JP6964314B2 - 組換え微生物、組換え微生物を用いたピリドキサミン又はその塩の製造方法、及び組換え微生物を用いたピリドキサール又はその塩の製造方法 - Google Patents
組換え微生物、組換え微生物を用いたピリドキサミン又はその塩の製造方法、及び組換え微生物を用いたピリドキサール又はその塩の製造方法 Download PDFInfo
- Publication number
- JP6964314B2 JP6964314B2 JP2019517729A JP2019517729A JP6964314B2 JP 6964314 B2 JP6964314 B2 JP 6964314B2 JP 2019517729 A JP2019517729 A JP 2019517729A JP 2019517729 A JP2019517729 A JP 2019517729A JP 6964314 B2 JP6964314 B2 JP 6964314B2
- Authority
- JP
- Japan
- Prior art keywords
- seq
- amino acid
- nucleotide sequence
- pyridoxamine
- recombinant microorganism
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images






Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P17/00—Preparation of heterocyclic carbon compounds with only O, N, S, Se or Te as ring hetero atoms
- C12P17/10—Nitrogen as only ring hetero atom
- C12P17/12—Nitrogen as only ring hetero atom containing a six-membered hetero ring
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/70—Vectors or expression systems specially adapted for E. coli
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0006—Oxidoreductases (1.) acting on CH-OH groups as donors (1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0012—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7)
- C12N9/0014—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7) acting on the CH-NH2 group of donors (1.4)
- C12N9/0016—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7) acting on the CH-NH2 group of donors (1.4) with NAD or NADP as acceptor (1.4.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1096—Transferases (2.) transferring nitrogenous groups (2.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y101/00—Oxidoreductases acting on the CH-OH group of donors (1.1)
- C12Y101/01—Oxidoreductases acting on the CH-OH group of donors (1.1) with NAD+ or NADP+ as acceptor (1.1.1)
- C12Y101/01065—Pyridoxine 4-dehydrogenase (1.1.1.65)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y104/00—Oxidoreductases acting on the CH-NH2 group of donors (1.4)
- C12Y104/01—Oxidoreductases acting on the CH-NH2 group of donors (1.4) with NAD+ or NADP+ as acceptor (1.4.1)
- C12Y104/01001—Alanine dehydrogenase (1.4.1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y104/00—Oxidoreductases acting on the CH-NH2 group of donors (1.4)
- C12Y104/01—Oxidoreductases acting on the CH-NH2 group of donors (1.4) with NAD+ or NADP+ as acceptor (1.4.1)
- C12Y104/01002—Glutamate dehydrogenase (1.4.1.2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y206/00—Transferases transferring nitrogenous groups (2.6)
- C12Y206/01—Transaminases (2.6.1)
- C12Y206/01001—Aspartate transaminase (2.6.1.1), i.e. aspartate-aminotransferase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y206/00—Transferases transferring nitrogenous groups (2.6)
- C12Y206/01—Transaminases (2.6.1)
- C12Y206/0103—Pyridoxamine--pyruvate transaminase (2.6.1.30)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y206/00—Transferases transferring nitrogenous groups (2.6)
- C12Y206/01—Transaminases (2.6.1)
- C12Y206/01031—Pyridoxamine--oxaloacetate transaminase (2.6.1.31)
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Enzymes And Modification Thereof (AREA)
Description
ピリドキサミン及びその塩は化学的に合成することが可能である。例えば国際公開第2006/066806号は、アラニンとギ酸を出発物質としてピリドキサミン二塩酸塩を化学的に合成する方法を開示している。また、国際公開第2005/077902号は、ピリドキシンからピリドキサミンを化学的に合成する方法を開示している。
特開平9−107985号公報はリゾビウム属に属しビタミンB6生産能を有する微生物を、好気的条件下、培地中で培養し、生成したビタミンB6を培養液から得るビタミンB6の製造方法を開示している。
一方、特定の種の微生物を用いた方法である国際公開第2007/142222号に記載の方法では、ピリドキサールからピリドキサミンへの変換効率が微生物種によって大きくばらついており、また全体としてはさほど高いものではなかった。また、国際公開第2007/142222号に記載のピリドキシン存在下でのアクレモニウム フシジオイデスの培養実験では、生成物の大部分がピリドキサミンではなくピリドキサールであった。さらに、特開平9−107985号公報の記載においては、生成したビタミンB6は、そのほとんどがピリドキソール(ピリドキシン)であり、ピリドキサミンの生成はわずかであった。
上記のように、ピリドキサミン又はその塩の化学的合成(例えば国際公開第2006/066806号や国際公開第2005/077902号)は、多くの反応及び精製工程を経るものであって、高い収率が得られていない。また、微生物の中にはピリドキサミン合成能を有するものもいるものの(例えば国際公開第2007/142222号や特開平9−107985号公報)、そうした微生物を新たに発見することは手間がかかり、また、ビタミンB6群の中でピリドキサミン又はその塩を選択的に合成することができておらず、高いピリドキサミン生産効率も得られていない。また、このように天然に存在する微生物種から特定の微生物種をスクリーニングするというアプローチからは、ビタミンB6を高生産するためにはどの酵素あるいは遺伝子が重要であるかという分子生物学的な情報は得られない。
特公昭43−5712号公報に記載の方法では、反応系にカルボニル試薬の添加が必須である。カルボニル試薬は、生成されたピリドキサールをカルボニル誘導体に変換するものである。しかし、ピリドキサールを中間体として次の反応に供するためには、カルボニル誘導体を生成する方法は適していないことが多い。
本発明者らは、ピリドキサールの工業的生産の観点から、ピリドキシンデヒドロゲナーゼをコードする遺伝子が導入された工業的に利用可能な組換え微生物の作製を試みた。このような遺伝子組換え微生物の宿主細胞は、コスト及び製造の容易さの観点から原核生物であることが好ましいが、例えば導入する遺伝子が真核生物の遺伝子である場合、真核生物において発現するタンパク質をコードする遺伝子を原核生物に導入し発現させることは多くの困難を伴い、しばしば活性を保持したままのタンパク質を発現させることができない。そこで、本開示の第2の態様では、導入されたピリドキシンデヒドロゲナーゼを有し発現する、組換え微生物を提供する。
<1>
ピリドキシンデヒドロゲナーゼをコードする遺伝子、ピリドキサールからピリドキサミンを合成する酵素活性を有するピリドキサミン合成酵素をコードする遺伝子、及び前記ピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有するアミノ酸再生酵素をコードする遺伝子を有し、
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子、前記ピリドキサミン合成酵素をコードする遺伝子及び前記アミノ酸再生酵素をコードする遺伝子のうち少なくとも2つは菌体外から導入された遺伝子であるか又は菌体に内在するがその発現が強化された遺伝子である、組換え微生物。
<2>
前記ピリドキサミン合成酵素が、ピリドキサミン−ピルビン酸トランスアミナーゼ、ピリドキサミン−オキサロ酢酸トランスアミナーゼ、アスパラギン酸トランスアミナーゼ、又はピリドキサミンリン酸トランスアミナーゼである、<1>に記載の組換え微生物。
<3>
前記ピリドキシンデヒドロゲナーゼが下記の部分アミノ酸配列(a)及び部分アミノ酸配列(b)のうち少なくとも1つを含み、且つピリドキシンデヒドロゲナーゼ活性を有する、<1>又は<2>に記載の組換え微生物。
(a) NX1X2EX3YG(配列番号97)
(ここで、X1は、V、C、I、A、M、S、G又はLを表し、
X2は、G又はAを表し、
X3は、F又はLを表す)
(b) X4X5X6KGX7(配列番号98)
(X4は、I、V、F又はLを表し、
X5は、S、T、N、C又はMを表し、
X6は、C、V、A、I、W又はFを表し、
X7は、G、A、S又はCを表す)
<4>
前記ピリドキシンデヒドロゲナーゼが酵素番号EC1.1.1.65で表される、<1>〜<3>のうちいずれか1つに記載の組換え微生物。
<5>
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子がSaccharomyces cerevisiaeに由来する、<1>〜<4>のうちいずれか1つに記載の組換え微生物。
<6>
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子が、
配列番号7及び配列番号53〜配列番号59のうちいずれかのヌクレオチド配列を有するか、
配列番号13及び配列番号75〜81のいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域を有するか、
配列番号7及び配列番号53〜配列番号59のうちいずれかのヌクレオチド配列と相補的なヌクレオチド配列を有するDNA、又は配列番号13及び配列番号75〜81のいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有する、<1>〜<5>のうちいずれか1つに記載の組換え微生物。
<7>
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子が、配列番号7のヌクレオチド配列を有するか、配列番号7のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有する、<1>〜<6>のうちいずれか1つに記載の組換え微生物。
<8>
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子がSchizosaccharomyces pombeに由来する、<1>〜<4>のうちいずれか1つに記載の組換え微生物。
<9>
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子が、配列番号8のヌクレオチド配列を有するか、配列番号8のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有する、<1>〜<4>及び<8>のうちいずれか1つに記載の組換え微生物。
<10>
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子が、配列番号1、配列番号2及び配列番号22〜配列番号28のうちいずれかのアミノ酸配列、又は配列番号1、配列番号2及び配列番号22〜28のアミノ酸配列のうち少なくとも1つと80%以上の配列同一性を有し且つピリドキシンデヒドロゲナーゼ活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、<1>〜<9>のうちいずれか1つに記載の組換え微生物。
<11>
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子が、配列番号1又は配列番号2のアミノ酸配列、又は配列番号1のアミノ酸配列及び配列番号2のアミノ酸配列のうち少なくとも1つと80%以上の配列同一性を有し且つピリドキシンデヒドロゲナーゼ活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、<1>〜<10>のうちいずれか1つに記載の組換え微生物。
<12>
前記ピリドキサミン合成酵素が、下記の部分アミノ酸配列(c)、部分アミノ酸配列(d)、部分アミノ酸配列(e)、部分アミノ酸配列(f)、部分アミノ酸配列(g)及び部分アミノ酸配列(h)のうち少なくとも1つを含み、且つピリドキサールからピリドキサミンを合成する酵素活性を有する、<1>〜<11>のうちいずれか1つに記載の組換え微生物。
(c) X8X9X10X11X12X13(配列番号99)
(X8は、L、M、I又はVを表し、
X9は、H又はQを表し、
X10は、G、C又はAを表し、
X11は、E又はDを表し、
X12は、P又はAを表し、
X13は、V、I、L又はAを表す)
(d) X14X15TPSGTX16X17(配列番号100)
(X14は、H又はSを表し、
X15は、D又はEを表し、
X16は、I、V又はLを表し、
X17は、N又はTを表す)
(e) X18DX19VSX20X21(配列番号101)
(X18は、V、I又はAを表し、
X19は、A、T又はSを表し、
X20は、S、A又はGを表し、
X21は、F、W、又はVを表す)
(f) X22X23X24KCX25GX26X27P(配列番号102)
(X22は、G又はSを表し、
X23は、P、S又はAを表し、
X24は、N、G、S、A又はQを表し、
X25は、L又はMを表し、
X26は、A、S、C又はGを表し、
X27は、P、T、S又はAを表す)
(g) X28X29X30X31SX32GX33X34(配列番号103)
(X28は、G又はDを表し、
X29は、V又はIを表し、
X30は、V、T、A、S、M、I又はLを表し、
X31は、F、M、L、I又はVを表し、
X32は、S、G、A、T、I、L又はHを表し、
X33は、R、M又はQを表し、
X34は、G、R、A、D、H又はKを表す)
(h) X35X36RX37X38HMGX39X40A(配列番号104)
(X35は、L又はVを表し、
X36は、T、I、V又はLを表し、
X37は、I、V又はLを表し、
X38は、G又はSを表し、
X39は、P、A又はRを表し、
X40は、T、V又はSを表す)
<13>
前記ピリドキサミン合成酵素が酵素番号EC2.6.1.30で表される、<1>〜<12>のうちいずれか1つに記載の組換え微生物。
<14>
前記ピリドキサミン合成酵素をコードする遺伝子がMesorhizobium lotiに由来する、<1>〜<13>のうちのいずれか1つに記載の組換え微生物。
<15>
前記ピリドキサミン合成酵素をコードする遺伝子が、
配列番号9及び配列番号60〜配列番号66のうちいずれかのヌクレオチド配列を有するか、
配列番号14のヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域又は配列番号82〜配列番号88のうちいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域を有するか、
配列番号9及び配列番号60〜配列番号66のうちいずれかのヌクレオチド配列と相補的なヌクレオチド配列を有するDNA、又は配列番号14のヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域若しくは配列番号82〜配列番号88のうちいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキサールからピリドキサミンを合成する酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、<1>〜<14>のうちいずれか1つに記載の組換え微生物。
<16>
前記ピリドキサミン合成酵素をコードする遺伝子が、配列番号9のヌクレオチド配列を有するか、配列番号9のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキサールからピリドキサミンを合成する酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、<1>〜<15>のうちいずれか1つに記載の組換え微生物。
<17>
前記ピリドキサミン合成酵素をコードする遺伝子が、配列番号3及び配列番号29〜配列番号35のうちいずれかのアミノ酸配列、又は配列番号3及び配列番号29〜配列番号35のアミノ酸配列のうち少なくとも1つと80%以上の配列同一性を有し且つピリドキサールからピリドキサミンを合成する酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、<1>〜<16>のうちいずれか1つに記載の組換え微生物。
<18>
前記ピリドキサミン合成酵素をコードする遺伝子が、配列番号3のアミノ酸配列、又は配列番号3のアミノ酸配列と80%以上の配列同一性を有し且つピリドキサールからピリドキサミンを合成する酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、<1>〜<17>のうちいずれか1つに記載の組換え微生物。
<19>
前記アミノ酸再生酵素がアラニンデヒドロゲナーゼである、<12>〜<18>のうちいずれか1つに記載の組換え微生物。
<20>
前記アラニンデヒドロゲナーゼが、ピルビン酸及びNH3からL−アラニンを生成する反応の際にNADPHを補酵素として利用可能である、<19>に記載の組換え微生物。
<21>
前記アラニンデヒドロゲナーゼが下記の部分アミノ酸配列(i)、部分アミノ酸配列(j)、部分アミノ酸配列(k)及び部分アミノ酸配列(l)のうち少なくとも1つを含み、且つピルビン酸及びNH3からL−アラニンを生成する活性を有する<19>又は<20>に記載の組換え微生物。
(i) EX41KX42X43EX44RX45X46(配列番号105)
(X41は、I、T、S、F、N又はVを表し、
X42は、N、M、A、V、L、T又はDを表し、
X43は、H、N、L又はQを表し、
X44は、Y、N又はFを表し、
X45は、V又はIを表し、
X46は、G又はAを表す)
(j) X47X48X49KVKEPX50(配列番号106)
(X47は、M又はLを表し、
X48は、I、L又はVを表し、
X49は、V、L、I又はMを表し、
X50は、Q、L、V、N又はIを表す)
(k) LX51TYLHLA(配列番号107)
(X51は、F又はYを表す)
(l) X52DX53AX54DQGG(配列番号108)
(X52は、V又はAを表し、
X53は、V又はIを表し、
X54は、I又はVを表す)
<22>
前記アミノ酸再生酵素をコードする遺伝子がShewanella sp. AC10に由来する、<19>〜<21>のうちいずれか1つに記載の組換え微生物。
<23>
前記アミノ酸再生酵素をコードする遺伝子が、
配列番号11及び配列番号67〜配列番号74のうちいずれかのヌクレオチド配列を有するか、
配列番号15及び配列番号89〜配列番号96のうちいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域を有するか、
配列番号11及び配列番号67〜配列番号74のうちいずれかのヌクレオチド配列と相補的なヌクレオチド配列を有するDNA、又は配列番号15及び配列番号89〜配列番号96のうちいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つアラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、<19>〜<22>のうちいずれか1つに記載の組換え微生物。
<24>
前記アミノ酸再生酵素をコードする遺伝子が、配列番号11のヌクレオチド配列を有するか、配列番号11のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つアラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、<19>〜<23>のうちいずれか1つに記載の組換え微生物。
<25>
前記アミノ酸再生酵素をコードする遺伝子が、配列番号5及び配列番号45〜配列番号52のうちいずれかのアミノ酸配列、又は配列番号5及び配列番号45〜配列番号52のアミノ酸配列のうち少なくとも1つと80%以上の配列同一性を有し且つアラニン再生酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、<19>〜<24>のうちいずれか1つに記載の組換え微生物。
<26>
前記アミノ酸再生酵素をコードする遺伝子が、配列番号5のアミノ酸配列、又は配列番号5のアミノ酸配列と80%以上の配列同一性を有し且つアラニン再生酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、<19>〜<25>のうちいずれか1つに記載の組換え微生物。
<27>
前記ピリドキサミン合成酵素が酵素番号EC2.6.1.31又はEC2.6.1.1で表される、<1>〜<11>のうちいずれか1つに記載の組換え微生物。
<28>
前記ピリドキサミン合成酵素をコードする遺伝子がEscherichia coliに由来する、<1>〜<11>及び<27>のうちいずれか1つに記載の組換え微生物。
<29>
前記ピリドキサミン合成酵素をコードする遺伝子が、配列番号10のヌクレオチド配列を有するか、配列番号10のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキサールからピリドキサミンを合成する酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、<1>〜<11>、<27>及び<28>のうちいずれか1つに記載の組換え微生物。
<30>
前記ピリドキサミン合成酵素をコードする遺伝子が、配列番号4のアミノ酸配列、又は配列番号4のアミノ酸配列と80%以上の配列同一性を有し且つピリドキサールからピリドキサミンを合成する酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、<1>〜<11>及び<27>〜<29>のうちいずれか1つに記載の組換え微生物。
<31>
前記アミノ酸再生酵素がグルタミン酸デヒドロゲナーゼである、<27>〜<30>のうちいずれか1つに記載の組換え微生物。
<32>
前記アミノ酸再生酵素をコードする遺伝子がEscherichia coliに由来する、<31>に記載の組換え微生物。
<33>
前記アミノ酸再生酵素をコードする遺伝子が、配列番号12のヌクレオチド配列を有するか、配列番号12のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つグルタミン酸再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、<31>又は<32>に記載の組換え微生物。
<34>
前記アミノ酸再生酵素をコードする遺伝子が、配列番号6のアミノ酸配列、又は配列番号6のアミノ酸配列と80%以上の配列同一性を有し且つグルタミン酸再生酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、<31>〜<33>のうちいずれか1つに記載の組換え微生物。
<35>
組換え大腸菌である、<1>〜<34>のうちいずれか1つに記載の組換え微生物。
<36>
<1>〜<35>のうちいずれか1つに記載の組換え微生物若しくは前記組換え微生物の培養物又は前記組換え微生物若しくは前記培養物の処理物と、ピリドキシン又はその塩とを接触させ、ピリドキサミン又はその塩を生産させる、ピリドキサミン又はその塩の製造方法。
<37>
前記組換え微生物若しくは前記組換え微生物の培養物又は前記組換え微生物若しくは前記培養物の処理物が、前記ピリドキシンデヒドロゲナーゼ、前記ピリドキサミン合成酵素、及び前記アミノ酸再生酵素を含む、<36>に記載の製造方法。
<38>
前記組換え微生物の処理物又は前記組換え微生物の培養物の処理物が、加熱処理、冷却処理、細胞の機械的破壊、超音波処理、凍結融解処理、乾燥処理、加圧又は減圧処理、浸透圧処理、細胞の自己消化、界面活性剤処理、酵素処理、細胞分離処理、精製処理及び抽出処理からなる群から選択される1つ以上を含む処理による処理物である、<36>又は<37>に記載の製造方法。
<39>
以下の(1)〜(7)に記載のポリヌクレオチドのうち少なくとも1つが導入された組換え微生物:
(1)配列番号1のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、
(2)配列番号1のアミノ酸配列において、1個〜50個のアミノ酸が欠失、付加又は置換されているアミノ酸配列からなるタンパク質をコードするポリヌクレオチドであって、前記タンパク質はピリドキシン若しくはその塩又はピリドキサール若しくはその塩の合成活性を有するものである、ポリヌクレオチド、
(3)配列番号1のアミノ酸配列と少なくとも60%の配列同一性を有するアミノ酸配列からなるタンパク質をコードするポリヌクレオチドであって、前記タンパク質はピリドキシン若しくはその塩又はピリドキサール若しくはその塩の合成活性を有するものである、ポリヌクレオチド、
(4)配列番号13のヌクレオチド配列からなるポリヌクレオチド、
(5)配列番号13のヌクレオチド配列において、1個〜150個のヌクレオチドが欠失、付加又は置換されているヌクレオチド配列からなるポリヌクレオチドであって、前記ポリヌクレオチドはピリドキシン若しくはその塩又はピリドキサール若しくはその塩の合成活性を有するタンパク質をコードするポリヌクレオチド、
(6)配列番号13のヌクレオチド配列と少なくとも60%の配列同一性を有するヌクレオチド配列からなるポリヌクレオチドであって、前記ポリヌクレオチドはピリドキシン若しくはその塩又はピリドキサール若しくはその塩の合成活性を有するタンパク質をコードするポリヌクレオチド、
(7)配列番号13のヌクレオチド配列と相補的なヌクレオチド配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドであって、前記ポリヌクレオチドはピリドキシン若しくはその塩又はピリドキサール若しくはその塩の合成活性を有するタンパク質をコードするポリヌクレオチド。
<40>
前記組換え微生物が組換え細菌である、<39>に記載の組換え微生物。
<41>
前記組換え微生物が組換え大腸菌である、<40>に記載の組換え微生物。
<42>
前記タンパク質が、EC番号1.1.1.65に分類される酵素である、<39>〜<41>のうちいずれか1つに記載の組換え微生物。
<43>
前記タンパク質が、Saccharomyces cerevisiaeに由来するピリドキシンデヒドロゲナーゼである、<39>〜<42>のうちいずれか1つに記載の組換え微生物。
<44>
<39>〜<43>のうちいずれか1つに記載の組換え微生物と、ピリドキシン若しくはその塩とを接触させることを含む、ピリドキサール若しくはその塩の製造方法。
<45>
<39>〜<43>のうちいずれか1つに記載の組換え微生物を培養して得られる培養物又は培養物の処理物と、ピリドキシン若しくはその塩とを接触させることを含む、ピリドキサール若しくはその塩の製造方法。
本開示の第2の態様によれば、導入されたピリドキシンデヒドロゲナーゼを有し発現する、組換え微生物が提供される。
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子、前記ピリドキサミン合成酵素をコードする遺伝子及び前記アミノ酸再生酵素をコードする遺伝子のうち少なくとも2つは菌体外から導入された遺伝子であるか又は元々菌体に内在するがその発現が強化された遺伝子である、組換え微生物(以下、第1の態様に係る組換え微生物という)を提供する。本開示の第2の態様(以下、単に第2の態様と称する)は、(1)配列番号1のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、(2)配列番号1のアミノ酸配列において、1個〜50個のアミノ酸が欠失、付加又は置換されているアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、(3)配列番号1のアミノ酸配列と少なくとも60%の配列同一性を有するアミノ酸配列からなるタンパク質をコードするポリヌクレオチド、(4)配列番号13のヌクレオチド配列からなるポリヌクレオチド、(5)配列番号13のヌクレオチド配列において、1個〜150個のヌクレオチドが欠失、付加又は置換されているヌクレオチド配列からなるポリヌクレオチド、(6)配列番号13のヌクレオチド配列と少なくとも60%の配列同一性を有するヌクレオチド配列からなるポリヌクレオチド、又は、(7)配列番号13のヌクレオチド配列と相補的なヌクレオチド配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドを導入された組換え微生物を提供する。「本開示」の記載は、第1の態様と第2の態様を包含する意味で用いられる。なお、本開示において導入とは、菌体内で発現可能となるように導入することをいう。また、「組換え」の語は「遺伝子組換え」と同義で用いられる。
これまで、化学的方法によっても生物学的方法によっても、ピリドキシン又はその塩からピリドキサミン又はその塩を高い生産効率で且つ安価に生産する方法は知られていなかった。ピリドキサミンの構造を以下に示す。
ピリドキシンデヒドロゲナーゼはピリドキサールレダクターゼあるいはピリドキシン−4−デヒドロゲナーゼとも呼ばれる酵素である。ピリドキシンデヒドロゲナーゼは酵素番号EC1.1.1.65で表される酵素であってもよく、他の酵素番号で表される酵素であってもよい。ピリドキシンデヒドロゲナーゼは、ピリドキサールに水素を加えてピリドキシンに変換する反応あるいはこれの逆反応を触媒する酵素活性を有する酵素である。なお、ピリドキサールやピリドキシンは周囲の環境により塩としても存在しうるが、表記の簡略化のため、本開示においては酵素の活性の記載については塩への言及は省略して表記する。ピリドキシンデヒドロゲナーゼは、ピリドキサールに水素を付加する際には、補酵素としてNADH(還元型ニコチンアミドアデニンジヌクレオチド)又はNADPH(還元型ニコチンアミドアデニンジヌクレオチドリン酸)を消費し、ピリドキシンから水素を引き抜く際には補酵素としてNAD+(酸化型ニコチンアミドアデニンジヌクレオチド)又はNADP+(酸化型ニコチンアミドアデニンジヌクレオチドリン酸)を消費する。一般的な条件下では、この酵素が触媒する反応の平衡はピリドキシン生成側に大きく偏っているため、ピリドキシンデヒドロゲナーゼをピリドキシンを消費する反応のために用いても高い収率は達成できないと考えられてきた。しかし、第1の態様においては、上記のように特定の他の酵素と組み合わせることにより、ピリドキシン又はその塩を原料としたピリドキサミン又はその塩の生産が高い生産効率で達成できるという予想外の効果が得られた。
第1の態様で用いられるピリドキサミン合成酵素とは、ピリドキサールからピリドキサミンを合成する酵素活性を有する任意の酵素を指す。なお、ピリドキサールやピリドキサミンは周囲の環境により塩としても存在しうるが、表記の簡略化のため、本開示においては酵素の活性の記載については塩への言及は省略して表記する。ピリドキサミン合成酵素の例としては、例えば、ピリドキサミン−ピルビン酸トランスアミナーゼ(例えば酵素番号EC2.6.1.30で表される酵素)、アスパラギン酸トランスアミナーゼ(例えばEC2.6.1.1で表される酵素)、ピリドキサミン−オキサロ酢酸トランスアミナーゼ(例えばEC2.6.1.31で表される酵素)及びピリドキサミンリン酸トランスアミナーゼ(例えばEC2.6.1.54で表される酵素)が挙げられる。
なおEC2.6.1.1で表されるアスパラギン酸トランスアミナーゼは、ピリドキサールリン酸を補酵素とするホロ酵素であり、アスパラギン酸のアミノ基を2−オキソグルタル酸に転移し、グルタミン酸とオキサロ酢酸を生成する酵素活性を有する。当該アスパラギン酸トランスアミナーゼはピリドキサールリン酸と結合していないアポ酵素の状態では、グルタミン酸又はアスパラギン酸のアミノ基をピリドキサールに転移しピリドキサミンを合成することが知られている(JOURNAL OF BIOLOGICAL CHEMISTRY, 1962年1月, Vol.237, No.1, p.127-132)。つまりEC2.6.1.1で表されるアスパラギン酸トランスアミナーゼのアポ酵素の形態がEC2.6.1.31のピリドキサミン−オキサロ酢酸トランスアミナーゼである。このため、アスパラギン酸トランスアミナーゼは、補酵素であるピリドキサールリン酸が存在しない又は量が少ない場合にはアポ酵素として存在し、ピリドキサミンを合成する。
ピリドキサミン合成酵素は、ピリドキサール又はその塩からピリドキサミン又はその塩を合成する際に、特定のアミノ酸のアミノ基部分を(=O)に酸化し、当該アミノ基を転移させてピリドキサミン又はその塩を生成する。例えば、ピリドキサミン−ピルビン酸トランスアミナーゼはL−アラニン及びD−アラニンのいずれも利用可能であり、ピリドキサミン−オキサロ酢酸トランスアミナーゼ及びアポ酵素の状態のアスパラギン酸トランスアミナーゼはD−アスパラギン酸、L−アスパラギン酸、D−グルタミン酸、及びL−グルタミン酸のいずれも利用可能であり、ピリドキサミンリン酸トランスアミナーゼはD−グルタミン酸を利用可能である。
第1の態様で用いられるアミノ酸再生酵素とは、前記ピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有する任意の酵素を指す。ピリドキサミンはアミノ基を有しており、ピリドキサミン合成の際にはアミノ基の提供源としてアミノ酸が消費されると考えられる。なお、アミノ酸は周囲の環境により塩としても存在しうるが、表記の簡略化のため、本開示においては酵素の活性の記載については塩への言及は省略して表記する。
例えば、前記ピリドキサミン合成酵素がL−アラニンを消費する場合には、L−アラニンを再生できる酵素がアミノ酸再生酵素として使用可能である。また、前記ピリドキサミン合成酵素がL−グルタミン酸又はL−アスパラギン酸を消費する場合には、L−グルタミン酸又はL−アスパラギン酸を再生できる酵素がアミノ酸再生酵素として使用可能である。
このようなアミノ酸再生酵素の例としては、アラニンデヒドロゲナーゼ(例えば酵素番号1.4.1.1で表される酵素)、グルタミン酸デヒドロゲナーゼ(例えば、酵素番号1.4.1.2、又は1.4.1.3、又は1.4.1.4で表される酵素)、アミノ酸配列の改変によりNADP+/NADPHを補酵素として利用可能となった改変型アラニンデヒドロゲナーゼなどが挙げられる。なお、第1の態様においては、NADH又はNADPHを補酵素として用いてピルビン酸及びNH3からL−アラニンを生成する酵素活性のことをアラニン再生酵素活性と呼び、NADH又はNADPHを補酵素として用いて2−オキソグルタル酸及びNH3からL−グルタミン酸を生成する酵素活性のことをグルタミン酸再生酵素活性と呼ぶことがある。
ピリドキサミン合成酵素との好ましい組み合わせとしては、ピリドキサミン合成酵素としてのピリドキサミン−ピルビン酸トランスアミナーゼと、アミノ酸再生酵素としてのアラニンデヒドロゲナーゼとの組み合わせ、及びピリドキサミン合成酵素としてのピリドキサミン−オキサロ酢酸トランスアミナーゼ又はアスパラギン酸トランスアミナーゼと、アミノ酸再生酵素としてのグルタミン酸デヒドロゲナーゼとの組み合わせ、が挙げられる。アミノ酸再生酵素は、菌体外から導入された遺伝子又は菌体に内在するがその発現が強化された遺伝子によってコードされるピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有することが好ましい。
例えば、アラニンデヒドロゲナーゼの場合は、ピルビン酸及びNH3からL−アラニンを生成する反応の際にNADPHを補酵素として利用可能であるアラニンデヒドロゲナーゼであることが好ましい。NADPHを補酵素として利用可能なアミノ酸再生酵素を使用した場合には、NADPHの消費によりNADPH/NADP+比が変化し、ピリドキシンデヒドロゲナーゼが触媒する反応の平衡や速度に影響を与えて、ピリドキサミン又はその塩の生産効率をより高めることができる。また、ほとんどのアラニンデヒドロゲナーゼは、NADH(還元型ニコチンアミドアデニンジヌクレオチド)を補酵素として利用可能であるが、NADPHを利用することができない。NADPHを補酵素として利用可能であるアラニンデヒドロゲナーゼを使用した場合には、上記のとおりNADPHの消費によりNADPH/NADP+比が変化し、ピリドキシンデヒドロゲナーゼが触媒する反応の平衡や速度に影響を与えて、ピリドキサミン又はその塩の生産効率をより高めることができる。また、前記アミノ酸再生酵素(例えばアラニンデヒドロゲナーゼ又はグルタミン酸デヒドロゲナーゼ)は、アミノ酸を再生する反応の際にNADH(還元型ニコチンアミドアデニンジヌクレオチド)及びNADPH(還元型ニコチンアミドアデニンジヌクレオチドリン酸)のどちらも補酵素として利用可能であることも好ましい。このようなアラニンデヒドロゲナーゼの例としては、後述の配列番号5及び配列番号45〜配列番号52のうちいずれかのアミノ酸配列を有するタンパク質が挙げられるが、これに限定されない。
あるいは、各酵素は、当該酵素活性を有する既知のアミノ酸配列(例えば、生物中に天然に存在する遺伝子によってコードされるアミノ酸配列)そのものを有するタンパク質、又は当該酵素活性を有する既知のアミノ酸配列(例えば、生物中に天然に存在する遺伝子によってコードされるアミノ酸配列)に対して80%以上、あるいは85%以上、あるいは90%以上、あるいは95%以上の配列同一性を有するアミノ酸配列を有し、所望の酵素活性(上述の酵素活性)を有するタンパク質であってもよい。ここで、配列同一性は例えばBLAST(登録商標、National Library of Medicine)プログラムを用いてデフォールトパラメータで評価することができる。
あるいは、ピリドキシンデヒドロゲナーゼは、配列番号1若しくは配列番号2のアミノ酸配列を有するタンパク質又は配列番号1のアミノ酸配列及び配列番号2のアミノ酸配列のうち少なくとも1つに対して例えば80%以上、又は85%以上、又は90%以上、又は95%以上の配列同一性を有するアミノ酸配列を有するタンパク質であってもよい。
上記のような、配列番号1のアミノ酸配列及び配列番号2のアミノ酸配列のうち1つ以上に類似するアミノ酸配列を有するタンパク質を使用する場合には、該タンパク質はピリドキシンデヒドロゲナーゼとしての活性を有しているべきである。ピリドキシンデヒドロゲナーゼ活性は、例えば、基質としてのピリドキシン及び必要なNAD+若しくはNADP+を含む水溶液に試験対象のタンパク質を加え、生成したピリドキサールを高速液体クロマトグラフィーで定量するか、あるいは生成したピリドキサールとトリスヒドロキシメチルアミノメタンのようなアミンとの間でシッフ塩基を形成させ415nm等での吸光度測定などで該シッフ塩基を定量することにより測定できる。
(a) NX1X2EX3YG(配列番号97)
(ここで、X1は、V、C、I、A、M、S、G又はLを表し、
X2は、G又はAを表し、
X3は、F又はLを表す)
(b) X4X5X6KGX7(配列番号98)
(X4は、I、V、F又はLを表し、
X5は、S、T、N、C又はMを表し、
X6は、C、V、A、I、W又はFを表し、
X7は、G、A、S又はCを表す)
図1−1及び図1−2には、配列番号1及び配列番号22〜配列番号28の配列のアラインメントを示す。図1−1及び図1−2中、ScPlrはSaccharomyces cerevisiaeのピリドキシンデヒドロゲナーゼを、SePlrはSaccharomyces eubayanusのピリドキシンデヒドロゲナーゼを、TdPlrはTorulaspora delbrueckiiのピリドキシンデヒドロゲナーゼを、ZbPlrはZygosaccharomyces bailiiのピリドキシンデヒドロゲナーゼを、KmPlrはKluyveromyces marxianusのピリドキシンデヒドロゲナーゼを、AoPlrはAspergillus oryzaeのピリドキシンデヒドロゲナーゼを、CaPlrはCandida albicansのピリドキシンデヒドロゲナーゼを、YlPlrはYarrowia lipolyticaのピリドキシンデヒドロゲナーゼを表す。部分アミノ酸配列(a)は、Saccharomyces cerevisiaeのピリドキシンデヒドロゲナーゼのN末端から55番目〜61番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当し、部分アミノ酸配列(b)は、Saccharomyces cerevisiaeのピリドキシンデヒドロゲナーゼのN末端から86番目〜91番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。本開示において配列同士のアラインメントは、例えばBLAST(登録商標、National Library of Medicine)プログラムを用いてデフォールトパラメータで行うことができる。部分アミノ酸配列(a)は、タンパク質のN末端から45番目〜71番目のアミノ酸残基の領域内に存在することが好ましく、N末端から46番目〜66番目のアミノ酸残基の領域内に存在することがより好ましい。部分アミノ酸配列(b)は、タンパク質のN末端から76番目〜101番目のアミノ酸残基の領域内に存在することが好ましく、N末端から79番目〜96番目のアミノ酸残基の領域内に存在することがより好ましい。
(a−1)X1X2NX3X4EX5YGX6X7(配列番号109)
X1は、F、L、I、M、Y又はVを表し、
X2は、 F、I、Y、L、W又はVを表し、
X3は、V、C、I、A、M、S、G又はLを表し、
X4は、G又はAを表し、
X5は、F又はLを表し、
X6は、P、K、R、G、E、T、A、N又はSを表し、
X7は、D、N、H、K、E、P、L又はIを表す。
部分アミノ酸配列(a−1)は、Saccharomyces cerevisiaeのピリドキシンデヒドロゲナーゼのN末端から53番目〜63番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(a−1)は、タンパク質のN末端から43番目〜73番目のアミノ酸残基の領域内に存在することが好ましく、N末端から45番目〜68番目のアミノ酸残基の領域内に存在することがより好ましい。
(b−1)X1X2X3X4X5X6X7X8KGX9(配列番号110)
X1は、R、K、A、S又はNを表し、
X2は、K、S、E、Q、D又はAを表し、
X3は、D、H、N、Y、E、K、C、Q又はRを表し、
X4は、V、T、I、M又はLを表し、
X5は、V、I、L、F、M又はTを表し、
X6は、I、V、F又はLを表し、
X7は、S、T、N、C又はMを表し、
X8は、C、V、A、I、W又はFを表し、
X9は、G、A、S又はCを表す。
部分アミノ酸配列(b−1)は、Saccharomyces cerevisiaeのピリドキシンデヒドロゲナーゼのN末端から81番目〜91番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(b−1)は、タンパク質のN末端から71番目〜101番目のアミノ酸残基の領域内に存在することが好ましく、N末端から74番目〜96番目のアミノ酸残基の領域内に存在することがより好ましい。
あるいは、ピリドキサミン合成酵素は、配列番号3のアミノ酸配列を有するタンパク質又は配列番号3のアミノ酸配列に対して例えば80%以上、又は85%以上、又は90%以上、又は95%以上の配列同一性を有するアミノ酸配列を有するタンパク質であってもよい。
上記のような、配列番号3のアミノ酸配列に類似するアミノ酸配列を有するタンパク質を使用する場合には、該タンパク質はピリドキサミン合成酵素としての活性(ピリドキサールからピリドキサミンを合成する酵素活性、第1の態様においてはピリドキサミン合成酵素活性ともいう)を有しているべきである。ピリドキサールからピリドキサミンを合成する酵素活性は、例えば、基質としてのピリドキサール及び必要なアミノ酸(配列番号3のアミノ酸配列に類似するアミノ酸配列を有するタンパク質の場合は例えばL−アラニン)を含む水溶液に試験対象のタンパク質を加え、生成したピリドキサミンの量を高速液体クロマトグラフィーなどで定量することにより測定できる。
(c) X8X9X10X11X12X13(配列番号99)
(X8は、L、M、I又はVを表し、
X9は、H又はQを表し、
X10は、G、C又はAを表し、
X11は、E又はDを表し、
X12は、P又はAを表し、
X13は、V、I、L又はAを表す)
(d) X14X15TPSGTX16X17(配列番号100)
(X14は、H又はSを表し、
X15は、D又はEを表し、
X16は、I、V又はLを表し、
X17は、N又はTを表す)
(e) X18DX19VSX20X21(配列番号101)
(X18は、V、I又はAを表し、
X19は、A、T又はSを表し、
X20は、S、A又はGを表し、
X21は、F、W、又はVを表す)
(f) X22X23X24KCX25GX26X27P(配列番号102)
(X22は、G又はSを表し、
X23は、P、S又はAを表し、
X24は、N、G、S、A又はQを表し、
X25は、L又はMを表し、
X26は、A、S、C又はGを表し、
X27は、P、T、S又はAを表す)
(g) X28X29X30X31SX32GX33X34(配列番号103)
(X28は、G又はDを表し、
X29は、V又はIを表し、
X30は、V、T、A、S、M、I又はLを表し、
X31は、F、M、L、I又はVを表し、
X32は、S、G、A、T、I、L又はHを表し、
X33は、R、M又はQを表し、
X34は、G、R、A、D、H又はKを表す)
(h) X35X36RX37X38HMGX39X40A(配列番号104)
(X35は、L又はVを表し、
X36は、T、I、V又はLを表し、
X37は、I、V又はLを表し、
X38は、G又はSを表し、
X39は、P、A又はRを表し、
X40は、T、V又はSを表す)
(c−1)X1X2X3X4X5X6X7X8X9X10X11X12X13X14X15X16X17X18X19(配列番号111)
X1が、V、L、I又はMを表し、
X2は、I、L又はVを表し、
X3は、L、M、I又はVを表し、
X4は、H又はQを表し、
X5は、G、C又はAを表し、
X6は、E又はDを表し、
X7は、P又はAを表し、
X8は、V、I、A又はLを表し、
X9は、L、M、P又はVを表し、
X10は、G又はAを表し、
X11は、L又はIを表し、
X12は、E又はQを表し、
X13は、A又はGを表し、
X14は、A又はVを表し、
X15は、A又はLを表し、
X16は、A、L、H又はYを表し、
X17は、S、G又はAを表し、
X18は、L、F、V又はAを表し、
X19は、I、F、V又はLを表す。
部分アミノ酸配列(c−1)は、Mesorhizobium lotiのピリドキサミン−ピルビン酸トランスアミナーゼのN末端から63番目〜81番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(c−1)は、タンパク質のN末端から53番目〜91番目のアミノ酸残基の領域内に存在することが好ましく、N末端から58番目〜86番目のアミノ酸残基の領域内に存在することがより好ましい。
(d−1)X1X2X3X4X5X6X7X8TPSGTX9X10X11X12X13X14 X15X16 (配列番号112)
X1は、V、I、L又はMを表し、
X2は、V又はIを表し、
X3は、S、A、V、C又はFを表し、
X4は、V、I、A、L又はTを表し、
X5は、C又はVを表し、
X6は、H、N又はAを表し、
X7は、H又はSを表し、
X8は、D又はEを表し、
X9は、I、V又はLを表し、
X10は、N又はTを表し、
X11は、P又はDを表し、
X12は、I、V、L又はAを表し、
X13は、D、N、E、A、G、Q、V、R又はPを表し、
X14は、A、E、Q又はDを表し、
X15は、I又はLを表し、
X16は、G又はAを表す。
部分アミノ酸配列(d−1)は、Mesorhizobium lotiのピリドキサミン−ピルビン酸トランスアミナーゼのN末端から138番目〜158番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(d−1)は、タンパク質のN末端から128番目〜168番目のアミノ酸残基の領域内に存在することが好ましく、N末端から133番目〜163番目のアミノ酸残基の領域内に存在することがより好ましい。
(e−1)X1X2X3X4X5X6DX7VSX8X9X10X11X12(配列番号113)
X1は、G、D又はA
X2は、A、G、K、T、Q、R又はEを表し、
X3は、Y、N、L又はFを表し、
X4は、L、F、M又はVを表し、
X5は、I、L又はYを表し、
X6は、V、A又はIを表し、
X7は、A、S又はTを表し、
X8は、S、A又はGを表し、
X9は、F、W又はVを表し、
X10は、G、A又はLを表し、
X11は、G又はSを表し、
X12は、M、V又はLを表す。
部分アミノ酸配列(e−1)は、Mesorhizobium lotiのピリドキサミン−ピルビン酸トランスアミナーゼのN末端から165番目〜179番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(e−1)は、タンパク質のN末端から155番目〜189番目のアミノ酸残基の領域内に存在することが好ましく、N末端から160番目〜184番目のアミノ酸残基の領域内に存在することがより好ましい。
(f−1)X1X2X3X4X5X6X7X8X9KCX10GX11X12PX13X14X15X16X17X18X19S(配列番号114)
X1は、A、S、V又はIを表し、
X2は、D、G又はAを表し、
X3は、I、L、F、V又はMを表し、
X4は、Y、F、L又はCを表し、
X5は、V又はIを表し、
X6は、T又はAを表し、
X7は、G又はSを表し、
X8は、P、S又はAを表し、
X9は、N、G、S、Q又はAを表し、
X10は、L又はMを表し、
X11は、A、S、C又はGを表し、
X12は、P、T、S又はAを表し、
X13は、G、A又はSを表し、
X14は、L又はVを表し、
X15は、T、S又はAを表し、
X16は、M、I、L、V又はFを表し、
X17は、M、L、V、A又はIを表し、
X18は、G、A、H又はSを表し、
X19は、V、I又はAを表す。
部分アミノ酸配列(f−1)は、Mesorhizobium lotiのピリドキサミン−ピルビン酸トランスアミナーゼのN末端から188番目〜211番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(f−1)は、タンパク質のN末端から178番目〜221番目のアミノ酸残基の領域内に存在することが好ましく、N末端から183番目〜216番目のアミノ酸残基の領域内に存在することがより好ましい。
(g−1)X1X2X3X4X5SX6GX7X8(配列番号115)
X1は、Y、F、H又はSを表し、
X2は、G又はDを表し、
X3は、V又はIを表し、
X4は、V、T、A、S、M、I又はLを表し、
X5は、F、M、L、I又はVを表し、
X6は、S、G、A、T、I、L又はHを表し、
X7は、R、M又はQを表し、
X8は、G、R、A、D、H又はKを表す。
部分アミノ酸配列(g−1)は、Mesorhizobium lotiのピリドキサミン−ピルビン酸トランスアミナーゼのN末端から328番目〜337番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(g−1)は、タンパク質のN末端から318番目〜347番目のアミノ酸残基の領域内に存在することが好ましく、N末端から323番目〜342番目のアミノ酸残基の領域内に存在することがより好ましい。
(h−1)X1X2X3X4X5RX6X7HMGX8X9AX10X11(配列番号116)
X1は、L、Q、K、A、F、Y又はWを表し、
X2は、G、N、H又はDを表し、
X3は、K、R又はNを表し、
X4は、L又はVを表し、
X5は、T、I、V又はLを表し、
X6は、I、V又はLを表し、
X7は、G又はSを表し、
X8は、P、A又はRを表し、
X9は、T、V又はSを表し、
X10は、Q、R、E、K、H、Y又はGを表し、
X11は、P又はGを表す。
部分アミノ酸配列(h−1)は、Mesorhizobium lotiのピリドキサミン−ピルビン酸トランスアミナーゼのN末端から340番目〜355番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(h−1)は、タンパク質のN末端から330番目〜365番目のアミノ酸残基の領域内に存在することが好ましく、N末端から335番目〜360番目のアミノ酸残基の領域内に存在することがより好ましい。
あるいは、ピリドキサミン合成酵素は、配列番号4のアミノ酸配列を有するタンパク質又は配列番号4のアミノ酸配列に対して例えば80%以上、又は85%以上、又は90%以上、又は95%以上の配列同一性を有するアミノ酸配列を有するタンパク質であってもよい。
上記のような、配列番号4のアミノ酸配列に類似するアミノ酸配列を有するタンパク質を使用する場合には、該タンパク質はピリドキサミン合成酵素としての活性(ピリドキサールからピリドキサミンを合成する酵素活性)を有しているべきである。ピリドキサールからピリドキサミンを合成する酵素活性は、例えば、基質としてのピリドキサール及び必要なアミノ酸(配列番号4のアミノ酸配列に類似するアミノ酸配列を有するタンパク質の場合は例えばL−グルタミン酸若しくはL−アスパラギン酸又はその塩)を含む水溶液に試験対象のタンパク質を加え、生成したピリドキサミンの量を高速液体クロマトグラフィーなどで定量することにより測定できる。
あるいは、アミノ酸再生酵素は、配列番号5のアミノ酸配列を有するタンパク質又は配列番号5のアミノ酸配列に対して例えば80%以上、又は85%以上、又は90%以上、又は95%以上の配列同一性を有するアミノ酸配列を有するタンパク質であってもよい。
上記のような、配列番号5のアミノ酸配列に類似するアミノ酸配列を有するタンパク質を使用する場合には、該タンパク質はアラニン再生酵素活性を有しているべきである。また、配列番号5のアミノ酸配列のN末端から198番目のAla残基又はこれに配列アラインメント上対応するAla残基を保持していることが好ましい。
上記のような、配列番号5、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、配列番号50、配列番号51及び配列番号52のアミノ酸配列に類似するアミノ酸配列を有するタンパク質を使用する場合には、該タンパク質はアラニン再生酵素活性を有しているべきである。また、配列番号5のアミノ酸配列のN末端から198番目のAla残基又はこれに配列アラインメント上対応するAla残基を保持していることが好ましい。
あるいは、アミノ酸再生酵素は、配列番号6のアミノ酸配列を有するタンパク質又は配列番号6のアミノ酸配列に対して例えば80%以上、又は85%以上、又は90%以上、又は95%以上の配列同一性を有するアミノ酸配列を有するタンパク質であってもよい。
上記のような、配列番号6のアミノ酸配列に類似するアミノ酸配列を有するタンパク質を使用する場合には、該タンパク質はグルタミン酸再生酵素活性を有しているべきである。
(i) EX41KX42X43EX44RX45X46(配列番号105)
(X41は、I、T、S、F、N又はVを表し、
X42は、N、M、A、V、L、T又はDを表し、
X43は、H、N、L又はQを表し、
X44は、Y、N又はFを表し、
X45は、V、又はIを表し、
X46は、G又はAを表す)
(j) X47X48X49KVKEPX50(配列番号106)
(X47は、M又はLを表し、
X48は、I、L又はVを表し、
X49は、V、L、I又はMを表し、
X50は、Q、L、V、N又はIを表す)
(k) LX51TYLHLA(配列番号107)
(X51は、F又はYを表す)
(l) X52DX53AX54DQGG(配列番号108)
(X52は、V又はAを表し、
X53は、V又はIを表し、
X54は、I又はVを表す)
(i−1)GX1PX2EX3KX4X5EX6RX7X8X9X10PX11X12X13X14X15X16(配列番号117)
X1は、V、I、L又はCを表し、
X2は、T、K又はRを表し、
X3は、I、V、T、S、F又はNを表し、
X4は、N、M、A、D、V、L又はTを表し、
X5は、H、N、Q又はLを表し、
X6は、Y、N又はFを表し、
X7は、V又はIを表し、
X8は、G又はAを表し、
X9は、M、L又はIを表し、
X10は、V、T、I又はSを表し、
X11は、S、A、T、Q、H、N、L又はGを表し、
X12は、S、A、N、G又はVを表し、
X13は、V又はAを表し、
X14は、R、K、N、Q、S、A、L又はHを表し、
X15は、E、Q、D、V又はAを表し、
X16は、L、A、V、F又はYを表す。
部分アミノ酸配列(i−1)は、Shewanella sp. AC10のアラニンデヒドロゲナーゼのN末端から4番目〜26番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(i−1)は、タンパク質のN末端から2番目〜36番目のアミノ酸残基の領域内に存在することが好ましく、N末端から3番目〜31番目のアミノ酸残基の領域内に存在することがより好ましい。
(j−1)X1X2X3X4KVKEPX5X6X7EX8X9X10(配列番号118)
X1は、D、E、Q又はKを表し、
X2は、M又はLを表し、
X3は、I、L又はVを表し、
X4は、V、L、I又はMを表し、
X5は、Q、L、I、V又はNを表し、
X6は、A、T、S、P、M、K、R、Q、V、I又はEを表し、
X7は、V、I、E、N、Q、T、A、D、H、M、N、V、A、S、I、D、G、W又はKを表し、
X8は、R、C、Y又はWを表し、
X9は、A、E、R、Q、K、T、S、M、N、V、P、G、C又はHを表し、
X10は、M、L、K、R、Q、E、W、F又はYを表す。
部分アミノ酸配列(j−1)は、Shewanella sp. AC10のアラニンデヒドロゲナーゼのN末端から69番目〜84番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(j−1)は、タンパク質のN末端から59番目〜94番目のアミノ酸残基の領域内に存在することが好ましく、N末端から64番目〜89番目のアミノ酸残基の領域内に存在することがより好ましい。
(k−1)X1X2X3X4X5X6LX7TYLHLAX8X9X10X11X12X13X14X15LX16X17X18X19(配列番号119)
X1は、L又はFを表し、
X2は、R、K、C、S、H、G又はQを表し、
X3は、H、E、P、S、D、R、K、Q又はAを表し、
X4は、D、G、Q、H、E、S、N又はMを表し、
X5は、Q又はHを表し、
X6は、I、L、V、T、C又はAを表し、
X7は、F又はYを表し、
X8は、P又はAを表し、
X9は、D、S、N、H又はEを表し、
X10は、L、M、P、R、V、Q、E又はKを表し、
X11は、P、A、V、Q、K、E、D、T、N、R、S又はMを表し、
X12は、Q、C又はLを表し、
X13は、T、A又はVを表し、
X14は、E、I、Q、A、R、K、T、D又はNを表し、
X15は、E、D、L、A、G、H、Y又はSを表し、
X16は、I、M、V、L、T又はKを表し、
X17は、T、K、S、D、H、E、A、N、R、G又はQを表し、
X18は、S、G、C、A又はKを表し、
X19は、G、K、R、Q又はDを表す。
部分アミノ酸配列(k−1)は、Shewanella sp. AC10のアラニンデヒドロゲナーゼのN末端から85番目〜111番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(k−1)は、タンパク質のN末端から75番目〜121番目のアミノ酸残基の領域内に存在することが好ましく、N末端から80番目〜116番目のアミノ酸残基の領域内に存在することがより好ましい。
(l−1)X1X2X3X4X5DX6AX7DQGGX8X9X10X11(配列番号120)
X1は、G、R又はSを表し、
X2は、S、A又はGを表し、
X3は、A又はVを表し、
X4は、I、L、M又はVを表し、
X5は、V又はAを表し、
X6は、V又はIを表し、
X7は、I又はVを表し、
X8は、C又はIを表し、
X9は、V、I、A、F、S又はCを表し、
X10は、E又はAを表し、
X11は、T又はDを表す。
部分アミノ酸配列(l−1)は、Shewanella sp. AC10のアラニンデヒドロゲナーゼのN末端から261番目〜277番目のアミノ酸残基にアラインメント上対応するアミノ酸残基に相当する。部分アミノ酸配列(l−1)は、タンパク質のN末端から251番目〜287番目のアミノ酸残基の領域内に存在することが好ましく、N末端から256番目〜282番目のアミノ酸残基の領域内に存在することがより好ましい。
ピリドキシンデヒドロゲナーゼをコードする遺伝子は、上述のピリドキシンデヒドロゲナーゼをコードする任意の遺伝子であってよい。ピリドキサミン合成酵素をコードする遺伝子は、上述のピリドキサミン合成酵素をコードする任意の遺伝子であってよい。アミノ酸再生酵素をコードする遺伝子は、上述のアミノ酸再生酵素をコードする任意の遺伝子であってよい。これらの遺伝子がコードする酵素は、当該酵素活性を有する既知のアミノ酸配列(例えば、生物中に天然に存在する遺伝子によってコードされるアミノ酸配列)に限られず、前記既知のアミノ酸配列とは異なる改変されたアミノ酸配列を有する酵素であってもよい。
ヌクレオチド配列を改変する方法としては、例えば、部位特異的変異法(Kramer,W. and frita,H.J., Methods in Enzymology,vol.154,P.350(1987))、リコンビナントPCR法(PCR Technology,Stockton Press(1989)、特定の部分のDNAを化学合成する方法、遺伝子をヒドロキシアミン処理する方法、遺伝子を保有する菌株を紫外線照射処理、又は、ニトロソグアニジンや亜硝酸などの化学薬剤で処理する方法、市販の突然変異導入キットを使用する方法などが挙げられる。
ハイブリダイゼーションでは、基準となるヌクレオチド配列と相補的なヌクレオチド配列又はその部分配列からなるDNAをプローブとして、対象とするDNAに対してハイブリダイゼーションを行い、ストリンジェントな条件下で洗浄後にプローブが対象とする核酸に有意にハイブリダイズしているかを確認する。プローブの長さは例えば連続した20ヌクレオチド以上、好ましくは50ヌクレオチド以上、さらに好ましくは100ヌクレオチド以上、さらに好ましくは200ヌクレオチド以上を用いることができる。基準となるヌクレオチド配列と同じヌクレオチド長を有し、全長に渡って相補的なDNAをプローブとして用いることも好ましい。ハイブリダイゼーションのための条件とは、特定のハイブリダイゼーションシグナルを検出するために当業者が一般的に用いている条件を例示できる。好ましくは、ストリンジェントなハイブリダイゼーション条件とストリンジェントな洗浄条件を意味する。例えば、6×SSC(saline sodium citrate)(1×SSCの組成:0.15M NaCl、0.015Mクエン酸ナトリウム、pH7.0)、0.5%SDS、5×デンハート及び100mg/mlニシン精子DNAを含む溶液中でプローブとともに55℃で一晩保温するという条件等が挙げられる。ついでフィルターを0.2×SSC中42℃で洗浄するなどを例示することができる。ストリンジェントな条件としては、フィルターの洗浄工程における0.1×SSC、50℃の条件であり、更にストリンジェントな条件としては、同工程における0.1×SSC、65℃の条件を挙げることができる。
あるいは、ピリドキシンデヒドロゲナーゼをコードする遺伝子は、例えば、配列番号7のヌクレオチド配列を有するDNAであってもよいし、配列番号7のヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、ピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、ピリドキシンデヒドロゲナーゼは、配列番号7のヌクレオチド配列を有するDNA又は配列番号7のヌクレオチド配列に対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、ピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、ピリドキシンデヒドロゲナーゼをコードする遺伝子は、例えば、配列番号8のヌクレオチド配列を有するDNAであってもよいし、配列番号8のヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、ピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、ピリドキシンデヒドロゲナーゼは、配列番号8のヌクレオチド配列を有するDNA又は配列番号8のヌクレオチド配列に対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、ピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、ピリドキシンデヒドロゲナーゼをコードする遺伝子は、例えば、配列番号7及び配列番号53〜配列番号59のうちいずれかのヌクレオチド配列を有するDNAであってもよいし、配列番号7及び配列番号53〜配列番号59のうちいずれかのヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、ピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、ピリドキシンデヒドロゲナーゼは、配列番号7及び配列番号53〜配列番号59のうちいずれかのヌクレオチド配列を有するDNA、又は配列番号7のヌクレオチド配列、配列番号53のヌクレオチド配列(Saccharomyces eubayanusのピリドキシンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号54のヌクレオチド配列(Torulaspora delbrueckiiのピリドキシンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号55のヌクレオチド配列(Zygosaccharomyces bailiiのピリドキシンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号57のヌクレオチド配列(Kluyveromyces marxianusのピリドキシンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号56のヌクレオチド配列(Aspergillus oryzaeのピリドキシンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号58のヌクレオチド配列(Candida albicansのピリドキシンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、及び配列番号59のヌクレオチド配列(Yarrowia lipolyticaのピリドキシンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)のうち少なくとも1つに対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、ピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、ピリドキシンデヒドロゲナーゼをコードする遺伝子は、例えば、配列番号13及び配列番号75〜配列番号81のうちいずれかのヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域を有するDNAであってもよいし、配列番号13及び配列番号75〜配列番号81のうちいずれかのヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、ピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、ピリドキシンデヒドロゲナーゼは、配列番号13及び配列番号75〜配列番号81のうちいずれかのヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域を有するDNA、又は配列番号13のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Saccharomyces cerevisiaeのピリドキシンデヒドロゲナーゼをコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号75のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Saccharomyces eubayanusのピリドキシンデヒドロゲナーゼをコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号76のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Torulaspora delbrueckiiのピリドキシンデヒドロゲナーゼをコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号77のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Zygosaccharomyces bailiiのピリドキシンデヒドロゲナーゼをコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号79のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Kluyveromyces marxianusのピリドキシンデヒドロゲナーゼをコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号78のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Aspergillus oryzaeのピリドキシンデヒドロゲナーゼをコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号80のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Candida albicansのピリドキシンデヒドロゲナーゼをコードする遺伝子のコドン最適化ヌクレオチド配列)、及び配列番号81のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Yarrowia lipolyticaのピリドキシンデヒドロゲナーゼをコードする遺伝子のコドン最適化ヌクレオチド配列)のうち少なくとも1つに対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、ピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、ピリドキサミン合成酵素をコードする遺伝子は、例えば、配列番号9のヌクレオチド配列を有するDNAであってもよいし、配列番号9のヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、ピリドキサミン合成酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、ピリドキサミン合成酵素は、配列番号9のヌクレオチド配列を有するDNA又は配列番号9のヌクレオチド配列に対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、ピリドキサミン合成酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、ピリドキサミン合成酵素をコードする遺伝子は、例えば、配列番号10のヌクレオチド配列を有するDNAであってもよいし、配列番号10のヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、ピリドキサミン合成酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、ピリドキサミン合成酵素は、配列番号10のヌクレオチド配列を有するDNA又は配列番号10のヌクレオチド配列に対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、ピリドキサミン合成酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、ピリドキサミン合成酵素をコードする遺伝子は、例えば、配列番号9及び配列番号60〜配列番号66のうちいずれかのヌクレオチド配列を有するDNAであってもよいし、配列番号9及び配列番号60〜配列番号66のうちいずれかのヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、ピリドキサミン合成酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、ピリドキサミン合成酵素は、配列番号9及び配列番号60〜配列番号66のうちいずれかのヌクレオチド配列を有するDNA、又は配列番号9のヌクレオチド配列、配列番号60(Mesorhizobium sp. YR577のピリドキサミン−ピルビン酸トランスアミナーゼをコードする遺伝子のヌクレオチド配列)のヌクレオチド配列、配列番号61(Pseudaminobacter salicylatoxidansのピリドキサミン−ピルビン酸トランスアミナーゼをコードする遺伝子のヌクレオチド配列)のヌクレオチド配列、配列番号62(Bauldia litoralisのピリドキサミン−ピルビン酸トランスアミナーゼをコードする遺伝子のヌクレオチド配列)のヌクレオチド配列、配列番号63(Skermanella stibiiresistensのピリドキサミン−ピルビン酸トランスアミナーゼをコードする遺伝子のヌクレオチド配列)のヌクレオチド配列、配列番号64(Rhizobium sp. AC44/96のピリドキサミン−ピルビン酸トランスアミナーゼをコードする遺伝子のヌクレオチド配列)のヌクレオチド配列、配列番号65(Erwinia toletanaのピリドキサミン−ピルビン酸トランスアミナーゼをコードする遺伝子のヌクレオチド配列)のヌクレオチド配列、及び配列番号66(Herbiconiux ginsengiのピリドキサミン−ピルビン酸トランスアミナーゼをコードする遺伝子のヌクレオチド配列)のヌクレオチド配列のうち少なくとも1つに対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、ピリドキサミン合成酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、ピリドキサミン合成酵素をコードする遺伝子は、例えば、配列番号14のヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域又は配列番号82〜配列番号88のうちいずれかのヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域を有するDNAであってもよいし、配列番号14のヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域又は配列番号82〜配列番号88のうちいずれかのヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、ピリドキサミン合成酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、アミノ酸再生酵素をコードする遺伝子は、例えば、配列番号11のヌクレオチド配列を有するDNAであってもよいし、配列番号11のヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、アラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、アミノ酸再生酵素は、配列番号11のヌクレオチド配列を有するDNA又は配列番号11のヌクレオチド配列に対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、アラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、アミノ酸再生酵素をコードする遺伝子は、例えば、配列番号12のヌクレオチド配列を有するDNAであってもよいし、配列番号12のヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、グルタミン酸再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、アミノ酸再生酵素は、配列番号12のヌクレオチド配列を有するDNA又は配列番号12のヌクレオチド配列に対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、グルタミン酸再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、アミノ酸再生酵素をコードする遺伝子は、例えば、配列番号11及び配列番号67〜配列番号74のうちいずれかのヌクレオチド配列を有するDNAであってもよいし、配列番号11及び配列番号67〜配列番号74のうちいずれかのヌクレオチド配列に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、アラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、アミノ酸再生酵素は、配列番号11及び配列番号67〜配列番号74のうちいずれかのヌクレオチド配列を有するDNA、又は配列番号11のヌクレオチド配列、配列番号67のヌクレオチド配列(Aeromonas hydrophilaのアラニンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号68のヌクレオチド配列(Rhizobium sp. LPU83のアラニンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号69のヌクレオチド配列(Pseudomonas mendocinaのアラニンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号70のヌクレオチド配列(Bradyrhizobium japonicumのアラニンデヒドロゲナーゼのうちの一つをコードする遺伝子のヌクレオチド配列)、配列番号71のヌクレオチド配列(Bradyrhizobium japonicumのアラニンデヒドロゲナーゼのうちの別の一つをコードする遺伝子のヌクレオチド配列)、配列番号72のヌクレオチド配列(Streptomyces aureofaciensのアラニンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、配列番号73のヌクレオチド配列(Anabaena cylindricaのアラニンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)、及び配列番号74のヌクレオチド配列(Bacillus subtilisのアラニンデヒドロゲナーゼをコードする遺伝子のヌクレオチド配列)のうち少なくとも1つに対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、アラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
あるいは、アミノ酸再生酵素をコードする遺伝子は、例えば、配列番号15及び配列番号89〜配列番号96のうちいずれかのヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域を有するDNAであってもよいし、配列番号15及び配列番号89〜配列番号96のうちいずれかのヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域に対してヌクレオチドの置換、欠失、挿入及びヌクレオチド配列の5’末端若しくは3’末端若しくはその両方への追加のヌクレオチドの付加のうち1つ以上を行ったヌクレオチド配列であって、アラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。ヌクレオチドの置換、欠失、挿入及びヌクレオチド配列のN末端若しくはC末端若しくはその両方への追加のヌクレオチドの付加の程度の例については、上記のとおりである。
あるいは、アミノ酸再生酵素は、配列番号15及び配列番号89〜配列番号96のうちいずれかのヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域を有するDNA、又は配列番号15のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Shewanella sp. AC10のアラニンデヒドロゲナーゼのアミノ酸配列に対してD198Aのアミノ酸置換を行ったアミノ酸配列をコードするコドン最適化ヌクレオチド配列)、配列番号89のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Aeromonas hydrophilaのアラニンデヒドロゲナーゼにNADP+対応アミノ酸置換を行ったアミノ酸配列をコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号90のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Rhizobium sp. LPU83のアラニンデヒドロゲナーゼにNADP+対応アミノ酸置換を行ったアミノ酸配列をコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号91のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Pseudomonas mendocinaのアラニンデヒドロゲナーゼにNADP+対応アミノ酸置換を行ったアミノ酸配列をコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号92のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Bradyrhizobium japonicumのアラニンデヒドロゲナーゼのうちの一つにNADP+対応アミノ酸置換を行ったアミノ酸配列をコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号93のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Bradyrhizobium japonicumのアラニンデヒドロゲナーゼのうちの別の一つにNADP+対応アミノ酸置換を行ったアミノ酸配列をコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号94のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Streptomyces aureofaciensのアラニンデヒドロゲナーゼにNADP+対応アミノ酸置換を行ったアミノ酸配列をコードする遺伝子のコドン最適化ヌクレオチド配列)、配列番号95のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Anabaena cylindricaのアラニンデヒドロゲナーゼにNADP+対応アミノ酸置換を行ったアミノ酸配列をコードする遺伝子のコドン最適化ヌクレオチド配列)、及び配列番号96のヌクレオチド配列の18番目のヌクレオチド〜3’末端までの領域(Bacillus subtilisのアラニンデヒドロゲナーゼにNADP+対応アミノ酸置換を行ったアミノ酸配列をコードする遺伝子のコドン最適化ヌクレオチド配列)のうち少なくとも1つに対して例えば80%以上、若しくは85%以上、若しくは90%以上、若しくは95%以上の配列同一性を有するヌクレオチド配列であって、アラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有するDNAであってもよい。
本開示に係る組換え微生物内において、ピリドキシンデヒドロゲナーゼをコードする遺伝子、ピリドキサミン合成酵素をコードする遺伝子、及び該ピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有するアミノ酸再生酵素をコードする遺伝子の各々は、菌体に内在する遺伝子であってもよいし、菌体に内在するがその発現を強化した遺伝子であってもよいし、菌体外から菌体内へと導入された遺伝子であってもよい。ただし、ピリドキシンデヒドロゲナーゼをコードする遺伝子、ピリドキサミン合成酵素をコードする遺伝子、及び該ピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有するアミノ酸再生酵素をコードする遺伝子のうち少なくとも2つは菌体外から導入された遺伝子であるか又は菌体に内在するがその発現が強化された遺伝子である。
言い換えると、ピリドキシンデヒドロゲナーゼをコードする遺伝子、ピリドキサミン合成酵素をコードする遺伝子、及び該ピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有するアミノ酸再生酵素をコードする遺伝子の各々は、組換え前の宿主微生物のゲノムに内在する遺伝子であってもよいし、宿主微生物のゲノムに内在するがプロモーターの置換等の操作を行うことによってその発現を強化した遺伝子であってもよいし、プラスミド等のベクターを用いて菌体外から菌体内へと導入された遺伝子であってもよい。第1の態様においては、前記3種類の酵素の組み合わせによるピリドキサミン生産能力を発揮すると共に、ピリドキシンデヒドロゲナーゼをコードする遺伝子、ピリドキサミン合成酵素をコードする遺伝子、及び該ピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有するアミノ酸再生酵素をコードする遺伝子のうち少なくとも2つについて、それぞれ、宿主微生物の細胞外から導入するか、あるいはプロモーターの置換等によって宿主微生物の細胞に内在する当該遺伝子の発現を強化することによって、当該遺伝子の発現を上昇させ、前記3種類の酵素の組み合わせによるピリドキサミン生産能力をさらに増強している。ここで、ピリドキシンデヒドロゲナーゼをコードする遺伝子、ピリドキサミン合成酵素をコードする遺伝子、及び該ピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有するアミノ酸再生酵素をコードする遺伝子の各々については、導入又は発現強化を行う場合には、宿主微生物の細胞外からの導入と、プロモーターの置換等による当該遺伝子発現の強化のうち、一方のみを行ってもよいし、両方を行ってもよい。
宿主微生物が例えば原核生物である場合には、新たに導入するプロモーターとして使用可能なプロモーターの例としては、大腸菌由来のtrpプロモーター、lacプロモーター、GAPDHプロモーター、ラムダファージ由来のPLプロモーター及びPRプロモーターや、枯草菌由来のグルコン酸合成酵素プロモーター(gnt)、アルカリプロテアーゼプロモーター(apr)、中性プロテアーゼプロモーター(npr)、α−アミラーゼプロモーター(amy)等が挙げられる。また、tacプロモーターのように独自に改変又は設計されたプロモーター配列も利用できる。
また、前記組換え微生物の選択を容易にするという観点より、選択マーカーとなりうる選択遺伝子をコードするヌクレオチド配列をさらに含んでいてもよい。
所望の酵素を産生するために必要となる制御領域としては、プロモーター配列(転写を制御するオペレーター配列を含む。)、リボゾーム結合配列(SD配列)、転写終結配列等を挙げることができる。
分泌シグナルとしては、宿主微生物となる酵母から所望の酵素を分泌できるものであれば、特に限定されない。分泌効率の観点より、αファクターシグナル配列、インベルターゼシグナル配列、酸ホスファターシグナル配列、グルコアミラーゼシグナル配列などを用いることが好ましい。
また、pUC18、pBR322,pUC100、pSL1180(ファルマシア インク社製)、pFB6及びアスペルギルス(Aspergillus)pRAX、トリコデルマ(Trichoderma)pTEX等のような一般的に用いられる他の発現ベクターを用いることもできる。
前記リボゾーム結合配列としては、例えば、16SリボゾームRNAの3’末端領域に相補的な配列のうち、4ヌクレオチド以上連続したコンセンサス配列をDNA合成により作成した配列などが挙げられる。
転写終結配列は必ずしも必要ではないが、ρ因子非依存性のもの、例えばリポプロテインターミネーター、trpオペロンターミネーター等が利用できる。
これら制御領域の発現ベクター上での配列順序は、特に制限されるものではないが、転写効率を考慮すると5’末端側上流からプロモーター配列、リボゾーム結合配列、目的酵素をコードする遺伝子、転写終結配列の順に並ぶことが望ましい。
また、2種類以上の宿主微生物内での自律複製が可能な発現ベクターの例として、pHV14、TRp7、YEp7及びpBS7等を発現ベクターとして利用することができる。
また、これらの酵素をコードするDNAは、組換え微生物から取り出すことができ、他の微生物に移入させることも可能である。また、このDNAを鋳型として用いて、PCRにより酵素をコードするDNA断片を増幅し、制限酵素等で処理した後、他のベクターDNA断片と結合させ、宿主微生物に新たに導入することも容易に実施できる。
一実施形態においては、ピリドキサミン又はその塩の製造方法は、第1の態様に係る組換え微生物、前記組換え微生物の培養物又は前記組換え微生物若しくは前記培養物の処理物と、ピリドキシン又はその塩とを接触させて、ピリドキサミン又はその塩を生産させることを含む。
ピリドキシンの塩の例としては、ピリドキシンと酸との塩が挙げられる。酸の例としては、塩酸、硫酸、硝酸、リン酸などが挙げられる。ピリドキシンの塩は、例えばピリドキシン塩酸塩である。
なお、このような組換え微生物、前記組換え微生物の培養物又は前記組換え微生物若しくは前記培養物の処理物を用いるピリドキサミン又はその塩の製造方法のことを、単に「組換え微生物を用いたピリドキサミン又はその塩の製造方法」とも称する。
培地に使用する成分としては、公知のものを用いることができる。例えば、肉エキス、酵母エキス、麦芽エキス、ペプトン、NZアミン及びジャガイモ等の有機栄養源、グルコース、マルトース、しょ糖、デンプン及び有機酸等の炭素源、硫酸アンモニウム、尿素及び塩化アンモニウム等の窒素源、リン酸塩、マグネシウム、カリウム及び鉄等の無機栄養源、ビタミン類を適宜組み合わせて使用できる。
なお、前記選択マーカーを含む発現ベクターにより形質転換した組換え微生物の培養においては、例えば、前記選択マーカーが薬剤耐性である場合には、それに対応する薬剤を含む培地を使用し、前記選択マーカーが栄養要求性である場合には、それに対応する栄養素を含まない培地を使用する。
培地のpHは、例えば4〜8の範囲で選べばよく、5〜8の範囲であってもよい。
培養温度は例えば20℃〜45℃であり、好ましくは24℃〜37℃である。培養は、微生物の種類に応じて、好気的に行ってもよいし、嫌気的に行ってもよい。
培養期間は例えば1日間〜7日間である。培養期間は、目的の酵素の産生量が最大になるように設定してもよい。
接触は基質としてのピリドキシン又はその塩を含む溶液中で行うことが好ましい。溶液のpHは、ピリドキシンデヒドロゲナーゼ、ピリドキサミン合成酵素、及びアミノ酸再生酵素の酵素活性が維持される限りは特に制限されないが、6.0〜9.0であることが好ましく、7.0〜8.5であることがより好ましい。溶液の温度も、溶液のpHは、ピリドキシンデヒドロゲナーゼ、ピリドキサミン合成酵素、及びアミノ酸再生酵素の酵素活性が維持される限りは特に制限されないが、20〜70℃であることが好ましく、25℃〜50℃であることがより好ましい。
溶液の媒体としては、水若しくは水性媒体、有機溶媒又は水若しくは水性媒体と有機溶媒の混合液が用いられる。水性媒体としては、例えばリン酸緩衝液、HEPES(N−2−ヒドロキシエチルピペラジン−N−エタンスルホン酸)緩衝液、トリス[トリス(ヒドロキシメチル)アミノメタン]塩酸緩衝液等の緩衝液が用いられる。有機溶媒としては反応を阻害しないものであればいずれでもよく、例えばアセトン、酢酸エチル、ジメチルスルホキシド、キシレン、メタノール、エタノール、ブタノール等が用いられる。
ピリドキシンを基質としてピリドキサールを選択的かつ高効率に生産する組換え微生物は、これまで知られていなかった。
ピリドキシン及びピリドキサールの構造を以下に示す。なお、本開示において、ピリドキシン及びピリドキサールの語は、特に塩を除外する旨が明示されない限り、それぞれその塩も含む意味で使用される。ピリドキシンの塩は、例えば、ピリドキシン塩酸塩、ピリドキシン硫酸塩、ピリドキシン硝酸塩、ピリドキシンリン酸塩を含む。ピリドキサールの塩は、例えば、ピリドキサール塩酸塩、ピリドキサール硫酸塩、ピリドキサール硝酸塩、ピリドキサールリン酸塩を含む。
[配列番号1のアミノ酸配列]
配列番号1のアミノ酸配列は、Saccharomyces cerevisiaeに由来するピリドキシンデヒドロゲナーゼである。ピリドキシンデヒドロゲナーゼは、以下に示す通り、ピリドキシンとNADP+とから、ピリドキサールとNADPHとH+とを生成する反応、及びその逆方向の反応を触媒する酸化還元酵素である。Saccharomyces cerevisiaeに由来するピリドキシンデヒドロゲナーゼは、国際生化学連合(I.U.B.)酵素委員会報告に準拠した酵素番号EC1.1.1.65に分類される。
[(1)配列番号1のアミノ酸配列からなるタンパク質をコードするポリヌクレオチド]
第2の態様に係る組換え微生物において宿主微生物に導入されるポリヌクレオチドは、配列番号1のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドであることが好ましい。配列番号1のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドであれば、いかなるコドンが使用されていてもよい。好ましくは、配列番号13のヌクレオチド配列からなるポリヌクレオチドが用いられる。
第2の態様に係る組換え微生物において宿主微生物に導入されるポリヌクレオチドは、配列番号1のアミノ酸配列において、1個〜50個のアミノ酸が欠失、付加又は置換されているアミノ酸配列からなるタンパク質をコードするポリヌクレオチドでもよい。前記タンパク質はピリドキシン又はピリドキサールの合成活性を有する。好ましくは、EC1.1.1.65に分類される酵素活性を有している。
ピリドキサール又はピリドキシンの検出は、公知の方法、例えば、高速液体クロマトグラフィー(HPLC)によって行うことができる。
第2の態様に係る組換え微生物において宿主微生物に導入されるポリヌクレオチドは、配列番号1のアミノ酸配列と少なくとも60%の配列同一性を有し、ピリドキシン又はピリドキサールの合成活性を有するタンパク質をコードするポリヌクレオチドでもよい。前記タンパク質は、好ましくは、EC1.1.1.65に分類される酵素活性を有している。
配列同一性は、少なくとも70%であることが好ましく、少なくとも80%又は少なくとも85%以上であることがさらに好ましく、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%又は100%であることが特に好ましい。
第2の態様に係る組換え微生物において宿主微生物に導入されるポリヌクレオチドは、配列番号13のヌクレオチド配列からなるポリヌクレオチドであることが好ましい。
配列番号13のヌクレオチド配列からなるポリヌクレオチドは、配列番号1のアミノ酸配列からなるタンパク質であるSaccharomyces cerevisiaeのピリドキシンデヒドロゲナーゼをコードしている。
第2の態様に係る組換え微生物において宿主微生物に導入されるポリヌクレオチドは、配列番号13のヌクレオチド配列において、1個〜150個のヌクレオチドが欠失、付加又は置換されているヌクレオチド配列からなるポリヌクレオチドでもよい。前記ポリヌクレオチドがコードするタンパク質は、ピリドキシン又はピリドキサールの合成活性を有しており、好ましくは、EC1.1.1.65に分類される酵素活性を有している。
第2の態様に係る組換え微生物において宿主微生物に導入されるポリヌクレオチドは、配列番号13のヌクレオチド配列と60%以上の配列同一性を有するヌクレオチド配列からなるポリヌクレオチドでもよい。前記ポリヌクレオチドがコードするタンパク質は、ピリドキシン又はピリドキサールの合成活性を有しており、前記タンパク質は、好ましくは、EC1.1.1.65に分類される酵素活性を有している。
配列同一性は、少なくとも70%であることが好ましく、少なくとも80%又は少なくとも85%以上であることがさらに好ましく、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%又は100%であることが特に好ましい。
第2の態様に係る組換え微生物において宿主微生物に導入されるポリヌクレオチドは、配列番号13のヌクレオチド配列と相補的なヌクレオチド配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドでもよい。前記ポリヌクレオチドがコードするタンパク質は、ピリドキシン又はピリドキサールの合成活性を有しており、前記タンパク質は、好ましくは、EC1.1.1.65に分類される酵素活性を有している。
ここで、ストリンジェントな条件とは、特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。ストリンジェントな条件は、Molecular Cloning 3rd(J.Sambrook et al.,Cold Spring Harbor Lab.Press,2001)の記載に基づいて、比較する配列に応じた条件を適宜設定することができる。この文献は、参照により本明細書に組み入れられるものとする。
典型的なストリンジェントな条件は、例えば、ストリンジェントなハイブリダイゼーション条件とストリンジェントな洗浄条件を意味する。例えば、6×SSC(1×SSCの組成:0.15M NaCl、0.015Mクエン酸ナトリウム、pH7.0)、0.5%SDS、5×デンハート及び100mg/mlニシン精子DNAを含む溶液中でプローブとともに55℃で一晩保温するという条件等が挙げられる。ついでフィルターを0.2×SSC中42℃で洗浄するなどを例示することができる。ストリンジェントな条件としては、フィルターの洗浄工程における0.1×SSC、50℃の条件であり、更にストリンジェントな条件としては、同工程における0.1×SSC、65℃の条件を挙げることができる。
また、配列番号13のポリヌクレオチド配列との配列同一性は、比較対象とされるポリヌクレオチド配列と最適なアライメントを行った場合に、同一であるヌクレオチドの数を配列番号13のポリヌクレオチドのヌクレオチド数である1055で除した値に100を乗じて%で表したものである。
最適なアラインメントとは、2つの配列間で一致するアミノ酸又はヌクレオチドの数が最も大きくなるアラインメントを指す。配列同士の最適なアラインメントは、公知の方法、例えば、BLAST(登録商標、National Library of Medicine)プログラムのデフォルトパラメータにおいてマスクフィルタリングをオフにして使用することによって行うことができる。
第2の態様に係る組換え微生物が、導入された遺伝子からピリドキシンを基質としてピリドキサールを合成する能力を有するタンパク質、又はピリドキサールを基質としてピリドキシンを合成する能力を有するタンパク質を発現していることは、公知の方法で確認すればよい。遺伝子を導入された組換え微生物を、基質としてのピリドキシン又はピリドキサールと接触させて酵素反応の結果物としてのピリドキシン又はピリドキサールの生成を検出することによって、前記タンパク質の発現を確認することができる。ピリドキシン又はピリドキサール合成の前後に別の反応を行う組換え微生物である場合には、当該前後の反応を勘案して基質としてのピリドキシン又はピリドキサールから、酵素反応の結果物としてのピリドキシン又はピリドキサールの生成がなされていることを理論的に確認することができる。
[宿主]
第2の態様に係る組換え微生物を調製する際に宿主となる微生物は、一般的にタンパク質の発現に用いられる微生物を宿主として使用することができる。宿主微生物の例としては、酵母、糸状菌、及び原核生物が挙げられる。酵母の例としては、Saccharomyces cerevisiaeなどのサッカロミセス(Saccharomyces)属の酵母、Schizosaccharomyces pombe等のシゾサッカロミセス(Schizosaccharomyces)属の酵母、ハンゼヌラ(Hansenula)属の酵母、及びピキア(Pichia)属の酵母、が挙げられる。Saccharomyces cerevisiaeを宿主とした場合には、内在性のピリドキシンデヒドロゲナーゼに加えて、前述の(1)〜(7)のいずれか1つのポリヌクレオチドを導入することによって、ピリドキサールを選択的に効率的に生産することができる微生物を得ることができる。
糸状菌の例としては、Trichoderma reesei若しくはviride等のトリコデルマ(Trichoderma)属糸状菌、Aspergillus niger若しくはoryzae等のアスペルギルス(Aspergillus)属糸状菌、Humicola insolens等のフミコラ(Humicola)属糸状菌、及びAcremonium cellulolyticus若しくはfusidioides等のアクレモニウム(Acremonium)属の糸状菌、が挙げられる。
細菌の例としては、エシェリヒア属(Escherichia)細菌、バチルス属(Bacillus)細菌、コリネバクテリウム(Corynebacterium)属細菌、ロドコッカス(Rhodococcus)属等が例示される。工業的な使用の実績が豊富な大腸菌(Escherichia coli)が特に好適に使用される。大腸菌以外の宿主としては、例えば、ブレビバチルス(Bacillus brevis)、Bacillus megaterium、Brevibacillus choshinensis、Corynebacterium glutamicum及びRhodococcus erythropolisが挙げられる。
ポリヌクレオチドを宿主に導入する方法は公知である。例えば「Molecular Cloning 3rd Edition」(J.Sambrookら;Cold Spring Harbor Laboratory Press,2001)等に記載されている分子生物学、生物工学、及び遺伝子工学の分野において公知の一般的な方法によって、目的のポリヌクレオチドを宿主に導入することができる。例えば、前記(1)〜(6)に記載のポリヌクレオチドを含む発現ベクターを用意し、この発現ベクターにより任意の宿主微生物を形質転換して組換え微生物を得ることができる。
宿主が最近である場合、宿主細菌の細胞に発現ベクター等の組換えDNAを導入する方法としては、カルシウム処理によるコンピテントセル法やエレクトロポレーション法などが用いることができる。このようにして得られた組換え細菌は、培養されることにより、多量のピリドキシンデヒドロゲナーゼを安定的に発現させることができる。
また、tacプロモーターのように独自に改変又は設計されたプロモーター配列も利用できる。
前記リボゾーム結合配列としては、例えば、16SリボゾームRNAの3’末端領域に相補的な配列のうち、4ヌクレオチド以上連続したコンセンサス配列をDNA合成により作成した配列などが挙げられる。
転写終結配列は必ずしも必要ではないが、ρ因子非依存性のもの、例えばリポプロテインターミネーター、trpオペロンターミネーター等が利用できる。
これら制御領域の発現ベクター上での配列順序は、特に制限されるものではないが、転写効率を考慮すると5’末端側上流からプロモーター配列、リボゾーム結合配列、目的蛋白質をコードする遺伝子、転写終結配列の順に並ぶことが望ましい。
また、2種類以上の宿主内での自律複製が可能な発現ベクターの例として、pHV14、TRp7、YEp7及びpBS7等を発現ベクターとして利用することができる。
第2の態様に係るピリドキサールの製造方法は、第2の態様に係る組換え微生物とピリドキシンとを接触させることを含む。上述した発現ベクターを所望の宿主微生物に導入して得られた組換え微生物は、前記発現ベクターが含むポリヌクレオチドがコードするタンパク質を生産する。生産されたタンパク質は、ピリドキシンを基質としてピリドキサールを生成する。
培地としては炭素源、窒素源、無機物及びその他の栄養素を適量含有する培地ならば合成培地又は天然培地のいずれでも使用可能である。培地に使用する成分としては、公知のものを用いることができる。例えば、肉エキス、酵母エキス、麦芽エキス、ペプトン、NZアミン及びジャガイモ等の有機栄養源、グルコース、マルトース、しょ糖、デンプン及び有機酸等の炭素源、硫酸アンモニウム、尿素及び塩化アンモニウム等の窒素源、リン酸塩、マグネシウム、カリウム及び鉄等の無機栄養源、ビタミン類を適宜組み合わせて使用できる。
なお、前記選択マーカーを含む発現ベクターにより形質転換した組換え微生物の培養においては、例えば、前記選択マーカーが薬剤耐性である場合には、それに対応する薬剤を含む培地を使用し、前記選択マーカーが栄養要求性である場合には、それに対応する栄養素を含まない培地を使用する。培地のpHは、pH4〜pH8の範囲で選べばよい。
培養は前記培地を含有する液体培地中で、組換え微生物を振とう培養、通気攪拌培養、連続培養、流加培養などの通常の培養方法を用いて行なうことができる。
培養条件は、宿主、培地、培養方法の種類により適宜選択すればよく、組換え微生物が生育し、第2の態様に係る組換え微生物を産生できる条件であれば特に制限はない。
培養温度は20℃〜45℃、好ましくは24℃〜37℃で好気的に培養を行う。
組換え微生物を培養して得られる培養物は、組換え微生物の培養から得られたものであれば特に限定はされない。培養物は、培地から回収した菌体であってもよいし、培地から回収した上述の(1)〜(7)のいずれかに記載のポリヌクレオチドがコードするタンパク質でもよい。あるいは、菌体を含む培地又は菌体を含まないが組換え微生物が生成する組換えタンパク質を含む培地をそのまま用いてもよい。
上記のような細胞の破砕あるいは溶解処理は、リゾチーム処理、凍結融解、超音波破砕などの公知の方法に従い組換え微生物の細胞膜を破壊することにより、行うことができる。
本開示において、組成物中の各成分の量について言及する場合、組成物中に各成分に該当する物質が複数存在する場合には、特に断らない限り、組成物中に存在する当該複数の物質の合計量を意味する。
ピリドキシン塩酸塩、ピリドキサール塩酸塩、ピリドキサミン二塩酸塩は高速液体クロマトグラフィーにより定量した。これらの分析条件は次の通りである。
カラム:Shodex(登録商標) Asahipak ODP−50 6E(昭和電工株式会社)
ガードカラム:Shodex(登録商標) Asahipak ODP−50G 6A(昭和電工株式会社)
カラム温度:30℃
ポンプ流速:1.0ml/min
溶離液:50mMリン酸緩衝液(pH2.0)
検出:UV254nm
Mesorhizobium loti MAFF303099由来ピリドキサミン−ピルビン酸アミノトランスフェラーゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化したものをGenScript社に委託して合成し、配列番号14のヌクレオチド配列を有する合成DNAを得た。該合成DNAを鋳型として用い配列番号18のヌクレオチド配列を有するオリゴヌクレオチド及び配列番号19のヌクレオチド配列を有するオリゴヌクレオチドをプライマーとしてPCRにより目的遺伝子を含むDNA断片を増幅した。この増幅されたDNA断片をEcoRI及びBamHIで処理し、得られたDNA断片と、pUC18(Takara製)のEcoRI及びBamHIによる処理物とを、ライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
Saccharomyces cerevisiae由来ピリドキサールレダクターゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化したものをGenScript社に委託して合成し、配列番号13のヌクレオチド配列を有する合成DNAを得た。該合成DNAを鋳型として用い配列番号16のヌクレオチド配列を有するオリゴヌクレオチド及び配列番号17のヌクレオチド配列を有するオリゴヌクレオチドをプライマーとしてPCRにより目的遺伝子を含むDNA断片を増幅した。この増幅されたDNA断片をSalI及びHindIIIで処理し、得られたDNA断片と、比較例1で作製したpUC18-ppatをSalI及びHindIIIで処理した処理物とを、ライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
Shewanella sp. AC10由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD198Aとなる変異を導入し、かつ大腸菌にコドン最適化したものをGenScript社に委託して合成し、配列番号15のヌクレオチド配列を有する合成DNAを得た。該合成DNAを鋳型として用い配列番号20のヌクレオチド配列を有するオリゴヌクレオチド及び配列番号21のヌクレオチド配列を有するオリゴヌクレオチドをプライマーとしてPCRにより目的遺伝子を含むDNA断片を増幅した。この増幅されたDNA断片をBamHI及びSalIで処理し、得られたDNA断片と、比較例1で作製したpUC18-ppatをBamHI及びSalIで処理した処理物とを、ライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。この形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
Shewanella sp. AC10由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD198Aとなる変異を導入し、かつ大腸菌にコドン最適化したものをGenScript社に委託して合成し、配列番号15のヌクレオチド配列を有する合成DNAを得た。該合成DNAを鋳型として用い配列番号20のヌクレオチド配列を有するオリゴヌクレオチド及び配列番号21のヌクレオチド配列を有するオリゴヌクレオチドをプライマーとしてPCRにより目的遺伝子を含むDNA断片を増幅した。この増幅されたDNA断片をBamHI及びSalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理した処理物とを、ライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。この形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
pUC18、比較例1〜3及び実施例1で作製したプラスミドのそれぞれで形質転換したDH5αを、500mlのバッフル付き三角フラスコ中のアンピシリン100μg/mlを含むLB培地100mlに接種し、30℃にて24時間振盪培養した。培養液を8000rpmで20分間遠心分離し、沈殿物として得られた菌体を水1mlに懸濁して菌体懸濁液を調製した。
ピリドキシン塩酸塩及びL−アラニンを水で溶解し、ピリドキシン塩酸塩(600mM)及びL−アラニン(600mM)を含む基質液を調製した。基質液のpHを、25%アンモニア水でpH8.0に調整した。基質液500μlと菌体懸濁液1000μlを混合し、37℃で24時間反応させた。反応液を一部採取し、上記に示す分析条件で分析した。結果を表1に示す。なお、表1に示す収率とは、基質液中のピリドキシン塩酸塩のモル量に対する得られたピリドキサミン二塩酸塩のモル量の比を表す。
実施例1で作製したプラスミド(pUC18-ppat-adh-plr)で形質転換したDH5αを、500mlのバッフル付き三角フラスコ中のアンピシリン100μg/mlを含むLB培地100mlに接種し、30℃にて24時間振盪培養した。培養液を8000rpmで20分間遠心分離し、沈殿物として得られた菌体を水0.5mlに懸濁して菌体懸濁液を調製した。
実施例1で作製したプラスミド(pUC18-ppat-adh-plr)を形質転換したDH5αを、500mlのバッフル付き三角フラスコ中のアンピシリン100μg/mlを含むLB培地100mlに接種し、30℃にて24時間振盪培養した。培養液を8000rpmで20分間遠心分離し、沈殿物として得られた菌体を水0.5mlに懸濁して菌体懸濁液を調製した。
ピリドキシン塩酸塩を水で溶解し、25%アンモニア水でpH8.0に調整して、ピリドキシン塩酸塩(600mM)を含む基質液を調製した。この基質液を500μl、菌体懸濁液500μl、及び残量の水を混合して1,500μlの反応液を調製し、L−アラニン粉末を飽和溶解度となるように添加し、37℃で24時間反応させた。反応液を一部採取し、上記に示す分析条件で分析することで、ピリドキシン塩酸塩、ピリドキサール塩酸塩、ピリドキサミン二塩酸塩を定量した。その結果、反応収率は39%であった。つまり、L−アラニンをより低濃度で用いた場合の方が高い収率を達成できていた。
Saccharomyces eubayanus由来ピリドキサールレダクターゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にSalI、3’末端にHindIIIを導入した遺伝子をeurofins社に委託して合成し、配列番号75のヌクレオチド配列を有する合成DNAを得た。該合成DNAをSalI/HindIIIで処理し、得られたDNA断片と、比較例3で作製したpUC18-ppat-adhをSalI及びHindIIIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体はLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号75のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-adh-Seplrと命名した。
Torulaspora delbrueckii由来ピリドキサールレダクターゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にSalI、3’末端にHindIIIを導入した遺伝子をeurofins社に委託して合成し、配列番号76のヌクレオチド配列を有する合成DNAを得た。該合成DNAをSalI/HindIIIで処理し、得られたDNA断片と、比較例3で作製したpUC18-ppat-adhをSalI及びHindIIIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号76のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-adh-Tdplrと命名した。
Zygosaccharomyces bailii 由来ピリドキサールレダクターゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にSalI、3’末端にHindIIIを導入した遺伝子をGenScript社に委託して合成し、配列番号77のヌクレオチド配列を有する合成DNAを得た。該合成DNAをSalI/HindIIIで処理し、得られたDNA断片と、比較例3で作製したpUC18-ppat-adhをSalI及びHindIIIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号77のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-adh-Zbplrと命名した。
Aspergillus oryzae 由来ピリドキサールレダクターゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にSalI、3’末端にHindIIIを導入した遺伝子をGenScript社に委託して合成し、配列番号78のヌクレオチド配列を有する合成DNAを得た。該合成DNAをSalI/HindIIIで処理し、得られたDNA断片と、比較例3で作製したpUC18-ppat-adhをSalI及びHindIIIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号78のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-adh-Aoplrと命名した。
Kluyveromyces marxianus 由来ピリドキサールレダクターゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にSalI、3’末端にHindIIIを導入した遺伝子をGenScript社に委託して合成し、配列番号79のヌクレオチド配列を有する合成DNAを得た。該合成DNAをSalI/HindIIIで処理し、得られたDNA断片と、比較例3で作製したpUC18-ppat-adhをSalI及びHindIIIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号79のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-adh-Kmplrと命名した。
Candida albicans由来ピリドキサールレダクターゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にSalI、3’末端にHindIIIを導入した遺伝子をeurofins社に委託して合成し、配列番号80のヌクレオチド配列を有する合成DNAを得た。該合成DNAをSalI/HindIIIで処理し、得られたDNA断片と、比較例3で作製したpUC18-ppat-adhをSalI及びHindIIIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号80のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-adh-Caplrと命名した。
Yarrowia lipolytica 由来ピリドキサールレダクターゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にSalI、3’末端にHindIIIを導入した遺伝子をGenScript社に委託して合成し、配列番号81のヌクレオチド配列を有する合成DNAを得た。該合成DNAをSalI/HindIIIで処理し、得られたDNA断片と、比較例3で作製したpUC18-ppat-adhをSalI及びHindIIIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号81のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-adh-Ylplrと命名した。
実施例2〜8で作製したプラスミドのそれぞれで形質転換したDH5αを、500mlのバッフル付き三角フラスコ中のアンピシリン100μg/mlを含むLB培地100mlに接種し、30℃にて24時間振盪培養した。培養液の40mlを取り、5000rpmで5分間遠心分離し、沈殿物として得られた菌体を水0.9mlに懸濁して菌体懸濁液を調製した。
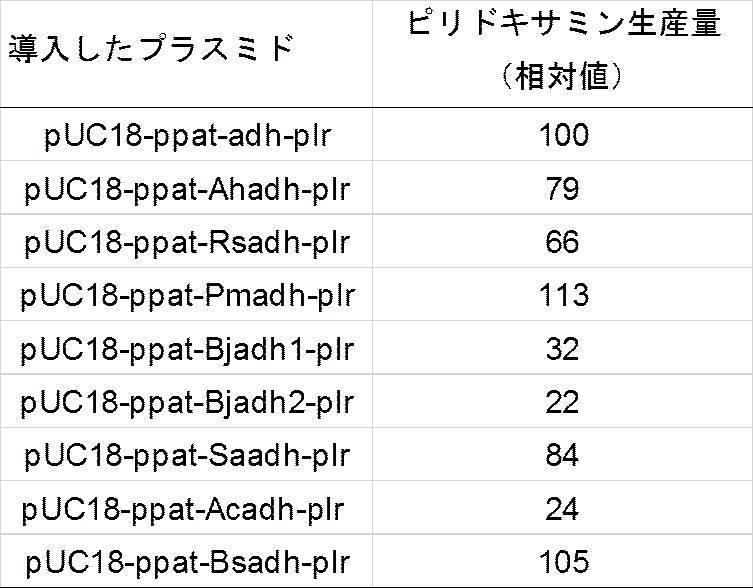
ピリドキシン塩酸塩及びL−アラニンを水で溶解し、ピリドキシン塩酸塩(500mM)及びL−アラニン(500mM)を含む基質液を調製した。基質液のpHを、25%アンモニア水でpH8.0に調整した。基質液400μl、水500μl及び菌体懸濁液100μlを混合し、37℃で1時間反応させた。反応液を一部採取し、上記した分析条件で分析した。結果を表3に示す。なお、表3に示すピリドキサミン生産量とは、実施例1で作製したppat-adh-plr発現株が生産するピリドキサミン二塩酸塩のモル量を100とした相対量で表す。
比較例2で作製したpUC18-ppat-plrをSalI及びHindIIIで処理し、回収したピリドキサールレダクターゼ遺伝子(plr)断片と、比較例3で作製したpUC-ppat-adhをBamHI及びSalIで処理し、回収したアラニンデヒドロゲナーゼ遺伝子(adh)断片とpUC18のBamHI及びHindIIIによる処理物とを、ライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出し、得られたプラスミドをpUC18-adh-plrと命名した。
Mesorhizobium sp. YR577由来ピリドキサミン−ピルビン酸アミノトランスフェラーゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にEcoRI、3’末端にBamHIを導入した遺伝子をeurofins社に委託して合成し、配列番号82のヌクレオチド配列を有する合成DNAを得た。該合成DNAをEcoRI/BamHIで処理し、得られたDNA断片と、比較例4で作製したpUC18-adh-plrをEcoRI及びBamHIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号82のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-Msppat-adh-plrと命名した。
Pseudaminobacter salicylatoxidans由来ピリドキサミン−ピルビン酸アミノトランスフェラーゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にEcoRI、3’末端にBamHIを導入した遺伝子をeurofins社に委託して合成し、配列番号83のヌクレオチド配列を有する合成DNAを得た。該合成DNAをEcoRI/BamHIで処理し、得られたDNA断片と、比較例4で作製したpUC18-adh-plrをEcoRI及びBamHIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号83のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-Psppat-adh-plrと命名した。
Bauldia litoralis由来ピリドキサミン−ピルビン酸アミノトランスフェラーゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にEcoRI、3’末端にBamHIを導入した遺伝子をeurofins社に委託して合成し、配列番号84のヌクレオチド配列を有する合成DNAを得た。該合成DNAをEcoRI/BamHIで処理し、得られたDNA断片と、比較例4で作製したpUC18-adh-plrをEcoRI及びBamHIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号84のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-Blppat-adh-plrと命名した。
Skermanella stibiiresistens由来ピリドキサミン−ピルビン酸アミノトランスフェラーゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にEcoRI、3’末端にBamHIを導入した遺伝子をeurofins社に委託して合成し、配列番号85のヌクレオチド配列を有する合成DNAを得た。該合成DNAをEcoRI/BamHIで処理し、得られたDNA断片と、比較例4で作製したpUC18-adh-plrをEcoRI及びBamHIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号85のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-Ssppat-adh-plrと命名した。
Rhizobium sp. AC44/96由来ピリドキサミン−ピルビン酸アミノトランスフェラーゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にEcoRI、3’末端にBamHIを導入した遺伝子をeurofins社に委託して合成し、配列番号86のヌクレオチド配列を有する合成DNAを得た。該合成DNAをEcoRI/BamHIで処理し、得られたDNA断片と、比較例4で作製したpUC18-adh-plrをEcoRI及びBamHIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号86のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-Rsppat-adh-plrと命名した。
Erwinia toletana由来ピリドキサミン−ピルビン酸アミノトランスフェラーゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にEcoRI、3’末端にBamHIを導入した遺伝子をeurofins社に委託して合成し、配列番号87のヌクレオチド配列を有する合成DNAを得た。該合成DNAをEcoRI/BamHIで処理し、得られたDNA断片と、比較例4で作製したpUC18-adh-plrをEcoRI及びBamHIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号87のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-Etppat-adh-plrと命名した。
Herbiconiux ginsengi由来ピリドキサミン−ピルビン酸アミノトランスフェラーゼ遺伝子のヌクレオチド配列を大腸菌にコドン最適化するようにデザインし、5’末端にEcoRI、3’末端にBamHIを導入した遺伝子をeurofins社に委託して合成し、配列番号88のヌクレオチド配列を有する合成DNAを得た。該合成DNAをEcoRI/BamHIで処理し、得られたDNA断片と、比較例4で作製したpUC18-adh-plrをEcoRI及びBamHIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号88のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-Hgppat-adh-plrと命名した。
実施例9〜15で作製したプラスミドのそれぞれで形質転換したDH5αを、500mlのバッフル付き三角フラスコ中のアンピシリン100μg/mlを含むLB培地100mlに接種し、30℃にて24時間振盪培養した。培養液の40mlを取り、5000rpmで5分間遠心分離し、沈殿物として得られた菌体を水0.9mlに懸濁して菌体懸濁液を調製した。
ピリドキシン塩酸塩及びL−アラニンを水で溶解し、ピリドキシン塩酸塩(500mM)及びL−アラニン(500mM)を含む基質液を調製した。基質液のpHを、25%アンモニア水でpH8.0に調整した。基質液400μl、水500μlと菌体懸濁液100μlを混合し、37℃で1時間反応させた。反応液を一部採取し、上記した分析条件で分析した。結果を表4に示す。なお、表4に示すピリドキサミン生産量とは、実施例1で作製したppat-adh-plr発現株が生産するピリドキサミン二塩酸塩のモル量を100とした相対量で表す。
Aeromonas hydrophila由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD198Aとなる変異を導入し、かつ大腸菌にコドン最適化するようにデザインし、さらに5’末端にBamHI、3’末端にSalIを導入したものをeurofins社に委託して合成し、配列番号89のヌクレオチド配列を有する合成DNAを得た。該合成DNAをBamHI/SalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号89のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-Ahadh-plrと命名した。
Rhizobium sp. LPU83由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD198Aとなる変異を導入し、かつ大腸菌にコドン最適化するようにデザインし、さらに5’末端にBamHI、3’末端にSalIを導入したものをeurofins社に委託して合成し、配列番号90のヌクレオチド配列を有する合成DNAを得た。該合成DNAをBamHI/SalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号90のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-Rsadh-plrと命名した。
Pseudomonas mendocina由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD198Aとなる変異を導入し、かつ大腸菌にコドン最適化するようにデザインし、さらに5’末端にBamHI、3’末端にSalIを導入したものをeurofins社に委託して合成し、配列番号91のヌクレオチド配列を有する合成DNAを得た。該合成DNAをBamHI/SalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号91のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-Pmadh-plrと命名した。
Bradyrhizobium japonicum由来アラニンデヒドロゲナーゼ遺伝子はBradyrhizobium japonicum中に2種存在する。これら2種のBradyrhizobium japonicum由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD198Aとなる変異を導入し、かつ大腸菌にコドン最適化するようにデザインし、さらに5’末端にBamHI、3’末端にSalIを導入したものをeurofins社に委託して合成し、配列番号92及び配列番号93のヌクレオチド配列を有する合成DNA2種を得た。配列番号92を含む該合成DNAをBamHI/SalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号92のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-Bjadh1-plrと命名した。
同様に配列番号93を含む該合成DNAをBamHI/SalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号93のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-Bjadh2-plrと命名した。
Streptomyces aureofaciens由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD198Aとなる変異を導入し、かつ大腸菌にコドン最適化するようにデザインし、さらに5’末端にBamHI、3’末端にSalIを導入したものをeurofins社に委託して合成し、配列番号94のヌクレオチド配列を有する合成DNAを得た。該合成DNAをBamHI/SalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号94のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-Saadh-plrと命名した。
Anabaena cylindrica由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD198Aとなる変異を導入し、かつ大腸菌にコドン最適化するようにデザインし、さらに5’末端にBamHI、3’末端にSalIを導入したものをeurofins社に委託して合成し、配列番号95のヌクレオチド配列を有する合成DNAを得た。該合成DNAをBamHI/SalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号95のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-Acadh-plrと命名した。
Bacillus subtilis由来アラニンデヒドロゲナーゼ遺伝子のヌクレオチド配列に、コードするアミノ酸配列においてD196Aとなる変異を導入し、かつ大腸菌にコドン最適化するようにデザインし、さらに5’末端にBamHI、3’末端にSalIを導入したものをeurofins社に委託して合成し、配列番号96のヌクレオチド配列を有する合成DNAを得た。該合成DNAをBamHI/SalIで処理し、得られたDNA断片と、比較例2で作製したpUC18-ppat-plrをBamHI及びSalIで処理したDNA断片とをライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションした。このライゲーション産物でエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。このようにして得られた形質転換体からプラスミドを抽出した。
通常のヌクレオチド配列の決定法に従い、プラスミドに導入されたDNA断片のヌクレオチド配列が配列番号96のヌクレオチド配列であることを確認した。得られたプラスミドをpUC18-ppat-Bsadh-plrと命名した。
実施例16〜22で作製したプラスミドのそれぞれで形質転換したDH5αを、500mlのバッフル付き三角フラスコ中のアンピシリン100μg/mlを含むLB培地100mlに接種し、30℃にて24時間振盪培養した。培養液の40mlを取り、5000rpmで5分間遠心分離し、沈殿物として得られた菌体を水0.9mlに懸濁して菌体懸濁液を調製した。
ピリドキシン塩酸塩及びL−アラニンを水で溶解し、ピリドキシン塩酸塩(500mM)及びL−アラニン(500mM)を含む基質液を調製した。基質液のpHを、25%アンモニア水でpH8.0に調整した。基質液400μl、水500μlと菌体懸濁液100μlを混合し、37℃で1時間反応させた。反応液を一部採取し、上記した分析条件で分析した。結果を表5に示す。なお、表5に示すピリドキサミン生産量とは、実施例1で作製したppat-adh-plr発現株が生産するピリドキサミン二塩酸塩のモル量を100とした相対量で表す。
Saccharomyces cerevisiaeのピリドキシンデヒドロゲナーゼ遺伝子(配列番号1)をコードするヌクレオチド配列を大腸菌にコドン最適化したDNAを、GenScript社に委託して作成した(配列番号13)。得られた合成DNAを鋳型として、配列番号18及び配列番号17に示すオリゴヌクレオチドをプライマーとして用いてポリメラーゼ連鎖反応(PCR)法により目的遺伝子を増幅した。増幅したDNAをEcoRI/HindIIIで処理し、得られたDNA断片と、pUC18(Takara社製)のEcoRI/HindIII処理物とを、ライゲーション ハイ(東洋紡株式会社製)を用いてライゲーションして発現ベクターを得た。発現ベクターを含むライゲーション産物によってエシェリヒア コリDH5α(東洋紡株式会社製)を形質転換した。形質転換体をLB寒天培地で培養し、アンピシリン耐性株の中から目的のプラスミドを有する株を選択した。
pUC18-plrによって形質転換した大腸菌DH5α株をLB液体培地2mL(アンピシリン100μg/mL含有) へ接種し、試験管で37℃にて24時間振盪培養した。1mLの培養液を13000rpmで5分間遠心分離し、得られた菌体を水1mLに懸濁して菌体懸濁液を得た。
ピリドキシン塩酸塩及びβ−NADP+をそれぞれ100mMとなるよう水に溶かして基質液を得た。100μLの基質液、100μLの1M Tris−HCl(pH8.0)、及び700μLの水を混合し、そこに100μLの菌体懸濁液を添加して反応を開始した。反応は37℃で1時間行った。
1時間の反応によって37.2μgのピリドキサールが得られ、OD660当たりのピリドキサール生成速度は2.1×10−3μmol/min/OD660であった。
・HPLCの分析条件
カラム:Shodex Asahipak ODP−50 6E(商品名、昭和電工株式会社製)
ガードカラム:Shodex Asahipak ODP-50G 6A(商品名、昭和電工株式会社製)
カラム温度:30℃
ポンプ流速:1.0mL/分
溶離液:50mMリン酸緩衝液(pH2.0)
検出:UV294nm。
Saccharomyces cerevisiae X−2180−1A(ATCC26486)を2mLのYPD液体培地へ接種し、試験管で30℃にて24時間振盪培養した。1mLの培養液を13000rpmで5分間遠心分離し、得られた菌体を1mLの水で懸濁して菌体懸濁液を得た。
基質液として、実施例24の基質液と同じ組成のものを調製し、実施例24の反応条件と同じ条件で反応を行った。
1時間の反応によってピリドキサールは得られなかった。
実施例24と同様に菌体懸濁液を調製し、ピリドキサール塩酸塩及びβ−NADPHをそれぞれ100mMとなるよう水に溶かして基質液を得た。100μLの基質液、100μLの1M Tris−HCl(pH8.0)、及び700μLの水を混合し、そこに100μLの菌体懸濁液を添加して反応を開始した。
反応終了後に反応液の一部を採取して高速液体クロマトグラフィー(HPLC)によって、実施例24と同様の条件で分析を行った。
1時間の反応によって815.5μgのピリドキシンが得られ、OD660当たりのピリドキシン生成速度は4.5×10−2μmol/min/OD660であった。
比較例5と同様に菌体懸濁液を調製し、基質液として実施例25の基質液と同じ組成のものを調製し、実施例25の反応条件と同じ条件で反応を行った。
反応終了後に反応液の一部を採取して高速液体クロマトグラフィー(HPLC)によって、実施例24と同様の条件で分析を行った。
1時間の反応によって10.9μgのピリドキシンが得られ、OD660当たりのピリドキシン生成速度は1.7×10−4μmol/min/OD660であった。
pUC18-plrによって形質転換した大腸菌DH5α株をLB液体培地100mL(アンピシリン100μg/mL含有) へ接種し、バッフル付三角フラスコで37℃にて24時間振盪培養した。40mLの培養液を8000rpmで20分間遠心分離し、得られた菌体を水400μLに懸濁して菌体懸濁液を得た。基質液として実施例24の基質液と同じ組成のものを調製し、実施例24の反応条件と同じ条件で反応を行った。反応は37℃で30分行った。
反応終了後に反応液の一部を採取して高速液体クロマトグラフィー(HPLC)によって、実施例24と同様の条件で分析を行った。
30分の反応によって262.4μgのピリドキサールが得られ、乾燥菌体重量当たりのピリドキサール生成速度は5.8μmol/min/g−dry cellであった。
スーパーカメリヤ ドライイースト(商品名、日清)を100mg/mlになるよう水で懸濁し、菌体懸濁液を得た。
基質液として、実施例24の基質液と同じ組成のものを調製し、実施例24の反応条件と同じ条件で反応を行った。
反応終了後に反応液の一部を採取して高速液体クロマトグラフィー(HPLC)によって、実施例24と同様の条件で分析を行った。
30分の反応によって34.1μgのピリドキサールが得られ、乾燥菌体重量当たりのピリドキサール生成速度は0.9μmol/min/g−dry cellであった。
実施例26と同様に菌体懸濁液を調製し、基質液として実施例25の基質液と同じ組成のものを調製し、実施例25の反応条件と同じ条件で反応を行った。
反応終了後に反応液の一部を採取して高速液体クロマトグラフィー(HPLC)によって、実施例24と同様の条件で分析を行った。
30分の反応によって1457.4μgのピリドキシンが得られ、乾燥菌体重量当たりのピリドキシン生成速度は33.5μmol/min/g−dry cellであった。
比較例7と同様に菌体懸濁液を調製し、基質液として実施例25の基質液と同じ組成のものを調製し、実施例25の反応条件と同じ条件で反応を行った。
反応終了後に反応液の一部を採取して高速液体クロマトグラフィー(HPLC)によって、実施例24と同様の条件で分析を行った。
30分の反応によって325.3μgnoピリドキシンが得られ、乾燥菌体重量当たりのピリドキシン生成速度は13.9μmol/min/g−dry cellであった。
本明細書に記載された全ての文献、特許出願、及び技術規格は、個々の文献、特許出願、及び技術規格が参照により取り込まれることが具体的かつ個々に記された場合と同程度に、本明細書中に参照により取り込まれる。
Claims (32)
- ピリドキシンデヒドロゲナーゼをコードする遺伝子、ピリドキサールからピリドキサミンを合成する酵素活性を有するピリドキサミン合成酵素をコードする遺伝子、及び前記ピリドキサミン合成酵素が消費するアミノ酸を再生する酵素活性を有するアミノ酸再生酵素をコードする遺伝子を有し、
前記ピリドキシンデヒドロゲナーゼをコードする遺伝子、前記ピリドキサミン合成酵素をコードする遺伝子及び前記アミノ酸再生酵素をコードする遺伝子のうち少なくとも2つは菌体外から導入された遺伝子であるか又は菌体に内在するがその発現が強化された遺伝
子である、組換え微生物。 - 前記ピリドキサミン合成酵素が、ピリドキサミン−ピルビン酸トランスアミナーゼ、ピリドキサミン−オキサロ酢酸トランスアミナーゼ、アスパラギン酸トランスアミナーゼ、又はピリドキサミンリン酸トランスアミナーゼである、請求項1に記載の組換え微生物。
- 前記ピリドキシンデヒドロゲナーゼが下記の部分アミノ酸配列(a)及び部分アミノ酸配列(b)のうち少なくとも1つを含み、且つピリドキシンデヒドロゲナーゼ活性を有する、請求項1又は請求項2に記載の組換え微生物。
(a)NX1X2EX3YG(配列番号97)
(ここで、X1は、V、C、I、A、M、S、G又はLを表し、
X2は、G又はAを表し、
X3は、F又はLを表す)
(b)X4X5X6KGX7(配列番号98)
(X4は、I、V、F又はLを表し、
X5は、S、T、N、C又はMを表し、
X6は、C、V、A、I、W又はFを表し、
X7は、G、A、S又はCを表す) - 前記ピリドキシンデヒドロゲナーゼが酵素番号EC1.1.1.65で表される、請求項1〜請求項3のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキシンデヒドロゲナーゼをコードする遺伝子がSaccharomycescerevisiaeに由来する、請求項1〜請求項4のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキシンデヒドロゲナーゼをコードする遺伝子が、
配列番号7及び配列番号53〜配列番号59のうちいずれかのヌクレオチド配列を有するか、
配列番号13及び配列番号75〜81のいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域を有するか、
配列番号7及び配列番号53〜配列番号59のうちいずれかのヌクレオチド配列と相補的なヌクレオチド配列を有するDNA、又は配列番号13及び配列番号75〜81のいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有する、請求項1〜請求項5のうちいずれか一項に記載の組換え微生物。 - 前記ピリドキシンデヒドロゲナーゼをコードする遺伝子がSchizosaccharomycespombeに由来する、請求項1〜請求項4のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキシンデヒドロゲナーゼをコードする遺伝子が、配列番号8のヌクレオチド配列を有するか、配列番号8のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキシンデヒドロゲナーゼ活性を有するタンパク質をコードするヌクレオチド配列を有する、請求項1〜請求項4及び請求項7のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキシンデヒドロゲナーゼをコードする遺伝子が、配列番号1、配列番号2及び配列番号22〜配列番号28のうちいずれかのアミノ酸配列、又は配列番号1、配列番号2及び配列番号22〜28のアミノ酸配列のうち少なくとも1つと80%以上の配列同一性を有し且つピリドキシンデヒドロゲナーゼ活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、請求項1〜請求項8のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキサミン合成酵素が、下記の部分アミノ酸配列(c)、部分アミノ酸配列(d)、部分アミノ酸配列(e)、部分アミノ酸配列(f)、部分アミノ酸配列(g)及び部分アミノ酸配列(h)のうち少なくとも1つを含み、且つピリドキサールからピリドキサミンを合成する酵素活性を有する、請求項1〜請求項9のうちいずれか一項に記載の組換え微生物。
(c)X8X9X10X11X12X13(配列番号99)
(X8は、L、M、I又はVを表し、
X9は、H又はQを表し、
X10は、G、C又はAを表し、
X11は、E又はDを表し、
X12は、P又はAを表し、
X13は、V、I、L又はAを表す)
(d)X14X15TPSGTX16X17(配列番号100)
(X14は、H又はSを表し、
X15は、D又はEを表し、
X16は、I、V又はLを表し、
X17は、N又はTを表す)
(e)X18DX19VSX20X21(配列番号101)
(X18は、V、I又はAを表し、
X19は、A、T又はSを表し、
X20は、S、A又はGを表し、
X21は、F、W、又はVを表す)
(f)X22X23X24KCX25GX26X27P(配列番号102)
(X22は、G又はSを表し、
X23は、P、S又はAを表し、
X24は、N、G、S、A又はQを表し、
X25は、L又はMを表し、
X26は、A、S、C又はGを表し、
X27は、P、T、S又はAを表す)
(g)X28X29X30X31SX32GX33X34(配列番号103)
(X28は、G又はDを表し、
X29は、V又はIを表し、
X30は、V、T、A、S、M、I又はLを表し、
X31は、F、M、L、I又はVを表し、
X32は、S、G、A、T、I、L又はHを表し、
X33は、R、M又はQを表し、
X34は、G、R、A、D、H又はKを表す)
(h)X35X36RX37X38HMGX39X40A(配列番号104)
(X35は、L又はVを表し、
X36は、T、I、V又はLを表し、
X37は、I、V又はLを表し、
X38は、G又はSを表し、
X39は、P、A又はRを表し、
X40は、T、V又はSを表す) - 前記ピリドキサミン合成酵素が酵素番号EC2.6.1.30で表される、請求項1〜請求項10のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキサミン合成酵素をコードする遺伝子がMesorhizobiumlotiに由来する、請求項1〜請求項11のうちのいずれか一項に記載の組換え微生物。
- 前記ピリドキサミン合成酵素をコードする遺伝子が、
配列番号9及び配列番号60〜配列番号66のうちいずれかのヌクレオチド配列を有するか、
配列番号14のヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域又は配列番号82〜配列番号88のうちいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域を有するか、
配列番号9及び配列番号60〜配列番号66のうちいずれかのヌクレオチド配列と相補的なヌクレオチド配列を有するDNA、又は配列番号14のヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域若しくは配列番号82〜配列番号88のうちいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキサールからピリドキサミンを合成する酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、請求項1〜請求項12のうちいずれか一項に記載の組換え微生物。 - 前記ピリドキサミン合成酵素をコードする遺伝子が、配列番号3及び配列番号29〜配列番号35のうちいずれかのアミノ酸配列、又は配列番号3及び配列番号29〜配列番号35のアミノ酸配列のうち少なくとも1つと80%以上の配列同一性を有し且つピリドキサールからピリドキサミンを合成する酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、請求項1〜請求項13のうちいずれか一項に記載の組換え微生物。
- 前記アミノ酸再生酵素がアラニンデヒドロゲナーゼである、請求項10〜請求項14のうちいずれか一項に記載の組換え微生物。
- 前記アラニンデヒドロゲナーゼが、ピルビン酸及びNH3からL−アラニンを生成する反応の際にNADPHを補酵素として利用可能である、請求項15に記載の組換え微生物。
- 前記アラニンデヒドロゲナーゼが下記の部分アミノ酸配列(i)、部分アミノ酸配列(j)、部分アミノ酸配列(k)及び部分アミノ酸配列(l)のうち少なくとも1つを含み、且つピルビン酸及びNH3からL−アラニンを生成する活性を有する請求項15又は請求項16に記載の組換え微生物。
(i)EX41KX42X43EX44RX45X46(配列番号105)
(X41は、I、T、S、F、N又はVを表し、
X42は、N、M、A、V、L、T又はDを表し、
X43は、H、N、L又はQを表し、
X44は、Y、N又はFを表し、
X45は、V、又はIを表し、
X46は、G又はAを表す)
(j)X47X48X49KVKEPX50(配列番号106)
(X47は、M又はLを表し、
X48は、I、L又はVを表し、
X49は、V、L、I又はMを表し、
X50は、Q、L、V、N又はIを表す)
(k)LX51TYLHLA(配列番号107)
(X51は、F又はYを表す)
(l)X52DX53AX54DQGG(配列番号108)
(X52は、V又はAを表し、
X53は、V又はIを表し、
X54は、I又はVを表す) - 前記アミノ酸再生酵素をコードする遺伝子がShewanellasp.AC10に由来する、請求項15〜請求項17のうちいずれか一項に記載の組換え微生物。
- 前記アミノ酸再生酵素をコードする遺伝子が、
配列番号11及び配列番号67〜配列番号74のうちいずれかのヌクレオチド配列を有するか、
配列番号15及び配列番号89〜配列番号96のうちいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域を有するか、
配列番号11及び配列番号67〜配列番号74のうちいずれかのヌクレオチド配列と相補的なヌクレオチド配列を有するDNA、又は配列番号15及び配列番号89〜配列番号96のうちいずれかのヌクレオチド配列における18番目のヌクレオチド〜3’末端までの領域と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つアラニン再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、請求項15〜請求項18のうちいずれか一項に記載の組換え微生物。 - 前記アミノ酸再生酵素をコードする遺伝子が、配列番号5及び配列番号45〜配列番号52のうちいずれかのアミノ酸配列、又は配列番号5及び配列番号45〜配列番号52のアミノ酸配列のうち少なくとも1つと80%以上の配列同一性を有し且つアラニン再生酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、請求項15〜請求項19のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキサミン合成酵素が酵素番号EC2.6.1.31又はEC2.6.1.1で表される、請求項1〜請求項9のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキサミン合成酵素をコードする遺伝子がEscherichiacoliに由来する、請求項1〜請求項9及び請求項21のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキサミン合成酵素をコードする遺伝子が、配列番号10のヌクレオチド配列を有するか、配列番号10のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つピリドキサールからピリドキサミンを合成する酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、請求項1〜請求項9、請求項21及び請求項22のうちいずれか一項に記載の組換え微生物。
- 前記ピリドキサミン合成酵素をコードする遺伝子が、配列番号4のアミノ酸配列、又は配列番号4のアミノ酸配列と80%以上の配列同一性を有し且つピリドキサールからピリドキサミンを合成する酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、請求項1〜請求項9及び請求項21〜請求項23のうちいずれか一項に記載の組換え微生物。
- 前記アミノ酸再生酵素がグルタミン酸デヒドロゲナーゼである、請求項21〜請求項24のうちいずれか一項に記載の組換え微生物。
- 前記アミノ酸再生酵素をコードする遺伝子がEscherichiacoliに由来する、請求項25に記載の組換え微生物。
- 前記アミノ酸再生酵素をコードする遺伝子が、配列番号12のヌクレオチド配列を有するか、配列番号12のヌクレオチド配列と相補的なヌクレオチド配列を有するDNAとストリンジェントな条件下でハイブリダイズし、且つグルタミン酸再生酵素活性を有するタンパク質をコードするヌクレオチド配列を有する、請求項25又は請求項26に記載の組換え微生物。
- 前記アミノ酸再生酵素をコードする遺伝子が、配列番号6のアミノ酸配列、又は配列番号6のアミノ酸配列と80%以上の配列同一性を有し且つグルタミン酸再生酵素活性を有するアミノ酸配列、をコードするヌクレオチド配列を有する、請求項25〜請求項27のうちいずれか一項に記載の組換え微生物。
- 組換え大腸菌である、請求項1〜請求項28のうちいずれか一項に記載の組換え微生物。
- 請求項1〜請求項29のうちいずれか一項に記載の組換え微生物若しくは前記組換え微生物の培養物又は前記組換え微生物若しくは前記培養物の処理物と、ピリドキシン又はその塩とを接触させ、ピリドキサミン又はその塩を生産させる、ピリドキサミン又はその塩の製造方法。
- 前記組換え微生物若しくは前記組換え微生物の培養物又は前記組換え微生物若しくは前記培養物の処理物が、前記ピリドキシンデヒドロゲナーゼ、前記ピリドキサミン合成酵素、及び前記アミノ酸再生酵素を含む、請求項30に記載の製造方法。
- 前記組換え微生物の処理物又は前記組換え微生物の培養物の処理物が、加熱処理、冷却処理、細胞の機械的破壊、超音波処理、凍結融解処理、乾燥処理、加圧又は減圧処理、浸透圧処理、細胞の自己消化、界面活性剤処理、酵素処理、細胞分離処理、精製処理及び抽出処理からなる群から選択される1つ以上を含む処理による処理物である、請求項30又は請求項31に記載の製造方法。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2017095571 | 2017-05-12 | ||
| JP2017095573 | 2017-05-12 | ||
| JP2017095571 | 2017-05-12 | ||
| JP2017095573 | 2017-05-12 | ||
| PCT/JP2018/018413 WO2018207931A1 (ja) | 2017-05-12 | 2018-05-11 | 組換え微生物、組換え微生物を用いたピリドキサミン又はその塩の製造方法、及び組換え微生物を用いたピリドキサール又はその塩の製造方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JPWO2018207931A1 JPWO2018207931A1 (ja) | 2020-03-19 |
| JP6964314B2 true JP6964314B2 (ja) | 2021-11-10 |
Family
ID=64105567
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2019517729A Active JP6964314B2 (ja) | 2017-05-12 | 2018-05-11 | 組換え微生物、組換え微生物を用いたピリドキサミン又はその塩の製造方法、及び組換え微生物を用いたピリドキサール又はその塩の製造方法 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US20200123581A1 (ja) |
| EP (1) | EP3623472A4 (ja) |
| JP (1) | JP6964314B2 (ja) |
| CN (1) | CN110678546A (ja) |
| WO (1) | WO2018207931A1 (ja) |
Families Citing this family (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN110804634B (zh) * | 2019-12-04 | 2022-02-25 | 深圳瑞德林生物技术有限公司 | 酶催化法制备2,4-二氨基丁酸的工艺 |
| CN112375725B (zh) * | 2021-01-18 | 2021-03-30 | 中国科学院天津工业生物技术研究所 | 一种生产维生素b6的代谢工程菌株及其构建方法与应用 |
Family Cites Families (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS431268Y1 (ja) | 1965-06-16 | 1968-01-22 | ||
| US5766894A (en) | 1995-09-30 | 1998-06-16 | Roche Vitamins Inc. | Production of vitamin B6 by fermentation |
| ID22473A (id) | 1998-04-15 | 1999-10-21 | Hoffmann La Roche | Produksi enzimatik vitamin b6 |
| CN1715413A (zh) * | 1999-06-25 | 2006-01-04 | Basf公司 | 编码代谢途径蛋白的谷氨酸棒杆菌基因 |
| WO2004035010A2 (en) | 2002-03-22 | 2004-04-29 | Basf Aktiengesellschaft | Methods and organisms for production of b6 vitamers |
| US7214799B2 (en) | 2004-02-09 | 2007-05-08 | Biostratum, Inc. | Methods for the synthesis of pyridoxamine |
| JP2007159401A (ja) * | 2004-05-07 | 2007-06-28 | Precision System Science Co Ltd | 改変タンパク質の作製方法 |
| WO2006066806A1 (en) | 2004-12-20 | 2006-06-29 | Dsm Ip Assets B.V. | Manufacture of vitamin b6 |
| JP4752026B2 (ja) * | 2005-03-04 | 2011-08-17 | 国立大学法人高知大学 | 新規なピリドキサール4−デヒドロゲナーゼ及びその利用 |
| WO2007142222A1 (ja) | 2006-06-09 | 2007-12-13 | Daiichi Fine Chemical Co., Ltd. | ピリドキサミン製造方法 |
| TW200914044A (en) * | 2007-07-12 | 2009-04-01 | Daiichi Sankyo Co Ltd | Anti human Sulf1 antibody |
| JP2009291158A (ja) * | 2008-06-06 | 2009-12-17 | Daiichi Fine Chemical Co Ltd | ピリドキサミン−ピルビン酸アミノトランスフェラーゼの変異酵素 |
| JP2014139139A (ja) * | 2011-04-28 | 2014-07-31 | Igaku Seibutsugaku Kenkyusho:Kk | ヒトテロメレース逆転写酵素に対するモノクローナル抗体 |
| JP6799910B2 (ja) | 2015-11-20 | 2020-12-16 | 旭化成株式会社 | 封止材用エポキシ樹脂組成物、及び封止材。 |
| JP2017095573A (ja) | 2015-11-20 | 2017-06-01 | 出光興産株式会社 | 印刷用インク、印刷膜、及びタッチパネル |
-
2018
- 2018-05-11 US US16/611,053 patent/US20200123581A1/en not_active Abandoned
- 2018-05-11 JP JP2019517729A patent/JP6964314B2/ja active Active
- 2018-05-11 EP EP18798161.8A patent/EP3623472A4/en not_active Withdrawn
- 2018-05-11 WO PCT/JP2018/018413 patent/WO2018207931A1/ja not_active Ceased
- 2018-05-11 CN CN201880029703.3A patent/CN110678546A/zh active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| US20200123581A1 (en) | 2020-04-23 |
| JPWO2018207931A1 (ja) | 2020-03-19 |
| EP3623472A1 (en) | 2020-03-18 |
| WO2018207931A1 (ja) | 2018-11-15 |
| CN110678546A (zh) | 2020-01-10 |
| EP3623472A4 (en) | 2021-01-27 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN113969269B (zh) | D-氨基酸氧化酶突变体及其在制备l-草铵膦中的应用 | |
| JP2017529847A (ja) | アクリル酸含有量がより少ないアミド化合物を産生する手段および方法 | |
| EP1756291B1 (en) | Enzymatic decarboxylation of 2-keto-l-gulonic acid to produce xylose | |
| JP5320692B2 (ja) | 酵母及びl−乳酸の製造方法 | |
| CN102197141A (zh) | 制备光学活性胺衍生物的方法 | |
| CN106868030A (zh) | 重组载体、含有其的工程菌及在产α-酮戊二酸的应用 | |
| WO2003027301A1 (fr) | Procede permettant de produire un alcool au moyen d'un micro-organisme | |
| CN118480524B (zh) | 一株l-苏氨酸转醛酶突变体及其高效合成氯霉素中间体 | |
| WO2005106005A1 (ja) | ヒドロキシカルボン酸類の生産方法 | |
| JP6964314B2 (ja) | 組換え微生物、組換え微生物を用いたピリドキサミン又はその塩の製造方法、及び組換え微生物を用いたピリドキサール又はその塩の製造方法 | |
| CN113969268A (zh) | Glu/Leu/Phe/Val脱氢酶突变体及其在制备L-草铵膦中的应用 | |
| TWI500767B (zh) | 高產量之異丙醇生產細菌 | |
| JP4809660B2 (ja) | 3−キヌクリジノン還元酵素およびこれを用いる(r)−3−キヌクリジノールの製造方法 | |
| JP4663631B2 (ja) | 放線菌由来のampデアミナーゼ及びその利用 | |
| CN110628835A (zh) | 一种利用pH自平衡催化体系制备木糖醇的方法 | |
| JP2011177029A (ja) | 光学活性なアミン誘導体の製造方法 | |
| JP2005278414A (ja) | 1,3−プロパンジオール及び3−ヒドロキシプロピオン酸を製造する方法 | |
| US20200149079A1 (en) | Recombinant microorganism and method of producing pyridoxamine or salt thereof using recombinant microorganism | |
| CN109897872B (zh) | 酶法制备(2s,3s)-n-叔丁氧羰基-3-氨基-1-氯-2-羟基-4-苯基丁烷 | |
| CN105051200A (zh) | 用于产生(r)-3-奎宁环醇的生物催化方法 | |
| CN120624386B (zh) | 一种醇脱氢酶突变体及其合成覆盆子酮的方法 | |
| US20050239180A1 (en) | Nucleotide sequence coding for a mannitol-2 dehydrogenase and method for the production of d-mannitol | |
| KR20200068144A (ko) | 디카르복시산 생합성 관련 효소 및 이를 이용한 디카르복시산 생산방법 | |
| JPWO2005123921A1 (ja) | 新規グリセロール脱水素酵素、その遺伝子、及びその利用法 | |
| CN121729482A (zh) | 直链型菊粉合成菌和直链型菊粉的制造方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20191108 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20191108 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20201027 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20201217 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210216 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20210330 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210519 |
|
| C60 | Trial request (containing other claim documents, opposition documents) |
Free format text: JAPANESE INTERMEDIATE CODE: C60 Effective date: 20210519 |
|
| C11 | Written invitation by the commissioner to file amendments |
Free format text: JAPANESE INTERMEDIATE CODE: C11 Effective date: 20210601 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20210810 |
|
| C21 | Notice of transfer of a case for reconsideration by examiners before appeal proceedings |
Free format text: JAPANESE INTERMEDIATE CODE: C21 Effective date: 20210817 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20210921 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20211011 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6964314 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| S531 | Written request for registration of change of domicile |
Free format text: JAPANESE INTERMEDIATE CODE: R313531 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |