ES2912400T3 - Modificación génica orientada mediante la reparación génica mediada por oligonucleótidos - Google Patents
Modificación génica orientada mediante la reparación génica mediada por oligonucleótidos Download PDFInfo
- Publication number
- ES2912400T3 ES2912400T3 ES14763072T ES14763072T ES2912400T3 ES 2912400 T3 ES2912400 T3 ES 2912400T3 ES 14763072 T ES14763072 T ES 14763072T ES 14763072 T ES14763072 T ES 14763072T ES 2912400 T3 ES2912400 T3 ES 2912400T3
- Authority
- ES
- Spain
- Prior art keywords
- nicases
- gron
- site
- plant
- dna
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8213—Targeted insertion of genes into the plant genome by homologous recombination
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Organic Chemistry (AREA)
- Chemical & Material Sciences (AREA)
- General Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Physics & Mathematics (AREA)
- Cell Biology (AREA)
- Plant Pathology (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Physiology (AREA)
- Botany (AREA)
- Developmental Biology & Embryology (AREA)
- Environmental Sciences (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Saccharide Compounds (AREA)
Abstract
Un procedimiento para introducir una mutación mediada por una oligonucleobase de reparación de genes (GRON) en una secuencia de ácido desoxirribonucleico (ADN) objetivo en una célula vegetal, que comprende: cultivar la célula bajo condiciones que aumenten uno o más procesos de reparación del ADN celular antes de, y/o coincidiendo con, la entrega de un GRON en la célula vegetal, en el que las condiciones que aumentan uno o más procesos celulares de reparación del ADN comprenden la introducción de una o más nicasas específicas de sitio en la célula vegetal, en el que una o más nicasas específicas del sitio introducen mellas en la cadena simple en la vecindad del sitio que se pretende convertir con el GRON.
Description
DESCRIPCIÓN
Modificación génica orientada mediante la reparación génica mediada por oligonucleótidos
CAMPO DE LA INVENCIÓN
La presente invención se refiere en general a procedimientos novedosos para mejorar la eficacia de la orientación de las modificaciones a lugares específicos en las secuencias genómicas o de otros nucleótidos. Además, esta invención se relaciona con el ADN objetivo que ha sido modificado, mutado o marcado por los enfoques divulgados en el presente documento. La invención también se refiere a células, tejidos y organismos que han sido modificados por los procedimientos de la invención.
ANTECEDENTES DE LA INVENCIÓN
La rotura de la doble cadena del ADN (DSB) potencia la recombinación homóloga en las células vivas y se ha aprovechado para la edición selectiva del genoma mediante el uso de endonucleasas diseñadas. El componente clave de las nucleasas diseñadas es el dominio de reconocimiento del ADN que es capaz de dirigir la nucleasa al sitio objetivo del genoma para una rotura de la doble cadena del ADN genómico. La reparación celular de DSB debido a la unión de extremos no homólogos (NHEJ) da lugar a deleciones/inserciones mutagénicas de un gen objetivo. Alternativamente, la DSB puede estimular la recombinación homóloga entre el locus objetivo endógeno y un fragmento de ADN homólogo introducido de forma exógena con la información genética deseada, un proceso que se denomina orientación de genes.
El procedimiento más prometedor que implica la edición de genes o del genoma son las nucleasas de dedos de zinc (ZFN) diseñadas a medida, un tipo de enzima híbrida que consiste en dominios de unión al ADN de proteínas de dedos de zinc y el dominio de nucleasa Fokl (FN). La tecnología ZFN implica principalmente el uso de proteínas híbridas derivadas de los dominios de unión al ADN de las proteínas de dedos de zinc (ZF) y el dominio de escisión inespecífico de la endonucleasa FokI. Los ZF pueden ensamblarse como módulos diseñados a medida para reconocer secuencias de ADN seleccionadas, tras la unión en el sitio preseleccionado, se produce un DSB por la acción del dominio de escisión de Fokl.
La endonucleasa FokI se aisló por primera vez de la bacteria Flavobacterium okeanokoites. Esta nucleasa de tipo IIS consiste en dos dominios separados, el dominio de unión al ADN N-terminal y el dominio de escisión del a Dn C-terminal. El dominio de unión al ADN funciona para el reconocimiento de una secuencia no palindrómica 5' -GGATG-375' -CATCC-3' mientras que el dominio catalítico escinde el ADN de doble cadena de forma no específica a una distancia fija de 9 y 13 nucleótidos corriente abajo del sitio de reconocimiento. El Fokl existe como un monómero inactivo en solución y se convierte en un atenuador activo tras la unión a su ADN objetivo y en presencia de algunos metales divalentes. Como complejo funcional, dos moléculas de Fokl, cada una de las cuales se une a una molécula de ADN de doble cadena, se dimerizan a través del dominio catalítico del ADN para la escisión efectiva de las dobles cadenas de ADN.
De manera similar, las nucleasas también pueden hacerse utilizando otras proteínas/dominios si son capaces de reconocer específicamente el ADN. Los efectores TAL pertenecen a un gran grupo de proteínas bacterianas que existen en varias cepas de Xanthomonas y que se translocan a las células del huésped mediante un sistema de secreción de tipo III, los llamados efectores de tipo III. Una vez en las células del huésped, se ha descubierto que algunos efectores TAL activan transcripcionalmente sus correspondientes genes objetivo del huésped, ya sea para la virulencia de la cepa (capacidad de causar la enfermedad) o la avirulencia (capacidad de desencadenar respuestas de resistencia del huésped), dependiendo del contexto genético del huésped. Cada efector contiene los motivos funcionales de localización nuclear y un potente dominio de activación de la transcripción que son característicos del activador de la transcripción eucariótico. Y cada efector también contiene una región central repetitiva que consiste en un número variable de unidades de repetición de 34 aminoácidos, y la región repetida como dominio de unión al ADN determina la especificidad biológica de cada efector.
Zhang et al., Plant Physiol. 161: 20-27, 2013, divulga el uso de las TALEN -nucleasas efectoras similares a activadores transcripcionales- que son endonucleasas diseñadas con base en la combinación de un dominio de unión al ADN similar al efector TAL con un dominio catalítico de Fokl. Mediante la ingeniería del dominio de unión al ADN, estas TALEN pueden diseñarse fácilmente para reconocer dominios específicos de unión al ADN. Utilizando protoplastos de tabaco como sistema modelo, se evaluó la actividad de TALEN usando un reportero de polinucleótido de fusión de una sola cadena que comprendía una secuencia codificadora de proteína amarilla fluorescente unida a un sitio de reconocimiento de TALEN. Este sistema reportero se introdujo en protoplastos, y se pudo medir un evento de escisión y reparación mediante la expresión de YFP funcional.
SUMARIO DE LA INVENCIÓN
El objeto de la presente invención es un procedimiento como se define en la reivindicación 1 para introducir una mutación mediada por una oligonucleobase de reparación de genes (GRON) en una secuencia de ácido desoxirribonucleico (ADN) objetivo en una célula vegetal. Las reivindicaciones dependientes se refieren a realizaciones preferidas de las mismas. El procedimiento de acuerdo con la presente invención para introducir una mutación mediada por una oligonucleobase reparadora de genes (GRON) en una secuencia de ácido desoxirribonucleico (ADN) objetivo en una célula vegetal comprende el cultivo de la célula vegetal en condiciones que aumentan uno o más procesos celulares de reparación del ADN antes de, y/o coincidiendo con, la entrega de un GRON en la célula vegetal. Las condiciones que aumentan uno o más procesos celulares de reparación del ADN comprenden la introducción de una o más nicasas específicas de sitio en la célula vegetal, en la que la una o más nicasas específicas de sitio introducen mellas de una sola cadena en la vecindad del sitio orientado que es objeto de conversión por el GRON.
Otras condiciones divulgadas en el presente documento que aumentan uno o más procesos de reparación del ADN celular comprenden: la introducción de uno o más sitios en el GRON o en el ADN de la célula vegetal que son objetivos para la reparación por escisión de bases, la introducción de uno o más sitios en el GRON o en el ADN de la célula vegetal que son objetivos para la unión de extremos no homólogos, la introducción de uno o más sitios en el GRON o en el a Dn de la célula vegetal que sean objetivos para la unión de extremos mediada por microhomología, la introducción de uno o más sitios en el GRON o en el ADN de la célula vegetal que sean objetivos para la recombinación homóloga, y la introducción de uno o más sitios en el GRON o en el ADN de la célula vegetal que sean objetivos para la reparación por empuje.
En ciertas realizaciones, la secuencia de ácido desoxirribonucleico (ADN) objetivo está dentro del genoma de la célula vegetal. La célula vegetal puede ser no transgénica o transgénica, y la secuencia de ADN objetivo puede ser un transgén o un gen endógeno de la célula vegetal.
En ciertas realizaciones, la una o más nicasas se seleccionan entre las nicasas TALEN, las nicasas de dedos de zinc y las nicasas meganucleas. En ciertas realizaciones, la una o más nicasas específicas del sitio están acopladas covalentemente al GRON.
Los procedimientos y composiciones descritos en el presente documento son aplicables a las plantas en general. Sólo a modo de ejemplo, una especie vegetal puede seleccionarse del grupo que consiste en canola, girasol, maíz, tabaco, remolacha azucarera, algodón, maíz, trigo, cebada, arroz, alfafa, cebada, sorgo, tomate, mango, melocotón, manzana, pera, fresa, plátano, melón, patata, zanahoria, lechuga, cebolla, soja, soja spp, caña de azúcar, guisante, garbanzo, guisante de campo, faba, lentejas, nabo, colinabo, coles de Bruselas, altramuz, coliflor, col rizada, habas, álamo, pino, eucalipto, uva, cítricos, triticale, alfalfa, centeno, avena, césped y hierbas forrajeras, lino, colza, mostaza, pepino, gloria de la mañana, bálsamo, pimienta, berenjena, caléndula, loto, col, margarita, clavel, tulipán, iris y lirio. También pueden aplicarse, en todo o en parte, a todos los demás sistemas biológicos, incluidos, entre otros, las bacterias, los hongos y las células de mamíferos, e incluso sus orgánulos (por ejemplo, las mitocondrias y los cloroplastos).
En ciertas realizaciones, los procedimientos comprenden además la regeneración de una planta que tiene una mutación introducida por el GRON a partir de la célula vegetal, y pueden comprender la recolección de semillas de la planta.
En realizaciones relacionadas, el procedimiento de acuerdo con la presente invención comprende además evaluar la eficiencia de conversión del GRON, como se define en la reivindicación 10. Otro tema de la presente invención es el uso, como se define en la reivindicación 8, de una o más nicasas para mejorar la conversión mutacional mediante oligonucleótidos de reparación de genes (GRON) como donantes, que comprende la introducción de la una o más nicasas específicas de sitio en una célula. De acuerdo con las realizaciones, la una o más nicasas se seleccionan entre las nicasas TALEN, las nicasas de dedos de zinc y las nicasas meganucleas.
