FR2851255A1 - Microorganisme a activite methionine synthase modifiee et procede de preparation de la methionine - Google Patents
Microorganisme a activite methionine synthase modifiee et procede de preparation de la methionine Download PDFInfo
- Publication number
- FR2851255A1 FR2851255A1 FR0305768A FR0305768A FR2851255A1 FR 2851255 A1 FR2851255 A1 FR 2851255A1 FR 0305768 A FR0305768 A FR 0305768A FR 0305768 A FR0305768 A FR 0305768A FR 2851255 A1 FR2851255 A1 FR 2851255A1
- Authority
- FR
- France
- Prior art keywords
- cystathionine
- strain
- methionine
- synthase activity
- enzyme
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
- ALUGUZNECOIQKE-UHFFFAOYSA-N CC(CCC(O)=N)=O Chemical compound CC(CCC(O)=N)=O ALUGUZNECOIQKE-UHFFFAOYSA-N 0.000 description 1
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1058—Directional evolution of libraries, e.g. evolution of libraries is achieved by mutagenesis and screening or selection of mixed population of organisms
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/20—Bacteria; Culture media therefor
- C12N1/205—Bacterial isolates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/01—Preparation of mutants without inserting foreign genetic material therein; Screening processes therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1085—Transferases (2.) transferring alkyl or aryl groups other than methyl groups (2.5)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/88—Lyases (4.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/12—Methionine; Cysteine; Cystine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12R—INDEXING SCHEME ASSOCIATED WITH SUBCLASSES C12C - C12Q, RELATING TO MICROORGANISMS
- C12R2001/00—Microorganisms ; Processes using microorganisms
- C12R2001/01—Bacteria or Actinomycetales ; using bacteria or Actinomycetales
- C12R2001/185—Escherichia
- C12R2001/19—Escherichia coli
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Bioinformatics & Computational Biology (AREA)
- Ecology (AREA)
- Crystallography & Structural Chemistry (AREA)
- Tropical Medicine & Parasitology (AREA)
- Virology (AREA)
- Enzymes And Modification Thereof (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
La présente invention concerne une souche de microorganisme produisant de l'acide 2-amino-4-(alkylmercapto)butyrique de formule générale (I)R-S-(CH2)2-CHNH2-COOH (I)dans laquelle R représente un radical alkyle linéaire ou ramifié comprenant de 1 à 18 atomes de carbones, éventuellement substitué par un ou groupe hydroxy, ou un radical aryle ou hétéroaryle comprenant un ou plusieurs atomes d'azote ou de soufre dans le cycle hétéroaromatique, sélectionné parmi les groupes phényle, pyridyle, pyrolyle, pyrazolyle, triazolyle, tetrazolyle, thiazolyle, ou thienyle,par métabolisme d'une source de carbone simple et d'une source de soufre comprenant un composé de formule générale (II) :R'-SH (II)dans laquelle R' représente un atome d'hydrogène ou R, R étant défini précédemment, et ses sels physiologiquement acceptables,ladite souche comprenant au moins une un gène codant pour une enzyme à activité «méthionine synthase» modifiée.L'invention concerne également un procédé de préparation d'acides aminés de formule générale (I).
Description
<Desc/Clms Page number 1>
Microorganisme à activité méthionine synthase modifiée et procédé de préparation de la méthionine
La présente invention se rapporte au domaine de la bioconversion et d'obtention d'acides aminés par fermentation de microorganismes. Elle se rapporte à une méthode de criblage et d'évolution dirigée permettant d'identifier une souche de microorganisme, éventuellement génétiquement modifié, possédant une enzyme à activité méthionine synthase modifiée, ladite souche produisant de l'acide 2Amino-4- (alkylmercapto)butyrique, en particulier de la L-méthionine (acide 2Amino-4- (méthylmercapto)butyrique). L'invention concerne également la souche de microorganisme, les enzymes améliorées et à leurs séquences codantes. L'invention concerne enfin un procédé de préparation de la méthionine par culture de ladite souche de microorganisme.
La présente invention se rapporte au domaine de la bioconversion et d'obtention d'acides aminés par fermentation de microorganismes. Elle se rapporte à une méthode de criblage et d'évolution dirigée permettant d'identifier une souche de microorganisme, éventuellement génétiquement modifié, possédant une enzyme à activité méthionine synthase modifiée, ladite souche produisant de l'acide 2Amino-4- (alkylmercapto)butyrique, en particulier de la L-méthionine (acide 2Amino-4- (méthylmercapto)butyrique). L'invention concerne également la souche de microorganisme, les enzymes améliorées et à leurs séquences codantes. L'invention concerne enfin un procédé de préparation de la méthionine par culture de ladite souche de microorganisme.
La DL-Methionine est produite industriellement par synthèse chimique. Le méthyl-mercaptan réagit avec l'acroléine pour produire le p-méthyl thiopropionaldéhyde, qui réagit avec l'hydrogène cyanide pour produire l'a-hydroxy- y-méthyl-thio-butyro-nitrile. Après traitement avec l'ammoniac, et une hydrolyse, on obtient la méthionine.
Tous les producteurs industriels de la DL-methionine utilisent les mêmes matières premières à savoir l'acroléine, le méthane thiol (méthyl-mercaptan), l'hydrogène cyanide et l' ammoniac ou l' ammonium carbonate. Le procédé de synthèse du mélange racémique peut être conduit en batch ou en continu.
Un procédé industriel combine à la synthèse chimique de la biocatalyse par l'utilisant l'amino acylase, enzyme produite par Aspergillus oryzas pour obtenir uniquement la L-méthionine à partir de la DL-méthionine.
Les brevets US 6,379,934 et EP 1 055 730 se rapportent à la production d'acides aminés en utilisant une souche de bactéries corynéformes, dans laquelle le gène accBC est amplifié. La méthionine est mentionnée, mais seule la préparation de L-lysine est exemplifiée.
Toutefois, la synthèse d'acides aminés soufrés culture de microorganisme reste difficile à mettre en #uvre à des niveaux susceptibles de conduire à une
<Desc/Clms Page number 2>
exploitation industrielle, notamment du fait de la complexité de leurs voies de biosynthèse et de nombreux mécanismes de régulation.
En effet, le métabolisme de la méthionine, est fortement régulé, la régulation de son métabolisme se faisant à plusieurs niveaux (Weissbach et al., 1991, Mol.
Microbiol., 5,1593-1597) : métabolisme carboné pour la fabrication de la L-sérine à partir du glycerate3P, de la L-homosérine à partir de l'aspartate et de l'acétyl-CoA métabolisme du soufre pour la fabrication de la L-cystéine à partir de la L- sérine, de l'acétyl-CoA et du sulfate du milieu de culture synthèse de la méthionine (Fig. 1) à partir de la L-homosérine, de la cystéine et d'acétyl-CoA ou succinyl-CoA.
La demande WO 93/17112 décrit les différentes enzymes impliquées dans la biosynthèse de la méthionine à partir de l'acide L-aspartique dans divers organismes.
Cette demande de brevet décrit également l'introduction dans un microorganisme de plusieurs gènes exogènes agissant vde manière séquentielle pour la synthèse de la méthionine, employant du mléthyl-mercaptan ou du sulfure d'hydrogène comme source de soufre.
La présente invention se rapporte à des souches de microorganismes en particulier des bactéries, notamment E. coli et les corynébactéries, produisant de l'acide 2-amino-4-(alkylmercapto)butyrique de formule générale (I) R-S-(CH2)2-CHNH2-COOH (I) dans laquelle R représente un radical alkyle linéaire ou ramifié comprenant de
1 à 18 atomes de carbones, éventuellement substitué par un ou des groupe (s) ou un radical aryle ou hétéroaryle comprenant un ou plusieurs atomes d'azote ou de soufre dans le cycle hétéroaromatique, sélectionné parmi les groupes phényle, pyridyle, pyrolyle, pyrazolyle, triazolyle, tetrazolyle, thiazolyle, ou thienyle, par métabolisme d'une source de carbone simple et d'une source de soufre comprenant un composé de formule générale (II) :
R'-SH (II) dans laquelle R' représente un atome d'hydrogène ou R, R étant défini précédemment, et ses sels physiologiquement acceptables,
1 à 18 atomes de carbones, éventuellement substitué par un ou des groupe (s) ou un radical aryle ou hétéroaryle comprenant un ou plusieurs atomes d'azote ou de soufre dans le cycle hétéroaromatique, sélectionné parmi les groupes phényle, pyridyle, pyrolyle, pyrazolyle, triazolyle, tetrazolyle, thiazolyle, ou thienyle, par métabolisme d'une source de carbone simple et d'une source de soufre comprenant un composé de formule générale (II) :
R'-SH (II) dans laquelle R' représente un atome d'hydrogène ou R, R étant défini précédemment, et ses sels physiologiquement acceptables,
<Desc/Clms Page number 3>
les dites souches présentant au moins une un gène codant pour une enzyme à activité méthionine synthase modifiée.
Par enzyme à activité méthionine synthase modifiée, on entend selon l'invention toute enzyme impliquée dans la biosynthèse des acides 2-amino-4- (alkylmercapto)butyriques de formule générale (I) par conversion d'un substrat dérivé de formule générale (III)
R''-O-(CH2)2-CHNH2-COOH (III) dans laquelle R" représente un radical acyle de préférence choisi parmi le radical succinyle ou le radical acétyle, soit par conversion directe du substrat en acide de formule générale (I), soit par conversion du substrat en homocystéine de formule générale (IV)
HS-(CH2)2-CHNH2-COOH (IV) laquelle est ensuite transformée en acide de formule générale (I) par une enzyme appropriée.
R''-O-(CH2)2-CHNH2-COOH (III) dans laquelle R" représente un radical acyle de préférence choisi parmi le radical succinyle ou le radical acétyle, soit par conversion directe du substrat en acide de formule générale (I), soit par conversion du substrat en homocystéine de formule générale (IV)
HS-(CH2)2-CHNH2-COOH (IV) laquelle est ensuite transformée en acide de formule générale (I) par une enzyme appropriée.
Les enzymes à activité méthionine synthase modifiée sont des enzymes modifiées par rapport aux enzymes natives pour leur permettre de réaliser de manière préférentielle la conversion directe du substrat de formule générale (III) en acide de formule générale (I) ou en homocystéine de formule générale (IV) au lieu de la réaction catalysée par l'enzyme native. Cette modification, appelée encore mutation, consiste essentiellement en ce que l'enzyme modifiée ait une plus grande affinité pour le composé soufré de formule générale (II) que pour son co-substrat naturel.
Par source de carbone simple, selon la présente invention, on entend des sources de carbone utilisables par l'homme du métier pour la croissance normale d'un microorganisme, d'une bactérie en particulier. On entend désigner notamment les différents sucres assimilables, tels le glucose, le galactose, le saccharose, le lactose ou les mélasses, ou les sous-produits de ces sucres. Une source de carbone simple tout particulièrement préférée est le glucose. Une autre source de carbone simple préférée est le saccharose.
Par sel physiologiquement acceptable, on entend selon l'invention les sels du composé de formule générale (I) qui n'affectent pas le métabolisme et la capacité de croissance de la souche de microorganisme selon l'invention, notamment les sels de métaux alcalins comme le sodium.
<Desc/Clms Page number 4>
Selon un mode préférentiel de réalisation de l'invention, R représente un radical alkyle linéaire ou ramifié comprenant de 1 à 4 atomes de carbone, en particulier choisi parmi les radicaux méthyle, éthyle, n-propyle, i-propyle, n-butyle, ibutyle ou t-butyle. De manière plus préférentielle, R représente le radical méthyle.
L'acide 2-amino-4- (alkylmercapto)butyrique formule générale (I) obtenu est de préférence la L-méthionine.
L'utilisation d'une source de carbone simple et d'un composé soufré de formule (II) pour la production d'acide de formule (I) par bioconversion présente a priori plusieurs avantages, notamment : la synthèse de l'acide (I), comme la méthionine, en une ou deux étapes à partie à partir d'O-acyl-L-homosérine, devient indépendante de la synthèse de la cystéine voire également du cycle du tétrahydrofolate ; les composés soufrés de formule (II), comme le méthyl-mercaptan, qui sont des matières premières généralement toxiques issues de la pétrochimie, peuvent être valorisés dans la synthèse d'acides aminés à haute valeur ajoutée.
L'invention est basée sur le fait que l'on peut obtenir, de façon dirigée, une modification de l'activité méthionine-synthase de la cystationine-y-synthase (EC 4.2.99.9 ; GenBank AAN83320, ou AAA24167) en présence de méthyl-mercaptan.
Cette enzyme de la voie de biosynthèse de méthionine, codée par le gène metB chez E. coli (Fig. 2) et C. glutamicum, présente une activité pour un large spectre de substrats (Flavin, M. ; C. (1967) Enzymatic synthesis of homoserine or méthionine directly from O-succinyl-homoserine.Biochim. Biophys. Acta 132 :
405).
405).
L'invention est aussi basée sur le fait que l'on peut obtenir, de façon dirigée, une modification de l'activité méthionine-synthase de l'O-acétyl-L-homosérine sulfhydrolase (ou O-acétyl-L-homosérine sulfhydrylase, C 4.2.99.10) en présence de méthylmercaptan. Cette enzyme de la voie de biosynthèse de méthionine, codée par le gène metY chez C. glutamicum (Genbank AF220150), présente une activité pour un large spectre de substrats (Smith IK, Thompson JF. (1969) Utilization of Smethylcysteine and methylmercaptan by methionineless mutants of Neurospora and the pathway of their conversion to méthionine. II. Enzyme studies. Biochim Biophys Actal84(l):130-8).
<Desc/Clms Page number 5>
L'invention concerne donc également un procédé de préparation des souches selon l'invention et' leur utilisation dans un procédé de préparation d'un acide 2amino-4- (alkylmercapto)butyrique de formule générale (I), de préférence de la Lméthionine.
Le procédé de préparation de souches selon l'invention consiste à obtenir, à partir d'une souche bactérienne initiale, une souche bactérienne génétiquement modifiée présentant au moins une modification dans un gène codant pour une enzyme à activité méthionine synthase , ledit procédé comprenant une étape consistant à soumettre ladite souche bactérienne initiale à une pression de sélection en présence du composé de formule (II) défini ci-dessus, afin de diriger une évolution dudit gène codant pour ladite enzyme à activité méthionine synthase dans ladite souche bactérienne, vers un gène codant pour une enzyme à activité méthionine synthase modifiée par rapport à ladite souche bactérienne initiale.
Une activité méthionine synthase est améliorée dans la souche (A) de microorganisme par rapport à la souche (I) initiale lorsque la production de méthionine dans les mêmes conditions de culture (dans un milieu contenant une quantité efficace de dérivé soufré de formule (II)) est supérieure pour la souche (A) que pour la souche (I). Cette amélioration est préférentiellement observée par étude de la quantité de méthionine produite. Dans certains cas, on peut observer cette amélioration par l'augmentation du taux de croissance de la bactérie (A) par rapport au taux de croissance de la bactérie (I), dans un milieu minimum ne contenant pas de méthionine.
La présente invention concerne également les souches à activité méthionine synthase améliorée susceptibles d'être obtenues par le procédé de sélection selon l'invention et comprenant au moins un gène codant pour une enzyme à activité méthionine synthase modifiée telle que définie précédemment et ci-après.
Selon un premier mode de réalisation de l'invention, l'enzyme à activité méthionine synthase modifiée permet la conversion directe du substrat de formule générale (III) en acide de formule générale (I). Dans ce cas, la source de soufre est un
<Desc/Clms Page number 6>
composé de formule générale (II) pour laquelle R' représente le radical R définit précédemment.
L'enzyme à activité méthionine synthase modifiée est choisie parmi les cystathionine-y-synthases et les acylhomosérine sulfhydrylases.
Selon un deuxième mode de réalisation de l'invention, l'enzyme à activité méthionine synthase modifiée permet la conversion du substrat de formule générale (III) en homocystéine de formule générale (IV)
HS-(CH2)2-CHNH2-COOH (IV) laquelle est ensuite transformée en acide de formule générale (I) par une enzyme appropriée. Dans ce cas, la source de source de soufre est un composé de formule générale (II) pour laquelle R' représente un atome d'hydrogène, préférentiellement du sulfure d'hydrogène. La source de soufre H2S peut-être introduite dans le milieu de culture ou bien être produite par la bactérie à partir d'une source de soufre simple, par exemples un sulfate.
