JP2004201677A - METHOD FOR PRODUCING gamma-GLUTAMYLCYCTEINE - Google Patents
METHOD FOR PRODUCING gamma-GLUTAMYLCYCTEINE Download PDFInfo
- Publication number
- JP2004201677A JP2004201677A JP2003399198A JP2003399198A JP2004201677A JP 2004201677 A JP2004201677 A JP 2004201677A JP 2003399198 A JP2003399198 A JP 2003399198A JP 2003399198 A JP2003399198 A JP 2003399198A JP 2004201677 A JP2004201677 A JP 2004201677A
- Authority
- JP
- Japan
- Prior art keywords
- yeast
- glutamylcysteine
- gene
- strain
- culture
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images



Landscapes
- Seasonings (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Alcoholic Beverages (AREA)
- Coloring Foods And Improving Nutritive Qualities (AREA)
- Bakery Products And Manufacturing Methods Therefor (AREA)
Abstract
Description
本発明は、γ−グルタミルシステインを生産する酵母菌株とその培養方法、及び同菌株の菌体を利用した飲食品に関するものである。γ−グルタミルシステインを含む素材及びそれから製造されるシステインを含む素材は、食品分野で有用である。 TECHNICAL FIELD The present invention relates to a yeast strain producing γ-glutamylcysteine, a method for culturing the yeast strain, and a food or drink using cells of the yeast strain. Materials containing γ-glutamylcysteine and materials containing cysteine produced therefrom are useful in the food field.
システインは、食品の風味改善などを目的に用いられている。システインの製法については蛋白分解法や半合成法などが知られているが、現在主に用いられている方法は蛋白分解法と半合成法である。システインを食品の風味改善に用いることを目的として、システイン含量の高い天然食品素材が求められているが、そのような天然食品素材は従来ほとんど知られていなかった。一方、γ−グルタミルシステインを含む酵母エキスを加熱または酵素処理すれば、システインを高含有する食品素材を得ることが可能であると報告されている(特許文献1)。 Cysteine is used for the purpose of improving the flavor of foods and the like. As for the production method of cysteine, a proteolysis method and a semi-synthesis method are known, but currently mainly used methods are a proteolysis method and a semi-synthesis method. For the purpose of using cysteine to improve the flavor of foods, natural food materials having a high cysteine content have been demanded, but such natural food materials have hitherto hardly been known. On the other hand, it has been reported that a food material having a high cysteine content can be obtained by heating or enzymatically treating a yeast extract containing γ-glutamylcysteine (Patent Document 1).
γ−グルタミルシステインは、γ−グルタミルシステイン合成酵素の働きによりシステインとグルタミン酸を基質にして合成される。また、グルタチオンはグルタチオン合成酵素の働きによりγ−グルタミルシステインとグリシンを基質にして合成される。グルタチオン合成酵素遺伝子を破壊した酵母は、γ−グルタミルシステインを蓄積することが報告されている(非特許文献1)。 γ-glutamylcysteine is synthesized using cysteine and glutamic acid as substrates by the action of γ-glutamylcysteine synthase. Glutathione is synthesized using γ-glutamylcysteine and glycine as substrates by the action of glutathione synthase. It has been reported that yeast in which the glutathione synthase gene has been disrupted accumulates γ-glutamylcysteine (Non-Patent Document 1).
これまでに、γ−グルタミルシステインを高含有する酵母は、特許文献1、非特許文献1、非特許文献2及び非特許文献3などで報告されている。しかしこれらの報告には、グルタチオン合成酵素遺伝子を破壊または弱化させた酵母を用いてγ−グルタミルシステインを高蓄積させるための培養条件に関する記載はない。 So far, yeasts containing high content of γ-glutamylcysteine have been reported in Patent Document 1, Non-patent Document 1, Non-Patent Document 2, Non-Patent Document 3, and the like. However, there is no description in these reports about culture conditions for highly accumulating γ-glutamylcysteine using yeast in which the glutathione synthase gene has been disrupted or weakened.
ところで、γ−グルタミルシステインの代謝産物であるグルタチオンを酵母に高蓄積させる培養方法に関する技術が開示されている(特許文献2等)。この報告には、酵母の培養途中にグルタチオンの構成アミノ酸であるシステインを添加するとグルタチオン蓄積量が上昇すると記載されている。その為、グルタチオン合成酵素遺伝子を破壊または弱化させた酵母の培養途中にシステインを添加して培養すればγ−グルタミルシステインを高蓄積できると考えられる。しかしながら、γ−グルタミルシステインを含む素材はシステインを含む素材を製造する目的で有用であるため、システイン含有素材を得る目的でγ−グルタミルシステインを含有する酵母の培養途中にシステインを添加することは、コスト的見地からは非実用的な方法である。 By the way, a technique relating to a culture method for highly accumulating glutathione, a metabolite of γ-glutamylcysteine, in yeast is disclosed (Patent Document 2 and the like). This report states that addition of cysteine, a constituent amino acid of glutathione, during the culture of yeast increases glutathione accumulation. Therefore, it is considered that γ-glutamylcysteine can be highly accumulated by adding cysteine during the cultivation of yeast in which the glutathione synthase gene has been destroyed or weakened. However, since a material containing γ-glutamylcysteine is useful for the purpose of producing a material containing cysteine, adding cysteine during the culturing of a yeast containing γ-glutamylcysteine for the purpose of obtaining a cysteine-containing material, This is impractical from a cost standpoint.
さらに、大竹らは、グルタチオン合成酵素遺伝子が破壊された酵母YL1株の培養途中に3mMのシステインを添加した時の酵母菌体中のγ−グルタミルシステイン含有量を報告している(非特許文献1)。この報告では、YL1株にシステインを添加して培養した場合のγ−グルタミルシステイン蓄積量は0.533%であり、システインを添加しないで培養した場合のγ−グルタミルシステイン含有量0.518%であったと記載されている。この結果からも、グルタチオン合成酵素遺伝子を破壊または弱化させた酵母の培養途中にシステインを添加して培養することは非実用的な方法であると考えられる。 Further, Otake et al. Report the γ-glutamylcysteine content in yeast cells when 3 mM cysteine was added during the culture of yeast YL1 strain in which the glutathione synthase gene was disrupted (Non-patent Document 1). ). In this report, the γ-glutamylcysteine accumulation when cysteine was added to the YL1 strain was 0.533%, and the γ-glutamylcysteine content when cultured without cysteine was 0.518%. ing. From these results, it is considered that it is an impractical method to add cysteine during the cultivation of the yeast in which the glutathione synthase gene has been destroyed or weakened, and culture the yeast.
また、酵母におけるMET25遺伝子の発現量を増大させると菌体内グルタチオン含有量が上昇することが報告されている。さらに、MET25遺伝子の発現量を増大させる方法については変異型MET4遺伝子(非特許文献4、特許文献3)、変異型MET30遺伝子(非特許文献5)を利用する方法等が報告されている。 It has also been reported that increasing the expression level of the MET25 gene in yeast increases the intracellular glutathione content. Furthermore, methods for increasing the expression level of the MET25 gene include a method using a mutant MET4 gene (Non-patent Document 4, Patent Document 3) and a method using a mutant MET30 gene (Non-patent Document 5).
MET25遺伝子の発現については、次のように考えられている(非特許文献6、非特許文献7)。MET4産物は、MET25遺伝子に発現に正の因子として働く。通常は、MET4産物は、MET30産物及び他の数種のタンパク質とともにSCFMET30複合体を形成し、ユビキチン化を受け、26Sプロテアソームによるタンパク分解系によりMET30産物とともに分解されるため、MET25遺伝子の発現は抑制されている。一方、SCFMET30複合体の機能が低下すると、MET4産物及びMET30産物は分解を受けず、MET25遺伝子が発現する。 The expression of the MET25 gene is considered as follows (Non-Patent Document 6, Non-Patent Document 7). The MET4 product acts as a positive factor on expression in the MET25 gene. Normally, the MET4 product forms an SCFMET30 complex with the MET30 product and several other proteins, undergoes ubiquitination, and is degraded together with the MET30 product by the 26S proteasome proteolytic system, thus suppressing the expression of the MET25 gene. Have been. On the other hand, when the function of the SCFMET30 complex is reduced, the MET4 product and the MET30 product are not degraded, and the MET25 gene is expressed.
上記の知見から、γ−グルタミルシステインを高含有する酵母においても、MET25遺伝子の発現量を増大させることにより、γ−グルタミルシステイン含有量が増大することが予測される。 From the above findings, it is predicted that, even in yeast having a high content of γ-glutamylcysteine, increasing the expression level of the MET25 gene will increase the γ-glutamylcysteine content.
また、清酒酵母における研究から、清酒酵母をパントテン酸カルシウム欠乏状態で培養すると、その対数増殖期に硫化水素を蓄積するとの報告がある。この報告では、システインからの硫化水素の発生に着目しており、この現象はパントテン酸欠乏状態でより促進されると記載されている。 In addition, studies on sake yeast have reported that when sake yeast is cultured in a calcium pantothenate-deficient state, hydrogen sulfide accumulates during its logarithmic growth phase. This report focuses on the generation of hydrogen sulfide from cysteine, and states that this phenomenon is more accelerated in pantothenic acid deficiency.
本発明は、上記の技術背景の下に、γ−グルタミルシステイン生産に適した酵母及びそれを用いたγ−グルタミルシステインの製造方法、並びに、同酵母菌株を利用したγ−グルタミルシステイン含有飲食品を提供することを課題とする。 The present invention provides a yeast suitable for γ-glutamylcysteine production and a method for producing γ-glutamylcysteine using the same, and γ-glutamylcysteine-containing food and drink using the yeast strain, under the above technical background. The task is to provide.
本発明者らは、上記課題を解決するために鋭意検討を行い、酵母のγ−グルタミルシステインの蓄積量が培養中に一定である必要はなく、集菌直前に必要量のγ−グルタミルシステインを含有していればよいと考えた。そして、γ−グルタミルシステイン産生能を有し、かつパントテン酸を要求する酵母を、その酵母が必要とするパントテン酸を必要最小限度添加して培養し、十分量の菌体含量を得た後、パントテン酸を欠乏した状態で培養す
ることにより、γ−グルタミルシステインの蓄積量が上昇することを見いだし、本発明を完成するに至った。
すなわち、本発明は以下のとおりである。
The present inventors have conducted intensive studies in order to solve the above-mentioned problems, and the accumulated amount of γ-glutamylcysteine in yeast does not need to be constant during culture, and the required amount of γ-glutamylcysteine is obtained immediately before collection. We thought that it should be contained. And yeast having a γ-glutamylcysteine-producing ability, and requesting pantothenic acid, and culturing by adding the necessary minimum amount of pantothenic acid required by the yeast to obtain a sufficient amount of bacterial cells, It has been found that culturing in a state deficient in pantothenic acid results in an increase in the amount of accumulated γ-glutamylcysteine, thereby completing the present invention.
That is, the present invention is as follows.