Otras realizaciones de la invención serán evidentes a partir de la siguiente descripción detallada, las realizaciones ejemplares y las reivindicaciones.
DESCRIPCIÓN DETALLADA DE LA INVENCIÓN
Definiciones
La invención debe entenderse de acuerdo con las siguientes definiciones.
Una oligonucleobase es un polímero de nucleobases, cuyo polímero puede hibridarse por emparejamiento de bases Watson-Crick a un ADN que tenga la secuencia complementaria.
Las nucleobases comprenden una base que es una purina, pirimidina o un derivado o análogo de las mismas. Las nucleobases incluyen las nucleobases peptídicas, las subunidades de los ácidos nucleicos peptídicos y las nucleobases de morfolina, así como los nucleósidos y los nucleótidos. Los nucleósidos son nucleobases que contienen una fracción de pentosefuranosilo, por ejemplo, un ribósido opcionalmente sustituido o un 2'-desoxirribósido. Los
nucleósidos pueden estar unidos por una de varias fracciones de enlace, que pueden contener o no un fósforo. Los nucleósidos unidos por enlaces fosfodiésteres no sustituidos se denominan nucleótidos.
Una cadena de oligonucleobase tiene un único terminal 5' y 3', que son las últimas nucleobases del polímero. Una cadena de oligonucleobase concreta puede contener nucleobases de todo tipo. Un compuesto de oligonucleobase es un compuesto que comprende una o más cadenas de oligonucleobase que son complementarias y se hibridan mediante el emparejamiento de bases Watson-Crick. Las nucleobases son de tipo desoxirribón o de tipo ribo. Las nucleobases de tipo ribo son nucleobases que contienen pentosefuranosilo y en las que el carbono 2' es un metileno sustituido por un hidroxilo, un alquiloxi o un halógeno. Las nucleobases de tipo desoxirribo son nucleobases distintas de las de tipo ribo e incluyen todas las nucleobases que no contienen una fracción de pentosefuranosilo.
Una cadena de oligonucleobase incluye genéricamente tanto cadenas de oligonucleobase como segmentos o regiones de cadenas de oligonucleobase. Una hebra de oligonucleobase tiene un extremo 3' y un extremo 5'. Cuando una hebra de oligonucleobase es coextensiva a una cadena, los extremos 3' y 5' de la cadena son también los extremos 3' y 5' de la cadena.
De acuerdo con la presente invención, los órganos vegetales incluyen, pero no se limitan a, hojas, tallos, raíces, yemas vegetativas, yemas florales, meristemos, embriones, cotiledones, endospermo, sépalos, pétalos, pistilos, carpelos, estambres, anteras, microsporas, polen, tubos polínicos, óvulos, ovarios y frutos, o secciones, rodajas o discos tomados de los mismos. Los tejidos vegetales incluyen, pero no se limitan, los tejidos de callo, los tejidos del suelo, los tejidos vasculares, los tejidos de almacenamiento, los tejidos meristemáticos, los tejidos de las hojas, los tejidos de los brotes, los tejidos de las raíces, los tejidos de las agallas, los tejidos de los tumores vegetales y los tejidos reproductivos. Las células vegetales incluyen, pero no se limitan a, células aisladas con paredes celulares, agregados de diferentes tamaños y protoplastos.
Dos polinucleótidos o polipéptidos son idénticos si la secuencia de nucleótidos o de residuos de aminoácidos, respectivamente, en las dos secuencias es la misma cuando se alinean para obtener la máxima correspondencia como se describe a continuación. Los términos "idéntico" o "porcentaje de identidad", en el contexto de dos o más ácidos nucleicos o secuencias polipeptídicas, se refieren a dos o más secuencias o subsecuencias que son iguales o tienen un porcentaje especificado de residuos de aminoácidos o nucleótidos que son iguales, cuando se comparan y alinean para obtener la máxima correspondencia en una ventana de comparación, según se mide utilizando uno de los siguientes algoritmos de comparación de secuencias o mediante alineación manual e inspección visual. En el caso de los polipéptidos cuyas secuencias difieren en sustituciones conservadoras, el porcentaje de identidad de la secuencia puede ajustarse al alza para corregir la naturaleza conservadora de la sustitución. Los medios para realizar este ajuste son bien conocidos por los expertos en la técnica. Por lo general, esto implica puntuar una sustitución conservadora como una falta de coincidencia parcial en lugar de completa, aumentando así el porcentaje de identidad de la secuencia. Así, por ejemplo, cuando un aminoácido idéntico recibe una puntuación de 1 y una sustitución no conservadora recibe una 'puntuación de cero, una sustitución conservadora recibe una puntuación entre cero y 1. La puntuación de las sustituciones conservadoras se calcula de acuerdo con , por ejemplo, el algoritmo de Meyers & Miller, Computer Applic. Biol. Sci. 4: 11-17 (1988) por ejemplo, tal como se implementa en el programa PC/GENE (Intelligenetics, Mountain View, Calif., EE. UU.).
Las frases "sustancialmente idénticas" y "porcentaje de identidad" en el contexto de dos ácidos nucleicos o polipéptidos, se refieren a secuencias o subsecuencias que tienen al menos 50 %, ventajosamente 60 %, preferiblemente 70 %, más preferiblemente 80 %, y más preferiblemente 90-95 % de identidad de nucleótidos o residuos de aminoácidos cuando se alinean para obtener la máxima correspondencia en una ventana de comparación, según se mide utilizando uno de los siguientes algoritmos de comparación de secuencias o mediante alineación manual e inspección visual. Esta definición también se refiere al complemento de una secuencia de prueba, que tiene una complementariedad sustancial de secuencia o subsecuencia cuando la secuencia de prueba tiene una identidad sustancial con una secuencia de referencia.
Un experto en la técnica reconocerá que dos polipéptidos también pueden ser "sustancialmente idénticos" si los dos polipéptidos son inmunológicamente similares. Así, la estructura general de la proteína puede ser similar mientras que la estructura primaria de los dos polipéptidos muestra una variación significativa. Por lo tanto, un procedimiento para medir si dos polipéptidos son sustancialmente idénticos implica medir la unión de anticuerpos monoclonales o policlonales a cada polipéptido. Dos polipéptidos son sustancialmente idénticos si los anticuerpos específicos para un primer polipéptido se unen a un segundo polipéptido con una afinidad de al menos un tercio de la afinidad para el primer polipéptido. Para la comparación de secuencias, normalmente una secuencia actúa como secuencia de referencia, con la que se comparan las secuencias de prueba. Cuando se utiliza un algoritmo de comparación de secuencias, se introducen las secuencias de prueba y de referencia en un ordenador, se designan las coordenadas de la subsecuencia, si es necesario, y se designan los parámetros del programa del algoritmo de secuencia. A continuación, el algoritmo de comparación de secuencias calcula el porcentaje de identidad de la secuencia o secuencias de prueba en relación con la secuencia de referencia, con base en los parámetros designados del programa.
La alineación óptima de las secuencias para su comparación puede realizarse, por ejemplo, mediante el algoritmo de
homología local de Smith & Waterman, 0.4dv. Appl. Math. 2:482 (I 98 I), mediante el algoritmo de alineación homológica de Needleman & Wunsch, J. Mol. Biol. 48:443 (1970), por el procedimiento de búsqueda de similitudes de Pearson & Lipman, Proc. Nat'I. Acad. Sci. USA 585:2444 (1988), mediante implementaciones informáticas de estos algoritmos (GAP, BESTFIT, FASTA y TFASTA en el paquete de software Wisconsin Genetics, Genetics Computer Group, 575 Science Dr., Madison, Wis.), mediante software para alineamientos como VECTOR NTI Versión #6 de InforMax, Inc. MD, USA, mediante los procedimientos descritos en ClustalW, Thompson, J. D., Higgins, D. G. y Gibson, T. J. (1994) CLUSTALW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position--specific gap penalties and weight matrix choice. Investigación de Ácidos Nucleicos, 22:4673-4680 o por inspección visual (véase en general Protocols in Molecular Biology, F. M. Ausubel et al., eds., Current Protocols, una empresa conjunta de Greene Publishing Associates, Inc. y John Wiley & Sons, Inc. (suplemento de 1995) (Ausubel)).
Ejemplos de algoritmos adecuados para determinar el porcentaje de identidad de secuencia y la similitud de secuencia son los algoritmos BLAST y BLAST 2.0, que se describen en Altschul et al. (1990) J. Mol. Biol. 215: 403-410 y Altschul et al. (1977) Nucleic Acids Res. 25: 33 89-3402respectivamente. El software para realizar análisis BLAST está disponible públicamente a través del National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/). Este algoritmo consiste en identificar primero los pares de secuencias de alta puntuación (HSP) mediante la identificación de palabras cortas de longitud W en la secuencia de consulta, que coinciden o satisfacen alguna puntuación umbral de valor positivo T cuando se alinean con una palabra de la misma longitud en una secuencia de la base de datos. T se denomina umbral de puntuación de palabras de la vecindad (Altschul et al, supra). Estas palabras vecinas iniciales actúan como semillas para iniciar las búsquedas para encontrar los HSP más largos que las contengan. A continuación, los aciertos de las palabras se extienden en ambas direcciones a lo largo de cada secuencia hasta donde pueda aumentar la puntuación de alineación acumulada. Las puntuaciones acumuladas se calculan utilizando, para las secuencias de nucleótidos, los parámetros M (puntuación de recompensa para un par de residuos coincidentes; siempre>0) y N (puntuación de penalización para los residuos no coincidentes; siempre<0). Para las secuencias de aminoácidos, se utiliza una matriz de puntuación para calcular la puntuación acumulada. La extensión de los aciertos de palabras en cada dirección se detiene cuando: la puntuación acumulada de la alineación cae en la cantidad X desde su valor máximo alcanzado; la puntuación acumulada llega a cero o menos, debido a la acumulación de una o más alineaciones de residuos con puntuación negativa; o se alcanza el final de cualquiera de las secuencias. Los parámetros del algoritmo BLAST W, T y X determinan la sensibilidad y la velocidad del alineamiento. El programa BLASTN (para secuencias de nucleótidos) utiliza por defecto una longitud de palabra (W) de 11, una expectativa (E) de 10, M=5, N=-4, y una comparación de ambas cadenas. Para las secuencias de aminoácidos, el programa BLASTP utiliza por defecto una longitud de palabra (W) de 3, una expectativa (E) de 10 y la matriz de puntuación BLOSUM62 (véase Henikoff& Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 (1989)). Además de calcular el porcentaje de identidad de la secuencia, el algoritmo BLAST también realiza un análisis estadístico de la similitud entre dos secuencias (véase, por ejemplo Karlin & Altschul, Proc. Nat'1. Acad. Sci. USA 90:5873-5787 (1993)). Una de las medidas de similitud que ofrece el algoritmo BLAST es la probabilidad de suma mínima (P(N)), que proporciona una indicación de la probabilidad de que se produzca una coincidencia entre dos secuencias de nucleótidos o aminoácidos por azar. Por ejemplo, un ácido nucleico se considera similar a una secuencia de referencia si la menor probabilidad de suma en una comparación del ácido nucleico de prueba con el ácido nucleico de referencia es inferior a aproximadamente 0,1, más preferiblemente inferior a aproximadamente 0,01, y más preferiblemente inferior a aproximadamente 0,001.
