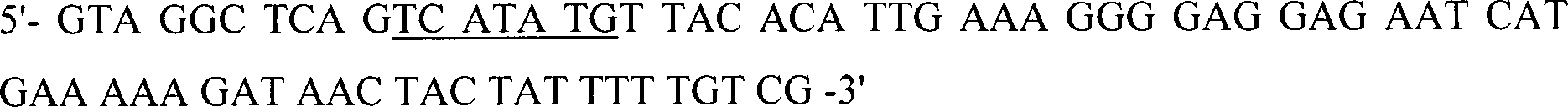-
Die
vorliegende Erfindung betrifft einen transformierten Bacillus-Wirt,
der in der Lage ist, ein Cellulose-Bindedomäne-Polypeptid zu codieren,
einen Bacillus-Expressionsvektor und ein Verfahren zur Produktion einer
Cellulose-Bindedomäne
in einer Bacillus-Wirtszelle.
-
HINTERGRUND DER ERFINDUNG
-
Die
Fokussierung auf die CBD als eine funktionelle Domäne hat die
Synthese der Domäne
als ein Einzeldomänenmolekül mit eingeschlossen.
Eine der ersten reinen CBDs wurde erhalten, indem sie durch automatische
Festphasensynthese (Kraulis P. et al. (1989) synthetisiert wurde.
-
Es
wurde gezeigt, dass CBDs in E. coli als funktionelle Einzeldomänen exprimiert
werden können,
siehe z. B.: Ong E. et al. (1993), wo offenbart ist, dass die Expression
in das Periplasma der Zellen unter Verwendung von E. coli zu einer
Ausbeute von 33 mg CBD pro Liter Kulturflüssigkeit führt.
-
Neuerdings
wurde auch eine Pilz-Doppel-CBD (ein Dimer) erfolgreich in E. coli
exprimiert, siehe Linder M. et al. (1996).
-
WO 96/13524 offenbart modifizierte
CBD Proteine, die in E. coli exprimiert werden.
-
Allerdings
ist die Expression der CBDs in E. coli keine echte extrazelluläre Expression
und führt
zu einer unzufriedenstellenden Ausbeute, die für die großtechnische Produktion von
CBD zu gering ist.
-
US 5,525,195 ,
US 5,536,655 ,
WO 91/17244 und
WO 91/10732 offenbaren die Expression
in einer Bacillus-Wirtszelle eines Endoglucanase-Enzyms, bei dem
die katalytisch aktive Domäne
operabel mit einer Cellulose-Bindedomäne verknüpft ist.
-
Demgemäß besteht
die Aufgabe der vorliegenden Erfindung darin, ein Verfahren zur
Produktion von CBD in einer hohen Ausbeute, vorzugsweise mittels
einer herkömmlichen
Fermentationstechnik, die die extrazelluläre Produktion der CBD umfasst,
die wiederum die Verwendung von CBD bei industriellen Anwendungen wirtschaftlich
machbar macht, bereitzustellen.
-
ZUSAMMENFASSUNG DER ERFINDUNG
-
Die
Erfinder haben nun gefunden, dass die Produktion von Cellulose-Bindedomänen (CBDs)
durch Expression in einem Bacillus-Wirt möglich ist.
-
Vor
der vorliegenden Erfindung war die Expression einer CBD ohne eine
katalytisch aktive Domäne im
Bacillus sehr unerwartet, da erstens Cellulose-Bindedomänen bekanntlich
Disulfid-Brücken
enthalten und da sie zweitens potentiell dem Abbau durch Proteasen,
die durch den Bacillus-Wirt produziert werden, unterliegen.
-
Demnach
betrifft die vorliegende Erfindung in ihrem ersten Aspekt einen
Bacillus-Wirt, der mit einem Vektor transformiert ist, der eine
DNA-Sequenz umfasst, die eine Cellulose-Bindedomäne codiert und zur Expression
der DNA-Sequenz in der Lage ist.
-
In
einem zweiten Aspekt betrifft die Erfindung einen Bacillus-Expressionsvektor,
der eine inserierte DNA-Sequenz trägt, die eine Cellulase-Bindedomäne trägt.
-
Weiterhin
betrifft die Erfindung in ihrem dritten Aspekt ein Verfahren zur
Herstellung eines Cellulose-Bindedomäne-Polypeptids in einer Bacillus-Wirtszelle,
wobei das Verfahren die Schritte umfasst
- – Wachsen
lassen in einem Nährmedium
unter Bedingungen, um Cellulose-Bindedomäne überzuproduzieren, von Bacillus-Wirtszellen,
die mit einer Expressionskassette transformiert wurden, welche als
funktionsfähig
verbundene Bestandteile einschließt:
a) eine regulatorische
Region zur Initiation der Transkription und Translation,
b)
eine DNA-Sequenz, die das Cellulose-Bindedomäne-Polypeptid codiert,
c)
eine regulatorische Region zur Termination der Transkription und
Translation, wobei die regulatorischen Regionen in dem Wirt funktionell
sind,
d) ein Selektionsmarkergen zum Selektionieren transformierter
Wirtszellen; und
- – Gewinnen
des Cellulose-Bindedomäne-Polypeptids.
-
AUSFÜHRLICHE BESCHREIBUNG DER ERFINDUNG
-
Eine
Cellulose-Bindedomäne
(CBD) ist ein Polypeptid, das eine hohe Affinität gegenüber wasserunlöslichen
Formen von Cellulose und Chitin, einschließlich kristalliner Formen besitzt
oder an sie bindet.
-
CBDs
kommen als integrale Teile von großen Proteinkomplexen vor, die
aus zwei oder mehr verschiedenen Polypeptiddomänen bestehen, beispielsweise
in hydrolytischen Enzymen (Hydrolasen, die typischerweise aus einer
katalytischen Domäne
bestehen, die die aktive Stelle für die Substrathydrolyse und
eine Kohlehydrat-Bindedomäne
oder Cellulose-Bindedomäne (CBD)
zum Binden an die unlösliche
Matrix enthält.
Solche Enzyme können
mehr als eine katalytische Domäne
und ein, zwei oder drei CBDs und gegebenenfalls eine oder mehrere
Polypeptidregionen umfassen, die die CBDs mit der (den) katalytischen
Domäne(n)
verknüpfen, wobei
letztere Regionen in der Regel als ein „Linker" bezeichnet werden. Beispiele für hydrolytische
Enzyme, die eine CBD umfassen, sind Cellulasen, Xylanasen, Mannanasen,
Arabinofuranosidasen, Acetylesterasen und Chitinasen. CBDs wurden
auch in Algen gefunden, z. B. in der Rotalge Porphyra purpurea als
ein nicht-hydrolytisches
Polysaccharide-Bindeprotein, siehe Peter Tomme et al. (1996). Allerdings
stammen die meisten der bekannten CBDs aus Cellulasen und Xylanasen.
-
In
diesem Zusammenhang soll der Begriff „Cellulose-Bindedomäne" wie von Tomme et
al., op. cit. definiert verstanden werden. Diese Definition klassifiziert
mehr als 120 Cellulose-Bindedomänen in 10
Familien (I-X), die verschiedene Funktionen oder Rollen im Zusammenhang
mit dem Mechanismus des Substratsbindens besitzen können. Während der
Arbeit, die zu der vorliegenden Erfindung führt, wurde allerdings eine
bisher unbekannte CBD-Familie gefunden, cf. Beispiel 8 nachstehend;
und es wird vermutet, dass in der Zukunft neue Familienstellvertreter
und zusätzliche
CBD-Familien auftreten werden.
-
In
dem Proteinkomplex ist typischerweise ein hydrolytisches Enzym,
eine CBD, an dem N- oder
C-Terminus lokalisiert oder ist intern.
-
Eine
monomere CBD besteht typischerweise aus mehr als etwa 30 und weniger
als etwa 250 Aminosäureresten.
Beispielsweise besteht eine in Familie I klassifizierte CBD aus
33–37
Aminosäureresten;
eine in Familie IIa klassifizierte CBD besteht 95–108 Aminosäureresten;
und eine in Familie VI klassifizierte CBD besteht aus 85–92 Aminosäureresten.
Demnach liegt das Molekulargewicht einer monomeren CBD typischerweise
im Bereich von etwa 4 kD bis etwa 40 kD, und in der Regel unter
etwa 35 kD.
-
CBDs
können
als ein Einzeldomänen-Polypeptid
oder als ein Dimer, ein Trimer oder ein Polymer; oder als ein Teil
eines Protein-Hybrids verwendbar sein.
-
Chimäre
Protein-Hybride
-
Chimäre Protein-Hybride
sind auf dem Fachgebiet bekannt, siehe z. B.
WO 90/00609 ,
WO 94/24158 und
WO 95/16782 , und umfassen eine Cellulose-Bindedomäne (CBD)
aus einem anderen Ursprung, vorzugsweise aus einem anderen mikrobiellen
Ursprung, als das Chimäre
Protein als solches, wobei die CBD als ein integraler Teil des Proteins
existiert. Typischerweise sind die Chimären Protein-Hybride Enzym-Hybride,
das heißt,
sie enthalten eine katalytische Domäne zusammen mit der Bindedomäne.