HS-(CH2)2-CHNH2-COOH (IV) laquelle est ensuite transformée en acide de formule générale (I) par une enzyme appropriée. Dans ce cas, la source de source de soufre est un composé de formule générale (II) pour laquelle R' représente un atome d'hydrogène, préférentiellement du sulfure d'hydrogène. La source de soufre H2S peut-être introduite dans le milieu de culture ou bien être produite par la bactérie à partir d'une source de soufre simple, par exemples un sulfate.
L'enzyme à activité méthionine synthase modifiée est avantageusement choisie parmi les cystathionine-y-synthases et les acylhomosérine sulfhydrylases.
L'homme du métier saura sélectionner d'autres enzymes à activité méthionine synthase modifiée en fonction de leur capacitén à évoluer pour effectuer la réaction enzymatique méthionine synthase modifiée telle que définie précédemment. On citera notamment les cystéines synthases A et B, codées par les gènes cysK et cysM chez les bactéries.
Pour les cystathionine-y-synthases modifiées définies précédemment et cidessous, le substrat est avantageusement la O-acétyl-L-homosérine ou la O-succinylL-homosérine, de préférence la O-succinyl-L-homosérine.
Pour les acylhomosérine sulfhydrylases modifiées définies précédemment et ci-dessous, le substrat est avantageusement la O-succinyl-L-homosérine ou la 0acétyl-L-homosérine, de préférence la O-acétyl-L-homosérine.
Pour ces deux enzymes, la modification consiste essentiellement en une mutation de manière à ce que la transformation du substrat de formule générale (III) se fasse de manière préférentielle avec le composé de formule générale (II) plutôt qu'avec la L-cystéine.
La mutation des enzymes peut être obtenue par la mise en #uvre du procédé de préparation de souches à activité méthionine synthase améliorée selon
<Desc/Clms Page number 7>
l'invention par culture sous pression de sélection en présence du composé de formule générale (II).
Dans ce cas, le gène comprenant la séquence codant pour la cystathionine-ysynthase ou l'acylhomosérine sulfhydrylase est # soit un gène natif, présent dans le génome de la souche initiale (I) où il est exprimé pour permettre la traduction de l'enzyme correspondante, # soit une un gène hétérologue comprenant une séquence codant pour une cystathionine-y-synthase ou une acylhomosérine sulfhydrylase, sous le contrôle d'éléments de régulation permettant son expression et sa traduction dans la souche initiale (I) où il aura été introduit.
La mutation peut également être obtenue par mutagénèse dirigée, # soit directement sur le gène natif présent naturellement dans la souche initiale (I), notamment par recombinaison homologue, # soit par des techniques usuelles de mutagenèse dirigée sur des séquences codant pour une cystathionine-y-synthase ou une acylhomosérine sulfhydrylase, introduite ensuite dans la souche initiale (I) sous le contrôle d'éléments de régulation permettant son expression et sa traduction dans ladite souche initiale (I) où il aura été introduit.
De tels éléments de régulation sont bien connus de l'homme du métier, et comprennent des séquences de régulation promotrice, ou promoteurs, en particulier des promoteurs dits promoteurs forts constitutifs chez les microorganismes. De préférence, le promoteur fort constitutif est choisi parmi pTAC-0 (SEQ ID N 1), pLAC-0 (SEQ ID N 2), pTRC-0 (SEQ ID N 3), pTHLA (SEQ ID N 4), promoteurs forts pour lesquels l'opérateur lac a été délété pour les rendre constitutifs.
De manière avantageuse, les cystathionine-y-synthases initiales ou à activité méthionine synthase non modifiée sont sélectionnées parmi les cystathionine-y-synthases correspondent au PFAM référence PF01053 et au COG référence CPG0626.
Les PFAM (Protein families database of alignments and Hidden Markov

Models ; http://www.sanger.ac.uk/Software/Pfam[) représentent une large collection
Models ; http://www.sanger.ac.uk/Software/Pfam[) représentent une large collection
<Desc/Clms Page number 8>
d'alignements de séquences protéiques. Chaque PFAM permet de visualiser des alignements multiples, de voir des domaines protéiques, d'évaluer la répartition entre les organismes, d'avoir accès à d'autres bases de données, de visualiser des structures connues de protéines.
Les COGs (Clusters of Orthologous Groups of proteins ; http://www.ncbi.nlm.nih.gov/COG/) sont obtenus en comparant les séquences protéiques issus de 43 génomes complètement séquences représentant 30 lignées phylogénétiques majeurs. Chaque COG est défini à partir d'au moins trois lignées ce qui permet ainsi d'identifier des domaines conservés anciens.
De manière préférentielle, la cystathionine-y-synthase est la cystathionine-ysynthase de E. coli K12, représentée sur la SEQ ID NO: 6, et les séquences homologues de cette séquence présentant une activité cystathionine-y-synthase et comprenant au moins 80% d'homologie, préférentiellement 90% d'homologie, plus préférentiellement 95% d'homologie avec la séquence d'acides aminés de la SEQ ID NO 6.
Les moyens d'identification des séquences homologues et de leur pourcentages d'homologie sont bien connus de l'homme du métier, comprenant notamment le programme BLAST, et notamment les programme BLASTP, qui peut être utilisé à partir du site http://www.ncbi.nlm.nih.gov/BLAST/ avec les paramètres indiqués par défaut sur ce site.
Selon un mode préférentiel de réalisation de l'invention, la cystathionine-y- synthase initiale, avant modification, comprend une séquence d'acide aminés cidessous dans sa partie C-terminale (zone conservée 1) X1-X2-X3-L-G-X4-X5-X6-X7-X8-X9 dans laquelle
XI représente A,G,S, de préférence A
X2 représente E, V,P,T, de préférence E
X3 représente S, T,N, de préférence S
X4 représente G,D,A,H,T, de préférence G
X5 représente V,A,T,H,N, de préférence V
X6 représente E,R,K,F, de préférence E
X7 représente S, T, de préférence S
X8 représente L,I,V,A, de préférence L et
XI représente A,G,S, de préférence A
X2 représente E, V,P,T, de préférence E
X3 représente S, T,N, de préférence S
X4 représente G,D,A,H,T, de préférence G
X5 représente V,A,T,H,N, de préférence V
X6 représente E,R,K,F, de préférence E
X7 représente S, T, de préférence S
X8 représente L,I,V,A, de préférence L et
<Desc/Clms Page number 9>
X9 représente I,V,A,T, de préférence I.
De manière préférentielle, la cystathionine-y-synthase à activité méthionine synthase modifiée, selon l'invention comprend la séquence d'acides aminés suivante dans sa partie C-terminale :
A-E-S-L-G-G-V-E-S
De manière avantageuse, la cystathionine y-synthase initiale, avant nodification, comprend également au moins une séquence d'acide aminés ci-dessous ians sa partie N-terminale (zone conservée 2) : X10-X11-Y-X12-R-X13-X14-X15-X16-X17-X18 dans laquelle
X10 représente H, Y,F,L,K, de préférence A
XI 1 représente E, D,K,R,V,I, de préférence Y
X12 représente S, A,T,P,G, de préférence S
XI 3 représente I,S,T,R,E,F,W,D, de préférence S
X14 représente S,G,A,I,E,N,K,P, de préférence G
XI 5 représente N, H,Q,S, de préférence N
X16 représente P, D,L, de préférence P XI 7 représente T, M,N,G,S, de préférence T et
XI 8 représente R, L,V,S,W,E, de préférence R.
A-E-S-L-G-G-V-E-S
De manière avantageuse, la cystathionine y-synthase initiale, avant nodification, comprend également au moins une séquence d'acide aminés ci-dessous ians sa partie N-terminale (zone conservée 2) : X10-X11-Y-X12-R-X13-X14-X15-X16-X17-X18 dans laquelle
X10 représente H, Y,F,L,K, de préférence A
XI 1 représente E, D,K,R,V,I, de préférence Y
X12 représente S, A,T,P,G, de préférence S
XI 3 représente I,S,T,R,E,F,W,D, de préférence S
X14 représente S,G,A,I,E,N,K,P, de préférence G
XI 5 représente N, H,Q,S, de préférence N
X16 représente P, D,L, de préférence P XI 7 représente T, M,N,G,S, de préférence T et
XI 8 représente R, L,V,S,W,E, de préférence R.
Les acides aminés XI à X9 correspondent aux résidus 324 à 334 de la séquence de cystathionine y-synthase de E. coli K12, représenté sur la SEQ ID NO 6.
Les acides aminés X10 à X18 correspondent aux résidus 44 à 54 de la séquence de cystathionine y-synthase de E. coli K12, représenté sur la SEQ ID NO 6.
Les positions des acides aminés employées ci-dessus et ci-après sont entendues comme étant des positions relatives faites par référence à la séquence de cystathionine-y-synthase de E. coli K12. L'homme du métier saura bien entendu retrouver les acides aminés correspondants sur d'autres séquences de cystathionine-y- synthases, par l'emploi d'outils usuels d'alignement de séquences. On citera notamment le programme BLAST, qui peut être utilisé à partir du site http://www.ncbi.nlm.nih.gov/BLAST/ avec les paramètres indiqués par défaut sur ce site. L'alignement de séquence pourra être effectué tant au niveau de la protéine (SEQ
<Desc/Clms Page number 10>
ID NO 6) que de sa séquence codante, comme par exemple la séquence codante représentée sur la SEQ ID NO 5.
On peut aussi avantageusement utiliser la recherche avancée de BlastP en affinant la recherche avec un motif (PHI-BLAST). Dans ce cas on pourra prendre le motif [AGS]-[EVPT]-[STN]-L-G-[GDAHT]-[VATHN]-[ERKF]-[ST]-[LIVA]- [IVAT] pour une première zone conservée et le motif x(2)-Y-[SATPG]-R-x(2)- [NHQS]-[PDL]-[TMNGS]-[ RLVSWE] pour une deuxième zone conservée où les lettres indiquées en gras correspondent aux acides aminés majoritairement présent à cette position dans la séquence, et où x correspond à n'importe quel acide aminé. Le chiffre 2 entre parenthèse signifie qu'il y a deux acides aminés indéterminés.
Pour l'alignement des séquences, on peut utiliser les programmes CLUSTALW (http://www.ebi.ac.uk/clustalw/) ou MULTALIN

((http://prodes.toulouse.inra.fr/multali Ici-bin/multalin.pl), avec les paramètres indiqués par défaut sur ces sites.
((http://prodes.toulouse.inra.fr/multali Ici-bin/multalin.pl), avec les paramètres indiqués par défaut sur ces sites.
Un tel alignement de séquences est représenté sur la figure 5 pour une sélection de différentes cystathionine-y-synthases.
Elles sont de préférence choisies parmi les cystathionine-y-synthase (CGS) suivantes : Q9ZMW7 O-succinylhomoserine (Thiol) -lyase, Helicobacter pylori P46807 O-succinylhomoserine (Thiol) -lyase, Mycobacterium leprae AA029646 Xylella fastidiosa Temeculal NP~638204 Xanthomonas campestris pv. campestris str. ATCC 33913 NP~358970 Streptococcus pneumoniae R6 NP~126586 O-succinylhomoserine (Thiol) -lyase, Pyrococcus abyssi NP-373671 Staphylococcus aureus subsp. aureus N315 NP~418374 [Escherichia coli K12 NP~601979 Corynebacterium glutamicum ATCC 13032 NP~343729 O-succinylhomoserine (thiol)-lyase, Sulfolobus solfataricus NP-786043 O-succinylhomoserine (thiol) -lyase, Lactobacillus plantarum WCFS1 NP 719586 Shewanella oneidensis MR-1 CAD30944 Streptomyces coelicolor A3(2) NP~696324 Bifidobacterium longum NCC2705 NP~457953 Salmonella enterica subsp. enterica serovar Typhi
<Desc/Clms Page number 11>
NP 539021 Brucella melitensis EAA30199 O-succinylhomoserine (Thiol) -lyase, Neurospora crassa BAC61028 Vibrio parahaemolyticus.
De préférence, la cystathionine-y-synthase modifiée telle que définie précédemment, comprend au moins une mutation dans sa partie C-terminale, et/ou au moins une mutation dans sa partie N-terminale.
Par mutation, on entend selon l'invention la substitution d'un acide aminé de la séquence native par un acide aminé différent.
De préférence, la mutation consiste à remplacer un acide aminé acide, lequel interagit avec le co-substrat cystéine pour l'enzyme non modifiée, par un acide aminé apolaire, sélectionné parmi les résidus glycine, alanine, leucine, isoleucine, valine, phénylalanine ou méthionine.
Ces acides aminés peuvent être identifiés par référence à la structure cristalline de la cystathionine-y-synthases de E. coli, décrite par Clausen & al.
(EMBOJ, Vol. 17, No. 23, pp 6827-6838, 1998).
De manière avantageuse, la mutation dans la partie C-terminale est introduite parmi les acides aminés acides de la zone conservée 1 telle que définie ci-dessus, en particulier au niveau du résidu X2.
La cystathionine-y-synthase à activité méthionine synthase modifiée selon l'invention comprend avantageusement la séquence d'acides aminés suivante dans sa partie C-terminale : X1-X2-X3-L-G-X4-X5-X6-X7-X8-X9 dans laquelle
X1, X3, X4, X5, X6, X7, X8 et X9 sont définis ci-dessus, et
X2 représente G, A,L,I,V,F,M, de préférence A.
X1, X3, X4, X5, X6, X7, X8 et X9 sont définis ci-dessus, et
X2 représente G, A,L,I,V,F,M, de préférence A.
De manière avantageuse, la mutation dans la partie N-terminale est introduite parmi les acides aminés acides de la zone conservée 2 telle que définie ci-dessus, en particulier au niveau du résidu X11 et/ou R48 et/ou X13.
De manière préférentielle, la cystathionine-y-synthase à activité méthionine synthase modifiée, selon l'invention comprend la séquence d'acides aminés suivante dans sa partie C-terminale :
A-A-S-L-G-G-V-E-S
A-A-S-L-G-G-V-E-S
<Desc/Clms Page number 12>
La cystathionine-y-synthase à activité méthionine synthase modifiée selon l'invention peut comprendre avantageusement la séquence d'acide aminés suivante dans sa partie N-terminale : X10-X11-Y-X12-X19-X13-X14-X15-X16-X17-X18 dans laquelle
X7, X9, X12, X14, X15, X16, X17 et X18 sont définis ci-dessus,
XI 1 est défini ci-dessus ou représente un acide aminé apolaire,
XI 3 est défini ci-dessus ou représente un acide aminé apolaire,
XI 9 est défini ci-dessus ou représente un acide aminé apolaire, et l'un au moins de Xl l, X13 et X19 représente un acide aminé apolaire tel que défini précédemment.
X7, X9, X12, X14, X15, X16, X17 et X18 sont définis ci-dessus,
XI 1 est défini ci-dessus ou représente un acide aminé apolaire,
XI 3 est défini ci-dessus ou représente un acide aminé apolaire,
XI 9 est défini ci-dessus ou représente un acide aminé apolaire, et l'un au moins de Xl l, X13 et X19 représente un acide aminé apolaire tel que défini précédemment.
Selon un mode préférentiel de réalisation de l'invention, la cystathionine-ysynthase à activité méthionine synthase modifiée est une cystathionine-y-synthase telle que définie précédemment, modifiée pour permettre la conversion directe de la O-succinyl-L-homosérine en L-methionine avec le méthyl-mercaptan comme source de soufre (composé de formule générale (II) dans lasquelle R représente le méthyle).
Selon un mode de réalisation préférentiel de l'invention, la cystathionine-ysynthase à activité méthionine synthase modifiée comprend la séquence d'acides aminés représentée sur la SEQ ID NO 8. Une séquence d'ADN codant pour cette enzyme modifiée est représentée sur la SEQ ID NO 7.
La présente invention concerne également les cystathionine-y-synthases modifiées telles que définies ci-dessus ainsi que les séquences d'acide nucléique codant pour ces cystathionine-y-synthases modifiées, notamment les séquences isolées, en particulier les séquences d'ADN et notamment la séquence représentée sur la SEQ ID NO 7, les vecteur de clonage et/ou d'expression comprenant les dites séquences, en particulier les vecteurs comprenant lesdites séquences d'acide nucléique sous le contrôle d'éléments de régulation nécessaires à l'expression et la transcription de la cystathionine-y-synthase modifiée dans un organisme hôte et les organismes hôtes transformés avec lesdits vecteurs.
<Desc/Clms Page number 13>
De manière avantageuse, les acylhomosérine sulfhydrylases initiales ou à activité méthionine synthase non modifiée sont chosies parmi les acylhomosérine sulfhydrylases correspondant au PFAM référence PF01053 et au COG référence COG2873.
Elles sont de préférence choisies parmi les acylhomosérine sulfhydrylases suivantes : NP~785969 O-acetylhomoserine (thiol) -lyase, Lactobacillus plantarum WCFS AAN68137 O-acetylhomoserine sulfhydrylase, Pseudomonas putida KT2440 NP~599886 O-acetylhomoserine sulfhydrylase, Corynebacterium glutamicum ATCC
13032 NP-712243 acetylhomoserine sulfhydrylase, Leptospira interrogans serovar lai str.
13032 NP-712243 acetylhomoserine sulfhydrylase, Leptospira interrogans serovar lai str.
56601 BAC46370 O-succinylhomoserine sulfhydrylase, Bradyrhizobium japonicum
USDA110 AA057279 O-succinylhomoserine sulfhydrylase, Pseudomonas syringae pv. tomato str. DC3000 NP-284520 O-succinylhomoserine sulfhydrolase [Neisseria meningitidis Z2491 AAA83435 O-succinylhomoserine sulfhydrylase (P. aeruginosa)
Selon un mode préférentiel de réalisation de l'invention, la souche à activité méthionine synthase améliorée comprend une inactivation d'au moins un gène endogène impliqué dans la voie de biosynthèse habituelle de la méthionine.
USDA110 AA057279 O-succinylhomoserine sulfhydrylase, Pseudomonas syringae pv. tomato str. DC3000 NP-284520 O-succinylhomoserine sulfhydrolase [Neisseria meningitidis Z2491 AAA83435 O-succinylhomoserine sulfhydrylase (P. aeruginosa)
Selon un mode préférentiel de réalisation de l'invention, la souche à activité méthionine synthase améliorée comprend une inactivation d'au moins un gène endogène impliqué dans la voie de biosynthèse habituelle de la méthionine.
Ceci permet de sélectionner les souches qui ont développé le métabolisme alternatif selon l'invention pour la production de méthionine. Il est à noter que l'on obtient alors des souches auxotrophes pour la méthionine (e.g. metE), qui survivent en raison de leur possibilité à produire cet acide aminé par une voie alternative. Il peut être nécessaire, dans le test de criblage selon l'invention, que de la méthionine soit présente initialement dans le milieu de culture afin de permettre une première croissance des microorganismes.
Dans un mode de réalisation préféré, la souche bactérienne comprend une inactivation d'au moins un gène endogène choisi parmi metB, metJ, metC, metE, metH.
<Desc/Clms Page number 14>
Une mutation du gène metJ a été proposée dans JP 2000157267-A/3, pour produire une quantité supérieure de méthionine (voir aussi GenBank E35587). Ce gène code pour une protéine de répression des gènes met B, E, L, J et R (chez Salmonella typhimurium). Son inactivation ou sa modification permet de diminuer le rétrocontrôle par la méthionine.
Le gène metC (GenBank M12858), code la cystathionine-ss-lyase (EC 4. 4.1.8), les gènes metE (GenBank AE000458) et metH (GenBank J04975) codent la méthionine synthase (EC 2. 1.1.13). La méthionine est un acide aminé essentiel à la vie cellulaire. L'inactivation d'un ou plusieurs de ces gènes revient à supprimer la voie habituelle de biosynthèse de la méthionine.
En utilisant les références données sur GenBank pour ces gènes qui sont bien connus, l'homme du métier est capable de déterminer les gènes équivalents dans d'autres souches bactériennes qu'E. coli. Ce travail de routine est avantageusement effectué en utilisant les séquences consensus pouvant être déterminées du fait de la synthèse de ces gènes pour d'autres microorganismes, et en dessinant des sondes dégénérées permettant de cloner le gène correspondant dans un autre organisme. Ces techniques de routine de biologie moléculaire sont bien connues dans l'art et sont décrites par exemple dans Sambrook et al. (1989 Molecular cloning: a laboratory manual. 2nd Ed. Cold Spring Harbor Lab., Cold Spring Harbor, New York.).
La souche à activité méthionine synthase modifiée selon l'invention comprenant une enzyme à activité méthionine synthase modifiée définie ci-dessus comprend de préférence au moins une inactivation du gène metE et/ou metH, et/ou du gène metC, et/ou du gène met B.
Lorsque l'enzyme à activité méthionine synthase modifiée définie cidessus permet d'effectuer la conversion directe du substrat de formule générale (III) en acide de formule générale (I), la souche selon l'invention comprend avantageusement au mois une inactivation du gène metE et/ou metH et/ou metB, de préférence au moins une inactivation du gène metE.
Lorsque l'enzyme à activité méthionine synthase modifiée définie cidessus permet d'effectuer la conversion du substrat de formule générale (III) en homocystéine de formule générale (IV)
HS-(CH2)2-CHNH2-COOH (IV)
HS-(CH2)2-CHNH2-COOH (IV)
<Desc/Clms Page number 15>
laquelle est ensuite transformée en acide de formule générale (I) par une enzyme appropriée, la souche selon l'invention comprend au moins une inactivation du gène metC et/ou metB. Elle peut également comprendre une inactivation du gène metE et/ou metH endogène. Dans ce cas, l'activité méthylase associée aux gènes metE et/ou metH est restaurée par l'introduction d'un gène codant pour une enzyme ayant la même activité. Cette enzyme peut avoir été sélectionnée et/ou modifiée pour permettre une amélioration des rendements de synthèse des acides aminés de formule générale (I).
Dans un mode de réalisation, ladite souche bactérienne peut comprendre également une modification de l'activité homosérine O-acyltransférase porté par le gène metA afin de lui conférer au choix une activité homosérine 0succinyltransférase (EC 2. 3.1.46) ou homosérine O-acétyltransférase (EC 2.3.1.11).
Dans un mode particulier, on pourra remplacer ou modifier le gène metA de E. coli, codant l'enzyme possédant l'activité homosérine O-succinyltransférase (Genbank AAN83396), afin d'obtenir une activité homosérine O-acétyltransférase. Il est connu de l'homme du métier que cette activité est codée par le gène metA de C. glutamicum (Genbank AF052652). Les protocoles permettant de remplacer le gène metA de E. coli par le gène metA de C. glutamicum, ou de modifier la séquence de metA de E. coli afin d'obtenir une activité homosérine O-acetyltransferase au lieu d'une activité homosérine 0-succinyltransférase sont connus de l'homme du métier.
De manière similaire on peut remplacer ou modifier le gène metA de C. glutamicum, codant une activité homosérine O-acetyltransférase, afin d'obtenir une activité homosérine O-succinyltransférase.
Toutes les modifications mentionnées ci-dessus peuvent être effectuées directement sur la souche objet de la pression de sélection, lorsque le procédé selon l'invention est mis en oeuvre. Alternativement, il est préférable de mettre en #uvre le procédé de criblage selon l'invention sur une souche ne présentant qu'un nombre restreint de modifications, d'obtenir une souche présentant une activité méthionine synthase en présence du composé de formule (II), en particulier de méthylmercaptan, et d'effectuer alors d'autres modifications telles que mentionnées, afin d'augmenter le 'bypass' de la voie classique de synthèse de la méthionine.
<Desc/Clms Page number 16>
L'homme du métier connaît les protocoles permettant de modifier le caractère génétique de microorganismes. La surexpression d'un gène peut être effectuée par changement du promoteur de ce gène in situ, par un promoteur fort ou inductible. De façon alternative, on introduit, dans la cellule, un plasmide réplicatif (simple ou multicopies) dans lequel le gène que l'on désire surexprimer est sous le contrôle du promoteur adéquat.
L'inactivation d'un gène se fait préférentiellement par recombinaison homologue. Le principe d'un protocole en est rappelé brièvement : on introduit dans la cellule un fragment linéaire, obtenu in vitro, comprenant les deux régions flanquant le gène, et au moins un gène de sélection entre ces deux régions (généralement un gène de résistance à un antibiotique), ledit fragment linéaire présentant donc un gène inactivé. On sélectionne les cellules ayant subi un événement de recombinaison et ayant intégré le fragment introduit par étalement sur milieu sélectif. On sélectionne ensuite les cellules ayant subi un événement de double recombinaison, dans lesquelles le gène natif a été remplacé par le gène inactivé. Ce protocole peut être amélioré en utilisant des systèmes de sélections positive et négative, afin d'accélérer la détection des événements de double recombinaison.
Dans un mode de réalisation préféré, ladite souche bactérienne est une souche d'E. coli.
Dans un autre mode de réalisation, ladite souche bactérienne est une souche de Corynebacterium, en particulier C. glutamicum.
Selon un mode préférentiel de réalisation de l'invention, la souche bactérienne est la souche E. coli K183, déposée à la CNCM le 2 avril 2003 sous le numéro I- 3005.Cette souche comprend un gène exprimant une cystathionine-y-synthase modifiée, l'enzyme comprenant la mutation E325A, décrite précédemment et une inactivation du gène metE.
Les souches de microorganismes selon l'invention possèdent une enzyme cystathionine-y-synthase et/ou l'acylhomosérine sulfhydrylase ; elles sont de préférence sélectionnées et améliorées par un procédé de criblage et d'évolution, qui est aussi un objet de l'invention. Les souches selon l'invention peuvent également être génétiquement modifiées (c'est-à-dire présenter une inactivation, une mutation
<Desc/Clms Page number 17>
et/ou la suractivation d'au moins un gène endogène), la modification étant effectuée préalablement ou non à la mise en #uvre du procédé de criblage.
Afin d'accélérer la sélection et l'évolution dirigée des souches pour la production de méthionine en présence de composé de formule (II), en particulier de méthyl-mercaptan, on peut effectuer les opérations ci-dessous. Le procédé est décrit pour le méthyl-mercaptan. Toutefois, l'homme du métier saura l'adapter avec tout autre composé de formule (II), en particulier l'H2S. a. Coupler la biosynthèse de la molécule d'intérêt à la croissance du microorganisme de telle sorte que la production de cette molécule est nécessaire pour une bonne croissance du microorganisme
Pour cette raison on peut faire le choix de détruire le gène metE codant la méthionine-synthase qui produit la méthionine à partir d'homocystéine. Ce faisant la souche devient auxotrophe pour la méthionine.
Pour cette raison on peut faire le choix de détruire le gène metE codant la méthionine-synthase qui produit la méthionine à partir d'homocystéine. Ce faisant la souche devient auxotrophe pour la méthionine.
Le microorganisme, pour vivre en milieu minimum contenant une source de carbone simple et du méthylmercaptan ou méthylmercaptide de sodium, doit donc optimiser la voie de synthèse de la L-méthionine à partir de l'O-acyl-L-homosérine et du méthylmercaptan ou méthylmercaptide de sodium. Une modélisation informatique montre que dans ces conditions il est possible de doubler les rendements théoriques en méthionine (Tableau 1).
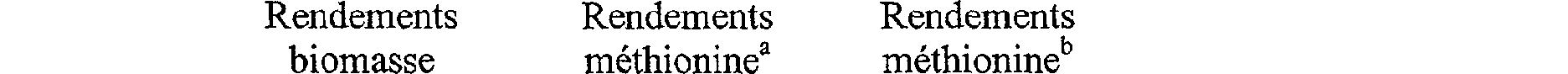
<tb>
<tb>
<tb>
Rendements <SEP> Rendements <SEP> Rendements
<tb> biomasse <SEP> méthionine' <SEP> méthionine
<tb>
<tb> biomasse <SEP> méthionine' <SEP> méthionine
<tb>
yy./s (g.g-1) y PIS (g.i1) Yp/sCg-g'1)

<tb>
<tb> 0 <SEP> 0,36 <SEP> 0,74
<tb> 0,11 <SEP> 0,30 <SEP> 0,62
<tb> 0,28 <SEP> 0,21 <SEP> 0,42
<tb> 0,44 <SEP> 0,12 <SEP> 0,24
<tb> 0,61 <SEP> 0 <SEP> 0 <SEP>
<tb>
Tableau 1 : rendements théoriques maximums pour la production de methionine (g de produit/g de glucose) par E. coli dans le cas d'une fermentation sur glucose (a) et d'une fermentation sur glucose et méthyl-mercaptan (b) avec un rendement en biomasse constant (cultures continues).
<tb> 0 <SEP> 0,36 <SEP> 0,74
<tb> 0,11 <SEP> 0,30 <SEP> 0,62
<tb> 0,28 <SEP> 0,21 <SEP> 0,42
<tb> 0,44 <SEP> 0,12 <SEP> 0,24
<tb> 0,61 <SEP> 0 <SEP> 0 <SEP>
<tb>
Tableau 1 : rendements théoriques maximums pour la production de methionine (g de produit/g de glucose) par E. coli dans le cas d'une fermentation sur glucose (a) et d'une fermentation sur glucose et méthyl-mercaptan (b) avec un rendement en biomasse constant (cultures continues).
Cependant, lorsque l'on souhaite produire un acide 2-amino-4- (alkylmercapto)butyrique différent de la L-méthionine, il est nécessaire de supplémenter le milieu en méthionine pour permettre la croissance du microorganisme.
<Desc/Clms Page number 18>
b. Supprimer les régulations, notamment les rétro-inhibitions soit au niveau des enzymes, soit au niveau des gènes afin que la voie de biosynthèse principale soit potentialisée
On peut ainsi supprimer le gène metJ codant une protéine répresseur. Par ailleurs, il a été montré que l'homosérine trans-succinylase, codée par le gène metA, était rétro-inhibée par la méthionine et la S-adénosylméthionine (Taylor et al., 1966, J. Biol. Chem., 241 : Il est donc souhaitable de remplacer cette enzyme par une enzyme insensible à la rétro-inhibition (Chater et al, 1970, J. Gen. Microbiol.
On peut ainsi supprimer le gène metJ codant une protéine répresseur. Par ailleurs, il a été montré que l'homosérine trans-succinylase, codée par le gène metA, était rétro-inhibée par la méthionine et la S-adénosylméthionine (Taylor et al., 1966, J. Biol. Chem., 241 : Il est donc souhaitable de remplacer cette enzyme par une enzyme insensible à la rétro-inhibition (Chater et al, 1970, J. Gen. Microbiol.
63 : 121-131).
Un autre objet de l'invention est un test de criblage-identification permettant d'obtenir un microorganisme produisant de l'acide 2-amino-4- (alkylmercapto)butyrique, notamment la L-méthionine, en métabolisant un composé soufré de formule générale (II), en particulier un alkyl-mercaptan ou l'H2S, notamment le méthyl-mercaptan.
Ainsi, la présente invention permet d'identifier des souches présentant des mutations dans leur génome, lesdites mutations permettant l'assimilation du composé de formule (II) par ladite souche, et la production dudit acide aminé de formule (I).
Lesdites modifications induisent donc une modification/augmentation de l'activité méthionine synthase de ladite souche. On peut alors accélérer la production de la souche produisant de la méthionine de façon autonome à partir d'une source de carbone simple et de composé de formule (II).
Un autre objet de l'invention est donc un procédé de criblage d'une souche bactérienne initiale, éventuellement génétiquement modifiée, possédant un gène codant pour une enzyme cystathionine-y-synthase ou acylhomosérine sulfhydrylase, en vue d'obtenir une souche bactérienne génétiquement modifiée produisant un acide aminé de formule (I), en particulier de la L-méthionine, et présentant une modification dans le gène de ladite enzyme, induisant une modification de l'activité méthionine synthase en présence d'un composé soufré de formule (II), présentant l'étape consistant à soumettre ladite souche bactérienne à une pression de sélection en présence du composé de formule (II), afin de diriger une évolution du gène codant pour ladite enzyme cystathionine-y-synthase ou acylhomosérine sulfhydrylase, dans ladite souche bactérienne, ladite évolution consistant en une mutation pour leur
<Desc/Clms Page number 19>
permettre de réaliser de manière préférentielle la conversion directe du substrat de formule générale (III) en acide aminé de formule générale (I) ou en homocystéine de formule générale (IV).
