발명의 상세한 설명
달리 지시되지 않는 한, 본 발명의 실행은 관련 기술 분야의 통상의 기술자의 능력 내에 있는, 화학, 분자생물학, 미생물학, 재조합 DNA 기술 및 화학적 방법의 통상적인 기법을 이용한다. 이같은 기법들이 문헌, 예를 들어, [M.R. Green, J. Sambrook, 2012, Molecular Cloning: A Laboratory Manual, Fourth Edition, Books 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY]; [Ausubel, F. M. et al. (1995 and periodic supplements; Current Protocols in Molecular Biology, ch. 9, 13, and 16, John Wiley & Sons, New York, N. Y.)]; [B. Roe, J. Crabtree, and A. Kahn, 1996, DNA Isolation and Sequencing: Essential Techniques, John Wiley & Sons]; [J. M. Polak and James O'D. McGee, 1990, In Situ Hybridisation: Principles and Practice, Oxford University Press]; [M. J. Gait (Editor), 1984, Oligonucleotide Synthesis: A Practical Approach, IRL Press]; 및 [D. M. J. Lilley and J. E. Dahlberg, 1992, Methods of Enzymology: DNA Structure Part A: Synthesis and Physical Analysis of DNA Methods in Enzymology, Academic Press]에 또한 설명되어 있다. 각각의 이러한 일반 교재가 본원에서 참고로 포함된다.
본 발명을 설명하기 전에, 본 발명의 이해를 도울 다수의 정의가 제공된다. 본원에서 인용된 모든 참고문헌은 전문이 참고로 포함된다. 달리 정의되지 않는 한, 모든 과학 기술 용어는 본 발명이 속하는 분야의 통상의 기술자가 통상적으로 이해하는 바와 동일한 의미를 갖는다.
본원에서 사용된 경우에, 용어 "포함하는"은 임의의 언급된 요소가 필수적으로 포함되고, 다른 요소 또한 임의적으로 포함될 수 있다는 것을 의미한다. "~로 본질적으로 이루어지는"은 임의의 언급된 요소가 필수적으로 포함되고, 열거된 요소의 기본 특성 및 신규 특성에 실질적으로 영향을 미칠 요소는 배제되며, 다른 요소가 임의적으로 포함될 수 있다는 것을 의미한다. "~로 이루어지는"은 열거된 것들 이외의 모든 요소가 배제된다는 것을 의미한다. 각각의 이러한 용어에 의해 정의된 실시양태들이 본 발명의 범주 내에 속한다.
본원에서 사용된 경우의 용어 "핵산"은 각각의 뉴클레오티드의 3' 및 5' 끝부분이 포스포디에스테르 결합에 의해 연결된, 뉴클레오티드들의 공유결합으로 연결된 단일 또는 이중 가닥 서열이다. 폴리뉴클레오티드는 데옥시리보뉴클레오티드 염기 또는 리보뉴클레오티드 염기로 구성될 수 있다. 핵산은 DNA 및 RNA를 포함할 수 있고, 합성에 의해 시험관 내에서 제작될 수 있거나 또는 천연 공급원으로부터 단리될 수 있다. 핵산은 변형된 DNA 또는 RNA, 예를 들어, 메틸화된 DNA 또는 RNA, 또는 번역-후 변형, 예를 들어, 7-메틸구아노신의 5'-캡핑, 3'-프로세싱 예컨대 절단 및 폴리아데닐화, 및 스플라이싱에 적용된 RNA를 추가로 포함할 수 있다. 핵산은 합성 핵산 (XNA), 예컨대 헥시톨 핵산 (HNA), 시클로헥센 핵산 (CeNA), 트레오스 핵산 (TNA), 글리세롤 핵산 (GNA), 잠금 핵산 (LNA) 및 펩티드 핵산 (PNA)을 또한 포함할 수 있다. 본원에서 "폴리뉴클레오티드"로 또한 지칭되는 핵산의 크기는 전형적으로 이중 가닥 폴리뉴클레오티드에 대한 염기쌍 (bp)의 개수로, 또는 단일 가닥 폴리뉴클레오티드의 경우에는 뉴클레오티드 (nt)의 개수로 표현된다. 1000개의 bp 또는 nt는 킬로베이스 (kb)와 같다. 뉴클레오티드 약 40개 미만의 길이의 폴리뉴클레오티드는 전형적으로 "올리고뉴클레오티드"로 칭해지고, 폴리머라제 연쇄 반응 (PCR)을 통해서와 같은 DNA 조작에서 사용하기 위한 프라이머를 포함할 수 있다.
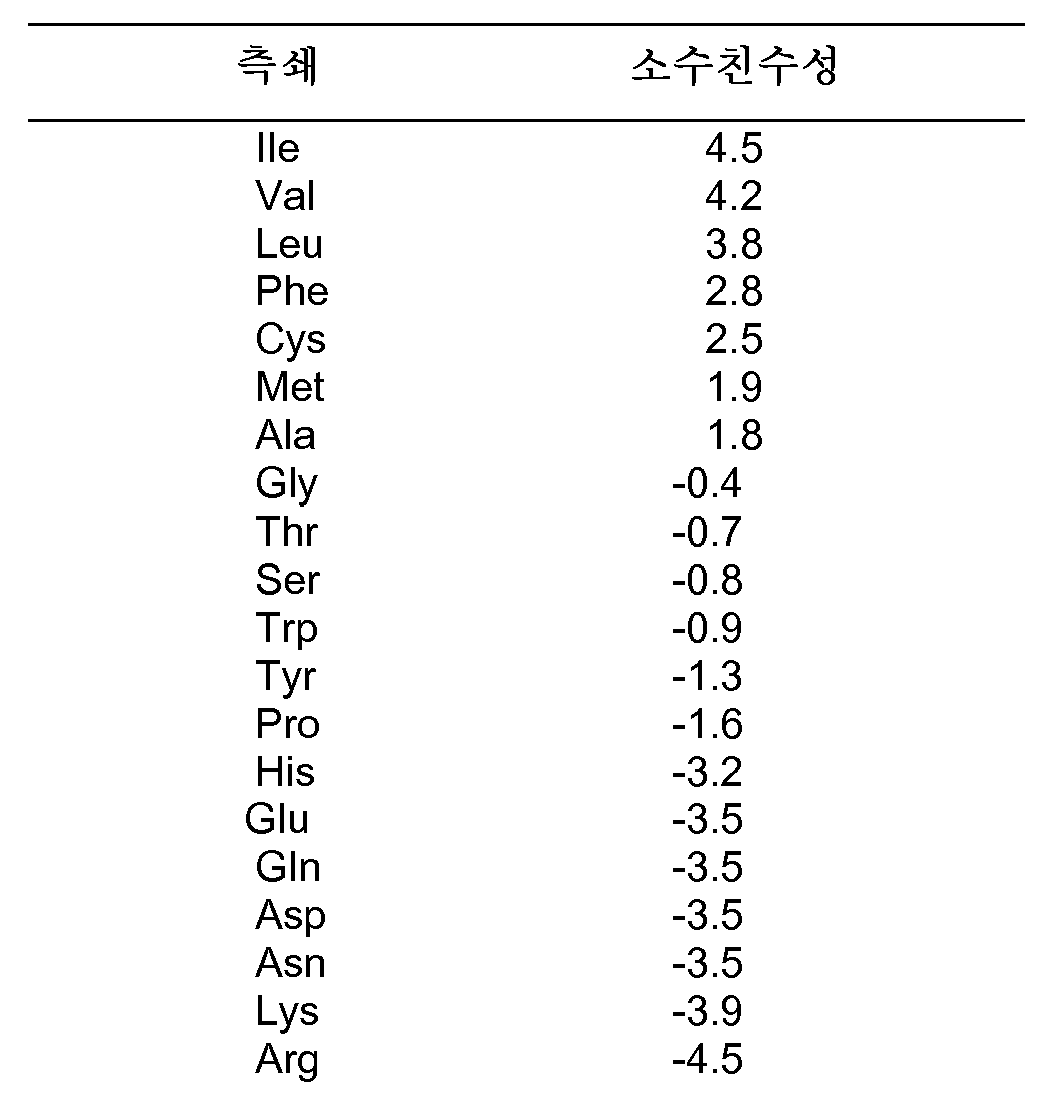
본 발명의 맥락에서의 용어 "아미노산"은 이의 가장 넓은 의미로 사용되고, 천연 발생 L α-아미노산 또는 잔기를 포함하도록 의도된다. 천연 발생 아미노산에 대한 통상적으로 사용되는 1-문자 및 3-문자 약어가 본원에서 사용된다: A=Ala; C=Cys; D=Asp; E=Glu; F=Phe; G=Gly; H=His; I=Ile; K=Lys; L=Leu; M=Met; N=Asn; P=Pro; Q=Gln; R=Arg; S=Ser; T=Thr; V=Val; W=Trp; 및 Y=Tyr (Lehninger, A. L., (1975) Biochemistry, 2d ed., pp. 71-92, Worth Publishers, New York). 일반적인 용어 "아미노산"은 D-아미노산, 레트로-인베르소(retro-inverso) 아미노산, 뿐만 아니라 화학적으로 변형된 아미노산 예컨대 아미노산 유사체, 일반적으로는 단백질 내로 혼입되지 않는 천연 발생 아미노산 예컨대 노르류신, 및 아미노산의 특성인 것으로 관련 기술 분야에 공지된 성질을 갖는 화학적으로 합성된 화합물, 예컨대 β-아미노산을 추가로 포함한다. 예를 들어, 천연 Phe 또는 Pro와 동일하게 펩티드 화합물의 형상적 제한을 허용하는, 페닐알라닌 또는 프롤린의 유사체 또는 모방체가 아미노산의 정의 내에 포함된다. 이같은 유사체 및 모방체는 본원에서 각각의 아미노산의 "기능성 등가물"로 지칭된다. 본원에 참고로 포함된 문헌 [Roberts and Vellaccio, The Peptides: Analysis, Synthesis, Biology, Gross and Meiehofer, eds., Vol. 5 p. 341, Academic Press, Inc., N.Y. 1983]에 아미노산의 기타 예가 열거되어 있다.
"폴리펩티드"는, 천연적으로 생산되었는지 또는 시험관 내에서 합성 수단에 의해 생산되었는지와 관계없이, 펩티드 결합에 의해 연결된 아미노산 잔기들의 중합체이다. 아미노산 잔기 약 12개 미만의 길이의 폴리펩티드는 전형적으로 "펩티드"로 지칭되고, 아미노산 잔기 약 12개 내지 약 30개 길이의 것은 "올리고펩티드"로 지칭될 수 있다. 본원에서 사용된 경우의 용어 "폴리펩티드"는 천연 발생 폴리펩티드의 생성물, 전구체 형태 또는 전구단백질을 나타낸다. 또한 폴리펩티드에 성숙 또는 번역-후 변형 프로세스가 진행될 수 있고, 이러한 프로세스는 글리코실화, 단백질분해성 절단, 지질화, 신호 펩티드 절단, 프로펩티드 절단, 인산화 등을 포함할 수 있지만 이에 제한되지 않는다. 본원에서 용어 "단백질"은 하나 이상의 폴리펩티드 쇄를 포함하는 거대분자를 지칭하도록 사용된다.
"생물학적 포어"는 막의 한쪽 측면에서 또 다른 측면으로의 분자 및 이온의 변위를 허용하는 채널 또는 구멍을 규정하는 막횡단 단백질 구조이다. 포어의 한쪽 측면에 인가된 전위 차에 의해 포어를 통한 이온성 종의 변위가 구동될 수 있다. "나노포어"는 분자 또는 이온이 통과하는 채널의 최소 직경이 대략적으로 나노미터 (10-9 미터)인 생물학적 포어이다.
본 발명의 모든 측면 및 실시양태에 대해, 폴리뉴클레오티드는 서열식별번호: 2에서 제시된 바와 같은 야생형 이. 콜라이 CsgG에 대한 적어도 50%, 60%, 70%, 80%, 90%, 95% 또는 99%의 완전한 서열 동일성을 갖는 폴리뉴클레오티드를 포함할 수 있다. 유사하게, 폴리펩티드는 서열식별번호: 1에서 제시된 바와 같은 야생형 이. 콜라이 CsgG에 대한 적어도 50%, 60%, 70%, 80%, 90%, 95% 또는 99%의 완전한 서열 동일성을 갖는 폴리펩티드를 포함할 수 있다. 폴리펩티드는 CsgG-유사 단백질의 특성인 PFAM 도메인 PF03783을 함유하는 폴리펩티드를 포함할 수 있다. 현재 공지된 CsgG 상동체 및 CsgG 구조의 목록을 http://pfam.xfam.Org//family/PF03783에서 확인할 수 있다. 따라서 서열 동일성은 또한 전장 폴리뉴클레오티드 또는 폴리펩티드의 단편 또는 일부분일 수 있다. 그러므로, 서열이 본 발명의 서열과의 단지 50%의 전체 서열 동일성을 가질 수 있지만, 특정 영역, 도메인 또는 서브유닛은 본 발명의 서열과 80%, 90%, 또는 많게는 99%의 서열 동일성을 공유할 수 있다. 본 발명에 따르면, 서열식별번호: 2의 핵산 서열에 대한 상동성은 단순히 서열 동일성에 제한되지 않는다. 다수의 핵산 서열이 외관상 낮은 서열 동일성을 가짐에도 불구하고 서로에 대해 생물학적으로 유의한 상동성을 실연할 수 있다. 본 발명에서, 상동성 핵산 서열은 낮은 엄격도의 조건 하에 서로 혼성화할 서열들인 것으로 간주된다 (M.R. Green, J. Sambrook, 2012, Molecular Cloning: A Laboratory Manual, Fourth Edition, Books 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY).
용어 "벡터"는 적합한 크기의 또 다른 핵산 (전형적으로는 DNA) 서열 단편이 내부로 통합될 수 있는, 선형 또는 원형인 DNA 분자를 나타내도록 사용된다. 이같은 DNA 단편(들)은 DNA 서열 단편에 의해 코딩되는 유전자의 전사를 제공하는 추가적인 분절을 포함할 수 있다. 추가적인 분절은 하기를 포함할 수 있지만, 이에 제한되지 않는다: 프로모터, 전사 종결인자, 인핸서, 내부 리보솜 진입 부위, 미번역 영역, 폴리아데닐화 신호, 선별성 마커, 복제 기원 등. 원핵생물 (예를 들어, 이. 콜라이에 대한 [베타]-락타마제 및 락토스 프로모터 시스템, 알칼리성 포스파타제, 트립토판 (trp) 프로모터 시스템, lac, tac, T3, T7 프로모터) 및 진핵생물 (예를 들어, 유인원 바이러스 40 초기 또는 후기 프로모터, 라우스 육종 바이러스 긴 말단 반복부 프로모터, 사이토메갈로바이러스 프로모터, 아데노바이러스 후기 프로모터, EG-1a 프로모터) 숙주에 대한 다양한 적절한 프로모터가 이용가능하다. 대개 발현 벡터는 플라스미드, 코스미드, 바이러스 벡터 및 효모 인공 염색체로부터 유래된다; 대개 벡터는 여러 공급원으로부터의 DNA 서열을 함유하는 재조합 분자이다. 본 발명의 구체적인 실시양태는 본원에 기술된 바와 같은 야생형 또는 변형된 CsgG 폴리펩티드를 코딩하는 발현 벡터를 제공한다. 예를 들어 상기 언급된 바와 같이 핵산 벡터 내에서, DNA 서열에 적용될 때의 용어 "작동가능하게 연결된"은, 이러한 서열들이 이들의 의도된 목적을 달성하기 위해 협동적으로 기능하도록 배열된다는 것을 가리키고, 즉 프로모터 서열이 전사 개시를 허용하여, 회합된 코딩 서열을 통해 멀게는 종결 서열까지 전사가 진행된다.
생물학적 포어의 막횡단 단백질 구조는 성질 면에서 단량체성 또는 올리고머성일 수 있다. 전형적으로, 포어는 중심축 둘레로 배열된 복수의 폴리펩티드 서브유닛을 포함함으로써, 나노포어가 있는 막에 대해 실질적으로 수직으로 뻗은, 단백질이 라이닝된 채널을 형성한다. 폴리펩티드 서브유닛의 개수는 제한되지 않는다. 전형적으로, 서브유닛의 개수는 5개 내지 30개이고, 적절하게는 서브유닛의 개수는 6개 내지 10개이다. 대안적으로, 퍼프린고리신(perfringolysin) 또는 관련된 대형 막 포어의 경우에서와 같이 서브유닛의 개수가 규정되지 않는다. 단백질이 라이닝된 채널을 형성하는 나노포어 내의 단백질 서브유닛의 일부분은 하나 이상의 막횡단 β-배럴, 및/또는 α-나선 섹션을 포함할 수 있는 2차 구조 모티프를 전형적으로 포함한다.
개시된 생성물 및 방법의 상이한 용도들이 관련 기술 분야에서의 구체적인 요구에 맞춰질 수 있다는 것을 이해하여야 한다. 본원에서 사용된 용어는 단지 본 발명의 특정 실시양태를 기술하는 것을 목적으로 하고, 제한적인 것으로 의도되지 않는다는 것을 또한 이해하여야 한다.
또한, 본 명세서 및 첨부된 특허청구범위에서 사용된 경우에, 단수 형태 ("a", "an", 및 "the")는 문맥적으로 명확하게 다르게 지시되지 않는 한 복수의 지시물을 포함한다. 따라서, 예를 들어, "폴리뉴클레오티드"를 언급하는 것은 2개 이상의 폴리뉴클레오티드를 포함하고, "폴리뉴클레오티드 결합 단백질"을 언급하는 것은 2개 이상의 이같은 단백질을 포함하며, "헬리카제"를 언급하는 것은 2개 이상의 헬리카제를 포함하고, "단량체"를 언급하는 것은 2개 이상의 단량체를 지칭하고, "포어"를 언급하는 것은 2개 이상의 포어를 포함하는 것 등이다.
상기 또는 하기에서 본원에서 인용된 모든 공보, 특허 및 특허 출원은 이에 의해 전문이 참고로 포함된다.
본 발명은, 부분적으로, 박테리아 아밀로이드 분비 채널 (CsgG), 이의 제작 방법, 및 핵산 시퀀싱 용도 및 분자 센싱에서의 이의 용도에 관한 것이다.
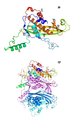
CsgG는 이. 콜라이의 외막 내에 존재하는 막 지질단백질이다 (유니프롯 등록 번호 P0AEA2; 유전자 ID: 12932538). 바깥쪽의 지질 막에서, CsgG는 9개의 CsgG 단량체 서브유닛의 올리고머 복합체를 포함하는 나노포어를 형성한다. II형 (지질단백질) 신호 서열에 의해, CsgG 전구단백질이 SEC 트랜스로콘(translocon)을 가로질러 변위되고, 이어서 성숙 CsgG (즉, II형 신호 서열이 절단된 CsgG)의 N-말단 Cys 잔기에서 트리아실화된다. 트리아실화 또는 "지질화"된 CsgG가 그람 음성 숙주의 외막으로 수송되고, 여기에서 9량체 포어로서 이중층 내로 삽입된다. 비-지질화 형태의 CsgG, 예를 들어 CsgGC1S는 프리-포어 형상 (도 42)의 가용성 단백질로서 원형질막 주위공간 내에 존재한다.
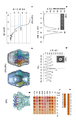
야생형 CsgG 나노포어의 X-선 구조 (Goyal et al., Nature, 2014, 516(7530), 250-3)는 이의 폭이 120 Å이고, 높이는 85 Å임을 나타낸다 (이하에서, 나노포어의 "폭"이라는 용어는 막 표면과 평행하는 이의 치수에 관련될 것이고, 나노포어의 "높이"라는 용어는 막에 대해 수직인 이의 치수에 관련될 것이다). CsgG 포어 복합체는 36-가닥의 β-배럴을 통해 막을 횡단하여, 내경이 40 Å인 채널을 제공한다 (도 1). CsgG 채널의 조립된 단량체는 보존된 12-잔기 루프 (C-루프, "CL"; 도 2)를 각각 소유하고, 이는 약 9.0 Å의 직경으로 채널 내의 협착부를 형성하도록 협력한다 (도 1 및 2). 야생형 CsgG 나노포어 내의 협착부는 CsgG 올리고머 내에 존재하는 각각의 CsgG 단량체의 아미노산 잔기 Tyr51, Asn55 및 Phe56의 측쇄에 의해 형성된 3개의 스태킹된 동심성 고리로 구성된다 (도 3). 이러한 잔기 번호매김은 N-말단의 아미노산 15개의 천연의 신호 서열이 결여된 성숙 단백질을 기초로 한다. 따라서, 성숙 단백질은 서열식별번호: 1의 잔기 16 내지 277에 상응한다. Tyr51은 서열식별번호: 1에서 위치 66에 있고, Asn은 서열식별번호: 1에서 위치 70에 있으며, Phe56은 서열식별번호: 1에서 위치 71에 있다.
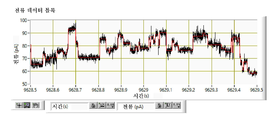
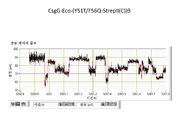
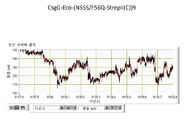
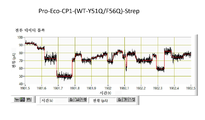
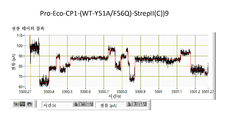
협착부는 CsgG 채널을 통한 이온 및 기타 분자의 통과를 제한하는 작용을 한다. 평면형 인지질 이중층에서 재구성된 CsgG의 단일-채널 전류 기록은 표준 전해질 조건 및 각각 +50 mV 또는 -50 mV의 전위를 사용하여 43.1 ± 4.5 pA (n = 33) 또는 -45.1 ± 4.0 pA (n = 13)의 정상(steady) 전류에 이르렀다 (도 5).
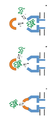
화학량론적 양의 원형질막 주위공간 인자 CsgE (유니프롯 등록 번호 POAE95)를 첨가함으로써 CsgG 채널을 통한 전류 흐름을 효과적으로 차단할 수 있다 (실시예 10 내지 12). 이론에 의해 제한되기를 원치 않으면서, 전류 증거는 CsgE가 CsgG 포어와 복합체를 형성하여 채널의 한쪽 끝부분을 캡핑하는 작용을 하는 메커니즘을 가리킨다. 표준 단일-채널 기록 기술을 사용하여 CsgG 채널을 통한 이온 흐름의 유의한 감소를 측정할 수 있다 (실시예 12 및 13, 도 6). 본 발명가들은 전류 흐름에 대한 측정된 파라미터 (최대 전류, 및 전류 변동을 모니터링하는 능력)가 나노포어를 본 발명의 한 실시양태에 따른 핵산 시퀀싱 및 분자 센싱 용도에서 사용하기에 적절하게 한다는 것을 발견하였다.
따라서, 부분적으로 본 발명은 나노포어를 통한 전류 흐름의 전기적 측정의 변동을 기초로 하는 핵산 시퀀싱에서의 CsgG 나노포어 단백질 복합체의 방법 및 용도에 관한 것이다.
핵산이 나노포어 시퀀싱에 특히 적절하다. DNA 및 RNA 내의 천연-발생 핵산 염기가 이의 물리적 크기에 의해 구별될 수 있다. 핵산 분자 또는 개별적인 염기가 나노포어의 채널을 통과할 때, 염기들 사이의 차별적인 크기가 채널을 통한 이온 흐름에서 직접적으로 상관되는 감소를 야기한다. 이온 흐름의 변동을 기록할 수 있다. 이온 흐름 변동을 기록하기 위한 적절한 전기적 측정 기술이, 예를 들어, WO 2000/28312 및 문헌 [D. Stoddart et al., Proc. Natl. Acad. Sci., 2010, 106, pp 7702-7] (단일 채널 기록 기기); 및, 예를 들어, WO 2009/077734 (다채널 기록 기술)에 기술되어 있다. 적절한 보정을 통해, 이온 흐름의 특징적인 감소를 사용하여, 채널을 횡단하는 특정 뉴클레오티드 및 연관된 염기를 실시간으로 확인할 수 있다.
전형적으로, 막횡단 채널 내의 가장 좁은 협착부의 크기가 핵산 시퀀싱 용도에 대한 나노포어의 적합성을 결정하는 것에서 핵심 요소이다. 협착부가 너무 작으면, 시퀀싱될 분자가 통과할 수 없을 것이다. 그러나, 채널을 통한 이온 흐름에 대한 최대 효과를 달성하기 위해, 이의 가장 좁은 지점에서 (즉, 협착부에서), 채널이 너무 크기 않아야 한다. 이상적으로, 임의의 협착부는 직경 면에서 통과하는 염기의 크기에 가능한 한 가까워야 한다. 핵산 및 핵산 염기의 시퀀싱을 위해, 적절한 협착부 직경은 나노미터 범위 (10-9 미터 범위)이다. 적절하게는, 직경이 0.5 내지 1.5 nm의 영역 내에 있어야 하고, 전형적으로는 직경이 0.7 내지 1.2 nm의 영역 내에 있다. 야생형 이. 콜라이 CsgG 내의 협착부는 직경이 약 9 Å (0.9 nm)이다. 본 발명가들은 CsgG 채널 내의 협착부의 크기 및 형상이 핵산 시퀀싱에 적합하다고 추정하였다.
핵산 시퀀싱과 관련된 용도를 위해, CsgG 나노포어는 야생형 형태로 사용될 수 있거나, 또는, 예컨대 특정 아미노산 잔기의 지정 돌연변이유발에 의해, 사용 시 나노포어의 원하는 성질을 추가로 강화하도록 추가로 변형될 수 있다. 예를 들어, 본 발명의 실시양태에서, 채널 내의 협착부의 개수, 크기, 형태, 배치 또는 배향을 변경시키도록 돌연변이가 구상된다. 폴리펩티드 서열 내의 특정한 표적화된 아미노산 잔기의 삽입, 치환 및/또는 결실을 초래하는 공지된 유전자 조작 기술에 의해, 돌연변이된 돌연변이체 CsgG 나노포어 복합체를 제조할 수 있다. 올리고머 CsgG 나노포어의 경우, 각각의 단량체 폴리펩티드 서브유닛, 또는 단량체 중 임의의 하나, 또는 모든 단량체에서 돌연변이가 이루어질 수 있다. 적절하게는, 본 발명의 한 실시양태에서, 기술된 돌연변이가 올리고머 단백질 구조 내의 모든 단량체 폴리펩티드에 대해 이루어진다.
본 발명의 실시양태에 따르면, 포어 내의 채널 협착부의 개수가 감소된 변형된 돌연변이체 CsgG 나노포어가 제공된다.
야생형 이. 콜라이 CsgG 포어는 2개의 채널 협착부를 포함한다 (도 1 참조). 이들은 C-루프 모티프뿐만 아니라 위치 54 내지 53으로부터의 추가적인 아미노산을 포함하는 더 넓은 구조의 일부분으로서, (i) 아미노산 잔기 Phe56 및 Asn55, 및 (ii) 아미노산 잔기 Tyr 51에 의해 형성된다 (도 2 및 3).
전형적인 나노포어 핵산 시퀀싱에서, 관심 핵산 서열의 개별적인 뉴클레오티드들이 순차적으로 나노포어의 채널을 통과할 때 뉴클레오티드에 의한 채널의 부분적인 차단으로 인해 개방-채널 이온 흐름이 감소된다. 상기 기술된 적절한 기록 기술을 사용하여 이온 흐름의 이러한 감소를 측정한다. 이러한 이온 흐름 감소를 채널을 통한 공지된 뉴클레오티드에 대한 측정된 이온 흐름의 감소에 대해 보정하여, 어떤 뉴클레오티드가 채널을 통과하는지를 결정하는 수단이 초래될 수 있고, 따라서, 순차적으로 행하는 경우, 나노포어를 통과하는 핵산의 뉴클레오티드 서열을 결정하는 방식이 초래될 수 있다. 개별적인 뉴클레오티드의 정확한 결정을 위해, 채널을 통한 이온 흐름의 감소를 단일 협착부 (또는 "판독 헤드")를 통과하는 개별적인 뉴클레오티드의 크기와 직접적으로 상관시키는 것이 전형적으로 요구되었다. 예를 들어, 회합된 폴리머라제의 작용을 통해 포어를 '관통'하는 무손상 핵산 중합체에 대해 시퀀싱이 수행될 수 있다는 것이 이해될 것이다. 대안적으로, 포어 가까이에서 표적 핵산으로부터 순차적으로 제거된 뉴클레오티드 트리포스페이트 염기의 통과에 의해 서열을 결정할 수 있다 (예를 들어 WO 2014/187924 참조).
2개 이상의 협착부가 존재하고 떨어져 놓인 경우, 각각의 협착부가 동시에 핵산 가닥 내의 별개의 뉴클레오티드들과 상호작용하거나 또는 이들을 "판독"할 수 있다. 이러한 상황에서, 채널을 통한 이온 흐름의 감소는 뉴클레오티드를 함유하는 모든 협착부의 조합된 흐름 제한의 결과일 것이다. 따라서, 일부 경우에, 이중 협착부는 복합적인 전류 신호에 이를 수 있다. 특정 상황에서, 2개의 이같은 판독 헤드가 존재할 때 하나의 협착부 또는 "판독 헤드"에 대한 전류 판독치를 개별적으로 결정할 수 없을 수 있다.
재조합 유전자 기술을 통해 CsgG의 야생형 포어 구조를 재조작하여, 채널 내에 단일 협착부가 남도록 2개의 협착부 중 하나를 넓히거나, 변경시키거나 또는 제거하고, 따라서 단일 판독 헤드를 규정할 수 있다. CsgG 올리고머 포어 내의 협착부 모티프는 야생형 단량체 이. 콜라이 CsgG 폴리펩티드 내의 위치 38 내지 63의 아미노산 잔기에 위치한다. 이러한 영역의 야생형 아미노산 서열이 서열식별번호: 3으로 제공된다. 이러한 영역을 고려하여, 아미노산 잔기 위치 50 내지 53, 54 내지 56 및 58 내지 59 중 임의의 것에서의 돌연변이가 본 발명의 검토 사항 내에서 구상된다. CsgG 상동체와의 서열 유사성을 기초로 (도 4), 아미노산 잔기 위치 38 내지 49, 53, 57, 및 61 내지 63은 고도로 보존적인 것으로 간주되고, 따라서 치환 또는 기타 변형에 덜 적절할 수 있다. 야생형 CsgG 구조의 채널 내의 Tyr51, Asn55, 및 Phe56의 측쇄의 핵심 위치화로 인해, 판독 헤드의 특성을 변형 또는 변경시키기 위해 이러한 위치에서의 돌연변이가 유리할 수 있다.
단량체 CsgG 단백질의 소정의 위치에서의 돌연변이는 이러한 위치의 야생형 아미노산이 임의의 다른 천연 또는 비천연 아미노산으로 치환되는 것을 초래할 수 있다. 본 발명의 한 실시양태에서, 협착부를 넓히거나 제거하는 것이 바람직하다; 적절하게는, 변형된 CsgG 단백질 내의 아미노산 측쇄는 자신이 교체하는 야생형 구조 내의 아미노산 측쇄보다 입체적으로 덜 방해하도록 선택될 것이다. 소정의 위치의 아미노산 잔기의 교체는 유사한 정전기 성질을 가질 수 있거나, 또는 상이한 정전기 성질을 가질 수 있다. 적절하게는, 채널의 성질 또는 2차 구조의 파괴를 최소화하기 위해 교체 아미노산 측쇄는 자신이 교체하는 야생형 구조 내의 아미노산 측쇄와 유사한 정전기 전하를 소유할 것이다.
교체 아미노산의 선택은 대형 다중 서열 정렬을 기초로 아미노산이 또 다른 아미노산으로 치환될 가능성을 계산하기 위한 표준 방법을 제공하는 BLOSUM62 매트릭스를 기초로 할 수 있다. 통상의 기술자는 인터넷에서 BLOSUM62 매트릭스의 예를 자유롭게 입수할 수 있다; 예를 들어 국립 생물공학 정보 센터(National Center for Biotechnology Information)의 웹사이트 (http://www.ncbi.nlm.nih.gov/Class/Structure/aa/aa_explorer.cgi)를 참조한다.
CsgG 채널에서 제1 협착부를 형성하는 작용을 하는, 야생형 CsgG 구조 내의 Tyr51에 대해, 임의의 아미노산으로의 치환이 제공된다. 특히, 본 발명의 특정 실시양태에서, Tyr51이 알라닌, 글리신, 발린, 류신, 이소류신, 아스파라긴, 글루타민, 및 페닐알라닌으로 치환될 수 있다 (서열식별번호: 39-318). 본 발명의 실시양태에서, Tyr51이 알라닌 또는 글리신으로 치환되는 것이 특히 적절하다 (서열식별번호: 39-108). 실시양태에서, 잔기 50 내지 53 (야생형 서열에서의 PYPA)이 글리신-글리신 (GG)으로 교체될 수 있다 (서열식별번호: 354-388).
CsgG 채널 내의 제2 협착부에 기여하는 Asn55에 대해, 임의의 아미노산으로의 치환이 제공된다. 특히, 본 발명의 특성 실시양태에서, Asn55이 알라닌, 글리신, 발린, 세린 또는 트레오닌으로 치환될 수 있다 (서열식별번호: 9-33, 44-68, 79-103 114-138, 149-173, 184-208, 219-243, 254-278, 289-313 및 324-348).
CsgG 채널 내의 제2 협착부의 일부분을 형성하는 Phe56에 대해, 임의의 아미노산으로의 치환이 제공된다. 특히, 본 발명의 특성 실시양태에서, Phe56이 알라닌, 글리신, 발린, 류신, 이소류신, 아스파라긴, 및 글루타민으로 치환될 수 있다 (서열식별번호: 5-13, 15-18, 20-23, 25-28, 30-33, 40-43, 45-48, 50-53, 55-58, 60-63, 65-68, 75-78, 80-83, 85-88, 90-93, 95-98, 100-103, 110-113, 115-118, 120-123, 125-128, 130-133, 135-138, 145-148, 150-153, 155-158, 160-163, 165-168, 170-173, 180-183, 185-188, 190-193, 195-198, 200-203, 205-208, 215-218, 220-223, 225-228, 230-233, 235-238, 240-243, 250-253, 255-258, 260-263, 265-268, 270-273, 275-578, 285-288, 290-293, 295-298, 300-303, 305-308, 310-313, 320-323, 325-328, 330-333, 335-338, 340-343 및 345-348). 본 발명의 실시양태에서, Phe56이 알라닌 및 글리신으로 치환되는 것이 특히 적절하다 (서열식별번호: 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145, 150, 155, 160, 165, 170, 175, 180, 185, 190, 195, 200, 205, 210, 215, 220, 225, 230, 235, 240, 245, 250, 255, 260, 265, 270, 275, 280, 285, 290, 295, 300, 305, 310, 315, 320, 325, 330, 335, 340, 345, 350, 355, 360, 365, 370, 375, 380, 385, 6, 11, 16, 21, 26, 31, 36, 41, 46, 51, 56, 61, 66, 71, 76, 81, 86, 91, 96, 101, 106, 111, 116, 121, 126, 131, 136, 141, 146, 151, 156, 161, 166, 171, 176, 181, 186, 191, 196, 201, 206, 211, 216, 221, 226, 231, 236, 241, 246, 251, 256, 261, 266, 271, 276, 281, 286, 291, 296, 301, 306, 311, 316, 321, 326, 331, 336, 341 346, 351, 356, 361, 366, 371, 376, 381 및 386).
소정의 돌연변이체 CsgG 단백질에서, Tyr51의 치환이 위치 55 및 56 중 하나에서와 동시에 수행될 수 있고 (서열식별번호: 44, 49, 54, 59, 64, 79, 84, 89, 94, 99, 114, 119, 124, 129, 134, 149, 154, 159, 164, 169, 184, 189, 194, 199, 204, 219, 224, 229, 234, 239, 254, 259, 264, 269, 274, 289, 294, 299, 304, 309, 359, 364, 369, 374, 379, 40, 41, 42, 75, 76, 77, 110, 111, 112, 145, 146, 147, 180, 181, 182, 215, 216, 217, 250, 251, 252, 285, 286 및 287), 여기서 채널 내의 적절한 치수의 적어도 1개의 협착부가 유지된다. 대안적으로는, Tyr51의 치환이 위치 Asn55 및 Phe56 양쪽 모두에서의 치환에 대해 서로 배타적이다 (서열식별번호: 39, 74, 109, 144, 179, 214, 249, 284, 354, 10, 11, 12, 15, 16, 17, 20, 21, 22, 25, 26, 27, 30, 31 및 32).
대안적으로, 야생형 CsgG 단백질 내의 Tyr51, Asn55 또는 Phe56 중 하나 이상이 결실될 수 있다 (서열식별번호: 319-353, 34-38, 69-73, 104-108, 139-143, 174-178, 209-213, 244-248, 279-283, 314-318, 384-388, 8, 13, 18, 23, 28, 33, 43, 48, 53, 58, 63, 68, 78, 83, 88, 93, 98, 103, 113, 118, 123, 128, 133, 138, 148, 153, 158, 163, 168, 173, 183, 188, 193, 198, 203, 208, 218, 223, 228, 233, 238, 243, 253, 258, 263, 268, 273, 278, 288, 293, 298, 303, 308 및 313). 채널 내의 적어도 1개의 협착부를 유지하기 위해, 소정의 실시양태에서, 아미노산 잔기 Tyr51의 결실이 아미노산 잔기 Asn55 및 Phe56 양쪽 모두에서의 결실에 대해 서로 배타적이다 (서열식별번호: 319-322, 324-327, 329-332, 334-337, 339-342, 344-347, 38, 73, 108, 143, 178, 213, 248, 283 및 318). 위치 53 및 54 및 48 및 49의 특정 이웃 아미노산 잔기가 또한 결실될 수 있다.
본 발명이 상기 변형들이 별개로 또는 임의의 조합으로 이루어질 수 있는 실시양태를 제공한다는 것을 이해하여야 한다.
Tyr51에서의 협착부 또는 Asn55/Tyr56에서의 협착부 중 하나의 제거는 CsgG 채널 내의 단일 협착부를 초래한다. 이론에 의해 제한되기를 원치 않으면서, 바람직할 수 있는 Tyr51에서의 협착부보다 Asn55/Tyr56에서의 협착부가 형상 안정성이 더 높을 것으로 가정된다. 그러나, Asn55/Tyr56 협착부는 뉴클레오티드에 비교하여 너무 높을 수 있다 (중심 포어 축을 따라 측정했을 때). 이는 변위되는 DNA 가닥 내의 개별적인 염기쌍의 불량한 해상도에 이를 수 있다.
그 반대가 Tyr51 협착부에 대해 사실일 것이다. Asn55/Tyr56 협착부의 제거 후, 올리고머 내의 잔여 Tyr51 잔기 고리는 천연 구조에서보다 형상적으로 덜 안정적일 수 있다. 그러나, Tyr51 협착부는 더 짧고 (중심 포어 축을 따라 측정했을 때), 개별적인 염기들을 구별할 수 있는 채널 내의 협착부를 제공할 가능성이 더 많다.
어느 한쪽의 실시양태에서, 단일한 좁은 협착부 (중심 포어 축을 따라 측정했을 때)가 존재하는 것은 포어가 핵산 시퀀싱 용도에 이용될 때 전류 판독의 복잡성을 감소시킬 것이다. 따라서, 포어를 통한 핵산 변위 동안 발생하는 관찰된 전류에서의 조정이 단일 협착부 또는 "판독 헤드"를 통한 별개의 뉴클레오티드들의 통과를 단독으로 반영할 것이다.
1개의 협착부의 효과적인 제거는 또한 포어 변이체의 개방-채널 전류를 증가시킬 수 있다. 더 높은 배경 전도도는 상이한 핵산 염기 쌍 신호들에 대한 더 양호하게 해상된 전류 차단 수준에 이르기 때문에, 증가된 개방-채널 전류가 유리할 것이다. 이러한 방식으로, 판독 헤드에 대한 변형이 핵산 시퀀싱 및 기타 분자 센싱 용도 양쪽 모두에 대한 생물학적 포어의 적절성을 개선시킬 수 있다.
대안적인 실시양태로서, 또는 상기 기술된 서열 변형에 더하여, Asn55/Phe56 협착부가 이의 높이 (중심 포어 축을 따라 측정했을 때)를 조율하도록 추가로 개조될 수 있는 것이 또한 제공된다. Asn55/Phe56 협착부의 이같은 추가 개조에 CsgG 채널 내의 Tyr51 또는 다른 위치의 돌연변이가 동반될 수 있거나 또는 그렇지 않을 수 있다. 적절하게는, Tyr51 잔기에 의해 형성된 협착부를 넓히거나 제거하는 돌연변이의 일부분으로서 Asn55/Phe56 협착부의 추가 개조가 구상된다.
야생형 형태에서, Asn55/Phe56 채널 협착부는 서로에 대해 수직으로 놓인 2개의 아미노산 고리로 구성된다. 그 결과, 협착부의 길이가 1 nm를 초과한다. 1 nm 길이의 협착부는 변위되는 핵산 가닥 내의 별개의 염기들에 대한 이온 흐름으로부터 생성된 전기적 신호의 해상을 허용하지 않을 수 있다. 전형적으로, 핵산 시퀀싱에 사용되는 공지된 나노포어의 협착부(들)는 전형적으로 길이가 1 nm 미만이다. 예를 들어, DNA 시퀀싱에 사용되는 MspA 나노포어는 협착부 높이가 0.6 nm이다 (중심 포어 축을 따라 측정했을 때; 문헌 [Manrao et al., Nature Biotechnology, 2012, 30(4), 349-353]).
Asn55/Phe56 협착부의 높이 (중심 포어 축을 따라 측정했을 때)를 감소시키기 위해, 2개의 잔기 중 하나가 치환 또는 결실되어 협착부의 상부 또는 하부가 넓어지는 것에 이를 수 있다.
CsgG 채널 내의 제2 협착부의 일부분을 형성하는 Asn55에 대해, 임의의 아미노산으로의 치환이 구상된다. 특히, 알라닌, 글리신, 발린, 세린 또는 트레오닌으로의 치환 (서열식별번호: 9-33, 44-68, 79-103 114-138, 149-173, 184-208, 219-243, 254-278, 289-313 및 324-348).
CsgG 채널 내의 제2 협착부의 일부분을 형성하는 Phe 56에 대해, 임의의 아미노산으로의 치환이 구상된다. 특히, 알라닌, 글리신, 발린, 류신, 이소류신, 아스파라긴, 및 글루타민으로의 치환 (서열식별번호: 5-13, 15-18, 20-23, 25-28, 30-33, 40-43, 45-48, 50-53, 55-58, 60-63, 65-68, 75-78, 80-83, 85-88, 90-93, 95-98, 100-103, 110-113, 115-118, 120-123, 125-128, 130-133, 135-138, 145-148, 150-153, 155-158, 160-163, 165-168, 170-173, 180-183, 185-188, 190-193, 195-198, 200-203, 205-208, 215-218, 220-223, 225-228, 230-233, 235-238, 240-243, 250-253, 255-258, 260-263, 265-268, 270-273, 275-578, 285-288, 290-293, 295-298, 300-303, 305-308, 310-313, 320-323, 325-328, 330-333, 335-338, 340-343 및 345-348). 본 발명의 실시양태에서, Phe56이 알라닌 및 글리신으로 치환되는 것이 특히 적절하다 (서열식별번호: 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145, 150, 155, 160, 165, 170, 175, 180, 185, 190, 195, 200, 205, 210, 215, 220, 225, 230, 235, 240, 245, 250, 255, 260, 265, 270, 275, 280, 285, 290, 295, 300, 305, 310, 315, 320, 325, 330, 335, 340, 345, 350, 355, 360, 365, 370, 375, 380, 385, 6, 11, 16, 21, 26, 31, 36, 41, 46, 51, 56, 61, 66, 71, 76, 81, 86, 91, 96, 101, 106, 111, 116, 121, 126, 131, 136, 141, 146, 151, 156, 161, 166, 171, 176, 181, 186, 191, 196, 201, 206, 211, 216, 221, 226, 231, 236, 241, 246, 251, 256, 261, 266, 271, 276, 281, 286, 291, 296, 301, 306, 311, 316, 321, 326, 331, 336, 341 346, 351, 356, 361, 366, 371, 376, 381 및 386).
야생형 CsgG 포어의 협착부의 최소 직경을 조율하는 변형이 또한 구상된다. CsgG 포어 내의 양쪽 협착부의 최소 직경은 약 0.9 nm (9 Å)이고, 이는 DNA 시퀀싱에 대한 유용성을 나타내는 공지된 MspA 나노포어 (Manrao et al., Nature Biotechnology, 2012, 30(4), 349-353) 내의 협착부에 대한 1.2 nm보다 더 작다. 최소 직경이 0.5 내지 1.5 nm인 변형된 CsgG 포어 내의 잔여 협착부를 제공하는 상기 돌연변이 중 임의의 것이 적절할 것이다.
전류 판독을 개선하기 위해 변위되는 핵산 가닥의 통과 및 비-공유결합 상호작용을 개선하도록 상기 열거된 변형 중 임의의 것이 협착부에서의 아미노산의 친수성 및 전하 분포를 이롭게 변경시킬 수 있다. 또한, 협착부를 통한 전해질 이온의 흐름을 최적화하고, 변위되는 핵산 가닥의 통과되는 뉴클레오티드들 사이의 더 양호한 판별을 달성하기 위해 상기 열거된 돌연변이 중 임의의 것이 채널 협착부에 가까운 친수성 및 전하 분포를 이롭게 변경시킬 수 있다.
채널 내강 내의 표면 전하 분포를 변화시키는 것을 초래할 수 있는 CsgG 단백질의 추가 변형이 구상된다. 본 발명의 한 실시양태에서, 채널 벽에 대한 변위되는 핵산의 원치 않는 정전기 흡착을 피하도록 이러한 변형이 이루어질 수 있다. 핵산은 음으로 하전되고, CsgG 채널 내강은 약간의 양성 전하를 함유하기 때문에 (도 1), 정전기 상호작용이 핵산 시퀀싱 동안 관통 또는 변위를 방해할 수 있는 것으로 가정된다. 적절하게는, 포어를 통한 핵산 변위의 효율, 따라서 전류 판독의 명확성을 추가로 개선하기 위해 양으로 하전된 아미노산 잔기 예컨대 리신, 히스티딘 및 아르기닌이 중성이거나 음으로 하전된 측쇄로 치환될 수 있다.
포어 협착부 내로의 핵산 가닥 (또는 개별적인 뉴클레오티드들)의 변위 또는 관통을 용이하게 하는 야생형 이. 콜라이 CsgG 또는 돌연변이체 CsgG의 채널 내강의 변형이 본 발명의 실시양태에 의해 또한 제공된다. 내부 협착부가 있는 CsgG 의 막-스패닝(spanning) 섹션은 중앙의 구멍을 특색으로 하는 뚜껑이 있는 배럴과 유사하다. 배럴 및 뚜껑에 가장 가까운 포어 내강 내로 추가적인 루프를 부가함으로써 관통이 용이해질 수 있다.
본 발명의 구체적 실시양태는 CsgG 포어가 공유결합으로 부착된 하나 이상의 단량체, 2량체 또는 올리고머로 구성될 수 있는 것을 제공한다. 비제한적인 예로서, 단량체들이 임의의 형상으로, 예컨대 이들의 말단 아미노산에 의해, 유전적으로 융합될 수 있다. 이러한 경우에, 1개의 단량체의 아미노 말단이 또 다른 단량체의 카르복시 말단에 융합될 수 있다.
본 발명의 실시양태에 따르면, 포어를 통한 분자의 통과에 대한 이로운 성질이 있을 수 있는 추가적인 보조 단백질을 수용하도록 CsgG 포어가 개조될 수 있는 것이 또한 제공된다. 포어에 대한 개조는 핵산-프로세싱 효소의 앵커링(anchoring)을 용이하게 할 수 있다. 핵산-프로세싱 효소는 DNA 또는 RNA 폴리머라제; 이소머라제; 토포이소머라제; 자이라제; 텔로머라제; 및 헬리카제를 포함할 수 있다. 이러한 효소 중 하나 이상이 나노포어와 회합되는 것은 포어 내로의 핵산 관통의 강화의 관점에서, 그리고 핵산 가닥이 포어를 통해 전이되는 속도를 제어하는 것에서 이점이 있을 수 있다 (Manrao et al., Nature Biotechnology, 2012, 30(4), 349-353). 포어를 통한 핵산 가닥의 변위 속도를 제어하는 것은 판독에 더욱 적절하고 더욱 균일한 이온 흐름의 전류 측정의 관점에서 반응 개선을 제공하는 장점이 있다.
본 발명의 실시양태에서, 나노포어의 막외 영역에서의 변형이 하나 이상의 결합/앵커링 부위의 제공을 통해 채널의 내강 내부에 또는 내강에 인접하여 적절한 핵산-프로세싱 효소, 예컨대 DNA-폴리머라제가 도킹되는 것을 용이하게 하는 것을 도울 수 있는 것으로 생각된다. 적절한 앵커링 부위는 효소의 정전기적 결합을 위한 정전기적 패치(patch); 공유결합 커플링을 허용하는 하나 이상의 시스테인 잔기; 및/또는 입체적 앵커를 제공하는 막횡단 채널 섹션의 변경된 내부 폭을 포함할 수 있다.
하기에서 더욱 상세하게 기재되는 구체적 실시양태에서 본 발명은 핵산 시퀀싱에서 사용하기 위한 CsgG 야생형 포어의 추가 개조를 제공한다.
본 발명의 실시양태에서, CsgG 포어의 막외 영역 (도 1에서 제시된 바와 같은 하부 부분)이 절단되거나 또는 제거되어, 채널 내강의 다른쪽 측면 상에서의 핵산 가닥의 퇴장을 용이하게 할 수 있다. 막외 영역의 절단 또는 제거는 또한 채널에서의 이온 흐름으로부터의 전기적 신호의 전류 해상도를 개선시킬 수 있다. 후자의 이점은 막외 영역에 의해 이온 흐름에 야기되는 저항성을 저하시킴으로써 초래된다. 본 발명의 CsgG 포어에서, 막횡단 채널, 내부 협착부, 및 캡 영역이 3개의 저항성 영역을 차례로 나타낸다. 이들 중 하나의 기여를 제거하거나 저하시키는 것은 개방-채널 전류를 증가시킬 것이고, 따라서 전류 해상도를 개선시킬 것이다. 이같은 변경은 α-나선 2 (α2), C-말단 a-나선 (αC) (도 1), 및/또는 이들의 조합의 결실을 포함할 수 있다.
본 발명의 추가 실시양태에서, 야생형 이. 콜라이 CsgG 포어의 바깥쪽 면 상의 막을 향한 아미노산이 변형되어, 막 내로의 포어의 삽입을 용이하게 할 수 있다. 특정 실시양태에서, 단일 아미노산 치환이 야생형 잔기를 적절한 더욱 소수성인 유사체로 교체할 수 있다. 예를 들어, 잔기 Ser136, Gly138, Gly140, Ala148, Ala 188 또는 Gly202 중 하나 이상이 Ala, Val, Leu, 또는 Ile로 변화될 수 있다. 추가적으로, 방향족 잔기 예컨대 타이로신 또는 트립토판이 소수성 막과 소수성 용매 사이의 계면에 위치하도록 적합한 야생형 아미노산을 치환할 수 있다. 예를 들어, 잔기 Leu 154 또는 Leu182 중 하나 이상이 Tyr, Phe 또는 Trp으로 치환될 수 있다.
본 발명의 실시양태에서, 단백질 포어의 열 안정성이 증가될 수 있다. 이는 시퀀싱 장치에서의 나노포어의 저장 기간의 유리한 증가를 초래한다. 실시양태에서, 베타-회전 서열의 변형 또는 단백질 표면에서의 정전기적 상호작용을 개선시키는 것에 의해 단백질 포어의 열 안정성의 증가가 달성된다. 한 실시양태에서, 인접한 β-헤어핀들 내부 또는 사이의 2개의 인접한 β-가닥을 교차하는 공유결합 디술피드 형성에 의해 막횡단 영역 내의 β-헤어핀이 안정화될 수 있다. 이같은 가닥-교차 시스테인 쌍의 예는 Val139-Asp203; Gly139-Gly205; Lys135-Thr207; Glu201-Ala141; Gly147-Gly189; Asp149-Gln187; Gln151-Glu185; Thr207-Glu185; Gly205-Gln187; Asp203-Gly189; Ala153-Lys135; Gly137-Gln151; Val139-Asp149 또는 Ala141-Gly147일 수 있다.
본 발명의 추가 실시양태에서, 문헌 [Biotechnol. J., 2011, 6(6), 650-659]에 기술된 방법에 따라 CsgG 단백질의 발현을 높은 수준 및 낮은 오차율로 허용하도록 폴리펩티드 서열의 코돈 용법이 변형될 수 있다. 이러한 변형은 mRNA의 임의의 2차 구조를 표적화할 수도 있다.
본 발명은 단백질 서열을 이의 프로테아제 안정성을 개선시키기 위해 변경하는 것을 또한 제공한다. 이는 가요성 루프 영역의 제거, 예를 들어 α-나선 2 (α2), C-말단 a-나선 (αC) (도 1), 및/또는 이들의 조합의 결실에 의해 달성될 수 있다.
본 발명의 실시양태에서, 가능한 단백질 응집을 피하도록 CsgG 폴리펩티드 서열/발현 시스템이 변경된다.
본 발명은 단백질의 폴딩 효율을 개선시키도록 폴리펩티드 서열을 변경시키는 것을 또한 제공한다. 적절한 기술이 문헌 [Biotechnol. J., 2011, 6(6), 650-659]에서 제공된다.
본 발명의 한 실시양태는 CsgG 단백질의 정제를 용이하게 하도록 생체친화력 태그(tag)를 교체 또는 부가하는 것을 추가로 제공한다. CsgG 포어의 공개된 구조는 StrepII 태그를 함유한다 (Goyal et al., Nature, 2014, 516(7530), 250-3). 본 발명의 실시양태는 다른 생체친화력 태그, 예컨대 금속 킬레이트 친화력 크로마토그래피를 통한 정제를 용이하게 하기 위한 히스티딘-태그를 포함한다. 본 발명의 대안적인 실시양태에서, 태그는 FLAG-태그 또는 에피토프 태그, 예컨대 Myc- 또는 HA-태그를 포함할 수 있다. 본 발명의 추가 실시양태에서, 나노포어가 비오틴 또는 이의 유사체 (예를 들어 데스티오비오틴)으로의 비오틴화에 의해 변형됨으로써, 스트렙타비딘과의 상호작용을 통한 정제를 용이하게 할 수 있다.
본 발명의 실시양태에서, 폴리펩티드의 순전하를 증가시키고 폴리아크릴아미드 겔 전기영동에서의 단백질 이동을 용이하게 하기 위해 단백질 말단에서의 음전하가 부가될 수 있다. 이는 CsgG의 이종올리고머가 관심 대상인 경우에 이러한 종의 분리 개선에 이를 수 있다 (문헌 [Howorka et al. Proc. Nat. Acad. Sci., 2001, 98(23), 12996-13001] 참조). 이종올리고머는 포어 당 1개의 시스테인 잔기를 도입하는데 유용할 수 있고, 이는 하기 기술된 바와 같은 적절한 핵산-프로세싱 효소, 예컨대 DNA-폴리머라제의 부착을 용이하게 하는데 유리할 수 있다.
본 발명은, 부분적으로, 나노포어를 통한 전류 흐름의 전기적 측정의 변동을 기초로 하는 분자 센싱에서의 야생형 또는 변형된 CsgG 나노포어의 용도에 관한 것이다.
CsgG 포어의 채널 내에 또는 채널의 한쪽 개구부에 인접하여 분자가 결합하는 것은 포어를 통한 개방-채널 이온 흐름에 대한 효과가 있을 것이다. 상기 기술된 핵산 시퀀싱 용도와 유사한 방식으로, 개방-채널 이온 흐름에서의 변동을 전류의 변화에 의해 적절한 측정 기술을 사용하여 측정할 수 있다 (예를 들어, WO 2000/28312 및 문헌 [D. Stoddart et al., Proc. Natl. Acad. Sci., 2010, 106, 7702-7] 또는 WO 2009/077734). 전류 감소에 의해 측정된 바와 같은 이온 흐름의 감소 정도는 포어 내부 또는 포어에 인접한 장애물의 크기에 관련된다. 따라서, 피분석물로 또한 지칭되는 관심 분자가 포어 내에 또는 포어 근처에서 결합하는 것은 검출가능 및 측정가능한 이벤트를 제공하고, 이에 의해 생물학적 센서의 기초를 형성한다. 나노포어 센싱을 위한 적절한 분자는 핵산; 단백질; 펩티드; 및 소형 분자 예컨대 제약, 독소 또는 시토카인을 포함한다.
생물학적 분자를 검출하는 것은 개인화된 약물 개발, 의학, 진단, 생명과학 연구, 환경 모니터링, 및 보안 및/또는 방위 산업에서 용도가 발견된다.
본 발명의 실시양태에서, 본원에 개시된 야생형 이. 콜라이 CsgG 나노포어 또는 변형된 이. 콜라이 CsgG 나노포어, 또는 이의 상동체는 분자 센서로서의 역할을 할 수 있다. 피분석물 검출을 위한 절차가 문헌 [Howorka et al. Nature Biotechnology (2012) Jun 7; 30(6):506-7]에 기술되어 있다. 검출될 피분석물 분자는 채널의 한쪽 면에, 또는 채널 자체의 내강 내에 결합할 수 있다. 센싱될 분자의 크기에 의해 결합 위치가 결정될 수 있다. 야생형 CsgG 포어가 센서로서 작용할 수 있거나, 또는 본 발명의 실시양태에서, 센싱될 분자의 결합 강도, 결합 위치 또는 결합 특이성을 증가시키도록 재조합 또는 화학적 방법을 통해 CsgG 포어가 변형될 수 있다. 전형적인 변형은 센싱될 분자의 구조에 대해 상보적인 특정 결합 모이어티를 부가하는 것을 포함한다. 피분석물 분자가 핵산을 포함하는 경우, 이러한 결합 모이어티는 시클로덱스트린 또는 올리고뉴클레오티드를 포함할 수 있거나; 소형 분자의 경우, 이는 공지된 상보적 결합 영역, 예를 들어 항체 또는 비-항체 분자의 항원 결합 부분 (단일쇄 가변 단편 (scFv) 영역 또는 T-세포 수용체 (TCR)로부터의 항원 인식 도메인을 포함함)일 수 있거나; 또는 단백질의 경우, 이는 표적 단백질에 대한 공지된 리간드일 수 있다. 이러한 방식으로, 야생형 이. 콜라이 CsgG 나노포어 또는 변형된 이. 콜라이 CsgG 나노포어, 또는 이의 상동체가 적절한 항원 (에피토프를 포함함)이 샘플 내에 존재하는 것을 검출하는 분자 센서로서 작용할 수 있게 될 수 있고, 이러한 항원은 수용체를 포함하는 세포 표면 항원, 고형 종양 또는 혈액암 세포 (예를 들어 림프종 또는 백혈병)의 마커, 바이러스 항원, 박테리아 항원, 원충 항원, 알레르기 항원, 알레르기 관련 분자, 알부민 (예를 들어 인간, 설치류 또는 소), 형광 분자 (플로오레세인을 포함함), 혈액형 항원, 소형 분자, 약물, 효소, 효소 또는 효소 기질의 촉매 부위, 및 효소 기질의 전이 상태 유사체를 포함한다.
공지된 유전자 조작 및 재조합 DNA 기술을 사용하여 변형이 달성될 수 있다. 임의의 개조의 위치를 정하는 것은 센싱될 분자의 성질, 예를 들어, 크기, 3차원 구조, 및 이의 생화학적 성질에 좌우될 것이다. 개조된 구조의 선택은 전산 구조 디자인을 이용할 수 있다. 예정되는 센싱 용도에 대해 각각 특이적으로 개조된 일련의 맞춤 CsgG 나노포어가 구상된다. 표면 플라즈몬 공명을 사용하여 분자성 상호작용을 검출하는 비아코어(BIAcore)® (비아코어 인크(BIAcore, Inc.), 뉴저지주 피스카타웨이)를 사용하여 단백질-단백질 상호작용 또는 단백질-소형 분자 상호작용의 결정 및 최적화를 연구할 수 있다 (www.biacore.com을 또한 참조한다).
한 실시양태에서, CsgG 포어는 단백질 N-말단의 지질화를 방지하기 위해 N-말단 Cys 잔기가 대안적인 아미노산으로 교체된 수용성 8량체 형태의 것일 수 있다. 대안적인 실시양태에서, 박테리아 지질화 경로에 의한 프로세싱을 피하기 위해 N-말단 리더 서열의 제거에 의해 단백질이 세포질에서 발현될 수 있다.
CsgG 단량체 가용성 단백질, 8량체 가용성 단백질 및 올리고머성 지질화 CsgG 포어의 제작 방법이 본원에 참고로 포함된 문헌 [Goyal et al., Nature, 2014; 516(7530): 250-3], 및 실시예 1 및 2에서 기술된다.
돌연변이체
CsgG
단량체
본 발명은 돌연변이체 CsgG 단량체를 제공한다. 이러한 돌연변이체 CsgG 단량체는 본 발명의 포어를 형성하는데 사용될 수 있다. 돌연변이체 CsgG 단량체는 단량체의 서열이 야생형 CsgG 단량체의 것과 상이하고 포어를 형성하는 능력을 유지하는 단량체이다. 포어를 형성하는 돌연변이체 단량체의 능력을 확인하는 방법이 관련 기술 분야에 널리 공지되어 있고, 하기에서 더욱 상세하게 논의된다.
돌연변이체 단량체는 개선된 폴리뉴클레오티드 판독 성질을 갖고, 즉 개선된 폴리뉴클레오티드 포착 및 뉴클레오티드 판별을 나타낸다. 특히, 돌연변이체 단량체로부터 구축된 포어는 야생형보다 더 쉽게 뉴클레오티드 및 폴리뉴클레오티드를 포착한다. 또한, 돌연변이체 단량체로부터 구축된 포어는 증가된 전류 범위 (이는 상이한 뉴클레오티드들을 판별하는 것을 더욱 쉽게 만든다), 및 감소된 상태 변동 (이는 신호-대-노이즈 비를 증가시킨다)을 나타낸다. 또한, 돌연변이체로부터 구축된 포어를 통해 폴리뉴클레오티드가 이동할 때 전류에 기여하는 뉴클레오티드의 개수가 감소된다. 이는 폴리뉴클레오티드가 포어를 통해 이동할 때의 관찰된 전류와 폴리뉴클레오티드 서열 사이의 직접적인 관계를 확인하는 것을 더욱 쉽게 만든다. 또한, 돌연변이체 단량체로부터 구축된 포어는 증가된 처리량을 나타낼 수 있고, 즉, 피분석물, 예컨대 폴리뉴클레오티드와 상호작용할 가능성이 더 많다. 이는 포어를 사용하여 피분석물을 특성화하는 것을 더욱 쉽게 만든다. 돌연변이체 단량체로부터 구축된 포어는 막 내로 더욱 쉽게 삽입될 수 있다.
본 발명의 돌연변이체 단량체는 서열식별번호: 390에서 제시된 서열의 변이체를 포함한다. 서열식별번호: 390은 에스케리키아 콜라이 균주 K-12 아계 MC4100으로부터의 야생형 CsgG 단량체이다. 서열식별번호: 390의 변이체는 서열식별번호: 390의 것과 다른 아미노산 서열을 갖고 포어를 형성하는 이의 능력을 유지하는 폴리펩티드이다. 포어를 형성하는 변이체의 능력을 관련 기술 분야에 공지된 임의의 방법을 사용하여 검정할 수 있다. 예를 들어, 변이체를 다른 적합한 서브유닛과 함께 양친매성 층 내로 삽입할 수 있고, 포어를 형성하도록 올리고머화되는 이의 능력을 결정할 수 있다. 서브유닛을 막, 예컨대 양친매성 층 내로 삽입하는 방법이 관련 기술 분야에 공지되어 있다. 예를 들어, 서브유닛이 막으로 확산되고, 막에 결합하여 기능성 상태로 조립되는 것에 의해 삽입되도록, 3-블럭 공중합체 막을 함유하는 용액 내에 서브유닛을 정제된 형태로 현탁시킬 수 있다
본원에서의 모든 논의에서, 아미노산에 대한 표준 1-문자 코드가 사용된다. 이는 하기와 같다: 알라닌 (A), 아르기닌 (R), 아스파라긴 (N), 아스파르트산 (D), 시스테인 (C), 글루탐산 (E), 글루타민 (Q), 글리신 (G), 히스티딘 (H), 이소류신 (I), 류신 (L), 리신 (K), 메티오닌 (M), 페닐알라닌 (F), 프롤린 (P), 세린 (S), 트레오닌 (T), 트립토판 (W), 타이로신 (Y) 및 발린 (V). 또한 표준 치환 표기법이 사용되고, 즉 Q42R은 위치 42의 Q가 R로 교체된다는 것을 의미한다.