Rotura de cadenas
De acuerdo con la presente invención, las condiciones que aumentan uno o más procesos celulares de reparación del ADN comprenden la introducción de una o más nicasas específicas de sitio en la célula vegetal, en la que la una o más nicasas específicas de sitio introducen mellas de una sola cadena en la vecindad del sitio que es objeto de conversión por el GRON. Aunque la siguiente divulgación abarca referencias a compuestos distintos de las nicasas específicas de sitio, cuyos compuestos introducen mellas de cadena simple o roturas de cadena doble en la secuencia de ADN objetivo, esta divulgación no forma parte del objeto de la presente invención.
La inclusión de compuestos que inducen roturas de cadena simple o doble, ya sea en el oligonucleótido o junto con el oligonucleótido, genera una lesión que se repara mediante la unión de extremos no homólogos (NHEJ), la unión de extremos mediada por microhomología (MMEJ) y la recombinación homóloga. A modo de ejemplo, la familia de antibióticos de la bleomicina, las nucleasas de dedo de zinc, la Fokl (o cualquier clase de enzima de restricción de tipo IIS) y otras nucleasas pueden acoplarse covalentemente al extremo 3' o 5' de los oligonucleótidos de reparación, con el fin de introducir roturas de cadena simple o doble en las proximidades del sitio que se pretende convertir con el oligonucleótido de reparación. Los antibióticos de la familia de la bleomicina son glicopéptidos que rompen el ADN, incluyen la bleomicina, la zeocina, la fleomicina, la tallysomicina y la pepleomicina, entre otros.
Los procedimientos divulgados en el presente documento no se limitan a la naturaleza o al tipo de reactivo modificador del ADN que se utiliza. Por ejemplo, estos reactivos modificadores del ADN liberan radicales que provocan la rotura de la cadena de ADN. Alternativamente, los reactivos alquilan el ADN para formar aductos que bloquearían la replicación y la transcripción. En otra alternativa, los reactivos generan enlaces cruzados o moléculas que inhiben las enzimas celulares que conducen a la rotura de la cadena. Los ejemplos de reactivos modificadores del ADN que se han unido a los oligonucleótidos para formar TFO incluyen, pero no se limitan a, los indolocarbazoles, la diimida de
naftalina (NDI), el transplatino, la bleomicina, los análogos del ciclopropirroloindol y las fenanodihidrodioxinas. En particular, los indolocarbazoles son inhibidores de la topoisomerasa I. La inhibición de estas enzimas da lugar a roturas de la cadena y a la formación de aductos proteicos del ADN [Arimondo et al., Bioorganic and Medicinal Chem. 8, 777, 2000]. NDI es un fotooxidante que puede oxidar las guaninas, lo que podría causar mutaciones en sitios de residuos de guanina [Nunez, et al., Biochemistry, 39, 6190, 2000]. Se ha demostrado que el transplatino reacciona con el ADN en un objetivo triplex cuando el TFO está unido al reactivo. Esta reacción provoca la formación de aductos de ADN que serían mutagénicos [Columbier, et al., Nucleic Acids Research, 24: 4519, 1996]. La bleomicina es un rompedor del ADN, ampliamente utilizado como mimético de la radiación. Se ha unido a oligonucleótidos y se ha demostrado que es activo como rompedor en ese formato [Sergeyev, Nucleic Acids Research 23, 4400, 1995; Kane, et al., Biochemistry, 34, 16715, 1995]. Los análogos del ciclopropirroindol se han unido a los TFO y han demostrado que alquilan el ADN en una secuencia objetivo triplex. El ADN alquilado contendría entonces aductos químicos que serían mutagénicos [Lukhtanov, et al., Nucleic Acids Research, 25, 5077, 1997]. Las fenantodihidrodioxinas son quinonas enmascaradas que liberan especies radicales tras la fotoactivación. Se han vinculado a los TFO y se ha demostrado que introducen roturas en el ADN dúplex en la fotoactivación [Bendinskas et al., Bioconjugate Chem. 9, 555, 1998].
De acuerdo con la presente invención, la estrategia para producir una disrupción génica dirigida es a través de la generación de roturas de ADN de una sola cadena causadas por nicasas específicas del sitio. Las endonucleasas se utilizan con mayor frecuencia para la interrupción selectiva de genes en organismos que tradicionalmente han sido refractarios a los procedimientos de orientación de genes más convencionales, tal como las algas, las plantas y los modelos de animales grandes, incluyendo los humanos. Por ejemplo, actualmente se están realizando ensayos clínicos en humanos con nucleasas de dedos de zinc para el tratamiento y la prevención de la infección por el VIH. Además, la ingeniería de endonucleasas se utiliza actualmente para intentar interrumpir los genes que producen fenotipos indeseables en los cultivos.
Los buscadores de objetivos de endonucleasas, también conocidas como meganucleasas, son endonucleasas específicas de secuencia que generan roturas de doble cadena en el ADN genómico con un alto grado de especificidad debido a sus grandes (por ejemplo, >14 pb) sitios de escisión. Si bien la especificidad de los buscadores de objetivos de endonucleasas para sus sitios objetivo permite dirigir con precisión las roturas de ADN inducidas, los sitios de escisión de los buscadores de objetivos de endonucleasas son raros y la probabilidad de encontrar un sitio de corte natural en un gen objetivo es baja.
Los buscadores de objetivos de endonucleasas diseñados se generan modificando la especificidad de los buscadores de objetivos de endonucleasas existentes. En uno de los enfoques, se introducen variaciones en la secuencia de aminoácidos de los buscadores de objetivos de endonucleasas de origen natural y, a continuación, se examinan los buscadores de objetivos de endonucleasas diseñados resultantes para seleccionar las proteínas funcionales que escinden un sitio de unión específico. En otro enfoque, los buscadores de objetivos de endonucleasas quiméricas se diseñan combinando los sitios de reconocimiento de dos buscadores de objetivos de endonucleasas diferentes para crear un nuevo sitio de reconocimiento compuesto por un medio sitio de cada buscador de objetivos de endonucleasas.
Una clase de endonucleasas artificiales son las endonucleasas de dedo de zinc. Las endonucleasas de dedo de zinc combinan un dominio de escisión inespecífico, normalmente el de la endonucleasa Fokl, con dominios de proteína de dedo de zinc diseñados para unirse a secuencias específicas de ADN. La estructura modular de las endonucleasas de dedos de zinc las convierte en una plataforma versátil para realizar roturas de doble cadena específicas en el genoma. Una de las limitaciones de las endonucleasas de dedo de zinc es que la baja especificidad para un sitio objetivo o la presencia de múltiples sitios objetivo en un genoma puede dar lugar a eventos de escisión fuera del objetivo. Dado que la endonucleasa Fokl se escinde como un dímero, una de las estrategias para evitar eventos de escisión fuera del objetivo ha sido diseñar dominios de dedos de zinc que se unen en sitios adyacentes de 9 pares de bases.
Las TALEN son nucleasas direccionables que se utilizan para inducir roturas de una o dos cadenas en sitios específicos del ADN, que luego son reparadas por mecanismos que pueden ser explotados para crear alteraciones de la secuencia en el sitio de escisión.
El bloque de construcción fundamental que se utiliza para diseñar la región de unión al ADN de las TALEN es un dominio de repetición altamente conservado derivado de los TALE naturales codificados por las proteobacterias Xanthomonas spp. La unión al ADN por parte de un TALEN está mediada por conjuntos de repeticiones de 33-35 aminoácidos altamente conservados que están flanqueados por dominios adicionales derivados de TALE en los extremos amino-terminal y carboxi-terminal de las repeticiones.
Estas repeticiones TALE se unen específicamente a una sola base de ADN, cuya identidad está determinada por dos residuos hipervariables que se encuentran típicamente en las posiciones 12 y 13 de la repetición, con el número de repeticiones en un conjunto correspondiente a la longitud del ácido nucleico objetivo deseado, la identidad de la repetición seleccionada para que coincida con la secuencia del ácido nucleico objetivo. El ácido nucleico objetivo está preferentemente entre 15 y 20 pares de bases para maximizar la selectividad del sitio objetivo. La escisión del ácido nucleico objetivo suele producirse en un plazo de 50 pares de bases desde la unión de TALEN. Se han descrito en la
técnica programas informáticos para el diseño del sitio de reconocimiento de TALEN. Véase, por ejemplo Cermak et al., Nucleic Acids Res. 2011 Julio; 39(12): e82.
Una vez diseñadas para que coincidan con la secuencia objetivo deseada, las TALENS pueden ser expresadas recombinantemente e introducidas en protoplastos como proteínas exógenas, o expresadas desde un plásmido dentro del protoplasto.
Estructura del GRON e introducción en las células vegetales
La oligonucleobase recombinante puede ser introducida en una célula vegetal utilizando cualquier procedimiento comúnmente utilizado en la técnica, incluyendo, pero no limitado a, microportadores (entrega biolística), microfibras (bigotes), electroporación, captación directa de ADN y microinyección. A continuación se describen ejemplos ilustrativos de una oligonucleobase recombinante.
La invención puede practicarse con oligonucleobases recombinantes que tengan las conformaciones y químicas descritas en las patentes Kmiec I y Kmiec II. Kmiec I enseña un procedimiento para introducir alteraciones genéticas específicas en un gen objetivo. Las oligonucleobases recombinantes de Kmiec I y/o Kmiec II contienen dos cadenas complementarias, una de las cuales contiene al menos un segmento de nucleótidos de tipo ARN (un "segmento de ARN") que están emparejados en sus bases con los nucleótidos de tipo ADN de la otra cadena.
Kmiec II divulga que los no-nucleótidos que contienen bases de purina y pirimidina pueden ser sustituidos por nucleótidos. Las Pat. de EE.Uu . Nos 5,756,325; 5,871,984; 5,760,012; 5,888,983; 5,795,972; 5,780,296; 5,945,339; 6,004,804; y 6,010,907; y en la Patente Internacional No PCT/US00/23457y en la Publicación Internacional de Patentes Nos. WO 98/49350 WO 99/07865 WO 99/58723 WO 99/58702 WO 99/40789 US 6,870,075;y la Solicitud de Patente Publicada en EE.UU.
