JP2014003979A - B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 - Google Patents
B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 Download PDFInfo
- Publication number
- JP2014003979A JP2014003979A JP2013138528A JP2013138528A JP2014003979A JP 2014003979 A JP2014003979 A JP 2014003979A JP 2013138528 A JP2013138528 A JP 2013138528A JP 2013138528 A JP2013138528 A JP 2013138528A JP 2014003979 A JP2014003979 A JP 2014003979A
- Authority
- JP
- Japan
- Prior art keywords
- polypeptide
- nucleic acid
- fragment
- fbsa
- fibrinogen
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images














































Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/315—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Streptococcus (G), e.g. Enterococci
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Gastroenterology & Hepatology (AREA)
- Veterinary Medicine (AREA)
- Peptides Or Proteins (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Oncology (AREA)
- Communicable Diseases (AREA)
- Public Health (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Apparatus Associated With Microorganisms And Enzymes (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
【課題】B群連鎖球菌(GBS)は新生児死亡率の主要原因である、B群連鎖球菌感染に対するワクチンなどの薬物の開発の手段、具体的には、このような薬物の製造に使用可能なGBSの新しい接着物因子を提供する。
【解決手段】B群連鎖球菌の接着因子をコードする核酸、B群連鎖球菌の接着因子。具体的には、このような接着因子であるポリペプチド、および特定のアミノ酸配列を含むポリペプチド、ならびにワクチンの製造におけるこのようなポリペプチドの使用。
【選択図】なし
【解決手段】B群連鎖球菌の接着因子をコードする核酸、B群連鎖球菌の接着因子。具体的には、このような接着因子であるポリペプチド、および特定のアミノ酸配列を含むポリペプチド、ならびにワクチンの製造におけるこのようなポリペプチドの使用。
【選択図】なし
Description
本発明は、細菌の接着因子をコードする単離された核酸分子、細菌の接着因子、およびさまざまなその使用に関する。
発明の背景
ストレプトコッカス・アガラクティエ(Streptococcus agalactiae)またはB群連鎖球菌(GBS)は新生児死亡率の主要原因である。GBSは、推定有病率が年に数千例であり、年間死亡率は米国では約10〜15%である(Schuchat, 1998)。米国における研究では、出生児1000人あたり1〜2例であり(Zangwill et al., 1992)、また欧州諸国では出生児1000人あたりの罹患率が0.24〜1.26例であることが報告されている(Carstensen et al., 1985; Faxelius et al., 1988)。米国では、最大30%の妊娠女性が、症状を示すことなく膣内または腸内に少なくとも一時的にGBSを有する(Schuchat, 1998)。このような女性から生まれた乳児は、分娩時にGBSのコロニーが形成される(Baker and Edwards, 1995)。感染した羊水または膣分泌物を吸引することでGBSが肺に達する場合がある。感染の一般的な徴候には、菌血症、肺炎、および髄膜炎などがある(Spellerberg, 2000)。GBSによる髄膜炎の犠牲とならなくとも、生存者は難聴、学習障害、ならびに運動障害、感覚障害、および認知障害などの神経学的な後遺症に悩まされることになる(Baker and Edwards, 1995)。現在、妊産婦を対象とした抗生物質による予防が、GBSによる新生児疾患の予防に推奨されている方法である(Baker et al., 1999)が、他の連鎖球菌種における抗生物質耐性株の出現に伴い、GBSに絡む類似の問題が生じる恐れがある。
ストレプトコッカス・アガラクティエ(Streptococcus agalactiae)またはB群連鎖球菌(GBS)は新生児死亡率の主要原因である。GBSは、推定有病率が年に数千例であり、年間死亡率は米国では約10〜15%である(Schuchat, 1998)。米国における研究では、出生児1000人あたり1〜2例であり(Zangwill et al., 1992)、また欧州諸国では出生児1000人あたりの罹患率が0.24〜1.26例であることが報告されている(Carstensen et al., 1985; Faxelius et al., 1988)。米国では、最大30%の妊娠女性が、症状を示すことなく膣内または腸内に少なくとも一時的にGBSを有する(Schuchat, 1998)。このような女性から生まれた乳児は、分娩時にGBSのコロニーが形成される(Baker and Edwards, 1995)。感染した羊水または膣分泌物を吸引することでGBSが肺に達する場合がある。感染の一般的な徴候には、菌血症、肺炎、および髄膜炎などがある(Spellerberg, 2000)。GBSによる髄膜炎の犠牲とならなくとも、生存者は難聴、学習障害、ならびに運動障害、感覚障害、および認知障害などの神経学的な後遺症に悩まされることになる(Baker and Edwards, 1995)。現在、妊産婦を対象とした抗生物質による予防が、GBSによる新生児疾患の予防に推奨されている方法である(Baker et al., 1999)が、他の連鎖球菌種における抗生物質耐性株の出現に伴い、GBSに絡む類似の問題が生じる恐れがある。
GBSは幼児の感染に加えて、高齢者および免疫不全者における重要な病原体でもあり、浸潤性GBS疾患の発症率は100,000人に約9人である(Farley et al., 1993)。このような感染のうち死亡率は30%にのぼる場合がある。
重要なGBS病原性決定因子は、宿主補体因子C3bの沈着を防ぐことで、同菌のオプソニン貧食を阻害する型特異的な莢膜多糖である(Rubens et al., 1987)。GBSには9種類の異なる莢膜血清型であるIa、Ib、およびII〜VIIIがこれまで同定されている(Wessels, 1997)。臨床的に重要な血清型の莢膜多糖を元に、GBSに対する多価混合ワクチンを開発する取り組みが進行中である(Paoletti et al., 1999; Baker et al., 1999; Baker et al., 2000; Paoletti and Kasper, 2002)。しかし、莢膜を含む混合ワクチンには、多数の血清型が必要であること、適切なタンパク質混合物を同定して検証する必要があること、またヒト組織との潜在的な交差反応を解明する必要があることなどの、克服すべき技術上の問題がいくつかある(Korzeniowska-Kowal et al., 2001)。GBSの細胞表面タンパク質の使用は、同細菌に対するワクチンの開発における、莢膜多糖に代わる魅力ある方法である。GBSの表面タンパク質Sip、Rib、α、およびβは、GBS感染に対する防御免疫をマウスにもたらすことが既にわかっている。(Madoff et al., 1992; Larsson et al., 1997; Larsson et al., 1999; Brodeur et al., 2000)。血清型V株の2種類の特有の表面タンパク質についてもGBS感染を防ぐことがマウスモデルで報告されている(Areschoug et al., 1999)。またGBSのC5aペプチダーゼに対する抗体は、マクロファージによる細菌死滅を引き起こすことがわかっている(Cheng et al., 2001)。
GBSとその宿主との相互作用は、コロニー形成ならびに上皮および内皮の表面への浸透、および免疫防御の回避が関与する複雑な過程である(Spellerberg, 2000)。連鎖球菌では、フィブリノーゲン結合が、宿主表面への接着(Courtney et al., 1994; Cheung et al., 1991; Ni et al., 1998; Pei and Flock, 2001)、および免疫系からの防御(Courtney et al., 1997; Them et al., 1998; Ringdahl et al., 2000)に重要な役割を果たすことが報告されている。したがって複数の研究で、血清群A、C、およびGの連鎖球菌におけるフィブリノーゲン結合の分子的基盤が説明されている(Fischetti, 1989; Meehan et al., 1998; Vasi et al., 2000)。
フィブリノーゲンは、血漿中に高濃度で見出される330 kDaの糖タンパク質である(Fuss et al., 2001; Mosesson et al., 2001)。フィブリノーゲンは、それぞれジスルフィド結合で連結された2つのAα鎖、Bβ鎖、およびγ鎖からなるヘキサマーである。フィブリノーゲンは止血に重要な役割を果たし、傷害部位における血小板接着および凝集に関与する。またフィブリノーゲンはトロンビンによって切断されて凝血の主要要素であるフィブリンとなる。フィブリノーゲンはオプソニン貧食にも関連する。活性化された補体因子C3bの結合を阻害することで、代替的な補体経路の活性化を阻害することが報告されている(Whitnack et al., 1984; Whitnack and Beachey, 1985)。新生児にみられる汎発性GBS感染に対する特有の感受性は、相対的な補体欠損と関連づけられている(Mills et al., 1979; Edwards et al., 1983)。したがって、GBSのフィブリノーゲン結合は、新生児における残存性補体活性の阻害に重要な役割を果たすと考えられる(Noel et al., 1991)。
複数の研究でGBSとヒトのフィブリノーゲンとの相互作用について報告されている(Schonbeck et al., 1981; Lammler et al., 1983; Chhatwal et al., 1984; Spellerberg et al., 2002)。しかしながら、GBSとフィブリノーゲン結合の分子的基盤は未だ不明である。
GBSは、上皮細胞および内皮細胞に結合して浸潤することがわかっている(Gibson et al., 1993; La Penta et al., 1997; Winram et al., 1998)。プロテアーゼであるトリプシンでGBSを処理すると、同細菌の接着性および浸潤性は消失する(Valentin-Weigand and Chhatwal, 1995; Winram et al., 1998)ので、GBSの付着因子(adhesin)およびインベーシン(invasin)はタンパク質性であることが示唆されている。付着因子およびインベーシンは細菌表面に位置し、GBSの病原性に重要であるので、GBSワクチンを開発する上で理想的な標的となる。
本発明の基礎となる問題は、細菌感染に対するワクチンなどの薬物の開発の手段を提供することである。具体的には、この問題は、このような薬物の製造に使用可能なGBSの新しい接着物因子を提供することである。
本発明の基礎となる問題は、第1の局面では、以下からなる群より選択される核酸配列を含む、単離された核酸分子(好ましくはフィブリノーゲン結合ポリペプチド、またはこのようなタンパク質もしくはこの断片をコードする核酸分子)によって解決される:
(a)SEQ ID NO 1〜SEQ ID NO 6からなる群より選択される核酸配列と少なくとも70%が同一な核酸;
(b)(a)の核酸に本質的に相補的な核酸;
(c)(a)または(b)の核酸の少なくとも15連続塩基を含む核酸;
(d)ストリンジェントなハイブリダイゼーション条件で、(a)、(b)、または(c)のポリヌクレオチドとアニールする核酸;ならびに
(e)遺伝コードの縮重を除いて、(a)、(b)、(c)、または(d)で定義された核酸とハイブリダイズする可能性がある核酸。
(a)SEQ ID NO 1〜SEQ ID NO 6からなる群より選択される核酸配列と少なくとも70%が同一な核酸;
(b)(a)の核酸に本質的に相補的な核酸;
(c)(a)または(b)の核酸の少なくとも15連続塩基を含む核酸;
(d)ストリンジェントなハイブリダイゼーション条件で、(a)、(b)、または(c)のポリヌクレオチドとアニールする核酸;ならびに
(e)遺伝コードの縮重を除いて、(a)、(b)、(c)、または(d)で定義された核酸とハイブリダイズする可能性がある核酸。
本発明の基礎となる問題は、第2の局面では、以下からなる群より選択される核酸配列を含む、単離された核酸分子(好ましくは接着因子またはこの断片をコードする核酸分子)によって解決される:
(a)Seq ID NO 7、Seq ID NO 8、Seq ID NO 9、またはSeq ID NO 10に記載された核酸配列と少なくとも70%が同一な核酸;
(b)(a)の核酸に本質的に相補的な核酸;
(c)(a)または(b)の核酸の少なくとも15連続塩基を含む核酸;
(d)ストリンジェントなハイブリダイゼーション条件で、(a)、(b)または(c)の核酸とアニールする核酸;ならびに
(e)遺伝コードの縮重を除いて、(a)、(b)、(c)、または(d)で定義された核酸とハイブリダイズする可能性がある核酸。
(a)Seq ID NO 7、Seq ID NO 8、Seq ID NO 9、またはSeq ID NO 10に記載された核酸配列と少なくとも70%が同一な核酸;
(b)(a)の核酸に本質的に相補的な核酸;
(c)(a)または(b)の核酸の少なくとも15連続塩基を含む核酸;
(d)ストリンジェントなハイブリダイゼーション条件で、(a)、(b)または(c)の核酸とアニールする核酸;ならびに
(e)遺伝コードの縮重を除いて、(a)、(b)、(c)、または(d)で定義された核酸とハイブリダイズする可能性がある核酸。
本発明の両局面の態様では、同一性は少なくとも80%、好ましくは少なくとも90%、より好ましくは100%である。
本発明の両局面の別の態様では、核酸はDNAである。
本発明の両局面のさらに別の態様では、核酸はRNAである。
本発明の両局面の好ましい態様では、核酸分子は細菌から単離される。
本発明の両局面のより好ましい態様では、細菌は連鎖球菌、ブドウ球菌、およびラクトコッカスからなる群より選択される種である。
本発明の両局面のさらにより好ましい態様では、細菌はStreptococcus agalactiae、ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)、およびストレプトコッカス・ミュータンス(Streptococcus mutans)からなる群より選択される種である。
本発明の両局面の最も好ましい態様では、細菌はStreptococcus agalactiaeである。
本発明の第1の局面の態様では、核酸分子は、16アミノ酸を含むアミノ酸モチーフの少なくとも1個のリピートを含むフィブリノーゲン結合タンパク質をコードする。
本発明の第2の局面の態様では、核酸分子は、上皮細胞と相互作用する接着因子をコードする。
本発明の第1の局面の好ましい態様では、コードされたフィブリノーゲン結合タンパク質は、特定されたものか、または本明細書に開示されたものであるアミノ酸モチーフのリピートを19個含む。
本発明の第1の局面のより好ましい態様では、このようなリピートは、SEQ ID NO 21〜SEQ ID NO 112からなる群より選択されるポリヌクレオチドにコードされている。
第3の局面では、本発明の基礎となる問題は、SEQ ID NO 21〜SEQ ID NO 21〜112からなる群より選択される核酸配列を含む、単離された核酸分子によって解決される。
第4の局面では、本発明の基礎となる問題は、
 のアミノ酸モチーフを含むポリペプチドをコードする、単離された核酸分子によって解決される。
のアミノ酸モチーフを含むポリペプチドをコードする、単離された核酸分子によって解決される。
本発明の第1〜4の局面の好ましい態様では、核酸はDNA、RNA、またはこの混合物であり、好ましくは核酸分子はゲノムDNAから単離される。
第5の局面では、本発明の基礎となる問題は、本発明の任意の局面の核酸分子を含むベクターによって解決される。
好ましい態様では、ベクターは、本発明の任意の局面の核酸分子にコードされたポリペプチドの組換え発現に適している。
第6の局面では、本発明の基礎となる問題は、本発明のベクターを含む細胞によって解決される。
好ましい態様では、このような細胞は宿主細胞である。
第7の局面では、本発明の基礎となる問題は、本発明の任意の局面の核酸分子にコードされているアミノ酸配列を含むポリペプチド、好ましくはフィブリノーゲン結合ポリペプチド、および/または接着因子、ならびにこのようなポリペプチドの断片によって解決される。
第8の局面では、本発明の基礎となる問題は、SEQ ID NO 11〜SEQ ID NO 20からなる群より選択されるアミノ酸配列を含むポリペプチド、好ましくはフィブリノーゲン結合ポリペプチド、および/または接着因子によって解決される。
本発明のこの局面の態様では、SEQ ID NO 11〜16のいずれか1つのアミノ酸配列を有するポリペプチド(好ましくはフィブリノーゲン結合ポリペプチド)、および/または接着因子はフィブリノーゲン結合タンパク質である。
本発明のこの局面の別の態様では、このようなポリペプチドは、上皮細胞と相互作用する接着因子である。さらにより好ましい態様では、上皮細胞はヒト上皮細胞である。
第9の局面では、本発明の基礎となる問題は、SEQ ID NO 113〜SEQ ID NO 205からなる群より選択されるアミノ酸配列を含むポリペプチドによって解決される。ある態様ではポリペプチドは、SEQ ID NO 113〜SEQ ID NO 225のいずれかのアミノ酸配列の少なくとも1つを、少なくとも1つの他のアミノ酸配列と組み合わせて含む。より好ましい、少なくとも1つの他のアミノ酸配列は、SEQ ID NO 113〜SEQ ID NO 205のいずれかのアミノ酸配列である。
第10の局面では、本発明の基礎となる問題は、
のアミノ酸モチーフを含むポリペプチドによって解決される。
第11の局面では、本発明の基礎となる問題は、本発明の任意の局面の核酸分子を発現させる段階を含む、本発明の任意の局面のポリペプチドの作製法によって解決される。
第12の局面では、本発明の基礎となる問題は、適切な宿主細胞を本発明のベクターで形質転換する段階またはトランスフェクトする段階段階を含む、本発明の任意の局面のポリペプチドおよびこの断片を発現する細胞の作製法によって解決される(形質転換された細胞またはトランスフェクトされた細胞は、ベクターに含まれるポリヌクレオチドにコードされたポリペプチドを発現する)。
第13の局面では、本発明の基礎となる問題は、本発明の任意の局面で定義されたポリペプチドもしくはこの断片、または本発明の任意の局面の核酸分子を含む薬学的組成物(特にワクチン)によって解決される。
好ましい態様では、薬学的組成物は、好ましくはポリカチオンポリマー、免疫刺激性デオキシヌクレオチド(ODN)、合成KLKペプチド、神経刺激性化合物、ミョウバン、フロインドの完全もしくは不完全アジュバント、またはこれらの組み合わせからなる群より選択される免疫刺激物質を含む。
好ましい態様では、免疫刺激物質は、ポリカチオンアニオンと免疫刺激性デオキシヌクレオチドの組み合わせ、または合成KLKペプチドと免疫刺激性デオキシヌクレオチドの組み合わせのいずれかである。
より好ましい態様では、ポリカチオンポリマーはポリカチオンペプチドであり、および/または神経刺激性化合物はヒト成長ホルモンである。
第14の局面では、本発明の基礎となる問題は、薬物の製造(特に対細菌感染ワクチンの製造)用の本発明の任意の局面のポリペプチド、またはこの断片の使用によって解決される。
好ましい態様では、細菌感染はStreptococcus agalactiaeの細菌感染である。
第15の局面では、本発明の基礎となる問題は、細菌感染を予防および治療するための薬剤の製造用の本発明の任意の局面のポリペプチドとフィブリノーゲンの結合を阻害する分子の使用によって解決される。好ましくは、細菌感染はStreptococcus agalactiaeの感染である。
別の態様では、このような分子は、本発明の任意の局面のポリペプチドに結合するフィブリノーゲン受容体抗体、フィブリノーゲン受容体ミモトープ(mimotope)、およびフィブリノーゲン受容体拮抗物質からなる群より選択される。
第16の局面では、本発明の基礎となる問題は、本発明の任意の局面のポリペプチドと上皮細胞(好ましくはヒト上皮細胞)との結合を阻害する分子の使用によって解決される。
第17の局面では、本発明の基礎となる問題は、本発明の任意の局面のポリペプチドまたはこの断片の少なくとも選択的部分に結合する抗体、または少なくとも抗体の有効部分によって解決される。
ある態様では、このような抗体はモノクローナル抗体である。
別の態様では、有効部分はFab断片を含む。
さらに別の態様では、このような抗体はキメラ抗体である。
好ましい態様では、このような抗体はヒト化抗体である。
第18の局面では、本発明の基礎となる問題は、本発明の抗体を産生するハイブリドーマ細胞系列によって解決される。
第19の局面では、本発明の基礎となる問題は、細菌感染(特にStreptococcus agalactiaeの感染)を治療または予防するための薬物の調製用の本発明の抗体の使用によって解決される。
第20の局面では、本発明の基礎となる問題は、本発明の任意の局面のポリペプチドまたはこの断片の活性を低下もしくは阻害する拮抗物質によって解決される。
第21の局面では、本発明の基礎となる問題は、以下の段階を含む、本発明の任意の局面のポリペプチドもしくはこの断片の活性を低下もしくは阻害することを可能とする拮抗物質を同定する方法によって解決される:
a)本発明の任意の局面の単離された、または固定化されたポリペプチドもしくはこの断片に候補拮抗物質を、候補拮抗物質とポリペプチドまたはこの断片の結合に応じた検出シグナルの提供を可能とする成分の存在下で、候補拮抗物質とポリペプチドまたはこの断片の結合を可能とする条件で接触させる段階;ならびに
b)拮抗物質とポリペプチドまたはこの断片の結合に応じて発生したシグナルの有無を、好ましくは、ポリペプチドもしくはこの断片の活性を阻害もしくは低下させることを可能とする化合物を示すシグナルの存在により検出する段階。
a)本発明の任意の局面の単離された、または固定化されたポリペプチドもしくはこの断片に候補拮抗物質を、候補拮抗物質とポリペプチドまたはこの断片の結合に応じた検出シグナルの提供を可能とする成分の存在下で、候補拮抗物質とポリペプチドまたはこの断片の結合を可能とする条件で接触させる段階;ならびに
b)拮抗物質とポリペプチドまたはこの断片の結合に応じて発生したシグナルの有無を、好ましくは、ポリペプチドもしくはこの断片の活性を阻害もしくは低下させることを可能とする化合物を示すシグナルの存在により検出する段階。
第22の局面では、本発明の基礎となる問題は、以下の段階を含む、本発明の任意の局面のポリペプチドまたはこの断片の活性を低下または阻害することを可能とする拮抗物質を同定する方法によって解決される:
(a)本発明の任意の局面のポリペプチドまたはこの断片を提供する段階;
(b)本発明の任意の局面のポリペプチドの相互作用パートナー(好ましくは本発明の抗体)を提供する段階;
(c)候補拮抗物質を提供する段階;
(d)ポリペプチド、ポリペプチドの相互作用パートナー、および候補拮抗物質を反応させる段階;ならびに
(e)候補拮抗物質がポリペプチドの活性を阻害するか、または低下させるか否かを判定する段階。
(a)本発明の任意の局面のポリペプチドまたはこの断片を提供する段階;
(b)本発明の任意の局面のポリペプチドの相互作用パートナー(好ましくは本発明の抗体)を提供する段階;
(c)候補拮抗物質を提供する段階;
(d)ポリペプチド、ポリペプチドの相互作用パートナー、および候補拮抗物質を反応させる段階;ならびに
(e)候補拮抗物質がポリペプチドの活性を阻害するか、または低下させるか否かを判定する段階。
第23の局面では、本発明の基礎となる問題は、以下の段階を含む、本発明のポリペプチドまたはこの断片の、相互作用パートナーに対する相互作用活性を低下させるか、もしくは阻害することを可能とする拮抗物質を同定する方法によって解決される:
(a)本発明のポリペプチドまたはこの断片を提供する段階;
(b)ポリペプチドまたはこの断片に対する相互作用パートナー(好ましくは本発明の抗体)を提供する段階;
(c)ポリペプチドまたはこの断片と相互作用パートナーの相互作用により、相互作用複合体の形成を可能とする段階;
(d)候補拮抗物質を提供する段階;
(e)候補拮抗物質と相互作用複合体間の競合反応を可能とする段階;ならびに
(f)候補拮抗物質が、ポリペプチドもしくはこの断片の、相互作用パートナーとの相互作用活性を阻害するか、または低下させるか否かを判定する段階。
(a)本発明のポリペプチドまたはこの断片を提供する段階;
(b)ポリペプチドまたはこの断片に対する相互作用パートナー(好ましくは本発明の抗体)を提供する段階;
(c)ポリペプチドまたはこの断片と相互作用パートナーの相互作用により、相互作用複合体の形成を可能とする段階;
(d)候補拮抗物質を提供する段階;
(e)候補拮抗物質と相互作用複合体間の競合反応を可能とする段階;ならびに
(f)候補拮抗物質が、ポリペプチドもしくはこの断片の、相互作用パートナーとの相互作用活性を阻害するか、または低下させるか否かを判定する段階。
第24の局面では、本発明の基礎となる問題は、本発明の第22の局面もしくは第23の局面の方法で同定される、または同定可能な拮抗物質によって解決される。
第25の局面では、本発明の基礎となる問題は、本発明の任意の局面のポリペプチドをコードするポリヌクレオチド配列の存在、またはポリペプチドの存在を判定する段階を含む、本発明の任意の局面のポリペプチドまたはこの断片の発現と関連する疾患のインビトロ診断法によって解決される。
第26の局面では、本発明の基礎となる問題は、本発明のポリペプチドもしくはこの断片をコードする核酸配列の存在、または本発明のポリペプチドもしくはこの断片の存在を判定する段階を含む、本発明のポリペプチドもしくはこの断片の発現と関連する疾患のインビトロ診断法によって解決される。
第27の局面では、本発明の基礎となる問題は、本発明の任意の局面の核酸分子、または本発明の任意の局面のポリペプチドの存在を判定する段階を含む、細菌感染(好ましくはStreptococcus agalactiaeの感染)のインビトロ診断法によって解決される。
後者の3つの本発明の任意の局面の好ましい態様では、ポリペプチドの存在は、好ましくは宿主生物に由来する試料を対象に判定される。
第28の局面では、本発明の基礎となる問題は、支持材料、および本発明の任意の局面のポリペプチド、または本発明の任意の局面の核酸分子が支持材料に固定化されたものを含むアフィニティ装置によって解決される。
第29の局面では、本発明の基礎となる問題は、ポリペプチドの相互作用パートナーを単離および/または精製、および/または同定するための、本発明の任意の局面のポリペプチドの使用によって解決される。
第30の局面では、本発明の基礎となる問題は、ポリペプチドに結合するペプチドを生成させるための、本発明の任意の局面の任意のポリペプチドの使用によって解決される。
好ましい態様では、このようなペプチドは、アンチカリン(anticaline)からなる群より選択される。
第31の局面では、本発明の基礎となる問題は、アプタマーおよびシュピーゲルマーからなる群より選択される機能性核酸の作製用の、本発明の任意の局面のポリペプチドの使用によって解決される。
第32の局面では、本発明の基礎となる問題は、リボザイム、アンチセンス核酸、およびsiRNAからなる群より選択される機能性リボ核酸の作製用の、本発明の任意の局面の核酸分子の使用によって解決される。
第33の局面では、本発明の基礎となる問題は、本発明のポリペプチドまたはこの断片の抗原としての使用によって解決される。
第34の局面では、本発明の基礎となる問題は、機能性核酸(好ましくはリボ核酸)の作製または生成用の、本発明の任意の局面の核酸の使用によって解決される(機能性リボ核酸は、リボザイム、アンチセンス核酸、およびsiRNAからなる群より選択される)。
第35の局面では、本発明の基礎となる問題は、抗体を生成させるか、もしくは作製するための、本発明のポリペプチドもしくはこの任意の断片の使用によって解決される。
本明細書で用いる「SEQ ID NO X〜SEQ ID NO Y」という表現は、XおよびYを含む、X〜Yに含まれる任意の配列番号の省略形である。
発明者らは驚くべきことにGBSのゲノムが、共通のアミノ酸モチーフを有する多様な接着因子を含むことを見出した。このアミノ酸モチーフは接着因子とフィブリノーゲンの結合に関連する。本明細書で用いるように、接着因子は好ましくは、微生物と基質の結合に関与するペプチドまたはタンパク質の因子である。好ましくは、このような微生物はGBSである。より好ましくは、基質はそれぞれフィブリノーゲンおよび宿主細胞である。本明細書で用いる接着因子は付着因子またはインベーシンの場合がある。共通のアミノ酸モチーフは、アミノ酸の1文字コードで
のように表すことができる。
上記の配列からわかるように、アミノ酸モチーフは合計16か所存在する。モチーフ位置の一部は、例えば1位または3位または4位などの別個のアミノ酸によって占められていることになる。15位または16位などの他の位置は、任意のアミノ酸(好ましくは天然のアミノ酸)で占められている場合がある。このような位置は、上記配列で「X」で記されている。さらに別のモチーフ位置は、さまざまなアミノ酸で占められる場合がある。このようなさまざまなアミノ酸は、上記モチーフで示されており、さまざまなアミノ酸は「/」で隔てられている。したがって、2位にN、S、またはTが存在する場合がある。当業者であれば、上記のアミノ酸配列が任意の順序をとることを理解するので、これは本発明の範囲に含まれる。
したがって本発明は1つの局面では、上記アミノ酸モチーフに関する。具体的には本発明は、このアミノ酸モチーフを含む任意のペプチドまたはポリペプチドに関する。「ペプチド」および「ポリペプチド」という表現は、特に明記されない限りにおいて同義的に用いられると理解される。
本明細書で用いる「ポリペプチド」は、後述するようにすべてのポリペプチドを含む。ポリペプチドの基本構造は周知であり、また当技術分野の数多くの教科書および他の出版物に記載されている。この文脈では、この表現は、ペプチド結合で直鎖状に相互に連結された2個もしくはこれ以上のアミノ酸を含む任意のペプチドまたはタンパク質を表現するように本明細書で用いられる。本明細書で用いるこの表現は、特に明記した部分を除いて、例えばペプチド、オリゴペプチド、およびオリゴマーと当技術分野で一般に呼ばれる短い鎖と、多くの種類のあるタンパク質と当技術分野で一般に呼ばれる長い鎖の両方を意味する。当然のことながら、ポリペプチドは、20の天然のアミノ酸と一般に呼ばれる20種類のアミノ酸以外のアミノ酸を含む場合があり、また末端のアミノ酸を含む多くのアミノ酸は、任意のポリペプチドにおいて、プロセシングおよび他の翻訳後修飾などの天然のプロセスだけでなく、当技術分野で周知の化学的修飾法による修飾を受ける場合がある。ポリペプチド中に天然に存在する共通の修飾は、本明細書で詳細に列挙するにはあまりに数が多すぎるが、基本的な教科書、またさらに詳しくはモノグラフ、ならびに大部の研究論文に詳しく記載されており、これらは当業者に周知である。本発明のポリペプチド中に存在する可能性のある既知の修飾を説明目的で挙げるとすれば、アセチル化、アシル化、ADPリボシル化、アミド化、フラビンの共有結合、ヘム基の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合形成、脱メチル化、共有結合の架橋形成、シスチン形成、ピログルタミン酸形成、ホルミル化、ガンマ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ素化、メチル化、ミリストイル化、酸化、プロテアーゼによるプロセシング、リン酸化、プレニル化、ラセミ化、セレノイル化、硫酸化、アルギニル化などの転移RNA介在性のタンパク質へのアミノ酸の付加、およびユビキチン化などがある。このような修飾は当業者に周知であり、また文献に詳細に記載されている。例えば複数の特に一般的な修飾、グリコシル化、脂質結合、硫酸化、グルタミン酸残基のガンマ-カルボキシル化、ヒドロキシル化、およびADPリボシル化は、例えばPROTEINS-STRUCTURE AND MOLECULAR PROPERTIES, 2nd Ed., T.E. Creighton, W.H. Freeman and Company, New York (1993)などの基本的な教科書で説明されている。この件に関しては例えばWold, F.のPosttranslational Protein Modifications: Perspectives and Prospects, pgs.1-12(POSTTRANSLATIONAL COVALENT MODIFICATION OF PROTEINS, B.C. Johnson, Ed., Academic Press, New York (1983))、SeifterらのMeth. Enzymol. 182: 626-646 (1990)、およびRattanらのProtein Synthesis: Posttranslational Modification and Aging, Ann. N.Y. Acad. Sci. 663: 48-62 (1992)などの多くの詳細な総説を参照することができる。当然のことながら、周知かつ上述したように、このようなポリペプチドは常に完全に直線状とは限らない。例えばポリペプチドは一般に、天然のプロセシングや、人間の操作による天然には起こらないプロセシングなどの翻訳後処理の結果である場合がある。環状、分岐状、および分岐環状のポリペプチドは、非翻訳的な天然のプロセスによって、また完全合成法によって合成される場合もある。修飾は、ペプチドの主鎖、アミノ酸側鎖、およびアミノ末端またはカルボキシ末端を含むポリペプチド上のどの位置でも生じる場合がある。実際にポリペプチド中のアミノ基もしくはカルボキシ基のいずれか、または両方を共有結合の修飾でブロックすることは、天然および合成のポリペプチドに一般的であり、またこのような修飾は、本発明のポリペプチドにも存在する場合がある。例えば、大腸菌または他の細胞で作られるポリペプチドのアミノ末端残基は、プロテアーゼによるプロセシングを受ける前は、ほぼ常にN-ホルミルメチオニンである。ペプチドの翻訳後修飾中にNH2末端のメチオニン残基は除去される。したがって本発明は、本発明のタンパク質のメチオニンを含むアミノ末端バリアントと、メチオニンを含まないアミノ末端バリアントの両方の使用を対象とする。ポリペプチド中にみられる修飾は、その作製法の関数である場合がある。例えば、クローン化された遺伝子を宿主で発現させることによって作製されるポリペプチドの場合、修飾の性質および規模の大半は、宿主細胞の翻訳後修飾能力によって決定され、修飾シグナルがポリペプチドアミノ酸配列中に存在する。例えば、よく知られているように、グリコシル化は、例えば大腸菌などの細菌宿主では起こらないことが多い。したがって、グリコシル化が望ましい場合は、グリコシル化の可能な宿主(一般に真核細胞)でポリペプチドを発現させるべきである。昆虫細胞は、哺乳類の細胞と同じ翻訳後グリコシル化を行うことがあるので、昆虫の細胞発現系が、特に天然のパターンのグリコシル化を有する哺乳類のタンパク質を効率的に発現させるために開発されている。似たようなことは他の修飾についても言える。当然のことながら同じ種類の修飾が、任意のポリペプチド中の複数の部位に同じ程度か、または多様な程度で存在する場合がある。また任意のポリペプチドは、多くの種類の修飾を含む場合がある。一般に、本明細書で用いる「ポリペプチド」という表現は、このようなあらゆる修飾、特に宿主細胞でポリヌクレオチドを発現させることによって組換え的に合成されたポリペプチドに存在する修飾を含む。
