpers in
tierischen Zellen verwendet werden.
-
(2) Herstellung einer cDNA, codierend
die V-Region eines Antikörpers,
der von einem nicht-menschlichen Tier stammt, und Analyse der Aminosäuresequenz
-
Die
cDNAs, codierend die VH-Region und die VL-Region eines Antikörpers, der
von einem nicht-menschlichen Tier stammt, wie eines Mausantikörpers, werden
wie folgt erhalten.
-
Aus
Hybridomzellen, die einen Mausantikörper oder dergleichen herstellen,
wird die mRNA extrahiert. Anschließend wird unter Verwendung
der mRNA eine cDNA synthetisiert. Die synthetisierte cDNA wird in
einen Vektor wie einen Phagen oder ein Plasmid inseriert, um eine
cDNA-Bank herzustellen. Unter Verwendung eines Teils der C-Region
oder der V-Region eines Mausantikörpers als Sonde werden jeweils
ein rekombinanter Phage oder ein rekombinantes Plasmid, enthaltend
die cDNA, welche die VH-Region codiert, und ein rekombinanter Phage
oder ein rekombinantes Plasmid, enthaltend die cDNA, welche die
VL-Region codiert, aus der Bank isoliert. Die vollständigen Nucleotidsequenzen
der VH-Region und der VL-Region
des Mausantikörpers von
Interesse auf dem rekombinanten Phagen oder dem rekombinanten Plasmid
werden bestimmt, und die vollständigen
Aminosäuresequenzen
der VH-Region und der VL-Region werden aus den Nucleotidsequenzen abgeleitet.
-
Das
nicht-menschliche Tier kann irgendein Tier wie eine Maus, eine Ratte,
ein Hamster oder ein Kaninchen sein, solange wie ein Hybridom davon
erzeugt werden kann.
-
Das
Verfahren zur Präparation
der Gesamt-RNA aus einem Hybridom schließt die Guanidin-Thiocyanat-Cäsiumtrifluoracetat-Methode
[Methods in Enzymol. 154 (1987), 3] und dergleichen ein. Das Verfahren
zur Präparation
der mRNA aus der Gesamt-RNA schließt eine Methode unter Verwendung
von Oligo(dT), immobilisiert an eine Cellulosesäule [Molecular Cloning: A Laborstory
Manual, Cold Spring Harbor Lab. Press, New York (1989)] und dergleichen
ein. Ein Kit für
die Präparation
der mRNA aus einer Hybridomzelle schließt den Fast Track mRNA-lsolationskit
(hergestellt von Invitrogen), den Quick Prep mRNA-Reinigungskit
(hergestellt von Pharmacia) und dergleichen ein.
-
Das
Verfahren zur Synthese einer cDNA und zur Herstellung einer cDNA-Bank schließt herkömmliche Verfahren
[Molecular Cloning: A Laborstory Manual, Cold Spring Harbor Lab.
Press, New York (1989); Current Protocols in Molecular Biology,
Anhang 1–34],
ein Verfahren unter Verwendung eines im Handel erhältlichen Kits
wie des Super ScriptTM Plasmid-Systems für die cDNA-Synthese
und der Plasmidclonierung (hergestellt von Gibco-BRL) oder des ZAP-cDNA-Kits
(hergestellt von Stratagene), und dergleichen ein.
-
Jeder
beliebiger Vektor, in den die cDNA inseriert werden kann, welche
unter Verwendung der mRNA, die aus einem Hybridom extrahiert wurde,
als Matrize synthetisiert wurde, um eine cDNA-Bank herzustellen, kann
verwendet werden, solange wie die cDNA inseriert werden kann. Beispiele
schließen
ZAP-Express [Strategies 5 (1992), 58], pBluescript II SK(+) [Nucleic
Acids Research 17 (1989), 9494], λzapII
(hergestellt von Stratagene), λgt10
und λgt11
[DNA Cloning: A Practical Approach 1 (1985), 49], Lambda BlueMid
(hergestellt von Clontech), λExCell
und pP7T3-18U (hergestellt von Pharmacia), pcD2 [Mol. Cell. Biol.
3 (1983), 280], pUC18 [Gene 33 (1985), 103] und dergleichen ein.
-
Es
kann ein beliebiges E. coli-Bakterium verwendet werden, um die cDNA-Bank, welche unter
Verwendung eines Phagen- oder Plasmidvektors konstruiert wurde,
einzuschleusen, solange wie die cDNA-Bank eingeschleust, exprimiert
und aufrechterhalten werden kann. Beispiele schließen XL1-Blue
MRF' [Strategies
5 (1992), 81], C600 [Genetics 39 (1954), 440], Y1088 und Y1090 [Science
222 (1983), 778], NM522 [J. Mol. Biol. 166 (1983), 1], K802 [J.
Mol. Biol. 16 (1966), 118], JM105 [Gene 38 (1985), 275] und dergleichen
ein.
-
Ein
Koloniehybridisierungs- oder Plaquehybridisierungsverfahren unter
Verwendung einer Isotop- oder Fluoreszenz-markierten Sonde kann
verwendet werden, um die cDNA-Clone, codierend die VH-Region und
die VL-Region eines Antikörpers,
welcher von einem nicht-menschlichen Tier stammt, in der cDNA-Bank zu
selektieren [Molecular Cloning: A Laborstory Manual, Cold Spring
Harbor Lab. Press, New York (1989)]. Die cDNAs, welche die VH-Region
und die VL-Region codieren, können
auch durch eine Polymerasekettenreaktion [PCR; Molecular Cloning:
A Laborstory Manual, Cold Spring Harbor Lab. Press, New York (1989);
Current Protocols in Molecular Biology, Anhang 1–34] mittels der Konstruktion
von Primern und unter Verwendung einer cDNA, welche aus der mRNA
hergestellt wurde oder aus einer cDNA-Bank isoliert wurde, als Matrize hergestellt
werden.
-
Die
Nucleotidsequenz der cDNA kann durch das Spalten der cDNA, welche
durch das vorstehende Verfahren selektiert wurde, mit geeigneten
Restriktionsenzymen und dergleichen, das Clonieren der Fragmente
in ein Plasmid wie pBluescript SK(–) (hergestellt von Stratagene),
das Durchführen
einer Umsetzung mittels eines herkömmlicherweise angewendeten
Nucleotid-Analyseverfahren wie der Didesoxy-Methode nach Sanger F. et al. [Proc.
Natl. Acad. Sci. USA 74 (1977), 5463] und dann das Analysieren der
Sequenz unter Verwendung eines automatischen Nucleotidsequenzanalysengerätes wie
ABI377 (hergestellt von Applied Biosystems) oder dergleichen bestimmt
werden.
-
Ob
die erhaltenen cDNAs die vollständigen
Aminosäuresequenzen
der VH-Region und
der VL-Region des Antikörpers
codieren, welche eine sekretorische Signalsequenz enthalten, kann
durch die Ableitung der vollständigen
Aminosäuresequenzen
der VH-Region und der VL-Region aus der bestimmten Nucleotidsequenz
und den Vergleich davon mit den vollständigen Aminosäuresequenzen
der VH-Region und
der VL-Region von bekannten Antikörpern [Sequences of Proteins
of Immunological Interest, US Dept. Health and Human Services (1991)]
bestätigt
werden. Die Länge
der sekretorischen Signalsequenz und der N-terminalen Aminosäuresequenz
kann durch den Vergleich der vollständigen Aminosäuresequenzen
der VH-Region und der VL-Region des Antikörpers, umfassend eine sekretorische
Signalsequenz, mit den vollständigen
Aminosäuresequenzen
der VH-Region und der VL-Region von bekannten Antikörpern [Sequences
of Proteins of Immunological Interest, US Dept. Health and Human
Services (1991)], wobei die Untergruppe, denen sie angehören, bekannt
sein kann, abgeleitet werden. Außerdem kann die Aminosäuresequenz
aller CDR-Regionen in der VH-Region und der VL-Region identifiziert
werden.
-
Ferner
kann die Neuartigkeit der Sequenz durch das Durchführen einer
Homologiesuche zu Sequenzen in einer beliebigen Datenbank, zum Beispiel
SWISS-PROT, PIR-Protein
oder dergleichen, unter Verwendung der vollständigen Aminosäuresequenzen
der VH-Region und der VL-Region zum Beispiel nach der BLAST-Methode
[J. Mol. Biol. 215 (1990), 403] oder dergleichen untersucht werden.
-
(3) Konstruktion eines Vektors für die Expression
des menschlichen chimären
Antikörpers
-
Ein
Vektor für
die Expression des menschlichen chimären Antikörpers kann durch die Clonierung
der cDNAs, codierend die VH-Region und die VL-Region eines Antikörpers, welcher
von einem nicht-menschlichen Tier stammt, stromaufwärts der
DNAs, welche die CH-Region und die CL-Region eines menschlichen
Antikörpers
codieren, auf dem Vektor für
die Expression eines humanisierten Antikörpers, wie unter Punkt 1(1) beschrieben,
konstruiert werden. Zum Beispiel werden unter Verwendung eines Plasmids,
umfassend die cDNAs, welche die VH-Region und die VL-Region eines Antikörpers codieren,
welcher von einem nicht-menschlichen Tier stammt, als Matrize die
VH-Region und die VL-Region des Antikörpers mittels PCR unter Verwendung
von Primern an dem 5'-terminalen
Ende und dem 3'-terminalen
Ende, umfassend die Erkennungssequenz eines geeigneten Restriktionsenzyms
und eine Nucleotidsequenz, welche die V-Region codiert, amplifiziert.
Das jeweilige amplifizierte Produkt wird in ein Plasmid wie pBluescript
SK(–)
(hergestellt von Stratagene) cloniert, und die Nucleotidsequenz
wird gemäß dem unter
Punkt 1(2) beschriebenen Verfahren bestimmt. Auf diese Weise wird
ein Plasmid erhalten, das die DNA-Sequenzen umfasst, welche die
Aminosäuresequenzen der
VH-Region und der VL-Region des gewünschten Antikörpers codieren.
Die cDNAs, welche die Aminosäuresequenzen
der VH-Region und der VL-Region des Antikörpers codieren, werden aus
dem Plasmid isoliert und zur Expression davon in einer geeigneten
Form stromaufwärts
der Gene, welche die C-Regionen der H-Kette und der L-Kette des menschlichen
Antikörpers
codieren, in den Vektor für
die Expression eines humanisierten Antikörpers, wie unter Punkt 1(1)
beschrieben, cloniert, um auf diese Weise einen Vektor für die Expression
des menschlichen chimären
Antikörpers
zu konstruieren.
-
(4) Konstruktion einer cDNA, welche die
V-Region eines menschlichen CDR-transplantierten Antikörpers codiert
-
Die
cDNAs, welche die VH-Region und die VL-Region eines menschlichen
CDR-transplantierten Antikörpers
codieren, können
wie folgt erhalten werden. Zuerst werden die Aminosäuresequenzen
von FR-Regionen in der VH-Region und der VL-Region eines menschlichen Antikörpers, in
welche die Aminosäuresequenzen
von CDR-Regionen in der VH-Region und der VL-Region eines Antikörpers transplantiert
werden, welche von einem Antikörper
abgeleitet sind, der von einem nicht-menschlichen Tier stammt, ausgewählt. Es
können beliebige
Aminosäuresequenzen
von FR-Regionen
in der VH-Region und der VL-Region eines menschlichen Antikörpers verwendet
werden, solange wie sie von einem menschlichen Antikörper stammen.
Beispiele schließen
Aminosäuresequenzen
von FR-Fragmenten in der VH-Region und der VL-Region von menschlichen Antikörpern, welche
in einer Datenbank wie der Proteindatenbank erfasst sind, sowie
Aminosäuresequenzen, welche
in den Untergruppen von FR-Regionen in der VH-Region und der VL-Region
von menschlichen Antikörpern
häufig
vorkommen [Sequences of Proteins of Immunological Interest, US Dept.
Health and Human Services (1991)], und dergleichen ein. Von diesen
Sequenzen werden für
die Herstellung eines menschlichen CDR-transplantierten Antikörpers mit
einer wirksamen Aktivität
vorzugsweise Aminosäuresequenzen
mit einer hohen Homologie (mindestens 60% oder mehr) mit den Aminosäuresequenzen
von FR-Regionen in der VH-Region und der VL-Region eines Zielantikörpers, welcher
von einem nicht-menschlichen Tier stammt, ausgewählt.
-
Anschließend werden
die Aminosäuresequenzen
von CDR-Regionen in der VH-Region und der VL-Region des Antikörpers, welcher
von einem nicht-menschlichen Tier stammt, in die ausgewählten Aminosäuresequenzen
von FR-Regionen in der VH-Region und der VL-Region eines menschlichen
Antikörpers transplantiert,
um die Aminosäuresequenzen
der VH-Region und der VL-Region eines menschlichen CDR-transplantierten
Antikörpers
zu konstruieren. Die konstruierten Aminosäure sequenzen werden unter Berücksichtigung
der Frequenz des Codongebrauchs, welche für Nucleotidsequenzen von Genen,
codierend Antikörper,
ermittelt wurde [Sequences of Proteins of Immunological Interest,
US Dept. Health and Human Services (1991)], in Nucleotidsequenzen
translatiert, und anschließend
werden die DNA-Sequenzen, welche die Aminosäuresequenzen der VH-Region
und der VL-Region
eines menschlichen CDR-transplantierten Antikörpers codieren, konstruiert.
Hierzu werden mehrere synthetische DNAs mit einer Länge von
etwa 100 Nucleotiden basierend auf den entworfenen Nucleotidsequenzen
synthetisiert. Im Anschluss daran wird unter Verwendung dieser DNAs
eine PCR durchgeführt.
In diesem Fall wird bevorzugt, dass im Hinblick auf die Reaktionseffizienz
der PCR und die Längen
der DNAs, welche synthetisiert werden können, jeweils 6 synthetische DNAs
für die
VH-Region und die VL-Region entworfen werden. Ferner können diese
DNAs ohne weiteres in den Vektor für die Expression eines humanisierten
Antikörpers,
welcher unter Punkt 1(1) konstruiert wurde, cloniert werden, indem
die Erkennungssequenz eines geeigneten Restriktionsenzyms in das
an beiden Enden vorhandene 5'-Ende
der synthetischen DNAs eingeführt
wird. Im Anschluss an die PCR wird das amplifizierte Produkt in
ein Plasmid wie pBluescript SK(–)
(hergestellt von Stratagene) cloniert, und die Nucleotidsequenzen werden
gemäß dem Verfahren,
welches unter Punkt 1(2) beschrieben ist, bestimmt, um ein Plasmid
zu erhalten, das die Nucleotidsequenzen enthält, welche die VH-Region und
die VL-Region des konstruierten menschlichen CDR-transplantierten
Antikörpers
codieren.
-
(5) Modifikation der Aminosäuresequenz
der V-Region des menschlichen CDR-transplantierten Antikörpers
-
Es
ist bekannt, dass, wenn ein menschlicher CDR-transplantierter Antikörper dadurch
hergestellt wird, dass lediglich nur CDRs aus der VH-Region und
der VL-Region eines Antikörpers,
welcher von einem nicht-menschlichen Tier stammt, in die FR-Regionen
in der VH-Region und der VL-Region eines menschlichen Antikörpers transplantiert
werden, die Antigenbindungsaktivität geringer ist als diejenige
des ursprünglichen Antikörpers, welcher
von einem nicht-menschlichen
Tier stammt [Bio/Technology 9 (1991), 266]. Als Grund hierfür wird in
Betracht gezogen, dass mehrere Aminosäurereste nicht nur in den CDR-Regionen,
sondern auch in den FR-Regionen direkt oder indirekt mit der Antigenbindungsaktivität in der
VH-Region und der VL-Region des ursprünglichen Antikörpers, der
von einem nicht-menschlichen Tier stammt, in Beziehung stehen, und
dass sie durch andere Aminosäurereste
anderer FR-Regionen in der VH-Region und der VL-Region eines menschlichen
Antikörpers
ersetzt werden. Um dieses Problem zu lösen, wird bei menschlichen CDR-transplantierten
Antikörpern
in den Aminosäuresequenzen
der FR-Regionen in der VH-Region und der VL-Region eines menschlichen
Antikörpers
ein Aminosäurerest,
welcher direkt mit der Bindung an ein Antigen in Beziehung steht,
oder ein Aminosäurerest,
welcher indirekt mit der Bindung an ein Antigen in Beziehung steht,
durch die Wechselwirkung mit einem Aminosäurerest in der CDR-Region oder
durch die Beibehaltung der dreidimensionalen Struktur eines Antikörpers identifiziert
und zu einem Aminosäurerest
modifiziert, der in dem ursprünglichen
Antikörper,
welcher von einem nicht-menschlichen Tier stammt, vorhanden ist,
wodurch die Antigenbindungsaktivität, die vermindert worden ist,
erhöht
wird [Bio/Technology 9 (1991), 266]. Bei der Herstellung eines menschlichen
CDR-transplantierten Antikörpers
ist die effiziente Identifizierung der Aminosäurereste, welche mit der Antigenbindungsaktivität in der
FR-Region in Beziehung stehen, besonders wichtig, wobei die dreidimensionale
Struktur eines Antikörpers
konstruiert und durch Röntgenkristallographie
[J. Mol. Biol. 112 (1977), 535], computergestützte Modellerstellung [Protein
Engineering 7 (1991), 1501] oder dergleichen analysiert wird. Obwohl
die Information der dreidimensionalen Struktur von Antikörpern bei
der Herstellung eines menschlichen CDR-transplantierten Antikörpers nützlich gewesen
ist, ist bisher noch kein Verfahren zur Herstellung eines menschlichen
CDR-transplantierten Antikörpers
entwickelt worden, welches auf alle Antikörper angewendet werden kann.
Daher müssen
gegenwärtig
verschiedene Versuche durchgeführt
werden, wobei zum Beispiel mehrere modifizierte Antikörper des
jeweiligen Antikörpers
hergestellt werden, um die Beziehung zwischen jedem der modifizierten
Antikörper
und dessen Antigenbindungsaktivität zu untersuchen.
-
Die
Modifikation der ausgewählten
Aminosäuresequenz
einer FR-Region in der VH-Region und der VL-Region eines menschlichen
Antikörpers
kann durch eine PCR unter Verwendung von verschiedenen synthetischen
DNAs für
die Modifikation erfolgen. Die Nucleotidsequenz des amplifizierten
Produkts, welches durch die PCR erhalten wurde, wird gemäß dem Verfahren,
das vorstehend unter Punkt 1(2) be schrieben wurde, bestimmt, um
zu überprüfen, ob
die gewünschte
Modifikation eingeführt
worden ist.
-
(6) Konstruktion eines Vektors für die Expression
des menschlichen CDR-transplantierten Antikörpers
-
Der
Vektor für
die Expression des menschlichen CDR-transplantierten Antikörpers kann
durch die Clonierung der cDNAs, codierend die VH-Region und die
VL-Region des menschlichen
CDR-transplantierten Antikörpers,
welche unter Punkt 1(4) und 1(5) konstruiert wurden, stromaufwärts der
DNAs, welche die CH-Region und die CL-Region des menschlichen Antikörpers codieren,
in den Vektor für
die Expression eines humanisierten Antikörpers, wie unter Punkt 1(1)
beschrieben, konstruiert werden. Wenn zum Beispiel Erkennungsstellen
für geeignete
Restriktionsenzyme in das 5'-Ende
der synthetischen DNAs an beiden Enden in die synthetischen DNAs
eingeführt
werden, welche bei der Konstruktion der VH-Region und der VL-Region
des menschlichen CDR-transplantierten Antikörpers unter Punkt 1(4) und
1(5) verwendet wurden, kann die Clonierung derart erfolgen, dass
die DNAs in einer geeigneten Form stromaufwärts der DNAs, welche die CH-Region und
die CL-Region des
menschlichen Antikörpers
codieren, in dem Vektor für
die Expression eines humanisierten Antikörpers, wie unter Punkt 1(1)
beschrieben, exprimiert werden.
-
(7) Transiente Expression der humanisierten
Antikörper
-
Um
die Antigenbindungsaktivität
der verschiedenen hergestellten humanisierten Antikörper wirksam zu
beurteilen, können
die humanisierten Antikörper
unter Verwendung des Expressionsvektors für den humanisierten Antikörper, wie
unter Punkt 1(3) und 1(6) beschrieben, oder eines modifizierten
Expressionsvektors davon transient exprimiert werden. Jede beliebige
Zelle kann als eine Wirtszelle verwendet werden, solange wie die
Wirtszelle einen humanisierten Antikörper exprimieren kann. Im Allgemeinen
werden COS-7-Zellen (ATCC CRL-1651) im Hinblick auf ihre hohe Expressionsmenge
verwendet [Methods in Nucleic Acids Res., CRC Press, S. 283 (1991)].