-
Chimäre Protein-Hybride
und Enzym-Hybride können
durch Transformieren eines DNA-Konstrukts, welches
mindestens ein DNA-Fragment umfasst, das die Cellulose-Bindedomäne (CBD)
codiert, welches, mit oder ohne einen Linker mit einer DNA-Sequenz
ligiert ist, die das Protein oder Enzym codiert, in eine Wirtszelle hergestellt
werden und durch Wachsenlassen der Wirtszelle, um das fusionierte
Gen zu exprimieren. Das rekombinante Fusionsprotein oder die Enzym-Hybride
können
durch die folgende Formel beschrieben werden: CBD-MR-X
-
Wobei
CBD die N-terminale oder die C-terminale Region einer Aminosäuresequenz
ist, die mindestens der Cellulose-Bindedomäne entspricht; MR die mittlere
Region (der Linker), und eine Bindung oder eine kurze Verknüpfungsgruppe
sein kann, vorzugsweise von etwa 2 bis etwa 100 Kohlenstoffatomen,
stärker
bevorzugt von zwei bis 40 Kohlenstoffatomen; oder vorzugsweise von
etwa 2 bis etwa 100 Aminosäuren,
stärker bevorzugt
von 2 bis 40 Aminosäuren;
und X eine N-terminale oder C-terminale Region eines Polypeptids
ist, welches durch die DNA-Sequenz codiert wird, die das Protein
oder Enzym codiert.
-
Das
rekombinante Fusionsprotein oder die Enzym-Hybride mit einer internen
CBD werden allerdings ebenfalls berücksichtigt.
-
Eine
DNA-Sequenz, die eine CBD aus einem gegebenen Organismus codiert,
kann herkömmlicherweise
unter Anwendung der PCR-Techniken und auch auf der Grundlage der
derzeitigen Erkenntnis erhalten werden, dass es möglich ist,
homologe Sequenzen aus anderen Organismen zu finden.
-
Es
wird davon ausgegangen, dass neue CBDs durch Klonieren von Cellulasen,
Xylanasen oder einem anderen Pflanzenzellwand-Abbauenzym und durch
Messen des Bindens an Cellulose gefunden werden können. Wenn
die Enzymaktivität
unter den nachstehend beschriebenen Standardbedingungen an Avicel
gebunden ist, kann angenommen werden, dass ein Teil des Gens eine
Bindedomäne
codiert.
-
Nach
Erhalt des DNA-Fragments, das eine CBD codiert, wird das DNA-Gen
in einen Vektor inseriert, der für
seine Expression in Bacillus spp geeignet ist.
-
Beispielsweise
kann die Celluloseaffinität
unter Verwendung von 10 g Avicel in einer 500 ml gepufferten Aufschlämmung (Puffer:
0,1 Natriumphosphat, pH 7,5) gemessen werden, welche langsam unter
Verwendung eines Löffels
gerührt
und 30 Minuten bei Raumtemperatur quellen gelassen wird. Anschließend wird
das Enzym in einem Verhältnis
von 1 Teil Cellulose-Bindedomäne
zu 150 Teilen Avicel zugesetzt. Dies erfolgt auf Eis, was innerhalb
5 bis 10 Minuten ein optimales Binden ergibt. Das Avicel kann sodann
gewaschen und zur Sichtbarmachung der gebundenen Proteine direkt
auf SDS-PAGE aufgebracht werden (da die Verwendung von SDS und Kochen
die gebundenen Proteine freisetzen wird). Alternativ wird die Aufschlämmung in
eine Säule
gepackt und gewaschen. Das gebundene Protein wird eluiert, entweder
in ionisiertem Wasser oder in einem Puffer mit hohem pH, wie Triethylamin
(pH 11,2; 1% Lösung),
wobei das pH-eluierte Protein schnell auf neutral eingestellt wird.
-
Mehrere
CBDs wurden bereits in E. coli exprimiert, allerdings wurde nicht
von einer Expression und Sezernierung aus Bacillus sp berichtet.
E. coli besitzt als ein Expressionswirt für heterologe Proteine mehrere Vorteile
gegenüber
Bacillus spp., zum einen, da E. coli einen periplasmatischen Raum
aufweist, wo ein ordnungsgemäßes Falten
von heterologen exprimierten Genen möglich ist (zur Übersicht
siehe beispielsweise Hockney, R. C. (1994)). Insbesondere das Oxidationspotential
und die Existenz von Disulfid-Oxidoreductasen in dem Periplasma
sind notwendig, wenn Proteine mit einer Funktionalität exprimiert
werden, die von ordnungsgemäß angeordneten
Disulfidbrücken
abhängig
ist. (Emmanuel Brun et al. (1995)). Überproduktion, Reinigung und
Charakterisierung der Cellulose-Bindedomäne der Erwinia chrysanthemi-sezernierten
Endoglucanase EGZ ist in Eur. J. Biochem 231, 142–148, und
Ong et al., (1993) offenbart. Weitere Beispiele für CBDs mit
Disulfid-Bindungen sind: die N-terminale CBD von Ce1B aus Pseudomonas
fluorescens subsp. cellulosa (NCIMB 10462) (siehe den Abgleich in
Tomme P. et al., op. cit., und die N-terminale CBD von CenA aus
Cellulomonas fimi (ATCC 484), N. R. Gilkes et al. (1991).
-
Weiterhin
wirkt das Periplasma von E. coli als Schutz des heterelog exprimierten
Proteins gegenüber der
Wirkung von Proteasen, die sowohl im Überstand als auch im Cytoplasma
vorhanden sind.
-
Es
ist auch bekannt, dass bei der Expression von sezernierten Proteinen
mit Disulfidbrücken
in Bacillus subtilis das Expressionsniveau signifikant abfällt. (van
den Berg et al. (1993)).
-
Ein
weiteres Problem bei heterologer Expression besteht im proteolytischen
Abbau des exprimierten Proteins. Bacillus subtilis exprimiert bekanntlich
mindestens 7 verschiedene extrazelluläre Proteasen (Hrsg. A. L. Sonenshein
et al. (1993)).
-
Insbesondere
für die
CBDs, die sehr stark hydrophobe Proteine sind, könnte die Translokation des Proteins
bei Expression in Bacillus subtilis schwerwiegend beeinträchtigt werden
und infolge der nachteiligen Auswirkungen, wenn das Protein aufgrund
seiner Hydrophobizität
an der Zellmembran verankert wird, sogar den Zelltod herbeiführen.
-
In
ihrem ersten Aspekt betrifft die vorliegende Erfindung einen Bacillus-Wirt,
der mit einem Vektor transformiert ist, der eine DNA-Sequenz umfasst,
die eine CBD codiert, und in der Lage ist, die Sequenz zu exprimieren.
Offensichtlich besteht das exprimierte Polypeptid im Wesentlichen
aus einer oder mehreren nicht katalytischen Domänen, d. h. das Polypeptid umfasst
keine katalytisch aktive Domäne.
-
Bei
einer bevorzugten Ausführungsform
besitzt die exprimierte CBD oder das CBD-enthaltende Polypeptid ein Molekulargewicht
(Mw), das gleich oder höher
ist als etwa 4 kD. Vorzugsweise ist das Mw gleich oder geringer
als etwa 35 kD, stärker
bevorzugt etwa 32 kD, noch stärker
bevorzugt etwa 30 kD, insbesondere etwa 25 kD.
-
Die
CBD kann in der Form eines Einzeldomänen-Polypeptids exprimiert
werden, d. h. ein Polypeptid, das eine CBD umfasst. Alternativ kann
die CBD in der Form eines Dimers oder Trimers oder sogar eines Polymers
exprimiert werden, d. h. ein Polypeptid oder Protein, das zwei,
drei oder sogar mehr als drei identische CBD-„Einheiten” einschließt.
-
Die
CBD kann auch als ein Teil eines Mehrdomänen-Polypeptids exprimiert
werden, wobei der Nicht-CBD-Teil eines solchen Polypeptids beispielsweise
eine, zwei oder sogar mehr Domänen
ohne katalytische Aktivität
ist.
-
Es
wird angenommen, dass fast jede CBD erfindungsgemäß exprimiert
werden kann, d. h. mittels eines tranformierten Bacillus-Wirts.
Vorzugsweise werden solche CBDs exprimiert, die aus einem Mikroorganismus
oder einer Pflanze, stärker
bevorzugt aus einem Bakterium oder aus einem Pilz, erhältlich sind.
-
Beispiele
für CBDs
aus Bakterien umfassen CBDs, die aus Spezies erhältlich sind, die zu einer der folgenden
Arten gehören:
Butyrivibrio,
Cellulomonas, Clostridium, Microbispora, Micromonospora, Pseudomonas,
Streptomyces, Thermomonospora, Bacillus, Caldocellum, Erwinia, Myxococcus,
Cellvibrio, Thermoanaerobacterium, und Thermotoga.