(1)γ−グルタミルシステイン生産能を有し、パントテン酸要求性を有し、かつ、パントテン酸量が制限された培地で培養したとき、乾燥酵母菌体当たりのγ−グルタミルシステイン含有量が経時的に上昇する酵母。
(2)細胞内のグルタチオン合成酵素活性が低下又は消失するように改変された(1)に記載の酵母。
(3)MET25遺伝子の発現が脱抑制されるように改変された(1)又は(2)に記載の酵母。
(4)コードされる蛋白質の569位のセリンがセリン以外の他のアミノ酸に置換する変異を有する変異型MET30遺伝子を保持することにより、MET25遺伝子の発現が脱抑制された(3)に記載の酵母。
(5)前記他のアミノ酸がフェニルアラニンである(4)に記載の酵母。
(6)サッカロマイセス属に属する(1)〜(5)のいずれかに記載の酵母。
(7)(1)〜(6)のいずれかに記載の酵母を、十分な量のパントテン酸を含む培地で培養して同酵母を増殖させる工程と、パントテン酸量が制限された培地で培養し、菌体内のγ−グルタミルシステイン含有量を上昇させる工程を含む、γ−グルタミルシステインを蓄積した酵母の製造方法。
(8)(1)〜(6)のいずれかに記載の酵母を好適な条件で培養して得られる培養物、もしくはγ−グルタミルシステインを含む前記培養物の分画物、又は熱処理によってシステインが生成したこれらの培養物もしくは分画物を含む飲食品。
(9)飲食品がアルコール飲料、パン食品、又は発酵食品調味料である(8)記載の飲食品。
(10)(1)〜(6)のいずれかに記載の酵母を好適な条件で培養して得られる培養物を用いて製造された酵母エキス。
(11)(1)〜(6)のいずれかに記載の酵母を好適な条件で培養して得られる培養物もしくはその分画物、又は加熱処理した前記培養物又は分画物を、飲食品原料に混合し、飲食品に加工することを特徴とする、γ−グルタミルシステイン又はシステイン含有飲食品の製造法。
(12)コードされる蛋白質の569位のセリンがフェニルアラニンに置換された変異を有する変異型MET30遺伝子を保持することにより、MET25遺伝子の発現が脱抑制された酵母。
(1) When cultured in a medium having a γ-glutamylcysteine-producing ability, having a requirement for pantothenic acid, and having a restricted amount of pantothenic acid, the γ-glutamylcysteine content per dry yeast cell increased over time. Yeast that rises.
(2) The yeast according to (1), wherein the yeast is modified so that intracellular glutathione synthetase activity is reduced or eliminated.
(3) The yeast according to (1) or (2), which has been modified so that expression of the MET25 gene is derepressed.
(4) The expression of the MET25 gene was derepressed by retaining a mutant MET30 gene having a mutation in which serine at position 569 of the encoded protein was substituted with another amino acid other than serine, thereby suppressing the expression of the MET25 gene. yeast.
(5) The yeast according to (4), wherein the other amino acid is phenylalanine.
(6) The yeast according to any one of (1) to (5), which belongs to the genus Saccharomyces.
(7) a step of culturing the yeast according to any one of (1) to (6) in a medium containing a sufficient amount of pantothenic acid to grow the yeast, and culturing the yeast in a medium in which the amount of pantothenic acid is restricted; And a method for producing a yeast in which γ-glutamylcysteine is accumulated, comprising a step of increasing the content of γ-glutamylcysteine in the cells.
(8) A culture obtained by culturing the yeast according to any of (1) to (6) under suitable conditions, a fraction of the culture containing γ-glutamylcysteine, or cysteine by heat treatment. A food or drink containing the produced culture or fraction thereof.
(9) The food or drink according to (8), wherein the food or drink is an alcoholic beverage, a bread food, or a fermented food seasoning.
(10) A yeast extract produced using a culture obtained by culturing the yeast according to any one of (1) to (6) under suitable conditions.
(11) A culture obtained by culturing the yeast according to any one of (1) to (6) under suitable conditions, or a fraction thereof, or the heat-treated culture or fraction, and A method for producing γ-glutamylcysteine or cysteine-containing food or drink, which is mixed with a raw material and processed into a food or drink.
(12) A yeast in which expression of the MET25 gene is desuppressed by retaining a mutant MET30 gene having a mutation in which serine at position 569 of the encoded protein is substituted with phenylalanine.
以下、本発明を詳細に説明する。 Hereinafter, the present invention will be described in detail.
<1>本発明の酵母
本発明の酵母は、γ−グルタミルシステイン生産能を有し、かつ、パントテン酸要求性を有する酵母である。本発明の酵母は、好ましくは、パントテン酸量が制限された培地で培養したとき、乾燥酵母菌体当たりのγ−グルタミルシステイン含有量が経時的に上昇する。
<1> Yeast of the present invention The yeast of the present invention is a yeast having a γ-glutamylcysteine-producing ability and having a requirement for pantothenic acid. Preferably, when the yeast of the present invention is cultured in a medium in which the amount of pantothenic acid is restricted, the γ-glutamylcysteine content per dry yeast cell increases with time.
本発明において、「γ−グルタミルシステイン生産能を有する」とは、酵母野生株よりも多量のγ−グルタミルシステインを菌体内に蓄積することをいい、好ましくは、酵母を十分な量のパントテン酸を含む培地で培養した後、パントテン酸量が制限された培地で培養したとき、乾燥酵母菌体当たり1%以上のγ−グルタミルシステインを蓄積することをいう。より好ましくは、さらに、乾燥酵母菌体当たりのグルタチオン蓄積量が0.1%以下であることをいう。 In the present invention, "having γ-glutamylcysteine-producing ability" refers to accumulating a larger amount of γ-glutamylcysteine in a bacterial cell than a yeast wild-type strain, and preferably, a yeast having a sufficient amount of pantothenic acid. This means that when cultured in a medium containing a limited amount of pantothenic acid after culturing in a culture medium containing 1% or more of γ-glutamylcysteine per dry yeast cell. More preferably, it means that the amount of accumulated glutathione per dry yeast cell is 0.1% or less.
乾燥酵母菌体当たりのγ−グルタミルシステイン又はグルタチオンの蓄積量は、例えば、105℃で4時間加熱した後の菌体重量に対するγ−グルタミルシステイン又はグルタチオンの含有量(%)をいう。 The accumulated amount of γ-glutamylcysteine or glutathione per dry yeast cell refers to, for example, the content (%) of γ-glutamylcysteine or glutathione with respect to the cell weight after heating at 105 ° C. for 4 hours.
γ−グルタミルシステイン生産能を有する酵母としては、例えば、細胞内のグルタチオン合成酵素活性が低下又は消失した酵母、もしくは、γ−グルタミルシステイン合成酵素活性が増強されるように改変された酵母、又は、細胞内のグルタチオン合成酵素活性が低下又は消失し、かつ、γ−グルタミルシステイン合成酵素活性が増強されるように改変された酵母が挙げられる。 Examples of yeast having γ-glutamylcysteine-producing ability include, for example, yeast in which intracellular glutathione synthetase activity has been reduced or disappeared, or yeast modified so that γ-glutamylcysteine synthetase activity has been enhanced, or A yeast modified so that intracellular glutathione synthetase activity decreases or disappears and γ-glutamylcysteine synthetase activity is enhanced is exemplified.
細胞内のグルタチオン合成酵素活性が低下又は消失した酵母は、例えば、グルタチオン合成酵素をコードする遺伝子(GSH2)の部分配列を欠失し、正常に機能する酵素を産生しないように改変した遺伝子、又は酵素活性が低下するような変異を有する遺伝子(以下、単に「変異型GSH2遺伝子」という)を含むDNAを用いた遺伝子置換により創製することができる。また、細胞内のグルタチオン合成酵素活性が低下又は消失した酵母は、実施例に記載したのと同様に、野生型酵母を、通常の変異処理、例えばUV照射、あるいはN−メチル−N−ニトロソグアニジン(NTG)、エチルメタンスルホネート(EMS)、亜硝酸、アクリジン等の変異剤による処理によって、取得することもできる。得られた変異株が目的の変異を有していることは、例えばPCR法等により確認することができる。 A yeast in which the intracellular glutathione synthetase activity is reduced or eliminated is, for example, a gene modified so that the partial sequence of the gene encoding glutathione synthase (GSH2) is deleted and a normally functioning enzyme is not produced, or It can be created by gene replacement using DNA containing a gene having a mutation that reduces the enzyme activity (hereinafter, simply referred to as “mutant GSH2 gene”). In addition, yeast in which the intracellular glutathione synthetase activity has been reduced or eliminated, as described in the Examples, wild-type yeast can be subjected to ordinary mutation treatment, for example, UV irradiation, or N-methyl-N-nitrosoguanidine. (NTG), ethyl methanesulfonate (EMS), nitrite, acridine or the like, and can be obtained by treatment with a mutagen. Whether the obtained mutant has the desired mutation can be confirmed by, for example, PCR.
前記のようなグルタチオン合成酵素活性が低下するような変異としては、例えば、370位のアルギニン残基を終止コドンに変更する変異が挙げられる。 Examples of the mutation that reduces glutathione synthase activity as described above include a mutation that changes an arginine residue at position 370 to a stop codon.
また、グルタチオン合成酵素活性が低下する他の変異としては以下の変異が挙げられる(国際公開03/046155号パンフレット参照)。
(1)47位のスレオニン残基をイソロイシン残基に置換する変異。
(2)387位のグリシン残基をアスパラギン酸残基に置換する変異。
(3)54位のプロリン残基がロイシン残基に置換する変異。
上記変異は、単独でも、任意の組合せでもよいが、(1)と(3)の組合せ、及び(2)と(3)の組合せが好ましい。
Other mutations that reduce glutathione synthase activity include the following mutations (see WO 03/046155).
(1) A mutation in which the threonine residue at position 47 is replaced with an isoleucine residue.
(2) A mutation in which the glycine residue at position 387 is replaced with an aspartic acid residue.
(3) A mutation in which the proline residue at position 54 is replaced with a leucine residue.
The above mutations may be used alone or in any combination, but a combination of (1) and (3) and a combination of (2) and (3) are preferred.
グルタチオン合成酵素遺伝子への目的の変異の導入は、合成オリゴヌクレオチドを用いた部位特異的変異法によって行うことができる。 Introduction of the desired mutation into the glutathione synthase gene can be performed by site-directed mutagenesis using a synthetic oligonucleotide.
前記遺伝子置換は、以下のようにして行うことができる。変異型GSH2遺伝子を含む組換えDNAで酵母を形質転換し、変異型GSH2遺伝子と染色体上のGSH2遺伝子との間で組換えを起こさせる。その際、組換えDNAには、宿主の栄養要求性等の形質にしたがって、マーカー遺伝子を含ませておくと操作がしやすい。また、前記組換えDNAは、制限酵素で切断する等により直鎖状にし、さらに、酵母で機能する複製制御領域を除いておくと、染色体に組換えDNAが組み込まれた株を効率よく取得することができる。 The gene replacement can be performed as follows. Yeast is transformed with the recombinant DNA containing the mutant GSH2 gene to cause recombination between the mutant GSH2 gene and the GSH2 gene on the chromosome. At that time, if the recombinant DNA contains a marker gene according to the traits such as auxotrophy of the host, the operation is easy. In addition, the recombinant DNA is made linear by cutting with a restriction enzyme or the like, and further, if a replication control region that functions in yeast is removed, a strain in which the recombinant DNA is integrated into the chromosome can be efficiently obtained. be able to.