Das Verfahren für
die Einschleusung des Expressionsvektors in COS-7-Zellen schließt die DEAE-Dextran-Methode
[Methods in Nucleic Acids Res., CRC Press, S. 283 (1991)], die Lipofektion-Methode
[Proc. Natl. Acad. Sci. USA 84 (1987), 7413] und dergleichen ein.
-
Nach
der Einschleusung des Vektors in die Zelle kann die Expressionsmenge
und die Antigenbindungsaktivität
des humanisierten Antikörpers
in dem Kulturüberstand
durch einen Enzymimmuntest [ELISA; Antibodies: A Laborstory Manual,
Cold Spring Harbor Laborstory, Kapitel 14 (1988); Monoclonal Antibodies: Principles
and Practice, Academic Press Limited (1996)] und dergleichen bestimmt
werden.
-
(8) Stabile Expression des humanisierten
Antikörpers
-
Eine
Transformante, welche einen humanisierten Antikörper stabil produziert, kann
dadurch erhalten werden, dass der Expressionsvektor für einen
humanisierten Antikörper,
beschrieben unter Punkt 1(3) und 1(6), in eine geeignete Wirtszelle
eingeschleust wird.
-
Das
Verfahren für
die Einschleusung des Expressionsvektors in eine Wirtszelle schließt eine
Elektroporation [ungeprüfte
veröffentlichte
japanische Patentanmeldung Nr.
257891/90 ;
Cytotechnology 3 (1990), 133] und dergleichen ein.
-
Jede
beliebige Zelle kann als die Wirtszelle, in welche der Vektor für die Expression
eines humanisierten Antikörpers
eingebracht werden soll, verwendet werden, solange wie sie einen
humanisierten Antikörper exprimieren
kann. Beispiele schließen
Maus-SP2/0-Ag14-Zellen (ATCC CRL-1581), Maus-P3X63-Ag8.653-Zellen
(ATCC CRL-1580), CHO-Zellen, worin das Dihydrofolatreduktase-Gen
(dhfr) nachweisbar ist [Proc. Natl. Acad. Sci. USA 77 (1980), 4216],
Ratten-YB2/3HL.P2.-G11.16Ag.20-Zellen (YB2/0-Zellen,
ATCC CRL-1662] und dergleichen ein.
-
Nach
der Einschleusung des Expressionsvektors in die Zelle werden die
Transformanten, welche einen humanisierten Antikörper stabil exprimieren, durch
die Züchtung
in einem Medium für
die Kultur von tierischen Zellen, umfassend ein Mittel wie G418-Sulfat
(G418, hergestellt von Sigma) oder dergleichen, selektiert [J. Immunol.
Methods 167 (1994), 271]. Das Medium für die Kultur von tierischen
Zellen schließt
PRMI-1640-Medium (hergestellt von Nissui Pharmaceuticals), GIT-Medium
(hergestellt von Nissui Pharmaceuticals), EX-CELL302-Medium (hergestellt
von JRH), IMDM-Medium (hergestellt von Gibco-BRL), Hybridoma-SFM-Medium
(hergestellt von Gibco-BRL) sowie Medien, welche durch die Zugabe
von verschiedenen Zusätzen
wie FBS zu diesen Medien erhalten werden, und dergleichen ein. Der
humanisierte Antikörper
kann durch Züchtung
der ausgewählten
Transformanten in einem Medium in dem Kulturmedium hergestellt und
angereichert werden. Die Menge und die Antigenbindungsaktivität des in
dem Kulturmedium exprimierten humanisierten Antikörpers kann
durch einen ELISA oder dergleichen gemessen werden. Die Produktionsmenge
des humanisierten Antikörpers
kann durch eine Transformation unter Verwendung des dhfr-Amplifikationssystems oder
dergleichen erhöht
werden [J. Immunol. Methods 167 (1994), 271].
-
Der
humanisierte Antikörper
kann aus dem Medium, in dem die Transformante gezüchtet wird,
unter Verwendung einer Protein A-Säule gereinigt werden [Antibodies:
A Laborstory Manual, Cold Spring Harbor Laborstory, Kapitel 8 (1988);
Monoclonal Antibodies: Principles and Practice, Academic Press Limited
(1996)]. Irgendwelche anderen herkömmliche Verfahren für die Proteinreinigung
können
ebenfalls angewendet werden. Zum Beispiel kann der humanisierte
Antikörper
durch eine Kombination von Gelfiltration, Ionenaustauschchromatographie,
Ultrafiltration und dergleichen gereinigt werden. Das Molekulargewicht
der H-Kette oder der L-Kette des gereinigten humanisierten Antikörpers oder
des Antikörpermoleküls insgesamt
wird durch eine Polyacrylamid-Gelelektrophorese [SDS-PAGE; Nature
227 (1970), 680], ein Western-Blot-Verfahren [Antibodies: A Laborstory
Manual, Cold Spring Harbor Laborstory, Kapitel 12 (1988); Monoclonal
Antibodies: Principles and Practice, Academic Press Limited (1996)]
und dergleichen bestimmt.
-
2. Herstellung eines Antikörperfragments
-
Das
Antikörperfragment
kann basierend auf dem unter Punkt 1 beschriebenen humanisierten
Antikörper
genetisch oder proteinchemisch hergestellt werden. Das Antikörperfragment
schließt
Fab, F(ab')2, Fab', scFv,
ein Diabody, dsFv, ein Peptid, das eine CDR-Region umfasst, und
dergleichen ein.
-
(1) Herstellung eines Fab-Fragments
-
Das
Fab-Fragment kann durch die Behandlung von IgG mit dem proteolytischen
Enzym Papain hergestellt werden. Wenn der ursprüngliche Antikörper einer
IgG-Subklasse angehört,
welche eine Protein A-Bindungsaktivität besitzt, können im
Anschluss an die Papainbehandlung einheitliche Fab-Fragmente gewonnen werden,
indem das Reaktionsgemisch über
eine Protein A-Säule
geleitet wird, wobei IgG-Moleküle und Fc-Fragmente
abgetrennt werden [Monoclonal Antibodies: Principles and Practice,
dritte Auflage (1995)]. Wenn der ursprüngliche Antikörper einer
IgG-Subklasse angehört, welche
keine Protein A-Bindungsaktivität besitzt,
können
die Fab-Fragmente durch eine Ionenaustauschchromatographie in einer
Fraktion, die bei einer niedrigen Salzkonzentration eluiert, gewonnen
werden [Monoclonal Antibodies: Principles and Practice, dritte Auflage
(1995)]. Ferner kann das Fab-Fragment auch unter Verwendung von
Escherichia coli genetisch hergestellt werden. Zum Beispiel kann
ein Fab-Expressionsvektor dadurch hergestellt werden, dass die DNA,
welche die V-Region des Antikörpers
codiert, beschrieben unter Punkt 1(2), 1(3) und 1(5), in einen Vektor
für die Expression
von Fab cloniert wird. Als Vektor für die Expression von Fab kann
ein beliebiger Vektor verwendet werden, solange wie eine DNA, welche
das Fab-Fragment codiert, inseriert und exprimiert werden kann.
Beispiele schließen
pIT106 [Science 240 (1988), 1041] und dergleichen ein. Das Fab-Fragment
kann in einem Einschlusskörper
oder im periplasmatischen Raum gebildet und angereichert werden,
indem der Fab-Expressionsvektor in ein geeignetes Escherichia coli-Bakterium
eingebracht wird. Das Fab-Fragment mit einer Bindungsaktivität kann durch
einen Umfaltungsprozess, der im Allgemeinen bei der Herstellung
eines Proteins verwendet wird, aus dem Einschlusskörper gewonnen
werden. Wenn das Fab-Fragment im periplasmatischen Raum exprimiert
wird, entweicht das Fab-Fragment
mit einer Bindungsaktivität
in den Kulturüberstand.
Die einheitlichen Fab-Fragmente
können
nach der Umfaltung oder unter Verwendung einer Antigen bindenden
Säule aus
dem Kulturmedium gereinigt werden [Antibody Engineering, A Practical
Guide, W. H. Freeman and Company (1992)].
-
(2) Herstellung eines F(ab')2-Fragments
-
Das
F(ab')2-Fragment
kann durch die Behandlung von IgG mit der Protease Pepsin hergestellt
werden. Im Anschluss an die Pepsinbehandlung können die einheitlichen F(ab')2-Fragmente
mittels eines ähnlichen Reinigungsverfahrens
wie im Falle des Fab-Fragments gewonnen werden [Monoclonal Antibodies:
Principles and Practice, dritte Auflage, Academic Press (1995)].
Ferner kann das F(ab')2-Fragment auch durch das Verfahren, welches
unter Punkt 2(3) beschrieben wird, wobei ein Fab'-Fragment mit einem Maleimid wie o-PDM oder
Bismaleimidhexan unter Bildung einer Thioetherbindung behandelt
wird, oder ein Verfahren, wobei das Fab'-Fragment mit DTNB unter Bildung einer
S-S-Bindung behandelt wird, hergestellt werden [Antibody Engineering,
A Practical Approach, IRL Press (1996)].
-
(3) Herstellung eines Fab'-Fragments
-
Das
Fab'-Fragment kann
durch die Behandlung des unter Punkt 2(2) beschriebenen F(ab')2-Fragments
mit einem Reduktionsmittel wie Dithiothreit hergestellt werden.
Ferner kann das Fab'-Fragment
unter Verwendung von Escherichia coli genetisch hergestellt werden.
Zum Beispiel kann ein Fab'-Expressionsvektor durch
die Clonierung der DNA, welche die V-Region des Antikörpers codiert,
wie unter Punkt 1(2), 1(4) und 1(5) beschrieben, in einen Vektor
für die
Expression von Fab' konstruiert
werden. Als Vektor für
die Expression von Fab' kann
ein beliebiger Vektor verwendet werden, solange wie eine DNA, welche
Fab' codiert, inseriert und
exprimiert werden kann. Beispiele schließen pAK19 [Bio/Technology 10
(1992), 163] und dergleichen ein. Das Fab'-Fragment kann gebildet und in einem
Einschlusskörper
oder im periplasmatischen Raum angereichert werden, indem der Fab'-Expressionsvektor
in ein geeignetes Escherichia coli-Bakterium eingebracht wird. Das
Fab'-Fragment mit
einer Bindungsaktivität
kann durch einen Umfaltungsprozess, der im Allgemeinen bei der Herstellung
eines Proteins verwendet wird, aus dem Einschlusskörper gewonnen
werden. Wenn das Fab'-Fragment im periplasmatischen
Raum exprimiert wird, kann es extrazellulär gewonnen werden, indem die Zellen
durch eine Behandlung wie einen partiellen Verdau mit Lysozym, einen
osmotischen Schock oder Beschallen aufgebrochen werden. Die einheitlichen
Fab'-Fragmente können nach
der Umfaltung oder unter Verwendung einer Protein G-Säule oder
dergleichen aus der Suspension der aufgebrochenen Zellen gereinigt werden
[Antibody Engineering, A Practical Approach, IRL Press (1996)].
-
(4) Herstellung eines scFv-Fragments
-
Das
scFv-Fragment kann unter Verwendung eines Phagens oder eines Escherichia
coli-Bakteriums genetisch hergestellt werden. Zum Beispiel wird
eine DNA, codierend das scFv-Fragment, durch die Ligierung der DNAs,
welche die VH-Region
und die VL-Region des Antikörpers
codieren, beschrieben unter Punkt 1(2), 1(4) und 1(5), über eine
DNA, codierend einen Polypeptidlinker, welcher eine Aminosäuresequenz
von 12 oder mehr Resten umfasst, hergestellt. Durch die Clonierung der
DNA in einen Vektor für
die Expression des scFv-Fragments kann ein scFv-Expressionsvektor konstruiert werden.
Als Vektor für
die Expression von scFv kann ein beliebiger Vektor verwendet werden,
solange wie die DNA, welche scFv codiert, inseriert und exprimiert
werden kann. Beispiele schließen
pCANTAB5E (hergestellt von Pharmacia), Phfa [Hum. Antibody Hybridoma
5 (1994), 48] und dergleichen ein. Der scFv-Expressionsvektor wird
in ein geeignetes Escherichia coli-Bakterium eingebracht, welches
mit einem Helferphagen infiziert wird, wodurch ein Phage erhalten
wird, welcher das scFv-Fragment in Verknüpfung mit dem Phagen-Oberflächenprotein
auf der Phagenoberfläche
exprimiert. Das scFv-Fragment kann auch gebildet und in einem Einschlusskörper oder
im periplasmatischen Raum eines Escherichia coli-Bakterium, in welches
der scFv-Expressionsvektor eingeschleust wird, angereichert werden.
Das scFv-Fragment, welches eine Bindungsaktivität hat, kann durch einen Umfaltungsprozess,
welcher im Allgemeinen bei der Herstellung eines Proteins verwendet
wird, aus dem Einschlusskörper
gewonnen werden. Wenn das scFv-Fragment im periplasmatischen Raum
exprimiert wird, kann es extrazellulär gewonnen werden, indem die
Zellen durch eine Behandlung wie einen partiellen Verdau mit Lysozym,
einen osmotischen Schock, Beschallen oder dergleichen aufgebrochen
werden. Die einheitlichen scFv-Fragmente können nach der Umfaltung oder
aus der aufgebrochenen Zellsuspension durch eine Kationenaustauschchromatographie
oder dergleichen gereinigt werden [Antibody Engineering, A Practical
Approach, IRL Press (1996)].
-
(5) Herstellung eines Diabody-Fragments
-
Ein
Diabody kann dadurch hergestellt werden, dass der Polypeptidlinker,
welcher verwendet wurde, um das vorstehende scFv-Fragment herzustellen,
durch amen Polypeptidlinker von etwa 3 bis 10 Resten ersetzt wird.
Ein divalenter Diabody kann hergestellt werden, wenn die VH-Region
und die VL-Region einer Antikörperspezies
verwendet werden. Ein Diabody mit zwei unterschiedlichen Spezifitäten kann
hergestellt werden, wenn die VH-Region und die VL-Region zweier
Antikörperspezies
verwendet werden [FEBS Letters 453 (1999), 164; Int. J. Cancer 77
(1998), 763].
-
(6) Herstellung eines dsFv-Fragments
-
Das
dsFv-Fragment kann unter Verwendung von Escherichia coli genetisch
hergestellt werden. Zuerst werden DNAs hergestellt, in denen ein
codierter Aminosäurerest
durch einen Cysteinrest ersetzt ist, indem eine Mutation in geeignete
Positionen in die DNAs, welche die VH-Region und die VL-Region des
Antikörpers codieren,
wie unter Punkt 1(2), 1(4) und 1(5) beschrieben, eingeführt wird.
Die Expressionsvektoren für
die VH-Region und die VL-Region können durch die Clonierung der
jeweiligen DNAs in einen Vektor für die Expression von dsFv hergestellt
werden. Als Vektor für
die Expression von dsFv kann ein beliebiger Vektor verwendet werden,
solange wie die DNA, welche das dsFv-Fragment codiert, inseriert
und exprimiert werden kann. Beispiele schließen pULI9 [Protein Engineering
7 (1994), 697] und dergleichen ein. Die Expressionsvektoren für die VH-Region
und die VL-Region
werden in ein geeignetes Escherichia coli-Bakterium eingeschleust,
wodurch die VH-Region und die VL-Region gebildet und in einem Einschlusskörper oder
im periplasmatischen Raum angereichert werden. Die VH-Region und
die VL-Region werden aus dem Einschlusskörper oder dem periplasmatischen
Raum gewonnen und gemischt, wobei durch einen Umfaltungsprozess,
welcher im Allgemeinen bei der Herstellung eines Proteins verwendet
wird, ein dsFv-Fragment mit einer Bindungsaktivität erhalten
werden kann. Nach der Umfaltung kann das Fragment durch Ionenaustauschchromatographie, Gelfiltration
oder dergleichen weiter gereinigt werden [Protein Engineering 7
(1994), 697].
-
(7) Herstellung eines Peptids, das eine
CDR-Region umfasst
-
Das
Peptid, welches eine CDR-Region umfasst, kann durch ein chemisches
Syntheseverfahren wie Fmoc oder tBoc hergestellt werden. Auch kann
eine DNA, codierend ein Peptid, das eine CDR-Region umfasst, hergestellt
werden, wobei die erhaltene DNA in einen geeigneten Expressionsvektor
cloniert wird, um das Peptid, welches eine CDR-Region umfasst, herzustellen.
Als Vektor für
die Expression kann ein beliebiger Vektor verwendet werden, solange
wie eine DNA, codierend ein Peptid, das eine CDR-Region umfasst,
inseriert und exprimiert werden kann. Beispiele schließen pLEX
(hergestellt von Invitrogen), pAX4a+ (hergestellt von Invitrogen)
und dergleichen ein. Der Expressionsvektor wird in ein geeignetes
Escherichia coli-Bakterium eingeschleust,
so dass das Peptid, welches eine CDR-Region umfasst, in einem Einschlusskörper oder
im periplasmatischen Raum gebildet und angereichert werden kann.
Das Peptid, welches eine CDR-Region umfasst, kann aus dem Einschlusskörper oder
dem periplasmatischen Raum gewonnen und durch Ionenaustauschchromatographie,
Gelfiltration oder dergleichen weiter gereinigt werden [Protein
Engineering 7 (1994), 697].
-
3. Beurteilung der Aktivität des humanisierten
Antikörpers
oder des Antikörperfragments
-
(1) Antigenbindungsaktivität
-
Die
FGF-8-Bindungsaktivität
des hergestellten humanisierten Antikörpers oder des Antikörperfragments
kann durch einen ELISA oder mittels Oberflächenplasmonresonanz [J. Immunol.
Methods 145 (1991), 229] oder dergleichen gemessen werden.
-
(2) Inhibitionsaktivität gegenüber der Funktion von FGF-8
-
Die
Inhibitionsaktivität
des hergestellten humanisierten Antikörpers oder des Antikörperfragments
gegenüber
der Funktion von FGF-8 kann z. B. durch einen Zellwachstumstest
unter Verwendung einer Zelllinie gemessen werden. Zum Beispiel kann
die Inhibitionsaktivität
des hergestellten humanisierten Antikörpers oder des Antikörperfragments
gegenüber
der Funktion von FGF-8 durch die Züchtung der Maus-Brustkrebszelllinie SC-3
[Proc. Natl. Acad. Sci. USA 89 (1992), 8928], der Maus-Fibroblastenzelllinie
NIH3T3 oder einer menschlichen Prostatakrebs-, Brustkrebs- oder
Eierstockkrebszelllinie als Zielzellen unter Verwendung eines Mediums,
enthaltend 1 bis 100 ng/ml FGF-8 oder Testosteron, für einen
Zeitraum von 24 bis 72 Stunden nach der Zugabe des humanisierten
Antikörpers
oder des Antikörperfragments
nach serieller Verdünnung
auf eine Endkonzentration von 0,001 bis 100 μg/ml in dem Medium, und im Anschluss
daran die Messung der Anzahl der lebensfähigen Zellen unter Verwendung
einer MTT [3-(4,5-Dimethyl-2-thiazonyl)-2,5-diphenyl-2H-tetrazoliumbromid]-Lösung, eines
Zellzählungskits,
eines WST-Kits oder dergleichen gemessen werden. Die Aktivität des humanisierten
Antikörpers
oder des Antikörperfragments
im Hinblick auf die Inhibition der Bindung von FGF-8 an den Zelloberflächenrezeptor
kann durch die Bolton-Hunter-Methode unter Verwendung von FGF-8,
markiert mit 125I [Biochem. J. 133 (1973),
527], oder dergleichen bei den vorstehenden Zelllinien gemessen
werden.
-
(3) In-vivo-Antitumorwirkung
-
Die
In-vivo-Antitumorwirkung des hergestellten humanisierten Antikörpers oder
des Antikörperfragments
kann zum Beispiel unter Verwendung des nachstehend veranschaulichten
Krebszellen-Transplantationssystem für Nacktmäuse beurteilt werden.