-
Beispiele
für CBDs
aus Pilzen umfassen CBDs, die aus Spezies erhältlich sind, die zu einer der
folgenden Arten gehören:
Agaricus, Dictyostelium, Fusarium, Humicola, Neocallimastix, Neurospora,
Limulus, Penicillium, Phanerochaete, und Trichoderma.
-
Beispiele
für CBDs,
die aus Pflanzen erhältlich
sind, sind CBDs aus Expansionen.
-
Der
erfindungsgemäße Bacillus-Wirt
ist ein neutralophiler, ein alkalophiler, ein mesophiler oder ein thermophiler
Wirt.
-
Beispiele
für Wirte,
die bei der vorliegenden Erfindung geeignet sind, sind Wirte aus
den Spezies Bacillus subtilis, Bacillus licheniformis, Bacillus
megaterium, Bacillus stearothermophilos, und Bacillus amyloliquefaciens.
Es wird allerdings davon ausgegangen, dass andere Bacillus-Spezies
ebenfalls geeignete Wirte zur Expression von CBDs sein können.
-
Wie
in weiteren Einzelheiten nachstehend beschrieben, wird der erfindungsgemäße Wirt
mit einem Vektor transformiert, der eine CBD umfasst, der eine CBD-codierende
DNA-Sequenz umfasst. Vorzugsweise ist der Vektor in ein Genom des
Wirts integriert, stärker
bevorzugt wurde er auf dem Genom amplifiziert.
-
Bei
einer anderen bevorzugten Ausführungsform
der Erfindung ist der Vektor als ein Expressionsplasmid, vorzugsweise
ein Multikopienplasmid, vorhanden.
-
In
einem zweiten Aspekt betrifft die vorliegende Erfindung einen Bacillus-Expressionsvektor,
der eine inserierte CBD-codierende DNA-Sequenz trägt. Vorzugsweise
umfasst die Expressionskassette des Vektors regulatorische Regionen
aus einem Bacillus sp., stärker
bevorzugt sind solche regulatorische Regionen endogen zu dem Wirt.
-
In
einem dritten Aspekt betrifft die vorliegende Erfindung ein Verfahren
zur Produktion eines CBD-Polypeptids, wobei das Verfahren die Schritte
umfasst:
- – Wachsen
lassen in einem Nährmedium
unter Bedingungen, um Cellulose-Bindedomäne überzuproduzieren, von Bacillus-Wirtszellen,
die mit einer Expressionskassette transformiert wurden, welche als
funktionsfähig
verbundene Bestandteile einschließt:
a) eine regulatorische
Region zur Initiation der Transkription und Translation,
b)
eine DNA-Sequenz, die das Cellulose-Bindedomäne-Polypeptid codiert,
c)
eine regulatorische Region zur Termination der Transkription und
Translation,
wobei die regulatorischen Regionen im Wirt funktionell
sind, und
d) ein Selektionsmarker-Gen zum Selektionieren transformierter
Wirtszellen; und
- – Gewinnen
des Cellulose-Bindedomäne-Polypeptids.
-
In
ihrem vierten Aspekt betrifft die Erfindung ein Verfahren zur Optimierung
der CBD-Expression
in einem Bacillus-Wirt, wobei das Verfahren die Schritte der Expression
in dem Wirt einer an ein Reportermolekül fusionierten CBD; und das
Aufzeichnen der Konzentration der exprimierten CBD in den Überstand
des fermentierten Wirts durch Messen der natürlichen Eigenschaft oder der
natürlichen
Eigenschaften des Reportermoleküls
umfasst.
-
Bei
einer bevorzugten Ausführungsform
ist das Reportermolekül
ein Green Fluorescent Protein, und die natürliche Eigenschaft ist die
Fluoreszenzemission.
-
In
ihrem fünften
und sechsten Aspekt betrifft die Erfindung ein Polypeptidhybrid,
bestehend im Wesentlichen aus einer oder mehreren Cellulose-Bindedomänen, die
an ein Green Fluorescent Protein fusioniert sind, und ein Verfahren
der Produktion eines solchen Hybrids durch Expression in einem Bacillus-Wirt,
Wachstum des transformierten Wirts unter Bedingungen, wobei die
transformierte Kultur im Wesentlichen frei von untransformierten
Zellen ist; Inkubation der transformierten Kultur in einem Nährmedium,
wobei das Hybrid überproduziert
wird; und Gewinnen des Hybrids.
-
EXPRESSION EINER CBD
-
Rekombinante Expressionsvektoren
-
Ein
rekombinanter Vektor, der ein DNA-Konstrukt umfasst, das die erfindungsgemäße CBD codiert, kann
jeder beliebige Vektor sein, der zweckmäßigerweise DNA-Rekombinationsverfahren
unterzogen werden kann, und die Wahl des Vektors hängt oft
von der Wirtszelle ab, in die er eingebracht werden soll. Das Einbringen
des Vektors in die Wirtszelle wird oft als die transformierte Wirtszelle
bezeichnet. Eine solche Transformation gibt die Einführung von
DNA in eine Wirtszelle unter Verwendung z. B. von Protoplasten,
natürlichen
kompetenten Zellen, Transfektion, Konjugation, Elektroporation,
oder von jedem gleichwertigen Verfahren an. Somit kann der Vektor
ein autonom replizierender Vektor sein, d. h. ein Vektor, der als
extrachromosomale Einheit existiert, dessen Replikation von der
chromosomalen Replikation unabhängig
ist, z. B. ein Plasmid.
-
Alternativ
kann der Vektor ein Vektor sein, der beim Einbringen in einer Wirtszelle
in das Wirtszellen-Genom zum Teil oder in seiner Gesamtheit integriert
und zusammen mit dem (den) Chromosom(en), in die er integriert worden
ist, repliziert wird.
-
Der
Vektor ist vorzugsweise ein Expressionsvektor, in dem die DNA-Sequenz,
die die erfindungsgemäße CBD codiert,
funktionsfähig
mit zusätzlichen
Segmenten verknüpft
ist, die zur Transkription der DNA erforderlich sind. Im Allgemeinen
stammt der Expressionsvektor von Plasmid- oder Virus-DNA ab oder
kann Elemente von beiden enthalten. Der Begriff „funktionsfähig verknüpft" gibt an, dass die
Segmente so angeordnet sind, dass sie für ihre beabsichtigten Zwecke
konzertiert funktionieren, z. B. startet die Transkription in einem Promotor
und verläuft über die
DNA-Sequenz, die die CBD codiert.
-
Der
Promotor kann jede beliebige DNA-Sequenz sein, die transkriptionale
Aktivität
in der Wirtszelle der Wahl zeigt, und kann von Genen stammen, die
Proteine codieren, die für
die Wirtszelle entweder homolog oder heterolog sind.
-
Beispiele
für geeignete
Promotoren zur Verwendung in bakteriellen Wirtszellen umfassen den
Promotor des Bacillus-stearothermophilus-maltogenen Amylase-Gens,
des Bacillus-licheniformis-Alpha-Amylase-Gens,
des Bacillus-amyloliquefaciens-Alpha-Amylase-G-Gens, des Bacillus-subtilis-alkalinischen
Protease-Gens oder des Bacillus pumilus-Xylosidasegens, oder den
Phagen Lambda PR- oder PL-Promotoren
oder die E. coli-Promotoren lac, trp oder tac.
-
Alternativ
ist die Konstruktion von Integrationsvektoren so möglich, dass
die DNA, die die CBD codiert, nur funktionell exprimiert wird, wenn
sie ordnungsgemäß in das
Wirtsgenom integriert ist, z. B. stromabwärts von einem residenten Promotor.
-
Die
DNA-Sequenz, die die erfindungsgemäße CBD codiert, kann, sofern
notwendig, auch mit einem geeigneten Terminator funktionsfähig verknüpft sein.
-
Der
erfindungsgemäße rekombinante
Vektor kann weiterhin eine DNA-Sequenz umfassen, die es dem Vektor
ermöglicht,
in der in Frage kommenden Wirtszelle zu replizieren.
-
Der
Vektor kann auch einen selektierbaren Marker, z. B. ein Gen, dessen
Produkt einen Defekt in der Wirtszelle komplementiert, oder ein
Gen, das Resistenz gegenüber
z. B. Antibiotika, wie Kanamycin, Chloramphenicol, Erythromycin,
Tetracyclin, Spectinomycin oder dergleichen oder Resistenz gegenüber Schwermetallen
oder Herbiziden codiert, einschließen.
-
Um
eine erfindungsgemäße CBD auf
den sekretorischen Weg der Wirtszellen zu lenken, kann eine sekretorische
Signalsequenz (auch als Leader-Sequenz, Prepro-Sequenz oder Pre-Sequenz bekannt)
in dem rekombinanten Vektor bereitgestellt sein. Die sekretorische
Signalsequenz ist mit der DNA-Sequenz verknüpft, die die CBD in dem korrekten
Leseraster codiert. Sekretorische Signalsequenzen sind allgemein
5' zu der DNA-Sequenz,
die die CBD codiert, positioniert. Die sekretorische Signalsequenz
kann diejenige sein, die normalerweise mit der CBD assoziiert ist,
oder kann aus einem Gen stammen, das ein anderes sezerniertes Protein
codiert.