酵母の形質転換は、プロトプラスト法、KU法、KUR法、エレクトロポレーション法等、通常酵母の形質転換に用いられる方法を採用することができる。
上記のようにして染色体に組換えDNAが組み込まれた株は、変異型GSH2遺伝子と染色体上にもともと存在するGSH2遺伝子との組換えを起こし、野生型GSH2遺伝子と変異型GSH2遺伝子との融合遺伝子2個が組換えDNAの他の部分(ベクター部分及びマーカー遺伝子)を挟んだ状態で染色体に挿入されている。
For the transformation of yeast, a method usually used for transformation of yeast, such as a protoplast method, a KU method, a KUR method, and an electroporation method, can be employed.
The strain in which the recombinant DNA has been integrated into the chromosome as described above undergoes recombination between the mutant GSH2 gene and the GSH2 gene originally present on the chromosome, and is a fusion gene of the wild-type GSH2 gene and the mutant GSH2 gene. Two are inserted into the chromosome with the other part of the recombinant DNA (vector part and marker gene) sandwiched.
融合遺伝子2個のうち、変異型GSH2遺伝子のみを残すために、2個のGSH2遺伝子の組換
えにより1コピーのGSH2遺伝子を、ベクター部分(マーカー遺伝子を含む)とともに染色体DNAから脱落させる。その際、野生型GSH2遺伝子が染色体DNA上に残され、変異型GSH2遺伝子が切り出される場合と、反対に変異型GSH2遺伝子が染色体DNA上に残され、野生型GSH2遺伝子が切り出される場合がある。いずれの場合もマーカー遺伝子が脱落するので、2回目の組換えが生じたことは、マーカー遺伝子に対応する表現形質によって確認することができる。また、目的とする遺伝子置換株は、PCRによりGSH2遺伝子を増幅し、その構造を調べることによって、選択することができる。
In order to leave only the mutant GSH2 gene among the two fusion genes, one copy of the GSH2 gene is dropped from the chromosomal DNA together with the vector portion (including the marker gene) by recombination of the two GSH2 genes. At that time, there are cases where the wild-type GSH2 gene is left on the chromosomal DNA and the mutant GSH2 gene is cut out, and conversely, where the mutant GSH2 gene is left on the chromosomal DNA and the wild-type GSH2 gene is cut out. In any case, since the marker gene is dropped, the occurrence of the second recombination can be confirmed by the phenotype corresponding to the marker gene. The target gene-replacement strain can be selected by amplifying the GSH2 gene by PCR and examining its structure.
尚、遺伝子置換に用いる変異型GSH2遺伝子は、グルタチオン合成酵素タンパク質全長をコードするものであってもよいが、欠失部位を含む限り、タンパク質の一部をコードする遺伝子断片であってもよい。 The mutant GSH2 gene used for gene replacement may encode the full-length glutathione synthase protein, or may be a gene fragment encoding a part of the protein as long as it contains a deletion site.
サッカロマイセス・セレビシエのグルタチオン合成酵素遺伝子(GSH2)は塩基配列が報告されており(Inoue et al., Biochim. Biophys. Acta, 1395 (1998) 315-320、GenBank
accession Y13804、配列番号1)、同塩基配列に基づいて作製したオリゴヌクレオチドをプライマーとするPCRにより、サッカロマイセス・セレビシエ染色体DNAから取得することができる。また、導入する遺伝子は、サッカロマイセス属以外の微生物に由来する遺伝子を用いることができる。
The nucleotide sequence of the glutathione synthase gene (GSH2) of Saccharomyces cerevisiae has been reported (Inoue et al., Biochim. Biophys. Acta, 1395 (1998) 315-320, GenBank
accession Y13804, SEQ ID NO: 1), and can be obtained from Saccharomyces cerevisiae chromosomal DNA by PCR using oligonucleotides prepared based on the same base sequence as primers. In addition, a gene derived from a microorganism other than Saccharomyces can be used as the gene to be introduced.
本発明に用いる変異型GSH2遺伝子は、前記47位、387位、54位の変異以外に、配列番号2に示すアミノ酸配列において、1又は数箇所の位置における1又は数個のアミノ酸の置換、欠失、挿入又は付加を含むアミノ酸配列を有するGSH2産物をコードするものであってもよい。ここで、「数個」とは、アミノ酸残基のタンパク質の立体構造における位置や種類によっても異なるが、具体的には例えば、前記「数個」は、2〜10個、好ましくは、2〜6個、より好ましくは2〜3個である。あるいは、変異型GSH2遺伝子は、GSH2産物のアミノ酸配列全体に対し、30〜40%以上、好ましくは55〜65%以上の相同性を有するタンパク質をコードするDNAであってもよい。 In the mutant GSH2 gene used in the present invention, in addition to the mutation at positions 47, 387, and 54, substitution or deletion of one or several amino acids at one or several positions in the amino acid sequence shown in SEQ ID NO: 2 It may encode a GSH2 product having an amino acid sequence containing a deletion, insertion or addition. Here, “several” differs depending on the position and type of the amino acid residue in the three-dimensional structure of the protein, but specifically, for example, the “several” is 2 to 10, preferably 2 to Six, more preferably two to three. Alternatively, the mutant GSH2 gene may be a DNA encoding a protein having 30 to 40% or more, preferably 55 to 65% or more homology to the entire amino acid sequence of the GSH2 product.
また、上記のような塩基の置換、欠失、挿入、付加、又は逆位等には、GSH2遺伝子を保持する微生物の個体差、種や属の違いに基づく場合などの天然に生じる変異(mutant又はvariant)も含まれる。 In addition, substitution, deletion, insertion, addition, or inversion of the base as described above includes mutations that occur naturally (mutant) such as those based on individual differences, species, and genera of microorganisms that carry the GSH2 gene. Or variant) is also included.
尚、GSH2遺伝子を破壊する場合は、遺伝子置換に用いる変異型GSH2遺伝子は必ずしも全長を含む必要はなく、遺伝子破壊を起こすのに必要な長さを有していればよい。また、GSH2遺伝子の取得に用いる微生物は、同遺伝子が、遺伝子破壊株の創製に用いる微生物の相同遺伝子と相同組換えを起こす程度の相同性を有していれば特に制限されない。 When the GSH2 gene is disrupted, the mutant GSH2 gene used for gene replacement does not necessarily need to include the full length, but may have a length necessary to cause gene disruption. The microorganism used for obtaining the GSH2 gene is not particularly limited as long as the gene has homology to homologous recombination with a homologous gene of the microorganism used for creating a gene-disrupted strain.
前記サッカロマイセス・セレビシエのGSH2遺伝子と相同組換えを起こし得るDNAとしては具体的には、配列番号1に示す塩基配列を有するDNAと、例えば70%以上、好ましくは80%以上、より好ましくは90%以上の相同性を有するDNAが挙げられる。より具体的には、配列番号1に示す塩基配列を有するDNAと、ストリンジェントな条件下でハイブリダイズするDNAが挙げられる。ストリンジェントな条件としては、60℃、1×SSC,0.1%SDS、好ましくは、0.1×SSC、0.1%SDSに相当する塩濃度で洗浄が行われる条件が挙げられる。 Specific examples of the DNA capable of causing homologous recombination with the GSH2 gene of Saccharomyces cerevisiae include a DNA having the nucleotide sequence shown in SEQ ID NO: 1, for example, 70% or more, preferably 80% or more, more preferably 90% or more. DNAs having the above homology are exemplified. More specifically, it includes a DNA having the nucleotide sequence shown in SEQ ID NO: 1 and a DNA that hybridizes under stringent conditions. Stringent conditions include conditions where washing is performed at a salt concentration corresponding to 60 ° C., 1 × SSC, 0.1% SDS, preferably 0.1 × SSC, 0.1% SDS.
また、グルタチオン合成酵素活性が低下又は消失した酵母は、実施例に記載したのと同様に、野生型酵母を、通常の変異処理、例えばUV照射、あるいはN−メチル−N−ニトロソグアニジン(NTG)、エチルメタンスルホネート(EMS)、亜硝酸、アクリジン等の変異剤による処理によって、取得することもできる。 Further, the yeast in which the glutathione synthetase activity has been reduced or eliminated can be obtained by subjecting a wild-type yeast to a normal mutation treatment, for example, UV irradiation, or N-methyl-N-nitrosoguanidine (NTG), as described in Examples. , Ethyl methanesulfonate (EMS), nitrous acid, acridine and the like.
また、酵母細胞内のγ−グルタミルシステイン合成酵素活性を増強する方法としては、同酵素遺伝子(例えば、Saccharomyces cerevisiaeのγ−グルタミルシステイン合成酵素遺伝子: GenBank Accession No. D90220)を挿入したプラスミドで酵母を形質転換し、同遺伝子の細胞内のコピー数を高めるか、染色体上のγ−グルタミルシステイン合成酵素遺伝子のプロモーターを強転写プロモーターで置換する方法(大竹康之ら、バイオサイエンスとインダストリー、第50巻第10号、第989〜994頁、1992年)が挙げられる。 In addition, as a method for enhancing γ-glutamylcysteine synthase activity in yeast cells, a yeast having a plasmid into which the same enzyme gene (for example, γ-glutamylcysteine synthase gene of Saccharomyces cerevisiae: GenBank Accession No. D90220) is inserted. Transformation to increase the intracellular copy number of the gene or to replace the γ-glutamylcysteine synthetase gene promoter on the chromosome with a strong transcription promoter (Yasuyuki Otake et al., Bioscience and Industry, Vol. 50, No. 10, No. 989-994, 1992).
細胞内のγ−グルタミルシステイン合成酵素活性及びグルタチオン合成酵素性は、それぞれ、Jacksonの方法(Jackson, R.C., Biochem. J., 111, 309 (1969))、及びGushimaらの方法(Gushima, T. et al., J. Appl. Biochem., 5, 210 (1983))によって測定することができる。 The intracellular γ-glutamylcysteine synthetase activity and glutathione synthase activity were measured by the method of Jackson (Jackson, RC, Biochem. J., 111, 309 (1969)) and the method of Gushima et al. et al., J. Appl. Biochem., 5, 210 (1983)).
本発明の酵母は、上記のようなγ−グルタミルシステイン生産能を有し、かつ、パントテン酸要求性を有する。本発明において「パントテン酸要求性」とは、酵母の非改変株、例えば野生株が増殖できるのに必要なパントテン酸の濃度よりも高い濃度のパントテン酸を増殖に必要とすることをいう。 The yeast of the present invention has the ability to produce γ-glutamylcysteine as described above, and has a requirement for pantothenic acid. In the present invention, “requiring pantothenic acid” means that a non-modified strain of yeast, for example, a wild strain, requires a higher concentration of pantothenic acid for growth than that required for growth.