-
a. Beurteilung der Antitumorwirkung für ein frühes Krebsmodell
-
Die
Maus-Brustkrebszelllinie SC-3 wird in einer Dichte von 1 × 107 Zellen/ml in PBS suspendiert und in einer
Dosis von 0,1 ml/Tier (1 × 106 Zellen/Tier) in 6 bis 8 Wochen alte, männliche
Nacktmäuse
subkutan transplantiert. Unmittelbar nach der Transplantation wird
der humanisierte Antikörper
oder das Antikörperfragment
in einer Dosis von 10 μg
bis 400 μg/Tier
in die Bauchhöhle
oder die Schwanzvene der Maus verabreicht. Der Antikörper wird
in einer Häufigkeit
von 1- bis 7mal pro Woche insgesamt 1- bis 10mal verabreicht. Die
Antitumorwirkung kann durch das regelmäßige Messen des Tumorvolumens
für 14
bis 60 Tage nach der Tumortransplantation und das Vergleichen des
Tumorvolumens mit demjenigen der Gruppe, an die der negative Kontrollantikörper verabreicht
wurde, oder der Gruppe, an die das Lösungsmittel verabreicht wurde,
beurteilt werden. Das Tumorvolumen (V) kann auch basierend auf der
folgenden Gleichung durch Messung der Länge (L), der Breite (W) und
der Höhe
(H) des Tumors unter Verwendung eines Messschiebers berechnet werden: V (mm3) = (L) × (W) × (H) × 0,5236
-
b. Beurteilung der Antitumorwirkung für ein fortgeschrittenes
Krebsmodell
-
SC-3-Zellen
werden in einer Dichte von 1 × 107 Zellen/ml in PBS suspendiert und in einer
Dosis von 0,1 ml/Tier (1 × 106 Zellen/Tier) in 6 bis 8 Wochen alte, männliche
Nacktmäuse
subkutan transplantiert. Wenn das Tumorvolumen etwa 100 bis 300
mm3 erreicht (ungefähr am 14. Tag nach der Tumortransplantation),
wird mit der Verabreichung des humanisierten Antikörpers oder
des Antikörperfragments
in einer Dosis von 10 μg bis
400 μg/Tier
in die Bauchhöhle
oder die Schwanzvene der Maus begonnen. Der Antikörper wird
in einer Häufigkeit
von 1- bis 7mal pro Woche insgesamt 1- bis 10mal verabreicht. Die
Antitumorwirkung kann in der gleichen Weise wie für das frühe Krebsmodell
von a. beurteilt werden.
-
(4) Verfahren zur Verwendung des humanisierten
Antikörpers
oder des Antikörperfragments
davon
-
Da
der humanisierte Antikörper
oder das Antikörperfragment
davon der vorliegenden Erfindung spezifisch an FGF-8 bindet, von
welchem angenommen wird, dass er ein Wachstumsfaktor für menschliche
Krebszellen ist, die ein Geschlechtshormon-abhängiges Wachstum zeigen, und
die Funktion von FGF-8 inhibiert, ist der humanisierte Antikörper oder
das Antikörperfragment
davon bei der Behandlung und der Diagnose von Krebserkrankungen
beim Menschen wie Prostatakrebs, Brustkrebs, Eierstockkrebs und
Hodenkrebs nützlich. Da
der Anteil der Aminosäuresequenzen,
welche von menschlichen Antikörpern
stammen, in dem humanisierten Antikörper oder dem Antikörperfragment
davon höher
ist als in Antikörpern,
welche von einem nicht-menschlichen Tier stammen, wird erwartet,
dass der humanisierte Antikörper
oder das Antikörperfragment
davon eine starke cytotoxische Aktivität im menschlichen Körper zeigt,
keine Immunogenität
hervorruft und eine lang anhaltende Wirkung hat.
-
Der
humanisierte Antikörper
oder das Antikörperfragment
davon der vorliegenden Erfindung kann allein verabreicht werden,
wobei aber im Allgemeinen bevorzugt wird, dass der humanisierte
Antikörper
oder das Antikörperfragment
in Form einer pharmazeutischen Zubereitung bereitgestellt wird,
welche durch das Mischen des humanisierten Antikörpers oder des Antikörperfragments
mit mindestens einem pharmazeutisch verträglichen Träger gemäß einem Verfahren, welches
auf dem technischen Gebiet der Pharmazie gut bekannt ist, hergestellt
wird.
-
Es
wird bevorzugt, einen Verabreichungsweg zu verwenden, der im Hinblick
auf die Durchführung
der beabsichtigten Behandlung am wirksamsten ist, wie eine orale
Verabreichung oder parenterale Verabreichung, z. B. eine intraorale
Verabreichung, tracheale Verabreichung, rektale Verabreichung, subkutane
Injektion, intramuskuläre
Injektion, intravenöse
Injektion und dergleichen. Die intravenöse Injektion wird für eine Antikörper- oder
eine Peptidformulierung bevorzugt.
-
Die
Dosierungsform schließt
Sprays, Kapseln, Tabletten, Granula, Sirupe, Emulsionen, Zäpfchen,
Injektionen, Salben, Pflaster und dergleichen ein.
-
Für die orale
Verabreichung geeignete Formulierungen schließen Emulsionen, Sirupe, Kapsein,
Tabletten, Pulver, Granula und dergleichen ein.
-
Flüssige Zubereitungen
wie Emulsionen und Sirupe können
unter Verwendung von Zusätzen
wie zum Beispiel Wasser; Sacchariden wie Saccharose, Sorbit oder
Fructose; Glykolen wie Polyethylenglykol oder Propylenglykol; Ölen wie
Sesamöl,
Olivenöl
oder Sojaöl;
Antiseptika wie p-Hydroxybenzoat; und Geschmacksstoffen wie Erdbeergeschmack
oder Pfefferminze hergestellt werden.
-
Kapseln,
Tabletten, Pulver, Granula und dergleichen können unter Verwendung von Zusätzen wie
zum Beispiel Füllstoffen
wie Lactose, Glucose, Saccharose oder Mannit; Sprengmitteln wie
Stärke
oder Natriumalginat; Gleitmitteln wie Magnesiumstearat oder Talkum;
Bindemitteln wie Polyvinylalkohol, Hydroxypropylcellulose oder Gelatine;
Tensiden wie Fettsäureestern;
und Weichmachern wie Glycerin hergestellt werden.
-
Für die parenterale
Verabreichung geeignete Formulierungen schließen Injektionen, Zäpfchen,
Sprays und dergleichen ein.
-
Injektionen
können
unter Verwendung eines Trägers
wie einer Salzlösung,
einer Glucoselösung
oder eines Gemisches davon hergestellt werden.
-
Zäpfchen können unter
Verwendung eines Trägers
wie Kakaobutter, gehärtetem
Fett oder einer Carbonsäure
hergestellt werden.
-
Sprays
können
auch aus dem Antikörper
selbst oder unter Verwendung eines Trägers oder dergleichen, welcher
die Mund- und Atemwegsschleimhaut der Patienten nicht reizt und
die Absorption des Antikörpers
oder des Antikörperfragments
davon durch das Dispergieren davon in Form von mikrofeinen Partikeln
erleichtern kann, hergestellt werden.
-
Der
Träger
schließt
Lactose, Glycerin und dergleichen ein. Abhängig von den Eigenschaften
des Antikörpers
oder des Antikörperfragments
und dem zu verwendenden Träger
können
Aerosole, trockene Pulver und dergleichen hergestellt werden. Die
für die
oralen Zubereitungen veranschaulichten Zusätze können auch zu den parenteralen
Zubereitungen zugegeben werden.
-
Die
Dosis und die Häufigkeit
der Verabreichung variieren abhängig
von der gewünschten
therapeutischen Wirkung, der Verabreichungsmethode, dem Behandlungszeitraum,
dem Alter, dem Körpergewicht
und dergleichen, wobei die Dosis im Allgemeinen für einen
Erwachsenen im Bereich von 0,01 mg/kg bis 20 mg/kg pro Tag liegt.
-
Kurze Beschreibung der Zeichnungen
-
1 zeigt
die Konstruktionsschritte für
Plasmid pKM1334CH-H5.
-
2 zeigt
die Konstruktionsschritte für
Plasmid pKM1334CH-L4.
-
3 zeigt
die Konstruktionsschritte für
Plasmid pKANTEX1334.
-
4 zeigt
die Neutralisierungsaktivitäten
für den
Anti-FGF-8-Mausanti körper
KM1334 und die chimären
Anti-FGF-8-Antikörper
KM3034 und KM3334 auf das FGF-8-abhängige Wachstum von Zellen der Maus-Brustkrebszelllinie
SC-3. Die x-Achse und die y-Achse zeigen die Antikörperkonzentration
(μg/ml)
bzw. das relative Wachstum (%), wenn für das Wachstum durch die Zugabe
von FGF-8 allein ein Wert von 100% definiert wird. "O", "☐", "Δ" und "x" zeigen
die Aktivitäten
von KM1334, KM3034, KM3334 bzw. KM2760 als eine negative Kontrolle.
-
5 zeigt
die Konstruktionsschritte für
Plasmid pKM1334HV0.
-
6 zeigt
die Konstruktionsschritte für
Plasmid pKANTEX1334HV0LV0.
-
7 zeigt
die Ergebnisse der ELISA-Messung der FGF-8-Bindungsaktivitäten des
chimären
Anti-FGF-8-Antikörpers
KM3334 und der Anti-FGF-8-CDR-transplantierten
Antikörper
HV0LV0, HV0LV6 und HV6LV6. Die x-Achse und die y-Achse zeigen die
Antikörperkonzentration
(μg/ml)
bzw. die Bindungsaktivität (OD415). "O", "☐", "Δ" und "x" zeigen
die Aktivitäten
von KM3334, HV0LV0, HV0LV6 bzw. HV6LV6.
-
8 zeigt
die Ergebnisse der BIAcore-2000-Messung der FGF-8-Bindungsaktivitäten des
chimären Anti-FGF-8-Antikörpers KM3334
und der Anti-FGF-8-CDR-transplantierten
Antikörper
HV0LV0, HV0LV6 und HV6LV6. Die x-Achse und die y-Achse zeigen die
Zeit (Sekunden) bzw. das Resonanzsignal (RU).
-
9 zeigt
die Konstruktionsschritte für
Plasmid pKM1334LV3-1.
-
10 zeigt die Konstruktionsschritte für Plasmid
pKM1334LV4-2.
-
11 zeigt die Konstruktionsschritte für Plasmid
pKM1334LV3-2.
-
12 zeigt die Ergebnisse der BIAcore-2000-Messung
der FGF-8-Bindungsaktivitäten
des chimären Anti-FGF-8-Antikörpers KM3034
und der Anti-FGF-8-CDR-transplantierten
Antikörper
HV0LV0/CHO, HV0LV2-1/CHO, HV0LV2-2/CHO, HV0LV3-1/CHO, HV0LV3-2/CHO,
HV0LV4-1/CHO, HV0LV4-2/CHO und HV0LV6/CHO. Die x-Achse und die y-Achse
zeigen die Zeit (Sekunden) bzw. das Resonanzsignal (RU).
-
13 zeigt die Neutralisierungsaktivitäten für den chimären Anti-FGF-8-Antikörper KM3034
und die Anti-FGF-8-CDR-transplantierten Antikörper HV0LV2-1/CHO, HV0LV3-1/CHO,
HV0LV4-1/CHO und HV0LV6/CHO auf das FGF-8-abhängige Wachstum von Zellen der
Maus-Brustkrebszelllinie SC-3. Die x-Achse und die y-Achse zeigen
die Antikörperkonzentration
(μg/ml)
bzw. das relative Wachstum (%), wenn für das Wachstum durch die Zugabe
von FGF-8 allein ein Wert von 100% definiert wird. "O", "☐", "Δ", "♢", "
" und "x" zeigen die Aktivitäten von KM3034, HV0LV2-1/CHO,
HV0LV3-1/CHO, HV0LV4-1/CHO, HV0LV6/CHO bzw. KM2760 als eine negative
Kontrolle.
-
14 zeigt die Konstruktionsschritte für Plasmid
pKM1334LV3-3.
-
15 zeigt die Ergebnisse der BIAcore-2000-Messung
der FGF-8-Bindungsaktivitäten
des chimären Anti-FGF-8-Antikörpers KM3034
und der Anti-FGF-8-CDR-transplantierten
Antikörper
HV0LV3-1/CHO, HV0LV3-3/CHO, HV0LV4-3/CHO und HV0LV6/CHO. Die x-Achse
und die y-Achse zeigen die Zeit (Sekunden) bzw. das Resonanzsignal
(RU).
-
16 zeigt die Reutralisierungsaktivitäten für den chimären Anti-FGF-8-Antikörper KM3034
und die Anti-FGF-8-CDR-transplantierten Antikörper HV0LV3-1/CHO, HV0LV3-3/CHO,
HV0LV4-3/CHO und HV0LV6/CHO auf das FGF-8-abhängige Wachstum von Zellen der
Maus-Brustkrebszelllinie SC-3. Die x-Achse und die y-Achse zeigen
die Antikörperkonzentration
(μg/ml)
bzw. das relative Wachstum (%), wenn für das Wachstum durch die Zugabe
von FGF-8 allein ein Wert von 100% definiert wird. "O", "Δ", "∎",
und "x" zeigen die Aktivitäten von KM3034, HV0LV3-1/CHO,
HV0LV3-3/CHO, HV0LV4-3/CHO, HV0LV6/CHO bzw. KM2760 als eine negative
Kontrolle.
-
Die
vorliegende Erfindung wird nachstehend unter Bezugnahme auf Beispiele
erläutert,
jedoch ist der Schutzumfang der vorliegenden Erfindung nicht darauf
begrenzt.
-
Beste Art der Durchführung der
Erfindung
-
Beispiel 1
-
Herstellung eines chimären Anti-FGF-8-Antikörpers:
-
1. Isolierung und Analyse der cDNA, codierend
die V-Region eines Anti-FGF-8-Mausantikörpers
-
(1) Präparation
der mRNA aus Hybridomzellen, die einen Anti-FGF-8-Mausantikörper herstellen
-
Aus
1 × 107 Zellen des Hybridoms KM1334 (FERM BP-5451),
das in der Lage ist, einen Mausantikörper gegen FGF-8 herzustellen
(nachstehend bezeichnet als "Anti-FGF-8-Mausantikörper"), wurden unter Verwendung
des mRNA-Präparationskits
Fast Track mRNA-Isolationskits (hergestellt von Invitrogen) gemäß den beiliegenden
Anweisungen des Herstellers etwa 8 μg mRNA präpariert.
-
(2) Herstellung von cDNA-Banken der H-Kette
und der L-Kette des Anti-FGF-8-Mausantikörpers
-
Aus
5 μg der
mRNA von KM1334, welche unter Punkt 1(1) in Beispiel 1 erhalten
wurde, wurde unter Verwendung des Time Saver cDNA-Synthesekits (hergestellt
von Amersham Pharmacia Biotech) gemäß den beiliegenden Anweisungen
des Herstellers eine cDNA mit einem EcoRI-NotI-Adapter an beiden
Enden synthetisiert. Anschließend
wurden die cDNA-Banken unter Verwendung des λZAPII-Clonierungskits (hergestellt von
Stratagene) hergestellt. Zuerst wurde die Gesamtmenge der cDNA in
20 μl sterilem
Wasser gelöst
und dann durch eine Agarosegelelektrophorese fraktioniert, wobei
jeweils etwa 0,1 μg
eines cDNA-Fragments von etwa 1,5 kb, entsprechend der H-Kette eines
Antikörpers
der IgG-Klasse, und eines cDNA-Fragments
von etwa 1,0 kb, entsprechend der L-Kette der κ-Klasse, gewonnen wurden. Anschließend wurden
jeweils 0,1 μg des
cDNA-Fragments von etwa 1,5 kb und 0,1 μg des cDNA-Fragments von etwa
1,0 kb mit 1 μg
des λZAPII-Vektors,
dessen Enden nach der Spaltung mit dem Restriktionsenzym EcoRI mit
alkalischer Phosphatase aus Kälberdarm
dephosphoryliert worden waren, gemäß den beiliegenden Anweisungen
des Herstellers ligiert.
-
Unter
Verwendung von Gigapack II Packaging Extracts Gold (hergestellt
von Stratagene) wurden gemäß den beiliegenden
Anweisungen des Herstellers 4 μl
jeder Reaktionslösung
nach der Ligierung in einem λ-Phagen
verpackt, und eine geeignete Menge davon wurde mittels Infektion
in Zellen von Escherichia coli XL1-Blue [Biotechniques 5 (1987), 376] eingebracht,
wobei etwa 8,1 × 104 und 5,5 × 104 Phagen-Clone
als cDNA-Bank für
die H-Kette bzw. cDNA-Bank für
die L-Kette von KM1334 erhalten wurden. Anschließend wurde jeder Phage gemäß dem üblichen
Verfahren [Molecular Cloning: A Laborstory Manual, Cold Spring Harbor Lab.
Press, New York (1989)] auf einer Nylonmembran fixiert.
-
(3) Clonierung der cDNA für die H-Kette
und die L-Kette des Anti-FGF-8-Mausantikörpers
-
Unter
Verwendung von ECL Direct Nucleic Acid Labelling and Detection Systems
(hergestellt von Amersham Pharmacia Biotech) wurden gemäß den beiliegenden
Anweisungen des Herstellers die Clone der cDNA-Bank für die H-Kette
und der cDNA-Bank für
die L-Kette von KM1334 auf der unter Punkt 1(2) in Beispiel 1 hergestellten
Nylonmembran unter Verwendung einer cDNA für die C-Region des Mausantikörpers [wobei die
H-Kette ein DNA-Fragment ist, das die Maus-Cγ1-cDNA enthält [J. Immunol. 146 (1991),
2010], und wobei die L-Kette ein DNA-Fragment ist, das die Maus-Cκ-cDNA enthält [Cell
22 (1980), 197]) als Sonde nachgewiesen, wobei jeweils 10 Phagen-Clone
als Clone für
die H-Kette und die L-Kette erhalten wurden, die eine starke Reaktion
mit der Sonde zeigten. Anschließend
wurde gemäß den beiliegenden
Anweisungen des Herstellers des λZAPII-Clonierungskits
(hergestellt von Stratagene) jeder Phagen-Clon durch ein in-vivo-Exzisionsverfahren
in ein Plasmid überführt. Die
Nucleotidsequenz der cDNA, die in jedem der auf diese Weise erhaltenen Plasmide
enthalten war, wurde mittels der Didesoxy-Methode [Molecular Cloning:
A Laborstory Manual, Cold Spring Harbor Lab. Press, New York (1989)]
unter Verwendung des Big Dye Terminator-Kits, Ver. 2 (hergestellt von
Applied Biosystems) bestimmt. Als Ergebnis wurde das Plasmid pKM1334H7-1,
enthaltend die vollständige
cDNA der funktionellen H-Kette, und das Plasmid pKM1334L7-1, enthaltend
die vollständige
cDNA der funktionellen L-Kette erhalten, wobei die cDNA am 5'-Ende die Sequenz
ATG enthält,
welche als ein Initiationscodon angesehen wird.
-
(4) Analyse der Aminosäuresequenz der V-Region des
Anti-FGF-8-Mausantikörpers
-
Die
in dem Plasmid pKM1334H7-1 enthaltene vollständige Nucleotidsequenz der
VH-Region ist in SEQ ID NO 1 dargestellt, und die davon abgeleitete
Aminosäuresequenz
ist in SEQ ID NO 2 dargestellt. Die in dem Plasmid pKM1334-L7-1 enthaltene vollständige Nucleotidsequenz
der VL-Region ist in SEQ ID NO 3 dargestellt, und die davon abgeleitete
vollständige
Aminosäuresequenz
ist in SEQ ID NO 4 dargestellt. Basierend auf einem Vergleich mit
Sequenzdaten von bekannten Mausantikörpern [Sequences of Proteins
of Immunological Interest, US Dept. Health and Human Services (1991)]
und einem Vergleich mit den Ergebnissen der Analyse der N-terminalen
Aminosäuresequenzen
der H-Kette und der L-Kette des gereinigten Anti-FGF-8-Mausantikörpers KM1334,
welche mit Hilfe eines automatischen Edman-Abbaus unter Verwendung des Protein-Sequenzierungsautomats
PPSQ-10 (hergestellt von Shimadzu Corporation) durchgeführt wurde, wurde
festgestellt, dass jede der auf diese Weise isolierten cDNA eine
cDNA vollständiger
Länge ist,
welche den Anti-FGF-8-Mausantikörper
KM1334, enthaltend eine sekretorische Signalsequenz, codiert, wobei
die Aminosäuresequenz
von Position 1 bis 19 in der H-Kette, dargestellt in SEQ ID NO 2,
und die Aminosäuresequenz
von Position 1 bis 19 in der L-Kette, dargestellt in SEQ ID NO 4,
die sekretorischen Signalsequenzen darstellen. Die Aminosäuresequenzen
der VH-Region und der VL-Region, mit Ausnahme der sekretorischen Signalsequenz,
sind in SEQ ID NO 40 bzw. SEQ ID NO 41 dargestellt.