-
Die
Verfahrensweisen, die zur Ligation der DNA-Sequenzen, die die vorliegende
CBD codieren, verwendet werden, der Promotor und gegebenenfalls
der Terminator und/oder die sekretorische Signalsequenz oder die
Verfahrensweisen, die zum Zusammenfügen dieser Sequenzen durch
geeignete PCR-Amplifikationschemata, und zu ihrer Insertion in geeignete
Vektoren, die die Information enthalten, die zur Replikation oder Integration
notwendig sind, verwendet werden, sind den Fachleuten gut bekannt.
(cf., z. B., Sambrook et al., op. cit.).
-
Das
Green Fluorescent Protein (GFP) ist zu einem breit eingesetzten
Reportermolekül
zur Aufzeichnung der Genexpression, für Tracer von Zellabstammungen
und als Fusions-Marker für
Proteine geworden (Crameri et al. (1996); Cubitt et al. (1995);
International Patent Application PCT/DK96/00051).
-
GFP
könnte
an CBDs fusioniert sein, um ein Fusionsprotein mit der Cellulose-Bindeeigenschaft
sowie den Fluoreszenz-Eigenschaften zu erzeugen. Die Expression
dieses Fusionsproteins könnte
zur Aufzeichnung der Expression von CBDs in Bacillus-Spezies verwendet
werden und könnte
dadurch zur Optimierung der Expressionsniveaus von gegebenen CBDs
verwendet werden.
-
BEISPIELE
-
MATERIALIEN UND METHODEN
-
Stämme:
-
- Bacillus agaradherens NCIMB Nr. 40482 umfasst das Endoglucanase-Enzym,
das die DNA-Sequenz
von Beispiel 8 codiert.
- E. coli: SJ2 (Diderichsen, B. et al. (1990)) Elektrokompetente
Zellen, die unter Verwendung eines Bio-Rad-GenePulserTM hergestellt
und transformiert wurden, wie von dem Hersteller empfohlen.
- B. subtilis PL2306. Dieser Stamm ist der B. subtilis DN1885
(Diderichsen, B. et al. (1990)), gespalten in die transkriptionale
Einheit des bekannten Bacillus-subtilis-Cellulase-Gens, was zu Cellulase-negativen
Zellen führt.
Weiterhin wurde der Stamm in die aprE- und nprE-Gene gespalten (aprE:
Stahl and Ferrari (1984)) und (nprE: Yang et al (1984)). Die Aufspaltungen
wurden im Wesentlichen wie in (Hrsg. Sonenshein et al. (1993), p.
618) beschrieben durchgeführt.
- B. subtilis PL2304. Dieser Stamm ist der B. subtilis Stamm DN1885
(Diderichsen, B., op. Cit.), gespalten in die transkriptionale Einheit
des Bacillus subtilis Cellulase-Gens, was zu Cellulase-negativen
Zellen führt.
Die Aufspaltung wurde im Wesentlichen wie in (Hrsg. A. L. Sonenshein,
op cit.) beschrieben durchgeführt.
B. subtilis ToC46 (Diderichsen, B. et. al., op. cit.).
-
Plasmide:
-
- pMB100, welches ein Derivativ von pDN1528 ist (S. Jorgensen
et al. (1991). Das Plasmid ist im Wesentlichen das gleiche wie pDN1528,
allerdings wurde eine SacI-Schnittstelle zu Klonierungszwecken zwischen
das Stop-Codon des amyL-Gens und seinem Terminator eingebaut.
- pDN1981 (P. L. Jorgensen et al. (1990))
-
Lösungen/Medien
-
- TY- und LB-Agar (wie beschrieben bei Ausubel, F. M. et al.,
1995).
- SB: 32 g Trypton, 20 g Hefeextrakt, 5 g NaCl und 5 ml 1 N NaOH
werden in sterilem Wasser bis auf Endvolumen von 1 Liter vermischt.
Die Lösung
wird durch Autoklavieren für
20 min bei 121°C
sterilisiert.
- 10% Avicel: 100 g Avicel (FLUKA, Schweiz) werden mit sterilem
Wasser bis zu einem Endvolumen von 1 Liter vermischt, und das 10%
Avicel wird durch Autoklavieren für 20 min bei 121°C sterilisiert.
- Stammlösung
von Kongorot (SIGMA, USA). 1% in ionisiertem Wasser.
- Puffer: 0,1 M Kaliumphosphat, pH 7,5.
-
Allgemeine molekularbiologische Verfahren:
-
- DNA-Manipulationen und -Transformationen wurden unter Verwendung
der molekularbiologischen Standardverfahren durchgeführt (Sambrook
et al. (1989) Molecular cloning: A laboratory manual, Cold Spring
Harbor lab., Cold Spring Harbor, NV; Ausubel, F. M. et al., 1995;
Harwood und Cutting, 1990).
-
Enzyme
für DNA-Manipulationen
wurden nach den Beschreibungen der Hersteller eingesetzt.
-
Primern
eingesetzt wurden, um die DNA-Fragmente zu erhalten, die die CBDs
aus den drei unterschiedlichen Bakterien codieren.
-
Gleichzeitig
wurden PCR-Primer konstruiert, um Extra-Codons hinzuzufügen, entsprechend
den Aminosäuren,
die der Signal-Sequenz von amyL vorangehen, welche verwendet wird,
um die CBDs zum Äußeren der
Bacillus-subtilis-Zelle zu lenken.
-
In-vitro-Amplifikation
der CBD von Cellulomonas-fimi(ATCC 484)-Cellulase CenA Ungefähr 100 bis 200
ng genomische DNA wurde in PCR-Puffer (10 mM Tris-HCl, pH 8,3, 50
mM KCl, 1,5 mM MgCl
2, 0,01% (w/v) Gelatine),
enthaltend 200 μM
von jedem dNTP, 1,5% DMSO (SIGMA, USA), 2,5 Einheiten von AmpliTaq-Polymerase
(Perkin-Elmer, Cetus, USA) und 100 pmol von jedem Primer: CELFIMO1U,
CELFIMO1D,
PCR-amplifiziert.
-
Die
Restriktionsstellen PstI und SacI sind unterstrichen.
-
Die
PCR-Reaktionen wurden unter Verwendung eines DNA-Thermal-Zyklers
(Landgraf, Deutschland) durchgeführt.
Eine Inkubation bei 94°C
für 5 min
und anschließende
Durchführung
von dreißig
PCR-Zyklen unter Verwendung eines Zyklusprofils mit Denaturierung
bei 94°C
für 1 min,
Annelieren bei 65°C
für 1 min,
und Extension bei 72°C
für 1 min.
Zehn-μl-Aliquote
des Amplifikationsprodukts wurden durch Elektrophorese in 1,5%-Agarose-Gelen
(NuSieve, FMC) mit ReadyLoad 100 bp DNA-Leiter (GibcoBRL, Denmark)
als ein Größenmarker
analysiert.
-
In-vitro-Amplifikation der CBD von Pseudomonas
fluorescens (NCIMB10462) CelB
-
Ungefähr 100 bis
200 ng genomische DNA wurde in HiFidelity
TM-PCR-Puffer
(Boehringer Mannheim, Deutschland), angereichert mit 200 μM von jedem
dNTP, 2,6 Einheiten von HiFidelity
TM Expand
Enzymgemisch und 300 pmol von jedem Primer: PSUPPER,
PSLOWER,
PCR-amplifiziert.
-
Die
Restriktionsstellen PstI und SacI sind unterstrichen.
-
Die
PCR-Reaktionen wurden unter Verwendung eines DNA-Thermal-Zyklers
(Landgraf, Deutschland) durchgeführt.
Eine Inkubation bei 94°C
für 2 min,
30 sec bei 60°C
und 45 sec bei 72°C
und anschließende Durchführung von
zehn PCR-Zyklen, unter Verwendung eines Zyklusprofils mit Denaturierung
bei 94°C
für 30 sec,
Annelieren bei 60°C
für 30
sec, und Extension bei 72°C
für 45
sec und zwanzig PCR-Zyklen mit Denaturierung bei 94°C für 30 sec,
60°C für 30 sec
und 72°C
für 45
sec (bei diesem Elongationsschritt wurden 20 sec zu jedem Zyklus
hinzugefügt).
Zehn-μl-Aliquote
des Amplifikationsprodukts wurden durch Elektrophorese in 1,5%-Agarose-Gelen
(NuSieve, FMC) mit ReadyLoad 100 bp DNA-Leiter (GibcoBRL, Denmark)
als ein Größenmarker
analysiert.
-
In-vitro-Amplifikation des CBD-Dimers
von Clostridium stercorarium (NCIMB 11754) XynA
-
Ungefähr 100 bis
200 ng genomische DNA wurde in HiFidelity
TM-PCR-Puffer
(Boehringer Mannheim, Deutschland), angereichert mit 200 μM von jedem
dNTP, 2,6 units von HiFidelity
TM Expand
Enzymgemisch und 300 pmol von jedem Primer: CLOST03U,
CLOST03D2,
PCR-amplifiziert.
-
Die
Restriktionsstellen PstI und SacI sind unterstrichen.
-
Die
PCR-Reaktionen wurden unter Verwendung eines DNA-Thermal-Zyklers
(Landgraf, Deutschland) durchgeführt.
Eine Inkubation bei 94°C
für 2 min,
30 sec bei 60°C
und 45 sec bei 72°C
und anschließende Durchführung von
zehn PCR-Zyklen unter Verwendung eines Zyklusprofils mit Denaturierung
bei 94°C
für 30 sec,
Annelieren bei 60°C
für 30
sec und Extension bei 72°C
für 45
sec und zwanzig PCR-Zyklen mit Denaturierung bei 94°C für 30 sec,
bei 60°C
für 30
sec und bei 72°C
für 45
sec (bei diesem Elongationsschritt wurden 20 sec zu jedem Zyklus
hinzugefügt).
Zehn-μl-Aliquote
des Amplifikationsprodukts wurden durch Elektrophorese in 1,5%-Agarose-Gelen
(NuSieve, FMC) mit ReadyLoad 100 bp DNA-Leiter (GibcoBRL, Denmark) als ein Größenmarker
analysiert.
-
Klonieren durch Polymerase-Kettenreaktion
(PCR):
-
Subklonieren von PCR-Fragmenten
-
Vierzig-μl-Aliquote
der PCR-Produkte, die wie vorstehend beschrieben erzeugt wurden,
wurden unter Verwendung eines QIAquick-PCR-Purification-Kit (Qiagen,
USA) gemäß den Anweisungen
des Herstellers aufgereinigt. Die aufgereinigte DNA wurde in 50 μl 10 mM Tris-HCl,
pH 8,5 eluiert, Fünfundzwanzig μl des aufgereinigten
PCR-Fragments wurden mit SacI und PstI verdaut, in 1,5% Agarose
mit niedriger Geliertemperatur (SeaPlaque GTG, FMC)-Gelen elektrophoresiert,
die relevanten Fragmente wurden aus den Gelen ausgeschnitten und
unter Verwendung des QIAquick Gel Extraction Kit (Qiagen, USA) nach
den Anweisungen des Herstellers gereinigt. Das isolierte DNA-Fragment
wurde anschließend
an PSTI-, SacI-verdautes pMB100 ligiert, und das Ligationsgemisch
wurde zur Transformation von B. subtilisi PL2306 verwendet.
-
Identifizierung und Charakterisierung
von positiven Klonen
-
Die
Zellen wurden auf LB-Agar-Platten ausgestrichen, die Chloramphenicol
(6 μg/ml),
0,4% Glucose und 10 mM Kaliumhydrogenphosphat enthielten, und bei
37°C über Nacht
inkubiert. Die Kolonien wurden am nächsten Tag wieder auf frische
LBPG-Chloramphenicol-Agar-Platten
ausgestrichen und über
Nacht bei 37°C inkubiert.
Die Einzelkolonien am nächsten
Tag von jedem Klon wurden in flüssiges
LB-Medium, das Chloramphenicol (6 μg/ml) enthielt, übergeführt und über Nacht
bei 37°C
unter Schütteln
bei 250 U/min inkubiert.
-
Die
Plasmide wurden aus den flüssigen
Kulturen unter Verwendung des QIAgen Plasmid Purification Mini Kit
(Qiagen, USA) gemäß den Anweisungen
des Herstellers extrahiert, allerdings wurde der Resuspensionspuffer
mit 1 mg/ml Hühnereiweiß-Lysozym
(SIGMA, USA) vor der Lyse der Zellen bei 37°C für 15 min angereichert. Fünf-μl-Proben
der Plasmide wurden mit PstI und SacI verdaut. Die Restriktionsverdaus
wurden durch Gel-Elektrophorese
auf einem 1,5%-Agarose-Gel (NuSieve, FMC) überprüft. Das Auftreten eines DNA-Fragments
der gleichen Größe wie aus
der PCR-Amplifikation gesehen, gab einen positiven Klon an. Drei Klone
wurden selektiert, die jeweils eine CBD aus den drei verschiedenen
vorstehend erwähnten
Bakterien darstellten: MB 144 (exprimiert C. fimi CenA-CBD) und MB203 (exprimiert
C. stercorarium XynA-Dimer-CBD) und MB207 (exprimiert P. fluorescens
ssp cellulosa CelB-CBD).
-
Nukleotid-Sequenzieren des
klonierten DNA-Fragments
-
Qiagen-aufgereinigte
Plasmid-DNA wurde mit dem Taq Deoxy Terminal Cycle Sequencing Kit
(Perkin Elmer, USA) unter Verwendung der gleichen Primer wie vorstehend
verwendet und unter Verwendung eines Applied Biosystems 373A Automatik-Sequenzierers
nach den Anweisungen des Herstellers sequenziert. Die Analyse der
Sequenzdaten wird nach bekannten Verfahren durchgeführt.
-
Expression, Sekretion und Funktionsanalyse
der klonierten CBDs
-
Die
Klone MB 144 (exprimiert C. fimi CenA-CBD), MB203 (exprimiert C.
stercorarium XynA-Dimer-CBD) und MB207 (exprimiert P. fluorescens
ssp cellulosa CelB-CBD), wurden alle 20 h in SB-Medium bei 37°C und 250
U/min inkubiert. 1 ml zellfreier Überstand wurde mit 200 μl 10% Avicel
vermischt. Das Gemisch wurde 1 h zur Inkubation bei 0°C stehengelassen.
Nach diesem Binden von CBD an Avicel wurde das Avicel mit CBD 5
min bei 5000 g zentrifugiert. Das Pellet wurde in 100 μl SDS-Page-Puffer
resuspendiert, 5 min bei 95°C
gekocht, bei 5000 g 5 min zentrifugiert, und 25 μl wurden auf 18% Laemmli Tris-Glycin, SDS-PAGE
NOVEX Gel (Novex, USA) geladen. Die Proben wurden in einer XcellTM-Mini-Zelle (NOVEX, USA) wie vom Hersteller
empfohlen elektrophoresiert, und sämtliche anschließenden Behandlungen
der Gele, einschließlich
Färben
mit Comassie, Entfärben
und Trocknen wurden wie vom Hersteller beschrieben durchgeführt.
-
Das
Auftreten von Proteinbanden der erwarteten Größen (MB144-Proteinbande ca.
12 kDa), (MB203-Proteinbande ca. 35 kDa) und (MB207-Proteinbande
ca. 12 kDa) gab die Expression in B. subtilis von funktionellen
CBDs an.
-
BEISPIEL 4
-
Expression und Reinigung von CBD-Dimer,
das aus C. stercorarium kloniert wurde.
-
Plasmid,
das aus MB203 isoliert wurde, wurde zur Transformation eines weiteren
Bacillus subtilis ToC46 verwendet, und somit ein neuer CBD-Dimer-exprimierender
Klon MB206 erhalten. Unter Verwendung dieses Stamms als Expressionswirt
für das
CBD-Dimer wurde der Klon in Schüttelkolben
inkubiert, die SB-Medium (6 μg/ml
Chloramphenicol) 20 h bei 37°C
und Schütteln
bei 250 U/min inkubiert.
-
1400
ml Kulturfluid-Überstand
wurden auf einem Eisbad gekühlt.
Er wurde über
ein Whatman Glasfilter F filtriert, und anschließend über ein 0,45 Micron Millipore
Typ HVLP steril filtriert.
-
50
Gramm Avicel wurden in 0,1 M Natriumphosphat-Puffer, pH 7,5, bei
Raumtemperatur 30 min suspendiert. Der Überstand wurde entfernt, und
die Avicel-Aufschlämmung
wurde auf 4°C
abgekühlt.
Der klare Überstand
wurde mit der Avicel-Aufschlämmung
30 min bei 4°C
vermischt.
-
Das
Avicel wurde 10 min absetzen gelassen und der Überstand entfernt. Der Avicel-Proteinkomplex wurde
in eine Säule
gepackt und mit 0,1 M Natriumphosphat-Puffer und anschließend mit
Puffer, der 0,5 M Natriumchlorid einschloss, gewaschen. Schließlich wurde
die CBD durch deionisiertes Wasser eluiert.
-
Ingesamt
wurden 78 ml, die CBD enthielten, eluiert. Die CBD wurde nach Zugabe
von festem Natriumchlorid auf einer Amiconzelle mit einer R81P-Membran
mit einer Ausschlussgrenze von 8 kD bis auf eine Endkonzentration
von 0,5 M konzentriert.
-
Die
konzentrierte CBD-Lösung
(30 ml) besaß eine
Extinktion bei 280 nm von 1,2. Der molare Extinktionskoeffizient
von MB 206 betrug 42000, entsprechend einer Proteinkonzentration
von 0,82, was zu insgesamt 25 mg hoch aufgereinigter, Doppel-CBD
führte.
Auf der Grundlage von SDS-PAGE besaß das Ausgangsmaterial etwa
0,1 mg pro ml 29 kD. Das aufgereinigte Endprodukt zeigte nur eine
einzige Bande auf SDS-PAGE.
-
BEISPIEL 5
-
Charakterisierung einer dimerisierten
Pilz-CBD, kloniert und exprimiert in Bacillus subtilis
-
Ein
CBD-Dimer von Pilzursprung wird durch Fusionieren der CBD, die von
der DNA-Sequenz
von Humicola insolens EGII codiert wird, mit der CBD, die von der
DNA-Sequenz von 43 kDa aus Humicola insolens codiert wird, konstruiert.
-
Die
DNA-Sequenz, die Humicola insolens EGII CBD codiert, und Linker
werden aus dem Plasmid, das die cDNA von EG II, die auch als CMC
3 (Dalbøge
and Heldt Hansen, 1994), unter Verwendung von Primern, die für die CBD-Region
spezifisch sind, PCR-amplifiziert, und weiterhin wird der Antisense-Primer
so konstruiert, dass dem PCR-Fragment ein Überhang verliehen wird, der
identisch zu dem DNA-Fragment ist, der die vorhergehende CBD codiert,
die CBD, die von dem Gen der 43 kDa Endoglucanase aus H. insolens
codiert wird, die ausführlich
in
EP-B-0 531 372 und
US 5,457,046 beschrieben
ist. Die DNA, die diese CBD codiert, wird aus genomischer DNA der
in
EP-B-0 531 372 beschriebenen
Humicola insulens PCR-amplifiziert.
-
Die
beiden Fragmente werden durch SOE-PCR (Higuchi et al. (1988)) unter
Verwendung der Primer: Nr.
22857
und des Primers Nr.
20622
kombiniert.
-
Dieses
PCR-Fragment wird, als ein PstI-SacI-Fragment, an pMB100 ligiert,
und das Ligationsgemisch wird zur Transformation von Bacillus subtilis
PL2306 verwendet.
-
Die
klonierte DNA, die im Wesentlichen CBD-Dimer codiert, kann in der
Sequenz CBD-EGII-CZ (327bp):
und in
der entsprechenden Aminosäure-Sequenz
(108 aa Reste):
gefunden werden.
-
Expression,
Sekretion und Funktionalität
der CBD sind wie vorstehend beschrieben gekennzeichnet.
-
BEISPIEL 6
-
Konstruktion der GFP-CBD-Fusion zur CBD-Expressionsoptimierung
-
Ungefähr 100 ng
Plasmid-DNA pMB144, das Plasmid wird wie vorstehend beschrieben
isoliert, werden in PCR-Puffer (10 mM Tris-HCl, pH 8,3, 50 mM KCl,
1,5 mM MgCl
2, 0,01% (w/v) Gelatine), enthaltend
200 μM von
jedem dNTP, 1,5% DMSO (SIGMA, USA), 2,5 Einheiten AmpliTaq-Polymerase
(Perkin-Elmer, Cetus, USA) und 100 pmol von jedem Primer: C-Fusion1:
C-Fusion2:
amplifiziert.
-
Die
Restriktionsstellen EcoRI und HindIII sind unterstrichen.
-
Die
PCR-Reaktionen werden unter Verwendung eines DNA-Thermal-Zyklers
(Landgraf, Deutschland) durchgeführt.
Eine Inkubation bei 94°C
für 5 min
und anschließende
Durchführung
von dreißig
PCR-Zyklen unter Verwendung eines Zyklusprofils mit Denaturatierung
bei 94°C
für 1 min,
Annelieren bei 60°C
für 1 min,
und Extension bei 72°C
für 1 min.
Zehn-μl-Aliquote
des Amplifikationsprodukts werden durch Elektrophorese in 0,7% Agarose-Gelen
(NuSieve, FMC) mit ReadyLoad 100 bp DNA-Leiter (GibcoBRL, Dänemark)
als Größenmarker
analysiert.
-
Das
Fragment wird aufgereinigt, mit EcoRI und HindIII verdaut, Gel-aufgereinigt
und mit dem Vektor pBR322 (Bolivar et al. (1977), Gene, 2, 95–113.) ligiert.
-
Das
Ligationsgemisch wird zur Transformation von SJ2-elektrokompetenter
E. coli. verwendet.
-
Identifizierung und Charakterisierung
von positiven Klonen
-
Die
transformierten Zellen werden auf LB-Agar-Platten ausgestrichen,
die Ampicillin (200 μg/ml)
enthalten und bei 37°C über Nacht
inkubiert. Am nächsten
Tag werden die Kolonien durch Wiederausstreichen von diesen auf
frische LB-Ampicillin-Agar-Platten gewonnen und bei 37°C über Nacht
inkubiert. Die Einzelkolonien am zweiten Tag von jedem Klon werden
in flüssiges
LB-Medium übergeführt, das
Ampicillin (200 μg/ml) enthält, und über Nacht
bei 37°C
unter Schütteln
bei 250 U/min inkubiert.
-
Die
Plasmide werden aus den Flüssigkulturen
unter Verwendung von QIAgen Plasmid Purification Mini Kit (Qiagen,
USA) gemäß den Anweisungen
des Herstellers extrahiert. Fünf-μl-Proben
werden mit HindIII und EcoRI verdaut. Die Restriktionsverdaus werden
durch Gel-Elektrophorese auf einem 0,7%-Agarose-Gel (NuSieve, FMC) überprüft.
-
Ein
Derivat von GFP wird aus der DNA-Konstruktion der Mutante F64L-S65T-GFP
kloniert, welche wie in der internationalen Patentanmeldung PCT/DK96/00051
konstruiert wurde.
-
Das
DNA-Fragment, das die F64L-S65T-GFP codiert, wird als ein BamHI-HindIII-Fragment
im Raster mit der CBD-codierenden DNA, die in pBR322 kloniert wird,
kloniert. Die Ligation, Transformation und Identifizierung eines
positiven Klons erfolgen im Wesentlichen wie vorstehend beschrieben.
-
Diese
Fusionskonstruktion wird als ein EcoRI-BamHI-Fragment aus dem E.
coli-Vektor in den Vektor pUB110 übergeführt (Gryczan et al. (1978)).
Bacillus subtilis PL2306 wird transformiert und positive Klone werden
durch ihre Fähigkeit
zur Fluoreszenz und durch die Existenz eines Avicel-bindenden F64L-S65T-GFP-CBD-Fusionspolypeptids
identifiziert.
-
Die
Wellenlänge
des zur Anregung des F64L-S65T-GFP verwendeten Lichts dieser Studie
beträgt
488 nm, dieses aktiviert das F64L-S65T-GFP zur Emission von Licht
bei 510–530
nm.
-
Die
Fluoreszenz des Überstands
wird durch Fluoreszenz-Spektroskopie gemessen und mit der Fluoreszenz
des Überstandes
nach Inkubation mit Avicel verglichen. Weiterhin kann das Fluoreszenzmolekül mit CBD
durch Binden des Fusionsproteins an Avicel, Entfernen von überschüssigem Überstand
und Überführen des
Avicels in Küvetten
zur Fluoreszenzmessung in einen Fluoreszenz-Spektrophotometer sichtbar
gemacht werden.
-
Durch
Vornehmen serieller Verdünnungen
des Avicel-gebundenen oder nicht gebundenen Fusionsproteins kann
das Expressionsniveau bestimmt werden, wodurch die Identifizierung
eines Bacillus-Klons möglich
gemacht wird, der relativ höhere
Mengen an CBD exprimiert.
-
BEISPIEL 7
-
Screening unter Verwendung CMC-Kongorot
-
Rekombinante
Bacillus-Klone, die CBDs exprimieren, können mittels des Expressionsniveaus
der CBD gescreent werden.
-
Um
Bacillus-Stämme
zu finden, die zur Expression einer gegebenen CBD optimal sind,
werden die Klone von Interesse in einem geeigneten Medium inkubiert,
z. B. wie vorstehend in TV beschrieben, und bei entsprechendem Wachstumsbedingungen
24 h inkubiert. Der Überstand
der Klone wird auf Agarose-CMC-Kongorot-Platten mit ausgestanzten
Löchern übergeführt, der Überstand
mit der CBD wird an die CMC 5 h bei 37°C binden gelassen. Nach dem
Waschen für
15 min mit 2% NaCl-Lösung
kann die CBD-Aktivität
als eine Clearing-Zone gesehen werden.
-
Der
Platten-Assay kann wie nachstehend beschrieben kombiniert werden.
-
Die
Präparation
des Gels zur Verwendung im CBD-Platten-Assay: 0,5% CMC und 0,7%
Agarose (CMC; Carboxymethylcellulose, 7LF von Hercules) (Agarose;
Litex HSA/HSB) werden durch Befeuchten der CMC und Agarose mit 96%
Alkohol hergestellt. 0,1 M Kaliumphosphat-Puffer, pH-7,5, wird zugesetzt
und das Gemisch auf 100°C
bis zur vollständigen
Auflösung
erhitzt. Die Lösung
wird auf 60°C
abkühlen
gelassen. Kongorot-Stammlösung wird
bis zu einer Endkonzentration von 5% zugesetzt, und die Platten
werden mit 15 ml auf eine Petri-Schale mit 9 cm Durchmesser gegossen.
-
Die
Probenapplikationslöcher
werden mit einem Stanzgerät
hergestellt.
-
BEISPIEL 8
-
Identifizierung einer neuen CBD, die eine
neue CBD-Familie definiert
-
Die
alkalische Cellulase, die in Bacillus subtilis, wie nachstehend
beschrieben, kloniert ist, wurde durch Inkubation des Klons für 20 h in
SB-Medium bei 37°C
und 250 U/min exprimiert. Es wurde gezeigt, dass die exprimierte
Cellulase eine CBD enthält,
durch ihre Fähigkeit,
spezifisch an Avicel zu binden.
-
Beim
Stehenlassen zur Inkubation für
weitere 20 h wurde die Cellulase proteolytisch gespalten, und zwei
spezifische Proteinbanden traten auf SDS-Page auf, wobei eine den
katalytischen Teil des ungefähren Molekulargewichts
(MW) von Cellulase von 35 kD und die andere einem vorgeschlagenen
Linker und CBD von ungefähr
MW 8 kD entsprach.
-
Es
wurde gefunden, dass die CBD der terminale Teil der Cellulase ist
und dass die CBD nicht mit einer der zuvor beschriebenen CBD-Familien
(Tomme et al., 1995, p. 142–161) übereinstimmt.
Demgemäß ist diese CBD
das erste Element einer neuen Familie.
-
Klonieren der alkalischen Cellulase aus
Bacillus agaradherens und Expression der alkalischen Endoglucanase in
Bacillus subtilis
-
Die
Präparation
des Gels zur Verwendung im CBD-Platten-Assay: 0,5% CMC und 0,7%
Agarose (CMC; Carboxymethylcellulose, 7LF von Hercules) (Agarose;
Litex HSA/HSB) werden durch Befeuchten der CMC und Agarose mit 96%
Alkohol hergestellt. 0,1 M Kaliumphosphat-Puffer, pH-7,5, wird zugesetzt
und das Gemisch auf 100°C
bis zur vollständigen
Auflösung
erhitzt. Die Lösung
wird auf 60°C
abkühlen
gelassen. Kongorot-Stammlösung wird
bis zu einer Endkonzentration von 5% zugesetzt, und die Platten
werden mit 15 ml auf eine Petri-Schale mit 9 cm Durchmesser gegossen.
-
Die
Probenapplikationslöcher
werden mit einem Stanzgerät
hergestellt.
-
BEISPIEL 8
-
Identifizierung einer neuen CBD, die eine
neue CBD-Familie definiert
-
Die
alkalische Cellulase, die in Bacillus subtilis, wie nachstehend
beschrieben, kloniert ist, wurde durch Inkubation des Klons für 20 h in
SB-Medium bei 37°C
und 250 U/min exprimiert. Es wurde gezeigt, dass die exprimierte
Cellulase eine CBD enthält,
durch ihre Fähigkeit,
spezifisch an Avicel zu binden.
-
Beim
Stehenlassen zur Inkubation für
weitere 20 h wurde die Cellulase proteolytisch gespalten, und zwei
spezifische Proteinbanden traten auf SDS-Page auf, wobei eine den
katalytischen Teil des ungefähren Molekulargewichts
(MW) von Cellulase von 35 kD und die andere einem vorgeschlagenen
Linker und CBD von ungefähr
MW 8 kD entsprach.
-
Es
wurde gefunden, dass die CBD der terminale Teil der Cellulase ist
und dass die CBD nicht mit einer der zuvor beschriebenen CBD-Familien
(Tomme et al., 1995, p. 142–161) übereinstimmt.
Demgemäß ist diese CBD
das erste Element einer neuen Familie.
-
Klonieren der alkalischen Cellulase aus
Bacillus agaradherens und Expression der alkalischen Endoglucanase in
Bacillus subtilis
-
Die
Nucleotid-Sequenz, die die alkalische Cellulase aus Bacillus agaradherens
(Hinterlegungsnr. NCIMB 40482) codiert, wurde durch PCR zum Einbringen
in ein Expressionsplasmid pDN1981 kloniert.
-
Die
PCR wurde im Wesentlichen, wie vorstehend beschrieben, auf 500 ng
genomischer DNA unter Verwendung der folgenden beiden Primer, enthaltend
die NdeI- und KpnI-Restriktionsstellen,
zum Einbringen der Endoglucanase-codierenden DNA-Sequenz in pDN1981
zur Expression durchgeführt: Primer
5: (Nr. 20887)
Primer
6: (Nr. 21318)
-
Nach
PCR-Cycling wurde das PCR-Fragment unter Verwendung eines QIAquick
PCR Column Kit (Qiagen, USA) nach den Anweisungen des Herstellers
aufgereinigt. Die aufgereinigte DNA wurde in 50 μl 10 mM Tris-HCl, pH 8,5, eluiert.
Verdaut mit NdeI und KpnI, aufgereinigt und mit verdautem pDN1981
ligiert. Das Ligationsgemisch wurde zur Transformation von B. subtilis
PL2304 verwendet. Kompetente Zellen wurden hergestellt und wie von
Yasbin et al. (1975) beschrieben transformiert.
-
Isolierung und Test von Bacillus
subtilis-Transformanten
-
Die
transformierten Zellen wurden auf LB-Agar-Platten, die 10 mg/ml
Kanamycin, 0,4% Glucose, 10 mM KH2PO4 und 0,1% AZCL HE-Cellulose (Megazyme, Australien)
enthalten, ausgestrichen und bei 37°C 18 h inkubiert. Endoglucanase-positive
Kolonien wurden als Kolonien identifiziert, die von einer blauen
Halo umgeben waren.
-
Jede
der positiven Transformanten wurde in 10 ml TV-Medium, das 10 mg/ml
Kanamycin enthielt, übergeimpft.
Nach 1 Tag Inkubation bei 37°C,
250 U/min wurden 50 ml Überstand
entnommen. Die Endoglucanase-Aktivität wurde durch Zugabe von 50
ml Überstand
zu den Löchern,
die in dem Agar der LB-Agar-Platten ausgestochen waren, die 0,1%
AZCL HE-Cellulose
enthielten, bestimmt.
-
Nach
16 h Inkubation bei 37°C
gaben blaue Halo, die die Löcher
umgaben, die Expression der Endoglucanase in Bacillus subtilis an.
-
BEISPIEL 9
-
Assay zum Selektieren der CBDs
-
Herstellung von Phosphorsäure-gequollener
Cellulose (PASC):
-
5
g Avicel wird mit Wasser befeuchtet und zu 150 ml eiskalter 85%iger
Phosphorsäure
zugesetzt und auf einem Eisbad 1 h schwach gerührt. Anschließend werden
500 ml kaltes Aceton unter Rühren
zugesetzt. Das gequollene Avicel (PASC) wird über einen Glasfilter-Trichter filtriert
und 3 Mal mit 100 ml eiskaltem Aceton und anschließend 2 Mal
mit 500 ml Wasser gewaschen. Das PASC wird anschließend in
500 ml Wasser suspendiert und bis zur Homogenität unter Verwendung eines Ultra
Thorax Homogenisators gemischt. Das PASC wird kalt gelagert.
-
CBD-Binden an Phosphorsäure-gequollene
Cellulose (PASC)-Selektion der CBDs:
-
400
ml 10 mg/ml PASC (hergestellt wie vorstehend beschrieben und mit
50 mM Natriumphosphat, pH 7, gewaschen) in 50 mM Natriumphosphat,
pH 7, in Eppendorf-Röhrchen wurden
mit 400 ml Cellulose-Bindedomäne
(Ce15A CBD oder MB206 Doppel-CBD),
verdünnt
in 50 mM Natriumphosphat, pH 7, gemischt. Die Konzentration der
CBD wurde variiert, z. B. von 0 mM bis ungefähr 8 mM für die CBD Ce15A. Eine Kontrollreihe
wurde ohne PASC eingeschlossen. Die Proben wurden 1 h bei Raumtemperatur
vor dem Zentrifugieren der Proben für 4 min bei 14000 g inkubiert.
500 μl des Überstandes
wurden in 2 ml Wasser verdünnt.
Die Menge an CBD, die in dem Überstand
vorhanden war (freie CBD) wurde anschließend durch Tryptophan-Fluoreszenz-Spektroskopie
auf einem Perkin-Elmer LS50 Lumineszenz-Spektrometer (Anregung bei
280 nm und Emission bei 340 nm) unter Verwendung der Fluoreszenzintensität der Proben
ohne PASC-Zugabe als Referenz (Standardkurve) gemessen. Die Menge
an gebundener CBD wurde dann wie folgt berechnet: Gesamt CBD (ohne
PASC-Zugabe) – freie
CBD. Somit wurde eine Bindungsisotherme durch Auftragen der Menge
an gebundener CBD pro g PASC als Funktion von freier CBD in Lösung in
mM, wie in 1 und 2 gezeigt, erhalten.
Die Daten konnten unter Verwendung eines einfachen Langmuir-Bindungsmodells
angepasst werden (Bothwell et al., 1995): E(gebunden) = (Amax·E(frei))/(Kd + E(frei)), wobei E(gebunden) die Menge
an gebundener CBD in mmol/g PASC und E(frei) die Menge an freier
CBD in mM ist. Amax ist die maximale Menge an
CBD, die an PASC gebunden werden kann, und Kd ist die Gleichgewichtskonstante
für das
Gleichgewicht E(gebunden) << E(frei). Je geringer
die Kd (Desorptionskonstante) ist, desto
stärker
ist somit die Bindung. Diese Konstanten werden nach Anpassen der
Daten an das Modell unter Verwendung der Algorithmen in GraphPad
Prizm erhalten. Die Desorptionskonstanten, die für Ce15A CBD und MB206 Doppel-CBD
gefunden werden, betragen 0,42 bzw. 0,76 mM (cf. 1 und 2).
-
Die
erfindungsgemäßen CBDs
zeigen Desorptionskonstanten unter 1 mM, stärker bevorzugt unter 0,1 mM
und am stärksten
bevorzugt unter 10 mM.
-
Literaturliste
-
- Bothwell, M. K. und Walker, L. P. (1995) Bioresource Technology
53: 21–29
- Kraulis P., G. M. Clore, M. Nilges, T. A. Jones, G. Pettersson,
J. Knowles and A. M. Gronenborn, 1989, "Determination of the three-dimensional
structure of the C terminal domain of cellobiohydrolase I from Trichoderma reesei.
A study using nuclear magnetic resonance and hybrid distance geometry-dynamical
simulated annealing" in
Biochemistry 28: 7241–7257.
- Ong E., N. R. Gilkes, R. C. Miller und D. G. Kilburn, 1993, "The cellulose-binding
domain (cbdcex) of an exoglucanase from Cellulomonas fimi – production
in Escherichia coli and characterization of the polypeptide" in Biotechnology
and Bioengineering. 42: 401–409.
- Linder M., I. Salovuori, L. Ruohonen und T. T. Teeri, 1996, "Characterization
of a double cellulose-binding domain – synergistic high-affinity
binding to crystalline cellulose" in
Journal of Biological Chemistry 271: 21268–21272.
- Peter Tomme et al. "Cellulose-Binding
Domains: Classification and Properties" in "Enzymatic
Degradation of Insoluble Carbohydrates", John N. Saddler and Michael H. Penner
(Eds.), ACS Symposium Series, No. 618, 1996.
- Hockney, R. C. (1994) TIBTECH, Bd. 12, p. 456–463.
- Emmanuel Brun et al. (1995).
- Eur. J. Biochem 231, 142–148.
- N. R. Gilkes et al. (1991) Eur. J. Biochem, 202: 367–377.
- Bertus van den Berg et al., (1993), Introduction of disulfide
bonds into Bacillus subtilis neutral protease. Protein Engineering,
Bd. 6 Nr. 5, S. 521–527.
- Hrsg. A. L. Sonenshein, J. A. Hoch und Richard Losick (1993)
Bacillus subtilis and other Gram-Positive Bacteria, American Society
for microbiology, S. 939 und S. 618.
- Andreas Crameri et al. (1996) Improved Green Flourescent Protein
by molecular evolution using DNA shuffling, Nature Biotechnology,
Bd. 14, S. 315–319.
- Andrew B. Cubitt et al. (1995) Understanding, improving and
using flourescent proteins, TIBS, Bd. 20. S. 448–455.
- Diderichsen, B., Wedsted, U., Hedegaard, L., Jensen, B. R.,
Sjøholm,
C. (1990) Cloning of aldB, which encodes alpha-acetolactate decarboxylase, an exoenzyme
from Bacillus brevis. J. Bacteriol., 172, 4315–4321.
- Stahl, M. L. und E. Ferrari (1984) Replacement of the Bacillus
subtilis subtilisin structural gene with an In vitro-derived deletion
mutation. J. Bacteriol. 158: 411–418.
- Yang, M. Y. et al (1984) Cloning ot the neutral protease gene
of Bacillus subtilis and the use of the cloned gene to create an
in vitro-derived deletion mutation. J. Bacteriol. 160: 16–21.
- S. Jørgensen
et al. (1991) Journal of Bacteriology, Bd. 173, No., S. 559–567.
- P. L. Jørgensen,
C. K. Hansen, G. B. Poulsen und B. Diderichsen (1990) In vivo genetic
engineering: homologues recombination as a tool for plasmid construction,
Gene, 96 S. 37–41.
- Ausubel, F. M. et al. (Hrsg.) "Current protocols in Molecular Biology". John Wiley and
Sons, 1995
- Sambrook et al. (1989) Molecular cloning: A laboratory manual,
cold Spring Harbor lab., Cold Spring Harbor, NY.
- Harwood, C. R., und Cutting, S. M. (eds.) "Molecular Biological Methods for Bacillus". John Wiley and
Sons, 1990.
- Pitcher, D. G., Saunders, N. A., Owen, R. J. (1989). Rapid extraction
of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl.
Microbiol., 8, 151–156).
- Dalbøge,
H. und H. P. Heldt Hansen. 1994. A novel method for efficient expression
cloning of fungal enzyme genes, Molecular & General Genetics. 243: 253–260.
- Higuchi et al. (1988) NAR 16: 7351–7367.
- Gryczan, T. J. et al. (1978) Journal of Bacteriology, 134, p.
318–329.
- Tomme P., R. A. Warren, R. C. Miller, Jr., D. G. Kilburn und
N. R. Gilkes. 1995. Cellulose-Binding Domains: Classification and
Properties. In: Saddler J. N. and M. H. Penner, eds. Enzymatic Degradation
of Insoluble Carbohydrates. Washington, DC, American Chemical Society.
page 142–161.
- Yasbin, R. E., Wilson, G. A. und Young, F. E. (1975) Transformation
and transfection in lysogenic strains of Bacillus subtilis: evidence
for selective induction of prophage in competent cells. J. Bacteriol,
121: 296–304.
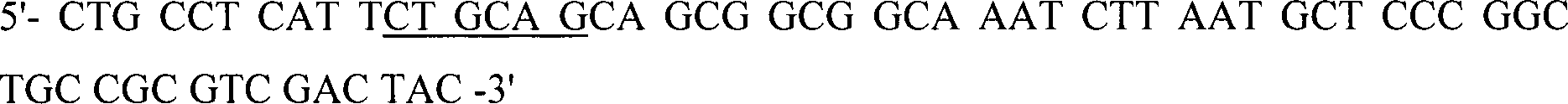 PCR-amplifiziert.
PCR-amplifiziert.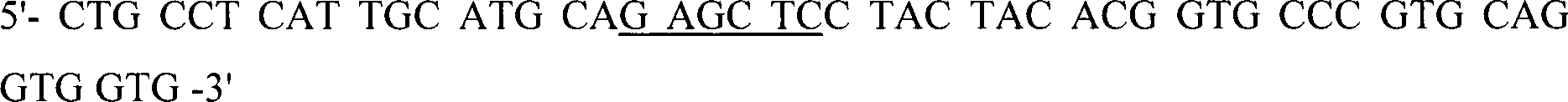
 PCR-amplifiziert.
PCR-amplifiziert.
 PCR-amplifiziert.
PCR-amplifiziert.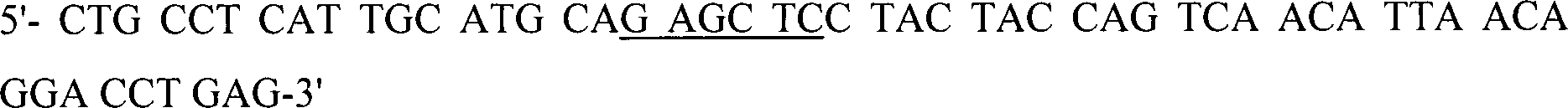
 kombiniert.
kombiniert.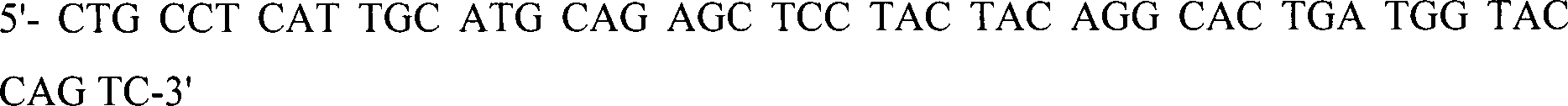
 gefunden werden.
gefunden werden.
 amplifiziert.
amplifiziert.