パントテン酸要求性を有する酵母変異株は、変異処理を行った酵母を、パントテン酸を含む培地及び含まない培地にレプリカし、パントテン酸を含まない培地でコロニーを形成せず、パントテン酸を含む培地でコロニーを形成する株を選択することによって、取得することができる。また、変異処理を行った酵母をパントテン酸を含まず、かつ、増殖する細胞に選択的に作用する抗生物質、例えばナイスタチンを添加した培地で培養することによって、パントテン酸要求性株を濃縮することができる。 A yeast mutant strain having a pantothenic acid auxotrophy, the yeast subjected to the mutation treatment is replicated in a medium containing and not containing pantothenic acid, and does not form a colony in a medium containing no pantothenic acid, and contains a medium containing pantothenic acid. By selecting a strain that forms a colony in step (1). In addition, by enriching a pantothenic acid-requiring strain by culturing the mutated yeast in a medium containing no pantothenic acid and selectively adding an antibiotic that acts on growing cells, for example, nystatin. Can be.
上記パントテン酸を含まない培地としては、具体的には下記の組成を有する培地が挙げられる。また、パントテン酸を含む培地としては、前記培地にパントテン酸塩を、0.1〜10mg/L、例えば0.4mg/L添加した培地が挙げられる。パントテン酸塩としては、パントテン酸カルシウムが挙げられる。また、固体培地の場合は、適当量の寒天を含む。 As the medium not containing pantothenic acid, a medium having the following composition is specifically mentioned. Examples of the medium containing pantothenic acid include a medium in which pantothenate is added to the above medium at 0.1 to 10 mg / L, for example, 0.4 mg / L. Pantothenates include calcium pantothenate. In the case of a solid medium, it contains an appropriate amount of agar.
上記のような性質を有する本発明の酵母は、十分な量のパントテン酸を含む培地で増殖することができ、かつ、パントテン酸量が制限された培地で培養したとき、乾燥酵母菌体当たりのγ−グルタミルシステイン含有量が経時的に上昇する。「十分な量のパントテン酸」とは、対数増殖期にある酵母が増殖することができるような量を意味する。この量は、通常は、0.1mg/L以上、好ましくは0.4mg/L以上である。また、パントテン酸量の上限は特に制限されないが、通常は、10mg/L以上であると過剰である。従って、パントテン酸量は通常、0.1〜10mg/Lである。 The yeast of the present invention having the above-mentioned properties can be grown in a medium containing a sufficient amount of pantothenic acid, and when cultured in a medium in which the amount of pantothenic acid is restricted, per yeast yeast cells The γ-glutamylcysteine content increases over time. By "sufficient amount of pantothenic acid" is meant an amount that allows yeast in logarithmic growth to grow. This amount is usually at least 0.1 mg / L, preferably at least 0.4 mg / L. Although the upper limit of the amount of pantothenic acid is not particularly limited, it is usually excessive when the amount is 10 mg / L or more. Therefore, the amount of pantothenic acid is usually 0.1 to 10 mg / L.
一方、「パントテン酸量が制限された」とは、十分な量のパントテン酸を含むときに対数増殖期にある酵母であっても、増殖できないか、増殖速度が低下するような量に培地中のパントテン酸量が制限されたことを意味する。この量は、通常は、0.1mg/L以下、好ましくは0.01mg/L以下である。尚、パントテン酸量が0mg/Lでもかまわない。 On the other hand, `` the amount of pantothenic acid is restricted '' means that even if the yeast is in the logarithmic growth phase when it contains a sufficient amount of pantothenic acid, it cannot be grown or the growth rate is reduced in the medium. Means that the amount of pantothenic acid was limited. This amount is usually 0.1 mg / L or less, preferably 0.01 mg / L or less. Incidentally, the amount of pantothenic acid may be 0 mg / L.
本発明において、「γ−グルタミルシステイン含有量が経時的に上昇する」とは、本発明の酵母を十分な量のパントテン酸を含む培地で増殖させた後、パントテン酸量が制限された培地に移して培養したとき、培地交換時での乾燥酵母菌体当たりのγ−グルタミルシステイン含有量に対し、培地交換後のγ−グルタミルシステイン含有量の最大値が好ましくは1.5倍以上、より好ましくは1.8倍以上、特に好ましくは2倍以上になることをいう。 In the present invention, "the γ-glutamylcysteine content increases with time" means that after the yeast of the present invention is grown on a medium containing a sufficient amount of pantothenic acid, the amount of pantothenic acid in the medium is restricted. When transferred and cultured, the maximum value of the content of γ-glutamylcysteine after the medium replacement is preferably 1.5 times or more, more preferably, to the γ-glutamylcysteine content per dry yeast cells at the time of medium replacement. Means 1.8 times or more, particularly preferably 2 times or more.
本発明の酵母は、MET25遺伝子の発現が脱抑制されるように改変されていてもよい。MET25遺伝子の発現が脱抑制されたとは、DOMINIQUEらの報告(MOLLECULAR AND CELLUAR BIOLOGY Dec, 1995, p6526-6534)に記載された条件において、MET25遺伝子の発現量がメチオニンにより抑制されないことを意味する。 The yeast of the present invention may be modified so that the expression of the MET25 gene is derepressed. The expression of the MET25 gene is derepressed means that the expression level of the MET25 gene is not suppressed by methionine under the conditions described in the report of DOMINIQUE et al. (MOLLECULAR AND CELLUAR BIOLOGY Dec, 1995, p6526-6534).
MET25遺伝子の発現を脱抑制させる方法として具体的には、例えば、コードされる蛋白質の569位のセリンがセリン以外の他のアミノ酸に置換する変異を有する変異型MET30
遺伝子を酵母に保持させる方法が挙げられる。前記他のアミノ酸としてはフェニルアラニンが挙げられる。このような性質を有する酵母は、後記実施例に示すように酵母を変異処理することによって取得することができるが、必要な変異が特定されたので、同変異を有する酵母は遺伝子工学的手法によって容易、かつ、確実に取得することができる。例えば、MET25遺伝子の発現が脱抑制された酵母は、上記変異を有する変異型MET30遺伝子を用いた遺伝子置換により創製することができる。遺伝子置換は、上記のGSH2遺伝子と同様にして行うことができる。また、変異型MET30遺伝子を保持する酵母は、同遺伝子を挿入したプラスミドで酵母を形質転換し、同遺伝子の細胞内のコピー数を高めることによっても創製することができる。さらに、変異型MET30遺伝子を保持する酵母は、実施例に記載したのと同様に、野生型酵母を、通常の変異処理、例えばUV照射、あるいはN−メチル−N−ニトロソグアニジン(NTG)、エチルメタンスルホネート(EMS)、亜硝酸、アクリジン等の変異剤による処理によって、取得することもできる。得られた変異株が目的の変異を有していることは、例えばPCR法等により確認することができる。なお、上記の、コードされる蛋白質の569位のセリンがフェニルアラニンに置換する変異を有する変異型MET30遺伝子を保持する酵母は、グルタチオンの生産にも使用することができる。
As a method of desuppressing the expression of the MET25 gene, specifically, for example, a mutant MET30 having a mutation in which serine at position 569 of the encoded protein is substituted with another amino acid other than serine
There is a method in which a gene is retained in yeast. Examples of the other amino acids include phenylalanine. Yeast having such properties can be obtained by subjecting yeast to mutation treatment as shown in Examples below, but since the necessary mutation has been identified, yeast having the mutation can be obtained by genetic engineering techniques. It can be obtained easily and reliably. For example, yeast in which the expression of the MET25 gene has been derepressed can be created by gene replacement using a mutant MET30 gene having the above mutation. Gene replacement can be performed in the same manner as in the GSH2 gene described above. In addition, yeast having the mutant MET30 gene can also be created by transforming yeast with a plasmid having the gene inserted therein and increasing the intracellular copy number of the gene. Furthermore, as described in Examples, the yeast having the mutant MET30 gene can be prepared by subjecting a wild-type yeast to a conventional mutation treatment, for example, UV irradiation, N-methyl-N-nitrosoguanidine (NTG), ethyl acetate, or the like. It can also be obtained by treatment with a mutating agent such as methanesulfonate (EMS), nitrous acid, and acridine. Whether the obtained mutant has the desired mutation can be confirmed by, for example, PCR. The yeast having the mutant MET30 gene having a mutation in which serine at position 569 of the encoded protein is substituted with phenylalanine can be used for producing glutathione.
本発明において、MET30遺伝子とは、MET4産物及び他の数種のタンパク質とともにSCFMET30複合体を形成し、MET25遺伝子の発現に関与するタンパク質をコードする遺伝子であり、例えば、配列番号3に示す塩基配列を有するサッカロマイセス・セレビシエ由来のMET30遺伝子、及びそのホモログが挙げられる。前記ホモログとしては、例えば配列番号3に記載の塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズするDNAが挙げられる。ここでいう「ストリンジェントな条件」とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。この条件を明確に数値化することは困難であるが、一例を示せば、相同性が高いDNA同士、例えば50%以上の相同性を有するDNA同士がハイブリダイズし、それより相同性が低いDNA同士がハイブリダイズしない条件、あるいは通常のサザンハイブリダイゼーションの洗いの条件である60℃、1×SSC,0.1%SDS、好ましくは、0.1×SSC、0.1%SDSに相当する塩濃度でハイブリダイズする条件が挙げられる。 In the present invention, the MET30 gene is a gene that forms a SCFMET30 complex with the MET4 product and several other proteins and encodes a protein involved in the expression of the MET25 gene. For example, the nucleotide sequence shown in SEQ ID NO: 3 MET30 gene derived from Saccharomyces cerevisiae and a homolog thereof. Examples of the homolog include a DNA that hybridizes with a DNA having the nucleotide sequence of SEQ ID NO: 3 under stringent conditions. The term "stringent conditions" used herein refers to conditions under which a so-called specific hybrid is formed and a non-specific hybrid is not formed. Although it is difficult to quantify these conditions clearly, as an example, DNAs having high homology, for example, DNAs having homology of 50% or more hybridize to each other, and DNAs having lower homology A salt corresponding to 60 ° C., 1 × SSC, 0.1% SDS, preferably 0.1 × SSC, 0.1% SDS at 60 ° C., which is a condition under which they do not hybridize with each other, or a washing condition for ordinary Southern hybridization. Conditions for hybridizing at a concentration are listed.
「569位のセリン」とは、配列番号4に示すMET30産物のアミノ酸配列において、569位に位置するセリン残基を意味する。また、アミノ酸配列におけるあるアミノ酸残基の位置は、その位置よりも上流側における配列中のアミノ酸残基の挿入、欠失等によって変化し得る。本発明において、「569位のセリン」とは、このように、アミノ酸配列中の絶対的位置が変わった場合であっても、配列番号4に示すアミノ酸配列における569位のセリン残基に相当するアミノ酸残基を含む。 “Serine at position 569” means a serine residue located at position 569 in the amino acid sequence of the MET30 product shown in SEQ ID NO: 4. In addition, the position of a certain amino acid residue in the amino acid sequence can be changed by insertion or deletion of an amino acid residue in the sequence upstream of the position. In the present invention, “serine at position 569” corresponds to the serine residue at position 569 in the amino acid sequence shown in SEQ ID NO: 4, even when the absolute position in the amino acid sequence is changed. Contains amino acid residues.