-
Anschließend wurde
die Neuheit der Aminosäuresequenzen
der VH-Region und der VL-Region des Anti-FGF-8-Mausantikörpers KM1334
untersucht. Mit Hilfe des GCG-Pakets (Version 9.1, entwickelt von
der Genetics Computer Group) als Sequenzanalyseprogramm wurde eine
Aminosäuresequenz-Datenbank
von bekannten Proteinen [PIR-Protein (Release 56.0)] nach der BLAST-Methode
[J. Mol. Biol. 215 (1990), 403] abgefragt. Als Ergebnis wurden im
Hinblick auf sowohl die H-Kette als auch die L-Kette keine vollständig identischen
Sequenzen gefunden, wodurch bestätigt
wurde, dass die VH-Region und die VL-Region des Anti-FGF-8-Mausantikörpers KM1334
neue Aminosäuresequenzen
sind.
-
Ferner
wurden die CDR-Regionen der VH-Region und der VL-Region des Anti-FGF-8-Mausantikörpers KM1334
durch einen Vergleich mit den Aminosäuresequenzen von bekannten
Antikörpern
identifiziert. Die Aminosäuresequenzen
von CDR1, -2 und -3 der VH-Region des Anti-FGF-8-Mausantikörpers KM1334 sind
in SEQ ID NO 5, 6 bzw. 7 dargestellt, und die Aminosäuresequenzen
von CDR1, -2 und -3 der VL-Region sind in SEQ ID NO 8, 9 bzw. 10
dargestellt.
-
2. Stabile Expression eines chimären Anti-FGF-8-Antikörpers unter
Verwendung von tierischen Zellen
-
(1) Konstruktion eines Plasmids, enthaltend
die cDNA für
die VH-Region des chimären
Anti-FGF-8-Antikörpers
-
Unter
Verwendung von 50 ng des Plasmids pKM1334H7-1, das unter Punkt 1(3)
von Beispiel 1 erhalten wurde, als Matrize und synthetischen DNA-Fragmenten
mit den in SEQ ID NO 11 und 12 dargestellten Nucleotidsequenzen
(hergestellt von GENSET), welche in einer Endkonzentration von 0,3 μMol/l als
Primer zugegeben wurden, wurde eine PCR, umfassend zunächst das
Erhitzen eines Gesamtvolumens von 50 μl des Gemisches bei 94°C für 2 Minuten
und dann 30 Reaktionszyklen bei 94°C für 15 Sekunden, 57°C für 30 Sekunden
und 68°C
für 1 Minute
für einen
Zyklus, gemäß den Anweisungen
des Herstellers, welche der KOD Plus-Polymerase (hergestellt von
TOYOBO) beiliegen, durchgeführt.
Das Reaktionsprodukt wurde gereinigt, in sterilem Wasser gelöst und dann
mit 10 Einheiten des Restriktionsenzyms EcoRI (hergestellt von Takara Shuzo)
bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
0,3 μg eines
EcoRI-Fragments von etwa 0,48 kb (wobei das 5'-Ende ein EcoRI-Ende ist und das 3'-Ende ein glattes Ende ist) gewonnen
wurden.
-
Anschließend wurden
3 μg des
Plasmids pBluescript SK(–)
mit 10 Einheiten des Restriktionsenzyms EcoRI (hergestellt von Takara
Shuzo) und 10 Einheiten des Restriktionsenzyms EcoRV (hergestellt
von Takara Shuzo) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
2 μg eines
EcoRI-EcoRV-Fragments von etwa 2,95 kb gewonnen wurden.
-
Im
Anschluss daran wurden 0,1 μg
des EcoRI-Fragments der cDNA für
die VH-Region und 0,1 μg
des EcoRI-EcoRV-Fragments des Plasmids pBluescript SK(–), welches
vorstehend erhalten wurde, zu einem Gesamtvolumen von 10 μl sterilem
Wasser zugegeben und unter Verwendung von Ligation High (hergestellt
von TOYOBO) ligiert. Unter Verwendung der so erhaltenen rekombinanten
Plasmid-DNA- Lösung wurden
Zellen von E. coli XL-1 Blue transformiert, wobei das in 1 dargestellte
Plasmid pKM1334CH-H5 erhalten wurde, welches die cDNA der VH-Region
des chimären
Anti-FGF-8-Antikörpers
enthält.
-
(2) Konstruktion eines Plasmids, enthaltend
die cDNA für
die VL-Region des chimären
Anti-FGF-8-Antikörpers
-
Unter
Verwendung von 50 ng des Plasmids pKM1334L7-1, das unter Punkt 1(3)
von Beispiel 1 erhalten wurde, als Matrize und synthetischen DNA-Fragmenten
mit den in SEQ ID NO 13 und 14 dargestellten Nucleotidsequenzen
(hergestellt von GENSET), welche in einer Endkonzentration von 0,3 μMol/l als
Primer zugegeben wurden, wurde eine PCR, umfassend zunächst das
Erhitzen eines Gesamtvolumens von 50 μl des Gemisches bei 94°C für 2 Minuten
und dann 30 Reaktionszyklen bei 94°C für 15 Sekunden, 57°C für 30 Sekunden
und 68°C
für 1 Minute
für einen
Zyklus, gemäß den Anweisungen
des Herstellers, welche der KOD Plus-Polymerase (hergestellt von
TOYOBO) beiliegen, durchgeführt.
Das Reaktionsprodukt wurde gereinigt, in sterilem Wasser gelöst und dann
mit 10 Einheiten des Restriktionsenzyms EcoRI (hergestellt von Takara Shuzo)
bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
0,3 μg eines
EcoRI-Fragments von etwa 0,45 kb (wobei das 5'-Ende das EcoRI-Ende ist und das 3'-Ende ein glattes
Ende ist) gewonnen wurden.
-
Anschließend wurden
0,1 μg des
EcoRI-Fragments der cDNA für
die VL-Region und 0,1 μg
des EcoRI-EcoRV-Fragments des Plasmids pBluescript SK(–), welche
jeweils vorstehend erhalten wurden, zu einem Gesamtvolumen von 10 μl sterilem
Wasser zugegeben und unter Verwendung von Ligation High (hergestellt von
TOYOBO) ligiert. Unter Verwendung der so erhaltenen rekombinanten
Plasmid-DNA-Lösung wurden
Zellen von E. coli XL-1 Blue transformiert, wobei das in 2 dargestellte
Plasmid pKM1334CH-L4 erhalten wurde, das die cDNA der VL-Region
des chimären
Anti-FGF-8-Antikörpers
enthält.
-
(3) Konstruktion des Expressionsvektors
pKANTEX1334 für
den chimären
Anti-FGF-8-Antikörper
-
Unter
Verwendung des Expressionsvektors pKANTEX93 für einen humanisierten Antikörper, beschrieben
in
WO 97/10354 , und
der Plasmide pKM1334CH-H5 und pKM1334CH-L4, die unter Punkt 2(1)
und 2(2) von Beispiel 1 erhalten wurden, wurde der Expressionsvektor
pKANTEX1334 für
einen chimären
Anti-FGF-8-Antikörper
wie folgt konstruiert.
-
Zunächst wurden
3 μg des
Plasmids pKM1334CH-H5, das unter Punkt 2(1) von Beispiel 1 erhalten wurde,
mit 10 Einheiten des Restriktionsenzyms NotI (hergestellt von New
England Biolabs) und 10 Einheiten des Restriktionsenzyms ApaI (hergestellt
von Takara Shuzo) gemischt und dann bei 37°C für 1 Stunde umgesetzt. Die Reaktionslösung wurde
durch eine Agarosegelelektrophorese fraktioniert, wobei etwa 0,2 μg eines NotI-Apal-Fragments
von etwa 0,48 kb gewonnen wurden.
-
Anschließend wurden
3 μg des
Expressionsvektors pKANTEX93 für
einen humanisierten Antikörper mit
10 Einheiten des Restriktionsenzyms ApaI (hergestellt von Takara
Shuzo) und 10 Einheiten des Restriktionsenzyms NotI (hergestellt
von New England Biolabs) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung wurde
durch eine Agarosegelelektrophorese fraktioniert, wobei etwa 2 μg eines ApaI-Notl-Fragments von etwa 12,8
kb gewonnen wurden.
-
Im
Anschluss daran wurden 0,1 μg
des NotI-ApaI-Fragments des Plasmids pKM1334CH-H5 und 0,1 μg des ApaI-NotI-Fragments
des Plasmids pKANTEX93 zu einem Gesamtvolumen von 10 μl sterilem
Wasser zugegeben und unter Verwendung von Ligation High (hergestellt
von TOYOBO) ligiert. Unter Verwendung der so erhaltenen rekombinanten
Plasmid-DNA-Lösung
wurden Zellen von E. coli XL-1 Blue transformiert, wobei das in 3 dargestellte
Plasmid pKANTEX1334H erhalten wurde.
-
Anschließend wurden
3 μg des
Plasmids pKM1334CH-L4, das unter Punkt 2(2) von Beispiel 1 erhalten wurde,
mit 10 Einheiten des Restriktionsenzyms EcoRI (hergestellt von Takara
Shuzo) und 10 Einheiten des Restriktionsenzyms BsiWI (hergestellt
von New England Biolabs) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
0,2 μg eines
EcoRI-BsiWI-Fragments von etwa 0,45 kb gewonnen wurden.
-
Im
Anschluss daran wurden 3 μg
des Plasmids pKANTEX1334H, welches vorstehend erhalten wurde, mit
10 Einheiten des Restriktionsenzyms EcoRI (hergestellt von Takara
Shuzo) und 10 Einheiten des Restriktionsenzyms BsiWI (hergestellt
von New England Biolabs) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
2 μg eines
EcoRI-BsiWI-Fragments von etwa 13,30 kb gewonnen wurden.
-
Anschließend wurden
0,1 μg des
EcoRI-BsiWI-Fragments des Plasmids pKM1334CH-L4 und 0,1 μg des EcoRI-BsiWI-Fragments
des Plasmids pKANTEX-1334H
zu einem Gesamtvolumen von 10 μl
sterilem Wasser zugegeben und unter Verwendung von Ligation High
(hergestellt von TOYOBO) ligiert. Unter Verwendung der so erhaltenen
rekombinanten Plasmid-DNA-Lösung
wurden Zellen von E. coli XL-1 Blue transformiert, wobei das in 3 dargestellte
Plasmid pKANTEX1334 erhalten wurde.
-
Unter
Verwendung von 400 ng des auf diese Weise erhaltenen Plasmids wurde
eine Analyse der Nucleotidsequenz des Plasmids durch die Didesoxy-Methode
[Molecular Cloning: A Laborstory Manual, Cold Spring Harbor Lab.
Press, New York (1989)] unter Verwendung des Big Dye Terminator-Kits,
Ver. 2 (hergestellt von Applied Biosystems) durchgeführt, wodurch
bestätigt
wurde, dass ein Plasmid erhalten wurde, in welches die DNA von Interesse
cloniert worden war.
-
(4) Stabile Expression des chimären Anti-FGF-8-Antikörpers unter
Verwendung von CHO/DG44-Zellen
-
Unter
Verwendung des Expressionsvektors pKANTEX1334 für den chimären Anti-FGF-8-Antikörper, welcher
unter Punkt (2(3) in Beispiel 1 erhalten wurde, wurde die Expression
des chimären
Anti-FGF-8-Antikörpers
in CHO/DG44-Zellen [Proc. Natl. Acad. Sci. USA 77 (1980), 4216]
wie folgt bewirkt.
-
Nach
dem Einbringen von 10 μg
des Plasmids pKANTEX1334 in 1,6 × 106 Zellen
von CHO/DG44-Zellen durch Elektroporation [Cytotechnology 3 (1990),
133] wurden die Zellen in 10 bis 30 ml IMDM-1xHT-Ergänzungslösung-dFBS(10)
[IMDM-Medium, enthaltend
10% dialysiertes fötales
Rinderserum (dFBS) und 1xHT-Ergänzungslösung (jeweils
hergestellt von Gibco)] suspendiert und zu 100 μl/Vertiefung in einer Mikrotiterplatte
mit 96 Vertiefungen (hergestellt von IWAKI) dispensiert. Nach Züchten bei
37°C für 24 Stunden
in einem Brutschrank mit 5% CO2 wurde das
Kulturmedium durch IMDM-dFBS(10), welches dadurch hergestellt wurde,
dass das 1xHT-Ergänzungslösung allein
weggelassen wurde, ausgetauscht, gefolgt von einem Züchtungszeitraum
von 1 bis 2 Wochen. Die Kulturüberstände wurden
aus Vertiefungen gewonnen, in denen resistente Kolonien wuchsen
und konfluent wurden, und die Antigenbindungsaktivität der chimären Anti-FGF-8-Antikörper in
den Überständen wurde
durch den ELISA gemessen, welcher unter Punkt 2(6) von Beispiel
1 erläutert
wird.
-
Zur
Erhöhung
der Expressionsmenge des Antikörpers
unter Verwendung des dhfr-Genamplifikationssystems wurden die Transformanten
aus Vertiefungen, für
die eine Expression von chimären
Anti-FGF-8-Antikörpern
in den Kulturüberständen nachgewiesen
wurde, auf eine Platte mit 24 Vertiefungen überimpft und während 2
Wochen in IMDM-dFBS(10)-Medium, enthaltend 50 nMol/l Methotrexat
(MTX; hergestellt von Sigma), welches ein Inhibitor des dhfr-Genprodukts
Dihydrofolatreduktase ist, gezüchtet.
Die MTX-Konzentration wurde auf 200 nMol/l und dann auf 500 nMol/l
weiter erhöht,
gefolgt von einem Züchtungszeitraum
von 2 Wochen für
jeden Erhöhungsschritt,
um Transformanten zu induzieren, welche eine Resistenz gegen 500
nMol/l MTX zeigen. Wenn die Transformanten in den Vertiefungen konfluent
wurden, wurde die Antigenbindungsaktivität der chimären Anti-FGF-8-Antikörper in
den Überständen durch
den ELISA gemessen, welcher unter Punkt 2(6) von Beispiel 1 erläutert wird.
Schließlich
wurden Transformanten erhalten, welche in der Lage sind, in dem
IMDM-dFBS(10)-Medium, enthaltend 500 nMol/l MTX, zu wachsen und
eine starke Expression der chimären
Anti-FGF-8-Antikörper
zeigen. Unter Verwendung der so erhaltenen Transformanten wurde
mit Hilfe des Grenzverdünnungsverfahrens
eine Einzelzellisolierung (Clonierung) durchgeführt, und ein transformierter Zellclon,
welche die höchste
Expression des chimären
Anti-FGF-8-Antikörpers
zeigte, wurde als KM3034 bezeichnet. KM3034 wurde am 26. Dezember
2001 auch als FERM BP-7836 bei der International Patent Organism
Depositary, National Institute of Advanced Industrial Science and
Technology (AIST Tsukuba Central 6, 1–1, Higashi 1-Chorre Tsukuba-shi,
Ibaraki-ken 305–8566,
Japan), hinterlegt.
-
(5) Stabile Expression des chimären Anti-FGF-8-Antikörpers unter
Verwendung von YB2/0-Zellen
-
Unter
Verwendung des Expressionsvektors pKANTEX1334 für den chimären Anti-FGF-8-Antikörper, welcher
unter Punkt 2(3) von Beispiel 1 erhalten wurde, wurde die Expression
des chimären
Anti-FGF-8-Antikörpers
in YB2/0-Zellen (ATCC CRL-1662) wie folgt bewirkt.
-
Nach
dem Einbringen von 10 μg
des Plasmids pKANTEX1334 in 4 × 106 Zellen von YB2/0-Zellen (ATCC CRL-1662)
durch Elektroporation [Cytotechnology 3 (1990), 133] wurden die
Zellen in 40 ml Hybridoma-SFM-FBS(5) [Hybridoma-SFM-Medium (hergestellt
von Gibco), enthaltend 5% fötales
Rinderserum (FBS; hergestellt von PAA Laborstories)] suspendiert
und zu 200 μl/Vertiefung
in einer Kulturplatte mit 96 Vertiefungen (hergestellt von Sumitomo
Bakelite) dispensiert. Nach Züchten
bei 37°C
für 24
Stunden in einem Brutschrank mit 5% CO2 wurde
G418 in einer Konzentration von 1 mg/ml zugegeben, gefolgt von einem
Züchtungszeitraum
von 1 bis 2 Wochen. Die Kulturüberstände wurden
aus Vertiefungen gewonnen, in denen Kolonien von Transformanten
mit einer G418-Resistenz wuchsen, und deren Vermehrung wurde bestätigt, und
die Antigenbindungsaktivität
der chimären
Anti-FGF-8-Antikörper
in den Überständen wurde
durch den ELISA gemessen, welcher unter Punkt 2(6) von Beispiel
1 erläutert
wird.
-
Zur
Erhöhung
der Expressionsmenge des Antikörpers
unter Verwendung des dhfr-Genamplifikationssystems wurden die Transformanten
aus Vertiefungen, für
die eine Expression von chimären
Anti-FGF-8-Antikörpern
in den Kulturüberständen nachgewiesen
wurde, in Hybridoma-SFM-FBS(5)-Medium, enthaltend 1 mg/ml G418 und
50 nMol/l MTX (hergestellt von Sigma) in einer Dichte von 1–2 × 105 Zellen/ml suspendiert und zu 1 ml in einer
Platte mit 24 Vertiefungen (hergestellt von Greiner) dispensiert.
Durch Züchtung
bei 37°C für 1 bis
2 Wochen in einem Brutschrank mit 5% CO2 wurden
Transformanten induziert, welche eine Resistenz gegen 50 nMol/l
MTX zeigen. Die Antigenbindungsaktivität der chimären Anti-FGF-8-Antikörper in
den Kulturüberständen aus
Vertiefungen, in denen eine Vermehrung der Transformanten beobachtet
wurde, wurde durch den ELISA gemessen, welcher unter Punkt 2(6)
von Beispiel 1 erläutert
wird.
-
Im
Hinblick auf die Transformanten aus Vertiefungen, für die eine
Expression der chimären
Anti-FGF-8-Antikörper
in den Kulturüberständen nachgewiesen
wurde, wurde die MTX-Konzentration nach der gleichen Methode erhöht, wie
vorstehend beschrieben, wobei die Transformante 5-D erhalten wurde,
welche in der Lage ist, in dem Hybridoma-SFM-FBS(5)-Medium, enthaltend
Endkonzentrationen von 1 mg/ml G418 und 200 nMol/l MTX, zu wachsen
und eine starke Expression des chimären Anti-FGF-8-Antikörpers zeigt.
Die so erhaltene Transformante wurde mit Hilfe des Grenzverdünnungsverfahrens
einer Clonierung unterzogen, wobei eine transformierte Zelllinie
erhalten wurde, welche die höchste
Expression des chimären Anti-FGF-8-Antikörpers zeigte.
Die so erhaltene transformierte Zelle wurde als KM3334 bezeichnet.
-
(6) Bindungsaktivität des Antikörpers gegenüber dem FGF-8-Peptid (ELISA)
-
Verbindung
1 (SEQ ID NO: 15) wurde als das menschliche FGF-8-Peptid ausgewählt, welches
mit dem Anti-FGF-8-Antikörper
reagieren kann. Für
die Verwendung bei einer Aktivitätsmessung
durch einen ELISA wird ein Konjugat davon mit Rinderserumalbumin
(BSA; hergestellt von Nacalai Tesque) hergestellt und als Antigen
verwendet. Hierfür
wurden 100 μl
einer Lösung
von 25 mg/ml SMCC [4-(N-Maleimidomethyl)cyclohexan-1-carbonsäure-N-Hydroxysuccinimidester]
(hergestellt von Sigma) und DMSO unter Rühren zu 900 μl einer PBS-Lösung, enthaltend
10 mg BSA, zugetropft, gefolgt von langsamem Rühren für 30 Minuten. Nach dem Aufgeben
von 1 ml der Reaktionslösung
auf eine Gelfiltrationssäule
wie eine NAP-10-Säule, welche
mit 25 ml PBS äquilibriert
worden war, wurde das Eluat, welches mit 1,5 ml PBS eluierte, als
BSA-SMCC-Lösung
verwendet. Die BSA-Konzentration einer jeden Fraktion wurde bezogen
auf die Absorption bei 280 nm gemessen. Anschließend wurden 200 μl DMSO zu
1,0 mg der Verbindung 1 zugegeben, welche dann durch Zugabe von 800 μl PBS vollständig gelöst wurde.
Im Anschluss daran wurde die vorstehende BSA-SMCC-Lösung (2,5
mg BSA) unter Rühren
zugegeben, gefolgt von langsamen Rühren bei Raumtemperatur für 3 Stunden.