한 실시양태에서, 본 발명의 돌연변이체 단량체는 하기 중 하나 이상을 포함하는 서열식별번호: 390의 변이체를 포함한다: (i) 하기 위치 N40, D43, E44, S54, S57, Q62, R97, E101, E124, E131, R142, T150 및 R192에서의 하나 이상의 돌연변이 (즉, 상기 위치 중 하나 이상에서의 돌연변이), 예컨대 하기 위치 N40, D43, E44, S54, S57, Q62, E101, E131 및 T150 또는 N40, D43, E44, E101 및 E131에서의 하나 이상의 돌연변이 (즉, 상기 위치 중 하나 이상에서의 돌연변이); (ii) Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이; (iii) Q42R 또는 Q42K; (iv) K49R; (v) N102R, N102F, N102Y 또는 N102W; (vi) D149N, D149Q 또는 D149R; (vii) E185N, E185Q 또는 E185R; (viii) D195N, D195Q 또는 D195R; (ix) E201N, E201Q 또는 E201R; (x) E203N, E203Q 또는 E203R; 및 (xi) 하기 위치 F48, K49, P50, Y51, P52, A53, S54, N55, F56 및 S57 중 하나 이상의 결실. 변이체는 (i) 내지 (xi)의 임의의 조합을 포함할 수 있다. 특히, 변이체는 하기를 포함할 수 있다: {i} {ii} {iii} {iv} {v} {vi} {vii} {viii} {ix} {x} {xi} {i,ii} {i,iii} {i,iv} {i,v} {i,vi} {i,vii} {i,viii} {i,ix} {i,x} {i,xi} {ii,iii} {ii,iv} {ii,v} {ii,vi} {ii,vii} {ii,viii} {ii,ix} {ii,x} {ii,xi} {iii,iv} {iii,v} {iii,vi} {iii,vii} {iii,viii} {iii,ix} {iii,x} {iii,xi} {iv,v} {iv,vi} {iv,vii} {iv,viii} {iv,ix} {iv,x} {iv,xi} {v,vi} {v,vii} {v,viii} {v,ix} {v,x} {v,xi} {vi,vii} {vi,viii} {vi,ix} {vi,x} {vi,xi} {vii,viii} {vii,ix} {vii,x} {vii,xi} {viii,ix} {viii,x} {viii,xi} {ix,x} {ix,xi} {x,xi} {i,ii,iii} {i,ii,iv} {i,ii,v} {i,ii,vi} {i,ii,vii} {i,ii,viii} {i,ii,ix} {i,ii,x} {i,ii,xi} {i,iii,iv} {i,iii,v} {i,iii,vi} {i,iii,vii} {i,iii,viii} {i,iii,ix} {i,iii,x} {i,iii,xi} {i,iv,v} {i,iv,vi} {i,iv,vii} {i,iv,viii} {i,iv,ix} {i,iv,x} {i,iv,xi} {i,v,vi} {i,v,vii} {i,v,viii} {i,v,ix} {i,v,x} {i,v,xi} {i,vi,vii} {i,vi,viii} {i,vi,ix} {i,vi,x} {i,vi,xi} {i,vii,viii} {i,vii,ix} {i,vii,x} {i,vii,xi} {i,viii,ix} {i,viii,x} {i,viii,xi} {i,ix,x} {i,ix,xi} {i,x,xi} {ii,iii,iv} {ii,iii,v} {ii,iii,vi} {ii,iii,vii} {ii,iii,viii} {ii,iii,ix} {ii,iii,x} {ii,iii,xi} {ii,iv,v} {ii,iv,vi} {ii,iv,vii} {ii,iv,viii} {ii,iv,ix} {ii,iv,x} {ii,iv,xi} {ii,v,vi} {ii,v,vii} {ii,v,viii} {ii,v,ix} {ii,v,x} {ii,v,xi} {ii,vi,vii} {ii,vi,viii} {ii,vi,ix} {ii,vi,x} {ii,vi,xi} {ii,vii,viii} {ii,vii,ix} {ii,vii,x} {ii,vii,xi} {ii,viii,ix} {ii,viii,x} {ii,viii,xi} {ii,ix,x} {ii,ix,xi} {ii,x,xi} {iii,iv,v} {iii,iv,vi} {iii,iv,vii} {iii,iv,viii} {iii,iv,ix} {iii,iv,x} {iii,iv,xi} {iii,v,vi} {iii,v,vii} {iii,v,viii} {iii,v,ix} {iii,v,x} {iii,v,xi} {iii,vi,vii} {iii,vi,viii} {iii,vi,ix} {iii,vi,x} {iii,vi,xi} {iii,vii,viii} {iii,vii,ix} {iii,vii,x} {iii,vii,xi} {iii,viii,ix} {iii,viii,x} {i,ii,iii,iv} {i,ii,iii,v} {i,ii,iii,vi} {i,ii,iii,vii} {i,ii,iii,viii} {i,ii,iii,ix} {i,ii,iii,x} {i,ii,iii,xi} {i,ii,iv,v} {i,ii,iv,vi} {i,ii,iv,vii} {i,ii,iv,viii} {i,ii,iv,ix} {i,ii,iv,x} {i,ii,iv,xi} {i,ii,v,vi} {i,ii,v,vii} {i,ii,v,viii} {i,ii,v,ix} {i,ii,v,x} {i,ii,v,xi} {i,ii,vi,vii} {i,ii,vi,viii} {i,ii,vi,ix} {i,ii,vi,x} {i,ii,vi,xi} {i,ii,vii,viii} {i,ii,vii,ix} {i,ii,vii,x} {i,ii,vii,xi} {i,ii,viii,ix} {i,ii,viii,x} {i,ii,viii,xi} {i,ii,ix,x} {i,ii,ix,xi} {i,ii,x,xi} {i,iii,iv,v} {i,iii,iv,vi} {i,iii,iv,vii} {i,iii,iv,viii} {i,iii,iv,ix} {i,iii,iv,x} {i,iii,iv,xi} {i,iii,v,vi} {i,iii,v,vii} {i,iii,v,viii} {i,iii,v,ix} {i,iii,v,x} {i,iii,v,xi} {i,iii,vi,vii} {i,iii,vi,viii} {i,iii,vi,ix} {i,iii,vi,x} {i,iii,vi,xi} {i,iii,vii,viii} {i,iii,vii,ix} {i,iii,vii,x} {i,iii,vii,xi} {i,iii,viii,ix} {i,iii,viii,x} {i,iii,viii,xi} {i,iii,ix,x} {i,iii,ix,xi} {i,iii,x,xi} {i,iv,v,vi} {i,iv,v,vii} {i,iv,v,viii} {i,iv,v,ix} {i,iv,v,x} {i,iv,v,xi} {i,iv,vi,vii} {i,iv,vi,viii} {i,iv,vi,ix} {i,iv,vi,x} {i,iv,vi,xi} {i,iv,vii,viii} {i,iv,vii,ix} {i,iv,vii,x} {i,iv,vii,xi} {i,iv,viii,ix} {i,iv,viii,x} {i,iv,viii,xi} {i,iv,ix,x} {i,iv,ix,xi} {i,iv,x,xi} {i,v,vi,vii} {i,v,vi,viii} {i,v,vi,ix} {i,v,vi,x} {i,v,vi,xi} {i,v,vii,viii} {i,v,vii,ix} {i,v,vii,x} {i,v,vii,xi} {i,v,viii,ix} {i,v,viii,x} {i,v,viii,xi} {i,v,ix,x} {i,v,ix,xi} {i,v,x,xi} {i,vi,vii,viii} {i,vi,vii,ix} {i,vi,vii,x} {i,vi,vii,xi} {i,vi,viii,ix} {i,vi,viii,x} {i,vi,viii,xi} {i,vi,ix,x} {i,vi,ix,xi} {i,vi,x,xi} {i,vii,viii,ix} {i,vii,viii,x} {i,vii,viii,xi} {i,vii,ix,x} {i,vii,ix,xi} {i,vii,x,xi} {i,viii,ix,x} {i,viii,ix,xi} {i,viii,x,xi} {i,ix,x,xi} {ii,iii,iv,v} {ii,iii,iv,vi} {ii,iii,iv,vii} {ii,iii,iv,viii} {ii,iii,iv,ix} {ii,iii,iv,x} {ii,iii,iv,xi} {ii,iii,v,vi} {ii,iii,v,vii} {ii,iii,v,viii} {ii,iii,v,ix} {ii,iii,v,x} {ii,iii,v,xi} {ii,iii,vi,vii} {ii,iii,vi,viii} {ii,iii,vi,ix} {ii,iii,vi,x} {ii,iii,vi,xi} {ii,iii,vii,viii} {ii,iii,vii,ix} {ii,iii,vii,x} {ii,iii,vii,xi} {ii,iii,viii,ix} {ii,iii,viii,x} {ii,iii,viii,xi} {ii,iii,ix,x} {ii,iii,ix,xi} {ii,iii,x,xi} {ii,iv,v,vi} {ii,iv,v,vii} {ii,iv,v,viii} {ii,iv,v,ix} {ii,iv,v,x} {ii,iv,v,xi} {ii,iv,vi,vii} {ii,iv,vi,viii} {ii,iv,vi,ix} {ii,iv,vi,x} {ii,iv,vi,xi} {ii,iv,vii,viii} {ii,iv,vii,ix} {ii,iv,vii,x} {ii,iv,vii,xi} {ii,iv,viii,ix} {ii,iv,viii,x} {ii,iv,viii,xi} {ii,iv,ix,x} {ii,iv,ix,xi} {ii,iv,x,xi} {ii,v,vi,vii} {ii,v,vi,viii} {ii,v,vi,ix} {ii,v,vi,x} {ii,v,vi,xi} {ii,v,vii,viii} {ii,v,vii,ix} {ii,v,vii,x} {ii,v,vii,xi} {ii,v,viii,ix} {ii,v,viii,x} {ii,v,viii,xi} {ii,v,ix,x} {ii,v,ix,xi} {ii,v,x,xi} {ii,vi,vii,viii} {ii,vi,vii,ix} {ii,vi,vii,x} {ii,vi,vii,xi} {ii,vi,viii,ix} {ii,vi,viii,x} {ii,vi,viii,xi} {ii,vi,ix,x} {ii,vi,ix,xi} {ii,vi,x,xi} {ii,vii,viii,ix} {ii,vii,viii,x} {ii,vii,viii,xi} {ii,vii,ix,x} {ii,vii,ix,xi} {ii,vii,x,xi} {ii,viii,ix,x} {ii,viii,ix,xi} {ii,viii,x,xi} {ii,ix,x,xi} {iii,iv,v,vi} {iii,iv,v,vii} {iii,iv,v,viii} {iii,iv,v,ix} {iii,iv,v,x} {iii,iv,v,xi} {iii,iv,vi,vii} {iii,iv,vi,viii} {iii,iv,vi,ix} {iii,iv,vi,x} {iii,iv,vi,xi} {iii,iv,vii,viii} {iii,iv,vii,ix} {iii,iv,vii,x} {iii,iv,vii,xi} {iii,iv,viii,ix} {iii,iv,viii,x} {iii,iv,viii,xi} {iii,iv,ix,x} {iii,iv,ix,xi} {iii,iv,x,xi} {iii,v,vi,vii} {iii,v,vi,viii} {iii,v,vi,ix} {iii,v,vi,x} {iii,v,vi,xi} {iii,v,vii,viii} {iii,v,vii,ix} {iii,v,vii,x} {iii,v,vii,xi} {iii,v,viii,ix} {iii,v,viii,x} {iii,v,viii,xi} {iii,v,ix,x} {iii,v,ix,xi} {iii,v,x,xi} {iii,vi,vii,viii} {iii,vi,vii,ix} {iii,vi,vii,x} {iii,vi,vii,xi} {iii,vi,viii,ix} {iii,vi,viii,x} {iii,vi,viii,xi} {iii,vi,ix,x} {iii,vi,ix,xi} {iii,vi,x,xi} {iii,vii,viii,ix} {iii,vii,viii,x} {iii,vii,viii,xi} {iii,vii,ix,x} {iii,vii,ix,xi} {iii,vii,x,xi} {iii,viii,ix,x} {iii,viii,ix,xi} {iii,viii,x,xi} {iii,ix,x,xi} {iv,v,vi,vii} {iv,v,vi,viii} {iv,v,vi,ix} {iv,v,vi,x} {iv,v,vi,xi} {iv,v,vii,viii} {iv,v,vii,ix} {iv,v,vii,x} {iv,v,vii,xi} {iv,v,viii,ix} {iv,v,viii,x} {iv,v,viii,xi} {iv,v,ix,x} {iv,v,ix,xi} {iv,v,x,xi} {iv,vi,vii,viii} {iv,vi,vii,ix} {iv,vi,vii,x} {iv,vi,vii,xi} {iv,vi,viii,ix} {iv,vi,viii,x} {iv,vi,viii,xi} {iv,vi,ix,x} {iv,vi,ix,xi} {iv,vi,x,xi} {iv,vii,viii,ix} {iv,vii,viii,x} {iv,vii,viii,xi} {iv,vii,ix,x} {iv,vii,ix,xi} {iv,vii,x,xi} {iv,viii,ix,x} {iv,viii,ix,xi} {iv,viii,x,xi} {iv,ix,x,xi} {v,vi,vii,viii} {v,vi,vii,ix} {v,vi,vii,x} {v,vi,vii,xi} {v,vi,viii,ix} {v,vi,viii,x} {v,vi,viii,xi} {v,vi,ix,x} {v,vi,ix,xi} {v,vi,x,xi} {v,vii,viii,ix} {v,vii,viii,x} {v,vii,viii,xi} {v,vii,ix,x} {v,vii,ix,xi} {v,vii,x,xi} {v,viii,ix,x} {v,viii,ix,xi} {v,viii,x,xi} {v,ix,x,xi} {vi,vii,viii,ix} {vi,vii,viii,x} {vi,vii,viii,xi} {vi,vii,ix,x} {vi,vii,ix,xi} {vi,vii,x,xi} {vi,viii,ix,x} {vi,viii,ix,xi} {vi,viii,x,xi} {vi,ix,x,xi} {vii,viii,ix,x} {vii,viii,ix,xi} {vii,viii,x,xi} {vii,ix,x,xi} {viii,ix,x,xi} {i,ii,iii,iv,v} {i,ii,iii,iv,vi} {i,ii,iii,iv,vii} {i,ii,iii,iv,viii} {i,ii,iii,iv,ix} {i,ii,iii,iv,x} {i,ii,iii,iv,xi} {i,ii,iii,v,vi} {i,ii,iii,v,vii} {i,ii,iii,v,viii} {i,ii,iii,v,ix} {i,ii,iii,v,x} {i,ii,iii,v,xi} {i,ii,iii,vi,vii} {i,ii,iii,vi,viii} {i,ii,iii,vi,ix} {i,ii,iii,vi,x} {i,ii,iii,vi,xi} {i,ii,iii,vii,viii} {i,ii,iii,vii,ix} {i,ii,iii,vii,x} {i,ii,iii,vii,xi} {i,ii,iii,viii,ix} {i,ii,iii,viii,x} {i,ii,iii,viii,xi} {i,ii,iii,ix,x} {i,ii,iii,ix,xi} {i,ii,iii,x,xi} {i,ii,iv,v,vi} {i,ii,iv,v,vii} {i,ii,iv,v,viii} {i,ii,iv,v,ix} {i,ii,iv,v,x} {i,ii,iv,v,xi} {i,ii,iv,vi,vii} {i,ii,iv,vi,viii} {i,ii,iv,vi,ix} {i,ii,iv,vi,x} {i,ii,iv,vi,xi} {i,ii,iv,vii,viii} {i,ii,iv,vii,ix} {i,ii,iv,vii,x} {i,ii,iv,vii,xi} {i,ii,iv,viii,ix} {i,ii,iv,viii,x} {i,ii,iv,viii,xi} {i,ii,iv,ix,x} {i,ii,iv,ix,xi} {i,ii,iv,x,xi} {i,ii,v,vi,vii} {i,ii,v,vi,viii} {i,ii,v,vi,ix} {i,ii,v,vi,x} {i,ii,v,vi,xi} {i,ii,v,vii,viii} {i,ii,v,vii,ix} {i,ii,v,vii,x} {i,ii,v,vii,xi} {i,ii,v,viii,ix} {i,ii,v,viii,x} {i,ii,v,viii,xi} {i,ii,v,ix,x} {i,ii,v,ix,xi} {i,ii,v,x,xi} {i,ii,vi,vii,viii} {i,ii,vi,vii,ix} {i,ii,vi,vii,x} {i,ii,vi,vii,xi} {i,ii,vi,viii,ix} {i,ii,vi,viii,x} {i,ii,vi,viii,xi} {i,ii,vi,ix,x} {i,ii,vi,ix,xi} {i,ii,vi,x,xi} {i,ii,vii,viii,ix} {i,ii,vii,viii,x} {i,ii,vii,viii,xi} {i,ii,vii,ix,x} {i,ii,vii,ix,xi} {i,ii,vii,x,xi} {i,ii,viii,ix,x} {i,ii,viii,ix,xi} {i,ii,viii,x,xi} {i,ii,ix,x,xi} {i,iii,iv,v,vi} {i,iii,iv,v,vii} {i,iii,iv,v,viii} {i,iii,iv,v,ix} {i,iii,iv,v,x} {i,iii,iv,v,xi} {i,iii,iv,vi,vii} {i,iii,iv,vi,viii} {i,iii,iv,vi,ix} {i,iii,iv,vi,x} {i,iii,iv,vi,xi} {i,iii,iv,vii,viii} {i,iii,iv,vii,ix} {i,iii,iv,vii,x} {i,iii,iv,vii,xi} {i,iii,iv,viii,ix} {i,iii,iv,viii,x} {i,iii,iv,viii,xi} {i,iii,iv,ix,x} {i,iii,iv,ix,xi} {i,iii,iv,x,xi} {i,iii,v,vi,vii} {i,iii,v,vi,viii} {i,iii,v,vi,ix} {i,iii,v,vi,x} {i,iii,v,vi,xi} {i,iii,v,vii,viii} {i,iii,v,vii,ix} {i,iii,v,vii,x} {i,iii,v,vii,xi} {i,iii,v,viii,ix} {i,iii,v,viii,x} {i,iii,v,viii,xi} {i,iii,v,ix,x} {i,iii,v,ix,xi} {i,iii,v,x,xi} {i,iii,vi,vii,viii} {i,iii,vi,vii,ix} {i,iii,vi,vii,x} {i,iii,vi,vii,xi} {i,iii,vi,viii,ix} {i,iii,vi,viii,x} {i,iii,vi,viii,xi} {i,iii,vi,ix,x} {i,iii,vi,ix,xi} {i,iii,vi,x,xi} {i,iii,vii,viii,ix} {i,iii,vii,viii,x} {i,iii,vii,viii,xi} {i,iii,vii,ix,x} {i,iii,vii,ix,xi} {i,iii,vii,x,xi} {i,iii,viii,ix,x} {i,iii,viii,ix,xi} {i,iii,viii,x,xi} {i,iii,ix,x,xi} {i,iv,v,vi,vii} {i,iv,v,vi,viii} {i,iv,v,vi,ix} {i,iv,v,vi,x} {i,iv,v,vi,xi} {i,iv,v,vii,viii} {i,iv,v,vii,ix} {i,iv,v,vii,x} {i,iv,v,vii,xi} {i,iv,v,viii,ix} {i,iv,v,viii,x} {i,iv,v,viii,xi} {i,iv,v,ix,x} {i,iv,v,ix,xi} {i,iv,v,x,xi} {i,iv,vi,vii,viii} {i,iv,vi,vii,ix} {i,iv,vi,vii,x} {i,iv,vi,vii,xi} {i,iv,vi,viii,ix} {i,iv,vi,viii,x} {i,iv,vi,viii,xi} {i,iv,vi,ix,x} {i,iv,vi,ix,xi} {i,iv,vi,x,xi} {i,iv,vii,viii,ix} {i,iv,vii,viii,x} {i,iv,vii,viii,xi} {i,iv,vii,ix,x} {i,iv,vii,ix,xi} {i,iv,vii,x,xi} {i,iv,viii,ix,x} {i,iv,viii,ix,xi} {i,iv,viii,x,xi} {i,iv,ix,x,xi} {i,v,vi,vii,viii} {i,v,vi,vii,ix} {i,v,vi,vii,x} {i,v,vi,vii,xi} {i,v,vi,viii,ix} {i,v,vi,viii,x} {i,v,vi,viii,xi} {i,v,vi,ix,x} {i,v,vi,ix,xi} {i,v,vi,x,xi} {i,v,vii,viii,ix} {i,v,vii,viii,x} {i,v,vii,viii,xi} {i,v,vii,ix,x} {i,v,vii,ix,xi} {i,v,vii,x,xi} {i,v,viii,ix,x} {i,v,viii,ix,xi} {i,v,viii,x,xi} {i,v,ix,x,xi} {i,vi,vii,viii,ix} {i,vi,vii,viii,x} {i,vi,vii,viii,xi} {i,vi,vii,ix,x} {i,vi,vii,ix,xi} {i,vi,vii,x,xi} {i,vi,viii,ix,x} {i,vi,viii,ix,xi} {i,vi,viii,x,xi} {i,vi,ix,x,xi} {i,vii,viii,ix,x} {i,vii,viii,ix,xi} {i,vii,viii,x,xi} {i,vii,ix,x,xi} {i,viii,ix,x,xi} {ii,iii,iv,v,vi} {ii,iii,iv,v,vii} {ii,iii,iv,v,viii} {ii,iii,iv,v,ix} {ii,iii,iv,v,x} {ii,iii,iv,v,xi} {ii,iii,iv,vi,vii} {ii,iii,iv,vi,viii} {ii,iii,iv,vi,ix} {ii,iii,iv,vi,x} {ii,iii,iv,vi,xi} {ii,iii,iv,vii,viii} {ii,iii,iv,vii,ix} {ii,iii,iv,vii,x} {ii,iii,iv,vii,xi} {ii,iii,iv,viii,ix} {ii,iii,iv,viii,x} {ii,iii,iv,viii,xi} {ii,iii,iv,ix,x} {ii,iii,iv,ix,xi} {ii,iii,iv,x,xi} {ii,iii,v,vi,vii} {ii,iii,v,vi,viii} {ii,iii,v,vi,ix} {ii,iii,v,vi,x} {ii,iii,v,vi,xi} {ii,iii,v,vii,viii} {ii,iii,v,vii,ix} {ii,iii,v,vii,x} {ii,iii,v,vii,xi} {ii,iii,v,viii,ix} {ii,iii,v,viii,x} {ii,iii,v,viii,xi} {ii,iii,v,ix,x} {ii,iii,v,ix,xi} {ii,iii,v,x,xi} {ii,iii,vi,vii,viii} {ii,iii,vi,vii,ix} {ii,iii,vi,vii,x} {ii,iii,vi,vii,xi} {ii,iii,vi,viii,ix} {ii,iii,vi,viii,x} {ii,iii,vi,viii,xi} {ii,iii,vi,ix,x} {ii,iii,vi,ix,xi} {ii,iii,vi,x,xi} {ii,iii,vii,viii,ix} {ii,iii,vii,viii,x} {ii,iii,vii,viii,xi} {ii,iii,vii,ix,x} {ii,iii,vii,ix,xi} {ii,iii,vii,x,xi} {ii,iii,viii,ix,x} {ii,iii,viii,ix,xi} {ii,iii,viii,x,xi} {ii,iii,ix,x,xi} {ii,iv,v,vi,vii} {ii,iv,v,vi,viii} {ii,iv,v,vi,ix} {ii,iv,v,vi,x} {ii,iv,v,vi,xi} {ii,iv,v,vii,viii} {ii,iv,v,vii,ix} {ii,iv,v,vii,x} {ii,iv,v,vii,xi} {ii,iv,v,viii,ix} {ii,iv,v,viii,x} {ii,iv,v,viii,xi} {ii,iv,v,ix,x} {ii,iv,v,ix,xi} {ii,iv,v,x,xi} {ii,iv,vi,vii,viii} {ii,iv,vi,vii,ix} {ii,iv,vi,vii,x} {ii,iv,vi,vii,xi} {ii,iv,vi,viii,ix} {ii,iv,vi,viii,x} {ii,iv,vi,viii,xi} {ii,iv,vi,ix,x} {ii,iv,vi,ix,xi} {ii,iv,vi,x,xi} {ii,iv,vii,viii,ix} {ii,iv,vii,viii,x} {ii,iv,vii,viii,xi} {ii,iv,vii,ix,x} {ii,iv,vii,ix,xi} {ii,iv,vii,x,xi} {ii,iv,viii,ix,x} {ii,iv,viii,ix,xi} {ii,iv,viii,x,xi} {ii,iv,ix,x,xi} {ii,v,vi,vii,viii} {ii,v,vi,vii,ix} {ii,v,vi,vii,x} {ii,v,vi,vii,xi} {ii,v,vi,viii,ix} {ii,v,vi,viii,x} {ii,v,vi,viii,xi} {ii,v,vi,ix,x} {ii,v,vi,ix,xi} {ii,v,vi,x,xi} {ii,v,vii,viii,ix} {ii,v,vii,viii,x} {ii,v,vii,viii,xi} {ii,v,vii,ix,x} {ii,v,vii,ix,xi} {ii,v,vii,x,xi} {ii,v,viii,ix,x} {ii,v,viii,ix,xi} {ii,v,viii,x,xi} {ii,v,ix,x,xi} {ii,vi,vii,viii,ix} {ii,vi,vii,viii,x} {ii,vi,vii,viii,xi} {ii,vi,vii,ix,x} {ii,vi,vii,ix,xi} {ii,vi,vii,x,xi} {ii,vi,viii,ix,x} {ii,vi,viii,ix,xi} {ii,vi,viii,x,xi} {ii,vi,ix,x,xi} {ii,vii,viii,ix,x} {ii,vii,viii,ix,xi} {ii,vii,viii,x,xi} {ii,vii,ix,x,xi} {ii,viii,ix,x,xi} {iii,iv,v,vi,vii} {iii,iv,v,vi,viii} {iii,iv,v,vi,ix} {iii,iv,v,vi,x} {iii,iv,v,vi,xi} {iii,iv,v,vii,viii} {iii,iv,v,vii,ix} {iii,iv,v,vii,x} {iii,iv,v,vii,xi} {iii,iv,v,viii,ix} {iii,iv,v,viii,x} {iii,iv,v,viii,xi} {iii,iv,v,ix,x} {iii,iv,v,ix,xi} {iii,iv,v,x,xi} {iii,iv,vi,vii,viii} {iii,iv,vi,vii,ix} {iii,iv,vi,vii,x} {iii,iv,vi,vii,xi} {iii,iv,vi,viii,ix} {iii,iv,vi,viii,x} {iii,iv,vi,viii,xi} {iii,iv,vi,ix,x} {iii,iv,vi,ix,xi} {iii,iv,vi,x,xi} {iii,iv,vii,viii,ix} {iii,iv,vii,viii,x} {iii,iv,vii,viii,xi} {iii,iv,vii,ix,x} {iii,iv,vii,ix,xi} {iii,iv,vii,x,xi} {iii,iv,viii,ix,x} {iii,iv,viii,ix,xi} {iii,iv,viii,x,xi} {iii,iv,ix,x,xi} {iii,v,vi,vii,viii} {iii,v,vi,vii,ix} {iii,v,vi,vii,x} {iii,v,vi,vii,xi} {iii,v,vi,viii,ix} {iii,v,vi,viii,x} {iii,v,vi,viii,xi} {iii,v,vi,ix,x} {iii,v,vi,ix,xi} {iii,v,vi,x,xi} {iii,v,vii,viii,ix} {iii,v,vii,viii,x} {iii,v,vii,viii,xi} {iii,v,vii,ix,x} {iii,v,vii,ix,xi} {iii,v,vii,x,xi} {iii,v,viii,ix,x} {iii,v,viii,ix,xi} {iii,v,viii,x,xi} {iii,v,ix,x,xi} {iii,vi,vii,viii,ix} {iii,vi,vii,viii,x} {iii,vi,vii,viii,xi} {iii,vi,vii,ix,x} {iii,vi,vii,ix,xi} {iii,vi,vii,x,xi} {iii,vi,viii,ix,x} {iii,vi,viii,ix,xi} {iii,vi,viii,x,xi} {iii,vi,ix,x,xi} {iii,vii,viii,ix,x} {iii,vii,viii,ix,xi} {iii,vii,viii,x,xi} {iii,vii,ix,x,xi} {iii,viii,ix,x,xi} {iv,v,vi,vii,viii} {iv,v,vi,vii,ix} {iv,v,vi,vii,x} {iv,v,vi,vii,xi} {iv,v,vi,viii,ix} {iv,v,vi,viii,x} {iv,v,vi,viii,xi} {iv,v,vi,ix,x} {iv,v,vi,ix,xi} {iv,v,vi,x,xi} {iv,v,vii,viii,ix} {iv,v,vii,viii,x} {iv,v,vii,viii,xi} {iv,v,vii,ix,x} {iv,v,vii,ix,xi} {iv,v,vii,x,xi} {iv,v,viii,ix,x} {iv,v,viii,ix,xi} {iv,v,viii,x,xi} {iv,v,ix,x,xi} {iv,vi,vii,viii,ix} {iv,vi,vii,viii,x} {iv,vi,vii,viii,xi} {iv,vi,vii,ix,x} {iv,vi,vii,ix,xi} {iv,vi,vii,x,xi} {iv,vi,viii,ix,x} {iv,vi,viii,ix,xi} {iv,vi,viii,x,xi} {iv,vi,ix,x,xi} {iv,vii,viii,ix,x} {iv,vii,viii,ix,xi} {iv,vii,viii,x,xi} {iv,vii,ix,x,xi} {iv,viii,ix,x,xi} {v,vi,vii,viii,ix} {v,vi,vii,viii,x} {v,vi,vii,viii,xi} {v,vi,vii,ix,x} {v,vi,vii,ix,xi} {v,vi,vii,x,xi} {v,vi,viii,ix,x} {v,vi,viii,ix,xi} {v,vi,viii,x,xi} {v,vi,ix,x,xi} {v,vii,viii,ix,x} {v,vii,viii,ix,xi} {v,vii,viii,x,xi} {v,vii,ix,x,xi} {v,viii,ix,x,xi} {vi,vii,viii,ix,x} {vi,vii,viii,ix,xi} {vi,vii,viii,x,xi} {vi,vii,ix,x,xi} {vi,viii,ix,x,xi} {vii,viii,ix,x,xi} {i,ii,iii,iv,v,vi} {i,ii,iii,iv,v,vii} {i,ii,iii,iv,v,viii} {i,ii,iii,iv,v,ix} {i,ii,iii,iv,v,x} {i,ii,iii,iv,v,xi} {i,ii,iii,iv,vi,vii} {i,ii,iii,iv,vi,viii} {i,ii,iii,iv,vi,ix} {i,ii,iii,iv,vi,x} {i,ii,iii,iv,vi,xi} {i,ii,iii,iv,vii,viii} {i,ii,iii,iv,vii,ix} {i,ii,iii,iv,vii,x} {i,ii,iii,iv,vii,xi} {i,ii,iii,iv,viii,ix} {i,ii,iii,iv,viii,x} {i,ii,iii,iv,viii,xi} {i,ii,iii,iv,ix,x} {i,ii,iii,iv,ix,xi} {i,ii,iii,iv,x,xi} {i,ii,iii,v,vi,vii} {i,ii,iii,v,vi,viii} {i,ii,iii,v,vi,ix} {i,ii,iii,v,vi,x} {i,ii,iii,v,vi,xi} {i,ii,iii,v,vii,viii} {i,ii,iii,v,vii,ix} {i,ii,iii,v,vii,x} {i,ii,iii,v,vii,xi} {i,ii,iii,v,viii,ix} {i,ii,iii,v,viii,x} {i,ii,iii,v,viii,xi} {i,ii,iii,v,ix,x} {i,ii,iii,v,ix,xi} {i,ii,iii,v,x,xi} {i,ii,iii,vi,vii,viii} {i,ii,iii,vi,vii,ix} {i,ii,iii,vi,vii,x} {i,ii,iii,vi,vii,xi} {i,ii,iii,vi,viii,ix} {i,ii,iii,vi,viii,x} {i,ii,iii,vi,viii,xi} {i,ii,iii,vi,ix,x} {i,ii,iii,vi,ix,xi} {i,ii,iii,vi,x,xi} {i,ii,iii,vii,viii,ix} {i,ii,iii,vii,viii,x} {i,ii,iii,vii,viii,xi} {i,ii,iii,vii,ix,x} {i,ii,iii,vii,ix,xi} {i,ii,iii,vii,x,xi} {i,ii,iii,viii,ix,x} {i,ii,iii,viii,ix,xi} {i,ii,iii,viii,x,xi} {i,ii,iii,ix,x,xi} {i,ii,iv,v,vi,vii} {i,ii,iv,v,vi,viii} {i,ii,iv,v,vi,ix} {i,ii,iv,v,vi,x} {i,ii,iv,v,vi,xi} {i,ii,iv,v,vii,viii} {i,ii,iv,v,vii,ix} {i,ii,iv,v,vii,x} {i,ii,iv,v,vii,xi} {i,ii,iv,v,viii,ix} {i,ii,iv,v,viii,x} {i,ii,iv,v,viii,xi} {i,ii,iv,v,ix,x} {i,ii,iv,v,ix,xi} {i,ii,iv,v,x,xi} {i,ii,iv,vi,vii,viii} {i,ii,iv,vi,vii,ix} {i,ii,iv,vi,vii,x} {i,ii,iv,vi,vii,xi} {i,ii,iv,vi,viii,ix} {i,ii,iv,vi,viii,x} {i,ii,iv,vi,viii,xi} {i,ii,iv,vi,ix,x} {i,ii,iv,vi,ix,xi} {i,ii,iv,vi,x,xi} {i,ii,iv,vii,viii,ix} {i,ii,iv,vii,viii,x} {i,ii,iv,vii,viii,xi} {i,ii,iv,vii,ix,x} {i,ii,iv,vii,ix,xi} {i,ii,iv,vii,x,xi} {i,ii,iv,viii,ix,x} {i,ii,iv,viii,ix,xi} {i,ii,iv,viii,x,xi} {i,ii,iv,ix,x,xi} {i,ii,v,vi,vii,viii} {i,ii,v,vi,vii,ix} {i,ii,v,vi,vii,x} {i,ii,v,vi,vii,xi} {i,ii,v,vi,viii,ix} {i,ii,v,vi,viii,x} {i,ii,v,vi,viii,xi} {i,ii,v,vi,ix,x} {i,ii,v,vi,ix,xi} {i,ii,v,vi,x,xi} {i,ii,v,vii,viii,ix} {i,ii,v,vii,viii,x} {i,ii,v,vii,viii,xi} {i,ii,v,vii,ix,x} {i,ii,v,vii,ix,xi} {i,ii,v,vii,x,xi} {i,ii,v,viii,ix,x} {i,ii,v,viii,ix,xi} {i,ii,v,viii,x,xi} {i,ii,v,ix,x,xi} {i,ii,vi,vii,viii,ix} {i,ii,vi,vii,viii,x} {i,ii,vi,vii,viii,xi} {i,ii,vi,vii,ix,x} {i,ii,vi,vii,ix,xi} {i,ii,vi,vii,x,xi} {i,ii,vi,viii,ix,x} {i,ii,vi,viii,ix,xi} {i,ii,vi,viii,x,xi} {i,ii,vi,ix,x,xi} {i,ii,vii,viii,ix,x} {i,ii,vii,viii,ix,xi} {i,ii,vii,viii,x,xi} {i,ii,vii,ix,x,xi} {i,ii,viii,ix,x,xi} {i,iii,iv,v,vi,vii} {i,iii,iv,v,vi,viii} {i,iii,iv,v,vi,ix} {i,iii,iv,v,vi,x} {i,iii,iv,v,vi,xi} {i,iii,iv,v,vii,viii} {i,iii,iv,v,vii,ix} {i,iii,iv,v,vii,x} {i,iii,iv,v,vii,xi} {i,iii,iv,v,viii,ix} {i,iii,iv,v,viii,x} {i,iii,iv,v,viii,xi} {i,iii,iv,v,ix,x} {i,iii,iv,v,ix,xi} {i,iii,iv,v,x,xi} {i,iii,iv,vi,vii,viii} {i,iii,iv,vi,vii,ix} {i,iii,iv,vi,vii,x} {i,iii,iv,vi,vii,xi} {i,iii,iv,vi,viii,ix} {i,iii,iv,vi,viii,x} {i,iii,iv,vi,viii,xi} {i,iii,iv,vi,ix,x} {i,iii,iv,vi,ix,xi} {i,iii,iv,vi,x,xi} {i,iii,iv,vii,viii,ix} {i,iii,iv,vii,viii,x} {i,iii,iv,vii,viii,xi} {i,iii,iv,vii,ix,x} {i,iii,iv,vii,ix,xi} {i,iii,iv,vii,x,xi} {i,iii,iv,viii,ix,x} {i,iii,iv,viii,ix,xi} {i,iii,iv,viii,x,xi} {i,iii,iv,ix,x,xi} {i,iii,v,vi,vii,viii} {i,iii,v,vi,vii,ix} {i,iii,v,vi,vii,x} {i,iii,v,vi,vii,xi} {i,iii,v,vi,viii,ix} {i,iii,v,vi,viii,x} {i,iii,v,vi,viii,xi} {i,iii,v,vi,ix,x} {i,iii,v,vi,ix,xi} {i,iii,v,vi,x,xi} {i,iii,v,vii,viii,ix} {i,iii,v,vii,viii,x} {i,iii,v,vii,viii,xi} {i,iii,v,vii,ix,x} {i,iii,v,vii,ix,xi} {i,iii,v,vii,x,xi} {i,iii,v,viii,ix,x} {i,iii,v,viii,ix,xi} {i,iii,v,viii,x,xi} {i,iii,v,ix,x,xi} {i,iii,vi,vii,viii,ix} {i,iii,vi,vii,viii,x} {i,iii,vi,vii,viii,xi} {i,iii,vi,vii,ix,x} {i,iii,vi,vii,ix,xi} {i,iii,vi,vii,x,xi} {i,iii,vi,viii,ix,x} {i,iii,vi,viii,ix,xi} {i,iii,vi,viii,x,xi} {i,iii,vi,ix,x,xi} {i,iii,vii,viii,ix,x} {i,iii,vii,viii,ix,xi} {i,iii,vii,viii,x,xi} {i,iii,vii,ix,x,xi} {i,iii,viii,ix,x,xi} {i,iv,v,vi,vii,viii} {i,iv,v,vi,vii,ix} {i,iv,v,vi,vii,x} {i,iv,v,vi,vii,xi} {i,iv,v,vi,viii,ix} {i,iv,v,vi,viii,x} {i,iv,v,vi,viii,xi} {i,iv,v,vi,ix,x} {i,iv,v,vi,ix,xi} {i,iv,v,vi,x,xi} {i,iv,v,vii,viii,ix} {i,iv,v,vii,viii,x} {i,iv,v,vii,viii,xi} {i,iv,v,vii,ix,x} {i,iv,v,vii,ix,xi} {i,iv,v,vii,x,xi} {i,iv,v,viii,ix,x} {i,iv,v,viii,ix,xi} {i,iv,v,viii,x,xi} {i,iv,v,ix,x,xi} {i,iv,vi,vii,viii,ix} {i,iv,vi,vii,viii,x} {i,iv,vi,vii,viii,xi} {i,iv,vi,vii,ix,x} {i,iv,vi,vii,ix,xi} {i,iv,vi,vii,x,xi} {i,iv,vi,viii,ix,x} {i,iv,vi,viii,ix,xi} {i,iv,vi,viii,x,xi} {i,iv,vi,ix,x,xi} {i,iv,vii,viii,ix,x} {i,iv,vii,viii,ix,xi} {i,iv,vii,viii,x,xi} {i,iv,vii,ix,x,xi} {i,iv,viii,ix,x,xi} {i,v,vi,vii,viii,ix} {i,v,vi,vii,viii,x} {i,v,vi,vii,viii,xi} {i,v,vi,vii,ix,x} {i,v,vi,vii,ix,xi} {i,v,vi,vii,x,xi} {i,v,vi,viii,ix,x} {i,v,vi,viii,ix,xi} {i,v,vi,viii,x,xi} {i,v,vi,ix,x,xi} {i,v,vii,viii,ix,x} {i,v,vii,viii,ix,xi} {i,v,vii,viii,x,xi} {i,v,vii,ix,x,xi} {i,v,viii,ix,x,xi} {i,vi,vii,viii,ix,x} {i,vi,vii,viii,ix,xi} {i,vi,vii,viii,x,xi} {i,vi,vii,ix,x,xi} {i,vi,viii,ix,x,xi} {i,vii,viii,ix,x,xi} {ii,iii,iv,v,vi,vii} {ii,iii,iv,v,vi,viii} {ii,iii,iv,v,vi,ix} {ii,iii,iv,v,vi,x} {ii,iii,iv,v,vi,xi} {ii,iii,iv,v,vii,viii} {ii,iii,iv,v,vii,ix} {ii,iii,iv,v,vii,x} {ii,iii,iv,v,vii,xi} {ii,iii,iv,v,viii,ix} {ii,iii,iv,v,viii,x} {ii,iii,iv,v,viii,xi} {ii,iii,iv,v,ix,x} {ii,iii,iv,v,ix,xi} {ii,iii,iv,v,x,xi} {ii,iii,iv,vi,vii,viii} {ii,iii,iv,vi,vii,ix} {ii,iii,iv,vi,vii,x} {ii,iii,iv,vi,vii,xi} {ii,iii,iv,vi,viii,ix} {ii,iii,iv,vi,viii,x} {ii,iii,iv,vi,viii,xi} {ii,iii,iv,vi,ix,x} {ii,iii,iv,vi,ix,xi} {ii,iii,iv,vi,x,xi} {ii,iii,iv,vii,viii,ix} {ii,iii,iv,vii,viii,x} {ii,iii,iv,vii,viii,xi} {ii,iii,iv,vii,ix,x} {ii,iii,iv,vii,ix,xi} {ii,iii,iv,vii,x,xi} {ii,iii,iv,viii,ix,x} {ii,iii,iv,viii,ix,xi} {ii,iii,iv,viii,x,xi} {ii,iii,iv,ix,x,xi} {ii,iii,v,vi,vii,viii} {ii,iii,v,vi,vii,ix} {ii,iii,v,vi,vii,x} {ii,iii,v,vi,vii,xi} {ii,iii,v,vi,viii,ix} {ii,iii,v,vi,viii,x} {ii,iii,v,vi,viii,xi} {ii,iii,v,vi,ix,x} {ii,iii,v,vi,ix,xi} {ii,iii,v,vi,x,xi} {ii,iii,v,vii,viii,ix} {ii,iii,v,vii,viii,x} {ii,iii,v,vii,viii,xi} {ii,iii,v,vii,ix,x} {ii,iii,v,vii,ix,xi} {ii,iii,v,vii,x,xi} {ii,iii,v,viii,ix,x} {ii,iii,v,viii,ix,xi} {ii,iii,v,viii,x,xi} {ii,iii,v,ix,x,xi} {ii,iii,vi,vii,viii,ix} {ii,iii,vi,vii,viii,x} {ii,iii,vi,vii,viii,xi} {ii,iii,vi,vii,ix,x} {ii,iii,vi,vii,ix,xi} {ii,iii,vi,vii,x,xi} {ii,iii,vi,viii,ix,x} {ii,iii,vi,viii,ix,xi} {ii,iii,vi,viii,x,xi} {ii,iii,vi,ix,x,xi} {ii,iii,vii,viii,ix,x} {ii,iii,vii,viii,ix,xi} {ii,iii,vii,viii,x,xi} {ii,iii,vii,ix,x,xi} {ii,iii,viii,ix,x,xi} {ii,iv,v,vi,vii,viii} {ii,iv,v,vi,vii,ix} {ii,iv,v,vi,vii,x} {ii,iv,v,vi,vii,xi} {ii,iv,v,vi,viii,ix} {ii,iv,v,vi,viii,x} {ii,iv,v,vi,viii,xi} {ii,iv,v,vi,ix,x} {ii,iv,v,vi,ix,xi} {ii,iv,v,vi,x,xi} {ii,iv,v,vii,viii,ix} {ii,iv,v,vii,viii,x} {ii,iv,v,vii,viii,xi} {ii,iv,v,vii,ix,x} {ii,iv,v,vii,ix,xi} {ii,iv,v,vii,x,xi} {ii,iv,v,viii,ix,x} {ii,iv,v,viii,ix,xi} {ii,iv,v,viii,x,xi} {ii,iv,v,ix,x,xi} {ii,iv,vi,vii,viii,ix} {ii,iv,vi,vii,viii,x} {ii,iv,vi,vii,viii,xi} {ii,iv,vi,vii,ix,x} {ii,iv,vi,vii,ix,xi} {ii,iv,vi,vii,x,xi} {ii,iv,vi,viii,ix,x} {ii,iv,vi,viii,ix,xi} {ii,iv,vi,viii,x,xi} {ii,iv,vi,ix,x,xi} {ii,iv,vii,viii,ix,x} {ii,iv,vii,viii,ix,xi} {ii,iv,vii,viii,x,xi} {ii,iv,vii,ix,x,xi} {ii,iv,viii,ix,x,xi} {ii,v,vi,vii,viii,ix} {ii,v,vi,vii,viii,x} {ii,v,vi,vii,viii,xi} {ii,v,vi,vii,ix,x} {ii,v,vi,vii,ix,xi} {ii,v,vi,vii,x,xi} {ii,v,vi,viii,ix,x} {ii,v,vi,viii,ix,xi} {ii,v,vi,viii,x,xi} {ii,v,vi,ix,x,xi} {ii,v,vii,viii,ix,x} {ii,v,vii,viii,ix,xi} {ii,v,vii,viii,x,xi} {ii,v,vii,ix,x,xi} {ii,v,viii,ix,x,xi} {ii,vi,vii,viii,ix,x} {ii,vi,vii,viii,ix,xi} {ii,vi,vii,viii,x,xi} {ii,vi,vii,ix,x,xi} {ii,vi,viii,ix,x,xi} {ii,vii,viii,ix,x,xi} {iii,iv,v,vi,vii,viii} {iii,iv,v,vi,vii,ix} {iii,iv,v,vi,vii,x} {iii,iv,v,vi,vii,xi} {iii,iv,v,vi,viii,ix} {iii,iv,v,vi,viii,x} {iii,iv,v,vi,viii,xi} {iii,iv,v,vi,ix,x} {iii,iv,v,vi,ix,xi} {iii,iv,v,vi,x,xi} {iii,iv,v,vii,viii,ix} {iii,iv,v,vii,viii,x} {iii,iv,v,vii,viii,xi} {iii,iv,v,vii,ix,x} {iii,iv,v,vii,ix,xi} {iii,iv,v,vii,x,xi} {iii,iv,v,viii,ix,x} {iii,iv,v,viii,ix,xi} {iii,iv,v,viii,x,xi} {iii,iv,v,ix,x,xi} {iii,iv,vi,vii,viii,ix} {iii,iv,vi,vii,viii,x} {iii,iv,vi,vii,viii,xi} {iii,iv,vi,vii,ix,x} {iii,iv,vi,vii,ix,xi} {iii,iv,vi,vii,x,xi} {iii,iv,vi,viii,ix,x} {iii,iv,vi,viii,ix,xi} {iii,iv,vi,viii,x,xi} {iii,iv,vi,ix,x,xi} {iii,iv,vii,viii,ix,x} {iii,iv,vii,viii,ix,xi} {iii,iv,vii,viii,x,xi} {iii,iv,vii,ix,x,xi} {iii,iv,viii,ix,x,xi} {iii,v,vi,vii,viii,ix} {iii,v,vi,vii,viii,x} {iii,v,vi,vii,viii,xi} {iii,v,vi,vii,ix,x} {iii,v,vi,vii,ix,xi} {iii,v,vi,vii,x,xi} {iii,v,vi,viii,ix,x} {iii,v,vi,viii,ix,xi} {iii,v,vi,viii,x,xi} {iii,v,vi,ix,x,xi} {iii,v,vii,viii,ix,x} {iii,v,vii,viii,ix,xi} {iii,v,vii,viii,x,xi} {iii,v,vii,ix,x,xi} {iii,v,viii,ix,x,xi} {iii,vi,vii,viii,ix,x} {iii,vi,vii,viii,ix,xi} {iii,vi,vii,viii,x,xi} {iii,vi,vii,ix,x,xi} {iii,vi,viii,ix,x,xi} {iii,vii,viii,ix,x,xi} {iv,v,vi,vii,viii,ix} {iv,v,vi,vii,viii,x} {iv,v,vi,vii,viii,xi} {iv,v,vi,vii,ix,x} {iv,v,vi,vii,ix,xi} {iv,v,vi,vii,x,xi} {iv,v,vi,viii,ix,x} {iv,v,vi,viii,ix,xi} {iv,v,vi,viii,x,xi} {iv,v,vi,ix,x,xi} {iv,v,vii,viii,ix,x} {iv,v,vii,viii,ix,xi} {iv,v,vii,viii,x,xi} {iv,v,vii,ix,x,xi} {iv,v,viii,ix,x,xi} {iv,vi,vii,viii,ix,x} {iv,vi,vii,viii,ix,xi} {iv,vi,vii,viii,x,xi} {iv,vi,vii,ix,x,xi} {iv,vi,viii,ix,x,xi} {iv,vii,viii,ix,x,xi} {v,vi,vii,viii,ix,x} {v,vi,vii,viii,ix,xi} {v,vi,vii,viii,x,xi} {v,vi,vii,ix,x,xi} {v,vi,viii,ix,x,xi} {v,vii,viii,ix,x,xi} {vi,vii,viii,ix,x,xi} {i,ii,iii,iv,v,vi,vii} {i,ii,iii,iv,v,vi,viii} {i,ii,iii,iv,v,vi,ix} {i,ii,iii,iv,v,vi,x} {i,ii,iii,iv,v,vi,xi} {i,ii,iii,iv,v,vii,viii} {i,ii,iii,iv,v,vii,ix} {i,ii,iii,iv,v,vii,x} {i,ii,iii,iv,v,vii,xi} {i,ii,iii,iv,v,viii,ix} {i,ii,iii,iv,v,viii,x} {i,ii,iii,iv,v,viii,xi} {i,ii,iii,iv,v,ix,x} {i,ii,iii,iv,v,ix,xi} {i,ii,iii,iv,v,x,xi} {i,ii,iii,iv,vi,vii,viii} {i,ii,iii,iv,vi,vii,ix} {i,ii,iii,iv,vi,vii,x} {i,ii,iii,iv,vi,vii,xi} {i,ii,iii,iv,vi,viii,ix} {i,ii,iii,iv,vi,viii,x} {i,ii,iii,iv,vi,viii,xi} {i,ii,iii,iv,vi,ix,x} {i,ii,iii,iv,vi,ix,xi} {i,ii,iii,iv,vi,x,xi} {i,ii,iii,iv,vii,viii,ix} {i,ii,iii,iv,vii,viii,x} {i,ii,iii,iv,vii,viii,xi} {i,ii,iii,iv,vii,ix,x} {i,ii,iii,iv,vii,ix,xi} {i,ii,iii,iv,vii,x,xi} {i,ii,iii,iv,viii,ix,x} {i,ii,iii,iv,viii,ix,xi} {i,ii,iii,iv,viii,x,xi} {i,ii,iii,iv,ix,x,xi} {i,ii,iii,v,vi,vii,viii} {i,ii,iii,v,vi,vii,ix} {i,ii,iii,v,vi,vii,x} {i,ii,iii,v,vi,vii,xi} {i,ii,iii,v,vi,viii,ix} {i,ii,iii,v,vi,viii,x} {i,ii,iii,v,vi,viii,xi} {i,ii,iii,v,vi,ix,x} {i,ii,iii,v,vi,ix,xi} {i,ii,iii,v,vi,x,xi} {i,ii,iii,v,vii,viii,ix} {i,ii,iii,v,vii,viii,x} {i,ii,iii,v,vii,viii,xi} {i,ii,iii,v,vii,ix,x} {i,ii,iii,v,vii,ix,xi} {i,ii,iii,v,vii,x,xi} {i,ii,iii,v,viii,ix,x} {i,ii,iii,v,viii,ix,xi} {i,ii,iii,v,viii,x,xi} {i,ii,iii,v,ix,x,xi} {i,ii,iii,vi,vii,viii,ix} {i,ii,iii,vi,vii,viii,x} {i,ii,iii,vi,vii,viii,xi} {i,ii,iii,vi,vii,ix,x} {i,ii,iii,vi,vii,ix,xi} {i,ii,iii,vi,vii,x,xi} {i,ii,iii,vi,viii,ix,x} {i,ii,iii,vi,viii,ix,xi} {i,ii,iii,vi,viii,x,xi} {i,ii,iii,vi,ix,x,xi} {i,ii,iii,vii,viii,ix,x} {i,ii,iii,vii,viii,ix,xi} {i,ii,iii,vii,viii,x,xi} {i,ii,iii,vii,ix,x,xi} {i,ii,iii,viii,ix,x,xi} {i,ii,iv,v,vi,vii,viii} {i,ii,iv,v,vi,vii,ix} {i,ii,iv,v,vi,vii,x} {i,ii,iv,v,vi,vii,xi} {i,ii,iv,v,vi,viii,ix} {i,ii,iv,v,vi,viii,x} {i,ii,iv,v,vi,viii,xi} {i,ii,iv,v,vi,ix,x} {i,ii,iv,v,vi,ix,xi} {i,ii,iv,v,vi,x,xi} {i,ii,iv,v,vii,viii,ix} {i,ii,iv,v,vii,viii,x} {i,ii,iv,v,vii,viii,xi} {i,ii,iv,v,vii,ix,x} {i,ii,iv,v,vii,ix,xi} {i,ii,iv,v,vii,x,xi} {i,ii,iv,v,viii,ix,x} {i,ii,iv,v,viii,ix,xi} {i,ii,iv,v,viii,x,xi} {i,ii,iv,v,ix,x,xi} {i,ii,iv,vi,vii,viii,ix} {i,ii,iv,vi,vii,viii,x} {i,ii,iv,vi,vii,viii,xi} {i,ii,iv,vi,vii,ix,x} {i,ii,iv,vi,vii,ix,xi} {i,ii,iv,vi,vii,x,xi} {i,ii,iv,vi,viii,ix,x} {i,ii,iv,vi,viii,ix,xi} {i,ii,iv,vi,viii,x,xi} {i,ii,iv,vi,ix,x,xi} {i,ii,iv,vii,viii,ix,x} {i,ii,iv,vii,viii,ix,xi} {i,ii,iv,vii,viii,x,xi} {i,ii,iv,vii,ix,x,xi} {i,ii,iv,viii,ix,x,xi} {i,ii,v,vi,vii,viii,ix} {i,ii,v,vi,vii,viii,x} {i,ii,v,vi,vii,viii,xi} {i,ii,v,vi,vii,ix,x} {i,ii,v,vi,vii,ix,xi} {i,ii,v,vi,vii,x,xi} {i,ii,v,vi,viii,ix,x} {i,ii,v,vi,viii,ix,xi} {i,ii,v,vi,viii,x,xi} {i,ii,v,vi,ix,x,xi} {i,ii,v,vii,viii,ix,x} {i,ii,v,vii,viii,ix,xi} {i,ii,v,vii,viii,x,xi} {i,ii,v,vii,ix,x,xi} {i,ii,v,viii,ix,x,xi} {i,ii,vi,vii,viii,ix,x} {i,ii,vi,vii,viii,ix,xi} {i,ii,vi,vii,viii,x,xi} {i,ii,vi,vii,ix,x,xi} {i,ii,vi,viii,ix,x,xi} {i,ii,vii,viii,ix,x,xi} {i,iii,iv,v,vi,vii,viii} {i,iii,iv,v,vi,vii,ix} {i,iii,iv,v,vi,vii,x} {i,iii,iv,v,vi,vii,xi} {i,iii,iv,v,vi,viii,ix} {i,iii,iv,v,vi,viii,x} {i,iii,iv,v,vi,viii,xi} {i,iii,iv,v,vi,ix,x} {i,iii,iv,v,vi,ix,xi} {i,iii,iv,v,vi,x,xi} {i,iii,iv,v,vii,viii,ix} {i,iii,iv,v,vii,viii,x} {i,iii,iv,v,vii,viii,xi} {i,iii,iv,v,vii,ix,x} {i,iii,iv,v,vii,ix,xi} {i,iii,iv,v,vii,x,xi} {i,iii,iv,v,viii,ix,x} {i,iii,iv,v,viii,ix,xi} {i,iii,iv,v,viii,x,xi} {i,iii,iv,v,ix,x,xi} {i,iii,iv,vi,vii,viii,ix} {i,iii,iv,vi,vii,viii,x} {i,iii,iv,vi,vii,viii,xi} {i,iii,iv,vi,vii,ix,x} {i,iii,iv,vi,vii,ix,xi} {i,iii,iv,vi,vii,x,xi} {i,iii,iv,vi,viii,ix,x} {i,iii,iv,vi,viii,ix,xi} {i,iii,iv,vi,viii,x,xi} {i,iii,iv,vi,ix,x,xi} {i,iii,iv,vii,viii,ix,x} {i,iii,iv,vii,viii,ix,xi} {i,iii,iv,vii,viii,x,xi} {i,iii,iv,vii,ix,x,xi} {i,iii,iv,viii,ix,x,xi} {i,iii,v,vi,vii,viii,ix} {i,iii,v,vi,vii,viii,x} {i,iii,v,vi,vii,viii,xi} {i,iii,v,vi,vii,ix,x} {i,iii,v,vi,vii,ix,xi} {i,iii,v,vi,vii,x,xi} {i,iii,v,vi,viii,ix,x} {i,iii,v,vi,viii,ix,xi} {i,iii,v,vi,viii,x,xi} {i,iii,v,vi,ix,x,xi} {i,iii,v,vii,viii,ix,x} {i,iii,v,vii,viii,ix,xi} {i,iii,v,vii,viii,x,xi} {i,iii,v,vii,ix,x,xi} {i,iii,v,viii,ix,x,xi} {i,iii,vi,vii,viii,ix,x} {i,iii,vi,vii,viii,ix,xi} {i,iii,vi,vii,viii,x,xi} {i,iii,vi,vii,ix,x,xi} {i,iii,vi,viii,ix,x,xi} {i,iii,vii,viii,ix,x,xi} {i,iv,v,vi,vii,viii,ix} {i,iv,v,vi,vii,viii,x} {i,iv,v,vi,vii,viii,xi} {i,iv,v,vi,vii,ix,x} {i,iv,v,vi,vii,ix,xi} {i,iv,v,vi,vii,x,xi} {i,iv,v,vi,viii,ix,x} {i,iv,v,vi,viii,ix,xi} {i,iv,v,vi,viii,x,xi} {i,iv,v,vi,ix,x,xi} {i,iv,v,vii,viii,ix,x} {i,iv,v,vii,viii,ix,xi} {i,iv,v,vii,viii,x,xi} {i,iv,v,vii,ix,x,xi} {i,iv,v,viii,ix,x,xi} {i,iv,vi,vii,viii,ix,x} {i,iv,vi,vii,viii,ix,xi} {i,iv,vi,vii,viii,x,xi} {i,iv,vi,vii,ix,x,xi} {i,iv,vi,viii,ix,x,xi} {i,iv,vii,viii,ix,x,xi} {i,v,vi,vii,viii,ix,x} {i,v,vi,vii,viii,ix,xi} {i,v,vi,vii,viii,x,xi} {i,v,vi,vii,ix,x,xi} {i,v,vi,viii,ix,x,xi} {i,v,vii,viii,ix,x,xi} {i,vi,vii,viii,ix,x,xi} {ii,iii,iv,v,vi,vii,viii} {ii,iii,iv,v,vi,vii,ix} {ii,iii,iv,v,vi,vii,x} {ii,iii,iv,v,vi,vii,xi} {ii,iii,iv,v,vi,viii,ix} {ii,iii,iv,v,vi,viii,x} {ii,iii,iv,v,vi,viii,xi} {ii,iii,iv,v,vi,ix,x} {ii,iii,iv,v,vi,ix,xi} {ii,iii,iv,v,vi,x,xi} {ii,iii,iv,v,vii,viii,ix} {ii,iii,iv,v,vii,viii,x} {ii,iii,iv,v,vii,viii,xi} {ii,iii,iv,v,vii,ix,x} {ii,iii,iv,v,vii,ix,xi} {ii,iii,iv,v,vii,x,xi} {ii,iii,iv,v,viii,ix,x} {ii,iii,iv,v,viii,ix,xi} {ii,iii,iv,v,viii,x,xi} {ii,iii,iv,v,ix,x,xi} {ii,iii,iv,vi,vii,viii,ix} {ii,iii,iv,vi,vii,viii,x} {ii,iii,iv,vi,vii,viii,xi} {ii,iii,iv,vi,vii,ix,x} {ii,iii,iv,vi,vii,ix,xi} {ii,iii,iv,vi,vii,x,xi} {ii,iii,iv,vi,viii,ix,x} {ii,iii,iv,vi,viii,ix,xi} {ii,iii,iv,vi,viii,x,xi} {ii,iii,iv,vi,ix,x,xi} {ii,iii,iv,vii,viii,ix,x} {ii,iii,iv,vii,viii,ix,xi} {ii,iii,iv,vii,viii,x,xi} {ii,iii,iv,vii,ix,x,xi} {ii,iii,iv,viii,ix,x,xi} {ii,iii,v,vi,vii,viii,ix} {ii,iii,v,vi,vii,viii,x} {ii,iii,v,vi,vii,viii,xi} {ii,iii,v,vi,vii,ix,x} {ii,iii,v,vi,vii,ix,xi} {ii,iii,v,vi,vii,x,xi} {ii,iii,v,vi,viii,ix,x} {ii,iii,v,vi,viii,ix,xi} {ii,iii,v,vi,viii,x,xi} {ii,iii,v,vi,ix,x,xi} {ii,iii,v,vii,viii,ix,x} {ii,iii,v,vii,viii,ix,xi} {ii,iii,v,vii,viii,x,xi} {ii,iii,v,vii,ix,x,xi} {ii,iii,v,viii,ix,x,xi} {ii,iii,vi,vii,viii,ix,x} {ii,iii,vi,vii,viii,ix,xi} {ii,iii,vi,vii,viii,x,xi} {ii,iii,vi,vii,ix,x,xi} {ii,iii,vi,viii,ix,x,xi} {ii,iii,vii,viii,ix,x,xi} {ii,iv,v,vi,vii,viii,ix} {ii,iv,v,vi,vii,viii,x} {ii,iv,v,vi,vii,viii,xi} {ii,iv,v,vi,vii,ix,x} {ii,iv,v,vi,vii,ix,xi} {ii,iv,v,vi,vii,x,xi} {ii,iv,v,vi,viii,ix,x} {ii,iv,v,vi,viii,ix,xi} {ii,iv,v,vi,viii,x,xi} {ii,iv,v,vi,ix,x,xi} {ii,iv,v,vii,viii,ix,x} {ii,iv,v,vii,viii,ix,xi} {ii,iv,v,vii,viii,x,xi} {ii,iv,v,vii,ix,x,xi} {ii,iv,v,viii,ix,x,xi} {ii,iv,vi,vii,viii,ix,x} {ii,iv,vi,vii,viii,ix,xi} {ii,iv,vi,vii,viii,x,xi} {ii,iv,vi,vii,ix,x,xi} {ii,iv,vi,viii,ix,x,xi} {ii,iv,vii,viii,ix,x,xi} {ii,v,vi,vii,viii,ix,x} {ii,v,vi,vii,viii,ix,xi} {ii,v,vi,vii,viii,x,xi} {ii,v,vi,vii,ix,x,xi} {ii,v,vi,viii,ix,x,xi} {ii,v,vii,viii,ix,x,xi} {ii,vi,vii,viii,ix,x,xi} {iii,iv,v,vi,vii,viii,ix} {iii,iv,v,vi,vii,viii,x} {iii,iv,v,vi,vii,viii,xi} {iii,iv,v,vi,vii,ix,x} {iii,iv,v,vi,vii,ix,xi} {iii,iv,v,vi,vii,x,xi} {iii,iv,v,vi,viii,ix,x} {iii,iv,v,vi,viii,ix,xi} {iii,iv,v,vi,viii,x,xi} {iii,iv,v,vi,ix,x,xi} {iii,iv,v,vii,viii,ix,x} {iii,iv,v,vii,viii,ix,xi} {iii,iv,v,vii,viii,x,xi} {iii,iv,v,vii,ix,x,xi} {iii,iv,v,viii,ix,x,xi} {iii,iv,vi,vii,viii,ix,x} {iii,iv,vi,vii,viii,ix,xi} {iii,iv,vi,vii,viii,x,xi} {iii,iv,vi,vii,ix,x,xi} {iii,iv,vi,viii,ix,x,xi} {iii,iv,vii,viii,ix,x,xi} {iii,v,vi,vii,viii,ix,x} {iii,v,vi,vii,viii,ix,xi} {iii,v,vi,vii,viii,x,xi} {iii,v,vi,vii,ix,x,xi} {iii,v,vi,viii,ix,x,xi} {iii,v,vii,viii,ix,x,xi} {iii,vi,vii,viii,ix,x,xi} {iv,v,vi,vii,viii,ix,x} {iv,v,vi,vii,viii,ix,xi} {iv,v,vi,vii,viii,x,xi} {iv,v,vi,vii,ix,x,xi} {iv,v,vi,viii,ix,x,xi} {iv,v,vii,viii,ix,x,xi} {iv,vi,vii,viii,ix,x,xi} {v,vi,vii,viii,ix,x,xi} {i,ii,iii,iv,v,vi,vii,viii} {i,ii,iii,iv,v,vi,vii,ix} {i,ii,iii,iv,v,vi,vii,x} {i,ii,iii,iv,v,vi,vii,xi} {i,ii,iii,iv,v,vi,viii,ix} {i,ii,iii,iv,v,vi,viii,x} {i,ii,iii,iv,v,vi,viii,xi} {i,ii,iii,iv,v,vi,ix,x} {i,ii,iii,iv,v,vi,ix,xi} {i,ii,iii,iv,v,vi,x,xi} {i,ii,iii,iv,v,vii,viii,ix} {i,ii,iii,iv,v,vii,viii,x} {i,ii,iii,iv,v,vii,viii,xi} {i,ii,iii,iv,v,vii,ix,x} {i,ii,iii,iv,v,vii,ix,xi} {i,ii,iii,iv,v,vii,x,xi} {i,ii,iii,iv,v,viii,ix,x} {i,ii,iii,iv,v,viii,ix,xi} {i,ii,iii,iv,v,viii,x,xi} {i,ii,iii,iv,v,ix,x,xi} {i,ii,iii,iv,vi,vii,viii,ix} {i,ii,iii,iv,vi,vii,viii,x} {i,ii,iii,iv,vi,vii,viii,xi} {i,ii,iii,iv,vi,vii,ix,x} {i,ii,iii,iv,vi,vii,ix,xi} {i,ii,iii,iv,vi,vii,x,xi} {i,ii,iii,iv,vi,viii,ix,x} {i,ii,iii,iv,vi,viii,ix,xi} {i,ii,iii,iv,vi,viii,x,xi} {i,ii,iii,iv,vi,ix,x,xi} {i,ii,iii,iv,vii,viii,ix,x} {i,ii,iii,iv,vii,viii,ix,xi} {i,ii,iii,iv,vii,viii,x,xi} {i,ii,iii,iv,vii,ix,x,xi} {i,ii,iii,iv,viii,ix,x,xi} {i,ii,iii,v,vi,vii,viii,ix} {i,ii,iii,v,vi,vii,viii,x} {i,ii,iii,v,vi,vii,viii,xi} {i,ii,iii,v,vi,vii,ix,x} {i,ii,iii,v,vi,vii,ix,xi} {i,ii,iii,v,vi,vii,x,xi} {i,ii,iii,v,vi,viii,ix,x} {i,ii,iii,v,vi,viii,ix,xi} {i,ii,iii,v,vi,viii,x,xi} {i,ii,iii,v,vi,ix,x,xi} {i,ii,iii,v,vii,viii,ix,x} {i,ii,iii,v,vii,viii,ix,xi} {i,ii,iii,v,vii,viii,x,xi} {i,ii,iii,v,vii,ix,x,xi} {i,ii,iii,v,viii,ix,x,xi} {i,ii,iii,vi,vii,viii,ix,x} {i,ii,iii,vi,vii,viii,ix,xi} {i,ii,iii,vi,vii,viii,x,xi} {i,ii,iii,vi,vii,ix,x,xi} {i,ii,iii,vi,viii,ix,x,xi} {i,ii,iii,vii,viii,ix,x,xi} {i,ii,iv,v,vi,vii,viii,ix} {i,ii,iv,v,vi,vii,viii,x} {i,ii,iv,v,vi,vii,viii,xi} {i,ii,iv,v,vi,vii,ix,x} {i,ii,iv,v,vi,vii,ix,xi} {i,ii,iv,v,vi,vii,x,xi} {i,ii,iv,v,vi,viii,ix,x} {i,ii,iv,v,vi,viii,ix,xi} {i,ii,iv,v,vi,viii,x,xi} {i,ii,iv,v,vi,ix,x,xi} {i,ii,iv,v,vii,viii,ix,x} {i,ii,iv,v,vii,viii,ix,xi} {i,ii,iv,v,vii,viii,x,xi} {i,ii,iv,v,vii,ix,x,xi} {i,ii,iv,v,viii,ix,x,xi} {i,ii,iv,vi,vii,viii,ix,x} {i,ii,iv,vi,vii,viii,ix,xi} {i,ii,iv,vi,vii,viii,x,xi} {i,ii,iv,vi,vii,ix,x,xi} {i,ii,iv,vi,viii,ix,x,xi} {i,ii,iv,vii,viii,ix,x,xi} {i,ii,v,vi,vii,viii,ix,x} {i,ii,v,vi,vii,viii,ix,xi} {i,ii,v,vi,vii,viii,x,xi} {i,ii,v,vi,vii,ix,x,xi} {i,ii,v,vi,viii,ix,x,xi} {i,ii,v,vii,viii,ix,x,xi} {i,ii,vi,vii,viii,ix,x,xi} {i,iii,iv,v,vi,vii,viii,ix} {i,iii,iv,v,vi,vii,viii,x} {i,iii,iv,v,vi,vii,viii,xi} {i,iii,iv,v,vi,vii,ix,x} {i,iii,iv,v,vi,vii,ix,xi} {i,iii,iv,v,vi,vii,x,xi} {i,iii,iv,v,vi,viii,ix,x} {i,iii,iv,v,vi,viii,ix,xi} {i,iii,iv,v,vi,viii,x,xi} {i,iii,iv,v,vi,ix,x,xi} {i,iii,iv,v,vii,viii,ix,x} {i,iii,iv,v,vii,viii,ix,xi} {i,iii,iv,v,vii,viii,x,xi} {i,iii,iv,v,vii,ix,x,xi} {i,iii,iv,v,viii,ix,x,xi} {i,iii,iv,vi,vii,viii,ix,x} {i,iii,iv,vi,vii,viii,ix,xi} {i,iii,iv,vi,vii,viii,x,xi} {i,iii,iv,vi,vii,ix,x,xi} {i,iii,iv,vi,viii,ix,x,xi} {i,iii,iv,vii,viii,ix,x,xi} {i,iii,v,vi,vii,viii,ix,x} {i,iii,v,vi,vii,viii,ix,xi} {i,iii,v,vi,vii,viii,x,xi} {i,iii,v,vi,vii,ix,x,xi} {i,iii,v,vi,viii,ix,x,xi} {i,iii,v,vii,viii,ix,x,xi} {i,iii,vi,vii,viii,ix,x,xi} {i,iv,v,vi,vii,viii,ix,x} {i,iv,v,vi,vii,viii,ix,xi} {i,iv,v,vi,vii,viii,x,xi} {i,iv,v,vi,vii,ix,x,xi} {i,iv,v,vi,viii,ix,x,xi} {i,iv,v,vii,viii,ix,x,xi} {i,iv,vi,vii,viii,ix,x,xi} {i,v,vi,vii,viii,ix,x,xi} {ii,iii,iv,v,vi,vii,viii,ix} {ii,iii,iv,v,vi,vii,viii,x} {ii,iii,iv,v,vi,vii,viii,xi} {ii,iii,iv,v,vi,vii,ix,x} {ii,iii,iv,v,vi,vii,ix,xi} {ii,iii,iv,v,vi,vii,x,xi} {ii,iii,iv,v,vi,viii,ix,x} {ii,iii,iv,v,vi,viii,ix,xi} {ii,iii,iv,v,vi,viii,x,xi} {ii,iii,iv,v,vi,ix,x,xi} {ii,iii,iv,v,vii,viii,ix,x} {ii,iii,iv,v,vii,viii,ix,xi} {ii,iii,iv,v,vii,viii,x,xi} {ii,iii,iv,v,vii,ix,x,xi} {ii,iii,iv,v,viii,ix,x,xi} {ii,iii,iv,vi,vii,viii,ix,x} {ii,iii,iv,vi,vii,viii,ix,xi} {ii,iii,iv,vi,vii,viii,x,xi} {ii,iii,iv,vi,vii,ix,x,xi} {ii,iii,iv,vi,viii,ix,x,xi} {ii,iii,iv,vii,viii,ix,x,xi} {ii,iii,v,vi,vii,viii,ix,x} {ii,iii,v,vi,vii,viii,ix,xi} {ii,iii,v,vi,vii,viii,x,xi} {ii,iii,v,vi,vii,ix,x,xi} {ii,iii,v,vi,viii,ix,x,xi} {ii,iii,v,vii,viii,ix,x,xi} {ii,iii,vi,vii,viii,ix,x,xi} {ii,iv,v,vi,vii,viii,ix,x} {ii,iv,v,vi,vii,viii,ix,xi} {ii,iv,v,vi,vii,viii,x,xi} {ii,iv,v,vi,vii,ix,x,xi} {ii,iv,v,vi,viii,ix,x,xi} {ii,iv,v,vii,viii,ix,x,xi} {ii,iv,vi,vii,viii,ix,x,xi} {ii,v,vi,vii,viii,ix,x,xi} {iii,iv,v,vi,vii,viii,ix,x} {iii,iv,v,vi,vii,viii,ix,xi} {iii,iv,v,vi,vii,viii,x,xi} {iii,iv,v,vi,vii,ix,x,xi} {iii,iv,v,vi,viii,ix,x,xi} {iii,iv,v,vii,viii,ix,x,xi} {iii,iv,vi,vii,viii,ix,x,xi} {iii,v,vi,vii,viii,ix,x,xi} {iv,v,vi,vii,viii,ix,x,xi} {i,ii,iii,iv,v,vi,vii,viii,ix} {i,ii,iii,iv,v,vi,vii,viii,x} {i,ii,iii,iv,v,vi,vii,viii,xi} {i,ii,iii,iv,v,vi,vii,ix,x} {i,ii,iii,iv,v,vi,vii,ix,xi} {i,ii,iii,iv,v,vi,vii,x,xi} {i,ii,iii,iv,v,vi,viii,ix,x} {i,ii,iii,iv,v,vi,viii,ix,xi} {i,ii,iii,iv,v,vi,viii,x,xi} {i,ii,iii,iv,v,vi,ix,x,xi} {i,ii,iii,iv,v,vii,viii,ix,x} {i,ii,iii,iv,v,vii,viii,ix,xi} {i,ii,iii,iv,v,vii,viii,x,xi} {i,ii,iii,iv,v,vii,ix,x,xi} {i,ii,iii,iv,v,viii,ix,x,xi} {i,ii,iii,iv,vi,vii,viii,ix,x} {i,ii,iii,iv,vi,vii,viii,ix,xi} {i,ii,iii,iv,vi,vii,viii,x,xi} {i,ii,iii,iv,vi,vii,ix,x,xi} {i,ii,iii,iv,vi,viii,ix,x,xi} {i,ii,iii,iv,vii,viii,ix,x,xi} {i,ii,iii,v,vi,vii,viii,ix,x} {i,ii,iii,v,vi,vii,viii,ix,xi} {i,ii,iii,v,vi,vii,viii,x,xi} {i,ii,iii,v,vi,vii,ix,x,xi} {i,ii,iii,v,vi,viii,ix,x,xi} {i,ii,iii,v,vii,viii,ix,x,xi} {i,ii,iii,vi,vii,viii,ix,x,xi} {i,ii,iv,v,vi,vii,viii,ix,x} {i,ii,iv,v,vi,vii,viii,ix,xi} {i,ii,iv,v,vi,vii,viii,x,xi} {i,ii,iv,v,vi,vii,ix,x,xi} {i,ii,iv,v,vi,viii,ix,x,xi} {i,ii,iv,v,vii,viii,ix,x,xi} {i,ii,iv,vi,vii,viii,ix,x,xi} {i,ii,v,vi,vii,viii,ix,x,xi} {i,iii,iv,v,vi,vii,viii,ix,x} {i,iii,iv,v,vi,vii,viii,ix,xi} {i,iii,iv,v,vi,vii,viii,x,xi} {i,iii,iv,v,vi,vii,ix,x,xi} {i,iii,iv,v,vi,viii,ix,x,xi} {i,iii,iv,v,vii,viii,ix,x,xi} {i,iii,iv,vi,vii,viii,ix,x,xi} {i,iii,v,vi,vii,viii,ix,x,xi} {i,iv,v,vi,vii,viii,ix,x,xi} {ii,iii,iv,v,vi,vii,viii,ix,x} {ii,iii,iv,v,vi,vii,viii,ix,xi} {ii,iii,iv,v,vi,vii,viii,x,xi} {ii,iii,iv,v,vi,vii,ix,x,xi} {ii,iii,iv,v,vi,viii,ix,x,xi} {ii,iii,iv,v,vii,viii,ix,x,xi} {ii,iii,iv,vi,vii,viii,ix,x,xi} {ii,iii,v,vi,vii,viii,ix,x,xi} {ii,iv,v,vi,vii,viii,ix,x,xi} {iii,iv,v,vi,vii,viii,ix,x,xi} {i,ii,iii,iv,v,vi,vii,viii,ix,x} {i,ii,iii,iv,v,vi,vii,viii,ix,xi} {i,ii,iii,iv,v,vi,vii,viii,x,xi} {i,ii,iii,iv,v,vi,vii,ix,x,xi} {i,ii,iii,iv,v,vi,viii,ix,x,xi} {i,ii,iii,iv,v,vii,viii,ix,x,xi} {i,ii,iii,iv,vi,vii,viii,ix,x,xi} {i,ii,iii,v,vi,vii,viii,ix,x,xi} {i,ii,iv,v,vi,vii,viii,ix,x,xi} {i,iii,iv,v,vi,vii,viii,ix,x,xi} {ii,iii,iv,v,vi,vii,viii,ix,x,xi} 또는 {i,ii,iii,iv,v,vi,vii viii,ix,x,xi}.