Ladite souche bactérienne initiale présente éventuellement une inactivation et/ou une suractivation, notamment par insertion d'un promoteur fort constitutif, d'au moins un gène endogène.
L'invention se rapporte également à une souche bactérienne présentant une modification dans le gène de l'enzyme cystathionine-y-synthase et/ou dans le gène de l'enzyme acylhomosérine sulfhydrylase, induisant une augmentation de l'activité méthionine synthase de ladite de ladite enzyme en présence du composé de formule (II), en particulier de méthyl-mercaptan. Une telle souche peut également présenter au moins une autre modification génétique, (inactivation, mutation ou surexpression d'un gène endogène), telle que mentionnée ci-dessus.
La souche selon l'invention est de préférence susceptible d'être obtenue par un procédé selon l'invention, et en particulier est obtenue par le procédé selon l'invention.
L'invention se rapporte également à un procédé de préparation d'un acide 2amino-4 (alkylmercapto)butyrique de formule (1) définie ci-dessus, dans lequel on cultive un microorganisme à activité méthionine synthase modifiée telle que défini précédemment, en présence d'un composé soufré de formule générale (II) dans un milieu approprié, ledit milieu approprié comprenant une source de carbone simple telle que définie précédemment. De préférence, ledit acide aminé de formule (I) est la méthionine, plus préférentiellement L-méthionine, et ledit composé soufré de formule (II) est le méthyl-mercaptan ou l'H2S.
Selon l'invention, les termes culture et fermentation sont employés indifféremment pour désigner la croissance de la bactérie sur un milieu de culture approprié comprenant une source de carbone simple.
La définition des conditions de culture des microorganismes selon l'invention (fermentation) est à la portée de l'homme du métier. On fermente notamment les bactéries à une température comprise entre 20 C et 55 C, de préférence entre 25 C et
<Desc/Clms Page number 20>
40 C, plus particulièrement d'environ 30 C pour C. glutamicum et d'environ 37 C pour E. coli.
La fermentation est généralement conduite en fermenteurs avec un milieu minéral de culture de composition connu défini et adapté en fonction des bactéries utilisées, contenant au moins une source de carbone simple ainsi que du composé soufré de formule (II).
En particulier, le milieu minéral de culture pour E. coli pourra ainsi être de composition identique ou similaire à un milieu M9 (Anderson, 1946, Proc. Natl.
Acad. Sci. USA 32 :120-128), un milieu M63 (Miller, 1992 ; A Short Course in Bacterial Genetics : Laboratory Manual and Handbook for Escherichia coli and Related Bacteria, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York) ou un milieu tel que défini par Schaefer et al. (1999, Anal. Biochem. 270 : 88- 96).
De manière analogue, le milieu minéral de culture pour C. glutamicum pourra ainsi être de composition identique ou similaire au milieu BMCG (Liebl et al., 1989, Appl. Microbiol. Biotechnol. 32: 205-210) à un milieu tel que défini par Riedel et al.
(2001, J. Mol. Microbiol. Biotechnol. 3 : 573-583).
Les milieux peuvent être supplémentés pour compenser les auxotrophies ; contiennent une concentration en carbone simple adaptée en fonction du mode de culture et de production, ainsi que du composé soufré de formule (II) en concentration adaptée en fonction de l'évolution de la souche et du mode de production retenu.
Après fermentation, l'acide aminé de formule (I) est récupéré selon les méthodes usuelles, et le cas échéant, purifié.
Les techniques de récupération puis de purification des acides aminés de formule (I) dans les milieux de culture sont bien connues de l'homme du métier.
La présente invention concerne également un rocédé de préparation d'un acide
2-amino-4(alkylmercapto)butyrique de formule (I) défini précédemment, dans lequel on fait réagir substrat dérivé de formule générale (III)
R''-O-(CH2)2-CHNH2-COOH (III) dans laquelle R" représente un radical acyle, de préférence choisi parmi le radical succinyle ou le radical acétyle,
2-amino-4(alkylmercapto)butyrique de formule (I) défini précédemment, dans lequel on fait réagir substrat dérivé de formule générale (III)
R''-O-(CH2)2-CHNH2-COOH (III) dans laquelle R" représente un radical acyle, de préférence choisi parmi le radical succinyle ou le radical acétyle,
<Desc/Clms Page number 21>
avec une enzyme à activité méthionine synthase modifiée telle que définie précédemment, dans un milieu réactionnel approprié comprenant un composé soufré de formule générale (II) défini précédemment.
Le milieu réactionnel approprié est un milieu usuel de réaction enzymatique, bien connu de l'homme du métier, en particulier un milieu aqueux dans lequel les substrats et l'enzyme sont en solution ou en suspension. Les conditions de mise en #uvre de la réaction sont bien connues de l'homme du métier, afin notamment d'éviter une dénaturation substantielle de l'enzyme.
Selon un mode particulier de réalisation de l'invention, l'enzyme à activité méthionine synthase modifiée est présente dans une bactérie inactivée ou dans un extrait cellulaire.
Selon un autre mode particulier de réalisation de l'invention, l'enzyme à activité méthionine synthase modifiée est une enzyme purifiée.
DESCRIPTION DES FIGURES Figure 1 : synthèse de la méthionine à partir de l'homosérine, chez les bactéries.
Figure 2 : Schéma de synthèse de la méthionine selon l'invention appliqué dans E. coli ; une stratégie équivalente est transposable chez de nombreux microorganismes dont C. glutamicum. Les stratégies metB* ou metY** nécessitent l'utilisation d'une souche initialement au moins A(metE) tandis que les stratégies metB** ou metY* nécessitent l'utilisation d'une souche initialement au moins A(metC).
Légende :
MetA : homosérine succinyltransferase ; pourra être remplacé par une isoforme insensible à la rétro-inhibition par la méthionine ou par une isoforme homoserine acetyltransferase éventuellement insensible à la rétro-inhibition par la méthionine.
MetA : homosérine succinyltransferase ; pourra être remplacé par une isoforme insensible à la rétro-inhibition par la méthionine ou par une isoforme homoserine acetyltransferase éventuellement insensible à la rétro-inhibition par la méthionine.
MetB : cystathionine y-synthase
MetB* : cystathionine y-synthase évoluée en méthionine synthase
MetB** : cystathionine y-synthase évoluée en homocysteine synthase
MetY* : O-acetyl-homosérine (de C. glutamicum) évoluée en homocysteine synthase
MetB* : cystathionine y-synthase évoluée en méthionine synthase
MetB** : cystathionine y-synthase évoluée en homocysteine synthase
MetY* : O-acetyl-homosérine (de C. glutamicum) évoluée en homocysteine synthase
<Desc/Clms Page number 22>
MetY** : O-acetyl-homosérine (de C. glutamicum) évoluée en méthionine synthase
La voie centrale représente la voie naturelle de synthèse de la méthionine chez E. coli. Les autres voies indiquées correspondent aux procédés selon l'invention.
La voie centrale représente la voie naturelle de synthèse de la méthionine chez E. coli. Les autres voies indiquées correspondent aux procédés selon l'invention.
Figure 3 : représentation d'un mécanisme de fermentation continue pour la sélection dirigée des souches selon l'invention. Nous utilisons par exemple la technologie Fedbacth-Pro de la société DASGIP. Il s'agit d'un système modulaire contrôlé par ordinateur permettant la fermentation en parallèle de microorganismes en ayant un control de l'alimentation en milieu, en pH et p02.
Figure 4 : de spectres de 13C-RMN, correspondant au carbone 5 de la méthionine, obtenus par HSQC sur un hydrolysat de la souche sauvage (haut) ou de la souche Kla-F optimisée (bas). On observe que le carbone 5 de la souche Kla-F n'est pas marqué au carbone 13 ce qui confirme qu'il provient du méthylmercaptide de sodium Figure 5 : de séquences non modifiées de cystathionine-y-synthases.
Alignement réalisé avec l'algorithme MULTALIN

(http://prodes.toulouse.inra.fr/multalin/ cgi-bin/multalin. pl) Figure 6 : Alignement de séquences non modifiées de acylhomosérine sulfhydrylases.
(http://prodes.toulouse.inra.fr/multalin/ cgi-bin/multalin. pl) Figure 6 : Alignement de séquences non modifiées de acylhomosérine sulfhydrylases.
Alignement réalisé avec l'algorithme MULTALIN.
Figure 7 : Cinétique de croissance de la population E. coli A(metC) lors de son ensemencement initial (culture N 1) et lors de son dixième repiquage (Repicage 10).
EXEMPLES
Exemple 1 : Construction de la souche A(metE)
L'inactivation du gène metE est réalisée en insérant une cassette de résistance à l'antibiotique chloramphénicol tout en délétant la majeure partie du gène concerné.
Exemple 1 : Construction de la souche A(metE)
L'inactivation du gène metE est réalisée en insérant une cassette de résistance à l'antibiotique chloramphénicol tout en délétant la majeure partie du gène concerné.
La technique utilisée est décrite par Datsenko, K.A. ; Wanner, B. L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products.
Proc. Natl. Acad. Sci. USA 97 : 6640-6645.
Deux oligonucléotides sont utilisés pour réaliser la stratégie: - DmetER de 100 bases (SEQ ID NO 9): tacccccgacgcaagttctgcgccgcctgcaccatgttcgccagtgccgcgcgggtttctggccagccgcgcg ttttcagCATATGAATATCCTCCTTAG
<Desc/Clms Page number 23>
avec : une région (caractères minuscules) homologue à la séquence (4012903 à 4012824) du gène metE (séquence 4010643 à 4012904, séquence de référence sur le

site http://Renolist.pasteur.fr/Colibri/) une région (caractères majuscules) pour l'amplification de la cassette de résistance au chloramphénicol du plasmide pKD3 (Datsenko, K.A. ; Wanner, B. L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97 : 6640-6645) - DmetEF de 100 bases (SEQ ID NO 10): tgacaatattgaatcacaccctcggtttccctcgcgttggcctgcgtcgcgagctgaaaaaagcgcaagaaagtt attggTGTAGGCTGGAGCTGCTTCG avec : une région (caractères minuscules) homologue à la séquence (4010644 à 4010723) du gène metE une région (caractères majuscules) pour l'amplification de la cassette de résistance au chloramphénicol portée par le plasmide pKD3
Les oligonucléotides DmetER et DmetEF sont utilisés pour amplifier la cassette de résistance au chloramphénicol à partir du plasmide pKD3. Le produit PCR obtenu est alors introduit par électroporation dans la souche MG1655 (pKD46) dans laquelle l'enzyme Red recombinase exprimée permet la recombinaison homologue.
site http://Renolist.pasteur.fr/Colibri/) une région (caractères majuscules) pour l'amplification de la cassette de résistance au chloramphénicol du plasmide pKD3 (Datsenko, K.A. ; Wanner, B. L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97 : 6640-6645) - DmetEF de 100 bases (SEQ ID NO 10): tgacaatattgaatcacaccctcggtttccctcgcgttggcctgcgtcgcgagctgaaaaaagcgcaagaaagtt attggTGTAGGCTGGAGCTGCTTCG avec : une région (caractères minuscules) homologue à la séquence (4010644 à 4010723) du gène metE une région (caractères majuscules) pour l'amplification de la cassette de résistance au chloramphénicol portée par le plasmide pKD3
Les oligonucléotides DmetER et DmetEF sont utilisés pour amplifier la cassette de résistance au chloramphénicol à partir du plasmide pKD3. Le produit PCR obtenu est alors introduit par électroporation dans la souche MG1655 (pKD46) dans laquelle l'enzyme Red recombinase exprimée permet la recombinaison homologue.
Les transformants résistants à l'antibiotique sont alors sélectionnés et l'insertion de la cassette de résistance est vérifiée par une analyse PCR avec les oligonucléotides metER et metEF.
MetER (SEQ ID NO 11) : ggtttaagcagtatggtgggaagaagtcgc (homologue à la séquence de 4012978 à 4012949)
MetEF (SEQ ID NO 12) : cccggggatgaataaacttgccgccttccc (homologue à la séquence de 4010567 à 4010596)
La cassette de résistance au chloramphénicol peut ensuite être éliminée. Le plasmide pCP20 porteur de la recombinase FLP agissant au niveau des sites FRT de la cassette de résistance au chloramphénicol, est alors introduit dans les souches recombinantes par électroporation. Après une série de culture à 42 C, la perte de la cassette de résistance à l'antibiotique est vérifiée par une analyse PCR avec les mêmes oligonucléotides que ceux utilisés précédemment.
MetEF (SEQ ID NO 12) : cccggggatgaataaacttgccgccttccc (homologue à la séquence de 4010567 à 4010596)
La cassette de résistance au chloramphénicol peut ensuite être éliminée. Le plasmide pCP20 porteur de la recombinase FLP agissant au niveau des sites FRT de la cassette de résistance au chloramphénicol, est alors introduit dans les souches recombinantes par électroporation. Après une série de culture à 42 C, la perte de la cassette de résistance à l'antibiotique est vérifiée par une analyse PCR avec les mêmes oligonucléotides que ceux utilisés précédemment.
<Desc/Clms Page number 24>
Exemple 2: Modification de la souche (metE) pour la production de méthionine à partir de méthyl-mercaptan
Pour optimiser E. coli pour la production de méthionine à partir du méthylmercaptan, on effectue une sélection dirigée en flacons.
Pour optimiser E. coli pour la production de méthionine à partir du méthylmercaptan, on effectue une sélection dirigée en flacons.
La souche E. coliK12 est préalablement rendue auxotrophe pour la méthionine en inactivant le gène metE, préparée selon les conditions de l'exemple 1 ne peut donc croître qu'en fabriquant sa propre méthionine, par l'utilisation du méthyl-mercaptan.
La mise en #uvre de cette technique permet la sélection d'une souche de Escherichia coli dont la cystathionine-y-synthase (EC 4. 2.99.9) a développé une activité méthionine-synthase modifiée en présence de méthyl-mercaptan.
La sélection dirigée est conduite en flacon en verre hermétiquement fermé contenant 50 mL de milieu minéral (Schaefer et al., 1999, Anal. Biochem. 270: 88- 96) en présence de 33 mM glucose, et du chloramphénicol à une concentration finale de 25 mg/1.
Les milieux de culture sont ensemencés avec la souche E. coli K12 metE à DO600nm définie. On ensemence avec une population de bactéries suffisamment importante pour que certaines bactéries possèdent potentiellement des mutations spontanées intéressantes dans le gène metB, permettant d'assimiler le méthylmercaptan. Cette population a été obtenue par culture de la souche auxotrophe en méthionine sur milieu minimum supplémenté en méthionine.
Trois flacons reçoivent alors 100 L d'une solution à 400 mg/L de sodium mercaptide, tandis qu'un quatrième flacon ne reçoit pas de sodium mercaptide. Les cultures sont réalisées sous agitation, à 37 C, pendant 6 jours, puis la DO600nm est mesurée. Les résultats sont résumés dans le tableau N 3.
Tableau 3. Mesure de la densité optique de milieux de cultures pour E. coli en présence (flacons 1-3) ou absence (flacon témoin) de sodium mercaptide.

<tb>
<tb>
<tb>
Flacon <SEP> 1 <SEP> Flacon <SEP> 2 <SEP> Flacon <SEP> 3 <SEP> Flacon <SEP> Témoin
<tb> DO600nm <SEP> à <SEP> T=0 <SEP> 0.34 <SEP> 0.34 <SEP> 0.34 <SEP> 0.34
<tb> DO600nm <SEP> à <SEP> T=6 <SEP> jours <SEP> 0.23 <SEP> 1.14 <SEP> 0.79 <SEP> 0.32
<tb>
<tb> DO600nm <SEP> à <SEP> T=0 <SEP> 0.34 <SEP> 0.34 <SEP> 0.34 <SEP> 0.34
<tb> DO600nm <SEP> à <SEP> T=6 <SEP> jours <SEP> 0.23 <SEP> 1.14 <SEP> 0.79 <SEP> 0.32
<tb>
<Desc/Clms Page number 25>
Ces résultats montrent que les flacons 2 et 3 ont permis de multiplier une souche capable d'utiliser le méthyl-mercaptan pour produire la méthionine nécessaire à sa croissance (augmentation de la densité optique).