Die Reaktionslösung
wurde über
Nacht bei 4°C
gegen PBS dialysiert. Anschließend
wurde Natriumazid in einer Endkonzentration von 0,05% zugegeben.
Das Gemisch wurde durch einen 0,22 μm-Filter gefiltert, und das
Filtrat wurde als BSA-Verbindung 1-Lösung verwendet.
-
Das
vorstehend hergestellte Konjugat wurde in 50 μ/Vertiefung in einer ELISA-Platte
mit 96 Vertiefungen (hergestellt von Greiner) in einer Konzentration
von 0,5 bis 1,0 μg/ml
dispensiert und über
Nacht bei 4°C stehen
gelassen, um eine Adsorption zu ermöglichen. Nach dem Waschen mit
PBS wurde eine PBS-Lösung, enthaltend
1% BSA (BSA-PBS) in 100 μ/Vertiefung
zugegeben und bei Raumtemperatur für 1 Stunde reagieren gelassen,
um verbleibende aktive Gruppen zu blockieren. Nach dem Waschen einer
jeden Vertiefung mit einer PBS-Lösung,
enthaltend 0,05% Tween (Tween-PBS), wurden die Kulturüberstände von
Transformanten oder gereinigte Antikörper in 50 μ/Vertiefung zugegeben und bei
Raumtemperatur für 1
Stunde reagieren gelassen. Nach der Reaktion wurde jede Vertiefung
mit Tween-PBS gewaschen.
Anschließend
wurde die Lösung eines
Peroxidase-markierten Ziege-anti-Mensch-IgG (H & L)-Antikörpers (hergestellt von American
Qualex), der mit BSA-PBS um das 3000- bis 6000fache verdünnt wurde,
als eine Sekundärantikörperlösung in
50 μ/Vertiefung
zugegeben und bei Raumtemperatur für 1 Stunde reagieren gelassen.
Nach der Reaktion und dem anschließenden Waschen mit Tween-PBS
wurde eine ABTS-Substratlösung
[eine Lösung
hergestellt durch Lösen
von 0,55 g 2,2'-Azinobis(3-ethylbenzothiazolin-6-sulfonsäure)ammonium
in 1 l 0,1 M Citratpuffer (pH 4,2) und Zugeben von 1 μl/ml Wasserstoffperoxid
unmittelbar vor der Verwendung) in 50 μ/Vertiefung für eine Farbentwicklung
zugegeben, und die Reaktion wurde durch Zugabe einer 5%igen SDS-Lösung in
50 μ/Vertiefung gestoppt.
Anschließend
wurde die Absorption bei 415 nm (OD415) gemessen.
-
3. Reinigung des chimären Anti-FGF-8-Antikörpers
-
(1) Züchtung
der von CHO/DG44-Zellen abstammenden Expressionszellen und Reinigung
des Antikörpers
-
Die
den chimären
Anti-FGF-8-Antikörper
exprimierende transformierte Zelllinie KM3034, welche unter Punkt
2(4) von Beispiel 1 erhalten wurde, wurde in IMDM-dFBS(10)-Medium,
enthaltend 500 nMol/MTX, in einer Dichte von 1–2 × 105 Zellen/ml
suspendiert und in 40 ml in einer 175 cm2-Flasche
(hergestellt von Greiner) dispensiert. Wenn die Zellen durch das
Züchten
in einem Brutschrank mit 5% 002 bei 37°C für 5 bis 7 Tage konfluent wurden,
wurde der Kulturüberstand
verworfen, und die Zellen wurden mit 20 ml PBS gewaschen. Nach dem
Verwerfen der PBS-Lösung und
der anschließenden
Zugabe von 40 ml EXCELL301-Medium (hergestellt von JRH) wurden die
Zellen in einem Brutschrank mit 5% CO2 bei
37°C für 7 bis
14 Tage gezüchtet, und
anschließend
wurde der Kulturüberstand
gewonnen. Unter Verwendung einer Prosep-A-Säule (hergestellt von Millipore)
wurde der chimäre
Anti-FGF-8-Antikörper gemäß den beiliegenden
Anweisungen des Herstellers aus dem Kulturüberstand gereinigt. Der so
erhaltene chimäre
Anti-FGF-8-Antikörper
wurde als KM3034 bezeichnet.
-
(2) Züchtung
der von YB2/0-Zellen abstammenden Expressionszellen und Reinigung
des Antikörpers
-
Die
den chimären
Anti-FGF-8-Antikörper
exprimierende transformierte Zelllinie KM3334, welche unter Punkt
2(5) von Beispiel 1 erhalten wurde, wurde in Hybridoma-SFM-Medium
(hergestellt von Gibco), enthaltend 200 nMol/ MTX und Daigo's GF21 (hergestellt
von Wako Pure Chemical Industries) in einer Konzentration von 5%,
bei 37°C
in einem Brutschrank mit 5% CO2 unter Verwendung
einer 175 cm2-Flasche (hergestellt von Greiner)
gezüchtet.
Unter Verwendung einer Prosep-A-Säule (hergestellt von Millipore)
wurde der chimäre
Anti-FGF-8-Antikörper
gemäß den beiliegenden
Anweisungen des Herstellers aus dem Kulturüberstand gereinigt, welcher
nach einem Züchtungszeitraum
von 8 bis 10 Tagen gewonnen worden war. Der so erhaltene chimäre Anti-FGF-8-Antikörper wurde
als KM3334 bezeichnet.
-
(4) Analyse des gereinigten chimären Anti-FGF-8-Antikörpers
-
Jeweils
etwa 4 μg
der in den jeweiligen tierischen Zellen exprimierten chimären Anti-FGF-8-Antikörper KM3034
und KM3334, die unter Punkt 3 von Beispiel 1 gereinigt und erhalten
wurden, wurden einer SDS-PAGE nach der bekannten Methode [Nature
227 (1970), 680] unterzogen, um deren Molekulargewicht und Reinheit
zu untersuchen. Bei jedem der gereinigten chimären Anti-FGF-8-Antikörper wurde
unter nichtreduzierenden Bedingungen eine einzelne Bande mit einem
Molekulargewicht von etwa 150 kD identifiziert und wurden unter
reduzierenden Bedingungen zwei Banden von etwa 50 kD und etwa 25
kD beobachtet. Diese Molekulargewichte entsprachen nahezu den Molekulargewichten,
welche aus den Nucleotidsequenzen der cDNA für die H-Kette und die L-Kette
des Antikörpers
abgeleitet wurden (H-Kette: etwa 49 kD, L-Kette: etwa 23 kD, gesamtes
Molekül:
etwa 144 kD), und stimmten auch mit den Berichten überein,
denen zufolge ein Antikörper vom
IgG-Typ unter nichtreduzierenden Bedingungen ein Molekulargewicht
von etwa 150 kD aufweist und unter reduzierenden Bedingungen aufgrund
der Spaltung einer S-S-Bindung in dem Molekül in H-Ketten mit einem Molekulargewicht
von etwa 50 kD und in L-Ketten mit einem Molekulargewicht von etwa
25 kD abgebaut wird [Antibodies: A Laborstory Manual, Cold Spring
Harbor Laborstory, Kapitel 14 (1988); Monoclonal Antibodies: Principles
and Practice, Academic Press Limited (1996)], wodurch bestätigt wurde,
dass jeder chimäre
Anti-FGF-8-Antikörper
als ein Antikörpermolekül in der
richtigen Struktur exprimiert und gereinigt wurde.
-
(5) Aktivitätsbeurteilung der gereinigten
chimären
Anti-FGF-8-Antikörper
-
Die
FGF-8 neutralisierende Aktivität
der gereinigten chimären
Anti-FGF-8-Antikörper wurde
durch die Messung der nachfolgenden Hemmwirkung gegenüber dem
FGF-8-abhängigen
Wachstum der Maus-Brustkrebszelllinie SC-3 [Proc. Natl. Acad. Sci.
USA 89 (1992), 8928] beurteilt. Hierfür wurden die SC-3-Zellen in einer
Dichte von 3,0 × 10
4 Zellen/ml in DMEM-Ham's F12 (1:1)-Medium (hergestellt von
Gibco), enthaltend 2% mit Aktivkohlenstoff behandeltes FBS, suspendiert
und in 150 μl
(4,5 × 10
3 Zellen/ml) in eine Platte mit 96 Vertiefungen überimpft.
Nach Züchten
bei 37°C
für 18
Stunden in einem Brutschrank mit 5% CO
2 wurde
das Medium unter Verwendung von 100 μl/Vertiefung eines Testmediums
ausgetauscht. Das Testmedium wurde durch das Lösen von 50 ng/ml FGF-8 (hergestellt
von R&D) und
des chimären
Anti-FGF-8-Antikörpers
in der jeweiligen verdünnten
Konzentration in DMEM-Ham's
F12 (1:1)-Medium, enthaltend 0,1% BSA, hergestellt. Der chimäre Antikörper KM2760
gegen den menschlichen Chemokin-Rezeptor CCR4, beschrieben in
WO 01/64754 , wurde als
ein negativer Kontrollantikörper
verwendet. Nach Züchten
bei 37°C
für 48
Stunden in einem Brutschrank mit 5% CO
2 wurde
das Medium durch ein frisch hergestelltes Testmedium ausgetauscht,
gefolgt von einem Züchtungszeitraum
von 48 Stunden. Anschließend
wurde WST-1-Reagens (hergestellt von Roche) in 10 μ/Vertiefung
zugegeben, gefolgt von leichtem Rühren, und nach Züchtung bei
37°C für 1 Stunde in
einem Brutschrank mit 5% CO
2 wurde der OD
450/650-Wert gemessen. In
4 entsprechen
die x-Achse und die y-Achse der Konzentration des zugegebenen Antikörpers bzw.
dem relativen Wachstum (%) bezogen auf das Wachstum nach der Zugabe
von 50 ng/ml FGF-8 allein. Das relative Wachstum (%), bezogen auf
das Wachstum nach der Zugabe von 50 ng/ml FGF-8 allein, wurde durch
die folgende Gleichung berechnet:
(Gleichung)
Relatives
Wachstum (%), bezogen auf das Wachstum nach Zugabe von FGF-8 = [(OD-Wert
nach der Zugabe von FGF-8 und des Antikörpers) – (OD-Wert vor der Zugabe von
FGF-8 und des Antikörpers)]/[(OD-Wert
nach der Zugabe von FGF-8 allein) – [(OD-Wert vor der Zugabe
von FGF-8 und des Antikörpers)] × 100
-
Wie
in 4 gezeigt, wurde für den Anti-FGF-8-Mausantikörper KM1334
und die chimären
Anti-FGF-8-Antikörper
KM3034 und KM3334 jeweils eine vergleichbare Hemmwirkung gegenüber dem
Wachstum von SC-3-Zellen beobachtet, so dass keine Verminderung
der Aktivität
durch die Erzeugung eines chimären
Antikörpers
festgestellt wurde.
-
Beispiel 2
-
Herstellung eines menschlichen CDR-transplantierten
Antikörpers
gegen FGF-8:
-
1. Konstruktion einer cDNA, codierend
die VH-Region und die VL-Region eines menschlichen CDR-transplantierten
Antikörpers
gegen FGF-8
-
(1) Konstruktion von Aminosäuresequenzen
für die
VH-Region und die VL-Region eines menschlichen CDR-transplantierten
Antikörpers
gegen FGF-8
-
Zuerst
wurde die Aminosäuresequenz
der VH-Region eines menschlichen CDR-transplantierten Antikörpers gegen
FGF-8 (Anti-FGF-8-CDR-transplantierter Antikörper) wie folgt konstruiert.
Für die
Transplantation der Aminosäuresequenz
einer CDR-Region aus der VH-Region des Anti-FGF-8-Mausantikörpers KM1334,
welche unter Punkt 1(4) von Beispiel 1 identifiziert wurde, wurde
die Aminosäuresequenz
einer FR-Region in der VH-Region eines menschlichen Antikörpers ausgewählt. Kabat
et al. haben die VH-Region von verschiedenen bekannten menschlichen
Antikörpern
basierend auf der Homologie der Aminosäuresequenzen in drei Untergruppen
(HSG I bis III) eingeteilt und dann die Konsensussequenzen in den
jeweiligen Untergruppen beschrieben [Sequences of Proteins of Immunological
Interest, US Dept. Health and Human Services (1991)]. Da diese Konsensussequenzen
möglicherweise
die Immunogenität
beim Menschen verringern, wurde beschlossen, die Aminosäuresequenz
für die
VH-Region eines Anti-FGF-8-CDR-transplantierten Antikörpers basierend
auf diesen Konsensussequenzen zu entwerfen. Um einen Anti-FGF-8-CDR-transplantierten
Antikörper
herzustellen, welcher eine höhere
Aktivität
besitzt, wurde im Hinblick auf die Konstruktion davon beschlossen,
die Aminosäuresequenz
einer FR-Region auszuwählen,
welche die größte Homologie
zu der Aminosäuresequenz
einer FR-Region in der VH-Region von KM1334 unter den Aminosäuresequenzen
von FR-Regionen in den Konsensussequenzen der drei Untergruppen
für die
VH-Region von menschlichen Antikörpern
hat. Die Ergebnisse der Homologiesuche sind in Tabelle 1 angegeben.
Wie in Tabelle 1 gezeigt, zeigte die Aminosäuresequenz einer FR-Region
in der VH-Region von KM1334 die größte Homologie zu der Untergruppe
I.
-
Tabelle 1
-
Homologie
zwischen der Aminosäuresequenz
einer FR-Region in der Konsensussequenz jeder Untergruppe der VH-Region
eines menschlichen Antikörpers
und der Aminosäuresequenz
einer FR-Region in der VH-Region von KM1334
| HSG
I | HSG
II | HSG
III |
| 79,3% | 51,7% | 59,8% |
-
Basierend
auf den vorstehenden Ergebnissen wurde die Aminosäuresequenz
HV.0 für
die VH-Region des Anti-FGF-8-CDR-transplantierten Antikörpers, welche
in SEQ ID NO: 16 dargestellt ist, durch die Transplantation der
Aminosäuresequenz
einer CDR-Region aus der VH-Region des Anti-FGF-8-Mausantikörpers KM1334
in eine geeignete Position der Aminosäuresequenz einer FR-Region
in der Konsensussequenz der Untergruppe I der VH-Region eines menschlichen
Antikörpers
konstruiert.
-
Anschließend wurde
die Aminosäuresequenz
für die
VL-Region eines Anti-FGF-8-CDR-transplantierten
Antikörpers
wie folgt konstruiert. Für
die Transplantation der Aminosäuresequenz
einer CDR-Region aus der VL-Region des Anti-FGF-8-Mausantikörpers KM1334,
welche unter Punkt 1(4) von Beispiel 1 identifiziert wurde, wurde
die Aminosäuresequenz
einer FR-Region in der VL-Region eines menschlichen Antikörpers ausgewählt. Kabat
et al haben die VL-Region von verschiedenen bekannten menschlichen
Antikörpern
basierend auf der Homologie der Aminosäuresequenzen in vier Untergruppen
(HSG I bis IV) eingeteilt und dann die Konsensussequenzen in den
jeweiligen Untergruppen beschrieben [Sequences of Proteins of Immunological
Interest, US Dept. Health and Human Services (1991)]. Entsprechend
wurde ähnlich
wie im Falle der VH-Region die Aminosäuresequenz einer FR-Region
mit der größten Homologie
zu der Aminosäuresequenz
der FR-Region in der VL-Region von KM1334 aus den Aminosäuresequenzen
für die
FR-Regionen in den Konsensussequenzen der vier Untergruppen der
VL-Region von menschlichen Antikörpern
ausgewählt.
-
Die
Ergebnisse der Homologiesuche sind in Tabelle 2 angegeben. Wie in
Tabelle 2 angegeben, zeigte die Aminosäuresequenz einer FR-Region
in der VL-Region von KM1334 die größte Homologie zu der Untergruppe
II.
-
Tabelle 2
-
Homologie
zwischen der Aminosäuresequenz
einer FR-Region in der Konsensussequenz jeder Untergruppe der VL-Region
eines menschlichen Antikörpers
und der Aminosäuresequenz
einer FR-Region in der VL-Region von KM1334
| HSG
I | HSG
II | HSG
III | HSG
IV |
| 66,3% | 83,8% | 66,2% | 73,8% |
-
Basierend
auf den vorstehenden Ergebnissen wurde die Aminosäuresequenz
LV.0 für
die VL-Region des Anti-FGF-8-CDR-transplantierten Antikörpers, welche
in SEQ ID NO: 17 dargestellt ist, durch die Transplantation der
Aminosäuresequenz
einer CDR-Region aus der VL-Region des Anti-FGF-8-Mausantikörpers KM1334
in eine geeignete Position der Aminosäuresequenz einer FR-Region
in der Konsensussequenz der Untergruppe II der VL-Region eines menschlichen
Antikörpers
konstruiert.
-
Die
Aminosäuresequenz
HV.0 für
die VH-Region und die Aminosäuresequenz
LV.0 für
die VL-Region des Anti-FGF-8-CDR-transplantierten Antikörpers, welche
auf diese Weise konstruiert wurden, sind Sequenzen, wobei nur die
CDR-Aminosäuresequenz
des Anti-FGF-8-Mausantikörpers
KM1334 in die Aminosäuresequenz
einer FR-Region in dem ausgewählten
menschlichen Antikörper
transplantiert wird. In vielen Fällen,
wie im Falle von menschlichen CDR-transplantierten Antikörpern, wird
die Bindungsaktivität
durch die Transplantation der Aminosäuresequenz einer CDR-Region
aus dem Mausantikörper
allein vermindert. Um eine solche Verminderung zu vermeiden, werden
von den Aminosäureresten
in der FR-Region,
welche sich zwischen einem menschlichen Antikörper und einem Mausantikörper unterscheiden,
diejenigen Aminosäurereste,
von denen angenommen wird, dass sie die Bindungsaktivität beeinflussen,
zusammen mit der Aminosäuresequenz der
CDR-Region transplantiert. Entsprechend wurde in diesem Beispiel auch
der Versuch unternommen, die Aminosäurereste in der FR-Region zu
identifizieren, welche wahrscheinlich die Bindungsaktivität beeinflussen.
-
Zuerst
wurde eine dreidimensionale Struktur der V-Region eines Antikörpers, der
die vorstehend konstruierte Aminosäuresequenz HV.0 für die VH-Region
und Aminosäuresequenz
LV.0 für
die VL-Region des Anti-FGF-8-CDR-transplantierten Antikörpers umfasst
(HV0LV0), mit Hilfe von computergestützten Modellerstellungstechniken
entworfen. Die Festlegung der Koordinaten der dreidimensionalen
Struktur erfolgte mit Hilfe der AbM-Software (entwickelt von Oxford
Molecular), und die Darstellung der dreidimensionalen Struktur erfolgte
unter Verwendung der Software Pro-Explore (entwickelt von Oxford
Molecular) oder RasMol (entwickelt von Glaxo) gemäß der jeweiligen
beiliegenden Anweisungen des Herstellers. Ein Computermodell der
dreidimensionalen Struktur der V-Region des Anti-FGF-8-Mausantikörpers KM1334
wurde ebenfalls in der gleichen Weise konstruiert. Außerdem wurde
ein Modell der dreidimensionalen Struktur eines modifizierten Antikörpers, umfassend
eine Aminosäuresequenz,
worin die Aminosäurereste,
welche sich von dem Anti-FGF-8-Mausantikörper KM1334 hinsichtlich der
Aminosäuresequenzen
der FR-Regionen
in der VH-Region und der VL-Region von HV0LV0 unterscheiden, durch
die Aminosäurereste
in Positionen, welche dem Anti-FGF-8-Mausantikörper KM1334 entsprechen, substituiert
wurden, konstruiert, und die dreidimensionalen Strukturen der V-Regionen
von dem Anti-FGF-8-Mausantikörper
KM1334, HV0LV0 und der modifizierten Struktur wurden miteinander
verglichen. Als Ergebnis wurden als Reste unter den Aminosäureresten
der FR-Regionen in HV0LV0, welche wahrscheinlich die Antikörperaktivität durch
Veränderung
der dreidimensionalen Struktur der Antigenbindungsregion beeinflussen,
Lys in Position 12, Lys in Position 13, Ala in Position 40, Pro
in Position 41, Met in Position 48, Val in Position 68, Ile in Position
70, Thr in Position 74, Thr in Position 76, Glu in Position 82, Arg
in Position 87 und Tyr in Position 95 für HV.0 und Ile in Position
2, Val in Position 3, Thr in Position 14, Pro in Position 15, Gln
in Position 50, Leu in Position 51 und Tyr in Position 92 für LV.0 ausgewählt und
für die
Modifikation von Aminosäureresten
verwendet. Durch den Austausch von mindestens einem oder mehreren
dieser ausgewählten
Aminosäurereste
durch die Aminosäurereste,
welche in dem Mausantikörper
KM1334 zu finden sind, wurden die VH-Region und die VL-Region von
menschlichen CDR-transplantierten Antikörpern mit verschiedenen Modifikationen
wie folgt konstruiert.