변이체가 (i) 및 (iii) 내지 (xi) 중 임의의 하나를 포함하면, 이는 Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 추가로 포함할 수 있다.
(i)에서, 변이체는 임의의 개수 및 조합의 N40, D43, E44, S54, S57, Q62, R97, E101, E124, E131, R142, T150 및 R192에서의 돌연변이를 포함할 수 있다. (i)에서, 변이체는 바람직하게는 하기 위치 N40, D43, E44, S54, S57, Q62, E101, E131 및 T150에서의 하나 이상의 돌연변이 (즉, 상기 위치 중 하나 이상에서의 돌연변이)를 포함한다. (i)에서, 변이체는 바람직하게는 하기 위치 N40, D43, E44, E101 및 E131에서의 하나 이상의 돌연변이 (즉, 상기 위치 중 하나 이상에서의 돌연변이)를 포함한다. (i)에서, 변이체는 바람직하게는 S54 및/또는 S57에서의 돌연변이를 포함한다. (i)에서, 변이체는 더욱 바람직하게는 (a) S54 및/또는 S57 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다. S54 및/또는 S57이 (xi)에서 결실되면, 이는 (i)에서 돌연변이될 수 없고, 반대도 마찬가지이다. (i)에서, 변이체는 바람직하게는 T150에서의 돌연변이, 예컨대 T150I를 포함한다. 대안적으로, 변이체는 바람직하게는 (a) T150 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다. (i)에서, 변이체는 바람직하게는 Q62에서의 돌연변이, 예컨대 Q62R 또는 Q62K를 포함한다. 대안적으로, 변이체는 바람직하게는 (a) Q62 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다. 변이체는 D43, E44, Q62 또는 이의 임의의 조합, 예컨대 D43, E44, Q62, D43/E44, D43/Q62, E44/Q62 또는 D43/E44/Q62에서의 돌연변이를 포함할 수 있다. 대안적으로, 변이체는 바람직하게는 (a) D43, E44, Q62, D43/E44, D43/Q62, E44/Q62 또는 D43/E44/Q62 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다.
(ii) 및 본 출원의 다른 곳에서, / 기호는 "및"을 의미하여, Y51/N55는 Y51및 N55이다. (ii)에서, 변이체는 바람직하게는 Y51/N55에서의 돌연변이를 포함한다. CsgG 내의 협착부가 잔기 Y51, N55 및 F56의 측쇄에 의해 형성된 3개의 스태킹된 동심성 고리로 구성된다는 것이 제안되었다 (Goyal et al., 2014, Nature, 516, 250-253). 따라서 (ii)에서의 이러한 잔기의 돌연변이는 폴리뉴클레오티드가 포어를 통해 이동할 때 전류에 기여하는 뉴클레오티드의 개수를 감소시킬 수 있고, 이에 의해, 관찰된 전류 (폴리뉴클레오티드가 포어를 통해 이동할 때)와 폴리뉴클레오티드 사이의 직접적인 관계를 확인하는 것을 더 용이하게 할 수 있다. 본 발명의 방법에서 유용한 변이체 및 포어와 관련하여 하기에서 논의된 방식 중 임의의 것으로 Y56이 돌연변이될 수 있다.
(v)에서, 변이체는 N102R, N102F, N102Y 또는 N102W를 포함할 수 있다. 변이체는 바람직하게는 (a) N102R, N102F, N102Y 또는 N102W 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다.
(xi)에서, 임의의 개수 및 조합의 K49, P50, Y51, P52, A53, S54, N55, F56 및 S57이 결실될 수 있다. 바람직하게는, K49, P50, Y51, P52, A53, S54, N55 및 S57 중 하나 이상이 결실될 수 있다. Y51, N55 및 F56 중 임의의 것이 (xi)에서 결실되면, 이는 (ii)에서 돌연변이될 수 없고, 반대도 마찬가지이다.
(i)에서, 변이체는 바람직하게는 하기 치환 N40R, N40K, D43N, D43Q, D43R, D43K, E44N, E44Q, E44R, E44K, S54P, S57P, Q62R, Q62K, R97N, R97G, R97L, E101N, E101Q, E101R, E101K, E101F, E101Y, E101W, E124N, E124Q, E124R, E124K, E124F, E124Y, E124W, E131D, R142E, R142N, T150I, R192E 및 R192N 중 하나 이상, 예컨대 N40R, N40K, D43N, D43Q, D43R, D43K, E44N, E44Q, E44R, E44K, S54P, S57P, Q62R, Q62K, E101N, E101Q, E101R, E101K, E101F, E101Y, E101W, E131D 및 T150I 중 하나 이상, 또는 N40R, N40K, D43N, D43Q, D43R, D43K, E44N, E44Q, E44R, E44K, E101N, E101Q, E101R, E101K, E101F, E101Y, E101W 및 E131D 중 하나 이상을 포함한다. 변이체는 임의의 개수 및 조합의 이러한 치환을 포함할 수 있다. (i)에서, 변이체는 바람직하게는 S54P 및/또는 S57P를 포함한다. (i)에서, 변이체는 바람직하게는 (a) S54P 및/또는 S57P 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다. Y51, N55 및 F56 중 하나 이상에서의 돌연변이는 하기 논의된 것들 중 임의의 것일 수 있다. (i)에서, 변이체는 바람직하게는 F56A/S57P 또는 S54P/F56A를 포함한다. 변이체는 바람직하게는 T150I를 포함한다. 대안적으로, 변이체는 바람직하게는 (a) T150I 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다.
(i)에서, 변이체는 바람직하게는 Q62R 또는 Q62K를 포함한다. 대안적으로, 변이체는 바람직하게는 (a) Q62R 또는 Q62K 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다. 변이체는 D43N, E44N, Q62R 또는 Q62K 또는 이의 임의의 조합, 예컨대 D43N, E44N, Q62R, Q62K, D43N/E44N, D43N/Q62R, D43N/Q62K, E44N/Q62R, E44N/Q62K, D43N/E44N/Q62R 또는 D43N/E44N/Q62K를 포함할 수 있다. 대안적으로, 변이체는 바람직하게는 (a) D43N, E44N, Q62R, Q62K, D43N/E44N, D43N/Q62R, D43N/Q62K, E44N/Q62R, E44N/Q62K, D43N/E44N/Q62R 또는 D43N/E44N/Q62K 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56에서의 돌연변이를 포함한다.
(i)에서, 변이체는 바람직하게는 D43N을 포함한다.
(i)에서, 변이체는 바람직하게는 E101R, E101S, E101F 또는 E101N을 포함한다.
(i)에서, 변이체는 바람직하게는 E124N, E124Q, E124R, E124K, E124F, E124Y, E124W 또는 E124D, 예컨대 E124N을 포함한다.
(i)에서, 변이체는 바람직하게는 R142E 및 R142N을 포함한다.
(i)에서, 변이체는 바람직하게는 R97N, R97G 또는 R97L을 포함한다.
(i)에서, 변이체는 바람직하게는 R192E 및 R192N을 포함한다.
(ii)에서, 변이체는 바람직하게는 F56N/N55Q, F56N/N55R, F56N/N55K, F56N/N55S, F56N/N55G, F56N/N55A, F56N/N55T, F56Q/N55Q, F56Q/N55R, F56Q/N55K, F56Q/N55S, F56Q/N55G, F56Q/N55A, F56Q/N55T, F56R/N55Q, F56R/N55R, F56R/N55K, F56R/N55S, F56R/N55G, F56R/N55A, F56R/N55T, F56S/N55Q, F56S/N55R, F56S/N55K, F56S/N55S, F56S/N55G, F56S/N55A, F56S/N55T, F56G/N55Q, F56G/N55R, F56G/N55K, F56G/N55S, F56G/N55G, F56G/N55A, F56G/N55T, F56A/N55Q, F56A/N55R, F56A/N55K, F56A/N55S, F56A/N55G, F56A/N55A, F56A/N55T, F56K/N55Q, F56K/N55R, F56K/N55K, F56K/N55S, F56K/N55G, F56K/N55A, F56K/N55T, F56N/Y51L, F56N/Y51V, F56N/Y51A, F56N/Y51N, F56N/Y51Q, F56N/Y51S, F56N/Y51G, F56Q/Y51L, F56Q/Y51V, F56Q/Y51A, F56Q/Y51N, F56Q/Y51Q, F56Q/Y51S, F56Q/Y51G, F56R/Y51L, F56R/Y51V, F56R/Y51A, F56R/Y51N, F56R/Y51Q, F56R/Y51S, F56R/Y51G, F56S/Y51L, F56S/Y51V, F56S/Y51A, F56S/Y51N, F56S/Y51Q, F56S/Y51S, F56S/Y51G, F56G/Y51L, F56G/Y51V, F56G/Y51A, F56G/Y51N, F56G/Y51Q, F56G/Y51S, F56G/Y51G, F56A/Y51L, F56A/Y51V, F56A/Y51A, F56A/Y51N, F56A/Y51Q, F56A/Y51S, F56A/Y51G, F56K/Y51L, F56K/Y51V, F56K/Y51A, F56K/Y51N, F56K/Y51Q, F56K/Y51S, F56K/Y51G, N55Q/Y51L, N55Q/Y51V, N55Q/Y51A, N55Q/Y51N, N55Q/Y51Q, N55Q/Y51S, N55Q/Y51G, N55R/Y51L, N55R/Y51V, N55R/Y51A, N55R/Y51N, N55R/Y51Q, N55R/Y51S, N55R/Y51G, N55K/Y51L, N55K/Y51V, N55K/Y51A, N55K/Y51N, N55K/Y51Q, N55K/Y51S, N55K/Y51G, N55S/Y51L, N55S/Y51V, N55S/Y51A, N55S/Y51N, N55S/Y51Q, N55S/Y51S, N55S/Y51G, N55G/Y51L, N55G/Y51V, N55G/Y51A, N55G/Y51N, N55G/Y51Q, N55G/Y51S, N55G/Y51G, N55A/Y51L, N55A/Y51V, N55A/Y51A, N55A/Y51N, N55A/Y51Q, N55A/Y51S, N55A/Y51G, N55T/Y51L, N55T/Y51V, N55T/Y51A, N55T/Y51N, N55T/Y51Q, N55T/Y51S, N55T/Y51G, F56N/N55Q/Y51L, F56N/N55Q/Y51V, F56N/N55Q/Y51A, F56N/N55Q/Y51N, F56N/N55Q/Y51Q, F56N/N55Q/Y51S, F56N/N55Q/Y51G, F56N/N55R/Y51L, F56N/N55R/Y51V, F56N/N55R/Y51A, F56N/N55R/Y51N, F56N/N55R/Y51Q, F56N/N55R/Y51S, F56N/N55R/Y51G, F56N/N55K/Y51L, F56N/N55K/Y51V, F56N/N55K/Y51A, F56N/N55K/Y51N, F56N/N55K/Y51Q, F56N/N55K/Y51S, F56N/N55K/Y51G, F56N/N55S/Y51L, F56N/N55S/Y51V, F56N/N55S/Y51A, F56N/N55S/Y51N, F56N/N55S/Y51Q, F56N/N55S/Y51S, F56N/N55S/Y51G, F56N/N55G/Y51L, F56N/N55G/Y51V, F56N/N55G/Y51A, F56N/N55G/Y51N, F56N/N55G/Y51Q, F56N/N55G/Y51S, F56N/N55G/Y51G, F56N/N55A/Y51L, F56N/N55A/Y51V, F56N/N55A/Y51A, F56N/N55A/Y51N, F56N/N55A/Y51Q, F56N/N55A/Y51S, F56N/N55A/Y51G, F56N/N55T/Y51L, F56N/N55T/Y51V, F56N/N55T/Y51A, F56N/N55T/Y51N, F56N/N55T/Y51Q, F56N/N55T/Y51S, F56N/N55T/Y51G, F56Q/N55Q/Y51L, F56Q/N55Q/Y51V, F56Q/N55Q/Y51A, F56Q/N55Q/Y51N, F56Q/N55Q/Y51Q, F56Q/N55Q/Y51S, F56Q/N55Q/Y51G, F56Q/N55R/Y51L, F56Q/N55R/Y51V, F56Q/N55R/Y51A, F56Q/N55R/Y51N, F56Q/N55R/Y51Q, F56Q/N55R/Y51S, F56Q/N55R/Y51G, F56Q/N55K/Y51L, F56Q/N55K/Y51V, F56Q/N55K/Y51A, F56Q/N55K/Y51N, F56Q/N55K/Y51Q, F56Q/N55K/Y51S, F56Q/N55K/Y51G, F56Q/N55S/Y51L, F56Q/N55S/Y51V, F56Q/N55S/Y51A, F56Q/N55S/Y51N, F56Q/N55S/Y51Q, F56Q/N55S/Y51S, F56Q/N55S/Y51G, F56Q/N55G/Y51L, F56Q/N55G/Y51V, F56Q/N55G/Y51A, F56Q/N55G/Y51N, F56Q/N55G/Y51Q, F56Q/N55G/Y51S, F56Q/N55G/Y51G, F56Q/N55A/Y51L, F56Q/N55A/Y51V, F56Q/N55A/Y51A, F56Q/N55A/Y51N, F56Q/N55A/Y51Q, F56Q/N55A/Y51S, F56Q/N55A/Y51G, F56Q/N55T/Y51L, F56Q/N55T/Y51V, F56Q/N55T/Y51A, F56Q/N55T/Y51N, F56Q/N55T/Y51Q, F56Q/N55T/Y51S, F56Q/N55T/Y51G, F56R/N55Q/Y51L, F56R/N55Q/Y51V, F56R/N55Q/Y51A, F56R/N55Q/Y51N, F56R/N55Q/Y51Q, F56R/N55Q/Y51S, F56R/N55Q/Y51G, F56R/N55R/Y51L, F56R/N55R/Y51V, F56R/N55R/Y51A, F56R/N55R/Y51N, F56R/N55R/Y51Q, F56R/N55R/Y51S, F56R/N55R/Y51G, F56R/N55K/Y51L, F56R/N55K/Y51V, F56R/N55K/Y51A, F56R/N55K/Y51N, F56R/N55K/Y51Q, F56R/N55K/Y51S, F56R/N55K/Y51G, F56R/N55S/Y51L, F56R/N55S/Y51V, F56R/N55S/Y51A, F56R/N55S/Y51N, F56R/N55S/Y51Q, F56R/N55S/Y51S, F56R/N55S/Y51G, F56R/N55G/Y51L, F56R/N55G/Y51V, F56R/N55G/Y51A, F56R/N55G/Y51N, F56R/N55G/Y51Q, F56R/N55G/Y51S, F56R/N55G/Y51G, F56R/N55A/Y51L, F56R/N55A/Y51V, F56R/N55A/Y51A, F56R/N55A/Y51N, F56R/N55A/Y51Q, F56R/N55A/Y51S, F56R/N55A/Y51G, F56R/N55T/Y51L, F56R/N55T/Y51V, F56R/N55T/Y51A, F56R/N55T/Y51N, F56R/N55T/Y51Q, F56R/N55T/Y51S, F56R/N55T/Y51G, F56S/N55Q/Y51L, F56S/N55Q/Y51V, F56S/N55Q/Y51A, F56S/N55Q/Y51N, F56S/N55Q/Y51Q, F56S/N55Q/Y51S, F56S/N55Q/Y51G, F56S/N55R/Y51L, F56S/N55R/Y51V, F56S/N55R/Y51A, F56S/N55R/Y51N, F56S/N55R/Y51Q, F56S/N55R/Y51S, F56S/N55R/Y51G, F56S/N55K/Y51L, F56S/N55K/Y51V, F56S/N55K/Y51A, F56S/N55K/Y51N, F56S/N55K/Y51Q, F56S/N55K/Y51S, F56S/N55K/Y51G, F56S/N55S/Y51L, F56S/N55S/Y51V, F56S/N55S/Y51A, F56S/N55S/Y51N, F56S/N55S/Y51Q, F56S/N55S/Y51S, F56S/N55S/Y51G, F56S/N55G/Y51L, F56S/N55G/Y51V, F56S/N55G/Y51A, F56S/N55G/Y51N, F56S/N55G/Y51Q, F56S/N55G/Y51S, F56S/N55G/Y51G, F56S/N55A/Y51L, F56S/N55A/Y51V, F56S/N55A/Y51A, F56S/N55A/Y51N, F56S/N55A/Y51Q, F56S/N55A/Y51S, F56S/N55A/Y51G, F56S/N55T/Y51L, F56S/N55T/Y51V, F56S/N55T/Y51A, F56S/N55T/Y51N, F56S/N55T/Y51Q, F56S/N55T/Y51S, F56S/N55T/Y51G, F56G/N55Q/Y51L, F56G/N55Q/Y51V, F56G/N55Q/Y51A, F56G/N55Q/Y51N, F56G/N55Q/Y51Q, F56G/N55Q/Y51S, F56G/N55Q/Y51G, F56G/N55R/Y51L, F56G/N55R/Y51V, F56G/N55R/Y51A, F56G/N55R/Y51N, F56G/N55R/Y51Q, F56G/N55R/Y51S, F56G/N55R/Y51G, F56G/N55K/Y51L, F56G/N55K/Y51V, F56G/N55K/Y51A, F56G/N55K/Y51N, F56G/N55K/Y51Q, F56G/N55K/Y51S, F56G/N55K/Y51G, F56G/N55S/Y51L, F56G/N55S/Y51V, F56G/N55S/Y51A, F56G/N55S/Y51N, F56G/N55S/Y51Q, F56G/N55S/Y51S, F56G/N55S/Y51G, F56G/N55G/Y51L, F56G/N55G/Y51V, F56G/N55G/Y51A, F56G/N55G/Y51N, F56G/N55G/Y51Q, F56G/N55G/Y51S, F56G/N55G/Y51G, F56G/N55A/Y51L, F56G/N55A/Y51V, F56G/N55A/Y51A, F56G/N55A/Y51N, F56G/N55A/Y51Q, F56G/N55A/Y51S, F56G/N55A/Y51G, F56G/N55T/Y51L, F56G/N55T/Y51V, F56G/N55T/Y51A, F56G/N55T/Y51N, F56G/N55T/Y51Q, F56G/N55T/Y51S, F56G/N55T/Y51G, F56A/N55Q/Y51L, F56A/N55Q/Y51V, F56A/N55Q/Y51A, F56A/N55Q/Y51N, F56A/N55Q/Y51Q, F56A/N55Q/Y51S, F56A/N55Q/Y51G, F56A/N55R/Y51L, F56A/N55R/Y51V, F56A/N55R/Y51A, F56A/N55R/Y51N, F56A/N55R/Y51Q, F56A/N55R/Y51S, F56A/N55R/Y51G, F56A/N55K/Y51L, F56A/N55K/Y51V, F56A/N55K/Y51A, F56A/N55K/Y51N, F56A/N55K/Y51Q, F56A/N55K/Y51S, F56A/N55K/Y51G, F56A/N55S/Y51L, F56A/N55S/Y51V, F56A/N55S/Y51A, F56A/N55S/Y51N, F56A/N55S/Y51Q, F56A/N55S/Y51S, F56A/N55S/Y51G, F56A/N55G/Y51L, F56A/N55G/Y51V, F56A/N55G/Y51A, F56A/N55G/Y51N, F56A/N55G/Y51Q, F56A/N55G/Y51S, F56A/N55G/Y51G, F56A/N55A/Y51L, F56A/N55A/Y51V, F56A/N55A/Y51A, F56A/N55A/Y51N, F56A/N55A/Y51Q, F56A/N55A/Y51S, F56A/N55A/Y51G, F56A/N55T/Y51L, F56A/N55T/Y51V, F56A/N55T/Y51A, F56A/N55T/Y51N, F56A/N55T/Y51Q, F56A/N55T/Y51S, F56A/N55T/Y51G, F56K/N55Q/Y51L, F56K/N55Q/Y51V, F56K/N55Q/Y51A, F56K/N55Q/Y51N, F56K/N55Q/Y51Q, F56K/N55Q/Y51S, F56K/N55Q/Y51G, F56K/N55R/Y51L, F56K/N55R/Y51V, F56K/N55R/Y51A, F56K/N55R/Y51N, F56K/N55R/Y51Q, F56K/N55R/Y51S, F56K/N55R/Y51G, F56K/N55K/Y51L, F56K/N55K/Y51V, F56K/N55K/Y51A, F56K/N55K/Y51N, F56K/N55K/Y51Q, F56K/N55K/Y51S, F56K/N55K/Y51G, F56K/N55S/Y51L, F56K/N55S/Y51V, F56K/N55S/Y51A, F56K/N55S/Y51N, F56K/N55S/Y51Q, F56K/N55S/Y51S, F56K/N55S/Y51G, F56K/N55G/Y51L, F56K/N55G/Y51V, F56K/N55G/Y51A, F56K/N55G/Y51N, F56K/N55G/Y51Q, F56K/N55G/Y51S, F56K/N55G/Y51G, F56K/N55A/Y51L, F56K/N55A/Y51V, F56K/N55A/Y51A, F56K/N55A/Y51N, F56K/N55A/Y51Q, F56K/N55A/Y51S, F56K/N55A/Y51G, F56K/N55T/Y51L, F56K/N55T/Y51V, F56K/N55T/Y51A, F56K/N55T/Y51N, F56K/N55T/Y51Q,F56K/N55T/Y51S, F56K/N55T/Y51G, F56E/N55R, F56E/N55K, F56D/N55R, F56D/N55K, F56R/N55E, F56R/N55D, F56K/N55E 또는 F56K/N55D를 포함한다.
(ii)에서, 변이체는 바람직하게는 Y51R/F56Q, Y51N/F56N, Y51M/F56Q, Y51L/F56Q, Y51I/F56Q, Y51V/F56Q, Y51A/F56Q, Y51P/F56Q, Y51G/F56Q, Y51C/F56Q, Y51Q/F56Q, Y51N/F56Q, Y51S/F56Q, Y51E/F56Q, Y51D/F56Q, Y51K/F56Q 또는 Y51H/F56Q를 포함한다.
(ii)에서, 변이체는 바람직하게는 Y51T/F56Q, Y51Q/F56Q 또는 Y51A/F56Q를 포함한다.
(ii)에서, 변이체는 바람직하게는 Y51T/F56F, Y51T/F56M, Y51T/F56L, Y51T/F56I, Y51T/F56V, Y51T/F56A, Y51T/F56P, Y51T/F56G, Y51T/F56C, Y51T/F56Q, Y51T/F56N, Y51T/F56T, Y51T/F56S, Y51T/F56E, Y51T/F56D, Y51T/F56K, Y51T/F56H 또는 Y51T/F56R을 포함한다.
(ii)에서, 변이체는 바람직하게는 Y51T/N55Q, Y51T/N55S 또는 Y51T/N55A를 포함한다.
(ii)에서, 변이체는 바람직하게는 Y51A/F56F, Y51A/F56L, Y51A/F56I, Y51A/F56V, Y51A/F56A, Y51A/F56P, Y51A/F56G, Y51A/F56C, Y51A/F56Q, Y51A/F56N, Y51A/F56T, Y51A/F56S, Y51A/F56E, Y51A/F56D, Y51A/F56K, Y51A/F56H 또는 Y51A/F56R을 포함한다.
(ii)에서, 변이체는 바람직하게는 Y51C/F56A, Y51E/F56A, Y51D/F56A, Y51K/F56A, Y51H/F56A, Y51Q/F56A, Y51N/F56A, Y51S/F56A, Y51P/F56A 또는 Y51V/F56A를 포함한다.
(xi)에서, 변이체는 바람직하게는 Y51/P52, Y51/P52/A53, P50 내지 P52, P50 내지 A53, K49 내지 Y51, K49 내지 A53의 결실 및 단일 프롤린 (P)으로의 교체, K49 내지 S54의 결실 및 단일 P로의 교체, Y51 내지 A53, Y51 내지 S54, N55/F56, N55 내지 S57, N55/F56의 결실 및 단일 P로의 교체, N55/F56의 결실 및 단일 글리신 (G)으로의 교체, N55/F56의 결실 및 단일 알라닌 (A)으로의 교체, N55/F56의 결실 및 단일 P로의 교체 및 Y51N, N55/F56의 결실 및 단일 P로의 교체 및 Y51Q, N55/F56 및 단일 P로의 교체 및 Y51S, N55/F56 및 단일 G로의 교체 및 Y51N, N55/F56의 결실 및 단일 G로의 교체 및 Y51Q, N55/F56의 결실 및 단일 G로의 교체 및 Y51S, N55/F56의 결실 및 단일 A로의 교체 및 Y51N, N55/F56의 결실 및 단일 A로의 교체/Y51Q, 또는 N55/F56의 결실 및 단일 A로의 교체 및 Y51S를 포함한다.
변이체는 더욱 바람직하게는 D195N/E203N, D195Q/E203N, D195N/E203Q, D195Q/E203Q, E201N/E203N, E201Q/E203N, E201N/E203Q, E201Q/E203Q, E185N/E203Q, E185Q/E203Q, E185N/E203N, E185Q/E203N, D195N/E201N/E203N, D195Q/E201N/E203N, D195N/E201Q/E203N, D195N/E201N/E203Q, D195Q/E201Q/E203N, D195Q/E201N/E203Q, D195N/E201Q/E203Q, D195Q/E201Q/E203Q, D149N/E201N, D149Q/E201N, D149N/E201Q, D149Q/E201Q, D149N/E201N/D195N, D149Q/E201N/D195N, D149N/E201Q/D195N, D149N/E201N/D195Q, D149Q/E201Q/D195N, D149Q/E201N/D195Q, D149N/E201Q/D195Q, D149Q/E201Q/D195Q, D149N/E203N, D149Q/E203N, D149N/E203Q, D149Q/E203Q, D149N/E185N/E201N, D149Q/E185N/E201N, D149N/E185Q/E201N, D149N/E185N/E201Q, D149Q/E185Q/E201N, D149Q/E185N/E201Q, D149N/E185Q/E201Q, D149Q/E185Q/E201Q, D149N/E185N/E203N, D149Q/E185N/E203N, D149N/E185Q/E203N, D149N/E185N/E203Q, D149Q/E185Q/E203N, D149Q/E185N/E203Q, D149N/E185Q/E203Q, D149Q/E185Q/E203Q, D149N/E185N/E201N/E203N, D149Q/E185N/E201N/E203N, D149N/E185Q/E201N/E203N, D149N/E185N/E201Q/E203N, D149N/E185N/E201N/E203Q, D149Q/E185Q/E201N/E203N, D149Q/E185N/E201Q/E203N, D149Q/E185N/E201N/E203Q, D149N/E185Q/E201Q/E203N, D149N/E185Q/E201N/E203Q, D149N/E185N/E201Q/E203Q, D149Q/E185Q/E201Q/E203Q, D149Q/E185Q/E201N/E203Q, D149Q/E185N/E201Q/E203Q, D149N/E185Q/E201Q/E203Q, D149Q/E185Q/E201Q/E203N, D149N/E185N/D195N/E201N/E203N, D149Q/E185N/D195N/E201N/E203N, D149N/E185Q/D195N/E201N/E203N, D149N/E185N/D195Q/E201N/E203N, D149N/E185N/D195N/E201Q/E203N, D149N/E185N/D195N/E201N/E203Q, D149Q/E185Q/D195N/E201N/E203N, D149Q/E185N/D195Q/E201N/E203N, D149Q/E185N/D195N/E201Q/E203N, D149Q/E185N/D195N/E201N/E203Q, D149N/E185Q/D195Q/E201N/E203N, D149N/E185Q/D195N/E201Q/E203N, D149N/E185Q/D195N/E201N/E203Q, D149N/E185N/D195Q/E201Q/E203N, D149N/E185N/D195Q/E201N/E203Q, D149N/E185N/D195N/E201Q/E203Q, D149Q/E185Q/D195Q/E201N/E203N, D149Q/E185Q/D195N/E201Q/E203N, D149Q/E185Q/D195N/E201N/E203Q, D149Q/E185N/D195Q/E201Q/E203N, D149Q/E185N/D195Q/E201N/E203Q, D149Q/E185N/D195N/E201Q/E203Q, D149N/E185Q/D195Q/E201Q/E203N, D149N/E185Q/D195Q/E201N/E203Q, D149N/E185Q/D195N/E201Q/E203Q, D149N/E185N/D195Q/E201Q/E203Q, D149Q/E185Q/D195Q/E201Q/E203N, D149Q/E185Q/D195Q/E201N/E203Q, D149Q/E185Q/D195N/E201Q/E203Q, D149Q/E185N/D195Q/E201Q/E203Q, D149N/E185Q/D195Q/E201Q/E203Q, D149Q/E185Q/D195Q/E201Q/E203Q, D149N/E185R/E201N/E203N, D149Q/E185R/E201N/E203N, D149N/E185R/E201Q/E203N, D149N/E185R/E201N/E203Q, D149Q/E185R/E201Q/E203N, D149Q/E185R/E201N/E203Q, D149N/E185R/E201Q/E203Q, D149Q/E185R/E201Q/E203Q, D149R/E185N/E201N/E203N, D149R/E185Q/E201N/E203N, D149R/E185N/E201Q/E203N, D149R/E185N/E201N/E203Q, D149R/E185Q/E201Q/E203N, D149R/E185Q/E201N/E203Q, D149R/E185N/E201Q/E203Q, D149R/E185Q/E201Q/E203Q, D149R/E185N/D195N/E201N/E203N, D149R/E185Q/D195N/E201N/E203N, D149R/E185N/D195Q/E201N/E203N, D149R/E185N/D195N/E201Q/E203N, D149R/E185Q/D195N/E201N/E203Q, D149R/E185Q/D195Q/E201N/E203N, D149R/E185Q/D195N/E201Q/E203N, D149R/E185Q/D195N/E201N/E203Q, D149R/E185N/D195Q/E201Q/E203N, D149R/E185N/D195Q/E201N/E203Q, D149R/E185N/D195N/E201Q/E203Q, D149R/E185Q/D195Q/E201Q/E203N, D149R/E185Q/D195Q/E201N/E203Q, D149R/E185Q/D195N/E201Q/E203Q, D149R/E185N/D195Q/E201Q/E203Q, D149R/E185Q/D195Q/E201Q/E203Q, D149N/E185R/D195N/E201N/E203N, D149Q/E185R/D195N/E201N/E203N, D149N/E185R/D195Q/E201N/E203N, D149N/E185R/D195N/E201Q/E203N, D149N/E185R/D195N/E201N/E203Q, D149Q/E185R/D195Q/E201N/E203N, D149Q/E185R/D195N/E201Q/E203N, D149Q/E185R/D195N/E201N/E203Q, D149N/E185R/D195Q/E201Q/E203N, D149N/E185R/D195Q/E201N/E203Q, D149N/E185R/D195N/E201Q/E203Q, D149Q/E185R/D195Q/E201Q/E203N, D149Q/E185R/D195Q/E201N/E203Q, D149Q/E185R/D195N/E201Q/E203Q, D149N/E185R/D195Q/E201Q/E203Q, D149Q/E185R/D195Q/E201Q/E203Q, D149N/E185R/D195N/E201R/E203N, D149Q/E185R/D195N/E201R/E203N, D149N/E185R/D195Q/E201R/E203N, D149N/E185R/D195N/E201R/E203Q, D149Q/E185R/D195Q/E201R/E203N, D149Q/E185R/D195N/E201R/E203Q, D149N/E185R/D195Q/E201R/E203Q, D149Q/E185R/D195Q/E201R/E203Q, E131D/K49R, E101N/N102F, E101N/N102Y, E101N/N102W, E101F/N102F, E101F/N102Y, E101F/N102W, E101Y/N102F, E101Y/N102Y, E101Y/N102W, E101W/N102F, E101W/N102Y, E101W/N102W, E101N/N102R, E101F/N102R, E101Y/N102R 또는 E101W/N102F를 포함한다.
폴리뉴클레오티드가 포어를 통해 이동할 때 더 적은 뉴클레오티드가 전류에 기여하는 포어를 형성하는 본 발명의 바람직한 변이체는 Y51A/F56A, Y51A/F56N, Y51I/F56A, Y51L/F56A, Y51T/F56A, Y51I/F56N, Y51L/F56N 또는 Y51T/F56N을 포함하거나, 더욱 바람직하게는 Y51I/F56A, Y51L/F56A 또는 Y51T/F56A를 포함한다. 상기 논의된 바와 같이, 이는 관찰된 전류 (폴리뉴클레오티드가 포어를 통해 이동할 때)와 폴리뉴클레오티드 사이의 직접적인 관계를 확인하는 것을 더 쉽게 만든다.
증가된 범위를 나타내는 포어를 형성하는 바람직한 변이체는 하기 위치에서의 돌연변이를 포함한다:
Y51, F56, D149, E185, E201 및 E203;
N55 및 F56;
Y51및 F56;
Y51, N55 및 F56; 또는
F56 및 N102.
증가된 범위를 나타내는 포어를 형성하는 바람직한 변이체는 하기를 포함한다:
Y51N, F56A, D149N, E185R, E201N 및 E203N;
N55S 및 F56Q;
Y51A 및 F56A;
Y51A 및 F56N;
Y51I 및 F56A;
Y51L 및 F56A;
Y51T 및 F56A;
Y51I 및 F56N;
Y51L 및 F56N;
Y51T 및 F56N;
Y51T 및 F56Q;
Y51A, N55S 및 F56A;
Y51A, N55S 및 F56N;
Y51T, N55S 및 F56Q; 또는
F56Q 및 N102R.
폴리뉴클레오티드가 포어를 통해 이동할 때 더 적은 뉴클레오티드가 전류에 기여하는 포어를 형성하는 바람직한 변이체는 하기 위치에서의 돌연변이를 포함한다:
N55 및 F56, 예컨대 N55X 및 F56Q [식 중, X는 임의의 아미노산이다]; 또는
Y51 및 F56, 예컨대 Y51X 및 F56Q [식 중, X는 임의의 아미노산이다].
증가된 처리량을 나타내는 포어를 형성하는 바람직한 변이체는 하기 위치에서의 돌연변이를 포함한다:
D149, E185 및 E203;
D149, E185, E201 및 E203; 또는
D149, E185, D195, E201 및 E203.
증가된 처리량을 나타내는 포어를 형성하는 바람직한 변이체는 하기를 포함한다:
D149N, E185N 및 E203N;
D149N, E185N, E201N 및 E203N;
D149N, E185R, D195N, E201N 및 E203N; 또는
D149N, E185R, D195N, E201R 및 E203N.
폴리뉴클레오티드의 포착이 증가된 포어를 형성하는 바람직한 변이체는 하기의 돌연변이를 포함한다:
D43N/Y51T/F56Q;
E44N/Y51T/F56Q;
D43N/E44N/Y51T/F56Q;
Y51T/F56Q/Q62R;
D43N/Y51T/F56Q/Q62R;
E44N/Y51T/F56Q/Q62R; 또는
D43N/E44N/Y51T/F56Q/Q62R.
바람직한 변이체는 하기의 돌연변이를 포함한다:
D149R/E185R/E201R/E203R 또는 Y51T/F56Q/D149R/E185R/E201R/E203R;
D149N/E185N/E201N/E203N 또는 Y51T/F56Q/D149N/E185N/E201N/E203N;
E201R/E203R 또는 Y51T/F56Q/E201R/E203R
E201N/E203R 또는 Y51T/F56Q/E201N/E203R;
E203R 또는 Y51T/F56Q/E203R;
E203N 또는 Y51T/F56Q/E203N;
E201R 또는 Y51T/F56Q/E201R;
E201N 또는 Y51T/F56Q/E201N;
E185R 또는 Y51T/F56Q/E185R;
E185N 또는 Y51T/F56Q/E185N;
D149R 또는 Y51T/F56Q/D149R;
D149N 또는 Y51T/F56Q/D149N;
R142E 또는 Y51T/F56Q/R142E;
R142N 또는 Y51T/F56Q/R142N;
R192E 또는 Y51T/F56Q/R192E; 또는
R192N 또는 Y51T/F56Q/R192N.
바람직한 변이체는 하기의 돌연변이를 포함한다:
Y51A/F56Q/E101N/N102R;
Y51A/F56Q/R97N/N102G;
Y51A/F56Q/R97N/N102R;
Y51A/F56Q/R97N;
Y51A/F56Q/R97G;
Y51A/F56Q/R97L;
Y51A/F56Q/N102R;
Y51A/F56Q/N102F;
Y51A/F56Q/N102G;
Y51A/F56Q/E101R;
Y51A/F56Q/E101F;
Y51A/F56Q/E101N; 또는
Y51A/F56Q/E101G
본 발명은 변이체가 T150에서의 돌연변이를 포함하는, 서열식별번호: 390에 제시된 서열의 변이체를 포함하는 돌연변이체 CsgG 단량체를 또한 제공한다. 증가된 삽입을 나타내는 포어를 형성하는 바람직한 변이체는 T150I를 포함한다. T150에서의 돌연변이, 예컨대 T150I는 상기 논의된 돌연변이 또는 돌연변이 조합 중 임의의 것과 조합될 수 있다.
본 발명은 실시예에서 개시된 변이체 내에 존재하는 돌연변이의 조합을 포함하는 서열식별번호: 390에 제시된 서열의 변이체를 포함하는 돌연변이체 CsgG 단량체를 또한 제공한다.
천연 발생 아미노산을 도입하거나 치환하는 방법은 관련 기술 분야에 널리 공지되어 있다. 예를 들어, 돌연변이체 단량체를 코딩하는 폴리뉴클레오티드 내의 관련된 위치에서 메티오닌에 대한 코돈 (ATG)을 아르기닌에 대한 코돈 (CGT)으로 교체함으로써 메티오닌 (M)이 아르기닌 (R)으로 치환될 수 있다. 그 후, 하기 논의된 바와 같이 이러한 폴리뉴클레오티드가 발현될 수 있다.
비-천연 발생 아미노산을 도입하거나 치환하는 방법 또한 관련 기술 분야에 널리 공지되어 있다. 예를 들어, 돌연변이체 단량체를 발현하는데 사용되는 IVTT 시스템 내에 합성 아미노아실-tRNA를 포함시킴으로써 비-천연 발생 아미노산이 도입될 수 있다. 대안적으로, 특정 아미노산에 대해 영양요구성인 이. 콜라이에서 이러한 특정 아미노산의 합성 (즉, 비-천연-발생) 유사체의 존재 하에 돌연변이체 단량체를 발현시킴으로써 비-천연 발생 아미노산이 도입될 수 있다. 돌연변이체 단량체가 부분적인 펩티드 합성을 사용하여 생산되는 경우 네이키드 라이게이션(naked ligation)에 의해 비-천연 발생 아미노산이 또한 생산될 수 있다.
본 발명의 기타 단량체
또 다른 실시양태에서, 본 발명은 변이체가 위치 Y51, N55 및 F56 중 하나 이상에서의 돌연변이를 포함하는, 서열식별번호: 390에 제시된 서열의 변이체를 포함하는 돌연변이체 CsgG 단량체를 제공한다. 변이체는 Y51; N55; F56; Y51/N55; Y51/F56; N55/F56; 또는 Y51/N55/F56에서의 돌연변이를 포함할 수 있다. 변이체는 바람직하게는 Y51, N55 또는 F56에서의 돌연변이를 포함한다. 변이체는 상기 논의된 위치 Y51, N55 및 F56 중 하나 이상에서의 임의로 조합된 특정 돌연변이 중 임의의 것을 포함할 수 있다. 하나 이상의 Y51, N55 및 F56이 임의의 아미노산으로 치환될 수 있다. Y51이 F, M, L, I, V, A, P, G, C, Q, N, T, S, E, D, K, H 또는 R, 예컨대 A, S, T, N 또는 Q로 치환될 수 있다. N55가 F, M, L, I, V, A, P, G, C, Q, T, S, E, D, K, H 또는 R, 예컨대 A, S, T 또는 Q로 치환될 수 있다. F56이 M, L, I, V, A, P, G, C, Q, N, T, S, E, D, K, H 또는 R, 예컨대 A, S, T, N 또는 Q로 치환될 수 있다.
변이체는 하기 중 하나 이상을 추가로 포함할 수 있다: (i) 하기 위치 (i) N40, D43, E44, S54, S57, Q62, R97, E101, E124, E131, R142, T150 및 R192에서의 하나 이상의 돌연변이 (즉, 상기 위치 중 하나 이상에서의 돌연변이); (iii) Q42R 또는 Q42K; (iv) K49R; (v) N102R, N102F, N102Y 또는 N102W; (vi) D149N, D149Q 또는 D149R; (vii) E185N, E185Q 또는 E185R; (viii) D195N, D195Q 또는 D195R; (ix) E201N, E201Q 또는 E201R; (x) E203N, E203Q 또는 E203R; 및 (xi) 하기 위치 F48, K49, P50, Y51, P52, A53, S54, N55, F56 및 S57 중 하나 이상의 결실. 변이체는 상기 논의된 (i) 및 (iii) 내지 (xi)의 조합 중 임의의 것을 포함할 수 있다. 변이체는 (i) 및 (iii) 내지 (xi)에 대해 상기 논의된 실시양태 중 임의의 것을 포함할 수 있다.
변이체
상기 논의된 특정 돌연변이에 더하여, 변이체는 기타 돌연변이를 포함할 수 있다. 서열식별번호: 390의 아미노산 서열의 전체 길이에 걸쳐, 변이체는 이러한 서열에 대해 아미노산 동일성을 기초로 바람직하게는 적어도 50% 상동성일 것이다. 더욱 바람직하게는, 변이체는 아미노산 동일성을 기초로 서열식별번호: 390의 아미노산 서열에 대해 전체 서열에 걸쳐 적어도 55%, 적어도 60%, 적어도 65%, 적어도 70%, 적어도 75%, 적어도 80%, 적어도 85%, 적어도 90%, 더욱 바람직하게는 적어도 95%, 97% 또는 99% 상동성일 수 있다. 100개 이상, 예를 들어 125, 150, 175 또는 200개 또는 이를 초과하는 개수의 연속된 아미노산의 신장물에 걸쳐 적어도 80%, 예를 들어 적어도 85%, 90% 또는 95%의 아미노산 동일성이 있을 수 있다 ("강한 상동성(hard homology)").
관련 기술 분야의 표준 방법을 사용하여 상동성을 결정할 수 있다. 예를 들어, UWGCG 패키지가 상동성을 계산하는데 사용될 수 있는 BESTFIT 프로그램 (예를 들어, 이의 디폴트 설정에서 사용됨)을 제공한다 (Devereux et al. (1984) Nucleic Acids Research 12, p387-395). 예를 들어 문헌 [Altschul S. F. (1993) J Mol Evol 36:290-300]; [Altschul, S.F et al. (1990) J Mol Biol 215:403-10]에 기술된 바와 같이, PILEUP 및 BLAST 알고리즘을 상동성을 계산하거나 서열을 정렬하는데 사용할 수 있다 (예컨대 (전형적으로는 디폴트 설정에서) 등가의 잔기 또는 상응하는 서열을 확인함). BLAST 분석을 수행하는 소프트웨어를 국립 생물공학 정보 센터 (http://www.ncbi.nlm.nih.gov/)를 통해 공개적으로 입수할 수 있다.
서열식별번호: 390은 에스케리키아 콜라이 균주 K-12 아계 MC4100으로부터의 야생형 CsgG 단량체이다. 서열식별번호: 390의 변이체는 또 다른 CsgG 상동체 내에 존재하는 치환 중 임의의 것을 포함할 수 있다. 바람직한 CsgG 상동체가 서열식별번호: 391 내지 395 및 414 내지 429에서 제시된다. 변이체는 서열식별번호: 390과 비교하여 서열식별번호: 391 내지 395 및 414 내지 429 내에 존재하는 치환 중 하나 이상의 조합을 포함할 수 있다.
상기에서 논의된 것들에 더하여 아미노산 치환, 예를 들어, 1, 2, 3, 4, 5, 10, 20 또는 30개까지의 치환이 서열식별번호: 390의 아미노산 서열에 이루어질 수 있다. 보존적 치환은 아미노산을 유사한 화학 구조, 유사한 화학 성질 또는 유사한 측쇄 부피의 다른 아미노산으로 교체한다. 도입된 아미노산은 자신이 교체하는 아미노산과 유사한 극성, 친수성, 소수성, 염기도, 산도, 중성도 또는 전하를 가질 수 있다. 대안적으로, 보존적 치환은 기존의 방향족 또는 지방족 아미노산의 위치에 방향족 또는 지방족인 또 다른 아미노산을 도입할 수 있다. 보존적 아미노산 변화는 관련 기술 분야에 널리 공지되어 있고, 하기 표 1에서 정의된 바와 같은 20개의 주요 아미노산의 성질에 따라 선택될 수 있다. 아미노산들이 유사한 극성을 갖는 경우, 이는 표 2의 아미노산 측쇄에 대한 소수친수성(hydropathy) 척도를 참조로 또한 결정될 수 있다.
<표 1>
아미노산의 화학적 성질
<표 2>
소수친수성 척도
서열식별번호: 390의 아미노산 서열의 하나 이상의 아미노산 잔기가 상기 기술된 폴리펩티드로부터 추가적으로 결실될 수 있다. 1, 2, 3, 4, 5, 10, 20 또는 30개 이상까지의 잔기가 결실될 수 있다.
변이체는 서열식별번호: 390의 단편을 포함할 수 있다. 이같은 단편은 포어 형성 활성을 유지한다. 단편은 적어도 50개, 적어도 100개, 적어도 150개, 적어도 200개 또는 적어도 250개의 아미노산의 길이일 수 있다. 이같은 단편을 사용하여 포어를 생산할 수 있다. 단편은 바람직하게는 서열식별번호: 390의 막 스패닝 도메인, 즉 K135-Q153 및 S183-S208을 포함한다.
하나 이상의 아미노산이 대안적으로 또는 추가적으로 상기 기술된 폴리펩티드에 부가될 수 있다. 확장물이 서열식별번호: 390 또는 이의 폴리펩티드 변이체 또는 단편의 아미노산 서열의 아미노 말단 또는 카르복시 말단에서 제공될 수 있다. 확장물은 아주 짧을 수 있고, 예를 들어, 1 내지 10개의 아미노산의 길이일 수 있다. 대안적으로, 확장물이 더 길 수 있고, 예를 들어, 50 내지 100개까지의 아미노산일 수 있다. 캐리어 단백질이 본 발명에 따른 아미노산 서열에 융합될 수 있다. 기타 융합 단백질이 하기에서 더욱 상세하게 논의된다.
상기 논의된 바와 같이, 변이체는 서열식별번호: 390의 것과 상이한 아미노산 서열을 갖고 포어를 형성하는 이의 능력을 유지하는 폴리펩티드이다. 전형적으로 변이체는 포어 형성을 담당하는 서열식별번호: 390의 영역을 포함한다. β-배럴을 함유하는 CsgG의 포어 형성 능력은 각각의 서브유닛 내의 β-시트에 의해 제공된다. 서열식별번호: 390의 변이체는 β-시트를 형성하는 서열식별번호: 390 내의 영역, 즉 K135-Q153 및 S183-S208을 전형적으로 포함한다. 생성된 변이체가 포어를 형성하는 이의 능력을 유지하는 한, 하나 이상의 변형이 β-시트를 형성하는 서열식별번호: 390 내의 영역에 이루어질 수 있다. 바람직하게는 서열식별번호: 390의 변이체는 이의 α-나선 및/또는 루프 영역 내에 하나 이상의 변형, 예컨대 치환, 부가 또는 결실을 포함한다.
예를 들어, 스트렙타비딘 태그의 부가 또는 세포로부터의 분비를 촉진하는 신호 서열의 부가 (단량체가 이같은 서열을 천연적으로 함유하지 않는 경우)에 의해, CsgG로부터 유래된 단량체가 이의 확인 또는 정제를 돕도록 변형될 수 있다. 기타 적절한 태그가 하기에서 더욱 상세하게 논의된다. 단량체가 시현성 표지로 표지될 수 있다. 시현성 표지는 단량체가 검출되게 하는 임의의 적절한 표지일 수 있다. 적절한 표지가 하기에서 기술된다.
CsgG로부터 유래된 단량체를 D-아미노산을 사용하여 또한 생산할 수 있다. 예를 들어, CsgG로부터 유래된 단량체가 L-아미노산 및 D-아미노산의 혼합물을 포함할 수 있다. 이는 이같은 단백질 또는 펩티드를 생산하는 분야에서 통상적이다.
CsgG로부터 유래된 단량체는 뉴클레오티드 판별을 용이하게 하는 하나 이상의 특이적 변형을 함유한다. CsgG로부터 유래된 단량체는, 변형이 포어 형성을 방해하지 않는 한, 다른 비-특이적 변형을 또한 함유할 수 있다. 다수의 비-특이적 측쇄 변형이 관련 기술 분야에 공지되어 있고, CsgG로부터 유래된 단량체의 측쇄에 이루어질 수 있다. 이같은 변형은, 예를 들어, 알데히드와 반응시킨 후 NaBH4로 환원시키는 것에 의한 아미노산의 환원적 알킬화, 메틸아세트이미데이트로의 아미딘화, 또는 아세트산 무수물로의 아실화를 포함한다.
CsgG로부터 유래된 단량체를 관련 기술 분야에 공지된 표준 방법을 사용하여 생산할 수 있다. CsgG로부터 유래된 단량체를 합성으로 또는 재조합 수단에 의해 제조할 수 있다. 예를 들어, 시험관내 번역 및 전사 (IVTT)에 의해 단량체를 합성할 수 있다. 포어 및 단량체를 생산하는 적절한 방법이 국제 출원 번호 PCT/GB09/001690 (WO 2010/004273으로 공개됨), PCT/GB09/001679 (WO 2010/004265로 공개됨) 또는 PCT/GB10/000133 (WO 2010/086603으로 공개됨)에 논의되어 있다. 포어를 막 내로 삽입하는 방법이 논의되어 있다.
일부 실시양태에서, 돌연변이체 단량체가 화학적으로 변형된다. 임의의 방식으로 임의의 부위에서 돌연변이체 단량체가 화학적으로 변형될 수 있다. 바람직하게는, 분자를 하나 이상의 시스테인에 부착하는 것 (시스테인 연결), 분자를 하나 이상의 리신에 부착하는 것, 분자를 하나 이상의 비-천연 아미노산에 부착하는 것, 에피토프의 효소 변형, 또는 말단 변형에 의해 돌연변이체 단량체가 화학적으로 변형된다. 이같은 변형을 수행하는 적절한 방법이 관련 기술 분야에 널리 공지되어 있다. 임의의 분자의 부착에 의해 돌연변이체 단량체가 화학적으로 변형될 수 있다. 예를 들어, 염료 또는 형광단의 부착에 의해 돌연변이체 단량체가 화학적으로 변형될 수 있다.
일부 실시양태에서, 돌연변이체 단량체가 단량체를 포함하는 포어와 표적 뉴클레오티드 또는 표적 폴리뉴클레오티드 서열 사이의 상호작용을 용이하게 하는 분자 어댑터로 화학적으로 변형된다. 어댑터의 존재는 포어 및 뉴클레오티드 또는 폴리뉴클레오티드 서열의 주객(host-guest) 화학을 개선하고, 이에 의해, 돌연변이체 단량체로부터 형성된 포어의 시퀀싱 능력을 개선한다. 주객 화학의 원리는 관련 기술 분야에 공지되어 있다. 어댑터는 포어의 물리적 또는 화학적 성질에 대한 효과가 있고, 이러한 효과는 포어와 뉴클레오티드 또는 폴리뉴클레오티드 서열의 상호작용을 개선한다. 어댑터는 포어의 배럴 또는 채널의 전하를 변경시키거나, 또는 특이적으로 뉴클레오티드 또는 폴리뉴클레오티드 서열과 상호작용하거나 이에 결합함으로써, 서열과 포어의 상호작용을 용이하게 할 수 있다.
분자 어댑터는 바람직하게는 고리형 분자, 시클로덱스트린, 혼성화할 수 있는 종, DNA 결합제 또는 인터칼레이터(interchelator), 펩티드 또는 펩티드 유사체, 합성 중합체, 방향족 평면 분자, 양으로 하전된 소형 분자, 또는 수소-결합이 가능한 소형 분자이다.
어댑터는 고리형일 수 있다. 고리형 어댑터는 바람직하게는 대칭성이 포어와 동일하다. CsgG가 전형적으로 중심축 둘레로 8개 또는 9개의 서브유닛을 갖기 때문에 어댑터는 바람직하게는 8중 또는 9중 대칭성을 갖는다. 이는 하기에서 더욱 상세하게 논의된다.
전형적으로 어댑터는 주객 화학을 통해 뉴클레오티드 또는 폴리뉴클레오티드 서열과 상호작용한다. 어댑터는 전형적으로 뉴클레오티드 또는 폴리뉴클레오티드 서열과 상호작용할 수 있다. 어댑터는 뉴클레오티드 또는 폴리뉴클레오티드 서열과 상호작용할 수 있는 하나 이상의 화학 기를 포함한다. 바람직하게는 하나 이상의 화학 기는 비-공유결합 상호작용, 예컨대 소수성 상호작용, 수소 결합, 반데르발스 힘, π-양이온 상호작용 및/또는 정전기력에 의해 뉴클레오티드 또는 폴리뉴클레오티드 서열과 상호작용한다. 뉴클레오티드 또는 폴리뉴클레오티드 서열과 상호작용할 수 있는 하나 이상의 화학 기는 바람직하게는 양으로 하전된다. 뉴클레오티드 또는 폴리뉴클레오티드 서열과 상호작용할 수 있는 하나 이상의 화학 기는 더욱 바람직하게는 아미노 기를 포함한다. 1급, 2급 또는 3급 탄소 원자에 아미노 기가 부착될 수 있다. 어댑터는 더욱 더 바람직하게는 아미노 기의 고리, 예컨대 6, 7 또는 8개의 아미노 기의 고리를 포함한다. 어댑터는 가장 바람직하게는 8개의 아미노 기의 고리를 포함한다. 양성자화된 아미노 기의 고리가 뉴클레오티드 또는 폴리뉴클레오티드 서열 내의 음으로 하전된 포스페이트 기와 상호작용할 수 있다.
어댑터를 포어 내에 정확하게 위치시키는 것이 어댑터와 돌연변이체 단량체를 포함하는 포어 사이의 주객 화학에 의해 용이해질 수 있다. 바람직하게는 어댑터는 포어 내의 하나 이상의 아미노산과 상호작용할 수 있는 하나 이상의 화학 기를 포함한다. 더욱 바람직하게는 어댑터는 비-공유결합 상호작용, 예컨대 소수성 상호작용, 수소 결합, 반데르발스 힘, π-양이온 상호작용 및/또는 정전기력을 통해 포어 내의 하나 이상의 아미노산과 상호작용할 수 있는 하나 이상의 화학 기를 포함한다. 포어 내의 하나 이상의 아미노산과 상호작용할 수 있는 화학 기는 전형적으로는 히드록실 또는 아민이다. 1급, 2급 또는 3급 탄소 원자에 히드록실 기가 부착될 수 있다. 히드록실 기는 포어 내의 하전되지 않은 아미노산과 수소 결합을 형성할 수 있다. 포어와 뉴클레오티드 또는 폴리뉴클레오티드 서열 사이의 상호작용을 용이하게 하는 임의의 어댑터가 사용될 수 있다.
적절한 어댑터는 시클로덱스트린, 고리형 펩티드 및 쿠커비투릴(cucurbituril)을 포함하지만, 이에 제한되지 않는다. 어댑터는 바람직하게는 시클로덱스트린 또는 이의 유도체이다. 시클로덱스트린 또는 이의 유도체는 문헌 [Eliseev, A. V., and Schneider, H-J. (1994) J. Am. Chem. Soc. 116, 6081-6088]에 기술된 것들 중 임의의 것일 수 있다. 어댑터는 더욱 바람직하게는 헵타키스-6-아미노-β-시클로덱스트린 (am7-βCD), 6-모노데옥시-6-모노아미노-β-시클로덱스트린 (am1-βCD) 또는 헵타키스-(6-데옥시-6-구아니디노)-시클로덱스트린 (gu7-βCD)이다. gu7-βCD 내의 구아니디노 기는 am7-βCD 내의 1급 아민보다 pKa가 훨씬 더 높고, 따라서 더 많이 양으로 하전된다. 포어 내의 뉴클레오티드의 체류 시간을 증가시키도록, 측정된 잔여 전류의 정확도를 증가시키도록, 뿐만 아니라 고온에서의 염기 검출률 또는 낮은 데이터 취득률을 증가시키도록 이러한 gu7-βCD 어댑터를 사용할 수 있다.
하기에서 더욱 상세하게 논의되는 바와 같이 숙신이미딜 3-(2-피리딜디티오)프로피오네이트 (SPDP) 가교제가 사용되는 경우, 어댑터는 바람직하게는 헵타키스(6-데옥시-6-아미노)-6-N-모노(2-피리딜)디티오프로파노일-β-시클로덱스트린 (am6amPDP1-βCD)이다.
더욱 적절한 어댑터는 γ-시클로덱스트린을 포함하고, 이는 9개의 당 단위를 포함한다 (따라서 9중 대칭성을 갖는다). γ-시클로덱스트린은 링커 분자를 함유할 수 있거나, 또는 상기 논의된 β-시클로덱스트린 예에서 사용된 변형된 당 단위를 모두 또는 더 많이 포함하도록 변형될 수 있다.
분자 어댑터는 바람직하게는 돌연변이체 단량체에 공유결합으로 부착된다. 관련 기술 분야에 공지된 임의의 방법을 사용하여 어댑터가 포어에 공유결합으로 부착될 수 있다. 전형적으로 어댑터는 화학적 연결을 통해 부착된다. 분자 어댑터가 시스테인 연결을 통해 부착되는 경우, 바람직하게는, 하나 이상의 시스테인이 치환에 의해 돌연변이체에, 예를 들어 배럴 내에, 도입되어 있다. 분자 어댑터가 돌연변이체 단량체 내의 하나 이상의 시스테인에 부착되는 것에 의해 돌연변이체 단량체가 화학적으로 변형될 수 있다. 하나 이상의 시스테인은 천연적으로 발생할 수 있고, 즉 서열식별번호: 390 내의 위치 1 및/또는 215에 있을 수 있다. 대안적으로, 다른 위치에서 도입된 하나 이상의 시스테인에 분자가 부착되는 것에 의해 돌연변이체 단량체가 화학적으로 변형될 수 있다. 위치 215의 시스테인이 예를 들어 치환에 의해 제거되어, 분자 어댑터가 위치 1의 시스테인 또는 또 다른 위치에서 도입된 시스테인이 아니라 이러한 위치에 부착되지 않는 것을 확실하게 할 수 있다.
인접한 잔기의 변형에 의해 시스테인 잔기의 반응성이 강화될 수 있다. 예를 들어, 측면의 아르기닌, 히스티딘 또는 리신 잔기의 염기성 기가 시스테인 티올 기의 pKa를 더욱 반응성인 S- 기의 것으로 변화시킬 것이다. 티올 보호기 예컨대 dTNB에 의해 시스테인 잔기의 반응성이 보호될 수 있다. 이들은 링커가 부착되기 전에 돌연변이체 단량체의 하나 이상의 시스테인 잔기와 반응될 수 있다.
이러한 분자는 돌연변이체 단량체에 직접적으로 부착될 수 있다. 바람직하게는 이러한 분자는 링커, 예컨대 화학적 가교제 또는 펩티드 링커를 사용하여 돌연변이체 단량체에 부착된다.
적절한 화학적 가교제가 관련 기술 분야에 널리 공지되어 있다. 바람직한 가교제는 2,5-디옥소피롤리딘-1-일 3-(피리딘-2-일디술파닐)프로파노에이트, 2,5-디옥소피롤리딘-1-일 4-(피리딘-2-일디술파닐)부타노에이트 및 2,5-디옥소피롤리딘-1-일 8-(피리딘-2-일디술파닐)옥타노에이트를 포함한다. 가장 바람직한 가교제는 숙신이미딜 3-(2-피리딜디티오)프로피오네이트 (SPDP)이다. 전형적으로, 분자가 이관능성 가교제에 공유결합으로 부착된 후, 분자/가교제 복합체가 돌연변이체 단량체에 공유결합으로 부착되지만, 이관능성 가교제가 단량체에 공유결합으로 부착된 후, 이관능성 가교제/단량체 복합체가 분자에 부착되는 것이 또한 가능하다.
링커는 바람직하게는 디티오트레이톨 (DTT)에 대해 저항성이다. 적절한 링커는 아이오도아세트아미드-계 및 말레이미드-계 링커를 포함하지만, 이에 제한되지 않는다.
다른 실시양태에서, 단량체가 폴리뉴클레오티드 결합 단백질에 부착될 수 있다. 이는 본 발명의 시퀀싱 방법에서 사용될 수 있는 모듈형 시퀀싱 시스템을 형성한다. 폴리뉴클레오티드 결합 단백질은 하기에서 논의된다.
폴리뉴클레오티드 결합 단백질은 바람직하게는 돌연변이체 단량체에 공유결합으로 부착된다. 관련 기술 분야에 공지된 임의의 방법을 사용하여 단백질이 단량체에 공유결합으로 부착될 수 있다. 단량체 및 단백질이 화학적으로 융합되거나 또는 유전적으로 융합될 수 있다. 전체 구축물이 단일 폴리뉴클레오티드 서열로부터 발현되면 단량체 및 단백질이 유전적으로 융합된다. 단량체가 폴리뉴클레오티드 결합 단백질에 유전적으로 융합되는 것이 국제 출원 번호 PCT/GB09/001679 (WO 2010/004265로 공개됨)에 논의되어 있다.
폴리뉴클레오티드 결합 단백질이 시스테인 연결을 통해 부착되는 경우, 바람직하게는, 하나 이상의 시스테인이 치환에 의해 돌연변이체에 도입되어 있다. 바람직하게는, 상동체들 사이에서 낮게 보존되는 루프 영역 내로 하나 이상의 시스테인이 도입되고, 낮게 보존되는 것은 돌연변이 또는 삽입이 허용될 수 있음을 가리킨다. 따라서, 이는 폴리뉴클레오티드 결합 단백질을 부착하는데 적절하다. 이같은 실시양태에서, 위치 251의 천연-발생 시스테인이 제거될 수 있다. 상기 기술된 바와 같은 변형에 의해 시스테인 잔기의 반응성이 강화될 수 있다.
직접적으로 또는 하나 이상의 링커를 통해 폴리뉴클레오티드 결합 단백질이 돌연변이체 단량체에 부착될 수 있다. 국제 출원 번호 PCT/GB10/000132 (WO 2010/086602로 공개됨)에 기술된 혼성화 링커를 사용하여 분자가 돌연변이체 단량체에 부착될 수 있다. 대안적으로, 펩티드 링커가 사용될 수 있다. 펩티드 링커는 아미노산 서열이다. 전형적으로 펩티드 링커의 길이, 가요성 및 친수성이 링커가 단량체 및 분자의 기능을 방해하지 않도록 디자인된다. 바람직한 가요성 펩티드 링커는 2 내지 20개, 예컨대 4, 6, 8, 10 또는 16개의 세린 및/또는 글리신 아미노산의 신장물이다. 더욱 바람직한 가요성 링커는 (SG)1, (SG)2, (SG)3, (SG)4, (SG)5 및 (SG)8 [식 중 S는 세린이고, G는 글리신이다]을 포함한다. 바람직한 강직성 링커는 2 내지 30개, 예컨대 4, 6, 8, 16 또는 24개의 프롤린 아미노산의 신장물이다. 더욱 바람직한 강직성 링커는 (P)12 [식 중 P는 프롤린이다]를 포함한다.
돌연변이체 단량체가 분자 어댑터 및 폴리뉴클레오티드 결합 단백질로 화학적으로 변형될 수 있다.
분자 (단량체가 이러한 분자로 화학적으로 변형됨)는 국제 출원 번호 PCT/GB09/001690 (WO 2010/004273으로 공개됨), PCT/GB09/001679 (WO 2010/004265로 공개됨) 또는 PCT/GB10/000133 ( WO 2010/086603으로 공개됨)에 개시된 바와 같이 링커를 통해 부착될 수 있거나 또는 직접적으로 단량체에 부착될 수 있다.
예를 들어 히스티딘 잔기 (his 태그), 아스파르트산 잔기 (asp 태그), 스트렙타비딘 태그, 플래그 태그, SUMO 태그, GST 태그 또는 MBP 태그의 부가에 의해, 또는 세포로부터 분비를 촉진하는 신호 서열의 부가 (폴리펩티드가 이같은 서열을 천연적으로 함유하지 않는 경우)에 의해, 본원에 기술된 단백질, 예컨대 본 발명의 돌연변이체 단량체 및 포어 중 임의의 것이 이의 확인 또는 정제를 돕도록 변형될 수 있다. 유전적 태그를 도입하는 것에 대한 대안은 태그를 단백질 상의 천연 위치 또는 조작된 위치 상으로 반응시키는 것이다. 이의 예는 단백질 외부 상에서 조작된 시스테인에 겔-이동 시약을 반응시키는 것일 것이다. 이는 헤모리신 이종-올리고머를 분리하는 방법으로서 실연되었다 (Chem Biol. 1997 Jul;4(7):497-505).
본원에 기술된 단백질, 예컨대 본 발명의 돌연변이체 단량체 및 포어 중 임의의 것이 시현성 표지로 표지될 수 있다. 시현성 표지는 단백질이 검출되게 하는 임의의 적절한 표지일 수 있다. 적절한 표지는 형광 분자, 방사성 동위원소, 예를 들어 125l, 35S, 효소, 항체, 항원, 폴리뉴클레오티드 및 리간드 예컨대 비오틴을 포함하지만, 이에 제한되지 않는다.
본원에 기술된 단백질, 예컨대 본 발명의 돌연변이체 단량체 및 포어 중 임의의 것을 합성으로 또는 재조합 수단에 의해 제조할 수 있다. 예를 들어, 시험관내 번역 및 전사 (IVTT)에 의해 단백질을 합성할 수 있다. 비-천연 발생 아미노산을 포함하도록 또는 단백질의 안정성을 증가시키도록 단백질의 아미노산 서열이 변형될 수 있다. 단백질이 합성 수단에 의해 생산되는 경우, 생산 동안 이같은 아미노산이 도입될 수 있다. 또한 합성 생산 또는 재조합 생산 후에 단백질이 변경될 수 있다.
단백질을 D-아미노산을 사용하여 또한 생산할 수 있다. 예를 들어, 단백질이 L-아미노산 및 D-아미노산의 혼합물을 포함할 수 있다. 이는 이같은 단백질 또는 펩티드를 생산하는 분야에서 통상적이다.
단백질은, 변형이 단백질의 기능을 방해하지 않는 한, 다른 비-특이적 변형을 또한 함유할 수 있다. 다수의 비-특이적 측쇄 변형이 관련 기술 분야에 공지되어 있고, 단백질(들)의 측쇄에 이루어질 수 있다. 이같은 변형은, 예를 들어, 알데히드와 반응시킨 후 NaBH4로 환원시키는 것에 의한 아미노산의 환원적 알킬화, 메틸아세트이미데이트로의 아미딘화, 또는 아세트산 무수물로의 아실화를 포함한다.
본 발명의 단량체 및 포어를 포함하는, 본원에 기술된 단백질 중 임의의 것을 관련 기술 분야에 공지된 표준 방법을 사용하여 생산할 수 있다. 관련 기술 분야의 표준 방법을 사용하여 단백질을 코딩하는 폴리뉴클레오티드 서열을 유도하고 복제할 수 있다. 단백질을 코딩하는 폴리뉴클레오티드 서열을 관련 기술 분야의 표준 방법을 사용하여 박테리아 숙주 세포에서 발현시킬 수 있다. 재조합 발현 벡터로부터 폴리펩티드의 원위치 발현에 의해 세포 내에서 단백질을 생산할 수 있다. 발현 벡터는 폴리펩티드의 발현을 제어하도록 유도성 프로모터를 임의적으로 보유한다. 이러한 방법들이 문헌 [Sambrook, J. and Russell, D. (2001). Molecular Cloning: A Laboratory Manual, 3rd Edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY]에 기술되어 있다.
재조합 발현 후에 또는 단백질 생산 생물로부터 임의의 단백질 액체 크로마토그래피 시스템에 의해 정제한 뒤에 단백질이 대규모로 생산될 수 있다. 전형적인 단백질 액체 크로마토그래피 시스템은 FPLC, AKTA 시스템, 바이오-캐드(Bio-Cad) 시스템, 바이오-래드 바이오로직(Bio-Rad BioLogic) 시스템 및 길슨(Gilson) HPLC 시스템을 포함한다.