本発明に用いる変異型MET30遺伝子は、配列番号4に示すアミノ酸配列において、569位のセリンがセリン以外の他のアミノ酸に置換したアミノ酸配列を有するタンパク質と同等の機能を有するタンパク質をコードする保存的バリアント(conservative variant)、すなわち569位以外の1又は数箇所の位置における1又は数個のアミノ酸の置換、欠失、挿入又は付加を含むアミノ酸配列を有するMET30産物をコードするものであってもよい。ここで、「数個」とは、アミノ酸残基のタンパク質の立体構造における位置や種類によっても異なるが、具体的には例えば、前記「数個」は、2〜10個、好ましくは、2〜6個、より好ましくは2〜3個である。あるいは、変異型MET30遺伝子は、MET30産物のアミノ酸配列全体に対し、30から40%以上、好ましくは55〜65%以上の相同性を有するタンパク質をコードするDNAであってもよい。 The constitutive MET30 gene used in the present invention is a conservative gene encoding a protein having a function equivalent to that of a protein having an amino acid sequence in which serine at position 569 is substituted with another amino acid other than serine in the amino acid sequence shown in SEQ ID NO: 4. It may encode a variant (conservative variant), that is, a MET30 product having an amino acid sequence containing substitution, deletion, insertion or addition of one or several amino acids at one or several positions other than position 569. . Here, “several” differs depending on the position and type of the amino acid residue in the three-dimensional structure of the protein, but specifically, for example, the “several” is 2 to 10, preferably 2 to Six, more preferably two to three. Alternatively, the mutant MET30 gene may be a DNA encoding a protein having 30 to 40% or more, preferably 55 to 65% or more homology to the entire amino acid sequence of the MET30 product.
また、上記のような塩基の置換、欠失、挿入、付加、又は逆位等には、MET30遺伝子を保持する微生物の個体差、種や属の違いに基づく場合などの天然に生じる変異(mutant又はvariant)も含まれる。 In addition, substitution, deletion, insertion, addition, or inversion of a base as described above includes mutations that occur naturally (mutant) such as those based on individual differences, species, and genera of microorganisms that carry the MET30 gene. Or variant) is also included.
本発明の酵母としては、γ−グルタミルシステインを生産することができるものであれば特に制限されないが、具体的にはサッカロマイセス・セレビシエ等のサッカロマイセス属、キャンディダ・ユティリス等のキャンディダ属、シゾサッカロマイセス・ポンベ等のシゾサッカロマイセス属等に属する酵母が挙げられる。また、本発明の酵母菌株は、1倍体でもよいが、2倍性またはそれ以上の倍数性を有することが、生育が良好である点で好ましい。 The yeast of the present invention is not particularly limited as long as it can produce γ-glutamylcysteine.Specifically, Saccharomyces genus such as Saccharomyces cerevisiae, Candida genus such as Candida utilis, and Shizo Examples include yeast belonging to the genus Schizosaccharomyces such as Saccharomyces pombe. Although the yeast strain of the present invention may be haploid, it is preferable that the yeast strain has a diploid or higher ploidy in terms of good growth.
2倍性またはそれ以上の倍数性を有する酵母は、それらを変異処理しγ−グルタミルシステインを産生する株を選択すること、或いはγ−グルタミルシステイン産生株の育種に用いた1倍体酵母と、野生型酵母の1倍体を接合させ、得られた2倍体酵母を胞子形成させることにより、グルタチオン合成酵素活性が弱化し、γ−グルタミルシステインを産生する株を選択し、相異なる接合型を有するγ−グルタミルシステイン産生1倍体酵母2株を接合させることによって、取得することができる。同様にして、3倍性又はそれ以上の倍数性を有する酵母を取得することができる。 Yeast having a diploid or higher polyploidy, mutating them and selecting a strain that produces γ-glutamylcysteine, or a haploid yeast used for breeding a γ-glutamylcysteine producing strain, Glutathione synthetase activity is weakened by mating haploids of wild-type yeast and forming spores of the resulting diploid yeast, and a strain that produces γ-glutamylcysteine is selected. Γ-glutamylcysteine-producing haploid yeast can be obtained by conjugation. Similarly, a yeast having a triploid or higher ploidy can be obtained.
上記のような酵母の育種、改変に関する操作は、「化学と生物 実験ライン31 酵母の実験技術」初版 廣川書店;「バイオマニュアルシリーズ10 酵母による遺伝子実験法」初版、羊土社、「METHODS in YEAST GENETICS 2000 Edition」Cold Spring Harbor Laboratory Press等に記載されている。
The operations related to breeding and modification of yeast described above are described in “Chemistry and Biology Experiment Line 31 Yeast Experimental Techniques”, first edition, Hirokawa Shoten; “
<2>本発明の酵母の利用
本発明の酵母を、十分な量のパントテン酸を含む培地で培養して同酵母を増殖させた後、パントテン酸量が制限された培地で培養し、菌体内のγ−グルタミルシステイン含有量を上昇させることにより、γ−グルタミルシステインが蓄積した酵母を製造することができる。
<2> Utilization of the yeast of the present invention After culturing the yeast of the present invention in a medium containing a sufficient amount of pantothenic acid and growing the yeast, the yeast is cultured in a medium in which the amount of pantothenic acid is restricted. By increasing the content of γ-glutamylcysteine in the above, a yeast in which γ-glutamylcysteine has accumulated can be produced.
好ましくは、前記パントテン酸の「十分な量」は、予め一定量の酵母菌体を得るのに必要なパントテン酸要求量を実験的に測定し、目的の量の菌体を得るために必要なパントテン酸量を算出することによって、決定することができる。 Preferably, the `` sufficient amount '' of the pantothenic acid is determined in advance by experimentally measuring the required amount of pantothenic acid necessary to obtain a certain amount of yeast cells, and necessary to obtain the desired amount of cells. It can be determined by calculating the amount of pantothenic acid.
酵母を増殖させる工程では、パントテン酸は初発培地に全量添加してもよいし、分割添加してもよい。使用する培地及び培養条件は、パントテン酸量を制御できるものであれば特に制限されず、通常の酵母エキスの製造等に用いられる培地及び条件を採用することができる。 In the step of growing the yeast, pantothenic acid may be added to the initial medium in its entirety or may be added in portions. The medium and culture conditions to be used are not particularly limited as long as the amount of pantothenic acid can be controlled, and the medium and conditions used for the production of ordinary yeast extract and the like can be used.
本発明の酵母の好ましい態様として、低下したグルタチオン合成酵素活性を有する酵母は、グルタチオンを含まない培地でも良好に生育することができるので、通常、工業的に用いられる培地を用いることができる。尚、必要に応じて、用いる酵母の形質にしたがって必要な栄養素を培地に添加する。 As a preferred embodiment of the yeast of the present invention, a yeast having reduced glutathione synthetase activity can grow well even in a medium containing no glutathione, and therefore, a culture medium usually used industrially can be used. If necessary, necessary nutrients are added to the medium according to the characteristics of the yeast to be used.
十分な量の酵母菌体が得られたら、パントテン酸量が制限された培地で培養する。パントテン酸の量を制限する方法としては、十分な量のパントテン酸を含む培地で酵母を培養した培養液又は酵母菌体を、制限された量のパントテン酸を含むか、又はパントテン酸を含まない培地に移す方法が挙げられる。また、培地交換によらずに、パントテン酸の分割添加を停止することによっても、パントテン酸量を制限することができる。パントテン酸量を制限する時期は、対数増殖期に行うことが好ましい。例えば、パン酵母では、対数増殖期又は定常期まで培養した培養液を、2%となるように栄養培地に接種し、30℃で8〜16時間、振とう培養すれば、対数増殖期の菌体が得られる。 When a sufficient amount of yeast cells is obtained, the cells are cultured in a medium in which the amount of pantothenic acid is restricted. As a method of limiting the amount of pantothenic acid, a culture solution or yeast cells obtained by culturing yeast in a medium containing a sufficient amount of pantothenic acid, containing a limited amount of pantothenic acid, or containing no pantothenic acid The method of transferring to a culture medium is mentioned. Also, the amount of pantothenic acid can be limited by stopping the divisional addition of pantothenic acid without depending on the medium exchange. It is preferable to limit the amount of pantothenic acid during the exponential growth phase. For example, in baker's yeast, a culture solution cultured to a logarithmic growth phase or a stationary phase is inoculated to a nutrient medium at 2% and shake-cultured at 30 ° C. for 8 to 16 hours. The body is obtained.
パントテン酸量が制限された培地で培養する間に、酵母菌体内のγ−グルタミルシステインの蓄積量は経時的に上昇する。目的とする蓄積量に達したら、培養を終了する。通常は、好適な条件下では、培養時間は10〜30時間、好ましくは15〜27時間である。 During culturing in a medium in which the amount of pantothenic acid is restricted, the amount of accumulated γ-glutamylcysteine in the yeast increases with time. When the target accumulation amount is reached, the culture is terminated. Usually, under suitable conditions, the culture time is between 10 and 30 hours, preferably between 15 and 27 hours.
上記のようにして得られる培養物又はその分画物は、γ−グルタミルシステインを含有する。培養物は、酵母菌体を含む培養液であってもよいし、それから採取された酵母菌体、菌体破砕物、又は菌体抽出物(酵母エキス)であってもよい。菌体破砕物又は酵母エキスから、γ−グルタミルシステインを含む画分を得てもよい。 The culture or fraction thereof obtained as described above contains γ-glutamylcysteine. The culture may be a culture solution containing yeast cells, or a yeast cell, a crushed cell, or a cell extract (yeast extract) collected therefrom. A fraction containing γ-glutamylcysteine may be obtained from a crushed bacterial cell or a yeast extract.
上記γ−グルタミルシステインを含む培養物又はその分画物を加熱することにより、γ−グルタミルシステインからシステインを遊離させることができる。
酵母エキス等の調製は、通常の酵母エキスの調製と同様にして行えばよい。酵母エキスは、酵母菌体を熱水抽出したものを処理したものでもよいし、酵母菌体を消化したものを処理したものでもよい。
By heating the culture containing γ-glutamylcysteine or a fraction thereof, cysteine can be released from γ-glutamylcysteine.
The preparation of the yeast extract and the like may be performed in the same manner as the preparation of a normal yeast extract. The yeast extract may be obtained by treating a yeast cell that has been extracted with hot water, or a digested yeast cell that has been treated.
上記γ−グルタミルシステイン又はシステインを含む培養物又はその分画物は、飲食品の製造に用いることができる。飲食品としては、アルコール飲料、パン食品、又は発酵食品調味料が挙げられる。熱処理によるγ−グルタミルシステインからシステインの生成は、飲食品の製造中、又は製造の後に行われてもよい。 The above-mentioned culture containing γ-glutamylcysteine or cysteine or a fraction thereof can be used for the production of foods and drinks. Foods and drinks include alcoholic beverages, bread foods, and fermented food seasonings. The production of cysteine from γ-glutamylcysteine by heat treatment may be performed during or after the production of the food or drink.
上記飲食品は、γ−グルタミルシステイン又はシステインを含む培養物又はその分画物を、飲食品原料に混合し、飲食品に加工することによって製造される。本発明の飲食品は、前記培養物又は分画物を使用すること以外は、通常の飲食品と同様の原料を用い、同様の方法によって製造することができる。このような原料としては、例えばアルコール飲料では、米、大麦、コーンスターチ等、パン食品では小麦粉、砂糖、食塩、バター、発酵用酵母菌等が、発酵食品調味料では大豆、小麦等が挙げられる。 The food or drink is produced by mixing a culture containing γ-glutamylcysteine or cysteine or a fraction thereof with a food or drink raw material and processing the mixture into a food or drink. The food and drink of the present invention can be produced by the same method using the same raw materials as ordinary food and drink except for using the culture or the fraction. Examples of such raw materials include rice, barley, corn starch and the like for alcoholic beverages, flour, sugar, salt, butter, yeast for fermentation and the like for bread foods, and soybean and wheat for fermented food seasonings.
以下、本発明を実施例によりさらに具体的に説明する。 Hereinafter, the present invention will be described more specifically with reference to examples.
[実施例1]
<1>グルタチオン合成酵素活性が低下した酵母の取得
食品用途に用いられる市販の2倍体のサッカロマイセス・セレビシエを常法にしたがい胞子形成させた。形成した胞子からランダムスポア法により1倍体酵母YN0001株(MATα)を取得した。YN0001株をEMSにより変異処理し、グルタチオン含有量が低下した変異株YN0002株(MATα)を取得した。4分子解析により、YN0002株のGSH2遺伝子が変異していることを確認した。具体的には、コードされるタンパク質の387番目のグリシン残基がアスパラギン酸残基に置換されていた。また、グルタチオン含有量が低下した変異株YN0003株(MATa)も取得した。
[Example 1]
<1> Acquisition of yeast having reduced glutathione synthase activity A commercially available diploid Saccharomyces cerevisiae used for food applications was sporulated in a conventional manner. A haploid yeast strain YN0001 (MATα) was obtained from the formed spores by a random pore method. The YN0001 strain was subjected to mutation treatment by EMS to obtain a mutant YN0002 strain (MATα) having a reduced glutathione content. Four-molecule analysis confirmed that the GSH2 gene of the YN0002 strain was mutated. Specifically, the glycine residue at position 387 of the encoded protein was replaced with an aspartic acid residue. In addition, a mutant strain YN0003 (MATa) having a reduced glutathione content was also obtained.
尚、前記変異処理は、死滅率が90%になるような条件で行なった。YN0001株を50mlのYPD培地で30℃で1日間振とう培養し、酵母菌体を集菌した。酵母菌体を0.2Mリン酸ナトリウムバッファー(pH7.5)で3回洗浄した。酵母菌体を0.2Mリン酸ナトリウムバッファー(pH7.5)9.2ml、40% D-グルコース0.5ml、EMS 0.3ml(ナカライテスク社Code155-19)を含む溶液に懸濁し、30℃で90分間振とう培養した。この懸濁液に、10%チオ硫酸ナトリウム(フィルター滅菌)を10ml加え、10分間室温に放置して変異剤を中和した。酵母菌体を集菌し、0.2Mリン酸ナトリウムバッファー(pH7.5)で洗浄した。 The mutation treatment was performed under such conditions that the mortality was 90%. The YN0001 strain was cultured with shaking at 30 ° C. for 1 day in 50 ml of YPD medium, and yeast cells were collected. The yeast cells were washed three times with a 0.2 M sodium phosphate buffer (pH 7.5). The yeast cells were suspended in a solution containing 9.2 ml of 0.2 M sodium phosphate buffer (pH 7.5), 0.5 ml of 40% D-glucose, and 0.3 ml of EMS (Nacalai Tesque Code155-19), and shaken at 30 ° C for 90 minutes. Culture was continued. To this suspension was added 10 ml of 10% sodium thiosulfate (filter sterilized), and the mixture was left at room temperature for 10 minutes to neutralize the mutagen. The yeast cells were collected and washed with a 0.2 M sodium phosphate buffer (pH 7.5).
前記YN0001株とYN0002株を各々YPD培地に植菌し、30℃で振とう培養した。培養産物をSD培地に2%植菌し、30℃で振とう培養した。対数増殖期における菌体内グルタチオン含有
量を測定した。その結果、YN0001株のグルタチオン含有量は0.52%であった。一方、YN0002株のグルタチオン含有量は0.006%以下であった。
The YN0001 strain and the YN0002 strain were each inoculated in a YPD medium and cultured at 30 ° C. with shaking. The culture product was inoculated into SD medium at 2% and cultured with shaking at 30 ° C. The intracellular glutathione content during the logarithmic growth phase was measured. As a result, the glutathione content of the YN0001 strain was 0.52%. On the other hand, the glutathione content of the YN0002 strain was 0.006% or less.
<2>MET30遺伝子変異株の取得
前記1倍体酵母YN0001株(MATα)を、前記と同様にしてEMSにて変異処理し、MET25遺伝子の発現がメチオニンによって抑制されない変異株AJ14819株(MATα)を取得した。本菌株は、プライベートナンバーAJ14819が付与され、2002年9月 11 日付けで独立行政法人 産業技術総合研究所 特許生物寄託センター(〒305-8566 日本国茨城県つくば市東1丁目1番地1中央第6)に寄託され、受託番号FERM P-19007が付与されている。さらに、同菌株は平成15年10月1日にブダペスト条約に基く国際寄託に移管され、受託番号FERM BP-08502が付与されている。なお、MET25遺伝子の発現がメチオニンによって抑制されるか否かは、セレンを含む培地での生育の有無によって判別することもできる(DOMINIQUEら、MOLLECULAR AND CELLUAR BIOLOGY Dec, 1995, p6526-6534)。
<2> Acquisition of MET30 Gene Mutant The haploid yeast YN0001 strain (MATα) was mutated by EMS in the same manner as described above, and a mutant AJ14819 strain (MATα) in which the expression of the MET25 gene was not suppressed by methionine was obtained. I got it. This strain has been granted a private number AJ14819, and deposited on September 11, 2002, National Institute of Advanced Industrial Science and Technology, Patent Organism Depositary (1-1-1, Higashi 1-1, Tsukuba, Ibaraki, Japan 305-8566, Japan) ) And assigned accession number FERM P-19007. Further, this strain was transferred to an international deposit based on the Budapest Treaty on October 1, 2003, and given the accession number FERM BP-08502. Whether or not the expression of the MET25 gene is suppressed by methionine can also be determined by the presence or absence of growth in a medium containing selenium (DOMINIQUE et al., MOLLECULAR AND CELLUAR BIOLOGY Dec, 1995, p6526-6534).
具体的には、以下のようにして変異株の選択を行った。変異処理を行なった酵母をYPD寒天培地にスプレッドした。スプレッドは寒天培地に生育してくる酵母が100株前後になるように行なった。YPD培地に生育してきた酵母をセレンを含む培地とセレンを含まない培地(前記DOMINIQUEらに記載の寒天培地)にレプリカした。セレンを含む培地では生育しないが、セレンを含まない培地で生育する酵母菌株を選択した。 Specifically, mutants were selected as follows. The mutated yeast was spread on a YPD agar medium. Spreading was performed so that about 100 strains of yeast grew on the agar medium. The yeast grown on the YPD medium was replicated on a selenium-containing medium and a selenium-free medium (agar medium described in DOMINIQUE et al.). A yeast strain that does not grow on a medium containing selenium but grows on a medium not containing selenium was selected.
選択した酵母のMET25遺伝子発現量が上昇している事を以下のようにして確認した。選択した酵母菌株及びYN0001株をSD培地で培養し、その対数増殖期に集菌した。菌体内に含まれるRNAを回収し、RNA中に含まれるMET25遺伝子の転写産物の量を内部標準としてACT1遺伝子を用いて定量した。定量は、定量PCRであるPCR5700(Applied Biosystems社)を用い、TaqMan One-Step RT-PCRキット(Applied Biosystems社)を用いて行なった。TaqMan
Probe(Applied Biosystems社)に、ACT1-986T(配列番号5)及びMET25-1077T(配列番号6)を用い、ACT1及びMET25遺伝子の増幅用にACT1-963F(配列番号7)とACT1-1039R(配列番号8)、及びMET251056F(配列番号9)とMET25-1134R(配列番号10)を用いた(以上、Applied Biosystems社)。このようにして、MET25遺伝子の発現量がYN0001株よりも2倍以上に上昇した酵母AJ14819株を取得した。
It was confirmed that the expression level of the MET25 gene in the selected yeast was increased as follows. The selected yeast strain and YN0001 strain were cultured in an SD medium and collected during the logarithmic growth phase. RNA contained in the cells was recovered, and the amount of the MET25 gene transcript contained in the RNA was quantified using the ACT1 gene as an internal standard. The quantification was performed using PCR5700 (Applied Biosystems), which is a quantitative PCR, using a TaqMan One-Step RT-PCR kit (Applied Biosystems). TaqMan
Using ACT1-986T (SEQ ID NO: 5) and MET25-1077T (SEQ ID NO: 6) in a Probe (Applied Biosystems), ACT1-963F (SEQ ID NO: 7) and ACT1-1039R (SEQ ID NO: 7) for amplification of ACT1 and MET25 genes No. 8), and MET251056F (SEQ ID NO: 9) and MET25-1134R (SEQ ID NO: 10) (Applied Biosystems). In this way, a yeast strain AJ14819 in which the expression level of the MET25 gene was at least twice as high as that of the YN0001 strain was obtained.
このようにして取得したAJ14819株の変異遺伝子を4分子解析により特定し、その遺伝子の配列を調べたところ、MET30遺伝子がコードするタンパク質の569位のセリン残基がフェニルアラニン残基に変異していた。このようにしてMET25遺伝子の発現がメチオニンによって抑制されない酵母AJ14819株を取得した。 The mutant gene of the AJ14819 strain thus obtained was identified by four-molecule analysis, and the sequence of the gene was examined. The serine residue at position 569 of the protein encoded by the MET30 gene was mutated to a phenylalanine residue. . Thus, the yeast strain AJ14819 in which the expression of the MET25 gene was not suppressed by methionine was obtained.