アミノ酸モチーフを含む任意のポリペプチドは、本発明のポリペプチドであるとみなされる。実施例で詳細に説明するように発明者らは、GBSが1コピーのアミノ酸モチーフだけでなく、数コピーのモチーフも含むいくつかの接着因子を含むことを明らかにした。したがって、複数のアミノ酸モチーフを含む、または複数のアミノ酸モチーフからなる任意のポリペプチドは本発明のポリペプチドである。例えば、本明細書でFbsAと記載される接着因子は、わずか1単位のアミノ酸モチーフから、多くて19コピーのモチーフを含む場合がある。
本発明の他の接着因子は、本明細書でPabA、PabB、PabC、およびPabDと記載される。「本発明のポリペプチド」という表現は、本発明のポリペプチドの任意の断片、誘導体、または類似体も含むと理解される。さらに好ましい本発明のポリペプチドは、SEQ ID 11〜20に対応するアミノ酸配列である。
本発明のポリペプチドの断片、誘導体、または類似体は、(i)1つもしくは複数のアミノ酸残基が、保存アミノ酸残基もしくは非保存アミノ酸残基(好ましくは保存アミノ酸残基)と置換されたもの(このような置換アミノ酸残基は、遺伝コードにコードされたものである場合もあればそうでない場合もある)、または(ii)1つもしくは複数のアミノ酸残基が置換基を含むもの、または(iii)成熟ポリペプチドが、ポリペプチドの半減期を延長させる化合物(例えばポリエチレングリコール)などの別の化合物と融合したもの、または(iv)リーダー配列もしくは分泌配列などの追加的なアミノ酸が成熟ポリペプチドと融合したもの、または成熟ポリペプチドもしくはプロタンパク質の配列の精製に用いられる配列である場合がある。このような断片、誘導体、および類似体は、本明細書に記述されることで、当技術分野に含まれるとみなされる。
この点に関して、本発明の特に好ましい態様は、配列表に記載されたポリペプチド、バリアント、類似体、誘導体、およびこれらの断片、ならびに断片のバリアント、類似体、および誘導体である。また、このようなポリペプチド、バリアント、類似体、誘導体、およびこれらの断片、ならびに断片のバリアント、類似体、および誘導体、ならびに異種ポリペプチドを含む融合ポリペプチドは本発明の対象となる。このような融合ポリペプチドおよびタンパク質、ならびにこれらをコードするポリヌクレオチドは、融合タンパク質をコードする組換えポリ核酸を作製および発現させる標準的な組換え法を含む標準的な手段で容易に作製することができる。
好ましいバリアントは、保存的アミノ酸置換が基準と異なるものである。このような置換は、ポリペプチド中の任意のアミノ酸が、同様の特徴をもつ他のアミノ酸と置換される置換である。典型的には、保存的置換にみられる置換は、脂肪族アミノ酸であるAla、Val、Leu、およびIle間の相互の置換であり、ヒドロキシル残基であるSerとThrの交換であり、酸性残基であるAspとGluの交換であり、アミド残基であるAsnとGln間の置換であり、塩基性残基LysとArgの交換であり、ならびに芳香族残基であるPheとTyrの置換である。
この点に関して、さらに特に好ましいのは、複数、少数、5〜10個、1〜5個、1〜3個、2個、1個、または0個のアミノ酸残基が任意の組み合わせで置換、欠失、付加される、配列表に記載された任意のポリペプチドのアミノ酸配列を有するバリアント、類似体、誘導体、および断片、ならびに断片のバリアント、類似体、および誘導体である。特に好ましいのは、本発明のポリペプチドの性質および活性を変化しないサイレントの置換、付加、および欠失である。またこの点に関して特に好ましいのは保存的置換である。アミノ酸配列を有する最も極めて好ましいポリペプチドが、置換を含まない状態で配列表に記載されている。
本発明のポリペプチドおよびポリヌクレオチドは好ましくは、単離された状態で、好ましくは均一に精製された状態で提供される。また本発明のポリペプチドは、好ましくは単離されたポリペプチドである。
本発明のポリペプチドは、配列表に記載された任意のポリペプチド(特に成熟ポリペプチド)、ならびに配列表記載のポリペプチドと少なくとも70%が同一なポリペプチド、配列表記載のポリペプチドと好ましくは少なくとも80%もしくは85%が同一なポリペプチド、またより好ましくは、配列表記載のポリペプチドと少なくとも90%の類似性(より好ましくは少なくとも90%の同一性)ポリペプチド、またさらにより好ましくは配列表記載のポリペプチドと少なくとも95%、96%、97%、98%、99%、または99.5%の類似性(さらにより好ましくは少なくとも95%、96%、97%、98%、99%、または99.5%の同一性)を有するポリペプチドを含み、ならびにこのようなポリペプチドの部分も含む(ポリペプチドのこのような部分は一般に、少なくとも5アミノ酸、またより好ましくは少なくとも10個、15個、または16個またはその倍数(multiples)を含む)。倍数は、好ましくは16アミノ酸のリピートの倍数である(16アミノ酸は、本明細書に開示されたアミノ酸モチーフに対応する)。
本発明のポリペプチドの断片または一部は、対応する完全長のポリペプチドをペプチド合成法で作製するために使用することができる。したがって、このような断片は、完全長のポリペプチドを作製するための中間体として使用することができる。
本発明のポリヌクレオチドの断片または一部を使用して完全長の本発明のポリヌクレオチドを合成することができる。
また本発明のこの局面の好ましい態様には、配列表に記載されたアミノ酸配列を有するポリペプチドの断片、ならびに配列表に記載されたポリペプチドのバリアントおよび誘導体の断片を含むポリペプチドがある。
本明細書で用いる断片は、前述のS. agalactiaeのポリペプチドのアミノ酸配列、およびこのバリアントまたは誘導体のアミノ酸配列の一部として全体的には同じであるが、必ずしもすべてが同じというわけではないポリペプチドである。
このような断片は「自立形(free-standing)」である場合があり、すなわち別のアミノ酸またはポリペプチドの一部、またはこれらとの融合体ではなく、一部もしくは領域を形成する、より大きなポリペプチドに含まれる場合がある。より大きなポリペプチドに含まれる場合は、議論の対象である断片は、最も好ましくは1つの連続領域を形成する。しかし複数の断片は、より1つの長いポリペプチドの内部に含まれる場合がある。例えば、ある好ましい態様は、宿主における発現用に設計され、断片のアミノ酸末端に融合された異種のプレポリペプチド領域およびプロポリペプチド領域、および断片のカルボキシル末端と融合された追加的な領域を有する前駆体ポリペプチドの内部に含まれる本発明のポリペプチドの断片に関する。したがって、本明細書で意図される意味の1つの局面における断片は、本発明のポリペプチドに由来する融合ポリペプチドまたは融合タンパク質の部分もしくは部分群を意味する。
本発明のポリペプチド断片の代表例は例えば、任意に選択されたポリペプチドにおいて、本明細書に記載されたアミノ酸配列のアミノ酸番号約45〜60、61〜76、77〜92、93〜108、109〜124、125〜140、141〜156、157〜172、173〜188、189〜204、205〜220、221〜236、237〜252、253〜268、269〜284、285〜300、301〜316、317〜332、333〜348、410〜414に由来する断片、または任意のリピートを、単独または表1および表2に挙げられたものの1つもしくは複数の組み合わせとして含む(任意選択でシグナルペプチドもしくはLPXTGモチーフが結合される)。
本明細書で用いる「約」という表現は、いずれかの末端で、または両方の末端で少数のアミノ酸、または5アミノ酸、4アミノ酸、3アミノ酸、2アミノ酸、または1アミノ酸が大きいかまたは小さい、特に記載された範囲を含む。
本発明の好ましい断片は例えば、配列表に記載されたアミノ酸配列を有するポリペプチドを含む切断型ポリペプチド、またはこのバリアントもしくは誘導体を含む。ただし、アミノ末端を含む一連の連続残基(すなわち連続する領域、部分、または一部)の欠失、カルボキシル末端を含む一連の連続した残基の欠失、または二重切断型変異体の場合であれば2つの一連の連続した残基(1つはアミノ末端を含み、1つはカルボキシル末端を含む)の欠失を除く。上記範囲の大きさをもつ断片は、切断型断片の好ましい態様でもあり、これは、一般に特に好ましい断片である。宿主細胞における本発明のポリペプチドの分解型も好ましい。
本発明のこの局面では、本発明のポリペプチドの構造または機能性の属性を特徴とする断片も好ましい。この点における本発明の好ましい態様は、本発明のポリペプチドのアルファヘリックスおよびアルファヘリックス形成領域、ベータシートおよびベータシート形成領域、ターンおよびターン形成領域、コイルおよびコイル形成領域、親水性領域、疎水性領域、アルファ両親媒性領域、ベータ両親媒性領域、可動性領域、表面形成領域、基質結合領域、および高抗原性指標領域、ならびにこれらの断片の組み合わせを含む断片を含む。
好ましい領域は、本発明のポリペプチドの活性に関連する領域である。この点に関して最も好ましいのは、類似の活性もしくは改善された活性を有する断片、または望ましくない活性が低下された断片を含む、本発明のポリペプチドの化学的、生物学的、または他の活性を有する断片である。特に好ましいのはS. agalactiaeのヒトにおける疾患を引き起こす能力、および/またはS. agalactiaeの上皮細胞(より好ましくはヒト上皮細胞)への接着および/または浸潤に介在可能な能力に本質的な機能をもたらす、例えばフィブリノーゲン(FbsAの場合)、または宿主細胞(PabA、PabB、PabC、およびPabDの場合)などに対する受容体活性を含む断片である。さらに好ましいポリペプチド断片は、動物(特にヒト)で抗原性または免疫原性決定因子を構成するか、または含む断片である。本明細書で用いられる宿主細胞は、実施例1に記載された細胞などの、天然宿主によるGBSの取り込み、または内在化アッセイ法におけるGBSの取り込みを可能とする細胞である。
本発明のポリペプチドは、このようなポリペプチドを含む試料中の生物または生物群の検出に使用することができる。好ましくは、このような検出は診断目的であり、より好ましくは疾患の診断目的であり、最も好ましくはグラム陽性菌(特に連鎖球菌、ブドウ球菌、およびラクトコッカスからなる群より選択される細菌)の存在もしくは数度と関連するか、または結びつく疾患の診断目的である。より好ましくは、微生物はStreptococcus agalactiae、Streptococcus pyogenes、Streptococcus pneumoniae、およびStreptococcus mutansからなる群より選択される。
本発明は、正常レベルおよび異常レベルの決定を含む、細胞および組織における本発明のポリペプチドのレベルを検出する定量アッセイ法や診断アッセイ法などの診断アッセイ法にも関する。したがって例えば、正常対照組織試料と比較してポリペプチドの過剰発現を検出する本発明の診断アッセイ法は、感染の存在を検出する(例えば感染生物を同定する)ために使用することができる。宿主に由来する試料中のポリペプチドのレベルを決定する際に使用可能なアッセイ法は当業者に周知である。このようなアッセイ法には、放射免疫アッセイ法、競合結合アッセイ法、ウェスタンブロット解析、およびELISAアッセイ法などがある。中でもELISAが好ましい場合が多い。ELISAアッセイ法では最初に、対象ポリペプチドに特異的な抗体(好ましくはモノクローナル抗体)を調製する。また一般に、モノクローナル抗体に結合するレポーター抗体を作製する。このようなレポーター抗体に、放射活性試薬、蛍光試薬、または(西洋ワサビペルオキシダーゼ酵素などの)酵素試薬などの検出用試薬を結合させる。
本発明のポリペプチドは、アレイの目的で、またはアレイと関連して使用される場合もある。具体的には、本発明のポリペプチドの少なくとも1つを支持体に固定化することができる。このような支持体は典型的には、1つもしくは複数の本発明のポリペプチド、および/またはこれとは異なるポリペプチドを用いることで多様性が作られるさまざまなポリペプチドを含む。上記のアレイ、ならびに任意のアレイの特徴は一般に、支持体上またはこの表面の特定の、または事前に決定された領域もしくは位置に特定のポリペプチドが固定化されているということである。このため、アレイの特定の位置または領域における任意の活性を特定のポリペプチドと相関させることができる。支持体上に固定化されるさまざまなポリペプチドの数は、わずか10個のポリペプチドから1000個の多様なポリペプチドに至る範囲をとりうる。1 cm2あたりのポリペプチドの密度は、好ましい態様では1 cm2あたりわずか10オリゴヌクレオチドから少なくとも400個の異種ポリヌクレオチドであり、具体的には少なくとも1 cm2あたり1000個の異種ポリペプチドである。
このようなアレイの作製法は当業者に周知であり、例えば米国特許第5,744,309号に記載されている。アレイは好ましくは、第1の表面を有する、平面状で多孔性または非多孔性の固相支持体を含む。本明細書に開示されたポリペプチドが、この表面上に固定化される。好ましい支持材料は、特にガラスまたはセルロースである。アレイが、本明細書に記載された任意の診断応用に使用されることも本発明に含まれる。本発明のポリペプチドのほか、本発明の核酸分子も、上述のアレイの作製に使用することができる。これは、抗体(特に好ましくは本明細書に記載されたモノクローナル抗体)で作製されたアレイにも応用される。
本明細書において本発明の核酸(分子)とも称される、本発明の単離された核酸分子は、本発明のアミノ酸モチーフおよびポリペプチドをコードする。本発明の核酸分子は、第1の変形形態では、SEQ ID No. 1〜10に記載された核酸配列を有する核酸分子と少なくとも70%が同一な核酸(分子)の場合がある。単離された核酸分子が、任意の本発明のポリペプチドをコードする任意の配列に関して少なくとも70%の類似性を有することも本発明に含まれる。好ましくは同一性は少なくとも80%であり、またより好ましくは同一性は少なくとも90%である。同一性は95%、96%、97%、98%、99%、または99.5%の場合もある。
当技術分野で周知で本明細書で用いられる同一性は、2つもしくはこれ以上のポリペプチド配列間の、または2つもしくはこれ以上のポリヌクレオチド配列間の比較によって決定される関連性である。当技術分野では同一性は、場合によっては対象配列間の一致率によって決定されるポリペプチドまたはポリヌクレオチドの配列間の配列関連性の程度も意味する。同一性は容易に計算することができる(Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; and Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991)。2つのポリヌクレオチド配列間、または2つのポリペプチド配列間の同一性を測定する方法がいくつか存在するが、同一性という表現は当業者に周知である(Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991; and Carillo, H., and Lipman, D., SIAM J. Applied Math., 48: 1073 (1988))。同一性を決定する好ましい方法は、検討対象の配列間で最大の一致率を与えるように設計されている。同一性を決定する方法は、コンピュータプログラムで体系化されている。2つの配列間の同一性を決定する好ましいコンピュータプログラム法には、GCGプログラムパッケージ(Devereux, J., et al., Nucleic Acids Research 12 (1): 387 (1984))、BLASTP、BLASTN、およびFASTA(Atschul, S.F. et al., J. Molec. Biol. 215: 403 (1990))などがあるがこれらに限定されない。
本発明の核酸は、第2の変形形態として、上記の第1の変形形態として記載された核酸と少なくとも本質的に相補的な核酸の場合がある。本明細書で用いる「相補的」という表現は、核酸鎖がワトソン-クリック塩基対合を介して第2の核酸鎖と塩基対を形成することを意味する。本明細書で用いる「本質的に相補的である」という表現は、塩基対合が、個々の鎖の塩基のすべてに対しては生じずに、ある程度の数もしくはある程度のパーセンテージの塩基が対合を形成しない状態か、または誤って対合した状態を意味する。正しく対合を形成する塩基のパーセンテージは、好ましくは少なくとも70%であり、より好ましくは80%であり、さらにより好ましくは90%であり、また最も好ましくは90%を上回る任意のパーセンテージである。70%が一致する塩基は相同性があるとみなされ、またこのような規模の一致性塩基対のハイブリダイゼーションがストリンジェントであるとみなされることは重要である。このようなストリンジェントなハイブリダイゼーションのハイブリダイゼーション条件は、Current Protocols in Molecular Biology, John Wiley and Sons, Inc., 1987に記載されている。具体的には、ハイブリダイゼーション条件には以下のような条件がある:
●例えば5×SSPE、5×デンハルト試薬、0.1% SDS、100 g/mL変性DNA中における68℃でのハイブリダイゼーション。
●0.2xSSC、0.1% SDSによる42℃における中ストリンジェンシーによる洗浄。
●0.1xSSC、0.1% SDSによる68℃における高ストリンジェンシーによる洗浄。
●例えば5×SSPE、5×デンハルト試薬、0.1% SDS、100 g/mL変性DNA中における68℃でのハイブリダイゼーション。
●0.2xSSC、0.1% SDSによる42℃における中ストリンジェンシーによる洗浄。
●0.1xSSC、0.1% SDSによる68℃における高ストリンジェンシーによる洗浄。
GC含量が50%であるゲノムDNAのTMは概算で96℃である。1%のミスマッチにつきTMは約1℃低下する。
また、本明細書に記載された別の任意のハイブリダイゼーション条件を原理的に適用することもできる。
本発明の核酸は、第3の変形形態として、概要を前述した本発明の核酸分子の第1および第2の変形形態の少なくとも15塩基の連続核酸を含む核酸の場合がある。好ましくは、このような塩基は、連続した一連の塩基を形成する。しかし、こうした連続が、数塩基で隔てられた2つもしくはこれ以上の部分からなることも本発明に含まれる。
本発明の核酸は、第4の変形形態として、ストリンジェントなハイブリダイゼーション条件で、本発明の上記の第1、第2、および第3の変形形態の任意の核酸とアニールする核酸の場合もある。ストリンジェントなハイブリダイゼーション条件は、典型的な条件が本明細書に記載されている。
また本発明の核酸は、第5の変形形態として、遺伝コードの縮重を除いて、本発明の任意の核酸と、概要を上述した第1、第2、第3、および第4の変形形態の本発明の核酸とハイブリダイズする場合のある核酸の場合もある。このような核酸の存在は、好ましくは本発明の核酸が、本発明のポリペプチドをコードする、したがって付着因子およびインベーシンをそれぞれコードするという事実を意味する。この種の核酸は、本発明の核酸分子の検出に、ひいては同分子の診断に、さらにはGBSなどの個々の微生物、およびこのような微生物が関与する任意の疾患または疾患状態の診断に特に有用である。好ましくはハイブリダイゼーションは、上述の第4の変形形態と関連して記載された、ストリンジェントな条件で生じるか、または実施される場合がある。
本明細書で用いるポリヌクレオチドは一般に、非修飾型のRNAもしくはDNA、または修飾型のRNAもしくはDNAの場合がある任意のポリリボヌクレオチドまたはポリデオキシリボヌクレオチドを意味する。したがって例えば、本明細書で用いる「ポリヌクレオチド」という表現は特に、1本鎖および2本鎖のDNA、1本鎖と2本鎖のRNAとの混合物であるDNA、ならびに1本鎖と2本鎖の領域の混合物であるRNA、1本鎖、またはより典型的には2本鎖もしくは3本鎖、または1本鎖および2本鎖領域の混合物の場合があるDNAおよびRNAを含むハイブリッド分子を意味する。また、本明細書で用いる「ポリヌクレオチド」という表現は、RNAもしくはDNAを、またはRNAとDNAの両方を含む3本鎖領域を意味する。このような領域中の鎖は、同じ分子に由来する場合があるほか、異なる分子に由来する場合がある。このような領域は、1つまたは複数の分子をすべて含む場合があるが、より典型的には、一部の分子の領域のみを含む。3重らせん領域の分子の1つはオリゴヌクレオチドの場合がある。本明細書で用いる「ポリヌクレオチド」という表現は、1つもしくは複数の修飾型の塩基を含む上述のDNAまたはRNAを含む。したがって、安定性または他の理由のために、DNAまたはRNAは、この表現が本明細書で意図される「ポリヌクレオチド」である。また、イノシンなどの一般的でない塩基、またはトリチル化された塩基などの修飾型塩基を含むDNAまたはRNAは、(わずか2つの例を挙げたに過ぎないが)本明細書で用いられるポリヌクレオチドである。当然のことながら、当業者に周知の多くの有用な目的のためにDNAおよびRNAには極めて多様な修飾が施される。本明細書で用いられる「ポリヌクレオチド」という表現は、このような化学的に、酵素学的に、または代謝的に修飾された形状のポリヌクレオチド、ならびにウイルスおよび(特に単純および複雑な細胞を含む)細胞に特徴的な化学的形状のDNAおよびRNAを含む。「ポリヌクレオチド」という表現は、オリゴヌクレオチドと呼ばれることの多い短いポリヌクレオチドも含む。「ポリヌクレオチド」および「核酸」または「核酸分子」という表現は、本明細書で互換的に用いられる場合がある。
本明細書に提供される情報、ならびにクローニング法および配列決定法、またはポリヌクレオチドおよびポリペプチドの合成法を始めとする既知の標準的な方法(例えばSambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989)を参照)を用いて、本発明に含まれる、S. agalactiaeのゲノム、および配列表に記載されたS. agalactiaeのコード領域のポリヌクレオチドおよびポリペプチドと比較して長い場合もあれば短い場合もある数多くの特有の断片を作製することができる。固有の断片であるためには、選択された任意のS. agalactiaeの断片とGenBankなどのコンピュータデータベース上のヌクレオチド配列の比較によって極めて容易に決定される、他の既知の核酸配列と区別するために十分な大きさがなければならない。このような比較探索法は当技術分野で標準的な方法である。多くの固有の断片がS. agalactiaeに特異的である。典型的には、プライマーまたはプローブとして有用な特有の断片の長さは、配列の特定のヌクレオチドの内容に依存して少なくとも約20〜25ヌクレオチドである。また断片は、例えば少なくとも約30ヌクレオチド、40ヌクレオチド、50ヌクレオチド、60ヌクレオチド、75ヌクレオチド、80ヌクレオチド、90ヌクレオチド、100ヌクレオチド、150ヌクレオチド、200ヌクレオチド、250ヌクレオチド、300ヌクレオチド、400ヌクレオチド、500ヌクレオチド、またはこれ以上の長さの場合がある。核酸断片は、意図された目的に応じて1本鎖、2本鎖、または3本鎖の場合がある。
上述のように、また後述されるように、本発明に含まれるS. agalactiaeのポリヌクレオチドおよびポリペプチドに対して修飾が施される場合がある。例えばヌクレオチドの置換は、核酸にコードされたポリペプチドに影響を及ぼさずに施される場合があるので、本発明のポリペプチドをコードする任意のポリヌクレオチドは本発明の範囲に含まれる。また特定のアミノ酸置換(および、特定のアミノ酸をコードする対応ヌクレオチドの置換)を中性とすることができることが当技術分野でわかっている(Robinson W.E. Jr. and Mitchell, W.m., AIDS 4: S141-S162 (1990))。このような変化は、(例えば遺伝的多型に起因する)対立遺伝子変異のように天然の状態で生じる場合があるほか、誘導的な点変異、欠失変異、挿入変異、および置換変異などの、人間の介入で(例えばクローン化されたDNA配列の変異導入によって)作られる場合がある。保存的アミノ酸置換、小さな内部欠失または挿入、および分子の末端における付加または欠失などの、アミノ酸配列上の小さな変化が一般に好ましい。置換は例えばDayhoffらのモデル(Atlas of Protein Sequence and Structure 1978, Nat'l Biomed. Res. Found., Washington D.C.)を元に設計することができる。このような修飾は、アミノ酸配列の変化を生じたり、サイレント変異を生じたり、制限酵素切断部位を修飾したり、または他の特異的な変異を提供したりする場合がある。同様に、このようなアミノ酸変化は、ポリペプチドおよびタンパク質をコードする異なる核酸を生じる。したがって、本発明のパラメータに含まれる別のポリヌクレオチドは、このような修飾の対象となる。
また、配列表に記載されたポリヌクレオチドの配列の一部は、読み枠(ORF)、すなわちS. agalactiaeのコード領域である。各読み枠にコードされたポリペプチドは推定可能なので、ポリペプチドの分子量を、当技術分野で周知のアミノ酸残基の分子量の値から計算することができる。任意の選択されたコード領域を、標準的なサブクローニング法などの標準的な手法で、任意の所望の調節配列(S. agalactiaeの調節配列か、または異種の調節配列かに限らない)に機能的に連結するか、または異種コード配列に連結して、本明細書にさらに記載されている融合タンパク質を作製することができる。
本発明のポリヌクレオチドは、クローン化によって得られるか、化学合成法、またはこれらの組み合わせによって作製されるmRNAもしくはcRNAなどのRNAの形状をとりうるほか、例えばcDNAおよびゲノムDNAを含むDNAの形状をとりうる。このようなDNAは3本鎖、2本鎖、または1本鎖の場合がある。1本鎖DNAは、センス鎖としても知られるコード鎖の場合があるほか、アンチセンス鎖としても知られる非コード鎖の場合もある。
本発明のS. agalactiaeのポリペプチドをコードするコード配列は、配列表に記載されたポリヌクレオチドのコード配列と同一な場合がある。これは、遺伝コードの冗長度(縮重)の結果としてさまざまな配列を有する、配列表に記載されたS. agalactiaeのポリペプチドをコードするポリヌクレオチドの場合もある。
本明細書に記載された、配列表に記載された配列を含むS. agalactiaeのポリペプチドをコードする本発明のポリヌクレオチドは、成熟ポリペプチドそのもののコード配列;成熟ポリペプチドのコード配列と、プレタンパク質の配列、プロタンパク質の配列、またはプレプロタンパク質の配列などの、リーダー配列もしくは分泌配列をコードする配列などの追加的なコード配列;例えば転写に役割を果たす、転写されるが翻訳されない配列(例えば終結シグナルを含む)、リボソーム結合、mRNA安定化エレメントなどの非コード性の5'側配列および3'側配列を含むがこれらに限定されない追加的な非コード配列とともに、前述の追加的なコード配列を含む場合もあれば含まない場合もある成熟ポリペプチドのコード配列、ならびに追加的な機能性を提供するアミノ酸などの追加的なアミノ酸をコードする追加的なコード配列を含む場合があるがこれらに限定されない。したがって例えば、ポリペプチドに、融合ポリペプチドの精製を促進するペプチドなどのマーカー配列を融合させることができる。本発明のこの局面のある態様では、マーカー配列は、多くが市販されている特にpQEベクター(Qiagen, Inc.)などの形で提供されるタグなどのヘキサヒスチジンペプチドである。例えばGentz et al., Proc. Natl. Acad. Sci., USA 86: 821-824 (1989)に記載されているように、ヘキサヒスチジンは融合タンパク質の簡便な精製を可能とする。HAタグは、融合タンパク質を作製するために使用することも可能であり、また例えばWilson et al., Cell 37: 767 (1984)に記載されているインフルエンザの赤血球凝集素タンパク質に由来するエピトープに対応する。本発明のポリヌクレオチドは、構造遺伝子およびこの天然に関連した遺伝的エレメントを含むポリヌクレオチドも含むがこれらに限定されない。
上記の記述に準じて、本明細書で用いる「ポリペプチドをコードするポリヌクレオチド」という表現は、本発明のポリペプチド、特に配列表に記載されたS. agalactiaeのアミノ酸配列を有するポリペプチドをコードする配列を含むポリヌクレオチドを含む。この表現は、ポリペプチドをコードする1つの連続領域または(例えば、組込型ファージもしくは挿入配列、またはエディティングによって分断される)非連続領域を、コード配列および/または非コード配列を含む場合もある追加的な領域とともに含むポリヌクレオチドを含む。
本発明はさらに、配列表に記載されたS. agalactiaeの推定アミノ酸配列を有するポリペプチドの断片、類似体および誘導体をコードする上述のポリヌクレオチドのバリアントに関する。ポリヌクレオチドのバリアントは、天然の対立遺伝子バリアントなどの天然のバリアントの場合があるほか、天然に存在すること知られていないバリアントの場合がある。このようなポリヌクレオチドの非天然のバリアントは、ポリヌクレオチド、細胞、または生物に適用される方法を含む変異導入法で作製することができる。
この点に関するバリアントは、前述のポリヌクレオチドとは、ヌクレオチドの置換、欠失、または付加が異なるバリアントである。置換、欠失、または付加には、1個もしくは複数のヌクレオチドが関与する場合がある。バリアントは、コード領域もしくは非コード領域、またはこの両方を変化させることができる。コード領域を変化させることで、保存的または非保存的なアミノ酸の置換、欠失、または付加を作ることができる。好ましいのは、複数、少数、5〜10残基、1〜5残基、1〜3残基、2残基、1残基、または0残基のアミノ酸が任意の組み合わせで置換、欠失、または付加された、配列表に記載されたS. agalactiae配列を有するバリアント、類似体、誘導体、もしくは断片、または断片のバリアント、類似体、または誘導体をコードするポリヌクレオチドである。中でも特に好ましいのは、配列表に記載されたS. agalactiaeのポリペプチドの性質および活性が変化しないサイレントの置換、付加、および欠失である。またこの点に関しては、保存的置換も特に好ましい。
本発明のさらに好ましい態様は、本発明のポリペプチド、具体的には、配列表記載のアミノ酸配列を有するポリペプチドをコードするポリヌクレオチドと、全長に対して少なくとも70%が同一なポリヌクレオチドであり、またこのようなポリヌクレオチドに相補的なポリヌクレオチドである。あるいは最も極めて好ましいのは、本発明のS. agalactiaeのポリペプチド、具体的には、配列表記載のポリペプチドをコードするポリヌクレオチドと、全長に対して少なくとも80%または少なくとも85%が同一な領域を含むポリヌクレオチドである(相補的ポリヌクレオチドを含む)。この点に関しては、上記ポリヌクレオチドと、全長に対して少なくとも90%、91%、92%、93%、94%、95%、または96%が同一であるポリヌクレオチドが特に好ましく、また特に好ましいポリペプチドの中でも、95%が同一であるポリヌクレオチドが特に好ましい。また少なくとも97%が同一なポリヌクレオチドが、少なくとも95%が同一なポリヌクレオチドの中でも極めて好ましく、少なくとも98%および少なくとも99%が同一なポリヌクレオチドが特に極めて好ましく、少なくとも99%または99.5%が同一なポリヌクレオチドがより好ましい。
さらに、この点に関して好ましい態様は、配列表に記載されたDNAにコードされた成熟ポリペプチドと実質的に同じ生物学的機能または活性を保持するポリペプチドをコードするポリヌクレオチドである。
本発明はさらに、上記の配列とハイブリダイズするポリヌクレオチドに関する。この点に関して本発明は特に、ストリンジェントな条件で上述のポリヌクレオチドとハイブリダイズするポリヌクレオチドに関する。ストリンジェントな条件は、典型的には選択的な条件である。本明細書で用いられる「ストリンジェントな条件」という表現は、少なくとも95%、また好ましくは少なくとも97%が同一な配列間でのみハイブリダイゼーションが起きることを意味する。特定の配列に関して、ストリンジェントな条件は当技術分野で周知であり、また本明細書で例示されるヌクレオチドの内容にしたがって経験的に決定することができる。例えば、ストリンジェントな条件の典型例は、GC含量が55%の48merについて42℃での50%ホルムアミドおよび750 mM NaCl中におけるハイブリダイゼーションと、これに続く55℃での15 mM NaClおよび0.1% SDSによる洗浄である。
例えば本発明のポリヌクレオチドアッセイ法に関して本明細書に追加的に記載されるように、上記の本発明のポリヌクレオチドを、RNA、cDNA、およびゲノムDNAに対するハイブリダイゼーション用プローブとして使用することで、本発明のポリペプチドをコードする完全長のcDNAクローンおよびゲノムクローンを単離することができるほか、本発明のポリヌクレオチドとの配列類似性の高い他の遺伝子のcDNAクローンおよびゲノムクローンを単離することができる。このようなプローブの長さは一般に少なくとも15塩基である。好ましくは、このようなプローブは少なくとも20塩基、少なくとも25塩基、または少なくとも30塩基であり、また少なくとも50塩基の場合がある。特に好ましいプローブは少なくとも30塩基であり、50塩基またはこれ未満(30塩基、35塩基、40塩基、45塩基、または50塩基など)である。
例えば、本発明のポリヌクレオチドのコード領域は、オリゴヌクレオチドプローブを合成するための既知DNA配列を用いたスクリーニングによって単離することができる。次に、本発明の遺伝子の配列と相補的な配列を有する標識オリゴヌクレオチドを用いて、cDNA、ゲノムDNA、またはmRNAのライブラリーのスクリーニングを行って、プローブとハイブリダイズするライブラリー中の分子を決定する。
本発明のポリヌクレオチドおよびポリペプチドは、本明細書では特にポリヌクレオチドアッセイ法と関連して詳述する疾患(特にヒトの疾患)の治療法および診断法の開発用の試薬および材料として使用することができる。
オリゴヌクレオチドの場合がある本発明のポリヌクレオチドを、本明細書に記載された方法(好ましくはPCR)に使用して、本明細書で同定されるS. agalactiaeの遺伝子が、血液などの感染組織内で全体的または部分的に存在するか、および/または転写されるか否かを判定することができる。このような配列が、病原体の感染段階、および感染の種類の診断にも有用でもあることが認識される。この目的および他の目的で、本明細書に記載された本発明の少なくとも1種類の核酸を含むアレイを使用することができる。
本発明のポリヌクレオチドは、成熟タンパク質+追加的なアミノ末端もしくはカルボキシ末端のアミノ酸、または成熟ポリペプチドの内部のポリペプチド(例えば、成熟型が複数のポリペプチド鎖から成る場合)をコードする場合がある。このような配列は、前駆体から成熟型へのタンパク質のプロセシングに役割を果たす場合があり、タンパク質の輸送を可能とする場合があり、タンパク質の半減期を延長もしくは短縮する場合があり、または特にアッセイ法もしくは産生に関するタンパク質の操作を容易にする場合がある。一般にはインビボの場合に、追加的なアミノ酸は、細胞内酵素によって成熟タンパク質から切り出される場合がある。
1つもしくは複数のプロ配列と融合したポリペプチドの成熟型を有する前駆体タンパク質は、不活性型のポリペプチドである場合がある。プロ配列が除去されると、このような不活性型の前駆体は一般に活性化される。プロ配列の一部または全体を除去することで活性化することができる。一般に、このような前駆体はプロタンパク質と呼ばれる。
本発明はさらに、融合ポリペプチドを機能的にコードするポリヌクレオチドを対象とする(融合ポリペプチドは、S. agalactiaeのポリペプチド、および別のS. agalactiaeまたは別の生物のポリペプチドに由来する1つもしくは複数のポリペプチド、または合成ポリアミノ酸配列の断片を含む)。このようなポリヌクレオチドは、適切な条件でS. agalactiaeの他のポリペプチドからのポリペプチドの切断を促進するアミノ酸配列をコードする場合もあればコードしない場合もある。
要約すると、本発明のポリヌクレオチドは好ましくは、成熟タンパク質、成熟タンパク質+リーダー配列(プレタンパク質と呼ばれる場合がある)、1つもしくは複数のプロ配列(プレタンパク質のリーダー配列ではない)を有する成熟タンパク質の前駆体、または一般に、活性型および成熟型のポリペプチドを生じるプロセシング段階で除去されるリーダー配列および1つもしくは複数のプロ配列を有するプロタンパク質の前駆体であるプレプロタンパク質をコードする場合がある。