구축물
본 발명은 단량체 중 적어도 1개가 본 발명의 돌연변이체 단량체인, 2개 이상의 공유결합으로 부착된 CsgG 단량체를 포함하는 구축물을 또한 제공한다. 본 발명의 구축물은 포어를 형성하는 이의 능력을 유지한다. 이는 상기 논의된 바와 같이 결정될 수 있다. 하나 이상의 본 발명의 구축물이 폴리뉴클레오티드를 특성화하기 위한, 예컨대 시퀀싱하기 위한 포어를 형성하는데 사용될 수 있다. 구축물은 적어도 2개, 적어도 3개, 적어도 4개, 적어도 5개, 적어도 6개, 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체를 포함할 수 있다. 구축물은 바람직하게는 2개의 단량체를 포함한다. 2개 이상의 단량체는 동일하거나 상이할 수 있다.
구축물 내의 적어도 1개의 단량체가 본 발명의 돌연변이체 단량체이다. 구축물 내의 2개 이상, 3개 이상, 4개 이상, 5개 이상, 6개 이상, 7개 이상, 8개 이상, 9개 이상 또는 10개 이상의 단량체가 본 발명의 돌연변이체 단량체일 수 있다. 바람직하게는 구축물 내의 모든 단량체가 본 발명의 돌연변이체 단량체이다. 돌연변이체 단량체들은 동일하거나 상이할 수 있다. 바람직한 실시양태에서, 구축물은 2개의 본 발명의 돌연변이체 단량체를 포함한다.
구축물 내의 본 발명의 돌연변이체 단량체들은 바람직하게는 대략적으로 동일한 길이이거나 또는 동일한 길이이다. 구축물 내의 본 발명의 돌연변이체 단량체의 배럴들은 바람직하게는 대략적으로 동일한 길이이거나 또는 동일한 길이이다. 길이는 아미노산의 개수 및/또는 길이 단위로 측정될 수 있다.
구축물은 본 발명의 돌연변이체 단량체가 아닌 1개 이상의 단량체를 포함할 수 있다. 본 발명의 돌연변이체 단량체가 아닌 CsgG 돌연변이체 단량체는 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429, 또는 상기 논의된 아미노산/위치 중 어느 것도 돌연변이되지 않은 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 비교 변이체를 포함하는 단량체를 포함한다. 구축물 내의 적어도 1개의 단량체가 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429, 또는 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429에 제시된 서열의 비교 변이체를 포함할 수 있다. 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 비교 변이체는 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429, 40 또는 41에 대해 이의 전체 서열에 걸쳐 아미노산 동일성을 기초로 적어도 50% 상동성이다. 더욱 바람직하게는, 비교 변이체는 아미노산 동일성을 기초로 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 아미노산 서열에 대해 전체 서열에 걸쳐 적어도 55%, 적어도 60%, 적어도 65%, 적어도 70%, 적어도 75%, 적어도 80%, 적어도 85%, 적어도 90%, 더욱 바람직하게는 적어도 95%, 97% 또는 99% 상동성일 수 있다.
구축물 내의 단량체들은 바람직하게는 유전적으로 융합된다. 전체 구축물이 단일 폴리뉴클레오티드 서열로부터 발현되면 단량체들이 유전적으로 융합된다. 구축물을 코딩하는 단일 폴리뉴클레오티드 서열을 형성하도록 단량체에 대한 코딩 서열들이 임의의 방식으로 조합될 수 있다.
단량체들은 임의의 형상으로 유전적으로 융합될 수 있다. 단량체들은 이의 말단 아미노산을 통해 융합될 수 있다. 예를 들어, 1개의 단량체의 아미노 말단이 또 다른 단량체의 카르복시 말단에 융합될 수 있다. 구축물 내의 (아미노 → 카르복시 방향의) 제2 및 후속 단량체는 이의 아미노 말단 끝부분 (각각이 이전 단량체의 카르복시 말단에 융합된다)에 메티오닌을 포함할 수 있다. 예를 들어, M이 (아미노 말단 메티오닌이 없는) 단량체이고, mM이 아미노 말단 메티오닌이 있는 단량체인 경우, 구축물은 M-mM, M-mM-mM 또는 M-mM-mM-mM의 서열을 포함할 수 있다. 전형적으로 이러한 메티오닌의 존재는 전체 구축물을 코딩하는 폴리뉴클레오티드 내의 제2 또는 후속 단량체를 코딩하는 폴리뉴클레오티드의 5' 끝부분의 개시 코돈 (즉, ATG)의 발현으로부터 초래된다. 구축물 내의 (아미노 → 카르복시 방향의) 제1 단량체 또한 메티오닌을 포함할 수 있다 (예를 들어 mM-mM, mM-mM-mM 또는 mM-mM-mM-mM).
2개 이상의 단량체가 직접적으로 함께 유전적으로 융합될 수 있다. 바람직하게는 단량체들이 링커를 사용하여 유전적으로 융합된다. 링커는 단량체의 이동성을 구속하도록 디자인될 수 있다. 바람직한 링커는 아미노산 서열 (즉 펩티드 링커)이다. 상기 논의된 펩티드 링커 중 임의의 것이 사용될 수 있다.
또 다른 바람직한 실시양태에서, 단량체들은 화학적으로 융합된다. 2개의 부분이, 예를 들어 화학적 가교제를 통해, 화학적으로 부착되면, 2개의 단량체가 화학적으로 융합된다. 상기 논의된 화학적 가교제 중 임의의 것이 사용될 수 있다. 본 발명의 돌연변이체 단량체 내로 도입된 하나 이상의 시스테인 잔기에 링커가 부착될 수 있다. 대안적으로, 구축물 내의 단량체 중 하나의 말단에 링커가 부착될 수 있다.
구축물이 상이한 단량체들을 함유하는 경우, 링커 농도를 단량체에 비해 크게 과량으로 유지시킴으로써 단량체들이 자신에게 가교되는 것을 방지할 수 있다. 대안적으로, 2개의 링커가 사용되는 "자물쇠 & 열쇠(lock and key)" 배열을 사용할 수 있다. 각각의 링커의 한쪽 끝부분만 함께 반응하여 더 긴 링커를 형성할 수 있고, 링커의 다른쪽 끝부분은 상이한 단량체와 각각 반응한다. 이같은 링커가 국제 출원 번호 PCT/GB10/000132 (WO 2010/086602로 공개됨)에 기술되어 있다.
폴리뉴클레오티드
본 발명은 본 발명의 돌연변이체 단량체를 코딩하는 폴리뉴클레오티드 서열을 또한 제공한다. 돌연변이체 단량체는 상기 논의된 것 중 임의의 것일 수 있다. 바람직하게는 폴리뉴클레오티드 서열은 뉴클레오티드 동일성을 기초로 서열식별번호: 389의 서열에 대해 전체 서열에 걸쳐 적어도 50%, 60%, 70%, 80%, 90% 또는 95% 상동성인 서열을 포함한다. 300개 이상, 예를 들어 375, 450, 525 또는 600개 또는 이를 초과하는 개수의 연속적인 뉴클레오티드의 신장물에 걸쳐 적어도 80%, 예를 들어 적어도 85%, 90% 또는 95%의 뉴클레오티드 동일성이 있을 수 있다 ("강한 상동성"). 상기 기술된 바와 같이 상동성을 계산할 수 있다. 폴리뉴클레오티드 서열은 유전자 코드의 축퇴성을 기초로 서열식별번호: 389과 상이한 서열을 포함할 수 있다.
본 발명은 본 발명의 유전적으로 융합된 구축물 중 임의의 것을 코딩하는 폴리뉴클레오티드 서열을 또한 제공한다. 바람직하게는 폴리뉴클레오티드는 서열식별번호: 389에 제시된 서열의 2개 이상의 변이체를 포함한다. 바람직하게는 폴리뉴클레오티드 서열은 뉴클레오티드 동일성을 기초로 서열식별번호: 389에 대해 전체 서열에 걸쳐 적어도 50%, 60%, 70%, 80%, 90% 또는 95% 상동성인 2개 이상의 서열을 포함한다. 600개 이상, 예를 들어 750, 900, 1050 또는 1200개 또는 이를 초과하는 개수의 연속적인 뉴클레오티드의 신장물에 걸쳐 적어도 80%, 예를 들어 적어도 85%, 90% 또는 95%의 뉴클레오티드 동일성이 있을 수 있다 ("강한 상동성"). 상기 기술된 바와 같이 상동성을 계산할 수 있다.
관련 기술 분야의 표준 방법을 사용하여 폴리뉴클레오티드 서열을 유도 및 복제할 수 있다. 야생형 CsgG를 코딩하는 염색체 DNA를 포어 생산 생물, 예컨대 에스케리키아 콜라이로부터 추출할 수 있다. 포어 서브유닛을 코딩하는 유전자를 특이적인 프라이머를 수반하는 PCR을 사용하여 증폭시킬 수 있다. 그 후, 증폭된 서열에 부위-지정 돌연변이유발을 진행시킬 수 있다. 적절한 부위-지정 돌연변이유발 방법이 관련 기술 분야에 공지되어 있고, 예를 들어, 조합 연쇄 반응을 포함한다. 널리 공지된 기술, 예컨대 문헌 [Sambrook, J. and Russell, D. (2001). Molecular Cloning: A Laboratory Manual, 3rd Edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY]에 기술된 것을 사용하여 본 발명의 구축물을 코딩하는 폴리뉴클레오티드를 제조할 수 있다.
그 후, 생성된 폴리뉴클레오티드 서열을 복제가능한 재조합 벡터 예컨대 클로닝 벡터 내로 혼입시킬 수 있다. 이러한 벡터를 사용하여 상용성 숙주 세포에서 폴리뉴클레오티드를 복제할 수 있다. 따라서, 폴리뉴클레오티드를 복제가능한 벡터 내로 도입하고, 벡터를 상용성 숙주 세포 내로 도입하고, 숙주 세포를 벡터의 복제를 초래하는 조건 하에 성장시킴으로써, 폴리뉴클레오티드 서열을 제조할 수 있다. 벡터를 숙주 세포로부터 회수할 수 있다. 폴리뉴클레오티드의 클로닝을 위한 적절한 숙주 세포가 관련 기술 분야에 공지되어 있고, 하기에서 더욱 상세하게 기술된다.
폴리뉴클레오티드 서열을 적절한 발현 벡터 내로 클로닝할 수 있다. 발현 벡터에서, 전형적으로 폴리뉴클레오티드 서열은 숙주 세포에 의한 코딩 서열의 발현을 제공할 수 있는 제어 서열에 작동가능하게 연결된다. 이같은 발현 벡터를 사용하여 포어 서브유닛을 발현시킬 수 있다.
용어 "작동가능하게 연결된"은 기술된 성분들이 이들이 이의 의도된 방식으로 기능하도록 허용하는 관계에 있는 병치를 지칭한다. 코딩 서열에 "작동가능하게 연결된" 제어 서열은 제어 서열과 상용성인 조건 하에 코딩 서열의 발현이 달성되는 방식으로 라이게이션된다. 동일하거나 상이한 폴리뉴클레오티드 서열의 다중 카피가 벡터 내로 도입될 수 있다.
그 후, 발현 벡터를 적절한 숙주 세포 내로 도입할 수 있다. 따라서, 폴리뉴클레오티드 서열을 발현 벡터 내로 삽입하고, 벡터를 상용성 박테리아 숙주 세포 내로 도입하고, 숙주 세포를 폴리뉴클레오티드 서열의 발현을 초래하는 조건 하에 성장시킴으로써 본 발명의 돌연변이체 단량체 또는 구축물을 생산할 수 있다. 재조합으로 발현된 단량체 또는 구축물이 숙주 세포 막에서 포어 내로 자가-조립될 수 있다. 대안적으로, 이러한 방식으로 생산된 재조합 포어가 숙주 세포로부터 제거되어, 또 다른 막 내로 삽입될 수 있다. 적어도 2개의 상이한 단량체 또는 구축물을 포함하는 숙주를 생산하는 경우, 상이한 단량체 또는 구축물들이 상기 기술된 바와 같은 상이한 숙주 세포들에서 발현되고, 숙주 세포로부터 제거되고, 별도의 막, 예컨대 토끼 세포 막 또는 합성 막에서 포어 내로 조립될 수 있다.
벡터는, 예를 들어, 복제 기원, 상기 폴리뉴클레오티드 서열의 발현을 위한 임의적인 프로모터, 및 임의적인 프로모터 조절인자가 제공된 플라스미드, 바이러스 또는 파지 벡터일 수 있다. 벡터는 하나 이상의 선별가능한 마커 유전자, 예를 들어 테트라사이클린 저항성 유전자를 함유할 수 있다. 발현 벡터에 이에 대해 디자인되는 숙주 세포와 상용성이도록 프로모터 및 기타 발현 조절 신호가 선택될 수 있다. T7, trc, lac, ara 또는 λL 프로모터가 전형적으로 사용된다.
숙주 세포는 전형적으로 단량체 또는 구축물을 높은 수준으로 발현한다. 폴리뉴클레오티드 서열로 형질전환되는 숙주 세포는 세포를 형질전환시키는데 사용된 발현 벡터와 상용성이도록 선택될 것이다. 숙주 세포는 전형적으로 박테리아성이고, 바람직하게는 에스케리키아 콜라이이다. λ DE3 용원이 있는 임의의 세포, 예를 들어 C41 (DE3), BL21 (DE3), JM109 (DE3), B834 (DE3), TUNER, 오리가미(Origami) 및 오리가미 B가 T7 프로모터를 포함하는 벡터를 발현할 수 있다. 상기 열거된 조건에 더하여, 문헌 [Cao et al., 2014, PNAS, Structure of the nonameric bacterial amyloid secretion channel, doi - 1411942111] 및 [Goyal et al., 2014, Nature, 516, 250-253 structural and mechanistic insights into the bacterial amyloid secretion channel CsgG]에 언급된 방법 중 임의의 것을 사용하여 CsgG 단백질을 발현시킬 수 있다.
본 발명은 본 발명의 돌연변이체 단량체 또는 본 발명의 구축물을 생산하는 방법을 또한 포함한다. 이러한 방법은 본 발명의 폴리뉴클레오티드를 적절한 숙주 세포에서 발현시키는 것을 포함한다. 폴리뉴클레오티드는 바람직하게는 벡터의 일부분이고, 바람직하게는 프로모터에 작동가능하게 연결된다.
포어
본 발명은 다양한 포어를 또한 제공한다. 상이한 뉴클레오티드들을 높은 감도로 판별할 수 있기 때문에 본 발명의 포어는 폴리뉴클레오티드 서열을 특성화하는데, 예컨대 시퀀싱하는데 이상적이다. 놀랍게도 포어는 DNA 및 RNA 내의 4개의 뉴클레오티드를 구별할 수 있다. 심지어 본 발명의 포어는 메틸화 뉴클레오티드와 비-메틸화 뉴클레오티드를 구별할 수 있다. 본 발명의 포어의 염기 해상도가 놀랍게 높다. 포어는 4개의 DNA 뉴클레오티드 모두의 거의 완전한 분리를 나타낸다. 추가로 포어는 포어 내에서의 체류 시간 및 포어를 통해 흐르는 전류를 기초로 데옥시시티딘 모노포스페이트 (dCMP)와 메틸-dCMP를 판별한다.
본 발명의 포어는 광범위한 조건 하에 상이한 뉴클레오티드들을 또한 판별할 수 있다. 특히, 포어는 핵산의 특성화, 예컨대 시퀀싱에 유리한 조건 하에 뉴클레오티드들을 판별할 것이다. 인가된 전위, 염 농도, 완충제, 온도, 및 첨가제 예컨대 요소, 베타인 및 DTT의 존재를 변경시킴으로써 본 발명의 포어가 상이한 뉴클레오티드들을 판별할 수 있는 정도를 제어할 수 있다. 이는, 특히 시퀀싱의 경우에, 포어의 기능을 미세-조율하는 것을 허용한다. 이는 하기에서 더욱 상세하게 논의된다. 본 발명의 포어는 뉴클레오티드 하나씩을 기초로 하는 것보다는 하나 이상의 단량체와의 상호작용으로부터 폴리뉴클레오티드 중합체를 확인하는데 또한 사용될 수 있다.
본 발명의 포어는 단리되거나, 실질적으로 단리되거나, 정제되거나 또는 실질적으로 정제될 수 있다. 임의의 다른 성분, 예컨대 지질 또는 기타 포어가 전혀 없으면 본 발명의 포어가 단리 또는 정제된 것이다. 포어의 의도된 용도를 방해하지 않을 담체 또는 희석제와 혼합되면 포어가 실질적으로 단리된 것이다. 예를 들어, 10% 미만, 5% 미만, 2% 미만 또는 1% 미만의 다른 성분, 예컨대 3-블럭 공중합체, 지질 또는 기타 포어를 포함하는 형태로 존재하면 포어가 실질적으로 단리되거나 또는 실질적으로 정제된 것이다. 대안적으로, 본 발명의 포어가 막 내에 존재할 수 있다. 적절한 막이 하기에서 논의된다.
본 발명의 포어는 개별적 또는 단일 포어로서 존재할 수 있다. 대안적으로, 본 발명의 포어는 2개 이상의 포어의 동종성 또는 이종성 집단 내에 존재할 수 있다.
동종-올리고머
포어
본 발명은 동일한 본 발명의 돌연변이체 단량체들을 포함하는 CsgG로부터 유래된 동종-올리고머 포어를 또한 제공한다. 동종-올리고머 포어는 본 발명의 돌연변이체 중 임의의 것을 포함할 수 있다. 본 발명의 동종-올리고머 포어는 폴리뉴클레오티드의 특성화, 예컨대 시퀀싱에 이상적이다. 본 발명의 동종-올리고머 포어는 상기 논의된 장점 중 임의의 것을 가질 수 있다.
동종-올리고머 포어는 임의 개수의 돌연변이체 단량체를 함유할 수 있다. 포어는 전형적으로 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 동일한 돌연변이체 단량체, 예컨대 7, 8, 9 또는 10개의 돌연변이체 단량체를 포함한다. 포어는 바람직하게는 8개 또는 9개의 동일한 돌연변이체 단량체를 포함한다. 바람직하게는, 1개 이상, 예컨대 2, 3, 4, 5, 6, 7, 8, 9 또는 10개의 돌연변이체 단량체가 상기 논의된 바와 같이 화학적으로 변형된다.
포어의 제조 방법이 하기에서 더욱 상세하게 논의된다.
이종-올리고머
포어
본 발명은 적어도 1개의 본 발명의 돌연변이체 단량체를 포함하는 CsgG로부터 유래된 이종-올리고머 포어를 또한 제공한다. 본 발명의 이종-올리고머 포어는 폴리뉴클레오티드의 특성화, 예컨대 시퀀싱에 이상적이다. 관련 기술 분야에 공지된 방법 (예를 들어 문헌 [Protein Sci. 2002 Jul; 11(7):1813-24])을 사용하여 이종-올리고머 포어를 제조할 수 있다.
이종-올리고머 포어는 포어를 형성하기 위해 충분한 단량체를 포함한다. 단량체는 임의의 유형의 것일 수 있다. 포어는 전형적으로 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함한다. 포어는 바람직하게는 8개 또는 9개의 단량체를 포함한다.
바람직한 실시양태에서, 모든 단량체 (예컨대 단량체 중 10, 9, 8 또는 7개)가 본 발명의 돌연변이체 단량체이고, 이들 중 적어도 1개가 다른 것들과 상이하다. 더욱 바람직한 실시양태에서, 포어는 8개 또는 9개의 본 발명의 돌연변이체 단량체를 포함하고, 이들 중 적어도 1개가 다른 것들과 상이하다. 이들은 모두 서로 상이할 수 있다.
포어 내의 본 발명의 돌연변이체 단량체들은 바람직하게는 대략 동일한 길이이거나 또는 동일한 길이이다. 포어 내의 본 발명의 돌연변이체 단량체의 배럴들은 바람직하게는 대략적으로 동일한 길이이거나 또는 동일한 길이이다. 길이는 아미노산의 개수 및/또는 길이 단위로 측정될 수 있다.
또 다른 바람직한 실시양태에서, 돌연변이체 단량체 중 적어도 1개가 본 발명의 돌연변이체 단량체가 아니다. 이러한 실시양태에서, 나머지 단량체들은 바람직하게는 본 발명의 돌연변이체 단량체이다. 따라서, 포어는 9, 8, 7, 6, 5, 4, 3, 2 또는 1개의 본 발명의 돌연변이체 단량체를 포함할 수 있다. 포어 내의 임의 개수의 단량체가 본 발명의 돌연변이체 단량체가 아닐 수 있다. 바람직하게는 포어는 7개 또는 8개의 본 발명의 돌연변이체 단량체 및 본 발명의 단량체가 아닌 단량체 1개를 포함한다. 본 발명의 돌연변이체 단량체들은 동일하거나 상이할 수 있다.
구축물 내의 본 발명의 돌연변이체 단량체들은 바람직하게는 대략적으로 동일한 길이이거나 또는 동일한 길이이다. 구축물 내의 본 발명의 돌연변이체 단량체의 배럴들은 바람직하게는 대략적으로 동일한 길이이거나 또는 동일한 길이이다. 길이는 아미노산의 개수 및/또는 길이 단위로 측정될 수 있다.
포어는 본 발명의 돌연변이체 단량체가 아닌 1개 이상의 단량체를 포함할 수 있다. 본 발명의 돌연변이체 단량체가 아닌 CsgG 단량체는 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429, 또는 본 발명과 관련하여 상기 논의된 아미노산/위치 중 어느 것도 돌연변이/치환되지 않은 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 비교 변이체를 포함하는 단량체를 포함한다. 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 비교 변이체는 전형적으로 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429에 대해 이의 전체 서열에 걸쳐 아미노산 동일성을 기초로 적어도 50% 상동성이다. 더욱 바람직하게는, 비교 변이체는 아미노산 동일성을 기초로 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 아미노산 서열에 대해 전체 서열에 걸쳐 적어도 55%, 적어도 60%, 적어도 65%, 적어도 70%, 적어도 75%, 적어도 80%, 적어도 85%, 적어도 90%, 더욱 바람직하게는 적어도 95%, 97% 또는 99% 상동성일 수 있다.
상기 논의된 모든 실시양태에서, 바람직하게는, 1개 이상, 예컨대 2, 3, 4, 5, 6, 7, 8, 9 또는 10개의 돌연변이체 단량체가 상기 논의된 바와 같이 화학적으로 변형된다.
포어의 제조 방법이 하기에서 더욱 상세하게 논의된다.
구축물을 함유하는
포어
본 발명은 적어도 1개의 본 발명의 구축물을 포함하는 포어를 또한 제공한다. 본 발명의 구축물은 단량체 중 적어도 1개가 본 발명의 돌연변이체 단량체인, CsgG로부터 유래된 2개 이상의 공유결합으로 부착된 단량체를 포함한다. 달리 말하면, 구축물은 1개를 초과하는 단량체를 함유하여야 한다. 포어는 포어를 형성하기 위해 충분한 구축물 및 단량체 (필요하다면)를 함유한다. 예를 들어, 8량체 포어는 (a) 각각 2개의 구축물을 포함하는 구축물 4개, (b) 각각 4개의 단량체를 포함하는 구축물 2개, 또는 (b) 2개의 단량체를 포함하는 구축물 1개, 및 구축물의 일부를 형성하지 않는 6개의 단량체를 포함할 수 있다. 예를 들어, 9량체 포어는 (a) 각각 2개의 구축물을 포함하는 구축물 4개, 및 구축물의 일부를 형성하지 않는 1개의 단량체, (b) 각각 4개의 단량체를 포함하는 구축물 2개, 및 구축물의 일부를 형성하지 않는 1개의 단량체, 또는 (b) 2개의 단량체를 포함하는 구축물 1개, 및 구축물의 일부를 형성하지 않는 7개의 단량체를 포함할 수 있다. 통상의 기술자는 구축물 및 단량체의 기타 조합을 구상할 수 있다.
포어 내의 단량체 중 적어도 2개가 본 발명의 구축물의 형태일 수 있다. 구축물, 및 따라서 포어는 적어도 1개의 본 발명의 돌연변이체 단량체를 포함한다. 전형적으로 포어는 총괄하여 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함한다 (이들 중 적어도 2개가 구축물 내에 있어야 한다). 바람직하게는 포어는 8개 또는 9개의 단량체를 포함한다 (이들 중 적어도 2개가 구축물 내에 있어야 한다).
구축물을 함유하는 포어는 동종-올리고머일 수 있거나 (즉, 동일한 구축물들을 포함함), 또는 이종-올리고머일 수 있다 (즉, 적어도 1개의 구축물이 다른 것들과 상이한 경우).
전형적으로 포어는 (a) 2개의 단량체를 포함하는 구축물 1개, 및 (b) 5, 6, 7 또는 8개의 단량체를 함유한다. 구축물은 상기 논의된 것들 중 임의의 것일 수 있다. 단량체는 본 발명의 돌연변이체 단량체, 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429를 포함하는 단량체, 및 상기 논의된 바와 같은 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 비교 변이체를 포함하는 돌연변이체 단량체를 포함하여, 상기 논의된 것들 중 임의의 것일 수 있다.
또 다른 전형적인 포어는 1개를 초과하는 본 발명의 구축물, 예컨대 2개, 3개 또는 4개의 본 발명의 구축물을 포함한다. 필요하다면, 이같은 포어는 포어를 형성하기 위해 충분한 추가 단량체 또는 구축물을 추가로 포함할 수 있다. 추가 단량체(들)는 본 발명의 돌연변이체 단량체, 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429를 포함하는 단량체, 및 상기 논의된 바와 같은 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 비교 변이체를 포함하는 돌연변이체 단량체를 포함하여, 상기 논의된 것들 중 임의의 것일 수 있다. 추가 구축물(들)은 상기 논의된 것들 중 임의의 것일 수 있거나, 또는 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429 또는 상기 논의된 바와 같은 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429의 비교 변이체를 포함하는 단량체를 각각 포함하는 2개 이상의 공유결합으로 부착된 CsgG 단량체를 포함하는 구축물일 수 있다.
추가적인 본 발명의 포어는 2개의 단량체를 포함하는 구축물만 포함하고, 예를 들어, 포어는 2개의 단량체를 포함하는 구축물 4, 5, 6, 7 또는 8개를 포함할 수 있다. 적어도 1개의 구축물이 본 발명의 구축물이고, 즉 적어도 1개의 구축물 내의 적어도 1개의 단량체, 바람직하게는 적어도 1개의 구축물 내의 각각의 단량체가 본 발명의 돌연변이체 단량체이다. 2개의 단량체를 포함하는 구축물 모두가 본 발명의 구축물일 수 있다.
본 발명의 따른 특정 포어는 2개의 단량체를 각각 포함하는 본 발명의 구축물 4개를 포함하고, 여기서 각각의 구축물 내의 적어도 1개의 단량체, 바람직하게는 각각의 구축물 내의 각각의 단량체가 본 발명의 돌연변이체 단량체이다. 각각의 구축물의 1개의 단량체만 포어의 채널에 기여하는 구조의 포어 내로 구축물이 올리고머화될 수 있다. 전형적으로는, 구축물의 다른 단량체는 포어 채널의 외부에 있을 것이다. 예를 들어, 본 발명의 포어는 채널이 7, 8, 9 또는 10개의 단량체를 포함하는, 2개의 단량체를 포함하는 구축물 7, 8, 9 또는 10개를 포함할 수 있다.
상기 기술된 바와 같이 구축물 내로 돌연변이가 도입될 수 있다. 돌연변이들이 교대성일 수 있고, 즉, 2개의 단량체 구축물 내의 각각의 단량체에 대해 돌연변이들이 상이하고, 구축물이 동종-올리고머로서 조립되어, 교대 변형이 초래된다. 달리 말하면, MutA 및 MutB를 포함하는 단량체들이 융합 및 조립되어 A-B:A-B:A-B:A-B 포어를 형성한다. 대안적으로, 돌연변이들이 이웃할 수 있고, 즉, 동일한 돌연변이들이 구축물 내의 2개의 단량체 내로 도입되고, 그 후 이들이 상이한 돌연변이체 단량체 또는 구축물과 함께 올리고머화된다. 달리 말하면, MutA를 포함하는 단량체들이 융합된 후, MutB-함유 단량체와 함께 올리고머화되어 A-A:B:B:B:B:B:B를 형성한다.
구축물을 함유하는 포어 내의 본 발명의 단량체 중 하나 이상이 상기 논의된 바와 같이 화학적으로 변형될 수 있다.
피분석물
특성화
본 발명은 표적 피분석물의 존재, 부재 또는 하나 이상의 특성을 결정하는 방법을 제공한다. 이러한 방법은 표적 피분석물이 포어에 대해, 예컨대 포어를 통해 이동하도록 표적 피분석물을 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉시키는 단계 및 피분석물이 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 이에 의해 피분석물의 존재, 부재 또는 하나 이상의 특성을 결정하는 단계를 수반한다. 표적 피분석물은 주형 피분석물 또는 관심 피분석물로 또한 칭해질 수 있다.
이러한 방법은 표적 피분석물이 포어를 통해 이동하도록 표적 피분석물을 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉시키는 것을 포함한다. 포어는 전형적으로 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함한다. 포어는 바람직하게는 8개 또는 9개의 동일한 단량체를 포함한다. 바람직하게는, 단량체 중 1개 이상, 예컨대 2, 3, 4, 5, 6, 7, 8, 9 또는 10개가 상기 논의된 바와 같이 화학적으로 변형된다.
CsgG 포어는 임의의 생물로부터 유래될 수 있다. CsgG 포어는 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429에 제시된 서열을 포함하는 단량체를 포함할 수 있다. CsgG 포어는 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429에 제시된 서열을 각각 포함하는, 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함할 수 있다 (즉, 포어는 동일한 생물로부터의 동일한 단량체들을 포함하는 동종-올리고머이다). CsgG는 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429에 제시된 서열을 각각 포함하는 단량체의 임의의 조합을 포함할 수 있다. 예를 들어, 포어는 서열식별번호: 390에 제시된 서열을 포함하는 단량체 7개 및 서열식별번호: 391에 제시된 서열을 포함하는 단량체 2개를 포함할 수 있다.
CsgG 돌연변이체는 임의 개수의 돌연변이체 단량체, 예컨대 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함할 수 있다. 돌연변이체 단량체는 서열식별번호: 390, 391, 392, 393, 394, 395, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428 또는 429에 제시된 서열의 비교 변이체를 포함할 수 있다. 상기에 비교 변이체가 논의되어 있다. 비교 변이체는 포어를 형성할 수 있어야 하고, 본 발명의 포어와 관련하여 상기에서 논의된 % 상동성 중 임의의 것을 가질 수 있다.
CsgG 돌연변이체는 바람직하게는 9개의 단량체를 포함하고, 단량체 중 적어도 1개가 (a) 하기 위치 N40, Q42, D43, E44, K49, Y51, S54, N55, F56, S57, Q62, E101, N102, E124, E131, R142, D149, T150, E185, R192, D195, E201 및 E203 중 하나 이상에서의 돌연변이, 예컨대 하기 위치 N40, Q42, D43, E44, K49, Y51, S54, N55, F56, S57, Q62, E101, N102, E131, D149, T150, E185, D195, E201 및 E203 중 하나 이상에서의 돌연변이 또는 N40, Q42, D43, E44, K49, Y51, N55, F56, E101, N102, E131, D149, T150, E185, D195, E201 및 E203 중 하나 이상에서의 하나 이상의 돌연변이; 및/또는 (b) 하기 위치 F48, K49, P50, Y51, P52, A53, S54, N55, F56 및 S57 중 하나 이상에서의 결실을 포함하는 서열식별번호: 390에 제시된 서열의 변이체이다. 변이체는 (a); (b); 또는 (a) 및 (b)를 포함할 수 있다. (a)에서, 임의 개수 및 조합의 위치 N40, Q42, D43, E44, K49, Y51, S54, N55, F56, S57, Q62, E101, N102, R124, E131, R142, D149, E185, R192, D195, E201 및 E203이 돌연변이될 수 있다. 상기 논의된 바와 같이, Y51, N55 및 F56 중 하나 이상을 돌연변이시키는 것은 폴리뉴클레오티드가 포어를 통해 이동할 때 전류에 기여하는 뉴클레오티드의 개수를 감소시킬 수 있고, 이에 의해, 폴리뉴클레오티드가 포어를 통해 이동할 때의 관찰된 전류와 폴리뉴클레오티드 사이의 직접적인 관계를 확인하는 것을 더 용이하게 할 수 있다.
(b)에서, 임의 개수 및 조합의 F48, K49, P50, Y51, P52, A53, S54, N55, F56 및 S57이 결실될 수 있다. 변이체는 본 발명의 돌연변이체 단량체와 관련하여 상기에서 논의된 특정 돌연변이/치환 또는 이의 조합 중 임의의 것을 포함할 수 있다.
본 발명의 방법에서 사용하기 위한 바람직한 변이체는 하기 치환 중 하나 이상을 포함한다: (a) F56N, F56Q, F56R, F56S, F56G, F56A 또는 F56K 또는 F56A, F56P, F56R, F56H, F56S, F56Q, F56I, F56L, F56T 또는 F56G; (b) N55Q, N55R, N55K, N55S, N55G, N55A 또는 N55T; (c) Y51L, Y51V, Y51A, Y51N, Y51Q, Y51S 또는 Y51G; (d) T150I; (e) S54P; 및 (f) S57P. 변이체는 임의 개수 및 조합의 (a) 내지 (f)를 포함할 수 있다.
본 발명의 방법에서 사용하기 위한 바람직한 변이체는 Q62R 또는 Q62K를 포함한다.
본 발명의 방법에서 사용하기 위한 바람직한 변이체는 D43, E44, Q62에서의 돌연변이 또는 이의 임의의 조합, 예컨대 D43, E44, Q62, D43/E44, D43/Q62, E44/Q62 또는 D43/E44/Q62를 포함한다.
변이체는 하기 위치에서의 돌연변이를 포함할 수 있다:
Y51, F56, D149, E185, E201 및 E203, 예컨대 Y51N, F56A, D149N, E185R, E201N 및 E203N;
N55, 예컨대 N55A 또는 N55S;
Y51, 예컨대 Y51N 또는 Y51T;
S54, 예컨대 S54P;
S57, 예컨대 S57P;
F56, 예컨대 F56N, F56Q, F56R, F56S, F56G, F56A 또는 F56K 또는 F56A, F56P, F56R, F56H, F56S, F56Q, F56I, F56L, F56T 또는 F56G;
Y51및 F56, 예컨대 Y51A 및 F56A, Y51A 및 F56N, Y51I 및 F56A, Y51L 및 F56A, Y51T 및 F56A, Y51T 및 F56Q, Y51I 및 F56N, Y51L 및 F56N 또는 Y51T 및 F56N, 바람직하게는, Y51I 및 F56A, Y51L 및 F56A 또는 Y51T 및 F56A, 더욱 바람직하게는 Y51T 및 F56Q, 더욱 바람직하게는 Y51X 및 F56Q [식 중 X는 임의의 아미노산이다];
N55 및 F56, 예컨대 N55X 및 F56Q [식 중 X는 임의의 아미노산이다];
Y51, N55 및 F56, 예컨대 Y51A, N55S 및 F56A, Y51A, N55S 및 F56N 또는 Y51T, N55S 및 F56Q;
S54 및 F56, 예컨대 S54P 및 F56A 또는 S54P 및 F56N;
F56 및 S57, 예컨대 F56A 및 S57P 또는 F56N 및 S57P;
D149, E185 및 E203, 예컨대 D149N, E185N 및 E203N;
D149, E185, E201 및 E203, 예컨대 D149N, E185N, E201N 및 E203N;
D149, E185, D195, E201 및 E203, 예컨대 D149N, E185R, D195N, E201N 및 E203N 또는 D149N, E185R, D195N, E201R 및 E203N;
F56 및 N102, 예컨대 F56Q 및 N102R;
(a) Q62, 예컨대 Q62R 또는 Q62K, 및 (b) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56, 예컨대 Y51T/F56Q/Q62R;
(i) D43, E44, Q62 또는 이의 임의의 조합, 예컨대 D43, E44, Q62, D43/E44, D43/Q62, E44/Q62 또는 D43/E44/Q62 및 (ii) Y51, N55 및 F56 중 하나 이상, 예컨대 Y51, N55, F56, Y51/N55, Y51/F56, N55/F56 또는 Y51/N55/F56, 예컨대 D43N/Y51T/F56Q, E44N/Y51T/F56Q, D43N/E44N/Y51T/F56Q, D43N/Y51T/F56Q/Q62R, E44N/Y51T/F56Q/Q62R 또는 D43N/E44N/Y51T/F56Q/Q62R; 또는
T150, 예컨대 T150I.
본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 하기 치환 중 하나 이상을 포함하는 서열식별번호: 390의 변이체를 포함한다: (a) F56N, F56Q, F56R, F56S, F56G, F56A 또는 F56K 또는 F56A, F56P, F56R, F56H, F56S, F56Q, F56I, F56L, F56T 또는 F56G; (b) N55Q, N55R, N55K, N55S, N55G, N55A 또는 N55T; (c) Y51L, Y51V, Y51A, Y51N, Y51Q, Y51S 또는 Y51G; (d) T150I; (e) S54P; 및 (f) S57P. 변이체는 임의 개수 및 조합의 (a) 내지 (f)를 포함할 수 있다. 바람직하게는 이러한 바람직한 포어에서 단량체들이 동일하다.
본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 하기를 포함하는 서열식별번호: 390의 변이체를 포함한다:
Y51N, F56A, D149N, E185R, E201N 및 E203N;
N55A;
N55S;
Y51N;
S54P;
S57P;
F56N, F56Q, F56R, F56S, F56G, F56A 또는 F56K;
F56A, F56P, F56R, F56H, F56S, F56Q, F56I, F56L, F56T 또는 F56G;
Y51A 및 F56A;
Y51A 및 F56N;
Y51I 및 F56A;
Y51L 및 F56A;
Y51T 및 F56A;
Y51T 및 F56Q;
Y51I 및 F56N;
Y51L 및 F56N;
Y51T 및 F56N;
N55S 및 F56Q;
Y51A, N55S 및 F56A;
Y51A, N55S 및 F56N;
Y51T, N55S 및 F56Q;
S54P 및 F56A;
S54P 및 F56N;
F56A 및 S57P;
F56N 및 S57P;
D149N, E185N 및 E203N;
D149N, E185N, E201N 및 E203N;
D149N, E185R, D195N, E201N 및 E203N;
D149N, E185R, D195N, E201R 및 E203N;
T150I;
F56Q 및 N102R;
F56 및 N102, 예컨대 F56Q 및 N102R;
Y51T/F56Q/Q62R;
D43N/Y51T/F56Q;
E44N/Y51T/F56Q;
D43N/E44N/Y51T/F56Q
D43N/Y51T/F56Q/Q62R;
E44N/Y51T/F56Q/Q62R; 또는
D43N/E44N/Y51T/F56Q/Q62R.
본 발명의 방법에서 사용하기 위한 CsgG 돌연변이체는 바람직하게는 9개의 단량체를 포함하고, 단량체 중 적어도 1개가 위치 Y51, N55 및 F56 중 하나 이상에서의 돌연변이를 포함하는 서열식별번호: 390에 제시된 서열의 변이체이다. 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 위치 Y51, N55 및 F56 중 하나 이상에서의 돌연변이를 포함하는 서열식별번호: 390의 변이체를 포함한다. 바람직하게는 이러한 바람직한 포어에서 단량체들이 동일하다. 변이체는 Y51; N55; F56; Y51/N55; Y51/F56; N55/F56; 또는 Y51/N55/F56에서의 돌연변이를 포함할 수 있다. 변이체는 상기 논의된 위치 Y51, N55 및 F56 중 하나 이상에서의 특정 돌연변이 중 임의의 것을 임의의 조합으로 포함할 수 있다. 하나 이상의 Y51, N55 및 F56이 임의의 아미노산으로 치환될 수 있다. Y51이 F, M, L, I, V, A, P, G, C, Q, N, T, S, E, D, K, H 또는 R, 예컨대 A, S, T, N 또는 Q로 치환될 수 있다. N55가 F, M, L, I, V, A, P, G, C, Q, T, S, E, D, K, H 또는 R, 예컨대 A, S, T 또는 Q로 치환될 수 있다. F56이 M, L, I, V, A, P, G, C, Q, N, T, S, E, D, K, H 또는 R, 예컨대 A, S, T, N 또는 Q로 치환될 수 있다. 변이체는 하기 중 하나 이상을 추가로 포함할 수 있다: (i) 하기 위치 (i) N40, D43, E44, S54, S57, Q62, R97, E101, E124, E131, R142, T150 및 R192에서의 하나 이상의 돌연변이 (즉, 상기 위치 중 하나 이상에서의 돌연변이); (iii) Q42R 또는 Q42K; (iv) K49R; (v) N102R, N102F, N102Y 또는 N102W; (vi) D149N, D149Q 또는 D149R; (vii) E185N, E185Q 또는 E185R; (viii) D195N, D195Q 또는 D195R; (ix) E201N, E201Q 또는 E201R; (x) E203N, E203Q 또는 E203R; 및 (xi) 하기 위치 F48, K49, P50, Y51, P52, A53, S54, N55, F56 및 S57 중 하나 이상의 결실. 변이체는 상기 논의된 (i) 및 (iii) 내지 (xi)의 조합 중 임의의 것을 포함할 수 있다. 변이체는 (i) 및 (iii) 내지 (xi)에 대해 상기 논의된 실시양태 중 임의의 것을 포함할 수 있다.
(1) 본 발명의 방법에서 사용하기 위한 바람직한 변이체 또는 (2) 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 Y51R/F56Q, Y51N/F56N, Y51M/F56Q, Y51L/F56Q, Y51I/F56Q, Y51V/F56Q, Y51A/F56Q, Y51P/F56Q, Y51G/F56Q, Y51C/F56Q, Y51Q/F56Q, Y51N/F56Q, Y51S/F56Q, Y51E/F56Q, Y51D/F56Q, Y51K/F56Q 또는 Y51H/F56Q를 포함하는 서열식별번호: 390의 변이체를 포함한다.
(1) 본 발명의 방법에서 사용하기 위한 바람직한 변이체 또는 (2) 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 Y51T/F56Q, Y51Q/F56Q 또는 Y51A/F56Q를 포함하는 서열식별번호: 390의 변이체를 포함한다.
(1) 본 발명의 방법에서 사용하기 위한 바람직한 변이체 또는 (2) 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 Y51T/F56F, Y51T/F56M, Y51T/F56L, Y51T/F56I, Y51T/F56V, Y51T/F56A, Y51T/F56P, Y51T/F56G, Y51T/F56C, Y51T/F56Q, Y51T/F56N, Y51T/F56T, Y51T/F56S, Y51T/F56E, Y51T/F56D, Y51T/F56K, Y51T/F56H 또는 Y51T/F56R을 포함하는 서열식별번호: 390의 변이체를 포함한다.
(1) 본 발명의 방법에서 사용하기 위한 바람직한 변이체 또는 (2) 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 Y51T/N55Q, Y51T/N55S 또는 Y51T/N55A를 포함하는 서열식별번호: 390의 변이체를 포함한다.
(1) 본 발명의 방법에서 사용하기 위한 바람직한 변이체 또는 (2) 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 Y51A/F56F, Y51A/F56L, Y51A/F56I, Y51A/F56V, Y51A/F56A, Y51A/F56P, Y51A/F56G, Y51A/F56C, Y51A/F56Q, Y51A/F56N, Y51A/F56T, Y51A/F56S, Y51A/F56E, Y51A/F56D, Y51A/F56K, Y51A/F56H 또는 Y51A/F56R을 포함하는 서열식별번호: 390의 변이체를 포함한다.
(1) 본 발명의 방법에서 사용하기 위한 바람직한 변이체 또는 (2) 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 Y51C/F56A, Y51E/F56A, Y51D/F56A, Y51K/F56A, Y51H/F56A, Y51Q/F56A, Y51N/F56A, Y51S/F56A, Y51P/F56A 또는 Y51V/F56A를 포함하는 서열식별번호: 390의 변이체를 포함한다.
(1) 본 발명의 방법에서 사용하기 위한 바람직한 변이체 또는 (2) 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 하기를 포함하는 서열식별번호: 390의 변이체를 포함한다:
D149R/E185R/E201R/E203R 또는 Y51T/F56Q/D149R/E185R/E201R/E203R;
D149N/E185N/E201N/E203N 또는 Y51T/F56Q/D149N/E185N/E201N/E203N;
E201R/E203R 또는 Y51T/F56Q/E201R/E203R
E201N/E203R 또는 Y51T/F56Q/E201N/E203R;
E203R 또는 Y51T/F56Q/E203R;
E203N 또는 Y51T/F56Q/E203N;
E201R 또는 Y51T/F56Q/E201R;
E201N 또는 Y51T/F56Q/E201N;
E185R 또는 Y51T/F56Q/E185R;
E185N 또는 Y51T/F56Q/E185N;
D149R 또는 Y51T/F56Q/D149R;
D149N 또는 Y51T/F56Q/D149N;
R142E 또는 Y51T/F56Q/R142E;
R142N 또는 Y51T/F56Q/R142N;
R192E 또는 Y51T/F56Q/R192E; 또는
R192N 또는 Y51T/F56Q/R192N.
(1) 본 발명의 방법에서 사용하기 위한 바람직한 변이체 또는 (2) 본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 하기를 포함하는 서열식별번호: 390의 변이체를 포함한다:
Y51A/F56Q/E101N/N102R;
Y51A/F56Q/R97N/N102G;
Y51A/F56Q/R97N/N102R;
Y51A/F56Q/R97N;
Y51A/F56Q/R97G;
Y51A/F56Q/R97L;
Y51A/F56Q/N102R;
Y51A/F56Q/N102F;
Y51A/F56Q/N102G;
Y51A/F56Q/E101R;
Y51A/F56Q/E101F;
Y51A/F56Q/E101N; 또는
Y51A/F56Q/E101G.
바람직하게는 이러한 바람직한 포어에서 단량체들이 동일하다.
본 발명의 방법에서 사용하기 위한 바람직한 포어는 적어도 7개, 적어도 8개, 적어도 9개 또는 적어도 10개의 단량체, 예컨대 7, 8, 9 또는 10개의 단량체를 포함하고, 이들 각각은 F56A, F56P, F56R, F56H, F56S, F56Q, F56I, F56L, F56T 또는 F56G를 포함하는 서열식별번호: 390의 변이체를 포함한다. 바람직하게는 이러한 바람직한 포어에서 단량체들이 동일하다.
CsgG 돌연변이체는 실시예에 개시된 서열식별번호: 390의 변이체 중 임의의 것을 포함할 수 있거나 또는 실시예에 개시된 포어 중 임의의 것일 수 있다.
CsgG 돌연변이체는 가장 바람직하게는 본 발명의 포어이다.
바람직하게는, 포어를 가로질러 전위가 인가되면서 단계 (a) 및 (b)가 수행된다. 하기에서 더욱 상세하게 논의된 바와 같이, 인가된 전위는 전형적으로 포어와 폴리뉴클레오티드 결합 단백질 사이에 복합체가 형성되는 것을 초래한다. 인가된 전위는 전압 전위일 수 있다. 대안적으로, 인가된 전위는 화학 전위일 수 있다. 이의 예는 양친매성 층을 가로지르는 염 구배를 사용하는 것이다. 염 구배가 문헌 [Holden et al., J Am Chem Soc. 2007 Jul 11;129(27):8650-5]에 개시되어 있다.
이러한 방법은 표적 피분석물의 존재, 부재 또는 하나 이상의 특성을 결정하기 위한 것이다. 방법은 적어도 1개의 피분석물의 존재, 부재 또는 하나 이상의 특성을 결정하기 위한 것일 수 있다. 방법은 2개 이상의 피분석물의 존재, 부재 또는 하나 이상의 특성을 결정하는 것에 관련될 수 있다. 방법은 임의 개수의 피분석물, 예컨대 2, 5, 10, 15, 20, 30, 40, 50, 100개 또는 이를 초과하는 개수의 피분석물의 존재, 부재 또는 하나 이상의 특성을 결정하는 것을 포함할 수 있다. 하나 이상의 피분석물의 임의 개수의 특성, 예컨대 1, 2, 3, 4, 5, 10개 또는 이를 초과하는 개수의 특성이 결정될 수 있다.
표적 피분석물은 바람직하게는 금속 이온, 무기 염, 중합체, 아미노산, 펩티드, 폴리펩티드, 단백질, 뉴클레오티드, 올리고뉴클레오티드, 폴리뉴클레오티드, 염료, 표백제, 약제, 진단제, 향락성 약물, 폭발물 또는 환경 오염물이다. 방법은 동일한 유형의 2개 이상의 피분석물, 예컨대 2개 이상의 단백질, 2개 이상의 뉴클레오티드 또는 2개 이상의 제약의 존재, 부재 또는 하나 이상의 특성을 결정하는 것에 관련될 수 있다. 대안적으로, 방법은 상이한 유형의 2개 이상의 피분석물, 예컨대 1개 이상의 단백질, 1개 이상의 뉴클레오티드 및 1개 이상의 제약의 존재, 부재 또는 하나 이상의 특성을 결정하는 것에 관련될 수 있다.
표적 피분석물은 세포로부터 분비될 수 있다. 대안적으로, 표적 피분석물은 세포 내부에 존재하는 피분석물일 수 있어서, 본 발명이 수행될 수 있기 전에 피분석물이 세포로부터 추출되어야 한다.
피분석물은 바람직하게는 아미노산, 펩티드, 폴리펩티드 및/또는 단백질이다. 아미노산, 펩티드, 폴리펩티드 또는 단백질은 천연 발생 또는 비-천연발생일 수 있다. 폴리펩티드 또는 단백질은 이들 내에 합성 또는 변형 아미노산을 포함할 수 있다. 아미노산에 대한 다수의 상이한 유형의 변형이 관련 기술 분야에 공지되어 있다. 적절한 아미노산 및 이의 변형이 상기에 있다. 본 발명의 목적을 위해, 표적 피분석물이 관련 기술 분야에서 입수가능한 임의의 방법으로 변형될 수 있는 것으로 이해되어야 한다.
단백질은 효소, 항체, 호르몬, 성장 인자 또는 성장 조절 단백질, 예컨대 시토카인일 수 있다. 시토카인은 인터루킨, 바람직하게는 IFN-1, IL-1, IL-2, IL-4, IL-5, IL-6, IL-10, IL-12 및 IL-13, 인터페론, 바람직하게는 IL-γ, 및 기타 시토카인 예컨대 TNF-α로부터 선택될 수 있다. 단백질은 박테리아 단백질, 진균 단백질, 바이러스 단백질 또는 기생충-유래 단백질일 수 있다.
표적 피분석물은 바람직하게는 뉴클레오티드, 올리고뉴클레오티드 또는 폴리뉴클레오티드이다. 뉴클레오티드 및 폴리뉴클레오티드가 하기에서 논의된다. 올리고뉴클레오티드는 전형적으로 50개 이하의 뉴클레오티드, 예컨대 40개 이하, 30개 이하, 20개 이하, 10개 이하 또는 5개 이하의 뉴클레오티드를 갖는 짧은 뉴클레오티드 중합체이다. 올리고뉴클레오티드는 무염기성 및 변형 뉴클레오티드를 포함하여 하기 논의된 뉴클레오티드 중 임의의 것을 포함할 수 있다.
표적 피분석물, 예컨대 표적 폴리뉴클레오티드는 하기 논의된 적절한 샘플 중 임의의 것 내에 존재할 수 있다.
전형적으로 포어는 하기 논의된 바와 같은 막 내에 존재한다. 하기 논의된 방법을 사용하여 표적 피분석물이 막으로 커플링 또는 전달될 수 있다.
하기 논의된 측정치 중 임의의 것을 사용하여 표적 피분석물의 존재, 부재 또는 하나 이상의 특성을 결정할 수 있다. 바람직하게는 방법은 피분석물이 포어에 대해, 예컨대 포어를 통해 이동하도록 표적 피분석물을 포어와 접촉시키는 단계 및 피분석물이 포어에 대해 이동할 때 포어를 통과하는 전류를 측정하고, 이에 의해 피분석물의 존재, 부재 또는 하나 이상의 특성을 결정하는 단계를 포함한다.
피분석물에 대해 특이적인 방식으로 전류가 포어를 통해 흐르면 (즉, 피분석물과 연관된 독특한 전류가 포어를 통해 흐르는 것으로 검출되면), 표적 피분석물이 존재한다. 뉴클레오티드에 대해 특이적인 방식으로 포어를 통해 전류가 흐르지 않으면, 피분석물이 부재한다. 피분석물이 포어를 통해 흐르는 전류에 영향을 미치는 방식을 결정하기 위해 피분석물의 존재 하에 대조군 실험을 수행할 수 있다.
본 발명은 포어를 통과하는 전류에 대한 피분석물의 상이한 효과를 기초로 유사한 구조의 피분석물들을 구별하는데 사용될 수 있다. 개별적인 피분석물을 이들이 포어와 상호작용할 때의 이들의 전류 진폭으로부터 단일 분자 수준에서 확인할 수 있다. 본 발명은 특정 피분석물이 샘플 내에 존재하는지 여부를 결정하는데 또한 사용될 수 있다. 본 발명은 샘플 내의 특정 피분석물의 농도를 측정하는데 또한 사용될 수 있다. CsgG 이외의 포어를 사용하는 피분석물 특성화가 관련 기술 분야에 공지되어 있다.
폴리뉴클레오티드 특성화
본 발명은 표적 폴리뉴클레오티드를 특성화하는, 예컨대 폴리뉴클레오티드를 시퀀싱하는 방법을 제공한다. 나노포어를 사용하여 폴리뉴클레오티드를 특성화 또는 시퀀싱하는 2가지 주요 전략, 즉 가닥 특성화/시퀀싱 및 엑소뉴클레아제 특성화/시퀀싱이 있다. 본 발명은 양쪽 방법에 관련될 수 있다.
가닥 시퀀싱에서, 인가된 전위를 따라 또는 이에 대항하여 DNA가 나노포어를 통해 변위된다. 이중 가닥 DNA 상에서 점진적으로 또는 진행성으로 작용하는 엑소뉴클레아제를 인가된 전위 하에 나머지 단일 가닥을 공급하도록 포어의 시스 측에서 사용할 수 있거나 또는 역 전위 하에 트랜스 측에서 사용할 수 있다. 유사하게, 이중 가닥 DNA를 푸는 헬리카제를 유사한 방식으로 또한 사용할 수 있다. 폴리머라제를 또한 사용할 수 있다. 인가된 전위에 대항하는 가닥 변위를 요구하는 시퀀싱 용도에 대한 가능성이 또한 존재하지만, 먼저 DNA가 역 전위 또는 전위 부재 하에 효소에 의해 "포획"되어야 한다. 그 후, 결합 후에 전위가 되돌아가면, 가닥이 시스에서 트랜스로 포어를 통과할 것이고, 전류 흐름에 의해, 쭉 뻗은 형상으로 유지될 것이다. 단일 가닥 DNA 엑소뉴클레아제 또는 단일 가닥 DNA 의존적 폴리머라제가 방금 변위된 단일 가닥을 제어된 단계적 방식으로, 트랜스에서 시스로, 인가된 전위에 대항하여 포어를 통해 되돌려 당기는 분자 모터로서 작용할 수 있다.
한 실시양태에서, 표적 폴리뉴클레오티드를 특성화하는 방법은 표적 서열을 포어 및 헬리카제 효소와 접촉시키는 것을 수반한다. 임의의 헬리카제가 방법에서 사용될 수 있다. 적절한 헬리카제가 하기에서 논의된다. 헬리카제는 포어에 대해 2가지 모드로 작동할 수 있다. 첫째로, 바람직하게는, 헬리카제가 인가된 전압으로부터 초래된 장을 따라 포어를 통해 표적 서열이 이동하는 것을 제어하도록 헬리카제를 사용하여 방법이 수행된다. 이러한 모드에서는, DNA의 5' 끝부분이 먼저 포어 내에 포착되고, 최종적으로 표적 서열이 이중층의 트랜스 측으로 변위될 때까지 장을 따라 표적 서열이 포어에 통과되도록 효소가 포어 내로의 DNA 이동을 제어한다. 대안적으로, 바람직하게는, 헬리카제 효소가 인가된 전압으로부터 초래된 장에 대항하여 포어를 통해 표적 서열이 이동하는 것을 제어하도록 방법이 수행된다. 이러한 모드에서는, DNA의 3' 끝부분이 먼저 포어 내에 포착되고, 최종적으로 이중층의 시스 측으로 되돌려 배출될 때까지 인가된 장에 대항하여 표적 서열이 포어 밖으로 당겨지도록 효소가 포어를 통한 DNA의 이동을 제어한다
엑소뉴클레아제 시퀀싱에서는, 엑소뉴클레아제가 개별적인 뉴클레오티드들을 표적 폴리뉴클레오티드의 한쪽 끝부분으로부터 방출시키고, 이러한 개별적인 뉴클레오티드들이 하기 논의된 바와 같이 확인된다. 또 다른 실시양태에서, 표적 폴리뉴클레오티드를 특성화하는 방법은 표적 서열을 포어 및 엑소뉴클레아제 효소와 접촉시키는 것을 수반한다. 하기 논의된 엑소뉴클레아제 효소 중 임의의 것이 이러한 방법에서 사용될 수 있다. 하기 논의된 바와 같이 효소가 포어에 공유결합으로 부착될 수 있다.
엑소뉴클레아제는 전형적으로 폴리뉴클레오티드의 한쪽 끝부분 상에 걸려서 서열을 이러한 끝부분으로부터 한번에 하나의 뉴클레오티드를 소화시키는 효소이다. 엑소뉴클레아제는 폴리뉴클레오티드를 5' → 3' 방향 또는 3' → 5' 방향으로 소화시킬 수 있다. 전형적으로, 엑소뉴클레아제가 결합하는 폴리뉴클레오티드의 끝부분은 사용된 효소의 선택을 통해 및/또는 관련 기술 분야에 공지된 방법을 사용하여 결정된다. 전형적으로, 폴리뉴클레오티드의 한쪽 끝부분의 히드록실 기 또는 캡 구조가 엑소뉴클레아제가 폴리뉴클레오티드의 특정 끝부분에 결합하는 것을 방지하거나 용이하게 하는데 사용될 수 있다.
방법은 상기 논의된 바와 같은 뉴클레오티드의 일부분의 특성화 또는 확인을 허용하는 속도로 폴리뉴클레오티드의 끝부분으로부터 뉴클레오티드가 소화되도록 폴리뉴클레오티드를 엑소뉴클레아제와 접촉시키는 것을 수반한다. 이를 행하기 위한 방법은 관련 기술 분야에 널리 공지되어 있다. 예를 들어, 에드만(Edman) 분해를 사용하여 폴리펩티드의 끝부분으로부터 단일 아미노산을 연속적으로 소화시켜, 이를 고성능 액체 크로마토그래피 (HPLC)를 사용하여 확인할 수 있다. 상응하는 방법이 본 발명에서 사용될 수 있다.
엑소뉴클레아제가 기능하는 속도는 전형적으로 야생형 엑소뉴클레아제에 대한 최적 속도보다 더 느리다. 본 발명의 방법에서의 엑소뉴클레아제의 활성의 적절한 속도는 초 당 0.5 내지 1000개의 뉴클레오티드, 초 당 0.6 내지 500개의 뉴클레오티드, 초 당 0.7 내지 200개의 뉴클레오티드, 초 당 0.8 내지 100개의 뉴클레오티드, 초 당 0.9 내지 50개의 뉴클레오티드 또는 초 당 1 내지 20개 또는 10개의 뉴클레오티드의 소화를 수반한다. 바람직하게는 속도는 초 당 1, 10, 100, 500 또는 1000개의 뉴클레오티드이다. 다양한 방식으로 엑소뉴클레아제 활성의 적절한 속도가 달성될 수 있다. 예를 들어, 감소된 최적 활성 속도의 변이체 엑소뉴클레아제가 본 발명에 따라 사용될 수 있다.
가닥 특성화 실시양태에서, 방법은 폴리뉴클레오티드가 포어에 대해, 예컨대 포어를 통해 이동하도록 폴리뉴클레오티드를 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉시키는 단계 및 폴리뉴클레오티드가 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 여기서 측정은 폴리뉴클레오티드의 하나 이상의 특성을 지시하며, 이에 의해 표적 폴리뉴클레오티드를 특성화하는 단계를 포함한다.
엑소뉴클레오티드 특성화 실시양태에서, 방법은 엑소뉴클레아제가 개별적인 뉴클레오티드를 표적 폴리뉴클레오티드의 한쪽 끝부분으로부터 소화하고 개별적인 뉴클레오티드가 포어에 대해, 예컨대 포어를 통해 이동하도록, 폴리뉴클레오티드를 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어, 및 엑소뉴클레아제와 접촉시키는 단계, 및 개별적인 뉴클레오티드가 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 여기서 측정은 개별적인 뉴클레오티드의 하나 이상의 특성을 지시하며, 이에 의해 표적 폴리뉴클레오티드를 특성화하는 단계를 포함한다.
개별적인 뉴클레오티드는 단일 뉴클레오티드이다. 개별적인 뉴클레오티드는 뉴클레오티드 결합에 의해 또 다른 뉴클레오티드 또는 폴리뉴클레오티드에 결합되지 않은 것이다. 뉴클레오티드 결합은 뉴클레오티드의 포스페이트 기 중 하나가 또 다른 뉴클레오티드의 당 기에 결합되는 것을 수반한다. 전형적으로, 개별적인 뉴클레오티드는 적어도 5개, 적어도 10개, 적어도 20개, 적어도 50개, 적어도 100개, 적어도 200개, 적어도 500개, 적어도 1000개 또는 적어도 5000개의 뉴클레오티드의 또 다른 폴리뉴클레오티드에 뉴클레오티드 결합에 의해 결합되지 않은 것이다. 예를 들어, 표적 폴리뉴클레오티드 서열, 예컨대 DNA 또는 RNA 가닥으로부터 개별적인 뉴클레오티드가 소화되었다. 뉴클레오티드는 하기 기술된 것들 중 임의의 것일 수 있다.
개별적인 뉴클레오티드는 임의의 방식으로 임의의 부위에서 포어와 상호작용할 수 있다. 바람직하게는 뉴클레오티드는 상기 논의된 바와 같은 어댑터를 통해 또는 이와 함께 가역적으로 포어에 결합한다. 가장 바람직하게는 뉴클레오티드는 이들이 막을 가로질러 포어를 통과할 때 어댑터를 통해 또는 이와 함께 가역적으로 포어에 결합한다. 또한 뉴클레오티드는 이들이 막을 가로질러 포어를 통과할 때 어댑터를 통해 또는 이와 함께 가역적으로 포어의 배럴 또는 채널에 결합할 수 있다.
개별적인 뉴클레오티드와 포어의 상호작용 동안, 전형적으로 뉴클레오티드는 포어를 통해 흐르는 전류에 이러한 뉴클레오티드에 대해 특이적인 방식으로 영향을 미친다. 예를 들어, 특정 뉴클레오티드는 포어를 통해 흐르는 전류를 특정한 평균 시간 동안 특정한 정도로 감소시킬 것이다. 달리 말하면, 포어를 통해 흐르는 전류가 특정 뉴클레오티드에 대해 독특하다. 포어를 통해 흐르는 전류에 대한 특정 뉴클레오티드의 효과를 결정하기 위해 대조군 실험을 수행할 수 있다. 그 후, 샘플 내의 특정 뉴클레오티드를 확인하거나 또는 특정 뉴클레오티드가 샘플 내에 존재하는지 여부를 결정하기 위해, 테스트 샘플 상에서 본 발명의 방법을 수행하는 것으로부터의 결과를 이같은 대조군 실험으로부터 유래된 것과 비교할 수 있다. 포어를 통해 흐르는 전류가 특정 뉴클레오티드를 지시하는 방식으로 영향을 받는 빈도를 사용하여, 샘플 내의 이러한 뉴클레오티드의 농도를 결정할 수 있다. 샘플 내의 상이한 뉴클레오티드들의 비를 또한 계산할 수 있다. 예를 들어, dCMP 대 메틸-dCMP의 비를 계산할 수 있다.
방법은 표적 폴리뉴클레오티드의 하나 이상의 특성을 측정하는 것을 수반한다. 표적 폴리뉴클레오티드는 주형 폴리뉴클레오티드 또는 관심 폴리뉴클레오티드로 또한 칭해질 수 있다.
이러한 실시양태 또한 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어를 사용한다. 표적 피분석물과 관련하여 상기 논의된 포어 및 실시양태 중 임의의 것이 사용될 수 있다.
폴리뉴클레오티드
폴리뉴클레오티드, 예컨대 핵산은 2개 이상의 뉴클레오티드를 포함하는 거대분자이다. 폴리뉴클레오티드 또는 핵산은 임의의 뉴클레오티드의 임의의 조합을 포함할 수 있다. 뉴클레오티드는 천연 발생 또는 인공일 수 있다. 폴리뉴클레오티드 내의 하나 이상의 뉴클레오티드가 산화 또는 메틸화될 수 있다. 폴리뉴클레오티드 내의 하나 이상의 뉴클레오티드가 손상될 수 있다. 예를 들어, 폴리뉴클레오티드가 피리미딘 이량체를 포함할 수 있다. 이같은 이량체는 전형적으로 자외선에 의한 손상과 연관되고, 피부 흑색종의 주요 원인이다. 폴리뉴클레오티드 내의 하나 이상의 뉴클레오티드가, 예를 들어 표지 또는 태그로, 변형될 수 있다. 적절한 표지가 하기에서 기술된다. 폴리뉴클레오티드는 하나 이상의 스페이서를 포함할 수 있다.
뉴클레오티드는 전형적으로 핵염기, 당 및 적어도 1개의 포스페이트 기를 함유한다. 핵염기 및 당은 뉴클레오시드를 형성한다.
핵염기는 전형적으로 헤테로고리형이다. 핵염기는 퓨린 및 피리미딘, 더욱 구체적으로는 아데닌 (A), 구아닌 (G), 티민 (T), 우라실 (U) 및 시토신 (C)을 포함하지만, 이에 제한되지 않는다.
당은 전형적으로 펜토스 당이다. 뉴클레오티드 당은 리보스 및 데옥시리보스를 포함하지만, 이에 제한되지 않는다. 바람직하게는 당은 데옥시리보스이다.
폴리뉴클레오티드는 바람직하게는 하기의 뉴클레오시드를 포함한다: 데옥시아데노신 (dA), 데옥시우리딘 (dU) 및/또는 티미딘 (dT), 데옥시구아노신 (dG) 및 데옥시시티딘 (dC).
뉴클레오티드는 전형적으로 리보뉴클레오티드 또는 데옥시리보뉴클레오티드이다. 뉴클레오티드는 전형적으로 모노포스페이트, 디포스페이트 또는 트리포스페이트를 함유한다. 뉴클레오티드는 3개를 초과하는 포스페이트, 예컨대 4 또는 5개의 포스페이트를 포함할 수 있다. 포스페이트는 뉴클레오티드의 5' 또는 3' 측면 상에 부착될 수 있다. 뉴클레오티드는 아데노신 모노포스페이트 (AMP), 구아노신 모노포스페이트 (GMP), 티미딘 모노포스페이트 (TMP), 우리딘 모노포스페이트 (UMP), 5-메틸시티딘 모노포스페이트, 5-히드록시메틸시티딘 모노포스페이트, 시티딘 모노포스페이트 (CMP), 고리형 아데노신 모노포스페이트 (cAMP), 고리형 구아노신 모노포스페이트 (cGMP), 데옥시아데노신 모노포스페이트 (dAMP), 데옥시구아노신 모노포스페이트 (dGMP), 데옥시티미딘 모노포스페이트 (dTMP), 데옥시우리딘 모노포스페이트 (dUMP), 데옥시시티딘 모노포스페이트 (dCMP) 및 데옥시메틸시티딘 모노포스페이트를 포함하지만, 이에 제한되지 않는다. 뉴클레오티드는 바람직하게는 AMP, TMP, GMP, CMP, UMP, dAMP, dTMP, dGMP, dCMP 및 dUMP로부터 선택된다.
뉴클레오티드는 무염기성일 수 있다 (즉, 핵염기가 결여될 수 있다). 또한 뉴클레오티드는 핵염기 및 당이 결여될 수 있다 (즉, C3 스페이서이다).
폴리뉴클레오티드 내의 뉴클레오티드들은 임의의 방식으로 서로 부착될 수 있다. 전형적으로 뉴클레오티드들은 핵산에서와 같이 이들의 당 및 포스페이트 기에 의해 부착된다. 피리디딘 이량체에서와 같이 뉴클레오티드들이 이들의 핵염기를 통해 연결될 수 있다.
폴리뉴클레오티드는 단일 가닥 또는 이중 가닥일 수 있다. 바람직하게는 폴리뉴클레오티드의 적어도 일부분이 이중 가닥이다.
폴리뉴클레오티드는 핵산, 예컨대 데옥시리보핵산 (DNA) 또는 리보핵산 (RNA)일 수 있다. 폴리뉴클레오티드는 1개의 DNA 가닥에 혼성화된 1개의 RNA 가닥을 포함할 수 있다. 폴리뉴클레오티드는 관련 기술 분야에 공지된 임의의 합성 핵산, 예컨대 펩티드 핵산 (PNA), 글리세롤 핵산 (GNA), 트레오스 핵산 (TNA), 잠금 핵산 (LNA) 또는 뉴클레오티드 측쇄가 있는 기타 합성 중합체일 수 있다. PNA 백본(backbone)은 펩티드 결합으로 연결된 반복되는 N-(2-아미노에틸)-글리신 단위로 구성된다. GNA 백본은 포스포디에스테르 결합으로 연결된 반복되는 글리콜 단위로 구성된다. TNA 백본은 포스포디에테스테르 결합으로 함께 연결된 반복되는 트레오스 당으로 구성된다. LNA는 리보스 모이어티 내의 2' 산소와 4' 탄소를 연결하는 여분의 가교가 있는 상기 논의된 바와 같은 리보뉴클레오티드로부터 형성된다. 가교(bridged) 핵산 (BNA)은 변형된 RNA 뉴클레오티드이다. 이는 구속(constrained) 또는 접근불능성 RNA로 또한 칭해질 수 있다. BNA 단량체는 "고정된" C3'-엔도(endo) 당 퍼커링(puckering)이 있는 5원, 6원 또는 심지어 7원의 가교 구조를 함유할 수 있다. 이러한 가교가 리보스의 2',4'-위치에서 합성으로 혼입되어 2',4 -BNA 단량체가 생산된다.
폴리뉴클레오티드는 가장 바람직하게는 리보핵산 (RNA) 또는 데옥시리보핵산 (DNA)이다.