20030084473divulgan otras moléculas recombinantes que pueden utilizarse para la presente invención. El término "oligonucleobase recombinante" se utiliza en el presente documento para denotar las moléculas que pueden utilizarse en los procedimientos de la presente invención e incluyen oligonucleótidos dúplex mixtos, moléculas que contienen no nucleótidos enseñadas en Kmiec II, oligodeoxinucleótidos monocatenarios y otras moléculas recombinantes enseñadas en las patentes y publicaciones de patentes mencionadas anteriormente.
En una realización, la oligonucleobase recombinante es un oligonucleótido dúplex mixto en la que los nucleótidos de tipo ARN del oligonucleótido dúplex mixto se hacen resistentes a la RNasa sustituyendo el 2'-hidroxilo por una funcionalidad de fluoro, cloro o bromo o colocando un sustituyente en el 2'-O. Los sustituyentes adecuados incluyen los sustituyentes enseñados por el Kmiec II. Los sustituyentes alternativos incluyen los sustituyentes enseñados por la Pat. de EE.UU. No. 5,334,711 (Sproat) y los sustituyentes enseñados por las publicaciones de patentes e P 629 387 y EP 679657 (colectivamente, las solicitudes Martin). Tal como se utiliza en el presente documento, un derivado 2'-fluoro, cloro o bromo de un ribonucleótido o un ribonucleótido que tenga un 2 '-Oh sustituido con un sustituyente descrito en las Solicitudes Martin o Sproat se denomina "Ribonucleótido 2'-sustituido" Tal como se utiliza en el presente documento, el término "nucleótido tipo ARN" indica un nucleótido 2'-hidroxilo o 2'-sustituido que está unido a otros nucleótidos de un oligonucleótido dúplex mixto mediante un enlace fosfodiéster no sustituido o cualquiera de los enlaces no naturales enseñados por Kmiec I o Kmiec II. Tal como se utiliza en el presente documento, el término "nucleótido de tipo desoxirribonucleótido" indica un nucleótido que tiene un 2'-H, que puede unirse a otros nucleótidos de un MDON mediante un enlace fosfodiéster no sustituido o cualquiera de los enlaces no naturales enseñados por Kmiec I o Kmiec II.
En una realización de la presente invención, la oligonucleobase recombinante es un oligonucleótido dúplex mixto que está unido únicamente por enlaces fosfodiéster no sustituidos. En realizaciones alternativas, el enlace es por fosfodiésteres sustituidos, derivados de fosfodiésteres y enlaces no basados en fósforo como enseña Kmiec II. En otra realización, cada nucleótido de tipo ARN en el oligonucleótido dúplex mixto es un nucleótido 2'-sustituido. Las realizaciones particularmente preferidas de los ribonucleótidos 2'-sustituidos son los ribonucleótidos 2'-fluoro, 2'-metoxi, 2'-propiloxi, 2'-aliloxi, 2'-hidroxiloxi, 2'-metoxiloxi, 2'-fluoropropiloxi y 2'-trifluoropropiloxi sustituidos. Las formas de realización más preferidas de los ribonucleótidos 2'-sustituidos son los nucleótidos 2'-fluoro, 2'-metoxi, 2'-metoxiloxi y 2'-aliloxi sustituidos. En otra realización, el oligonucleótido dúplex mixto está unido por enlaces fosfodiéster no sustituidos.
Aunque los oligonucleótidos dúplex mixtos que tienen un solo tipo de nucleótido de tipo ARN 2'-sustituido se sintetizan más convenientemente, los procedimientos de la invención pueden practicarse con oligonucleótidos dúplex mixtos que tienen dos o más tipos de nucleótidos de tipo ARN. La función de un segmento de ARN puede no verse afectada por una interrupción causada por la introducción de un desoxinucleótido entre dos trinucleótidos de tipo ARN, por lo que el término segmento de a Rn abarca dicho "segmento de ARN interrumpido" Un segmento de ARN ininterrumpido se denomina segmento de ARN contiguo. En una realización alternativa, un segmento de ARN puede contener nucleótidos alternados resistentes a la RNasa y 2'-OH no sustituidos. Los oligonucleótidos dúplex mixtos tienen preferentemente menos de 100 nucleótidos y más preferentemente menos de 85 nucleótidos, pero más de 50 nucleótidos. La primera y la segunda cadena están emparejadas con bases Watson-Crick. En una realización, las cadenas del oligonucleótido dúplex mixto están unidas covalentemente por un enlazador, tal como un hexa, penta o
tetranucleótido de una sola cadena, de modo que las cadenas primera y segunda son segmentos de una sola cadena de oligonucleótidos que tiene un solo extremo 3' y un solo extremo 5'. Los extremos 3' y 5' pueden protegerse mediante la adición de un "capuchón de horquilla" por el que los nucleótidos terminales 3' y 5' se emparejan Watson-Crick con nucleótidos adyacentes. Un segundo capuchón de horquilla puede, además, colocarse en la unión entre las cadenas primera y segunda, distante de los extremos 3' y 5', de modo que se estabilice el emparejamiento Watson-Crick entre las cadenas primera y segunda.
Las cadenas primera y segunda contienen dos regiones que son homólogas con dos fragmentos del gen de la ACCasa objetivo, es decir, tienen la misma secuencia que el gen objetivo. Una región homóloga contiene los nucleótidos de un segmento de ARN y puede contener uno o más nucleótidos de tipo ADN de conexión del segmento de ADN y también puede contener nucleótidos de tipo ADN que no están dentro del segmento de ADN intermedio. Las dos regiones de homología están separadas por, y cada una es adyacente a, una región que tiene una secuencia que difiere de la secuencia del gen objetivo, denominada "región heteróloga" La región heteróloga puede contener uno, dos o tres nucleótidos no coincidentes. Los nucleótidos no coincidentes pueden ser contiguos o, alternativamente, pueden estar separados por uno o dos nucleótidos homólogos con el gen objetivo. Alternativamente, la región heteróloga también puede contener una inserción o uno, dos, tres o de cinco o menos nucleótidos. Alternativamente, la secuencia del oligonucleótido dúplex mixto puede diferir de la secuencia del gen objetivo sólo por la supresión de uno, dos, tres o cinco o menos nucleótidos del oligonucleótido dúplex mixto. En este caso, se considera que la longitud y la posición de la región heteróloga es la longitud de la deleción, aunque ningún nucleótido del oligonucleótido dúplex mixto esté dentro de la región heteróloga. La distancia entre los fragmentos del gen objetivo que son complementarios a las dos regiones homólogas es idéntica a la longitud de la región heteróloga cuando se pretende una sustitución o sustituciones. Cuando la región heteróloga contiene una inserción, las regiones homólogas están así separadas en el oligonucleótido dúplex mixto más que sus fragmentos homólogos complementarios en el gen, y lo contrario es aplicable cuando la región heteróloga codifica una deleción.
Los segmentos de ARN de los oligonucleótidos dúplex mixtos son cada uno una parte de una región homóloga, es decir, una región que es idéntica en secuencia a un fragmento del gen objetivo, cuyos segmentos juntos contienen preferentemente al menos 13 nucleótidos de tipo ARN y preferentemente de 16 a 25 nucleótidos de tipo ARN o aún más preferentemente 18-22 nucleótidos de tipo ARN o más preferentemente 20 nucleótidos. En una realización, los segmentos de ARN de las regiones homológicas están separados y adyacentes, es decir, "conectados por" un segmento de ADN intermedio. En una realización, cada nucleótido de la región heteróloga es un nucleótido del segmento de ADN intermedio. Un segmento de ADN intermedio que contiene la región heteróloga de un oligonucleótido dúplex mixto se denomina "segmento mutador"
El cambio que se va a introducir en el gen objetivo está codificado por la región heteróloga. El cambio que se va a introducir en el Agene puede ser un cambio en una o más bases de la secuencia del gen que cambie el aminoácido nativo en esa posición por el aminoácido deseado.
En otra realización de la presente invención, la oligonucleobase recombinante es un vector mutacional de oligodeoxinucleótidos de una sola cadena o SSOMV, que se divulga en la Solicitud de Patente Internacional PCT/US00/23457. La secuencia del SSOMV se basa en los mismos principios que los vectores mutacionales descritos en las Pat. de EE.UU. Nos.
5,756,325; 5,871,984; 5,760,012; 5,888,983; 5,795,972; 5,780,296; 5,945,339; 6,004,804; y6,010,907y en la Publicación Internacional Nos. WO 98/49350 WO 99/07865 WO 99/58723 WO 99/58702 WO 99/40789 US 6,870,075y la Solicitud de Patente Publicada en EE.UU. 20030084473. La secuencia del SSOMV contiene dos regiones homólogas a la secuencia objetivo separadas por una región que contiene la alteración genética deseada denominada región mutante. La región mutante puede tener una secuencia de la misma longitud que la secuencia que separa las regiones homólogas en la secuencia objetivo, pero con una secuencia diferente. Una región mutante de este tipo provocará una sustitución.
Los nucleótidos del SSOMV son desoxirribonucleótidos que están unidos por enlaces fosfodiéster no modificados, excepto que el enlace internucleotídico terminal 3' y/o terminal 5' o alternativamente los dos enlaces internucleotídicos terminal 3' y/o terminal 5' pueden ser un fosforotioato o un fosfoamidato. Tal y como se utiliza en el presente documento, un enlace internucleotídico es el enlace entre los nucleótidos del SSOMV y no incluye el enlace entre el nucleótido del extremo 3' o el nucleótido del extremo 5' y un sustituyente de bloqueo, véase supra. En una realización específica, la longitud del SSOMV está entre 21 y 55 desoxinucleótidos y las longitudes de las regiones de homología son, en consecuencia, una longitud total de al menos 20 desoxinucleótidos y al menos dos regiones de homología deben tener cada una longitudes de al menos 8 desoxinucleótidos.
El SSOMV puede ser diseñado para ser complementario a la cadena codificante o no codificante del gen objetivo. Cuando la mutación deseada es una sustitución de una sola base, se prefiere que los dos nucleótidos mutadores sean una pirimidina. En la medida en que sea coherente con la consecución del resultado funcional deseado, se prefiere que tanto el nucleótido mutador como el nucleótido objetivo de la cadena complementaria sean pirimidinas. Se prefieren particularmente los SSOMV que codifican mutaciones de transversión, es decir, un nucleótido mutante C o T no coincide, respectivamente, con un nucleótido C o T en la cadena complementaria.