-
Genauer
wurde die in SEQ ID NO: 18 dargestellte Aminosäuresequenz, worin 6 Reste,
umfassend Lys in Position 12, Lys in Position 13, Ala in Position
40, Pro in Position 41, Met in Position 48 und Tyr in Position 95,
durch die Reste Ala, Arg, Arg, Ser, Ile bzw. Phe, welche in dem
Mausantikörper
KM1334 vorhanden sind, ersetzt wurden, für die VH-Region konstruiert.
Die in SEQ ID NO: 19 dargestellte Aminosäuresequenz, worin 6 Reste,
umfassend Ile in Position 2, Thr in Position 14, Pro in Position
15, Gln in Position 50, Leu in Position 51 und Tyr in Position 92,
durch die Reste Val, Ser, Leu, Lys, Val bzw. Phe, welche in dem
Mausantikörper KM1334
vorhanden sind, ersetzt wurden, für die VL-Region konstruiert.
-
(2) Konstruktion einer cDNA, codierend
die VH-Region des Anti-FGF-8-CDR-transplantierten Antikörpers
-
Die
cDNA, codierend die Aminosäuresequenz
HV.0 für
die VH-Region des Anti-FGF-8-CDR-transplantierten Antikörpers, welche
unter Punkt 1(1) von Beispiel 2 konstruiert wurde, wurde mittels
der PCR-Methode wie folgt konstruiert.
-
Zuerst
wurde die konstruierte Aminosäuresequenz
durch die Ligierung der sekretorischen Signalsequenz für die H-Kette
des Anti-FGF-8-Mausantikörpers
KM1334, welche in SEQ ID NO: 2 dargestellt ist, zu der vollständigen Aminosäuresequenz
eines Antikörpers
ergänzt.
Anschließend
wurde die Aminosäuresequenz
in die genetischen Codons translatiert. Wenn zwei oder mehrere genetische
Codons für
einen Aminosäurerest vorhanden
waren, wurde das entsprechende genetische Codon unter Berücksichtigung
des Codongebrauchs, der für
die Nucleotidsequenzen von Antikörpergenen
ermittelt wurde, festgelegt [Sequences of Proteins of Immunological
Interest, US Dept. Health and Human Services (1991)]. Die Nucleotidsequenz
der cDNA, codierend die vollständige
Aminosäuresequenz
der V-Region des Antikörpers,
wurde konstruiert, indem die auf diese Weise bestimmten genetischen
Codons miteinander ligiert wurden, wobei anschließend komplementäre Nucleotidsequenzen
von Primern für
die Verwendung bei einer PCR-Amplifikation (enthaltend Restriktionsenzym-Erkennungssequenzen
für die
Clonierung in einen Expressionsvektor für einen humanisierten Antikörper) an
das 5'-Ende und
das 3'-Ende angehängt wurden.
Die auf diese Weise konstruierte Nucleotidsequenz wurde in Sequenzen
von jeweils 141 Basen beginnend von dem 5'-terminalen Ende unterteilt, wobei die
angrenzenden Nucleotidsequenzen derart konstruiert wurden, dass
die Enden eine überlappende
Sequenz von etwa 20 Basen aufweisen. Diese Sequenzen wurden in einer
abwechselnden Reihenfolge von Sense-Sequenz und Antisense-Sequenz synthetisiert.
Insbesondere wurden 4 synthetische Oligonucleotide mit der SEQ ID
NO: 20 bis 23 synthetisiert (hergestellt von GENSET).
-
Jedes
Oligonucleotid wurde zu 50 μl
einer Reaktionslösung
zugegeben, um eine Endkonzentration von 0,1 μMol/l zu erhalten. Die PCR wurde
unter Verwendung von 0,5 μMol/l
des M13-Primers RV (hergestellt von Takara Shuzo), 0,5 μMol/l des
M13-Primers M4 (hergestellt von Takara Shuzo) und 2,5 Einheiten
der KOD-Polymerase (hergestellt von TOYOBO) gemäß den Anweisungen des Herstellers,
die der KOD-Polymerase beigefügt
waren, durchgeführt.
Die Reaktionsbedingungen beinhalteten das Erhitzen der Reaktionslösung bei
94°C für 5 Minuten
und anschließend
25 Reaktionszyklen bei 94°C
für 30
Sekunden, bei 50°C
für 30 Sekunden
und bei 74°C
für 60
Sekunden für
einen Zyklus, und weiterhin das Erhitzen bei 74°C für 5 Minuten. Die Reaktionslösung wurde
mit Ethanol gefällt.
Das Präzipitat
wurde in sterilem Wasser gelöst
und mit 10 Einheiten des Restriktionsenzyms EcoRI (hergestellt von
Takara Shuzo) und 10 Einheiten des Restriktionsenzyms SpeI (hergestellt
von Takara Shuzo) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
0,3 μg eines
EcoRI-Spel-Fragments
von etwa 0,47 kb gewonnen wurden.
-
Anschließend wurden
3 μg des
Plasmids pBluescript II SK(–)
(hergestellt von Stratagene) mit 10 Einheiten des Restriktionsenzyms
EcoRI (hergestellt von Takara Shuzo) und 10 Einheiten des Restriktionsenzyms SpeI
(hergestellt von Takara Shuzo) bei 37°C für 1 Stunde umgesetzt. Die Reaktionslösung wurde
durch eine Agarosegelelektrophorese fraktioniert, wobei etwa 2,9 μg eines EcoRI-SpeI-Fragments von etwa
2,95 kb gewonnen wurden.
-
Im
Anschluss daran wurden 0,1 μg
des EcoRI-SpeI-Fragments des PCR-Produkts
der VH-Region des Anti-FGF-8-CDR-transplantierten Antikörpers und
0,1 μg des
EcoRI-SpeI-Fragments des Plasmids pBluescript II SK(–), welche
beide vorstehend erhalten wurden, zu einem Gesamtvolumen von 10 μl sterilem
Wasser zugegeben und unter Verwendung von Ligation High (hergestellt
von TOYOBO) ligiert. Unter Verwendung der so erhaltenen rekombinanten
Plasmid-DNA-Lösung
wurden Zellen von E. coli DH5α (hergestellt
von TOYOBO) transformiert. Aus jeweils 10 Clonen der Transformanten
wurde die Plasmid-DNA präpariert,
und anschließend
wurde eine Nucleotidsequenzanalyse nach der Didesoxy-Methode [Molecular Cloning:
A Laborstory Manual, Cold Spring Harbor Lab. Press, New York (1989)]
unter Verwendung des Big Dye Terminator-Kits, Ver. 2 (hergestellt
von Applied Biosystems) durchgeführt.
Als Ergebnis der Nucleotidsequenzanalyse wurde das in 5 gezeigte
Plasmid pKM1334HV0 erhalten, welches die Nucleotidsequenz von Interesse
enthält.
-
In
der gleichen Weise wie vorstehend beschrieben, wurde auch eine cDNA,
codierend die Aminosäuresequenz
HV.6 der VH-Region des Anti-FGF-8-CDR-transplantierten Antikörpers, welche
unter Punkt 1(1) von Beispiel 2 entworfen wurde, mittels PCR unter
Verwendung von 4 synthetischen Oligonucleotiden mit der SEQ ID NO:
24 bis 27 (hergestellt von GENSET) als synthetische DNAs konstruiert.
Als Ergebnis wurde das Plasmid pKM1334HV6 erhalten, welches die
HV.6 codierende cDNA enthält.
-
(3) Konstruktion einer cDNA, codierend
die VL-Region des Anti-FGF-8-CDR-transplantierten Antikörpers
-
Die
cDNA, codierend die Aminosäuresequenz
LV.0 für
die VL-Region des Anti-FGF-8-CDR-transplantierten Antikörpers, welche
unter Punkt 1(1) von Beispiel 2 entworfen wurde, wurde in der gleichen
Weise wie im Falle der VH-Region mittels PCR konstruiert. In diesem
Fall jedoch wurde die in SEQ ID NO: 4 dargestellte Sequenz der L-Kette
des Anti-FGF-8-Mausantikörpers
als sekretorische Signalsequenz verwendet und wurden die 4 synthetischen
Oligonucleotide mit SEQ ID NO: 28 bis 31 (hergestellt von GENSET)
als synthetische DNAs verwendet. Als Ergebnis wurde das Plasmid
pKM1334LV0 erhalten, welches die LV.0 codierende cDNA enthält.
-
In
der gleichen Weise wie vorstehend beschrieben, wurde auch eine cDNA,
codierend die Aminosäuresequenz
LV.6 der VL-Region des Anti-FGF-8-CDR-transplantierten Antikörpers, welche
unter Punkt 1(1) von Beispiel 2 konstruiert wurde, mittels PCR unter
Verwendung von 4 synthetischen Oligonucleotiden mit der SEQ ID NO:
32 bis 35 (hergestellt von GENSET) als synthetische DNAs konstruiert.
Als Ergebnis wurde das Plasmid pKM1334LV6 erhalten, welches die
LV.6 codierende cDNA enthält.
-
2. Konstruktion eines Expressionsvektors
für den
Anti-FGF-8-CDR-transplantierten Antikörper
-
Unter
Verwendung des Vektors pKANTEX93 für die Expression eines humanisierten
Antikörpers,
beschrieben in
WO 97/10354 ,
und der Plasmide pKM1334HV0 und pKM1334LV4, welche unter Punkt 1(2)
und 1(3) von Beispiel 2 erhalten wurden, wurde der Expressionsvektor
pKANTEX1334HV0LV0 für
den Anti-FGF-8-CDR-transplantierten
Antikörper
wie folgt konstruiert.
-
Zuerst
wurden 3 μg
des Plasmids pKM1334HV0, welches unter Punkt 1(2) von Beispiel 2
erhalten wurde, mit 10 Einheiten des Restriktionsenzyms ApaI (hergestellt
von Takara Shuzo) und 10 Einheiten des Restriktionsenzyms NotI (hergestellt
von Takara Shuzo) gemischt und bei 37°C für 1 Stunde umgesetzt. Die Reaktionslösung wurde
durch eine Agarosegelelektrophorese fraktioniert, wobei etwa 0,3 μg eines ApaI-NotI-Fragments
von etwa 0,47 kb gewonnen wurden.
-
Anschließend wurden
3 μg des
Expressionsvektors pKANTEX93 für
einen humanisierten Antikörper mit
10 Einheiten des Restriktionsenzyms ApaI (hergestellt von Takara
Shuzo) und 10 Einheiten des Restriktionsenzyms NotI (hergestellt
von Takara Shuzo) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
2 μg eines
ApaI-NotI-Fragments von etwa 12,8 kb gewonnen wurden.
-
Im
Anschluss daran wurden 0,1 μg
des ApaI-NotI-Fragments des Plasmids pKM1334HV0 und 0,1 μg des ApaI-NotI-Fragments
des Plasmids pKANTEX93 zu einem Gesamtvolumen von 10 μl sterilem
Wasser zugegeben und unter Verwendung von Ligation High (hergestellt
von TOYOBO) ligiert. Unter Verwendung der so erhaltenen rekombinanten
Plasmid-DNA-Lösung
wurden Zellen von E. coli DH5α transformiert,
wobei das in 6 gezeigte Plasmid pKANTEX1334HV0
erhalten wurde.
-
Anschließend wurden
3 μg des
Plasmids pKM1334LV0, welches unter Punkt 1(3) von Beispiel 2 erhalten
wurde, mit 10 Einheiten des Restriktionsenzyms EcoRI (hergestellt
von Takara Shuzo) und 10 Einheiten des Restriktionsenzyms BsiWI
(hergestellt von New England Biolabs) bei 37°C für 1 Stunde umgesetzt. Die Reaktionslösung wurde
durch eine Agarosegelelektrophorese fraktioniert, wobei etwa 0,3 μg eines EcoRI-BsiWI-Fragments
von etwa 0,45 kb gewonnen wurden.
-
Im
Anschluss daran wurden 3 μg
des Plasmids pKM1334HV0, das vorstehend erhalten wurde, mit 10 Einheiten
des Restriktionsenzyms EcoRI (hergestellt von Takara Shuzo) und
10 Einheiten des Restriktionsenzyms BsiWI (hergestellt von New England
Biolabs) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
2 μg eines
EcoRI-BsiWI-Fragments
von etwa 13,30 kb gewonnen wurden.
-
Als
nächstes
wurden 0,1 μg
des EcoRI-BsiWI-Fragments. des Plasmids pKM1334LV0 und 0,1 μg des EcoRI-BsiWI-Fragments
des Plasmids pKANTEX1334-HV0
zu einem Gesamtvolumen von 10 μl
sterilem Wasser zugegeben und unter Verwendung von Ligation High
(hergestellt von TOYOBO) ligiert. Unter Verwendung der so erhaltenen
rekombinanten Plasmid-DNA-Lösung
wurden Zellen des E. coli-Stamms
DH5α transformiert,
wobei das in 6 gezeigte Plasmid pKANTEX1334-HV0LV0 erhalten wurde.
-
Unter
Verwendung von 400 ng des so erhaltenen Plasmids wurde die Nucleotidsequenz
hiervon mit Hilfe der Didesoxy-Methode [Molecular Cloning: A Laborstory
Manual, Cold Spring Harbor Lab. Press, New York (1989)] unter Verwendung
des Big Dye Terminator-Kits, Ver. 2 (hergestellt von Applied Biosystems)
bestimmt, wodurch bestätigt
wurde, dass ein Plasmid erhalten wurde, in welches die DNA von Interesse
cloniert worden war.
-
In
der gleichen Weise wie vorstehend beschrieben, wurde auch der Expressionsvektor pKANTEX1334HV0LV6
unter Verwendung des Plasmids pKM1334HV0, welches unter Punkt 1(2)
von Beispiel 2 erhalten wurde, und des Plasmids pKANTEX1334LV6,
welches unter Punkt 1(3) von Beispiel 2 erhalten wurde, konstruiert.
-
In
der gleichen Weise wie vorstehend beschrieben, wurde außerdem der
Expressionsvektor pKANTEX1334HV6LV6 unter Verwendung des Plasmids
pKM-1334HV6, welches
unter Punkt 1(2) von Beispiel 2 erhalten wurde, und des Plasmids
pKANTEX1334LV6, welches unter Punkt 1(3) von Beispiel 2 erhalten wurde,
konstruiert.
-
3. Stabile Expression des Anti-FGF-8-CDR-transplantierten
Antikörpers
unter Verwendung von YB2/0-Zellen
-
Unter
Verwendung der Expressionsvektoren pKANTEX1334HV0LV0, pKANTEX1334HV0LV6
und pKANTEX1334HV6LV6 für
den Anti-FGF-8-CDR-transplantierten Antikörper, welche unter Punkt 2
von Beispiel 2 erhalten wurden, wurde die stabile Expression der
verschiedenen Anti-FGF-8-CDR-transplantierten Antikörper in
YB2/0-Zellen gemäß dem unter
Punkt 2(5) von Beispiel 1 beschriebenen Verfahren bewirkt.
-
4. Reinigung der Anti-FGF-8-CDR-transplantierten
Antikörper
-
Die
Züchtung
der Transformanten, erhalten unter Punkt 3 von Beispiel 2, welche
von den YB2/0-Zellen erhalten wurden und welche die verschiedenen
Anti-FGF-8-CDR-transplantierten
Antikörper
exprimieren, sowie die Reinigung der Anti-FGF-8-CDR-transplantierten Antikörper aus
den Kulturüberständen wurden
gemäß dem unter
Punkt 3(2) von Beispiel 1 beschriebenen Verfahren durchgeführt. Der
Antikörper
von einer Transformante, in die pKANTEX1334HV0LV0 eingeführt wurde,
wurde als HV0LV0 bezeichnet; der Antikörper von einer Transformante,
in die pKANTEX1334HV0LV6 eingeführt
wurde, wurde als HV0LV6 bezeichnet; und der Antikörper von
einer Transformante, in die pKANTEX1334HV6LV6 eingeführt wurde,
wurde als HV6LV6 bezeichnet.
-
5. Analyse der gereinigten Anti-FGF-8-CDR-transplantierten
Antikörper
-
Die
SDS-PAGE unter Verwendung der verschiedenen Anti-FGF-8-CDRtransplantierten
Antikörper, welche
unter Punkt 4 von Beispiel 2 erhalten wurden, wurde gemäß dem unter
Punkt 4 von Beispiel 1 beschriebenen Verfahren durchgeführt. Als
Ergebnis wurde bestätigt,
dass jeder Antikörper
als ein Antikörpermolekül in der
richtigen Struktur exprimiert und gereinigt wurde.
-
6. Messung der Bindungsaktivität der Anti-FGF-8-CDR-transplantierten
Antikörper
gegenüber
FGF-8 (ELISA)
-
Die
Aktivität
der verschiedenen Anti-FGF-8-CDR-transplantierten Antikörper, welche
unter Punkt 4 von Beispiel 2 erhalten wurden, im Hinblick auf die
Bindung an FGF-8 wurde durch den ELISA, welcher unter Punkt 2(6)
von Beispiel 1 beschrieben wurde, gemessen. Der chimäre Anti-FGF-8-Antikörper KM3334,
welcher unter Punkt 3(2) von Beispiel 1 aus YB2/0-Zellen erhalten
wurde, wurde als eine positive Kontrolle verwendet. Die Ergebnisse
sind in 7 dargestellt. Wie in 7 dargestellt,
zeigte jeder der Anti-FGF-8-CDR-transplantierten Antikörper eine
FGF-8-Bindungsaktivität,
welche mit derjenigen von KM3334 vergleichbar war, so dass keine
wesentliche Verringerung der Bindungsaktivität durch die CDR-Transplantation
beobachtet wurde.
-
7. Messung der Bindungsaktivität der Anti-FGF-8-CDR-transplantierten
Antikörper
gegenüber
FGF-8
-
Um
die Aktivität
der verschiedenen Anti-FGF-8-CDR-transplantierten Antikörper, welche
unter Punkt 4 von Beispiel 2 erhalten wurden, im Hinblick auf die
Bindung an FGF-8 genauer zu untersuchen, wurden die Aktivitäten der
verschiedenen Anti-FGF-8-CDR-transplantierten Antikörpern hinsichtlich
der Bindung an FGF-8 gemessen und unter Verwendung des BIAcore-Systems
2000 (hergestellt von BIACORE) wie folgt verglichen. Der chimäre Anti-FGF-8-Antikörper KM3334,
welcher unter Punkt 3(2) von Beispiel 1 aus YB2/0-Zellen erhalten
wurde, wurde als eine positive Kontrolle verwendet.
-
Im
Folgenden wurde HBS-EP (hergestellt von BIACORE) als Puffer für die Verdünnung der
Proben und während
der Messung verwendet. Zuerst wurde die Sensorspitze CM5 (hergestellt
von BIACORE) justiert, und anschließend wurde FGF-8 (hergestellt
von R&D), gelöst in einer
Konzentration von 31,25 μg/ml
in 10 mMol/l Acetatpuffer (pH 4,0), durch ein Amin-Kopplungsverfahren
auf der Oberfläche
der Sensorspitze immobilisiert. Die immobilisierte Menge entsprach
4,498 RU.
-
Jeweils
60 μl der
Antikörperlösung wurden
in einer Durchflussrate von 20 μl/Minute
in die den immobilisierten FGF-8 enthaltende Durchflusszelle eingebracht,
und anschließend
wurde die Dissoziationsreaktion während 3 Minuten überwacht.
Im Anschluss an die Dissoziationsreaktion wurde die Spitzenoberfläche durch die
kontinuierliche Zugabe von zweimal 20 μl 10 mMol/l Glycin-HCl-Puffer
(pH 1,5) zu der Durchflusszelle regeneriert. Dieser Zyklus wurde
für Antikörperlösungen mit
unterschiedlichen Konzentrationen (50 bis 0,068 μg/ml) durchgeführt, wobei
für jede
Konzentration ein Sensorgramm erhalten wurde. Das Sensorgramm für jeden
Antikörper
wurde durch Subtraktion des Sensorgramms, welches unter Verwendung
des chimären
Antikörpers
KM871 gegen CD3 [Cancer Immunol. Immunother. 36 (1993), 373] als
eine negative Kontrolle erhalten wurde, in ein Sensorgramm für die spezifische
Reaktion umgewandelt. Das Sensorgramm für 50 μg/ml jedes Antikörpers ist
in 8 dargestellt. Wie aus dem Sensorgramm ersichtlich
ist, wurde zum Zeitpunkt der Dissoziationsreaktion jedes Antikörpers praktisch
keine Dissoziation beobachtet, so dass es schwierig war, eine exakte
Dissoziationskonstante zu erhalten. Demgemäß erfolgte der Vergleich der
Bindungsaktivität
der verschiedenen Antikörper
durch den Vergleich der Stärke
der Bindung [Resonanzsignal (RU)] zum Zeitpunkt der Bindungsreaktion.