폴리뉴클레오티드는 임의의 길이일 수 있다. 예를 들어, 폴리뉴클레오티드는 적어도 10개, 적어도 50개, 적어도 100개, 적어도 150개, 적어도 200개, 적어도 250개, 적어도 300개, 적어도 400개 또는 적어도 500개의 뉴클레오티드 또는 뉴클레오티드 쌍의 길이일 수 있다. 폴리뉴클레오티드는 1000개 이상의 뉴클레오티드 또는 뉴클레오티드 쌍, 5000개 이상의 뉴클레오티드 또는 뉴클레오티드 쌍의 길이, 또는 100000개 이상의 뉴클레오티드 또는 뉴클레오티드 쌍의 길이일 수 있다.
임의 개수의 폴리뉴클레오티드를 연구할 수 있다. 예를 들어, 본 발명의 방법은 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 50, 100개 또는 이를 초과하는 개수의 폴리뉴클레오티드를 특성화하는 것에 관련될 수 있다. 2개 이상의 폴리뉴클레오티드가 특성화되는 경우, 이들은 상이한 폴리뉴클레오티드, 또는 동일한 폴리뉴클레오티드의 2개의 예일 수 있다.
폴리뉴클레오티드는 천연 발생 또는 인공일 수 있다. 예를 들어, 제작된 올리고뉴클레오티드의 서열을 입증하는데 방법이 사용될 수 있다. 전형적으로는 시험관 내에서 방법이 수행된다.
샘플
전형적으로 폴리뉴클레오티드는 임의의 적절한 샘플 내에 존재한다. 전형적으로 본 발명은 폴리뉴클레오티드를 함유하는 것으로 알려진 또는 이를 함유하는 것으로 추정되는 샘플 상에서 수행된다. 대안적으로, 샘플 내의 폴리뉴클레오티드의 존재가 알려진 또는 예상되는 폴리뉴클레오티드의 신원을 확증하기 위해 샘플 상에서 본 발명이 수행될 수 있다.
샘플은 생물학적 샘플일 수 있다. 임의의 생물 또는 미생물로부터 수득 또는 추출된 샘플을 사용하여 시험관 내에서 본 발명이 수행될 수 있다. 생물 또는 미생물은 전형적으로 고세균, 원핵생물 또는 진핵생물이고, 전형적으로 5가지 계 중 하나에 속한다: 식물계, 동물계, 진균계, 모네라계 및 원생생물계. 임의의 바이러스로부터 수득 또는 추출된 샘플 상에서 시험관 내에서 본 발명이 수행될 수 있다. 샘플은 바람직하게는 유체 샘플이다. 샘플은 전형적으로 환자의 체액을 포함한다. 샘플은 소변, 림프, 타액, 점액 또는 양수일 수 있지만, 바람직하게는 혈액, 혈장 또는 혈청이다.
전형적으로, 샘플은 인간 기원의 것이지만, 대안적으로, 또 다른 포유동물 예컨대 상업적으로 사육되는 동물 예컨대 말, 소, 양, 어류, 닭 또는 돼지로부터의 것일 수 있거나 또는 대안적으로 애완동물 예컨대 고양이 또는 개의 것일 수 있다. 대안적으로, 샘플은 식물 기원의 것, 예컨대 상업 작물, 예컨대 곡류, 콩과식물, 과일 또는 채소, 예를 들어, 밀, 보리, 귀리, 카놀라, 옥수수, 대두, 벼, 대황, 바나나, 사과, 토마토, 감자, 포도, 담배, 콩, 렌틸, 사탕수수, 코코아, 목화로부터 수득된 샘플일 수 있다.
샘플은 비-생물학적 샘플일 수 있다. 비-생물학적 샘플은 바람직하게는 유체 샘플이다. 비-생물학적 샘플의 예는 수술 유체, 물 예컨대 음용수, 해수 또는 강물, 및 실험실 테스트용 시약을 포함한다.
전형적으로 샘플은, 예를 들어 원심분리에 의해 또는 원치 않는 분자 또는 세포, 예컨대 적혈구를 여과하는 막을 통과하는 것에 의해, 본 발명에서 사용되기 전에 프로세싱된다. 이는 채취 직후에 측정될 수 있다. 또한 전형적으로 샘플이 검정법 전에, 바람직하게는 -70℃ 미만에서, 보관될 수 있다.
특성화
폴리뉴클레오티드의 2, 3, 4 또는 5개 또는 이를 초과하는 특성을 측정하는 것이 방법에서 수반될 수 있다. 바람직하게는 하나 이상의 특성은 (i) 폴리뉴클레오티드의 길이, (ii) 폴리뉴클레오티드의 신원, (iii) 폴리뉴클레오티드의 서열, (iv) 폴리뉴클레오티드의 2차 구조 및 (v) 폴리뉴클레오티드가 변형되었는지 여부로부터 선택된다. (i) 내지 (v)의 임의의 조합, 예컨대 {i}, {ii}, {iii}, {iv}, {v}, {i,ii}, {i,iii}, {i,iv}, {i,v}, {ii,iii}, {ii,iv}, {ii,v}, {iii,iv}, {iii,v}, {iv,v}, {i,ii,iii}, {i,ii,iv}, {i,ii,v}, {i,iii,iv}, {i,iii,v}, {i,iv,v}, {ii,iii,iv}, {ii,iii,v}, {ii,iv,v}, {iii,iv,v}, {i,ii,iii,iv}, {i,ii,iii,v}, {i,ii,iv,v}, {i,iii,iv,v}, {ii,iii,iv,v} 또는 {i,ii,iii,iv,v}를 본 발명에 따라 측정할 수 있다. 상기 열거된 조합들 중 임의의 것을 포함하여, (i) 내지 (v)의 상이한 조합들을 제1 폴리뉴클레오티드에 대해 제2 폴리뉴클레오티드와 비교하여 측정할 수 있다.
(i)의 경우, 예를 들어 폴리뉴클레오티드와 포어 사이의 상호작용의 횟수 또는 폴리뉴클레오티드와 포어 사이의 상호작용의 지속기간을 결정함으로써 폴리뉴클레오티드의 길이를 측정할 수 있다.
(ii)의 경우, 다수의 방식으로 폴리뉴클레오티드의 신원을 측정할 수 있다. 폴리뉴클레오티드 서열의 측정과 함께 또는 폴리뉴클레오티드 서열의 측정 없이 폴리뉴클레오티드의 신원을 측정할 수 있다. 전자는 간단하다; 폴리뉴클레오티드를 시퀀싱하고 이에 의해 확인한다. 후자는 여러 방식으로 행해질 수 있다. 예를 들어, 폴리뉴클레오티드 내의 특정 모티프의 존재를 (폴리뉴클레오티드의 나머지 서열을 측정하지 않으면서) 측정할 수 있다. 대안적으로, 특정한 전기 및/또는 광학 신호를 방법에서 측정하는 것으로 폴리뉴클레오티드를 특정 공급원으로부터 유래되는 것으로서 확인할 수 있다.
(iii)의 경우, 이전에 기술된 바와 같이 폴리뉴클레오티드의 서열을 결정할 수 있다. 적절한 시퀀싱 방법 방법, 특히 전기적 측정을 사용하는 방법이 문헌 [Stoddart D et al., Proc Natl Acad Sci, 12; 106(19):7702-7], [Lieberman KR et al., J Am Chem Soc. 2010; 132(50):17961-72], 및 국제 출원 WO 2000/28312에 기술되어 있다.
(iv)의 경우, 다양한 방식으로 2차 구조를 측정할 수 있다. 예를 들어, 방법이 전기적 측정을 수반하는 경우, 체류 시간의 변화 또는 포어를 통해 흐르는 전류의 변화를 사용하여 2차 구조를 측정할 수 있다. 이는 단일-가닥 및 이중-가닥 폴리뉴클레오티드의 영역이 구별되게 한다.
(v)의 경우, 임의의 변형의 존재 또는 부재를 측정할 수 있다. 바람직하게는 방법은 폴리뉴클레오티드가 메틸화에 의해, 산화에 의해, 손상에 의해, 하나 이상의 단백질로, 또는 하나 이상의 표지, 태그 또는 스페이서로 변형되었는지 여부를 결정하는 것을 포함한다. 특정 변형은 포어와의 특정 상호작용을 초래할 것이고, 이를 하기 기술된 방법을 사용하여 측정할 수 있다. 예를 들어, 포어와 각각의 뉴클레오티드의 상호작용 동안 포어를 통해 흐르는 전류를 기초로 메틸시토신을 시토신과 구별할 수 있다.
표적 폴리뉴클레오티드가 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉된다. 포어는 전형적으로 막 내에 존재한다. 적절한 막이 하기에서 논의된다. 포어가 막 내에 존재하는 막/포어 시스템을 연구하는데 적절한 임의의 기구를 사용하여 방법을 수행할 수 있다. 막횡단 포어 센싱에 적절한 임의의 기구를 사용하여 방법을 수행할 수 있다. 예를 들어, 기구는 수용액을 포함하는 챔버 및 챔버를 2개의 섹션으로 분리하는 장벽을 포함한다. 전형적으로 장벽은 포어를 함유하는 막이 형성된 개구부가 있다. 대안적으로, 장벽은 포어가 존재하는 막을 형성한다.
국제 출원 번호 PCT/GB08/000562 (WO 2008/102120)에 기술된 기구를 사용하여 방법을 수행할 수 있다.
다양한 유형의 측정이 이루어질 수 있다. 이는 비제한적으로 전기적 측정 및 광학적 측정을 포함한다. 가능한 전기적 측정은 전류 측정, 임피던스 측정, 터널링(tunnelling) 측정 (Ivanov AP et al., Nano Lett. 2011 Jan 12;11(1):279-85), 및 FET 측정 (국제 출원 WO 2005/124888)을 포함한다. 광학적 측정이 전기적 측정과 조합될 수 있다 (Soni GV et al., Rev Sci Instrum. 2010 Jan;81(1):014301). 측정은 막횡단 전류 측정 예컨대 포어를 통해 흐르는 이온 전류의 측정일 수 있다.
문헌 [Stoddart D et al., Proc Natl Acad Sci, 12;106(19):7702-7], [Lieberman KR et al., J Am Chem Soc. 2010;132(50):17961-72], 및 국제 출원 WO 2000/28312에 기술된 바와 같이 표준 단일 채널 기록 기기를 사용하여 전기적 측정이 이루어질 수 있다. 대안적으로, 예를 들어 국제 출원 WO 2009/077734 및 국제 출원 WO 2011/067559에 기술된 바와 같이, 다채널 시스템을 사용하여 전기적 측정이 이루어질 수 있다.
바람직하게는, 막을 가로질러 전위가 인가되면서 방법이 수행된다. 인가된 전위는 전압 전위일 수 있다. 대안적으로, 인가된 전위는 화학 전위일 수 있다. 이의 예는 막, 예컨대 양친매성 층을 가로지르는 염 구배를 사용하는 것이다. 염 구배가 문헌 [Holden et al., J Am Chem Soc. 2007 Jul 11; 129(27):8650-5]에 개시되어 있다. 일부 경우에, 폴리뉴클레오티드가 포어에 대해 이동할 때 포어를 통과하는 전류가 폴리뉴클레오티드의 서열을 추정 또는 결정하는데 사용된다. 이는 가닥 시퀀싱이다.
방법은 폴리뉴클레오티드가 포어에 대해 이동할 때 포어를 통과하는 전류를 측정하는 것을 수반할 수 있다. 따라서, 방법에서 사용된 기구는 전위를 인가할 수 있고 막 및 포어를 가로지르는 전기적 신호를 측정할 수 있는 전기 회로를 또한 포함할 수 있다. 패치(patch) 클램프 또는 전압 클램프를 사용하여 방법을 수행할 수 있다. 바람직하게는 방법은 전압 클램프의 사용을 수반한다.
본 발명의 방법은 폴리뉴클레오티드가 포어에 관해 이동할 때 포어를 통과하는 전류를 측정하는 것을 포함할 수 있다. 막횡단 단백질 포어를 통한 이온 전류를 측정하기 위한 적절한 조건이 관련 기술 분야에 공지되어 있고, 실시예에 개시되어 있다. 전형적으로는, 막 및 포어를 가로질러 전압이 인가되면서 방법이 수행된다. 사용되는 전압은 전형적으로 +5 V 내지 -5 V, 예컨대 +4 V 내지 -4 V, +3 V 내지 -3 V 또는 +2 V 내지 -2 V이다. 사용되는 전압은 전형적으로 -600 mV 내지 +600 mV 또는 -400 mV 내지 +400 mV이다. 바람직하게는 사용되는 전압은 하한은 -400 mV, -300 mV, -200 mV, -150 mV, -100 mV, -50 mV, -20 mV 및 0 mV으로부터 선택되고 상한은 독립적으로 +10 mV, + 20 mV, +50 mV, +100 mV, +150 mV, +200 mV, +300 mV 및 +400 mV로부터 선택되는 범위 내일 수 있다. 더욱 바람직하게는 사용되는 전압은 100 mV 내지 240 mV의 범위 내이고, 가장 바람직하게는 120 mV 내지 220 mV의 범위 내이다. 증가된 인가 전위를 사용함으로써 포어에 의한 상이한 뉴클레오티드들 사이의 판별을 증가시키는 것이 가능하다.
전형적으로는, 임의의 전하 캐리어, 예컨대 금속 염, 예를 들어 알칼리금속 염, 할라이드 염, 예를 들어, 클로라이드 염, 예컨대 알칼리금속 클로라이드 염의 존재 하에 방법이 수행된다. 전하 캐리어는 이온성 액체 또는 유기 염, 예를 들어 테트라메틸 암모늄 클로라이드, 트리메틸페닐 암모늄 클로라이드, 페닐트리메틸 암모늄 클로라이드, 또는 1-에틸-3-메틸 이미다졸륨 클로라이드를 포함할 수 있다. 상기 논의된 예시적인 기구에서, 염은 챔버 내의 수용액 내에 존재한다. 염화칼륨 (KCl), 염화나트륨 (NaCl), 염화세슘 (CsCl), 또는 페로시안화칼륨 및 페리시안화칼륨의 혼합물이 전형적으로 사용된다. KCl, NaCl, 및 페로시안화칼륨 및 페리시안화칼륨의 혼합물이 바람직하다. 전하 캐리어는 막에 걸쳐 비대칭성일 수 있다. 예를 들어, 전하 캐리어의 유형 및/또는 농도가 막의 각각의 측면 상에서 상이할 수 있다.
염 농도는 포화일 수 있다. 염 농도는 3 M 이하일 수 있고, 전형적으로는 0.1 내지 2.5 M, 0.3 내지 1.9 M, 0.5 내지 1.8 M, 0.7 내지 1.7 M, 0.9 내지 1.6 M 또는 1 M 내지 1.4 M이다. 바람직하게는 염 농도는 150 mM 내지 1 M이다. 바람직하게는, 적어도 0.3 M, 예컨대 적어도 0.4 M, 적어도 0.5 M, 적어도 0.6 M, 적어도 0.8 M, 적어도 1.0 M, 적어도 1.5 M, 적어도 2.0 M, 적어도 2.5 M 또는 적어도 3.0 M의 염 농도를 사용하여 방법이 수행된다. 높은 염 농도는 높은 신호 대 노이즈 비를 제공하고, 정상 전류 변동의 배경에 대해 뉴클레오티드의 존재를 지시하는 전류가 확인되게 한다.
전형적으로는 완충제의 존재 하에 방법이 수행된다. 상기 논의된 예시적인 기구에서, 완충제는 챔버 내의 수용액 내에 존재한다. 임의의 완충제가 본 발명의 방법에서 사용될 수 있다. 전형적으로, 완충제는 포스페이트 완충제이다. 기타 적절한 완충제는 HEPES 및 트리스-HCl 완충제이다. 전형적으로는, 4.0 내지 12.0, 4.5 내지 10.0, 5.0 내지 9.0, 5.5 내지 8.8, 6.0 내지 8.7 또는 7.0 내지 8.8 또는 7.5 내지 8.5의 pH에서 방법이 수행된다. 사용되는 pH는 바람직하게는 약 7.5이다.
0℃ 내지 100℃, 15℃ 내지 95℃, 16℃ 내지 90℃, 17℃ 내지 85℃, 18℃ 내지 80℃, 19℃ 내지 70℃, 또는 20℃ 내지 60℃에서 방법이 수행될 수 있다. 전형적으로는 실온에서 방법이 수행된다. 임의적으로는, 효소 기능을 지지하는 온도, 예컨대 약 37℃에서 방법이 수행된다.
폴리뉴클레오티드 결합 단백질
바람직하게는 가닥 특성화 방법은 폴리뉴클레오티드 결합 단백질이 포어에 대한, 예컨대 포어를 통한 폴리뉴클레오티드의 이동을 제어하도록 폴리뉴클레오티드를 폴리뉴클레오티드 결합 단백질과 접촉시키는 것을 포함한다.
더욱 바람직하게는, 방법은 (a) 폴리뉴클레오티드 결합 단백질이 포어에 대한, 예컨대 포어를 통한 폴리뉴클레오티드의 이동을 제어하도록 폴리뉴클레오티드를 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어, 및 폴리뉴클레오티드 결합 단백질과 접촉시키는 단계, 및 (b) 폴리뉴클레오티드가 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 여기서 측정은 폴리뉴클레오티드의 하나 이상의 특성을 지시하며, 이에 의해 폴리뉴클레오티드를 특성화하는 단계를 포함한다.
더욱 바람직하게는, 방법은 (a) 폴리뉴클레오티드 결합 단백질이 포어에 대한, 예컨대 포어를 통한 폴리뉴클레오티드의 이동을 제어하도록 폴리뉴클레오티드를 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어, 및 폴리뉴클레오티드 결합 단백질과 접촉시키는 단계, 및 (b) 폴리뉴클레오티드가 포어에 대해 이동할 때 포어를 통한 전류를 측정하고, 여기서 전류는 폴리뉴클레오티드의 하나 이상의 특성을 지시하며, 이에 의해 폴리뉴클레오티드를 특성화하는 단계를 포함한다.
폴리뉴클레오티드 결합 단백질은 폴리뉴클레오티드에 결합할 수 있고 포어를 통한 이의 이동을 제어할 수 있는 임의의 단백질일 수 있다. 단백질이 폴리뉴클레오티드에 결합하는지 여부를 결정하는 것은 관련 기술 분야에서 간단하다. 전형적으로 이러한 단백질은 폴리뉴클레오티드와 상호작용하고, 폴리뉴클레오티드의 적어도 하나의 성질을 변형시킨다. 이러한 단백질은 폴리뉴클레오티드를 절단하여 개별적인 뉴클레오티드 또는 더 짧은 뉴클레오티드 쇄, 예컨대 디- 또는 트리뉴클레오티드를 형성시킴으로써 폴리뉴클레오티드를 변형시킬 수 있다. 이러한 단백질은 특정한 위치로 폴리뉴클레오티드를 배향시키거나 또는 이를 이동시킴으로써, 즉 이의 이동을 제어함으로써 폴리뉴클레오티드를 변형시킬 수 있다.
폴리뉴클레오티드 결합 단백질은 바람직하게는 폴리뉴클레오티드 취급 효소로부터 유래된다. 폴리뉴클레오티드 취급 효소는 폴리뉴클레오티드와 상호작용할 수 있고 폴리뉴클레오티드의 적어도 하나의 성질을 변형시킬 수 있는 폴리펩티드이다. 이러한 효소는 폴리뉴클레오티드를 절단하여 개별적인 뉴클레오티드 또는 더 짧은 뉴클레오티드 쇄, 예컨대 디- 또는 트리뉴클레오티드를 형성시킴으로써 폴리뉴클레오티드를 변형시킬 수 있다. 이러한 효소는 특정한 위치로 폴리뉴클레오티드를 배향시키거나 또는 이를 이동시킴으로써 폴리뉴클레오티드를 변형시킬 수 있다. 폴리뉴클레오티드 취급 효소는 폴리뉴클레오티드에 결합할 수 있고 포어를 통한 이의 이동을 제어할 수 있는 한 효소 활성을 나타낼 필요가 없다. 예를 들어, 효소는 이의 효소 활성을 제거하도록 변형될 수 있거나, 또는 효소가 효소로서 작용하는 것을 방지하는 조건 하에 사용될 수 있다. 이같은 조건들이 하기에서 더욱 상세하게 논의된다.
폴리뉴클레오티드 취급 효소는 바람직하게는 핵산분해 효소로부터 유래된다. 효소의 구축물에서 사용되는 폴리뉴클레오티드 취급 효소는 더욱 바람직하게는 효소 분류(Enzyme Classification) (EC) 군 3.1.11, 3.1.13, 3.1.14, 3.1.15, 3.1.16, 3.1.21, 3.1.22, 3.1.25, 3.1.26, 3.1.27, 3.1.30 및 3.1.31 중 임의의 것의 구성원으로부터 유래된다. 효소는 국제 출원 번호 PCT/GB10/000133 (WO 2010/086603으로 공개됨)에 개시된 것 중 임의의 것일 수 있다.
바람직한 효소는 폴리머라제, 엑소뉴클레아제, 헬리카제 및 토포이소머라제, 예컨대 자이라제이다. 적절한 효소는 이. 콜라이로부터의 엑소뉴클레아제 I (서열식별번호: 399), 이. 콜라이로부터의 엑소뉴클레아제 III 효소 (서열식별번호: 401), 티. 써모필루스로부터의 RecJ (서열식별번호: 403) 및 박테리오파지 람다 엑소뉴클레아제 (서열식별번호: 405), TatD 엑소뉴클레아제 및 이의 변이체를 포함하지만, 이에 제한되지 않는다. 서열식별번호: 403에 제시된 서열 또는 이의 변이체를 포함하는 3개의 서브유닛이 상호작용하여 3량체 엑소뉴클레아제를 형성한다. 이러한 엑소뉴클레아제들이 본 발명의 엑소뉴클레아제 방법에서 또한 사용될 수 있다. 폴리머라제는 파이로파지(PyroPhage)® 3173 DNA 폴리머라제 (루시젠® 코포레이션(Lucigen® Corporation)에서 시판됨), SD 폴리머라제 (바이오론(Bioron)®에서 시판됨) 또는 이의 변이체일 수 있다. 효소는 바람직하게는 Phi29 DNA 폴리머라제 (서열식별번호: 397) 또는 이의 변이체이다. 토포이소머라제는 바람직하게는 모이어티 분류 (EC) 군 5.99.1.2 및 5.99.1.3 중 임의의 것의 구성원이다.
가장 바람직하게는, 헬리카제, 예컨대 Hel308 Mbu (서열식별번호: 406), Hel308 Csy (서열식별번호: 407), Hel308 Tga (서열식별번호: 408), Hel308 Mhu (서열식별번호: 409), Tral Eco (서열식별번호: 410), XPD Mbu (서열식별번호: 411) 또는 이의 변이체로부터 효소가 유래된다. 임의의 헬리카제가 본 발명에서 사용될 수 있다. 헬리카제는 Hel308 헬리카제, RecD 헬리카제, 예컨대 Tral 헬리카제 또는 TrwC 헬리카제, XPD 헬리카제 또는 Dda 헬리카제일 수 있거나 또는 이로부터 유래될 수 있다. 헬리카제는 국제 출원 번호 PCT/GB2012/052579 (WO 2013/057495로 공개됨); PCT/GB2012/053274 (WO 2013/098562로 공개됨); PCT/GB2012/053273 (WO2013098561로 공개됨); PCT/GB2013/051925 (WO 2014/013260으로 공개됨); PCT/GB2013/051924 (WO 2014/013259로 공개됨); PCT/GB2013/051928 (WO 2014/013262로 공개됨) 및 PCT/GB2014/052736에 개시된 헬리카제, 변형된 헬리카제 또는 헬리카제 구축물 중 임의의 것일 수 있다.
바람직하게는 헬리카제는 서열식별번호: 413 (Trwc Cba)에 제시된 서열 또는 이의 변이체, 서열식별번호: 406 (Hel308 Mbu)에 제시된 서열 또는 이의 변이체, 또는 서열식별번호: 412 (Dda)에 제시된 서열 또는 이의 변이체를 포함한다. 변이체는 막횡단 포어에 대해 하기에 논의된 방식 중 임의의 것으로 천연 서열과 상이할 수 있다. 서열식별번호: 412의 바람직한 변이체는 (a) E94C 및 A360C 또는 (b) E94C, A360C, C109A 및 C136A를 포함하고, 이어서 (ΔM1)G1G2 (즉, M1 결실 후의 G1 및 G2 부가)를 임의적으로 포함한다.
임의 개수의 헬리카제가 본 발명에 따라 사용될 수 있다. 예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10개 또는 이를 초과하는 개수의 헬리카제가 사용될 수 있다. 일부 실시양태에서, 상이한 개수의 헬리카제가 사용될 수 있다.
바람직하게는 본 발명의 방법은 폴리뉴클레오티드를 2개 이상의 헬리카제와 접촉시키는 것을 포함한다. 전형적으로 2개 이상의 헬리카제는 동일한 헬리카제이다. 2개 이상의 헬리카제는 상이한 헬리카제일 수 있다.
2개 이상의 헬리카제는 상기 언급된 헬리카제의 임의의 조합일 수 있다. 2개 이상의 헬리카제는 2개 이상의 Dda 헬리카제일 수 있다. 2개 이상의 헬리카제는 1개 이상의 Dda 헬리카제 및 1개 이상의 TrwC 헬리카제일 수 있다. 2개 이상의 헬리카제는 동일한 헬리카제의 상이한 변이체들일 수 있다.
바람직하게는 2개 이상의 헬리카제가 서로 부착된다. 더욱 바람직하게는 2개 이상의 헬리카제가 서로 공유결합으로 부착된다. 헬리카제들은 임의의 순서로 임의의 방법을 사용하여 부착될 수 있다. 본 발명에서 사용하기 위한 바람직한 헬리카제 구축물이 국제 출원 번호 PCT/GB2013/051925 (WO 2014/013260으로 공개됨); PCT/GB2013/051924 (WO 2014/013259로 공개됨); PCT/GB2013/051928 (WO 2014/013262로 공개됨) 및 PCT/GB2014/052736에 기술되어 있다.
서열식별번호: 397, 399, 401, 403, 405, 406, 407, 408, 409, 410, 411, 412 또는 413의 변이체는 서열식별번호: 397, 399, 401, 403, 405, 406, 407, 408, 409, 410, 411, 412 또는 413의 것과 상이한 아미노산 서열을 갖고 폴리뉴클레오티드 결합 능력을 유지하는 효소이다. 관련 기술 분야에 공지된 임의의 방법을 사용하여 이를 측정할 수 있다. 예를 들어, 변이체를 폴리뉴클레오티드와 접촉시킬 수 있고, 폴리뉴클레오티드에 결합하고 이를 따라 이동하는 변이체의 능력을 측정할 수 있다. 변이체는 폴리뉴클레오티드의 결합을 용이하게 하고/하거나 높은 염 농도 및/또는 실온에서의 이의 활성을 용이하게 하는 변형을 포함할 수 있다. 변이체는 폴리뉴클레오티드에 결합하지만 (즉, 폴리뉴클레오티드 결합 능력을 유지하지만), 헬리카제처럼 기능하지는 않도록 (즉, 이동을 용이하게 하는 모든 필요한 성분, 예를 들어 ATP 및 Mg2 +가 제공되었을 때 폴리뉴클레오티드를 따라 이동하지 않도록) 변형될 수 있다. 이같은 변형이 관련 기술 분야에 공지되어 있다. 예를 들어, 전형적으로 헬리카제 내의 Mg2 + 결합 도메인의 변형은 헬리카제처럼 기능하지 않는 변이체를 초래한다. 이러한 유형의 변이체는 분자 브레이크처럼 작용할 수 있다 (하기 참조).
서열식별번호: 397, 399, 401, 403, 405, 406, 407, 408, 409, 410, 411, 412 또는 413의 아미노산 서열의 전체 길이에 걸쳐, 바람직하게는 변이체는 이러한 서열에 대해 아미노산 동일성을 기초로 적어도 50% 상동성일 것이다. 더욱 바람직하게는, 변이체 폴리펩티드는 아미노산 동일성을 기초로 서열식별번호: 397, 399, 401, 403, 405, 406, 407, 408, 409, 410, 411, 412 또는 413의 아미노산 서열에 대해 전체 서열에 걸쳐 적어도 55%, 적어도 60%, 적어도 65%, 적어도 70%, 적어도 75%, 적어도 80%, 적어도 85%, 적어도 90%, 더욱 바람직하게는 적어도 95%, 97% 또는 99% 상동성일 수 있다. 200개 이상, 예를 들어 230, 250, 270, 280, 300, 400, 500, 600, 700, 800, 900 또는 1000개 또는 이를 초과하는 개수의 연속된 아미노산의 신장물에 걸쳐 적어도 80%, 예를 들어 적어도 85%, 90% 또는 95%의 아미노산 동일성이 있을 수 있다 ("강한 상동성"). 상기 기술된 바와 같이 상동성이 결정된다. 변이체는 상기 서열식별번호: 390을 참조로 상기 논의된 방식 중 임의의 것으로 야생형 서열과 상이할 수 있다. 효소는 포어에 공유결합으로 부착될 수 있다. 임의의 방법이 효소를 포어에 공유결합으로 부착시키는데 사용될 수 있다.
바람직한 분자 브레이크는 TrwC Cba-Q594A (돌연변이 Q594A가 있는 서열식별번호: 413)이다. 이러한 변이체는 헬리카제처럼 기능하지 않는다 (즉, 폴리뉴클레오티드에 결합하지만, 이동을 용이하게 하는 모든 필요한 성분, 예를 들어 ATP 및 Mg2+가 제공되었을 때 폴리뉴클레오티드를 따라 이동하지 않는다).
가닥 시퀀싱에서, 인가된 전위를 따라 또는 이에 대항하여 폴리뉴클레오티드가 포어를 통해 변위된다. 이중 가닥 폴리뉴클레오티드 상에서 점진적으로 또는 진행성으로 작용하는 엑소뉴클레아제를 인가된 전위 하에 나머지 단일 가닥을 공급하도록 포어의 시스 측에서 사용할 수 있거나 또는 역 전위 하에 트랜스 측에서 사용할 수 있다. 유사하게, 이중 가닥 DNA를 푸는 헬리카제를 유사한 방식으로 또한 사용할 수 있다. 폴리머라제를 또한 사용할 수 있다. 인가된 전위에 대항하는 가닥 변위를 요구하는 시퀀싱 용도에 대한 가능성이 또한 존재하지만, 먼저 DNA가 역 전위 또는 전위 부재 하에 효소에 의해 "포획"되어야 한다. 그 후, 결합 후에 전위가 되돌아가면, 가닥이 시스에서 트랜스로 포어를 통과할 것이고, 전류 흐름에 의해, 쭉 뻗은 형상으로 유지될 것이다. 단일 가닥 DNA 엑소뉴클레아제 또는 단일 가닥 DNA 의존적 폴리머라제가 방금 변위된 단일 가닥을 제어된 단계적 방식으로, 트랜스에서 시스로, 인가된 전위에 대항하여 포어를 통해 되돌려 당기는 분자 모터로서 작용할 수 있다.
임의의 헬리카제가 방법에서 사용될 수 있다. 헬리카제는 포어에 대해 2가지 모드로 작동할 수 있다. 첫째로, 바람직하게는, 헬리카제가 인가된 전압으로부터 초래된 장을 따라 포어를 통해 폴리뉴클레오티드를 이동시키도록, 헬리카제를 사용하여 방법이 수행된다. 이러한 모드에서는, 폴리뉴클레오티드의 5' 끝부분이 먼저 포어 내에 포착되고, 최종적으로 폴리뉴클레오티드가 막의 트랜스 측으로 변위될 때까지 장을 따라 폴리뉴클레오티드가 포어에 통과되도록 헬리카제가 폴리뉴클레오티드를 포어 내로 이동시킨다. 대안적으로, 바람직하게는, 헬리카제가 인가된 전압으로부터 초래된 장에 대항하여 포어를 통해 폴리뉴클레오티드를 이동시키도록 방법이 수행된다. 이러한 모드에서는, 폴리뉴클레오티드의 3' 끝부분이 먼저 포어 내에 포착되고, 최종적으로 막의 시스 측으로 되돌려 배출될 때까지 인가된 장에 대항하여 폴리뉴클레오티드가 포어 밖으로 당겨지도록 헬리카제가 폴리뉴클레오티드를 포어를 통해 이동시킨다.
방법은 반대 방향으로 또한 수행될 수 있다. 폴리뉴클레오티드의 3' 끝부분이 먼저 포어에 포착되고, 최종적으로 폴리뉴클레오티드가 막의 트랜스 측으로 변위될 때까지 장을 따라 폴리뉴클레오티드가 포어에 통과되도록 헬리카제가 폴리뉴클레오티드를 포어 내로 이동시킬 수 있다.
헬리카제에 이동을 용이하게 하는 필수 성분이 제공되지 않거나 또는 헬리카제가 이의 이동을 방해 또는 방지하도록 변형된 경우, 이는 폴리뉴클레오티드에 결합할 수 있고, 인가된 장에 의해 폴리뉴클레오티드가 포어 내로 당겨질 때 폴리뉴클레오티드의 이동을 느리게 하는 브레이크로서 작용할 수 있다. 불활성 모드에서, 폴리뉴클레오티드가 3' 또는 5'을 아래로 하여 포착되는지 여부는 중요하지 않고, 폴리뉴클레오티드를 트랜스 측을 향해 포어 내로 당기는 것은 인가된 장이고, 이때 효소는 브레이크로서 작용한다. 불활성 모드일 때, 헬리카제에 의한 폴리뉴클레오티드의 이동 제어는 래칭(ratcheting), 슬라이딩(sliding) 및 브레이킹(braking)을 포함하는 다수의 방식으로 기술될 수 있다. 헬리카제 활성이 결여된 헬리카제 변이체 또한 이러한 방식으로 사용될 수 있다.
임의의 순서로 폴리뉴클레오티드를 폴리뉴클레오티드 결합 단백질 및 포어와 접촉시킬 수 있다. 폴리뉴클레오티드가 폴리뉴클레오티드 결합 단백질, 예컨대 헬리카제, 및 포어와 접촉될 때, 폴리뉴클레오티드가 먼저 단백질과의 복합체를 형성하는 것이 바람직하다. 포어를 가로질러 전압이 인가되면, 폴리뉴클레오티드/단백질 복합체가 포어와 복합체를 형성하고, 포어를 통한 폴리뉴클레오티드의 이동을 제어한다.
폴리뉴클레오티드 결합 단백질을 사용하는 방법에서의 임의의 단계는 전형적으로 폴리뉴클레오티드 결합 단백질의 작용을 용이하게 하는 효소 보조인자 및 유리 뉴클레오티드 또는 유리 뉴클레오티드 유사체의 존재 하에 수행된다. 유리 뉴클레오티드는 상기 논의된 개별적인 뉴클레오티드 중 임의의 것 중 하나 이상일 수 있다. 유리 뉴클레오티드는 아데노신 모노포스페이트 (AMP), 아데노신 디포스페이트 (ADP), 아데노신 트리포스페이트 (ATP), 구아노신 모노포스페이트 (GMP), 구아노신 디포스페이트 (GDP), 구아노신 트리포스페이트 (GTP), 티미딘 모노포스페이트 (TMP), 티미딘 디포스페이트 (TDP), 티미딘 트리포스페이트 (TTP), 우리딘 모노포스페이트 (UMP), 우리딘 디포스페이트 (UDP), 우리딘 트리포스페이트 (UTP), 시티딘 모노포스페이트 (CMP), 시티딘 디포스페이트 (CDP), 시티딘 트리포스페이트 (CTP), 고리형 아데노신 모노포스페이트 (cAMP), 고리형 구아노신 모노포스페이트 (cGMP), 데옥시아데노신 모노포스페이트 (dAMP), 데옥시아데노신 디포스페이트 (dADP), 데옥시아데노신 트리포스페이트 (dATP), 데옥시구아노신 모노포스페이트 (dGMP), 데옥시구아노신 디포스페이트 (dGDP), 데옥시구아노신 트리포스페이트 (dGTP), 데옥시티미딘 모노포스페이트 (dTMP), 데옥시티미딘 디포스페이트 (dTDP), 데옥시티미딘 트리포스페이트 (dTTP), 데옥시우리딘 모노포스페이트 (dUMP), 데옥시우리딘 디포스페이트 (dUDP), 데옥시우리딘 트리포스페이트 (dUTP), 데옥시시티딘 모노포스페이트 (dCMP), 데옥시시티딘 디포스페이트 (dCDP) 및 데옥시시티딘 트리포스페이트 (dCTP)를 포함하지만, 이에 제한되지 않는다. 유리 뉴클레오티드는 바람직하게는 AMP, TMP, GMP, CMP, UMP, dAMP, dTMP, dGMP 또는 dCMP로부터 선택된다. 유리 뉴클레오티드는 바람직하게는 아데노신 트리포스페이트 (ATP)이다. 효소 보조인자는 구축물이 기능하게 하는 인자이다. 효소 보조인자는 바람직하게는 2가 금속 양이온이다. 2가 금속 양이온은 바람직하게는 Mg2 +, Mn2 +, Ca2 + 또는 Co2 +이다. 효소 보조인자는 가장 바람직하게는 Mg2+이다.
헬리카제
(들) 및 분자 브레이크(들)
바람직한 실시양태에서, 방법은 하기 단계를 포함한다:
(a) 폴리뉴클레오티드에 부착되는 하나 이상의 헬리카제 및 하나 이상의 분자 브레이크를 폴리뉴클레오티드에 제공하는 단계;
(b) 폴리뉴클레오티드를 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉시키고, 하나 이상의 헬리카제 및 하나 이상의 분자 브레이크가 결집되고 양쪽 모두가 포어에 대한, 예컨대 포어를 통한 폴리뉴클레오티드의 이동을 제어하도록 포어를 가로질러 전압을 인가하는 단계;
(c) 폴리뉴클레오티드가 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 여기서 측정은 폴리뉴클레오티드의 하나 이상의 특성을 지시하며, 이에 의해 폴리뉴클레오티드를 특성화하는 단계.
이러한 유형의 방법이 국제 출원 PCT/GB2014/052737에 상세하게 논의되어 있다.
하나 이상의 헬리카제는 상기 논의된 것 중 임의의 것일 수 있다. 하나 이상의 분자 브레이크는 폴리뉴클레오티드에 결합하고 포어를 통한 폴리뉴클레오티드의 이동을 느리게 하는 임의의 화합물 또는 분자일 수 있다. 바람직하게는 하나 이상의 분자 브레이크는 폴리뉴클레오티드에 결합하는 하나 이상의 화합물을 포함한다. 이러한 하나 이상의 화합물은 바람직하게는 하나 이상의 거대고리(macrocycle)이다. 적절한 거대고리는 시클로덱스트린, 칼릭사렌(calixarene), 고리형 펩티드, 크라운 에테르, 쿠커비투릴, 필라라렌(pillararene), 이의 유도체 및 이의 조합물을 포함하지만, 이에 제한되지 않는다. 시클로덱스트린 또는 이의 유도체는 문헌 [Eliseev, A. V., and Schneider, H-J. (1994) J. Am. Chem. Soc. 116, 6081-6088]에 기술된 것들 중 임의의 것일 수 있다. 이러한 작용제는 더욱 바람직하게는 헵타키스-6-아미노-β-시클로덱스트린 (am7-βCD), 6-모노데옥시-6-모노아미노-β-시클로덱스트린 (am1-βCD) 또는 헵타키스-(6-데옥시-6-구아니디노)-시클로덱스트린 (gu7-βCD)이다.
하나 이상의 분자 브레이크는 바람직하게는 하나 이상의 단일 가닥 결합 단백질 (SSB)이다. 더욱 바람직하게는 하나 이상의 분자 브레이크는 음의 순전하가 없는 카르복시-말단 (C-말단) 영역을 포함하는 단일-가닥 결합 단백질 (SSB), 또는 (ii) 이의 C-말단 영역에 C-말단 영역의 음의 순전하를 감소시키는 하나 이상의 변형을 포함하는 변형된 SSB이다. 가장 바람직하게는 하나 이상의 분자 브레이크는 국제 출원 번호 PCT/GB2013/051924 (WO 2014/013259로 공개됨)에 개시된 SSB 중 하나이다.
하나 이상의 분자 브레이크는 바람직하게는 하나 이상의 폴리뉴클레오티드 결합 단백질이다. 폴리뉴클레오티드 결합 단백질은 폴리뉴클레오티드에 결합할 수 있고 포어를 통한 이의 이동을 제어할 수 있는 임의의 단백질일 수 있다. 단백질이 폴리뉴클레오티드에 결합하는지 여부를 결정하는 것은 관련 기술 분야에서 간단하다. 전형적으로 이러한 단백질은 폴리뉴클레오티드와 상호작용하고, 폴리뉴클레오티드의 적어도 하나의 성질을 변형시킨다. 이러한 단백질은 폴리뉴클레오티드를 절단하여 개별적인 뉴클레오티드 또는 더 짧은 뉴클레오티드 쇄, 예컨대 디- 또는 트리뉴클레오티드를 형성시킴으로써 폴리뉴클레오티드를 변형시킬 수 있다. 이러한 모이어티는 특정한 위치로 폴리뉴클레오티드를 배향시키거나 또는 이를 이동시킴으로써, 즉 이의 이동을 제어함으로써 폴리뉴클레오티드를 변형시킬 수 있다.
폴리뉴클레오티드 결합 단백질은 바람직하게는 폴리뉴클레오티드 취급 효소로부터 유래된다. 하나 이상의 분자 브레이크가 상기 논의된 폴리뉴클레오티드 취급 효소 중 임의의 것으로부터 유래될 수 있다. 분자 브레이크로서 작용하는 Phi29 폴리머라제 (서열식별번호: 396)의 변형판이 미국 특허 번호 5,576,204에 개시되어 있다. 바람직하게는 하나 이상의 분자 브레이크가 헬리카제로부터 유래된다.
헬리카제로부터 유래된 임의 개수의 분자 브레이크가 사용될 수 있다. 예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10개 또는 이를 초과하는 개수의 헬리카제가 분자 브레이크로서 사용될 수 있다. 2개 이상의 헬리카제가 분자 브레이크로서 사용되는 경우, 전형적으로 2개 이상의 헬리카제는 동일한 헬리카제이다. 2개 이상의 헬리카제는 상이한 헬리카제일 수 있다.
2개 이상의 헬리카제는 상기 언급된 헬리카제의 임의의 조합일 수 있다. 2개 이상의 헬리카제는 2개 이상의 Dda 헬리카제일 수 있다. 2개 이상의 헬리카제는 1개 이상의 Dda 헬리카제 및 1개 이상의 TrwC 헬리카제일 수 있다. 2개 이상의 헬리카제는 동일한 헬리카제의 상이한 변이체들일 수 있다.
바람직하게는 2개 이상의 헬리카제가 서로 부착된다. 더욱 바람직하게는 2개 이상의 헬리카제가 서로 공유결합으로 부착된다. 헬리카제들은 임의의 순서로 임의의 방법을 사용하여 부착될 수 있다. 바람직하게는, 헬리카제로부터 유래된 하나 이상의 분자 브레이크는 폴리뉴클레오티드 결합 도메인 내의 개구부 (적어도 하나의 형상 상태에서 폴리뉴클레오티드가 이를 통해 헬리카제로부터 풀릴 수 있음)의 크기가 감소되도록 변형된다. 이는 WO 2014/013260에 개시되어 있다.
본 발명에서 사용하기 위한 바람직한 헬리카제 구축물이 국제 출원 번호 PCT/GB2013/051925 (WO 2014/013260로 공개됨); PCT/GB2013/051924 (WO 2014/013259로 공개됨); PCT/GB2013/051928 (WO 2014/013262로 공개됨) 및 PCT/GB2014/052736에 기술되어 있다.
하나 이상의 헬리카제가 활성 모드로 사용되는 경우 (즉, 하나 이상의 헬리카제에 이동을 용이하게 하는 모든 필요한 성분, 예를 들어 ATP 및 Mg2 +가 제공되었을 때), 바람직하게는 하나 이상의 분자 브레이크는 (a) 불활성 모드로 사용되거나 (즉, 이동을 용이하게 하는 필수 성분의 부재 하에 사용되거나 또는 활성 이동이 불가능할 수 있거나), (b) 하나 이상의 분자 브레이크가 하나 이상의 헬리카제에 대해 반대 방향으로 이동하는 활성 모드로 사용되거나, 또는 (c) 하나 이상의 분자 브레이크가 하나 이상의 헬리카제와 동일한 방향으로 하나 이상의 헬리카제보다 더 느리게 이동하는 활성 모드로 사용된다.
하나 이상의 헬리카제가 불활성 모드로 사용되는 경우 (즉, 하나 이상의 헬리카제에 이동을 용이하게 하는 모든 필요한 성분, 예를 들어 ATP 및 Mg2 +가 제공되지 않거나 또는 하나 이상의 헬리카제가 활성 이동이 불가능할 때), 바람직하게는 하나 이상의 분자 브레이크는 (a) 불활성 모드로 사용되거나 (즉, 이동을 용이하게 하는 필수 성분의 부재 하에 사용되거나 또는 활성 이동이 불가능할 수 있거나), 또는 (b) 하나 이상의 분자 브레이크가 포어를 통해 폴리뉴클레오티드와 동일한 방향으로 폴리뉴클레오티드를 따라 이동하는 활성 모드로 사용된다.
하나 이상의 헬리카제 및 하나 이상의 분자 브레이크는 이들이 결집되고 양쪽 모두가 포어를 통한 폴리뉴클레오티드의 이동을 제어하도록 임의의 위치에서 폴리뉴클레오티드에 부착될 수 있다. 하나 이상의 헬리카제 및 하나 이상의 분자 브레이크는 적어도 1개의 뉴클레오티드만큼 떨어지고, 예컨대 적어도 5개, 적어도 10개, 적어도 50개, 적어도 100개, 적어도 500개, 적어도 1000개, 적어도 5000개, 적어도 10,000개, 적어도 50,000개의 뉴클레오티드 또는 이를 초과하는 개수만큼 떨어진다. 방법이 한쪽 끝부분의 Y 어댑터 및 다른 쪽 끝부분의 헤어핀 루프 어댑터가 제공된 이중 가닥 폴리뉴클레오티드를 특성화하는 것에 관련되는 경우, 하나 이상의 헬리카제는 바람직하게는 Y 어댑터에 부착되고, 하나 이상의 분자 브레이크는 바람직하게는 헤어핀 루프 어댑터에 부착된다. 이러한 실시양태에서, 바람직하게는 하나 이상의 분자 브레이크는 폴리뉴클레오티드에 결합하지만 헬리카제처럼 기능하지 않도록 변형된 하나 이상의 헬리카제이다. Y 어댑터에 부착된 하나 이상의 헬리카제는, 바람직하게는, 하기에 더욱 상세하게 논의된 바와 같은 스페이서에서 실속된다. 헤어핀 루프 어댑터에 부착된 하나 이상의 분자 브레이크는, 바람직하게는, 스페이서에서 실속되지 않는다. 바람직하게는, 하나 이상의 헬리카제가 헤어핀 루프에 도달할 때 하나 이상의 헬리카제 및 하나 이상의 분자 브레이크가 결집된다. 하나 이상의 헬리카제는 Y 어댑터가 폴리뉴클레오티드에 부착되기 전에 또는 Y 어댑터가 폴리뉴클레오티드에 부착된 후에 Y 어댑터에 부착될 수 있다. 하나 이상의 분자 브레이크는 헤어핀 루프 어댑터가 폴리뉴클레오티드에 부착되기 전에 또는 헤어핀 루프 어댑터가 폴리뉴클레오티드에 부착된 후에 헤어핀 루프 어댑터에 부착될 수 있다.
하나 이상의 헬리카제 및 하나 이상의 분자 브레이크는 바람직하게는 서로 부착되지 않는다. 하나 이상의 헬리카제 및 하나 이상의 분자 브레이크는 더욱 바람직하게는 서로 공유결합으로 부착되지 않는다. 하나 이상의 헬리카제 및 하나 이상의 분자 브레이크는 바람직하게는 국제 출원 번호 PCT/GB2013/051925 (WO 2014/013260으로 공개됨); PCT/GB2013/051924 (WO 2014/013259로 공개됨); PCT/GB2013/051928 (WO 2014/013262로 공개됨) 및 PCT/GB2014/052736에 기술된 바와 같이 부착되지 않는다.
스페이서
국제 출원 번호 PCT/GB2014/050175에 논의된 바와 같이 하나 이상의 헬리카제가 하나 이상의 스페이서에서 실속될 수 있다. 이러한 국제 출원에 개시된 임의 형상의 하나 이상의 헬리카제 및 하나 이상의 스페이서가 본 발명에서 사용될 수 있다.
인가된 전위로부터 초래된 장을 따라 폴리뉴클레오티드의 일부분이 포어에 진입하고 포어를 통해 이동하는 경우, 폴리뉴클레오티드가 포어를 통해 이동할 때 하나 이상의 헬리카제가 포어에 의해 스페이서를 지나서 이동한다. 이는 폴리뉴클레오티드 (하나 이상의 스페이서를 포함함)는 포어를 통해 이동하고, 하나 이상의 헬리카제는 포어의 상부에 남기 때문이다.
하나 이상의 스페이서는 바람직하게는 폴리뉴클레오티드의 일부분이고, 예를 들어, 이는 폴리뉴클레오티드 서열에 개재된다. 바람직하게는 하나 이상의 스페이서는 폴리뉴클레오티드에 혼성화된 하나 이상의 차단 분자, 예컨대 과속방지턱의 일부분이 아니다.
폴리뉴클레오티드 내에 임의 개수의 스페이서, 예컨대 1, 2, 3, 4, 5, 6, 7, 8, 9, 10개 또는 이를 초과하는 개수의 스페이서가 있을 수 있다. 바람직하게는 폴리뉴클레오티드 내에 2, 4 또는 6개의 스페이서가 있다. 폴리뉴클레오티드의 상이한 영역들 내에 하나 이상의 스페이서가 있을 수 있고, 예컨대 Y 어댑터 및/또는 헤어핀 루프 어댑터 내에 하나 이상의 스페이서가 있을 수 있다.
하나 이상의 스페이서는 하나 이상의 헬리카제가 활성 모드에서도 극복할 수 없는 에너지 장벽을 각각 제공한다. 헬리카제의 견인을 감소시킴으로써 (예를 들어, 폴리뉴클레오티드 내의 뉴클레오티드로부터 염기를 제거함으로써), 또는 하나 이상의 헬리카제의 이동을 물리적으로 차단함으로써 (예를 들어, 부피가 큰 화학 기를 사용함), 하나 이상의 스페이서가 하나 이상의 헬리카제를 실속시킬 수 있다.
하나 이상의 스페이서는 하나 이상의 헬리카제를 실속시키는 임의의 분자 또는 분자 조합을 포함할 수 있다. 하나 이상의 스페이서는 하나 이상의 헬리카제가 폴리뉴클레오티드를 따라 이동하는 것을 방지하는 임의의 분자 또는 분자 조합을 포함할 수 있다. 막횡단 포어 및 인가된 전위의 부재 하에 하나 이상의 헬리카제가 하나 이상의 스페이서에서 실속되었는지 여부를 결정하는 것은 간단하다. 예를 들어, 스페이서를 지나 이동하고 상보적인 DNA 가닥을 치환하는 헬리카제의 능력을 PAGE로 측정할 수 있다.
하나 이상의 스페이서는 전형적으로 선형 분자, 예컨대 중합체를 포함한다. 하나 이상의 스페이서는 전형적으로 폴리뉴클레오티드와 상이한 구조를 갖는다. 예를 들어, 폴리뉴클레오티드가 DNA인 경우, 하나 이상의 스페이서는 전형적으로 DNA가 아니다. 특히, 폴리뉴클레오티드가 데옥시리보핵산 (DNA) 또는 리보핵산 (RNA)인 경우, 하나 이상의 스페이서는 바람직하게는 펩티드 핵산 (PNA), 글리세롤 핵산 (GNA), 트레오스 핵산 (TNA), 잠금 핵산 (LNA), 또는 뉴클레오티드 측쇄가 있는 합성 중합체를 포함한다. 하나 이상의 스페이서는 폴리뉴클레오티드와 반대되는 방향의 하나 이상의 뉴클레오티드를 포함할 수 있다. 예를 들어, 폴리뉴클레오티드가 5'→3' 방향일 때 하나 이상의 스페이서는 3'→5' 방향의 하나 이상의 뉴클레오티드를 포함할 수 있다. 뉴클레오티드는 상기 논의된 것들 중 임의의 것일 수 있다.
하나 이상의 스페이서는 바람직하게는 하나 이상의 니트로인돌, 예컨대 하나 이상의 5-니트로인돌, 하나 이상의 이노신, 하나 이상의 아크리딘, 하나 이상의 2-아미노퓨린, 하나 이상의 2-6-디아미노퓨린, 하나 이상의 5-브로모-데옥시우리딘, 하나 이상의 역전 티미딘 (역전 dT), 하나 이상의 역전 디데옥시-티미딘 (ddT), 하나 이상의 디데옥시-시티딘 (ddC), 하나 이상의 5-메틸시티딘, 하나 이상의 5-히드록시메틸시티딘, 하나 이상의 2'-O-메틸 RNA 염기, 하나 이상의 이소-데옥시시티딘 (이소-dC), 하나 이상의 이소-데옥시구아노신 (이소-dG), 하나 이상의 iSpC3 기 (즉, 당 및 염기가 결여된 뉴클레오티드), 하나 이상의 광-절단성 (PC) 기, 하나 이상의 헥산디올 기, 하나 이상의 스페이서 9 (iSp9) 기, 하나 이상의 스페이서 18 (iSp18) 기, 중합체 또는 하나 이상의 티올 연결부를 포함한다. 하나 이상의 스페이서는 이러한 기들의 임의의 조합을 포함할 수 있다. 다수의 이러한 기들이 IDT® (인테그레이티드 DNA 테크놀러지즈(Integrated DNA Technologies)®)에서 시판된다.
하나 이상의 스페이서는 임의 개수의 이러한 기들을 함유할 수 있다. 예를 들어, 2-아미노퓨린, 2-6-디아미노퓨린, 5-브로모-데옥시우리딘, 역전 dT, ddT, ddC, 5-메틸시티딘, 5-히드록시메틸시티딘, 2'-O-메틸 RNA 염기, 이소-dC, 이소-dG, iSpC3 기, PC 기, 헥산디올 기 및 티올 연결부의 경우, 하나 이상의 스페이서는 바람직하게는 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12개 또는 이를 초과하는 개수를 포함한다. 하나 이상의 스페이서는 바람직하게는 2, 3, 4, 5, 6, 7, 8개 또는 이를 초과하는 개수의 iSp9 기를 포함한다. 하나 이상의 스페이서는 바람직하게는 2, 3, 4, 5 또는 6개 또는 이를 초과하는 개수의 iSp18 기를 포함한다. 가장 바람직한 스페이서는 4개의 iSp18 기이다.
중합체는 바람직하게는 폴리펩티드 또는 폴리에틸렌 글리콜 (PEG)이다. 폴리펩티드는 바람직하게는 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12개 또는 이를 초과하는 개수의 아미노산을 포함한다. PEG는 바람직하게는 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12개 또는 이를 초과하는 개수의 단량체 단위를 포함한다.
바람직하게는 하나 이상의 스페이서는 하나 이상의 무염기성(abasic) 뉴클레오티드 (즉, 핵염기가 결여된 뉴클레오티드), 예컨대 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12개 또는 이를 초과하는 개수의 무염기성 뉴클레오티드를 포함한다. 무염기성 뉴클레오티드에서 핵염기가 -H (idSp) 또는 -OH로 교체될 수 있다. 1개의 이상의 인접한 뉴클레오티드로부터 핵염기를 제거함으로써 무염기성 스페이서가 폴리뉴클레오티드 내로 삽입될 수 있다. 예를 들어, 3-메틸아데닌, 7-메틸구아닌, 1,N6-에테노아데닌 이노신 또는 히포크산틴을 포함하도록 폴리뉴클레오티드가 변형될 수 있고, 인간 알킬아데닌 DNA 글리코실라제 (hAAG)를 사용하여 이러한 뉴클레오티드들로부터 핵염기가 제거될 수 있다. 대안적으로, 우라실을 포함하도록 폴리뉴클레오티드가 변형될 수 있고, 우라실-DNA 글리코실라제 (UDG)로 핵염기가 제거될 수 있다. 한 실시양태에서, 하나 이상의 스페이서는 어떠한 무염기성 뉴클레오티드도 포함하지 않는다.
각각의 선형 분자 스페이서에 의해 (즉, 이러한 스페이서 이전에) 또는 이러한 스페이서 상에서 하나 이상의 헬리카제가 실속될 수 있다. 선형 분자 스페이서가 사용되는 경우, 바람직하게는, 하나 이상의 헬리카제가 이를 지나 이동하여야 하는 각각의 스페이서의 끝부분에 인접하여 폴리뉴클레오티드의 이중 가닥 영역이 폴리뉴클레오티드에 제공된다. 전형적으로 이중 가닥 영역은 인접한 스페이서 상에서 하나 이상의 헬리카제를 실속시키는 것을 돕는다. 방법이 약 100 mM 이하의 염 농도에서 수행되는 경우에 이중 가닥 영역(들)의 존재가 특히 바람직하다. 각각의 이중 가닥 영역은 전형적으로 적어도 10개, 예컨대 적어도 12개의 뉴클레오티드의 길이이다. 본 발명에서 사용되는 폴리뉴클레오티드가 단일 가닥인 경우, 더 짧은 폴리뉴클레오티드를 스페이서에 인접한 영역에 혼성화시킴으로써 이중 가닥 영역이 형성될 수 있다. 더 짧은 폴리뉴클레오티드는 전형적으로는 폴리뉴클레오티드와 동일한 뉴클레오티드로부터 형성되지만, 상이한 뉴클레오티드로부터 형성될 수 있다. 예를 들어, 더 짧은 폴리뉴클레오티드가 LNA로부터 형성될 수 있다.
선형 분자 스페이서가 사용되는 경우, 바람직하게는, 하나 이상의 헬리카제가 이를 지나 이동하여야 하는 끝부분의 반대편인 각각의 스페이서의 끝부분에서 차단 분자가 폴리뉴클레오티드에 제공된다. 이는 하나 이상의 헬리카제가 각각의 스페이서 상에서 실속되어 유지되는 것을 확실히 하는 것을 도울 수 있다. 또한 이는 하나 이상의 헬리카제가 용액에서 확산되어 떨어지는 경우에 하나 이상의 헬리카제를 폴리뉴클레오티드 상에서 유지시키는 것을 도울 수 있다. 차단 분자는 물리적으로 하나 이상의 헬리카제를 실속시키는 하기 논의된 화학 기 중 임의의 것일 수 있다. 차단 분자는 폴리뉴클레오티드의 이중 가닥 영역일 수 있다.
바람직하게는 하나 이상의 스페이서는 물리적으로 하나 이상의 헬리카제를 실속시키는 하나 이상의 화학 기를 포함한다. 바람직하게는 이러한 하나 이상의 화학 기는 하나 이상의 펜던트(pendant) 화학 기이다. 폴리뉴클레오티드 내의 하나 이상의 핵염기에 하나 이상의 화학 기가 부착될 수 있다. 폴리뉴클레오티드 백본에 하나 이상의 화학 기가 부착될 수 있다. 임의 개수의 이러한 화학 기, 예컨대 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12개 또는 이를 초과하는 개수가 존재할 수 있다. 적절한 기는 형광단, 스트렙타비딘 및/또는 비오틴, 콜레스테롤, 메틸렌 블루, 디니트포페놀 (DNP), 디곡시게닌 및/또는 항-디곡시게닌 및 디벤질시클로옥틴 기를 포함하지만, 이에 제한되지 않는다.
폴리뉴클레오티드 내의 상이한 스페이서들이 상이한 실속 분자들을 포함할 수 있다. 예를 들어, 1개의 스페이서는 상기 논의된 선형 분자 중 하나를 포함할 수 있고, 또 다른 스페이서는 물리적으로 하나 이상의 헬리카제를 실속시키는 하나 이상의 화학 기를 포함할 수 있다. 스페이서는 상기 논의된 선형 분자 및 물리적으로 하나 이상의 헬리카제를 실속시키는 하나 이상의 화학 기 중 임의의 것, 예컨대 하나 이상의 무염기성 물질 및 형광단을 포함할 수 있다.
폴리뉴클레오티드의 유형 및 본 발명의 방법이 수행되는 조건에 따라, 적절한 스페이서가 디자인될 수 있다. 대부분의 헬리카제는 DNA에 결합하여 이를 따라 이동하고, 따라서 DNA가 아닌 임의의 것을 사용하여 이를 실속시킬 수 있다. 적절한 분자가 상기에서 논의된다.
본 발명의 방법은 바람직하게는 유리 뉴클레오티드의 존재 및/또는 헬리카제 보조인자의 존재 하에 수행된다. 이는 하기에서 더욱 상세하게 논의된다. 막횡단 포어 및 인가된 전위의 부재 하에, 하나 이상의 스페이서는, 바람직하게는, 유리 뉴클레오티드의 존재 및/또는 헬리카제 보조인자의 존재 하에 하나 이상의 헬리카제를 실속시킬 수 있다.
본 발명의 방법이 하기 논의된 바와 같이 유리 뉴클레오티드 및 헬리카제 보조인자의 존재 하에 수행되는 경우 (하나 이상의 헬리카제가 활성 모드이도록), 하나 이상의 헬리카제가 막횡단 포어와 접촉되고 전위가 인가되기 전에 하나 이상의 헬리카제가 폴리뉴클레오티드 상에서 실속되는 것을 확실히 하기 위해 하나 이상의 더 긴 스페이서가 전형적으로 사용된다. 유리 뉴클레오티드 및 헬리카제 보조인자의 부재 하에 (하나 이상의 헬리카제가 불활성 모드이도록), 하나 이상의 더 짧은 스페이서가 사용될 수 있다.
하나 이상의 헬리카제를 실속시키는 하나 이상의 스페이서의 능력에 염 농도가 또한 영향을 미친다. 막횡단 포어 및 인가된 전위의 부재 하에, 바람직하게는, 하나 이상의 스페이서가 약 100 mM 이하의 염 농도에서 하나 이상의 헬리카제를 실속시킬 수 있다. 본 발명의 방법에서 사용된 염 농도가 더 높을수록, 전형적으로 사용되는 하나 이상의 스페이서의 길이가 더 짧고, 반대도 마찬가지이다.
특색의 바람직한 조합이 하기 표 3에서 제시된다.
<표 3>
2개 이상의 헬리카제를 스페이서를 지나 이동시키는 것에 방법이 관련될 수 있다. 이같은 경우에, 뒤따르는 헬리카제가 포어 및 인가된 전위의 부재 하에 앞선 헬리카제를 스페이서를 지나서 미는 것을 방지하기 위해 스페이서의 길이가 전형적으로 증가된다. 2개 이상의 헬리카제를 하나 이상의 스페이서를 지나 이동시키는 것에 방법이 관련되는 경우, 상기 논의된 스페이서 길이가 적어도 적어도 1.5배, 예컨대 2배, 2.5배 또는 3배 증가될 수 있다. 예를 들어, 2개 이상의 헬리카제를 하나 이상의 스페이서를 지나 이동시키는 것에 방법이 관련되는 경우, 상기 표 3의 세번째 열의 스페이서 길이가 1.5배, 2배, 2.5배 또는 3배 증가될 수 있다.
막
본 발명의 포어는 막 내에 존재할 수 있다. 본 발명의 방법에서, 전형적으로 폴리뉴클레오티드는 막 내의 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉된다. 임의의 막이 본 발명에 따라 사용될 수 있다. 적절한 막이 관련 기술 분야에 널리 공지되어 있다. 막은 바람직하게는 양친매성 층이다. 양친매성 층은 친수성 및 친지성 성질 양쪽 모두를 갖는 양친매성 분자, 예컨대 인지질로부터 형성된 층이다. 양친매성 분자는 합성 또는 천연 발생일 수 있다. 비-천연 발생의 양친매성 물질 및 단층을 형성하는 양친매성 물질이 관련 기술 분야에 공지되어 있고, 예를 들어, 블록 공중합체를 포함한다 (Gonzalez-Perez et al., Langmuir, 2009, 25, 10447-10450). 블록 공중합체는 2개 이상의 단량체 서브유닛이 함께 중합되어 단일 중합체 쇄를 생성하는 중합체 물질이다. 전형적으로 블록 공중합체는 각각의 단량체 서브유닛이 기여하는 성질을 갖는다. 그러나, 블록 공중합체는 개별적인 서브유닛으로부터 형성된 중합체가 보유하지 않는 독특한 성질을 가질 수 있다. 수성 매질에 있을 때 단량체 서브유닛 중 하나는 소수성 (즉, 친지성)인 한편, 다른 서브유닛(들)은 친수성이도록 블록 공중합체가 조작될 수 있다. 이러한 경우에, 블록 공중합체는 양친매성 성질을 보유할 수 있고, 생물학적 막을 모방하는 구조를 형성할 수 있다. 블록 공중합체는 2블록일 수 있지만 (2개의 단량체 서브유닛으로 이루어짐), 또한 2개를 초과하는 단량체 서브유닛으로부터 구축되어, 양친매성 물질로서 거동하는 더욱 복합적인 배열을 형성할 수 있다. 공중합체는 3블록, 4블록 또는 5블록 공중합체일 수 있다. 막은 바람직하게는 3블록 공중합체 막이다.
고세균 양극성 테트라에테르 지질은 지질이 단층 막을 형성하도록 구축된 천연 발생 지질이다. 일반적으로 이러한 지질은 가혹한 생물학적 환경에서 생존하는 극한성 생물, 호열성 생물, 호염성 생물 및 호산성 생물에서 발견된다. 이들의 안정성은 최종 이중층의 융합된 성질로부터 유래되는 것으로 여겨진다. 일반적인 친수성-소수성-친수성 모티프를 갖는 3블록 중합체를 생성시킴으로써 이러한 생물체를 모방하는 블록 공중합체를 구축하는 것이 간단한다. 이러한 물질은 지질 이중층과 유사하게 거동하고 소포에서 층상 막까지의 광범위한 상 거동을 포괄하는 단량체 막을 형성할 수 있다. 이러한 3블록 공중합체로부터 형성된 막은 생물학적 지질 막에 비해 여러 장점을 갖는다. 3블록 공중합체는 합성되기 때문에, 정확한 구축을 주의 깊게 제어하여, 막을 형성하고 포어 및 다른 단백질과 상호작용하는데 요구되는 정확한 쇄 길이 및 성질을 제공할 수 있다.
지질 부재료(sub-material)로 분류되지 않는 서브유닛으로부터 블록 공중합체가 구축될 수도 있다; 예를 들어 소수성 중합체가 실록산 또는 기타 비-탄화수소계 단량체로부터 제조될 수 있다. 또한 블록 공중합체의 친수성 하위-섹션(sub-section)이 낮은 단백질 결합 성질을 보유할 수 있고, 이는 미가공 생물학적 샘플에 노출될 때 고도로 저항성인 막의 생성을 허용한다. 이러한 머리 기 단위 또한 고전적이지 않은 지질 머리 기로부터 유도될 수 있다.
또한 3블록 공중합체 막은 생물학적 지질 막과 비교하여 기계적 및 환경적 안정성이 증가되고, 예를 들어, 조작 온도 또는 pH 범위가 훨씬 더 높다. 블록 공중합체의 합성 성질은 중합체를 기초로 하는 막을 광범위한 용도에 맞추는 플랫폼을 제공한다.
막은 가장 바람직하게는 국제 출원 번호 PCT/GB2013/052766 또는 PCT/GB2013/052767에 개시된 막 중 하나이다.
양친매성 분자는 폴리뉴클레오티드의 커플링을 용이하게 하도록 화학적으로 변형되거나 관능화될 수 있다.
양친매성 층은 단층 또는 이중층일 수 있다. 전형적으로 양친매성 층은 평면형이다. 양친매성 층은 곡선형일 수 있다. 양친매성 층은 지지될 수 있다.
전형적으로 양친매성 막은 천연적으로 이동성이어서, 본질적으로 지질 확산 속도가 약 10-8 cm s-1인 2차원 유체로서 작용한다. 이는 포어 및 커플링된 폴리뉴클레오티드가 전형적으로 양친매성 막 내에서 이동할 수 있다는 것을 의미한다.
막은 지질 이중층일 수 있다. 지질 이중층은 세포막의 모델이고, 광범위한 실험 연구를 위한 우수한 플랫폼으로서의 역할을 한다. 예를 들어, 단일-채널 기록에 의한 막 단백질의 시험관내 연구에 지질 이중층이 사용될 수 있다. 대안적으로, 지질 이중층이 광범위한 물질의 존재를 검출하는 바이오센서로서 사용될 수 있다. 지질 이중층은 임의의 지질 이중층일 수 있다. 적절한 지질 이중층은 평면형 지질 이중층, 지지된 이중층 또는 리포솜을 포함하지만, 이에 제한되지 않는다. 지질 이중층은 바람직하게는 평면형 지질 이중층이다. 적절한 지질 이중층이 국제 출원 번호 PCT/GB08/000563 (WO 2008/102121로 공개됨), 국제 출원 번호 PCT/GB08/004127 (WO 2009/077734로 공개됨) 및 국제 출원 번호 PCT/GB2006/001057 (WO 2006/100484로 공개됨)에 개시되어 있다.
지질 이중층을 형성시키는 방법이 관련 기술 분야에 공지되어 있다. 수성 용액/공기 계면에 대해 수직인 개구부의 양쪽 측면을 지나 지질 단층이 이러한 계면 상에 보유되는 문헌 [Montal and Mueller, Proc. Natl. Acad. Sci. USA., 1972; 69: 3561-3566]의 방법에 의해 지질 이중층이 통상적으로 형성된다. 먼저 지질을 유기 용매에 용해시킨 후, 이러한 용매의 액적이 개구부의 양쪽 측면 상의 수성 용액의 표면 상에서 증발되게 함으로써, 일반적으로 지질이 수성 전해질 용액의 표면 상에 부가된다. 유기 용매가 증발되면, 이중층이 형성될 때까지 개구부의 양쪽 측면 상의 용액/공기 계면이 개구부를 지나 물리적으로 위아래로 이동된다. 개구부를 가로질러 막 내에서 또는 구멍을 가로질러 오목부(recess) 내로 평면형 지질 이중층이 형성될 수 있다.
단백질 포어 삽입에 적절한 양호한 품질의 지질 이중층을 형성시키는 비용 효율적이고 비교적 간단한 방법이기 때문에 몬탈(Montal) & 뮐러(Mueller)의 방법은 대중적이다. 이중층 형성을 위한 기타 통상적인 방법은 팁-침지(tip-dipping), 이중층 페인팅, 및 리포솜 이중층의 패치-클램핑(patch-clamping)을 포함한다.
팁-침지 이중층 형성은 개구부 표면 (예를 들어, 피펫 팁)을 지질 단층을 보유하는 테스트 용액의 표면 상에 접촉시키는 것을 수반한다. 또 다시, 유기 용매에 용해된 지질의 액적을 용액 표면에서 증발되게 함으로써 지질 단층이 용액/공기 계면에서 먼저 생성된다. 그 후, 이중층이 랭뮤어-쉐퍼(Langmuir-Schaefer) 공정에 의해 형성되고, 용액 표면에 대해 개구부를 이동하도록 기계적 오토메이션을 필요로 한다.
페인팅된 이중층의 경우, 수성 테스트 용액에 함침된 개구부에 유기 용매에 용해된 지질의 액적이 직접적으로 적용된다. 페인트브러시 또는 등가물을 사용하여 지질 용액이 개구부 너머로 얇게 확산된다. 용매의 박층화가 지질 이중층의 형성을 초래한다. 그러나, 이중층으로부터 완전히 용매를 제거하는 것이 어렵고, 결과적으로, 이러한 방법에 의해 형성된 이중층은 덜 안정적이고, 전기화학적 측정 동안 노이즈의 경향이 더 크다.
패치-클램핑은 생물학적 세포 막의 연구에서 통상적으로 사용된다. 세포 막이 흡인에 의해 피펫의 끝부분에 클램핑되고, 막의 패치가 개구부 너머로 부착되게 된다. 리포솜을 클램핑하고, 그 후 리포솜이 파열되어 피펫의 개구부 너머로 지질 이중층 실링(sealing)을 남기는 것에 의해 지질 이중층을 생산하기 위해 이러한 방법이 개조되었다. 이러한 방법은 안정적이고, 거대하며 단층인 리포솜, 및 유리 표면이 있는 물질에서의 소형 개구부의 제작을 필요로 한다.
초음파처리, 압출 또는 모자파리(Mozafari) 방법 (Colas et al. (2007) Micron 38:841-847)에 의해 리포솜이 형성될 수 있다.
바람직한 실시양태에서, 국제 출원 번호 PCT/GB08/004127 (WO 2009/077734로 공개됨)에 기술된 바와 같이 지질 이중층이 형성된다. 유리하게는, 이러한 방법에서, 건조된 지질로부터 지질 이중층이 형성된다. 가장 바람직한 실시양태에서, WO2009/077734 (PCT/GB08/004127)에 기술된 바와 같이 구멍을 가로질러 지질 이중층이 형성된다.
2개의 반대되는 지질 층으로부터 지질 이중층이 형성된다. 2개의 지질 층은 이들의 소수성 꼬리 기가 서로를 향하여 소수성 내부를 형성하도록 배열된다. 지질의 친수성 머리 기는 이중층의 각각의 측면 상에서 수성 환경을 향해 밖으로 향한다. 액체 불규칙 상 (유체 층상), 액체 규칙 상, 고체 규칙 상 (층상 겔 상, 상호교차 겔 상) 및 평면형 이중층 결정 (층상 서브-겔 상, 층상 결정질 상)을 포함하지만 이에 제한되지 않는 다수의 지질 상에서 이중층이 존재할 수 있다.
지질 이중층을 형성하는 임의의 지질 조성물이 사용될 수 있다. 필요한 성질, 예컨대 표면 전하, 막 단백질을 지지하는 능력, 패킹(packing) 밀도 또는 기계적 성질을 갖는 지질 이중층이 형성되도록 지질 조성물이 선택된다. 지질 조성물은 하나 이상의 상이한 지질을 포함할 수 있다. 예를 들어, 지질 조성물은 100가지까지의 지질을 함유할 수 있다. 바람직하게는 지질 조성물은 1 내지 10가지의 지질을 함유한다. 지질 조성물은 천연 발생 지질 및/또는 인공 지질을 포함할 수 있다.
지질은 전형적으로 머리 기, 계면 모이어티, 및 동일하거나 상이할 수 있는 2개의 소수성 꼬리 기를 포함한다. 적절한 머리 기는 중성 머리 기, 예컨대 디아실글리세리드 (DG) 및 세라마이드 (CM); 쯔비터이온성 머리 기, 예컨대 포스파티딜콜린 (PC), 포스파티딜에탄올아민 (PE) 및 스핑고미엘린 (SM); 음으로 하전된 머리 기, 예컨대 포스파티딜글리세롤 (PG); 포스파티딜세린 (PS), 포스파티딜이노시톨 (PI), 포스파트산 (PA) 및 카르디올리핀 (CA); 및 양으로 하전된 머리 기, 예컨대 트리메틸암모늄-프로판 (TAP)을 포함하지만, 이에 제한되지 않는다. 적절한 계면 모이어티는 천연-발생 계면 모이어티, 예컨대 글리세롤계 또는 세라마이드계 모이어티를 포함하지만, 이에 제한되지 않는다. 적절한 소수성 꼬리 기는 포화 탄화수소 쇄, 예컨대 라우르산 (n-도데카놀산), 미리스트산 (n-테트라데카논산), 팔미트산 (n-헥사데칸산), 스테아르산 (n-옥타데칸산) 및 아라키드산 (n-에이코사노산); 불포화 탄화수소 쇄, 예컨대 올레산 (시스-9-옥타데칸산); 및 분지형 탄화수소 쇄, 예컨대 피타노일을 포함하지만, 이에 제한되지 않는다. 불포화 탄화수소 쇄에서의 쇄 길이 및 이중 결합의 위치 및 개수가 변할 수 있다. 분지형 탄화수소 쇄에서의 쇄 길이 및 분지, 예컨대 메틸 기의 위치 및 개수가 변할 수 있다. 소수성 꼬리 기는 에테르 또는 에스테르로서 계면 모이어티에 연결될 수 있다. 지질은 미콜산일 수 있다.
지질이 화학적으로 변형될 수도 있다. 지질의 머리 기 또는 꼬리 기가 화학적으로 변형될 수 있다. 머리 기가 화학적으로 변형된 적절한 지질은 PEG-변형 지질, 예컨대 1,2-디아실-sn-글리세로-3-포스포에탄올아민-N-[메톡시(폴리에틸렌 글리콜)-2000]; 관능화된 PEG 지질, 예컨대 1,2-디스테아로일-sn-글리세로-3 포스포에탄올아민-N-[비오티닐(폴리에틸렌 글리콜)2000]; 및 접합용으로 변형된 지질, 예컨대 1,2-디올레오일-sn-글리세로-3-포스포에탄올아민-N-(숙시닐) 및 1,2-디팔미토일-sn-글리세로-3-포스포에탄올아민-N-(비오티닐)을 포함하지만, 이에 제한되지 않는다. 꼬리 기가 화학적으로 변형된 적절한 지질은 중합가능한 지질, 예컨대 1,2-비스(10,12-트리코사디이노일)-sn-글리세로-3-포스포콜린; 플루오린화 지질, 예컨대 1-팔미토일-2-(16-플루오로팔미토일)-sn-글리세로-3-포스포콜린; 중수소화 지질, 예컨대 1,2-디팔미토일-D62-sn-글리세로-3-포스포콜린; 및 에테르-연결 지질, 예컨대 1,2-Di-O-피타닐-sn-글리세로-3-포스포콜린을 포함하지만, 이에 제한되지 않는다. 폴리뉴클레오티드의 커플링을 용이하게 하도록 지질이 화학적으로 변형되거나 관능화될 수 있다.
전형적으로 양친매성 층, 예를 들어 지질 조성물은 층의 성질에 영향을 미칠 하나 이상의 첨가제를 포함한다. 적절한 첨가제는 지방산, 예컨대 팔미트산, 미리스트산 및 올레산; 지방 알콜, 예컨대 팔미틱 알콜, 미리스틱 알콜 및 올레익 알콜; 스테롤, 예컨대 콜레스테롤, 에르고스테롤, 라노스테롤, 시토스테롤 및 스트그마스테롤; 리소인지질, 예컨대 1-아실-2-히드록시-sn-글리세로-3-포스포콜린; 및 세라마이드를 포함하지만, 이에 제한되지 않는다.
또 다른 바람직한 실시양태에서, 막은 고체 상태 층을 포함한다. 마이크로전자 물질, 절연 물질 예컨대 Si3N4, Al2O3, 및 SiO, 유기 및 무기 중합체 예컨대 폴리아미드, 플라스틱 예컨대 테프론(Teflon)® 또는 엘라스토머 예컨대 2-성분 부가-경화 실리콘 고무, 및 유리를 포함하지만 이에 제한되지 않는 유기 및 무기 물질 양쪽 모두로부터 고체 상태 층이 형성될 수 있다. 그래핀으로부터 고체 상태 층이 형성될 수 있다. 적절한 그래핀 층이 국제 출원 번호 PCT/US2008/010637 (WO 2009/035647로 공개됨)에 개시되어 있다. 막이 고체 상태 층을 포함하는 경우, 전형적으로 포어는 고체 상태 층 내에, 예를 들어 고체 상태 층 내의 홀, 웰, 갭, 채널, 트렌치 또는 슬릿 내에 함유된 양친매성 막 또는 층에 존재한다. 통상의 기술자는 적절한 고체 상태/양친매성 하이브리드 시스템을 제조할 수 있다. 적절한 시스템이 WO 2009/020682 및 WO 2012/005857에 개시되어 있다. 상기 논의된 양친매성 막 또는 층 중 임의의 것이 사용될 수 있다.
전형적으로, 방법은 (i) 포어를 포함하는 인공 양친매성 층, (ii) 포어를 포함하는 단리된 천연-발생 지질 이중층, 또는 (iii) 포어가 삽입되어 있는 세포를 사용하여 수행된다. 전형적으로 방법은 인공 양친매성 층, 예컨대 인공 3블록 공중합체 층을 사용하여 수행된다. 이러한 층은 포어에 더하여 다른 막횡단 및/또는 막내 단백질, 뿐만 아니라 다른 분자를 포함할 수 있다. 적절한 기구 및 조건이 하기에서 논의된다. 본 발명의 방법은 전형적으로 시험관 내에서 수행된다.
커플링
바람직하게는, 폴리뉴클레오티드가 포어를 포함하는 막에 커플링된다. 방법은 폴리뉴클레오티드를 포어를 포함하는 막에 커플링시키는 것을 포함할 수 있다. 바람직하게는, 하나 이상의 앵커를 사용하여 폴리뉴클레오티드가 막에 커플링된다. 임의의 공지된 방법을 사용하여 폴리뉴클레오티드가 막에 커플링될 수 있다.
각각의 앵커는 폴리뉴클레오티드에 커플링 (또는 결합)되는 기 및 막에 커플링 (또는 결합)되는 기를 포함한다. 각각의 앵커는 폴리뉴클레오티드 및/또는 막에 공유결합으로 커플링 (또는 결합)될 수 있다. Y 어댑터 및/또는 헤어핀 루프 어댑터가 사용되는 경우, 바람직하게는 이러한 어댑터(들)를 사용하여 폴리뉴클레오티드가 막에 커플링된다.
임의 개수의 앵커, 예컨대 2, 3, 4 또는 이를 초과하는 개수의 앵커를 사용하여 폴리뉴클레오티드가 막에 커플링될 수 있다. 예를 들어, 각각 폴리뉴클레오티드 및 막 양쪽 모두에 별도로 커플링 (또는 결합)되는 2개의 앵커를 사용하여 폴리뉴클레오티드가 막에 커플링될 수 있다.
하나 이상의 앵커는 하나 이상의 헬리카제 및/또는 하나 이상의 분자 브레이크를 포함할 수 있다.
막이 양친매성 층, 예컨대 공중합체 막 또는 지질 이중층인 경우, 바람직하게는, 하나 이상의 앵커는 막 내에 존재하는 폴리펩티드 앵커 및/또는 막 내에 존재하는 소수성 앵커를 포함한다. 소수성 앵커는 바람직하게는 지질, 지방산, 스테롤, 탄소 나노튜브, 폴리펩티드, 단백질 또는 아미노산, 예를 들어 콜레스테롤, 팔미테이트 또는 토코페롤이다. 바람직한 실시양태에서, 하나 이상의 앵커는 포어가 아니다.
하나 이상의 앵커를 형성하도록 막의 성분, 예컨대 양친매성 분자, 공중합체 또는 지질이 화학적으로 변형되거나 관능화될 수 있다. 적절한 화학적 변형 및 막 성분을 관능화시키는 적절한 방식이 하기에서 더욱 상세하게 논의된다. 임의 비율의 막 성분, 예를 들어 적어도 0.01 %, 적어도 0.1 %, 적어도 1 %, 적어도 10%, 적어도 25%, 적어도 50% 또는 100%가 관능화될 수 있다.
폴리뉴클레오티드가 막에 직접적으로 커플링될 수 있다. 폴리뉴클레오티드를 막에 커플링시키는데 사용된 하나 이상의 앵커는 바람직하게는 링커를 포함한다. 하나 이상의 앵커는 하나 이상, 예컨대 2, 3, 4개 또는 이를 초과하는 개수의 링커를 포함할 수 있다. 1개의 링커를 사용하여 1개를 초과하는, 예컨대 2, 3, 4개 또는 이를 초과하는 개수의 폴리뉴클레오티드를 막에 커플링시킬 수 있다.
바람직한 링커는 중합체, 예컨대 폴리뉴클레오티드, 폴리에틸렌 글리콜 (PEG), 다당류 및 폴리펩티드를 포함하지만, 이에 제한되지 않는다. 이러한 링커는 선형, 분지형 또는 원형일 수 있다. 예를 들어, 링커는 원형 폴리뉴클레오티드일 수 있다. 폴리뉴클레오티드가 원형 폴리뉴클레오티드 링커 상의 상보적인 서열에 혼성화할 수 있다.
하나 이상의 앵커 또는 하나 이상의 링커는 절단되어 파괴될 수 있는 성분, 예컨대 제한 부위 또는 광-불안정 기를 포함할 수 있다.
관능화된 링커 및 이들이 분자를 커플링시킬 수 있는 방식이 관련 기술 분야에 공지되어 있다. 예를 들어, 말레이미드로 관능화된 링커는 단백질 내의 시스테인 잔기와 반응하여 이에 부착될 것이다. 본 발명의 맥락에서, 이러한 단백질은 막 내에 존재할 수 있거나, 또는 폴리뉴클레오티드에 커플링 (또는 결합)되는데 사용될 수 있다 이는 하기에서 더욱 상세하게 논의된다.
"자물쇠 & 열쇠" 배열을 사용하여 폴리뉴클레오티드의 가교를 피할 수 있다. 각각의 링커의 한쪽 끝부분만 함께 반응하여 더 긴 링커를 형성할 수 있고, 링커 각각의 다른쪽 끝부분은 각각 폴리뉴클레오티드 또는 막과 반응한다. 이같은 링커가 국제 출원 번호 PCT/GB10/000132 (WO 2010/086602로 공개됨)에 기술되어 있다.
하기 논의된 시퀀싱 실시양태에서 링커 사용이 선호된다. 포어와 상호작용할 때 언커플링(uncoupling)되지 않는다 (즉, 단계 (b) 또는 (e)에서 언커플링되지 않는다)는 점에서 폴리뉴클레오티드가 영구적으로 막에 직접적으로 커플링되면, 막과 포어 사이의 거리로 인해 시퀀싱 시행이 폴리뉴클레오티드 끝부분까지 계속될 수 없기 때문에 일부 서열 데이터가 손실될 것이다. 링커가 사용되면, 폴리뉴클레오티드가 완료까지 프로세싱될 수 있다.
커플링은 영구적 또는 안정적일 수 있다. 달리 말하면, 포어와 상호작용할 때 폴리뉴클레오티드가 막에 커플링되어 남도록 커플링될 수 있다.
커플링은 일시적일 수 있다. 달리 말하면, 포어와 상호작용할 때 폴리뉴클레오티드가 막으로부터의 커플링으로부터 벗어나도록 커플링될 수 있다.
특정 용도, 예컨대 앱타머 검출의 경우, 일시적인 성질의 커플링이 선호된다. 영구적이거나 안정적인 링커가 폴리뉴클레오티드의 5' 또는 3' 끝부분 중 하나에 직접적으로 부착되고, 링커가 막과 막횡단 포어의 채널 사이의 거리보다 짧으면, 시퀀싱 시행이 폴리뉴클레오티드 끝부분까지 계속될 수 없기 때문에 일부 서열 데이터가 손실될 것이다. 커플링이 일시적이면, 커플링된 끝부분이 무작위로 막으로부터 자유로워졌을 때, 폴리뉴클레오티드가 완료까지 프로세싱될 수 있다. 영구적/안정적 또는 일시적 연결을 형성하는 화학 기가 하기에서 더욱 상세하게 논의된다. 콜레스테롤 또는 지방 아실 쇄를 사용하여 폴리뉴클레오티드가 양친매성 층 또는 3블록 공중합체 막에 일시적으로 커플링될 수 있다. 탄소 원자 6 내지 30개 길이의 임의의 지방 아실 쇄, 예컨대 헥사데칸산이 사용될 수 있다.
바람직한 실시양태에서, 폴리뉴클레오티드, 예컨대 핵산이 양친매성 층 예컨대 3블록 공중합체 막 또는 지질 이중층에 커플링된다. 합성 지질 이중층에 핵산을 커플링시키는 것이 다양한 상이한 테더링(tethering) 전략으로 기존에 수행되었다. 이들이 하기 표 4에서 요약된다.
<표 4>
변형된 포스포르아미디트를 합성 반응에서 사용하여 합성 폴리뉴클레오티드 및/또는 링커가 관능화될 수 있고, 이는 적절한 앵커링 기, 예컨대 콜레스테롤, 토코페롤, 팔미테이트, 티올, 지질 및 비오틴 기를 직접적으로 부가하는 것에 대해 쉽게 상용성이다. 이러한 상이한 부착 화학들은 폴리뉴클레오티드에 부착하는 것에 대한 한 벌의 선택권을 제공한다. 각각의 상이한 변형 기는 폴리뉴클레오티드를 약간 상이한 방식으로 커플링시키고, 폴리뉴클레오티드에 대한 상이한 체류 시간을 막에 제공하도록 커플링이 항상 영구적이지는 않다. 일시적 커플링의 장점이 상기에 논의되어 있다.
상보적인 반응성 기 또는 앵커링 기가 폴리뉴클레오티드에 부가될 수 있는 한, 폴리뉴클레오티드를 링커 또는 관능화된 막에 커플링시키는 것이 다수의 다른 수단에 의해서 또한 달성될 수 있다. 폴리뉴클레오티드의 한쪽 끝부분에 반응성 기를 부가하는 것이 기존에 보고되었다. T4 폴리뉴클레오티드 키나제 및 ATPγS를 사용하여 티올 기가 ssDNA 또는 dsDNA의 5'에 부가될 수 있다 (Grant, G. P. and P. Z. Qin (2007). "A facile method for attaching nitroxide spin labels at the 5' terminus of nucleic acids." Nucleic Acids Res 35(10): e77). T4 폴리뉴클레오티드 키나제 및 γ-[2-아지도에틸]-ATP 또는 γ-[6-아지도헥실]-ATP를 사용하여 아지드 기가 ssDNA 또는 dsDNA의 5'-포스페이트에 부가될 수 있다. 티올 또는 클릭(Click)화학을 사용하여, 티올, 아이오도아세트아미드 OPSS 또는 말레이미드 기 (티올에 대해 반응성) 또는 DIBO (디벤조시클로옥시틴) 또는 알킨 기 (아지드에 대해 반응성)를 함유하는 테더가 폴리뉴클레오티드에 공유결합으로 부착될 수 있다. 변형된 올리고뉴클레오티드가 ssDNA의 3'에 혼입되도록 말단 트랜스퍼라제를 사용하여 더욱 다양한 선택의 화학 기, 예컨대 비오틴, 티올 및 형광단이 부가될 수 있다 (Kumar, A., P. Tchen, et al. (1988). "Nonradioactive labeling of synthetic oligonucleotide probes with terminal deoxynucleotidyl transferase." Anal Biochem 169(2): 376-82). 스트렙타비딘/비오틴 및/또는 스트렙타비딘/데스티오비오틴 커플링이 임의의 기타 폴리뉴클레오티드에 사용될 수 있다. 하기의 실시예에서 어떻게 스트렙타비딘/비오틴 및 스트렙타비딘/데스티오비오틴을 사용하여 폴리뉴클레오티드를 막에 커플링시킬 수 있는지가 기술된다. 말단 트랜스퍼라제를 적절하게 변형된 뉴클레오티드와 함께 사용하여 앵커가 폴리뉴클레오티드에 직접적으로 부가될 수 있는 것이 또한 가능할 수 있다 (예를 들어, 콜레스테롤 또는 팔미테이트).
바람직하게는, 하나 이상의 앵커는 혼성화를 통해 폴리뉴클레오티드를 막에 커플링시킨다. 하나 이상의 앵커에서의 혼성화는 상기 논의된 바와 같은 일시적인 방식으로 커플링을 허용한다. 혼성화는 하나 이상의 앵커의 임의의 부분에, 예컨대 하나 이상의 앵커와 폴리뉴클레오티드 사이에, 하나 이상의 앵커 내에, 또는 하나 이상의 앵커와 막 사이에 존재할 수 있다. 예를 들어, 링커는 함께 혼성화된 2개 이상의 폴리뉴클레오티드, 예컨대 3, 4 또는 5개의 폴리뉴클레오티드를 포함할 수 있다. 하나 이상의 앵커가 폴리뉴클레오티드에 혼성화할 수 있다. 하나 이상의 앵커가 폴리뉴클레오티드에 직접적으로, 또는 폴리뉴클레오티드에 부착된 Y 어댑터 및/또는 리더 서열에 직접적으로, 또는 폴리뉴클레오티드에 부착된 헤어핀 루프 어댑터 (하기 논의된 바와 같음)에 직접적으로 혼성화할 수 있다. 대안적으로, 하나 이상의 앵커가 폴리뉴클레오티드, 폴리뉴클레오티드에 부착된 Y 어댑터 및/또는 리더 서열, 또는 폴리뉴클레오티드에 부착된 헤어핀 루프 어댑터 (하기 논의된 바와 같음)에 혼성화된 1개 이상, 예컨대 2개 또는 3개의 중간체 폴리뉴클레오티드 (또는 "부목(splint)"에 혼성화될 수 있다.
하나 이상의 앵커는 단일 가닥 또는 이중 가닥 폴리뉴클레오티드를 포함할 수 있다. 앵커의 한 부분이 단일 가닥 또는 이중 가닥 폴리뉴클레오티드에 라이게이션될 수 있다. 짧은 ssDNA 조각의 라이게이션이 T4 RNA 리가제 I을 사용하여 보고되었다 (Troutt, A. B., M. G. McHeyzer-Williams, et al. (1992). "Ligation-anchored PCR: a simple amplification technique with single-sided specificity." Proc Natl Acad Sci U S A 89(20): 9823-5). 대안적으로, 단일 가닥 또는 이중 가닥 폴리뉴클레오티드가 이중 가닥 폴리뉴클레오티드에 라이게이션된 후, 2개의 가닥이 열 변성 또는 화학 변성에 의해 분리될 수 있다. 이중 가닥 폴리뉴클레오티드에, 단일 가닥 폴리뉴클레오티드 조각을 듀플렉스의 한쪽 또는 양쪽 끝부분에 부가하거나 또는 이중 가닥 폴리뉴클레오티드를 한쪽 또는 양쪽 끝부분에 부가할 수 있다. 단일 가닥 폴리뉴클레오티드를 이중 가닥 폴리뉴클레오티드에 부가하는 경우, 이는 단일 가닥 폴리뉴클레오티드의 다른 영역에 대한 라이게이션에 대해서와 같이 T4 RNA 리가제 I을 사용하여 달성될 수 있다. 이중 가닥 폴리뉴클레오티드를 이중 가닥 폴리뉴클레오티드에 부가하는 경우, 라이게이션은 상보적인 3' dA / dT 꼬리가 폴리뉴클레오티드 및 부가되는 폴리뉴클레오티드 상에 각각 있는 "평활-말단(blunt-ended)"일 수 있거나 (콘카테머(concatemer) 또는 이량체 형성을 방지하기 위해 다수의 샘플 프렙(prep) 용도에 대해 일상적으로 행해지는 바와 같음), 또는 폴리뉴클레오티드의 제한 소화 및 상용성 어댑터의 라이게이션에 의해 생성된 "점착성 말단(sticky-end)"을 사용할 수 있다. 그 후, 듀플렉스가 용융되면, 각각의 단일 가닥은 단일 가닥 폴리뉴클레오티드가 라이게이션에 사용된 경우에는 5' 또는 3' 변형이, 이중 가닥 폴리뉴클레오티드가 라이게이션에 사용된 경우에는 5' 끝부분, 3' 끝부분 또는 양쪽 모두에서의 변형이 있을 것이다.
폴리뉴클레오티드가 합성 가닥이면, 하나 이상의 앵커가 폴리뉴클레오티드의 화학적 합성 동안 혼입될 수 있다. 예를 들어, 반응성 기가 부착되어 있는 프라이머를 사용하여 폴리뉴클레오티드가 합성될 수 있다.
아데닐화 폴리뉴클레오티드는 아데노신-모노포스페이트가 폴리뉴클레오티드의 5'-포스페이트에 부착되어 있는, 라이게이션 반응에서의 중간체이다. 다양한 키트, 예컨대 NEB의 5' DNA 아데닐화 키트가 이러한 중간체의 생성을 위해 입수가능하다. 이러한 반응에서 ATP를 변형된 뉴클레오티드 트리포스페이트로 치환함으로써, 반응성 기 (예컨대 티올, 아민, 비오틴, 아지드 등)를 폴리뉴클레오티드의 5'에 부가하는 것이 가능할 수 있다. 5' DNA 아데닐화 키트를 적절하게 변형된 뉴클레오티드와 함께 사용하여 앵커가 폴리뉴클레오티드에 직접적으로 부가될 수 있는 것이 또한 가능하다 (예를 들어 콜레스테롤 또는 팔미테이트).
게놈 DNA의 절편의 증폭을 위한 통상적인 기술은 폴리머라제 연쇄 반응 (PCR)을 사용하는 것이다. 여기에서, 2개의 합성 올리고뉴클레오티드 프라이머를 사용하여, DNA의 동일한 절편의 다수의 카피가 생성될 수 있고, 이때 각각의 카피에 대해, 듀플렉스 내의 각각의 가닥의 5'이 합성 폴리뉴클레오티드일 것이다. 폴리머라제를 사용함으로써 단일 또는 다중 뉴클레오티드가 단일 또는 이중 가닥 DNA의 3' 끝부분에 부가될 수 있다. 사용될 수 있는 폴리머라제의의 예는 말단 트랜스퍼라제, 클레나우(Klenow) 및 이. 콜라이 폴리(A) 폴리머라제를 포함하지만, 이에 제한되지 않는다. 이러한 반응에서 ATP를 변형된 뉴클레오티드 트리포스페이트로 치환함으로써, 앵커, 예컨대 콜레스테롤, 티올, 아민, 아지드, 비오틴 또는 지질이 이중 가닥 폴리뉴클레오티드 내로 혼입될 수 있다. 따라서, 증폭된 폴리뉴클레오티드의 각각의 카피는 앵커를 함유할 것이다.
이상적으로는, 폴리뉴클레오티드를 관능화시켜야 하지 않으면서 폴리뉴클레오티드가 막에 커플링된다. 이는 하나 이상의 앵커, 예컨대 폴리뉴클레오티드 결합 단백질 또는 화학 기를 막에 커플링시키고, 하나 이상의 앵커가 폴리뉴클레오티드와 상호작용하게 함으로써, 또는 막을 관능화시킴으로써 달성될 수 있다. 본원에 기술된 방법 중 임의의 것에 의해 하나 이상의 앵커가 막에 커플링될 수 있다. 특히, 하나 이상의 앵커가 하나 이상의 링커, 예컨대 말레이미드 관능화 링커를 포함할 수 있다.
이러한 실시양태에서, 폴리뉴클레오티드는 전형적으로 RNA, DNA, PNA, TNA 또는 LNA이고, 이중 또는 단일 가닥일 수 있다. 이러한 실시양태는 게놈 DNA 폴리뉴클레오티드에 특히 적합하다.
하나 이상의 앵커는 단일 또는 이중 가닥 폴리뉴클레오티드, 폴리뉴클레오티드 내의 특정 뉴클레오티드 서열, 또는 폴리뉴클레오티드 내의 변형된 뉴클레오티드의 패턴, 또는 폴리뉴클레오티드 상에 존재하는 임의의 기타 리간드에 커플링되거나, 이에 결합하거나 또는 이와 상호작용하는 임의의 기를 포함할 수 있다.
앵커에서 사용하기 위한 적절한 결합 단백질은 이. 콜라이 단일 가닥 결합 단백질, P5 단일 가닥 결합 단백질, T4 gp32 단일 가닥 결합 단백질, TOPO V dsDNA 결합 영역, 인간 히스톤 단백질, 이. 콜라이 HU DNA 결합 단백질 및 기타 고세균, 원핵생물 또는 진핵생물 단일 가닥 또는 이중 가닥 폴리뉴클레오티드 (또는 핵산) 결합 단백질 (하기에 열거된 것들을 포함함)을 포함하지만, 이에 제한되지 않는다.
특정 뉴클레오티드 서열은 전사 인자, 리보솜, 엔도뉴클레아제, 토포이소머라제 또는 복제 개시 인자가 인식하는 서열일 수 있다. 변형된 뉴클레오티드의 패턴은 메틸화 또는 손상 패턴일 수 있다.
하나 이상의 앵커는 폴리뉴클레오티드에 커플링되거나, 이에 결합하거나, 이에 삽입(intercalation)되거나 또는 이와 상호작용하는 임의의 기를 포함할 수 있다. 이러한 기는 정전기, 수소 결합 또는 반데르발스 상호작용을 통해 폴리뉴클레오티드에 삽입되거나 또는 이와 상호작용할 수 있다. 이같은 기는 리신 단량체, 폴리-리신 (이는 ssDNA 또는 dsDNA와 상호작용할 것이다), 에티듐 브로마이드 (이는 dsDNA에 삽입될 것이다), 유니버설(universal) 염기 또는 유니버설 뉴클레오티드 (이는 임의의 폴리뉴클레오티드와 혼성화할 수 있다) 및 오스뮴 착물 (이는 메틸화 염기에 반응할 수 있다)를 포함한다. 따라서, 막에 부착된 하나 이상의 유니버설 뉴클레오티드를 사용하여 폴리뉴클레오티드가 막에 커플링될 수 있다. 하나 이상의 링커를 사용하여 각각의 유니버설 뉴클레오티드가 막에 커플링될 수 있다. 유니버설 뉴클레오티드는 바람직하게는 하기 핵염기 중 하나를 포함한다: 히포크산틴, 4-니트로인돌, 5-니트로인돌, 6-니트로인돌, 포르밀인돌, 3-니트로피롤, 니트로이미다졸, 4-니트로피라졸, 4-니트로벤즈이미다졸, 5-니트로인다졸, 4-아미노벤즈이미다졸 또는 페닐 (C6-방향족 고리). 유니버설 뉴클레오티드는 더욱 바람직하게는 하기 뉴클레오시드 중 하나를 포함한다: 2'-데옥시이노신, 이노신, 7-데아자-2'-데옥시이노신, 7-데아자-이노신, 2-아자-데옥시이노신, 2-아자-이노신, 2-O'-메틸이노신, 4-니트로인돌 2'-데옥시리보뉴클레오시드, 4-니트로인돌 리보뉴클레오시드, 5-니트로인돌 2'-데옥시리보뉴클레오시드, 5-니트로인돌 리보뉴클레오시드, 6-니트로인돌 2'-데옥시리보뉴클레오시드, 6-니트로인돌 리보뉴클레오시드, 3-니트로피롤 2'-데옥시리보뉴클레오시드, 3-니트로피롤 리보뉴클레오시드, 히포크산틴의 무고리(acyclic) 당 유사체, 니트로이미다졸 2'-데옥시리보뉴클레오시드, 니트로이미다졸 리보뉴클레오시드, 4-니트로피라졸 2'-데옥시리보뉴클레오시드, 4-니트로피라졸 리보뉴클레오시드, 4-니트로벤즈이미다졸 2'-데옥시리보뉴클레오시드, 4-니트로벤즈이미다졸 리보뉴클레오시드, 5-니트로인다졸 2'-데옥시리보뉴클레오시드, 5-니트로인다졸 리보뉴클레오시드, 4-아미노벤즈이미다졸 2'-데옥시리보뉴클레오시드, 4-아미노벤즈이미다졸 리보뉴클레오시드, 페닐 C-리보뉴클레오시드, 페닐 C-2'-데옥시리보실 뉴클레오시드, 2'-데옥시네불라린, 2'-데옥시이소구아노신, K-2'-데옥시리보스, P-2'-데옥시리보스 및 피롤리딘. 유니버설 뉴클레오티드는 더욱 바람직하게는 2'-데옥시이노신을 포함한다. 유니버설 뉴클레오티드는 더욱 바람직하게는 IMP 또는 dIMP이다. 유니버설 뉴클레오티드는 가장 바람직하게는 dPMP (2'-데옥시-P-뉴클레오시드 모노포스페이트) 또는 dKMP (N6-메톡시-2,6-디아미노퓨린 모노포스페이트)이다.
후그스틴(Hoogsteen) 수소 결합 (2개의 핵염기가 수소 결합에 의해 함께 유지됨) 또는 역 후그스틴 수소 결합 (1개의 핵염기가 다른 핵염기에 대해 180° 회전됨)을 통해 하나 이상의 앵커가 폴리뉴클레오티드에 커플링 (또는 결합)될 수 있다. 예를 들어, 폴리뉴클레오티드와 후그스틴 수소 결합 또는 역 후그스틴 수소 결합을 형성하는 하나 이상의 뉴클레오티드, 하나 이상의 올리고뉴클레오티드 또는 하나 이상의 폴리뉴클레오티드를 하나 이상의 앵커가 포함할 수 있다. 이러한 유형의 수소 결합은 제3 폴리뉴클레오티드 가닥이 이중 가닥 나선을 휘감아 트리플렉스를 형성할 수 있게 한다. 이중 가닥 듀플렉스와 함께 트리플렉스를 형성함으로써 하나 이상의 앵커가 이중 가닥 폴리뉴클레오티드에 커플링 (또는 결합)될 수 있다.
이러한 실시양태에서, 막 성분의 적어도 1%, 적어도 10%, 적어도 25%, 적어도 50% 또는 100%가 관능화될 수 있다.
하나 이상의 앵커가 단백질을 포함하는 경우, 예를 들어 막과 상용성인 외부의 소수성 영역이 단백질에 이미 있으면, 앵커가 추가 관능화 없이 막 내로 직접적으로 앵커링될 수 있다. 이같은 단백질의 예는 막횡단 단백질, 막내 단백질 및 막 단백질을 포함하지만, 이에 제한되지 않는다. 대안적으로, 막과 상용성인 소수성 영역이 유전적으로 융합된 단백질이 발현될 수 있다. 이같은 소수성 단백질 영역이 관련 기술 분야에 공지되어 있다.
바람직하게는, 하나 이상의 앵커가 막과의 접촉 전에 폴리뉴클레오티드와 혼합되지만, 하나 이상의 앵커가 막과 접촉되고, 이어서 폴리뉴클레오티드와 접촉될 수 있다.
또 다른 측면에서, 특정 결합 기에 의해 인식될 수 있도록, 상기 기술된 방법을 사용하여 폴리뉴클레오티드가 관능화될 수 있다. 구체적으로, 폴리뉴클레오티드가 리간드 예컨대 비오틴 (스트렙타비딘에 결합하기 위해), 아밀로스 (말토스 결합 단백질 또는 융합 단백질에 결합하기 위해), Ni-NTA (폴리-히스티딘 또는 폴리-히스티딘 태그가 부착된 단백질에 결합하기 위해) 또는 펩티드 (예컨대 항원)로 관능화될 수 있다.
바람직한 실시양태에 따르면, 포어 내로 우선적으로 관통되는 리더 서열에 폴리뉴클레오티드가 부착되었을 때 폴리뉴클레오티드를 막에 커플링시키는데 하나 이상의 앵커가 사용될 수 있다. 하기에서 리더 서열이 더욱 상세하게 논의된다. 바람직하게는, 포어 내로 우선적으로 관통되는 리더 서열에 폴리뉴클레오티드가 부착된다 (예컨대 라이게이션된다). 이같은 리더 서열은 동종중합체 폴리뉴클레오티드 또는 무염기 영역을 포함할 수 있다. 전형적으로 리더 서열은 직접적으로 또는 하나 이상의 중간체 폴리뉴클레오티드 (또는 부목)을 통해 하나 이상의 앵커에 혼성화되도록 디자인된다. 이같은 경우에, 하나 이상의 앵커는 리더 서열 내의 서열 또는 하나 이상의 중간체 폴리뉴클레오티드 (또는 부목) 내의 서열에 대해 상보적인 폴리뉴클레오티드 서열을 전형적으로 포함한다. 이같은 경우에, 하나 이상의 부목은 리더 서열 내의 서열에 대해 상보적인 폴리뉴클레오티드 서열을 전형적으로 포함한다.
화학적 부착에서 사용되는 분자의 예는 EDC (1-에틸-3-[3-디메틸아미노프로필]카르보디이미드 히드로클로라이드)이다. 시판되는 키트 (써모 피어스(Thermo Pierce), 파트(Part) 번호 22980)를 사용하여 반응성 기가 폴리뉴클레오티드의 5'에 부가될 수도 있다. 적절한 방법은 히스티딘 잔기 및 Ni-NTA를 사용하는 일시적인 친화력 부착, 뿐만 아니라 반응성 시스테인, 리신 또는 비-천연 아미노산에 의한 더욱 강건한 공유결합 부착을 포함하지만, 이에 제한되지 않는다.
이중 가닥 폴리뉴클레오티드
폴리뉴클레오티드는 이중 가닥일 수 있다. 폴리뉴클레오티드가 이중 가닥이면, 바람직하게는 방법은 접촉 단계 전에 가교 모이어티, 예컨대 헤어핀 루프를 폴리뉴클레오티드의 한쪽 끝부분에 라이게이션시키는 것을 추가로 포함한다. 그 후, 폴리뉴클레오티드가 본 발명에 따른 포어와 접촉될 때 또는 접촉되기 전에 폴리뉴클레오티드의 2개의 가닥이 분리될 수 있다. 포어를 통한 폴리뉴클레오티드 이동이 폴리뉴클레오티드 결합 단백질, 예컨대 헬리카제, 또는 분자 브레이크에 의해 제어될 때 2개의 가닥이 분리될 수 있다.
이러한 방식으로 이중 가닥 구축물 상의 양쪽 가닥을 연결하고 조사하는 것은 특성화의 효율 및 정확도를 증가시킨다.
가교 모이어티는 표적 폴리뉴클레오티드의 2개의 가닥을 연결할 수 있다. 전형적으로 가교 모이어티는 표적 폴리뉴클레오티드의 2개의 가닥을 공유결합으로 연결시킨다. 가교 모이어티는 표적 폴리뉴클레오티드의 2개의 가닥을 연결시킬 수 있는 임의의 것일 수 있고, 단 가교 모이어티는 막횡단 포어를 통한 단일 가닥 폴리뉴클레오티드의 이동을 방해하지 않는다.
관련 기술 분야에 공지된 임의의 적절한 수단에 의해 가교 모이어티가 표적 폴리뉴클레오티드에 연결될 수 있다. 가교 모이어티가 별도로 합성되어, 표적 폴리뉴클레오티드에 화학적으로 부착되거나 또는 효소에 의해 라이게이션될 수 있다. 대안적으로, 표적 폴리뉴클레오티드의 프로세싱에서 가교 모이어티가 생성될 수 있다.
가교 모이어티는 표적 폴리뉴클레오티드의 한쪽 끝부분에서 또는 이에 인접하여 표적 폴리뉴클레오티드에 연결된다. 바람직하게는 가교 모이어티는 표적 폴리뉴클레오티드의 끝부분으로부터 10개의 뉴클레오티드 이내에서 표적 폴리뉴클레오티드에 연결된다.
적절한 가교 모이어티는 중합체 링커, 화학 링커, 폴리뉴클레오티드 또는 폴리펩티드를 포함하지만, 이에 제한되지 않는다. 바람직하게는, 가교 모이어티는 DNA, RNA, 변형된 DNA (예컨대 무염기 DNA), RNA, PNA, LNA 또는 PEG를 포함한다. 가교 모이어티는 더욱 바람직하게는 DNA 또는 RNA이다.
가교 모이어티는 가장 바람직하게는 헤어핀 루프 또는 헤어핀 루프 어댑터이다. 관련 기술 분야에 공지된 방법을 사용하여 적절한 헤어핀 어댑터가 디자인될 수 있다. 헤어핀 루프는 임의의 길이일 수 있다. 헤어핀 루프는 전형적으로 110개 이하의 뉴클레오티드, 예컨대 100개 이하의 뉴클레오티드, 90개 이하의 뉴클레오티드, 80개 이하의 뉴클레오티드, 70개 이하의 뉴클레오티드, 60개 이하의 뉴클레오티드, 50개 이하의 뉴클레오티드, 40개 이하의 뉴클레오티드, 30개 이하의 뉴클레오티드, 20개 이하의 뉴클레오티드 또는 10개 이하의 뉴클레오티드의 길이이다. 바람직하게는 헤어핀 루프는 약 1 내지 110개, 2 내지 100개, 5 내지 80개 또는 6 내지 50개의 뉴클레오티드의 길이이다. 루프가 어댑터의 차별적 선택성에서 수반되면 더 긴 헤어핀 루프 길이, 예컨대 50 내지 110개의 뉴클레오티드가 바람직하다. 유사하게, 루프가 하기 논의된 바와 같은 선택성 결합에서 수반되지 않으면 더 짧은 헤어핀 루프 길이, 예컨대 1 내지 5개의 뉴클레오티드가 바람직하다.
제1 및/또는 제2 폴리뉴클레오티드의 한쪽 끝부분, 즉 5' 또는 3' 끝부분에서 헤어핀 어댑터가 라이게이션될 수 있다. 관련 기술 분야에 공지된 임의의 방법을 사용하여 헤어핀 어댑터가 제1 및/또는 제2 폴리뉴클레오티드에 라이게이션될 수 있다. 리가제, 예컨대 T4 DNA 리가제, 이. 콜라이 DNA 리가제, Taq DNA 리가제, Tma DNA 리가제 및 9°N DNA 리가제를 사용하여 헤어핀 어댑터가 라이게이션될 수 있다.
관련 기술 분야에 공지된 임의의 방법을 사용하여 폴리뉴클레오티드의 2개의 가닥이 분리될 수 있다. 예를 들어, 폴리뉴클레오티드 결합 단백질에 의해 또는 탈혼성화를 선호하는 조건 (탈혼성화를 선호하는 조건의 예는 고온, 높은 pH, 및 수소 결합 또는 염기 쌍 형성을 파괴할 수 있는 작용제 예컨대 포름아미드 및 요소의 첨가를 포함하지만 이에 제한되지 않는다)을 사용하여 이들이 분리될 수 있다.
바람직하게는 헤어핀 어댑터는 선택가능한 결합 모이어티를 포함한다. 이는 제1 및/또는 제2 폴리뉴클레오티드가 정제 또는 단리될 수 있게 한다. 선택가능한 결합 모이어티는 이의 결합 성질을 기초로 선택될 수 있는 모이어티이다. 따라서, 바람직하게는 선택가능한 결합 모이어티는 표면에 특이적으로 결합하는 모이어티이다. 본 발명에서 사용된 임의의 다른 모이어티보다 더 큰 정도로 표면에 결합하면, 선택가능한 결합 모이어티가 표면에 특이적으로 결합하는 것이다. 바람직한 실시양태에서, 이러한 모이어티는 본 발명에서 사용된 다른 모이어티가 결합하지 않는 표면에 결합한다.
적절한 선택적 결합 모이어티가 관련 기술 분야에 공지되어 있다. 바람직한 선택적 결합 모이어티는 비오틴, 폴리뉴클레오티드 서열, 항체, 항체 단편, 예컨대 Fab 및 ScSv, 항원, 폴리뉴클레오티드 결합 단백질, 폴리 히스티딘 꼬리 및 GST 태그를 포함하지만, 이에 제한되지 않는다. 가장 바람직한 선택적 결합 모이어티는 비오틴 및 선택가능한 폴리뉴클레오티드 서열이다. 비오틴은 아비딘으로 코팅된 표면에 특이적으로 결합한다. 선택가능한 폴리뉴클레오티드 서열은 상동성 서열로 코팅된 표면에 특이적으로 결합 (즉, 혼성화)한다. 대안적으로, 선택가능한 폴리뉴클레오티드 서열은 폴리뉴클레오티드 결합 단백질로 코팅된 표면에 특이적으로 결합한다.
헤어핀 어댑터 및/또는 선택가능한 결합 모이어티는 절단, 니킹(nicking), 분할 또는 가수분해될 수 있는 영역을 포함할 수 있다. 이같은 영역은 정제 및/또는 단리 후에 제1 및/또는 제2 폴리뉴클레오티드가 이들이 결합된 표면으로부터 제거될 수 있게 하도록 디자인될 수 있다. 적절한 영역이 관련 기술 분야에 공지되어 있다. 적절한 영역은 RNA 영역, 데스티오비오틴 및 스트렙타비딘을 포함하는 영역, 디술피드 결합, 및 광분할성(photocleavable) 영역을 포함하지만, 이에 제한되지 않는다.
바람직하게는 이중 가닥 표적 폴리뉴클레오티드는 가교 모이어티, 예컨대 헤어핀 루프 또는 헤어핀 루프 어댑터의 반대쪽 끝부분에 리더 서열을 포함한다. 하기에서 리더 서열이 더욱 상세하게 논의된다.
라운드-더-코너(Round the corner) 시퀀싱
바람직한 실시양태에서, 표적 이중 가닥 폴리뉴클레오티드에 가교 모이어티, 예컨대 헤어핀 루프 또는 헤어핀 루프 어댑터가 한쪽 끝부분에서 제공되고, 방법은 폴리뉴클레오티드의 양쪽 가닥이 포어를 통해 이동하도록 폴리뉴클레오티드를 포어와 접촉시키는 단계 및 폴리뉴클레오티드의 양쪽 가닥이 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 여기서 측정은 폴리뉴클레오티드의 가닥들의 하나 이상의 특성을 지시하며, 이에 의해 표적 이중 가닥 폴리뉴클레오티드를 특성화하는 단계를 포함한다. 또 다른 바람직한 실시양태에서, 표적 이중 가닥 폴리뉴클레오티드에 가교 모이어티, 예컨대 헤어핀 루프 또는 헤어핀 루프 어댑터가 한쪽 끝부분에서 제공되고, 방법은 폴리뉴클레오티드의 양쪽 가닥이 소화되어 개별적인 뉴클레오티드를 형성하도록 폴리뉴클레오티드를 포어 및 엑소뉴클레아제와 접촉시키는 단계를 포함한다. 상기 논의된 실시양태들 중 임의의 것이 이러한 실시양태에 동일하게 적용된다.
리더 서열
가닥 특성화/시퀀싱 방법에서의 접촉 단계 전에, 바람직하게는 방법은 포어 내로 우선적으로 관통되는 리더 서열을 폴리뉴클레오티드에 부착시키는 것을 포함한다. 리더 서열은 본 발명의 방법을 용이하게 한다. 리더 서열은 포어 내로 우선적으로 관통되고, 이에 의해 포어를 통한 폴리뉴클레오티드의 이동을 용이하게 하도록 디자인된다. 또한 리더 서열은 폴리뉴클레오티드를 상기 논의된 바와 같은 하나 이상의 앵커에 연결시키는데 사용될 수 있다.
리더 서열은 전형적으로 중합체를 포함한다. 이러한 중합체는 바람직하게는 음으로 하전된다. 이러한 중합체는 바람직하게는 폴리뉴클레오티드, 예컨대 DNA 또는 RNA, 변형된 폴리뉴클레오티드 (예컨대 무염기 DNA), PNA, LNA, 폴리에틸렌 글리콜 (PEG) 또는 폴리펩티드이다. 리더는 바람직하게는 폴리뉴클레오티드를 포함하고, 더욱 바람직하게는 단일 가닥 폴리뉴클레오티드를 포함한다. 리더 서열은 상기 논의된 폴리뉴클레오티드 중 임의의 것을 포함할 수 있다. 단일 가닥 리더 서열은 가장 바람직하게는 단일 가닥의 DNA, 예컨대 폴리 dT 절편을 포함한다. 리더 서열은 바람직하게는 하나 이상의 스페이서를 포함한다.
리더 서열은 임의의 길이일 수 있지만, 전형적으로 뉴클레오티드 10 내지 150개의 길이, 예컨대 뉴클레오티드 20 내지 150개의 길이이다. 전형적으로 리더의 길이는 방법에서 사용된 막횡단 포어에 좌우된다.
리더 서열은 바람직하게는 하기에서 정의되는 바와 같은 Y 어댑터의 일부분이다.
이중 커플링
본 발명의 방법은 이중 가닥 폴리뉴클레오티드의 이중 커플링을 수반할 수 있다. 바람직한 실시양태에서, 본 발명의 방법은 하기 단계를 포함한다:
(a) 이중 가닥 폴리뉴클레오티드에 Y 어댑터를 한쪽 끝부분에서, 가교 모이어티 어댑터, 예컨대 헤어핀 루프 어댑터를 다른쪽 끝부분에서 제공하고, 폴리뉴클레오티드를 막에 커플링시키기 위한 하나 이상의 제1 앵커를 Y 어댑터가 포함하고, 폴리뉴클레오티드를 막에 커플링시키기 위한 하나 이상의 제2 앵커를 가교 모이어티 어댑터가 포함하며, 막에 대한 가교 모이어티 어댑터의 커플링의 강도가 막에 대한 Y 어댑터의 커플링의 강도보다 더 큰 단계;
(b) 단계 (a)에서 제공된 폴리뉴클레오티드를 폴리뉴클레오티드가 막에 대해, 예컨대 막을 통해 이동하도록 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉시키는 단계; 및
(c) 폴리뉴클레오티드가 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 여기서 측정은 폴리뉴클레오티드의 하나 이상의 특성을 지시하며, 이에 의해 표적 폴리뉴클레오티드를 특성화하는 단계.
이러한 유형의 방법이 영국 출원 번호 1406147.7에 상세하게 논의되어 있다.
이중 가닥 폴리뉴클레오티드에 Y 어댑터가 한쪽 끝부분에서, 가교 모이어티 어댑터가 다른쪽 끝부분에서 제공된다. Y 어댑터 및/또는 가교 모이어티 어댑터는 전형적으로 폴리뉴클레오티드 어댑터이다. 이들은 상기 논의된 폴리뉴클레오티드 중 임의의 것으로부터 형성될 수 있다.
전형적으로 Y 어댑터는 (a) 이중 가닥 영역 및 (b) 다른쪽 끝부분의 단일 가닥 영역 또는 상보적이지 않은 영역을 포함한다. 단일 가닥 영역을 포함하면 Y 어댑터가 오버행이 있는 것으로 기술될 수 있다. Y 어댑터 내에 비-상보적 영역이 존재하는 것은 어댑터에게 이의 Y 형상을 제공하는데, 이중 가닥 부분과 달리 2개의 가닥이 전형적으로 서로 혼성화되지 않기 때문이다. Y 어댑터는 하나 이상의 제1 앵커를 포함한다. 앵커는 상기에서 더욱 상세하게 논의되어 있다.
바람직하게는 Y 어댑터는 포어 내로 우선적으로 통과되는 리더 서열을 포함한다. 이는 상기에서 논의되어 있다.
바람직하게는 가교 모이어티 어댑터는 상기 논의된 바와 같은 선택가능한 결합 모이어티를 포함한다. 가교 모이어티 어댑터 및/또는 선택가능한 결합 모이어티는 상기 논의된 바와 같은 절단, 니킹, 분할 또는 가수분해될 수 있는 영역을 포함할 수 있다.
하나 이상의 헬리카제 및 하나 이상의 분자 브레이크가 상기 논의된 바와 같이 사용되면, Y 어댑터는 바람직하게는 하나 이상의 헬리카제를 포함하고, 가교 모이어티 어댑터는 바람직하게는 하나 이상의 분자 브레이크를 포함한다.
관련 기술 분야에 공지된 임의의 방법을 사용하여 Y 어댑터 및/또는 가교 모이어티 어댑터가 폴리뉴클레오티드에 라이게이션될 수 있다. 어댑터 중 하나 또는 양쪽 모두가 리가제, 예컨대 T4 DNA 리가제, 이. 콜라이 DNA 리가제, Taq DNA 리가제, Tma DNA 리가제 및 9°N DNA 리가제를 사용하여 라이게이션될 수 있다. 대안적으로, 하기 논의된 본 발명의 방법을 사용하여 어댑터가 폴리뉴클레오티드에 부가될 수 있다.
바람직한 실시양태에서, 방법의 단계 a)는 Y 어댑터를 한쪽 끝부분에, 가교 모이어티 어댑터를 다른쪽 끝부분에 포함하도록 이중 가닥 폴리뉴클레오티드를 변형시키는 것을 포함한다. 임의의 변형 방식이 사용될 수 있다. 바람직하게는 방법은 본 발명에 따라 이중 가닥 폴리뉴클레오티드를 변형시키는 것을 포함한다. 이는 하기에서 더욱 상세하게 논의된다. 임의의 방식으로 변형 및 특성화 방법이 조합될 수 있다.
막에 대한 가교 모이어티 어댑터의 커플링 (또는 결합)의 강도가 막에 대한 Y 어댑터의 커플링 (또는 결합)의 강도보다 더 크다. 이는 임의의 방식으로 측정될 수 있다. 커플링 (또는 결합) 강도를 측정하는 적절한 방법이 영국 출원 번호 1406147.7의 실시예에 개시되어 있다.
가교 모이어티 어댑터의 커플링 (또는 결합)의 강도는 바람직하게는 Y 어댑터의 커플링 (또는 결합)의 강도의 적어도 1.5배, 예컨대 앵커 어댑터의 커플링 (또는 결합)의 강도의 적어도 2배, 적어도 3배, 적어도 4배, 적어도 5배 또는 적어도 10배이다. 막에 대한 가교 모이어티 어댑터의 친화력 상수 (Kd)은 바람직하게는 Y 어댑터의 친화력 상수의 적어도 1.5배, 예컨대 Y 어댑터의 커플링의 강도의 적어도 2배, 적어도 3배, 적어도 4배, 적어도 5배 또는 적어도 10배이다.
여러 방식으로 가교 모이어티 어댑터가 Y 어댑터보다 더 강하게 막에 커플링 (또는 결합)된다. 예를 들어, 가교 모이어티 어댑터가 Y 어댑터보다 더 많은 앵커를 포함할 수 있다. 예를 들어, 가교 모이어티 어댑터는 2개, 3개 또는 이를 초과하는 개수의 제2 앵커를 포함할 수 있는 반면, Y 어댑터는 1개의 제1 앵커를 포함할 수 있다.
막에 대한 하나 이상의 제2 앵커의 커플링 (또는 결합)의 강도가 막에 대한 하나 이상의 제1 앵커의 커플링 (또는 결합)의 강도보다 더 클 수 있다. 가교 모이어티 어댑터에 대한 하나 이상의 제2 앵커의 커플링 (또는 결합)의 강도가 Y 어댑터에 대한 하나 이상의 제1 앵커의 커플링 (또는 결합)의 강도보다 더 클 수 있다. 하나 이상의 제1 앵커 및 하나 이상의 제2 앵커가 이들의 각각의 어댑터에 혼성화를 통해 부착될 수 있고, 혼성화 강도가 하나 이상의 제1 앵커에서보다 하나 이상의 제2 앵커에서 더 크다. 이러한 실시양태들의 임의의 조합이 본 발명에서 또한 사용될 수 있다. 관련 기술 분야에 공지된 방법을 사용하여 커플링 (또는 결합) 강도를 측정할 수 있다.
바람직하게는 하나 이상의 제2 앵커는 막에 커플링 (또는 결합)되는 하나 이상의 제1 앵커 내의 하나 이상의 기보다 더 큰 강도로 막에 커플링 (또는 결합)되는 하나 이상의 기를 포함한다. 바람직한 실시양태에서, 가교 모이어티 어댑터/하나 이상의 제2 앵커는 콜레스테롤을 사용하여 막에 커플링 (또는 결합)되고, Y 어댑터/하나 이상의 제1 앵커는 팔미테이트를 사용하여 막에 커플링 (또는 결합)된다. 콜레스테롤은 팔미테이트보다 더 강하게 3블록 공중합체 막 및 지질 막에 결합한다. 대안적인 실시양태에서, 가교 모이어티 어댑터/하나 이상의 제2 앵커는 모노-아실 종, 예컨대 팔미테이트를 사용하여 막에 커플링 (또는 결합)되고, Y 어댑터/하나 이상의 제1 앵커는 디아실 종, 예컨대 디팔미토일포스파티딜콜린을 사용하여 막에 커플링 (또는 결합)된다.
헤어핀 루프 및 리더 서열의 부가
제공 전에, 이중 가닥 폴리뉴클레오티드가 MuA 트랜스포사제(transposase), 및 집단 내의 기질의 일부는 리더 서열을 포함하는 Y 어댑터이고 집단 내의 기질의 일부는 헤어핀 루프 어댑터인 이중 가닥 MuA 기질 집단과 접촉될 수 있다. 트랜스포사제는 이중 가닥 폴리뉴클레오티드 피분석물을 단편화하고, MuA 기질을 단편의 한쪽 또는 양쪽 끝부분에 라이게이션시킨다. 이는 리더 서열을 한쪽 끝에, 헤어핀 루프를 다른 쪽 끝에 포함하는 복수의 변형된 이중 가닥 폴리뉴클레오티드를 생산한다. 그 후, 변형된 이중 가닥 폴리뉴클레오티드를 본 발명의 방법을 사용하여 연구할 수 있다.
바람직하게는 집단 내의 각각의 기질이 적어도 하나의 유니버설 뉴클레오티드 오버행을 포함하여, 트랜스포사제가 주형 폴리뉴클레오티드를 단편화하고, 기질을 이중 가닥 단편의 한쪽 또는 양쪽 끝부분에 라이게이션시키고, 이에 의해 복수의 단편/기질 구축물을 생산하며, 방법은 오버행을 구축물 내의 단편에 라이게이션시키고, 이에 의해 복수의 변형된 이중 가닥 폴리뉴클레오티드를 생산하는 단계를 추가로 포함한다. 적절한 유니버설 뉴클레오티드가 상기에서 논의되어 있다. 오버행은 바람직하게는 5개의 뉴클레오티드의 길이이다.
대안적으로, 바람직하게는 집단 내의 각각의 기질이 (i) 적어도 하나의 오버행 및 (ii) 적어도 하나의 오버행과 동일한 가닥 내의, 주형 폴리뉴클레오티드 내에 존재하지 않는 뉴클레오시드를 포함하는 적어도 하나의 뉴클레오티드를 포함하여, 트랜스포사제가 주형 폴리뉴클레오티드를 단편화하고, 기질을 이중 가닥 단편의 한쪽 또는 양쪽 끝부분에 라이게이션시키고, 이에 의해 복수의 단편/기질 구축물을 생산하며, 방법은 (a) 이러한 적어도 하나의 뉴클레오티드를 선택적으로 제거하는 것에 의해 구축물로부터 오버행을 제거하고, 이에 의해, 단일 가닥 갭을 포함하는 복수의 이중 가닥 구축물을 생산하는 단계, 및 (b) 구축물 내의 단일 가닥 갭을 복구하고, 이에 의해 복수의 변형된 이중 가닥 폴리뉴클레오티드를 생산하는 단계를 추가로 포함한다. 전형적으로 폴리뉴클레오티드는 데옥시아데노신 (dA), 데옥시우리딘 (dU) 및/또는 티미딘 (dT), 데옥시구아노신 (dG) 및 데옥시시티딘 (dC) 뉴클레오시드를 포함한다. 폴리뉴클레오티드 내에 존재하지 않는 뉴클레오시드는 바람직하게는 무염기, 아데노신 (A), 우리딘 (U), 5-메틸우리딘 (m5U), 시티딘 (C) 또는 구아노신 (G)이거나 또는 요소, 5,6 디히드록시티민, 티민 글리콜, 5-히드록시-5 메틸히단토인, 우라실 글리콜, 6-히드록시-5, 6-디히드로티민, 메틸타르트로닐우레아, 7,8-디히드로-8-옥소구아닌 (8-옥소구아닌), 8-옥소아데닌, fapy-구아닌, 메틸-fapy-구아닌, fapy-아데닌, 아플라톡신 B1-fapy-구아닌, 5-히드록시-시토신, 5-히드록시-우라실, 3-메틸아데닌, 7-메틸구아닌, 1,N6-에테노아데닌, 히포크산틴, 5-히드록시우라실, 5-히드록시메틸우라실, 5-포르밀우라실 또는 시스-syn-시클로부탄 피리미딘 이량체를 포함한다. 적어도 하나의 뉴클레오티드는 바람직하게는 오버행으로부터 뉴클레오티드 10개 이하이다. 적어도 하나의 뉴클레오티드는 오버행 내의 제1 뉴클레오티드이다. 바람직하게는 오버행 내의 모든 뉴클레오티드가 주형 폴리뉴클레오티드 내에 존재하지 않는 뉴클레오시드를 포함한다.
이러한 MuA를 기초로 하는 방법이 국제 출원 번호 PCT/GB2014/052505에 개시되어 있다. 이는 또한 영국 출원 번호 1406147.7에 상세하게 논의되어 있다.
이중 가닥 폴리뉴클레오티드 및 MuA 트랜스포사제와 접촉되기 전에 MuA 기질 Y 어댑터에 하나 이상의 헬리카제가 부착될 수 있다. 대안적으로, 이중 가닥 폴리뉴클레오티드 및 MuA 트랜스포사제와 접촉되기 전에 MuA 기질 Y 어댑터에 하나 이상의 헬리카제가 부착될 수 있다.
이중 가닥 폴리뉴클레오티드 및 MuA 트랜스포사제와 접촉되기 전에 MuA 기질 헤어핀 루프에 하나 이상의 분자 브레이크가 부착될 수 있다. 대안적으로, 이중 가닥 폴리뉴클레오티드 및 MuA 트랜스포사제와 접촉되기 전에 MuA 기질 헤어핀 루프에 하나 이상의 분자 브레이크가 부착될 수 있다.
언커플링
본 발명의 방법은 다중 표적 폴리뉴클레오티드를 특성화하고 적어도 제1 표적 폴리뉴클레오티드를 언커플링시키는 것을 수반할 수 있다.
바람직한 실시양태에서, 본 발명은 2개 이상의 표적 폴리뉴클레오티드를 특성화하는 것을 수반한다. 방법은 하기 단계를 포함한다:
(a) 제1 샘플 내의 제1 폴리뉴클레오티드를 제공하는 단계;
(b) 제2 샘플 내의 제2 폴리뉴클레오티드를 제공하는 단계;
(c) 제1 샘플 내의 제1 폴리뉴클레오티드를 하나 이상의 앵커를 사용하여 막에 커플링시키는 단계;
(d) 폴리뉴클레오티드가 포어에 대해, 예컨대 포어를 통해 이동하도록 제1 폴리뉴클레오티드를 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉시키는 단계;
(e) 제1 폴리뉴클레오티드가 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 이러한 치수가 제1 폴리뉴클레오티드의 하나 이상의 특성을 지시하며, 이에 의해 제1 폴리뉴클레오티드를 특성화하는 단계;
(f) 제1 폴리뉴클레오티드를 막으로부터 언커플링시키는 단계;
(g) 제2 샘플 내의 제2 폴리뉴클레오티드를 하나 이상의 앵커를 사용하여 막에 커플링시키는 단계;
(h) 제2 폴리뉴클레오티드가 포어에 대해, 예컨대 포어를 통해 이동하도록 제2 폴리뉴클레오티드를 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 접촉시키는 단계; 및
(i) 제2 폴리뉴클레오티드가 포어에 대해 이동할 때 하나 이상의 측정을 수행하고, 이러한 치수가 제2 폴리뉴클레오티드의 하나 이상의 특성을 지시하며, 이에 의해 제2 폴리뉴클레오티드를 특성화하는 단계.
이러한 유형의 방법이 영국 출원 번호 1406155.0에 상세하게 논의되어 있다.
단계 (f) (즉, 제1 폴리뉴클레오티드의 언커플링)가 단계 (g) 전에 (즉, 제2 폴리뉴클레오티드를 막에 커플링시키기 전에) 수행될 수 있다. 단계 (g)가 단계 (f) 전에 수행될 수 있다. 제1 폴리뉴클레오티드가 언커플링되기 전에 제2 폴리뉴클레오티드가 막에 커플링되면, 바람직하게는 단계 (f)는 선택적으로 제1 폴리뉴클레오티드를 막으로부터 언커플링시키는 것 (즉, 제1 폴리뉴클레오티드는 언커플링시키지만 제2 폴리뉴클레오티드는 언커플링시키지 않음)을 포함한다. 통상의 기술자는 선택적 언커플링이 달성되는 시스템을 디자인할 수 있다. 단계 (f) 및 (g)가 동시에 수행될 수 있다. 이는 하기에서 더욱 상세하게 논의된다.
단계 (f)에서, 바람직하게는 제1 폴리뉴클레오티드의 적어도 10%가 막으로부터 언커플링된다. 예를 들어, 제1 폴리뉴클레오티드의 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60%, 적어도 70%, 적어도 80%, 적어도 90% 또는 적어도 95%가 막으로부터 언커플링된다. 바람직하게는, 모든 제1 폴리뉴클레오티드가 막으로부터 언커플링된다. 막으로부터 언커플링되는 제1 폴리뉴클레오티드의 양을 포어를 사용하여 결정할 수 있다. 이는 실시예에서 개시된다.
제1 폴리뉴클레오티드 및 제2 폴리뉴클레오티드는 서로 상이할 수 있다. 대안적으로, 제1 및 제2 폴리뉴클레오티드는 상이한 폴리뉴클레오티드일 수 있다. 이같은 경우에, 제2 폴리뉴클레오티드를 첨가하기 전에 제1 샘플의 적어도 일부분을 제거하는 것이 필요하지 않을 수 있다. 이는 하기에서 더욱 상세하게 논의된다. 방법이 3개 이상의 폴리뉴클레오티드를 연구하는 것에 관련되면, 이들 모두가 서로 상이할 수 있거나 또는 이들 중 일부가 서로 상이할 수 있다.
제1 폴리뉴클레오티드 및 제2 폴리뉴클레오티드는 동일한 폴리뉴클레오티드의 2개의 예일 수 있다. 제1 폴리뉴클레오티드는 제2 폴리뉴클레오티드와 동일할 수 있다. 이는 교정을 허용한다. 방법이 3개 이상의 폴리뉴클레오티드를 연구하는 것에 관련되면, 이들 모두가 동일한 폴리뉴클레오티드의 3개 이상의 예일 수 있거나, 또는 이들 중 일부가 동일한 폴리뉴클레오티드의 별개의 예일 수 있다.
제1 샘플 및 제2 샘플이 서로 상이할 수 있다. 예를 들어, 제1 샘플은 인간으로부터 유래될 수 있고, 제2 샘플은 바이러스로부터 유래될 수 있다. 제1 및 제2 샘플이 서로 상이한 경우, 이들은 동일한 제1 및 제2 폴리뉴클레오티드를 함유할 수 있거나 또는 이를 함유하는 것으로 추정될 수 있다. 방법이 3개 이상의 샘플을 연구하는 것에 관련되면, 이들 모두가 서로 상이할 수 있거나 또는 이들 중 일부가 서로 상이할 수 있다.
바람직하게는 제1 샘플 및 제2 샘플은 동일한 샘플의 2개의 예이다. 바람직하게는 제1 샘플은 제2 샘플과 동일하다. 이는 교정을 허용한다. 방법이 3개 이상의 샘플을 연구하는 것에 관련되면, 이들 모두가 동일한 샘플의 3개 이상의 예일 수 있거나, 또는 이들 중 일부가 동일한 샘플의 별개의 예일 수 있다.
임의 개수의 폴리뉴클레오티드를 연구할 수 있다. 예를 들어, 본 발명의 방법은 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 50, 100개 또는 이를 초과하는 개수의 폴리뉴클레오티드를 특성화하는 것에 관련될 수 있다. 3개 이상의 폴리뉴클레오티드가 본 발명의 방법을 사용하여 연구되는 경우, 제2 폴리뉴클레오티드 또한 막으로부터 언커플링되고, 필수 개수의 단계가 제3 폴리뉴클레오티드에 부가된다. 이는 4개 이상의 폴리뉴클레오티드에 대해서도 마찬가지이다.
본 발명의 방법은 제1 폴리뉴클레오티드를 막으로부터 언커플링시키는 것을 수반한다. 3개 이상의 폴리뉴클레오티드가 연구되는 경우 본 발명의 방법은 제2 폴리뉴클레오티드를 막으로부터 언커플링시키는 것을 수반할 수 있다.
임의의 공지된 방법을 사용하여 제1 폴리뉴클레오티드가 막으로부터 언커플링될 수 있다. 바람직하게는 제1 폴리뉴클레오티드는 단계 (f)에서 막 횡단 포어를 사용하여 막으로부터 언커플링되지 않는다. 바람직하게는 제1 폴리뉴클레오티드는 전압 또는 인가된 전위를 사용하여 막으로부터 언커플링되지 않는다.
바람직하게는 단계 (f)는 하나 이상의 앵커를 막으로부터 제거함으로써 제1 폴리뉴클레오티드를 막으로부터 언커플링시키는 것을 포함한다. 이러한 앵커가 제거되면, 제2 폴리뉴클레오티드가 다른 (또는 별도의) 앵커를 사용하여 막에 커플링된다. 제2 폴리뉴클레오티드를 커플링시키는데 사용되는 앵커는 제1 폴리뉴클레오티드를 커플링시키는데 사용된 것과 동일한 유형의 앵커 또는 상이한 유형의 앵커일 수 있다.
더욱 바람직하게는 단계 (f)는 하나 이상의 앵커를 하나 이상의 앵커에 대한 친화력이 이러한 앵커가 막에 대해 지니는 친화력보다 더 높은 작용제와 접촉시키는 것을 포함한다. 분자의 특이적 결합 능력을 결정하기 위한 경쟁적 결합 또는 면역방사측정 검정법에 대한 다양한 프로토콜이 관련 기술 분야에 널리 공지되어 있다 (예를 들어 문헌 [Maddox et al., J. Exp. Med. 158, 1211-1226, 1993]을 참조한다). 이러한 작용제는 앵커(들)를 막으로부터 제거하고, 이에 의해 제1 폴리뉴클레오티드를 언커플링시킨다. 바람직하게는 작용제는 당이다. 하나 이상의 앵커가 막에 대해 지니는 것보다 더 높은 친화력으로 하나 이상의 앵커에 결합하는 임의의 당이 사용될 수 있다. 당은 하기에 논의된 바와 같은 시클로덱스트린 또는 이의 유도체일 수 있다.
하나 이상의 앵커가 소수성 앵커, 예컨대 콜레스테롤를 포함하면, 작용제는 바람직하게는 시클로덱스트린 또는 이의 유도체 또는 지질이다. 시클로덱스트린 또는 이의 유도체는 문헌 [Eliseev, A. V., and Schneider, H-J. (1994) J. Am. Chem. Soc. 116, 6081-6088]에 개시된 것들 중 임의의 것일 수 있다. 작용제는 더욱 바람직하게는 헵타키스-6-아미노-β-시클로덱스트린 (am7-βCD), 6-모노데옥시-6-모노아미노-β-시클로덱스트린 (am1-βCD) 또는 헵타키스-(6-데옥시-6-구아니디노)-시클로덱스트린 (gu7-βCD)이다. 본원에 개시된 지질 중 임의의 것이 사용될 수 있다.
앵커가 스트렙타비딘, 비오틴 또는 데스티오비오틴을 포함하면, 작용제는 바람직하게는 비오틴, 데스티오비오틴 또는 스트렙타비딘이다. 비오틴 및 데스티오비오틴 양쪽 모두 스트렙타비딘이 막에 결합하는 것보다 더 높은 친화력으로 스트렙타비딘에 결합하고, 반대도 마찬가지이다. 비오틴은 데스티오비오틴보다 스트렙타비딘에 대한 친화력이 더 강하다. 따라서, 스트렙타비딘을 포함하는 앵커가 비오틴 또는 스트렙타비딘을 사용하여 막으로부터 제거될 수 있고, 반대도 마찬가지이다.
앵커가 단백질을 포함하면, 바람직하게는 작용제는 이러한 단백질에 특이적으로 결합하는 항체 또는 이의 단편이다. 우선적이거나 높은 친화력으로 단백질에 결합하지만 기타 또는 상이한 단백질에는 결합하지 않거나 또는 낮은 친화력으로만 결합하면, 항체가 이러한 단백질에 대해 특이적으로 결합하는 것이다. 1 × 10-6 M 이하, 더욱 바람직하게는 1 × 10-7 M 이하, 5 × 10-8 M 이하, 더욱 바람직하게는 1 × 10-8 M 이하, 더욱 바람직하게는 5 × 10-9 M 이하의 Kd로 결합하면 항체가 우선적이거나 높은 친화력으로 결합하는 것이다. 1 × 10-6 M 이상, 더욱 바람직하게는 1 × 10-5 M 이상, 더욱 바람직하게는 1 × 10-4 M 이상, 더욱 바람직하게는 1 × 10-3 M 이상, 더욱 더 바람직하게는 1 × 10-2 M 이상의 Kd로 결합하면 항체가 낮은 친화력으로 결합하는 것이다. 임의의 방법을 사용하여 직접적인 결합 또는 특이적인 결합을 검출할 수 있다. 항체가 단백질에 결합하는 것을 정량적으로 측정하는 방법이 관련 기술 분야에 널리 공지되어 있다. 항체는 모노클로날 항체 또는 폴리클로날 항체일 수 있다. 적절한 항체 단편은 Fv, F(ab') 및 F(ab')2 단편, 뿐만 아니라 단일쇄 항체를 포함하지만, 이에 제한되지 않는다. 또한, 항체 또는 이의 단편은 키메라 항체 또는 이의 단편, CDR-그래프트(grafted) 항체 또는 이의 단편 또는 인간화 항체 또는 이의 단편일 수 있다.
바람직하게는 단계 (f)는 하나 이상의 앵커를 하나 이상의 앵커가 막에 커플링되는 능력을 감소시키는 작용제와 접촉시키는 것을 포함한다. 예를 들어, 이러한 작용제는 하나 이상의 앵커의 구조 및/또는 소수성을 방해하고, 이에 의해 막에 커플링되는 앵커의 능력을 감소시킬 수 있다. 앵커가 콜레스테롤을 포함하면, 작용제는 바람직하게는 콜레스테롤 데히드로게나제이다. 앵커가 지질을 포함하면, 작용제는 바람직하게는 포스포리파제이다. 앵커가 단백질을 포함하면, 작용제는 바람직하게는 프로테이나제 또는 요소이다. 적절한 앵커 및 작용제의 기타 조합이 관련 기술 분야의 기술자에게 명백할 것이다.
바람직하게는 단계 (f)는 제1 폴리뉴클레오티드를 하나 이상의 앵커로부터 분리시킴으로써 제1 폴리뉴클레오티드를 막으로부터 언커플링시키는 것을 포함한다. 이는 임의의 방식으로 행해질 수 있다. 예를 들어, 링커를 포함하는 앵커에서 링커가 절단될 수 있다. 이러한 실시양태는 혼성화를 통한 연결을 수반하는 앵커에 대해 특히 적용가능하다. 이같은 앵커가 상기에서 논의되어 있다.
더욱 바람직하게는 단계 (f)는 제1 폴리뉴클레오티드 및 하나 이상의 앵커를 하나 이상의 앵커에 결합하는 것에 대해 제1 폴리뉴클레오티드와 경쟁하는 작용제와 접촉시킴으로써 제1 폴리뉴클레오티드를 막으로부터 언커플링시키는 것을 포함한다. 경쟁적 결합을 결정 및 측정하는 방법이 관련 기술 분야에 공지되어 있다. 바람직하게는 이러한 작용제는 하나 이상의 앵커에 혼성화하는 것에 대해 제1 폴리뉴클레오티드와 경쟁하는 폴리뉴클레오티드이다. 예를 들어, 제1 폴리뉴클레오티드가 혼성화를 수반하는 하나 이상의 앵커를 사용하여 막에 커플링되면, 하나 이상의 앵커를 혼성화 부위에서 또한 혼성화되는 폴리뉴클레오티드와 접촉시킴으로써 폴리뉴클레오티드가 언커플링될 수 있다. 전형적으로 폴리뉴클레오티드 작용제는 제1 폴리뉴클레오티드 및 하나 이상의 앵커의 농도보다 더 높은 농도로 첨가된다. 대안적으로, 폴리뉴클레오티드 작용제는 제1 폴리뉴클레오티드보다 더 강하게 하나 이상의 앵커에 혼성화될 수 있다.
더욱 바람직하게는 단계 (f)는 (i) 제1 폴리뉴클레오티드 및 하나 이상의 앵커를 요소, 트리스(2-카르복시에틸)포스핀 (TCEP), 디티오트레이톨 (DTT), 스트렙타비딘 또는 비오틴, UV 광, 효소 또는 결합제와 접촉시키거나; (ii) 제1 폴리뉴클레오티드 및 하나 이상의 앵커를 가열하거나; 또는 (iii) pH를 변경시키는 것을 포함한다. 요소, 트리스(2-카르복시에틸)포스핀 (TCEP) 또는 디티오트레이톨 (DTT)은 앵커를 파괴하고 제1 폴리뉴클레오티드를 막으로부터 분리시킬 수 있다. 앵커가 스트렙타비딘-비오틴 연결을 포함하면, 스트렙타비딘 작용제가 비오틴에 결합하는 것에 대해 경쟁할 것이다. 앵커가 스트렙타비딘-데스티오비오틴 연결을 포함하면, 비오틴 작용제가 스트렙타비딘에 결합하는 것에 대해 경쟁할 것이다. UV 광은 광-불안정 기를 파괴하는데 사용될 수 있다. 효소 및 결합제는 앵커를 절단하거나, 파괴하거나 또는 푸는데 사용될 수 있다. 바람직한 효소는 엑소뉴클레아제, 엔도뉴클레아제 또는 헬리카제를 포함하지만, 이에 제한되지 않는다. 바람직한 결합제는 효소, 항체 또는 이의 단편, 또는 단일-가닥 결합 단백질 (SSB)을 포함하지만, 이에 제한되지 않는다. 하기 논의된 효소 또는 상기 논의된 항체 중 임의의 것이 사용될 수 있다. 열 및 pH 또한 혼성화 및 기타 연결을 파괴하는데 사용될 수 있다.
제1 폴리뉴클레오티드를 하나 이상의 앵커로부터 분리시킴으로써 제1 폴리뉴클레오티드가 막으로부터 언커플링되면, 하나 이상의 앵커는 막 내에 남을 것이다. 바람직하게는 단계 (g)는 제1 폴리뉴클레오티드로부터 분리된 하나 이상의 앵커를 사용하여 제2 폴리뉴클레오티드를 막에 커플링시키는 것을 포함한다. 예를 들어, 막 내에 남은 하나 이상의 앵커에 혼성화되는 하나 이상의 폴리뉴클레오티드가 제2 폴리뉴클레오티드에 제공될 수 있다. 대안적으로, 바람직하게는 단계 (g)는 제1 폴리뉴클레오티드로부터 분리된 것과 별개의 하나 이상의 앵커 (즉, 하나 이상의 다른 앵커)를 사용하여 제2 폴리뉴클레오티드를 막에 커플링시키는 것을 포함한다. 하나 이상의 별개의 앵커는 제1 폴리뉴클레오티드를 막에 커플링시키는데 사용된 앵커와 동일한 유형일 수 있거나 또는 상이한 유형의 앵커일 수 있다. 바람직하게는 단계 (g)는 제1 폴리뉴클레오티드로부터 분리된 하나 이상의 앵커와 상이한 하나 이상의 앵커를 사용하여 제2 폴리뉴클레오티드를 막에 커플링시키는 것을 포함한다.
바람직한 실시양태에서, 단계 (f) 및 (g)는 제2 폴리뉴클레오티드가 하나 이상의 앵커에 결합하는 것에 대해 제1 폴리뉴클레오티드와 경쟁하고 제1 폴리뉴클레오티드를 교체하도록 막을 제2 폴리뉴클레오티드와 접촉시킴으로써 제1 폴리뉴클레오티드를 막으로부터 언커플링시키는 것을 포함한다. 예를 들어, 제1 폴리뉴클레오티드가 혼성화를 수반하는 하나 이상의 앵커를 사용하여 막에 커플링되면, 하나 이상의 앵커 내의 혼성화 부위에 또한 혼성화되는 폴리뉴클레오티드에 부착된 제2 폴리뉴클레오티드와 앵커를 접촉시킴으로써 제1 폴리뉴클레오티드가 언커플링될 수 있다. 전형적으로 제2 폴리뉴클레오티드는 제1 폴리뉴클레오티드 및 하나 이상의 앵커의 농도보다 더 높은 농도로 첨가된다. 대안적으로, 제2 폴리뉴클레오티드는 제1 폴리뉴클레오티드보다 더 강하게 하나 이상의 앵커에 혼성화될 수 있다.
제거 또는 세정
제1 폴리뉴클레오티드가 단계 (f)에서 막으로부터 언커플링되지만, 이는 필수적으로 제거 또는 세정되지 않는다. 제2 폴리뉴클레오티드가 제1 폴리뉴클레오티드로부터 쉽게 구별될 수 있으면, 제1 폴리뉴클레오티드를 제거하는 것이 필요하지 않다.
단계 (f)와 (g) 사이에, 방법은 제1 샘플의 적어도 일부를 막으로부터 제거하는 것을 추가로 포함한다. 제1 샘플의 적어도 10%가 제거될 수 있고, 예컨대 제1 샘플의 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60%, 적어도 70%, 적어도 80% 또는 적어도 90%가 제거될 수 있다.
더욱 바람직하게는 방법은 모든 제1 샘플을 막으로부터 제거하는 것을 추가로 포함한다. 이는 임의의 방식으로 행해질 수 있다. 예를 들어, 제1 폴리뉴클레오티드가 언커플링된 후, 막을 완충제로 세정할 수 있다. 적절한 완충제가 하기에서 논의된다.
변형된 폴리뉴클레오티드
특성화 전에, 폴리머라제가 표적 폴리뉴클레오티드를 주형으로 사용하여 변형된 폴리뉴클레오티드를 형성시키는 조건 하에 폴리뉴클레오티드를 폴리머라제 및 유리 뉴클레오티드 집단과 접촉시킴으로써 표적 폴리뉴클레오티드가 변형될 수 있고, 여기서 폴리머라제는 변형된 폴리뉴클레오티드를 형성시킬 때 표적 폴리뉴클레오티드 내의 뉴클레오티드 종 중 하나 이상을 상이한 뉴클레오티드 종으로 교체한다. 그 후, 변형된 폴리뉴클레오티드에 폴리뉴클레오티드에 부착되는 하나 이상의 헬리카제 또는 폴리뉴클레오티드에 부착되는 하나 이상의 분자 브레이크가 제공될 수 있다. 이러한 유형의 변형이 영국 출원 번호 1403096.9에 기술되어 있다. 상기에서 논의된 폴리머라제 중 임의의 것이 사용될 수 있다. 폴리머라제는 바람직하게는 클레나우 또는 9°노스(North)이다.
폴리머라제가 주형 폴리뉴클레오티드를 주형으로 사용하여 변형된 폴리뉴클레오티드를 형성시키는 조건 하에 주형 폴리뉴클레오티드가 폴리머라제와 접촉된다. 이같은 조건은 관련 기술 분야에 공지되어 있다. 예를 들어, 전형적으로 폴리뉴클레오티드가 시판되는 폴리머라제 완충제, 예컨대 뉴 잉글랜드 바이오랩스(New England Biolabs)®로부터의 완충제에서 폴리머라제와 접촉된다. 바람직하게는 온도는 클레나우에 대해서는 20 내지 37℃, 9°노스에 대해서는 60 내지 75℃이다. 전형적으로 프라이머 또는 3' 헤어핀이 폴리머라제 확장에 대한 핵화(nucleation) 지점으로서 사용된다.
막횡단 포어를 사용하는 폴리뉴클레오티드의 특성화, 예컨대 시퀀싱은 k개 (k는 양의 정수이다)의 뉴클레오티드로 구성된 중합체 단위 (즉, 'k량체')를 분석하는 것을 전형적으로 수반한다. 이는 국제 출원 번호 PCT/GB2012/052343 (WO 2013/041878로 공개됨)에 논의되어 있다. 상이한 k량체들에 대한 전류 측정치 사이에 명확한 분리가 있는 것이 바람직하지만, 이러한 측정치 중 일부가 중첩되는 것이 통상적이다. 특히, k량체 내의 중합체 단위의 개수가 높으면, 즉 k 값이 높으면, 폴리뉴클레오티드에 대한 정보, 예를 들어, 폴리뉴클레오티드의 기저 서열의 추정치를 유도하는 것에 해롭게, 상이한 k량체들에 의해 생성된 측정치를 해상하는 것이 어려워질 수 있다.
표적 폴리뉴클레오티드 내의 하나 이상의 뉴클레오티드 종을 변형된 폴리뉴클레오티드 내의 상이한 뉴클레오티드 종으로 교체함으로써, 변형된 폴리뉴클레오티드는 표적 폴리뉴클레오티드 내의 것과 상이한 k량체를 함유한다. 변형된 폴리뉴클레오티드 내의 상이한 k량체는 표적 폴리뉴클레오티드 내의 k량체와 상이한 전류 측정치를 생성시킬 수 있고, 따라서 변형된 폴리뉴클레오티드는 표적 폴리뉴클레오티드와 상이한 정보를 제공한다. 변형된 폴리뉴클레오티드로부터의 추가적인 정보가 표적 폴리뉴클레오티드를 특성화하는 것을 더 쉽게 만들 수 있다. 일부 경우에, 변형된 폴리뉴클레오티드 자체가 특성화하기 더 쉬울 수 있다. 예를 들어, 변형된 폴리뉴클레오티드가 전류 측정치 사이의 분리가 명확하거나 또는 분리가 증가된 k량체 또는 노이즈가 감소된 k량체를 포함하도록 디자인될 수 있다.
바람직하게는 폴리머라제는 변형된 폴리뉴클레오티드를 형성할 때 표적 폴리뉴클레오티드 내의 뉴클레오티드 종 중 2개 이상을 상이한 뉴클레오티드 종으로 교체한다. 폴리머라제는 표적 폴리뉴클레오티드 내의 2개 이상의 뉴클레오티드 종 각각을 별개의 뉴클레오티드 종으로 교체할 수 있다. 폴리머라제는 표적 폴리뉴클레오티드 내의 2개 이상의 뉴클레오티드 종 각각을 동일한 뉴클레오티드 종으로 교체할 수 있다.
표적 폴리뉴클레오티드가 DNA이면, 변형물 내의 상이한 뉴클레오티드 종은 전형적으로 아데닌, 구아닌, 티민, 시토신 또는 메틸시토신과 상이한 핵염기를 포함하고/하거나 데옥시아데노신, 데옥시구아노신, 티미딘, 데옥시시티딘 또는 데옥시메틸시티딘과 상이한 뉴클레오시드를 포함한다. 표적 폴리뉴클레오티드가 RNA이면, 변형된 폴리뉴클레오티드 내의 상이한 뉴클레오티드 종은 전형적으로 아데닌, 구아닌, 우라실, 시토신 또는 메틸시토신과 상이한 핵염기를 포함하고/하거나 아데노신, 구아노신, 우리딘, 시티딘 또는 메틸시티딘과 상이한 뉴클레오시드를 포함한다. 상이한 뉴클레오티드 종은 상기 논의된 유니버설 뉴클레오티드 중 임의의 것일 수 있다.
폴리머라제는 하나 이상의 뉴클레오티드 종을 하나 이상의 뉴클레오티드 종에 부재하는 화학 기 또는 원자를 포함하는 상이한 뉴클레오티드 종으로 교체할 수 있다. 이러한 화학 기는 프로피닐 기, 티오 기, 옥소 기, 메틸 기, 히드록시메틸 기, 포르밀 기, 카르복시 기, 카르보닐 기, 벤질 기, 프로파르길 기 또는 프로파르길아민 기일 수 있다.
폴리머라제는 하나 이상의 뉴클레오티드 종을 하나 이상의 뉴클레오티드 종에 존재하는 화학 기 또는 원자가 결여된 상이한 뉴클레오티드 종으로 교체할 수 있다. 폴리머라제는 뉴클레오티드 종 중 하나 이상을 전기음성도가 변경된 상이한 뉴클레오티드 종으로 교체할 수 있다. 전기음성도가 변경된 상이한 뉴클레오티드 종은 바람직하게는 할로겐 원자를 포함한다.
바람직하게는 방법은 변형된 폴리뉴클레오티드 내의 하나 이상의 상이한 뉴클레오티드 종으로부터 핵염기를 선택적으로 제거하는 것을 추가로 포함한다.
피분석물
전달
바람직하게는, 피분석물을 막으로 전달하는 미세입자에 표적 피분석물이 부착된다. 이러한 유형의 전달이 영국 출원 번호 1418469.1에 개시되어 있다. 임의 유형의 미세입자 및 부착 방법이 사용될 수 있다.
기타 특성화 방법
또 다른 실시양태에서, 폴리머라제에 의해 표적 폴리뉴클레오티드에 부가된 후 방출되는 표지된 종을 검출함으로서 폴리뉴클레오티드가 특성화된다. 폴리머라제가 폴리뉴클레오티드를 주형으로 사용한다. 각각의 표지된 종은 각각의 뉴클레오티드에 대해 특이적이다. 포스페이트로 표지된 종이 폴리머라제에 의해 폴리뉴클레오티드에 순차적으로 부가되도록 폴리뉴클레오티드가 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어, 폴리머라제 및 표지된 뉴클레오티드와 접촉되고, 포스페이트 종은 각각의 뉴클레오티드에 대해 특이적인 표지를 함유한다. 표지된 종을 이들이 뉴클레오티드로부터 방출되기 전에 (즉, 이들이 표적 폴리뉴클레오티드에 부가되어 있을 때) 또는 이들이 뉴클레오티드로부터 방출된 후에 포어를 사용하여 검출할 수 있다.
폴리머라제는 상기 논의된 것들 중 임의의 것일 수 있다. 포스페이트로 표지된 종이 포어를 사용하여 검출되고, 이에 의해 폴리뉴클레오티드를 특성화한다. 이러한 유형의 방법이 유럽 출원 번호 13187149.3 (EP 2682460으로 공개됨)에 개시되어 있다. 상기에 논의된 실시양태 중 임의의 것이 이러한 방법에 동일하게 적용된다.
표지된 종의 예는 중합체, 폴리에틸렌 글리콜, 당, 시클로덱스트린, 형광단, 약물, 대사산물, 펩티드를 포함하지만, 이에 제한되지 않는다. 이같은 태그의 비제한적인 예를 문헌 [Kumar et al. Sci Rep. 2012;2:684. Epub 2012 Sep 21]의 연구에서 확인할 수 있다.
센서 형성 방법
본 발명은 표적 폴리뉴클레오티드를 특성화하기 위한 센서의 형성 방법을 또한 제공한다. 이러한 방법은 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 폴리뉴클레오티드 결합 단백질, 예컨대 헬리카제 또는 엑소뉴클레아제 사이의 복합체를 형성시키는 것을 포함한다. 포어 및 단백질을 표적 폴리뉴클레오티드의 존재 하에 접촉시킨 후, 포어를 가로질러 전위를 인가함으로써 복합체가 형성될 수 있다. 인가된 전위는 상기 기술된 바와 같이 화학 전위 또는 전압 전위일 수 있다. 대안적으로, 포어를 단백질에 공유결합으로 부착시킴으로써 복합체가 형성될 수 있다. 공유결합 부착 방법이 관련 기술 분야에 공지되어 있고, 예를 들어, 국제 출원 번호 PCT/GB09/001679 (WO 2010/004265로 공개됨) 및 PCT/GB10/000133 (WO 2010/086603으로 공개됨)에 개시되어 있다. 이러한 복합체는 표적 폴리뉴클레오티드를 특성화하기 위한 센서이다. 바람직하게는 방법은 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 헬리카제 사이의 복합체를 형성시키는 것을 포함한다. 상기에 논의된 실시양태 중 임의의 것이 본 발명의 센서에 동일하게 적용된다.
본 발명은 표적 폴리뉴클레오티드를 특성화하기 위한 센서를 또한 제공한다. 이러한 센서는 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어와 폴리뉴클레오티드 결합 단백질 사이의 복합체를 포함한다. 상기에 논의된 실시양태 중 임의의 것이 본 발명의 센서에 동일하게 적용된다.
키트
본 발명은 표적 폴리뉴클레오티드를 특성화하기 위한 키트를 또한 제공한다. 이러한 키트는 CsgG 포어 또는 그의 돌연변이체, 예컨대 본 발명의 포어, 및 막의 성분들을 포함한다. 바람직하게는 이러한 성분들로부터 막이 형성된다. 바람직하게는 포어가 막 내에 존재한다. 키트는 상기 개시된 막 중 임의의 것, 예컨대 양친매성 층 또는 3블록 공중합체 막의 성분들을 포함할 수 있다.
키트는 폴리뉴클레오티드 결합 단백질을 추가로 포함할 수 있다. 상기 논의된 폴리뉴클레오티드 결합 단백질 중 임의의 것이 사용될 수 있다.
키트는 폴리뉴클레오티드를 막에 커플링시키기 위한 하나 이상의 앵커를 추가로 포함할 수 있다.
키트는 바람직하게는 이중 가닥 폴리뉴클레오티드를 특성화하기 위한 것이고, 바람직하게는 Y 어댑터 및 헤어핀 루프 어댑터를 포함한다. Y 어댑터는 바람직하게는 하나 이상의 헬리카제가 부착되어 있고, 헤어핀 루프 어댑터는 바람직하게는 하나 이상의 분자 브레이크가 부착되어 있다. Y 어댑터는 바람직하게는 폴리뉴클레오티드를 막에 커플링시키기 위한 하나 이상의 제1 앵커를 포함하고, 헤어핀 루프 어댑터는 바람직하게는 폴리뉴클레오티드를 막에 커플링시키기 위한 하나 이상의 제2 앵커를 포함하며, 바람직하게는 막에 대한 헤어핀 루프 어댑터의 커플링의 강도가 막에 대한 Y 어댑터의 커플링의 강도보다 더 크다.
본 발명의 키트는 상기 언급된 실시양태 중 임의의 것이 수행될 수 있게 하는 하나 이상의 시약 또는 기기를 추가적으로 포함할 수 있다. 이같은 시약 또는 기기는 하기 중 하나 이상을 포함한다: 적절한 완충제(들) (수성 용액), 대상체로부터 샘플을 수득하는 수단 (예컨대 도관, 또는 바늘을 포함하는 기기), 폴리뉴클레오티드를 증폭 및/또는 발현시키는 수단, 또는 전압 또는 패치 클램프 기구. 시약은 유체 샘플에 시약이 재현탁되도록 키트 내에 건조 상태로 존재할 수 있다. 또한, 키트가 본 발명의 방법에서 사용될 수 있게 하는 설명서 또는 어떤 생물에 대해 방법이 사용될 수 있는지에 관한 상세사항을 키트가 임의적으로 포함할 수 있다.
기구
본 발명은 표적 피분석물, 예컨대 표적 폴리뉴클레오티드를 특성화하기 위한 기구를 또한 제공한다. 이러한 기구는 복수의 CsgG 포어 또는 그의 돌연변이체 및 복수의 막을 포함한다. 바람직하게는 복수의 포어가 복수의 막 내에 존재한다. 바람직하게는 포어 및 막의 개수가 동일하다. 바람직하게는, 단일 포어가 각각의 막 내에 존재한다.
바람직하게는 기구는 본 발명의 방법을 수행하기 위한 설명서를 추가로 포함한다. 기구는 피분석물 분석을 위한 임의의 통상적인 기구, 예컨대 어레이 또는 칩일 수 있다. 본 발명의 방법과 관련하여 상기에 논의된 실시양태 중 임의의 것이 본 발명의 기구에 동일하게 적용가능하다. 기구는 본 발명의 키트 내에 존재하는 특색 중 임의의 것을 추가로 포함할 수 있다.
바람직하게는 기구는 본 발명의 방법을 수행하도록 설정된다.
바람직하게는 기구는 하기를 포함한다:
복수의 포어 및 막을 지지할 수 있고, 포어 및 막을 사용하여 피분석물 특성화를 수행하도록 작동가능할 수 있는 센서 장치; 및
특성화를 수행하기 위한 물질의 전달을 위한 적어도 1개의 포트(port).
대안적으로, 바람직하게는 기구는 하기를 포함한다:
복수의 포어 및 막을 지지할 수 있고, 포어 및 막을 사용하여 피분석물 특성화를 수행하도록 작동가능할 수 있는 센서 장치; 및
특성화를 수행하기 위한 물질을 보유하기 위한 적어도 1개의 저장소.
더욱 바람직하게는 기구는 하기를 포함한다:
막 및 복수의 포어 및 막을 지지할 수 있고, 포어 및 막을 사용하여 피분석물 특성화를 수행하도록 작동가능할 수 있는 센서 장치;
특성화를 수행하기 위한 물질을 보유하기 위한 적어도 1개의 저장소;
적어도 1개의 저장소로부터 센서 장치로 물질을 제어가능하게 공급하도록 구성된 유체 공학 시스템; 및
각각의 샘플을 받기 위한 하나 이상의 용기로서, 하나 이상의 용기로부터 센서 장치로 샘플을 선택적으로 공급하도록 상기 유체 공학 시스템이 구성되어 있는 용기.
이러한 기구는 국제 출원 번호 번호 PCT/GB08/004127 (WO 2009/077734로 공개됨), PCT/GB10/000789 (WO 2010/122293으로 공개됨), 국제 출원 번호 PCT/GB10/002206 (WO 2011/067559로 공개됨) 또는 국제 출원 번호 PCT/US99/25679 (WO 00/28312로 공개됨)에 기술된 것들 중 임의의 것일 수 있다.
하기 실시예는 본 발명을 설명한다.
실시예
실시예
1:
CsgG의
클로닝
및 계통
외막에 국소화되고 C-말단에 StrepII-태그가 부착된 CsgG (pPG1) 및 원형질막 주위공간의 C-말단에 StrepII-태그가 부착된 CsgGC1S (pPG2)의 생산을 위한 발현 구축물이 문헌 [Goyal, P. et al., Acta Crystallogr. F. Struct. Biol. Cryst. Commun. 2013, 69, 1349-1353]에 기술되어 있다. 셀레노메티오닌 표지화를 위해, StrepII-태그가 부착된 CsgGC1S가 수율 증가로 인해 세포질에서 발현되었다. 따라서, 프라이머 5'-TCT TTA AC CGC CCC GCC TAA AG-3' (정방향) (서열식별번호: 437) 및 5'-CAT TTT TTG CCC TCG TTA TC-3' (역방향) (서열식별번호: 438)으로의 역 PCR을 사용하여 N-말단 신호 펩티드를 제거하도록 pPG2가 변경되었다 (pPG3). 표현형 검정법을 위해, 이. 콜라이 BW25141의 csgG 결실 돌연변이체 (이. 콜라이 NVG2)를 문헌 [Datsenko, K. A. et al., Proc. Natl Acad. Sci. USA 97, 6640-6645 (2000)]에 기술된 방법에 의해 구축하였다 (프라이머 5'-AAT AAC TCA ACC GAT TTT TAA GCC CCA GCT TCA TAA GGA AAA TAA TCG TGT AGG CTG GAG CTG CTT C-3' (서열식별번호: 439) 및 5'-CGC TTA AAC AGT AAA ATG CCG GAT GAT AAT TCC GGC TTT TTT ATC TGC ATA TGA ATA TCC TCC TTA G-3' (서열식별번호: 440)를 사용함). Cys 접근성 검정법 및 채널 협착부의 표현형 프로빙(probing)에 사용된 다양한 CsgG 치환 돌연변이체가 trc 프로모터의 제어 하의 csgG를 함유하는 pTRC99a 벡터인 pMC2에서 출발하여 부위-지정 돌연변이유발 (퀵체인지(QuikChange) 프로토콜; 스트라타진(Stratagene))에 의해 구축되었다 (Robinson, L. S., et al., Mol. Microbiol., 2006, 59, 870-881).
실시예
2: 단백질 발현 및 정제
문헌 [Robinson, L. S., et al., Mol. Microbiol., 2006, 59, 870-881]에 기술된 바와 같이 CsgG 및 CsgGC1S가 발현 및 정제되었다. 간략하게, pPG1로 형질감염된 이. 콜라이 BL 21 (DE3)에서 CsgG를 재조합으로 생산하고, 단리된 외막으로부터 완충제 A (50 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM EDTA, 1 mM 디티오트레이톨 (DTT)) 내의 1% n-도데실-β-D-말토시드 (DDM)를 사용하여 추출하였다. StrepII-태그가 부착된 CsgG를 5 ml 스트렙-탁틴 세파로스 칼럼 (이바 게엠베하(Iba GmbH)) 상에 로딩하고, 0.5% 테트라에틸렌 글리콜 모노옥틸 에테르 (C8E4; 어피메트릭스(Affymetrix)) 및 4 mM 라우릴디메틸아민-N-옥시드 (LDAO; 어피메트릭스)가 보충된 20 칼럼 부피의 완충제 A로 세정하여 세제를 교환하였다. 2.5 mM D-데스티오비오틴을 첨가하여 단백질을 용출시키고, 결정화 실험을 위해 5 mg/ml로 농축하였다. 셀레노메티오닌 표지화를 위해, pPG3으로 형질전환되고 40 mg/ℓ의 L-셀레노메티오닌이 보충된 M9 최소 배지에서 성장된 Met 영양요구성 균주 B834 (DE3)에서 CsgGC1S를 생산하였다. 세포 펠릿을 컴플리트 프로테아제 인히비터 칵테일(cOmplete Protease Inhibitor Cocktail) (로슈(Roche))이 보충된 50 mM 트리스-HCl pH 8.0, 150 mM NaCl, 1 mM EDTA, 5 mM DTT에 재현탁시키고, 20 × 103 lb/in2으로 작동되는 TS 시리즈 세포 파괴기 (컨스탄트 시스템즈 엘티디(Constant Systems Ltd))에 통과시켜 파괴시켰다. 표지된 CsgGC1S를 문헌 [Robinson, L. S., et al., Mol. Microbiol., 2006, 59, 870-881]에 기술된 바와 같이 정제하였다. 셀레노메티오닌의 산화를 방지하도록 정제 과정 전반에 걸쳐 DTT (5 mM)가 첨가되었다.
pNH27를 보유하는 이. 콜라이 NEBC2566 세포에서 CsgE를 생산하였다 (Nenninger, A. A. et al., Mol. Microbiol. 2011, 81, 486-499). 25 mM 트리스-HCl pH 8.0, 150 mM NaCl, 25 mM 이미다졸, 5% (v/v) 글리세롤 내의 세포 용해물을 HisTrap FF (GE 헬스케어) 상에 로딩하였다. 20 mM 트리스-HCl pH 8.0, 150 mM NaCl, 5% (v/v) 글리세롤 완충제 내의 500 mM 이미다졸로의 선형 구배로 CsgE-his를 용출시켰다. CsgE를 함유하는 분획에 250 mM (NH4)2SO4를 보충하고, 20 mM 트리스-HCl pH 8.0, 100 mM NaCl, 250 mM (NH4)2SO4, 5% (v/v) 글리세롤로 평형화된 5 ml HiTrap 페닐 HP 칼럼 (GE 헬스케어)에 적용하였다. 20 mM 트리스-HCl pH 8.0, 10 mM NaCl, 5% (v/v) 글리세롤로의 선형 구배를 적용하여 용출시켰다. CsgE를 함유하는 분획을 20 mM 트리스-HCl pH 8.0, 100 mM NaCl, 5% (v/v) 글리세롤로 평형화된 수퍼로스 6 프렙 그레이드(Superose 6 Prep Grade) 10/600 (GE 헬스케어) 칼럼 상에 로딩하였다.
실시예
3: 용액 내에서의 올리고머 상태 평가
각각 약 0.5 mg의 세제-가용화 CsgG (0.5% C8E4, 4 mM LDAO) 및 CsgGC1S를 25 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM DTT, 4 mM LDAO 및 0.5% C8E4 (CsgG) 또는 25 mM 트리스-HCl pH 8.0, 200 mM NaCl (CsgGC1S)로 평형화된 수퍼덱스(Superdex) 200 10/300 GL 분석용 겔 여과 칼럼에 적용하고, 0.7 ml/분으로 러닝시켰다. 소 티로글로불린, 소 γ-글로불린, 닭 오브알부민, 말 미오글루불린 및 비타민 B12 (바이오-래드)로 칼럼 용출 부피를 보정하였다 (도 7). 막-추출 CsgG인 20 ㎍의 세제-가용화 단백질을 문헌 [Swamy, M., et al., Sci. STKE 2006, pl4, [http://dx.doi.org/10.1126/stke.3452006pl4(2006)]]에 기술된 절차를 사용하여 3-10% 청색 네이티브 PAGE 상에 또한 러닝시켰다 (도 7). 네이티브마크(NativeMark) (라이프 테크놀러지즈(Life Technologies)) 미염색 단백질 표준물 (7 ㎕)이 분자 질량 추정에 사용되었다. 성숙 CsgG는 C9 대칭성이 있는 별개의 9량체 포어 형성 입자, 뿐만 아니라 9량체 포어의 꼬리-대-꼬리 이량체 (즉, D9 대칭성이 있는 18량체)로서 주로 발견된다. 나노포어 센싱 용도의 목적을 위해, 선호되는 단백질 상태는 단일 9량체 포어이다. 나노포어 센싱 용도를 위한 지질 이중층 내로의 삽입 또는 크기 배제 크로마토그래피 전에 샘플을 가열함으로써 D9 18량체 포어에 대비하여 9량체 포어의 집단이 증가될 수 있다.
실시예
4: 결정화, 데이터 수집 및 구조 결정
셀레노메티오닌-표지화 CsgGC1S를 3.8 mg/ml로 농축하고, 100 mM 아세트산나트륨 pH 4.2, 8% PEG 4000 및 100 mM 말론산나트륨 pH 7.0을 함유하는 용액에 대한 시팅-드롭(sitting-drop) 증기 확산에 의해 결정화시켰다. 15% 글리세롤이 보충된 결정화 완충제에서 결정을 인큐베이션하고, 액체 질소에서 급속-냉동시켰다. 세제-가용화 CsgG를 5 mg/ml로 농축하고, 100 mM 트리스-HCl pH 8.0, 8% PEG 4000, 100 mM NaCl 및 500 mM MgCl2를 함유하는 용액에 대한 행잉-드롭(hanging-drop) 증기 확산에 의해 결정화시켰다. 결정을 액체 질소에서 급속-냉동시키고, 결정화 용액 내에 존재하는 세제에 의해 동결보호시켰다. 결정 상태의 최적화 및 회절 품질이 양호한 결정에 대한 스크리닝을 위해, 프록시마(Proxima)-1 및 프록시마-2a (솔레일(Soleil), 프랑스), PX-I (스위스 라이트 소스(Swiss Light Source), 스위스), I02, I03, I04 및 I24 (다이아몬드 라이트 소스(Diamond Light Source), 영국) 및 ID14eh2, ID23eh1 및 ID23eh2 (ESRF, 프랑스) 빔라인(beamline)에서 결정을 분석하였다. CsgGC1S 및 CsgG의 구조 결정에 사용된 최종 회절 데이터가 각각 I04 및 I03 빔라인에서 수집되었다 (표 5).
<표 5>
CsgGC1S에 대한 회절 데이터를 Xia2 및 XDS 패키지 (Winter, G., J. Appl. Cryst., 2010, 43, 186-190; Kabsch, W., Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125-132)를 사용하여 프로세싱하였다. CsgGC1S의 결정은 비대칭 단위 내의 16개의 단백질 카피를 함유하는, 단위 셀 치수가 a = 101.3 Å, b = 103.6 Å, c = 141.7 Å, α = 111.3°, β = 90.5°, γ = 118.2°인 공간 군 P1에 속했다. 구조 결정 및 정밀화를 위해, 0.9795 Å 파장에서 수집된 데이터를 최고 해상도 쉘에서의 Ι/ σΙ 차단값 2를 기초로 2.8 Å에서 절단하였다. 단일 비정상 분산(single anomalous dispersion) (SAD) 실험으로부터 계산된 실험적 위상을 사용하여 구조를 해명하였다. ShelxC 및 ShelxD (Sheldrick, G. M., Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 479-485)를 사용함으로써 총 92개의 셀레늄 부위가 비대칭 단위 내에 위치하였고, 정밀화되어, 샤프(Sharp) (Bricogne, G., Acta Crystallogr. D Biol. Crystallogr. 2003, 59, 2023-2030) (위상력(phasing power) 0.79, 성능 지수 (FOM) 0.25)로의 위상 계산에 사용되었다. 패럿(Parrot) (Cowtan, K., Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 470-478) (도 7; FOM 0.85)을 사용하여 비-결정학적 대칭(non-crystallographic symmetry) (NCS)에 의해 실험적 위상을 밀도를 변형시키고 평균화하였다. 버커니어(Buccaneer) (Cowtan, K., Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 1002-1011)로 초기 모델을 구축하고, 반복된 라운드의 휘닉스 리파인(Phenix refine) (Adams, P. D. et al. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213-221)으로의 최대 우도 정밀화 및 수동 검사 및 쿠트(Coot) (Emsley, P. et al., Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486-501)에서의 모델 (재)구축으로 정밀화하였다. 최종 구조는 1.34의 몰브로비티(molprobity) (Davis, I. W. et al., Nucleic Acids Res. 35 (Suppl 2), W375-W383) 점수로 16개의 CsgGC1S 쇄에 속하는 3,700개의 잔기 내의 28,853개의 원자를 함유하였다 (도 7); 잔기의 98%가 라마찬드란(Ramachandran) 플롯의 선호 영역 내에 놓였다 (99.7%가 허용 영역에 놓였다). 전자 밀도 지도는 가능한 용매 분자에 상응하는 명확한 밀도를 나타내지 않았고, 따라서 물 분자 또는 이온이 내부에 구축되지 않았다. 초기 및 후기 정밀화 단계에서 엄격한 NCS 구속(restraint) 및 국소적인 NCS 구속을 각각 사용하여, 16중 NCS 평균화가 정밀화 전반에 걸쳐 유지되었다.
CsgG에 대한 회절 데이터가 0.9763 Å 파장에서 단일 결정으로부터 수집되었고, XDS 패키지 (Winter, G., J. Appl. Cryst. 2010, 43, 186-190; Kabsch, W., Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125-132)를 사용하여, 비대칭 단위 내의 18개의 CsgG 카피 및 72% 용매 함량을 포함하는, 단위 셀 치수가 a = 161.7 Å, b = 372.3 Å, c = 161.8 Å 및 β = 92.9°인 공간 군 C2 내에 인덱싱 및 스케일링되었다. 구조 결정 및 정밀화를 위한 회절 데이터를 역 셀 방향 a*, b* 및 c*를 따라 3.6 Å, 3.7 Å 및 3.8 Å의 해상도 한계로 타원형으로 절단하고, 회절 비등방성 서버(Diffraction Anisotropy Server) (Strong, M. et al., Proc. Natl Acad. Sci. USA 2006, 103, 8060-8065)로 비등방성으로 스케일링하였다. CsgGC1S 단량체를 사용한 분자 교체가 성공적이지 않은 것으로 증명되었다. 자가 회전 함수의 분석에서 비대칭 단위 내의 D 9 대칭이 밝혀졌다 (제시되지 않음). CsgGC1S 구조를 기초로, C8 올리고머에서 C9 올리고머가 된 후, 프로토머간 각도가 45°에서 40°로 변할 때 입자 원주에서의 프로토머간 호는 대략적으로 동일하게 유지되어 약 4 Å의 계산된 반경 증가를 제공한다고 가정하여, 9량체 검색 모델이 생성되었다. 계산된 9량체를 검색 모델로 사용하여, 2개의 카피를 함유하는 분자 교체 해법이 페이저(Phaser) (McCoy, A. J. et al., J. Appl. Cryst. 2007, 40, 658-674)로 발견되었다. 밀도-변형 및 및 NCS-평균화 전자 밀도 지도 (패럿 [Cowtan, K., Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 470-478]; 도 8)의 검사는 TM1 및 TM2의 수동 구축 및 단백질 코어 내의 인접한 잔기의 리모델링, 뿐만 아니라 CsgGC1S 모델에서 상실되었고 α1 나선을 N-말단 지질 앵커에 연결한 잔기 2-18의 구축을 허용하였다. 각각 초기 및 최종 정밀화 라운드에 대한 부스터(Buster)-TNT (Smart, O. S. et al., Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 368-380) 및 Refmac5 (Murshudov, G. N. et al., Acta Crystallogr. D Biol. Crystallogr., 2011, 67, 355-367)로 CsgG 모델의 정밀화를 수행하였다. 정밀화 전반에 걸쳐 18중 국소 NCS 구속을 적용하였고, 프로스마트(Prosmart) (Nicholls, R. A., Long, F. & Murshudov, G. N., Acta Crystallogr. D Biol. Crystallogr. 2011, 68, 404-417)에 의해 생성된 수소-결합 구속 및 시그마 0.01의 젤리-바디(jelly-body) 정밀화를 사용하여 Refmac5를 시행하였다. 최종 구조는 2.79의 몰프로비티 점수로 18개의 CsgG 쇄에 속하는 4,451개의 잔기 내의 34,165개의 원자를 함유하였다 (도 7); 잔기의 93.0%가 라마찬드란 플롯의 선호 영역 내에 놓였다 (99.3%가 허용 영역 내에 놓였다). N-말단 지질 앵커에 상응하는 명확한 전자 밀도가 분간될 수 없었다.
실시예
5: 콩고
레드
검정법
콩고 레드 결합의 분석을 위해, 라이소제니 브로쓰(Lysogeny Broth) (LB)에서 37℃에서 성장된 철야 박테리아 배양물을 0.5의 D600에 도달할 때까지 LB 배지에 희석하였다. 5 ㎕ 샘플을 앰피실린 (100 mg/ℓ), 콩고 레드 (100 mg/ℓ) 및 0.1 % (w/v) 이소프로필 β-D-티오갈락토시드 (IPTG)가 보충된 LB 한천 플레이트 상에 스폿팅하였다. 플레이트를 실온 (20 내지 22℃)에서 48시간 동안 인큐베이션하여, 컬리 발현을 유도하였다. 48시에 콜로니 형태 및 염료 결합의 전개를 관찰하였다.
실시예
6: 시스테인 접근성 검정법
부위-지정 돌연변이유발을 사용하여 pMC2에서 시스테인 돌연변이체가 생성되었고, 이. 콜라이 LSR12에서 발현되었다 (Chapman, M. R. et al., Science, 2002, 295, 851-855). 철야로 성장된 박테리아 배양물을 1 mM IPTG 및 100 mg/ℓ 앰피실린을 함유하는 LB 한천 플레이트 상에 스폿팅하였다. 플레이트를 실온에서 인큐베이션하고, 48시간 후 세포를 긁어서 1 ml의 PBS에 재현탁시키고, D600을 사용하여 표준화하였다. 초음파처리로 세포를 용해시키고, 4℃에서 20초 동안 3,000 g로 원심분리하여, 피괴되지 않은 세포를 세포 용해물 및 현탁된 막으로부터 제거하였다. 상청액 내의 단백질을 실온에서 1시간 동안 15 mM 메톡시폴리에틸렌 글리콜-말레이미드 (MAL-PEG 5 kDa)로 표지하였다. 100 mM DTT로 반응을 정지시키고, 50.4 Ti 로터에서 4℃에서 20분 동안 40,000 r.p.m. (~100,000g)으로 원심분리하여 전체 막을 펠릿화시켰다. 펠릿을 1% 소듐 라우로일 사르코시네이트로 세정하여 세포질 막을 가용화시키고, 다시 원심분리하였다. 생성된 외막을 1% DDM을 함유하는 PBS를 사용하여 재현탁시키고 가용화하였다. 니켈 비드로의 금속-친화력 풀-다운(pull-down)을 SDS-PAGE 및 항-His 웨스턴 블롯에 사용하였다. 빈 pMC2 벡터가 있는 이. 콜라이 LSR12 세포를 음성 대조군으로서 사용하였다.
실시예
7: ATR-
FTIR
분광학
액체-질소-냉동 머큐리 카드뮴 텔루라이드 검출기 및 골든 게이트(Golden Gate) 반사율 부속품 (스펙액(Specac))이 장착된, 건식 공기로 연속적으로 퍼징되는 이퀴녹스(Equinox) 55 적외선 분광계 (브루커(Bruker)) 상에서 ATR-FTIR 측정을 수행하였다. 내부 반사 요소는 다이아몬드 결정 (2 mm × 2 mm)이었고, 빔 입사각은 45°였다. 각각의 정제된 단백질 샘플 (1 ㎕)을 결정 표면에 도포하고, 기체 질소 흐름 하에 건조시켜 필름을 형성시켰다. 2 cm-1 해상도에서 기록된 각각의 스펙트럼은 개선된 신호-대-노이즈 비를 위한 128개의 누적물의 평균이었다. 모든 스펙트럼을 수증기 기여 차감으로 처리하고, 애퍼디제이션(apodization)에 의해 4 cm-1의 최종 해상도에서 평활화(smoothing)하고, 아미드 I 밴드 (1,700-1,600 cm-1)의 면적에 대해 표준화하여 이들의 비교를 허용하였다 (Goormaghtigh, E.; Ruysschaert, J. M., Spectrochim. Acta, 1994, 50A, 2137-2144).
실시예
8: 음성 염색
EM
및 대칭 결정
음성 염색 EM을 사용하여 CsgG, CsgGC1S 및 CsgE의 용액 내에서의 올리고머화 상태를 모니터링하였다. CsgE, CsgGC1S 및 앰피폴(amphipol)-결합 CsgG를 0.05 mg/ml의 농도로 조정하고, 글로우-방전 탄소-코팅 구리 그리드(grid) (CF-400; 일렉트론 마이크로스코피 사이언시즈(Electron Microscopy Sciences))에 적용하였다. 1분 인큐베이션 후, 샘플을 블롯팅하고, 세정하고, 2% 우라닐 아세테이트에서 염색하였다. 120 kV의 전압, ×49,000의 배율 및 800 내지 2,000 nm의 디포커스(defocus)에서 작동되는 테크나이 T12 바이오트윈 LaB6(Tecnai T12 BioTWIN LaB6) 현미경에서 영상을 수집하였다. 콘트라스트 전달 함수(contrast transfer function) (CTF), 위상 플리핑(flipping) 및 입자 선택을 냉동-EM에 대해 기술된 바와 같이 수행하였다. 막-추출 CsgG의 경우, 18량체 입자 (총 1,780개)를 9량체 및 상면도와 별도로 분석하였다. 정제된 CsgE의 경우, 총 2,452개의 입자를 분석하였다. 3차원 모델을 하기의 CsgG-CsgE 냉동-EM 분석에 대해 기술된 바와 같이 수득하고, 여러 라운드의 다중-기준 정렬 (MRA), 다중-통계 분석 (MSA) 및 앵커 세트 정밀화에 의해 정밀화하였다. 모든 경우에, 표준화 및 중심화 후, 냉동-EM 섹션에 기술된 바와 같이 IMAGIC-4D를 사용하여 영상을 분류하였다. 특징적인 도에 상응하는 최상 클래스를 각각의 입자 세트에 대해 선택하였다. 클래스 당 약 20개의 영상으로 최상 클래스 평균을 사용하여 CsgG 상면도의 대칭 결정을 수행하였다. IMAGIC를 사용하여 회전식 자기상관 함수를 계산하고, 플롯팅하였다.
실시예
9: 음성 염색
EM
및 대칭 결정
음성 염색 EM을 사용하여 CsgG, CsgGC1S 및 CsgE의 용액 내에서의 올리고머화 상태를 모니터링하였다. CsgE, CsgGC1S 및 앰피폴-결합 CsgG를 0.05 mg/ml의 농도로 조정하고, 글로우-방전 탄소-코팅 구리 그리드 (CF-400; 일렉트론 마이크로스코피 사이언시즈)에 적용하였다. 1분 인큐베이션 후, 샘플을 블롯팅하고, 세정하고, 2% 우라닐 아세테이트에서 염색하였다. 120 kV의 전압, ×49,000의 배율 및 800 내지 2,000 nm의 디포커스에서 작동되는 테크나이 T12 바이오트윈 LaB6 현미경에서 영상을 수집하였다. 콘트라스트 전달 함수 (CTF), 위상 플리핑 및 입자 선택을 냉동-EM에 대해 기술된 바와 같이 수행하였다. 막-추출 CsgG의 경우, 18량체 입자 (총 1,780개)를 9량체 및 상면도와 별도로 분석하였다. 정제된 CsgE의 경우, 총 2,452개의 입자를 분석하였다. 3차원 모델을 하기의 CsgG-CsgE 냉동-EM 분석에 대해 기술된 바와 같이 수득하고, 여러 라운드의 다중-기준 정렬 (MRA), 다중-통계 분석 (MSA) 및 앵커 세트 정밀화에 의해 정밀화하였다. 모든 경우에, 표준화 및 중심화 후, 냉동-EM 섹션에 기술된 바와 같이 IMAGIC-4D를 사용하여 영상을 분류하였다. 특징적인 도에 상응하는 최상 클래스를 각각의 입자 세트에 대해 선택하였다. 클래스 당 약 20개의 영상으로 최상 클래스 평균을 사용하여 CsgG 상면도의 대칭 결정을 수행하였다. IMAGIC를 사용하여 회전식 자기상관 함수를 계산하고, 플롯팅하였다.
실시예
10:
CsgG
-
CsgE
복합체 형성
CsgG-CsgE 복합체 형성을 위해, 120 ml의 CsgG (0.5% C8E4, 4 mM LDAO, 25 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM DTT 내의 24 mg/ml 단백질)를 300 ml의 세제-탈안정화 리포솜 (1 mg/ml, 1,2-디미리스토일-sn-글리세로-3-포스포콜린 (DMPC) 및 0.4% LDAO)에 첨가하고, 얼음 상에서 5분 동안 인큐베이션한 후, 90 ml의 A8-35 앰피폴(Amphipol) (100 mg/ml 모액)을 첨가함으로써, 정제된 CsgG 내의 가용화 세제를 앰피폴 A8-35 (애너트레이스(Anatrace))로 교환하였다. 추가로 15분 동안 얼음 상에서 인큐베이션한 후, 샘플을 수퍼로스 6 10/300 GL (GE 헬스케어) 칼럼 상에 로딩하고, 200 mM NaCl, 2.5% 크실리톨, 25 mM 트리스-HCl pH 8, 0.2 mM DTT에서 겔 여과를 수행하였다. 200 mM NaCl, 2.5% 크실리톨, 25 mM 트리스-HCl pH 8, 0.2 mM DTT 내의 동일한 부피의 정제된 단량체 CsgE를 앰피폴-가용화 CsgG에 CsgE 및 CsgG에 대해 각각 15 및 5 mM의 최종 단백질 농도로 첨가하고, 샘플을 125 V로 18℃에서 0.5× TBE 완충제 내의 4.5% 네이티브 PAGE 상에 러닝시켰다. 제2의 변성 차원을 위해, CsgG-CsgE 복합체에 상응하는 밴드를 동일한 겔 상에서 병렬로 러닝된 미염색 레인으로부터 절단하고, 5분 동안 래믈리(Laemmli) 완충제 (60 mM 트리스-HCl pH 6.8, 2% SDS, 10% 글리세롤, 5% 2-메르캅토에탄올, 0.01 % 브로모페놀 블루)에서 비등시키고, 4-20% SDS-PAGE 상에 러닝시켰다. 정제된 CsgE 및 CsgG를 대조군 샘플로서 복합체와 나란히 러닝시켰다. 겔을 시각적 검사를 위해 인스턴트블루 쿠마시(InstantBlue Coomassie)로, 또는 SDS-PAGE 상에서의 CsgE 및 CsgG 밴드의 형광 검출 (타이푼(Typhoon) FLA 9000)에 의한 CsgG-CsgE 복합체의 화학량론 평가를 위해 사이프로 오렌지(SYPRO orange)로 염색하였고, 0.97의 CsgG/CsgE 비가 산출되었다.
실시예
11:
CsgG
-
CsgE
냉동-
EM
냉동-전자 현미경검사를 사용하여 C9 CsgG-CsgE 복합체의 용액 내에서의 구조를 결정하였다. 상기 기술된 바와 같이 제조된 CsgG-CsgE 복합체를 HisTrap FF (GE 헬스케어)에 결합시키고 이로부터 용출시켜, 결합되지 않은 CsgG를 제거하였고, 용출 시 복합체를 20 mA에서 30초 동안 글로우 방전된 퀀티포일(Quantifoil) R2/2 탄소 코팅 그리드 (퀀티포일 마이크로 툴스 게엠베하(Quantifoil Micro Tools GmbH))에 즉각적으로 적용하였다. 샘플을 자동화 시스템 (라이카(Leica))를 사용하여 액체 질소에서 플런지(plunge)-냉동시키고, 200 kV의 전압, 저용량 조건 하에서의 ×50,000의 공칭 배율, 및 1.4-3 mm의 디포커스 범위에서 작동되는 FEI F20 현미경 하에 관찰하였다. 팔콘(Falcon) II 검출기 상에서 영상 프레임을 기록하였다. 검체 수준에서의 픽셀 크기는 픽셀 당 1.9 Å이었다. CTFFIND3 (Mindell, J. A. & Grigorieff, N., J. Struct. Biol. 2003, 142, 334-347)을 사용하여 CTF 파라미터를 평가하였고, 스파이더(SPIDER) (Shaikh, T. R. et al., Nature Protocols, 2008, 3, 1941-1974)에서 위상 플리핑을 행하였다. 복서(BOXER) (EMAN2; Tang, G. et al., J. Struct. Biol., 2007, 157, 38-46)를 사용하여 CTF-수정 마이크로그래프로부터 입자가 자동적으로 선택되었다. 비점 수차가 10%를 초과하는 영상은 폐기되었다. 75개의 마이크로그래프로부터 총 1,221개의 입자가 선택되었고, 128-픽셀 × 128-픽셀 상자 내로 윈도우화되었다. 영상을 동일한 평균 및 표준 편차에 대해 표준화하고, ~200 Å의 저해상도 차단값으로 하이-패스(high-pass) 필터링하였다. 이들을 중심화한 후, 제1 라운드의 MSA에 적용하였다. IMAGIC-4D (이미지 사이언스 소프트웨어(Image Science Software))의 무기준(reference free) 분류를 사용하여 초기 기준 세트를 수득하였다. C9 원통형 입자의 특징적인 측면도에 상응하는 최상 클래스가 MRA에 대한 기준으로 사용되었다. CsgG-CsgE 복합체에 대해, 각도 재구성 (이미지 사이언스 소프트웨어)에 의해 배향이 결정된 복합체의 1,221개의 입자를 포괄하는 최상의 125개의 특징적인 도 (양호한 콘트라스트 및 윤곽이 뚜렷한 특색을 지님)로부터 최초의 3차원 모델이 계산되었다. 반복된 라운드의 MRA, MSA 및 앵커 세트 정밀화에 의해 3차원 지도를 정밀화하였다. 0.5 기준 수준에 따라 푸리에(Fourier) 쉘 상관관계 (FSC)에 의해 해상도가 24 Å인 것으로 추정되었다 (도 5).
지도 및 도면의 가시화를 UCSF 키메라(Chimera) (Pettersen, E. F. et al., J. Comput. Chem., 2004, 25, 1605-1612)에서 수행하였다.
실시예 12: CsgG 및 CsgG + CsgE 포어의 단일-채널 전류 분석
음의 장 전위 하에, CsgG 포어는 2가지 전도도 상태를 나타낸다. 도 5는 정상 전도도 형태 (+50, 0 및 -50 mV에서 측정됨) 및 저-전도도 형태 (0, +50 및 -50 mV에서 측정됨) 각각의 대표적인 단일-채널 전류 트레이스를 나타낸다. 전체 관찰 시간 동안 양쪽 상태 사이의 전환이 관찰되지 않았고 (n=22), 이는 전도도 상태가 긴 수명 (초 내지 분 시간 규모)을 갖는다는 것을 가리킨다. 왼쪽 아래의 패널은 +50 및 -50 mV에서 획득된 정상 및 저-전도도 형태의 CsgG 포어에 대한 전류 막대그래프를 나타낸다 (n=33). 정규 전도도 및 저-전도도의 CsgG 포어에 대한 I-V 곡선이 오른쪽 아래 패널에서 제시된다. 데이터는 4회 이상의 독립적인 기록으로부터의 평균 및 표준 편차를 나타낸다. 저-전도도 형태의 성질 또는 생리학적 존재는 미지이다.
도 6은 보조 인자 CsgE로 적정된 CsgG 채널의 전기생리학의 결과를 나타낸다. 이러한 플롯은 CsgE 농도의 함수로서 개방 채널, 중간체 채널 및 폐쇄 채널의 분율을 나타낸다. 개방 상태 및 폐쇄 상태의 CsgG가 도 6에서 도해된다 (각각 0 nM 및 100 nM CsgE). CsgE 농도를 10 nM 초과로 증가시키는 것은 CsgG 포어의 폐쇄에 이른다. 이러한 효과가 +50 mV (왼쪽) 및 -50 mV (오른쪽)에서 발생하여, 포어 차단이 CsgG 포어 내로의 CsgE (계산된 pI 4.7)의 전기영동에 의해 야기된다는 가능성을 배제시킨다. 드문 (<5%) 중간체 상태는 전도도가 개방 채널의 대략 절반이다. 이는 CsgE에 의해 유도된 불완전한 CsgG 채널 폐쇄를 나타낼 수 있다; 대안적으로, 이는 막에 매립된 포어에 전해질 용액으로부터의 잔여 CsgG 단량체가 결합하는 것에 야기된 CsgG 2량체의 일시적인 형성을 나타낼 수 있다. 단일-채널 전류 트레이스의 전체 지점 막대그래프 분석으로부터 3가지 상태에 대한 분율이 수득되었다. 막대그래프로 3개까지의 상태에 대한 피크 면적이 산출되었고, 소정의 상태에 대한 분율이 상응하는 피크 면적을 기록 내의 모든 다른 상태의 합계로 나눠서 수득되었다. 음의 장 전위 하에, CsgG에 대한 관찰 (a 참조)와 유사하게, 2개의 개방 전도도 상태가 분별된다. 양쪽 모두의 개방 채널 변동이 더 높은 CsgE 농도에 의해 차단되기 때문에, 도 6에서의 '개방' 트레이스 (0 nM)는 양쪽 전도도 형태를 조합한다. 플롯 내의 데이터는 3회의 독립적인 기록으로부터의 평균 및 표준 편차를 나타낸다.
결정 구조, 크기-배제 크로마토그래피 및 EM은 세제로 추출된 CsgG 포어가 더 높은 단백질 농도에서 비-천연의 꼬리-대-꼬리로 스태킹된 2량체 (예를 들어, 2개의 9량체 예컨대 D9 입자, 도 7)를 형성한다는 것을 나타낸다. 단일-채널 기록에서 이러한 이량체들이 또한 관찰될 수 있다. 위쪽의 패널은 +50, 0 및 -50 mV (왼쪽에서 오른쪽으로)에서의 스태킹된 CsgG 포어의 단일 채널 전류 트레이스를 나타낸다. 왼쪽 아래의 패널은 +50 및 -50 mV에서 기록된 2량체 CsgG 포어의 전류 막대그래프를 나타낸다. +50 및 -50 mV에서의 각각 +16.2±1.8 및 -16.0±3.0 pA (n=15)의 실험적 전도도는 23 pA의 이론적으로 계산된 값에 근접한다. 오른쪽 아래의 패널은 스태킹된 CsgG 포어에 대한 I-V 곡선을 나타낸다. 데이터는 6회의 독립적인 기록으로부터의 평균 및 표준 편차를 나타낸다.
스태킹된 CsgG 포어에 결합하고 이를 차단하는 CsgE의 능력을 전기생리학으로 테스트하였다. +50 mV (위쪽) 및 -50 mV (아래쪽)에서의 10 또는 100 nM CsgE의 존재 하에서의 스태킹된 CsgG 포어의 단일-채널 전류 트레이스가 제시된다. 이러한 전류 트레이스는 다른 경우에는 포화성인 농도의 CsgE가 스태킹된 CsgG 2량체에 대한 포어 폐쇄에 이르지 않는다는 것을 가리킨다. 이러한 관찰은 CsgG-CsgE 접촉 구역을 EM 및 부위-지정 돌연변이유발에 의해 분별된 바와 같이 나선 2 및 CsgG 원형질막 주위공간 공동의 입 부분에 맵핑하는 것 (도 5 및 6)과 잘 일치한다.
실시예
13:
담즙산
염 독성 검정법
담즙산 염을 함유하는 한천 플레이트 상에서의 성장 감소에 의해 외막 투과성을 연구하였다. pLR42 (Nenninger, A. A. et al., Mol. Microbiol., 2011, 81, 486-499) 및 pMC2 (Robinson, L. S. et al., Mol. Microbiol., 2006, 59, 870-881) 양쪽 모두 (또는 유래된 나선 2 돌연변이체)를 보유하는 이. 콜라이 LSR12 (Chapman, M. R. et al., Science, 2002, 295, 851-855) 세포의 10배 단계 희석물 (5 ml)을 0.2% (w/v) L-아라비노스와 함께 또는 L-아라비노스 없이 100 mg/ℓ 앰피실린, 25 mg/ℓ 클로람페니콜, 1 mM IPTG를 함유하는 맥콘키 한천 플레이트 상에 스폿팅하였다. 37℃에서 철야로 인큐베이션한 후, 콜로니 성장을 검사하였다.
실시예
14: 단일-채널 전류 기록
나니온(Nanion)으로부터의 오르비트(Orbit) 16 키트로의 병렬식 고해상도 전기 기록을 사용하여 단일-채널 전류 기록을 수행하였다. 간략하게, 1,2-디피타노일-sn-글리세로-3-포스포콜린 (아반티 폴라 리피즈(Avanti Polar Lipids))의 수평 이중층이 16-채널 다중전극 공동 어레이 (MECA) 칩 (아이오네라(Ionera))의 미세공동(microcavity) (피코리터 이하의 부피)에서 형성되었다 (del Rio Martinez, J. M., ACS NanoSmall, 2014, 5, 8080-8088). 이중층 위 및 아래의 시스 및 트랜스 공동 양쪽 모두 1.0 M KCl, 25 mM 트리스-HCl pH 8.0을 함유하였다. 채널을 막 내로 삽입하기 위해, 25 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM DTT, 0.5% C8E4, 5 mM LDAO에 용해된 CsgG를 90-300 nM의 최종 농도로 시스 구획에 첨가하였다. CsgG 채널과 CsgE의 상호작용을 테스트하기 위해, 25 mM 트리스-HCl pH 8.0, 150 mM NaCl에 용해된 후자 단백질의 용액을 0.1, 1, 10 및 100 nM의 최종 농도로 시스 구획에 첨가하였다. 3 kHz의 로우-패스(low-pass) 필터링 주파수 및 10 kHz의 샘플링 주파수로 테셀라 트리톤(Tecella Triton) 16-채널 증폭기를 사용하여 +50 mV 및 -50 mV (시스 측면 기준)의 유지 전위에서 막횡단 전류를 기록하였다. p클램프(pClamp) 스위트 (몰레큘러 디바이시즈)의 클램프피트(Clampfit)를 사용하여 전류 트레이스를 분석하였다. 오리진(Origin) 8.6 (마이크로칼(Microcal)) (Movileanu, L, Nature Biotechnol., 2000, 18, 1091-1095)을 사용하여 플롯이 생성되었다.
측정된 전류를 3개의 분절 (막횡단 섹션, 원형질막 주위공간 전정, 및 둘을 연결하는 내부 채널 협착부)로 구성되는 것으로 모델링된 CsgG X-선 구조의 포어 치수를 기초로 계산된 것과 비교하였다. 처음 2개의 분절은 원뿔형 형상인 것으로 모델링된 한편, 협착부는 원통형으로서 나타났다. 상응하는 저항 R 1, R 2 및 R 3 각각이 하기와 같이 계산되었다:
R 1 = L 1/(πD 1 d 1 κ)
R 2 = L 2/(πD 2 d 2 κ)
R 3 = L 3/(πd 1 d 2 κ)
[식 중, L 1, L 2 및 L 3은 각각 3.5, 4.0 및 2.0 nm로 측정된 분절의 축 길이이고, D 1, d 1, D 2 및 d 2는 각각 4.0, 0.8, 3.4 및 0.8 nm로 측정된 분절 1 및 2의 최대 및 최소 직경이다]. 전도도 κ의 값은 10.6 S/m이다. R 1, R 2 및 R 3 및 전압 V = 50 mV를 하기 식에 삽입함으로써 전류가 계산되었다:
I = V/(R 1 + R 2 + R 3)
접근(access) 저항이 예상 전류를 유의하게 변경시키는 것으로 발견되지 않았다.
단일 채널 전류 기록 예컨대 상기 기술된 것들을 피분석물의 존재 하에 작성함으로써, 채널이 바이오센서의 역할을 가정하게 허용할 수 있다.
실시예
15: 모델
폴리알라닌
쇄로의
CsgG
협착부의
분자 동역학 시뮬레이션
도 9에서 C-말단에서 N-말단 방향으로 제시된, 확장된 형상의 채널을 관통하는 폴리알라닌 쇄로 CsgG 협착부를 모델링하였다. CsgG 수송체를 통한 기질 통과는 자체적으로 서열 특이적이지 않다 (Nenninger, A. A. et al., Mol. Microbiol., 2011, 81, 486-499; Van Gerven, N. et al., Mol. Microbiol. 2014, 91, 1022-1035 2014). 명확하게 하기 위해, 통과하는 폴리펩티드 사슬의 추정 상호작용을 모델링하는데 폴리알라닌 쇄를 사용하였다. 모델링된 구역은 잔기 47-58을 각각 포함하는 9개의 동심성 CsgG C-루프로 구성된다. 협착부에 라이닝된 측쇄가 Asn 55 및 Phe 56이 표시된 막대 표현으로 제시된다. 협착부 내부의 폴리알라닌 잔기 (+1 내지 +5로 표지된 잔기)로부터 10 Å 이내의 용매 분자 (물)가 적색 점으로 제시된다. 도 9c는 도 9b에서와 같이 위치하고 명확성을 위해 C-루프가 제거된, 폴리알라닌 쇄의 모델링된 용매화를 나타낸다 (제시된 용매 분자는 전체 폴리알라닌 쇄로부터 10 Å 이내의 것이다). Asn 55 및 Phe 56의 고리의 높이에서, 폴리알라닌 쇄의 용매화가 펩티드 백본과 아미드-클램프 측쇄를 가교하는 단일 물 쉘로 감소된다. Tyr 51 고리 내의 대부분의 측쇄가, CsgG (및 CsgGC1S) X-선 구조에서 관찰된, 내부를 향하는 중심-지시 위치와 비교하여, 용매를 향해 회전하였다. 이러한 모델은 AMBER99SB-ILDN54 (Lindorff-Larsen, K. et al., Proteins 2010, 78, 1950-1958) 역장을 사용한 GROMACS (Pronk, S. et al., Bioinformatics, 2013, 29, 845-854) 및 위치적으로 제한된 C-루프의 단부의 잔기 (Gln 47 및 Thr 58)의 Ca 원자로의 40 ns 전체-원자 명시적 용매 분자 동역학 시뮬레이션의 결과이다.
실시예
16: 핵산 시퀀싱을 위한
CsgG
나노포어의
용도
Phi29 DNA 폴리머라제 (DNAP)를 막 내에 위치하는 돌연변이체 또는 야생형 CsgG 나노포어와 함께 분자 모터로 사용하여, 포어를 통한 올리고머 프로브 DNA 가닥의 제어된 이동을 허용할 수 있다. 포어를 가로질러 전압이 인가될 수 있고, 나노포어의 한쪽 측면 상의 염 용액 내의 이온의 이동으로부터 전류가 생성될 수 있다. 프로브 DNA가 포어를 통해 이동할 때, 포어를 통한 이온 흐름이 DNA와 관련되어 변한다. 이러한 정보는 서열 의존적인 것으로 나타났고, 프로브의 서열이 전류 측정치 예컨대 상기 실시예 14에서 기술된 것으로부터 정확하게 판독되도록 허용한다.
실시예 17
이러한 실시예는 CsgG 내에서의 DNA 거동을 연구하도록 시행된 시뮬레이션을 기술한다.
물질 및 방법
조향식 분자 동역학 시뮬레이션을 수행하여, DNA 변위에 대한 CsgG-Eco 및 다양한 돌연변이체의 에너지 장벽의 규모를 연구하였다. GROMOS 53a6 역장 및 SPC 물 모델과 함께 GROMACS 패키지 버전 4.0.5를 사용하여 시뮬레이션을 수행하였다. 단백질 데이터 뱅크의 등록 코드 4UV3로부터 CsgG-Eco (서열식별번호: 390)의 구조를 취하였다. CsgG-Eco 돌연변이체의 모델을 만들기 위해, PyMOL을 사용하여 야생형 단백질 구조를 돌연변이시켰다. 연구된 돌연변이체는 CsgG-Eco-(F56A) (돌연변이 F56A가 있는 서열식별번호: 390), CsgG-Eco-(F56A-N55S) (돌연변이 F56A/N55S가 있는 서열식별번호: 390) 및 CsgG-Eco-(F56A-N55S-Y51A) (돌연변이 F56A/N55S/Y51A가 있는 서열식별번호: 390)였다.
그 후, DNA를 포어 내에 놓았다. 2가지 상이한 시스템을 설정하였다:
i. 단일 구아닌 뉴클레오티드를 협착부 영역 바로 위 (잔기 56 고리에서 약 5-10 옹스트롬 위)에서 포어 내에 놓았다.
ii. 단일 가닥의 DNA (ssDNA)를 5' 끝부분을 포어의 베타-배럴 측면을 향하게 하여 포어 축을 따라 놓았다. 이러한 설정에서, ssDNA가 전체 길이의 포어에 예비-관통되었다.
그 후, 시뮬레이션 박스를 용매화하고, 최대 경사(steepest descent) 알고리즘을 사용하여 에너지를 최소화하였다.
300 K로 베렌드센(Berendse) 써모스탯 및 베렌드센 바로스탯을 사용하여, 각각의 시스템을 NPT 앙상블에서 시뮬레이션하였다. 시뮬레이션 동안, 포어의 백본에 구속이 적용되었다.
DNA를 포어를 통하여 당기기 위해, 단일 구아닌 시뮬레이션에서 인 원자에 인력(pulling force)을 인가하였다. ssDNA 시뮬레이션에서는, 가닥의 5' 끝부분의 인 원자에 인력을 인가하였다. 상기 언급된 DNA 인 원자와 포어 축에 대해 평행하게 일정 속도로 이동하는 허점 사이에 스프링을 연결함으로써, 인력을 일정한 속도로 인가하였다. 이러한 스프링이 어떠한 형상도 갖지 않거나 또는 어떠한 유체역학적 드래그(drag)도 받지 않는다는 것을 주의한다. 스프링 상수는 5 kJmol-1Å-2와 동일하였다.
결과
단일 G 변위
도 12에 나타난 바와 같이, 인력 대 시간의 플롯은 야생형 CsgG-Eco 포어의 페닐알라닌 잔기 F56의 고리 내로의 뉴클레오티드 진입에 큰 장벽이 있다는 것을 나타낸다. 연구된 CsgG-Eco 돌연변이체들에 대해서는 구아닌 변위에 대한 유의한 장벽이 관찰되지 않았다.
ssDNA
변위
ssDNA 변위에 대해, 포어 당 2회의 시뮬레이션이 시행되었고, 각각의 시행은 인가된 당기는 속도가 상이하였다 (100 A/ns 및 10 A/ns). 당기는 속도가 더 빠른 시뮬레이션을 도해하는 도 13에 나타난 바와 같이, ssDNA 변위를 가능하게 하기 위해 CsgG 야생형 포어가 가장 큰 인력을 요구하였다. 당기는 속도가 더 느린 시뮬레이션을 도해하는 도 14에 나타난 바와 같이, ssDNA 변위를 가능하게 하기 위해 CsgG-Eco (야생형, 서열식별번호: 390) 및 CsgG-Eco-(F56A) 포어 양쪽이 가장 큰 힘의 인가를 요구하였다. CsgG 및 MspA 기준선 포어를 통한 ssDNA 변위에 요구되는 인력 사이의 비교는 유사한 수준의 ssDNA 변위를 허용하는데 CsgG 포어의 돌연변이가 요구된다는 것을 시사한다.
실시예
18
이러한 실시예는 여러 CsgG 돌연변이체의 특성화를 기술한다.
물질 및 방법
실험을 설정하기 전에, DNA 구축물 X (최종 농도 0.1 nM, 구축물 X의 만화적인 표현 및 설명에 대해서 도 22를 참조한다)를 실온에서 5분 동안 T4 Dda - E94C/C109A/C136A/A360C (돌연변이 E94C/C109A/C136A/A360C가 있는 서열식별번호: 412이고, 나노포어 시스템에 10 nM의 최종 농도로 첨가되며, 완충제 (151.5 mM KCl, 25 mM 인산칼륨, 5% 글리세롤, pH 7.0, 1 mM EDTA) 내에서 제공됨)와 함께 예비 인큐베이션하였다. 5분 후, TMAD (100 μM)를 예비-혼합물에 첨가하고, 혼합물을 추가로 5분 동안 인큐베이션하였다. 최종적으로, MgCl2 (나노포어 시스템에 1.5 mM의 최종 농도로 첨가됨), ATP (나노포어 시스템에 1.5 mM의 최종 농도로 첨가됨), KCl (나노포어 시스템에 500 mM의 최종 농도로 첨가됨) 및 인산칼륨 완충제 (나노포어 시스템에 25 mM의 최종 농도로 첨가됨)를 예비-혼합물에 첨가하였다.
완충제 (25 mM 인산칼륨 완충제, 150 mM 페로시안화칼륨 (II), 150 mM 페리시안화칼륨 (III), pH 8.0) 내의 블록 공중합체에 삽입된 다양한 단일 CsgG 나노포어로부터 전기적 치수를 획득하였다. 블록 공중합체에 삽입된 단일 포어를 달성한 후, 완충제 (2 mL, 25 mM 인산칼륨 완충제, 150 mM 페로시안화칼륨 (II), 150 mM 페리시안화칼륨 (III), pH 8.0)를 시스템에 유동시켜 임의의 과량의 CsgG 나노포어를 제거하였다. 그 후, 150 uL의 500 mM KCl, 25 mM 인산칼륨, 1.5 mM MgCl2, 1.5 mM ATP, pH 8.0을 시스템에 유동시켰다. 10분 후, 150 uL의 500 mM KCl, 25 mM 인산칼륨, 1.5 mM MgCl2, 1.5 mM ATP, pH 8.0을 시스템에 유동시키고 나서, 효소 (T4 Dda -E94C/C109A/C136A/A360C, 10 nM 최종 농도), DNA 구축물 X (0.1 nM 최종 농도), 연료 (MgCl2 1.5 mM 최종 농도, ATP 1.5 mM 최종 농도) 예비 혼합물 (총 150 ㎕)를 단일 나노포어 실험 시스템 내로 유동시켰다. - 120 mV에서 실험을 수행하고, 헬리카제-제어 DNA 이동을 모니터링하였다.
결과
증가된
범위를 나타내는
포어
(도 15 내지 17, 및 27 내지 39)
CsgG-Eco-(StrepII(C)) (서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 범위가 ~ 10 pA인 한편 (도 15(a) 참조), 하기의 CsgG-Eco 포어 돌연변이체들은 증가된 전류 범위를 나타냈다:
1 - CsgG-Eco-(Y51N-F56A-D149N-E185R-E201N-E203N-StrepII(C))9 (돌연변이 Y51N/F56A/D149N/E185R/E201N/E203N이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 30 pA의 범위를 나타냈다 (도 15(b) 참조).
2 - CsgG-Eco-(N55A-StrepII(C))9 (돌연변이 N55A가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 35 pA의 범위를 나타냈다 (도 15(c) 참조).
3 - CsgG-Eco-(N55S-StrepII(C))9 (돌연변이 N55S가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 40 pA의 범위를 나타냈다 (도 16(a) 참조).
4 - CsgG-Eco-(Y51N-StrepII(C))9 (돌연변이 Y51N이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 40 pA의 범위를 나타냈다 (도 16(b) 참조).
5 - CsgG-Eco-(Y51A-F56A-StrepII(C))9 (돌연변이 Y51A/F56A가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 30 pA의 범위를 나타냈다 (도 16(c) 참조).
6 - CsgG-Eco-(Y51A-F56N-StrepII(C))9 (돌연변이 Y51A/F56N이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 20 pA의 범위를 나타냈다 (도 17(a) 참조).
7 - CsgG-Eco-(Y51A-N55S-F56A-StrepII(C))9 (돌연변이 Y51A/N55S/F56A가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 30 pA의 범위를 나타냈다 (도 17(b) 참조).
8 - CsgG-Eco-(Y51A-N55S-F56N-StrepII(C))9 (돌연변이 Y51A/N55S/F56N이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 30 pA의 범위를 나타냈다 (도 17(c) 참조).
13 - CsgG-Eco-(F56H-StrepII(C))9 (돌연변이 F56H가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 35 pA의 범위를 나타냈다 (도 27 참조).
14 - CsgG-Eco-(F56Q-StrepII(C))9 (돌연변이 F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 40 pA의 범위를 나타냈다 (도 28).
15 - CsgG-Eco-(F56T-StrepII(C))9 (돌연변이 F56T가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 35 pA의 범위를 나타냈다 (도 29 참조).
16 - CsgG-Eco-(S54P/F56A-StrepII(C))9 (돌연변이 S54P/F56A가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 35 pA의 범위를 나타냈다 (도 30 참조).
17 - CsgG-Eco-(Y51T/F56A-StrepII(C))9 (돌연변이 Y51T/F56A가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 30 pA의 범위를 나타냈다 (도 31 참조).
18 - CsgG-Eco-(F56P-StrepII(C))9 (돌연변이 F56P가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 30 pA의 범위를 나타냈다 (도 32 참조).
19 - CsgG-Eco-(F56A-StrepII(C))9 (돌연변이 F56A가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 40 pA의 범위를 나타냈다 (도 33 참조).
20 - CsgG-Eco-(Y51T/F56Q-StrepII(C))9 (돌연변이 Y51T/F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 30 pA의 범위를 나타냈다 (도 34 참조).
21 - CsgG-Eco-(N55S/F56Q-StrepII(C))9 (돌연변이 N55S/F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 35 pA의 범위를 나타냈다 (도 35 참조).
22 - CsgG-Eco-(Y51T/N55S/F56Q-StrepII(C))9 (돌연변이 Y51T/N55S/F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 35 pA의 범위를 나타냈다 (도 36 참조).
23 - CsgG-Eco-(F56Q/N102R-StrepII(C))9 (돌연변이 F56Q/N102R이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 30 pA의 범위를 나타냈다 (도 37 참조).
24 - CsgG-Eco-(Y51Q/F56Q-StrepII(C))9 (돌연변이 Y51Q/F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 40 pA의 범위를 나타냈다 (도 38 참조).
25 - CsgG-Eco-(Y51A/F56Q-StrepII(C))9 (돌연변이 Y51A/F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 ~ 35 pA의 범위를 나타냈다 (도 39 참조).
증가된
처리량을 나타내는
포어
(도 18 및 19 참조)
도 18 및 19로부터 볼 수 있듯이, 하기의 돌연변이체 포어 (하기의 9-12)는 4시간 내에 채널 당 다수의 헬리카제 제어 DNA 이동 (도 18 및 19에서 X로 표지됨)을 나타낸 반면, 도 18(a)에서 제시된 CsgG-Eco-(StrepII(C)) (서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)는 4시간 내에 채널 당 단지 1회 또는 2회의 헬리카제 제어 DNA 이동 (도 18(a)에서 X로 표지됨)을 빈번하게 나타냈고, 대신 장기간의 차단 영역 (도 18(a)에서 Y로 표지됨)을 나타냈다.
9 - CsgG-Eco-(D149N-E185N-E203N-StrepII(C))9 (돌연변이 D149N/E185N/E203N이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다) (도 18(b))
10 - CsgG-Eco-(D149N-E185N-E201N-E203N-StrepII(C))9 (돌연변이 D149N/E185N/E201N/E203N이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다) (도 18(c))
11 - CsgG-Eco-(D149N-E185R-D195N-E201N-E203N)-StrepII(C))9 (돌연변이 D149N/E185R/D195N/E201N/E203N이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다) (도 19(a))
12 - CsgG-Eco-(D149N-E185R-D195N-E201R-E203N)-StrepII(C))9 (돌연변이 D149N/E185R/D195N/E201R/E203N이 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다) (도 19(b))
증가된
삽입을 나타내는
포어
(도 20 및 21)
도 20 및 21을 비교함으로써 볼 수 있듯이, CsgG-Eco-(StrepII(C)) (서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다) 포어 (도 20에서 제시됨)와 비교하여 도 21에서 제시된 돌연변이체 포어 CsgG-Eco-(T150I-StrepII(C))9 (돌연변이 T150I가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)가 증가된 포어 개수 (~4-5배)로 막 내에 존재하였다. 도 20 및 21의 화살표는 4시간 실험에서 블록 공중합체 내로 삽입된 CsgG-Eco 나노포어의 개수를 도해한다 (도 20의 130-140 및 도 21의 1-11 각각은 별개의 나노포어 실험에 상응하였다). CsgG-Eco-(StrepII(C)) (서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)의 경우, 3개의 실험이 1개의 나노포어의 삽입을 나타낸 반면, 돌연변이체 포어 (CsgG-Eco-(T150I-StrepII(C))9)의 경우는, 각각의 실험이 적어도 1개의 나노포어의 삽입을 나타냈고, 여러 실험은 다중 포어 삽입을 나타냈다.
실시예
19
이러한 실시예는 CsgG 포어를 정제하기 위해 개발된 이. 콜라이 정제 방법을 기술한다.
물질 및 방법
폴리펩티드 Pro-CsgG-Eco-(StrepII(C)) (서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착되고, Pro는 서열식별번호: 436이고, N-말단에서 부착된다)를 코딩하는 DNA가 진스크립트 USA 인크(GenScript USA Inc.)에서 합성되었고, 앰피실린 저항성 유전자를 함유하는 pT7 벡터 내로 클로닝되었다. pT7 벡터의 단백질 발현을 이소프로필 β-D-1-티오갈락토피라노시드 (IPTG)에 의해 유도하였다. DNA 용액의 농도를 400 ng/uL로 조정하였다. DNA (1 ㎕) 를 사용하여, Lemo21 (DE3) 수용성 이. 콜라이 세포 (50 ㎕, NEB, 카탈로그 번호 C2528H)를 형질전환시켰다. 형질전환 전에, CsgG 유전자를 Lemo21 (DE3) 세포 (진 브리지스 게엠베하(Gene Bridges GmbH), 독일)로부터 녹아웃(knock out)시켰다. 그 후, 세포를 앰피실린 (0.1 mg/mL)을 함유하는 LB 한천 상에 플레이팅하고, 약 16시간 동안 37℃에서 인큐베이션하였다.
앰피실린을 함유하는 LB 플레이트 상에서 성장된 박테리아 콜로니 내로 CsgG 플라스미드가 혼입되었다. 한 이같은 콜로니를 사용하여, 카르베니실린 (0.1 mg/mL)을 함유하는 LB 배지의 출발 배양물 (100 mL)에 접종하였다. OD600이 1.0 - 1.2에 도달할 때까지 출발 배양물을 진탕하면서 37℃에서 성장시켰다. 출발 배양물을 사용하여, 카르베니실린 (0.1 mg/mL) 및 람노스 (500 μM)를 함유하는 신선한 500 mL의 LB 배지에 0.1의 O.D. 600으로 접종하였다. OD600이 0.6에 도달할 때까지 배양물을 진탕하면서 37℃에서 성장시켰다. 그 후, 배양물의 온도를 18℃로 조정하고, IPTG (0.2 mM 최종 농도)를 첨가하여 유도를 개시시켰다. 진탕하면서 18℃에서 약 18시간 동안 유도를 수행하였다.
유도 후, 6,000g에서 30분 동안의 원심분리로 배양물을 펠릿화하였다. 펠릿을 프로테아제 억제제 (머크 밀리포어(Merck Millipore) 539138), 벤조나제 뉴클레아제 (시그마(Sigma) E1014) 및 1× 버그버스터(bugbuster) (머크 밀리포어 70921) pH 8.0를 함유하는 50 mM 트리스, 300 mM NaCl (펠릿 1 그램 당 약 10 mL의 완충제)에 재현탁시켰다. 충분히 균질할 때까지 현탁액을 잘 혼합한 후, 4℃에서 약 5시간 동안 샘플을 롤러 혼합기로 전달하였다. 20,000g에서 45분 동안의 원심분리로 용해물을 펠릿화시키고, 상청액을 0.22 μM PES 주사기 필터를 통해 여과하였다. CsgG를 함유한 상청액 (샘플 1로 알려짐)을 칼럼 크로마토그래피에 의한 정제를 위해 진행시켰다.
샘플 1을 5 mL 스트렙 트랩 칼럼 (GE 헬스케어)에 적용하였다. 10 칼럼 부피의 안정적인 기준선이 유지될 때까지 칼럼을 25 mM 트리스, 150 mM NaCl, 2 mM EDTA, 0.01 % DDM pH 8로 세정하였다. 그 후, 칼럼을 25 mM 트리스, 2M NaCl, 2 mM EDTA, 0.01 % DDM pH 8로 세정한 후, 150 mM 완충제로 되돌렸다. 10 mM 데스티오비오틴으로 용출을 수행하였다. CsgG 단백질의 스트렙 트랩 (GE 헬스케어) 정제의 크로마토그래피 트레이스의 예가 도 23에서 제시된다. 용출 피크가 E1로 표지된다. 도 24는 초기 스트렙 정제 후의 CsgG-Eco 단백질의 전형적인 SDS-PAGE 가시화의 예를 나타낸다. 레인 1-3은 화살표로 지시된 바와 같은 CsgG 단백질을 함유한 주요 용출 피크 (도 23에서 E1로 표지됨)를 나타낸다. 레인 4-6은 오염물을 함유한 주요 용출 피크 (도 23에서 E1로 표지됨)의 꼬리의 용출 분획을 나타낸다.
용출 피크를 풀링하고, 15분 동안 65℃로 가열하여, 열에 불안정한 오염된 단백질을 제거하였다. 가열된 용액을 10분 동안 20,000g에서 원심분리하고, 펠릿을 폐기하였다. 상청액을 25 mM 트리스, 150 mM NaCl, 2 mM EDTA, 0.01% DDM, 0.1% SDS pH 8에서 120 mL 세파덱스(Sephadex) S200 칼럼 (GE 헬스케어) 상에서의 겔 여과에 적용하였다. 단백질의 낮은 트립토판 성분으로 인해 220 nM에서 모니터링을 수행하였다. 약 55 mL 부피에서 샘플이 용출되었다 (도 25는 크기 배제 컬럼 트레이스를 나타내고, 55 mL 샘플 피크가 별표로 표지된다). 이러한 용출 피크를 4-20% TGX (도 26 참조, 바이오 래드) 상에서 러닝시켜, 관심 포어인 CsgG-Eco-(StrepII(C)) (서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다)의 존재를 확인하였다. 확인된 분획을 풀링하고, 50 kD 아미콘(Amicon) 스핀 칼럼에 의해 농축하였다.
실시예
20
이러한 실시예는 CsgG-Eco-(Y51T/F56Q)-StrepII(C))9 (돌연변이 Y51T/F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다; 포어 돌연변이체 20번)와 T4 Dda -(E94C/C109A/C136A/A360C) (돌연변이 E94C/C109A/C136A/A360C 및 그 후의 (ΔM1)G1G2가 있는 서열식별번호: 412) 사이의 상호작용을 연구하기 위해 시행된 시뮬레이션을 설명한다.
시뮬레이션 방법
GROMOS 53a6 역장 및 SPC 물 모델과 함께 GROMACS 패키지 버전 4.0.5를 사용하여 시뮬레이션을 수행하였다.
CsgG-Eco-(Y51T/F56Q)-StrepII(C))9 (돌연변이 Y51T/F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다; 포어 돌연변이체 20번) 모델은 단백질 데이터 뱅크의 등록 번호 4UV3 및 4Q79에서 확인된 CsgG의 결정 구조를 기초로 하였다. PyMOL을 사용하여 관련된 돌연변이를 만들었다. 그 후, 생성된 포어 모델을 최대 경사 알고리즘을 사용하여 에너지를 최소화하였다. T4 Dda -(E94C/C109A/C136A/A360C) (돌연변이 E94C/C109A/C136A/A360C 및 그 후의 (ΔM1)G1G2가 있는 서열식별번호: 412) 모델은 단백질 데이터 뱅크의 등록 번호 3UPU에서 확인된 Dda1993 구조를 기초로 하였다. 또 다시, PyMOL을 사용하여 관련된 돌연변이를 만들고, 모델을 최대 경사 알고리즘을 사용하여 에너지를 최소화하였다.
그 후 T4 Dda -(E94C/C109A/C136A/A360C) (돌연변이 E94C/C109A/C136A/A360C 및 그 후의 (ΔM1)G1G2가 있는 서열식별번호: 412) 모델을 상기 CsgG-Eco-(Y51T/F56Q)-StrepII(C))9 (돌연변이 Y51T/F56Q가 있는 서열식별번호: 390이고, 여기서 StepII(C)는 서열식별번호: 435이고, C-말단에서 부착된다; 포어 돌연변이체 20번) 위에 놓았다. 상이한 초기 효소 형상으로 3회의 시뮬레이션을 수행하였다 (시행 1 내지 3 (0 ns), 도 40 참조).
모든 효소 형상에서, DNA의 5' 끝부분이 포어를 가리키도록 효소가 배향되었고, 시뮬레이션 동안 효소가 구속되지 않았다. 포어 백본이 구속되었고, 시뮬레이션 박스를 용매화하였다. 300 K로 베렌드센 써모스탯 및 베렌드센 바로스탯을 사용하여, 40 ns 동안 NPT 앙상블에서 시스템을 시뮬레이션하였다.
효소와 포어 사이의 접촉을 GROMACS 분석 소프트웨어 및 또한 국소적으로 작성된 코드 양쪽 모두를 사용하여 분석하였다. 하기의 표는 포어 및 효소 아미노산 양쪽 모두에 대해 관찰된 접촉 수를 나타낸다. 표 6-8은 효소 상의 아미노산 접촉 지점과 상호작용하는 포어 상의 아미노산 접촉 지점을 나타낸다. 3회의 시뮬레이션 중 2회에서, 효소가 포어의 상부 상에서 틸팅(tilting)된다 (도 40 및 41의 시행 2 및 3 (20, 30 및 40 ns) 참조). 시행 1은 효소가 틸팅되지 않았음을 나타내고, 따라서 표 6에서 상호작용이 높은 것으로 나타난 지점들이 포어 캡 상에서의 효소 안정성을 증가시키기 위해 최적화될 수 있다.
<표 6>
표 6 = 시행 1 효소 및 포어 접촉 상호작용
<표 7>
표 7 = 시행 2 효소 및 포어 접촉 상호작용
<표 8>
표 8 = 시행 3 효소 및 포어 접촉 상호작용
실시예
21: 채널
협착부
내에 핵산을 포착하는
CsgG
나노포어의
능력
나노포어를 핵산 시퀀싱에 사용하는 것은 나노포어에 의한 단일 가닥 DNA의 포착 및 관통을 필요로 한다. 이러한 실시예에서, 단일 가닥 DNA 오버행을 보유하는 DNA 헤어핀의 존재 하에 CsgG WT 단백질의 단일 채널 전류 트레이스를 추적하였다. 도 56에 제시된 예시적인 트레이스는 + 50 mV 또는 -50 mV 간격 (화살표로 지시됨)에서 측정된 전위에 반응하여 변하는 전류를 나타낸다. 마지막 +50 mV 분절에서의 하향 전류 차단은 단일-가닥 헤어핀 끝부분이 내부 포어 협착부 내로 관통하여 거의 완전한 전류 차단에 이르는 것을 나타낸다. -50 mV로의 전기장 역전은 포어의 전기영동성 차단 해제를 초래한다. 새로운 +50 mV 에피소드는 또 다시 DNA 헤어핀 결합 및 포어 차단을 초래한다. +50 mV 분절에서, 헤어핀 구조의 언폴딩은 전류 차단의 역전에 의해 지시되는 전류 차단의 종료에 이를 수 있다. 서열 3' GCGGGGAGCGTATTAGAGTTGGATCGGATGCAGCTGGCTACTGACGTCATGACGTCAGTAGCCAGCATGCATCCGATC-5' (서열식별번호: 441)의 헤어핀이 챔버의 시스 측면으로 10 nM의 최종 농도로 첨가되었다.
실시예
22. 이러한
실시예는
내부
협착부가
변경된 돌연변이체
CsgG
포어의
생성을 기술한다.
이러한 실시예에서, 협착부 루프 내의 서열 PYPA (잔기 50-53)가 GG로 교체된 돌연변이체 CsgG 포어인 CsgG-ΔPYPA에 대해 안정성 및 채널 성질이 실연된다. 이러한 돌연변이체에 대해, 위치 38 내지 63의 협착부 모티프가 서열식별번호: 354에 상응한다. 센싱 용도 동안 측정된 전도도가 포어에 결합하거나 이를 관통하는 기질의 성질에 대해 가장 민감한 곳인 포어 판독 헤드, 즉 포어의 가장 좁은 부분의 복잡성을 감소시키기 위해, 이러한 돌연변이로 포어 협착부로부터 Y51이 제거되고 협착부 루프가 짧아질 것으로 예상된다. 도 57에 제시된 바와 같이, PYPA 서열의 교체는 CsgG 9량체의 안정성을 유지하고, 전도도가 증가된 포어를 초래한다.
나니온 (독일 뮌헨)으로부터의 오르비트 16 키트로의 병렬식 고해상도 전기 기록을 사용하여 단일-채널 전류 기록으로 CsgG-ΔPYPA 포어 돌연변이체를 분석하였다. 간략하게, 1,2-디피타노일-sn-글리세로-3-포스포콜린 (아반티 폴라 리피즈)의 수평 이중층이 16-채널 다중전극 공동 어레이 (MECA) 칩 (아이오네라, 독일 프라이부르크)의 미세공동 (피코리터 이하의 부피)에서 형성되었다51. 이중층 위 및 아래의 시스 및 트랜스 공동 양쪽 모두 1.0 M KCl, 25 mM 트리스-HCl pH 8.0을 함유하였다. 채널을 막 내로 삽입하기 위해, 25 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM DTT, 0.5% C8E4, 5 mM LDAO에 용해된 CsgG를 100 nM의 최종 농도로 시스 구획에 첨가하였다. 3 kHz의 로우-패스 필터링 주파수 및 10 kHz의 샘플링 주파수로 테셀라 트리톤 16-채널 증폭기를 사용하여 +50 mV 및 -50 mV (시스 측면 기준)의 유지 전위에서 막횡단 전류를 기록하였다. p클램프 스위트 (몰레큘러 디바이시즈, 미국)의 클램프피트를 사용하여 전류 트레이스를 분석하였다.
실시예
23:
이러한 실시예는 아밀로이드 분비 채널 CsgG에 대한 구조적 및 기계적 통찰을 기술한다.
컬리는 박테로이데테스(Bacteroidetes) 및 프로테오박테리아(Proteobacteria) (주로 a 및 c 클래스)에 의해 형성된 균막(pellicle) 바이오필름 내의 세포외 매트릭스의 주요 단백질 성분을 구성하는 기능성 아밀로이드 섬유이다1-3. 이는 병원성 균주에 피트니스 장점을 제공하고, 균혈증 동안 강한 염증유발성 반응을 유도한다1 ,4, 5. 컬리 형성은 외막 지질단백질 CsgG 및 2개의 가용성 보조 단백질 CsgE 및 CsgF를 포함하는 전용 단백질 분비 기구를 필요로 한다6 , 7. 본원에서, 본 발명가들은 이의 천연 막-추출 형상, 뿐만 아니라 지질화되지 않은 가용성 형태의 에스케리키아 콜라이 CsgG의 X-선 구조를 보고한다. CsgG는 9개의 안티코돈-결합-도메인-유사 단위로 구성된 올리고머 수송 복합체를 형성하고, 이러한 단위들은 이중층을 횡단하고 원형질막 주위공간 내의 케이지-유사 전정에 연결되는 36-가닥의 β-배럴을 초래한다. 막횡단 및 원형질막 주위공간 도메인은 쭉 뻗은 폴리펩티드 기질을 분비 포어를 통해 안내할 수 있는 3개의 스태킹된 동심성 페닐알라닌, 아스파라긴 및 타이로신 고리로 구성된 0.9-nm 채널 협착부에 의해 분리된다. 특이성 인자 CsgE는 분비 채널의 원형질막 주위공간 면에 결합하고 이를 폐쇄하는 9량체 어댑터를 형성하여, 24,000 Å3 예비-협착부 챔버를 생성시킨다. 본 발명가들의 구조적, 기능적 및 전기생리학적 분석은 CsgG가 확산을 기초로 하고 엔트로피에 의해 구동되는 수송 메커니즘을 사용할 것으로 예상되는, 관문화되지 않은 비-선택적 단백질 분비 채널이라는 것을 시사한다.
컬리는 인간의 퇴행성 질환으로부터 가장 잘 알려진 아밀로이드 원섬유의 구조적 및 물리적 특성을 갖는 박테리아 표면 부속물이다7 - 9. 그러나, 박테리아 아밀로이드 예컨대 컬리의 역할은 바이오필름 형성을 용이하게 하는 것이다4 , 10. 단백질 미스폴딩(misfolding)의 생성물인 병원성 아밀로이드와 달리, 컬리 형성은 에스케리키아 콜라이에서 2개의 전용 오페론인 csgBAC (컬리 특이적 유전자 BAC) 및 csgDEFG에 의해 코딩되는 단백질들에 의해 조정된다 (도 46)6 , 7. 분비 후, CsgB가 CsgA 서브유닛을 컬리 원섬유 내로 핵화시킨다7 ,11, 12. CsgA 및 CsgB의 분비 및 세포외 침착은 각각 CsgE 및 CsgF인 2개의 보조 인자, 뿐만 아니라 외막 내에 위치하는 262-잔기 지질단백질인 CsgG에 의존적이다13 - 16. 가수분해가 가능한 에너지 공급원 또는 이온 구배가 외막에 없기 때문에, CsgG는 대안적으로 에너지화된 수송 메커니즘을 통해 작동되어야 하는 특수 클래스의 단백질 변위체(translocator)에 속한다. 구조 모델의 부재 하에, 어떻게 CsgG가 조절된 방식으로 생물학적 막을 가로질러 고도로 안정적인 아밀로이드-유사 원섬유의 분비 및 조립을 촉진하는지에 대한 동적 작용은 지금까지 수수께끼로 남았다.
외막 내로의 삽입 전에, 지질단백질은 지질단백질 국소화 (Lol) 경로에 의해 원형질막 주위공간을 가로질러 인도된다17. 본 발명가들은 지질화되지 않은 CsgG (CsgGC1S)가 포어-형성 단백질 및 독소에서 관찰되는 프리-포어 형태18와 유사한 가용성 원형질막 주위공간 중간체로서 단리될 수 있다는 것을 관찰하였다. CsgGC1S는 주로 단량체로서 확인되었고, 추가적으로 미량의 별개의 올리고머 복합체가 있었다 (도 47)19. 가용성 CsgGC1S 올리고머가 결정화되었고, 이의 구조가 2.8 Å로 결정되어, 꼬리-대-꼬리 상호작용으로 연결된 2개의 고리-형상 8량체 복합체 (C8)로 이루어진, 8중 2면각 대칭 (D8)이 있는 16량체 입자가 밝혀졌다 (도 47 및 도 46). CsgGC1S 프로토머는 아미노 및 카르복시 말단에 2개의 α-나선 (각각 αN 및 αC)이 확장된 안티코돈-결합 도메인 (ABD)-유사 폴드를 나타낸다 (도 42 및 도 48a-c). 추가적인 CsgG-특이적 요소는 β1 및 α1을 연결하는 확장된 루프, β3-β4 및 β5-α3을 연결하는 루프 내의 2개의 삽입물, 및 인접한 단량체들 사이에 패킹됨으로써 CsgG 올리고머화에 관여되는 확장된 α2 나선이다 (도 42b). β3-β5 시트의 후면과 확장된 β1-α1 루프 사이에서 추가적인 프로토머간 접촉부가 형성된다 (도 48d, e).
CsgGC1S 구조에서, α1을 N-말단 지질 앵커에 연결할 잔기 1-17이 무질서하고, 명확한 막횡단 (TM) 도메인이 분간될 수 없다 (도 42). CsgGC1S 및 천연의 막-추출 CsgG의 감쇠 전반사 푸리에 변환 적외선 분광학 (ATR-FTIR)에서, 후자에 β-시트 영역 (1,625-1,630 cm- 1)에서의 더 높은 흡수 및 랜덤 코일 및 α-나선 영역 (각각 1,645-1,650 cm-1 및 1,656 cm- 1)에서의 동반적인 감소가 있다는 것이 밝혀졌고 (도 43a), 이는 막-회합 CsgG가 β-배럴 도메인을 함유한다는 것을 시사한다. β-가닥 형성에 대한 후보 서열 신장물이 β3-β4 (잔기 134-154) 및 β5-α3 (잔기 184-204)을 연결하는 불량하게 질서화된 확장된 루프들에서 발견된다; 이들의 결실은 컬리 형성 상실을 초래하였다 (도 43b). 세제-추출 CsgG의 결정 구조에서, 양쪽 영역이 2개의 인접한 β-헤어핀으로 형상적으로 재배열되어, β3-β4 (TM1) 및 β5-α3 (TM2)에 의해 형성된 β-시트를 확장한다는 것이 입증되었다 (도 43c). CsgG 올리고머에서의 이들의 병치는 복합적인 36-가닥 β-배럴을 초래하였다 (도 43d). 결정화된 CsgGC1S 올리고머는 D8 대칭을 나타낸 반면, CsgG 구조는 D9 대칭을 나타냈고, 이때 CsgG 프로토머는 중심축에 대한 5° 회전 및 방사축을 따른 4 Å 전이를 제외하고는 등가의 프로토머간 접촉부를 유지하였다 (도 47). 이러한 관찰은 막-추출 CsgG에 대한 C9 및 D9 대칭을 독점적으로 발견한 단일-입자 전자 현미경에 의해 밝혀진 용액 내에서의 올리고머 상태에서 일치되었다 (도 47). 지질화되지 않은 샘플에 단량체가 우세하게 존재하는 것 및 막-결합 단백질과의 대칭 부조화는 막 삽입 전에, CsgG가 LolA에 결합된 단량체 형태로 외막에 표적화된다는 것과 C8 및 D8 입자가 CsgGC1S의 고도로 농축된 용액의 인공물이라는 것을 나타낸다. 추가로, 본 발명가들은 D9 복합체가 아니라 C9 9량체가 생리학적으로 관련된 입자를 형성한다는 것을 나타내는데, 단리된 이. 콜라이 외막에서, 관찰된 꼬리-대-꼬리 이량체화에 의해 둘러싸인 잔기에서의 시스테인 치환이 말레이미드폴리에틸렌 글리콜 (PEG, 5 kDa)로의 표지화에 대해 접근가능하기 때문이다 (도 49).
따라서, CsgG는 120 Å 폭 및 85 Å 높이의 9량체 수송 복합체를 형성한다. 이러한 복합체는 내경이 40 Å인 36-가닥 β-배럴을 통해 외막을 횡단한다 (도 43e). 인접한 프로토머를 감싸는 18-잔기 링커에 의해 코어 도메인으로부터 N-말단 지질 앵커가 분리된다 (도 48d). 전자 밀도 지도 상에서 N-말단 Cys 상의 디아실글리세롤- 및 아미드-연결 아실 쇄가 해상되지 않지만, Leu 2의 위치를 기초로, 지질 앵커는 β-배럴의 외벽에 플랭킹될 것으로 예상된다. 원형질막 주위공간 측면에서, 수송체는 용매에 접근가능한 내경 35 Å 및 높이 40 Å의 대형 공동을 형성하고, 이는 원형질막 주위공간에 나선 2에 의해 형성된 50 Å의 입을 개방한다 (도 43e). 이의 정점에서, 이러한 원형질막 주위공간 전정이 β1을 α1에 연결하는 보존된 12-잔기 루프 (C-루프)에 의해 TM 채널로부터 분리되고 (도 43e 및 44a, b), 이러한 루프는 분비 도관을 9.0 Å의 용매-배제 직경으로 협착시킨다 (도 44a, c). 이러한 포어 치수들은 5개의 잔기가 협착부의 높이에 스패닝되면서 1개 또는 2개 (예를 들어, 루프화 구조)의 쭉 뻗은 폴리펩티드 분절이 있는 것과 상용성일 것이다 (도 50). 협착부의 내강 라이닝은 잔기 Tyr 51, Asn 55 및 Phe 56의 측쇄에 의해 형성된 3개의 스태킹된 동심성 고리로 구성된다 (도 44a, b). 탄저균 PA63 독소에서, 위상적으로 등가인 동심성 Phe 고리 (Φ-클램프로 지칭됨)가 변위 채널의 입구에 라이닝되고, 폴리펩티드 포착 및 통과를 촉매한다20 - 22. CsgG-유사 변위체들의 다중 서열 정렬은 Phe 56의 절대적인 보존 및 Asn 55의 Ser 또는 Thr으로의 보존적 변동을 나타낸다 (도 51). Phe 56 또는 Asn 55가 Ala로 돌연변이되면 컬리 생산이 거의 상실되는 반면 (도 44d), Asn 55→Ser 치환은 야생형 분비 수준을 유지하고, 이는 CsgG 협착부에서 Φ-클램프의 스태킹된 형상에 이어서 수소-결합 공여체/수용체가 필요하다는 것을 함께 시사한다 (도 44b 및 도 51).
평면형 인지질 이중층에서 재구성된 CsgG의 단일-채널 전류 기록은 표준 전해질 조건 및 각각 +50 mV 또는 -50 mV의 전위를 사용하여 43.1 ± 4.5 pA (n = 33) 또는 -45.1 ± 4.0 pA (n = 13)의 정상 전류에 이르렀다 (도 44e, f 및 도 52). 관찰된 전류는 X-선 구조에서 나타난 중심 협착부의 치수 및 간단한 3-분절 포어 모델을 기초로 계산된 46.6 pA의 예상값과 잘 일치하였다 (도 44c). 음의 전기장 전위 하에 제2의 저-전도도 형상이 또한 관찰될 수 있었다 (-26.2±3.6 pA (n= 13); 도 52). 그러나, 이러한 종이 생리학적 조건 하에 존재하는지 여부는 명확하지 않다.
본 발명가들의 구조 데이터 및 단일-채널 기록은 CsgG가 관문화되지 않은 펩티드 확산 채널이라는 것을 시사한다. 모델 펩티드 확산 채널인 PA63에서, 폴리펩티드 통과는 변위 채널에서의 확산 단계를 수정하는 ΔpH-구동 브라운 래칫(Brownian ratchet)에 의존적이다20 - 22. 그러나, 이같은 양자 구배가 외막에 존재하지 않아, 대안적인 구동력을 필요로 한다. 상승된 농도에서 CsgG가 원형질막 주위공간 폴리펩티드의 비-선택적인 확산성 누출을 용이하게 할 수 있지만, 분비는 천연 조건 하에 CsgA에 대해 특이적이고, 원형질막 주위공간 인자 CsgE를 필요로 한다16,23. 과량의 CsgE의 존재 하에, 정제된 CsgG는 네이티브 PAGE 상에서 더욱 느리게 이동하는 종을 형성한다 (도 45a). SDS-PAGE 분석은 이러한 새로운 종이 1:1 화학량론으로 존재하는 CsgG-CsgE 복합체로 이루어진다는 것을 나타낸다 (도 45b). 풀-다운 친화력 정제에 의해 단리된 CsgG-CsgE의 냉동 전기 현미경검사 (냉동-EM) 가시화에서, 단일-입자 EM 및 크기-배제 크로마토그래피에서 또한 관찰된 C9 CsgE 올리고머와 크기 및 형상 면에서 유사하게, CsgG 9량체에 상응하는 9중 대칭 입자, 및 원형질막 주위공간 전정에 대한 입구에서의 추가적인 캡핑 밀도가 밝혀졌다 (도 45c-e 및 도 53). 관찰된 CsgG-CsgE 접촉부 계면의 위치가 CsgG 나선 2에서의 점 돌연변이를 차단함으로써 확인되었다 (도 53). 캡핑 기능과 일치하게, 단일-채널 기록은 CsgE가 변위체에 결합하는 것이 이의 이온 전도도의 특이적인 침묵화에 이르렀음을 나타냈다 (도 45f 및 도 52). 채널의 이러한 CsgE 캡핑은 CsgE 노나머 결합의 기능에서의 양단식(all-or-none) 반응인 것으로 보였다. 포화 시에는 CsgE 결합이 채널의 완전한 차단을 유도한 반면, 약 10 nM에서는 CsgE 결합과 해리 이벤트 사이에서의 평형이 간헐적으로 차단된 또는 완전히 개방된 변위체를 초래하였다. 1 nM 이하에서는, 단량체 CsgE와 단기적으로 마주치는 것으로부터 일시적인 (<1 ms) 부분적 차단 이벤트가 일어났을 수 있다.
따라서, CsgG 및 CsgE는 ~24,000 Å3의 중심 공동을 에워싸는 케이지화 복합체를 형성하는 것으로 보이고, 이는 외관 면에서 GroEL 샤페로닌 및 GroES 코-샤페로닌에서 나타난 기질 결합 공동 및 캡슐화 뚜껑 구조를 연상시킨다24. CsgG-CsgE 에워싸기는 129-잔기 CsgA의 완전한 또는 부분적인 포착과 상용성일 것이다. 변위 기질의 케이지화가 최근에 ABC 독소에서 관찰되었다25. 언폴딩된 폴리펩티드가 공간적으로 속박되면 이의 입체형상 공간이 감소되어, 엔트로피 전위가 생성되고, 이는 샤페로닌의 경우 폴리펩티드 폴딩에 유리한 것으로 나타났다24 , 26. 유사하게, 본 발명가들은 컬리 수송에서, CsgG 전정 내의 컬리 서브유닛의 국소적으로 높은 농도 및 입체형상적 속박이 변위 채널에 걸친 엔트로피성 자유 에너지를 생성시킬 것으로 추측한다 (도 45g). 협착부 내로 포착되면, 분비 채널 근처에서의 CsgE-매개 속박 및/또는 기질 포착으로부터 생성된 엔트로피 전위에 의해 수정된 브라운 확산에 의해 폴리펩티드 쇄가 점진적으로 밖으로 이동할 것으로 예상된다. 예비-협착부 공동에서의 완전한 속박에 대해, 언폴딩된 129-잔기 폴리펩티드가 벌크 용매로 탈출하는 것은 ~80 kcal/mol (약 340 kJ/mol)까지의 엔트로피성 자유 에너지 방출에 상응할 것이다 (참조문헌 27). CsgG-CsgE-CsgA 복합체의 조립 동안 방출되는 결합 에너지 및 원형질막 주위공간 내의 이미 낮춰진 CsgA 엔트로피에 의해 기질 도킹 및 속박의 초기 엔트로피 비용이 적어도 부분적으로 보상될 것이다. 이론적 근거로, 분비 복합체에 CsgA가 동원되는 것의 3가지 잠재적 경로가 구상될 수 있다 (도 54).
컬리-유도 바이오필름은 병원성 엔테로박테리아세아에(Enterobacteriaceae)에서 피트니스 및 발병 인자를 형성한다4 , 5. 이들의 독특한 분비 및 조립 성질 또한 (생물)공학 용도에 대해 급속하게 관심을 얻고 있다23 ,28, 29. 2가지 주요 분비 성분에 대한 본 발명가들의 구조적 특성화 및 생화학적 연구는 가수분해가 가능한 에너지 공급원, 막 전위 또는 이온 구배의 부재 하에서의 언폴딩된 단백질 기질의 막 변위에 대한 반복 메커니즘의 잠정적 모델을 제공한다 (도 45e 및 도 54). 제안된 분비 모델에서의 기여 인자의 전체적인 확인 및 탈구축은 수송 역학이 단일 분자 수준에서 정확하게 추적되도록 허용하기 위해 변위체의 시험관내 재구성을 필요로 할 것이다.
방법
클로닝 및 계통. 외막에 국소화되고 C-말단에 StrepII-태그가 부착된 CsgG (pPG1) 및 원형질막 주위공간의 C-말단에 StrepII-태그가 부착된 CsgGC1S (pPG2)의 생산을 위한 발현 구축물이 참고문헌 19에 기술되어 있다. 셀레노메티오닌 표지화를 위해, StrepII-태그가 부착된 CsgGC1S가 수율 증가로 인해 세포질에서 발현되었다. 따라서, 프라이머 5'-TCTTTAACCGCCCCGCCTAAAG-3' (정방향) 및 5'-CATTTTTTGCCCTCGTTATC-3' (역방향)으로의 역 PCR을 사용하여 N-말단 신호 펩티드를 제거하도록 pPG2가 변경되었다 (pPG3). 표현형 검정법을 위해, 이. 콜라이 BW25141의 csgG 결실 돌연변이체 (이. 콜라이 NVG2)를 참고문헌 30에 기술된 방법에 의해 구축하였다 (프라이머 5'-AATAACTCAACC GAT TTT TAA GCC CCA GCT TCA TAA GGA AAA TAA TCG TGT AGG CTG GAG CTG CTT C-3' 및 5'-CGC TTA AAC AGT AAA ATG CCG GAT GAT AAT TCC GGC TTT TTT ATC TGC ATA TGA ATA TCC TCC TTA G-3'를 사용함). Cys 접근성 검정법 및 채널 협착부의 표현형 프로빙에 사용된 다양한 CsgG 치환 돌연변이체가 trc 프로모터의 제어 하의 csgG를 함유하는 pTRC99a 벡터인 pMC2에서 출발하여 부위-지정 돌연변이유발 (퀵체인지 프로토콜; 스트라타진)에 의해 구축되었다14.
단백질 발현 및 정제. 19에 기술된 바와 같이 CsgG 및 CsgGC1S가 발현 및 정제되었다. 간략하게, pPG1로 형질감염된 이. 콜라이 BL 21 (DE3)에서 CsgG를 재조합으로 생산하고, 단리된 외막으로부터 완충제 A (50 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM EDTA, 1 mM 디티오트레이톨 (DTT)) 내의 1% n-도데실-b-D-말토시드 (DDM)를 사용하여 추출하였다. StrepII-태그가 부착된 CsgG를 5 ml 스트렙-탁틴 세파로스 칼럼 (이바 게엠베하) 상에 로딩하고, 0.5% 테트라에틸렌 글리콜 모노옥틸 에테르 (C8E4; 어피메트릭스) 및 4 mM 라우릴디메틸아민-N-옥시드 (LDAO; 어피메트릭스)가 보충된 20 칼럼 부피의 완충제 A로 세정하여 세제를 교환하였다. 2.5 mM D-데스티오비오틴을 첨가하여 단백질을 용출시키고, 결정화 실험을 위해 5 mg/ml로 농축하였다. 셀레노메티오닌 표지화를 위해, pPG3으로 형질전환되고 40 mg/ℓ의 L-셀레노메티오닌이 보충된 M9 최소 배지에서 성장된 Met 영양요구성 균주 B834 (DE3)에서 CsgGC1S를 생산하였다. 세포 펠릿을 컴플리트 프로테아제 인히비터 칵테일 (로슈)이 보충된 50 mM 트리스-HCl pH 8.0, 150 mM NaCl, 1 mM EDTA, 5 mM DTT에 재현탁시키고, 20 × 103 lb/in2으로 작동되는 TS 시리즈 세포 파괴기 (컨스탄트 시스템즈 엘티디)에 통과시켜 파괴시켰다. 표지된 CsgGC1S를 19에 기술된 바와 같이 정제하였다. 셀레노메티오닌의 산화를 방지하도록 정제 과정 전반에 걸쳐 DTT (5 mM)가 첨가되었다.
pNH27를 보유하는 이. 콜라이 NEBC2566 세포에서 CsgE를 생산하였다 (참조문헌 16). 25 mM 트리스-HCl pH 8.0, 150 mM NaCl, 25 mM 이미다졸, 5% (v/v) 글리세롤 내의 세포 용해물을 HisTrap FF (GE 헬스케어) 상에 로딩하였다. 20 mM 트리스-HCl pH 8.0, 150 mM NaCl, 5% (v/v) 글리세롤 완충제 내의 500 mM 이미다졸로의 선형 구배로 CsgE-his를 용출시켰다. CsgE를 함유하는 분획에 250 mM (NH4)2SO4를 보충하고, 20 mM 트리스-HCl pH 8.0, 100 mM NaCl, 250 mM (NH4)2SO4, 5% (v/v) 글리세롤로 평형화된 5 ml HiTrap 페닐 HP 칼럼 (GE 헬스케어)에 적용하였다. 20 mM 트리스-HCl pH 8.0, 10 mM NaCl, 5% (v/v) 글리세롤로의 선형 구배를 적용하여 용출시켰다. CsgE를 함유하는 분획을 20 mM 트리스-HCl pH 8.0, 100 mM NaCl, 5% (v/v) 글리세롤로 평형화된 수퍼로스 6 프렙 그레이드 10/600 (GE 헬스케어) 칼럼 상에 로딩하였다.
용액 내에서의 올리고머 상태 평가. 각각 약 0.5 mg의 세제-가용화 CsgG (0.5% C8E4, 4 mM LDAO) 및 CsgGC1S를 25 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM DTT, 4 mM LDAO 및 0.5% C8E4 (CsgG) 또는 25 mM 트리스-HCl pH 8.0, 200 mM NaCl (CsgGC1S)로 평형화된 수퍼덱스 200 10/300 GL 분석용 겔 여과 칼럼에 적용하고, 0.7 ml/분으로 러닝시켰다. 소 티로글로불린, 소 γ-글로불린, 닭 오브알부민, 말 미오글루불린 및 비타민 B12 (바이오-래드)로 칼럼 용출 부피를 보정하였다 (도 47). 막-추출 CsgG인 20 ㎎의 세제-가용화 단백질을 참고문헌 31에 기술된 절차를 사용하여 3-10% 청색 네이티브 PAGE 상에 또한 러닝시켰다 (도 47). 네이티브마크 (라이프 테크놀러지즈) 미염색 단백질 표준물 (7 ㎖)이 분자 질량 추정에 사용되었다.
결정화, 데이터 수집 및 구조 결정. 셀레노메티오닌-표지화 CsgGC1S를 3.8 mg/ml로 농축하고, 100 mM 아세트산나트륨 pH 4.2, 8% PEG 4000 및 100 mM 말론산나트륨 pH 7.0을 함유하는 용액에 대한 시팅-드롭 증기 확산에 의해 결정화시켰다. 15% 글리세롤이 보충된 결정화 완충제에서 결정을 인큐베이션하고, 액체 질소에서 급속-냉동시켰다. 세제-가용화 CsgG를 5 mg/ml로 농축하고, 100 mM 트리스-HCl pH 8.0, 8% PEG 4000, 100 mM NaCl 및 500 mM MgCl2를 함유하는 용액에 대한 행잉-드롭 증기 확산에 의해 결정화시켰다. 결정을 액체 질소에서 급속-냉동시키고, 결정화 용액 내에 존재하는 세제에 의해 동결보호시켰다. 결정 상태의 최적화 및 회절 품질이 양호한 결정에 대한 스크리닝을 위해, 프록시마-1 및 프록시마-2a (솔레일, 프랑스), PX-I (스위스 라이트 소스, 스위스), I02, I03, I04 및 I24 (다이아몬드 라이트 소스, 영국) 및 ID14eh2, ID23eh1 및 ID23eh2 (ESRF, 프랑스) 빔라인에서 결정을 분석하였다. CsgGC1S 및 CsgG의 구조 결정에 사용된 최종 회절 데이터가 각각 I04 및 I03 빔라인에서 수집되었다 (데이터 수집 및 정밀화 통계에 대해 도 55a를 참조한다). CsgGC1S에 대한 회절 데이터를 Xia2 및 XDS 패키지32 , 33를 사용하여 프로세싱하였다. CsgGC1S의 결정은 비대칭 단위 내의 16개의 단백질 카피를 함유하는, 단위 셀 치수가 a=101.3 Å, b=103.6 Å, c=141.7 Å, α=111.3°, β=90.5°, γ=118.2°인 공간 군 P1에 속했다. 구조 결정 및 정밀화를 위해, 0.9795 Å 파장에서 수집된 데이터를 최고 해상도 쉘에서의 Ι/δΙ 차단값 2를 기초로 2.8 Å에서 절단하였다. 단일 비정상 분산 (SAD) 실험으로부터 계산된 실험적 위상을 사용하여 구조를 해명하였다. ShelxC 및 ShelxD34를 사용함으로써 총 92개의 셀레늄 부위가 비대칭 단위 내에 위치하였고, 정밀화되어, 샤프35 (위상력 0.79, 성능 지수 (FOM) 0.25)로의 위상 계산에 사용되었다. 패럿36 (도 55; FOM 0.85)을 사용하여 비-결정학적 대칭 (NCS)에 의해 실험적 위상을 밀도를 변형시키고 평균화하였다. 버커니어37로 초기 모델을 구축하고, 반복된 라운드의 휘닉스 리파인38으로의 최대 우도 정밀화 및 수동 검사 및 쿠트39에서의 모델 (재)구축으로 정밀화하였다. 최종 구조는 1.34의 몰브로비티40 점수로 16개의 CsgGC1S 쇄에 속하는 3,700개의 잔기 내의 28,853개의 원자를 함유하였다 (도 47); 잔기의 98%가 라마찬드란 플롯의 선호 영역 내에 놓였다 (99.7%가 허용 영역에 놓였다). 전자 밀도 지도는 가능한 용매 분자에 상응하는 명확한 밀도를 나타내지 않았고, 따라서 물 분자 또는 이온이 내부에 구축되지 않았다. 초기 및 후기 정밀화 단계에서 엄격한 NCS 구속 및 국소적인 NCS 구속을 각각 사용하여, 16중 NCS 평균화가 정밀화 전반에 걸쳐 유지되었다.
CsgG에 대한 회절 데이터가 0.9763 Å 파장에서 단일 결정으로부터 수집되었고, XDS 패키지32 , 33를 사용하여, 비대칭 단위 내의 18개의 CsgG 카피 및 72% 용매 함량을 포함하는, 단위 셀 치수가 a=161.7 Å, b=372.3 Å, c=161.8 Å 및 β=92.9°인 공간 군 C2 내에 인덱싱 및 스케일링되었다. 구조 결정 및 정밀화를 위한 회절 데이터를 역 셀 방향 a*, b* 및 c*를 따라 3.6 Å, 3.7 Å 및 3.8 Å의 해상도 한계로 타원형으로 절단하고, 회절 비등방성 서버41로 비등방성으로 스케일링하였다. CsgGC1S 단량체를 사용한 분자 교체가 성공적이지 않은 것으로 증명되었다. 자가 회전 함수의 분석에서 비대칭 단위 내의 D9 대칭이 밝혀졌다 (제시되지 않음). CsgGC1S 구조를 기초로, C8 올리고머에서 C9 올리고머가 된 후, 프로토머간 각도가 45°에서 40°로 변할 때 입자 원주에서의 프로토머간 호는 대략적으로 동일하게 유지되어 약 4 Å의 계산된 반경 증가를 제공한다고 가정하여, 9량체 검색 모델이 생성되었다. 계산된 9량체를 검색 모델로 사용하여, 2개의 카피를 함유하는 분자 교체 해법이 페이저42로 발견되었다. 밀도-변형 및 및 NCS-평균화 전자 밀도 지도 (패럿36; 도 55)의 검사는 TM1 및 TM2의 수동 구축 및 단백질 코어 내의 인접한 잔기의 리모델링, 뿐만 아니라 CsgGC1S 모델에서 상실되었고 α1 나선을 N-말단 지질 앵커에 연결한 잔기 2-18의 구축을 허용하였다. 각각 초기 및 최종 정밀화 라운드에 대한 부스터-TNT43 및 Refmac5 (참조문헌 44)로 CsgG 모델의 정밀화를 수행하였다. 정밀화 전반에 걸쳐 18중 국소 NCS 구속을 적용하였고, 프로스마트45에 의해 생성된 수소-결합 구속 및 시그마 0.01의 젤리-바디 정밀화를 사용하여 Refmac5를 시행하였다. 최종 구조는 2.79의 몰프로비티 점수로 18개의 CsgG 쇄에 속하는 4,451개의 잔기 내의 34,165개의 원자를 함유하였다 (도 47); 잔기의 93.0%가 라마찬드란 플롯의 선호 영역 내에 놓였다 (99.3%가 허용 영역 내에 놓였다). N-말단 지질 앵커에 상응하는 명확한 전자 밀도가 분간될 수 없었다.
콩고 레드 검정법. 콩고 레드 결합의 분석을 위해, 라이소제니 브로쓰 (LB)에서 37℃에서 성장된 철야 박테리아 배양물을 0.5의 D600에 도달할 때까지 LB 배지에 희석하였다. 5 ㎕ 샘플을 앰피실린 (100 mg/ℓ), 콩고 레드 (100 mg/ℓ) 및 0.1 % (w/v) 이소프로필 β-D-티오갈락토시드 (IPTG)가 보충된 LB 한천 플레이트 상에 스폿팅하였다. 플레이트를 실온 (20 내지 22℃)에서 48시간 동안 인큐베이션하여, 컬리 발현을 유도하였다. 48시에 콜로니 형태 및 염료 결합의 전개를 관찰하였다.
시스테인 접근성 검정법. 부위-지정 돌연변이유발을 사용하여 pMC2에서 시스테인 돌연변이체가 생성되었고, 이. 콜라이 LSR12에서 발현되었다 (참고문헌 7). 철야로 성장된 박테리아 배양물을 1 mM IPTG 및 100 mg/ℓ 앰피실린을 함유하는 LB 한천 플레이트 상에 스폿팅하였다. 플레이트를 실온에서 인큐베이션하고, 48시간 후 세포를 긁어서 1 ml의 PBS에 재현탁시키고, D600을 사용하여 표준화하였다. 초음파처리로 세포를 용해시키고, 4℃에서 20초 동안 3,000 g로 원심분리하여, 피괴되지 않은 세포를 세포 용해물 및 현탁된 막으로부터 제거하였다. 상청액 내의 단백질을 실온에서 1시간 동안 15 mM 메톡시폴리에틸렌 글리콜-말레이미드 (MAL-PEG 5 kDa)로 표지하였다. 100 mM DTT로 반응을 정지시키고, 50.4 Ti 로터에서 4℃에서 20분 동안 40,000 r.p.m. (~100,000g)으로 원심분리하여 전체 막을 펠릿화시켰다. 펠릿을 1% 소듐 라우로일 사르코시네이트로 세정하여 세포질 막을 가용화시키고, 다시 원심분리하였다. 생성된 외막을 1% DDM을 함유하는 PBS를 사용하여 재현탁시키고 가용화하였다. 니켈 비드로의 금속-친화력 풀-다운을 SDS-PAGE 및 항-His 웨스턴 블롯에 사용하였다. 빈 pMC2 벡터가 있는 이. 콜라이 LSR12 세포를 음성 대조군으로서 사용하였다.
ATR- FTIR 분광학. 액체-질소-냉동 머큐리 카드뮴 텔루라이드 검출기 및 골든 게이트 반사율 부속품 (스펙액)이 장착된, 건식 공기로 연속적으로 퍼징되는 이퀴녹스 55 적외선 분광계 (브루커) 상에서 ATR-FTIR 측정을 수행하였다. 내부 반사 요소는 다이아몬드 결정 (2 mm × 2 mm)이었고, 빔 입사각은 45°였다. 각각의 정제된 단백질 샘플 (1 ㎕)을 결정 표면에 도포하고, 기체 질소 흐름 하에 건조시켜 필름을 형성시켰다. 2 cm-1 해상도에서 기록된 각각의 스펙트럼은 개선된 신호-대-노이즈 비를 위한 128개의 누적물의 평균이었다. 모든 스펙트럼을 수증기 기여 차감으로 처리하고, 애퍼디제이션에 의해 4 cm-1의 최종 해상도에서 평활화하고, 아미드 I 밴드 (1,700-1,600 cm- 1)의 면적에 대해 표준화하여 이들의 비교를 허용하였다46.
음성 염색 EM 및 대칭 결정. 음성 염색 EM을 사용하여 CsgG, CsgGC1S 및 CsgE의 용액 내에서의 올리고머화 상태를 모니터링하였다. CsgE, CsgGC1S 및 앰피폴-결합 CsgG를 0.05 mg/ml의 농도로 조정하고, 글로우-방전 탄소-코팅 구리 그리드 (CF-400; 일렉트론 마이크로스코피 사이언시즈)에 적용하였다. 1분 인큐베이션 후, 샘플을 블롯팅하고, 세정하고, 2% 우라닐 아세테이트에서 염색하였다. 120 kV의 전압, ×49,000의 배율 및 800 내지 2,000 nm의 디포커스에서 작동되는 테크나이 T12 바이오트윈 LaB6 현미경에서 영상을 수집하였다. 콘트라스트 전달 함수 (CTF), 위상 플리핑 및 입자 선택을 냉동-EM에 대해 기술된 바와 같이 수행하였다. 막-추출 CsgG의 경우, 18량체 입자 (총 1,780개)를 9량체 및 상면도와 별도로 분석하였다. 정제된 CsgE의 경우, 총 2,452개의 입자를 분석하였다. 3차원 모델을 하기의 CsgG-CsgE 냉동-EM 분석에 대해 기술된 바와 같이 수득하고, 여러 라운드의 다중-기준 정렬 (MRA), 다중-통계 분석 (MSA) 및 앵커 세트 정밀화에 의해 정밀화하였다. 모든 경우에, 표준화 및 중심화 후, 냉동-EM 섹션에 기술된 바와 같이 IMAGIC-4D를 사용하여 영상을 분류하였다. 특징적인 도에 상응하는 최상 클래스를 각각의 입자 세트에 대해 선택하였다. 클래스 당 약 20개의 영상으로 최상 클래스 평균을 사용하여 CsgG 상면도의 대칭 결정을 수행하였다. IMAGIC를 사용하여 회전식 자기상관 함수를 계산하고, 플롯팅하였다.
CsgG - CsgE 복합체 형성. CsgG-CsgE 복합체 형성을 위해, 120 ㎕의 CsgG (0.5% C8E4, 4 mM LDAO, 25 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM DTT 내의 24 mg/ml 단백질)를 300 ㎕의 세제-탈안정화 리포솜 (1 mg/ml, 1,2-디미리스토일-sn-글리세로-3-포스포콜린 (DMPC) 및 0.4% LDAO)에 첨가하고, 얼음 상에서 5분 동안 인큐베이션한 후, 90 ml의 A8-35 앰피폴 (100 mg/ml 모액)을 첨가함으로써, 정제된 CsgG 내의 가용화 세제를 앰피폴 A8-35 (애너트레이스)로 교환하였다. 추가로 15분 동안 얼음 상에서 인큐베이션한 후, 샘플을 수퍼로스 6 10/300 GL (GE 헬스케어) 칼럼 상에 로딩하고, 200 mM NaCl, 2.5% 크실리톨, 25 mM 트리스-HCl pH 8, 0.2 mM DTT에서 겔 여과를 수행하였다. 200 mM NaCl, 2.5% 크실리톨, 25 mM 트리스-HCl pH 8, 0.2 mM DTT 내의 동일한 부피의 정제된 단량체 CsgE를 앰피폴-가용화 CsgG에 CsgE 및 CsgG에 대해 각각 15 및 5 μM의 최종 단백질 농도로 첨가하고, 샘플을 125 V로 18℃에서 0.5× TBE 완충제 내의 4.5% 네이티브 PAGE 상에 러닝시켰다. 제2의 변성 차원을 위해, CsgG-CsgE 복합체에 상응하는 밴드를 동일한 겔 상에서 병렬로 러닝된 미염색 레인으로부터 절단하고, 5분 동안 래믈리 완충제 (60 mM 트리스-HCl pH 6.8, 2% SDS, 10% 글리세롤, 5% 2-메르캅토에탄올, 0.01 % 브로모페놀 블루)에서 비등시키고, 4-20% SDS-PAGE 상에 러닝시켰다. 정제된 CsgE 및 CsgG를 대조군 샘플로서 복합체와 나란히 러닝시켰다. 겔을 시각적 검사를 위해 인스턴트블루 쿠마시로, 또는 SDS-PAGE 상에서의 CsgE 및 CsgG 밴드의 형광 검출 (타이푼 FLA 9000)에 의한 CsgG-CsgE 복합체의 화학량론 평가를 위해 사이프로 오렌지로 염색하였고, 0.97의 CsgG/CsgE 비가 산출되었다.
CsgG - CsgE 냉동- EM. 냉동-전자 현미경검사를 사용하여 C9 CsgG-CsgE 복합체의 용액 내에서의 구조를 결정하였다. 상기 기술된 바와 같이 제조된 CsgG-CsgE 복합체를 HisTrap FF (GE 헬스케어)에 결합시키고 이로부터 용출시켜, 결합되지 않은 CsgG를 제거하였고, 용출 시 복합체를 20 mA에서 30초 동안 글로우 방전된 퀀티포일 R2/2 탄소 코팅 그리드 (퀀티포일 마이크로 툴스 게엠베하)에 즉각적으로 적용하였다. 샘플을 자동화 시스템 (라이카)를 사용하여 액체 질소에서 플런지-냉동시키고, 200 kV의 전압, 저용량 조건 하에서의 ×50,000의 공칭 배율, 및 1.4-3 ㎛의 디포커스 범위에서 작동되는 FEI F20 현미경 하에 관찰하였다. 팔콘 II 검출기 상에서 영상 프레임을 기록하였다. 검체 수준에서의 픽셀 크기는 픽셀 당 1.9 Å이었다. CTFFIND3 (참조문헌 47)을 사용하여 CTF 파라미터를 평가하였고, 스파이더48에서 위상 플리핑을 행하였다. 복서 (EMAN2; 참조문헌 49)를 사용하여 CTF-수정 마이크로그래프로부터 입자가 자동적으로 선택되었다. 비점 수차가 10%를 초과하는 영상은 폐기되었다. 75개의 마이크로그래프로부터 총 1,221개의 입자가 선택되었고, 128-픽셀 × 128-픽셀 상자 내로 윈도우화되었다. 영상을 동일한 평균 및 표준 편차에 대해 표준화하고, ~200 Å의 저해상도 차단값으로 하이-패스 필터링하였다. 이들을 중심화한 후, 제1 라운드의 MSA에 적용하였다. IMAGIC-4D (이미지 사이언스 소프트웨어)의 무기준 분류를 사용하여 초기 기준 세트를 수득하였다. C9 원통형 입자의 특징적인 측면도에 상응하는 최상 클래스가 MRA에 대한 기준으로 사용되었다. CsgG-CsgE 복합체에 대해, 각도 재구성 (이미지 사이언스 소프트웨어)에 의해 배향이 결정된 복합체의 1,221개의 입자를 포괄하는 최상의 125개의 특징적인 도 (양호한 콘트라스트 및 윤곽이 뚜렷한 특색을 지님)로부터 최초의 3차원 모델이 계산되었다. 반복된 라운드의 MRA, MSA 및 앵커 세트 정밀화에 의해 3차원 지도를 정밀화하였다. 0.5 기준 수준에 따라 푸리에 쉘 상관관계 (FSC)에 의해 해상도가 24 Å인 것으로 추정되었다 (도 52). 지도 및 도면의 가시화를 UCSF 키메라50에서 수행하였다.
담즙산 염 독성 검정법. 담즙산 염을 함유하는 한천 플레이트 상에서의 성장 감소에 의해 외막 투과성을 연구하였다. pLR42 (참조문헌 16) 및 pMC2 (참조문헌 14) 양쪽 모두 (또는 유래된 나선 2 돌연변이체)를 보유하는 이. 콜라이 LSR12 (참조문헌 7) 세포의 10배 단계 희석물 (5 ㎕)을 0.2% (w/v) L-아라비노스와 함께 또는 L-아라비노스 없이 100 ㎍/ℓ 앰피실린, 25 ㎍/ℓ 클로람페니콜, 1 mM IPTG를 함유하는 맥콘키 한천 플레이트 상에 스폿팅하였다. 37℃에서 철야로 인큐베이션한 후, 콜로니 성장을 검사하였다.
단일-채널 전류 기록. 나니온으로부터의 오르비트 16 키트로의 병렬식 고해상도 전기 기록을 사용하여 단일-채널 전류 기록을 수행하였다. 간략하게, 1,2-디피타노일-sn-글리세로-3-포스포콜린 (아반티 폴라 리피즈)의 수평 이중층이 16-채널 다중전극 공동 어레이 (MECA) 칩 (아이오네라)의 미세공동 (피코리터 이하의 부피)에서 형성되었다51. 이중층 위 및 아래의 시스 및 트랜스 공동 양쪽 모두 1.0 M KCl, 25 mM 트리스-HCl pH 8.0을 함유하였다. 채널을 막 내로 삽입하기 위해, 25 mM 트리스-HCl pH 8.0, 500 mM NaCl, 1 mM DTT, 0.5% C8E4, 5 mM LDAO에 용해된 CsgG를 90-300 nM의 최종 농도로 시스 구획에 첨가하였다. CsgG 채널과 CsgE의 상호작용을 테스트하기 위해, 25 mM 트리스-HCl pH 8.0, 150 mM NaCl에 용해된 후자 단백질의 용액을 0.1, 1, 10 및 100 nM의 최종 농도로 시스 구획에 첨가하였다. 3 kHz의 로우-패스 필터링 주파수 및 10 kHz의 샘플링 주파수로 테셀라 트리톤 16-채널 증폭기를 사용하여 +50 mV 및 -50 mV (시스 측면 기준)의 유지 전위에서 막횡단 전류를 기록하였다. p클램프 스위트 (몰레큘러 디바이시즈)의 클램프피트를 사용하여 전류 트레이스를 분석하였다. 오리진 8.6 (마이크로칼))52을 사용하여 플롯이 생성되었다.
측정된 전류를 3개의 분절 (막횡단 섹션, 원형질막 주위공간 전정, 및 둘을 연결하는 내부 채널 협착부)로 구성되는 것으로 모델링된 CsgG X-선 구조의 포어 치수를 기초로 계산된 것과 비교하였다. 처음 2개의 분절은 원뿔형 형상인 것으로 모델링되었고, 협착부는 원통형으로서 나타났다. 상응하는 저항 R1, R2 및 R3 각각이 하기와 같이 계산되었다:
R1 = L1/(πD1d1κ)
R2 = L2/(πD2d2κ)
R3 = L3/(πD1d2κ)
[식 중, L1, L2 및 L3은 각각 3.5, 4.0 및 2.0 nm로 측정된 분절의 축 길이이고, D1, d1, D2 및 d2는 각각 4.0, 0.8, 3.4 및 0.8 nm로 측정된 분절 1 및 2의 최대 및 최소 직경이다]. 전도도 κ의 값은 10.6 S/m이다. R1, R2 및 R3 및 전압 V = 50 mV를 하기 식에 삽입함으로써 전류가 계산되었다:
I = V/(R1 + R2 + R3)
접근 저항이 예상 전류를 유의하게 변경시키는 것으로 발견되지 않았다.
참고문헌