本明細書で用いる「単離された」という表現は、「人の手によって」天然の状態から分離されることを意味する。すなわち、これが天然の状態で起きる場合は、その当初の環境から変化もしくは除去されるか、または変化および除去される。例えば、生きている生物に天然の状態で存在する天然のポリヌクレオチドまたはポリペプチドは「単離されていない」が、天然の状態で共存する材料から分離される同じポリヌクレオチドまたはポリペプチドは、本明細書で用いられる表現のように「単離されている」。単離の一部、または単離後に、このようなポリヌクレオチドを、例えば変異を導入する目的で、融合タンパク質を形成させる目的で、ならびに宿主で発現させるために、DNAなどの他のポリヌクレオチドに連結させることができる。単離されたポリヌクレオチドは単独で、またはベクターなどの他のポリヌクレオチドに連結した状態で、培養物中の、または生物体全体の宿主細胞に導入することができる。培養物中に、または生物体の宿主細胞に導入されても、このようなDNAは、本明細書で用いられる表現のように単離することができる。というのは、これらは天然の状態または環境に存在しなくなるからである。同様に、ポリヌクレオチドおよびポリペプチドは、培地製剤、ポリヌクレオチドもしくはポリペプチドの(例えば細胞内への)導入用の溶液、例えば、天然の組成物ではなく、内部に単離されたポリヌクレオチドもしくはポリペプチドが、本明細書で用いられる表現の意味において残る化学的もしくは酵素学的な反応用の組成物もしくは溶液などの組成物中に存在する場合がある。
本発明の核酸は化学的に合成することができる。あるいは本発明の核酸は、さまざまな微生物から、当業者に周知の方法で単離することができる。適切な供給源は例えばStreptococcus agalactiae、Streptococcus pyogenes、Streptococcus mutans、およびStreptococcus pneumoniaeである。
本発明の核酸は、核酸、および同核酸を含む生物体または試料の検出に使用することができる。好ましくは、このような検出は診断目的であり、より好ましくは疾患の診断目的であり、最も好ましくはS. agalactiaeの存在もしくは量と関連した、または結びついた疾患の診断目的である。
真核生物(本明細書では「個体」とも呼ぶ)、特に哺乳類(特にヒト)に感染したS. agalactiae菌は、DNAレベルでさまざまな手法で検出することができる。S. agalactiaeの株間で変動する核酸の領域に注目することで、S. agalactiaeの特定の株を区別するための好ましい候補が得られる。さらに、S. agalactiaeと他の生物との間で変動する核酸領域に注目することで、S. agalactiaeと他の生物を区別するための好ましい候補が得られる。診断用の核酸は、感染個体の骨、血液、筋肉、軟骨、および皮膚などの細胞および組織から得られる場合がある。検出にはゲノムDNAを直接使用することが可能なほか、解析に先立ち酵素を用いてPCRで増幅することができる(Saiki et al., Nature、 324: 163-166 (1986))。RNAまたはcDNAを同様に使用することもできる。一例として、本発明のポリヌクレオチドの核酸形成部分に相補的なPCR用プライマーを用いて、その存在および/または発現の同定および解析を行うことができる。哺乳類(特にヒト)に存在するS. agalactiaeの株の特性解析をPCRで、原核生物遺伝子の遺伝子型の解析を元に行うことができる。例えば欠失および挿入を、標準配列の遺伝子型と比較時の増幅産物の大きさの変化によって検出することができる。点突然変異は、増幅DNAと放射標識RNAと、あるいは放射標識アンチセンスDNA配列とハイブリダイズさせることで同定することができる。好ましくは、一致した配列をRNase Aによる切断によって、または溶解温度の差によって一致しなかった2本鎖と区別することができる。
標準遺伝子と、変異を有する遺伝子間の配列の差は、直接的なDNA配列決定によっても明らかとなる。また、クローン化されたDNAセグメントを、特定のDNAセグメント検出するためのプローブとして使用することができる。このような方法の感度は、PCRまたは別の増幅法を適切に用いることで大きく高めることができる。例えば、配列決定用プライマーを、修飾型PCRで生成した2本鎖PCR産物または1本鎖テンプレート分子とともに使用することができる。配列の決定は、放射標識ヌクレオチドを用いる従来の手順で、または蛍光タグを用いる自動配列決定法で実施される。
DNA配列の差に基づく遺伝的特性解析は、変性剤を含む場合もあれば含まない場合もあるゲル上のDNA断片の電気泳動移動度の変化を検出することで行うことができる。小さな配列の欠失および挿入は高解像度ゲル電気泳動で可視化することができる。さまざまな配列のDNA断片は、変性ホルムアミド勾配ゲルで区別することができる。この場合、異なるDNA断片の移動度は、特定の溶解温度または部分溶解温度にしたがってゲル上で異なる位置に遅れて現れる(例えばMyers et al., Science, 230: 1242 (1985)を参照)。
特定の位置における配列の変化も、ヌクレアーゼ(例えばRNase)保護アッセイ法、およびS1保護法、または化学的切断法で明らかにすることができる(例えばCotton et al., Proc. Natl. Acad. Sci., USA, 85: 4397-4401 (1985))。
したがって、特定のDNA配列の検出は、ハイブリダイゼーション、RNase保護、化学的切断、直接DNA配列決定、または制限酵素の使用(例えば制限酵素切断断片長多型;RFLP)、およびゲノムDNAのサザンブロッティングなどの方法で達成することができる。
より従来型のゲル電気泳動およびDNA配列決定に加えて、変異をインサイチュー解析で検出することもできる。
本発明の遺伝子に変異または多型を有する細胞は、例えば血清型の決定を可能とするさまざまな手法によってDNAレベルで検出することもできる。例えばRT-PCRで変異を検出することができる。例えばGeneScanなどの自動検出システムによるRT-PCRの使用が特に好ましい。RNAまたはcDNAも同じ目的でPCRまたはRT-PCRに使用することができる。一例として、本発明のポリペプチドをコードする核酸に相補的なPCR用プライマーを変異の同定および解析に使用することができる。このようなプライマーを使用することで、個体から単離された遺伝子を増幅することができる(次に、このような遺伝子を対象に、さまざまな手法でDNA配列を明らかにする)。このようにしてDNA配列上の変異を診断することができる。
本発明は、個体から単離された、または個体に由来する試料を対象に、配列表に記載されたポリヌクレオチドの配列を有するポリヌクレオチドの発現レベルの上昇を決定する段階を含む、S. agalactiaeの感染から生じる疾患を診断する方法を提供する。ポリヌクレオチドの配列は、例えばPCR、RT-PCR、RNase保護法、ノーザンブロッティング、他のハイブリダイゼーション法、および本明細書に記載されたアレイなどの、ポリヌクレオチドを定量する当技術分野で周知の方法の任意の1つの方法で測定することができる。
本発明は、本発明のポリヌクレオチドまたはポリヌクレオチドを含むベクター、本発明のベクターで遺伝子工学的手法で作製された宿主細胞、および組換え法による本発明のポリペプチドの産生にも関する。
細胞に遺伝子工学的手法で本発明のポリヌクレオチドを取り込ませてポリペプチドを発現させることができる。宿主細胞へのポリヌクレオチドの導入は、リン酸カルシウムトランスフェクション、DEAEデキストランを用いるトランスフェクション、トランスベクション、マイクロインジェクション、陽イオン性脂質介在型トランスフェクション、エレクトロポレーション、形質導入、スクレープローディング、遺伝子銃による導入、感染、または他の方法で実施することができる。これらの方法は、Davis et al., BASIC METHODS IN MOLECULAR BIOLOGY, (1986) and Sambrook et al., MOLECULAR CLONONG: A LABORATORY MANUAL, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)などの多くの標準的な実験マニュアルに記載されている。
細胞内のポリヌクレオチドコンストラクトを従来の方法で使用して、組換え配列にコードされた遺伝子産物を産生させることができる。あるいは本発明のポリペプチドは、従来のペプチド合成装置で合成的に作製することができる。
成熟タンパク質は、哺乳類細胞、酵母、細菌、または他の細胞で、適切なプロモーターの制御下で発現させることができる。無細胞翻訳系で、本発明のDNAコンストラクトに由来するRNAを用いてこのようなタンパク質を産生させることもできる。原核生物および真核生物の宿主に使用される適切なクローニング用および発現用ベクターは、Sambrook et al., MOLECULAR CLONING: A LABORATORY MANUAL, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor. N.Y. (1989)に記載されている。
本発明のこの局面では、ベクターは例えば、プラスミドベクター、1本鎖もしくは2本鎖のファージベクター、1本鎖もしくは2本鎖のRNAもしくはDNAウイルスベクター、1本鎖もしくは2本鎖のRNAもしくはDNAウイルスベクターの場合がある。プラスミドは本明細書では一般に、当業者に周知の標準的な命名法に準じて、小文字のpに続いて、および/または大文字および/または数字が続くように表される。本明細書に開示された出発分子となるプラスミドは市販されているか、公的に入手可能であるか、または入手可能なプラスミドを元に本明細書に記載された周知の公開された手順の常用の応用法で構築することができる。本発明で使用可能な多くのプラスミド、ならびに他のクローニング用および発現用のベクターは周知であり、当業者であれば容易に入手できる。
好ましいベクターは、特定の局面では、本発明のポリヌクレオチドおよびポリペプチドの発現用ベクターである。一般に、このようなベクターは、発現対象のポリヌクレオチドに動作可能に連結された、宿主における発現に有効なシス作用性の制御領域を含む。適切なトランス作用性因子は、宿主によって提供されるか、相補性ベクターによって提供されるか、または宿主への導入時にベクターそのものによって提供される。
この点に関して、特定の好ましい態様では、ベクターは特異的な発現を可能とする。特異的な発現は、誘導型の発現か、ある種の細胞のみにおける発現、または誘導的および細胞特異的な発現の両方の場合がある。特に好ましい誘導型ベクターは、操作が容易な環境因子(温度や栄養添加物など)によって発現させるために誘導可能なベクターである。原核生物および真核細胞に用いられる構成的および誘導的な発現ベクターを含む、本発明のこの局面に適切なさまざまなベクターは当業者に周知であり、常用されている。
極めて多様な発現ベクターを本発明のポリペプチドを発現させるために使用することができる。このようなベクターは特に、いずれも本発明のこの局面における発現に使用することが可能な、染色体、エピソーム、およびウイルス由来ベクター、例えば細菌のプラスミドに由来するベクター、バクテリオファージに由来するベクター、トランスポゾンに由来するベクター、酵母エピソームに由来するベクター、挿入配列に由来するベクター、酵母染色体エレメントに由来するベクター、バキュロウイルスなどのウイルスに由来するベクター、SV40などのパポバウイルス、ワクシニアウイルス、アデノウイルス、鶏痘(fowl pox)ウイルス、仮性狂犬病ウイルス、およびレトロウイルス、ならびにコスミドやファジミドなどのプラスミドおよびバクテリオファージの遺伝的エレメントに由来するベクターなどを含む。一般に、宿主でポリペプチドを発現させるためのポリヌクレオチドの維持、増殖、発現に適切な任意のベクターを、この点に関する発現に使用することができる。
適切なDNA配列をベクターに、例えばSambrook et al., MOLECULAR CLONING, A LABORATORY MANUAL, 2nd Ed.; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989)に記載されている方法などの、さまざまな周知および常用の任意の手法で挿入することができる。
発現ベクターのDNA配列は例えば、mRNAの転写を誘導するプロモーターを含む適切な発現制御配列と動作可能に連結される。このようなプロモーターの代表例には、ファージラムダのPLプロモーター、大腸菌のlac、trp、およびtacプロモーター、SV40の初期および後期プロモーター、ならびにレトロウイルスのLTRプロモーターが含まれるがこれらに限定されない。
一般に発現コンストラクトは、転写の開始および終結に関与する部位を含み、また転写領域では、翻訳に関与するリボソーム結合部位を含む。コンストラクトで発現される成熟転写物のコード部分は、翻訳対象のポリペプチドの末端に適切に位置する翻訳開始のAUG、またはGUGやUUGなどの他のコドンを開始コドンおよび終結コドンに含む。
また発現コンストラクトは、発現を調節ならびに誘導する制御領域を含む場合がある。一般に、いくつかの広く実施されている手順では、このような領域は、転写(転写因子、リプレッサー結合部位、および終結など)を制御することで作用する。
増殖用および発現用のベクターは一般に、選択マーカーおよび、例えばSambrook et al., MOLECULAR CLONONG、 A LABORATORY MANUAL, 2nd Ed.; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989)に記載された領域などの増幅領域を含む。
ベクターの宿主となる適切な細胞の代表例には、連鎖球菌、ブドウ球菌、大腸菌、ストレプトマイセス、および枯草菌細胞などの細菌細胞;酵母細胞およびコウジカビ属の細胞などの真菌細胞;ショウジョウバエのS2細胞およびSpodopteraのSf9細胞などの昆虫細胞;CHO細胞、COS細胞、HeLa細胞、C127細胞、3T3細胞、BHK細胞、293細胞、およびボーズメラノーマ細胞などの動物細胞;ならびに植物細胞がある。
市販されている以下のベクターを例として紹介する。細菌における使用に好ましいベクターには、Qiagen社から入手可能なpQE70、pQE60、およびpQE-9、Stratagene社から入手可能なpBSベクター、Phagescriptベクター、Bluescriptベクター、pNH8A、pNH16a、pNH18A、pNH46A、ならびにPharmacia社から入手可能なptrc99a、pKK223-3、Pkk233-3、pDR540、pRIT5、およびpBR322(ATCC 37017)などがある。好ましい真核ベクターには、Stratagene社から入手可能なpWLNEO、pSV2CAT、pOG44、PXT1、およびpSG、ならびにPharmacia社から入手可能なpSVK3、pBPV、pMSG、およびpSVLなどがある。これらのベクターは、本発明のこの局面で使用するために当業者が入手可能な多くの市販の周知のベクターの説明目的でのみ列挙した。当然のことながら、宿主における本発明のポリヌクレオチドまたはポリペプチドの例えば導入、維持、増殖、または発現に適した他の任意のプラスミドまたはベクターを本発明のこの局面で使用することができる。
プロモーター領域は、クロラムフェニコールアセチルトランスフェラーゼ(「CAT」)転写単位などのプロモーター領域を欠くレポーター転写単位を、制限酵素切断部位すなわち候補プロモーター断片(プロモーターを含む場合がある断片)の導入部位の下流に含むベクターを用いて任意の所望の遺伝子から選択することができる。よく知られているように、cat遺伝子の上流の制限酵素切断部位における、プロモーター含有断片のベクターへの導入は、標準的なCATアッセイ法で検出可能なCAT活性の産生を可能とする。pKK232-8やpCM7などの、この目的に適したベクターは周知であり容易に入手できる。本発明のポリヌクレオチドを発現させるためのプロモーターは、周知かつ容易に入手可能なプロモーターを含むだけでなく、レポーター遺伝子を用いて従来の手法で容易に得られるプロモーターも含む。
本発明のポリヌクレオチドおよびポリペプチドの発現に適切な既知の原核生物プロモーターは、大腸菌のlacIおよびlacZのプロモーター、T3およびT7プロモーター、gptプロモーター、ラムダのPR、PLプロモーター、ならびにtrpプロモーターである。
この点に適した既知の真核生物プロモーターは、CMVの極初期プロモーター、HSVチミジンキナーゼのプロモーター、SV40の初期および後期プロモーター、ラウス肉腫ウイルス「(RSV)」などのレトロウイルスのLTRのプロモーター、ならびにマウスのメタロチオネイン-Iプロモーターなどのメタロチオネインプロモーターである。
組換え発現ベクターは例えば、複製起点、下流の構造配列の転写を誘導する高度に発現される遺伝子に好ましくは由来するプロモーター、およびベクターへの曝露後に、ベクターを含む細胞の単離を可能とする選択マーカーを含む。
本発明のポリペプチドの異種構造配列をコードする本発明のポリヌクレオチドは一般に、発現用のプロモーターに使用可能に連結されるように、標準的な手段でベクターに挿入される。このようなポリヌクレオチドは、転写開始部位が、発現対象ポリペプチドの翻訳を開始するAUGの5'側に来るように位置させる。一般に、開始コドン(通常はAUG)から始まる他の読み枠は、リボソーム結合部位と開始コドンの間にはない。また一般に、ポリペプチドの末端に翻訳停止コドンが位置し、真核宿主における使用のためにポリアデニル化シグナルがコンストラクト内に位置する。転写される領域の3'端に適切に配置される転写終結シグナルは、ポリヌクレオチドコンストラクトに含まれる場合がある。
翻訳されたタンパク質の、小胞体の内腔への、細胞膜周辺腔への、または細胞外環境への分泌のために、適切な分泌シグナルを、発現されるポリペプチドに組み入れることができる。
このようなシグナルは、ポリペプチドに対して内因的な場合があるほか、異種シグナルである場合がある。
ポリペプチドは、融合タンパク質のような修飾型として発現される場合があり、また分泌シグナルだけでなく、異種の他の機能性領域を含む場合もある。したがって例えば、追加的なアミノ酸(特に荷電アミノ酸)の領域を、精製時または後の処理および保存時における、宿主細胞内における安定性および維持を改善するために、ポリペプチドのN末端またはC末端に追加することができる。また精製を容易にする領域をポリペプチドに追加することができる。このような領域は、ポリペプチドの最終的な調製に先立ち除去することができる。分泌または排出を可能とすることの、安定性を改善すること、または精製を促進することなどを目的とした、ペプチド部分のポリペプチドへの付加は、当技術分野における周知かつ常用の手法である。好ましい融合タンパク質は、ポリペプチドの可溶化または精製に有用な免疫グロブリンに由来する異種領域を含む。例えばEP-A-O 464 533(カナダ特許では2045869に相当)は、別のタンパク質またはこの一部と免疫グロブリン分子の定常領域のさまざまな部分を含む融合タンパク質について開示している。拮抗物質を同定する薬剤開発では例えば、高処理能スクリーニングアッセイ法を実施する目的で、タンパク質が抗体のFc部分と融合されている。これについてはD. Bennett et al., Journal of Molecular Recognition, Vol.8, 52-58 (1995) and K. Johanson et al., The Journal of Biological Chemistry, Vol.270, No.16、 pp 9459-9471 (1995)を参照されたい。
細胞は典型的には後に遠心して回収し、物理的または化学的な手段で破壊し、結果として得られる粗抽出物を後の精製用に保持する。
タンパク質の発現に使用される微生物細胞は、凍結融解サイクリング、超音波処理、機械的破壊、または細胞溶解剤の使用を含む従来の任意の方法で破壊することができる。これらの方法は当業者に周知である。
哺乳類の発現ベクターは、複製起点、適切なプロモーター、およびエンハンサーなどの発現配列を含む場合があり、また任意の必要なリボソーム結合部位、ポリアデニル化領域、スプライシングのドナーおよびアクセプター部位、転写終結配列、ならびに発現に有用または必要な5'隣接非転写配列も含む。
ポリペプチドは、硫酸アンモニウムまたはエタノール沈殿、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水的相互作用クロマトグラフィー、アフィニティクロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィーなどの周知の方法で、組換え細胞培養物から回収および精製することができる。最も好ましくは、高処理能液体クロマトグラフィーで精製を行う。単離および/または精製時にポリペプチドが変性する場合には、タンパク質を再び折りたたむ周知の手法で活性構造を再生することができる。
本発明のポリペプチドは、化学合成で、ならびに生物工学的な手段で作製することができる。後者は、当業者に周知の条件における、本発明の核酸を含むベクターによる宿主細胞のトランスフェクションまたは形質転換、ならびにトランスフェクトされた宿主細胞または形質転換された宿主細胞の培養を含む。このような作製法は、作製対象のポリペプチドを精製または単離することを目的とした精製段階を含む場合もある。好ましい態様では、このようなベクターは本発明のベクターである。
別の局面では本発明は、本発明の任意のポリペプチド、誘導体、またはこの断片に対する抗体に関する。本発明は例えば、モノクローナル抗体およびポリクローナル抗体、キメラ抗体、1本鎖抗体、およびヒト化抗体、ならびにFab断片、またはFab発現ライブラリーの産物を含む。抗体がキメラの場合があること、すなわち異なる部分が異なる種に由来すること、または少なくとも個々の配列が異なる種に由来する可能性があることは本発明に含まれる。
本発明の配列に対応するポリペプチドに対する抗体は、ポリペプチドの動物への直接注入によって、またはポリペプチドを動物(好ましくは非ヒト)に投与することで得られる。このようにして得られた抗体は後に、対象ポリペプチドそのものと結合する。このようにして、ポリペプチドの断片のみをコードする配列であっても、天然のポリペプチド全体と結合する抗体の作製に使用することができる。このような抗体は後に、このようなポリペプチドを発現する組織に由来するポリペプチドを単離するために使用することができる。
モノクローナル抗体の調製に関しては、細胞系列の連続培養によって産生される抗体を提供する当技術分野で周知の任意の手法を使用することができる。例には、Kohler, G. and Milstein, C., Nature 256: 495-497 (1975); Kozbor et al., Immunology Today 4: 72 (1983); Cole et al., pg.77-96 in MONOCLONAL ANTIBODIES AND CANCER THERAPY, Alan R. Liss, Inc. (1985); U.S. Patent No. 5,545,403; U.S. Patent No. 5,545,405; U.S. Patent No. 5,654,403; U.S. Patent No. 5,792,838; U.S. Patent No. 5,316,938; U.S. Patent No. 5,633,162; U.S. Patent No. 5,644,036; U.S. Patent No. 5,858,725に記載されている手法などの多様な手法がある。
1本鎖抗体の作製に関して記載されている手法(U.S. Patent No. 4,946,778)を、本発明の免疫原性ポリペプチド産物に対する1本鎖抗体を産生させるために応用することができる。またトランスジェニックマウス、または他の哺乳類などの他の生物を使用して、本発明の免疫原性ポリペプチド産物に対するヒト化抗体を発現させることができる。
あるいはファージディスプレイ法で、抗Fabの保持に関してスクリーニングされたヒトに由来するPCRで増幅されたリンパ球のv-遺伝子のレパートリーか、または未感作ライブラリーのいずれかに由来するポリペプチドに対する結合活性を有する抗体遺伝子を選択することができる(McCafferty, J. et al., (1990), Nature 348, 552-554; Marks, J. et al., (1992) Biotechnology 10, 779-783)。これらの抗体の親和性は、鎖のシャッフリングによって改善することもできる(Clackson, T. et al., (1991) Nature 352, 624-628)。
2つの抗原結合ドメインが存在すると想定すると、個々のドメインは、異なるエピトープに対応する場合がある(「二重特異性」抗体と呼ばれる)。
上述の抗体を用いてポリペプチドを発現するクローンを単離または同定することができるほか、アフィニティクロマトグラフィーによる単離および/または精製を目的とした抗体の固相支持体への結合によって本発明のポリペプチドを精製することができる。
したがって特に、本発明のポリペプチドに対する抗体を用いて、感染(特に細菌感染、また特にS. agalactiaeの感染)を阻害および/または治療することができる。
ポリペプチド誘導体は、本発明の特定の局面を形成する抗原的に、エピトープ的に、または免疫学的に等価な誘導体を含む。本明細書で用いる「抗原的に等価の誘導体」という表現は、本発明のタンパク質またはポリペプチドに対し、病原体と哺乳類宿主間の直接的な物理的相互作用に干渉する特定の抗体によって特異的に認識されるポリペプチドまたはこの等価物を含む。本明細書で用いる「免疫学的に等価の誘導体」という表現は、脊椎動物で、病原体と哺乳類宿主間の直接的な物理的相互作用に干渉するように作用する抗体を生じる適切な剤形で使用されるペプチドまたはこの等価物を含む。
抗原的もしくは免疫学的に等価の誘導体などのポリペプチド、またはこの融合タンパク質は、マウスまたはラットやニワトリなどの他の動物を免疫化するための抗原として使用することができる。融合タンパク質は、ポリペプチドに安定性をもたらす場合がある。抗原は例えば、免疫原性担体タンパク質、例えばウシ血清アルブミン(BSA)、またはキーホールリンペットヘモシアニン(KLH)とコンジュゲーションによって連結させることができる。あるいは、複数のコピーのタンパク質もしくはポリペプチド、またはその抗原的もしくは免疫学的に等価のポリペプチドを含む多数の抗原ペプチドは、免疫原性を改善して担体の使用を回避するように十分な抗原性を有する場合がある。
好ましくは、抗体またはこの誘導体は、個体における免疫原性を低下させるように修飾される。例えば個体がヒトの場合、抗体は最も好ましくは、ハイブリドーマ由来抗体の相補性決定領域が、例えばJones, P. et al., (1986), Nature 321, 522-525 or Tempest at al., (1991) Biotechnology 9, 266-273に記載された手順で、ヒトモノクローナル抗体に移植されるように「ヒト化」することができる。
遺伝的免疫化(genetic immunization)における本発明のポリヌクレオチドの使用には好ましくは、プラスミドDNAの筋肉への直接注入(Wolff et al., (1992) Hum. Mol. Genet. 1, 363; Manthorpe et al., (1963) Hum. Gene Ther. 4, 419)、特定のタンパク質担体と複合体を形成したDNAの導入(Wu et al., (1989) J Biol. Chem. 264, 16985)、DNAとリン酸カルシウムの共沈殿(Benvenisty & Reshef (1986) PNAS 83, 9551)、さまざまな形状のリポソームへのDNAの封入(Kaneda et al., (1989) Science 243, 375)、粒子衝撃法(Tang et al., (1992) Nature 356, 152; Eisenbraun et al., (1993) DNA Cell. Biol. 12, 791)、およびクローン化されたレトロウイルスベクターを用いるインビボ感染(Seeger et al., (1984) PNAS 81, 5849)などの適切な導入法を利用する。
別の局面では本発明は、本発明の任意のポリペプチドに結合するペプチド、およびこのようなペプチドの作製法に関する。ペプチドの作製法は、本発明のポリペプチドの使用を特徴とし、基本的な段階は当業者に周知である。
このようなペプチドは、ファージディスプレイやリボソームディスプレイなどの従来技術の方法で作製することができる。ファージディスプレイの場合は基本的には、ファージの形状をしたペプチドのライブラリーを作製し、標的分子(この場合は本発明のポリペプチド)をライブラリーに接触させる。標的分子と結合するペプチドを次に、好ましくは標的分子との複合体として個々の反応物から除く。結合特性が、塩濃度ほかの特別に設けられる実験条件に少なくともある程度は依存することは当業者に周知である。高親和性すなわち大きな力で標的分子と結合するポリヌクレオチドを、ライブラリーの非結合要素と分離した後に、また任意選択で標的分子とペプチドの複合体から標的分子を除去した後にも、個々のペプチドの特徴を続いて明らかにすることができる。任意選択で、特製解析に先立ち、例えばペプチドをコードするファージを増幅させるなどして増幅段階を実施する。特製解析には好ましくは、標的結合ペプチドの配列決定などがある。しかし基本的には、ペプチドは長さで制限されず、好ましくは約8〜20アミノ酸の長さのペプチドが好ましくは個々の方法で得られる。ライブラリーの大きさは約1012〜1018、好ましくは108〜1015の異なるペプチドであるが、これらに限定されない。
標的結合ポリペプチドの特定の形状は、いわゆる「anticaline」である。これは特にドイツ特許出願DE 197 42 706に記載されている。
別の局面では本発明は、任意の本発明のポリペプチドと相互作用する機能性核酸、およびこのような機能性核酸の作製法に関する(作製法は、本発明のポリペプチドの使用を特徴とし、基本的な段階は当業者に周知である)。機能性核酸は好ましくはアプタマーおよびシュピーゲルマーである。
アプタマーは1本鎖または2本鎖のいずれかであり、標的分子と特異的に相互作用するD-核酸である。アプタマーの作製または選択は、例えば欧州特許EP 0 533 838に記載されている。基本的には、以下の段階を実施する。最初に核酸の混合物、すなわち潜在的なアプタマーを提供する(個々の核酸は典型的には、複数の、好ましくは少なくとも8つの続くランダムヌクレオチドのセグメントを含む)。次にこの混合物に標的分子を接触させる。核酸は、候補混合物と比較して、標的に対する高親和性に基づいて、またはこれに対するより強い力に基づいて標的分子と結合する。次に結合核酸を混合物の残分と分離する。任意選択で、このようにして得られた核酸を例えばポリメラーゼ連鎖反応で増幅する。これらの段階を、標的と特異的に結合する核酸の比率が高い混合物が最終的に得られるまで複数回繰返すことができる(これから、最終的な結合核酸を次に任意選択で選択する)。このような特異的に結合する核酸がアプタマーと呼ばれる。個々の核酸の混合物のアプタマー試料を生成または同定する方法の任意の段階において、その配列を標準的な手段で決定可能なことは明らかである。アプタマーが例えば、アプタマー生成の技術分野の当業者に周知の特定の官能基の導入などによって安定化可能であることは本発明に含まれる。このような修飾は例えば、ヌクレオチドの糖部分の2'位におけるアミノ酸基の導入に存在する場合がある。アプタマーは現在、治療薬として使用されている。しかしながら、このように選択または生成されたアプタマーを標的の検証に、および/または薬物(好ましくは小分子を元にした薬物)開発用のリード物質として使用可能なことも本発明に含まれる。これは実際には、標的分子とアプタマーとの間の特異的な相互作用が、標的とアプタマーとの複合体からアプタマーを置換することに伴い、個々の薬剤候補が、標的とアプタマーとの相互作用の特異的な阻害を可能とし、このとき相互作用が特異的な場合には、候補薬剤が少なくとも原理的には、標的を阻害するのに、ひいてはそのような標的を含む個々の系におけるその生物学的利用能または活性を低下させるのに適していると想定される候補薬剤によって阻害される競合アッセイ法で行われる。次に、このようにして得られた小分子を対象に、さらに誘導体化および修飾を行い、物理的、化学的、生物学的、および/または(毒性、特異性、生分解性、および生物学的利用能などの)医学的な特徴を最適化する。
シュピーゲルマーと、この生成または作製は類似の原理に基づく。シュピーゲルマーの作製はWO 98/08856に記載されている。シュピーゲルマーはL-核酸である。つまりシュピーゲルマーは、アプタマーがD-ヌクレオチドから構成されるのとは異なりL-ヌクレオチドからなる。シュピーゲルマーは、生物系で極めて高い安定性を有すること、またアプタマーと同等に、対象となる標的分子に対して特異的に相互作用することを特徴とする。シュピーゲルマーが生成する過程でD-核酸の非均質な集団が作られ、この集団に、標的分子の光学対掌体(この場合は例えば、本発明のポリペプチドの天然のL-エナンチオマーのD-エナンチオマー)を接触させる。次に、標的分子の光学対掌体と相互作用しないD-核酸を分離する。しかし、標的分子の光学対掌体と相互作用するD-核酸は分離され、任意選択で決定され、および/または配列が決定され、続いて対応するL-核酸が、D-核酸から得られた核酸配列情報を元に合成される。標的分子の光学対掌体と相互作用する前述のD-核酸と、配列に関して同一なこれらのL-核酸は、その光学対掌体とではなく、天然の標的分子と特異的に相互作用する。アプタマー生成法と類似して、さまざまな段階を複数回繰返すことで、標的分子の光学対掌体と特異的に相互作用する核酸を濃縮することもできる。
別の局面では本発明は、本発明の任意の核酸分子と相互作用する機能性核酸、およびこのような機能性核酸の作製法に関する(この方法は、本発明の核酸分子およびこれらの個々の配列の使用、および当業者に周知の基本的な段階を特徴とする)。機能性核酸は好ましくは、リボザイム、アンチセンスオリゴヌクレオチド、およびsiRNAである。
リボザイムは触媒活性のある核酸であり、好ましくはRNAからなり、基本的に2つの部分を含む。第1の部分は触媒活性を示し、第2の部分は標的核酸(この場合は、本発明のポリペプチドをコードする核酸)との特異的な相互作用に関与する。標的核酸とリボザイムの第2の部分との相互作用に伴い、典型的には、2つのハイブリッド形成鎖上の本質的に相補的な連続塩基におけるハイブリダイゼーションおよびワトソン-クリック塩基対形成によって触媒活性部分が活性化される場合がある(つまり、リボザイムの触媒活性がホスホジエステラーゼ活性の場合は、分子内または分子間に標的核酸を触媒することになる)。次に、最終的に標的核酸ならびに標的核酸に由来するタンパク質の分解に至る、標的核酸のさらなる分解が起こる可能性がある。リボザイム、リボザイムの使用法、および設計原理は当業者に周知であり、例えばDohertyおよびDoudna((2001) Ribozyme structures and mechanism. Annu. Rev. Biophys. Biomolstruct. 30, 457-475)、ならびにLewinおよびHauswirth(Ribozyme Gene Therapy: Applications for molecular medicine. 2001, 7: 221-8)に記載されている。
薬物製造用の、または診断薬としてのアンチセンスオリゴヌクレオチドの活性および設計はそれぞれ類似の作用機序に基づく。基本的にアンチセンスオリゴヌクレオチドは、塩基相補性を元に、標的RNA(好ましくはmRNA)とハイブリダイズするのでRNase Hを活性化する。RNase Hは、ホスホジエステルおよびホスホロチオエート結合DNAの両方によって活性化される。しかし、ホスホジエステル結合で結合されたDNAは、細胞ヌクレアーゼによって容易に分解される(例外はホスホロチオエート結合DNA)。このような耐性をもつ非天然のDNA誘導体は、RNAとのハイブリダイゼーション時にRNase Hを阻害しない。言い替えるとアンチセンスポリヌクレオチドは、DNA-RNAハイブリッド複合体としてのみ有効である。この種類のアンチセンスオリゴヌクレオチドの例は特に、米国特許US 5,849,902およびUS 5,989,912に記載されている。言い替えると、個々の核酸配列が原理的に推定される標的タンパク質に由来する標的分子の核酸配列(この場合は、本発明のポリペプチドに対する核酸分子)を元に、または核酸配列(特にmRNA)を明らかにすることで、適切なアンチセンスオリゴヌクレオチドを、塩基相補性の原理に基づいて設計することができる。
特に好ましいのは、短いホスホロチオエートDNA(3〜9塩基)のアンチセンスオリゴヌクレオチドである。最小3残基のDNA塩基が、細菌のRNase Hに活性化に必要であり、最小5塩基が、哺乳類のRNase Hの活性化に必要である。このようなキメラオリゴヌクレオチドでは、RNase Hの基質を形成しない修飾ヌクレオチドからなる「アーム」のハイブリッドの形成によって隣接する、RNase Hの基質を形成する中心領域が存在する。キメラオリゴヌクレオチドのハイブリッド形成アームは、2'-O-メチルや2'-フルオロなどによって修飾することができる。別の方法では、アーム中にメチルホスホン酸またはホスホラミデート結合が用いられている。本発明の実施に有用なアンチセンスオリゴヌクレオチドの別の態様は、P-メトキシオリゴヌクレオチド、部分P-メトキシオリゴデオキシリボヌクレオチド、またはP-メトキシオリゴヌクレオチドである。
本発明で特に重要で有用な点は、上記2つの米国特許で説明されているアンチセンスオリゴヌクレオチドである。このようなオリゴヌクレオチドは天然の5'3'-連結ヌクレオチドを含まない。むしろこのオリゴヌクレオチドは、2種類のタイプのヌクレオチドを有する。1つはRNase Hを活性化する2'-デオキシホスホロチオエートであり、もう1つはRNase Hを活性化しない2'-修飾ヌクレオチドである。2'-修飾ヌクレオチド間の連結は、ホスホジエステル、ホスホロチオエート、またはP-エトキシホスホジエステルの場合がある。RNase Hの活性化は、3〜5個の2'-デオキシホスホロチオエートヌクレオチド(細菌のRNase Hの活性化)、および5〜10個の2'-デオキシホスホロチオエートヌクレオチド(真核生物、特に哺乳類のRNase Hの活性化)を含む連続的なRNase H活性化領域によって達成される。分解は、ヌクレアーゼ耐性の高い5'および3'の末端塩基を作製することで、また任意選択で3'末端にブロッキング基を配置させることによって保護される。
具体的にはアンチセンスオリゴヌクレオチドは、5'端および3'端を、また2'-修飾型ホスホジエステルヌクレオチドおよび2'-修飾型P-アルキルオキシホスホトリエステルヌクレオチドからなる群より独立に選択される11〜59個の5'3'-連結ヌクレオチドを含む(5'端のヌクレオシドは、3〜10個の連続したホスホロチオエート結合デオキシリボヌクレオチドのRNase H活性化領域に結合され、また同オリゴヌクレオチドの3'端は、インバーテッドデオキシリボヌクレオチド(inverted deoxyribonucleotide)、1〜3個のホスホロチオエート2'-修飾型リボヌクレオチドの連続、ビオチン基、およびP-アルキルオキシホスホトリエステルヌクレオチドからなる群より選択される。
また、5'端のヌクレオシドがRNase H活性化領域に結合されていないが3'端ヌクレオシドが上述のようなアンチセンスオリゴヌクレオチドを使用することができる。また5'端は、このようなオリゴヌクレオチドの3'末端ではなく特定のグループから選択される。
本発明の核酸ならびにポリペプチドは、ワクチンとして使用可能なほか、ワクチンの製造に使用可能である。好ましくは、このようなワクチンは、GBSに起因する、GBSに関連する、またはGBSが関与する疾患の予防または治療が目的である。本発明の別の局面は、個体を感染(特に細菌感染、また最も特異的には連鎖球菌感染)から防御する抗体を作製するために適した本発明のポリペプチド、またはこの断片もしくはバリアントを個体に接種する段階を含む、個体(特に哺乳類)で免疫応答を誘導する方法に関する。
本発明のさらに別の局面は、遺伝子療法または他の方法によって、疾患が個体で既に確立しているか否かにかかわらず個体を疾患から防ぐために、抗体、または細胞介在性T細胞応答(サイトカイン産生T細胞または細胞傷害性T細胞のいずれか)を産生させる免疫応答を誘導することを目的として、ポリペプチド、またはこの断片もしくはバリアントをインビボで発現させるためにポリペプチドまたはこの断片もしくはバリアントを機能的にコードする核酸を導入する段階を含む、個体で免疫応答を誘導する方法に関する。遺伝子を投与する1つの方法は、対象遺伝子を、粒子ほかのコーティングとして所望の細胞内に加速して導入することである。
本発明の別の局面は、体内で免疫応答を誘導する能力のある宿主に免疫学的組成物を導入時に、このような宿主で免疫応答を誘導する免疫学的組成物に関する(組成物は、本発明のポリペプチドの抗原ををコードして発現する組換えDNAを含む)。免疫応答は治療的または予防的に使用することが可能であり、また抗体免疫、またはCTLもしくはCD4+ T細胞を生じるような細胞免疫の状態をとりうる。
本発明のポリペプチドまたはこの断片を補助タンパク質と融合することができる。補助タンパク質は、そのものが抗体を産生する場合があるほか、第1のタンパク質を安定化させる能力と、免疫原性および防御的性質をもつ融合タンパク質を産生する能力も有する。この融合組換えタンパク質は好ましくはさらに、タンパク質を可溶化し、タンパク質の産生および精製を促進する比較的大きな補助タンパク質であるグルタチオン-S-トランスフェラーゼ(GST)やベータ-ガラクトシダーゼなどの抗原性補助タンパク質を含む。また補助タンパク質は、全身性の免疫系の刺激を妨げるという意味においてアジュバントとして作用する場合がある。補助タンパク質は、第1のタンパク質のアミノ末端またはカルボキシ末端のいずれかに結合させることができる。
本発明は、Sato, Y. et al., Science 273: 352 (1996)に記載されたような、本発明のポリペプチドまたはポリヌクレオチド、および免疫刺激性のDNA配列を含む組成物、特にワクチン組成物、ならびに方法を提供する。
また本発明は、S. agalactiae感染の動物モデルにおける遺伝学的免疫実験などに使用されるようなDNAコンストラクト中で細菌の細胞表面タンパク質の非可変性領域をコードすることがわかっている、記載されたポリヌクレオチドまたはこの特定の断片を用いる方法を提供する。このような断片は特に、予防的または治療的な免疫応答を誘導可能なタンパク質エピトープを同定する際に有用である。このような方法では、哺乳類(特にヒト)におけるS. agalactiae感染の予防薬剤または治療法の開発上、耐性を良好に示すか、または感染を除外可能な動物の必須臓器から特定の価数のモノクローナル抗体の調製が後に可能となる場合がある。
このようなポリペプチドは例えば、損傷組織に対する細菌の接着を妨げることで細菌の侵入を防御する特定の抗体を産生させるために宿主へのワクチン接種時の抗原として使用することができる。組織損傷の例には例えば、機械的な損傷、化学的な損傷、または熱による損傷によって引き起こされる、または留置装置の設置によって、または口腔、乳腺、尿道、もしくは膣などの粘膜における創傷によって引き起こされる皮膚もしくは結合組織における創傷などがある。
本発明は、免疫原性組換えタンパク質と適切な担体をともにを含むワクチン製剤も含む。このようなタンパク質は胃で分解される可能性があるので、好ましくは例えば、皮下投与、筋肉内投与、静脈内投与、または皮内投与などの経路で非経口的に投与される。非経口投与に適した製剤には、抗酸化剤、緩衝剤、静菌剤、ならびに製剤を個体の体液(好ましくは血液)と等張にする溶質を含む場合がある水性および非水性の滅菌済み注射溶液;ならびに懸濁剤または濃化剤を含む場合がある水性および非水性の無菌性懸濁物などがある。製剤は例えば、密封されたアンプルおよびバイアルなどの単位投与容器または複数回投与容器で提供される場合があるほか、使用直前に滅菌済みの液体担体の添加だけを必要とする、凍結乾燥状態で保存される場合がある。ワクチン製剤は、当技術分野で周知の水中油型系および他の系などの、製剤の免疫原性を高めるためのアジュバント系を含む場合もある。投与量は、ワクチンの特定の活性に依存し、また常用の実験法で容易に決定することができる。
ワクチンが、本発明のポリペプチドおよび/または核酸分子以外に、生物学的または薬学的に活性のある他の化合物を含むことも本発明に含まれる。好ましくはワクチン組成物は、少なくとも1種類のポリカチオンペプチドを含む。本発明で使用されるポリカチオン化合物は、WO 97/30721に記載された特徴的な作用を示す任意のポリカチオン化合物の場合がある。好ましいポリカチオン化合物は、塩基性ポリペプチド、有機ポリカチオン、塩基性ポリアミノ酸またはこの混合物から選択される。これらのポリアミノ酸は、少なくとも4アミノ酸残基の鎖長を有するべきである(WO 97/30721)。特に好ましいのは、8アミノ酸残基、特に20残基を超えるアミノ酸残基、またはこの混合物の塩基性アミノ酸が20%を超える(特に50%を超える)ポリリシン、ポリアルギニン、およびポリペプチドのような物質である。他の好ましいポリカチオン、およびこの薬学的組成物は、WO 97/30721(例えばポリエチレンイミン)、およびWO 99/38528に記載されている。好ましくは、これらのポリペプチドは、20〜500残基(特に30〜200残基)のアミノ酸を含む。
ポリカチオン化合物は、化学的または組換え的に作製することができるほか、天然の供給源に由来する場合がある。
陽イオン性(ポリ)ペプチドは、文献(Ganz et al., 1999; Hancock, 1999)で説明されているように抗菌性を有する場合もある。このような(ポリ)ペプチドは、原核生物または動物もしくは植物に由来する場合があるほか、化学的または組換え的に作製することができる(WO 02/13857)。ペプチドは、ディフェンシン群のものである場合もある(WO 02/13857)。このようなペプチドの配列は例えばAntimicrobial Sequences Database(http://www.bbcm.univ.trieste.it/~tossi/pag2.html)に記載されている。
このような宿主防御ペプチドもしくは防御物質は、本発明のポリカチオンポリマーの好ましい形状でもある。一般に、最終産物として、適応免疫系の活性化(または抑制)を可能とする、好ましくはAPC(樹状細胞を含む)が関与する化合物をポリカチオンポリマーとして使用する。
本発明でポリカチオン物質としての使用に特に好ましいのは、カテリシジン(cathelicidin)に由来する抗菌性ペプチド、またはこの誘導体(参照として本明細書に組み入れられるWO 02/13857)、特に哺乳類のcathelicidin、好ましくはヒト、ウシまたはマウスに由来するcathelicidinに由来する抗菌性ペプチドである。
天然の供給源に由来するポリカチオン化合物には、HIV-REVまたはHIV-TAT(陽イオン性ペプチド派生物、アンテナペディアペプチド、キチンのキトサンもしくは他の誘導体)、または生化学的もしくは組換え的な産生によるこれらのペプチドもしくはタンパク質に由来する他のペプチドなどがある。他の好ましいポリカチオン化合物は、カテリン、またはカテリンの関連物質もしくは派生物質である。例えばマウスのカテリンは、
のアミノ酸配列を有するペプチドである。関連カテリン物質または誘導カテリン物質は、少なくとも15〜20アミノ酸残基の、カテリンの配列の全体または一部を含む。誘導は、天然のアミノ酸と、20種類の標準アミノ酸に含まれないアミノ酸との置換または修飾を含む場合がある。さらに他の陽イオン性残基を、このようなカテリン分子に導入することができる。このようなカテリン分子は、抗原と結合させることが好ましい。このようなカテリン分子は驚くべきことに、別のアジュバントを添加することなく、抗原のアジュバントとしても有効であることがわかっている。したがって、このようなカテリン分子を、さらなる免疫活性物質を使用してもしなくてもワクチン製剤で効率的なアジュバントとして使用することが可能である。
本発明で使用される別の好ましいポリカチオン物質は、3〜7残基の疎水性アミノ酸のリンカーで隔てられた少なくとも2個のKLKモチーフを含む合成ペプチドである(参照として本明細書に組み入れられるWO 02/32451)。
本発明の薬学的組成物は、さらに免疫刺激性核酸を含む場合がある。免疫刺激性核酸は例えば、中性または人工的なCpG含有核酸、非脊椎動物に由来する、または非メチル化シトシングアニンジヌクレオチド(CpG)を特定の塩基内容で含む短いオリゴヌクレオチド(ODN)状の短鎖連続核酸(例えばWO 96/02555)である。あるいは、例えばWO 01/93903に記載されたイノシンおよびシチジンに基づく核酸、またはデオキシイノシン残基および/またはデオキシウリジン残基を含むデオキシ核酸(参照として本明細書に組み入れられるWO 01/93905およびPCT/EP 02/05448)も、好ましくは本発明の免疫刺激性核酸として使用することができる。好ましくは、さまざまな免疫刺激性核酸の混合物を本発明で使用することができる。
前述の任意のポリカチオン化合物が、前述の任意の免疫刺激性核酸と組み合わされることも本発明に含まれる。好ましくは、このような組み合わせは、参照として本明細書に組み入れられるWO 01/93905、WO 02/32451、WO 01/54720、WO 01/93903、WO 02/13857、およびPCT/EP 02/05448、ならびにオーストリア特許出願A 1924/2001に記載されているものの1つである。
加えて、または、あるいは、このようなワクチン組成物は、本発明のポリペプチド/核酸分子のほかに神経刺激性化合物を含む場合がある。好ましくは神経刺激性化合物は、例えばWO 01/24822に記載されているヒト成長因子である。また好ましくは神経刺激性化合物は、前述の任意のポリカチオン化合物および/または免疫刺激性核酸と組み合わされる。
別の局面では本発明は薬学的組成物に関する。薬学的組成物は例えば、本明細書に記載されたワクチンである。また薬学的組成物は、以下の任意の化合物、またはこの組み合わせを含む薬学的組成物である:本発明の核酸、本発明のポリペプチド、本発明のベクター、本発明の細胞、本発明の抗体、本発明の機能性核酸、および本発明のanticalineなどの結合ペプチド、本明細書に記載された方法でスクリーニングされる任意の作動物質および拮抗物質。これと関連して、任意のこれらの化合物を、非滅菌担体もしくは滅菌済みの担体、または被験者への投与に適切な薬学的担体などの細胞、組織、もしくは生物に使用する担体と組み合わせて使用することができる。このような組成物は例えば、培地添加物、または治療的に有効量の本発明のポリペプチド、および薬学的に許容される担体または賦形剤を含む。このような担体は、生理食塩水、緩衝食塩水、デキストロース、水、グリセロール、エタノール、およびこれらの組み合わせを含む場合があるがこれらに限定されない。このような製剤は、投与様式に適したものであるべきである。
薬学的組成物は例えば、局所投与、経口投与、肛門投与、経膣投与、静脈内投与、腹腔内投与、筋肉内投与、皮下投与、経鼻投与、または皮内経路などを含む、任意の有効で簡便な様式で投与することができる。
薬学的組成物は一般に、特定の適応症の治療または予防に有効な量が投与される。一般に組成物は、少なくとも約10 μg/kg体重の活性薬剤量が投与される。多くの場合、組成物は1回または複数の用量が、1日あたり約8 mg/kg体重を超えない量で投与される。好ましくは、多くの場合は投与量は1日あたり約10 μg/kg〜約1 mg/kg体重である。特に哺乳類(特にヒト)への投与に関しては、活性薬剤の1日投与量レベルが0.01 mg/kg〜10 mg/kg(典型的には約1 mg/kg)であることが予想される。例えば用量は1日あたり1 mg/kgの場合がある。当然のことながら最適投与量は、適応症、重症度、投与経路、複雑な状態などを考慮して、個々の治療様式および適応症に標準的な方法で決定される。いずれにせよ医師は、個体に最も適切な、また特定の個人の年齢、体重、および反応によって変動する実際の投与量を決定することになる。上記の投与量は、平均的な場合の典型的な量である。個別の場合で、高い投与量または低い投与量が有効な場合がありうることは言うまでもない。そして、そのような量は本発明の範囲内である。
治療において、または予防目的で、活性薬剤を注射用組成物として、例えば滅菌済みの水性分散液(好ましくは等張液)として個体に投与することができる。
あるいは、このような組成物は、局所投与用に、例えば外用薬、クリーム、ローション、眼軟膏、点眼薬、点耳薬、洗口液、薬剤含浸包帯(impregnated dressing)、ならびに縫合糸およびエアゾル状に製剤化することが可能であり、また例えば保存剤、薬剤の調製を補助する溶媒、ならびに外用薬およびクリーム中の軟化剤を含む適切な従来の添加物を含む場合がある。このような局所用製剤は、適合性のある従来の担体、例えばクリーム、または軟膏基剤、およびローション用のエタノールまたはオレイルアルコールを含む場合がある。このような担体は、製剤の重量の約1〜約98%を占める場合があり、より一般的には、製剤の重量の最大約80%を占める。
薬学的組成物は、留置装置と連結して投与することができる。留置装置には、外科的インプラント、補綴装置、およびカテーテル、すなわち個体の身体に導入されて対象位置に一定期間留まる装置類などがある。このような装置には例えば、人工関節、心臓弁、ペースメーカー、代用血管、血管カテーテル、脳脊髄液シャント、尿管カテーテル、連続式携行式腹膜潅流(CAPD)カテーテルなどがある。
本発明の組成物を留置装置の挿入の直前に、関連細菌に対する全身作用を達成するために注入して投与することができる。処置は、手術後に装置の体内留置時間(in-body time)に継続することができる。また組成物は、連鎖球菌感染を予防するための任意の外科的処置のための術中の覆い(perioperative cover)を拡大するために使用することもできる。
整形外科医の多くは、人工関節使用者は、菌血症を生じる恐れのある歯科治療前に抗生物質による予防を考慮すべきであると考えている。後期深部(late deep)感染は、時に人工関節の喪失につながり、またかなりの罹患率および死亡率を伴う重篤な合併症である。したがって、このような状況では、予防的な抗生物質の代替物としての活性薬剤の使用を延長することが可能な場合がある。
上記の治療に加えて、本発明の組成物を一般に、創傷組織に露出したマトリックスタンパク質に対する細菌の接着を予防するための、および抗生物質による予防に代わる方法、または関連する方法としての、歯科治療における予防的使用のための創傷治療薬剤として使用することができる。
あるいは本発明の組成物を、挿入直前に留置装置を浸すために使用することができる。活性薬剤は、創傷または留置装置を浸す際に好ましくは1 μg/ml〜10 mg/mlの濃度で使用する。
ワクチン組成物は、好都合には注射可能な形状である。免疫応答を高めるために、従来のアジュバントを使用することができる。ワクチン接種に適切な単位用量は0.5〜5 μg/kgの抗原であり、またこのような用量が好ましくは1〜3回、また1〜3週の間隔をおいて投与される。
所定の用量範囲で、適切な個体への投与を除外するような、本発明の化合物による有害な毒物学的作用が観察されるべきではない
上記抗体は、本発明のポリペプチドを含む細菌の存在を検出するための診断薬として使用することができる。
別の態様では本発明は、1種類もしくは複数の本発明の前述の組成物の内容物で満たされた1つまたは複数の容器を含む、診断用および薬学的なパックおよびキットに関する。このような内容物は、有用な量、投与量、製剤、または配合剤で存在する場合がある。このような容器には、ヒトに投与する製品の製造、使用もしくは販売に関連した当局による承認を示す、薬物もしくは生物学的製剤の製造、使用、または販売を管轄する政府機関による、処方形状に関する注意書きを添付することができる。
本発明と関連して例えば、薬学的組成物またはワクチンの用途などの本明細書に開示された任意の疾患関連の用途は特に、グラム陽性菌、具体的には連鎖球菌、ブドウ球菌、およびラクトコッカスからなる群より選択される細菌によって引き起こされたり、関連したり、または関与する疾患または疾患状態である。より好ましくは、このような微生物はS. agalactiae、S. pyogenes、S. pneumoniae、およびS. mutansからなる群より選択される。これと関連してS. agalactiaeが、本明細書に開示された株を始めとする複数の株を含むことは重要である。また疾患は、特に糖尿病や肝不全などの慢性疾患患者、妊娠女性、胎児、および新生児からなる群より選択される任意の患者でみられる疾患の場合がある。本発明の予防および/または治療の対象となる細菌感染と関連する、原因となる、または関与する疾患には、新生児敗血症、肺炎、および髄膜炎、また成体の場合であれば敗血症や軟組織感染などがある。妊娠関連感染は、敗血症、羊膜炎、尿路感染、および死産である。
さらに別の態様では本発明は、本発明の任意のポリペプチドまたは核酸を用いるスクリーニング法に関する。このようなスクリーニング法は当業者には周知であり、作動物質または拮抗物質がスクリーニングされるように設計することができる。好ましくは本発明の場合は、本発明の任意のポリペプチドと相互作用パートナーとの結合を阻害または予防する拮抗物質をスクリーニングする。このような相互作用パートナーは、天然の相互作用パートナーまたは非天然の相互作用パートナーである場合がある。好ましい相互作用パートナーは、FbsAの場合はフィブリノーゲンまたはこの断片であり、またはPabA、PabB、PabC、およびPabDの場合は、上皮細胞(好ましくはヒト上皮細胞)を含む任意の宿主細胞である。
本発明はまた、結合分子との相互作用などの、本発明のポリペプチドまたはポリヌクレオチドの機能を促進する化合物(作動物質)、または阻害する化合物(拮抗物質)をスクリーニングして同定する方法を提供する。スクリーニング法には、高処理能のスクリーニング法が含まれる。
例えば、作動物質または拮抗物質をスクリーニングするために、本発明のポリヌクレオチドおよび核酸のそれぞれの相互作用パートナー、合成反応混合物、膜、細胞エンベロープ、もしくは細胞壁などの細胞区画、またはこれらの任意の調製物を、本発明のポリペプチドと結合する分子を発現する細胞から調製することができる。このような調製物を、作動物質または拮抗物質の場合がある候補分子の存在下もしくは非存在下で標識ポリペプチドとインキュベートする。候補分子が、結合分子と結合する能力は、標識リガンドの結合の低下を反映する。関連なく結合する分子、すなわちポリペプチドの機能性効果を誘導することなく結合する分子は、良好な拮抗物質である可能性が極めて高い。ポリペプチドと同じか、または密接に関連する、良好に結合して機能性効果を引き出す分子は良好な作動物質である。
潜在的な作動物質および拮抗物質の機能的作用は例えば、候補分子と細胞、または適切な細胞調製物との相互作用によりレポーター系の活性を決定すること、ならびにこうした作用を、ポリペプチドと同じ効果を誘発する本発明のポリペプチドまたは分子による作用と比較することで測定できる。この点に関して有用な場合のあるレポーター系には、産物に変換される比色標識された基質、機能性ポリペプチドの活性の変化に反応するレポーター遺伝子、および当技術分野で周知の結合アッセイ法などがあるがこれらに限定されない。
拮抗物質のアッセイ法の別の例は、本発明のポリペプチドおよび潜在的な拮抗物質を、膜に結合した状態の結合分子、組換え結合分子、天然の基質もしくはリガンド、または基質もしくはリガンド模倣物と、競合阻害アッセイ法に適切な条件で混合する競合アッセイ法である。ポリペプチドは、結合分子と結合したり、または産物に変換されたりするポリペプチド分子の数を正確に決定して、潜在的な拮抗物質の有効性を評価することができるように、放射活性化合物または比色化合物によって標識することができる。
潜在的な拮抗物質には、本発明のポリペプチドと結合することで、その活性を阻害するか、または消失させる有機小分子、ペプチド、ポリペプチド、および抗体などがある。潜在的な拮抗物質はまた、本発明のポリペプチドの機能性活性を誘導することなく、結合分子の同じ部位に結合する密接に関連するタンパク質または抗体などの有機小分子、ペプチド、ポリペプチドである場合がある。
潜在的な拮抗物質は、ポリペプチドの結合部位に結合して占有することで、正常な生物学的活性を妨げるような細胞の結合分子との結合を妨げる小分子を含む。小分子の例には有機小分子、ペプチド、またはペプチド様分子などがあるがこれらに限定されない。
他の潜在的な拮抗物質には、アンチセンス分子がある(これらの分子に関する記述に関してはOkano, J. Neurochem. 56: 560 (1991); OLIGODEOXYNUCLEOTIDES AS ANTISENSE INHIBITORS OF GENE EXPRESSION; CRC Press, Boca Raton, FL (1988)を参照)。
好ましい潜在的な拮抗物質には、本発明のポリペプチドの誘導体などがある。
本明細書で用いられるように、本発明のポリペプチドの活性は、任意の相互作用パートナーとの結合能力、または任意の相互作用パートナーとの結合能力の規模である。
特定の局面では本発明は、感染の後遺症に関連する、病原体と哺乳類宿主間の初期の物理的相互作用に干渉する本発明のポリペプチド、ポリヌクレオチド、または阻害剤の用途を提供する。特に本発明の分子は、以下に使用することができる:(i)留置装置上における哺乳類の細胞外マトリックスタンパク質との、または創傷内の細胞外マトリックスタンパク質とS. agalactiaeとの接着の予防;(ii)例えば、哺乳類のチロシンキナーゼのリン酸化の開始によるタンパク質が介在する哺乳類細胞の浸潤の阻害(Rosenshire et al., Infect. Immun. 60: 2211 (1992);(iii)組織損傷が介在する、哺乳類の細胞外マトリックスタンパク質と細菌タンパク質間における細菌の接着の阻害;(iv)留置装置の移植以外によって、または他の外科的手法によって開始された感染における病的変化の正常な進行の阻害。
本明細書で提供される個々のDNAコード配列は、抗菌化合物の発見および開発に使用することができる。コードされたタンパク質を発現させることで、抗菌剤のスクリーニングの標的として使用することができる。また、コードされたタンパク質のアミノ末端領域をコードするDNA配列、または個々のmRNAのShine-Delgarno配列もしくは他の翻訳促進配列を、対象コード配列の発現を制御するためのアンチセンス配列を構築するために使用することができる。
拮抗物質および作動物質を例えば、連鎖球菌(特にS. agalactiae)の感染に起因する疾患(敗血症など)を阻害するために使用することができる。
さらに別の局面で本発明は、少なくとも1つの支持材料と、支持材料に結合された本発明の任意のポリペプチドを含むアフィニティ装置に関する。標的細胞、または標的分子もしくはこれらの相互作用パートナーに対する本発明のポリペプチドの特異性のため、ポリペプチドは、結合に対する条件が適合した場合に、支持試料にアプライされた任意の種類の試料からの相互作用パートナーの選択的な除去を可能とする。試料は発酵ブロス、細胞デブリ、細胞の調製、組織の調製、器官の調製、血液、尿、リンパ液などを含むがこれらに限定されない、生物学的試料または医学的試料の場合がある。
ポリペプチドをマトリックスに共有結合または非共有結合で結合させることができる。適切な支持材料は当業者に周知であり、またセルロース、シリコン、ガラス、アルミニウム、常磁性ビーズ、デンプン、およびデキストランからなる群より選択することができる。
本発明をさらに、特徴、態様、および利点を引き出すことが可能な以下の図面、実施例、および配列表で説明する。実施例は説明目的のみで提供され、開示を制限する意図はないと理解される。
実施例1:実験手順
以下の材料および方法が、特に明記しない限りにおいて、本明細書に記載された実施例の全体で用いられることは重要である。
以下の材料および方法が、特に明記しない限りにおいて、本明細書に記載された実施例の全体で用いられることは重要である。
菌株および培養条件
GBS 6313株(血清型III)およびSS1169株(血清型V)は標準株であり、文献に記載されている(Wibawan and Lammler, 1992)。GBS 706 S2株(血清型Ia)、33H1A株(血清型Ib)、および176 H4A株(血清型II)はG.S. Chhatwal(GBF Braunschweig)から提供された。GBS O90R株(ATCC 12386)は、血清型Ia株O90の派生株である。血清群Ia、Ib、II、III、およびVのすべてのGBS株は臨床単離菌であり、感染新生児から単離され、IV群のGBS株は乳腺炎のウシから単離された(Chhatwal et al., 1984)。大腸菌DH5α(Hanahan, 1985)をクローニング目的で使用し、大腸菌BL21(Dubendorff and Studier, 1991)をFbsA融合タンパク質産生用の宿主として使用した。アルカリホスファターゼ陰性の大腸菌株CC118(Manoil and Beckwith, 1985)をpHRM104誘導体の宿主として、またGBSに由来するシグナルペプチドのコード配列のスクリーニングに使用した。
GBS 6313株(血清型III)およびSS1169株(血清型V)は標準株であり、文献に記載されている(Wibawan and Lammler, 1992)。GBS 706 S2株(血清型Ia)、33H1A株(血清型Ib)、および176 H4A株(血清型II)はG.S. Chhatwal(GBF Braunschweig)から提供された。GBS O90R株(ATCC 12386)は、血清型Ia株O90の派生株である。血清群Ia、Ib、II、III、およびVのすべてのGBS株は臨床単離菌であり、感染新生児から単離され、IV群のGBS株は乳腺炎のウシから単離された(Chhatwal et al., 1984)。大腸菌DH5α(Hanahan, 1985)をクローニング目的で使用し、大腸菌BL21(Dubendorff and Studier, 1991)をFbsA融合タンパク質産生用の宿主として使用した。アルカリホスファターゼ陰性の大腸菌株CC118(Manoil and Beckwith, 1985)をpHRM104誘導体の宿主として、またGBSに由来するシグナルペプチドのコード配列のスクリーニングに使用した。
GBSは、1%酵母エキスを含むTodd-Hewitt酵母ブロス(THY)で37℃で培養した。大腸菌はルリアブロス(LB)で37℃で成長させ、コスミドpTEX5236またはプラスミドpET28aまたはpHRM104を含むクローンを、クロラムフェニコール(15 μg/ml)、カナマイシン(50 μg/ml)またはエリスロマイシン(300 μ
g/ml)の存在下で選択した。アルカリホスファターゼ分泌性大腸菌CC118クローンのスクリーニングは、80 μ/mlのX-リン酸(Sigma)を含むLBプレート上で行った。
g/ml)の存在下で選択した。アルカリホスファターゼ分泌性大腸菌CC118クローンのスクリーニングは、80 μ/mlのX-リン酸(Sigma)を含むLBプレート上で行った。
抗体、酵素、ペプチド、およびヒトタンパク質
アフィニティ精製したウサギ抗フィブリノーゲンおよびペルオキシダーゼ標識抗ウサギ抗体はDako-Biochemicals社から入手した。ペルオキシダーゼ標識ヤギ抗マウス抗体はDianova社から購入した。モノクローナル抗ヒスチジンタグ抗体はRoche Diagnostics社から入手した。精製ウサギ抗フィブロネクチン抗体、トリプシン、プロナーゼ、ビトロネクチン、ラミニン、IgG、フィブロネクチン、およびフィブリノーゲンはSigma-Aldrich社から購入した。フィブリノーゲン(Sigma)は、ゼラチン-セファロースカラムを通過させて、残存する混入性のフィブロネクチンを調製時に除いた。フィブリノーゲン調製物の純度を、SDS-PAGEとクーマシー染色、および抗フィブロネクチン抗体を用いたウェスタンブロッティングで確認した。スポット膜解析用、および阻害実験用の合成ペプチドを文献(Frank and Overwin, 1996)に記載された手順で合成した。
アフィニティ精製したウサギ抗フィブリノーゲンおよびペルオキシダーゼ標識抗ウサギ抗体はDako-Biochemicals社から入手した。ペルオキシダーゼ標識ヤギ抗マウス抗体はDianova社から購入した。モノクローナル抗ヒスチジンタグ抗体はRoche Diagnostics社から入手した。精製ウサギ抗フィブロネクチン抗体、トリプシン、プロナーゼ、ビトロネクチン、ラミニン、IgG、フィブロネクチン、およびフィブリノーゲンはSigma-Aldrich社から購入した。フィブリノーゲン(Sigma)は、ゼラチン-セファロースカラムを通過させて、残存する混入性のフィブロネクチンを調製時に除いた。フィブリノーゲン調製物の純度を、SDS-PAGEとクーマシー染色、および抗フィブロネクチン抗体を用いたウェスタンブロッティングで確認した。スポット膜解析用、および阻害実験用の合成ペプチドを文献(Frank and Overwin, 1996)に記載された手順で合成した。
クローニング目的に使用するプラスミドおよびコスミド
GBS 6313由来のコスミド遺伝子ライブラリー(Reinscheid et al., 2001)を、GBSに由来するfbsA遺伝子の単離に使用した。低コピー数コスミドpTEX5236も、fbsAを含むコスミドのSau3Aによる部分切断後のfbsA遺伝子のサブクローニングに使用した。プラスミドpET28a(Novagen)を、ヘキサヒスチジンタグで標識したFbsA、PabA、PabB、PabC、およびPabD融合タンパク質の合成に使用した(以下の手順で構築)。シグナルペプチドおよび膜貫通ドメインのコード領域を含まない切断型fbsA遺伝子をGBS 6313の染色体DNAからプライマー
および
を用いたPCRで増幅した。fbsAのリピートコード領域をプライマー
および
で増幅した。fbsAの非リピート領域をプライマー
および
で増幅した。クローニングに使用したNcoIおよびBamHI制限酵素切断部位に下線を付した。シグナルペプチドのコード領域を欠くpabA、pabB、pabC、およびpabD遺伝子の増幅(膜貫通ドメインが存在する場合)をプライマー
で行った。
GBS 6313由来のコスミド遺伝子ライブラリー(Reinscheid et al., 2001)を、GBSに由来するfbsA遺伝子の単離に使用した。低コピー数コスミドpTEX5236も、fbsAを含むコスミドのSau3Aによる部分切断後のfbsA遺伝子のサブクローニングに使用した。プラスミドpET28a(Novagen)を、ヘキサヒスチジンタグで標識したFbsA、PabA、PabB、PabC、およびPabD融合タンパク質の合成に使用した(以下の手順で構築)。シグナルペプチドおよび膜貫通ドメインのコード領域を含まない切断型fbsA遺伝子をGBS 6313の染色体DNAからプライマー
クローニングに使用したNcoI、XhoI、およびBamHI、HindIIIの各制限酵素切断部位に下線を付した。PCR産物およびプラスミドpET28aを指定の制限酵素で切断し、連結し、大腸菌BL21を形質転換した。9個の内部配列を有するfbsAを有するプラスミドpETfbsA-9を、pETfbsA-19のXbaIによる部分切断によって構築した後に再連結し、大腸菌BL21を形質転換した。
GBS染色体断片のプラスミドライブラリーを、基本的に文献(Pearce et al., 1993)に記載されている手順でプラスミドpHRM104中に構築した。簡単に説明すると、GBS 6313の染色体DNAを45秒間の超音波処理で分離し、得られた断片をクレノウポリメラーゼで平滑末端化し、SmaIで切断したpHRM104に連結し、この連結混合物で大腸菌CC118を形質転換した。形質転換体を、エリスロマイシンおよびX-リン酸を含む寒天プレートにプレーティングし、3日間インキュベートした。
サザン解析およびブロット解析
GBSの染色体DNAを文献(Pospiech, 1995)に記載された手順で調製した。プラスミドpHRM104中の挿入物のジゴキシゲニン標識プローブを、プライマー
によるPCRで得た。同じプライマーを、pHRM104誘導体中の挿入物の配列決定にも使用した。遺伝子fbsA、pabA/B、およびpabC/Dのそれぞれのジゴキシゲニン標識プローブを、プライマー
によるPCRで得た。GBSのさまざまな臨床単離菌における遺伝子fbsA、pabA/B、およびpabC/Dの分布を解析するために、個々の染色体DNAをHindIII、BstEII、またはNcoIで切断し、fbsA、pabA/B、またはpabB/Cのそれぞれに特異的なプローブとハイブリダイズさせた。標識、ハイブリダイゼーション、洗浄、およびサザンブロッティングによる検出を、Dig標識および検出キット(Roche Diagnostics)を用いて、製造業者の指示書通りに行った後に化学発光法で検出した。
GBSの染色体DNAを文献(Pospiech, 1995)に記載された手順で調製した。プラスミドpHRM104中の挿入物のジゴキシゲニン標識プローブを、プライマー
さまざまなGBS株に由来するfbsAのPCR増幅および配列決定
GBS 706 S2株、33H1A株、176 H4A株、O90R株、およびSS1169株の染色体からfbsA遺伝子を、プライマー
によるPCRで増幅後にPCR産物の配列を決定した。6313株に由来するfbsA遺伝子のヌクレオチド配列を、pTEXfbsAの2.6 kbの挿入物の配列を決定して得た。
GBS 706 S2株、33H1A株、176 H4A株、O90R株、およびSS1169株の染色体からfbsA遺伝子を、プライマー
fbsA欠失変異体の構築
温度感受性プラスミドpG+host6(Appligene)を用いて、GBS 6313株、706 S2株、およびO90R株の各fbsA遺伝子の標的欠失を行った。fbsA遺伝子に隣接する2つの断片を、GBS 6313の染色体DNAを対象としたプライマー対
によるPCRで増幅した。プライマーfbsA_del2およびfbsA_del3中の相補的DNA配列をイタリックで表し、またプライマーfbsA_del1およびfbsA_del4中のBamHIおよびHindIII制限酵素切断部位に下線を付した。fbsA隣接PCR産物を等量で混合し、プライマーfbsA_del1およびfbsA_del4によるクロスオーバーPCRを行った。結果として得られたPCR産物は、1つのDNA断片上にfbsA隣接領域を含んでいた。クロスオーバーPCRの産物およびプラスミドpG+host6をBamHIおよびHindIIIで切断し、連結し、大腸菌DH5αを形質転換した。結果として得られたプラスミドpG+ΔfbsAで、GBS 6313株、706 S2株、およびO90R株をそれぞれ形質転換し、エリスロマイシン寒天上で30℃で成長させて形質転換体を選択した。pG+ΔfbsAが染色体に組込まれた細胞を、文献(Maguin et al., 1996)に記載された手順で、エリスロマイシンによる選択による39℃における形質転換体の成長によって選択した。各株に由来するこのような組込み産物のうち4つを、プラスミドpG+ΔfbsAの切り出しを促して所望のfbsA欠失を染色体上に残すためにエリスロマイシンによる選択を行うことなく液体培地で3日間、30℃で連続継代した。連続継代物を希釈して寒天上にプレーティングし、1個のコロニーを対象にエリスロマイシン感受性について検討を行ってpG+ΔfbsA切除体を同定した。親株にあたるGBS 6313株、706 S2株、およびO90R株、および各株に由来する10個のエリスロマイシン感受性GBS切除体の各染色体DNAを対象に、HindIII切断後に、プライマーfbsA_del3およびfbsA_del4を用いて得られたジゴキシゲニン標識fbsA隣接断片を用いてサザンブロッティングを行った。
温度感受性プラスミドpG+host6(Appligene)を用いて、GBS 6313株、706 S2株、およびO90R株の各fbsA遺伝子の標的欠失を行った。fbsA遺伝子に隣接する2つの断片を、GBS 6313の染色体DNAを対象としたプライマー対
pabAおよびpabB欠失変異体の構築
遺伝子pabAおよびpabAの各欠失変異体をGBS 6313で、fbsA欠失変異体の構築について記載した方法で構築した。pabA欠失変異体の構築にはプライマー対
を使用した。クロスオーバーPCR後に、結果として得られたPCR断片、およびベクターpG+host6をHindIIIおよびEcoRIで切断後に連結し、プラスミドpG+ΔpabAを得た。pabB欠失用のプライマー対として
を使用した。BamHIおよびSalI制限酵素切断部位に下線を付した。クロスオーバーPCRで得られた断片、およびベクターpG+host6をBamHIおよびSalIで切断して連結し、プラスミドpG+ΔpabBを得た。次にプラスミドpG+ΔpabAおよびpG+ΔpabBでGBS 6313を形質転換した。pabAおよびpabBの欠失変異体を作製する手順は、fbsA欠失変異体の構築時の手順と同じである。
遺伝子pabAおよびpabAの各欠失変異体をGBS 6313で、fbsA欠失変異体の構築について記載した方法で構築した。pabA欠失変異体の構築にはプライマー対
一般的なDNAの操作
制限酵素による切断、PCR、連結、エレクトロポレーションによる形質転換、およびサザンブロッティングなどの、DNA操作に関する従来の手法はSambrookらの文献(Sambrook et al., 1989)に記載された手順で実施した。
制限酵素による切断、PCR、連結、エレクトロポレーションによる形質転換、およびサザンブロッティングなどの、DNA操作に関する従来の手法はSambrookらの文献(Sambrook et al., 1989)に記載された手順で実施した。
可溶性125I標識フィブリノーゲンとGBSの結合
精製ヒトフィブリノーゲンを125IでクロマチンT法(Hunter and Greenwood, 1962)で放射標識した。標識フィブリノーゲンとGBSの結合を、基本的に文献(Chhatwal et al., 1983)に記載された手順で実施した。簡単に説明すると、GBSの一晩培養物を遠心して沈殿を得た後に、0.02% Tween 20 (PBST)を添加したリン酸緩衝食塩水で2回洗浄し、600 nmにおける透過率が10%になるように分光光度計で調節した。全体で0.2 mlの細菌の懸濁物に、23 ngのフィブリノーゲンを含む20 μlの125I標識フィブリノーゲンを添加した。室温で1時間のインキュベーション後に連鎖球菌を遠心して沈降させ、1 mlのPBSTで洗浄した。最後にペレットの放射活性をガンマカウンター(Packard Instruments)で測定した。細菌に結合した状態のフィブリノーゲンの量を、細菌に添加した総放射標識フィブリノーゲンのパーセンテージとして算出した。阻害実験では、23 ngの放射標識フィブリノーゲンと、0.2 mlのGBS (T=10%)の結合を、さまざまな量のFbsA融合タンパク質、Bsp融合タンパク質、または合成ペプチドの存在下で決定した。各実験を3通りで少なくとも3回繰返した。
精製ヒトフィブリノーゲンを125IでクロマチンT法(Hunter and Greenwood, 1962)で放射標識した。標識フィブリノーゲンとGBSの結合を、基本的に文献(Chhatwal et al., 1983)に記載された手順で実施した。簡単に説明すると、GBSの一晩培養物を遠心して沈殿を得た後に、0.02% Tween 20 (PBST)を添加したリン酸緩衝食塩水で2回洗浄し、600 nmにおける透過率が10%になるように分光光度計で調節した。全体で0.2 mlの細菌の懸濁物に、23 ngのフィブリノーゲンを含む20 μlの125I標識フィブリノーゲンを添加した。室温で1時間のインキュベーション後に連鎖球菌を遠心して沈降させ、1 mlのPBSTで洗浄した。最後にペレットの放射活性をガンマカウンター(Packard Instruments)で測定した。細菌に結合した状態のフィブリノーゲンの量を、細菌に添加した総放射標識フィブリノーゲンのパーセンテージとして算出した。阻害実験では、23 ngの放射標識フィブリノーゲンと、0.2 mlのGBS (T=10%)の結合を、さまざまな量のFbsA融合タンパク質、Bsp融合タンパク質、または合成ペプチドの存在下で決定した。各実験を3通りで少なくとも3回繰返した。
FITC標識GBSと固定化フィブリノーゲンの結合
テラサキプレートをヒトフィブリノーゲンでコーティングし、FITC標識細菌と固定化フィブリノーゲンの結合を、Podbielskiら(Podbielski et al., 1999)の手順で測定した。簡単に説明すると、10 μlの100 μg/mlのヒトフィブロネクチンフィブリノーゲン、ラミニン、およびコラーゲンIおよびIVの各ストック溶液を各ウェルに添加し、湿室で室温で一晩インキュベートした。次にこのマイクロタイタープレートをPBSで洗浄し、残存する緩衝液を十分除去した。GBSのFITC標識を、指数増殖期(OD600:0.5)の培養物、および定常期(OD600:1.5)の培養物を対象に実施した。12 mlの細菌培養物を遠心して沈殿させ、12 mlのPBSで洗浄し、2 mlのFITC溶液(1 mg/ml FITC、溶媒は50 mM 炭酸ナトリウム緩衝液、pH 9.2)に再懸濁した。暗所での20分間のインキュベーション後に、遠心して細胞を沈殿させ、PBSで2回洗浄し、20秒間の超音波処理を行って細菌の鎖構造を破壊した。細菌懸濁物をPBSでOD600が1.0となるように調整し、ボルテックスミキサーで激しく攪拌した後に、使用時まで暗所で保存した。10 μlのFITC標識GBS懸濁物を、さまざまなヒトタンパク質でコーティングした各テラサキウェルに添加した。37℃で60分間のインキュベーション後に、非結合細菌をPBSで5回の洗浄して除去し、結合細菌を0.5%グルタルアルデヒドで5分間かけて固定した。このプレートを最後にPBSで2回洗浄し、各ウェルの蛍光を自動Cyto Fluor II蛍光リーダー(PerSeptive Biosystems)で決定した(励起波長=485 nm、検出波長=530 nm)。500 μlのFITC標識細菌と37℃で60分間インキュベートし、細菌をPBSで3回洗浄し、細胞を500 μlのPBSに再懸濁し、コーティングを施していないテラサキマイクロタイタープレート上の10 μlの懸濁物のアリコートの蛍光を測定して細菌のFITC標識の効率を決定した。各アッセイ物を3通りで測定し、少なくとも4回繰返した。
テラサキプレートをヒトフィブリノーゲンでコーティングし、FITC標識細菌と固定化フィブリノーゲンの結合を、Podbielskiら(Podbielski et al., 1999)の手順で測定した。簡単に説明すると、10 μlの100 μg/mlのヒトフィブロネクチンフィブリノーゲン、ラミニン、およびコラーゲンIおよびIVの各ストック溶液を各ウェルに添加し、湿室で室温で一晩インキュベートした。次にこのマイクロタイタープレートをPBSで洗浄し、残存する緩衝液を十分除去した。GBSのFITC標識を、指数増殖期(OD600:0.5)の培養物、および定常期(OD600:1.5)の培養物を対象に実施した。12 mlの細菌培養物を遠心して沈殿させ、12 mlのPBSで洗浄し、2 mlのFITC溶液(1 mg/ml FITC、溶媒は50 mM 炭酸ナトリウム緩衝液、pH 9.2)に再懸濁した。暗所での20分間のインキュベーション後に、遠心して細胞を沈殿させ、PBSで2回洗浄し、20秒間の超音波処理を行って細菌の鎖構造を破壊した。細菌懸濁物をPBSでOD600が1.0となるように調整し、ボルテックスミキサーで激しく攪拌した後に、使用時まで暗所で保存した。10 μlのFITC標識GBS懸濁物を、さまざまなヒトタンパク質でコーティングした各テラサキウェルに添加した。37℃で60分間のインキュベーション後に、非結合細菌をPBSで5回の洗浄して除去し、結合細菌を0.5%グルタルアルデヒドで5分間かけて固定した。このプレートを最後にPBSで2回洗浄し、各ウェルの蛍光を自動Cyto Fluor II蛍光リーダー(PerSeptive Biosystems)で決定した(励起波長=485 nm、検出波長=530 nm)。500 μlのFITC標識細菌と37℃で60分間インキュベートし、細菌をPBSで3回洗浄し、細胞を500 μlのPBSに再懸濁し、コーティングを施していないテラサキマイクロタイタープレート上の10 μlの懸濁物のアリコートの蛍光を測定して細菌のFITC標識の効率を決定した。各アッセイ物を3通りで測定し、少なくとも4回繰返した。
融合タンパク質の調製および精製
さまざまなFbsA融合タンパク質、ならびに融合タンパク質PabA、PabB、PabC、PabD、およびBsp(Reinscheid et al., 2002)を、組換え大腸菌BL21を用いて、培養物の光学密度が1.0に到達後に1 mM IPTGを添加して合成した。細胞をフレンチプレスで破壊し、融合タンパク質の精製をNi2+アフィニティクロマトグラフィーでQiagen社の指示書に従って行った。次にPabA、PabB、およびPabC融合タンパク質を、20 mM Tris/HCl、pH 8.5で透析し、MonoQ陰イオン交換カラム(Amersham/Pharmacia)にロードした。0 M〜1.0 MのNaCl(溶媒は20 mM Tris/HCl)の直線勾配を用いて融合タンパク質をカラムから溶出した。PabDをさらに精製するために、融合タンパク質を20 mM Tris/HCl緩衝液で透析し、MonoS陽イオン交換カラム(Amersham/Pharmacia)にロードした。0 M〜1.0 M NaCl(溶媒は20 mM Tris/HCl緩衝液)の直線勾配でPabDを溶出した。最後にすべての融合タンパク質をPBSで透析し、-20℃で保存した。
さまざまなFbsA融合タンパク質、ならびに融合タンパク質PabA、PabB、PabC、PabD、およびBsp(Reinscheid et al., 2002)を、組換え大腸菌BL21を用いて、培養物の光学密度が1.0に到達後に1 mM IPTGを添加して合成した。細胞をフレンチプレスで破壊し、融合タンパク質の精製をNi2+アフィニティクロマトグラフィーでQiagen社の指示書に従って行った。次にPabA、PabB、およびPabC融合タンパク質を、20 mM Tris/HCl、pH 8.5で透析し、MonoQ陰イオン交換カラム(Amersham/Pharmacia)にロードした。0 M〜1.0 MのNaCl(溶媒は20 mM Tris/HCl)の直線勾配を用いて融合タンパク質をカラムから溶出した。PabDをさらに精製するために、融合タンパク質を20 mM Tris/HCl緩衝液で透析し、MonoS陽イオン交換カラム(Amersham/Pharmacia)にロードした。0 M〜1.0 M NaCl(溶媒は20 mM Tris/HCl緩衝液)の直線勾配でPabDを溶出した。最後にすべての融合タンパク質をPBSで透析し、-20℃で保存した。
フィブリノーゲン結合コロニーのスクリーニング
コスミドを含む大腸菌クローンを、テトラサイクリンを含む2枚のLBプレートに移して一晩インキュベートした。翌日、1枚のプレート上のコロニーを、ニトロセルロース上に6時間かけて移した。同フィルター上の細胞をクロロホルム蒸気で20分間かけて溶解後に、1 mg/mlリゾチームおよび1 mM PMSFを含むPBSで一晩インキュベートした。この膜を10%のスキムミルク(溶媒はリン酸緩衝食塩水(PBS))で一晩かけてブロック処理した後に、後述する手順でヒトフィブリノーゲンとの結合をみるためにプローブで処理した。
コスミドを含む大腸菌クローンを、テトラサイクリンを含む2枚のLBプレートに移して一晩インキュベートした。翌日、1枚のプレート上のコロニーを、ニトロセルロース上に6時間かけて移した。同フィルター上の細胞をクロロホルム蒸気で20分間かけて溶解後に、1 mg/mlリゾチームおよび1 mM PMSFを含むPBSで一晩インキュベートした。この膜を10%のスキムミルク(溶媒はリン酸緩衝食塩水(PBS))で一晩かけてブロック処理した後に、後述する手順でヒトフィブリノーゲンとの結合をみるためにプローブで処理した。
ウェスタンブロッティングトおよびスポット膜解析
ウェスタンブロット実験では、タンパク質をSDS-PAGEで分離し、ニトロセルロースに電気的にブロットした。次にこの膜を10%スキムミルク(溶媒PBS)で一晩ブロック処理した。スポット膜実験では、16アミノ酸からなるペプチドを合成し、文献に記載された手順で等量のペプチドをセルロースペーパー上に直接スポットした(Frank and Overwin, 1996)。ブロッキングは20 mlのカゼインベースのブロッキング緩衝液(Genosys Biotechnologies, Cambridge, England)、80 mlのTris緩衝食塩水(TBS)、0.05% tween 20、および5 gのショ糖を含む膜ブロッキング溶液(MBS)で行った。フィブリノーゲン結合に関するプローブ処理は、後述する手順で実施した。
ウェスタンブロット実験では、タンパク質をSDS-PAGEで分離し、ニトロセルロースに電気的にブロットした。次にこの膜を10%スキムミルク(溶媒PBS)で一晩ブロック処理した。スポット膜実験では、16アミノ酸からなるペプチドを合成し、文献に記載された手順で等量のペプチドをセルロースペーパー上に直接スポットした(Frank and Overwin, 1996)。ブロッキングは20 mlのカゼインベースのブロッキング緩衝液(Genosys Biotechnologies, Cambridge, England)、80 mlのTris緩衝食塩水(TBS)、0.05% tween 20、および5 gのショ糖を含む膜ブロッキング溶液(MBS)で行った。フィブリノーゲン結合に関するプローブ処理は、後述する手順で実施した。
ウェスタンブロッティング、スポット膜解析、およびコロニーブロッティングによるフィブリノーゲン結合の検出
一晩ブロック処理した膜を、2 μg/mlのヒトフィブリノーゲンと1時間インキュベートした。ウェスタンブロット実験およびコロニーブロット実験では、フィブリノーゲンおよび抗体をPBSで希釈し、スポット膜解析ではMBSで希釈した。PBSで3回洗浄した後に、膜を抗フィブリノーゲン抗体(1:1000;溶媒はPBSまたはMBS)で1時間インキュベートした。インキュベーション後に0.05% tween 20 (PBST)を含むPBSで3回洗浄し、PBSで2回洗浄した。次に、この膜をペルオキシダーゼ標識抗ウサギIgG(1:1000 in PBSまたはMBS)で1時間インキュベートした。PBSTによる3回の洗浄と、PBSによる2回の洗浄後に、結合状態のフィブリノーゲンをECLキット(Amersham/Pharmacia)で化学発光により検出した。対照実験では、固定化されたタンパク質およびペプチドと使用抗体との間で交差反応は認められなかった。
一晩ブロック処理した膜を、2 μg/mlのヒトフィブリノーゲンと1時間インキュベートした。ウェスタンブロット実験およびコロニーブロット実験では、フィブリノーゲンおよび抗体をPBSで希釈し、スポット膜解析ではMBSで希釈した。PBSで3回洗浄した後に、膜を抗フィブリノーゲン抗体(1:1000;溶媒はPBSまたはMBS)で1時間インキュベートした。インキュベーション後に0.05% tween 20 (PBST)を含むPBSで3回洗浄し、PBSで2回洗浄した。次に、この膜をペルオキシダーゼ標識抗ウサギIgG(1:1000 in PBSまたはMBS)で1時間インキュベートした。PBSTによる3回の洗浄と、PBSによる2回の洗浄後に、結合状態のフィブリノーゲンをECLキット(Amersham/Pharmacia)で化学発光により検出した。対照実験では、固定化されたタンパク質およびペプチドと使用抗体との間で交差反応は認められなかった。
オプソニン貧食アッセイ法
食作用に対する耐性を文献(Podbielski et al., 1996)に記載された手順で測定した。簡単に説明すると、成長させたGBS培養物を103コロニー形成単位/mlに調整した。100 μlの懸濁物を300 μlのヘパリン処理ヒト血液に添加し、反応混合物を周回式回転で37℃で3時間インキュベートした。インキュベーション前後のアリコートを段階希釈し、THY寒天にプレーティングして一晩培養した。各株について、インキュベーション前と、ヒト血液との3時間のインキュベーション後におけるコロニー形成単位の比を算出した。各実験は3通りで3回実施した。
食作用に対する耐性を文献(Podbielski et al., 1996)に記載された手順で測定した。簡単に説明すると、成長させたGBS培養物を103コロニー形成単位/mlに調整した。100 μlの懸濁物を300 μlのヘパリン処理ヒト血液に添加し、反応混合物を周回式回転で37℃で3時間インキュベートした。インキュベーション前後のアリコートを段階希釈し、THY寒天にプレーティングして一晩培養した。各株について、インキュベーション前と、ヒト血液との3時間のインキュベーション後におけるコロニー形成単位の比を算出した。各実験は3通りで3回実施した。
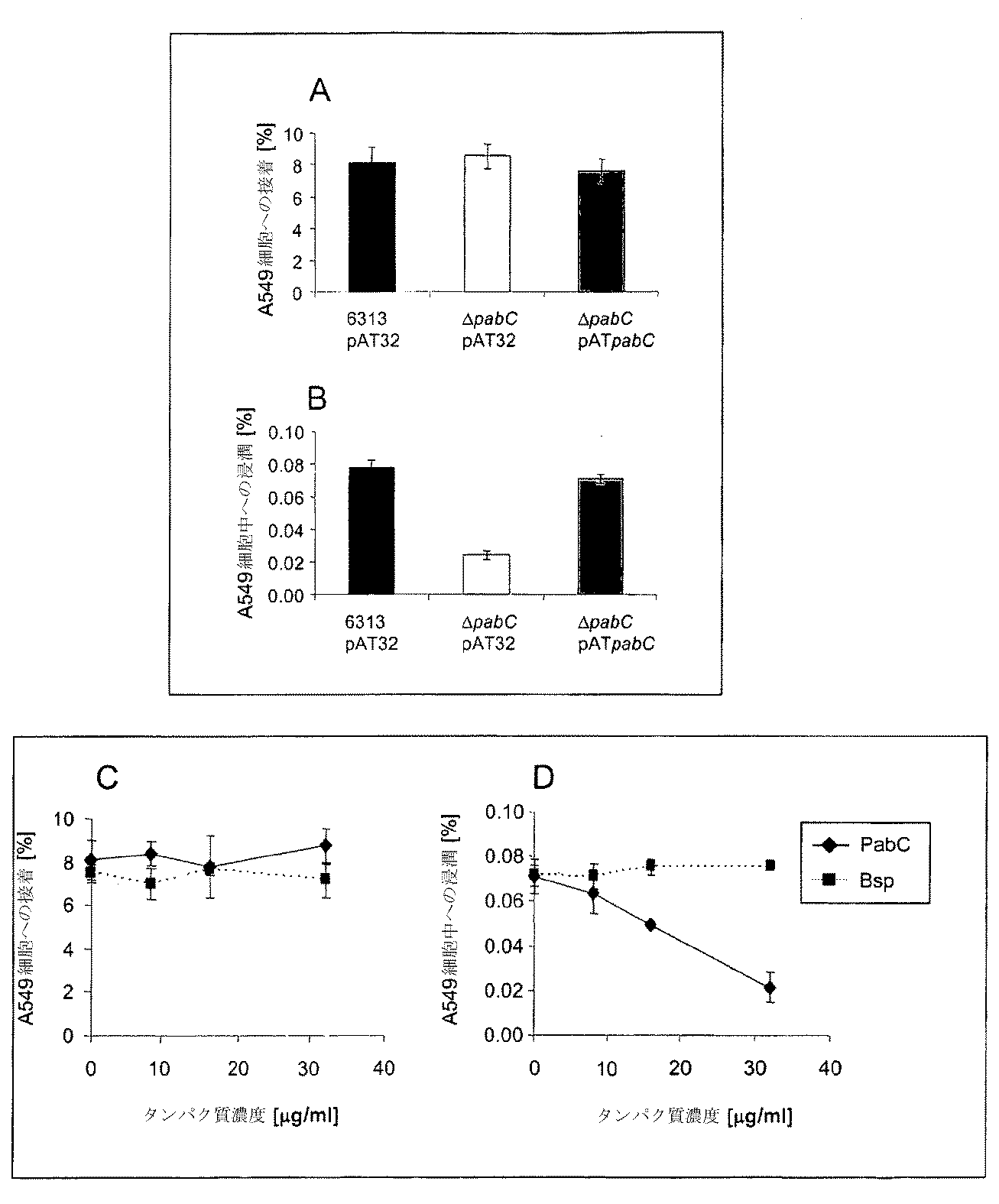
上皮細胞との接着および内在化のアッセイ法
GBSの上皮細胞に対する接着、および上皮細胞への内在化を、基本的に文献(Caparon et al., 1991; Rubens et al., 1992)に記載された手順で調べた。簡単に説明すると、A549細胞を24ウェル組織培養プレートに1ウェルあたり細胞が約4×105個となるように移し、10%ウシ胎児血清を添加したRPMI(Gibco BRL)組織培地で一晩培養した。培地を1 mlの新鮮な培地と交換後に、細胞に1ウェルあたり5×106個の連鎖球菌を感染させ、37℃で2時間インキュベートした。非接着性細菌はPBSで3回洗浄して除去した。接着アッセイ法では、次にトリプシン/EDTAを添加して上皮細胞をウェルから遊離させ、300 μlの滅菌水を添加して溶解した。溶解物の段階希釈物をTHY寒天プレートにプレーティングして接着性細菌を定量した。内在化アッセイ法では、上皮細胞を2時間の感染後に、ペニシリンG(10 U)、およびストレプトマイシン(0.01 mg)を添加した組織培地中でさらに2時間インキュベートして、細胞外に存在する細菌を死滅させた。PBSで3回洗浄した後に、トリプシン/EDTAを添加して上皮細胞をウェルから遊離させ、300 μlの滅菌水に溶解した。細胞内細菌の量を、溶解物の段階希釈物をTHY寒天プレートにプレーティングすることで定量した。各実験を3通りで少なくとも3回繰返した。
GBSの上皮細胞に対する接着、および上皮細胞への内在化を、基本的に文献(Caparon et al., 1991; Rubens et al., 1992)に記載された手順で調べた。簡単に説明すると、A549細胞を24ウェル組織培養プレートに1ウェルあたり細胞が約4×105個となるように移し、10%ウシ胎児血清を添加したRPMI(Gibco BRL)組織培地で一晩培養した。培地を1 mlの新鮮な培地と交換後に、細胞に1ウェルあたり5×106個の連鎖球菌を感染させ、37℃で2時間インキュベートした。非接着性細菌はPBSで3回洗浄して除去した。接着アッセイ法では、次にトリプシン/EDTAを添加して上皮細胞をウェルから遊離させ、300 μlの滅菌水を添加して溶解した。溶解物の段階希釈物をTHY寒天プレートにプレーティングして接着性細菌を定量した。内在化アッセイ法では、上皮細胞を2時間の感染後に、ペニシリンG(10 U)、およびストレプトマイシン(0.01 mg)を添加した組織培地中でさらに2時間インキュベートして、細胞外に存在する細菌を死滅させた。PBSで3回洗浄した後に、トリプシン/EDTAを添加して上皮細胞をウェルから遊離させ、300 μlの滅菌水に溶解した。細胞内細菌の量を、溶解物の段階希釈物をTHY寒天プレートにプレーティングすることで定量した。各実験を3通りで少なくとも3回繰返した。
競合試験では、50 μgの精製融合タンパク質、または1 μgのフィブリノーゲンを含む1 mlの新鮮な組織培地をA549細胞に添加した後に、GBS 6313を細胞に感染させた。
タンパク質でコーティングしたラテックスビーズとA549細胞の相互作用
約108個のラテックスビーズ(直径3 μm、Sigma)をPBSで3回洗浄後に、500 μlのPBSに溶解した300 μgの融合タンパク質またはBSAで4℃で一晩かけてコーティングした。コーティング処理後のビーズをPBSで1回洗浄した後に、200 μlの10 mg/ml BSA(溶媒PBS)で室温で1時間かけてブロック処理した。ビーズをPBSで2回、RPMI+10% FCSで1回洗浄後に、1 mlのRPMI+10% FCSに再懸濁した。24ウェルプレート中の約4×105個のA549細胞に300 μlのビーズを添加した。細胞を37℃で1時間(5% CO2)インキュベートし、PBSで5回洗浄し、3%グルタルアルデヒドおよび5%ホルムアルデヒド(溶媒はカコジル酸緩衝液)を含む溶液で、氷上で45分間かけて固定した。この試料をカコジル酸緩衝液で洗浄し、一連の勾配のアセトンで脱水し、CO2で臨界点乾燥を行った。次に試料を、厚さ10 nmの金フィルムでコーティングし、走査型電子顕微鏡を用いて文献記載の手順で観察した(Reinscheid et al., 2001)。
約108個のラテックスビーズ(直径3 μm、Sigma)をPBSで3回洗浄後に、500 μlのPBSに溶解した300 μgの融合タンパク質またはBSAで4℃で一晩かけてコーティングした。コーティング処理後のビーズをPBSで1回洗浄した後に、200 μlの10 mg/ml BSA(溶媒PBS)で室温で1時間かけてブロック処理した。ビーズをPBSで2回、RPMI+10% FCSで1回洗浄後に、1 mlのRPMI+10% FCSに再懸濁した。24ウェルプレート中の約4×105個のA549細胞に300 μlのビーズを添加した。細胞を37℃で1時間(5% CO2)インキュベートし、PBSで5回洗浄し、3%グルタルアルデヒドおよび5%ホルムアルデヒド(溶媒はカコジル酸緩衝液)を含む溶液で、氷上で45分間かけて固定した。この試料をカコジル酸緩衝液で洗浄し、一連の勾配のアセトンで脱水し、CO2で臨界点乾燥を行った。次に試料を、厚さ10 nmの金フィルムでコーティングし、走査型電子顕微鏡を用いて文献記載の手順で観察した(Reinscheid et al., 2001)。
ビオチン化ペプチドの合成
Rinkアミド樹脂(PepChem, Tubingen, Germany)上で、SyroII合成装置(Multisyntech, Witten, Germany)を用いて標準F-moc化学法でペプチドを小規模合成した(4 mgの樹脂;平行して最大288個)。配列をアセンブルした後に、ペプチドをFmoc-イプシロン-アミノヘキサン酸(リンカーとして使用)、およびビオチン(Sigma, St. Louis, MO;正常アミノ酸のように活性化)で延長させた。93% TFA、5%トリエチルシラン、および2%水で1時間処理してペプチドを樹脂から切り離した。ペプチドを真空下で乾燥し、アセトニトリル/水(1:1)で3回凍結乾燥した。正確な質量の存在を、Reflex III MALDI-TOF(Bruker, Bremen Germany)による質量分析で検証した。このペプチドを、それ以上精製することなく使用した。
Rinkアミド樹脂(PepChem, Tubingen, Germany)上で、SyroII合成装置(Multisyntech, Witten, Germany)を用いて標準F-moc化学法でペプチドを小規模合成した(4 mgの樹脂;平行して最大288個)。配列をアセンブルした後に、ペプチドをFmoc-イプシロン-アミノヘキサン酸(リンカーとして使用)、およびビオチン(Sigma, St. Louis, MO;正常アミノ酸のように活性化)で延長させた。93% TFA、5%トリエチルシラン、および2%水で1時間処理してペプチドを樹脂から切り離した。ペプチドを真空下で乾燥し、アセトニトリル/水(1:1)で3回凍結乾燥した。正確な質量の存在を、Reflex III MALDI-TOF(Bruker, Bremen Germany)による質量分析で検証した。このペプチドを、それ以上精製することなく使用した。
酵素結合免疫アッセイ法(ELISA)
ビオチン標識ペプチド(10 μg/ml)でストレプトアビジンELISAプレート(EXICON)を製造業者の指示書に従ってコーティングした。血清を2種類の希釈率(200倍および1,000倍)で検討した。
ビオチン標識ペプチド(10 μg/ml)でストレプトアビジンELISAプレート(EXICON)を製造業者の指示書に従ってコーティングした。血清を2種類の希釈率(200倍および1,000倍)で検討した。
特異性の高い西洋ワサビペルオキシダーゼ(HRP)結合抗ヒトIgG二次抗体、または抗ヒトIgA二次抗体(Southern Biotech)を製造業者の推奨(希釈率1,000倍)に従って使用した。抗原-抗体複合体を、自動ELISAリーダー(TECAN SUNRISE)によるOD405nmの読み値を元に、基質(ABTS)から発色産物への変換を測定することで定量した。マニュアルによるコーティング後に、ペプチドプレートを、一体型のリーダー(GENIOS, TECAN)を付属したGemini 160 ELISAロボット(TECAN)で処理して解析した。
実施例2:シグナルペプチドタギングスクリーンによるS. agalactiaeの新接着物の同定
結果
血清型IIIのGBS 6313株を、放射標識ヒトビトロネクチン、ラミニン、フィブロネクチンフィブリノーゲン、およびIgGとの相互作用に関して結合実験で検討した。6313株は、約50%の総フィブリノーゲンをその表面に蓄積した。検討した他のタンパク質のうち、有意な量(>5%)でGBS 6313と相互作用したものはなかった。細菌をトリプシンまたはプロナーゼのいずれかで処理したところ、結合状態のフィブリノーゲンの量が5%以下に減少したことから、GBS 6313のフィブリノーゲン結合構造がタンパク質性の性質をもつことがわかる。
結果
血清型IIIのGBS 6313株を、放射標識ヒトビトロネクチン、ラミニン、フィブロネクチンフィブリノーゲン、およびIgGとの相互作用に関して結合実験で検討した。6313株は、約50%の総フィブリノーゲンをその表面に蓄積した。検討した他のタンパク質のうち、有意な量(>5%)でGBS 6313と相互作用したものはなかった。細菌をトリプシンまたはプロナーゼのいずれかで処理したところ、結合状態のフィブリノーゲンの量が5%以下に減少したことから、GBS 6313のフィブリノーゲン結合構造がタンパク質性の性質をもつことがわかる。
GBS 6313の大腸菌コスミド遺伝子ライブラリーを、フィブリノーゲン結合大腸菌クローンの存在を確認するためのコロニーブロッティングでスクリーニングし、ヒトフィブリノーゲンと強力な相互作用を示すクローンを同定した。コスミドのSau3Aによる部分切断と、2〜3 kbの断片のプラスミドpTEX5236へのサブクローニングの結果、大腸菌DH5αにフィブリノーゲン結合をもたらす2.6 kbの挿入物を有するプラスミドpTEXfbsAが単離された。pTEXfbsAの挿入物の配列を決定し、得られた配列の解析から1329 bpの1つの読み枠が同定された。fbsAと命名したこの読み枠は、S. agalactiaeのフィブリノーゲン結合タンパク質をコードする(図1)。fbsA遺伝子の前方には、典型的なリボソーム結合部位
が存在し、後方には転写ターミネーター(ΔG°=-18 kcal/mol)に類似の配列が存在する。fbsAのコード領域の解析から、推定FbsAタンパク質には、連鎖球菌の表面局在タンパク質(図1)に典型的な特徴、すなわちN末端における35アミノ酸のシグナルペプチド配列(Nielsen et al., 1997)と、C末端における細胞壁アンカーモチーフ(LPKTG)(Schneewind et al., 1993)があることがわかった。fbsA遺伝子は、442アミノ酸(Mr 51319)の一次翻訳産物をコードする。これは翻訳後にプロセシングを受けて378アミノ酸(Mr 44260)の成熟タンパク質になると推定される。FbsAの最も顕著な特徴は、その高度の反復性である。FbsAは、ほぼ同一な16アミノ酸からなる完全なリピートを19個有する。19個中14個は、配列モチーフ
からなり、2個(3および10)はR14K置換を有し、3個(2、9、および19)は、A11V置換およびR14K置換の両方を有する。
血清型Ia、Ib、II、III、IV、およびVの臨床GBS単離物を対象としたサザンブロット実験を行い、GBSのfbsAの存在を解析した。サザンブロット解析で27株中25株でfbsA遺伝子が検出されたことから(図2)、fbsAがさまざまな血清型のGBSに広く分布することがわかった。興味深いことに、サザンブロット解析では、fbsA遺伝子の大きさは個々の株間でかなり変動していた。大きさの変動の分子的基盤を明らかにするために、GBS 706 S2株(血清型Ia)、33H1A株(血清型Ib)、O176 H4A株(血清型II)、SS1169株(血清型V)、およびO90R株(血清型Ia株に由来する莢膜変異体)のfbsA遺伝子をPCRで増幅して配列を決定した。得られた配列の解析から、GBS 6313株のfbsAと高い同一性を示す各PCR産物の1つの読み枠の存在が明らかとなった(図3〜7)。推定FbsAタンパク質の解析から、全ての対象のN末端に推定シグナルペプチドが、またC末端に推定細胞壁アンカーが同定された。サザンブロット実験から推定されるように、1つのタンパク質の大きさはかなり異なる。fbsAの一次翻訳産物は、706 S2株では410アミノ酸であり(図3)、33H1A株では346アミノ酸であり(図4)、176 H4A株では186アミノ酸であり(図5)、O90R株では298アミノ酸であり(図6)、またSS1169株では618アミノ酸である(図7)。図8に示すように、1つのFbsAタンパク質間の異なる大きさは主に、個々のタンパク質中のリピート数の差による。図8は、推定FbsAタンパク質の個々のリピートが、そのアミノ酸組成物に差を示すことも示す。したがって、さまざまなGBS株のfbsA遺伝子は、1つのリピートコード単位の数が多様であり、また組成に柔軟性があるようである。
実施例3:FbsAはStreptococcus agalactiaeのフィブリノーゲン受容体である
結果
FbsAの機能性解析では、シグナルペプチドおよび膜透過性領域を欠く切断型FbsAポリペプチド(FbsA-19)を、大腸菌BL21でヘキサヒスチジン融合タンパク質として合成し、アフィニティクロマトグラフィーで精製した。ウェスタンブロット実験では、FbsA-19がヒトフィブリノーゲンと結合することが判明し(図9)、FbsAがGBSのフィブリノーゲン受容体であることが確認された。FbsAタンパク質のフィブリノーゲン結合領域の位置を確認するために、FbsAのN末端およびC末端の領域を、FbsA-NおよびFbsA-C融合タンパク質として合成し、フィブリノーゲン結合に関して検討を行った。図9に示すように、FbsA-Nについてはフィブリノーゲン結合が認められたがFbsA-Cでは認められず、FbsAのN末端のリピートがフィブリノーゲン結合に関与することがわかった。
結果
FbsAの機能性解析では、シグナルペプチドおよび膜透過性領域を欠く切断型FbsAポリペプチド(FbsA-19)を、大腸菌BL21でヘキサヒスチジン融合タンパク質として合成し、アフィニティクロマトグラフィーで精製した。ウェスタンブロット実験では、FbsA-19がヒトフィブリノーゲンと結合することが判明し(図9)、FbsAがGBSのフィブリノーゲン受容体であることが確認された。FbsAタンパク質のフィブリノーゲン結合領域の位置を確認するために、FbsAのN末端およびC末端の領域を、FbsA-NおよびFbsA-C融合タンパク質として合成し、フィブリノーゲン結合に関して検討を行った。図9に示すように、FbsA-Nについてはフィブリノーゲン結合が認められたがFbsA-Cでは認められず、FbsAのN末端のリピートがフィブリノーゲン結合に関与することがわかった。
125I標識フィブリノーゲンを用いた競合阻害実験では、さまざまなタンパク質を対象に、放射標識フィブリノーゲンとGBSの結合に干渉する能力の検討を行った。対照として、GBSの非フィブリノーゲン結合表面タンパク質Bsp(Reinscheid et al., 2002)を対象に、フィブリノーゲンとGBSの結合の阻害について検討を行った。図10に示すように、高濃度のBspの添加では、GBSのフィブリノーゲンの結合に作用は認められなかった。しかし、高濃度の精製FbsA-19は、125I標識フィブリノーゲンとGBS 6313細胞の結合を実質的に阻害した。FbsAのリピートの数がフィブリノーゲン結合に影響するか否かを解析するために、リピートが9個しかないFbsA誘導体(FbsA-9)を対象に、GBSとフィブリノーゲンの結合を阻害する能力に関して検討を行った。興味深いことに、FbsA-19について得られたフィブリノーゲン結合の競合阻害を得るためには、有意に高い濃度のFbsA-9を使用しなければならなかった。この結果は、多数のリピートは、フィブリノーゲンに対するFbsAの親和性を高めるか、および/または高量のフィブリノーゲンとFbsAの結合を支持することを意味する。
分子レベルにおけるFbsAとフィブリノーゲンの相互作用の特性をさらに明らかにするために、FbsAに由来する合成ペプチドを対象にヒトフィブリノーゲンとの相互作用を検討した。最初に発明者らは、1つのFbsAのリピート単位
を対象に、ヒトフィブリノーゲンと相互作用する能力を解析した。ドットブロット実験では合成ペプチドとヒトフィブリノーゲンの強力な相互作用が認められたが、同一量のアミノ酸を含むが順序は異なるランダムなペプチドでは、フィブリノーゲンとの結合は認められなかった(図11)。この結果から、1つのFbsAのリピート単位が、特異的な対ヒトフィブリノーゲン結合能力を有することがわかる。フィブリノーゲンとの結合に不可欠なリピート領域中のアミノ酸を同定するため発明者らは、異なる位置に1つのアラニンの置換を含むペプチドを合成した。フィブリノーゲンとの相互作用に関するこれらのペプチドの検討により(図11)、リピート配列のN2、V3、L4、R6、およびR7がフィブリノーゲンとの結合に不可欠なことがわかった。またG1、R9、およびR14とアラニンの置換は、リピート単位とヒトフィブリノーゲンの相互作用を有意に低下させた。
16個のアミノ酸配列モチーフによるフィブリノーゲン結合の包括的解析を実施して、リピート領域内における推定保存的置換を同定することとした。このため配列モチーフ
に由来する合成ペプチドを合成し、膜上に直接スポットした。すべてのペプチドが1つのアミノ酸置換に関して相互に異なるようにした。このようにして、リピート中の全アミノ酸を、タンパク質をコードする20種類のアミノ酸と連続的に置き換えた。フィブリノーゲン結合に関する個々のスポットの検証の結果、フィブリノーゲンとリピート単位の相互作用に関して複雑な結果が得られた(図12)。G1と他の任意のアミノ酸の置換は、リピートのフィブリノーゲン結合を低下させたが、結合が完全に消失するわけではなかった。N2SおよびN2Tの置換は、フィブリノーゲン結合に影響しなかったが、N2における他の任意のアミノ酸との置換は、フィブリノーゲン結合を有意に低下させた。V3およびL4は、他のアミノ酸と置換時に、必ず結合機能の有意な低下がみられた。フィブリノーゲン結合はE5A、E5M、およびE5Qの置換による影響を受けなかったが、この位置に他の任意のアミノ酸が来ると、フィブリノーゲン結合は低下した。R6における置換は、フィブリノーゲン結合の喪失を顕著に引き起こしたが、R6A、R6K、およびR6Wの置換を有するペプチドは、わずかな結合活性を保持した。しかしR7と他の任意のアミノ酸との置換では、フィブリノーゲン結合は消失した。Q8は、結合に作用を及ぼすことなく、いくつかのアミノ酸と置換可能であったが、R9はKまたはWとだけ、結合に影響することなく置換可能であった。D10A、D10E、D10N、およびD10Qの置換は、フィブリノーゲン結合に影響を及ぼさず、A11F、A11I、A11L、A11V、およびA11Yの変化についても同じ結果が得られた。E12およびN13は結合に影響を及ぼすことなく、さまざまなアミノ酸との置換が可能であった。対照的にR14K置換だけが、ペプチドのフィブリノーゲン結合を保持していた。またS15およびQ16は、結合機能を喪失することなく、他のいくつかのアミノ酸と置換可能であった。スポット膜実験の結果から、
のフィブリノーゲン結合モチーフの存在を想定することができる。このコンセンサスモチーフは、他の生物のフィブリノーゲン結合タンパク質中には同定できなかったことから、新しいタイプのフィブリノーゲン結合部位であることがわかる。
スポット膜解析の結果から、2種類の異なる合成ペプチドを対象に、GBSのフィブリノーゲン結合阻害能力の検討を行った。1つのペプチド(pep_FbsA)はオリジナルのリピート単位配列
であり、もう1つのペプチド(pep_R6A)はR6A置換を有するものである。スポット膜解析で後者のペプチドは、フィブリノーゲン結合が有意に低下していることがわかった。競合阻害実験では、両方のペプチドを対象に、放射標識フィブリノーゲンとGBSの結合の阻害に関して検討を行った(図13)。160 μMのpep_FbsAは、フィブリノーゲン結合を80%阻害したが、同濃度のpep_R6Aは、フィブリノーゲン結合を20%しか阻害しなかった。以上の結果から、FbsAのリピート単位の可溶型がフィブリノーゲン結合を有することがわかる。さらに、2つのペプチド間にみられるフィブリノーゲン結合の阻害の差から、スポット膜解析の結果が確認され、またR6がフィブリノーゲン結合に重要な役割を果たすことがわかる。
GBSのフィブリノーゲン結合に対するFbsAの寄与を解析するために、GBS 6313株、706 S2株、およびO90R株のそれぞれについてfbsA欠失変異体を作製した。サザンブロット解析の結果、個々の株でfbsAが適切に欠失していること(データは示していない)が判明したので、これらを6313ΔfbsA、706 S2ΔfbsA、およびO90RΔfbsAと命名した。次に変異体および親株を対象に、可溶性フィブリノーゲンと固定化フィブリノーゲンとの結合に関して検討を行った。GBS 6313株、706 S2株、およびO90R株は、125I標識可溶性フィブリノーゲンと約50%、8%、および12%の結合を示したが、各fbsA変異株の結合率は2%未満であった。同様に、FITC標識細菌を用いた結合実験では、GBS 6313株、706 S2株、およびO90R株に由来する全菌の約45%、15%、および24%が固定化フィブリノーゲンと結合したが、固定化フィブリノーゲンと相互作用する各fbsA変異株は2%未満であった。以上の結果からFbsAが、GBS 6313株、706 S2株、およびO90R株のそれぞれにおける主要なフィブリノーゲン結合タンパク質であり、またFbsAが、細菌と可溶性フィブリノーゲンおよび固定化フィブリノーゲンの両方に対する結合に関与すると結論できる。
実施例4:FbsAは上皮細胞との接着および同細胞への浸潤に寄与し、オプソニン貧食を阻害する
結果
GBSによるオプソニン貧食防護に関するFbsAの重要性を解析するために、GBS 6313株および6313ΔfbsA株を対象に、ヒト全血を対象とした従来の殺菌アッセイ法による生存の検討を行った。ヘパリン処理したヒト血液は、2つの株について100±30コロニー形成単位(cfu)で播種後に両株で成長が認められたが、3時間の培養後に6313株は2500±500 cfu/アッセイ法に成長したものの、6313ΔfbsA株は800±100 cfu/アッセイ法しか成長しなかった。この結果は、FbsAがオプソニン化の予防に関与することを意味する。
結果
GBSによるオプソニン貧食防護に関するFbsAの重要性を解析するために、GBS 6313株および6313ΔfbsA株を対象に、ヒト全血を対象とした従来の殺菌アッセイ法による生存の検討を行った。ヘパリン処理したヒト血液は、2つの株について100±30コロニー形成単位(cfu)で播種後に両株で成長が認められたが、3時間の培養後に6313株は2500±500 cfu/アッセイ法に成長したものの、6313ΔfbsA株は800±100 cfu/アッセイ法しか成長しなかった。この結果は、FbsAがオプソニン化の予防に関与することを意味する。
GBS 6313株、706 S2株、およびO90R株、および各fbsA欠失変異体についても、ヒト肺上皮細胞系列A549に対する接着能力および浸潤能力に関して検討を行った。図14Aに示すように、A549細胞に対するfbsA欠失変異体の接着は、親株と比較して有意に低下した。同様に、fbsA欠失変異体がA549細胞に浸潤する能力も劇的に低下した(図14B)。この作用をさらに詳細に解析するために、GBS 6313がA549細胞に接着する能力および浸潤する能力を、1 μg/mlの外部添加フィブリノーゲンの存在下で定量した。フィブリノーゲンの添加は、A549細胞に対するGBS 6313の接着および浸潤の90%の低下を招いた。総合すると以上の結果は、GBSではFbsAとフィブリノーゲンの結合が、細菌とヒト上皮細胞の接着および同細胞への浸潤に重要な役割を果たすことを示している。
実施例5:FbsAはヒトで免疫原性が高い
結果
患者に由来する5つの血清を、5種類のペプチド(野生型:
アラニン変異体ペプチド:
図11参照)に対する抗体の存在に関して解析した。リピート領域の野生型配列に加えて、アラニン置換を有する4種類のペプチドが、フィブリノーゲン結合活性を欠くものとして選択された。フィブリノーゲンが結合抗体に干渉するか否かを評価するために、ペプチドのフィブリノーゲン結合活性の除去を調べることとした。すべてのペプチドを、N末端にビオチンタグが付くように合成し、ストレプトアビジンでコーティングされたELISAプレートにおけるコーティング試薬として使用した。
結果
患者に由来する5つの血清を、5種類のペプチド(野生型:
ELISA解析をGemini 160 ELISAロボットで実施した。IgA抗体およびIgG抗体のレベルを、全5種類のペプチドを含む所定の血清に関して示した(図15)。解析用に選択した5つの血清のうち、主に1つが、解析対象ペプチドと極めて高い反応性を示した。野生型と変異体ペプチドの比較から、変異体ペプチド2、3、および4が、IgA抗体とIgG抗体の両方と類似の反応性を有するが、野生型ペプチドおよびペプチド5は、すべての血清に十分認識されないことがわかった。野生型ペプチドに関しては、これはおそらく、ペプチドに対する抗体結合と競合する可能性のあるヒト血清中におけるフィブリノーゲンの存在によって説明される。ペプチド5における変異は、抗体の結合を変化させたことにより反応性を低下させていた可能性がある。興味深いことにペプチドの反応性はIgA抗体と極めて高く、IgGとはそれほど高くはなかった。この結果は、ヒトにおける抗体応答が主に、コロニー形成の予防に特に重要なIgA抗体の産生に関与することを意味する。以上の結果は、FbsAタンパク質が感染時にインビボで発現されること、また細胞表面でヒト抗体と結合しうることを強く意味する。
実施例6:シグナルペプチドタギングスクリーンによる他のS. agalactiaeの接着の同定
結果
GBSに由来する他の付着因子およびインベーシンを同定するために、GBS 6313の染色体DNAを超音波処理して断片化し、得られた断片をクレノウポリメラーゼで処理して平滑末端化し、次にプラスミドpHRM104に連結し、大腸菌CC118を形質転換した。X-リン酸を含むLBプレートによるスクリーニング後に、4個のコロニーの周囲に広範囲の青色のハロが認められた。これらのクローンのプラスミドを単離し、挿入領域の配列を決定した。得られた配列の解析から、4つの不完全な読み枠(それぞれがシグナルペプチドコード配列で開始される)が同定された。これらの遺伝子は、「B群連鎖球菌に由来する潜在的な付着因子」(potential adhesins from group B Streptococcus)であることから、それぞれpabA、pabB、pabC、およびpabDと命名した。ジゴキシゲニン標識プローブで4つの不完全な遺伝子をPCRで増幅した。DNAプローブを、大腸菌のGBS 6313コスミド遺伝子バンクのスクリーニングに使用し、pabAとpabBプローブの両方とハイブリダイズした1つの大腸菌クローン、およびpabCとpabDプローブの両方とハイブリダイズした1つの大腸菌クローンが同定された。これらのクローンからコスミドDNAを単離し、遺伝子pabA〜Dの全配列を決定した。得られた配列情報の解析の結果から、pabA遺伝子がpabB遺伝子の前方に位置し(図16)、pabC遺伝子がpabD遺伝子に先行する(図17)ことがわかった。遺伝子pabA、pabB、pabC、およびpabDは、それぞれ901アミノ酸、674アミノ酸、643アミノ酸、および182アミノ酸のタンパク質をコードする。文献(Nielsen et al., 1997)に記載された方法により、32アミノ酸、29アミノ酸、26アミノ酸、および23アミノ酸の推定シグナルペプチドをそれぞれタンパク質PabA、PabB、PabC、およびPabDであると推定することができる(図16および17)。またタンパク質PabAおよびPabBは、個々のC末端において配列IPMTGおよび配列IPQTGをそれぞれ有し、グラム陽性菌の細胞壁アンカーモチーフとの高い同一性が認められた。サザンブロット解析により、遺伝子pabA〜Dが、検討した35の臨床GBS単離菌の90〜95%で検出されたことから、GBSにこれらの遺伝子が広く分布することがわかる。
結果
GBSに由来する他の付着因子およびインベーシンを同定するために、GBS 6313の染色体DNAを超音波処理して断片化し、得られた断片をクレノウポリメラーゼで処理して平滑末端化し、次にプラスミドpHRM104に連結し、大腸菌CC118を形質転換した。X-リン酸を含むLBプレートによるスクリーニング後に、4個のコロニーの周囲に広範囲の青色のハロが認められた。これらのクローンのプラスミドを単離し、挿入領域の配列を決定した。得られた配列の解析から、4つの不完全な読み枠(それぞれがシグナルペプチドコード配列で開始される)が同定された。これらの遺伝子は、「B群連鎖球菌に由来する潜在的な付着因子」(potential adhesins from group B Streptococcus)であることから、それぞれpabA、pabB、pabC、およびpabDと命名した。ジゴキシゲニン標識プローブで4つの不完全な遺伝子をPCRで増幅した。DNAプローブを、大腸菌のGBS 6313コスミド遺伝子バンクのスクリーニングに使用し、pabAとpabBプローブの両方とハイブリダイズした1つの大腸菌クローン、およびpabCとpabDプローブの両方とハイブリダイズした1つの大腸菌クローンが同定された。これらのクローンからコスミドDNAを単離し、遺伝子pabA〜Dの全配列を決定した。得られた配列情報の解析の結果から、pabA遺伝子がpabB遺伝子の前方に位置し(図16)、pabC遺伝子がpabD遺伝子に先行する(図17)ことがわかった。遺伝子pabA、pabB、pabC、およびpabDは、それぞれ901アミノ酸、674アミノ酸、643アミノ酸、および182アミノ酸のタンパク質をコードする。文献(Nielsen et al., 1997)に記載された方法により、32アミノ酸、29アミノ酸、26アミノ酸、および23アミノ酸の推定シグナルペプチドをそれぞれタンパク質PabA、PabB、PabC、およびPabDであると推定することができる(図16および17)。またタンパク質PabAおよびPabBは、個々のC末端において配列IPMTGおよび配列IPQTGをそれぞれ有し、グラム陽性菌の細胞壁アンカーモチーフとの高い同一性が認められた。サザンブロット解析により、遺伝子pabA〜Dが、検討した35の臨床GBS単離菌の90〜95%で検出されたことから、GBSにこれらの遺伝子が広く分布することがわかる。
実施例7:PapA〜DはGBSのヒト上皮細胞に対する接着および浸潤に寄与する
結果
4種類のタンパク質の上皮細胞に対するGBSの接着における重要性を解析するために、シグナルペプチドのコード配列および細胞壁アンカーモチーフを欠く遺伝子pabAおよびpabBを大腸菌発現ベクターpET28aにクローン化し、ヘキサヒスチジンタグをPabAおよびPabB融合タンパク質のC末端に配した。これと平行して、シグナルペプチドのコード配列を欠く遺伝子pabCおよびpabDをpET28aにクローン化し、C末端にヒスチジンタグを付した融合タンパク質PabCおよびPabDを合成した。大腸菌DH5αにおけるプラスミドの構築後に、同コンストラクトで大腸菌BL21(DE)を形質転換し、融合タンパク質の合成をIPTGを添加して誘導した。次にさまざまな融合タンパク質をNi2+-アフィニティクロマトグラフィーで精製した。タンパク質PabA、PabB、およびPabCをさらに陽イオン交換クロマトグラフィーで精製し、またPabDタンパク質を精製して、陰イオン交換クロマトグラフィーで均一とした。精製タンパク質でラテックスビーズをコーティングし、ビーズをヒト肺上皮細胞系列A549と相互作用させた。対照として、ウシ血清アルブミン(BSA)でコーティングされたビーズもA549細胞と結合させた。図18に示すように、BSAでコーティングされたビーズについては、肺上皮細胞との相互作用は認められなかったが、タンパク質PabA、PabB、PabC、またはPabDでコーティングされたビーズの場合は、A549細胞との有意な結合が認められた。この結果は、タンパク質PabA、PabB、PabC、およびPabDが、細菌と宿主細胞の結合に関与することを意味する。競合実験で、GBS 6313とA549細胞の接着、および細菌の同細胞系列への浸潤を、精製PabA、PabB、PabC、またはPabD融合タンパク質の非存在下および存在下で定量した。図19に示すように、PabA、PabC、およびPabDの添加は、GBS 6313とのA549細胞の接着能力および同細胞への浸潤能力を有意に低下させた。驚くべきことにPabBの添加は、GBS 6313とA549細胞の接着および同細胞への浸潤を高めた。この結果も、PabA、PabB、PabC、およびPabDがGBSの付着因子であるという考えを支持している。
結果
4種類のタンパク質の上皮細胞に対するGBSの接着における重要性を解析するために、シグナルペプチドのコード配列および細胞壁アンカーモチーフを欠く遺伝子pabAおよびpabBを大腸菌発現ベクターpET28aにクローン化し、ヘキサヒスチジンタグをPabAおよびPabB融合タンパク質のC末端に配した。これと平行して、シグナルペプチドのコード配列を欠く遺伝子pabCおよびpabDをpET28aにクローン化し、C末端にヒスチジンタグを付した融合タンパク質PabCおよびPabDを合成した。大腸菌DH5αにおけるプラスミドの構築後に、同コンストラクトで大腸菌BL21(DE)を形質転換し、融合タンパク質の合成をIPTGを添加して誘導した。次にさまざまな融合タンパク質をNi2+-アフィニティクロマトグラフィーで精製した。タンパク質PabA、PabB、およびPabCをさらに陽イオン交換クロマトグラフィーで精製し、またPabDタンパク質を精製して、陰イオン交換クロマトグラフィーで均一とした。精製タンパク質でラテックスビーズをコーティングし、ビーズをヒト肺上皮細胞系列A549と相互作用させた。対照として、ウシ血清アルブミン(BSA)でコーティングされたビーズもA549細胞と結合させた。図18に示すように、BSAでコーティングされたビーズについては、肺上皮細胞との相互作用は認められなかったが、タンパク質PabA、PabB、PabC、またはPabDでコーティングされたビーズの場合は、A549細胞との有意な結合が認められた。この結果は、タンパク質PabA、PabB、PabC、およびPabDが、細菌と宿主細胞の結合に関与することを意味する。競合実験で、GBS 6313とA549細胞の接着、および細菌の同細胞系列への浸潤を、精製PabA、PabB、PabC、またはPabD融合タンパク質の非存在下および存在下で定量した。図19に示すように、PabA、PabC、およびPabDの添加は、GBS 6313とのA549細胞の接着能力および同細胞への浸潤能力を有意に低下させた。驚くべきことにPabBの添加は、GBS 6313とA549細胞の接着および同細胞への浸潤を高めた。この結果も、PabA、PabB、PabC、およびPabDがGBSの付着因子であるという考えを支持している。
この作用をさらに解析するために、遺伝子pabAおよびpabAをそれぞれ、GBS 6313の染色体から切り出した。結果として得られた変異体を対象に、上皮細胞に対する接着および浸潤を検討した。親GBS 6313株と比較して両変異株では、約A549細胞に対する接着および浸潤が50%低下することがわかった(図20)。
総合すると以上の結果は、タンパク質PabA、PabB、PabC、およびPabDがそれぞれGBSの上皮細胞に対する接着、および同細胞への浸潤に役割を果たすことを示唆している。
タンパク質PabA、PabB、およびPabDが、マウスで免疫応答を惹起するか否かを検討するために、精製したPabA、PabB、およびPabD融合タンパク質をマウスの皮下免疫に用いた。対象マウスに3週間後にブースター免疫を行い、初回免疫から6週間後に血清を回収した。PabA、PabB、およびPabDの各融合タンパク質の段階希釈物をニトロセルロースにブロットし、さまざまなタンパク質に対するマウス血清でプローブ処理した。図21に示すように、融合タンパク質PabA、PabB、およびPabDは、個々の抗血清によって高感度で検出されたことから、これら3種類のタンパク質のマウスにおける高い免疫原性がわかる。
実施例8:実験手順II
菌株、上皮細胞、および成長条件
細胞系列A549(ATCC CCL-185)およびHEL299(ATCC CCL-137)をAmerican Type Culture Collectionから入手した。A549はI型肺胞細胞のいくつかの特徴を示すヒト肺癌細胞である。HEL299はヒト繊維芽細胞細胞系列である。A549細胞とHEL299細胞を、10%ウシ胎児血清を添加したRPMIまたはDMEM組織培養培地(いずれもGibco BRL)で増殖させた。組織培養物を湿潤大気中(5% CO2)で37℃でインキュベートした。
菌株、上皮細胞、および成長条件
細胞系列A549(ATCC CCL-185)およびHEL299(ATCC CCL-137)をAmerican Type Culture Collectionから入手した。A549はI型肺胞細胞のいくつかの特徴を示すヒト肺癌細胞である。HEL299はヒト繊維芽細胞細胞系列である。A549細胞とHEL299細胞を、10%ウシ胎児血清を添加したRPMIまたはDMEM組織培養培地(いずれもGibco BRL)で増殖させた。組織培養物を湿潤大気中(5% CO2)で37℃でインキュベートした。
S. agalactiaeのfbsA欠失変異体の作製
S. agalactiaeのO176 H4A株、およびSS1169株のfbsA遺伝子を、文献(Schubert et al., 2002)に記載された手順で欠失させた。簡単に説明すると、温度感受性プラスミドpG+ΔfbsAでS. agalactiae株をエレクトロポレーション法で形質転換し、エリスロマイシン寒天上で30℃で成長させて形質転換体を選択した。pG+ΔfbsAが染色体に組込まれた細胞を、形質転換体を39℃で成長させてエリスロマイシン選択により、文献(Maguin et al., 1996)に記載された手順で選択した。染色体組込み株を液体培地で30℃で5日間、エリスロマイシン選択を行うことなく継代培養してプラスミドpG+ΔfbsAの切り出しを促し、所望のfbsA欠失を染色体上に残すようにした。継代培養物の希釈物を寒天上にプレーティングし、エリスロマイシン感受性に関して1個のコロニーを検討し、pG+ΔfbsA切除体を同定した。エリスロマイシン感受性のS. agalactiae切除体の染色体DNAをHindIIIで切断後に、ジゴキシゲニン標識fbsAフランキング断片を用いて、文献(Schubert et al., 2002)に記載された手順でサザンブロッティングで検討した。
S. agalactiaeのO176 H4A株、およびSS1169株のfbsA遺伝子を、文献(Schubert et al., 2002)に記載された手順で欠失させた。簡単に説明すると、温度感受性プラスミドpG+ΔfbsAでS. agalactiae株をエレクトロポレーション法で形質転換し、エリスロマイシン寒天上で30℃で成長させて形質転換体を選択した。pG+ΔfbsAが染色体に組込まれた細胞を、形質転換体を39℃で成長させてエリスロマイシン選択により、文献(Maguin et al., 1996)に記載された手順で選択した。染色体組込み株を液体培地で30℃で5日間、エリスロマイシン選択を行うことなく継代培養してプラスミドpG+ΔfbsAの切り出しを促し、所望のfbsA欠失を染色体上に残すようにした。継代培養物の希釈物を寒天上にプレーティングし、エリスロマイシン感受性に関して1個のコロニーを検討し、pG+ΔfbsA切除体を同定した。エリスロマイシン感受性のS. agalactiae切除体の染色体DNAをHindIIIで切断後に、ジゴキシゲニン標識fbsAフランキング断片を用いて、文献(Schubert et al., 2002)に記載された手順でサザンブロッティングで検討した。
ヘキサヒスチジンタグ融合タンパク質の調製
タンパク質FbsA-19は、S. agalactiae 6313の完全長FbsAタンパク質であり、16アミノ酸の19個のリピート単位をN末端に含むが、タンパク質FbsA-Nは、N末端に19個のFbsA-19のリピートを含むがC末端では切断型を含む(Schubert et al., 2002)。Bspタンパク質は、細菌の形態形成に役割を果たすS. agalactiaeの表面タンパク質であり(Reinscheid et al., 2002)、本研究における対照として使用した。培養物が1.0の光学密度に到達した時点で、1 mM IPTGを添加することで融合タンパク質を組換え大腸菌BL21中で合成させた。細胞をフレンチプレスで破壊し、Ni2+アフィニティクロマトグラフィーでQiagen社の指示書にしたがって融合タンパク質を精製した。
タンパク質FbsA-19は、S. agalactiae 6313の完全長FbsAタンパク質であり、16アミノ酸の19個のリピート単位をN末端に含むが、タンパク質FbsA-Nは、N末端に19個のFbsA-19のリピートを含むがC末端では切断型を含む(Schubert et al., 2002)。Bspタンパク質は、細菌の形態形成に役割を果たすS. agalactiaeの表面タンパク質であり(Reinscheid et al., 2002)、本研究における対照として使用した。培養物が1.0の光学密度に到達した時点で、1 mM IPTGを添加することで融合タンパク質を組換え大腸菌BL21中で合成させた。細胞をフレンチプレスで破壊し、Ni2+アフィニティクロマトグラフィーでQiagen社の指示書にしたがって融合タンパク質を精製した。
接着および浸潤アッセイ法。
A549細胞とHEL299細胞に対するS. agalactiaeの接着、および両細胞への内在化を、基本的にA549細胞に関する実施例1記載の手順で調べた。一部の実験では、A549細胞を、さまざまな量のFbsAタンパク質、またはFbsA由来ペプチドとRPMI培地中で30分間プレインキュベートし、次にPBSで3回洗浄した。
A549細胞とHEL299細胞に対するS. agalactiaeの接着、および両細胞への内在化を、基本的にA549細胞に関する実施例1記載の手順で調べた。一部の実験では、A549細胞を、さまざまな量のFbsAタンパク質、またはFbsA由来ペプチドとRPMI培地中で30分間プレインキュベートし、次にPBSで3回洗浄した。
FbsAでコーティングしたラテックスビーズの走査型電子顕微鏡による観察
約1×109個のラテックスビーズ(直径3 μm、Sigma)を、25 mM 2-N-モルホリノエタンスルホン酸(MES)、pH 6.8で3回洗浄した。この半分を500 μg/mlのFbsA融合タンパク質を含む1.0 mlのMES緩衝液に再懸濁し、残りの半分を1.0 mlのMES緩衝液に再懸濁した。このビーズを、周回式回転で4℃で一晩インキュベートした。ビーズを遠心して沈殿させた後に、上清中の残りのタンパク質の量をBradfordタンパク質アッセイ法キット(BioRad)で決定した。ビーズをMES緩衝液で1回洗浄し、10 mg/ml BSA(溶媒はMES緩衝液)で室温で1時間かけてブロック処理を行った。ビーズをMES緩衝液で2回洗浄し、RPMI+10% FCSで1回洗浄し、RPMI+10% FCSに再懸濁した。24ウェルプレートにコンフルエントのA549細胞を、1ウェルあたり2×108個のビーズとなるように総量1.0 mlを播種した。ビーズの単層混合物を大気(5% CO2)中で37℃で2時間インキュベートした。細胞をPBSで5回洗浄し、走査型電子顕微鏡による観察用に3%パラホルムアルデヒドおよび4%グルタルアルデヒド(溶媒は0.1%カコジル酸緩衝液)で固定した。走査型電子顕微鏡(Zeiss DSM 962)による観察を行った。
約1×109個のラテックスビーズ(直径3 μm、Sigma)を、25 mM 2-N-モルホリノエタンスルホン酸(MES)、pH 6.8で3回洗浄した。この半分を500 μg/mlのFbsA融合タンパク質を含む1.0 mlのMES緩衝液に再懸濁し、残りの半分を1.0 mlのMES緩衝液に再懸濁した。このビーズを、周回式回転で4℃で一晩インキュベートした。ビーズを遠心して沈殿させた後に、上清中の残りのタンパク質の量をBradfordタンパク質アッセイ法キット(BioRad)で決定した。ビーズをMES緩衝液で1回洗浄し、10 mg/ml BSA(溶媒はMES緩衝液)で室温で1時間かけてブロック処理を行った。ビーズをMES緩衝液で2回洗浄し、RPMI+10% FCSで1回洗浄し、RPMI+10% FCSに再懸濁した。24ウェルプレートにコンフルエントのA549細胞を、1ウェルあたり2×108個のビーズとなるように総量1.0 mlを播種した。ビーズの単層混合物を大気(5% CO2)中で37℃で2時間インキュベートした。細胞をPBSで5回洗浄し、走査型電子顕微鏡による観察用に3%パラホルムアルデヒドおよび4%グルタルアルデヒド(溶媒は0.1%カコジル酸緩衝液)で固定した。走査型電子顕微鏡(Zeiss DSM 962)による観察を行った。
実施例9:fbsA遺伝子およびタンパク質は、フィブリノーゲン結合に関してさまざまなS. agalactiae株に必要である
結果
血清型IIIのS. agalactiae 6313株では、FbsAタンパク質は同株のフィブリノーゲン結合に不可欠なことがわかった(実施例3)。fbsA遺伝子は、S. agalactiae 6313株、706 S2株(血清型Ia)、および莢膜変異体O90R株で欠失させてある(実施例3)。さまざまな血清型のS. agalactiae株のフィブリノーゲン結合に関するFbsAの重要性をさらに検討するために、S. agalactiae O176 H4A株(血清型II)、およびSS 1169株(血清型V)のゲノムからfbsA遺伝子を欠失させた。サザンブロット解析で、前述の株のゲノムにおいてfbsAが欠失されていることを確認し(データは示していない)、個々の変異株を、元株の名称に接尾辞ΔfbsAをつけて命名した。次に、さまざまなS. agalactiaeにおけるフィブリノーゲン結合タンパク質の合成におけるfbsA遺伝子の重要性をウェスタンブロット解析で明らかにした。S. agalactiae 6313株、O90R株、706 S2株、O176 H4A株、およびSS1169株、およびこれらの各fbsA欠失変異株の等量の培養上清をSDS-PAGEで分離し、ニトロセルロースにブロットし、次にフィブリノーゲン結合タンパク質の存在を検討した。図22に示すように、S. agalactiae 6313株および706 S2株では、培養上清中に有意な量のフィブリノーゲンタンパク質の存在が認められるが、S. agalactiae O90R株、O176 H4A株、およびSS1169株では、培養上清中に少量のフィブリノーゲン結合タンパク質しか認められない。またフィブリノーゲン結合タンパク質の大きさも、異なる株間では有意に異なる。しかしFbsAは、さまざまなS. agalactiae株におけるリピートの単位の数が多様な、高度に反復性のタンパク質である。使用したS. agalactiae株を、fbsA遺伝子中のリピート単位の数の有意差が明らかになったことから、さらなる研究用に選択した。さまざまな株のfbsA遺伝子の配列からFbsAタンパク質を、S. agalactiae 6313株、O90R株、706 S2株、O176 H4A株、およびSS1169株のそれぞれに関して分子量が51 kDa、34 kDa、47 kDa、20 kDa、および71 kDaと推定した。これらの株の培養上清中のフィブリノーゲン結合タンパク質の観察された大きさは、さまざまな株におけるFbsAタンパク質の推定サイズに良好に対応する(図22)。さまざまなfbsA欠失変異株の培養上清で、フィブリノーゲン結合タンパク質は検出できなかった。この結果は、多様な株に由来する培養上清中に観察されたフィブリノーゲン結合タンパク質がFbsAタンパク質であること、またFbsAが、検討した全株の培養上清における主要なフィブリノーゲン結合タンパク質であることを意味する。
結果
血清型IIIのS. agalactiae 6313株では、FbsAタンパク質は同株のフィブリノーゲン結合に不可欠なことがわかった(実施例3)。fbsA遺伝子は、S. agalactiae 6313株、706 S2株(血清型Ia)、および莢膜変異体O90R株で欠失させてある(実施例3)。さまざまな血清型のS. agalactiae株のフィブリノーゲン結合に関するFbsAの重要性をさらに検討するために、S. agalactiae O176 H4A株(血清型II)、およびSS 1169株(血清型V)のゲノムからfbsA遺伝子を欠失させた。サザンブロット解析で、前述の株のゲノムにおいてfbsAが欠失されていることを確認し(データは示していない)、個々の変異株を、元株の名称に接尾辞ΔfbsAをつけて命名した。次に、さまざまなS. agalactiaeにおけるフィブリノーゲン結合タンパク質の合成におけるfbsA遺伝子の重要性をウェスタンブロット解析で明らかにした。S. agalactiae 6313株、O90R株、706 S2株、O176 H4A株、およびSS1169株、およびこれらの各fbsA欠失変異株の等量の培養上清をSDS-PAGEで分離し、ニトロセルロースにブロットし、次にフィブリノーゲン結合タンパク質の存在を検討した。図22に示すように、S. agalactiae 6313株および706 S2株では、培養上清中に有意な量のフィブリノーゲンタンパク質の存在が認められるが、S. agalactiae O90R株、O176 H4A株、およびSS1169株では、培養上清中に少量のフィブリノーゲン結合タンパク質しか認められない。またフィブリノーゲン結合タンパク質の大きさも、異なる株間では有意に異なる。しかしFbsAは、さまざまなS. agalactiae株におけるリピートの単位の数が多様な、高度に反復性のタンパク質である。使用したS. agalactiae株を、fbsA遺伝子中のリピート単位の数の有意差が明らかになったことから、さらなる研究用に選択した。さまざまな株のfbsA遺伝子の配列からFbsAタンパク質を、S. agalactiae 6313株、O90R株、706 S2株、O176 H4A株、およびSS1169株のそれぞれに関して分子量が51 kDa、34 kDa、47 kDa、20 kDa、および71 kDaと推定した。これらの株の培養上清中のフィブリノーゲン結合タンパク質の観察された大きさは、さまざまな株におけるFbsAタンパク質の推定サイズに良好に対応する(図22)。さまざまなfbsA欠失変異株の培養上清で、フィブリノーゲン結合タンパク質は検出できなかった。この結果は、多様な株に由来する培養上清中に観察されたフィブリノーゲン結合タンパク質がFbsAタンパク質であること、またFbsAが、検討した全株の培養上清における主要なフィブリノーゲン結合タンパク質であることを意味する。
さまざまなS. agalactiae株、およびこれらのfbsA変異株を対象に、その表面における125I標識フィブリノーゲン結合の検討を行った。S. agalactiae 6313では、放射標識フィブリノーゲンのかなりの結合が認められた。しかし、O90R株および706 S2株では、ヒトフィブリノーゲンの中程度の結合が認められ、またO176 H4A株およびSS1169株ではヒトフィブリノーゲンの弱い結合が認められた。さまざまな株にみられたフィブリノーゲン結合の差は、これらの株のFbsAタンパク質のフィブリノーゲン結合リピートの数と相関しなかった。しかしながらfbsA欠失変異体では、フィブリノーゲン結合は1〜3%に低下した。FITC標識細菌を用いた結合実験では同様に、6313株、O90R株、706 S2株、O176 H4A株、およびSS1169株の全細菌の約45%、18%、14%、4%、および7%が固定化フィブリノーゲンと結合したが、固定化フィブリノーゲンと結合した各fbsA変異株は2%未満であった(図23)。以上の結果からさらにFbsAが、解析対象のS. agalactiae株における主要なフィブリノーゲン結合タンパク質であること、またFbsAが、細菌と可溶性フィブリノーゲンと固定化フィブリノーゲンの両方に対する結合に関与することがわかる。
実施例10:fbsA遺伝子およびタンパク質はS. agalactiaeのヒト細胞との効率的な結合、および同細胞への内在化に必要である
結果
S. agalactiae 6313株、O90R株、706 S2株、O176 H4A株、およびSS1169株、ならびにこれらの同質遺伝的なfbsA変異株を対象に、ヒト肺上皮細胞系列A549に対する接着能力と浸潤能力に関して検討を行った。図24に示すように、S. agalactiae 6313株は、その多くがA549細胞と結合して同細胞に浸潤したが、O90R株、706 S2株、およびSS1169株は、A549細胞に対する中程度の接着と同細胞への内在化を示すことがわかった。対照的に、S. agalactiae O176 H4A株は、極めて少数がA549細胞に接着して同細胞に浸潤した。さまざまな株がA549細胞に対して接着して浸潤する際の当初の差と関係なく、個々の株のfbsA遺伝子の欠失は、A549細胞との接着および同細胞への浸潤を、異なる株間で極めて低いが類似の値に低下させた。A549細胞への内在化がほとんどみられないことが既にわかっているO176 H4A株だけは、fbsA遺伝子の欠失は、細菌のA549細胞への内在化に低下はみられなかった。以上の結果はfbsA遺伝子が、S. agalactiaeとヒト上皮細胞との接着および同細胞への内在化に重要な役割を果たすことを意味する。fbsA遺伝子が、S. agalactiaeとさまざまな細胞系列の結合に果たす役割を評価するために発明者らは、ヒト繊維芽細胞細胞系列HEL299を対象に、S. agalactiae 6313およびこのfbsA欠失変異体の接着および内在化の解析を行った。図25に示すように、6313ΔfbsA株とHEL299細胞の結合、また細菌の同細胞系列への内在化は約90%低下した。この結果から、S. agalactiaeとさまざまなヒト細胞との接着および内在化にfbsA遺伝子が全般的に重要であることが示唆される。
結果
S. agalactiae 6313株、O90R株、706 S2株、O176 H4A株、およびSS1169株、ならびにこれらの同質遺伝的なfbsA変異株を対象に、ヒト肺上皮細胞系列A549に対する接着能力と浸潤能力に関して検討を行った。図24に示すように、S. agalactiae 6313株は、その多くがA549細胞と結合して同細胞に浸潤したが、O90R株、706 S2株、およびSS1169株は、A549細胞に対する中程度の接着と同細胞への内在化を示すことがわかった。対照的に、S. agalactiae O176 H4A株は、極めて少数がA549細胞に接着して同細胞に浸潤した。さまざまな株がA549細胞に対して接着して浸潤する際の当初の差と関係なく、個々の株のfbsA遺伝子の欠失は、A549細胞との接着および同細胞への浸潤を、異なる株間で極めて低いが類似の値に低下させた。A549細胞への内在化がほとんどみられないことが既にわかっているO176 H4A株だけは、fbsA遺伝子の欠失は、細菌のA549細胞への内在化に低下はみられなかった。以上の結果はfbsA遺伝子が、S. agalactiaeとヒト上皮細胞との接着および同細胞への内在化に重要な役割を果たすことを意味する。fbsA遺伝子が、S. agalactiaeとさまざまな細胞系列の結合に果たす役割を評価するために発明者らは、ヒト繊維芽細胞細胞系列HEL299を対象に、S. agalactiae 6313およびこのfbsA欠失変異体の接着および内在化の解析を行った。図25に示すように、6313ΔfbsA株とHEL299細胞の結合、また細菌の同細胞系列への内在化は約90%低下した。この結果から、S. agalactiaeとさまざまなヒト細胞との接着および内在化にfbsA遺伝子が全般的に重要であることが示唆される。
細菌の接着および内在化におけるFbsAタンパク質の役割を評価するために、FbsA-19融合タンパク質による真核細胞の前処理が、S. agalactiae 6313の接着および浸潤に及ぼす作用を評価した。タンパク質FbsA-19は6313株に由来するFbsAタンパク質であり、19個のリピート単位を有する。図26に示すように、A549細胞を高量のFbsA-19タンパク質で前処理すると、S. agalactiae 6313と同細胞系列の接着および浸潤が実質的に阻害された。重要なことに発明者らは、HEL299細胞と細菌の接着および浸潤の低下が相関することも見出した。
FbsAタンパク質がフィブリノーゲンと結合することについては既に報告した(実施例3)。したがって発明者らは、S. agalactiae 6313のフィブリノーゲンによるプレインキュベーションが、細菌とA549細胞の接着および同細胞への浸潤に及ぼす作用を検討した。発明者らはS. agalactiae 6313を、0.1 μg/ml〜1.0 μg/mlのフィブリノーゲンでプレインキュベートしたところ、細菌とA549細胞の接着および同細胞への浸潤の用量依存性が阻害されることを認めた(データは示していない)。しかし、細菌を顕微鏡で観察したところ、高量のフィブリノーゲンの使用時には細菌が凝集することがわかった。したがって、観察されたフィブリノーゲンによる細菌の接着および浸潤の阻害は、菌表面におけるFbsAタンパク質のブロッキングか、またはFbsAタンパク質中の複数のフィブリノーゲン結合部位の存在に起因する菌の凝集のいずれかに帰することができる。発明者らはまた、S. agalactiae 6313の接着および浸潤に対するフィブロネクチンの影響を検討した。しかし、10 μg/mlのフィブロネクチンでも、細菌の接着および内在化に対する阻害作用は誘導されなかった(データは示していない)。
実施例11:FbsAでコーティングしたラテックスビーズはA549細胞と接着する
結果
FbsAが、S. agalactiaeと宿主細胞の相互作用に役割を果たすことは既に明らかとなった。FbsAと真核細胞の相互作用に、さらに別の因子が必要か否かを調べるために、ラテックスビーズをFbsA-19タンパク質でコーティングしてヒトA549細胞との相互作用を検討した。対照として、BSAでコーティングしたラテックスビーズについてもA549細胞との相互作用に関して解析を行った。走査型電子顕微鏡による観察で、BSAでコーティングしたラテックスビーズとA549細胞との結合はほとんど認められなかったが、FbsA-19でコーティングしたビーズはA549細胞と多数が結合した(図27)。FbsA-19でコーティングしたビーズと形質膜との結合は、微繊毛と、初期仮足形成に似た構造体との接触が特徴的であった(図27C)。場合によっては仮足がビーズ表面を囲むように見えたことから、ビーズが最終的に内在化されたことがわかる(図27D)。しかしながら、FbsA-19でコーティングされたビーズの内在化は、どちらかと言えばまれに認められるだけであり、FbsA-19が通常はS. agalactiaeまたはFbsA-19でコーティングしたビーズの真核細胞への取り込みのきっかけとならないことがわかる。
結果
FbsAが、S. agalactiaeと宿主細胞の相互作用に役割を果たすことは既に明らかとなった。FbsAと真核細胞の相互作用に、さらに別の因子が必要か否かを調べるために、ラテックスビーズをFbsA-19タンパク質でコーティングしてヒトA549細胞との相互作用を検討した。対照として、BSAでコーティングしたラテックスビーズについてもA549細胞との相互作用に関して解析を行った。走査型電子顕微鏡による観察で、BSAでコーティングしたラテックスビーズとA549細胞との結合はほとんど認められなかったが、FbsA-19でコーティングしたビーズはA549細胞と多数が結合した(図27)。FbsA-19でコーティングしたビーズと形質膜との結合は、微繊毛と、初期仮足形成に似た構造体との接触が特徴的であった(図27C)。場合によっては仮足がビーズ表面を囲むように見えたことから、ビーズが最終的に内在化されたことがわかる(図27D)。しかしながら、FbsA-19でコーティングされたビーズの内在化は、どちらかと言えばまれに認められるだけであり、FbsA-19が通常はS. agalactiaeまたはFbsA-19でコーティングしたビーズの真核細胞への取り込みのきっかけとならないことがわかる。
実施例12:遺伝子pabCおよびpabDは同時に転写され、またS. agalactiaeの臨床株で保存されている
結果
pabC遺伝子を含む領域のゲノム構成を図31に示す。4つの遺伝子の重複領域を増幅するのに適したRT-PCRおよびオリゴヌクレオチドを用いて、pabC遺伝子とgbs0851(pabD)遺伝子は1つの転写物として転写されるが、RNAポリメラーゼはmetK遺伝子およびgbs0853遺伝子(図28)に関しては独立した転写物を生じることがわかった。この結果から、pabC遺伝子およびgbs0851遺伝子の産物が、S. agalactiaeと同じか、または類似のプロセスに必要とされる機能を示す可能性があることがわかる。
結果
pabC遺伝子を含む領域のゲノム構成を図31に示す。4つの遺伝子の重複領域を増幅するのに適したRT-PCRおよびオリゴヌクレオチドを用いて、pabC遺伝子とgbs0851(pabD)遺伝子は1つの転写物として転写されるが、RNAポリメラーゼはmetK遺伝子およびgbs0853遺伝子(図28)に関しては独立した転写物を生じることがわかった。この結果から、pabC遺伝子およびgbs0851遺伝子の産物が、S. agalactiaeと同じか、または類似のプロセスに必要とされる機能を示す可能性があることがわかる。
PabCおよびgbs0851をコードする遺伝子が、GBSのさまざまな血清型および臨床単離菌で保存されているか否かを判定するために、33種類のさまざまなS. agalactiae株の染色体DNAを単離し、遺伝子全体を増幅する特定のプライマーでPCR解析を行った。2つの遺伝子pabCおよびgbs0851は、検討した全株に存在することがわかった。図29は一例として、GBSの最も優勢な血清型(Ia、Ib、III、およびV)を対象としたPCRの結果を示す。gbs0851遺伝子は全株から同じ長さで増幅されたことから、遺伝子の配列および大きさが保存されていることがわかる。pabC遺伝子のPCRの結果、驚くべきことに、解析対象株に依存して2つの異なる大きさの産物が増幅されることがわかった。また大きさの差は同じ血清型の株でも認められた。S. agalactiae 6313(血清型III)、S. agalactiae NEM316(血清型III)、およびS. agalactiae 2003V_R(血清型V)のPabCタンパク質のアミノ酸配列の比較を図30に示す。この図から、S. agalactiae 6313およびS. agalactiae NEM316のPabCタンパク質は同一であるが、S. agalactiae 2003 V_RのPabCには明瞭な差があることがわかる。PabCの配列の多様性は、全体的に同タンパク質のN末端部分に帰することができるが、C末端部分は全3種類の血清型でほぼ同一である。大きさに関して認められた差は、図29のPCRの結果とも一致する。別のPCR実験では、大きさの差が、遺伝子の3'末端側ではなく5'側における配列の多様性に基づくことが確認された(データは示していない)。
実施例13:S. agalactiaeのPabCはヒトフィブリノーゲンと結合し、真核細胞への浸潤に関与する
最初の実験では、PabCがフィブリノーゲンのαサブユニットと結合することが判明した(図31A、C)。PabCのどの領域がフィブリノーゲン結合に関与するかということを詳細に明らかにするために、PabCのタンパク質全体、ならびにN末端部分およびC末端部分を、ヒスチジンタグ融合タンパク質として発現させた。フィブリノーゲンの添加後に、フィブリノーゲン抗体との結合を検出した(図31B)。この実験で、PabCの保存されたC末端部分そのものにはフィブリノーゲン結合活性はないが、N末端部分は、完全長タンパク質と同等の規模で同活性を十分発揮することがわかった。
最初の実験では、PabCがフィブリノーゲンのαサブユニットと結合することが判明した(図31A、C)。PabCのどの領域がフィブリノーゲン結合に関与するかということを詳細に明らかにするために、PabCのタンパク質全体、ならびにN末端部分およびC末端部分を、ヒスチジンタグ融合タンパク質として発現させた。フィブリノーゲンの添加後に、フィブリノーゲン抗体との結合を検出した(図31B)。この実験で、PabCの保存されたC末端部分そのものにはフィブリノーゲン結合活性はないが、N末端部分は、完全長タンパク質と同等の規模で同活性を十分発揮することがわかった。
ウェスタンブロッティングの結果を確認するために、同様に精製したPabCタンパク質誘導体を対象に捕捉ELISAアッセイ法を行った(図32)。この目的で、1ウェルあたり2 μgのフィブリノーゲンで4℃で一晩かけてコーティングを行った。高濃度のPabC誘導体の結合活性を、ヒスチジンタグ抗体をベースとしたペルオキシダーゼアッセイ法で定量した。捕捉ELISA実験で、PabCタンパク質のN末端部分がフィブリノーゲン結合領域を有するという結果が確認された。
図19から、比較的高濃度のPabCが、S. agalactiaeとA549真核細胞との接着、および同細胞への浸潤を阻害可能であることがわかる。低濃度の組換えPabCタンパク質を用いることでPabCが、GBSに対する接着よりも浸潤を促進する可能性が極めて高いことがわかる(図33C、D)。確認的な結果は、pabC欠失変異体に関して得られた(図33A、B)。得られた結果から、A549細胞への浸潤が減少することもわかった。以上の結果からPabCがS. agalactiaeに浸潤体として作用して真核細胞のコロニーが形成される可能性があることが示唆される。
以下に、本明細書で参照した全ての出版物および文献を記載する。これらの参考文献の全記述が参照として本明細書に組み入れられると理解される。
参考文献リスト
本明細書、請求項、および/または図面に開示された本発明の特徴は、いずれも別個に、また任意に組み合わせることで、本発明をさまざまな形状で実現するための材料となる。
本明細書で参照される図面について詳細に説明する。
(図1)血清型IIIのGBS 6313株のfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図2)GBSのさまざまな臨床単離菌におけるfbsA遺伝子の存在を判定するためのサザンブロット解析を示す。血清型Ia、Ib、II、III、IV、およびVの多様なGBS株の染色体DNAをそれぞれHindIIIで切断し、大きさで分離し、ナイロン膜にブロッティングし、ジゴキシゲニン標識fbsA特異的DNAプローブとハイブリダイズさせた。
(図3)血清型IaのGBS 706 S2株のfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図4)血清型IbのGBS 33H1A株のfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図5)血清型IIのGBS 176 H4A株のFbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図6)GBSの莢膜変異体O90RのfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図7)血清型VのGBS SS1169株のfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図8)GBS 6313株(血清型III)、706 S2株(血清型Ia)、33H1A株(血清型Ib)、O176 H4A株(血清型II)、O90R株(血清型Iaに由来)、およびSS1169株(血清型V)の各FbsAタンパク質の図による比較。シグナルペプチド(黒いボックス)、壁透過性領域(WSR;垂直線ボックス)、細胞壁アンカーモチーフ(LPKTG)、および膜透過性領域(MSR;斜線ボックス)の位置を示す。各タンパク質のリピート数をそれぞれ示す。灰色のボックスは配列モチーフ
のリピートを示し、水平線ボックスはR14K置換を有するリピートを示し、点線ボックスはA11VとR14Kの両置換を有するリピートの位置を示す。E12D置換を有するリピートを、GBS 33H1A株およびSS1169株のFbsAタンパク質の下部に示す。33H1A株のFbsAの上部には、1つのA11V置換を有するリピートを示す。
(図9)FbsA中にフィブリノーゲン結合ドメインを同定するための、切断型FbsA誘導体のウェスタンブロット解析の結果を示す。成熟FbsAタンパク質(FbsA-19)、FbsAのN末端リピート含有領域(FbsA-N)、またはC末端部分(FbsA-C)を示すヘキサヒスチジンタグ融合タンパク質をSDS-PAGEで分離し、ニトロセルロースにブロットし、ヒトフィブリノーゲンに対する結合を検討した。さまざまなコンストラクトにコードされた3種類のタンパク質のフィブリノーゲン結合活性を、FbsAの図の下部に示す。
(図10)精製した融合タンパク質FbsA-19、FbsA-9、およびBspのそれぞれによる、GBS 6313に対するフィブリノーゲン結合の競合阻害を示す。FbsA-9は、リピートドメインに9個のリピートを含む点がFbsA-19と異なる。結合アッセイ法は、125I標識フィブリノーゲンを用いて、さまざまな濃度の各融合タンパク質の存在下で実施した。各実験は少なくとも3通りで実施した。
(図11)FbsAのリピート単位に由来する合成ペプチドによる、フィブリノーゲン結合のスポット膜解析の結果を示す。フィブリノーゲン結合は、FbsAリピートモチーフ
を含むペプチド、およびスクランブル配列
を含むペプチドを用いて検討した。リピートモチーフが、1つのアミノ酸がアラニンと置換されている点で異なる合成ペプチドを、フィブリノーゲン結合のプローブとして用いた。スポット膜の脇に各合成ペプチドの配列を示す。太字および下線を付した文字は、リピートモチーフ内におけるアミノ酸置換を示す。
(図12)フィブリノーゲン結合リピート単位のスポット膜解析の結果を示す。合成ペプチドをフィブリノーゲン結合に関して検討した(フィブリノーゲン結合リピートの個々のアミノ酸を、それぞれ20アミノ酸と置換した)。垂直方向の太字は、FbsAに由来するフィブリノーゲン結合配列
を示す。水平方向の文字は、個々の位置において、当初のアミノ酸の代わりに合成ペプチド中に導入されたアミノ酸を示す。
(図13)合成ペプチドによるGBS 6313に対するフィブリノーゲン結合の競合阻害を示す。結合アッセイ法は、FbsAに由来するリピート単位を有するさまざまな濃度のペプチドpep_FbsA(SEQ ID 211)、およびリピート単位中にR6A置換を有するpep_R6Aの存在下で125I標識フィブリノーゲンを用いて実施した。各実験は少なくとも3回実施した。
(図14)GBS 6313株、706 S2株、およびO90R株、およびこれらの各fbsA欠失変異体の真核細胞との接着(A)および同細胞への浸潤(B)を示す。値は、3通りで実施した少なくとも4回の独立した実験の結果を示す。またエラーバーを示す。
(図15)ヒト血清とFbsAペプチドとのペプチドELISAを示す。5種類のビオチン化ペプチド(野生型:
アラニン変異体ペプチド:
図11も参照)で、ストレプトアビジンでコーティングされたELISAプレートをコーティングし、GBS感染者から採取した5種類の血清を用いて解析を行った。患者の血清を1:200および1:1,000に希釈してアプライした。IgG抗体(A)およびIgA抗体(B)の検出は、西洋ワサビペルオキシダーゼを結合させた二次抗ヒト抗体でABTSを基質として行った。
(図16)GBS 6313のpabA/Bコード領域のDNA配列、ならびに推定PabA(nt 319〜2964)タンパク質、およびPabB(nt 3087〜5111)タンパク質を示す。推定リボソーム結合部位(RBS)に下線を付した。太字およびイタリックは、推定PabAおよびPabBタンパク質の推定シグナルペプチドを示し、太字および下線部は、グラム陽性菌の細胞壁アンカーモチーフに同一性の高い領域を示す。
(図17)GBS 6313のpabC/Dコード領域のDNA配列、ならびに推定PabC(nt 487〜2394)タンパク質、およびPabD(nt 2461〜3006)タンパク質を示す。推定リボソーム結合部位(RBS)に下線を付した。太字およびイタリックは、推定PabCおよびPabDタンパク質の推定シグナルペプチドを示す。
(図18)PabA、PabB、PabC、PabDでそれぞれコーティングしたラテックスビーズと2時間インキュベートしたA549細胞の走査型電子顕微鏡写真を示す。BSAでコーティングしたラテックスビーズを対照とした使用した。
(図19)100 μg/mlのPabA、PabB、PabC、またはPabD融合タンパク質の存在下における、GBS 6313のA549細胞との接着、および同細胞へ浸潤を示す。GBS 6313のA549細胞との接着(A)、および同細胞への内在化(B)は、任意の値を100%とし、さまざまな融合タンパク質の存在下で得た結果を、この値と関連づけた。各実験を3通りで少なくとも3回実施した。
(図20)GBS 6313ならびにpabAおよびpabB欠失変異体と真核細胞の接着および内在化を示す。GBS 6313のA549細胞との接着(A)、および同細胞への内在化(B)は、任意の値を100%とし、GBS変異体6313ΔpabAおよび6313ΔpabBに関して得られた結果を、この値と関連づけた。各実験を3通りで少なくとも3回実施した。
(図21)個々の抗原を検出する段階における感度に関する抗PabA、抗PabB、および抗PabD抗血清の検討を示す。融合タンパク質PabA、PabB、およびPabDの段階希釈物をニトロセルロース上にスポットし、各タンパク質に対するマウス血清の1:1000希釈物でプローブ処理した。結合状態の抗体を、抗マウスHRP結合抗体で標識し、化学発光法で可視化した。
(図22)フィブリノーゲン結合タンパク質の存在に関する、さまざまなS. agalactiae株、およびこれらの同質遺伝的なfbsA欠失変異体の培養上清のウェスタンブロット解析の結果を示す。さまざまなS. agalactiae株、およびこれらのfbsA欠失変異体の濃縮培養上清に由来する15 μgのタンパク質をSDS-PAGEで大きさによって分離し、ニトロセルロースにブロットし、ヒトフィブリノーゲンとの相互作用を検討した。ブロットをウサギ抗フィブリノーゲン抗体とインキュベートした後に、ワサビペルオキシダーゼを結合させたヤギ抗ウサギ抗体でインキュベートすることで、結合状態のフィブリノーゲンを検出した。フィブリノーゲン抗体複合体の検出には化学発光法を用いた。
(図23)固定化フィブリノーゲンに対する、さまざまなS. agalactiae株およびこれらのfbsA欠失変異体の結合を示す。同等の細胞数のさまざまな株を、テラサキプレートに固定化されたフィブリノーゲンとインキュベートした。フィブリノーゲンと結合した細菌の数を、アッセイ系への入力菌数と関連づけた。
(図24)さまざまなS. agalactiae株、およびこれらの同質遺伝的なfbsA変異株と、肺上皮細胞系列A549の接着および同細胞への内在化を示す。同等の数の細菌を使用してA549細胞に感染させ、A549細胞に接着した菌数(A)、および同細胞に内在化した菌数(B)を入力菌数と関連づけた。
(図25)S. agalactiae 6313株および6313ΔfbsAと繊維芽細胞細胞系列HEL299との接着および同細胞への内在化を示す。HEL299細胞にS. agalactiaeをMOIが10:1となるように感染させ、細胞と接着した菌の数と、細胞へ内在化した菌の数を入力菌数と関連づけた。
(図26)S. agalactiaeのA549細胞に対する接着に及ぼすFbsAタンパク質の影響を示す。接着アッセイ法を、さまざまな量の精製FbsA融合タンパク質の存在下で実施し、細菌と接着した細胞の数を入力菌数と関連づけた。
(図27)ヒトA549細胞に対する、FbsAでコーティングされたラテックスビーズの結合を示す。ラテックスビーズをBSA(A)、またはFbsA融合タンパク質(B〜D)のいずれかでコーティングし、コーティングされたビーズと肺上皮細胞系列A549の相互作用を走査型電子顕微鏡で解析した。
(図28)S. agalactiae のpabCコード領域の転写構成を示す。図の上部の名称は、S. agalactiae 6313の全RNAとのPCR(A)、全RNAとのRT-PCR(B)、または染色体DNAとのPCR(C)においてプライマー対がアニールした遺伝子を示す。
(図29)pabC遺伝子およびpabD遺伝子の存在に関する、GBS株のPCR解析の結果を示す。1、S. agalactiae 1137株(Ia);2、S. agalactiae A90/14株(Ib);3、S. agalactiae 6313株(III);4、S. agalactiae 4416 S3株(III); 5、S. agalactiae 4357株(V);6、S. agalactiae 4327株(V)をPCRに使用した。
(図30)S. agalactiae 6313、S. agalactiae NEM316、およびS. agalactiae 2003V_RのPabCタンパク質のアミノ酸配列の比較を示す。
(図31)S. agalactiaeのpabCコード領域の制限酵素切断地図(A)、ならびにフィブリノーゲン結合に関するPabCおよびGbs0851融合タンパク質のウェスタンブロット解析の結果(B)を示す。融合タンパク質をSDS-PAGEで大きさにより分離し、ニトロセルロース膜にトランスファーし、フィブリノーゲン結合をウェスタンブロッティングで検討した。結合状態のフィブリノーゲンを、ウサギ抗フィブリノーゲン抗体と、これに続くペルオキシダーゼ標識ヤギ抗ウサギ抗体で検出後に化学発光で可視化した。PabCおよびGbs0851=完全長の融合タンパク質、PabC-N=PabCのN末端の388アミノ酸、PabC-C=PabCのC末端の222アミノ酸。(C)は、ウェスタンブロット解析による、ヒトフィブリノーゲン中のFbsA結合部位およびPabC結合部位の同定を示す。ヒトフィブリノーゲンをSDS-PAGEで大きさによって分離し、クーマシー染色(左のレーン)を行うか、またはニトロセルロースにトランスファーし、FbsA結合またはPabC結合をウェスタンブロッティングで検討した。結合状態の融合タンパク質を、マウス抗ヒスチジンタグ抗体と、これに続くペルオキシダーゼ結合ヤギ抗マウスIgG fab断片で検出し、化学発光で可視化した。
(図32)捕捉ELISAアッセイ法による、固定化フィブリノーゲンと組換えPabC融合タンパク質の結合を示す。マイクロタイタープレートのウェルを一定量のヒトフィブリノーゲンでコーティング後に、さまざまなPabC融合タンパク質を濃度を高めながら添加した。結合状態の融合タンパク質を、マウス抗ヒスチジンタグ抗体およびペルオキシダーゼ結合ヤギ抗マウスIgG fab断片で検出した。発色を、テトラメチル-ベンジジン基質を添加して開始させ、H2SO4を添加して停止させた。マイクロタイターウェルの吸光度を450 nmで読み取った。値は、3回の独立した実験(各3通りで試行)の平均値である。
(図33)S. agalactiae 6313 pAT32株、ΔpabC pAT32株およびΔpabC pATpabC株のそれぞれと肺上皮細胞系列A549との接着(A)、および同細胞への浸潤(B)を示す。細菌の接着および浸潤は以下のように計算した:接着=接着菌数/アッセイ法の総菌数×100。浸潤=内在化菌数/アッセイ法の総菌数×100。各実験を3通りで少なくとも3回実施した。さまざまな量のPabCおよびBsp融合タンパク質の存在下におけるS. agalactiae 6313と真核細胞の接着(C)および浸潤(D)を示す。図32の説明に記載されているように、細菌の接着および浸潤を計算した。各実験を3通りで少なくとも3回実施した。
(図1)血清型IIIのGBS 6313株のfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図2)GBSのさまざまな臨床単離菌におけるfbsA遺伝子の存在を判定するためのサザンブロット解析を示す。血清型Ia、Ib、II、III、IV、およびVの多様なGBS株の染色体DNAをそれぞれHindIIIで切断し、大きさで分離し、ナイロン膜にブロッティングし、ジゴキシゲニン標識fbsA特異的DNAプローブとハイブリダイズさせた。
(図3)血清型IaのGBS 706 S2株のfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図4)血清型IbのGBS 33H1A株のfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図5)血清型IIのGBS 176 H4A株のFbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図6)GBSの莢膜変異体O90RのfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図7)血清型VのGBS SS1169株のfbsAコード領域のDNA配列、および推定FbsAタンパク質を示す。推定リボソーム結合部位(RBS)に下線を付し、潜在的な転写ターミネーターを逆平行矢印で示す。推定FbsAタンパク質中で、太字およびイタリックは推定シグナルペプチド配列を示し、太字および下線部は細胞壁アンカーモチーフLPKTGを示す。FbsA中のリピートに数字を付し、矢印でマークした。
(図8)GBS 6313株(血清型III)、706 S2株(血清型Ia)、33H1A株(血清型Ib)、O176 H4A株(血清型II)、O90R株(血清型Iaに由来)、およびSS1169株(血清型V)の各FbsAタンパク質の図による比較。シグナルペプチド(黒いボックス)、壁透過性領域(WSR;垂直線ボックス)、細胞壁アンカーモチーフ(LPKTG)、および膜透過性領域(MSR;斜線ボックス)の位置を示す。各タンパク質のリピート数をそれぞれ示す。灰色のボックスは配列モチーフ
(図9)FbsA中にフィブリノーゲン結合ドメインを同定するための、切断型FbsA誘導体のウェスタンブロット解析の結果を示す。成熟FbsAタンパク質(FbsA-19)、FbsAのN末端リピート含有領域(FbsA-N)、またはC末端部分(FbsA-C)を示すヘキサヒスチジンタグ融合タンパク質をSDS-PAGEで分離し、ニトロセルロースにブロットし、ヒトフィブリノーゲンに対する結合を検討した。さまざまなコンストラクトにコードされた3種類のタンパク質のフィブリノーゲン結合活性を、FbsAの図の下部に示す。
(図10)精製した融合タンパク質FbsA-19、FbsA-9、およびBspのそれぞれによる、GBS 6313に対するフィブリノーゲン結合の競合阻害を示す。FbsA-9は、リピートドメインに9個のリピートを含む点がFbsA-19と異なる。結合アッセイ法は、125I標識フィブリノーゲンを用いて、さまざまな濃度の各融合タンパク質の存在下で実施した。各実験は少なくとも3通りで実施した。
(図11)FbsAのリピート単位に由来する合成ペプチドによる、フィブリノーゲン結合のスポット膜解析の結果を示す。フィブリノーゲン結合は、FbsAリピートモチーフ
(図12)フィブリノーゲン結合リピート単位のスポット膜解析の結果を示す。合成ペプチドをフィブリノーゲン結合に関して検討した(フィブリノーゲン結合リピートの個々のアミノ酸を、それぞれ20アミノ酸と置換した)。垂直方向の太字は、FbsAに由来するフィブリノーゲン結合配列
(図13)合成ペプチドによるGBS 6313に対するフィブリノーゲン結合の競合阻害を示す。結合アッセイ法は、FbsAに由来するリピート単位を有するさまざまな濃度のペプチドpep_FbsA(SEQ ID 211)、およびリピート単位中にR6A置換を有するpep_R6Aの存在下で125I標識フィブリノーゲンを用いて実施した。各実験は少なくとも3回実施した。
(図14)GBS 6313株、706 S2株、およびO90R株、およびこれらの各fbsA欠失変異体の真核細胞との接着(A)および同細胞への浸潤(B)を示す。値は、3通りで実施した少なくとも4回の独立した実験の結果を示す。またエラーバーを示す。
(図15)ヒト血清とFbsAペプチドとのペプチドELISAを示す。5種類のビオチン化ペプチド(野生型:
(図16)GBS 6313のpabA/Bコード領域のDNA配列、ならびに推定PabA(nt 319〜2964)タンパク質、およびPabB(nt 3087〜5111)タンパク質を示す。推定リボソーム結合部位(RBS)に下線を付した。太字およびイタリックは、推定PabAおよびPabBタンパク質の推定シグナルペプチドを示し、太字および下線部は、グラム陽性菌の細胞壁アンカーモチーフに同一性の高い領域を示す。
(図17)GBS 6313のpabC/Dコード領域のDNA配列、ならびに推定PabC(nt 487〜2394)タンパク質、およびPabD(nt 2461〜3006)タンパク質を示す。推定リボソーム結合部位(RBS)に下線を付した。太字およびイタリックは、推定PabCおよびPabDタンパク質の推定シグナルペプチドを示す。
(図18)PabA、PabB、PabC、PabDでそれぞれコーティングしたラテックスビーズと2時間インキュベートしたA549細胞の走査型電子顕微鏡写真を示す。BSAでコーティングしたラテックスビーズを対照とした使用した。
(図19)100 μg/mlのPabA、PabB、PabC、またはPabD融合タンパク質の存在下における、GBS 6313のA549細胞との接着、および同細胞へ浸潤を示す。GBS 6313のA549細胞との接着(A)、および同細胞への内在化(B)は、任意の値を100%とし、さまざまな融合タンパク質の存在下で得た結果を、この値と関連づけた。各実験を3通りで少なくとも3回実施した。
(図20)GBS 6313ならびにpabAおよびpabB欠失変異体と真核細胞の接着および内在化を示す。GBS 6313のA549細胞との接着(A)、および同細胞への内在化(B)は、任意の値を100%とし、GBS変異体6313ΔpabAおよび6313ΔpabBに関して得られた結果を、この値と関連づけた。各実験を3通りで少なくとも3回実施した。
(図21)個々の抗原を検出する段階における感度に関する抗PabA、抗PabB、および抗PabD抗血清の検討を示す。融合タンパク質PabA、PabB、およびPabDの段階希釈物をニトロセルロース上にスポットし、各タンパク質に対するマウス血清の1:1000希釈物でプローブ処理した。結合状態の抗体を、抗マウスHRP結合抗体で標識し、化学発光法で可視化した。
(図22)フィブリノーゲン結合タンパク質の存在に関する、さまざまなS. agalactiae株、およびこれらの同質遺伝的なfbsA欠失変異体の培養上清のウェスタンブロット解析の結果を示す。さまざまなS. agalactiae株、およびこれらのfbsA欠失変異体の濃縮培養上清に由来する15 μgのタンパク質をSDS-PAGEで大きさによって分離し、ニトロセルロースにブロットし、ヒトフィブリノーゲンとの相互作用を検討した。ブロットをウサギ抗フィブリノーゲン抗体とインキュベートした後に、ワサビペルオキシダーゼを結合させたヤギ抗ウサギ抗体でインキュベートすることで、結合状態のフィブリノーゲンを検出した。フィブリノーゲン抗体複合体の検出には化学発光法を用いた。
(図23)固定化フィブリノーゲンに対する、さまざまなS. agalactiae株およびこれらのfbsA欠失変異体の結合を示す。同等の細胞数のさまざまな株を、テラサキプレートに固定化されたフィブリノーゲンとインキュベートした。フィブリノーゲンと結合した細菌の数を、アッセイ系への入力菌数と関連づけた。
(図24)さまざまなS. agalactiae株、およびこれらの同質遺伝的なfbsA変異株と、肺上皮細胞系列A549の接着および同細胞への内在化を示す。同等の数の細菌を使用してA549細胞に感染させ、A549細胞に接着した菌数(A)、および同細胞に内在化した菌数(B)を入力菌数と関連づけた。
(図25)S. agalactiae 6313株および6313ΔfbsAと繊維芽細胞細胞系列HEL299との接着および同細胞への内在化を示す。HEL299細胞にS. agalactiaeをMOIが10:1となるように感染させ、細胞と接着した菌の数と、細胞へ内在化した菌の数を入力菌数と関連づけた。
(図26)S. agalactiaeのA549細胞に対する接着に及ぼすFbsAタンパク質の影響を示す。接着アッセイ法を、さまざまな量の精製FbsA融合タンパク質の存在下で実施し、細菌と接着した細胞の数を入力菌数と関連づけた。
(図27)ヒトA549細胞に対する、FbsAでコーティングされたラテックスビーズの結合を示す。ラテックスビーズをBSA(A)、またはFbsA融合タンパク質(B〜D)のいずれかでコーティングし、コーティングされたビーズと肺上皮細胞系列A549の相互作用を走査型電子顕微鏡で解析した。
(図28)S. agalactiae のpabCコード領域の転写構成を示す。図の上部の名称は、S. agalactiae 6313の全RNAとのPCR(A)、全RNAとのRT-PCR(B)、または染色体DNAとのPCR(C)においてプライマー対がアニールした遺伝子を示す。
(図29)pabC遺伝子およびpabD遺伝子の存在に関する、GBS株のPCR解析の結果を示す。1、S. agalactiae 1137株(Ia);2、S. agalactiae A90/14株(Ib);3、S. agalactiae 6313株(III);4、S. agalactiae 4416 S3株(III); 5、S. agalactiae 4357株(V);6、S. agalactiae 4327株(V)をPCRに使用した。
(図30)S. agalactiae 6313、S. agalactiae NEM316、およびS. agalactiae 2003V_RのPabCタンパク質のアミノ酸配列の比較を示す。
(図31)S. agalactiaeのpabCコード領域の制限酵素切断地図(A)、ならびにフィブリノーゲン結合に関するPabCおよびGbs0851融合タンパク質のウェスタンブロット解析の結果(B)を示す。融合タンパク質をSDS-PAGEで大きさにより分離し、ニトロセルロース膜にトランスファーし、フィブリノーゲン結合をウェスタンブロッティングで検討した。結合状態のフィブリノーゲンを、ウサギ抗フィブリノーゲン抗体と、これに続くペルオキシダーゼ標識ヤギ抗ウサギ抗体で検出後に化学発光で可視化した。PabCおよびGbs0851=完全長の融合タンパク質、PabC-N=PabCのN末端の388アミノ酸、PabC-C=PabCのC末端の222アミノ酸。(C)は、ウェスタンブロット解析による、ヒトフィブリノーゲン中のFbsA結合部位およびPabC結合部位の同定を示す。ヒトフィブリノーゲンをSDS-PAGEで大きさによって分離し、クーマシー染色(左のレーン)を行うか、またはニトロセルロースにトランスファーし、FbsA結合またはPabC結合をウェスタンブロッティングで検討した。結合状態の融合タンパク質を、マウス抗ヒスチジンタグ抗体と、これに続くペルオキシダーゼ結合ヤギ抗マウスIgG fab断片で検出し、化学発光で可視化した。
(図32)捕捉ELISAアッセイ法による、固定化フィブリノーゲンと組換えPabC融合タンパク質の結合を示す。マイクロタイタープレートのウェルを一定量のヒトフィブリノーゲンでコーティング後に、さまざまなPabC融合タンパク質を濃度を高めながら添加した。結合状態の融合タンパク質を、マウス抗ヒスチジンタグ抗体およびペルオキシダーゼ結合ヤギ抗マウスIgG fab断片で検出した。発色を、テトラメチル-ベンジジン基質を添加して開始させ、H2SO4を添加して停止させた。マイクロタイターウェルの吸光度を450 nmで読み取った。値は、3回の独立した実験(各3通りで試行)の平均値である。
(図33)S. agalactiae 6313 pAT32株、ΔpabC pAT32株およびΔpabC pATpabC株のそれぞれと肺上皮細胞系列A549との接着(A)、および同細胞への浸潤(B)を示す。細菌の接着および浸潤は以下のように計算した:接着=接着菌数/アッセイ法の総菌数×100。浸潤=内在化菌数/アッセイ法の総菌数×100。各実験を3通りで少なくとも3回実施した。さまざまな量のPabCおよびBsp融合タンパク質の存在下におけるS. agalactiae 6313と真核細胞の接着(C)および浸潤(D)を示す。図32の説明に記載されているように、細菌の接着および浸潤を計算した。各実験を3通りで少なくとも3回実施した。
Claims (37)
- 以下からなる群より選択される核酸配列を含む、好ましくはフィブリノーゲン結合ポリペプチドまたはその断片をコードする単離された核酸分子:
(a)SEQ ID NO 1〜SEQ ID NO 6からなる群より選択される核酸配列と少なくとも70%が同一な核酸;
(b)(a)の核酸と本質的に相補的な核酸;
(c)(a)または(b)の核酸の少なくとも15の連続する塩基を含む核酸;
(d)ストリンジェントなハイブリダイゼーション条件で、(a)、(b)、または(c)のポリヌクレオチドとアニールする核酸;ならびに
(e)遺伝コードの縮重を除いて、(a)、(b)、(c)、または(d)で定義された核酸とハイブリダイズする可能性のある核酸。 - 以下からなる群より選択される核酸配列を含む、好ましくは接着因子またはその断片をコードする単離された核酸分子:
(a)Seq ID NO 7、 Seq ID NO 8、Seq ID NO 9、またはSeq ID NO 10に記載された核酸配列と少なくとも70%が同一な核酸;
(b)(a)の核酸と本質的に相補的な核酸;
(c)(a)または(b)の核酸の少なくとも15の連続する塩基を含む核酸;
(d)ストリンジェントなハイブリダイゼーション条件で、(a)、(b)または(c)の核酸とアニールする核酸;ならびに
(e)遺伝コードの縮重を除いて、(a)、(b)、(c)、または(d)で定義された核酸とハイブリダイズする可能性のある核酸。 - 同一性が少なくとも80%、好ましくは少なくとも90%、より好ましくは100%である、請求項1または2のどちらかに記載の単離された核酸分子。
- 核酸分子が、16アミノ酸を含むアミノ酸モチーフの少なくとも1個のリピートを含むフィブリノーゲン結合タンパク質をコードする、請求項1または3のどちらかに記載の単離された核酸分子。
- コードされたフィブリノーゲン結合タンパク質が、請求項7および15のどちらかに記載で特定されるアミノ酸モチーフのリピートを19個含む、請求項4記載の単離された核酸分子。
- 核酸分子が、上皮細胞、好ましくはヒト上皮細胞と相互作用する接着因子をコードする、請求項2または3のどちらかに記載の単離された核酸分子。
- ポリペプチドが
のアミノ酸モチーフを含むポリペプチドをコードする単離された核酸分子。 - 核酸がDNA、RNA、またはこの混合物であり、好ましくは核酸分子がゲノムDNAから単離される、請求項1〜7のいずれか一項記載の核酸。
- 請求項1〜8のいずれか一項記載の核酸分子を含むベクター。
- ベクターが、請求項1〜8のいずれか一項記載の核酸分子のいずれかにコードされたポリペプチドの組換え体発現に適合された、請求項8記載のベクター。
- 請求項9または10のどちらかに記載のベクターを含む、好ましくは宿主細胞である細胞。
- 請求項1〜8のいずれか一項記載の核酸分子にコードされたアミノ酸配列を含むポリペプチド、好ましくはフィブリノーゲン結合ポリペプチド、および/または接着因子、ならびに該ポリペプチドの断片。
- Seq ID NO 11〜20からなる群より選択されるアミノ酸配列を含むポリペプチド、好ましくはフィブリノーゲン結合ポリペプチド、および/または接着因子。
- Seq ID NO 113〜205からなる群より選択されるアミノ酸配列を含むポリペプチド、好ましくはフィブリノーゲン結合ポリペプチド、および/または接着因子。
- のアミノ酸モチーフを含むポリペプチド、好ましくはフィブリノーゲン結合ポリペプチド、および/または接着因子。
- 請求項1〜8のいずれか一項記載の核酸分子を発現させる段階を含む、請求項12〜15のいずれか一項記載のポリペプチド、またはその断片の作製法。
- 適切な宿主細胞を請求項9〜10のいずれか一項記載のベクターで形質転換する段階またはトランスフェクトする段階を含む、請求項12〜15のいずれか一項記載のポリペプチドまたはその断片を発現する細胞の作製法。
- 請求項12〜15のいずれか一項で定義されたポリペプチドもしくはその断片、または請求項1〜8のいずれか一項記載の核酸分子を含む、特にワクチンである薬学的組成物。
- 好ましくは、ポリカチオンポリマー、免疫刺激性デオキシヌクレオチド(ODN)、合成KLKペプチド、向神経活性化合物、ミョウバン、フロインドの完全アジュバントもしくは不完全アジュバント、またはこれらの組み合わせからなる群より選択される免疫刺激物質を含むことを特徴とする、請求項18記載の薬学的組成物。
- 薬物の製造、特に対細菌感染ワクチン製造のための、請求項12〜15のいずれか一項記載のポリペプチド、またはその断片の使用。
- 請求項12〜15のいずれか一項記載のポリペプチドまたはその断片の少なくとも選択的な部分と結合する抗体、または少なくともその有効部分。
- モノクローナル抗体、ポリクローナル抗体、キメラ抗体、ヒト化抗体、およびこれらの各断片からなる群より選択される、請求項21記載の抗体。
- 抗体を作製するための、請求項12〜15のいずれか一項記載のポリペプチド、またはその断片の使用。
- 細菌感染、特にストレプトコッカス・アガラクティエ(Streptococcus agalactiae)感染を治療または予防するための薬物を調製するための、請求項21または22のどちらかに記載の抗体の使用。
- 以下の段階を含む、請求項12〜15のいずれか一項記載のポリペプチドもしくはその断片の活性を低下もしくは阻害することが可能な拮抗物質、または請求項12〜15のいずれか一項記載のポリペプチドと結合可能な拮抗物質を同定する方法:
(a)請求項12〜15のいずれか一項記載の、単離または固定化されたポリペプチドもしくはその断片と候補拮抗物質とを、候補拮抗物質と該ポリペプチドもしくはその断片との結合を可能とする条件で、候補拮抗物質と該ポリペプチドもしくはその断片との結合に応じた検出シグナルを提供可能な成分の存在下で接触させる段階;ならびに
(b)拮抗物質と対象ポリペプチドもしくはその断片との結合に応じて発生したシグナルの有無、好ましくは、ポリペプチドもしくはその断片の活性の阻害もしくは低下を可能とする化合物の存在を示すシグナルの存在を検出する段階。 - 以下の段階を含む、請求項12〜15のいずれか一項記載のポリペプチドもしくはその断片の、活性の低下もしくは阻害を可能とする拮抗物質を同定する方法:
(a)請求項12〜15のいずれか一項記載のポリペプチド、またはその断片を提供する段階;
(b)請求項12〜15のいずれか一項記載のポリペプチドの相互作用パートナー、好ましくは請求項21または22のどちらかに記載の抗体を提供する段階;
(c)候補拮抗物質を提供する段階;
(d)ポリペプチド、ポリペプチドの相互作用パートナー、および候補拮抗物質を反応させる段階;ならびに
(e)候補拮抗物質がポリペプチドの活性を阻害するか、または低下させるか否かを判定する段階。 - 以下の段階を含む、相互作用パートナーに対する、請求項12〜15のいずれか一項記載のポリペプチドもしくはその断片の、相互作用活性の低下もしくは阻害を可能とする拮抗物質を同定する方法:
(a)請求項12〜15のいずれか一項記載のポリペプチドもしくはその断片を提供する段階;
(b)該ポリペプチドもしくはその断片に対する相互作用パートナー、好ましくは請求項21または22のどちらかに記載の抗体を提供する段階;
(c)該ポリペプチドもしくはその断片と相互作用パートナーとを相互作用させて、相互作用複合体を形成させる段階;
(d)候補拮抗物質を提供する段階;
(e)候補拮抗物質と相互作用複合体との間に競合反応が起こることを可能とする段階;ならびに
(f)候補拮抗物質が、ポリペプチドもしくはその断片と相互作用パートナーとの相互作用活性を阻害するか、または低下させるか否かを判定する段階。 - 請求項26または27のどちらかに記載の方法で同定されたか、または同定可能な拮抗物質。
- 先行する請求項のいずれか一項記載の核酸分子の存在、または先行する請求項のいずれか一項記載のポリペプチドの存在を判定する段階を含む、細菌感染、好ましくはStreptococcus agalactiae感染のインビトロ診断法。
- 請求項1〜8のいずれか一項記載のポリペプチドもしくはその断片をコードする核酸配列の存在、または請求項12〜15のいずれか一項記載のポリペプチドもしくはその断片の存在を判定する段階を含む、請求項12〜15のいずれか一項記載のポリペプチドもしくはその断片の発現に関連する疾患をインビトロで診断する方法。
- 支持材料、および支持材料に固定された、前記請求項のいずれか一項記載のポリペプチド、または前記請求項のいずれか一項記載の核酸分子を含むアフィニティ装置。
- ポリペプチドもしくはその断片の相互作用パートナーを単離および/または精製および/または同定するための、前記請求項のいずれか一項記載のポリペプチドもしくはその断片の使用。
- ポリペプチドに対するペプチド結合を生じるための、前記請求項のいずれか一項記載の任意のポリペプチドの使用。
- ペプチドがアンチカリン(anticaline)を含む群から選択される、請求項33記載の使用。
- アプタマーおよびシュピーゲルマーからなる群より選択される機能性核酸の作製または生成のための、前記請求項のいずれか一項記載のポリペプチドの使用。
- 前記請求項のいずれか一項記載のポリペプチドの抗原としての使用。
- リボザイム、アンチセンス核酸、およびsiRNAからなる群より選択される機能性リボ核酸の作製または生成のための、請求項1〜8のいずれか一項記載の核酸の使用。
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP02023141.1 | 2002-10-15 | ||
| EP02023141 | 2002-10-15 | ||
| EP03006393 | 2003-03-20 | ||
| EP03006393.7 | 2003-03-20 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2010051266A Division JP2010207219A (ja) | 2002-10-15 | 2010-03-09 | B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2014003979A true JP2014003979A (ja) | 2014-01-16 |
Family
ID=32109135
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005501294A Expired - Fee Related JP5116971B2 (ja) | 2002-10-15 | 2003-10-15 | B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 |
| JP2010051266A Pending JP2010207219A (ja) | 2002-10-15 | 2010-03-09 | B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 |
| JP2013138528A Pending JP2014003979A (ja) | 2002-10-15 | 2013-07-02 | B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 |
Family Applications Before (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005501294A Expired - Fee Related JP5116971B2 (ja) | 2002-10-15 | 2003-10-15 | B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 |
| JP2010051266A Pending JP2010207219A (ja) | 2002-10-15 | 2010-03-09 | B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 |
Country Status (7)
| Country | Link |
|---|---|
| US (3) | US7485710B2 (ja) |
| EP (3) | EP1551869A2 (ja) |
| JP (3) | JP5116971B2 (ja) |
| CN (2) | CN102174533A (ja) |
| AU (2) | AU2003274011B2 (ja) |
| CA (1) | CA2502414A1 (ja) |
| WO (1) | WO2004035618A2 (ja) |
Families Citing this family (21)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DK1328543T3 (da) * | 2000-10-27 | 2009-11-23 | Novartis Vaccines & Diagnostic | Nukleinsyrer og proteiner fra streptococcus-gruppe A & B |
| AU2003260102A1 (en) * | 2002-08-26 | 2004-03-11 | Chiron Corporation | Conserved and specific streptococcal genomes |
| WO2004035618A2 (en) | 2002-10-15 | 2004-04-29 | Intercell Ag | Nucleic acids coding for adhesion factor of group b streptococcus, adhesion factors of group b streptococcus and further uses thereof |
| JP4875490B2 (ja) | 2003-07-31 | 2012-02-15 | ノバルティス バクシンズ アンド ダイアグノスティックス,インコーポレーテッド | StreptococcusPyogenesについての免疫原組成物 |
| US8945589B2 (en) * | 2003-09-15 | 2015-02-03 | Novartis Vaccines And Diagnostics, Srl | Immunogenic compositions for Streptococcus agalactiae |
| WO2005076827A2 (en) * | 2004-02-04 | 2005-08-25 | Universita' Degli Studi Di Pavia | Fbsa compositions for use in promoting fibrinogen aggregation and potentiating fibrin polymerization |
| EP1784211A4 (en) * | 2004-07-29 | 2010-06-30 | Novartis Vaccines & Diagnostic | IMMUNOGENIC COMPOSITIONS FOR GRAMPOSITIVE BACTERIA SUCH AS STREPTOCOCCUS AGALACTIAE |
| WO2006042027A2 (en) * | 2004-10-08 | 2006-04-20 | Novartis Vaccines And Diagnostics Inc. | Immunogenic and therapeutic compositions for streptococcus pyogenes |
| EP1828231A2 (en) * | 2004-12-22 | 2007-09-05 | Novartis Vaccines and Diagnostics, Inc. | Group b streptococcus |
| DE112004003030B4 (de) * | 2004-12-23 | 2014-04-30 | Daimler Ag | Rückhaltevorrichtung für einen Insassen eines Fahrzeugs |
| US8401159B2 (en) * | 2005-11-30 | 2013-03-19 | On-Q Telecom Systems Co., Inc. | Data provision to a virtual personal assistant for handling calls in a communication system |
| DK2054431T3 (da) * | 2006-06-09 | 2012-01-02 | Novartis Ag | Konformere af bakterielle adhæsiner |
| WO2008108830A2 (en) * | 2006-10-30 | 2008-09-12 | Novartis Ag | Immunogenic and therapeutic compositions for streptococcus pyogenes |
| CA2675379A1 (en) * | 2007-01-12 | 2008-07-17 | Intercell Ag | Protective proteins of s. agalactiae, combinations thereof and methods of using the same |
| CA2699513C (en) | 2007-09-12 | 2018-03-13 | Novartis Ag | Gas57 mutant antigens and gas57 antibodies |
| PL2235046T3 (pl) * | 2007-12-21 | 2012-12-31 | Novartis Ag | Zmutowane formy streptolizyny O |
| WO2009094391A1 (en) * | 2008-01-23 | 2009-07-30 | Xencor, Inc. | Optimized cd40 antibodies and methods of using the same |
| US10738338B2 (en) | 2016-10-18 | 2020-08-11 | The Research Foundation for the State University | Method and composition for biocatalytic protein-oligonucleotide conjugation and protein-oligonucleotide conjugate |
| CN108840914B (zh) * | 2018-08-13 | 2022-07-01 | 内蒙古民族大学 | 一种具有免疫原性的多肽、其抗体的制备方法以及用途 |
| CN111690584A (zh) * | 2020-06-16 | 2020-09-22 | 中国水产科学研究院珠江水产研究所 | 重组乳酸乳球菌和罗非鱼无乳链球菌病疫苗 |
| CN121108327B (zh) * | 2025-11-14 | 2026-01-27 | 北京市华信行生物科技有限公司 | B族链球菌表面免疫原性蛋白单克隆抗体及其应用 |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2002034771A2 (en) * | 2000-10-27 | 2002-05-02 | Chiron Srl | Nucleic acids and proteins from streptococcus groups a & b |
Family Cites Families (29)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4946778A (en) | 1987-09-21 | 1990-08-07 | Genex Corporation | Single polypeptide chain binding molecules |
| EP0786469B1 (en) | 1990-06-11 | 2006-03-01 | Gilead Sciences, Inc. | Methods of use of nucleic acid ligands |
| ES2120949T4 (es) | 1990-06-28 | 2011-12-29 | Sanofi-Aventis Deutschland Gmbh 50% | Proteinas de fusión con porciones de inmunoglobulinas, su preparación y empleo. |
| US5858725A (en) | 1990-10-10 | 1999-01-12 | Glaxo Wellcome Inc. | Preparation of chimaeric antibodies using the recombinant PCR strategy |
| GB9022543D0 (en) | 1990-10-17 | 1990-11-28 | Wellcome Found | Antibody production |
| GB9022545D0 (en) | 1990-10-17 | 1990-11-28 | Wellcome Found | Culture medium |
| GB9022547D0 (en) | 1990-10-17 | 1990-11-28 | Wellcome Found | Purified immunoglobulin |
| GB9122820D0 (en) | 1991-10-28 | 1991-12-11 | Wellcome Found | Stabilised antibodies |
| ATE509102T1 (de) | 1994-07-15 | 2011-05-15 | Univ Iowa Res Found | Immunomodulatorische oligonukleotide |
| FR2735478B1 (fr) * | 1995-06-13 | 1997-08-22 | Pasteur Institut | Molecules polypeptidiques de stade pre-erythrocytaire du paludisme |
| RS50101B (sr) | 1996-02-24 | 2009-01-22 | Boehringer Ingelheim International Gmbh., | Farmaceutski preparati za imunomodulaciju |
| JP2001504448A (ja) | 1996-08-30 | 2001-04-03 | フュルステ,イェンス,ペーター | 核酸の鏡面対称選択および進化 |
| US5910441A (en) * | 1996-09-16 | 1999-06-08 | The Rockefeller University | DNA encoding fibronectin and fibrinogen binding protein from group A streptococci |
| US5989912A (en) | 1996-11-21 | 1999-11-23 | Oligos Etc. Inc. | Three component chimeric antisense oligonucleotides |
| US5849902A (en) | 1996-09-26 | 1998-12-15 | Oligos Etc. Inc. | Three component chimeric antisense oligonucleotides |
| DE19742706B4 (de) | 1997-09-26 | 2013-07-25 | Pieris Proteolab Ag | Lipocalinmuteine |
| DE19803453A1 (de) | 1998-01-30 | 1999-08-12 | Boehringer Ingelheim Int | Vakzine |
| US7098182B2 (en) * | 1998-07-27 | 2006-08-29 | Microbial Technics Limited | Nucleic acids and proteins from group B streptococcus |
| GB9816335D0 (en) * | 1998-07-27 | 1998-09-23 | Cortecs Uk Ltd | Proteins |
| WO2000006736A2 (en) | 1998-07-27 | 2000-02-10 | Microbial Technics Limited | Nucleic acids and proteins from group b streptococcus |
| ES2275499T3 (es) * | 1999-03-19 | 2007-06-16 | Glaxosmithkline Biologicals S.A. | Vacuna contra streptococcus pneumoniae. |
| AT408721B (de) | 1999-10-01 | 2002-02-25 | Cistem Biotechnologies Gmbh | Pharmazeutische zusammensetzung enthaltend ein antigen |
| AT409085B (de) | 2000-01-28 | 2002-05-27 | Cistem Biotechnologies Gmbh | Pharmazeutische zusammensetzung zur immunmodulation und herstellung von vakzinen |
| AU2001281812B2 (en) | 2000-06-08 | 2005-04-07 | Valneva Austria Gmbh | Immunostimulatory oligodeoxynucleotides |
| AT410173B (de) | 2000-06-08 | 2003-02-25 | Cistem Biotechnologies Gmbh | Antigene zusammensetzung |
| US20040170642A1 (en) | 2000-08-17 | 2004-09-02 | Jorg Fritiz | Vaccine which comprises at least one antigen and a cathelididin derived antimicrobial peptide of a derivative thereof |
| AT410635B (de) | 2000-10-18 | 2003-06-25 | Cistem Biotechnologies Gmbh | Vakzin-zusammensetzung |
| FR2824074A1 (fr) * | 2001-04-26 | 2002-10-31 | Pasteur Institut | Sequence du genome streptococcus agalactiae, application au developpement de vaccins, d'outils de diagnostic, et a l'identification de cibles therapeutiques |
| WO2004035618A2 (en) | 2002-10-15 | 2004-04-29 | Intercell Ag | Nucleic acids coding for adhesion factor of group b streptococcus, adhesion factors of group b streptococcus and further uses thereof |
-
2003
- 2003-10-15 WO PCT/EP2003/011436 patent/WO2004035618A2/en not_active Ceased
- 2003-10-15 CN CN2011100420771A patent/CN102174533A/zh active Pending
- 2003-10-15 EP EP03757983A patent/EP1551869A2/en not_active Ceased
- 2003-10-15 EP EP10010866A patent/EP2314603A3/en not_active Withdrawn
- 2003-10-15 AU AU2003274011A patent/AU2003274011B2/en not_active Ceased
- 2003-10-15 CN CN200380101524XA patent/CN1705679B/zh not_active Expired - Fee Related
- 2003-10-15 US US10/531,659 patent/US7485710B2/en not_active Expired - Fee Related
- 2003-10-15 EP EP10010867A patent/EP2314604A3/en not_active Withdrawn
- 2003-10-15 JP JP2005501294A patent/JP5116971B2/ja not_active Expired - Fee Related
- 2003-10-15 CA CA002502414A patent/CA2502414A1/en not_active Abandoned
-
2009
- 2009-01-30 US US12/363,229 patent/US7960533B2/en not_active Expired - Fee Related
-
2010
- 2010-03-09 JP JP2010051266A patent/JP2010207219A/ja active Pending
- 2010-03-16 AU AU2010201016A patent/AU2010201016B2/en not_active Ceased
-
2011
- 2011-05-24 US US13/114,615 patent/US8318908B2/en not_active Expired - Fee Related
-
2013
- 2013-07-02 JP JP2013138528A patent/JP2014003979A/ja active Pending
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2002034771A2 (en) * | 2000-10-27 | 2002-05-02 | Chiron Srl | Nucleic acids and proteins from streptococcus groups a & b |
Non-Patent Citations (1)
| Title |
|---|
| JPN6014051342; Mol. Microbiol., 2002, Vol.45, p.1499-1513 * |
Also Published As
| Publication number | Publication date |
|---|---|
| JP5116971B2 (ja) | 2013-01-09 |
| CA2502414A1 (en) | 2004-04-29 |
| EP1551869A2 (en) | 2005-07-13 |
| WO2004035618A3 (en) | 2004-09-30 |
| WO2004035618A2 (en) | 2004-04-29 |
| AU2003274011B2 (en) | 2010-03-04 |
| US7960533B2 (en) | 2011-06-14 |
| EP2314604A2 (en) | 2011-04-27 |
| US8318908B2 (en) | 2012-11-27 |
| JP2010207219A (ja) | 2010-09-24 |
| EP2314603A2 (en) | 2011-04-27 |
| CN1705679A (zh) | 2005-12-07 |
| CN1705679B (zh) | 2011-04-06 |
| EP2314604A3 (en) | 2011-05-25 |
| AU2010201016A1 (en) | 2010-04-08 |
| EP2314603A3 (en) | 2011-05-18 |
| US7485710B2 (en) | 2009-02-03 |
| AU2003274011A1 (en) | 2004-05-04 |
| US20100113340A1 (en) | 2010-05-06 |
| JP2006520583A (ja) | 2006-09-14 |
| US20060115479A1 (en) | 2006-06-01 |
| US20110287013A1 (en) | 2011-11-24 |
| CN102174533A (zh) | 2011-09-07 |
| AU2010201016B2 (en) | 2012-06-14 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2014003979A (ja) | B群連鎖球菌の接着因子をコードする核酸、b群連鎖球菌の接着因子、およびその使用 | |
| JP2000512487A (ja) | 新規化合物 | |
| JP2000509974A (ja) | 新規化合物 | |
| TW201021845A (en) | Prevention, treatment and diagnosis of P. gingivalis infection | |
| JP2001508650A (ja) | 新規細菌ポリペプチドおよびポリヌクレオチド | |
| US20130243779A1 (en) | Peptides protective against e. faecalis, methods and uses relating thereto | |
| AU2004226262A1 (en) | Staphylococcus epidermidis antigens | |
| JPH11103871A (ja) | 新規化合物 | |
| US20100221277A1 (en) | Chlamydia pneumoniae antigens | |
| JPH10201484A (ja) | 新規FtsL | |
| JP2000509984A (ja) | 新規化合物 | |
| JPH11151091A (ja) | 新規aroA | |
| JPH11103870A (ja) | 新規化合物 | |
| JP2000500326A (ja) | フィブロネクチン結合タンパク質b化合物 | |
| JP2002272487A (ja) | 新規pcrA | |
| JP2002253274A (ja) | 新規化合物 | |
| JP2000509985A (ja) | 新規化合物 | |
| JP2000504207A (ja) | スタフィロコッカス・アウレウス由来の2成分シグナルトランスダクション応答レギュレーターポリペプチド | |
| JPH10290693A (ja) | 新規フィブロネクチン結合蛋白化合物 | |
| JP2001505421A (ja) | 新規FabD | |
| JP2002119292A (ja) | 新規valS | |
| JPH10229886A (ja) | spo/rel |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20141203 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20150513 |