Además del oligodeoxinudeótido, el SSOMV puede contener un sustituyente de bloqueo 5' que se une a los carbonos terminales 5' a través de un enlazador. La química del enlazador no es crítica, salvo su longitud, que debe ser preferiblemente de al menos 6 átomos y que el enlazador debe ser flexible. Se puede utilizar una variedad de sustituyentes no tóxicos tales como la biotina, el colesterol u otros esteroides o un colorante fluorescente catiónico no intercalante. Particularmente preferidos como reactivos para hacer SSOMV son los reactivos vendidos como Cy3™ y Cy5™ por Glen Research, Sterling VA (ahora GE Healthcare), que son fosforoamiditas bloqueadas que al incorporarse a un oligonucleótido dan lugar a los colorantes 3,3,3',3'-tetrametil N,N'-isopropil sustituidos indomonocarbocianina e indodicarbocianina, respectivamente. El Cy3 es el más preferido. Cuando la indocarbocianina está sustituida por N-oxialquilo, puede unirse convenientemente al terminal 5' del oligodeoxinucleótido a través de un fosfodiéster con un fosfato terminal 5'. La química del enlazador del colorante entre el colorante y el oligodeoxinucleótido no es crítica y se elige por conveniencia sintética. Cuando se utiliza la fosforamidita Cy3 disponible en el mercado, la modificación 5' resultante consiste en un sustituto de bloqueo y un enlazador juntos que son un N-hidroxipropilo, N'-fosfatidilpropilo 3,3,3',3'-tetrametil indomonocarbocianina.
En una realización preferida, el colorante de indocarbocianina está tetra sustituido en las posiciones 3 y 3' de los anillos de indol. Sin limitación en cuanto a la teoría, estas sustituciones impiden que el colorante sea un colorante intercalante. La identidad de los sustituyentes en estas posiciones no es crítica. El SSOMV puede tener además un sustituto de bloqueo 3'. De nuevo, la química del sustituto de bloqueo 3' no es crítica.
En otra realización preferida, el oligonucleótido recombinante es un oligodeoxinucleótido monocatenario que tiene un nucleótido en el extremo 3', un nucleótido en el extremo 5', que tiene al menos 25 desoxinucleótidos y no más de 65 desoxinucleótidos, y que tiene una secuencia que comprende al menos dos regiones de al menos 8 desoxinucleótidos cada una, respectivamente idénticas a al menos dos regiones del gen cromosómico objetivo, cuyas regiones juntas tienen una longitud de al menos 24 nucleótidos, y cuyas regiones están separadas por al menos un nucleótido en la secuencia del gen cromosómico objetivo o en la secuencia del oligodesoxinucleótido, o ambas, de manera que la secuencia del oligodesoxinucleótido no es idéntica a la secuencia del gen cromosómico objetivo. Véase Patente de EE.UU 6,271,360.
Microportadores y Microfibras
El uso de microportadores metálicos (microesferas) para introducir grandes fragmentos de ADN en células vegetales con paredes celulares de celulosa mediante la penetración de proyectiles es bien conocido por los expertos en la técnica (en adelante, entrega biolística). La Pat. de EE.UU. Nos. 4,945,050; 5,100,792 y 5,204,253 describen técnicas generales de selección de microportadores y dispositivos para proyectarlos. La Pat. de EE.UU. Nos 5,484,956 y 5,489,520 describen la preparación de maíz transgénico fértil mediante el bombardeo con microproyectiles de tejido de callo de maíz. Las técnicas biológicas también se utilizan para transformar embriones de maíz inmaduros.
Las condiciones específicas para el uso de microportadores en los procedimientos de la presente invención se describen en la Publicación Internacional WO 99/07865. En una técnica ilustrativa, se añaden microportadores fríos (60 mg/ml), oligonucleótidos dúplex mezclados (60 mg/ml) 2,5 M CaChy 0,1 M de espermidina en ese orden; la mezcla se agita suavemente, por ejemplo, mediante vórtex, durante 10 minutos y se deja reposar a temperatura ambiente durante 10 minutos, tras lo cual los microportadores se diluyen en 5 volúmenes de etanol, se centrifugan y se resuspenden en etanol al 100 %. Se pueden obtener buenos resultados con una concentración en la solución adherente de 8-10 pg/pl de microportadores, 14-17 pg/ml de oligonucleótido dúplex mixto, 1,1-1,4 M de CaC2 y 18-22 mM de espermidina. Los resultados óptimos se observaron en las condiciones de 8 pg/pl de microportadores, 16,5 pg/ml de oligonucleótido dúplex mixto, 1,3 M de CaCl y 21 mM de espermidina.
Las oligonucleobases recombinantes también pueden introducirse en las células vegetales para la práctica de la presente invención utilizando microfibras para penetrar la pared y la membrana celular. La Pat. de EE.UU. No.
5,302,523 de Coffee et al. describe el uso de fibras de carburo de silicio de 30.veces.0.5 pm y 10.veces.0.3 pm para facilitar la transformación de cultivos de maíz en suspensión de Black Mexican Sweet. Cualquier técnica mecánica que pueda utilizarse para introducir ADN para la transformación de una célula vegetal utilizando microfibras puede utilizarse para entregar oligonucleobases recombinantes para su uso en la fabricación de los presentes mutantes de la ACCasa. Una técnica ilustrativa para la entrega de microfibras de una oligonucleobase recombinante es la siguiente: Las microfibras estériles (2 pg) se suspenden en 150 pl de medio de cultivo vegetal que contiene aproximadamente10 pg de un oligonucleótido dúplex mixto. Se deja sedimentar un cultivo en suspensión y se mezclan en vórtex volúmenes iguales de células empaquetadas y de la suspensión estéril de fibras/nucleótidos durante 10 minutos y se colocan en placas. Los medios selectivos se aplican inmediatamente o con un retraso de hasta aproximadamente 120 horas, según convenga para el rasgo concreto.
Electroporación
En una realización alternativa, las oligonucleobases recombinantes pueden ser entregadas a la célula vegetal por electroporación de un protoplasto derivado de una parte de la planta de acuerdo con las técnicas bien conocidas por
un experto en la técnica. Véase, por ejemplo Gallois et al., 1996, en Methods in Molecular Biology 55:89-107, Humana Press, Totowa, N.J. Kipp et al., 1999, en Methods in Molecular Biology 133:213-221, Humana Press, Totowa, N.J.
Las oligonucleobases recombinantes también pueden introducirse en las microsporas por electroporación. Tras la liberación de la tétrada, la microspora es uninucleada y de paredes finas. Comienza a agrandarse y desarrolla un germoporo antes de que se forme la exina. Una microspora en esta etapa es potencialmente más susceptible de ser transformada con ADN exógeno que otras células vegetales. Además, el desarrollo de las microsporas puede alterarse in vitro para producir embriones haploides o callos embriogénicos que pueden regenerarse en plantas (Coumans et al., Plant Cell Rep. 7:618-621, 1989 Datta et al., Plant Sci. 67:83-88, 1990 Maheshwari et al., Am. J Bot. 69:865-879, 1982 Schaeffer, Adv. En Cell Culture 7:161-182, 1989 Swanson et al., Plant Cell Rep. 6:94-97, 1987). Así, las microsporas transformadas pueden regenerarse directamente en plantas haploides o en plantas fértiles dihaploides tras la duplicación cromosómica por procedimientos estándar. Véase también la solicitud copendiente U.S. Ser. No 09/680,858 titulada Compositions and Methods for Plant Genetic Modification.
Los procedimientos de electroporación de microesporas se describen en Jardinaud et al., Plant Sci. 93:177-184, 1993 y Fennell y Hauptman, Plant Cell Reports 11:567-570, 1992. Los procedimientos de electroporación de MDON en protoplastos de plantas también pueden adaptarse para su uso en la electroporación de microesporas.
Bigotes y microinyección
En otra realización alternativa, la oligonucleobase recombinante puede ser entregada a la célula vegetal por medio de bigotes o microinyección de la célula vegetal. La llamada técnica de los bigotes se realiza esencialmente como se describe en Frame et al., 1994, Plant J. 6:941-948. La oligonucleobase recombinante se añade a los bigotes y se utiliza para transformar las células vegetales. La oligonucleobase recombinante puede coincubarse con plásmidos que comprenden secuencias que codifican proteínas capaces de formar complejos de recombinasa en las células vegetales, de manera que se catalice la recombinación entre el oligonucleótido y la secuencia objetivo.
Selección de plantas
En diversas realizaciones, las plantas divulgadas en el presente documento pueden ser de cualquier especie de planta dicotiledónea, monocotiledónea o gimnosperma, incluyendo cualquier especie de planta leñosa que crezca como árbol o arbusto, cualquier especie herbácea, o cualquier especie que produzca frutos, semillas o verduras comestibles, o cualquier especie que produzca flores coloridas o aromáticas. Por ejemplo, la planta puede seleccionarse de una especie de planta del grupo que consiste en canola, girasol, maíz, tabaco, remolacha azucarera, algodón, maíz, trigo, cebada, arroz, alfafa sorgo, tomate, mango, melocotón, manzana, pera, fresa, plátano, melón, patata, zanahoria, lechuga, cebolla, soja, soya spp, caña de azúcar, guisante, garbanzo, guisante de campo, faba, lentejas, nabo, colinabo, coles de Bruselas, altramuz, coliflor, col rizada, habas, álamo, pino, eucalipto, uva, cítricos, triticale, alfalfa, centeno, avena, césped y hierbas forrajeras, lino, colza, mostaza, pepino, gloria de la mañana, bálsamo, pimiento, berenjena, caléndula, loto, col, margarita, clavel, tulipán, iris, lirio y plantas productoras de frutos secos, en la medida en que no se mencionen específicamente.
Las plantas y las células vegetales pueden someterse a pruebas de resistencia o tolerancia a un herbicida utilizando procedimientos comúnmente conocidos en la técnica, por ejemplo, cultivando la planta o la célula vegetal en presencia de un herbicida y midiendo la tasa de crecimiento en comparación con la tasa de crecimiento en ausencia del herbicida.
Tal como se utiliza en el presente documento, el crecimiento sustancialmente normal de una planta, órgano vegetal, tejido vegetal o célula vegetal se define como una tasa de crecimiento o tasa de división celular de la planta, órgano vegetal, tejido vegetal o célula vegetal que es al menos 35 %, al menos 50 %, al menos 60 % o al menos 75 % de la tasa de crecimiento o tasa de división celular en una planta, órgano vegetal, tejido vegetal o célula vegetal correspondiente que expresa la proteína AHAS de tipo silvestre.
Tal como se utiliza en el presente documento, el desarrollo sustancialmente normal de una planta, órgano vegetal, tejido vegetal o célula vegetal se define como la ocurrencia de uno o más eventos de desarrollo en la planta, órgano vegetal, tejido vegetal o célula vegetal que son sustancialmente iguales a los que ocurren en una planta, órgano vegetal, tejido vegetal o célula vegetal correspondiente que expresa la proteína de tipo silvestre.
En ciertas realizaciones, los órganos vegetales proporcionados en el presente documento incluyen, pero no se limitan a, hojas, tallos, raíces, yemas vegetativas, yemas florales, meristemos, embriones, cotiledones, endosperma, sépalos, pétalos, pistilos, carpelos, estambres, anteras, microsporas, polen, tubos de polen, óvulos, ovarios y frutos, o secciones, rodajas o discos tomados de los mismos. Los tejidos vegetales incluyen, pero no se limitan, los tejidos de callo, los tejidos del suelo, los tejidos vasculares, los tejidos de almacenamiento, los tejidos meristemáticos, los tejidos de las hojas, los tejidos de los brotes, los tejidos de las raíces, los tejidos de las agallas, los tejidos de los tumores vegetales y los tejidos reproductivos. Las células vegetales incluyen, pero no se limitan a, células aisladas con paredes celulares, agregados de diferentes tamaños y protoplastos.
Las plantas son sustancialmente "tolerantes" a un herbicida relevante cuando son sometidas a este y proporcionan
una curva dosis/respuesta que se desplaza hacia la derecha cuando se compara con la proporcionada por una planta similar no tolerante sometida. Estas curvas de dosis/respuesta tienen la "dosis" trazada en el eje X y el "porcentaje de muerte", el "efecto herbicida", etc., trazado en el eje Y. Las plantas tolerantes necesitarán más herbicida que las plantas similares no tolerantes para producir un efecto herbicida determinado. Las plantas que son sustancialmente "resistentes" al herbicida presentan pocas, si hay, lesiones necróticas, líticas, cloróticas o de otro tipo, cuando se las somete al herbicida en las concentraciones y dosis que suelen emplear los agroquímicos para matar las malas hierbas en el campo. Las plantas resistentes a un herbicida son también tolerantes al herbicida.
Generación de plantas
Se conoce el cultivo de tejidos de diversas especies vegetales y la regeneración de plantas a partir de ellos. Por ejemplo, la propagación de un cultivar de canola por cultivo de tejidos se describe en cualquiera de los siguientes, pero no se limita a ninguno de ellos: Chuong et al., "A Simple Culture Method for Brassica hypocotyls Protoplasts", Plant Cell Reports 4:4-6, 1985 Barsby, T. L., et al., "A Rapid and Efficient Alternative Procedure for the Regeneration of Plants from Hypocotyl Protoplasts of Brassica napus," Plant Cell Reports (Spring, 1996) Kartha, K., et al., "In vitro Plant Formation from Stem Explants of Rape," Physiol. Plant, 31:217-220, 1974 Narasimhulu, S., et al., "Species Specific Shoot Regeneration Response of Cotyledonary Explants of Brassicas," Plant Cell Reports (Spring 1988) Swanson, E., "Microspore Culture in Brassica," Methods in Molecular Biology, Vol. 6, Chapter 17, p. 159, 1990.
La reproducción adicional de la variedad puede producirse mediante el cultivo de tejidos y la regeneración. El cultivo de tejidos de diversos tejidos de soja y la regeneración de plantas a partir de los mismos es bien conocido y ampliamente publicado. Por ejemplo, se puede hacer referencia a Komatsuda, T. et al., "Genotype X Sucrose Interactions for Somatic Embryogenesis in Soybeans", Crop Sci. 31:333-337, 1991 Stephens, P. A., et al., "Agronomic Evaluation of Tissue-Culture-Derived Soybean Plants", Theor. Appl. Genet. 82:633-635, 1991 Komatsuda, T. et al., "Maturation and Germination of Somatic Embryos as Affected by Sucrose and Plant Growth Regulators in Soybeans Glycine gracilis Skvortz and Glycine max (L.) Merr." Plant Cell, Tissue and Organ Culture, 28:103-113, 1992 Dhir, S. et al., "Regeneration of Fertile Plants from Protoplasts of Soybean (Glycine max L. Merr.); Genotypic Differences in Culture Response," Plant Cell Reports 11:285-289, 1992 Pandey, P. et al., "Plant Regeneration from Leaf and Hypocotyl Explants of Glycine wightii (W. and A.) VERDC. var. longicauda," Japan J. Breed. 42:1-5, 1992y Shetty, K., et al., "Stimulation of In Vitro Shoot Organogenesis in Glycine max (Merrill.) by Allantoin and Amides," Plant Science 81:245-251, 1992. Las divulgaciones de Pat. de EE.UU. No. 5,024,944 expedido el 18 de junio de 1991 a Collins et al.y Pat. de EE.UU. No. 5,008,200 expedido el 16 de abril de 1991 a Ranch et al.
Los siguientes ejemplos ilustran la práctica de la presente invención, pero no deben interpretarse como una limitación de su alcance.
Ejemplo 1: Mejora drástica de la conversión de un gen de la proteína azul fluorescente (BFP) en células transgénicas de Arabidopsis thaliana a una proteína verde fluorescente (g Fp ) mediante la introducción de una mutación de un solo nucleótido dirigida a través de oligoucleótidos de reparación de genes (GRON) en combinación con un par de nucleasas de efecto similar al activador de la transcripción (TALEN) que escinde cerca del cambio de base dirigido.
Se creó una línea de Arabidopsis con múltiples copias de un gen de proteína azul fluorescente mediante procedimientos conocidos por los expertos en la técnica (véase, por ejemplo, Clough y Brent, 1998). Con esta línea se establecieron cultivos de tejidos meristemáticos derivados de la raíz, que se utilizaron para el aislamiento y cultivo de protoplastos (véase, por ejemplo, Mathur et al., 1995). El suministro de GRON a los protoplastos se logró mediante la captación de GRON por parte del polietilenglicol (PEG) en los protoplastos. Se utilizó un procedimiento en formato de 96 pocillos, similar al descrito por Fujiwara y Kato (2007). A continuación se describe brevemente el protocolo. Los volúmenes indicados son los aplicados a pocillos individuales de una placa de 96 pocillos.
1. Mezclar 6,25 pl de la mezcla GRON/TALEN (80 pM BFP4 Coding/41mer GRON) con 25 pl de protoplastos derivados de tejido meristemático de raíces transgénicas de Arabidopsis a 5*106 células/ml en cada pocillo de una placa de 96 pocillos.
2. Se añadieron 31, 25 pl de una solución de PEG al 40% y se mezclaron los protoplastos.
3. Las células tratadas se incubaron en hielo durante 30 minutos.
4. A cada pocillo se añadieron 200 pl de solución W5 y se mezclaron las células.
5. Las placas se dejaron incubar en hielo durante 30 minutos para que los protoplastos se depositaran en el fondo de cada pocillo.
6. Se retiraron 200 pl del medio por encima de los protoplastos asentados.
7. Se añadieron 85 pl de medio de cultivo (MSAP, véase Mathur et al., 1995).
8. Las placas se incubaron a temperatura ambiente en la oscuridad durante 48 horas. La concentración final de GRON tras añadir el medio de cultivo es de 8 pM.
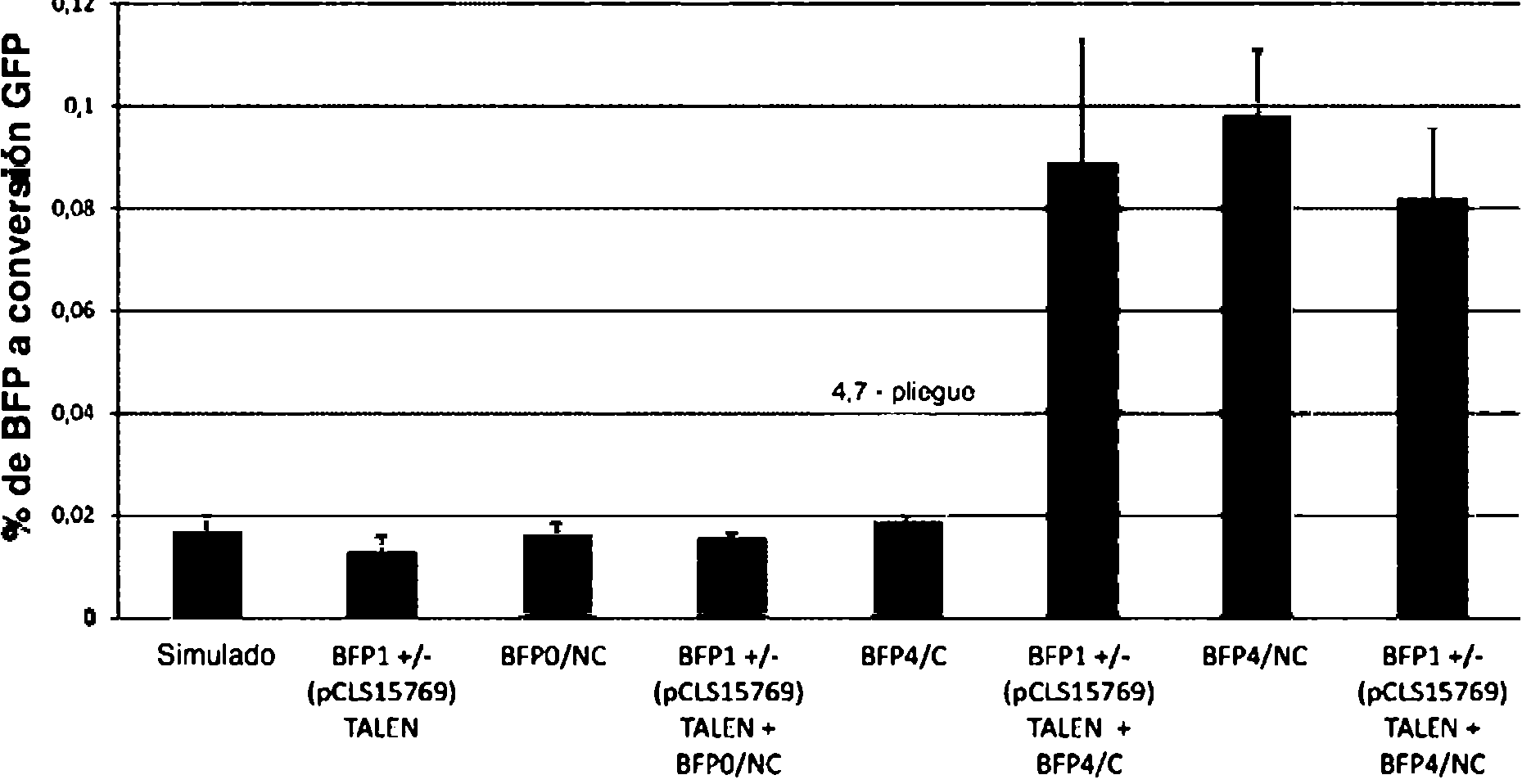
Utilizando este protocolo, se introdujeron plásmidos TALEN a diferentes concentraciones junto con GRON. Cuarenta y ocho horas después de la administración de GRON se analizaron las muestras mediante citometría de flujo para detectar los protoplastos cuya fluorescencia verde y amarilla es diferente de la de los protoplastos de control. La
fluorescencia verde se debe a la introducción de una mutación dirigida en el gen BFP, que da lugar a la síntesis de GFP. Los resultados se muestran en la Figura 1.

Figura 1. Los protoplastos derivados del tejido meristemático de la raíz se trataron con plásmidos TALEN a diversas concentraciones junto con GRON dirigidos a una mutación en el gen BFP, provocando una conversión en un gen GFP. La expresión de GFP se midió por citometría de flujo 48 h después de la administración de GRON/TALEN
Para todos los Ejemplos 2-11 siguientes, se aplica la siguiente Leyenda:
GRONs
Diseños de orientación BFP->GFP.
BFP->GFP H66Y CAC->TAC.
BFP4/C/41/5'Cy3/3'idC
VCCCTCGTGACCACCTTCACCTACGGCGTGCAGTGCTTCAGCH BFP4/NC/41/5'Cy3/3'idC
VGCTGAAGCACTGCACGCCGTAGGTGAAGGTGGTCACGAGGGH
Diseños de control sin objetivo de BFP.
BFP H66 - CAC
BFP0/C/41/5'3PS/3'3PS
VCCCTCGTGACCACCTTCACCCACGGCGTGCAGTGCTTCAGCH BFP/NC/41/5'3PS/3'3PS
VGCTGAAGCACTGCACGCCGTGGGTGAAGGTGGTCACGAGGGH
TALENs
Elector TAL
Dominio de unión a ADN

CTTCATGTGGTCGGGGTAGCGGCTGAAGCACTGCACGCCGTGGGTGAAGGTGGTCAC GAGGGTGGGCCAGGGCACGGG A

El pCLS14165 tiene ambos brazos TAL (diseñados según Zhang et al., 2013) en un solo plásmido con cada brazo que se une a la secuencia subrayada y se une a un monómero Fokl. Esta combinación produce una rotura de doble cadena (DSB) como se muestra a continuación en el ensayo de fusión de una sola cadena (SSA) realizado de la misma manera que los de Zhang et al. (2013)

El pCLS 15771 es una nicasa con una mutación (D450A) en el dominio Fokl del brazo izquierdo. El brazo derecho de esta construcción es el de pCLS14165.
EL pCLS 15769 es una nicasa con una mutación (D450A) en el dominio Fokl del brazo derecho. El brazo izquierdo de esta construcción es el de pCLS14165.
Los GRONs y TALENs fueron probados como se muestra a continuación. Los tratamientos de control consistentes en GRONs no dirigidos (BFP0/C o BFP0/NC) y TALENs solos, así como los tratamientos simulados con la solución de PEG al 40% que carece de GRONs o del plásmido TALEN, no tuvieron una actividad de conversión significativa.
En este sistema, el diseño BFP4/NC GRON solo es mejor que el diseño BFP4/C GRON solo. La combinación de estos con el TALEN DSB (pCLS 14165) mejora ambos con la mejor actividad y mejora de pliegues (en muchos casos >2 órdenes de magnitud) con el BFP4/C GRON. También se observan mejoras significativas en la combinación de GRON con pares TALEN de nicasa y se espera que sean más beneficiosas al minimizar los daños colaterales cuando las mutaciones se dirigen a varios genes/loci/alelos simultáneamente.
Ejemplo 2: GRON más TALEN nicasa.

Ejemplo 3: GRON más TALEN nicasa.
Ejemplo 4 : GRON más TALEN nicasa.

Ejemplo 6: GRON más TALEN nicasa.
0,07

Ejemplo 7 : GRON más TALEN nicasa.

Ejemplo 8 : GRON más TALEN DSB.

Simulado BFP1+/+ BFPO/NC BFP4/C BFP1 /+ BFP4/NC BFP1 /+ (PCLS14165) (pCLS14165) (PCLS14165) TALEN TALEN TALEN BFP4/C BFP4/NC
Ejemplo 9: GRON más TALEN.

Ejemplo 10: GRON más TALEN nicasa.
Ejemplo 11: GRON más TALEN DSB.

Referencias
Clough, S.J., y Bent, A.F. (1998). Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735-743.
Mathur, J., Szabados, L, and Koncz, C. (1995) A simple method for isolation, liquid culture, transformation and regeneration of Arabidopsis thaliana protoplasts. Plant Cell Rep. 14, 221-226
Fujikawa Y, Kato N (2007) Split luciferase complementaron assay to study protein-protein interactions in Arabidopsis protoplasts. Planta J 52: 185-195
Zhang Y, Zhang F, Li X, Baller JA, Qi Y, Starker CG, Bogdanove AJ, Voytas DF. (2013) Transcription activator-like effector nucleases enable efficient plant genome engineering. Plant Physiol. 161(1):20-7.
Claims (14)
1. Un procedimiento para introducir una mutación mediada por una oligonucleobase de reparación de genes (GRON) en una secuencia de ácido desoxirribonucleico (ADN) objetivo en una célula vegetal, que comprende:
cultivar la célula bajo condiciones que aumenten uno o más procesos de reparación del ADN celular antes de, y/o coincidiendo con, la entrega de un GRON en la célula vegetal,
en el que las condiciones que aumentan uno o más procesos celulares de reparación del ADN comprenden la introducción de una o más nicasas específicas de sitio en la célula vegetal,
en el que una o más nicasas específicas del sitio introducen mellas en la cadena simple en la vecindad del sitio que se pretende convertir con el GRON.
2. El procedimiento de la reivindicación 1, en el que la una o más nicasas se seleccionan entre las nicasas TALEN, las nicasas de dedos de zinc y las nicasas meganucleas.
3. El procedimiento de la reivindicación 1 o 2, en el que la una o más nicasas específicas del sitio están acopladas covalentemente al GRON.
4. El procedimiento de una de las reivindicaciones 1 a 3, en el que la célula es una célula vegetal de una especie seleccionada del grupo que consiste en canola, girasol, maíz, tabaco, remolacha azucarera, algodón, maíz, trigo, cebada, arroz, alfafa, cebada, sorgo, tomate, mango, melocotón, manzana, pera, fresa, plátano, melón, patata, zanahoria, lechuga, cebolla, soja, soya spp, caña de azúcar, guisante garbanzo, guisante, faba, lentejas, nabo, colinabo, coles de Bruselas, altramuz, coliflor, col rizada, habas, álamo, pino, eucalipto, uva, cítricos, triticale, alfalfa, centeno, avena, césped y hierbas forrajeras, lino, colza, mostaza, pepino, gloria de la mañana, bálsamo, pimiento, berenjena, caléndula, loto, col, margarita, clavel, tulipán, iris y lirio.
5. El procedimiento de una de las reivindicaciones 1-4, en el que la célula es transgénica.
6. El procedimiento de la reivindicación 5, en el que la secuencia de ADN objetivo es un gen endógeno de la célula.
7. El procedimiento de una de las reivindicaciones 1-6, en el que el procedimiento comprende además regenerar una planta que tiene una mutación introducida por el GRON a partir de la célula vegetal.
8. Uso de una o más nicasas para mejorar la conversión mutacional mediante oligonucleótidos reparadores de genes (GRONs) como donantes, comprendiendo la introducción de la una o más nicasas específicas de sitio en una célula.
9. El uso de la reivindicación 8, en el que la una o más nicasas se seleccionan entre nicasas TALEN, nicasas de dedos de zinc y nicasas meganucleas.
10. El procedimiento de la reivindicación 1, que comprende además la evaluación de la eficiencia de las conversiones del GRON, comprendiendo el procedimiento:
introducir el GRON en un protoplasto vegetal que exprese una secuencia de ADN objetivo que codifique la proteína azul fluorescente, en el que el GRON esté configurado para mutar la secuencia de ADN objetivo en un sitio predeterminado para provocar una conversión de la proteína azul fluorescente por la que la proteína azul fluorescente emita fluorescencia a una longitud de onda diferente, en el que antes de introducir el GRON o coincidiendo con él, el ADN dentro del protoplasto se ponga en contacto con dicha una o más nicasas específicas del sitio que introduzcan mellas de una sola cadena en la secuencia de ADN objetivo dentro del sitio al que apunta el GRON; y
determinar la eficacia de la conversión.
11. El procedimiento de la reivindicación 10, en el que la una o más nicasas de sitio específico se seleccionan entre las nicasas TALEN, las nicasas de dedo de zinc y las nicasas meganucleas.
12. El procedimiento de la reivindicación 10 u 11, en el que la secuencia de ADN objetivo está presente en el ADN cromosómico del protoplasto
13. El procedimiento de la reivindicación 10 u 11, en el que la secuencia de ADN objetivo está presente en un plásmido.
14. El procedimiento de una de las reivindicaciones 10-13, en el que la una o más nicasas específicas de sitio proporcionan una eficiencia de conversión aumentada en relación con una eficiencia determinada para el GRON en ausencia de la una o más nicasas.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201361801320P | 2013-03-15 | 2013-03-15 | |
| PCT/US2014/029621 WO2014144987A2 (en) | 2013-03-15 | 2014-03-14 | Methods and compositions for increasing efficiency of increased efficiency of targeted gene modification using oligonucleotide-mediated gene repair |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2912400T3 true ES2912400T3 (es) | 2022-05-25 |
Family
ID=51538403
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES14763072T Active ES2912400T3 (es) | 2013-03-15 | 2014-03-14 | Modificación génica orientada mediante la reparación génica mediada por oligonucleótidos |
Country Status (24)
| Country | Link |
|---|---|
| US (2) | US11434494B2 (es) |
| EP (1) | EP2966984B1 (es) |
| JP (1) | JP2016516408A (es) |
| KR (1) | KR102304487B1 (es) |
| CN (1) | CN105338805A (es) |
| AU (2) | AU2014233465A1 (es) |
| BR (1) | BR112015022848B1 (es) |
| CA (1) | CA2905135C (es) |
| CL (1) | CL2015002650A1 (es) |
| DK (1) | DK2966984T3 (es) |
| EA (1) | EA201591447A1 (es) |
| ES (1) | ES2912400T3 (es) |
| HK (1) | HK1220864A1 (es) |
| HR (1) | HRP20220521T1 (es) |
| HU (1) | HUE058351T2 (es) |
| IL (1) | IL241145B (es) |
| LT (1) | LT2966984T (es) |
| MY (1) | MY191390A (es) |
| PL (1) | PL2966984T3 (es) |
| PT (1) | PT2966984T (es) |
| RS (1) | RS63188B1 (es) |
| SI (1) | SI2966984T1 (es) |
| UA (1) | UA121099C2 (es) |
| WO (1) | WO2014144987A2 (es) |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2905135C (en) | 2013-03-15 | 2022-10-25 | Cibus Us Llc | Targeted gene modification using oligonucleotide-mediated gene repair |
| CA2997535A1 (en) * | 2015-07-23 | 2017-01-26 | Mayo Foundation For Medical Education And Research | Editing mitochondrial dna |
| EP3456825A4 (en) | 2016-05-13 | 2020-03-18 | Kaneka Corporation | Plant genome editing method |
| CN109152343A (zh) * | 2016-05-13 | 2019-01-04 | 株式会社钟化 | 转化植物的制备方法 |
| US11499158B2 (en) | 2016-05-13 | 2022-11-15 | Kaneka Corporation | Method for modifying plant |
| CN106755067A (zh) * | 2016-12-05 | 2017-05-31 | 天津吉诺沃生物科技有限公司 | 通过瞬时表达对植物基因进行定点插入的方法及获得的瞬时表达细胞、组织和突变植株 |
| US11859219B1 (en) | 2016-12-30 | 2024-01-02 | Flagship Pioneering Innovations V, Inc. | Methods of altering a target nucleotide sequence with an RNA-guided nuclease and a single guide RNA |
| WO2020159470A1 (en) | 2019-01-28 | 2020-08-06 | Mayo Foundation For Medical Education And Research | Mitochondrial genome editing methods |
Family Cites Families (37)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5100792A (en) | 1984-11-13 | 1992-03-31 | Cornell Research Foundation, Inc. | Method for transporting substances into living cells and tissues |
| US4945050A (en) | 1984-11-13 | 1990-07-31 | Cornell Research Foundation, Inc. | Method for transporting substances into living cells and tissues and apparatus therefor |
| US5024944A (en) | 1986-08-04 | 1991-06-18 | Lubrizol Genetics, Inc. | Transformation, somatic embryogenesis and whole plant regeneration method for Glycine species |
| US5008200A (en) | 1988-05-08 | 1991-04-16 | United Agriseeds, Inc. | Propagating multiple whole fertile plants from immature leguminous |
| US5302523A (en) | 1989-06-21 | 1994-04-12 | Zeneca Limited | Transformation of plant cells |
| US5550318A (en) | 1990-04-17 | 1996-08-27 | Dekalb Genetics Corporation | Methods and compositions for the production of stably transformed, fertile monocot plants and cells thereof |
| US5484956A (en) | 1990-01-22 | 1996-01-16 | Dekalb Genetics Corporation | Fertile transgenic Zea mays plant comprising heterologous DNA encoding Bacillus thuringiensis endotoxin |
| US5204253A (en) | 1990-05-29 | 1993-04-20 | E. I. Du Pont De Nemours And Company | Method and apparatus for introducing biological substances into living cells |
| DE4216134A1 (de) | 1991-06-20 | 1992-12-24 | Europ Lab Molekularbiolog | Synthetische katalytische oligonukleotidstrukturen |
| IT230274Y1 (it) | 1993-06-11 | 1999-06-02 | Silc Spa | Pannolone assorbente sagomato per incontinenza |
| DE733059T1 (de) | 1993-12-09 | 1997-08-28 | Univ Jefferson | Verbindungen und verfahren zur ortsspezifischen mutation in eukaryotischen zellen |
| PT679657E (pt) | 1994-04-27 | 2003-11-28 | Novartis Ag | Nucleosidos e oligonucleotidos com grupos 2'-eter |
| US5780296A (en) | 1995-01-17 | 1998-07-14 | Thomas Jefferson University | Compositions and methods to promote homologous recombination in eukaryotic cells and organisms |
| US5888983A (en) | 1996-05-01 | 1999-03-30 | Thomas Jefferson University | Method and oligonucleobase compounds for curing diseases caused by mutations |
| US5760012A (en) | 1996-05-01 | 1998-06-02 | Thomas Jefferson University | Methods and compounds for curing diseases caused by mutations |
| US5731181A (en) | 1996-06-17 | 1998-03-24 | Thomas Jefferson University | Chimeric mutational vectors having non-natural nucleotides |
| US6524613B1 (en) | 1997-04-30 | 2003-02-25 | Regents Of The University Of Minnesota | Hepatocellular chimeraplasty |
| EP0979311A1 (en) | 1997-04-30 | 2000-02-16 | Of The University Of Minnesota Regents | $i(IN VIVO) USE OF RECOMBINAGENIC OLIGONUCLEOBASES TO CORRECT GENETIC LESIONS IN HEPATOCYTES |
| US7094606B2 (en) | 1997-08-05 | 2006-08-22 | Arntzen Charles J | Use of mixed duplex oligonucleotides to effect localized genetic changes in plants |
| US6004804A (en) | 1998-05-12 | 1999-12-21 | Kimeragen, Inc. | Non-chimeric mutational vectors |
| US6010907A (en) | 1998-05-12 | 2000-01-04 | Kimeragen, Inc. | Eukaryotic use of non-chimeric mutational vectors |
| US6271360B1 (en) | 1999-08-27 | 2001-08-07 | Valigen (Us), Inc. | Single-stranded oligodeoxynucleotide mutational vectors |
| AR025996A1 (es) | 1999-10-07 | 2002-12-26 | Valigen Us Inc | Plantas no transgenicas resistentes a los herbicidas. |
| EP1152058A1 (en) * | 2000-05-03 | 2001-11-07 | Institut Curie | Methods and compositions for effecting homologous recombination |
| WO2003013226A2 (en) | 2001-08-09 | 2003-02-20 | Cibus Genetics | Non-transgenic herbicide resistant plants |
| CN108130336A (zh) * | 2007-10-05 | 2018-06-08 | 赛布斯欧洲公司 | 芸苔属中突变的乙酰羟酸合酶基因 |
| KR20140050759A (ko) * | 2007-12-21 | 2014-04-29 | 키진 엔.브이. | 식물 원형질체 내로 폴리에틸렌 글리콜 매개 돌연변이 뉴클레오염기의 도입을 이용한 개선된 돌연변이 생성방법 |
| CA2720903C (en) * | 2008-04-14 | 2019-01-15 | Sangamo Biosciences, Inc. | Linear donor constructs for targeted integration |
| WO2011078662A1 (en) | 2009-12-21 | 2011-06-30 | Keygene N.V. | Dsrna for improved genetic modification of plant dna |
| AU2011265733B2 (en) * | 2010-06-14 | 2014-04-17 | Iowa State University Research Foundation, Inc. | Nuclease activity of TAL effector and Foki fusion protein |
| US9512444B2 (en) * | 2010-07-23 | 2016-12-06 | Sigma-Aldrich Co. Llc | Genome editing using targeting endonucleases and single-stranded nucleic acids |
| AU2012249390B2 (en) | 2011-04-27 | 2017-03-30 | Amyris, Inc. | Methods for genomic modification |
| JP2014520551A (ja) * | 2011-07-11 | 2014-08-25 | セルラー ダイナミクス インターナショナル, インコーポレイテッド | 細胞のリプログラミング方法およびゲノムの改変方法 |
| AU2013266968B2 (en) * | 2012-05-25 | 2017-06-29 | Emmanuelle CHARPENTIER | Methods and compositions for RNA-directed target DNA modification and for RNA-directed modulation of transcription |
| CA2890160A1 (en) * | 2012-10-31 | 2014-05-08 | Cellectis | Coupling herbicide resistance with targeted insertion of transgenes in plants |
| CA2905135C (en) | 2013-03-15 | 2022-10-25 | Cibus Us Llc | Targeted gene modification using oligonucleotide-mediated gene repair |
| US9680858B1 (en) | 2013-09-09 | 2017-06-13 | BitSight Technologies, Inc. | Annotation platform for a security risk system |
-
2014
- 2014-03-14 CA CA2905135A patent/CA2905135C/en active Active
- 2014-03-14 SI SI201431952T patent/SI2966984T1/sl unknown
- 2014-03-14 PT PT147630727T patent/PT2966984T/pt unknown
- 2014-03-14 US US14/777,410 patent/US11434494B2/en active Active
- 2014-03-14 IL IL241145A patent/IL241145B/en unknown
- 2014-03-14 BR BR112015022848-8A patent/BR112015022848B1/pt active IP Right Grant
- 2014-03-14 EP EP14763072.7A patent/EP2966984B1/en active Active
- 2014-03-14 KR KR1020157026632A patent/KR102304487B1/ko active Active
- 2014-03-14 WO PCT/US2014/029621 patent/WO2014144987A2/en not_active Ceased
- 2014-03-14 HK HK16108753.2A patent/HK1220864A1/zh unknown
- 2014-03-14 MY MYPI2015703120A patent/MY191390A/en unknown
- 2014-03-14 UA UAA201508559A patent/UA121099C2/uk unknown
- 2014-03-14 HU HUE14763072A patent/HUE058351T2/hu unknown
- 2014-03-14 EA EA201591447A patent/EA201591447A1/ru unknown
- 2014-03-14 ES ES14763072T patent/ES2912400T3/es active Active
- 2014-03-14 HR HRP20220521TT patent/HRP20220521T1/hr unknown
- 2014-03-14 AU AU2014233465A patent/AU2014233465A1/en not_active Abandoned
- 2014-03-14 CN CN201480024708.9A patent/CN105338805A/zh active Pending
- 2014-03-14 JP JP2016503169A patent/JP2016516408A/ja active Pending
- 2014-03-14 DK DK14763072.7T patent/DK2966984T3/da active
- 2014-03-14 RS RS20220373A patent/RS63188B1/sr unknown
- 2014-03-14 PL PL14763072T patent/PL2966984T3/pl unknown
- 2014-03-14 LT LTEPPCT/US2014/029621T patent/LT2966984T/lt unknown
-
2015
- 2015-09-11 CL CL2015002650A patent/CL2015002650A1/es unknown
-
2020
- 2020-08-17 AU AU2020220059A patent/AU2020220059B2/en active Active
-
2022
- 2022-09-05 US US17/902,970 patent/US20230141977A1/en active Pending
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2912400T3 (es) | Modificación génica orientada mediante la reparación génica mediada por oligonucleótidos | |
| JP6947784B2 (ja) | オリゴヌクレオチド仲介型遺伝子修復を使用した標的遺伝子修飾の効率を高めるための方法および組成物 | |
| US20230407322A1 (en) | Methods and compositions for increasing efficiency of targeted gene modification using oligonucleotide-mediated gene repair | |
| ES2531882T3 (es) | EPSPS mutantes | |
| ES2974625T3 (es) | Procedimientos y composiciones para aumentar la eficacia de la modificación génica dirigida mediante reparación génica mediada por oligonucleótidos | |
| AU2014378946B2 (en) | Modified plants | |
| CN111819285B (zh) | 防碎基因和突变 | |
| CN121195071A (zh) | 乙酰辅酶a羧化酶突变体 | |
| BR122022013574B1 (pt) | Método para a introdução de uma mutação mediada por oligonucleobase de reparação de gene em uma sequência de ácido desoxirribonucleico alvo em um genoma de protoplasto vegetal | |
| NZ711146B2 (en) | Methods and Compositions for Increasing Efficiency of Targeted Gene Modification Using Oligonucleotide-Mediated Gene Repair | |
| BR112016021092B1 (pt) | Método para provocar uma alteração genética em uma célula vegetal | |
| NZ711145B2 (en) | Methods and compositions for increasing efficiency of targeted gene modification using oligonucleotide-mediated gene repair |