KR20150132096A - 플러스-센스 단일 가닥 rna 바이러스 벡터 및 킷트를 사용하여 폴리펩티드를 생산하기 위한 방법 - Google Patents
플러스-센스 단일 가닥 rna 바이러스 벡터 및 킷트를 사용하여 폴리펩티드를 생산하기 위한 방법 Download PDFInfo
- Publication number
- KR20150132096A KR20150132096A KR1020157020304A KR20157020304A KR20150132096A KR 20150132096 A KR20150132096 A KR 20150132096A KR 1020157020304 A KR1020157020304 A KR 1020157020304A KR 20157020304 A KR20157020304 A KR 20157020304A KR 20150132096 A KR20150132096 A KR 20150132096A
- Authority
- KR
- South Korea
- Prior art keywords
- sense single
- virus
- plus
- viral vector
- stranded rna
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Withdrawn
Links
Images















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8202—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation by biological means, e.g. cell mediated or natural vector
- C12N15/8205—Agrobacterium mediated transformation
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/00041—Use of virus, viral particle or viral elements as a vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/00041—Use of virus, viral particle or viral elements as a vector
- C12N2770/00043—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/40011—Tymoviridae
- C12N2770/40041—Use of virus, viral particle or viral elements as a vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/40011—Tymoviridae
- C12N2770/40041—Use of virus, viral particle or viral elements as a vector
- C12N2770/40043—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
Landscapes
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Zoology (AREA)
- Organic Chemistry (AREA)
- Chemical & Material Sciences (AREA)
- Molecular Biology (AREA)
- General Engineering & Computer Science (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Microbiology (AREA)
- Plant Pathology (AREA)
- Cell Biology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Physics & Mathematics (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
본 발명은 플러스-센스 단일 가닥 RNA 바이러스 벡터들을 포함하는 킷트들과 마찬가지로 이들 벡터들의 혼합물들 및 이들의 용도, 및 킷트들과 벡터들을 사용하여 둘 또는 그 이상의 폴리펩티드들을 동시적으로 식물 또는 식물 조직 또는 식물 세포 내에서 생산하기 위한 방법에 관한 것이다.
Description
본 발명은 플러스-센스 단일 가닥 RNA 바이러스 벡터들을 포함하는 킷트들과 마찬가지로 이들 벡터들의 혼합물들 및 이들의 용도, 및 킷트들과 벡터들을 사용하여 둘 또는 그 이상의 폴리펩티드들을 동시적으로 식물 또는 식물 조직 또는 식물 세포 내에서 생산하기 위한 방법에 관한 것이다.
식물들 내에서의 단백질들의 재조합 생산을 위하여, 바이러스 벡터들의 사용이 유전자 이식 식물들의 생산에 대한 장래성 있는 대안을 대표한다. 효율적인 복제로 인하여 바이러스 벡터들은 그들의 유전체들이 식물 세포들 내에 높은 농도들로 존재함에 따라 대상의 단백질들의 높은 수율을 허용한다. 게다가, 숙주 식물의 특정의 발달 단계가 수행되고 그리고 보다 높은 생물질(biomass)이 획득가능하게 될 때까지 감염 및 후속의 생산을 기다리는 것이 가능하다. 그러나, 동일 세포 내에서의 서로 다른 재조합 단백질들의 생산은 동일한 유전체로부터 파생된 서로 다른 바이러스 벡터들을 사용하는 경우에 문제를 나타낸다. 이는 특히 하나 이상의 폴리펩티드쇄들을 포함하는 복합 단백질들(complex proteins)이 생산되어야 하는 경우에 문제가 된다. 2개의 서로 다른 외래 단백질들(foreign proteins)에 대한 코딩 시퀀스(coding sequences)들을 수반하는 하나의 바이러스 종(viral species)들로부터 파생된 2개의 벡터들이 하나의 식물 내에 공-접종(co-inoculated)되는 경우, 감염의 과정 동안에 2개의 바이러스 벡터들이 공간적으로 분리된다. 이러한 딜레마를 도 1에 나타내었다. 단지 원래 감염된 세포들의 영역 내에서, 두 벡터들이 검출될 수 있는 일부 세포들이 발견될 수 있다. 전신적으로(systemically) 감염된 식물들 내에서, 벡터들 중의 단지 하나가 존재할 수 있거나, 또는 단지 하나 또는 다른 하나의 벡터가 존재하는 영역들이 발견될 수 있다. 따라서, 바이러스 벡터에 의하여 동일 식물 세포 내에서의 서로 다른 단백질들의 발현은 여전히 도전의 과제이다. 게다가, 많은 식물 바이러스들이 기계적으로 또는 곤충들의 도움으로 전염된다. 따라서, 식물들을 전신적으로 감염시킬 수 있는 능력을 갖는 전체 바이러스 유전체들은 환경 내로 해방될 수 있다.
상기 문제점들을 극복하기 위한 현존하는 접근법들은 매우 신속하고 그리고 높은 생산을 목표로 하는 담배모자이크바이러스(TMV: Tobacco Mosaic Virus) 및 감자 바이러스 X(PVX: Potato Virus X)와 같은 비-경쟁적(non-competitive) 바이러스 벡터들을 사용한다(문헌 US 2009/0111145 A1; EP 1 686 176 A1)(도 2 참조). 그러나, 보다 복잡한 단백질들을 생산하기 위해서는, 부가의 비-경쟁적 바이러스 벡터들이 필요하게 될 것이며, 이는 감염된 식물에 대한 별도의 부담을 나타낼 수 있다.
따라서, 본 발명의 목적은 식물 세포들 내에서 동시적으로 둘 또는 그 이상의 폴리펩티드들의 재조합 생산을 허용하고 그리고 동시에 생물안정성 봉쇄(biosafety containment)를 확실하게 하는 획득가능한 킷트들 및 방법들을 만드는 것이다.
하나의 구체예에 있어서, 상기 목적은
a) 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및
b) 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터를 포함하고,
여기에서
(i) 제1 플러스-센스 단일 가닥 바이러스 벡터 및 제2 플러스-센스 단일 가닥 바이러스 벡터들이 서로 다른 식물 바이러스들로부터 파생되고, 그리고
(ii) 제1 벡터가 파생되는 바이러스의 피복 단백질(coat protein) ORF가 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 내에서 완전히 결실되고(deleted), 그리고
(iii) 제2 벡터가 파생되는 바이러스의 피복 단백질 ORF가 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터 내에서 완전히 결실되고, 그리고
(iv) 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터가 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF를 포함하고, 그리고
(v) 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터가 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF를 포함하고, 그리고
(vi) 제1 플러스-센스 단일 가닥 바이러스 벡터 및 제2 플러스-센스 단일 가닥 바이러스 벡터들이 식물 세포들 내에서 복제될 수 있는 RNA 복제단위(replicon)를 포함하는 킷트에 의해서 해결된다.
놀랍게도 이러한 제1 및 제2 벡터가 단일 세포들의 전신적인 공-감염을 가능하게 하고, 그리고, 실시예 1에서 나타난 바와 같이, 두 유전자 산물들이 제공된다.
피복 단백질 ORF의 완전한 결실 및 개개 다른 바이러스의 피복 단백질 ORF의 삽입으로 인하여, 전신적인 공-감염 및 공-발현을 허용하는 킷트가 제조된다. 동시에, 단일 벡터의 환경 내로의 해방(liberation) 이후 건강한 식물의 전신적 감염이 불가능하기 때문에 봉쇄 시스템이 수득된다. 조합된 전신적 감염으로 인하여, 전 식물 부분들이 개별적으로 감염되어야 할 필요가 없기 때문에 감염에 요구되는 장치들에 관한 복잡도(complexity)가 훨씬 낮다. 바람직하게는, 피복 단백질들이 호혜적으로 교환된다; 즉, 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 피복 단백질 ORF가 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF로 치환되고, 그리고 그 역으로도 같다. 이러한 치환이 전체 바이러스 유전체가 전체 바이러스 유전체 기구(viral genome organization)를 교란하지 않고 그리고 과도한 부가의 유전체 부하(genome load)를 생성하지 않기 때문에 이러한 치환이 바람직하다.
따라서, 킷트의 바람직한 구체예에 있어서, 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 피복 단백질 ORF가 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF로 치환된다.
보다 바람직한 구체예에 있어서, 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 피복 단백질 ORF가 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF로 치환된다.
상기 설명된 바와 같이, 본 발명의 킷트들은 비-경쟁적 바이러스 벡터(non-competitive viral vectors)들을 제공한다. 따라서, 또 다른 바람직한 구체예에 있어서, 제1 및 제2 플러스-센스 단일 가닥 바이러스 벡터들이 파생되는 바이러스들은 서로 다른 바이러스 형(virus types)들, 바람직하게는 서로 다른 바이러스 속(virus genera)들, 보다 바람직하게는 서로 다른 바이러스 과(virus families)들에 속한다.
비-경쟁적 TMV- 및 PVX-바이러스들을 이용하는 것이 특히 바람직하다. TMV- 및 PVX-바이러스들에 기초하는 이러한 킷트가 실시예 1에서 성공적으로 사용되었다.
TMV 유전체는 적어도 4개의 단백질들 즉: 126 및 183-kDa(킬로달톤) 복제효소 단백질(replicase proteins)들, 30-kDa 세포-대-세포 이동 단백질(MP: movement protein) 및 17.5-kDa 피복 단백질(CP: coat protein)들을 인코딩한다. MP 및 CP들은 하위 유전체 RNA들로 인코딩되며, 이들은 유전체 RNA의 3' 말단과 공-말단(co-terminal)들이다(도 2 참조).
상기 PVX 유전체는 적어도 5개의 단백질들 즉: RNA-의존성 RNA-중합효소(polymerase)(RdRp), 삼중 유전자 블럭 단백질들 1-3(triple gene block proteins 1-3; TGBp25, TGBp12, TGBp8) 및 25kDa 피복 단백질(CP)들을 인코딩한다. TGB 단백질들 및 CP들은 하위 유전체 RNA들로 인코딩되며, 이들은 유전체 RNA의 3' 말단과 공-말단들이다(도 2 참조).
포텍스바이러스(Potexviruses)들 및 토바모바이러스(Tobamoviruses)들은 플러스-센스 단일 가닥 유전체를 수반하는 식물 RNA 바이러스들이다. 이들 유전체들은 모노파르티타(monopartite)이다.
따라서, 본 발명의 바이러스 벡터들은 RNA 복제단위이거나 또는 이를 포함하는 RNA일 수 있거나 또는 RNA 복제단위를 인코딩하는 DNA일 수 있다.
RNA 복제단위는 본 발명의 킷트들의 바이러스 벡터들의 인자들을 포함한다. 더욱이 유전인자들이 전형적으로 복제 및 발현을 위하여 복제단위 상에 제공될 수 있다. RNA 복제단위이기 위하여, 즉, 식물 세포 내에서의 자율적인 복제를 위하여는, RNA 복제단위가 RdRp 또는 그의 기능성 유도체를 인코딩한다.
RNA 복제단위는 바람직하게는 RdRp에 결합하기 위하여 그리고 RNA 복제단위를 복제하기 위하여 부가적으로 바이러스, 특히 토바모바이러스 또는 포텍스바이러스의 5'- 또는 3'-미번역 영역들 및 RNA 복제단위의 5'- 또는 3'-미번역 영역들 내에 프로모터-시퀀스(promoter-sequences)들을 갖는다.
상기 RNA 복제단위는, 적용가능한 경우, 부가적으로 CP(피복 단백질), MP(이동 단백질(들)) 또는 TGB(삼중 유전체 블럭 단백질들) 또는 기능성 이종 ORF 단백질(functional heterologous ORF proteins)들의 발현을 위하여 하위 유전체 프로모터들을 가질 수 있다.
만일 바이러스 벡터가 DNA인 경우, 이는 전형적으로 생체외(in vitro) 또는 식물들 내에서의 생체내(in vivo)에서의 RNA 복제단위의 전사에 의한 감염성 RNA 생성을 허용하기 위한 프로모터를 가질 것이다. 식물들 내에서 DNA로부터의 RNA 복제단위의 전사를 허용하기 위한 프로모터의 하나의 실시예는 식물 생명공학에서 광범위하게 사용되는 콜리플라워모자이크바이러스(Cauliflower mosaic virus)의 35S 프로모터이다.
바이러스 벡터들에서 사용된 RdRp는 그 시퀀스가 원래의 RdRp의 적어도 50%, 바람직하게는 적어도 60%, 보다 바람직하게는 적어도 80%, 그리고 가장 바람직하게는 적어도 90%의 시퀀스 동일성(sequence identity)을 갖는 단백질을 인코딩하는 한 바이러스성 RdRp의 기능성 변종(functional variant)으로 고려될 수 있다.
본 발명의 바이러스 벡터들을 공학화(engineering) 하기 위하여 사용될 수 있는 플러스-센스 단일 가닥 RNA 바이러스들의 목록이 이하에서 제공된다:
과: 브로모비리대(Bromoviridae),
속: 알파모바이러스(Alfamovirus), 모식종(Type species): 알팔파모자이크바이러스(alfalfa mosaic virus),
속: 아일라바이러스(Ilarvirus), 모식종: 담배줄무늬바이러스(tobacco streak virus),
속: 브로모바이러스(Bromovirus), 모식종: 브롬모자이크바이러스(brome mosaic virus),
속: 쿠쿠모바이러스(Cucumovirus), 모식종: 오이모자이크바이러스(cucumber mosaic virus);
과: 클로스테로비리대(Closteroviridae),
속: 클로스테로바이러스(Closterovirus), 모식종: 사탕무황화바이러스(beet yellows virus),
속: 크리니바이러스(Crinivirus), 모식종: 상추감염성황화바이러스(Lettuce infectious yellows virus),
과: 코모비리대(Comoviridae),
속: 코모바이러스(Comovirus), 모식종: 동부모자이크바이러스(cowpea mosaic virus),
속: 파바바이러스(Fabavirus), 모식종: 잠두위조바이러스 1(broad bean wilt virus 1),
속: 네포바이러스(Nepovirus), 모식종: 담배둥근무늬바이러스(tobacco ringspot virus);
과: 포티비리대(Potyviridae),
속: 포티바이러스(Potyvirus), 모식종: 감자 바이러스 Y(potato virus Y), 자두발진바이러스(plum pox virus); 담배식각바이러스(tobacco etch virus); 토끼풀황화정맥바이러스(clover yellow vein virus); 담배정맥반점형성바이러스(tobacco vein mottling virus);
속: 라이모바이러스(Rymovirus), 모식종: 독보리모자이크바이러스(ryegrass mosaic virus),
속: 바이모바이러스(Bymovirus), 모식종: 보리황화모자이크바이러스(barley yellow mosaic virus);
과: 세퀴비리대(Sequiviridae),
속: 세퀴바이러스(Sequivirus), 모식종: 파스닙황화얼룩바이러스(parsnip yellow fleck virus),
속: 와이카바이러스(Waikavirus), 모식종: 벼툰그로구형바이러스(rice tungro spherical virus);
과: 톰부스비리대(Tombusviridae),
속: 카르모바이러스(Carmovirus), 모식종: 카네이션반점바이러스(carnation mottle virus),
속: 디안토바이러스(Dianthovirus), 모식종: 카네이션둥근무늬바이러스(carnation ringspot virus),
속: 마클로바이러스(Machlomovirus), 모식종: 옥수수백색반점바이러스(maize chlorotic mottle virus),
속: 네크로바이러스(Necrovirus), 모식종: 담배괴사바이러스(tobacco necrosis virus),
속: 톰부스바이러스(Tombusvirus), 모식종: 토마토덤불성장위축바이러스(tomato bushy stunt virus),
ssRNA 바이러스들의 미할당 속들:
속: 카필로바이러스(Capillovirus), 모식종: 사과가지천공바이러스(apple stem grooving virus);
속: 칼라바이러스(Carlavirus), 모식종: 카네이션잠재바이러스(carnation latent virus);
속: 에나모바이러스(Enamovirus), 모식종: 완두융기생장모자이크바이러스(pea enation mosaic virus),
속: 퓨로바이러스(Furovirus), 모식종: 토양전염성 밀모자이크바이러스(soil-borne wheat mosaic virus),
속: 호르데이바이러스(Hordeivirus), 모식종: 보리반엽모자이크바이러스(barley stripe mosaic virus),
속: 이대오바이러스(Idaeovirus), 모식종: 산딸기덤불위축바이러스(raspberry bushy dwarf virus);
속: 루테오바이러스(Luteovirus), 모식종: 보리황화위축바이러스(barley yellow dwarf virus);
속: 마라피바이러스(Marafivirus), 모식종: 옥수수희박줄무늬바이러스(maize rayado fino virus);
속: 포텍스바이러스(Potexvirus), 모식종: 감자 바이러스 X(potato virus X);
속: 소베모바이러스(Sobemovirus), 모식종: 서던빈모자이크바이러스(Southern bean mosaic virus),
속: 테누이바이러스(Tenuivirus), 모식종: 벼줄무늬바이러스(rice stripe virus),
속: 토바모바이러스(Tobamovirus), 모식종: 담배모자이크바이러스(tobacco mosaic virus),
속: 토브라바이러스(Tobravirus), 모식종: 담배얼룩바이러스(tobacco rattle virus),
속: 트리코바이러스(Trichovirus), 모식종: 사과잎반점바이러스(apple chlorotic leaf spot virus);
속: 티모바이러스(Tymovirus), 모식종: 무황색모자이크바이러스(turnip yellow mosaic virus);
속: 움브라바이러스(Umbravirus), 모식종: 당근반점바이러스(carrot mottle virus);
따라서, 또 다른 바람직한 구체예에 있어서, 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스는 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이다.
또 다른 바람직한 구체예에 있어서, 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스는 토바모바이러스, 특히 TMV 바이러스(담배모자이크바이러스)이다.
실시예 1에서, 제1 및 제2 플러스-센스 단일 가닥 바이러스 벡터들은 이종 ORF를 포함한다. 따라서, 이러한 제1 및 제2 플러스-센스 단일 가닥 바이러스 벡터들을 사용하여 하나의 식물 세포 내에서 2개의 서로 다른 이종 단백질들을 생산하는 것이 가능하다(도 3 참조). 실시예 1에서 사용된 바와 같이, 바람직한 벡터들은 구조 PVX-CP(TMV)-ORF 및 TMV-CP(PVX)-ORF를 갖는다.
그러나, 이들 벡터들이 이러한 이종 ORF들을 포함할 것이 요구되는 것은 아니다. 오히려, 플러스-센스 단일 가닥 바이러스 벡터들이 하나의 바람직한 구체예에 있어서 단지 상보 벡터(complementing vectors)들로서 사용될 수 있다. 이러한 구체예에 있어서 벡터들 둘 중의 어느 하나 또는 둘 다는 이종 ORF를 포함하지 않는다. 바람직한 바이러스 벡터 조합들은 하기의 구조들 즉: (a) PVX-CP(TMV) 및 TMV-CP(PVX); (b) PVX-CP(TMV)-ORF 및 TMV-CP(PVX); (c) PVX-CP(TMV) 및 TMV-CP(PVX)-ORF 및 PVX-CP(TMV)-ORF 및 TMV-CP(PVX)-ORF을 갖는 벡터 조합(vector combinations)들이다(도 3에 나타낸 바와 같음).
따라서, 본 발명의 다른 바람직한 구체예에 있어서, 제1 플러스-센스 단일 가닥 바이러스 벡터 및/또는 제2 플러스-센스 단일 가닥 바이러스 벡터는 기능성 이종 ORF를 포함한다. ORF는 동일하거나 또는 서로 다를 수 있으며, 바람직하게는 서로 다를 수 있다. 이러한 구체예에 있어서, 벡터들이 상보 벡터들로서 사용되고 그리고 거기에 더해 대상의 이종 ORF를 발현하는 데 사용된다.
식물 내 또는 식물 조직 내에서 바이러스 벡터들로부터 발현되는 것이기 위해서는, 기능성 이종 ORF를 포함하는 바이러스 벡터들은 하나 또는 그 이상의 하위 유전체 프로모터들 및 리보솜 결합부위(ribosome binding site) 및/또는 내부리보솜진입부위(IRES: internal ribosome entry site) 등고 같은 발현에 요구되는 다른 시퀀스들을 포함한다.
따라서 본 발명의 다른 바람직한 구체예에 있어서, 제1 플러스-센스 단일 가닥 바이러스 벡터 및/또는 제2 플러스-센스 단일 가닥 바이러스 벡터는 기능성 이종 ORF를 포함하지 않는다. 이 구체예에 있어서, 킷트는 상보 바이러스 벡터 킷트로서 사용되고, 이는 이하에서 기술되는 바와 같이 피복 단백질을 포함하지 않는, 기능성 이종 ORF(들)을 포함하는 부가의 바이러스 벡터(들)를 부가하는 데 사용될 수 있다.
놀랍게도, 상보 벡터쌍 PVX-CP(TMV)-ORF 및 TMV-CP(PVX)-ORF 및 피복-단백질-결핍 벡터(coat-protein-deficient vector) PVX-ΔCP-ORF를 사용하여 실시예 1에서 기술되는 바와 같은 모든 ORF들의 발현을 달성하는 것이 가능하다는 것이 발견되었다. 따라서, 놀랍게도 상보 벡터들 및 이종 ORF(들)을 운반하는 하나 또는 그 이상의 부가의 벡터들에 기초하여 부가의 이종 ORF들이 동일한 식물 세포들 내에서 발현될 수 있다는 것이 발견되었으며, 여기에서 부가의 벡터들은 피복 단백질의 완전 결실을 갖는다. 완전 결실로 인하여, 바이러스의 유전체 부하가 상당히 낮아지고 그리고 부가의 벡터들의 피복 단백질 결핍은 부가의 봉쇄를 포함한다(도 3 및 도 4).
"PVX-CP(TMV)-ORF"는 PVX에 기초하는 바이러스 벡터로서 이해되고, 여기에서 원래의 피복 단백질은 TMV의 피복 단백질로 치환되고, 그리고 여기에서 벡터는 기능성 이종 ORF를 포함한다.
"TMV-CP(PVX)-ORF"는 TMV에 기초하는 바이러스 벡터로서 이해되고, 여기에서 원래의 피복 단백질은 PVX의 피복 단백질로 치환되고, 그리고 여기에서 벡터는 기능성 이종 ORF를 포함한다.
"PVX-ΔCP-ORF"는 PVX에 기초하는 바이러스 벡터로서 이해되고, 여기에서 원래의 피복 단백질 ORF는 완전히 결실되고, 그리고 여기에서 벡터는 기능성 이종 ORF를 포함한다(또한 도 4 참조).
"PVX-ΔCP-ΔMPs-ORF"는 PVX에 기초하는 바이러스 벡터로서 이해되고, 여기에서 원래의 피복 단백질 ORF는 완전히 결실되고, 그리고 여기에서 바이러스 벡터는 기능성 이동 단백질들이 존재하지 않고, 그리고 여기에서 벡터는 기능성 이종 ORF를 포함한다(또한 도 5 참조). 바람직한 구체예에 있어서, 이동 단백질들은 완전히 결실된다.
"PVX-ORF"는 PVX에 기초하는 바이러스 벡터로서 이해되고, 여기에서 벡터는 기능성 이종 ORF를 포함한다.
"TMV-ORF"는 TMV에 기초하는 바이러스 벡터로서 이해되고, 여기에서 벡터는 기능성 이종 ORF를 포함한다.
"기능성 이종 ORF(functional heterologous ORF)"는 오픈리딩프레임(open reading frame ; 개방형 해독틀)로서 이해되고, 이는 개개 미변성된 바이러스 내에는 존재하지 않고 그리고 이는 발현되어 기능성 유전자 산물을 수득할 수 있다. 이러한 유전자 산물은 RNA 또는 단백질일 수 있고, 바람직하게는 단백질일 수 있다. 바람직하게는, 기능성 이종 ORF는 사용된 모든 바이러스 벡터들에 대하여 이종이다. 전형적으로 기능성 이종 ORF는 적절한 하위 유전체 프로모터들 또는 리보솜-결합 부위들 또는 IRES의 통제 하에 있다.
따라서, 하나의 특히 바람직한 구체예에 있어서, 이 부가의 바이러스 벡터는 하기의 특징들을 갖는 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터이다:
특히 바람직한 구체예에 있어서, 킷트는 부가적으로
a) (i) 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)은 적어도 하나의 기능성 이종 ORF를 포함하고, 그리고
(ii) 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)은 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터와 동일한 식물 바이러스로부터 파생되고, 그리고
(iii)
적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 파생되는 바이러스의 피복 단백질 ORF가 적어도 하나의 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터(들) 내에서 완전히 결실되는
적어도 하나의 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터 및/또는
b) (i) 적어도 하나의 부가의 플러스-센스 단일 가닥 바이러스 벡터(들)가 적어도 하나의 기능성 이종 ORF를 포함하고, 그리고
(ii) 적어도 하나의 부가의 플러스-센스 단일 가닥 바이러스 벡터(들)가 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터와 동일한 식물 바이러스로부터 파생되는
적어도 하나의 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터를 포함한다.
게다가, 부가적으로 제3 플러스-센스 단일 가닥 바이러스 벡터(들)의 이동 단백질(들)이 결실되고, 그에 의하여 추가로 바이러스 유전자 크기를 감소시키는 것이 가능하다.
따라서, 특히 바람직한 구체예에 있어서, 적어도 하나의 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)은 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 파생되는 바이러스의 기능성 이동 단백질 ORF(들)이 존재하지 않고, 바람직하게는 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 파생되는 바이러스의 기능성 이동 단백질 ORF(들)이 완전히 결실된다(또한 도 4 참조).
따라서, 바람직한 구체예에 있어서, 제3 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스는 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이고 그리고 제3 플러스-센스 단일 가닥 바이러스 벡터가 포텍스바이러스의 기능성 이동 단백질 ORF(들)이 존재하지 않는다.
실시예 1에 나타난 바와 같이, 제3 바이러스 벡터로서 PVX-ΔCP-ORF를 사용하여 상보화(complementation)가 달성되었다. 이러한 벡터는 본 발명에 따른 킷트를 위한 바람직한 벡터이다.
추가의 구체예에 있어서, PVX의 이동 단백질들이 결실될 수 있다. 이러한 벡터는 구조 PVX-ΔCP-ΔTGB-ORF를 갖는다.
이러한 결실의 생성이 실시예 1에서 기술된다.
따라서, 특히 바람직한 구체예에 있어서, 제3 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스는 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이다. 심지어 구조 PVX-ΔCP-ΔTGB-ORF 및/또는 PVX-ΔCP-ORF의 벡터들이 보다 바람직하다.
따라서, 본 발명의 특히 바람직한 구체예에 있어서, 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이고, 그리고 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스가 토바모바이러스, 특히 TMV 바이러스(담배모자이크바이러스)이고, 그리고 제3 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 것이 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이다.
심지어 보다 바람직한 구체예에 있어서, 킷트는
(A)
(i) 구조 PVX-CP(TMV) 및 TMV-CP(PVX)의 벡터들; 또는
(ii) 구조 PVX-CP(TMV)-ORF 및 TMV-CP(PVX)의 벡터들; 또는
(iii) 구조 PVX-CP(TMV) 및 TMV-CP(PVX)-ORF의 벡터들 또는
(iv) 구조 PVX-CP(TMV)-ORF 및 TMV-CP(PVX-ORF)의 벡터들 그리고
(B) 구조 PVX-ΔCP-ΔTGB-ORF 또는 PVX-ΔCP-ORF의 벡터
를 포함하거나 또는 내포한다.
실시예 1에서, 또한 상보 벡터들이 이종 ORF를 포함한다. 그러나 단지 상보 벡터들의 바이러스 벡터 크기를 감소시키기 위하여 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 이종 ORF를 포함하는 것 또한 가능하다. 또한, 과도한 바이러스 유전자 크기를 감소시키기 위하여 하나의 바이러스 벡터가 단지 하나의 이종 ORF를 포함하는 것이 바람직하다.
또 다른 바람직한 구체예에 있어서, 적어도 하나의 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)은 기술된 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터의 특징들을 포함하고, 그리고 적어도 하나의 기능성 이종 ORF를 포함하고, 여기에서 벡터들 중의 하나 또는 둘 다 기능성 이종 ORF(들)을 포함하는 경우에서 적어도 하나의 기능성 이종 ORF가 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터의 기능성 이종 ORF(들)과는 다르다.
이러한 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)은 제1 또는 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터 내의 기능성 이종 ORF를 치환(기능성 이종 ORF가 존재하는 경우에) 하거나 또는 도입(기능성 이종 ORF가 존재하지 않는 경우에) 하는 것에 의하여 수득가능하다.
따라서, 바람직한 구체예에 있어서 바람직하게는 적어도 하나의 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)은 기능성 이종 ORF에 의하여 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터와는 다르다.
또 다른 바람직한 구체예에 있어서, 적어도 하나의 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)은 제1 플러스-센스 단일 가닥 바이러스 벡터 또는 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 것과 동일한 바이러스로부터 파생되고, 그리고 적어도 하나의 기능성 이종 ORF를 포함한다. 특히 바람직한 구체예에 있어서, 이러한 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터를 수득하기 위하여 적어도 하나의 기능성 이종 ORF를 도입하는 데 야생형 바이러스 벡터(wt viral vector)가 사용될 수 있다. 다른 특별히 바람직한 구체예에 있어서, 적어도 하나의 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)은 제1 플러스-센스 단일 가닥 바이러스 벡터 또는 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 피복 단백질, 이동 단백질(들) 및 RNA-의존성 RNA 중합효소 그리고 적어도 하나의 기능성 이종 ORF를 포함한다. 이러한 적어도 하나의 기능성 이종 ORF는 제1 플러스-센스 단일 가닥 바이러스 벡터 또는 제2 플러스-센스 단일 가닥 바이러스 벡터 내에서와 동일하거나 또는 서로 다른 기능성 이종 ORF일 수 있으며(이들이 이러한 기능성 이종 ORF를 포함하는 경우), 바람직하게는 서로 다를 수 있다.
따라서, 바람직하게는, 2 또는 그 이상의 서로 다른 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터들은 서로 다른 기능성 이종 ORF(들)을 포함한다.
추가의 바람직한 구체예에 있어서, 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)은 하나의 기능성 이종 ORF를 포함한다.
상기 바이러스 벡터들은 천연(natural)의 플러스-센스 단일 가닥 바이러스 상에, 예를 들면, 이러한 바이러스, 특히 토바모바이러스 또는 포텍스바이러스로부터의 유전 성분들을 사용하는 것에 의하여 구축될 수 있다. 본 발명의 바이러스 벡터들 또는 본 발명의 킷트들은 이종 ORF 시퀀스들을 바이러스를 인코딩하는 바이러스 벡터들 내로 삽입하고 그리고 기술한 바와 같은 CP를 인코딩하는 시퀀스를 치환하는 것에 의하여 수득될 수 있다. 그러나, 포텍스바이러스의 RdRp에 대하여 또는 포텍스바이러스의 5' 또는 3' 미-번역된 영역들에 대한 것과 같이 천연의 바이러스의 여러 유전 성분들에 대하여 여러 변형들이 이루어질 수 있다.
본 발명의 킷트들의 바이러스 벡터들은 혼합되어 이러한 바이러스 벡터들의 혼합물들을 수득할 수 있다. 이러한 혼합물들은, 예를 들면, 건조되거나 또는 동결-건조된 고체 등과 같은 고체 형태 또는 액체 형태, 예를 들면, 수용액(aqueous solution), 현탁액(suspension) 또는 분산액(dispersion) 또는 겔(gel)과 같은 것일 수 있다. 이러한 혼합물들은 식물, 식물 세포들 또는 식물 조직들을 감염시키는 데 사용될 수 있다.
따라서, 본 발명은 또한 하기의 벡터들 a) 내지 c)들 즉:
a) 앞서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터,
b) 앞서 정의된 바와 같은 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터,
c) 앞서 정의된 바와 같은 적어도 하나의 부가의 및/또는 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터
중의 2, 3 또는 그 이상의 서로 다른 벡터들을 포함하는 혼합물에 관한 것이다.
실시예 1에서, PVX-ΔCP-ORF, PVX-CP(TMV)-ORF 및 TMV-CP(PVX)-ORF의 혼합물이 감염을 위하여 성공적으로 사용되었다.
바람직한 구체예에 있어서, 혼합물은
(i) 앞서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터, 및
(ii) 앞서 정의된 바와 같은 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터, 및
(iii) 앞서 정의된 바와 같은 선택적으로 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터
를 포함한다.
추가의 바람직한 구체예에 있어서, 혼합물은
(i) 앞서 정의된 바와 같은 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터, 및
(ii) 앞서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및/또는 앞서 정의된 바와 같은 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터
를 포함한다.
본 발명의 킷트들 및 혼합물들은 앞서 정의된 바와 같은 벡터들로 식물들 또는 식물 세포들을 성공적으로 감염시키는 데 사용될 수 있다.
따라서 본 발명은 또한 앞서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터 및 앞서 정의된 바와 같은 선택적으로 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)를 포함하는 식물 또는 식물 세포에 관한 것이다.
최초로, 식물 RNA 바이러스 벡터들을 사용하여 하나 이상의 조직 내에서 하나 이상의 식물 세포 내에서 동시적으로 전신적으로 벡터들의 서로 다른 이종 ORF들의 동시의 공-발현을 수득하는 것이 이제 가능하다.
따라서, 아마도 하나 이상의 조직 내에서 하나 이상의 식물 세포가 앞서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 벡터 및 선택적으로 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)를 포함한다.
따라서, 다른 바람직한 구체예에 있어서, 하나 이상의 조직 내의 하나 이상의 식물 세포는 여기에서 정의된 바와 같은 적어도 2개의 서로 다른 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)의 ORF들에 의하여 인코딩되는 이종 폴리펩티드들을 포함한다.
본 발명의 킷트들 및 혼합물들은 이종 단백질들의 발현을 위하여 사용될 수 있다. 특히, 킷트들은 상업적 응용을 위한 바이러스 벡터들을 사용하여 최초로 전신적으로 감염된 식물들 내에서 헤테로올리고머 단백질(heterooligomeric proteins)들의 봉쇄 생산(contained production)을 허용한다.
따라서, 본 발명은 하나의 구체예에 있어서 적어도 하나의 식물 세포에
(i) 여기에서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터, 및
(ii) 여기에서 정의된 바와 같은 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터, 및
(iii) 여기에서 정의된 바와 같은 선택적으로 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터
를 제공하는 것을 포함하며, 여기에서
a) 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 기능성 이종 ORF를 포함하지 않는 경우, 적어도 2개의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 적어도 하나의 식물 세포에 제공되고, 그리고
b) 여기에서 (i) 내지 (iii)에 따른 바이러스 벡터들 중의 적어도 2개가 서로 다른 이종 ORF들을 포함하는
식물 또는 식물 조직 또는 식물 세포 내에서 헤테로올리고머 폴리펩티드 및/또는 2 또는 그 이상의 폴리펩티드들을 생산하기 위한 방법에 관한 것이다.
특히 바람직한 구체예에 있어서, 방법에는 식물, 식물 조직 또는 식물 세포로부터 헤테로올리고머 폴리펩티드 및/또는 2 또는 그 이상의 폴리펩티드들을 단리하는 것이 후속된다.
식물들 또는 식물 세포들로부터 단백질들을 분리하기 위한 방법은 당해 기술분야에서 공지되어 있다. 하나의 방법에 있어서, 대상의 단백질은 국제공개특허공보 제WO 03/020938호에서 기술된 바와 같이 식물 세포벽 매질(apoplast)로부터 단리될 수 있다.
놀랍게도, 모든 벡터들로의 전신적 감염이 킷트들의 벡터들을 사용하여 달성될 수 있다. 따라서, 바람직한 구체예에 있어서, 식물의 전신적 감염이 달성된다.
상기 방법은 특히 헤테로올리고머 단백질들을 생산하는 데 유용하며, 이는 이는 하나의 세포 내에서 생산되어야 하고 그리고 이러한 세포 내에서 조립되어 기능성 단백질을 수득하여야 한다. 이러한 헤테로올리고머 단백질들의 예들로는 면역글로블린, 특히 항체 또는 항체 단편 또는 인터류킨-12 등과 같은 특정의 인터류킨들이 있다.
바이러스 벡터들의 제공(provision)을 위하여 식물 세포들 내에서의 바이러스 벡터들의 아그로박테리움-매개 전달(Agrobacterium-mediated delivery)이 사용될 수 있다. 프로토플라스트(protoplasts)들의 미세투사폭격(microprojectile bombardment), 전기천공(electroporation) 또는 피이지-매개 형질전환(PEG-mediated transformation)의 수단에 의한 핵산 시퀀스의 세포들 내로의 직접도입 등과 같은 식물들의 안정한 형질전환을 위하여 통상적으로 사용되는 여러 다른 방법들이 또한 식물 세포들 내로의 바이러스 벡터들의 제공을 위하여 사용될 수 있다. 아그로박테리움-매개 식물 형질전환이 바람직하다. 따라서, 아그로박테리움(US 5,591,616 ; US 4,940,838 ; US 5,464,763), 입자 또는 미세투사폭격(US5,100,792; EP00444882 B1; EP 00434616 B1)에 의해 운반되는 티아이-플라스미드 벡터(Ti-plasmid vector)에 의하는 것과 같은 여러 기술들로 식물 세포들 내로 이종 뉴클레오티드 시퀀스가 형질전환될 수 있다. 특히, 예를 들어 미세주입(microinjection)(WO 09/209696; WO 09/400583A1; EP 175966 B1), 전기천공(EP00564595 B1; EP00290395 B1; WO 08/706614 A1) 등 다른 식물 형질전환 방법들이 또한 사용될 수 있다. 형질전환 방법의 선택은 그 중에서도 형질전환되어야 할 식물 종들에 의존적이다. 예를 들면, 외떡잎 형질전환을 위하여는 미세투사폭격이 바람직한 반면에, 쌍떡잎에 대하여는, 아그로박테리움-매개 형질전환이 대체로 더 나은 결과들을 제공한다.
본 발명은 바람직하게는 보다 고등의 다-세포 식물들로 실행된다. 본 발명에서 사용되기에 바람직한 식물들에는 임의의 식물 종들이 포함되며 작물학적으로 그리고 원예학적으로 중요한 종들이 선호된다. 본 발명에서 사용되기 위한 통상의 농작물(crop plants)들에는 알팔파(alfalfa), 보리(barley), 콩류(beans), 카놀라(canola), 동부콩(cowpeas), 면(cotton), 옥수수(corn), 토끼풀(clover), 연(lotus), 렌틸콩(lentils), 루핀(lupine), 기장(millet), 귀리(oats), 완두콩(peas), 땅콩(peanuts), 벼(rice), 호밀(rye), 전동싸리(sweet clover), 해바라기(sunflower), 스위트피(sweetpea), 대두(soybean), 수수(sorghum) 트리티케일(triticale), 얌콩(yam beans), 벨벳콩(velvet beans), 살갈퀴(vetch), 밀(wheat), 등나무(wisteria) 및 견과 식물(nut plants)들이 포함된다. 본 발명을 실행하기에 선호되는 식물종들에는 대표적으로 화본과(Graminae), 국화과(Compositae), 가지과(Solanacea) 및 장미과(Rosaceae)들이 포함되나 이들로 제한되는 것은 아니다. 게다가, 특정된 것들과 마찬가지로 하기의 속들로부터의 식물들이 본 발명에서 사용하기에 바람직한 종들로는 애기장대속(Arabidopsis), 겨이삭속(Agrostis), 부추속(Allium), 금어초속(Antirrhinum), 셀러리속(Apium), 땅콩속(Arachis), 아스파라거스속(Asparagus), 아트로파속(Atropa), 귀리속(Avena), 대속(Bambusa), 유채속(Brassica), 참새귀리속(Bromus), 브로왈리아속(Browaalia), 동백나무속(Camellia), 대마속(Cannabis), 고추속(Capsicum), 병아리콩속(Cicer), 청명아주속(Chenopodium), 치코리움속(Chichorium), 감귤속(Citrus), 코페아속(Coffea), 의이인속(Coix), 오이속(Cucumis), 커큐비타속(Curcubita), 우산잔디속(Cynodon), 오리새속(Dactylis), 독말풀속(Datura), 당근속(Daucus), 디기탈리스속(Digitalis), 마속(Dioscorea), 기름야자속(Elaeis), 왕바랭이속(Eleusine), 김의털속(Festuca), 딸기속(Fragaria), 제라늄속(Geranium), 글라이신속(Glycine), 돼지감자속(Helianthus), 헤테로칼리스속(Heterocallis), 파라고무나무속(Hevea), 보리속(Hordeum), 사리풀잎속(Hyoscyamus), 고구마속(Ipomoea), 왕고들빼기속(Lactuca), 렌즈콩속(Lens), 나리속(Lilium), 아마속(Linum), 독보리속(Lolium), 연속(Lotus), 토마토속(Lycopersicon), 꿀풀속(Majorana), 꽃사과속(Malus), 망고속(Mangifera), 닥풀속(Manihot), 개자리속(Medicago), 아프리카해란초속(Nemesia), 담배속(Nicotiana), 잠두속(Onobrychis), 벼속(Oryza), 기장속(Panicum), 펠라고니움속(Pelargonium), 갈대속(Pennisetum), 페투니아속(Petunia), 완두속(Pisum), 강남콩속(Phaseolus), 큰조아재비속(Phleum), 포아풀속(Poa), 살구속(Prunus), 젓가락풀속(Ranunculus), 무속(Raphanus), 까치밥나무속(Ribes), 피마자속(Ricinus), 나무딸기속(Rubus), 사탕수수속(Saccharum), 샐피글로시스속(Salpiglossis), 호밀속(Secale), 세네시오속(Senecio), 강아지풀속(Setaria), 겨자속(Sinapis), 가지속(Solanum), 수수속(Sorghum), 잔디속(Stenotaphrum), 테오브로마속(Theobroma), 붉은토끼풀속(Trifolium), 호로파속(Trigonella), 소맥속(Triticum), 갈퀴속(Vicia), 포도속(Vigna), 포도속(Vitis), 옥수수속(Zea) 및 올리레아에속(Olyreae), 벼아과(Pharoideae) 및 많은 다른 속들이 있다.
바람직한 식물들로는 야생담배(Nicotiana benthamiana) 및 재배담배(Nicotiana tabacum) 등과 같은 담배종(Nicotiana species)들이 있고; 담배종들 이외의 바람직한 식물종들로는 페투니아(Petunia hybrida), 배추(Brassica campestris), 갓(B. juncea), 냉이(cress), 아루굴라(arugula), 겨자(mustard), 딸기(Strawberry), 시금치(spinach), 시금치딸기(Chenopodium capitatum), 알팔파(alfalfa), 상추(lettuce), 해바라기, 감자 및 오이들이 있다.
바람직한 포텍스바이러스들에는 PVX, 대나무모자이크바이러스(bamboo mosaic virus), 파파야모자이크바이러스(papaya mosaic virus), 비름모자이크바이러스(alternanthera mosaic virus), 토끼풀황화모자이크바이러스, 플랜틴바이러스 X(plantain virus X), 흰토끼풀모자이크바이러스(white clover mosaic virus) 및감자누른모자이크바이러스(potato aucuba mosaic virus), 특히 감자바이러스 X(PVX)들이 있다.
본 발명의 주요 응응예는 식물, 식물 잎 또는 식물 조직 또는 식물 배양물(plant culture) 내에서의 대상의 단백질의 생산이다. 본 발명의 공정이 식물들 내에서 수행되는 경우, 담배종과 같이 인간 또는 동물 먹이 사슬에 들어가지 않는 식물들이 바람직하다. 인간 또는 동물 먹이 사슬에 들어가지 않는 식물들은 노지에서 재배되고 그리고 바이러스 벡터들로 감염된 이후 특정의 기간 내에 수확될 수 있다. 바람직하게는, 전체 식물들 또는 식물 부분들이 봉쇄된 환경, 예를 들면, 온실 또는 소정의 수준의 발현을 제공하는 데 필요한 배양 기간에 대해 설계된 챔버에 한정될 수 있다.
다른 구체예에 있어서, 본 발명은 PVX 바이러스 벡터에 관한 것으로서, 여기에서
a) PVX 바이러스의 기능성 피복 폴리펩티드 ORF가 완전히 결실되고, 그리고
b) PVX 바이러스 벡터가 식물 세포들 내에서 복제할 수 있는 RNA 복제단위를 포함하고, 그리고
c) PVX 바이러스 벡터가 선택적으로 적어도 하나의 기능성 이종 ORF를 포함하고, 그리고
d) PVX 바이러스 벡터는 선택적으로 다른 바이러스의 기능성 피복 폴리펩티드 ORF를 포함한다.
상기 PVX 피복 단백질의 완전 결실이 성공적인 공-감염을 허용한다는 것이 발견되었다. 당해 기술분야의 현재 기술에 있어서, 벡터들에서의 피복 단백질의 결실들의 결과들을 가져왔고, 여기에서 피복 단백질의 3' 말단은 여전히 존재하였다. 이러한 벡터들이 문헌들 Komarova et al. (2006, Biochemistry (Mosc); 71: 846-850) 및 Tyulkina et al. (2011, Acta naturae; 3: 73-82)에서 개시되었다. 그러나, ORF의 이러한 부분 결실은 공감염을 달성하기에는 불충분하다. 따라서, 본 발명의 신규한 PVX 벡터들은 놀랍게도 공감염, 특히 상기한 본 발명의 킷트들의 상보 벡터들로의 공감염에 유용하다.
본 발명의 다른 구체예에 있어서, 환경의 오염의 위험 없이 식물들의 감염을 위하여 피복-단백질 결핍 바이러스 벡터들을 사용하는 것 또한 가능하다. 이 구체예에 있어서, 동일한 백본(backbone)을 갖는 적어도 2개의 바이러스 벡터들이 사용되며, 이들은 원래의 피복 단백질이 결핍된다. 피복 단백질 결핍으로 인하여, 전신적 감염 및/또는 환경의 오염이 불가능하다. 2 또는 그 이상의 서로 다른 이종 ORF들의 발현을 위한 동일한 백본은 벡터들의 간단한 구성을 허용한다. 게다가, 피복 단백질의 결실은 바이러스 유전자 크기의 과도한 증가 없이 이종 ORF의 도입을 허용한다. 게다가, 벡터들이 식물 세포들 내에서 복제될 수 있기 때문에, 아그로박테리움 투메파시엔스(A. tumefaciens) 세포들을 사용하는 단순한 일시적인 발현에 비하여 보다 높은 단백질 발현 수율들이 달성될 수 있다(실시예 2).
따라서, 다른 구체예에 있어서, 본 발명은 식물 또는 식물 조직 또는 식물 세포에 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들을 제공하는 것을 포함하고,
a) 여기에서 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 동일한 바이러스로부터 파생되고, 그리고
b) 여기에서 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 서로 다른 기능성 이종 ORF들을 포함하고, 그리고
c) 여기에서 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 상기 벡터들이 파생되는 바이러스의 기능성 피복 단백질 ORF가 결실되고, 그리고
d) 여기에서 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 식물 세포들 내에서 복제할 수 있는 RNA 복제단위를 포함하는
식물 또는 식물 조직 또는 식물 세포 내에서 헤테로올리고머 폴리펩티드 및/또는 2 또는 그 이상의 폴리펩티드들을 생산하는 방법에 관한 것이다.
특히 바람직한 구체예에 있어서, 피복 단백질들은 완전히 결실된다. 따라서, 바람직한 구체예에 있어서, 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스의 피복 단백질 ORF는 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들에서 완전히 결실된다.
다른 바람직한 구체예에 있어서, 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이다.
보다 더 바람직한 구체예에 있어서, 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스의 피복 단백질 ORF는 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들 내에서 완전히 결실되고, 그리고 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이다.
상기 벡터들을 식물 또는 식물 조직 또는 식물 세포에 제공하는 다양한 방법들이 공지되어 있다. 특히, 이들은 아그로감염(agroinfection)에 의해, DNA, 특히 바이러스 유전자의 cDNA를 포함하는 DNA로의 형질주입(transfection)에 의해, RNA, 특히 바이러스 유전자에 대응하는 RNA로의 형질주입에 의해 제공될 수 있다. 이러한 방법들은 예를 들면 문헌들 Fischer, R., et al. (2004; Plant-based production of biopharmaceuticals. Curr Opin Plant Biol 7, 152-158); Komarova, T.V. et al. (2010; Transient expression systems for plant-derived biopharmaceuticals. Expert Rev Vaccines 9, 859-876) 및 Lico, C. et al. (2012; The use of plants for the production of therapeutic human peptides. Plant Cell Rep 31, 439-451)에 기술되어 있다.
달리, 바이러스 유전자들의 복제된 cDNA들이 생체외에서 RNA를 전사하는 데 사용될 수 있으며, 계속해서 이는 감염에 사용될 수 있다.
달리, 바이러스 유전자들의 복제된 cDNA들은 플라스미드들 또는 DNA 단편 등과 같은 적절한 벡터 구조물들 내로 복제될 수 있으며, 이들은 유전자의 전사를 허용한다. 특히, 이러한 구조물은 적절한 프로모터 및 종결부위(terminator) 시퀀스들을 포함한다.
아그로감염의 경우에 있어서, 바이러스 유전자의 cDNA는 T-DNA 내로 도입되고, 계속해서 이는 식물들에 제공된다. 다시 이러한 T-DNA는 추가로 조절 DNA 성분들을 포함하며, 이는 유전자의 전사를 허용한다. 특히, 이러한 T-DNA 구조는 적절한 프로모터 및 종결부위 시퀀스들을 포함한다.
서로 다른 기능성 이종 ORF들을 포함하는 개개 바이러스 벡터들을 포함하는 서로 다른 아그로박테리움 세포 배양물들이 혼합될 수 있고 그리고 감염을 위하여 식물들에 투여될 수 있다.
따라서, 하나의 구체예에 있어서, 본 발명은 아그로박테리움 세포들, 특히 아그로박테리움 투메파시엔스(Agrobacterium tumefaciens) 세포들의 혼합물에 관한 것이고,
a) 여기에서 복수의 아그로박테리움 세포들이 적어도 하나의 기능성 이종 ORF를 포함하는 적어도 하나의 플러스-센스 단일 가닥 RNA 바이러스 벡터를 포함하고,
i) 여기에서 적어도 하나의 플러스-센스 단일 가닥 RNA 바이러스 벡터가 기능성 피복 단백질 ORF가 결핍되고,
ii) 여기에서 적어도 하나의 플러스-센스 단일 가닥 RNA 바이러스 벡터가 식물 세포들 내에서 복제될 수 있는 RNA 복제단위를 포함하고,
그리고
b) 여기에서 적어도 2개의 아그로박테리움 세포들이 서로 다른 기능성 이종 ORF들을 포함하는 서로 다른 플러스-센스 단일 가닥 RNA 벡터들을 포함하고, 그리고
c) 여기에서 서로 다른 기능성 이종 ORF들을 포함하는 서로 다른 플러스-센스 단일 가닥 RNA 벡터들이 동일 바이러스로부터 파생된다.
본 발명의 바람직한 구체예에 있어서, 적어도 2개, 3개, 4개, 5개 또는 그 이상의 아그로박테리움 세포들이 서로 다른 플러스-센스 단일 가닥 RNA 벡터들을 포함하는 혼합물 내에 2개 3개, 4개, 5개 또는 2개, 3개, 4개, 5개 이상 또는 그 이상, 심지어 보다 바람직하게는 2개의 아그로박테리움 세포들을 포함한다. 심지어 보다 바람직하게는, 혼합물 내의 2개의 아그로박테리움 세포들 각각은 하나의 플러스-센스 단일 가닥 RNA 벡터를 포함한다.
보다 바람직한 구체예에 있어서, 상기 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스는 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이다.
보다 바람직한 구체예에 있어서, 상기 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스의 피복 단백질 ORF가 플러스-센스 단일 가닥 RNA 바이러스 벡터들 내에서 완전히 결실된다.
본 발명의 또 다른 바람직한 구체예에 있어서, 상기 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이고 그리고 상기 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스의 피복 단백질 ORF가 플러스-센스 단일 가닥 RNA 바이러스 벡터 내에서 완전히 결실된다.
본 발명을 사용하여 센스 또는 안티센스 방향(orientation) 내에서 발현될 수 있는 대상의 단백질들 또는 그의 단편들에는 전분 변성 효소들(starch modifying enzyme: 전분 합성효소(starch synthase), 전분 포스포릴화 효소(starch phosphorylation enzyme), 탈분지 효소(debranching enzyme), 전분 분지 효소(starch branching enzyme), 전분 분지 효소 II, 과립 결합 전분 합성효소(granule bound starch synthase)), 슈크로스 인산 합성효소(sucrose phosphate synthase), 슈크로스 가인산분해효소(sucrose phosphorylase), 폴리갈락투로나아제(polygalacturonase), 폴리푸룩탄 수크라아제(polyfructan sucrase), ADP 글루코스 파이로포스포릴라아제(ADP glucose pyrophosphorylase), 사이클로덱스트린 글리코실트랜스퍼라아제(cyclodextrin glycosyltransferase), 프룩토실 트랜스퍼라아제(fructosyl transferase), 글리코겐 합성효소(glycogen synthase), 펙틴 에스테르 가수분해효소(pectin esterase), 아프로티닌(aprotinin), 아비딘(avidin), 박테리아 레반슈크라아제(bacterial levansucrase), 대장균 glgA 단백질(E.coli glgA protein), MAPK4 및 상동체들(orthologues), 질소 동화/대사 효소(nitrogen assimilation/methabolism enzyme), 글루타민 합성효소(glutamine synthase), 식물 오스모틴(plant osmotin), 2S 알부민(2S albumin), 토마틴(thaumatin), 위치-특이적 재조합효소/삽입효소(site-specific recombinase/integrase ; FLP, Cre, R 재조합효소, Int, SSVI 삽입효소 R, 삽입효소 phiC31 또는 그의 활성 단편(active fragment) 또는 변종), 이소펜테닐 트랜스퍼라아제(isopentenyl transferase), Sca M5 (대두 칼모듈린(soybean calmodulin)), 콜레옵테란 타입 독소(coleopteran type toxin) 또는 살충 활성 단편(insecticidally active fragment), 유비퀴틴 결합 효소(E2) 융합 단백질들(ubiquitin conjugating enzyme (E2) fusion proteins), 지질을 대사하는 효소들(enzymes that metabolise lipids), 아미노산들(amino acids), 당류(sugars), 핵산들 및 다당류들(nucleic acids and polysaccharides), 과산화물 제거효소(superoxide dismutase), 단백질가수분해효소(protease)의 비활성 효소전구체 형태(inactive proenzyme form), 식물 프로테인 독소(plant protein toxins), 섬유 생산 식물들에서 섬유를 변화시키는 특성들(traits altering fiber in fiber producing plants), 곤충병원성균(Bacillus thuringiensis)로부터의 콜레옵테란 활성형 독소(Coleopteran active toxin ; Bt2 독소, 살충 결정 단백질(ICP: insecticidal crystal protein), CrylC 독소, 델타 내독소(delta endotoxin), 폴리펩티드 독소(polyopeptide toxin), 독소전구체(protoxin) 등), 곤충 특이적 독소(insect specific toxin AalT), 셀룰로오스 분해 효소(cellulose degrading enzymes), 아시도테르무스 셀룰로티쿠스(Acidothermus celluloticus)로부터의 E1 셀룰라아제(E1 cellulase), 리그닌 변성 효소(lignin modifying enzymes), 신나모일 알코올 탈수소효소(cinnamoyl alcohol dehydrogenase), 트레할로스-6-인산 합성효소(trehalose-6-phosphate synthase), 사이토키닌 대사 경로(cytokinin metabolic pathway)의 효소, HMG-CoA 환원효소, 대장균 무기 피로인산가수분해효소(E. coli inorganic pyrophosphatase), 종자 저장 단백질(seed storage protein), 에르위니아 허비콜라 라이코펜 합성효소(Erwinia herbicola lycopen synthase), ACC 산화효소(ACC oxidase), pTOM36 인코딩 단백질, 피타아제(phytase), 케토가수분해효소(ketohydrolase), 아세토아세틸 코엔자임 A 환원효소(acetoacetyl CoA reductase), PHB(폴리하이드록시부타노에이트) 합성효소(PHB(polyhydroxybutanoate) synthase), 아실기 운반 단백질(acyl carrier protein), 나핀(napin), EA9, 비-고급 식물 피토엔 합성효소(non-higher plant phytoene synthase), pTOM5 인코딩 단백질, 에틸렌 수용기(ETR: ethylene receptor), 가소성 피루베이트 인산 이인산화제(plastidic pyruvate phosphate dikinase), 네마토드-유도가능 경막 기공 단백질(nematode-inducible transmembrane pore protein), 식물 세포의 광합성 또는 플라스티드 기능을 향상시키는 특성들(trait enhancing photosynthetic or plastid function of the plant cell), 스틸벤 합성효소(stilbene synthase), 페놀을 히드록실화할 수 있는 효소(enzyme capable of hydroxylating phenols), 카테콜 2산소화효소(catechol dioxygenase), 카테콜 2,3-2산소화효소(catechol 2,3-dioxygenase), 클로로뮤코네이트 사이클로아이소머라아제(chloromuconate cycloisomerase), 안트라닐레이트 합성효소(anthranilate synthase), 브라시카 AGL15 단백질(Brassica AGL15 protein), 푸룩토스 1,6-2인산가수분해효소(FBPase: fructose 1,6-biphosphatase), AMV RNA3, PVY 복제효소, PLRV 복제효소, 포티바이러스 피복 단백질, CMV 피복 단백질, TMV 피복 단백질, 루테오바이러스 복제효소(luteovirus replicase), MDMV 전령 RNA(MDMV messenger RNA), 돌연변이 제미니바이러스 복제효소(mutant geminiviral replicase), 캘리포니아만월계수 C12:0 선호 아실-ACP 티오에스테라아제(Umbellularia californica C12:0 preferring acyl-ACP thioesterase), 식물 C10 또는 C12:0 선호 아실-ACP 티오에스테라아제(plant C10 or C12:0 preferring acyl-ACP thioesterase), C14:0 선호 아실-ACP 티오에스테라아제(luxD: C14:0 preferring acyl-ACP thioesterase), 식물 합성효소인자 A(plant synthase factor A), 식물 합성효소인자 B, 6-불포화효소(6-desaturase), 식물 세포들 내의 지방산들의 퍼옥시좀-산화(peroxysomal-oxidation)에서 효소 활성을 갖는 단백질, 아실-코엔자임 A 산화효소(acyl-CoA oxidase), 3-케토아실-코엔자임 A 티올라아제(3-ketoacyl-CoA thiolase), 지방분해효소(lipase), 옥수수 아세틸-코엔자임 A-카르복실화효소(maize acetyl-CoA-carboxylase), 5-엔올피루빌시키메이트-3-인산 합성효소(ESEP: 5-enolpyruvylshikimate-3-phosphate synthase), 포스피노트리킨 아세틸 트랜스퍼라아제(BAR, PAT: phosphinothricin acetyl transferase), CP4 단백질, ACC 탈아미노효소(ACC deaminase), 리보자임(ribozyme), 전사후 개열 부위를 갖는 단백질(protein having posttranslational cleavage site), Gal4 전사 활성화제의 DNA-결합 도메인과 전사 활성화 도메인으로 이루어지는 단백질 융합물(protein fusion consisting of a DNA-binding domain of Gal4 transcriptional activator and a transcriptional activation domain), 융합 단백질을 지질상 내로 표적화할 수 있는 대상의 단백질을 수반하는 올레오신 단백질의 전사 융합물(translational fusion of oleosin protein with protein of interest capable of targeting the fusion protein into the lipid phase), 설폰아미드 내성을 부여하는 DHPS 유전자(DHPS gene conferring sulfonamide resistance), 박테리아 니트릴라아제(bacterial nitrilase), 2,4-D 모노옥시게나아제(2,4-D monooxygenase), 아세톨아세테이트 합성효소(ALS: acetolactate synthase) 또는 아세토하이드록시산 합성효소(AHAS: acetohydroxyacid synthase), 폴리갈락투로나아제(polygalacturonase), 박테리아 니트릴라아제(bacterial nitrilase), 플라스티드의 내막 내에 체류하는 숙성 인산 운반체 단백질(mature phosphate translocator protein)의 아미노 말단 소수성 영역과 막 내로 표적화되어야 할 대상의 단백질의 융합물 등이 포함된다.
임의의 인간 또는 동물 단백질이 본 발명의 시스템을 사용하여 발현될 수 있다. 이러한 대상의 단백질들의 예들에는 그 중에서도 하기의 약제학적 대상의 단백질들 즉: 면역 반응 단백질들(immune response proteins ; 항체(antibodies), 단쇄 항체(single chain antibodies), T 세포 수용기(T cell receptors) 등), 항원(antigens), 콜로니자극인자(colony stimulating factors), 릴렉신(relaxins), 폴리펩티드 호르몬(polypeptide hormones), 사이토카인(cytokines) 및 이들의 수용기들, 인터페론(interferons), 성장인자(growth factors) 및 응고인자(coagulation factors), 효소 활성 리소좀 효소(enzymatically active lysosomal enzyme), 섬유소용해성 폴리펩티드(fibrinolytic polypeptides), 혈액응고인자(blood clotting factors), 트립시노겐(trypsinogen), 1-안티트립신(AAT: 1-antitrypsin)와 마찬가지로 단백질들의 융합물, 돌연변이 버전 및 합성 유도체들과 같은 기능-보존 단백질들이 포함된다.
실시예로 나타낸 바와 같이 그리고 설명된 바와 같이, 킷트들, 혼합물들, 벡터들, 식물들 및 식물 세포들이 특히 식물 내에서 폴리펩티드들, 특히 헤테로올리고머 폴리펩티드들의 발현에 유용하다.
따라서, 본 발명은 하나의 구체예에 있어서 폴리펩티드들, 특히 헤테로올리고머 폴리펩티드들을 생산하기 위한 본 발명의 킷트의 또는 본 발명의 혼합물의 또는 본 발명의 PVX 바이러스 벡터의 또는 본 발명의 식물 또는 식물 세포의 또는 본 발명의 식물의 또는 본 발명의 아그로박테리움 세포들의 혼합물의 용도에 관한 것이다.
용어 "이동 단백질(들)(movement protein(s))" 또는 "MP(s)"는 숙련된 자에게 공지되어 있고 그리고 토바모바이러스들 내의 MP ORF와 같은 세포-대-세포 이동에 요구되는 단백질(들)로서 이해된다.
용어 "삼중 유전자 블럭 단백질들(triple gene block proteins)" 또는 "TGB"는 숙련된 자에게 공지되어 있고 그리고 포텍스바이러스들 내의 TGB ORF들과 같은 세포-대-세포 이동에 요구되는 단백질들서 이해된다.
용어 "피복 단백질(coat protein)"은 숙련된 자에게 공지되어 있고 그리고 바이러스 피복을 구축하는 단백질(들)로서 이해된다.
도 1은 바이러스 벡터 백본 딜레마를 나타낸다. 개개 바이러스 벡터들 PVX 또는 TMV(동일한 백본을 갖는) 중의 어느 하나는 단일 식물 세포를 공-감염 시킬 수 없다; 이들은 전신적 이동 동안 분리되고 그리고 단지 잎들 상의 분리된 패치들 내에서 외래 유전자들을 발현할 것이다. RdRp: RNA-의존성 RNA-중합효소; CP: 피복 단백질; ORF: 오픈리딩프레임; MP: 이동 단백질, TGB: 삼중 유전자 블럭. 화살표는 하위 유전체 프로모터들을 나타낸다.
도 2는 천연의 헬핑페어(natural helping pair) TMV 및 PVX를 나타낸다. 두 바이러스들은 단일 세포를 공-감염시킬 수 있고, 함께 이동하고 그리고 식물들을 전신적으로 감염시킬 수 있다. RdRp: RNA-의존성 RNA-중합효소; CP: 피복 단백질; ORF: 오픈리딩프레임; MP: 이동 단백질, TGB: 삼중 유전자 블럭. 화살표는 하위 유전체 프로모터들을 나타낸다.
도 3은 제1 및 제2 플러스 가닥 바이러스 벡터를 포함하는 본 발명의 킷트를 나타낸다. 이들 바이러스 벡터들에 있어서, 피복 단백질들이 상호호환적으로 교환되었다. 두 바이러스들은 단일 세포를 공감염시킬 수 있고 그리고 식물들을 전신적으로 감염시킬 수 있다. 동시에, 둘 중의 어느 하나도 단독으로는 식물들을 전신적으로 감염시킬 수 없어 봉쇄를 확실하게 할 수 있다. RdRp: RNA-의존성 RNA-중합효소; CP: 피복 단백질; ORF: 오픈리딩프레임; MP: 이동 단백질, TGB: 삼중 유전자 블럭. 화살표는 하위 유전체 프로모터들을 나타낸다. A는 유전자 내로 삽입된 부가의 ORF들을 수반하지 않는 상보쌍(complementing pair) PVX-CP(TMV) 및 TMV-CP(PVX)를 나타낸다. B: 하나의 ORF가 상보쌍의 PVX 벡터 내에 삽입되었고, C: 하나의 ORF가 상보쌍의 TMV 벡터 내에 삽입되었고, D: ORF1이 상보쌍의 PVX 벡터 내에 그리고 ORF2가 TMV 벡터 내에 삽입되었다.
도 4는 PVX에 기초하는 본 발명의 킷트의 제3 바이러스 벡터의 모식적인 도면을 나타낸다. 바이러스 벡터는 피복 단백질을 포함하지 않고; 특히 피복 단백질이 완전히 결실되어 있다. 게다가, 바이러스 벡터는 이종 ORF(PVX-ΔCP-ORFx)를 포함한다. RdRp: RNA-의존성 RNA-중합효소; ORF: 오픈리딩프레임; TGB: 삼중 유전자 블럭. 화살표는 하위 유전체 프로모터들을 나타낸다.
도 5는 본 발명의 킷트의 제3 바이러스 벡터의 모식적인 도면을 나타낸다. 바이러스 벡터는 피복 단백질과 이동 단백질들 둘 다를 포함하지 않는다. 게다가, 바이러스 벡터는 이종 ORF(PVX-ΔCP-ΔTGB-ORFx)를 포함한다. RdRp: RNA-의존성 RNA-중합효소; ORF: 오픈리딩프레임. 화살표는 하위 유전체 프로모터들을 나타낸다.
도 6은 A 및 B에서 바이러스 벡터들 PVX-mBananaΔCP, PVX-mCherry-CPTMV 및 TMV-GFP-CPPVX들로 전신적으로 공-감염된 식물로부터의 식물의 잎을 나타낸다. 사진들은 가시화를 위하여 정상 광(normal light), GFP에 대하여 자외-광(UV-light), 녹색광 하에서 mCherry에 대하여 적색 필터(red filter) 그리고 mBanana에 대하여 황색 필터로 촬영하였다.
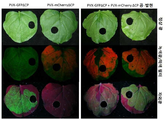
도 7은 서로 다른 바이러스 벡터들 단독 또는 조합을 수반하는 잎들로부터의 식물 수액(plant sap)의 SDS-PAGE(소듐도데실설페이트-폴리아크릴아미드 겔 전기영동: Sodiumdodecylsulfate-polyacrylamide gel electrophoresis)을 나타내고 있다. SDS-PAGE에 대하여는, 겔 내의 형광 단백질들을 직접적으로 가시화시키기 위하여 탐침들은 가열되지 않았다. 겔은 쿠마씨 염색(Coomassie staining) 이전에 자외-광, 녹색광과 적색 필터, 청색광과 황색 필터 하에서 그리고 쿠마씨 염색 이후에 정상 광 하에서 관찰되었다. M: P7711S 래더(P7711Sladder ; 뉴잉글랜드 바이오랩스(NEB); 야생담배(N. benthamiana): 미감염/접종된 식물로부터의 식물 수액; PVX201: 정제된 PVX201 입자들(1㎍), TMV: 정제된 TMV 입자들(1㎍).
도 8은 서로 다른 바이러스 벡터들 단독 또는 조합을 수반하는 잎들로부터의 식물 수액의 웨스턴 블럿(Western blots)들을 나타내고 있다. 이들 분석들을 위하여는, SDS-PAGE 이전에 탐침들이 가열되었다. 웨스턴 블럿들을 피복 단백질들을 검출하는 폴리클로날 PVX 또는 TMV 항체 및 알칼리포스파타아제(alkaline phosphatase)로 표지된 염소-항-토끼 항체(goat-anti-rabbit antibody)와 함께 배양시켰다. M: P7711S 래더(뉴잉글랜드 바이오랩스); 야생담배(N. benthamiana): 미감염/접종된 식물로부터의 식물 수액; PVX mCherry-2A-CP: mCherry-overcoat 입자로의 감염으로부터의 식물 수액; TMV-GFP: TMV 발현 GFP로의 감염으로부터의 식물 수액.
도 9는 피복 단백질 결핍 PVX 벡터들을 발현하는 야생담배(N. benthamiana) 잎들을 나타내고 있다. 잎들을 PVX-GFPΔCP, PVX-mCherryΔCP 또는 두 벡터들로 접종시켰다. 접종 후 4일차(dpi)에서 사진들을 촬영하였다.
도 10은 PVX-GFPΔCP 및 PVX-mCherryΔCP를 공-발현하는 야생담배(N. benthamiana) 식물을 나타내고 있다. 접종 후 4, 5 및 6일차(dpi)에서 사진들을 촬영하였다.
도 11은 야생담배(N. benthamiana)의 서로 다른 바이러스 벡터들 단독 또는 조합으로 접종된 잎들을 나타내고 있다. 가시화를 위하여 정상 광, GFP에 대하여는 자외-광, mCherry에 대하여는 녹색광 및 적색 필터 그리고 mBanana에 대하여는 청색광 및 황색 필터 하에서 사진들을 촬영하였다. 접종 후 13일차(dpi)에서 식물 부분들을 수확하였다.
도 12는 형광 단백질들의 특정의 여기를 수반하는 상보쌍으로 감염된 식물 잎들의 현미경 사진들을 나타내고 있다. A 내지 D: mCherry(텍사스 레드 필터) 및 GFP(GFP 필터)의 여기를 수반하는 상보쌍, E 내지 H: mCherry 및 mBanana(YFP 필터)의 여기를 수반하는 상보쌍; I 내지 L: mCherry 및 mBanana의 여기를 수반하는 PVX-mBananaΔCP를 수반하는 상보쌍; A, E, I: 투과된 광을 나타낸 세포들, B, F, J: mCherry의 여기, C: GFP의 여기, G, K: mBanana의 여기, D, H, L: 나타낸 감염의 사진들의 중첩.
도 13은 상보쌍 및 피복 단백질가 결핍된 PVX 또는 TMV 중의 하나인 제3 벡터로 감염된 식물 잎들을 나타내고 있다. PVX 또는 TMV 둘 중의 하나에 기초하였고 그리고 피복 단백질이 결핍된 mBanana를 발현하는 제3 벡터로 상보쌍 PVX-mCherry-CPTMV 및 TMV-GFP-CPPVX의 공-접종들을 시험하였다. 접종 후 7일차(dpi)(접종된 잎들) 및 24일차(dpi)(전신적으로 감염된 잎들)에서의 잎들을 나타내었고 그리고 서로 다른 형광 단백질들에 대한 특정의 여기 조건들이 적용되었다.
도 14는 상보쌍 및 피복 단백질이 결핍된 PVX 또는 TMV 둘 중의 하나인 제3 벡터로 전신적 감염된 잎들의 SDS-PAGE 및 웨스턴 블럿들을 나타내고 있다. 겔들을 형광 단백질들의 가시화를 위한 쿠마씨 염색 이전에 적색 필터(좌상부)를 수반하는 녹색광, 청색광과 황색 필터(중상부) 및 자외광(우상부) 하에서 나타내었다. 하부에 대하여는 쿠마씨 염색된 겔(좌측) 및 PVX의 CP(α-PVX) 및 TMV의 CP(α-TMV)에 대한 웨스턴 블럿들을 나타내었다. M: P7711S 단백질 래더(뉴잉글랜드 바이오랩스), Nb: 미감염된 식물 야생담배(N. benthamiana), 1: 26일차(dpi)에서의 상보쌍, 2: 26일차(dpi)에서의 PVX-mBananaΔCP를 수반하는 상보쌍, 3: 33일차(dpi)에서의 상보쌍, 4: 33일차(dpi)에서의 PVX-mBananaΔCP를 수반하는 상보쌍, 5: 33일차(dpi)에서의 TMV-mBananaΔCP를 수반하는 상보쌍. P: PVX201 정제(1㎍), T: TMV 정제(1㎍).
도 15는 형광 단백질들의 여기를 수반하는 PVX 기반 CP 결핍 벡터들로의 감염들의 식물 잎들의 현미경 사진들을 나타내고 있다. A 내지 D: 미감염된 식물 야생담배(N. benthamiana). E 내지 H: PVX-GFPΔCP로 감염된 식물, I 내지 L: plant infected with PVX-mCherryΔCP로 감염된 식물, M 내지 P: PVX-GFPΔCP 및 PVX-mCherryΔCP로 공-감염된 식물. A, E, I, M: 투과된 광을 나타낸 세포들, B, F, J, N: mCherry(텍사스 레드 필터)에 대한 여기; C, G, K, O: GFP(GFP 필터)의 여기, D, H, L, P: 나타낸 감염의 사진들의 중첩.
도 2는 천연의 헬핑페어(natural helping pair) TMV 및 PVX를 나타낸다. 두 바이러스들은 단일 세포를 공-감염시킬 수 있고, 함께 이동하고 그리고 식물들을 전신적으로 감염시킬 수 있다. RdRp: RNA-의존성 RNA-중합효소; CP: 피복 단백질; ORF: 오픈리딩프레임; MP: 이동 단백질, TGB: 삼중 유전자 블럭. 화살표는 하위 유전체 프로모터들을 나타낸다.
도 3은 제1 및 제2 플러스 가닥 바이러스 벡터를 포함하는 본 발명의 킷트를 나타낸다. 이들 바이러스 벡터들에 있어서, 피복 단백질들이 상호호환적으로 교환되었다. 두 바이러스들은 단일 세포를 공감염시킬 수 있고 그리고 식물들을 전신적으로 감염시킬 수 있다. 동시에, 둘 중의 어느 하나도 단독으로는 식물들을 전신적으로 감염시킬 수 없어 봉쇄를 확실하게 할 수 있다. RdRp: RNA-의존성 RNA-중합효소; CP: 피복 단백질; ORF: 오픈리딩프레임; MP: 이동 단백질, TGB: 삼중 유전자 블럭. 화살표는 하위 유전체 프로모터들을 나타낸다. A는 유전자 내로 삽입된 부가의 ORF들을 수반하지 않는 상보쌍(complementing pair) PVX-CP(TMV) 및 TMV-CP(PVX)를 나타낸다. B: 하나의 ORF가 상보쌍의 PVX 벡터 내에 삽입되었고, C: 하나의 ORF가 상보쌍의 TMV 벡터 내에 삽입되었고, D: ORF1이 상보쌍의 PVX 벡터 내에 그리고 ORF2가 TMV 벡터 내에 삽입되었다.
도 4는 PVX에 기초하는 본 발명의 킷트의 제3 바이러스 벡터의 모식적인 도면을 나타낸다. 바이러스 벡터는 피복 단백질을 포함하지 않고; 특히 피복 단백질이 완전히 결실되어 있다. 게다가, 바이러스 벡터는 이종 ORF(PVX-ΔCP-ORFx)를 포함한다. RdRp: RNA-의존성 RNA-중합효소; ORF: 오픈리딩프레임; TGB: 삼중 유전자 블럭. 화살표는 하위 유전체 프로모터들을 나타낸다.
도 5는 본 발명의 킷트의 제3 바이러스 벡터의 모식적인 도면을 나타낸다. 바이러스 벡터는 피복 단백질과 이동 단백질들 둘 다를 포함하지 않는다. 게다가, 바이러스 벡터는 이종 ORF(PVX-ΔCP-ΔTGB-ORFx)를 포함한다. RdRp: RNA-의존성 RNA-중합효소; ORF: 오픈리딩프레임. 화살표는 하위 유전체 프로모터들을 나타낸다.
도 6은 A 및 B에서 바이러스 벡터들 PVX-mBananaΔCP, PVX-mCherry-CPTMV 및 TMV-GFP-CPPVX들로 전신적으로 공-감염된 식물로부터의 식물의 잎을 나타낸다. 사진들은 가시화를 위하여 정상 광(normal light), GFP에 대하여 자외-광(UV-light), 녹색광 하에서 mCherry에 대하여 적색 필터(red filter) 그리고 mBanana에 대하여 황색 필터로 촬영하였다.
도 7은 서로 다른 바이러스 벡터들 단독 또는 조합을 수반하는 잎들로부터의 식물 수액(plant sap)의 SDS-PAGE(소듐도데실설페이트-폴리아크릴아미드 겔 전기영동: Sodiumdodecylsulfate-polyacrylamide gel electrophoresis)을 나타내고 있다. SDS-PAGE에 대하여는, 겔 내의 형광 단백질들을 직접적으로 가시화시키기 위하여 탐침들은 가열되지 않았다. 겔은 쿠마씨 염색(Coomassie staining) 이전에 자외-광, 녹색광과 적색 필터, 청색광과 황색 필터 하에서 그리고 쿠마씨 염색 이후에 정상 광 하에서 관찰되었다. M: P7711S 래더(P7711Sladder ; 뉴잉글랜드 바이오랩스(NEB); 야생담배(N. benthamiana): 미감염/접종된 식물로부터의 식물 수액; PVX201: 정제된 PVX201 입자들(1㎍), TMV: 정제된 TMV 입자들(1㎍).
도 8은 서로 다른 바이러스 벡터들 단독 또는 조합을 수반하는 잎들로부터의 식물 수액의 웨스턴 블럿(Western blots)들을 나타내고 있다. 이들 분석들을 위하여는, SDS-PAGE 이전에 탐침들이 가열되었다. 웨스턴 블럿들을 피복 단백질들을 검출하는 폴리클로날 PVX 또는 TMV 항체 및 알칼리포스파타아제(alkaline phosphatase)로 표지된 염소-항-토끼 항체(goat-anti-rabbit antibody)와 함께 배양시켰다. M: P7711S 래더(뉴잉글랜드 바이오랩스); 야생담배(N. benthamiana): 미감염/접종된 식물로부터의 식물 수액; PVX mCherry-2A-CP: mCherry-overcoat 입자로의 감염으로부터의 식물 수액; TMV-GFP: TMV 발현 GFP로의 감염으로부터의 식물 수액.
도 9는 피복 단백질 결핍 PVX 벡터들을 발현하는 야생담배(N. benthamiana) 잎들을 나타내고 있다. 잎들을 PVX-GFPΔCP, PVX-mCherryΔCP 또는 두 벡터들로 접종시켰다. 접종 후 4일차(dpi)에서 사진들을 촬영하였다.
도 10은 PVX-GFPΔCP 및 PVX-mCherryΔCP를 공-발현하는 야생담배(N. benthamiana) 식물을 나타내고 있다. 접종 후 4, 5 및 6일차(dpi)에서 사진들을 촬영하였다.
도 11은 야생담배(N. benthamiana)의 서로 다른 바이러스 벡터들 단독 또는 조합으로 접종된 잎들을 나타내고 있다. 가시화를 위하여 정상 광, GFP에 대하여는 자외-광, mCherry에 대하여는 녹색광 및 적색 필터 그리고 mBanana에 대하여는 청색광 및 황색 필터 하에서 사진들을 촬영하였다. 접종 후 13일차(dpi)에서 식물 부분들을 수확하였다.
도 12는 형광 단백질들의 특정의 여기를 수반하는 상보쌍으로 감염된 식물 잎들의 현미경 사진들을 나타내고 있다. A 내지 D: mCherry(텍사스 레드 필터) 및 GFP(GFP 필터)의 여기를 수반하는 상보쌍, E 내지 H: mCherry 및 mBanana(YFP 필터)의 여기를 수반하는 상보쌍; I 내지 L: mCherry 및 mBanana의 여기를 수반하는 PVX-mBananaΔCP를 수반하는 상보쌍; A, E, I: 투과된 광을 나타낸 세포들, B, F, J: mCherry의 여기, C: GFP의 여기, G, K: mBanana의 여기, D, H, L: 나타낸 감염의 사진들의 중첩.
도 13은 상보쌍 및 피복 단백질가 결핍된 PVX 또는 TMV 중의 하나인 제3 벡터로 감염된 식물 잎들을 나타내고 있다. PVX 또는 TMV 둘 중의 하나에 기초하였고 그리고 피복 단백질이 결핍된 mBanana를 발현하는 제3 벡터로 상보쌍 PVX-mCherry-CPTMV 및 TMV-GFP-CPPVX의 공-접종들을 시험하였다. 접종 후 7일차(dpi)(접종된 잎들) 및 24일차(dpi)(전신적으로 감염된 잎들)에서의 잎들을 나타내었고 그리고 서로 다른 형광 단백질들에 대한 특정의 여기 조건들이 적용되었다.
도 14는 상보쌍 및 피복 단백질이 결핍된 PVX 또는 TMV 둘 중의 하나인 제3 벡터로 전신적 감염된 잎들의 SDS-PAGE 및 웨스턴 블럿들을 나타내고 있다. 겔들을 형광 단백질들의 가시화를 위한 쿠마씨 염색 이전에 적색 필터(좌상부)를 수반하는 녹색광, 청색광과 황색 필터(중상부) 및 자외광(우상부) 하에서 나타내었다. 하부에 대하여는 쿠마씨 염색된 겔(좌측) 및 PVX의 CP(α-PVX) 및 TMV의 CP(α-TMV)에 대한 웨스턴 블럿들을 나타내었다. M: P7711S 단백질 래더(뉴잉글랜드 바이오랩스), Nb: 미감염된 식물 야생담배(N. benthamiana), 1: 26일차(dpi)에서의 상보쌍, 2: 26일차(dpi)에서의 PVX-mBananaΔCP를 수반하는 상보쌍, 3: 33일차(dpi)에서의 상보쌍, 4: 33일차(dpi)에서의 PVX-mBananaΔCP를 수반하는 상보쌍, 5: 33일차(dpi)에서의 TMV-mBananaΔCP를 수반하는 상보쌍. P: PVX201 정제(1㎍), T: TMV 정제(1㎍).
도 15는 형광 단백질들의 여기를 수반하는 PVX 기반 CP 결핍 벡터들로의 감염들의 식물 잎들의 현미경 사진들을 나타내고 있다. A 내지 D: 미감염된 식물 야생담배(N. benthamiana). E 내지 H: PVX-GFPΔCP로 감염된 식물, I 내지 L: plant infected with PVX-mCherryΔCP로 감염된 식물, M 내지 P: PVX-GFPΔCP 및 PVX-mCherryΔCP로 공-감염된 식물. A, E, I, M: 투과된 광을 나타낸 세포들, B, F, J, N: mCherry(텍사스 레드 필터)에 대한 여기; C, G, K, O: GFP(GFP 필터)의 여기, D, H, L, P: 나타낸 감염의 사진들의 중첩.
실시예들
실시예
1: 벡터들의 생성 및 상보 바이러스 벡터들을 수반하는 여러 이종 단백질들의 공-발현
오버랩 발현 중합효소 연쇄반응에 의한 유전자 스플라이싱 방법(gene splicing by overlap extension(SOE) PCR method)의 도움으로 상보 벡터들을 생성하였다. TMV의 피복 단백질을 수반하는 PVX의 구축을 위하여 3개의 PCT 산물들이 생성되었다. PCR 1에서 mCherry 유전자가 PVX 유전체 내로 삽입되고(PVX-mCherry) 그리고 PVX 피복 단백질의 하위 유전체 프로모터가 융합을 위한 오버랩 시퀀스(overlapping sequence)를 포함하는 프라이머들 mCherry-ClaI 및 SOE-TMVCP rv로 증폭되었다. PCR 2는 벡터 pJL24(US 2010/0071085 Al)로부터의 하위 유전체 프로모터의 일부 및 TMV 피복 단백질의 ORF를 2개의 오버랩 시퀀스들, PCR 산물 1에 상보인 5'-말단 및 PCR 산물 3에 상보인 3'-말단을 포함하는 프라이머들 SOE-TMVCP fw 및 SOE-TMVCP2 rv로 증폭시켰다. PCR 3에 대하여는 플라스미드 백본의 부분들을 포함하는 PVX 유전체의 3' 부분(3' part)이 벡터 PVX-mCherry로부터의 프라이머들 SOE-TMVCP2-fw 및 M13 universe로 생성되었다. 프라이머가 결핍된 4번째 PCR에서 모든 PCR 산물들이 오버랩 시퀀스들로 인하여 융합되고 그리고 다시 PCR 5에서 프라이머들 mCherry-ClaI 및 M13 universe들로 증폭되었다. 계속해서 최종 PCR 산물을 ClaI 및 SalI로 절단시키고 그리고 동일한 효소들로 절단되고 그리고 송아지 내장 포스파타제(CIP: calf intestinal phosphatase)로 탈포스포릴화된 PVX 벡터 내로 결찰시켰다. PVX 벡터는 또한 이원 pTRAc 벡터(binary pTRAc vector)의 플라스미드 백본으로 이루어진다(문헌 Mclean, 2007, J Gen Virol 88, 1460-1469). UK3 균주(UK3 strain)의 PVX 유전체가 T-DNA의 좌측 경계(left border)와 우측 경계 사이에서 일체화된다. 피복 단백질의 하위 유전체 프로모터가 복제되고 그리고 제한효소들 NheI, ClaI 및 SmaI을 갖는 다클로닝 부위(multiple cloning site)가 일체화된다(pPVX201 특허 WO96/12027). 최종 벡터에서 구조 pPVX-mCherry-CPTMV는 형광 단백질 mCherry 및 TMV의 피복 단백질에 대한 이종 유전자들과 마찬가지로 포텍스바이러스 RNA-의존성 RNA-중합효소 및 삼중 유전자 블럭 단백질들에 대하여 코딩한다. mCherry 및 TMV 피복 단백질의 발현을 위한 하위 유전체 프로모터들은 복제된 PVX 피복 단백질의 sg 프로모터들이다. 하기 표 1은 상보 벡터들의 구축을 위하여 사용된 DNA 올리고머들이다.

TMV 벡터의 구축을 위하여 2개의 PCR 산물들이 생성되었다. PCR 1에서 녹색 형광 단백질의 시퀀스가 증폭되어 플라스미드 pJL24 상의 프라이머들 PacI-GFP-TMV and SOE2-CP rv(표 1)를 수반하는 TMV의 5'-말단 및 피복 단백질 하위 유전체 프로모터의 일부에 있는 PacI 제한 부위를 첨가하였다. 제2 PCR에서 PVX의 피복 단백질 시퀀스가 프라이머들 SOE2-CP fw 및 SOE2-CP-NotI로 증폭되어 5'-말단에서 TMV의 하위 유전체 프로모터의 일부 및 3'-말단에서 NotI 제한 부위를 수반하는 구조물을 생성하였다. 2개의 PCR 산물들을 프라이머들 없이 제3 PCR에서 융합시키고 그리고 제4 PCR에서 프라이머들 PacI-GFP-TMV 및 SOE2-CP NotI로 증폭시켰다. 최종 PCR 산물을 효소들 PacI 및 NotI로 절단하고 그리고 아가로스 겔(agarose gel) 상에서 정제하였다. 표적 벡터 pTRBOG(US 2010/0071085 Al)를 동일한 제한 효소들로 처리하고 그리고 CIP로 탈포스포릴화시켰다. PCR 단편을 TMV 벡터 내로 결찰시키고 그리고 플라스미드 pTMV-GFP-CPPVX 내에서 결과를 야기시켰다.
완전한 피복 단백질 융합물들을 수반하는 PVX 벡터들의 구축을 위하여 2A 시퀀스를 수반하는 N-말단 mBanana 피복 단백질을 갖는 현존하는 PVX 벡터들이 사용되었다. 예를 들면 PVX-mBanana-2A-CP는 아그로감염에 맞는 PVX 벡터이다. 플라스미드의 백본은 이원 pTRAc 벡터이다. UK3 균주의 PVX 유전체는 T-DNA의 좌측 경계 및 우측 경계 사이에서 일체화된다. 선택된 벡터들은 다른 형광 단백질들의 수족구병 바이러스(FMDV: Food and Mouth Disease Virus)의 2A 시퀀스, 예를 들면 황색 형광 단백질 mBanana와의 피복 단백질 융합물을 갖는다. PVX 유전체의 3' 부분이 PCR로 증폭되었고 그리고 또한 융합 벡터들 내의 2A 시퀀스의 시작에 존재하는 제한 부위 BspEI이 프라이머들 2ADelCPfw(5'-AATCCGGATAACTACGTCTACATAACCG-3' (시퀀스 동정 번호 11)) 및 M13 universe(5'-GTTGTAAAACGACGGCCAGT-3' (시퀀스 동정 번호 ))에 첨가되었다. 프라이머 M13은 벡터 백본 내측 PVX 유전체 외측을 결합하고 그리고 프라이머 2ADelCpfw는 피복 단백질 코딩 시퀀스의 다운스트림을 직접 결합하고 그리고 BspEI 부위를 첨가한다. 산물을 pCR2.1-Topo 벡터(미합중국 칼스배드 소재 라이프테크놀로지스사(Lifetechnologies)) 내로 서브클론(subcloned) 시키고 대장균(E. coli SCS110) 내에서 증폭시켜 비-메틸화 플라스미드 DNA를 생성하였다. pCR2.1 벡터를 BspEI 및 XhoI로 절단시켰다. 피복 단백질 융합물들을 수반하는 표적 벡터들을 동일한 효소들로 처리하여 PVX 유전체의 2A 시퀀스 및 3'-말단을 결실시켰다. 계속해서 완전한 피복 단백질 시퀀스를 수반하지 않는 3'-말단을 PVX 유전체 내로 결찰시키고 그리고 시퀀싱에 의해 확인하였다.
본 발명에서 신규한 봉쇄 특성들을 갖는 식물 바이러스 발현 시스템이 개발되었다. 이 시스템에서 2개의 서로 다른 바이러스 벡터들이 상호 피복 단백질 교환에 의하여 서로의 결함성 기능(defective function)을 보상한다. 이들 수단들에 의하여 어떤 바이러스 벡터도 단독으로 전신적으로 식물을 감염시킬 수 없다. 이 실시예에서 기능성 이종 ORF로서 GFP 및 PVX의 피복 단백질을 수반하는 TMV 벡터("제2 벡터"로 나타냄) 및 기능성 이종 ORF로서 mCherry 및 TMV의 CP를 수반하는 PVX 벡터("제1 벡터"로 나타냄) 및 기능성 이종 ORF로서 mBanana 및 CP의 완전 결실을 수반하는 PVX 벡터("제3 벡터"로 나타냄)들이 생성되었다.
PVX 기반 벡터들에 대하여는 아그로박테리움 투메파시엔스 균주 GV3101:pMP90RK 그리고 TMV 기반 벡터들에 대하여는 아그로박테리움 투메파시엔스 균주 GV2260들 내로 바이러스 벡터들을 형질전환시켰다. 아그로박테리움들을 26℃에서 항생제들 카르베니실린(carbenicillin: 100㎎/ℓ), 리팜피신(rifampicin: 50㎎/ℓ) 및 가나마이신(GV3101에 대하여는 50㎎/ℓ 그리고 GV2260에 대하여는 25㎎/ℓ)을 수반하는 YEB 배지(0.5% 소고기 추출물, 0.1% 효모 추출물, 0.5% 펩톤, 0.5% 슈크로스, 2mM MgSO4)에서 성장시켰다. 24시간 후 배양물들에 10μM MES(pH 5.6), 10μM 글루코스 및 20μM 아세토시린곤(acetosyringone)을 보충하고 그리고 하루 더 배양시켰다. 계속해서 배양물들을 2배의 침투성 매질(infiltration media: 100g/ℓ 슈크로스, 3.6g/ℓ 글루코스, 8.6g/ℓ 무라시게와 스쿠그(MS: Murashige and Skoog) 염들, pH 5.6)로 OD600 = 1로 고정시키고, 200μM 아세토시린곤으로 보충하고 그리고 실온에서 30분 동안 배양시켰다. 2 또는 그 이상의 서로 다른 구조물들의 공-접종을 위하여 배양물들을 혼합시켜 각 배양물이 1의 OD600을 가질 수 있도록 하였다. 혼합물들을 바늘 없는 주사기로 약 4주령의 야생담배 잎들 내로 접종시켰다. 식물들을 추가로 온실(phytochamber) 내에서 26℃에서 12시간 동안 일정한 광(25000 내지 30000lux(룩스))으로 그리고 12시간 20℃ 암조건(dark)으로 배양시켰다.
상기 식물들을 선택된 형광 단백질에 대한 특정의 조건들 하에서 매일 모니터링하였다(표 2). GFP 가시화를 위해서는 소형 자외선 램프(handheld UV lamp: 7000㎼, 독일 칼/라인(Kehl/Rhein) 소재, 노보디렉트사(Novodirect))가, mCherry에 대하여는 녹색 LCD 램프(독일 웨츨라 소재, 라이카사(Leica) KL2500)가 그리고 mBanana에 대하여는 고강도 청색 LED 램프(미합중국 뉴욕 소재, 스펙트로닉스 코포레이션(Spectronics corporation), Optimax™450 Spectroline)가 사용되었다. 니콘 쿨픽스 5400 카메라(독일국 뒤셀도르프 소재, 니콘독일사(Nikon Deutschland))를 사용하여 사진들을 촬영하였다.
| 형광 단백질 | 최대 여기(㎚) | 여기 방법 | 최대 방출(㎚) |
| GFP | 395/475 | 자외선 램프(260㎚) | 508 |
| mCherry | 587 | 녹색 램프(515㎚) | 610 |
| mBanana | 540 | 청색 램프(450㎚) | 553 |
도 11에서, 바이러스 벡터들 PVX-mBananaΔCP, PVX-mCherry-CPTMV, TMV-GFP-CPPVX 및 이들의 조합들을 발현하는 접종된 잎들을 나타내었다. TMV 벡터는 그의 피복 단백질을 수반함이 없이 여전히 세포-대-세포 이동을 할 수 있어, 접종된 잎이 이미 GFP의 양호한 발현을 나타내고 있다. 다른 바이러스로부터의 피복 단백질의 상보를 수반하는 벡터들의 조합에서 그 효과는 이미 접종된 잎에서 가시화되고 있다. PVX 벡터들은 TMV 벡터와의 조합에서 보다 더 강한 그들의 이종 ORF를 발현하고 있다. 놀랍게도 또한 피복 단백질이 결핍된 제2 PVX 벡터(mBanana)가 제1 PVX 벡터(mCherry)와 동일한 세포들 내에서 명백하게 발현되었다.
접종 후 26일차(dpi)에서 식물들이 2 또는 심지어 3개의 서로 다른 바이러스 벡터들로 전신적으로 감염되었다. 이는 각각 다른 선천적인 피복 단백질 결핍을 상보하고 그리고 다음으로 추가의 결함성 바이러스 벡터들의 전신적인 이동 능력을 입증하고 있다.
도 6a 및 도 6b에 바이러스 벡터들 TMV-GFP-CPPVX 및 PVX-mCherry-CPTMV + PVX-mBananaΔCP로 공-접종된 전신적으로 감염된 식물로부터의 잎이 나타나 있다.
총 가용성 단백질들의 단리를 위하여 잎들을 서로 다른 시점들에서 수확하고 그리고 2배 용적들의 PBS(pH 7.4)로 균질화시켰다. 4℃에서 13000rpm에서 10분 동안의 원심분리 동안 불용성 식물 부분들이 분리되었다. 가용성 단백질들의 총량을 Roti®Quant 시약(독일국 칼스루헤 소재, 로쓰사(Roth))을 사용하여 브래드포드 분석(Bradford assay)으로 측정하였다.
형광의 정량화를 위하여 단리된 총 가용성 단백질이 사용되었다. 50㎍의 총 단백질을 100㎕의 PBS로 희석시키고 그리고 형광 프로파일들을 마이크로적정 플레이트 판독기(microtiter plate reader: 스위스 만도르프 소재, 테칸그룹리미티드(TECAN Group Ltd.)의 효소결합 면역흡착 분석-판독기(ELISA-Reader Infinite M200))로 측정하였다. 쿠마씨 브릴리언트 블루 염색(Coomassie Brilliant Blue staining) 이전에 형광 단백질들을 또한 SDS 겔들 내에서 가시화시켰다.
단백질 분석을 위하여 불연속 SDS-PAGE 및 웨스턴 블럿팅이 사용되었다. 식물 수액을 5배의 환원 샘플 완충제(reducing sample buffer: 62.5mM 트리스-염산(Tris-HCl), pH 6.8, 30% 글리신, 4% SDS, 10% 2-머캅토에탄올, 0.05% 브로모페놀 블루)로 보충시키고 그리고 직접적으로 겔들 내의 형광의 가시화를 위하여 겔들 상에 적재하거나 또는 웨스턴 블럿팅을 위하여 5 내지 10분 동안 가열시켰다. 계속해서 탐침들을 12% SDS 겔 상으로 적재하고 그리고 전기영동 후 쿠마씨 브릴리언트 블루로 염색시키거나 또는 웨스턴 블럿 분석을 위하여 니트로셀룰로오스 막 상에 블럿팅시켰다. 막들을 1시간 동안 PBS 중의 5% 탈지유(skimmed milk)로 블로킹시켰고 그리고 계속해서 PVX(독일국 브라운스바이그 소재, DSMZ) 또는 TMV(스위스 라이나흐 소재, 바이오레바 악티엔게젤샤프트(Bioreba AG))의 피복 단백질에 대한 폴리클로날 항체로 적어도 2시간 동안 그리고 실온에서 배양시켰다. 2차 항체로서 모노클로날 알칼라인 포스파타아제-결합 염소 항-토끼 항체(monoclonal alkaline phosphatase-conjugated goat anti-rabbit antibody: 독일국 함부르크 소재, 다이아노바사(Dianova))가 사용되었고 그리고 그 신호를 니트로블루 테트라졸리움 클로라이드/5-브로모-4-클로로-3'-인돌리포스페이트 파라-톨루이딘 염(NBT/BCIP: nitroblue tetrazolium chloride/5-bromo-4-chloro-3'-indolyphosphate p-toluidine salt: 독일국 칼스루헤 소재, 로쓰사)로 가시화시켰다.
SDS-PAGE들에서 이종 단백질들의 성공적인 공-발현이 다시 확인되었다(도 7). 서로 다른 형광 단백질들이 공-감염된 잎들로부터의 식물 수액 중에서 특정의 여기 하에서 쉽게 가시화되었다. mCherry 및 mBanana에 대하여는 이들 2개의 형광 단백질들의 이량체화(dimerization)가 또한 가시화되어 단백질 단량체의 크기의 2배의 단백질 밴드(protein band)를 형성한다. 바이러스 벡터들로부터의 서로 다른 피복 단백질들의 발현 및 그들의 원래의 선천적인 피복 단백질의 결핍들이 TMV 또는 PVX 피복 단백질에 대한 폴리클로날 항체들에 대한 웨스턴 블럿 분석에서 입증되었다(도 8).
이들 데이터는 바이러스들 중의 원래의 피복 단백질의 결핍 및 외래 피복 단백질의 발현과 마찬가지로 공-감염들에서의 피복 단백질 기능의 이종성 상보를 확증한다.
동일한 세포들 내에서의 2개의 서로 다른 PVX 기반 벡터들에 의한 mCherry 및 mBanana의 공-발현을 확증하기 위하여, 감염된 잎들을 형광 현미경 바이오레보 비지-9000(fluorescence microscope Biorevo BZ-9000: 독일국 뉴젠버그 소재, 케이엔스사(Keyence))으로 분석하였다. 상보성 쌍으로 감염된 잎들은 동일 세포들 내에서 mCherry 및 GFP의 명백한 공-발현을 나타내었고(도 12a 내지 도 12d) 그리고 mBanana에 대한 발현에 대한 신호를 나타내지 않았다(도 12e 내지 도 12h). 상보쌍 및 PVX-mBananaΔCP로 공-감염된 잎들에서 mBanana 및 mCherry의 명백한 공-발현을 볼 수 있다(도 12i 내지 도 12l). 이들 영역들에서 GFP 발현이 또한 확증되었다(데이터는 나타내지 않음).
본 발명자들은 추가로 결핍 CP 유전자를 수반하는 제2 TMV 벡터가 본 발명자들의 상보쌍과 함께 식물들을 공-감염시킬 수 있는 지의 여부를 분석하였다. 그러나, 감염은 감염된 잎들에서 단지 2개의 서로 다른 PVX-기반 벡터들의 공-발현 만을 나타내었다(도 13). mBanana 발현이 모든 침투된 잎들 내에서 관측될 수 있기 때문에, 침투된 잎들은 처음에는 상보쌍이 제3 TMV 또는 PVX 벡터와 함께 공-발현되는 것으로 보였다. 전신적으로 감염된 잎들에서 단지 부가의 PVX 벡터와의 조합 만이 모든 3가지 형광 단백질들의 공-발현을 나타내었다. TMV-mBananaΔCP와의 조합에서 상보쌍 만이 동일한 영역들 내에서 발견될 수 있었다. 이 조합에서 mBanana 발현의 점들을 거의 발견할 수 없었으며, 이는 절대 GFP 발현과 중첩되지 않는다. 따라서, PVX에 의하여 발현된 CPTMV가 CP 결핍 TMV 벡터를 상보할 수 있고 그리고 접종된 잎의 확산을 허용하나, 그러나 2개의 서로 다른 TMV-기반 벡터들의 공-감염은 관측되지 않았다.
이들 발견들은 SDS-PAGE 및 웨스턴 블럿 분석에서 확증되었다(도 14). 겔들 내의 형광 단백질들의 가시화는 전신적 감염된 잎들 중의 상보쌍에 의한 mCherry 및 GFP의 공-발현 및 PVX-mBananaΔCP 벡터와의 공-감염들에서의 mBanana의 공-발현을 나타내었다. 게다가 세포들 내에서 mBanana가 겔들 내에서 보다 낮은 밴드를 나타내지 않고 mCherry 및 mBanana가 공-발현되는 경우에 이들의 이량체화가 관측되었다. 부가의 TMV-mBananaΔCP로 감염된 식물들에서 소량의 mBanana가 이들 잎들에서 감소된 양의 GFP 및 mCherry를 수반하는 잎들에서 발견될 수 있다. 이는 이들 벡터들의 공-발현의 손실로 설명될 수 있다. 상보쌍은 이들 잎들에서 GFP 및 mCherry를 공-발현하나 그러나 mBanana를 발현하는 제2 TMV 벡터로부터 분리되고, 이는 전체 저감된 양의 발현된 재조합 단백질들을 야기한다. 웨스턴 블럿은 상보쌍에 의한 바이러스 벡터들의 CP들의 발현을 확증하였다.
요약하면 본 발명자들은 제2 TMV 벡터가 상보쌍을 수반함이 없이 식물들을 공-감염시킬 수 없는 반면에 상보쌍의 피복 단백질이 결핍된 부가의 PVX-기반 벡터와의 공-발현을 확증하였다. 이들 벡터들에 대하여 본 발명자들은 처음으로 전신적 감염된 잎들에서 동일한 RNA 바이러스에 기초하는 2개의 바이러스 벡터들의 공-발현을 보였다.
전신적 이동 및 재조합 단백질들의 동시적 생산을 가능하게 하고 그리고 동시에 생물안전성 봉쇄를 보증하는 피복 단백질 상보의 가능성이 본 발명의 잇점으로 명백하게 보여지고 있다.
실시예
2: 이동-결핍
PVX
벡터들로의 단백질 생산
하나의 바이러스 종들로부터 파생된 바이러스 벡터들로의 2 또는 그 이상의 재조합 단백질들의 발현을 위하여, 감자 바이러스 X(PVX) 벡터들을 구축하였으며 이들에는 피복 단백질이 결핍된다. 공지의 벡터들이 피복 단백질이 보존되는 C-말단 코딩 영역의 일부인 것과는 대조적으로 이들 벡터들에서 전체 피복 단백질 오픈리딩프레임이 결실되었다.
상기 바이러스 벡터들을 아그로박테리움 투메파시엔스 균주 GV3101:pMP90RK 내로 형질전환시키고 그리고 26℃에서 항생제들 카르베니실린(100㎎/ℓ), 리팜피신(50㎎/ℓ) 및 가나마이신(50㎎/ℓ)을 수반하는 YEB 배지(0.5% 소고기 추출물, 0.1% 효모 추출물, 0.5% 펩톤, 0.5% 슈크로스, 2mM MgSO4)에서 성장시켰다. 24시간 후 배양물들에 10μM MES(pH 5.6), 10μM 글루코스 및 20μM 아세토시린곤(acetosyringone)을 보충하고 그리고 하루 더 배양시켰다. 계속해서 배양물들을 2배의 침투성 매질(100g/ℓ 슈크로스, 3.6g/ℓ 글루코스, 8.6g/ℓ 무라시게와 스쿠그(MS) 염들, pH 5.6)로 OD600 = 1로 고정시키고, 200μM 아세토시린곤으로 보충하고 그리고 실온에서 30분 동안 배양시켰다. 2 또는 그 이상의 서로 다른 구조물들의 공-접종을 위하여 배양물들을 혼합시켜 각 배양물이 1의 OD600을 가질 수 있도록 하였다. 혼합물들을 바늘 없는 주사기로 약 4주령의 야생담배 잎들 내로 접종시켰다. 식물들을 추가로 온실 내에서 26℃에서 12시간 동안 일정한 광(25000 내지 30000lux(룩스))으로 그리고 12시간 20℃ 암조건으로 배양시켰다.
상기 식물들을 선택된 형광 단백질에 대한 특정의 조건들 하에서 매일 모니터링하였다(표 3). GFP 가시화를 위해서는 소형 자외선 램프(7000㎼, 독일 칼/라인 소재, 노보디렉트사)가, mCherry에 대하여는 녹색 LCD 램프(독일 웨츨라 소재, 라이카사 KL2500)가 사용되었다. 니콘 쿨픽스 5400 카메라(독일국 뒤셀도르프 소재, 니콘독일사)를 사용하여 사진들을 촬영하였다.
| 형광 단백질 | 최대 여기(㎚) | 여기 방법 | 최대 방출(㎚) | 관측 |
| GFP | 395/475 | 자외선 램프(260㎚) | 508 | |
| mCherry | 587 | 녹색 램프(515㎚) | 610 | 적색 필터 |
그 결과들은 명백하게 피복 단백질의 C-말단 코딩 영역이 PVX 벡터의 복제를 위하여 엄격히 요구되지 않는다는 것을 나타내고 있다. GFP 및 mCherry들이 접종된 식물 세포들 내에서 생산되었다(도 9). 공-접종된 잎들에서 2개의 서로 다른 재조합 단백질들의 공-발현이 명백하게 가시화되고 그리고 현미경 연구들에서 세포 수준에서 입증되었다(데이터는 나타내지 않음).
접종 후 4일차에서 두 형광 단백질들의 양호한 공-발현이 가시화되었고 그리고 발현은 이 효소 조합에 대하여 접종 후 6일차에서 최고의 공-발현 수준들에 도달하였다(도 10). 그러나 서로 다른 재조합 단백질의 서로 다른 요건들로 인하여 다른 생산 시간들이 또한 가능하거나 및/또는 적절하다.
총 가용성 단백질들의 단리를 위하여 잎들을 서로 다른 시점들에서 수확하고 그리고 2배 용적들의 PBS(pH 7.4)로 균질화시켰다. 4℃에서 13000rpm에서 10분 동안의 원심분리 동안 불용성 식물 부분들이 분리되었다. 가용성 단백질들의 총량을 Roti®Quant 시약(독일국 칼스루헤 소재, 로쓰사)을 사용하여 브래드포드 분석으로 측정하였다.
형광의 정량화를 위하여 단리된 총 가용성 단백질이 사용되었다. 50㎍의 총 단백질을 100㎕의 PBS로 희석시키고 그리고 형광 프로파일들을 마이크로적정 플레이트 판독기(스위스 만도르프 소재, 테칸그룹리미티드(TECAN Group Ltd.)의 효소결합 면역흡착 분석-판독기(ELISA-Reader Infinite M200))로 측정하였다. 쿠마씨 브릴리언트 블루 염색 이전에 형광 단백질들을 또한 SDS 겔들 내에서 가시화시켰다.
단백질 분석을 위하여 불연속 SDS-PAGE 및 웨스턴 블럿팅이 사용되었다(문헌 Laemmli, 1970 참조). 식물 수액을 5배의 환원 샘플 완충제(62.5mM 트리스-염산, pH 6.8, 30% 글리신, 4% SDS, 10% 2-머캅토에탄올, 0.05% 브로모페놀 블루)로 보충시키고 그리고 직접적으로 겔들 내의 형광의 가시화를 위하여 겔들 상에 적재하거나 또는 웨스턴 블럿팅을 위하여 5 내지 10분 동안 가열시켰다. 계속해서 탐침들을 12% SDS 겔 상으로 적재하고 그리고 전기영동 후 쿠마씨 브릴리언트 블루로 염색시키거나 또는 웨스턴 블럿 분석을 위하여 니트로셀룰로오스 막 상에 블럿팅시켰다. 막들을 1시간 동안 PBS 중의 5% 탈지유로 블로킹시켰고 그리고 계속해서 PVX(독일국 브라운스바이그 소재, DSMZ)의 피복 단백질에 대한 폴리클로날 항체로 적어도 2시간 동안 그리고 실온에서 배양시켰다. 2차 항체로서 모노클로날 알칼라인 포스파타아제-결합 염소 항-토끼 항체(독일국 함부르크 소재, 다이아노바사(Dianova))가 사용되었고 그리고 그 신호를 니트로블루 테트라졸리움 클로라이드/5-브로모-4-클로로-3'-인돌리포스페이트 파라-톨루이딘 염(NBT/BCIP: 독일국 칼스루헤 소재, 로쓰사)로 가시화시켰다.
SDS-PAGE 및 웨스턴 블럿 분석으로 PVX 벡터들이 어떠한 피복 단백질도 발현하지 않는다는 것이 입증될 수 있었다(데이터는 나타내지 않음). 비록 이전 간행물들이 PVX 벡터의 복제를 위하여 피복 단백질의 C-말단 코딩 영역에 대한 요건들이 나타나 있음에도 불구하고, 본 발명에서는 피복 단백질 코딩 시퀀스가 완전히 결실될 수 있다는 것을 보일 수 있었다. 대조적으로 2개의 서로 다른 그리고 보다 잠재적인 PVXΔCP 벡터들의 양호한 공-발현을 볼 수 있었다.
동일한 세포들 내에서의 2개의 서로 다른 PVX 기반 벡터들의 공-발현을 확증하기 위하여, 감염된 잎들을 형광 현미경 바이오레보 비지-9000(독일국 뉴젠버그 소재, 케이엔스사)으로 분석하였다. 사진들은 미-감염된 야생담배 식물에 대하여 어떠한 신호들도 확증되지 않았다(도 15a 내지 도 15d). PVX-GFPΔCP 또는 PVX-mCherryΔCP로의 감염들에 있어서 단지 발현된 형광 단백질만 볼 수 있었다(GFP에 대하여는 E 내지 H, mCherry에 대하여는 I 내지 L). 이들 벡터들의 공-감염들에서 서로 다른 형광 단백질들의 공-발현이 동일한 셀들 내에서 관측될 수 있다(M 내지 P).
요약하면 본 발명자들은 아그로침투된 잎들(agroinfiltrated leaves)에서 그의 CP가 결핍된 서로 다른 PVX-기반 벡터들의 공-발현을 확증하였다. 이러한 별견들은 PVX 벡터들이 CP 기능들의 상실(knockout) 이후 공-감염시킬 수 있다는 것을 나타내고 있다. 이는 벡터들에서 MP 및 CP 유전자들의 결실 이후조차에서도 공-감염을 할 수 없는 앞서 언급된 공지의 PVX-기반 벡터들 및 또한 TMV-기반 벡터들과는 대조적이다(문헌 Julve et al., 2013 참조).
피복 단백질 결실로 인하여, 봉쇄가 보장된다. 게다가, 본 발명은 하나의 셀 내에서의 2 또는 그 이상의 부적합성(incompatibility)을 극복하고 그리고 그에 따라 동일 식물 세포들 내의 서로 다른 바이러스 벡터들 상에 위치된 2 또는 그 이상의 이종 ORF들의 동시적 발현을 허용한다.
문헌들:
Fedorkin, O.N., Merits, A., Lucchesi, J., Solovyev, A.G., Saarma, M., Morozov, S.Y., Makinen, K., 2000. Complementation of the movement-deficient mutations in potato virus X: potyvirus coat protein mediates cell-to-cell trafficking of C-terminal truncation but not deletion mutant of potexvirus coat protein. Virology 270, 31-42.
Julve, J.M., Gandia, A., Fernandez-Del-Carmen, A., Sarrion-Perdigones, A., Castelijns, B., Granell, A., Orzaez, D., 2013. A coat-independent superinfection exclusion rapidly imposed in Nicotiana benthamiana cells by tobacco mosaic virus is not prevented by depletion of the movement protein. Plant Mol Biol 81, 553-564.
Komarova, T.V., Skulachev, M.V., Zvereva, A.S., Schwartz, A.M., Dorokhov, Y.L., Atabekov, J.G., 2006. New viral vector for efficient production of target proteins in plants. Biochemistry (Mosc) 71, 846-850.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685.
SEQUENCE LISTING
<110> Fraunhofer-Gesellschaft zur F?derung der angewandten
Forschung e.V.
Rheinisch-Westf?ische Technische Hochschule Aachen
<120> Kits comprising plus-sense single stranded RNA viral vectors and
methods for producing polypeptides using the kits
<130> F68470PC
<150> EP 13 160 627.9
<151> 2013-03-22
<160> 11
<170> PatentIn version 3.5
<210> 1
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> M13 universe primer
<400> 1
gttgtaaaac gacggccagt 20
<210> 2
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> mCherry-ClaI primer
<400> 2
tagcatcgat atggtgagca ag 22
<210> 3
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> PacI-GFP-TMV primer
<400> 3
tcattaatta aatggctagc 20
<210> 4
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> SOE2-CPfw primer
<400> 4
agtacgtttt aatcaatatg tcagcaccag ctagcac 37
<210> 5
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> SOE2-CP-rv primer
<400> 5
tgctagctgg tgctgacata ttgattaaaa cgtactc 37
<210> 6
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> SOE2-CP-NotI-rv primer
<400> 6
aatagcggcc gctatggtgg tggtag 26
<210> 7
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> SOE-TMVCP-fw primer
<400> 7
attgatactc gaaagatgcc ttatacaatc 30
<210> 8
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> SOE-TMVCP-rv primer
<400> 8
attgtataag gcatctttcg agtatcaatg 30
<210> 9
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> SOE-TMVCP2-fw primer
<400> 9
aactccggct acttaactac gtctacataa c 31
<210> 10
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> SOE-TMVCP2-rv primer
<400> 10
agacgtagtt aagtagccgg agttg 25
<210> 11
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> 2ADelCPfw primer
<400> 11
aatccggata actacgtcta cataaccg 28
Claims (19)
- a) 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및
b) 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터를 포함하되,
(i) 상기 제1 플러스-센스 단일 가닥 바이러스 벡터 및 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 서로 다른 식물 바이러스들로부터 파생되고,
(ii) 상기 제1 벡터가 파생되는 바이러스의 피복 단백질 ORF가 상기 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 내에서 완전히 결실되고,
(iii) 상기 제2 벡터가 파생되는 바이러스의 피복 단백질 ORF가 상기 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터 내에서 완전히 결실되고,
(iv) 상기 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터가 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF를 포함하고,
(v) 상기 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터가 상기 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF를 포함하며,
(vi) 상기 제1 플러스-센스 단일 가닥 바이러스 벡터 및 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 식물 세포들 내에서 복제될 수 있는 RNA 복제단위(replicon)를 포함하는, 킷트.
- 제 1 항에 있어서,
a) 상기 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 피복 단백질 ORF가 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF로 치환되고/되거나,
b) 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 피복 단백질 ORF가 상기 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 기능성 피복 단백질 ORF로 치환되는, 킷트. - 제 1 항 또는 제 2 항에 있어서,
상기 제1 및 제2 플러스-센스 단일 가닥 바이러스 벡터들이 파생되는 바이러스들이 서로 다른 바이러스 형(type), 바람직하게는 서로 다른 바이러스 속(genus), 보다 바람직하게는 서로 다른 바이러스 과(family)에 속하는, 킷트. - 제 1 항 내지 제 3 항 중 어느 한 항에 있어서,
a) 상기 제1 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이고/이거나,
b) 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스가 토바모바이러스, 특히 TMV 바이러스(담배 모자이크 바이러스)인, 킷트.
- 제 1 항 내지 제 4 항 중 어느 한 항에 있어서,
a) 상기 제1 플러스-센스 단일 가닥 바이러스 벡터 및/또는 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 기능성 이종 ORF를 포함하고/하거나,
b) 상기 제1 플러스-센스 단일 가닥 바이러스 벡터 및/또는 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 기능성 이종 ORF를 포함하지 않는, 킷트. - 제 1 항 내지 제 5 항 중 어느 한 항에 있어서,
a) 적어도 하나의 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터, 및/또는
b) 적어도 하나의 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터를 더 포함하되,
(i) 상기 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 적어도 하나의 기능성 이종 ORF를 포함하고,
(ii) 상기 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 상기 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터와 동일한 식물 바이러스로부터 파생되고,
(iii) 상기 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 파생되는 바이러스의 피복 단백질 ORF가 상기 적어도 하나의 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터(들) 내에서 완전히 결실되며,
(i) 상기 적어도 하나의 부가의 플러스-센스 단일 가닥 바이러스 벡터(들)가 적어도 하나의 기능성 이종 ORF를 포함하고,
(ii) 상기 적어도 하나의 부가의 플러스-센스 단일 가닥 바이러스 벡터(들)가 상기 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터와 동일한 식물 바이러스로부터 파생되는, 킷트. - 제 6 항에 있어서,
(i) 상기 적어도 하나의 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)에 상기 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 파생되는 바이러스의 기능성 이동 단백질 ORF(들)가 결핍되고, 바람직하게는 상기 적어도 하나의 제3 플러스-센스 단일 가닥 바이러스 벡터(들)가 파생되는 바이러스의 기능성 이동 단백질 ORF(들)가 완전히 결실되고/되거나,
(ii) 상기 제3 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이고/이거나,
(iii) 상기 적어도 하나의 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)가:
a) 청구항 제1항에 따른 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터의 특징들을 포함하고,
b) 적어도 하나의 기능성 이종 ORF를 포함하며,
c) 상기 제1 및 제2 플러스-센스 단일 가닥 바이러스 벡터들 중 하나 또는 둘 모두가 기능성 이종 ORF(들)를 포함하는 경우, 상기 적어도 하나의 기능성 이종 ORF가 상기 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터의 기능성 이종 ORF(들)과 상이하고,
바람직하게는 상기 제3 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이고 상기 제3 플러스-센스 단일 가닥 바이러스 벡터에 상기 포텍스바이러스의 기능성 이동 단백질 ORF(들)가 결핍되며, 또는
바람직하게는 상기 적어도 하나의 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)가 기능성 이종 ORF에 의해 상기 제1 또는 제2 플러스-센스 단일 가닥 바이러스 벡터와 상이하고, 또는
바람직하게는 상기 적어도 하나의 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)가
a) 상기 제1 플러스-센스 단일 가닥 바이러스 벡터 또는 상기 제2 플러스-센스 단일 가닥 바이러스 벡터가 파생되는 바이러스의 피복 단백질, 이동 단백질(들) 및 RNA-의존성 RNA 중합효소를 포함하며,
b) 적어도 하나의 기능성 이종 ORF를 포함하는, 킷트. - 제 6 항 또는 제 7 항에 있어서,
a) 둘 이상의 서로 다른 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 서로 다른 기능성 이종 ORF(들)을 포함하고/하거나,
b) 상기 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)가 하나의 기능성 이종 ORF를 포함하는, 킷트. - 혼합물로서,
a) 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터,
b) 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터,
c) 청구항 제 6 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터
중 2, 3 또는 그보다 많은 서로 다른 벡터들을 포함하는, 혼합물. - 제 9 항에 있어서,
a)가
(i) 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터,
(ii) 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터 및
(iii) 선택적으로 청구항 제 6 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터를 포함하거나, 또는
b)가
(i) 청구항 제 6 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터 및
(ii) 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및/또는 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터를 포함하는, 혼합물. - a) 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터와, 선택적으로 청구항 제 6 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)를 포함하는, 식물 또는 식물 세포.
- 제 11 항에 있어서,
하나보다 많은 조직 내의 하나보다 많은 식물 세포가
a) 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 벡터와, 선택적으로 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 벡터(들)를 포함하고/하거나,
b) 청구항 제 6 항 내지 제 8 항 중의 하나에서 정의된 바와 같은 적어도 2개의 서로 다른 제3 또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터(들)의 ORF들로 인코딩되는 이종 폴리펩티드들을 포함하는, 식물. - 식물, 또는 식물 조직, 또는 식물 세포 내에 헤테로올리고머 폴리펩티드 및/또는 둘 이상의 폴리펩티드를 생산하는 방법으로서, 적어도 하나의 식물 세포에:
(i) 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터,
(ii) 청구항 제 1 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터 및
(iii) 선택적으로 청구항 제 6 항 내지 제 8 항에서 정의된 바와 같은 적어도 하나의 제3 및/또는 부가의 플러스-센스 단일 가닥 RNA 바이러스 벡터들을 제공하는 단계를 포함하되,
a) 상기 제1 플러스-센스 단일 가닥 RNA 바이러스 벡터 및 상기 제2 플러스-센스 단일 가닥 RNA 바이러스 벡터가 기능성 이종 ORF를 포함하지 않는다면 적어도 하나의 식물 세포에 적어도 2개의 제3 플러스-센스 단일 가닥 RNA 바이러스 벡터가 제공되며,
b) (i) 내지 (iii)에 따른 바이러스 벡터들 중 적어도 2개가 서로 다른 이종 ORF들을 포함하는, 방법. - 제 13 항에 있어서,
a) 후속하여 상기 식물, 식물 조직, 또는 식물 세포로부터 헤테로올리고머 폴리펩티드 및/또는 둘 이상의 폴리펩티드들을 단리하고/하거나,
b) 상기 식물의 전신적 감염이 달성되고/되거나,
c) 상기 헤테로올리고머 폴리펩티드가 면역글로블린, 특히 항체 또는 항체 단편인, 방법. - 식물, 또는 식물 조직, 또는 식물 세포 내에 헤테로올리고머 폴리펩티드 및/또는 둘 이상의 폴리펩티드를 생산하는 방법으로서,
식물, 또는 식물 조직, 또는 식물 세포에 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들을 제공하는 단계를 포함하되,
a) 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 동일한 바이러스로부터 파생되고,
b) 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 서로 다른 기능성 이종 ORF들을 포함하고,
c) 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들에 상기 벡터들이 파생되는 바이러스의 기능성 피복 단백질 ORF가 결핍되며,
d) 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 식물 세포들 내에서 복제할 수 있는 RNA 복제단위를 포함하는, 방법. - 제 15 항에 있어서,
a) 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 아그로감염에 의해, DNA, 특히 바이러스 유전자의 cDNA를 포함하는 DNA를 이용한 형질주입에 의해, RNA, 특히 바이러스 유전자에 상응하는 RNA를 이용한 형질주입에 의해, 또는 바이러스 입자들을 이용한 감염에 의해 제공되고/되거나,
b) 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이고/이거나,
c) 상기 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스의 피복 단백질 ORF가 적어도 2개의 플러스-센스 단일 가닥 RNA 바이러스 벡터들 내에서 완전히 결실되는, 방법. - 아그로박테리움 세포들, 특히 아그로박테리움 투메파시엔스 세포들(Agrobacterium tumefaciens cells)의 혼합물로서,
a) 복수의 아그로박테리움 세포들이 적어도 하나의 기능성 이종 ORF를 포함하는 적어도 하나의 플러스-센스 단일 가닥 RNA 바이러스 벡터를 포함하되,
i) 상기 적어도 하나의 플러스-센스 단일 가닥 RNA 바이러스 벡터에 기능성 피복 단백질 ORF가 결핍되고,
ii) 상기 적어도 하나의 플러스-센스 단일 가닥 RNA 바이러스 벡터가 식물 세포들 내에서 복제할 수 있는 RNA 복제단위를 포함하며,
b) 적어도 2개의 아그로박테리움 세포들이 서로 다른 기능성 이종 ORF들을 포함하는 서로 다른 플러스-센스 단일 가닥 RNA 벡터들을 포함하고,
c) 상기 서로 다른 플러스-센스 단일 가닥 RNA 벡터들이 동일한 바이러스로부터 파생되는 서로 다른 기능성 이종 ORF들을 포함하며,
바람직하게는
i) 상기 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스가 포텍스바이러스, 특히 PVX 바이러스(감자 바이러스 X)이고/이거나,
ii) 상기 플러스-센스 단일 가닥 RNA 바이러스 벡터들이 파생되는 바이러스의 피복 단백질 ORF가 상기 플러스-센스 단일 가닥 RNA 바이러스 벡터들 내에서 완전히 결실되는, 혼합물. - PVX 바이러스 벡터로서,
a) PVX 바이러스의 기능성 피복 폴리펩티드 ORF가 완전히 결실되고,
b) 상기 PVX 바이러스 벡터가 식물 세포들 내에서 복제할 수 있는 RNA 복제단위를 포함하고,
c) 상기 PVX 바이러스 벡터가 선택적으로 적어도 하나의 기능성 이종 ORF를 포함하며,
d) 상기 PVX 바이러스 벡터가 선택적으로 서로 다른 바이러스의 기능성 피복 폴리펩티드 ORF를 포함하는, PVX 바이러스 벡터. - 폴리펩티드, 특히 헤테로올리고머 폴리펩티드를 생산하기 위한 제 1 항 내지 제 8 항 중의 어느 하나에 따른 킷트, 또는 제 9 항 또는 제 10 항에 따른 혼합물, 또는 제 18 항에 따른 PVX 바이러스 벡터, 또는 제 11 항에 따른 식물 또는 식물 세포, 또는 제 12 항에 따른 식물, 또는 제 17 항에 따른 아그로박테리움 세포들의 혼합물의 사용.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP13160627 | 2013-03-22 | ||
| EP13160627.9 | 2013-03-22 | ||
| PCT/EP2014/055730 WO2014147235A1 (en) | 2013-03-22 | 2014-03-21 | Kits comprising plus-sense single stranded rna viral vectors and methods for producing polypeptides using the kits |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20150132096A true KR20150132096A (ko) | 2015-11-25 |
Family
ID=47913231
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020157020304A Withdrawn KR20150132096A (ko) | 2013-03-22 | 2014-03-21 | 플러스-센스 단일 가닥 rna 바이러스 벡터 및 킷트를 사용하여 폴리펩티드를 생산하기 위한 방법 |
Country Status (8)
| Country | Link |
|---|---|
| US (1) | US10190128B2 (ko) |
| EP (1) | EP2976428B1 (ko) |
| JP (1) | JP2016514465A (ko) |
| KR (1) | KR20150132096A (ko) |
| AU (1) | AU2014234251A1 (ko) |
| CA (1) | CA2904472A1 (ko) |
| WO (1) | WO2014147235A1 (ko) |
| ZA (1) | ZA201504333B (ko) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN110885796A (zh) * | 2018-08-20 | 2020-03-17 | 山东农业大学 | 一种抗马铃薯x病毒的弱毒疫苗、制备方法及其应用 |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP7011327B2 (ja) * | 2017-02-15 | 2022-02-10 | 国立研究開発法人農業・食品産業技術総合研究機構 | 植物ウイルスベクターを利用したゲノム編集植物の生産方法 |
| EP3880828A4 (en) | 2018-11-13 | 2022-08-03 | University of Maryland, College Park | PLANT VECTORS, COMPOSITIONS AND USES RELATED THERETO |
| JPWO2020226176A1 (ko) * | 2019-05-09 | 2020-11-12 | ||
| US20220090107A1 (en) * | 2019-05-23 | 2022-03-24 | Nomad Bioscience Gmbh | Rna viral rna molecule for gene editing |
| CN115052979A (zh) | 2019-11-12 | 2022-09-13 | 马里兰大学派克分院 | 植物载体、组合物及其相关用途 |
Family Cites Families (21)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| NL8300698A (nl) | 1983-02-24 | 1984-09-17 | Univ Leiden | Werkwijze voor het inbouwen van vreemd dna in het genoom van tweezaadlobbige planten; agrobacterium tumefaciens bacterien en werkwijze voor het produceren daarvan; planten en plantecellen met gewijzigde genetische eigenschappen; werkwijze voor het bereiden van chemische en/of farmaceutische produkten. |
| CA1280081C (en) | 1984-09-24 | 1991-02-12 | Calgene, Inc. | Plant cell microinjection technique |
| US5100792A (en) | 1984-11-13 | 1992-03-31 | Cornell Research Foundation, Inc. | Method for transporting substances into living cells and tissues |
| AU7360087A (en) | 1986-04-30 | 1987-11-24 | Boyce Thompson Institute For Plant Research Inc. | Electric field mediated dna transformation of plant cells and organelles |
| ES2061720T3 (es) | 1987-05-05 | 1994-12-16 | Sandoz Ag | Transformacion de tejido vegetal. |
| ATE130371T1 (de) | 1989-12-19 | 1995-12-15 | Ciba Geigy Ag | Verfahren und vorrichtung zur genetischen transformation von zellen. |
| CA2036935A1 (en) | 1990-02-26 | 1991-08-27 | Paul Christou | Plant transformation process with early identification of germ line transformation events |
| ES2260886T3 (es) | 1990-11-23 | 2006-11-01 | Bayer Bioscience N.V. | Procedimiento para transformar plantas monocotiledoneas. |
| US5384253A (en) | 1990-12-28 | 1995-01-24 | Dekalb Genetics Corporation | Genetic transformation of maize cells by electroporation of cells pretreated with pectin degrading enzymes |
| WO1994000583A1 (en) | 1992-06-23 | 1994-01-06 | South Dakota State University | Transformation of plants by direct injection of dna |
| ATE398679T1 (de) | 1992-07-07 | 2008-07-15 | Japan Tobacco Inc | Verfahren zur transformation einer monokotyledon pflanze |
| CA2202761A1 (en) | 1994-10-18 | 1996-04-25 | Sean Nicholas Chapman | Method of producing a chimeric protein |
| EP1147178A1 (en) | 1999-02-05 | 2001-10-24 | Thomas Jefferson University | Production of biomedical peptides and proteins in plants using transcomplementation systems |
| KR20020013508A (ko) * | 1999-03-09 | 2002-02-20 | 유니버시티 오브 플로리다 | 외래 서열을 발현시키기위한 다성분 rna 벡터 시스템 |
| DE10143205A1 (de) | 2001-09-04 | 2003-03-20 | Icon Genetics Ag | Verfahren zur Proteinproduktion in Pflanzen |
| ES2531125T3 (es) * | 2003-02-03 | 2015-03-10 | Ibio Inc | Sistema para la expresión de genes en plantas |
| EP1686176A1 (en) | 2005-01-28 | 2006-08-02 | Icon Genetics AG | Production of antibodies in plants with plus-sense single-stranded viral RNA vectors |
| EP1897953A1 (en) | 2006-09-06 | 2008-03-12 | Icon Genetics GmbH | Potexvirus-derived replicon |
| EP2118274A4 (en) | 2007-01-29 | 2010-09-01 | Univ Ohio State Res Found | GENE EXPRESSION SYSTEM IN PLANTS WITH A VIRUS-BASED EXPRESSION VECTOR |
| RU2370280C2 (ru) * | 2007-10-08 | 2009-10-20 | Институт физико-химической биологии им. А.Н.Белозерского | СПОСОБ СУПЕРПРОДУКЦИИ В РАСТЕНИИ АНТИТЕЛ ПРОТИВ ОНКОГЕНА HER2/neu |
| EP2418283A1 (en) * | 2010-08-07 | 2012-02-15 | Nomad Bioscience GmbH | Process of transfecting plants |
-
2014
- 2014-03-21 US US14/764,112 patent/US10190128B2/en not_active Expired - Fee Related
- 2014-03-21 WO PCT/EP2014/055730 patent/WO2014147235A1/en not_active Ceased
- 2014-03-21 AU AU2014234251A patent/AU2014234251A1/en not_active Abandoned
- 2014-03-21 KR KR1020157020304A patent/KR20150132096A/ko not_active Withdrawn
- 2014-03-21 EP EP14711768.3A patent/EP2976428B1/en active Active
- 2014-03-21 CA CA2904472A patent/CA2904472A1/en not_active Abandoned
- 2014-03-21 JP JP2016503677A patent/JP2016514465A/ja active Pending
-
2015
- 2015-06-15 ZA ZA2015/04333A patent/ZA201504333B/en unknown
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN110885796A (zh) * | 2018-08-20 | 2020-03-17 | 山东农业大学 | 一种抗马铃薯x病毒的弱毒疫苗、制备方法及其应用 |
| CN110885796B (zh) * | 2018-08-20 | 2022-04-08 | 山东农业大学 | 一种抗马铃薯x病毒的弱毒疫苗、制备方法及其应用 |
Also Published As
| Publication number | Publication date |
|---|---|
| US10190128B2 (en) | 2019-01-29 |
| CA2904472A1 (en) | 2014-09-25 |
| WO2014147235A1 (en) | 2014-09-25 |
| JP2016514465A (ja) | 2016-05-23 |
| EP2976428A1 (en) | 2016-01-27 |
| EP2976428B1 (en) | 2019-11-27 |
| AU2014234251A1 (en) | 2015-06-18 |
| US20160145635A1 (en) | 2016-05-26 |
| ZA201504333B (en) | 2017-07-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR101515044B1 (ko) | 식물 바이러스 기반 유도성 발현 시스템 | |
| EP2061890B1 (en) | Potexvirus- derived replicon | |
| US10287602B2 (en) | RNA virus-derived plant expression system | |
| EP1564295A1 (en) | RNA virus-derived plant expression system | |
| EP2976428B1 (en) | Kits comprising plus-sense single stranded rna viral vectors and methods for producing polypeptides using the kits | |
| AU2004291658B2 (en) | RNA virus-derived plant expression system | |
| US9267143B2 (en) | RNA virus-derived plant expression system | |
| CN101454456B (zh) | 植物基于病毒的诱导型表达系统 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PA0105 | International application |
Patent event date: 20150724 Patent event code: PA01051R01D Comment text: International Patent Application |
|
| N231 | Notification of change of applicant | ||
| PN2301 | Change of applicant |
Patent event date: 20150908 Comment text: Notification of Change of Applicant Patent event code: PN23011R01D |
|
| PG1501 | Laying open of application | ||
| PC1203 | Withdrawal of no request for examination | ||
| WITN | Application deemed withdrawn, e.g. because no request for examination was filed or no examination fee was paid |