L'activité méthionine synthase améliorée observée provient d'une modification dans le gène de la cystathionine y-synthase de la souche E. coli K12 #metE, contenue dans les flacons 2 et 3.
La population bactérienne du Flacon 2 peut alors être utilisée pour améliorer davantage l'activité méthionine synthase en présence de méthyl-mercaptan, en utilisant un procédé de criblage et amélioration par fermentation en étage, ou en recommençant le procédé en flacon tel qu'il vient d'être décrit.
Exemple 3 : Criblage et amélioration par fermentation en étage
Pour optimiser E. coli pour la production de méthionine à partir du méthylmercaptan, on effectue une sélection.
Pour optimiser E. coli pour la production de méthionine à partir du méthylmercaptan, on effectue une sélection.
La mise en #uvre de cette technique permet la sélection d'une souche de Escherichia coli dont la cystathionine-y-synthase (EC 4. 2.99.9) a développé une activité méthionine-synthase améliorée en présence de méthyl-mercaptan.
Alternativement, on peut utiliser une souche obtenue selon l'exemple 2.
La sélection dirigée est conduite dans un système en continu en étage (Figure 3).
Le premier fermenteur produit les bactéries à une vitesse proche du taux de croissance maximum. Les bactéries passent en continu de ce fermenteur dans un second fermenteur caractérisé par un taux de dilution plus faible et un milieu avec le crible de sélection (ici, le méthyl-mercaptan).
La pression de sélection, imposée à la bactérie dans le second fermenteur, est établie par la concentration en méthyl-mercaptan. Des cycles successifs de sélection permettent d'appliquer aux bactéries des cribles de plus en plus fort par des concentrations croissantes en méthyl-mercaptan.
Pour chaque concentration, la souche sélectionnée dans le second fermenteur est celle qui a évolué pour métaboliser la totalité du méthyl-mercaptan (méthylmercaptan résiduel nul dans le fermenteur).
<Desc/Clms Page number 26>
Dans ce cas, on recommence la sélection en utilisant le fermenteur n 2 comme fermenteur de croissance et le fermenteur n 1comme fermenteur de crible, présentant du méthyl-mercaptan de concentration plus forte qu'à l'étape précédente.
On effectue différents cycles de sélection pour obtenir une souche fermentant le méthyl-mercaptan avec une vitesse élevée. L'analyse de cette souche permet de définir les mutations dans le gène de la cystathionine-y-synthase.
Exemple 4 :
La population d'E. coli K12 AmetE issue du flacon 2 de l'exemple 2 a subi des repiquages successifs en flacon. La nouvelle population obtenue Kla-F est mise en culture dans un milieu minimum (Schaefer et al., 1999, Anal. Biochem. 270 : 88-96) contenant 2,5 g.l-1 de glucose entièrement marqué au carbone 13 et du méthylmercaptide de sodium (200 ppm) non enrichi en carbone 13. Cette population est auxotrophe pour la méthionine en l'absence de méthylmercaptide de sodium.
La population d'E. coli K12 AmetE issue du flacon 2 de l'exemple 2 a subi des repiquages successifs en flacon. La nouvelle population obtenue Kla-F est mise en culture dans un milieu minimum (Schaefer et al., 1999, Anal. Biochem. 270 : 88-96) contenant 2,5 g.l-1 de glucose entièrement marqué au carbone 13 et du méthylmercaptide de sodium (200 ppm) non enrichi en carbone 13. Cette population est auxotrophe pour la méthionine en l'absence de méthylmercaptide de sodium.
Après la culture, les cellules sont récupérées, lavées puis hydrolysées par HC1 6N pendant 24 heures à 107 C. Une analyse par RMN bidimensionnelle est alors réalisée (HSQC). Cette analyse permet d'étudier le carbone 5 de la méthionine, qui provient, soit de la L-cystéine, produite à partir du glucose présent dans la solution (voie classique), soit du méthylmercaptide de sodium lorsque la nouvelle voie métabolique selon l'invention est utilisée.
L'expérience est conduite de manière similaire avec la souche sauvage E. coli K12 (produisant la méthionine à partir du glucose), en absence de méthylmercaptide de sodium.
La figure 4 montre deux spectres 1D, issus de deux acquisitions distinctes, superposés pour une meilleure lecture. Ces spectres 1D sont extraits de spectres RMN à deux dimension type HSQC (corrélation entre protons et carbone 13). Les spectres RMN à deux dimensions ont été obtenus sur un hydrolysat acide des bactéries.
L'échantillon analysé est un hydrolysat total ; cependant du fait de la sensibilité de la RMN et des temps d'acquisition utilisés, on détecte essentiellement les acides aminés, les sucres, les bases et le glycérol. Chaque carbone (couplé à un proton) de chaque acide aminé donne une résonance magnétique nucléaire.
<Desc/Clms Page number 27>
Le carbone 5 de la méthionine (c'est à dire le groupe méthyl terminal) présente un déplacement chimique d'environ 14,7 ppm. La figure 4 présente la zone de déplacement chimique centrée autour de 14,7 ppm pour les deux souches.
On remarque que dans le cas du spectre supérieur, le signal du carbone 5 est fort, indiquant que le carbone 5 est marqué au carbone 13. En conséquence, ce carbone 5 provient du glucose marqué introduit comme substrat dans le milieu de culture.
On remarque par contre que le même signal est très faible dans le spectre inférieur (souche Kla-F). Cela signifie que le carbone 5 n'est pratiquement pas marqué. Pourtant les autres carbones de la molécule sont fortement marqués (résultats non présentés). Le carbone 5 non marqué ne provient donc pas du glucose mais du méthyl-mercaptan.
On peut donc conclure que la souche Kla-F produit de la méthionine à partir de succinyl-L-homosérine et de méthylmercaptide de sodium.
Exemple 5 :
La population Kla-F subit 14 nouveaux cycles de repiquage successifs en flacon. On obtient ainsi la population Kl 44 que l'on étale alors sur milieu minimum gélosé contenant du glucose pour seule source de carbone. Les boites inoculées sont placées en condition aerobie dans une jarre anaérobie dans laquelle est introduit un tube contenant du méthylmercaptide de sodium dissout dans de l'eau, lajarre est alors placée dans un incubateur à 37 C. La température d'ébullition du methylmercaptide de sodium étant de 5 C, l'atmosphère de la jarre anaérobie s'enrichie en methylmercaptan. Après 4 jours, les clones apparaissent sur les boites ; ils correspondent aux bactéries capables de produire de la méthionine en présence de méthylmercaptan. Dix clones sont isolés, dont le clone K176. Le clone K176 est multiplié en culture liquide et des stocks glycérols sont réalisés portant le numéro K183.
La population Kla-F subit 14 nouveaux cycles de repiquage successifs en flacon. On obtient ainsi la population Kl 44 que l'on étale alors sur milieu minimum gélosé contenant du glucose pour seule source de carbone. Les boites inoculées sont placées en condition aerobie dans une jarre anaérobie dans laquelle est introduit un tube contenant du méthylmercaptide de sodium dissout dans de l'eau, lajarre est alors placée dans un incubateur à 37 C. La température d'ébullition du methylmercaptide de sodium étant de 5 C, l'atmosphère de la jarre anaérobie s'enrichie en methylmercaptan. Après 4 jours, les clones apparaissent sur les boites ; ils correspondent aux bactéries capables de produire de la méthionine en présence de méthylmercaptan. Dix clones sont isolés, dont le clone K176. Le clone K176 est multiplié en culture liquide et des stocks glycérols sont réalisés portant le numéro K183.
Le clone K183 est envoyé au séquençage en même temps que la souche E. coli K12 A(metE) initiale. Pour chaque clone la séquence des gènes metJ et metB (SEQ ID N 7) est déterminée. La séquence obtenue pour metB permet d'observer la présence d'une alanine en position 325 (SEQ ID N 8) en remplacement d'un
<Desc/Clms Page number 28>
glutamate (SEQ ID N 6). Le gène metJ ne présente aucune mutation. Cette souche a été déposé à la CNCM le 2 avril 2003, sous le numéro 1-3005.
Exemple 6 :
Le clone Kl 83 est cultivé en flacon, en milieu minimum avec glucose et methylmercaptide de sodium pour seule source de carbone. En parallèle, une culture est réalisée dans des conditions identiques avec la souche E. coli Kl2 sauvage. On observe que la consommation de glucose par g de biomasse est deux fois plus importante que pour une souche de E. coli sauvage. La surconsommation est probablement due en partie à la production d'acétate.

Le clone Kl 83 est cultivé en flacon, en milieu minimum avec glucose et methylmercaptide de sodium pour seule source de carbone. En parallèle, une culture est réalisée dans des conditions identiques avec la souche E. coli Kl2 sauvage. On observe que la consommation de glucose par g de biomasse est deux fois plus importante que pour une souche de E. coli sauvage. La surconsommation est probablement due en partie à la production d'acétate.
<tb>
<tb>
<tb>
Souches <SEP> Rendement <SEP> Biomasse <SEP> Rendement <SEP> Acétate
<tb> MBMOI <SEP> 0.45 <SEP> <0.002
<tb> K183 <SEP> 0.24 <SEP> 0.36 <SEP>
<tb>
comparaison au rendement de biomasse entre la souche E. coll sauvage et le clone évolué K 18 3 :
Rendement Biomasse exprimé en g(biomasse)/g(glucose)
Rendement Acétate exprimé en g(acetate)/g(glucose).
<tb> MBMOI <SEP> 0.45 <SEP> <0.002
<tb> K183 <SEP> 0.24 <SEP> 0.36 <SEP>
<tb>
comparaison au rendement de biomasse entre la souche E. coll sauvage et le clone évolué K 18 3 :
Rendement Biomasse exprimé en g(biomasse)/g(glucose)
Rendement Acétate exprimé en g(acetate)/g(glucose).
L'analyse des métabolites intracellulaires et extracellulaire de ces deux cultures montre notamment :
En intracellulaire, une augmentation de l'alanine, du pyruvate, du ketobutyrate et de 2 ketoisocaproate et une diminution de la concentration en tryptophane, norvaline, norleucine, leucine et methionine.
En intracellulaire, une augmentation de l'alanine, du pyruvate, du ketobutyrate et de 2 ketoisocaproate et une diminution de la concentration en tryptophane, norvaline, norleucine, leucine et methionine.
En extracellulaire, une accumulation de glutamate, d'isoleucine, thréonine, valine et 2-ketoisocaproate et une diminution de pyruvate, norleucine, tryptophane.
Exemple 7 : de l'activité spécifique méthionine synthase des souches MBM01 et Kl 83 en présence de méthylmercaptan.
Afin de montrer l'amélioration de l'activité méthionine synthase dans la souche K183 par rapport à la souche sauvage (MBM01), des réactions enzymatiques sont réalisées en utilisant des extraits acellulaires préparés à partir de cultures des souches Kl 83 et MBM01 réalisées sur riche (milieu BH1, commercialisé par DIFCO,
<Desc/Clms Page number 29>
avec 2,5 g/L de glucose) en absence de méthylmercaptan. Les extraits protéiques sont désalés sur PD 10 et réservés sur glace.
Conditions réactionnelles et traitement de l'échantillon - Préparer sur la glace une solution de sodium methanethiolate diluée 10 fois (100 ul de solution 3M plus 900 l d'eau mQ) - Préparer sur la glace les mélanges réactionnels constitué de 20 L de tampon phosphate 500 mM pH 6.5, 10 l Pyridoxal phosphate 2,5 mM, 16 L 0- succinylhomosérine 25 mM, 10 L Sodium methanethiolate 0,3 M, 24 L eau milliQ.
- placer les tubes à 37 C (thermomixer sous la hotte) et ajouter l'extrait protéique (20 l) pour démarrer la réaction.
- Pour arrêter la réaction (0 à 30 minutes), placer les tubes sur glace et ajouter 400 l d'acétone à -20 C.
- placer les tubes à -20 C pendant 30 minutes - puis ouvrir les tubes sous la hotte pendant 10 minutes pour évaporer le méthanethiol et l'acétone (les maintenir sur glace) - centrifuger 5 minutes à 10000 g (petite centri), transvaser le surnageant (100 L) dans des tubes éppendorf et diluer pour un volume final de 1 mL.
Mesure de l'activité méthionine synthase en détectant la quantité de succinate libéré du succinylhomosérine : - Dix l de l'échantillon ci-dessus obtenu sont analysés par chromatographie ionique en utilisant un Appareil Dionex DX-500 équipé d'une précolonne AG-11
2 mm et d'une colonne AS-11 2 mm, d'un suppresseur ASRS Ultra, d'une boucle injection 10 l. Un gradient est alors appliqué : 0 - 7 min 0,5 mM KOH ; min injection ; 7 - 9,5 min 0,5 mM KOH ; 13 min 0,5 - 5 mM KOH ; 25 min 5 - 38,3mM KOH.
2 mm et d'une colonne AS-11 2 mm, d'un suppresseur ASRS Ultra, d'une boucle injection 10 l. Un gradient est alors appliqué : 0 - 7 min 0,5 mM KOH ; min injection ; 7 - 9,5 min 0,5 mM KOH ; 13 min 0,5 - 5 mM KOH ; 25 min 5 - 38,3mM KOH.
Mesure de l'activité méthionine synthase en détectant la quantité de méthionine synthétisée en présence de methylmercaptan
L'analyse est réalisée par GC-MS ce qui nécessite de silyler les échantillons préalablement à l'injection. Pour cela chaque échantillon reçoit un standard interne (serinel3C) qui permet de valider la qualité de la silylation. Les échantillons sont ensuite lyophilisés pendant la nuit.
L'analyse est réalisée par GC-MS ce qui nécessite de silyler les échantillons préalablement à l'injection. Pour cela chaque échantillon reçoit un standard interne (serinel3C) qui permet de valider la qualité de la silylation. Les échantillons sont ensuite lyophilisés pendant la nuit.
La dérivation est réalisée en appliquant le protocole suivant :
<Desc/Clms Page number 30>
Ajouter 400 l de la solution d'hydroxylamine (0,250g +/- 0,002 g dissous dans 10 ml de Pyridine) à l'aide d'une pipette automatique de 1 ml et fermer correctement les tubes. Mélanger à l'aide d'un Vortex pendant 2 fois 10 secondes.
Concentrer le milieu réactionnel au fond du tube par centrifugation (maxi 1 minute à 5000 g) et laisser réagir 1 heure 30 à 30 C. Ouvrir les tubes et ajouter 1000 l de solution de BSTFA à l'aide d'une pipette automatique de 1 ml et compléter avec 100 l de Pyridine (pipette automatique de 200 l). Refermer les tubes, vortexer pendant 10 secondes et mettre à incuber respectivement pendant 60 minutes à 60 C dans le cas des dérivés TBDMS et 30 minutes à 70 C pour le BSTFA. Si nécessaire filtrer les échantillons sur filtre à usage unique avec membrane PTFE de 0,22 m ou centrifuger à 5000 g pendant 5 minutes. Transférer dans un flacon de 1,5 ml, sertir et injecter en CPG.
Les analyses ont été réalisées avec l'appareil Agilent technologies GC6890/MSD5973 équipé d'une colonne apolaire (HP-5MS, Bios Analytique). Le gaz vecteur est l'hélium à débit constant de 1 ml. min-1. L'injection de 1 l d'échantillon a lieu en mode splitless avec un débit de purge de 50 ml.min-1 pendant 0,85 min. Le profil de température est le suivant : la température initiale de 90 C est maintenue pendant 2 minutes puis augmente jusqu'à 320 C avec une pente de 10 C.min-1. Cette température est maintenue pendant 6 minutes. La détection se fait par spectrométrie de masse avec ionisation par impact électronique en mode balayage dans une gamme variant de m/z = 40 à 550 amu. Le délai de passage de solvant est réglé à 3,10 minutes.
Résultats
Dans ces conditions, une activité méthionine synthase a pu être dosée dans les échantillons incubés avec le méthanethiol, via la quantification d'une part, du succinate par chromatographie ionique et d'autre part, de la méthionine par GC-MS.
Dans ces conditions, une activité méthionine synthase a pu être dosée dans les échantillons incubés avec le méthanethiol, via la quantification d'une part, du succinate par chromatographie ionique et d'autre part, de la méthionine par GC-MS.
<Desc/Clms Page number 31>

<tb>
<tb>
<tb>
Activité <SEP> spécifique
<tb> Souche <SEP> Culture <SEP> Extrait <SEP> Concentration <SEP> (mUI/mg <SEP> protéines)
<tb> ouc <SEP> ure <SEP> protéique <SEP> en <SEP> protéines <SEP> Dosage <SEP> Dosage
<tb> Succinate <SEP> Méthionine
<tb> MBM01 <SEP> FB <SEP> 137/P2 <SEP> Z63 <SEP> 3,43 <SEP> 0,30 <SEP> 0,23
<tb> K183 <SEP> FB <SEP> 140/P2 <SEP> Z64 <SEP> 3,62 <SEP> 1,40 <SEP> 1,72
<tb>
<tb> Souche <SEP> Culture <SEP> Extrait <SEP> Concentration <SEP> (mUI/mg <SEP> protéines)
<tb> ouc <SEP> ure <SEP> protéique <SEP> en <SEP> protéines <SEP> Dosage <SEP> Dosage
<tb> Succinate <SEP> Méthionine
<tb> MBM01 <SEP> FB <SEP> 137/P2 <SEP> Z63 <SEP> 3,43 <SEP> 0,30 <SEP> 0,23
<tb> K183 <SEP> FB <SEP> 140/P2 <SEP> Z64 <SEP> 3,62 <SEP> 1,40 <SEP> 1,72
<tb>
On observe donc que l'activité méthionine synthase en présence de methylmercaptan est renforcée dans la souche évoluée par rapport à la souche sauvage confirmant que la cystathionine y-synthase mutée (E325A) a une activité méthionine synthase modifiée.
Exemple 8 : Construction de la souche A(metC, metJB) puis sélection d'un gène metY évolué.
Construction des souches MG1655 (AmetC::Cm) et MG1655 (#metC)
Pour inactiver le gène metC la stratégie de recombinaison homologue décrite par Datsenko & Wanner (2000) est utilisée. Cette stratégie permet l'insertion d'une cassette de résistance à l'antibiotique chloramphénicol tout en délétant la majeure partie du gène concerné. Pour cela, nous avons utilisé deux oligonucléotides :
Pour metC : - DmetCR de 100 bases (SEQ ID NO 13): ccggcgtccagatcggcaatcagatcgtcgacatcttccagaccaatatgcaggcgaatcaaggtcccgctaa aatcgatCATATGAATATCCTCCTTAG avec une région (caractères minuscules) homologue à la séquence (3151419 à 3151359) du gène metC (séquence 3150251 à 3151438, séquence de référence sur le site http://genolist.pasteur.fr/Colibri/) une région (caractères majuscules) pour l'amplification de la cassette de résistance au chloramphénicol (séquence de référence dans la publication Datsenko, K. A. et Wanner, B. L., 2000, PNAS, 97 : 6640-6645) - DmetCF de 100 bases (SEQ ID NO 14) : cggacaaaaagcttgatactcaactggtgaatgcaggacgcagcaaaaaatacactctcggcgcggtaaatag cgtgattTGTAGGCTGGAGCTGCTTCG avec
Pour inactiver le gène metC la stratégie de recombinaison homologue décrite par Datsenko & Wanner (2000) est utilisée. Cette stratégie permet l'insertion d'une cassette de résistance à l'antibiotique chloramphénicol tout en délétant la majeure partie du gène concerné. Pour cela, nous avons utilisé deux oligonucléotides :
Pour metC : - DmetCR de 100 bases (SEQ ID NO 13): ccggcgtccagatcggcaatcagatcgtcgacatcttccagaccaatatgcaggcgaatcaaggtcccgctaa aatcgatCATATGAATATCCTCCTTAG avec une région (caractères minuscules) homologue à la séquence (3151419 à 3151359) du gène metC (séquence 3150251 à 3151438, séquence de référence sur le site http://genolist.pasteur.fr/Colibri/) une région (caractères majuscules) pour l'amplification de la cassette de résistance au chloramphénicol (séquence de référence dans la publication Datsenko, K. A. et Wanner, B. L., 2000, PNAS, 97 : 6640-6645) - DmetCF de 100 bases (SEQ ID NO 14) : cggacaaaaagcttgatactcaactggtgaatgcaggacgcagcaaaaaatacactctcggcgcggtaaatag cgtgattTGTAGGCTGGAGCTGCTTCG avec
<Desc/Clms Page number 32>
une région (caractères minuscules) homologue à la séquence (3150255 à 3150334) du gène metC une région (caractères majuscules) pour l'amplification de la cassette de résistance au chloramphénicol.
Les oligonucléotides DmetCR et DmetCF sont utilisés pour amplifier la cassette de résistance au chloramphénicol à partir du plasmide pKD3. Le produit PCR obtenu est ensuite introduit par électroporation dans la souche MG1655 (pKD46) dans laquelle l'enzyme Red recombinase exprimée permet la recombinaison homologue. Les transformants résistants à l'antibiotique sont alors sélectionnés et l'insertion de la cassette de résistance est vérifiée par une analyse PCR avec les oligonucléotides metCR et metCF. La souche retenue est nommée MG1655 (metC::Cm).
MetCR (SEQ ID NO 15) : cgtccgggacgccttgatcccggacgcaac (homologue à la séquence de 3151522 à 3151493)
MetCF (SEQ ID NO 16) : gcgtttacgcagtaaaaaagtcaccagcacgc (homologue à la séquence de 3150118 à 3150149)
La cassette de résistance au chloramphénicol peut ensuite être éliminée. Le plasmide pCP20 porteur de la recombinase FLP agissant au niveau des sites FRT de la cassette de résistance au chloramphénicol, est alors introduit dans les souches recombinantes par électroporation. Après une série de culture à 42 C, la perte de la cassette de résistance à l'antibiotique est vérifiée par une analyse PCR avec les mêmes oligonucléotides que ceux utilisés précédemment. La souche retenue est nommée MG1655 (AmetC).
MetCF (SEQ ID NO 16) : gcgtttacgcagtaaaaaagtcaccagcacgc (homologue à la séquence de 3150118 à 3150149)
La cassette de résistance au chloramphénicol peut ensuite être éliminée. Le plasmide pCP20 porteur de la recombinase FLP agissant au niveau des sites FRT de la cassette de résistance au chloramphénicol, est alors introduit dans les souches recombinantes par électroporation. Après une série de culture à 42 C, la perte de la cassette de résistance à l'antibiotique est vérifiée par une analyse PCR avec les mêmes oligonucléotides que ceux utilisés précédemment. La souche retenue est nommée MG1655 (AmetC).
Construction de la souche MG1655 (AmetB-AmetJ)
Pour déléter les gènes metB et metJ, nous avons inséré une cassette de résistance à l'antibiotique chloramphénicol tout en délétant la majeure partie des gènes concernés. Pour cela, nous avons utilisé deux oligonucléotides.
Pour déléter les gènes metB et metJ, nous avons inséré une cassette de résistance à l'antibiotique chloramphénicol tout en délétant la majeure partie des gènes concernés. Pour cela, nous avons utilisé deux oligonucléotides.
Pour metBJ : - MetJR de 30 bases (SEQ ID NO 18): ggtacagaaaccagcaggctgaggatcagc homologue à la séquence (4125431 à 4125460) en aval du gène metJ (séquence 4125975 à 4125581, séquence de référence sur le site http ://genolist.pasteur.fr/Colibri/)
<Desc/Clms Page number 33>
- DmetJBF de 100 bases (SEQ ID NO 17) : tatgcagctgacgacctttcgcccctgcctgcgcaatcacactcatttttaccccttgtttgcagcccggaagccat tttcaggcaccagagtaaacatt avec une partie homologue (caractères majuscule) à la séquence (4126217 à 4126197) entre les gènes metJ et metB (séquence 4126252 à 4127412) contenant la région promotrice de l'opéron metBLF. une partie homologue (caractères minuscule) à la séquence (4127460 à 4127380) correspondant au début du gène metL (séquence 4127415 à 4129847) et à la fin du gène metB.
Ces deux oligonucléotides ont été utilisés pour amplifier la région concernée sur l'ADN chromosomique de MG1655 (#metJ : :Cm) (figure 8).
Le produit PCR obtenu est alors introduit par électroporation dans la souche MG1655 (pKD46) dans laquelle l'enzyme Red recombinase exprimée permet la recombinaison homologue. Les transformants résistants à l'antibiotique sont alors sélectionnés et la délétion de gène metB par recombinaison homologue est vérifiée par une analyse PCR avec les oligonucléotides MetJR et MetLR.
MetJR (SEQ ID NO 18) : ggtacagaaaccagcaggctgaggatcagc (homologue à la séquence 4125431 à 4125460)
MetLR (SEQ ID NO 19) : aaataacacttcacatcagccagactactgccaccaaattt (homologue à la séquence de 4127500 à 4127460)
L'événement de recombinaison homologue peut avoir lieu à deux endroits (figure 9).
MetLR (SEQ ID NO 19) : aaataacacttcacatcagccagactactgccaccaaattt (homologue à la séquence de 4127500 à 4127460)
L'événement de recombinaison homologue peut avoir lieu à deux endroits (figure 9).
Seul le cas B de la figure 9 (ligne inférieure) correspond à la souche souhaitée MG1655 (#metB-#metJ : :Cm) où les gènes metJ et metB ont été éliminés et le promoteur de l'opéron metBLF replacé devant metL.
La cassette de résistance au chloramphénicol peut ensuite être éliminée. Le plasmide pCP20 porteur de la recombinase FLP agissant au niveau des sites FRT de la cassette de résistance au chloramphénicol, est alors introduit dans les souches recombinantes par électroporation. Après une série de culture à 42 C, la perte de la cassette de résistance à l'antibiotique est vérifiée par une analyse PCR avec les mêmes oligonucléotides que ceux utilisés précédemment (MetLR et MetJR).
<Desc/Clms Page number 34>
Construction des souches MG1655 #(metC : :Cm, metJB) et MG1655 A(metC, metJB)
Pour déléter le gène metC (séquence 3150251 à 3151438, séquence de référence sur le site http://genolist.pasteur.fr/Colibri/) de la souche MG1655 (AmetB- AmetJ), nous avons utilisé la technique de transduction avec le phage P1. Le protocole suivi est réalisé en deux étapes avec la préparation du lysat de phage sur la souche MG1655 (#metC::Cm) puis transduction dans la souche MG1655 (AmetB- AmetJ).
Pour déléter le gène metC (séquence 3150251 à 3151438, séquence de référence sur le site http://genolist.pasteur.fr/Colibri/) de la souche MG1655 (AmetB- AmetJ), nous avons utilisé la technique de transduction avec le phage P1. Le protocole suivi est réalisé en deux étapes avec la préparation du lysat de phage sur la souche MG1655 (#metC::Cm) puis transduction dans la souche MG1655 (AmetB- AmetJ).
Préparation du lysat de phage P1: - Ensemencement par lOOul d'une culture de nuit de la souche MG1655 (#metC::Cm) de 10 ml de LB + Cm 30ug/ml + glucose 0,2% + CaCl2 5 mM - Incubation 30 min à 37 C sous agitation - Ajout de 100 l de lysat de phage Pl préparé sur la souche sauvage MG1655 (environ 1.109 phage/ml) - Agitation à 37 C pendant 3 heures jusqu'à la lyse totale des cellules - Ajout de 200 l de chloroforme et vortex - Centrifugation 10 min à 4500g pour éliminer les débris cellulaires - Transfert du surnageant dans un tube stérile et ajout de 200 l de chloroforme - Conservation du lysat à 4 C
Transduction - Centrifugation 10 min à 1500g de 5 ml d'une culture de nuit de la souche
MG1655 (#meB-#metJ) en milieu LB - Suspension du culot cellulaire dans 2,5 ml de MgS04 10 mM, CaCl2 5 mM - - Tubes témoins : 100 l cellules
100 l phages P1 de la souche MG1655 (AmetC::Cm) - Tube test: 100 l de cellules + 100 l de phages P1 de la souche MG1655 (#metC::Cm) - Incubation 30 min à 30 C sans agitation - Ajout de 100 l de citrate de sodium 1 M dans chaque tube puis vortex - Ajout de 1 ml de LB - Incubation 1 heure à 37 C sous agitation - Etalement sur des boîtes LB + Cm 30 ug/ml après centrifugation 3 min à 7000 rpm des tubes.
Transduction - Centrifugation 10 min à 1500g de 5 ml d'une culture de nuit de la souche
MG1655 (#meB-#metJ) en milieu LB - Suspension du culot cellulaire dans 2,5 ml de MgS04 10 mM, CaCl2 5 mM - - Tubes témoins : 100 l cellules
100 l phages P1 de la souche MG1655 (AmetC::Cm) - Tube test: 100 l de cellules + 100 l de phages P1 de la souche MG1655 (#metC::Cm) - Incubation 30 min à 30 C sans agitation - Ajout de 100 l de citrate de sodium 1 M dans chaque tube puis vortex - Ajout de 1 ml de LB - Incubation 1 heure à 37 C sous agitation - Etalement sur des boîtes LB + Cm 30 ug/ml après centrifugation 3 min à 7000 rpm des tubes.
<Desc/Clms Page number 35>
- Incubation à 37 C jusqu'au lendemain
Vérification de la souche
Les transformants résistants à l'antibiotique sont alors sélectionnés et l'insertion de la région contenant (metC::Cm) est vérifiée par une analyse PCR avec les oligonucléotides MetCR et MetCF d'une part et MetJR et MetLR d'autre part afin de vérifier également la souche délétée des gènes metB et metJ. La souche retenue est dénommée MG1655 A(metC : :Cm, metJB).
Vérification de la souche
Les transformants résistants à l'antibiotique sont alors sélectionnés et l'insertion de la région contenant (metC::Cm) est vérifiée par une analyse PCR avec les oligonucléotides MetCR et MetCF d'une part et MetJR et MetLR d'autre part afin de vérifier également la souche délétée des gènes metB et metJ. La souche retenue est dénommée MG1655 A(metC : :Cm, metJB).
MetCR (SEQ ID NO 21) : cgtccgggacgccttgatcccggacgcaac (homologue à la séquence de 3151522 à 3151493)
MetCF (SEQ ID NO 22) : gcgtttacgcagtaaaaaagtcaccagcacgc (homologue à la séquence de 3150118 à 3150149)
Comme précédemment, la cassette de résistance au chloramphénicol peut ensuite être éliminée. Le plasmide pCP20 porteur de la recombinase FLP agissant au niveau des sites FRT de la cassette de résistance au chloramphénicol, est alors introduit dans les souches recombinantes par électroporation. Après une série de culture à 42 C, la perte de la cassette de résistance à l'antibiotique est vérifiée par une analyse PCR avec les mêmes oligonucléotides que ceux utilisés précédemment (MetCR et MetCF d'une part et MetJR et Met LR d'autre part). La souche retenue est dénommée MG1655 A(metC, metJB).
MetCF (SEQ ID NO 22) : gcgtttacgcagtaaaaaagtcaccagcacgc (homologue à la séquence de 3150118 à 3150149)
Comme précédemment, la cassette de résistance au chloramphénicol peut ensuite être éliminée. Le plasmide pCP20 porteur de la recombinase FLP agissant au niveau des sites FRT de la cassette de résistance au chloramphénicol, est alors introduit dans les souches recombinantes par électroporation. Après une série de culture à 42 C, la perte de la cassette de résistance à l'antibiotique est vérifiée par une analyse PCR avec les mêmes oligonucléotides que ceux utilisés précédemment (MetCR et MetCF d'une part et MetJR et Met LR d'autre part). La souche retenue est dénommée MG1655 A(metC, metJB).
Construction du plasmide pTopo-metY
Parallèlement un plasmide permettant l'expression du gène metY de C. glutamicum sera construit. Par exemple, ce gène peut être amplifié par PCR à partir d'ADN chromosomique de C. glutamicum puis introduit dans un plasmide. On pourra choisir d'amplifier par PCR le gène metY et éventuellement son promoteur naturel.
Parallèlement un plasmide permettant l'expression du gène metY de C. glutamicum sera construit. Par exemple, ce gène peut être amplifié par PCR à partir d'ADN chromosomique de C. glutamicum puis introduit dans un plasmide. On pourra choisir d'amplifier par PCR le gène metY et éventuellement son promoteur naturel.
Dans un mode de réalisation préféré, on clonera le gène metY sous le contrôle d'un promoteur permettant une expression dans E. coli. Les vecteurs utilisés pourront être choisis parmi les vecteurs pUC, pBluescript, pTopo, pCR-Script, ... Dans un mode de réalisation particulier, on pourrait utiliser le plasmide pSL191, décrit par Hwang et al.
(2002) J. Bact. 184(5) : 1277-1286, qui comprend le gène metY sous le contrôle de son promoteur naturel cloné dans un vecteur navette C. glutamicum - E. coli pMT 1.
La souche Escherichia coli A(metC, metJB) précédemment obtenue est transformée avec un plasmide portant le gène metY de C. glutamicum. La
<Desc/Clms Page number 36>
transformation de la souche pouvant être réalisée selon l'une des techniques connues de l'homme du métier.
La souche obtenue sera ensuite inoculée dans un Erlen Meyer contenant 10% de son volume en milieu minimum avec glucose pour seule source de carbone. La faible activité succinylhomoserine sulfhydrylase initialement portée par l'enzyme MetY devrait limiter la croissance de la population bactérienne du fait d'une limitation en méthionine synthétisée. Des repiquages sont réalisés tous les 4 jours pendant 1 moins. L'amélioration de l'activité succinylhomoserine sulfhydrylase portée par la proteine MetY devrait se traduire par une synthèse accrue en méthionine ;on devrait donc observer une amélioration du taux de croissance de la population bactérienne entre chaque repiquage, ce qui imposera le cas échéant d'augmenter la fréquence de repiquage afin de maintenir les microorganismes dans une phase de division. On évitera en effet de maintenir trop longtemps les cultures dans une phase stationnaire. Lorsque le taux de croissance se stabilisera d'un repiquage à un autre, on considérera que la souche a suffisamment évoluée et la sélection de trois clones sera réalisée. Leur séquençage permettra de déterminer la séquence évoluée metY*. Dans cette première étape, l'évolution du gène metY aura permis de modifier l'activité O-acetyl-homosérine sulfhydrylase en une activité 0succinyl-homosérine sulfhydrylase permettant de produire de l'homocysteine à partir d'O-succinylhomosérine et d'H2S ; deux substrats étant produits par la bactérie.
Afin d'optimiser le processus d'évolution de metY, une démarche similaire peut-être réalisée en utilisant d'autres mutants d'Escherichia coli, on pourra notamment utiliser le mutant A(metC, metB).
Exemple 9: Procédé de culture Fed-Batch pour la production et la purification de méthionine.
Préculture :
La préculture est réalisée pendant une nuit en fiole de 500 ml contenant 50 ml de milieu minimum type M9 modifié, complété avec 2.5 g/1 de glucose. Les cellules sont récupérées par centrifugation et reprises dans 5 ml de milieu minimum type M9 modifié.
La préculture est réalisée pendant une nuit en fiole de 500 ml contenant 50 ml de milieu minimum type M9 modifié, complété avec 2.5 g/1 de glucose. Les cellules sont récupérées par centrifugation et reprises dans 5 ml de milieu minimum type M9 modifié.
Culture en fermenteur.
<Desc/Clms Page number 37>
La culture est réalisée dans un fermenteur de volume utile de 300ml de type Fedbatch-pro DASGIP.
Le fermenteur est rempli avec 145 ml de milieu minimum type M9 modifié et inoculer avec les 5 ml de préculture. Soit une D0600nm d'inoculation comprise entre 0. 5 et 1.2.
La température de la culture est maintenue entre 30 et 37 c et le pH est ajusté en permanence à une valeur comprise entre 6. 5 et 8. Il est partiellement régulé par l'ajout d'une solution CH3SNa. Une solution de soude 2N peut le cas échéant compléter la régulation. L'agitation est maintenue à entre 200 et 400 rpm pendant la phase de batch et est augmentée jusqu'à 1000 rpm en fin de fed-batch. La teneur en 02 dissout est maintenue entre 30% et 40% de saturation en utilisant un contrôleur de gaz. Dés que la OD 600nm à une valeur comprise entre 2. 5 et 3 nous commençons le fed-batch par ajout du milieu de fed à un débit initial compris entre 0. 3 et 0.5 ml/h avec une augmentation progressive jusqu'à des débits compris entre 2. 5 et 3.5 ml/h. après ce stade le débit est maintenu constant pendant un temps compris entre 24h et 32h . Le milieu de fed est constitué sur la base d'un milieu M9 modifié complémenté par une concentration en glucose comprise entre 300 et 500g/1 de glucose. Dans le même temps nous complémentons le milieu avec une solution de CH3SNa (solution entre 1 et 5M) afin de permettre la croissance de la bactérie tout en régulant le pH.
Dés que la concentration cellulaire atteint une concentration comprise entre 20 et 50 g/1 nous remplaçons le milieu de fed par un milieu minimum type M9 limité en phosphore. La solution de méthyl-mercaptan est remplacée par une injection directe de CH3SH sous forme gazeux dans le fermenteur. Le débit du gaz est adapté au débit de la solution de fed dans des rapports molaires avec le substrat carboné de 1 à 3. Dés que la concentration cellulaire est comprise entre 50 et 80 g/1 la fermentation est arrêtée. Le mout de fermentation est ajusté à un pH compris entre 7. 5 et 9 par une solution de NaOH puis chauffé à entre 60 et 80 c . Le mout est ensuite filtré sur des modules UF. La température du jus est maintenue entre 60 et 80 C, puis le jus est concentré avant passage sur charbon pour décoloration (en colonne ou en bacth). Le jus décoloré est de nouveau filtré pour retirer les dernières particules avant d'être acidifié par HC1 concentré jusqu'à un pH inférieur à 2.28 (pKl de la méthionine).
Les cristaux methionine.HCl ainsi formés sont récupérés par filtration, puis l'HCl est éliminé par évaporation pour purifier la L-Méthionine.
<Desc/Clms Page number 38>
Exemple 10: Production de méthionine avec une souche A(metC)
La construction de la souche A(metC) est décrite dans l'exemple 7. Dans un mode particulier de l'invention la souche E. coli A(metC) est mise en culture en flacon (voir exemple 2) contenant un milieu minimum avec glucose comme seule source de carbone. Le milieu ne contient ni methylmercaptan, ni H2S. Des repiquages sont réalisés et les taux de croissances sont déterminés pour chaque repiquage. On observe une très nette amélioration du taux de croissance de la souche A(metC) sur milieu minimum (Voir Figure n 7) suggérant que l'activité homocysteine synthase portée par la cystathionine y-synthase s'est fortement améliorée en présence d'H2S endogène.

La construction de la souche A(metC) est décrite dans l'exemple 7. Dans un mode particulier de l'invention la souche E. coli A(metC) est mise en culture en flacon (voir exemple 2) contenant un milieu minimum avec glucose comme seule source de carbone. Le milieu ne contient ni methylmercaptan, ni H2S. Des repiquages sont réalisés et les taux de croissances sont déterminés pour chaque repiquage. On observe une très nette amélioration du taux de croissance de la souche A(metC) sur milieu minimum (Voir Figure n 7) suggérant que l'activité homocysteine synthase portée par la cystathionine y-synthase s'est fortement améliorée en présence d'H2S endogène.
<tb>
<tb>
<tb>
Cycles <SEP> de <SEP> repiquage <SEP> N <SEP> mesuré <SEP> (h-1)
<tb> 1 <SEP> 0,05
<tb> 3 <SEP> 0,37
<tb> 5 <SEP> 0,39
<tb> 10 <SEP> 0,44
<tb> 12 <SEP> 0,44
<tb>
Dépôt de matériel biologique
La a Souche K183 été déposé le 02 Avril 2003 à la Collection Nationale de Cultures de Microorganismes (CNCM), 25 rue du Docteur Roux, 75724 Paris Cedex 15, France, selon les dispositions du Traité de Budapest, sous le Numéro d'ordre I- 3005.
<tb> 1 <SEP> 0,05
<tb> 3 <SEP> 0,37
<tb> 5 <SEP> 0,39
<tb> 10 <SEP> 0,44
<tb> 12 <SEP> 0,44
<tb>
Dépôt de matériel biologique
La a Souche K183 été déposé le 02 Avril 2003 à la Collection Nationale de Cultures de Microorganismes (CNCM), 25 rue du Docteur Roux, 75724 Paris Cedex 15, France, selon les dispositions du Traité de Budapest, sous le Numéro d'ordre I- 3005.
Claims (35)
1. Souche de microorganisme produisant de l'acide 2-amino-4- (alkylmercapto)butyrique de formule générale (I)
R-S-(CH2)2-CHNH2-COOH (I) dans laquelle R représente un radical alkyle linéaire ou ramifié comprenant de 1 à 18 atomes de carbones, éventuellement substitué par un ou des groupe (s) ou un radical aryle ou hétéroaryle comprenant un ou plusieurs atomes d'azote ou de soufre dans le cycle hétéroaromatique, sélectionné parmi les groupes phényle, pyridyle, pyrolyle, pyrazolyle, triazolyle, tetrazolyle, thiazolyle, ou thienyle, par métabolisme d'une source de carbone simple et d'une source de soufre comprenant un composé de formule générale (II) :
R'-SH (II) dans laquelle R' représente un atome d'hydrogène ou R, R étant défini précédemment, et ses sels physiologiquement acceptables, ladite souche comprenant au moins une un gène codant pour une enzyme à activité méthionine synthase modifiée.
2. Souche selon la revendication 1, caractérisée en ce que l'enzyme à activité méthionine synthase modifiée effectue la conversion directe d'un substrat dérivé de formule générale (III)
R"-O-(CH2)2-CHNH2-COOH (III) dans laquelle R" représente un radical acyle, de préférence choisi parmi le radical succinyle ou le radical acétyle, en acide aminé de formule générale (I).
3. Souche selon la revendication 2, caractérisée en ce que la source de soufre est un composé de formule générale (II) pour laquelle R' représente le radical
R.
4. Souche selon la revendication 1, caractérisée en ce que l'enzyme à activité méthionine synthase modifiée effectue la conversion d'un substrat dérivé de formule générale (III)
R"-O-(CH2)2-CHNH2-COOH (III) dans laquelle R" représente un radical acyle, de préférence choisi parmi le radical succinyle ou le radical acétyle, en homocystéine de formule générale (IV)
<Desc/Clms Page number 40>
HS-(CH2)2-CHNH2-COOH (IV) laquelle est ensuite transformée en acide aminé de formule générale (I) par une enzyme appropriée.
5. Souche selon la revendication 4, caractérisée en ce que la source de soufre est un composé de formule générale (II) pour laquelle R' représente un atome d'hydrogène.
6. Souche selon l'une des revendications 1 à 5, caractérisée en ce que R représente un atome d'hydrogène ou un radical alkyle linéaire ou ramifié comprenant de 1 à 4 atomes de carbone.
7. Souche selon l'une des revendications 1 à 6, caractérisée en ce l'acide 2-amino-4-(alkylmercapto)butyrique de formule générale (I) obtenu est la Lméthionine.
8. Souche selon l'une des revendications 1 à 7, caractérisée en ce que le microorganisme est choisi parmi les bactéries.
9. Souche selon la revendication 8, caractérisée en ce que la bactérie est choisie parmi E. coli et les corynébactéries.
10. Souche selon l'une des revendications 1 à 9, caractérisée en ce l'enzyme à activité méthionine synthase modifiée est choisie parmi les cystathionine-y-synthases et les acylhomosérine sulfhydrylases à activité méthionine synthase modifiée.
11. Souche selon la revendication 10, caractérisée en ce que la modification de l'enzyme consiste en une mutation de manière à ce que la transformation du substrat de formule générale (III) se fasse de manière préférentielle avec le composé de formule générale (II) plutôt qu'avec la L-cystéine.
12. Souche selon l'une des revendications 10 et 11, caractérisée en ce que la cystathionine-y-synthases à activité méthionine synthase non modifiée est sélectionnées parmi les cystathionine-y-synthases correspondant au PFAM référence PF01053 et au COG référence CPG0626.
13. Souche selon la revendication 12, caractérisée en ce que la cystathionine-y-synthase à activité méthionine synthase non modifiée est la cystathionine-y-synthase de E. coli K12, représentée sur la SEQ ID NO : 6, et les séquences homologues de cette séquence.
<Desc/Clms Page number 41>
14. Souche selon l'une des revendications 12 ou 13, caractérisée en ce que la cystathionine-y-synthase à activité méthionine synthase non modifiée, comprend la séquence d'acide aminés suivante dans sa partie C-terminale (zone conservée 1) X1-X2-X3-L-G-X4-X5-X6-X7-X8-X9
Dans laquelle
XI représente A, G,S, de préférence A
X2 représente E, V,P,T, de préférence E
X3 représente S,T,N, de préférence S
X4 représente G, D,A,H,T, de préférence G
X5 représente V, A,T,H,N, de préférence V
X6 représente E, R,K,F, de préférence E
X7 représente S, T, de préférence S
X8 représente L,I,V,A, de préférence L et
X9 représente I,V,A,T, de préférence I. correspondant aux résidus 324 à 334 de la séquence de cystathionine-ysynthase de E. coli K12, représentée sur la SEQ ID NO 6.
15. Souche selon l'une des revendications 12 à 14, caractérisée en ce que la cystathionine-y-synthase à activité méthionine synthase non modifiée, comprend la séquence d'acide aminés suivante dans sa partie N-terminale (zone conservée 2) : X10-X11-Y-X12-R-X13-X14-X15-X16-X17-X18
Dans laquelle
XI 0 représente A, H, Y,F,L,K, de préférence A
X11 représente Y, E, D,K,R,V,I, de préférence Y
XI 2 représente S, A,T,P,G, de préférence S
XI 3 représente I,S,T,R,E,F,W,D, de préférence S
XI 4 représente S, G,A,I,E,N,K,P, de préférence G
XI 5 représente N, H,Q,S, de préférence N
XI 6 représente P, D,L, de préférence P
XI 7 représente T, M,N,G,S, de préférence T et
XI 8 représente R,L,V,S,W,E, de préférence R.
<Desc/Clms Page number 42>
correspondant aux résidus 44 à 54 de la séquence de cystathionine-y-synthase de E. coli K12, représentée sur la SEQ ID NO 6.
16. Souche selon l'une des revendications 12 à 15, caractérisée en ce que la cystathionine-y-synthase modifiée comprend au moins une mutation dans sa partie C-terminale, et/ou au moins une mutation dans sa partie N-terminale.
17. Souche selon la revendication 16, caractérisée en ce que la mutation consiste à remplacer un acide aminé acide, lequel interagit avec le co-substrat cystéine pour l'enzyme non modifiée, par un acide aminé apolaire, sélectionné parmi les résidus glycine, alanine, leucine, isoleucine, valine, phénylalanine ou méthionine.
18. Souche selon la revendication 17, caractérisée en ce que la mutation dans la partie C-terminale de la cystathionine-y-synthase est introduite parmi les acides aminés acides de la zone conservée 1 telle que définie dans la revendicationl4, en particulier au niveau du résidu X2.
19. Souche selon la revendication 18, caractérisée en ce que la cystathionine-y-synthase à activité méthionine synthase modifiée comprend la séquence d'acides aminés suivante dans sa partie C-terminale : X1-X2-X3-L-G-X4-X5-X6-X7-X8-X9 dans laquelle
XI, X3, X4, X5, X6, X7, X8 et X9 sont définis ci-dessus, et
X2 représente G, A,L,I,V,F,M, de préférence A. correspondant aux résidus 324 à 334 de la séquence de cystathionine-ysynthase de E. coli K12, représentée sur la SEQ ID NO 8.
20. Souche selon la revendication 19, caractérisée en ce que la cystathionine-y-synthase à activité méthionine synthase modifiée comprend la séquence d'acides aminés suivante dans sa partie C-terminale :
A-A-S-L-G-G-V-E-S correspondant aux résidus 324 à 332 de la séquence de cystathionine-ysynthase de E. coli K12, représentée sur la SEQ ID NO 6.
21. Souche selon la revendication 20, caractérisée en ce que la cystathionine-y-synthase à activité méthionine synthase modifiée comprend la séquence d'acides aminés représentée sur la SEQ ID NO 8.
<Desc/Clms Page number 43>
22. Souche selon l'une des revendications 15 à 20, caractérisée en ce que la mutation dans la partie N-terminale de la cystathionine-y-synthase est introduite parmi les acides aminés acides de la zone conservée 2 telle que définie ci-dessus, en particulier au niveau du résidu XI 1 et/ou R et/ou XI 3.
23. Souche selon la revendication 22, caractérisé en ce que la cystathionine-y-synthase à activité méthionine synthase modifiée comprend la séquence d'acide aminés suivante dans sa partie N-terminale : X10-X11-Y-X12-X19-X13-X14-X15-X16-X17-X18 dans laquelle
X7, X9, X12, X14, X15, X16, X17 et X18 sont définis ci-dessus, XI 1 est défini dans la revendication 15 ou représente un acide aminé apolaire,
XI 3 est défini dans la revendication 15 ou représente un acide aminé apolaire,
X19 est R ou représente un acide aminé apolaire, et l'un au moins de X11, X13 et X19 représente un acide aminé apolaire, les acides aminés apolaires étant parmi choisis indépendamment parmi les résidus glycine, alanine, leucine, isoleucine, valine, phénylalanine ou méthionine.
24. Souche E. coli Kl 83 à activité méthionine synthase modifiée, déposée à la CNCM le 2 avril 2003 sous le numéro 1-3005.
25. Cystathionine-y-synthase à activité méthionine synthase modifiée telle que définie dans l'une des revendications 10 à 23.
26. Séquence d'acides nucléiques codant pour une cystathionine-ysynthase à activité méthionine synthase modifiée selon la revendication 25.
27. Vecteur de clonage et/ou d'expression comprenant une séquence d'acide nucléique selon la revendication 26, sous le contrôle d'éléments de régulation nécessaires à l'expression et la transcription de la cystathionine-y-synthase à activité méthionine synthase modifiée dans un organisme hôte.
28. Procédé de criblage d'une souche bactérienne initiale possédant un gène codant pour une enzyme cystathionine-y-synthase ou acylhomosérine sulfhydrylase, en vue d'obtenir une souche bactérienne génétiquement modifiée produisant un acide aminé de formule (I) défini selon l'une des revendications 1, 6 ou 7, et présentant une modification dans le gène de ladite enzyme, induisant une modification de l'activité méthionine synthase en présence d'un composé soufré
<Desc/Clms Page number 44>
de formule (II) défini selon l'une des revendications 1, 3, 5,6 ou 7, présentant l'étape consistant à soumettre ladite souche bactérienne à une pression de sélection en présence du composé de formule (II), afin de diriger une évolution du gène codant pour ladite enzyme cystathionine-y-synthase ou acylhomosérine sulfhydrylase, dans ladite souche bactérienne, ladite évolution consistant en une mutation pour leur permettre de réaliser de manière préférentielle la conversion directe du substrat de formule générale (III) défini selon la revendications 2, en acide aminé de formule générale (I) ou en homocystéine de formule générale (IV) défini selon la revendications 4.
29. Procédé de préparation d'un acide 2-amino-4(alkylmercapto)butyrique de formule (I) défini selon l'une des revendications 1 à 7, dans lequel on cultive un microorganisme à activité méthionine synthase modifiée défini selon l'une des revendications 1 à 24 ou obtenu par le procédé de sélection selon la revendication 28, en présence d'un composé soufré de formule générale (II) défini selon l'une des revendications 1 à 7, dans un milieu approprié comprenant une source de carbone simple.
30. Procédé selon la revendication 29, caractérisé en ce que l'acide aminé de formule (I) est la méthionine, plus préférentiellement L-méthionine.
31. Procédé selon la revendication 30, caractérisé en ce que le composé soufré de formule (II) est le méthyl-mercaptan ou l'H2S.
32. Procédé de préparation d'un acide 2-amino-4(alkylmercapto)butyrique de formule (I) défini selon l'une des revendications 1 à 7, dans lequel on fait réagir substrat dérivé de formule générale (III) R"-O-(CH2)2-CHNH2-COOH (III) dans laquelle R" représente un radical acyle, de préférence choisi parmi le radical succinyle ou le radical acétyle, avec une enzyme à activité méthionine synthase modifiée telle que définie dans l'une des revendications 1 à 23, dans un milieu réactionnel approprié comprenant un composé soufré de formule générale (II) défini dans l'une des revendications 1 à 7.
33. Procédé selon la revendication 32, caractérisé en ce que l'enzyme à activité méthionine synthase modifiée est présente dans une bactérie inactivée ou dans un extrait cellulaire.
<Desc/Clms Page number 45>
34. Procédé selon la revendication 32, caractérisé en ce que l'enzyme à activité méthionine synthase modifiée est une enzyme purifiée.
35. Procédé selon l'une des revendications 32 à 34, caractérisé en ce que l'enzyme est une cystathionine-y-synthase à activité méthionine synthase modifiée telle que définie dans l'une des revendications 10 à 23.
Priority Applications (9)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR0305768A FR2851255B1 (fr) | 2003-02-18 | 2003-05-14 | Microorganisme a activite methionine synthase modifiee et procede de preparation de la methionine |
| MXPA05008857A MXPA05008857A (es) | 2003-02-18 | 2004-02-17 | Procedimiento de preparacion de microorganismos evolucionados que permite la creacion o la modificacion de vias metabolicas. |
| EP10183261A EP2348107A3 (fr) | 2003-02-18 | 2004-02-17 | Procédé de préparation de microorganismes évolués permettant la création ou la modification de voies métaboliques |
| BRPI0407600-1A BRPI0407600A (pt) | 2003-02-18 | 2004-02-17 | processos de preparação de microorganismos evoluìdos e de uma proteìna evoluìda e de biotransformação, gene evoluìdo, proteìna evoluìda, e utilização de um microorganismo evoluìdo ou de uma proteìna evoluìda |
| PCT/FR2004/000354 WO2004076659A2 (fr) | 2003-02-18 | 2004-02-17 | Procede de preparation de microorganismes evolues permettant la creation ou la modification de voies metaboliques |
| US10/546,139 US20060270013A1 (en) | 2003-02-18 | 2004-02-17 | Method for the production of evolved microorganisms which permit the generation or modification of metabolic pathways |
| JP2006502150A JP2006517796A (ja) | 2003-02-18 | 2004-02-17 | 代謝経路の生成または改変を可能とする進化した微生物の生産方法 |
| EP04711626.4A EP1597364B1 (fr) | 2003-02-18 | 2004-02-17 | Procede de preparation de microorganismes evolues permettant la creation ou la modification de voies metaboliques |
| US10/781,499 US7745195B2 (en) | 2003-02-18 | 2004-02-18 | Method for the preparation of an evolved microorganism for the creation or the modification of metabolic pathways |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR0301924A FR2851256A1 (fr) | 2003-02-18 | 2003-02-18 | Procede de criblage et d'evolution dirigee de souches produisant de la methionine par nouvelle voie metabolique |
| FR0305768A FR2851255B1 (fr) | 2003-02-18 | 2003-05-14 | Microorganisme a activite methionine synthase modifiee et procede de preparation de la methionine |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| FR2851255A1 true FR2851255A1 (fr) | 2004-08-20 |
| FR2851255B1 FR2851255B1 (fr) | 2007-05-25 |
Family
ID=32773884
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| FR0305768A Expired - Fee Related FR2851255B1 (fr) | 2003-02-18 | 2003-05-14 | Microorganisme a activite methionine synthase modifiee et procede de preparation de la methionine |
Country Status (1)
| Country | Link |
|---|---|
| FR (1) | FR2851255B1 (fr) |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2013029690A1 (fr) * | 2011-09-02 | 2013-03-07 | Arkema France | Procédé de préparation de l-méthionine |
| EP3363908A4 (fr) * | 2015-10-13 | 2019-04-10 | CJ Cheiljedang Corporation | Variant de l'o-acétylhomosérine sulfhydrylase et procédé de production de l-méthionine l'utilisant |
| CN118374391A (zh) * | 2024-04-19 | 2024-07-23 | 杭州优泽生物科技有限公司 | 利用适应性进化筛选得到的高产o-琥珀酰-l-高丝氨酸的大肠杆菌 |
Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4071405A (en) * | 1975-06-19 | 1978-01-31 | Ajinomoto Co., Inc. | Method of producing sulfur containing L-amino acids |
| WO1993017112A1 (fr) * | 1992-02-20 | 1993-09-02 | Genencor International, Inc. | Biosynthese de methionine au moyen d'une source reduite de soufre |
| JP2000157267A (ja) * | 1998-11-24 | 2000-06-13 | Ajinomoto Co Inc | 変異型metJ遺伝子及びL−メチオニンの製造法 |
| WO2002018613A1 (fr) * | 2000-09-02 | 2002-03-07 | Degussa Ag | SEQUENCES NUCLEOTIDIQUES CODANT LE GENE metY |
| WO2002051231A1 (fr) * | 2000-12-22 | 2002-07-04 | Basf Aktiengesellschaft | Genes de corynebacterie |
-
2003
- 2003-05-14 FR FR0305768A patent/FR2851255B1/fr not_active Expired - Fee Related
Patent Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4071405A (en) * | 1975-06-19 | 1978-01-31 | Ajinomoto Co., Inc. | Method of producing sulfur containing L-amino acids |
| WO1993017112A1 (fr) * | 1992-02-20 | 1993-09-02 | Genencor International, Inc. | Biosynthese de methionine au moyen d'une source reduite de soufre |
| JP2000157267A (ja) * | 1998-11-24 | 2000-06-13 | Ajinomoto Co Inc | 変異型metJ遺伝子及びL−メチオニンの製造法 |
| WO2002018613A1 (fr) * | 2000-09-02 | 2002-03-07 | Degussa Ag | SEQUENCES NUCLEOTIDIQUES CODANT LE GENE metY |
| WO2002051231A1 (fr) * | 2000-12-22 | 2002-07-04 | Basf Aktiengesellschaft | Genes de corynebacterie |
Non-Patent Citations (8)
| Title |
|---|
| ARNOLD F H ET AL: "OPTIMIZING INDUSTRIAL ENZYMES BY DIRECTED EVOLUTION", ADVANCES IN BIOCHEMICAL ENGINEERING, BIOTECHNOLOGY, SPRINGER, BERLIN, DE, vol. 58, 1997, pages 1 - 14, XP000891392, ISSN: 0724-6145 * |
| DATABASE EMBL [online] 1 October 2001 (2001-10-01), KAWASHIMA T ET AL: "Cystathionine beta lyase / O-succinylhomoserine lyase.", XP002274157, Database accession no. Q97CC7 * |
| DATABASE EMBL [online] 13 June 1985 (1985-06-13), DUCHANGE N ET AL: "E.coli metB and metL (5' end) genes coding for cystathione gamma-synthase and aspartokinase II-homoserine dehydrogenase II.", XP002274156, Database accession no. K01546 * |
| DATABASE WPI Section Ch Week 200043, Derwent World Patents Index; Class B05, AN 2000-485354, XP002265367 * |
| FLAVIN M ET AL: "Enzymatic synthesis of homocysteine or methionine directly from O-succinyl-homoserine.", BIOCHIMICA ET BIOPHYSICA ACTA. NETHERLANDS 15 MAR 1967, vol. 132, no. 2, 15 March 1967 (1967-03-15), pages 400 - 405, XP008028828, ISSN: 0006-3002 * |
| MAY OLIVER ET AL: "Inverting enantioselectivity by directed evolution of hydantoinase for improved production of L-methionine", NATURE BIOTECHNOLOGY, NATURE PUBLISHING, US, vol. 18, no. 3, March 2000 (2000-03-01), pages 317 - 320, XP002154849, ISSN: 1087-0156 * |
| SMITH I K ET AL: "Utilization of S-methylcysteine and methylmercaptan by methionineless mutants of Neurospora and the pathway of their conversion to methionine. II. Enzyme studies.", BIOCHIMICA ET BIOPHYSICA ACTA. NETHERLANDS 17 JUN 1969, vol. 184, no. 1, 17 June 1969 (1969-06-17), pages 130 - 138, XP008028840, ISSN: 0006-3002 * |
| WEISSBACH H ET AL: "REGULATION OF METHIONINE SYNTHESIS IN ESCHERICHIA COLI", MOLECULAR MICROBIOLOGY, BLACKWELL SCIENTIFIC, OXFORD, GB, vol. 5, no. 7, 1987, pages 1593 - 1597, XP001037155, ISSN: 0950-382X * |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2013029690A1 (fr) * | 2011-09-02 | 2013-03-07 | Arkema France | Procédé de préparation de l-méthionine |
| US9365877B2 (en) | 2011-09-02 | 2016-06-14 | Cj Cheiljedang Corporation | Preparation of process of L-methionine |
| EP3363908A4 (fr) * | 2015-10-13 | 2019-04-10 | CJ Cheiljedang Corporation | Variant de l'o-acétylhomosérine sulfhydrylase et procédé de production de l-méthionine l'utilisant |
| CN118374391A (zh) * | 2024-04-19 | 2024-07-23 | 杭州优泽生物科技有限公司 | 利用适应性进化筛选得到的高产o-琥珀酰-l-高丝氨酸的大肠杆菌 |
Also Published As
| Publication number | Publication date |
|---|---|
| FR2851255B1 (fr) | 2007-05-25 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| EP1597364B1 (fr) | Procede de preparation de microorganismes evolues permettant la creation ou la modification de voies metaboliques | |
| JP5497628B2 (ja) | 組成物およびメチオニン生産方法 | |
| EP1704230B1 (fr) | Microorganisme evolue pour la production de 1,2-propanediol | |
| TWI654305B (zh) | 生產o-磷絲胺酸之微生物及使用該微生物生產o-磷絲胺酸或l-半胱胺酸之方法 | |
| US20060148041A1 (en) | Method for fermentative production of amino acids and amino acid derivatives of the phosphoglycerate family | |
| CN106906176A (zh) | 提高nadph可用性用于甲硫氨酸产生 | |
| JP2008529485A (ja) | 低いγ脱離活性を有する発現酵素を含む微生物 | |
| KR20160044038A (ko) | 메티오닌 유출이 증진된 메티오닌 생산용 미생물 | |
| EP3039121A1 (fr) | Micro-organisme modifié pour production améliorée d'alanine | |
| JP2019523271A (ja) | N−アセチルホモセリン | |
| KR20140135622A (ko) | 발효에 의한 감마-글루타밀시스테인 및 이 디펩티드의 유도체의 과다 생산을 위한 미생물 및 방법 | |
| RU2723714C2 (ru) | Способ и микроорганизм для ферментативного продуцирования метионина с улучшенным выходом метионина | |
| WO2024223833A1 (fr) | Micro-organisme génétiquement modifié pour la production d'ectoïne | |
| JP2009501547A (ja) | 微生物におけるメチオニン生産を改善するためのバチルス属metI遺伝子の使用 | |
| FR2851255A1 (fr) | Microorganisme a activite methionine synthase modifiee et procede de preparation de la methionine | |
| KR101770150B1 (ko) | 메티오닌 히드록시 유사체 (mha)의 발효 생산 | |
| KR101214632B1 (ko) | 타우린 생성용 재조합 변이 미생물 및 이를 이용한 타우린의 제조방법 | |
| JP2020503053A (ja) | O−ホスホセリンを生産するエシェリキア属微生物及びこれを用いたo−ホスホセリンまたはl−システインを生産する方法 | |
| FR2851256A1 (fr) | Procede de criblage et d'evolution dirigee de souches produisant de la methionine par nouvelle voie metabolique | |
| FR2862067A1 (fr) | Procede de preparation de microorganismes evolues permettant la creation ou la modification de voies metaboliques | |
| FR2854902A1 (fr) | Microorganisme a activite cysteine synthase modifiee et procede de preparation de la cysteine | |
| CN103710319B (zh) | 组合物及甲硫氨酸的生产方法 | |
| EP1137784A1 (fr) | Methode d'isolement et de selection de genes codant pour des enzymes, et milieu de culture approprie | |
| WO2024162450A1 (fr) | Procédé de production d'acide l-glutamique | |
| CN102703398A (zh) | 组合物及甲硫氨酸的生产方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| AS | Court action brought to claim ownership of the patent | ||
| ST | Notification of lapse |
Effective date: 20160129 |