<3>パントテン酸カルシウム要求性を有する酵母の取得
前記1倍体酵母YN0001株(MATα)を、前記と同様にしてEMSにて変異処理した。変異処理した酵母からパントテン酸カルシウム要求性酵母を取得するために、パントテン酸カルシウムを含まず、かつ、ナイスタチン(10μg/ml)を添加した培地で前記酵母を30℃で2時間培養し、培養液をYPD寒天培地にスプレッドした。生育してきた変異酵母をパントテン酸カルシウムを含まない寒天培地とパントテン酸カルシウムを含む寒天培地(各々前記表1に示す組成を有する)にレプリカし、前者の寒天培地では生育できないが、後者の寒天培地では生育できる酵母を選択した。このようにして、パントテン酸カルシウムを要求する酵母Pa0001株(MATa)を取得した。
<3> Obtaining yeast having a requirement for calcium pantothenate The haploid yeast strain YN0001 (MATα) was subjected to mutation treatment by EMS in the same manner as described above. In order to obtain a calcium pantothenate-requiring yeast from the mutated yeast, the yeast was cultured at 30 ° C. for 2 hours in a medium not containing calcium pantothenate and to which nystatin (10 μg / ml) was added. Was spread on a YPD agar medium. The grown mutant yeast is replicated on an agar medium not containing calcium pantothenate and an agar medium containing calcium pantothenate (each having the composition shown in Table 1 above), and cannot be grown on the former agar medium, but on the latter agar medium. Then we selected yeasts that could grow. In this way, yeast Pa0001 strain (MATa) requiring calcium pantothenate was obtained.
<4>変異型GSH2遺伝子及び変異型MET30遺伝子を有し、かつ、パントテン酸カルシウム要求性を有する酵母(GMP株;Diploid gsh2 met30 pa-)の取得
常法に従い、AJ14819株とPa0001株を接合させ、2倍体を取得した。取得した2倍体を胞子形成させ、ランダムスポア法により、変異型MET30遺伝子を有し、パントテン酸カル
シウム要求性を示す1倍体酵母MP株(MATa)を取得した。次に、YN0002株とMP株を接合させ2倍体を取得した。取得した2倍体を胞子形成させ、ランダムスポア法により、変異型GSH2遺伝子、変異型MET30遺伝子を有し、パントテン酸カルシウム要求性を示す1倍体酵母GMP-1(MATα)、GMP-2(MATa)株を取得した。GMP-1株とGMP-2株を接合させ、2倍体酵母GMP株を取得した。
<4> Acquisition of a yeast (GMP strain; Diploid gsh2 met30 pa-) having a mutant GSH2 gene and a mutant MET30 gene and having a requirement for calcium pantothenate According to a conventional method, the AJ14819 strain and the Pa0001 strain are conjugated. A diploid was obtained. The obtained diploid was sporulated, and a haploid yeast MP strain (MATa) having a mutant MET30 gene and exhibiting a requirement for calcium pantothenate was obtained by a random pore method. Next, the YN0002 strain and the MP strain were conjugated to obtain a diploid. The obtained diploid is sporulated, and haploid yeasts GMP-1 (MATα), GMP-2 (GMP-1) having a mutant GSH2 gene and a mutant MET30 gene and exhibiting a requirement for calcium pantothenate are obtained by a random pore method. MATa) strain. The GMP-1 strain and the GMP-2 strain were conjugated to obtain a diploid yeast GMP strain.
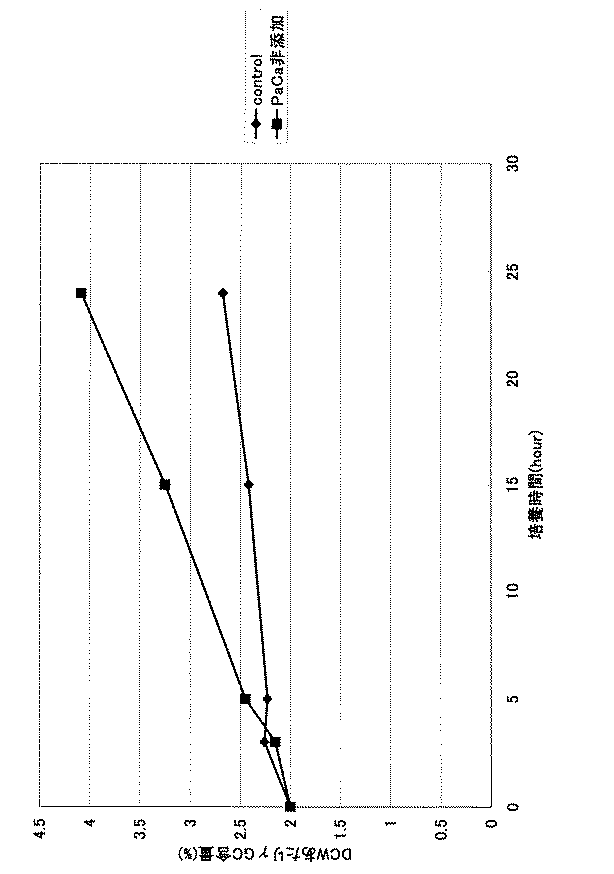
<5>GMP株を用いたγ−グルタミルシステインの製造
GMP株をYPD培地(試験管4ml)に植菌し、30℃で1日振とう培養した。培養液をパントテン酸カルシウム0.4mg/dlを含む培地に植菌し、30℃で振とう培養した。対数増殖期に培地を採取し、菌体濃度(乾燥酵母重量)が60mg/dlになるように、パントテン酸カルシウムを含まない培地、及び0.4mg/Lのパントテン酸カルシウムを含む培地(前記表1)に、それぞれ添加し、培養を行った。乾燥酵母菌体あたりのγ−グルタミルシステイン含有量を経時的に測定した。結果を図1に示す。パントテン酸カルシウム濃度が低い培地で培養した場合は、高濃度で含む培地で培養した場合に比べて、γ−グルタミルシステイン含有量が経時的に増加した。
<5> Production of γ-glutamylcysteine using GMP strain
The GMP strain was inoculated into a YPD medium (4 ml of a test tube) and cultured with shaking at 30 ° C. for 1 day. The culture was inoculated into a medium containing 0.4 mg / dl of calcium pantothenate and cultured at 30 ° C. with shaking. The medium was collected during the logarithmic growth phase, and a medium containing no calcium pantothenate and a medium containing 0.4 mg / L calcium pantothenate so that the cell concentration (dry yeast weight) was 60 mg / dl (see Table 1 above). ), And cultivation was performed. The content of γ-glutamylcysteine per dried yeast cells was measured over time. The results are shown in FIG. When cultured in a medium with a low concentration of calcium pantothenate, the content of γ-glutamylcysteine increased with time as compared with the case of culturing in a medium containing a high concentration of pantothenate.
以上の結果より、GMP株のγ−グルタミルシステイン含有量が、パントテン酸カルシウムの不足により上昇することが示された。 From the above results, it was shown that the γ-glutamylcysteine content of the GMP strain increased due to lack of calcium pantothenate.
[比較例1] 変異型GSH2遺伝子及び変異型MET30遺伝子を有する酵母の取得
常法に従い、前記MET30遺伝子変異株AJ14819株と、市販の酵母から得た1倍体酵母Pa0001株を接合させ、2倍体を取得した。この2倍体を胞子形成させ、ランダムスポア法により、変異型MET30遺伝子を有する、1倍体酵母M株(MATa)を取得した。次に、このM株と、前記GSH2遺伝子変異株YN0002株を接合させ2倍体を取得した。この2倍体を胞子形成させ、ランダムスポア法により、変異型GSH2遺伝子及び変異型MET30遺伝子を有する1倍体酵母GM-1株(MATα)、GM-2株(MATa)を取得した。GM-1株とGM-2株を接合させ、2倍体酵母GM株を取得した。
[Comparative Example 1] Obtaining yeast having mutant GSH2 gene and mutant MET30 gene According to a conventional method, the MET30 gene mutant AJ14819 strain was conjugated with a haploid yeast Pa0001 strain obtained from a commercially available yeast, and doubled. I got the body. The diploid was sporulated, and a haploid yeast M strain (MATa) having a mutant MET30 gene was obtained by a random-spore method. Next, the M strain was conjugated to the GSH2 gene mutant YN0002 strain to obtain a diploid. The diploid was sporulated, and haploid yeast strains GM-1 (MATα) and GM-2 (MATa) having a mutant GSH2 gene and a mutant MET30 gene were obtained by a random pore method. The GM-1 strain and the GM-2 strain were conjugated to obtain a diploid yeast GM strain.
[実施例2] GMP株及びGM株によるγ−グルタミルシステインの製造
GM株及びGMP株を各々YPD培地に植菌し、30℃で振とう培養した。培養液をパントテン酸カルシウム0.4mg/dlを含む培地に植菌し、30℃で振とう培養した。対数増殖期に集菌し、菌体濃度(乾燥酵母重量)が60mg/dlになるように、パントテン酸カルシウムを含まない培地に植菌した。30℃で振とう培養し、乾燥酵母菌体当たりのγ−グルタミルシステイン含有量を経時的に測定した。結果を図2に示す。
[Example 2] Production of γ-glutamylcysteine by GMP strain and GM strain
Each of the GM strain and the GMP strain was inoculated in a YPD medium and cultured at 30 ° C. with shaking. The culture was inoculated into a medium containing 0.4 mg / dl of calcium pantothenate and cultured at 30 ° C. with shaking. The cells were collected during the logarithmic growth phase, and inoculated in a medium containing no calcium pantothenate so that the cell concentration (dry yeast weight) was 60 mg / dl. After shaking culture at 30 ° C., the content of γ-glutamylcysteine per dried yeast cells was measured with time. FIG. 2 shows the results.
その結果、GMP株では、パントテン酸カルウシウムを欠乏させた状態で培養を継続するに従い、乾燥酵母菌体あたりのγ−グルタミルシスイテン含有量が上昇することが示された。 As a result, it was shown that in the GMP strain, the γ-glutamylcysitene content per dry yeast cell increases as the culture is continued in a state in which the calcium pantothenate is deficient.
[実施例3]
次に、GMP株のグルタチオン合成酵素活性を更に低下させた場合の影響を検討した。
[Example 3]
Next, the effect of further reducing the glutathione synthetase activity of the GMP strain was examined.
<6>グルタチオン合成酵素遺伝子破壊カセットの作成
まず、グルタチオン合成酵素遺伝子破壊カセットを以下のようにして作成した。
Kpn Iで切断したpAUR123ベクター(宝酒造 code3602)を鋳型として、GSH2-AUR1-C-F(配列番号11)とGSH2-AUR1-C-R(配列番号12)をプライマーに用いてPCRを行った。PCR産物はAUR1-C遺伝子の両端にGSH2遺伝子のORFのN末端側配列及びGSH2遺伝子のORFのC末端側配列を含むため、同産物を用いて、グルタチオン合成酵素遺伝子を破壊することが可能である。PCRの条件は以下の通りである。
<6> Preparation of cassette for disrupting glutathione synthase gene First, a cassette for disrupting glutathione synthase gene was prepared as follows.
Using pAUR123 vector (Takara Shuzo code3602) cut with KpnI as a template, PCR was performed using GSH2-AUR1-CF (SEQ ID NO: 11) and GSH2-AUR1-CR (SEQ ID NO: 12) as primers. Since the PCR product contains the N-terminal sequence of the ORF of the GSH2 gene and the C-terminal sequence of the ORF of the GSH2 gene at both ends of the AUR1-C gene, the glutathione synthase gene can be disrupted using the same product. is there. The PCR conditions are as follows.