Als Ergebnis, wie in 8 gezeigt, zeigte der chimäre Antikörper KM3334
die stärkste
Bindungsreaktion, und der CDR-transplantierte Antikörper HV0LV6
zeigte eine starke Bindungsreaktion, welche mit derjenigen von KM3334
vergleichbar war. Dagegen zeigten die CDR-transplantierten Antikörper HV0LV0 und
HV6LV6 eine etwas geringere Bindungsreaktion als KM3334 und HV0LV6.
Die vorstehenden Ergebnisse zeigen, dass ein Vergleich der Bindungsaktivitäten zwischen
Antikörpern,
welcher durch einen ELISA nicht möglich ist, unter Verwendung
des BIAcore-Systems durchgeführt
kann und dass die Bindungsaktivität des CDR-transplantierten
Antikörpers
durch die Modifikation der 6 Aminosäurereste in der FR-Region der VL-Region
wieder das gleiche Niveau wie die Bindungsaktivität des chimären Antikörpers erreicht.
Ferner wurde gezeigt, dass die Modifikation der 6 Aminosäurereste
in der FR-Region der VH-Region keinen Einfluss auf die Erhöhung der
Bindungsaktivität
hatte.
-
Der
aus YB2/0-Zellen erhaltene CDR-transplantierte Antikörper HV0LV6,
der eine starke Bindungsreaktion zeigte, welche mit derjenigen des
chimären
Antikörpers
KM3334 vergleichbar war, wurde als KM8037 bezeichnet; und die von
YB2/0-Zellen abgeleitete transformierte Zelllinie, welche eine hohe
Expression von KM8037 zeigt, wurde ebenfalls als KM8037 bezeichnet.
Die transformierte Zelllinie KM8037 ist auch am 20. Juni 2002 als
FERM BP-8084 bei der International Patent Organism Depositary, National
institute of Advanced Industrial Science and Technology (AIST Tsukuba
Central 6, 1–1,
Higashi 1-Chorre Tsukuba-shi, Ibaraki-ken 305–8566, Japan), hinterlegt worden.
-
Beispiel 3
-
Herstellung eines Anti-FGF-8-CDR-transplantierten
Antikörpers
mit einer geringeren Immunogenität
(1)
-
Anhand
der Ergebnisse von Beispiel 2 wurde gezeigt, dass der Anti-FGF-8-CDR-transplantierte
Antikörper,
der die Modifikation der 6 Aminosäurereste, welche von dem Mausantikörper KM1334
stammen, in der FR-Region der VL-Region enthält, eine Bindungsaktivität zeigt,
welche mit derjenigen des entsprechenden chimären Antikörpers vergleichbar ist. Demgemäß wurde
der Einfluss dieser sechs Reste auf die Wiederherstellung der Aktivität weiter
untersucht, wobei wie folgt Anti-FGF-8-CDR-transplantierte Antikörper hergestellt
wurden, welche eine ausreichende Aktivität aufweisen und die eine geringere
Anzahl an Aminosäureresten
enthalten, welche von dem Mausantikörper stammen, wobei erwartet
wird, dass diese Antikörper
eine weiter verminderte Immunogenität haben.
-
1. Konstruktion einer Aminosäuresequenz
für die
VL-Region
-
Im
Hinblick auf die vorstehenden 6 Aminosäurereste wurden Aminosäuresequenzen
für 6 VL-Regionen
mit den folgenden Modifikationen konstruiert.
-
In
LV.4-1 wurden 4 Reste, umfassend Ile in Position 2, Gln in Position
50, Leu in Position 51 und Tyr in Position 92, durch die Reste Val,
Lys, Val bzw. Phe, welche den Aminosäureresten entsprechen, die
in dem Mausantikörper
KM1334 vorhanden sind, ersetzt.
-
In
LV.4-2 wurden 4 Reste, umfassend Ile in Position 2, Thr in Position
14, Pro in Position 15 und Tyr in Position 92, durch die Reste Val,
Ser, Leu bzw. Phe, welche den Aminosäureresten entsprechen, die
in dem Mausantikörper
KM1334 vorhanden sind, ersetzt.
-
In
LV.3-1 wurden 3 Reste, umfassend Ile in Position 2, Leu in Position
51 und Tyr in Position 92, durch die Reste Val, Val bzw. Phe, welche
den Aminosäureresten
entsprechen, die in dem Mausantikörper KM1334 vorhanden sind,
ersetzt.
-
In
LV.3-2 wurden 3 Reste, umfassend Thr in Position 14, Pro in Position
15 und Tyr in Position 92, durch die Reste Ser, Leu bzw. Phe, welche
den Aminosäureresten
entsprechen, die in dem Mausantikörper KM1334 vorhanden sind,
ersetzt.
-
In
LV.2-1 wurden 2 Reste, umfassend Leu in Position 51 und Tyr in Position
92, durch die Reste Val bzw. Phe, welche den Aminosäureresten
entsprechen, die in dem Mausantikörper KM1334 vorhanden sind, ersetzt.
-
In
LV.2-2 wurden 2 Reste, umfassend Ile in Position 2 und Tyr in Position
92, durch die Reste Val bzw. Phe, welche den Aminosäureresten
entsprechen, die in dem Mausantikörper KM1334 vorhanden sind,
ersetzt.
-
2. Konstruktion einer cDNA, codierend
die VL-Region
-
Die
cDNAs, codierend die Aminosäuresequenzen
für die
VL-Region des jeweiligen Anti-FGF-8-CDR-transplantierten Antikörpers, welche
unter Punkt 1 von Beispiel 3 entworfen wurden, wurden wie folgt
konstruiert.
-
(1) Konstruktion einer cDNA, codierend
LV.4-1
-
Die
LV.4-1 codierende cDNA wurde gemäß dem Verfahren,
welches unter Punkt 1(3) von Beispiel 2 beschrieben wurde, unter
Verwendung von vier synthetischen Oligonucleotiden mit der SEQ ID
NO: 29, 32, 34 und 35 als synthetische DNA-Fragmente (hergestellt von GENSET) konstruiert.
Als Ergebnis wurde das Plasmid pKM1334LV4-1 erhalten, welches die
LV.4-1 codierende cDNA enthält.
-
(2) Konstruktion einer cDNA, codierend
LV.3-1
-
Unter
Verwendung von 50 ng des Plasmids pKM1334LV6, welches unter Punkt
1(3) von Beispiel 2 erhalten wurde, als Matrize, des M13-Primers
RV (hergestellt von Takara Shuzo) und der synthetischen DNA mit der
in SEQ ID NO: 38 dargestellten Nucleotidsequenz (hergestellt von
GENSET), welche als Primer in einer Endkonzentration von 0,3 μMol/l zugegeben
wurden, wurde eine PCR, umfassend zuerst das Erhitzen bei 94°C für 2 Minuten
und dann 35 Reaktionszyklen bei 94°C für 15 Sekunden, bei 50°C für 30 Sekunden
und bei 68°C
für 1 Minuten
für einen
Zyklus, gemäß den Anweisungen
des Herstellers, welche der KOD-Polymerase (hergestellt von TOYOBO)
beigefügt
waren, durchgeführt.
Die Reaktionslösung
wurde gereinigt, und das Produkt wurde in sterilem Wasser gelöst. Anschließend wurde
das Produkt mit 10 Einheiten des Restriktionsenzyms KpnI (hergestellt
von Takara Shuzo) und 10 Einheiten des Restriktionsenzyms SpeI (hergestellt
von Takara Shuzo) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektro phorese fraktioniert, wobei etwa
0,3 μg eines
KpnI-SpeI-Fragments von etwa 0,22 kb gewonnen wurden.
-
Anschließend wurden
3 μg des
Plasmids pKM1334LV4-1, welches unter Punkt 2(1) von Beispiel 3 erhalten
wurde, mit 10 Einheiten des Restriktionsenzyms KpnI (hergestellt
von Takara Shuzo) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa 0,2 μg eines KpnI-KpnI-Fragments
von etwa 0,21 kb gewonnen wurden.
-
Im
Anschluss daran wurden 3 μg
des Plasmids pBluescript II SK(–)
mit 10 Einheiten des Restriktionsenzyms KpnI (hergestellt von Takara
Shuzo) und 10 Einheiten des Restriktionsenzyms SpeI (hergestellt
von Takara Shuzo) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
2 μg eines
KpnI-SpeI-Fragments von etwa 2,95 kb gewonnen wurden.
-
Zu
einem Gesamtvolumen von 10 μl
sterilen Wasser wurden 0,1 μg
des Kpnl-Spei-Fragments der VL-cDNA, 0,1 μg des KpnI-KpnI-Fragments des
Plasmids pKM1334LV4-1 und 0,1 μg
des KpnI-Spei-Fragments des Plasmids pBluescript 11 SK(–), welche
jeweils vorstehend erhalten wurden, zugegeben und unter Verwendung
von Ligation High (hergestellt von TOYOBO) ligiert. Unter Verwendung
der so erhaltenen rekombinanten Plasmid-DNA-Lösung wurden Zellen von E. coli
DH5α transformiert,
wobei das in 9 dargestellte Plasmid pKM1334LV3-1
erhalten wurde, welches die LV.3-1 codierende cDNA enthält.
-
(3) Konstruktion einer cDNA, codierend
LV.2-1
-
Das
Plasmid pKM1334LV2-1, enthaltend eine cDNA, codierend LV.2-1, wurde
durch ein ähnliches
Verfahren erhalten, wie unter Punkt 2(1) von Beispiel 2 beschrieben
wurde, außer
dass das Plasmid pKM1334LV0, welches unter Punkt 1(3) von Beispiel
2 erhalten wurde, anstelle des Plasmids pKM1334LV4-1 verwendet wurde.
-
(4) Konstruktion einer cDNA, codierend
LV.2-2
-
Das
Plasmid pKM1334LV2-2, enthaltend eine cDNA, codierend LV.2-2, wurde
durch ein ähnliches
Verfahren erhalten, wie unter Punkt 2(1) von Beispiel 2 beschrieben
wurde, außer
dass die in SEQ ID NO: 37 dargestellte synthetische DNA anstelle
der in SEQ ID NO: 36 dargestellten synthetischen DNA als Primer
verwendet wurde.
-
(5) Konstruktion einer cDNA, codierend
LV.4-2
-
Zuerst
wurden 3 μg
des Plasmids pKM1334LV2-2, welches unter Punkt 2(4) von Beispiel
3 erhalten wurde, mit 10 Einheiten des Restriktionsenzyms KpnI (hergestellt
von Takara Shuzo) gemischt und bei 37°C für 1 Stunde umgesetzt. Die Reaktionslösung wurde
durch eine Agarosegelelektrophorese fraktioniert, wobei etwa 2 μg eines KpnI-KpnI-Fragments
von etwa 3,16 kb gewonnen wurden.
-
Anschließend wurden
3 μg des
Plasmids pKM1334LV6, welches unter Punkt 1(3) von Beispiel 2 erhalten
wurde, mit 10 Einheiten des Restriktionsenzyms KpnI (hergestellt
von Takara Shuzo) bei 37°C
für 1 Stunde umgesetzt.
Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
0,2 μg eines
KpnI-KpnI-Fragments von etwa 0,21 kb gewonnen wurden.
-
Zu
einem Gesamtvolumen von 10 μl
sterilem Wasser wurden 0,1 μg
des KpnI-KpnI-Fragments des Plasmids pKM1334LV2-2 und 0,1 μg des KpnI-KpnI-Fragments
des Plasmids pKM1334LV6, welche jeweils vorstehend erhalten wurden,
zugegeben und unter Verwendung von Ligation High (hergestellt von
TOYOBO) ligiert. Unter Verwendung der so erhaltenen rekombinanten
Plasmid-DNA-Lösung
wurden Zellen von E. coli DH5α transformiert,
wobei das in 10 dargestellte Plasmid pKM1334LV4-2
erhalten wurde, welches die LV.4-2 codierende cDNA enthält.
-
(6) Konstruktion einer cDNA, codierend
LV.3-2
-
Zunächst wurden
3 μg des
Plasmids pKM1334LV4-2, welches unter Punkt 2(5) von Beispiel 3 erhalten wurde,
mit 10 Einheiten der Restriktionsenzyme Tth111I (hergestellt von
Takara Shuzo) und XmnI (hergestellt von New England Biolabs) gemischt
und bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
2 μg eines
Tth111I-XmnI-Fragments
von etwa 2,24 kb gewonnen wurden.
-
Anschließend wurden
3 μg des
Plasmids pKM1334LV0, welches unter Punkt 1(3) von Beispiel 2 erhalten
wurde, mit 10 Einheiten der Restriktionsenzyme Tth111I (hergestellt
von Takara Shuzo) und XmnI (hergestellt von New England Biolabs)
bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
1 μg eines
Tth111I-XmnI-Fragments von etwa 1,11 kb gewonnen wurden.
-
Zu
einem Gesamtvolumen von 10 μl
sterilem Wasser wurden 0,1 μg
des Tth111I-XmnI-Fragments des Plasmids pKM1334LV4-2 und 0,1 μg des Tth111I-XmnI-Fragments des
Plasmids pKM1334LV0, welche jeweils vorstehend erhalten wurden,
zugegeben und unter Verwendung von Ligation High (hergestellt von
TOYOBO) ligiert. Unter Verwendung der so erhaltenen rekombinanten
Plasmid-DNA-Lösung wurden
Zellen von E. coli DH5α transformiert,
wobei das in 11 dargestellte Plasmid pKM1334LV3-2
erhalten wurde, welches die LV.3-2 codierende cDNA enthält.
-
3. Konstruktion eines Expressionsvektors
für einen
Anti-FGF-8-CDR-transplantierten Antikörper
-
Verschiedene
Expressionsvektoren für
einen Anti-FGF-8-CDR-transplantierten Antikörper, welche eine VL-cDNA enthalten,
wurden durch das Ersetzen des EcoRI-BsiWI-Fragments, welches die
VL-cDNA des Expressionsvektors pKANTEX1334HV0LV6 enthält, erhalten
unter Punkt 2 von Beispiel 2, durch die jeweiligen verschiedenen
EwRI-BsiWI-Fragmente, enthaltend eine VL-cDNA, welche unter Punkt
2 von Beispiel 3 erhalten wurden, konstruiert. Insbesondere wurden
die 6 Vektoren pKANTEX1334HV0LV4-1, pKANTEX1334HV0LV4-2, pKANTEX1334HV0LV3-1,
pKANTEX1334HV0LV3-2, pKANTEX1334HV0LV2-1 und pKANTEX1334HV0LV2-2,
konstruiert.
-
4. Stabile Expression der Anti-FGF-8-CDR-transplantierten
Antikörper
unter Verwendung von CHO/DG44-Zellen
-
Die
stabile Expression der verschiedenen Anti-FGF-8-CDR-transplantierten
Antikörper
in CHO/DG44-Zellen wurde gemäß dem unter
Punkt 2(4) von Beispiel 1 beschriebenen Verfahren unter Verwendung
der Expressionsvektoren pKANTEX1334HV0LV0 und pKANTEX1334HV0LV6,
welche unter Punkt 2 von Beispiel 2 erhalten wurden, und der verschiedenen
Expressionsvektoren für
einen Anti-FGF-8-CDR-transplantierten
Antikörper,
welche unter Punkt 3 von Beispiel 3 erhalten wurden, bewirkt.
-
5. Reinigung der Anti-FGF-8-CDR-transplantierten
Antikörper
-
Die
Züchtung
der Transformanten, erhalten unter Punkt 4 von Beispiel 3, welche
von den CHO/DG44-Zellen erhalten wurden und welche die verschiedenen
Anti-FGF-8-CDR-transplantierten Antikörper exprimieren, sowie die
Reinigung der Anti-FGF-8-CDR-transplantierten Antikörper aus
den Kulturüberständen wurden
gemäß dem unter
Punkt 3(1) von Beispiel 1 beschriebenen Verfahren durchgeführt. Der
Antikörper von
einer Transformante, in die pKANTEX1334HV0LV0 eingeführt wurde,
wurde als HV0LV0/CHO bezeichnet; der Antikörper von einer Transformante,
in die pKANTEX1334HV0LV6 eingeführt
wurde, wurde als HV0LV6/CHO bezeichnet; der Antikörper von
einer Transformante, in welche pKANTEX1334HV0LV4-1 eingeführt wurde,
wurde als HV0LV4-1/CHO bezeichnet; der Antikörper von einer Transformante,
in welche pKANTEX1334HV0LV4-2 eingeführt wurde, wurde als HV0LV4-2/CHO
bezeichnet; der Antikörper
von einer Transformante, in welche pKANTEX1334HV0LV3-1 eingeführt wurde,
wurde als HV0LV3-1/CHO bezeichnet; der Antikörper von einer Transformante,
in die pKANTEX1334HV0LV3-2 eingeführt wurde, wurde als HV0LV3-2/CHO
bezeichnet; der Antikörper
von einer Transformante, in die pKANTEX1334HV0LV2-1 eingeführt wurde,
wurde als HV0LV2-1/CHO bezeichnet; und der Antikörper von einer Transformante,
in die pKANTEX1334HV0LV2-2 eingeführt wurde, wurde als HV0LV2-2/CHO
bezeichnet.
-
6. Analyse der gereinigten Anti-FGF-8-CDR-transplantierten
Antikörper
-
Die
SDS-PAGE unter Verwendung der verschiedenen Anti-FGF-8-CDR-transpiantierten
Antikörper, welche
unter Punkt 5 von Beispiel 3 erhalten wurden, wurde gemäß dem unter
Punkt 4 von Beispiel 1 beschriebenen Verfahren durchgeführt. Als
Ergebnis wurde bestätigt,
dass jeder Antikörper
als ein Antikörpermolekül in der
richtigen Struktur exprimiert und gereinigt wurde.
-
7. Messung der Bindungsaktivität der Anti-FGF-8-CDR-transplantierten
Antikörper
gegenüber
FGF-8 (BIAcore-Biosensor)
-
Um
die Aktivität
der verschiedenen Anti-FGF-8-CDR-transplantierten Antikörper, welche
unter Punkt 5 von Beispiel 3 erhalten wurden, im Hinblick auf die
Bindung an FGF-8 genauer zu untersuchen, wurden die Aktivitäten der
verschiedenen Anti-FGF-8-CDR-transplantierten Antikörper hinsichtlich
der Bindung an FGF-8 gemessen und unter Verwendung des BIAcore-Systems
2000 (hergestellt von BIACORE) wie folgt verglichen. Der chimäre Anti-FGF-8-Antikörper KM3034,
welcher unter Punkt 3(1) von Beispiel 1 aus CHO/DG44-Zellen erhalten
wurde, wurde als eine positive Kontrolle verwendet.
-
Im
Folgenden wurde HBS-EP (hergestellt von Pharmacia) als Pufferlösung für die Verdünnung der Proben
und während
der Messung verwendet. Zuerst wurde die Sensorspitze SA (hergestellt
von BIACORE) justiert, und anschließend wurden 5 μl einer am
C-Terminus mit Biotin markierten Verbindung 1 (ein N-terminales
Peptid von FGF-8; SEQ ID NO: 15), zubereitet in Form einer Lösung von
0,05 μg/ml,
in einer Durchflussrate von 20 μl/Minute
zugegeben. Im Anschluss daran wurde die Spitzenoberfläche durch
die kontinuierliche Zugabe von zweimal 5 μl 10 mMol/l Glycin-HCl-Puffer
(pH 1,5) gewaschen. Die immobilisierte Menge des FGF-8-Peptids entsprach
35 RU.
-
Jeweils
60 μl der
Antikörperlösung wurden
in einer Durchflussrate von 20 μl/Minute
in die das immobilisierte FGF-8-Peptid enthaltende Durchflusszelle
eingebracht, und anschließend
wurde die Dissoziationsreaktion während 3 Minuten überwacht.
Im Anschluss an die Dissoziationsreaktion wurde die Spitzenoberfläche durch
die kontinuierliche Zugabe von zweimal 20 μl 10 mMol/l Glycin-HCl-Puffer
(pH 1,5) regeneriert. Dieser Zyklus wurde für Antikörperlösungen mit unterschiedlichen
Konzentrationen (50 bis 1,85 μg/ml)
durchgeführt, wobei
für jede
Konzentration ein Sensorgramm erhalten wurde. Das Sensorgramm jedes
Antikörpers
wurde durch Subtraktion des Sensorgramms, welches unter Verwendung
des chimären
Antikörpers
KM871 gegen CD3 [Cancer Immunol. Immunother. 36 (1993), 373], als
eine negative Kontrolle erhalten wurde, in ein Sensorgramm für die spezifische
Reaktion überführt.
-
Das
Sensorgramm für
16,7 μg/ml
jedes Antikörpers
ist in 12 gezeigt. Wie aus dem Sensorgramm ersichtlich
ist, wurde zum Zeitpunkt der Dissoziationsreaktion jedes Antikörpers praktisch
keine Dissoziation beobachtet, so dass es schwierig war, eine exakte
Dissoziationskonstante zu erhalten. Demgemäß wurde die Bindungsaktivität der verschiedenen
Antikörper
durch den Vergleich der Stärke
der Bindung [Resonanzsignal (RU)] zum Zeitpunkt der Bindungsreaktion
durchgeführt.
-
Als
Ergebnis, wie in 12 gezeigt, zeigten die chimären Antikörper KM3034
und HV0LV6/CHO die stärkste
Bindungsreaktion, gefolgt der Reihe nach von HV0LV3-1/CHO, HV0LV4-1/CHO
und HV0LV2-1/CHO. Dagegen zeigten HV0LV3-2/CHO, HV0LV2-2/CHO und
HV0LV4-2/CHO eine geringe Bindungsreaktion, und HV0LV0/CHO zeigte
die geringste Bindungsreaktion. Diese Ergebnisse stimmten mit den
Ergebnissen unter Verwendung der Anti-FGF-8-CDR-transplantierten
Antikörper
von YB2/0-Zellen, welche unter Punkt 7 von Beispiel 2 beschrieben
wurden, überein.
-
8. Messung der Neutralisierungsaktivität der Anti-FGF-8-CDR-transplantierten
Antikörper
gegenüber
FGF-8
-
Die
Beurteilung der Neutralisierungsaktivität der vier Anti-FGF-8-CDRtransplantierten
Antikörper HV0LV6/CHO,
HV0LV3-1/CHO, HV0LV4-1/CHO und HV0LV2-1/CHO, deren starke Bindungsreaktion
gegenüber
FGF-8 unter Punkt 7 von Beispiel 3 bestätigt wurde, erfolgte gemäß dem unter
Punkt 5 von Beispiel 2 beschriebenen Verfahren. Der chimäre Anti-FGF-8-Antikörper KM3034
aus CHO/DG44-Zellen, welcher unter Punkt 3(1) von Beispiel 1 erhalten
wurde, wurde als eine positive Kontrolle verwendet, und der chimäre Antikörper KM2760
gegen das menschliche Chemokin CCR4, beschrieben in
WO 01/64754 , wurde als eine negative
Kontrolle verwendet. Die Ergebnisse sind in
13 gezeigt.
Wie in
13 gezeigt, zeigte HV0LV6/CHO eine
FGF-8-Neutralisierungsaktivität,
welche mit derjenigen des chimären
Antikörpers.
KM3034 vergleichbar war; und HV0LV3-1/CHO zeigte die nächst höhere FGF-8-Neutralisierungsaktivität. HV0LV4-1/CHO
zeigte eine etwas geringere FGF-8-Neutralisierungsaktivität als HV0LV3-1/CHO;
und HV0LV2-1/CHO zeigte die geringste Neutralisierungsaktivität.
-
Beispiel 4
-
Herstellung eines Anti-FGF-8-CDR-transplantierten
Antikörpers
mit einer geringeren Immunogenität
(2)
-
Anhand
der Ergebnisse von Beispiel 4 wurde gezeigt, dass von den Modifikationen
der 6 Aminosäurereste
von LV6 die Modifikation an Position 51 für die Wiederherstellung der
Aktivität
essentiell ist. Die Ergebnisse deuteten auch darauf hin, dass die
Modifikation an Position 2 einen geringen Einfluss auf die Wiederher stellung
der Aktivität
durch die alleinige Modifikation hat, aber dass sie zusammenwirkend
zu der Wiederherstellung der Aktivität durch die Kombination davon
mit der Modifikation an Position 51 beiträgt. Im Hinblick auf die Modifikationen
an den Positionen 14 und 15 wurde deutlich, dass sie ebenfalls zusammenwirkend
zu der Wiederherstellung der Aktivität durch die Kombination davon
mit der Modifikation an Position 51 beitragen. Dagegen wurde deutlich,
dass der Einfluss der Modifikation an Position 50 gering ist. Um
zu untersuchen, ob die Modifikation an Position 2 oder die Modifikation
an Position 14 oder 15 einen größeren Einfluss
auf die Wiederherstellung der Aktivität hat, und um den Einfluss
der Modifikation an Position 92 zu untersuchen, wurden daher noch
einmal Anti-.FGF-8-CDR-transplantierte Antikörper hergestellt.
-
1. Erneute Konstruktion von
Aminosäuresequenzen
für die
VL-Region
-
Es
wurden Aminosäuresequenzen
für zwei
VL-Regionen mit den folgenden Modifikationen, welche sich jeweils
von den Aminosäureresten
von LV.0 unterscheiden, konstruiert.
-
In
LV.4-3 wurden 4 Reste, umfassend Thr in Position 14, Pro in Position
15, Leu in Position 51 und Tyr in Position 92, durch die Reste Ser,
Leu, Val bzw. Phe, welche den Aminosäureresten entsprechen, die
in dem Mausantikörper
KM1334 vorhanden sind, ersetzt.
-
In
LV.3-3 wurden 3 Reste, umfassend Thr in Position 14, Pro in Position
15 und Leu in Position 51, durch die Reste Ser, Leu bzw. Val, welche
den Aminosäureresten
entsprechen, die in dem Mausantikörper KM1334 vorhanden sind,
ersetzt.
-
2. Konstruktion einer cDNA, codierend
die VL-Region
-
Die
cDNAs, welche die jeweiligen unter Punkt 1 von Beispiel 4 konstruierten
Aminosäuresequenzen für die VL-Region
der Anti-FGF-8-CDR-transplantierten Antikörper codieren, wurden wie folgt
konstruiert.
-
(1) Konstruktion einer cDNA, codierend
LV.4-3
-
Diese
cDNA wurde gemäß dem unter
Punkt 2(5) von Beispiel 3 beschriebenen Verfahren konstruiert. Jedoch
wurde das Plasmid pKM1334LV2-1, welches unter Punkt 2(3) von Beispiel
3 erhalten wurde, anstelle des Plasmids pKM1334LV2-2 verwendet,
und das Plasmid pKM1334LV3-2, welches unter Punkt 2(6) von Beispiel
3 erhalten wurde, wurde anstelle des Plasmids pKM1334LV6 verwendet.
Als Ergebnis wurde das Plasmid pKM1334LV4-3 erhalten, welches die
LV.4-3 codierende cDNA enthält.
-
(2) Konstruktion einer cDNA, codierend
LV.3-3
-
Zunächst wurden
3 μg des
Plasmids pKM1334LV4-3, welches unter Punkt 2(1) von Beispiel 4 erhalten wurde,
mit 10 Einheiten des Restriktionsenzyms BamHI (hergestellt von Takara
Shuzo) und 10 Einheiten des Restriktionsenzyms SpeI (hergestellt
von Takara Shuzo) gemischt und bei 37°C für 1 Stunde umgesetzt. Die Reaktionslösung wurde
durch eine Agarosegelelektrophorese fraktioniert, wobei etwa 2,5 μg eines BamHI-SpeI-Fragments
von etwa 3,23 kb gewonnen wurden.
-
Anschließend wurden
3 μg des
Plasmids pKM1334LV0, welches unter Punkt 1(3) von Beispiel 2 erhalten
wurde, mit 10 Einheiten des Restriktionsenzyms BamHI (hergestellt
von Takara Shuzo) und 10 Einheiten des Restriktionsenzyms SpeI (hergestellt
von Takara Shuzo) bei 37°C
für 1 Stunde
umgesetzt. Die Reaktionslösung
wurde durch eine Agarosegelelektrophorese fraktioniert, wobei etwa
0,15 μg
eines BamHI-SpeI-Fragments von etwa 0,13 kb gewonnen wurden.
-
Zu
einem Gesamtvolumen von 10 μl
sterilem Wasser wurden 0,1, μg
des BamHI-SpeI-Fragments des Plasmids pKM1334LV4-3 und 0,1 μg des BamHI-SpeI-Fragments des Plasmids
pKM1334LV0, welche jeweils vorstehend erhalten wurden, zugegeben
und unter Verwendung von Ligation High (hergestellt von TOYOBO) ligiert.
Unter Verwendung der so erhaltenen rekombinanten Plasmid-DNA-Lösung wurden
Zellen von E. coli DH5α transformiert,
wobei das in 14 dargestellte Plasmid pKM1334LV3-3
erhalten wurde, welches die LV.3-3 codierende cDNA enthält.
-
3. Konstruktion von Expressionsvektoren
für einen
Anti-FGF-8-CDR-transplantierten Antikörper
-
Die
Expressionsvektoren für
einen Anti-FGF-8-CDR-transplantierten Antikörper, welche die verschiedenen
VL-cDNAs enthalten, wurden durch das Ersetzen des EcoRI-BsiWI-Fragments,
das die VL-cDNA des Expressionsvektors pKANTEX1334HV0LV6 enthält, erhalten
unter Punkt 2 von Beispiel 2, durch die EcoRI-BsiWI- Fragmente, enthaltend
die verschiedenen VL-cDNAs, welche unter Punkt 2 von Beispiel 4
konstruiert wurden, konstruiert. Genauer wurden die zwei Vektoren
pKANTEX1334HV0LV4-3 und pKANTEX1334HV0LV3-3 konstruiert.
-
4. Stabile Expression der Anti-FGF-8-CDR-transplantierten
Antikörper
unter Verwendung von CHO/DG44-Zellen
-
Die
stabile Expression der verschiedenen Anti-FGF-8-CDR-transplantierten
Antikörper
in CHO/DG44-Zellen wurde gemäß dem unter
Punkt 2(4) von Beispiel 1 beschriebenen Verfahren unter Verwendung
der Expressionsvektoren für
die verschiedenen Anti-FGF-8-CDR-transplantierten Antikörper, welche
unter Punkt 3 von Beispiel 4 erhalten wurden, bewirkt.
-
5. Reinigung der Anti-FGF-8-CDR-transplantierten
Antikörper
-
Die
Züchtung
der Transformanten, erhalten unter Punkt 4 von Beispiel 4, welche
von den CHO/DG44-Zellen erhalten wurden und welche die verschiedenen
Anti-FGF-8-CDR-transplantierten Antikörper exprimieren, sowie die
Reinigung der Anti-FGF-8-CDR-transplantierten Antikörper aus
den Kulturüberständen wurden
gemäß dem unter
Punkt 3(1) von Beispiel 1 beschriebenen Verfahren durchgeführt. Der
Antikörper von
einer Transformante, in die pKANTEX1334HV0LV4-3 eingeführt wurde,
wurde als HV0LV4-3/CHO bezeichnet; und der Antikörper von einer Transformante,
in die pKANTEX1334HV0LV3-3 eingeführt wurde, wurde als HV0LV3-3/CHO
bezeichnet.
-
6. Analyse der gereinigten Anti-FGF-8-CDR-transplantierten
Antikörper
-
Die
SDS-PAGE unter Verwendung der verschiedenen Anti-FGF-8-CDRtransplantierten
Antikörper, welche
unter Punkt 5 von Beispiel 4 erhalten wurden, wurde gemäß dem unter
Punkt 4 von Beispiel 1 beschriebenen Verfahren durchgeführt. Als
Ergebnis wurde bestätigt,
dass jeder Antikörper
als ein Antikörpermolekül in der
richtigen Struktur exprimiert und gereinigt wurde.
-
7. Messung der Bindungsaktivität der Anti-FGF-8-CDR-transplantierten
Antikörper
gegenüber
FGF-8
-
Die
FGF-8-Bindungsaktivitäten
der Anti-FGF-8-CDR-transplantierten Antikörper HV0LV6/CHO und HV0LV3-1/CHO,
welche unter Punkt 5 von Beispiel 3 erhalten wurden, und der Anti-FGF-8-CDR-transplantierten
Antikörper
HV0LV4-3/CHO und HV0LV3-3/CHO, welche unter Punkt 5 von Beispiel
4 erhalten wurden, wurden gemäß dem unter
Punkt 7 von Beispiel 3 beschriebenen Verfahren gemessen. Der chimäre Anti-FGF-8-Antikörper KM3034
aus CHO/DG44-Zellen, welcher unter Punkt 3(1) von Beispiel 1 erhalten
wurde, wurde als eine positive Kontrolle verwendet.
-
Das
Sensorgramm für
16,7 μg/ml
jedes Antikörpers
ist in 15 gezeigt. Wie aus dem Sensorgramm ersichtlich
ist, wurde zum Zeitpunkt der Dissoziationsreaktion jedes Antikörpers praktisch
keine Dissoziation beobachtet, so dass es schwierig war, eine exakte
Dissoziationskonstante zu erhalten. Demgemäß wurde die Bindungsaktivität der jeweiligen
Antikörper
durch den Vergleich der Stärke
der Bindung [Resonanzsignal (RU)] zum Zeitpunkt der Bindungsreaktion
durchgeführt.
Als Ergebnis, wie in 15 gezeigt, zeigte der chimäre Antikörper KM3034
die stärkste
Bindungsreaktion; und HV0LV4-3/CHO zeigte eine Bindungsreaktion,
welche stärker
als diejenige von HV0LV3-1/CHO war und welche mit derjenigen von
HV0LV3-3/CHO vergleichbar war. Die Bindungsreaktion von HV0LV3-3/CHO
war geringer als diejenige von HV0LV3-1. Anhand der vorstehenden
Ergebnisse wurde deutlich, dass im Hinblick auf die Stärke der
Bindungsreaktion die Modifikation an Position 14 oder 15 stärker kooperativ
wirksam ist als die Modifikation an Position 2 und dass die Modifikation
an Position 92 für
die Wiederherstellung der Aktivität essentiell ist.
-
8. Messung der Neutralisierungsaktivität der Anti-FGF-8-CDR-transplantierten
Antikörper
gegenüber
FGF-8
-
Die
Beurteilung der Neutralisierungsaktivität für die Anti-FGF-8-CDR-transplantierten
Antikörper HV0LV6/CHO
und HV0LV3-1/CHO, welche unter Punkt 5 von Beispiel 3 erhalten wurden,
und der Anti-FGF-8-CDR-transplantierten Antikörper HV0LV4-3/CHO und HV0LV3-3/CHO,
welche unter Punkt 5 von Beispiel 4 erhalten wurden, wurde gemäß dem unter
Punkt 5 von Beispiel 2 beschriebenen Verfahren durchgeführt. Der
chimäre
Anti-FGF-8-Antikörper
KM3034 aus CHO/DG44-Zellen, welcher unter Punkt 3(1) von Beispiel
1 erhalten wurde, wurde als eine positive Kontrolle verwendet, und
der chimäre
Antikörper
KM2760 gegen das menschliche Chemokin CCR4, beschrieben in
WO 01/64754 , wurde als
eine negative Kontrolle verwendet. Die Ergebnisse sind in
16 gezeigt. Wie in
16 dargestellt,
zeigten HV0LV6/CHO und HV0LV3-1/CHO eine FGF-8-Neutralisierungsaktivität, welche
mit derjenigen des chimären
Antikörpers KM3034
vergleichbar war. Dagegen zeigten HV0LV4-3/CHO und HV0LV3-3/CHO
nahezu die gleiche Neutralisierungsaktivität, wobei die Aktivität etwa der
Hälfte
der Aktivität
des chimären
Antikörpers
KM3034 entsprach. Die Neutralisierungsaktivität von HV0LV3-1/CHO und HV0LV4-3/CHO
zeigte keine Korrelation zu der Stärke der Bindungsreaktion unter
Verwendung des BIAcore-Systems, wobei deutlich wurde, dass der Aminosäurerest
in Position 2 und die Aminosäurereste
in den Positionen 14 und 15 unabhängig voneinander die Bindungsaktivität gegenüber FGF-8
und die FGF-8-Neutralisierungsaktivität gegenüber Zellen beeinflussen.
-
Basierend
auf den vorstehenden Ergebnissen der jeweiligen Beurteilungen wurde
der aus CHO/DG44-Zellen erhaltene CDR-transplantierte Antikörper HV0LV6/CHO,
welcher eine starke Bindungsreaktion zeigte und welcher eine mit
dem chimären
Antikörper
KM3034 vergleichbare FGF-8-Neutralisierungsaktivität aufwies,
als KM8034 bezeichnet; und in gleicher Weise wurde der von CHO/DG44-Zellen erhaltene transformierte
Zellclon, welcher eine starke Expression von KM8034 zeigt, als KM8034
bezeichnet. Der aus CHO/DG44-Zellen erhaltene CDR-transplantierte
Antikörper
HV0LV4-3/CHO, der eine starke Bindungsreaktion zeigte, welche mit
derjenigen von KM8034 vergleichbar war, wurde als KM8035 bezeichnet;
und in gleicher Weise wurde die von CHO/DG44-Zellen abgeleitete
transformierte Zelllinie, welcher eine starke Expression von KM8035
zeigt, als KM8035 bezeichnet. Die VL-Aminosäuresequenz LV.4-3 von KM8035
ist in SEQ ID NO: 38 dargestellt. Ferner ist die transformierte
Zelllinie KM8035 am 20. Juni 2002 als FERM BP-8082 bei der International
Patent Organism Depositary, National Institute of Advanced Industrial
Science and Technology (AIST Tsukuba Central 6, 1–1, Higashi
1-Chorre Tsukubashi, Ibaraki-ken 305–8566, Japan), hinterlegt worden. Weiterhin
wurde der aus CHO/DG44-Zellen erhaltene CDR-transplantierte Antikörper HV0LV3-1/CHO,
der eine starke Bindungsreaktion zeigte, welche mit derjenigen von
KM8034 vergleichbar war, als KM8036 bezeichnet; und in gleicher
Weise wurde die von CHO/DG44-Zellen abgeleitete transformierte Zelllinie,
welcher eine starke Expression von KM8036 zeigt, als KM8036 bezeichnet.
Die VL-Aminosäuresequenz
LV.3-1 von KM8036 ist in SEQ ID NO: 39 dargestellt. Die transformierte
Zelllinie KM8036 ist am 20. Juni 2002 als FERM BP-8083 bei der International
Patent Organism Depositary, National Institute of Advanced Industrial
Science and Technology (AST Tsukuba Central 6, 1–1, Higashi 1-Chorre Tsukuba-shi,
Ibaraki-ken 305–8566,
Japan), hinterlegt worden.
-
Da
der Anti-FGF-8-CDR-transplantierte Antikörper KM8034 eine starke Bindungsreaktion
und eine mit dem chimären
Antikörper
KM3034 vergleichbare FGF-8-Neutralisierungsaktivität zeigt,
und der Antikörper
eine geringere Immunogenität
bei Menschen als der chimäre
Antikörper
aufweist, wird erwartet, dass der Antikörper eine bessere therapeutische
Wirkung als der chimäre
Antikörper
zeigt. Obwohl die Möglichkeit
besteht, dass die Anti-FGF-8-CDR-transplantierten Antikörper KM8035
und KM8036 im Vergleich zu KM8034 eine etwas geringere Bindungsaktivität und FGF-8-Neutralisierungsaktivität aufweisen,
wird eine geringere Immunogenität als
bei KM8034 erwartet, da die Aminosäurereste in der FR-Region der
V-Region, welche von dem Mausantikörper KM1334 stammen, 3 Reste
bzw. 4 Reste umfassen.
-
Gewerbliche Anwendbarkeit
-
Die
vorliegende Erfindung stellt einen humanisierten Antikörper oder
ein Antikörperfragmente
davon bereit, wobei der humanisierte Antikörper oder das Antikörperfragmente
davon spezifisch mit FGF-8 reagiert, welcher wahrscheinlich ein
Wachstumsfaktor für
Prostatakrebs, Brustkrebs, Eierstockkrebs und dergleichen ist, und
die Funktion von FGF-8 inhibiert.
-
Freier
Text der Sequenzprotokolle:
- SEQ ID NO: 11 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 12 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 13 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 14 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 20 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 21 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 22 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 23 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 24 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 25 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 26 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 27 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 28 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 29 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 30 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 31 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 32 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 33 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 34 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 35 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 36 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
- SEQ ID NO: 37 – Beschreibung
der künstlichen
Sequenz: Synthetische DNA
SEQUENZPROTOKOLL