Kpn Iで切断したpAUR123 1μl
10×PCR Buffer 5μl
dNTP 4μl
10pmol/μl GSH2-AUR1-C-Fプライマー 1μl
10pmol/μl GSH2-AUR1-C-Rプライマー 1μl
KOD Dash DNA polymerase 0.5μl
MilliQ 37.5μl
Total 50μl
94℃2min ×1cycle→94℃40sec、54℃40sec、74℃1min ×30cycles →4℃ ∞
(KOD Dash はTOYOBO 製 codeLDP-101)
1 μl of pAUR123 cut with Kpn I
10 × PCR Buffer 5μl
dNTP 4μl
10 pmol/μl GSH2-AUR1-CF primer 1μl
10 pmol/μl GSH2-AUR1-CR primer 1μl
KOD Dash DNA polymerase 0.5μl
MilliQ 37.5μl
Total 50μl
94 ℃ 2min × 1cycle → 94 ℃ 40sec 、 54 ℃ 40sec 、 74 ℃ 1min × 30cycles → 4 ℃ ∞
(KOD Dash is codeLDP-101 made by TOYOBO)
<7>AJ14861株の取得
上記のようにして作製したグルタチオン合成酵素遺伝子破壊カセットを用いて、以下のようにしてGMP株のグルタチオン合成酵素遺伝子の破壊を試みた。即ち、GMP株をYPD培地で培養し、その対数増殖期に集菌した。1Mソルビトール溶液で2回洗浄した菌体を、以下の組成の溶液に懸濁し、5℃で1時間放置した。
組成: 0.1 M LiCl
10 mM DTT
10 mM Tris-HCl(pH7.5)
1 mM EDTA
<7> Acquisition of AJ14861 strain Using the glutathione synthase gene disruption cassette prepared as described above, the glutathione synthase gene of the GMP strain was disrupted as follows. That is, the GMP strain was cultured in the YPD medium, and collected during the logarithmic growth phase. The cells washed twice with a 1 M sorbitol solution were suspended in a solution having the following composition, and allowed to stand at 5 ° C for 1 hour.
Composition: 0.1 M LiCl
10 mM DTT
10 mM Tris-HCl (pH7.5)
1 mM EDTA
その後、1Mソルビトール溶液で2回洗浄した。このようにして調整した菌体に上述のようにして作製したPCR産物を混合し、エレクトロポレーションを行った。(「バイオマニュアルシリーズ10 酵母による遺伝子実験法」初版、羊土社)その後、混合物をYPD培地に植菌し、30℃で16時間振とう培養した。培養産物を、選択マーカーであるオーレオバシジンA(宝酒造 code9000)を0.2μg/ml含有するYPD寒天培地にスプレッドし3日間30℃で培養した。(GMP株のオーレオバシジンAに対する最小生育阻止濃度は0.05μg/ml)生育してきたコロニーを0.2μg/mlのオーレオバシジンAを含有するYPD寒天培地に塗布し、オーレオバシジンAに対する耐性能が獲得されているコロニーを選抜した。このようにして、AJ14861株を取得した。なお、本株は、プライベートナンバーAJ14861が付与され、2003年11月19日付けで、独立行政法人 産業技術総合研究所 特許生物寄託センター(〒305-8566
日本国茨城県つくば市東1丁目1番地1中央第6)に、ブダペスト条約に基いて国際寄託されており、受託番号FERM BP-08553が付与されている。
Then, it was washed twice with a 1 M sorbitol solution. The thus-prepared bacterial cells were mixed with the PCR product prepared as described above, and electroporation was performed. ("
It has been deposited internationally under the Budapest Treaty at 1-1-1 Higashi, 1-chome, Tsukuba, Ibaraki, Japan, and is assigned the accession number FERM BP-08553.
<8>AJ14861株を用いたγ−グルタミルシステインの製造
AJ14861株をYPD培地(試験管4ml)に植菌し、30℃で1日振とう培養した。培養液をパントテン酸カルシウム0.4mg/dlを含む培地に植菌し、30℃で振とう培養した。対数増殖期に培地を採取し、菌体濃度(乾燥酵母重量)が60mg/dlになるように、パントテン酸カルシウムを含まない培地、及び0.4mg/Lのパントテン酸カルシウムを含む培地(前記表1)に、それぞれ添加し、培養を行った。乾燥酵母菌体あたりのγ−グルタミルシステイン含有量を経時的に測定した。結果を図3に示す。パントテン酸カルシウム濃度が低い培地で培養した場合は、高濃度で含む培地で培養した場合に比べて、γ−グルタミルシステイン含有量が経時的に増加した。
<8> Production of γ-glutamylcysteine using AJ14861 strain
The AJ14861 strain was inoculated into a YPD medium (4 ml in a test tube) and cultured with shaking at 30 ° C. for 1 day. The culture was inoculated into a medium containing 0.4 mg / dl of calcium pantothenate and cultured at 30 ° C. with shaking. The medium was collected during the logarithmic growth phase, and a medium containing no calcium pantothenate and a medium containing 0.4 mg / L calcium pantothenate so that the cell concentration (dry yeast weight) was 60 mg / dl (see Table 1 above). ), And cultivation was performed. The content of γ-glutamylcysteine per dried yeast cells was measured over time. The results are shown in Figure 3. When cultured in a medium with a low concentration of calcium pantothenate, the content of γ-glutamylcysteine increased with time as compared with the case of culturing in a medium containing a high concentration of pantothenate.
以上の結果より、AJ14861株のγ−グルタミルシステイン含有量が、パントテン酸カルシウムの不足により上昇することが示された。 From the above results, it was shown that the γ-glutamylcysteine content of AJ14861 strain was increased due to lack of calcium pantothenate.
本発明により、グルタチオン合成酵素活性が低下又は消失し、MET25遺伝子の発現が脱抑制され、パントテン酸要求性を示す酵母が提供される。本発明の酵母を、好適な条件で培養することにより、γ−グルタミルシステインを高含有する酵母培養液が提供される。本発明の酵母及び酵母培養液は、γ−グルタミルシステイン含有飲食品又はシステイン含有飲食品の製造に利用することができる。 INDUSTRIAL APPLICABILITY The present invention provides a yeast showing reduced or eliminated glutathione synthetase activity, derepression of MET25 gene expression, and showing pantothenic acid requirement. By culturing the yeast of the present invention under suitable conditions, a yeast culture containing high content of γ-glutamylcysteine is provided. The yeast and the yeast culture solution of the present invention can be used for producing γ-glutamylcysteine-containing food or drink or cysteine-containing food or drink.
Claims (12)
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2003399198A JP4352877B2 (en) | 2002-12-13 | 2003-11-28 | Method for producing γ-glutamylcysteine |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2002361918 | 2002-12-13 | ||
| JP2003399198A JP4352877B2 (en) | 2002-12-13 | 2003-11-28 | Method for producing γ-glutamylcysteine |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2004201677A true JP2004201677A (en) | 2004-07-22 |
| JP4352877B2 JP4352877B2 (en) | 2009-10-28 |
Family
ID=32828634
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2003399198A Expired - Lifetime JP4352877B2 (en) | 2002-12-13 | 2003-11-28 | Method for producing γ-glutamylcysteine |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP4352877B2 (en) |
Cited By (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2008069173A1 (en) | 2006-12-04 | 2008-06-12 | Ajinomoto Co., Inc. | Process for producing seasoning |
| JP2008189573A (en) * | 2007-02-02 | 2008-08-21 | Fancl Corp | Sleep inducer, stress insomnia improving agent |
| WO2009102050A1 (en) | 2008-02-15 | 2009-08-20 | Ajinomoto Co., Inc. | Intestinal immune system stimulator |
| WO2009110624A1 (en) | 2008-03-04 | 2009-09-11 | 味の素株式会社 | γ-GLUTAMULCYSTEINE-PRODUCING YEAST, AND METHOD FOR PRODUCTION OF YEAST EXTRACT |
| WO2010116833A1 (en) | 2009-04-08 | 2010-10-14 | Ajinomoto Co., Inc. | Novel yeast having increased content of sulfur-containing compound, screening method thereof, and culturing method thereof |
| JP2013212063A (en) * | 2012-04-02 | 2013-10-17 | Ajinomoto Co Inc | YEAST CONTAINING γ-Glu-X |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP5857973B2 (en) | 2010-04-12 | 2016-02-10 | 味の素株式会社 | Yeast extract containing γ-Glu-X or γ-Glu-X-Gly and method for producing the same |
-
2003
- 2003-11-28 JP JP2003399198A patent/JP4352877B2/en not_active Expired - Lifetime
Cited By (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2008069173A1 (en) | 2006-12-04 | 2008-06-12 | Ajinomoto Co., Inc. | Process for producing seasoning |
| JP2008189573A (en) * | 2007-02-02 | 2008-08-21 | Fancl Corp | Sleep inducer, stress insomnia improving agent |
| WO2009102050A1 (en) | 2008-02-15 | 2009-08-20 | Ajinomoto Co., Inc. | Intestinal immune system stimulator |
| WO2009110624A1 (en) | 2008-03-04 | 2009-09-11 | 味の素株式会社 | γ-GLUTAMULCYSTEINE-PRODUCING YEAST, AND METHOD FOR PRODUCTION OF YEAST EXTRACT |
| WO2010116833A1 (en) | 2009-04-08 | 2010-10-14 | Ajinomoto Co., Inc. | Novel yeast having increased content of sulfur-containing compound, screening method thereof, and culturing method thereof |
| JP2012523230A (en) * | 2009-04-08 | 2012-10-04 | 味の素株式会社 | Yeast having increased sulfur-containing compound content, screening method thereof, and culture method |
| JP2013212063A (en) * | 2012-04-02 | 2013-10-17 | Ajinomoto Co Inc | YEAST CONTAINING γ-Glu-X |
Also Published As
| Publication number | Publication date |
|---|---|
| JP4352877B2 (en) | 2009-10-28 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4453781B2 (en) | DNA encoding γ-glutamylcysteine synthetase | |
| JP4273689B2 (en) | γ-glutamylcysteine-producing yeast | |
| JPWO2009110624A1 (en) | Method for producing γ-glutamylcysteine-producing yeast and yeast extract | |
| EP1512747B1 (en) | Gene encoding glutathione synthetase from Candida utilis | |
| JP4352877B2 (en) | Method for producing γ-glutamylcysteine | |
| JP5315600B2 (en) | Y-glutamylcysteine-producing yeast and use thereof | |
| CN114657079A (en) | Yeast with high content of Abu, gamma-Glu-Abu and/or gamma-Glu-Abu-Gly | |
| CN100439490C (en) | Method for producing gamma-glutamylcysteine | |
| JP4228644B2 (en) | Method for producing γ-glutamylcysteine | |
| CN112639117A (en) | Method for producing glutathione | |
| EP1452585A1 (en) | Gamma-glutamylcysteine-producing yeast and method of screening the same | |
| JP2005211068A (en) | Bacillus natto with high glutaminate polypeptidase activity and fermented soybean improved in threading property | |
| JP4778740B2 (en) | Phytase enzyme low activity natto and long-term stable natto |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20060627 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20090421 |
|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20090515 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20090601 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20090601 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20090707 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20090720 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 4352877 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120807 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120807 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120807 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120807 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130807 Year of fee payment: 4 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |