일반적인 기술 및 정의
달리 구체적으로 정의되지 않는 한, 본원에서 사용되는 모든 기술 및 과학 용어는 당업자에 의해 일반적으로 이해되는 의미와 동일한 의미를 갖는 것으로 받아들여야 한다(예를 들어, 효소, 발효, 접종).
용어 "및/또는", 예를 들어, "X 및/또는 Y"는 "X 및 Y" 또는 "X 또는 Y" 중 어느 하나를 의미하는 것으로 이해되어야 하고, 둘 모두의 의미에 대한 또는 어느 하나의 의미에 대한 명백한 지지를 제공하는 것으로 받아들여야 한다.
본 명세서 전반에 걸쳐, 단어 "포함하다(복수 주어)", 또는 변형어, 예컨대, "포함하다(단수 주어)" 또는 "포함하는"은 명시된 요소, 정수 또는 단계, 또는 요소들, 정수들 또는 단계들의 군의 포함이되, 임의의 다른 요소, 정수 또는 단계, 또는 요소들, 정수들 또는 단계들의 군의 배제가 아님을 암시하는 것으로 이해될 것이다.
본원에서 사용되는 용어 "약"은, 달리 반대로 명시되지 않는 한, 지정된 값의 +/- 10%, 더욱 바람직하게는 +/- 5%, 더욱 더 바람직하게는 +/- 1%를 지칭한다.
"대립형질"은 세포, 개별 식물 내 또는 집단 내 유전자 서열(유전자와 같은)의 하나의 특이적 형태를 지칭하며, 이러한 특이적 형태는 유전자의 서열 내 적어도 하나, 및 종종 하나보다 많은 변이체 부위의 서열에서 동일한 유전자의 다른 형태와 상이하다. 상이한 대립형질 간에 상이한 이러한 변이체 부위에서의 서열은 "변이", 다형성", 또는 "돌연변이"로 칭해진다.
브라시카케아이
당업자는 본원에 기재된 바와 같은 방법이 글루코시놀레이트/들을 포함하는 임의의 브라시카케아이 재료로부터 이소티오시아네이트 함유 프로덕트를 생산하기에 적합하다는 것을 인지할 것이다. 본원에서 사용되는 "브라시카케아이"는 겨자과, 십자화과 또는 양배추과로 흔히 지칭되는 패밀리 브라시카케아이의 구성원을 지칭한다. 당업자는 재료가 하나보다 많은 브라시카케아이로부터 유래될 수 있다는 것을 인지할 것이다.
일 구현예에서, 브라시카케아이는 브라시카(Brassica) 또는 카르다마인(Cardamine) 속으로부터 선택된다. 일 구현예에서, 브라시카는 브라시카 발레아리카(Brassica balearica), 브라시카 카리나타(Brassica carinata), 브라시카 에론가테(Brassica elongate), 브라시카 프루티쿨로사(Brassica fruticulosa), 브라시카 힐라리오니스(Brassica hilarionis), 브라시카 준키아(Brassica juncea), 브라시카 나푸스(Brassica napus), 브라시카 나리노사(Brassica narinosa), 브라시카 니그라(Brassica nigra), 브라시카 올레라케아(Brassica oleracea), 브라시카 페르비리디스(Brassica perviridis), 브라시카 라파(Brassica rapa), 브라시카 루페스트리스(Brassica rupestris), 브라시카 세프티케프스(Brassica septiceps), 및 브라시카 토우르네포르티(Brassica tournefortii)로부터 선택된다.
일 구현예에서, 브라시카는 브라시카 올레라케아이다.
일 구현예에서, 브라시카는 브라시카 올레라케아 변종 올레라케아(야생 양배추), 브라시카 올레라케아 변종 카피테이트(capitate)(양배추), 브라시카 라파 아종 키넨시스(chinensis)(청경채), 브라시카 라파 아종 페키넨시스(pekinensis)(배추), 브라시카 나포브라시카(Brassica napobrassica)(루타바가), 브라시카 라파 변종 라파(rapa)(터닙), 브라시카 올레라케아 변종 알보그라브라(alboglabra)(카이-란), 브라시카 올레라케아 변종 비리디스(viridis)(콜라드 그린), 브라시카 올레라케아 변종 론가타(longata)(저지 양배추), 브라시카 올레라케아 변종 아세팔라(acephala)(엽목단), 브라시카 올레라케아 변종 사베리카(sabellica)(케일), 브라시카 올레라케아 변종 팔미포리아(palmifolia)(라키나토 케일), 브라시카 올레라케아 변종 라모세(ramose)(페르페추얼 케일), 브라시카 올레라케아 변종 메둘로사(medullosa)(매로우 양배추), 브라시카 올레라케아 변종 코스타타(costata)(트론추다 케일), 브라시카 올레라케아 변종 겜미페라(gemmifera)(브루셀 싹양배추), 브라시카 올레라케아 변종 곤질로데스(gongylodes)(콜라비), 브라시카 올레라케아 변종 이탈리카(italica)(브로콜리), 브라시카 올레라케아 변종 보트라이티스(botrytis)(콜리플라워, 로마네스코 브로콜리, 브로콜리 디 토르볼레), 브라시카 올레라케아 변종 모트라이티스 x 이탈리카(botrytis x italica)(브로코플라워), 및 브라시카 올레라케아 변종 이탈리카 x 알보글라브라(italica Х alboglabra)(브로콜리니)로부터 선택된다.
일 구현예에서, 브라시카는 브라시카 올레라케아, 변종 이탈리카(브로콜리)이다.
일 구현예에서, 브라시카케아이는 카르다마인 히르수타(Cardamine hirsuta)(황새냉이), 이베리스 셈페르비렌스(Iberis sempervirens)(서양말냉이), 시나피스 아르벤시스(Sinapis arvensis)(들갓), 아르모라시아루스티카나(Armoracia rusticana)(호스래디시), 프린글레아 안티스코르부티카(Pringlea antiscorbutica)(케르겔렌 양배추), 탈라스피 아르벤세(Thlaspi arvense)(말냉이), 람파누스 라파니스트룸(Raphanus raphanistrum) 아종 사티부스(sativus)(래디시), 에루카 사티바(Eruca sativa)(로켓), 아나스타티카 히에로춘티카(Anastatica hierochuntica)(안산수), 크람베 마리티마(Crambe maritima)(갯배추), 카키레 마리티마(Cakile maritima)(씨로켓), 카프셀라 부르사-파스토리스(Capsella bursa-pastoris)(냉이), 스위트 알리숨, 아라비도프시스 탈리아나(Arabidopsis thaliana)(애기장대), 나스투르티움 오피키날레(Nasturtium officinale)(물냉이), 시나피스 알바(Sinapis alba)(백겨자), 에로필라 베르나(Erophila verna)(꽃다지), 라파누스 라파니스트룸(야생 래디시), 이사티스 틴크토리아(Isatis tinctoria)(대청), 및 나스투르티움 마이크로필룸(Nasturtium microphyllum)(개갓냉이)로부터 선택된다.
일 구현예에서, 브라시카케아이는 고수준의 하나 이상의 글루코시놀레이트/들을 갖는다. 일 구현예에서, 브라시카케아이는 고수준의 하나 이상의 글루코시놀레이트/들을 갖도록 선별적으로 사육되었다. 일 구현예에서, "고수준"의 글루코시놀레이트는 해당하는 브라시카케아이에서 문헌[Verkerk et al. (2009)]의 표 2에 나타나 있는 글루코시놀레이트보다 높은 수준의 글루코시놀레이트를 포함할 수 있다. 일 구현예에서, 고수준의 글루코시놀레이트는 3400 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 4000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 5000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 8000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 10,000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 12,000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 15,000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 18,000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 20,000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 25,000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 고수준의 글루코시놀레이트는 30,000 μmol/kg 건조 중량보다 높은 수준의 글루코시놀레이트이다. 일 구현예에서, 브라시카케아이는 유전자 변형되거나 고수준의 하나 이상의 글루코시놀레이트/들을 갖도록 생물학적 또는 비생물학적 스트레스에 주어졌었다. 당업자는 브라시카케아이가 당업자에게 공지된 임의의 방법에 의해 변형될 수 있다는 것을 인지할 것이다.
일 구현예에서, 글루코시놀레이트는 글루코라파닌 (4-메틸설피닐부틸)이다. 일 구현예에서, 글루코시놀레이트는 글루코브라시신 (3-인돌릴메틸)이다.
본원에서 사용되는 "브라시카케아이 재료"은, 잎, 줄기, 꽃, 꽃송이, 종자 및 뿌리 또는 이들의 혼합을 포함하지만, 이로 제한되지 않는, 글루코시놀레이트를 포함하는 브라시카케아이의 임의의 부분을 지칭한다.
당업자는 본원에 기재된 바와 같은 방법이, 이로 제한되지는 않지만, 예를 들어, 적어도 30 kg, 또는 적어도 50 kg, 또는 적어도 80 kg, 또는 적어도 100 kg, 또는 적어도 1,000 kg, 또는 적어도 2,000 kg, 또는 적어도 5,000 kg, 또는 적어도 8,000 kg, 또는 적어도 10,000 kg, 또는 적어도 15,000 kg, 또는 적어도 20,000 kg의 상이한 부피의 브라시카케아이 재료와 사용하기에 적합하다는 것을 인지할 것이다.
일 구현예에서, 브라시카케아이 재료는 세척되었다. 본원에서 사용되는 "세척"은 육안의 토양 및 오염물을 제거하는 것이다. 일 구현예에서, 브라시카케아이 재료는 살균되었다. 본원에서 사용되는 "살균된"은 브라시카케아이 재료에 대한 병원체의 감소를 지칭한다.
일 구현예에서, 브라시카케아이는 다른 식물 재료와 혼합된다. 일 구현예에서, 다른 식물 재료는 채소 또는 과일 재료이다. 일 구현예에서, 채소는 당근 또는 비트이다.
글루코시놀레이트
본원에서 사용되는 "글루코시놀레이트"는 문헌[Halkier et al. (2006)]에 기재된 바와 같이 아미노산으로부터 유래된 다양한 R 기와 더불어 황 원자를 통해 (Z)-N-하이드록시미노설페이트 에스테르에 연결된 β-D-글루코피라노스 잔기로 이루어진 화학 구조를 공유하는 적어도 브라시카케아이 패밀리에서 발견되는 이차 대사물을 지칭한다. 글루코시놀레이트의 예는 문헌[Halkier et al. (2006)] 및 문헌[Agerbirk et al. (2012)]에 제공되어 있다. 글루코시놀레이트의 가수분해로 이소티오시아네이트, 니트릴, 에피티오니트릴, 티오시아네이트 및 옥사졸리딘-2-티온이 생산될 수 있다(도 1a). 다수의 글루코시놀레이트가 해충 및 질병에 대항하는 식물 방어 메카니즘에 관여한다.
글루코시놀레이트는 브라시카케아이에서 저장소에 저장된다. 본원에서 사용되는 "저장소"는 글루코시놀레이트가 존재하고 미로시나제가 존재하지 않는 브라시카케아이 내 장소이다.
"티오글루코시다제", "시니그라제", 또는 "시니그리나제"로도 지칭되는 본원에서 사용되는 "미로시나제"는 티오-연결 글루코스를 절단할 수 있는 식물 방어 메카니즘에 관여하는 효소의 패밀리(EC 3.2.1.147)를 지칭한다. 미로시나제는 글루코시놀레이트의 가수분해를 촉매작용하여 이소티오시아네이트의 생산을 야기한다. 미로시나제는 미로신 세포라 불리는 특정 이형세포의 액포에서 미로신 과립으로 종종 저장되지만, 또한 단백질체 또는 액포에서, 그리고 멤브레인에 결합하는 경향이 있는 세포액 효소로서 보고되었다. 따라서, 일 구현예에서, 미로시나제는 브라시카케아이에서 미로신 세포에 저장된다.
일 구현예에서, 본원에 기재된 바와 같은 전처리는 글루코시놀레이트에 대한 미로시나제의 접근을 개선한다. 본원에서 사용되는 "접근을 개선하다" 또는 "접근이 개선된다"는 이소티오시아네이트의 생산을 가능하게 하는 미로시나제 효소에 대한 글루코시놀레이트의 이용 가능성을 증가시키는 것을 지칭한다. 일 구현예에서, 접근은 글루코시놀레이트 저장소로부터 글루코시놀레이트의 방출에 의해 개선된다. 일 구현예에서, 글루코시놀레이트 저장소는 기계적으로 파열되거나(즉, 침연에 의해) 효소적으로 분해된다. 일 구현예에서, 글루코시놀레이트는 하나 이상의 다당류 분해 효소, 예를 들어, 셀룰라제, 헤미셀룰라제, 펙티나제 및/또는 글리코시다제의 활성에 의해 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 접근은 미로시나제가 글루코시놀레이트 저장소에 들어갈 수 있게 함으로써 개선된다. 일 구현예에서, 접근은 미로신 세포로부터 미로시나제의 방출에 의해 개선된다. 일 구현예에서, 글루코시놀레이트의 약 10% 내지 약 90%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 20% 내지 약 80%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 30% 내지 약 70%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 40% 내지 약 60%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 45% 내지 약 55%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 10%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 20%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 30%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 40%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 50%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 60%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 70%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 80%가 글루코시놀레이트 저장소로부터 방출된다. 일 구현예에서, 글루코시놀레이트의 약 90%가 글루코시놀레이트 저장소로부터 방출된다.
일 구현예에서, 브라시카케아이 재료는 지방족, 인돌 또는 방향족 글루코시놀레이트로부터 선택된 하나 이상의 글루코시놀레이트/들을 포함한다.
일 구현예에서, 지방족 글루코시놀레이트는 글루코라파닌 (4-메틸설피닐부틸 또는 글루코라파닌), 시니그린 (2-프로페닐), 글루코나핀 (3-부테닐), 글루코브라시카나핀 (4-펜테닐), 프로고이트린 (2(R)-2-하이드록시-3-부테닐, 에피프로고이트린 (2(S)-2-하이드록시-3-부테닐), 글루코나폴레이페린 (2-하이드록시-4-펜테닐), 글루코이베르비린 (3-메틸티오프로필, 글루코에루신 (4-메틸티오부틸), 데하이드로에루신 (4-메틸티오-3-부테닐, 글루코이베린 (3-메틸설피닐프로필), 글루코라페닌 (4-메틸설피닐-3-부테닐), 글루코알리신 (5-메틸설피닐펜테닐), 및 글루코에리솔린 (3-메틸설포닐부틸, 4-머캅토부틸) 중 하나 이상으로부터 선택된다..
일 구현예에서, 인돌 글루코시놀레이트는 글루코브라시신 (3-인돌릴메틸), 4-하이드록시글루코브라시신 (4-하이드록시-3-인돌릴메틸), 4-메톡시글루코브라시신 (4-메톡시-3-인돌릴메틸), 및 네오글루코브라시신 (1-메톡시-3-인돌릴메틸) 중 하나 이상으로부터 선택된다.
일 구현예에서, 인돌 글루코시놀레이트는 글루코트로페올린 (벤질) 및 글루코나스투르티인 (2-페닐에틸) 중 하나 이상으로부터 선택된다.
일 구현예에서, 브라시카케아이 재료는 벤질글루코시놀레이트, 알릴글루코시놀레이트 및 4-메틸설피닐부틸로부터 선택된 하나 이상의 글루코시놀레이트/들을 포함한다. 일 구현예에서, 글루코시놀레이트는 글루코라파닌 (4-메틸설피닐부틸)이다. 일 구현예에서, 글루코시놀레이트는 글루코브라시신 (3-인돌릴메틸)이다.
일 구현예에서, 본원에 기재된 바와 같은 전처리는 전처리 전 브라시카케아이 재료의 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 증가시킨다.
본원에서 사용되는 "추출 가능한 글루코시놀레이트 함량"은 이소티오시아네이트로의 전환을 위한 브라시카케아이 재료에서 접근 가능한 글루코시놀레이트의 수준을 지칭한다. 니트릴 및 다른 화합물로의 전환을 배제하고, 1 몰의 글루코시놀레이트로부터 이소티오시아네이트의 예상 최대 수율은 1 몰의 이소티오시아네이트이다(1 몰의 글루코시놀레이트가 최대로 1 몰의 이소티오시아네이트, 1 몰의 글루코스 및 1 몰의 설페이트 이온으로 전환될 수 있다). 따라서, 일례에서, 상업적 브로콜리 재배종의 추출 가능한 글루코라파닌 함량은 3400 μmol의 글루코라파닌/kg dw이고, 상업적 브로콜리 재배종으로부터 설포라판의 예상 최대 수율은 3400 μmol 설포라판 /kg dw이다.
이소티오시아네이트
본원에서 사용되는 "이소티오시아네이트"는 글루코시놀레이트 및 이의 생체활성 유도체에 대한 미로시나에 활성의 산물인 일반 구조식 R-N=C=S를 갖는 황 함유 식물화학물질을 지칭한다. 일 구현예에서, 이소티오시아네이트는 설포라판 (1-이소티오시아네이토-4-메틸설피닐부탄)이다. 일 구현예에서, 이소티오시아네이트는 알릴 이소티오시아네이트 (3-이소티오시아네이토-1-프로펜)이다. 일 구현예에서, 이소티오시아네이트는 벤질 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트는 펜에틸 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트는 3-부테닐 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트는 5-비닐-1,3-옥사졸리딘-2-티온이다. 일 구현예에서, 이소티오시아네이트는 3-(메틸티오)프로필 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트는 3-(메틸설피닐)-프로필 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트는 4-(메틸티오)-부틸 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트는 1-메톡시인돌-3-카비놀 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트는 2-페닐에틸 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트는 이베린이다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 하나 이상의 이소티오시아네이트 생체활성 유도체/들 또는 이의 올리고머를 추가로 포함한다. 일 구현예에서, 이소티오시아네이트 생체활성 유도체는 본원에 기재된 바와 같은 임의의 이소티오시아네이트의 유도체이다. 일 구현예에서, 이소티오시아네이트 생체활성 유도체는 설포라판의 유도체이다. 일 구현예에서, 이소티오시아네이트 생체활성 유도체는 이베린이다. 일 구현예에서, 이소티오시아네이트 생체활성 유도체는 알릴 이소티오시아네이트이다. 일 구현예에서, 이소티오시아네이트 생체활성 유도체는 인돌-3-카리비놀이다. 일 구현예에서, 이소티오시아네이트 생체활성 유도체는 메톡시-인돌-3-카비놀이다. 일 구현예에서, 이소티오시아네이트 생체활성 유도체는 아스코르비겐이다. 일 구현예에서, 이소티오시아네이트 생체활성 유도체는 네오아스코르비겐이다.
전처리
본원에서 사용되는 "전처리" 또는 "전처리하는"은 글루코시놀레이트 저장소로부터 글루코시놀레이트를 방출시키거나 이의 방출을 돕고/돕거나 미로시나제가 브라시카케아이 재료에서 글루코시놀레이트 저장소에 들어가는 것을 가능하게 한다. 일 구현예에서, 전처리는 미로시나제에 대한 글루코시놀레이트의 노출을 증가시켜 미로시나제로 하여금 글루코시놀레이트를 이소티오시아네이트로 전환시키게 할 수 있다.
일 구현예에서, 전처리는 내인성 미로시나제 활성을 유지하면서 에피티오스페시피어 단백질(ESP)을 감소시킨다. 본원에서 사용되는 "에피티오스페시피어 단백질" 또는 "ESP"는 미로시나제 활성을 이소티오시아네이트 생산이 아니라 니트릴의 생산 쪽으로 유도하는 단백질을 지칭한다. ESP 생산(mRNA 또는 단백질) 또는 활성을 감소시키거나 억제하는 것은 이소티오시아네이트의 생산을 증가시킬 수 있다.
본원에서 사용되는 "에피티오페시피어 단백질 감소시키다"는 ESP의 단백질 생산 또는 활성을 감소시키는 것을 지칭한다. 일 구현예에서, ESP를 감소시키는 것은 고온에서 ESP를 불활성화시키는(예를 들어, 변성시키는) 것을 포함한다. 일 구현예에서, ESP는 약 50℃ 내지 약 80℃의 온도에서 변성된다.
본원에서 사용되는 "내인성 미로시나제 활성을 유지하는"은 비처리된 대조에 비해 미로시나제 활성을 유의하게 감소시키는 것을 의미하지 않는다. 일 구현예에서, 내인성 미로시나제 활성은 약 5% 이상 감소되지 않는다. 일 구현예에서, 내인성 미로시나제 활성은 약 10% 이상 감소되지 않는다. 일 구현예에서, 내인성 미로시나제 활성은 약 15% 이상 감소되지 않는다. 일 구현예에서, 내인성 미로시나제 활성은 약 20% 이상 감소되지 않는다. 일 구현예에서, 내인성 미로시나제 활성은 약 30% 이상 감소되지 않는다. 일 구현예에서, 내인성 미로시나제 활성은 약 40% 이상 감소되지 않는다. 일 구현예에서, 내인성 미로시나제 활성은 약 50% 이상 감소되지 않는다.
일 구현예에서, 전처리는 i) 가열; ii) 침연; iii) 마이크로웨이빙; iv) 고주파 음파(초음파)에 대한 노출, 또는 v) 펄스 전기장 가공 중 하나 이상을 포함하고, 여기서 브라시카케아이 재료의 온도는 전처리 동안 약 75℃를 초과하지 않는다.
일 구현예에서, 브라시카케아이 재료는 연료 기반 가열 시스템, 전기 기반 가열 시스템(즉, 오븐 또는 옴 가열), 무선 주파수 가열, 고압 열 가공 또는 스팀 기반 가열 시스템(스팀의 간접 또는 직접 적용)에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 밀봉된 패키지에서(예를 들어, 레토르트 파우치에서) 가열된다. 일 구현예에서, 브라시카케아이 재료는 오븐, 워터 배쓰, 바이오리엑터, 스토브, 워터 블랜처(water blancher), 또는 스팀 블랜처(steam blancher)에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 고압 열 가열을 통해 가열된다. 일 구현예에서, 브라시카케아이 재료는 옴 가열을 통해 가열된다. 일 구현예에서, 브라시카케아이 재료는 무선 주파수 가열을 통해 가열된다. 일 구현예에서, 브라시카케아이 재료는 물에서 블랜칭된다. 일 구현예에서, 브라시카케아이 재료는 고압 열 가공을 통해 가열된다. 일 구현예에서, 브라시카케아이 재료는 고압 열 가공을 위해 밀봉된 패키지에 넣어진다.
일 구현예에서, 전처리는 브라시카케아이 재료를 약 50℃ 내지 약 70℃까지 가열하는 것을 포함한다. 일 구현예에서, 전처리는 브라시카케아이 재료를 약 50℃ 내지 약 65℃까지 가열하는 것을 포함한다. 일 구현예에서, 전처리는 브라시카케아이 재료를 약 50℃ 내지 약 60℃까지 가열하는 것을 포함한다. 일 구현예에서, 가열은 브라시카케아이 재료를 약 55℃ 내지 약 70℃까지 가열하는 것을 포함한다. 일 구현예에서, 가열은 브라시카케아이 재료를 약 60℃ 내지 약 70℃까지 가열하는 것을 포함한다. 일 구현예에서, 가열은 브라시카케아이 재료를 약 65℃ 내지 약 70℃까지 가열하는 것을 포함한다. 일 구현예에서, 브라시카케아이 재료는 약 30 초 동안 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 1 분 동안 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 2 분 동안 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 3 분 동안 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 4 분 동안 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 5 분 동안 가열된다.
일 구현예에서, 브라시카케아이 재료는 약 60℃에서 약 1 분 동안 밀봉된 패키지에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 60℃에서 약 2 분 동안 밀봉된 패키지에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 60℃에서 약 3 분 동안 밀봉된 패키지에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 65℃에서 약 4 분 동안 밀봉된 패키지에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 65℃에서 약 1 분 동안 밀봉된 패키지에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 65℃에서 약 2 분 동안 밀봉된 패키지에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 65℃에서 약 3 분 동안 밀봉된 패키지에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 65℃에서 약 4 분 동안 밀봉된 패키지에서 가열된다.
일 구현예에서, 브라시카케아이 재료는 약 60℃에서 약 1 분 동안 물에서 가열된다. 일 구현예에서, 브라시카케아이 재료는 약 60℃에서 약 2 분 동안 물에서 가열된다.
일 구현예에서, 가열은 브라시카케아이 재료를 스티밍하는 것을 포함한다. 일 구현예에서, 전처리는 브라시카케아이 재료를 스티밍하는 것을 포함한다. 일 구현예에서, 브라시카케아이 재료는 약 50℃ 내지 약 70℃의 온도로 스티밍된다. 일 구현예에서, 브라시카케아이 재료는 약 60℃ 내지 약 70℃의 온도로 스티밍된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 30 초 동안 스티밍된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 1 분 동안 스티밍된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 2 분 동안 스티밍된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 3 분 동안 스티밍된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 4 분 동안 스티밍된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 5 분 동안 스티밍된다.
일 구현예에서, 전처리는 브라시카케아이 재료를 침연시키는 것을 포함한다. 본원에서 사용되는 "침연시키는", "침연된" 또는 "침연시키다"는 브라시카케아이 재료를 더 작은 조각으로 쪼개는 것을 지칭한다. 일 구현예에서, 침연은 미로시나제의 이의 기질 글루코시놀레이트에 대한 접근을 가능하게 하도록 브라시카케아이 재료의 적어도 약 30% 내지 약 90%의 세포를 탈구획화시키는 것을 포함한다. 일 구현예에서, 침연은 브라시카케아이 재료의 적어도 약 40% 내지 약 90%의 세포를 탈구획화시키는 것을 포함한다. 일 구현예에서, 침연은 브라시카케아이 재료의 적어도 약 50% 내지 약 90%의 세포를 탈구획화시키는 것을 포함한다. 일 구현예에서, 침연은 브라시카케아이 재료의 적어도 약 60% 내지 약 90%의 세포를 탈구획화시키는 것을 포함한다. 일 구현예에서, 침연은 브라시카케아이 재료의 적어도 약 70% 내지 약 90%의 세포를 탈구획화시키는 것을 포함한다. 당업자는 세포를 탈구획화시키는 것이 세포벽을 파괴하여 개방하고, 세포 내 소기관의 구획화를 방해하는 것을 포함한다는 것을 인지할 것이다.
일 구현예에서, 브라시카케아이 재료는 블렌더, 그라인더 또는 분쇄기로 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 80%가 약 2 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 80%가 약 1 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 80%가 약 0.5 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 80%가 약 0.25 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 80%가 약 0.1 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 80%가 약 0.05 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 80%가 약 0.025 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 80%가 약 0.01 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 50% 내지 약 90%가 약 2 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 60% 내지 약 80%가 약 2 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 50% 내지 약 90%가 약 1 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 브라시카케아이 재료의 적어도 약 60% 내지 약 80%가 약 1 mm 이하의 크기가 되도록 침연된다. 일 구현예에서, 브라시카케아이 재료는 침연 동안 약 50℃ 내지 약 70℃의 온도로 가열된다. 일 구현예에서, 브라시카케아이 재료는 침연 동안 약 55℃ 내지 약 70℃의 온도로 가열된다. 일 구현예에서, 브라시카케아이 재료는 침연 동안 약 60℃ 내지 약 70℃의 온도로 가열된다. 일 구현예에서, 브라시카케아이 재료는 침연 동안 약 65℃ 내지 약 70℃의 온도로 가열된다.
일 구현예에서, 전처리는 브라시카케아이 재료를 가열하고 침연시키는 것을 포함한다. 일 구현예에서, 전처리로 퓨레가 생성된다. 본원에서 사용되는 "퓨레"는 크림형 페이스트 또는 액체의 컨시스턴시(consistency)로 배합된 브라시카케아이 재료를 지칭한다.
당업자는 "마이크로웨이브" 또는 "마이크로웨이빙"이 브라시카케아이 재료와 같은 물질을 물질에 마이크로파 방사선을 통과시킴으로써 가열한다는 것을 인지할 것이다. 일 구현예에서, 전처리는 브라시카케아이 재료를 마이크로웨이빙하는 것을 포함한다. 일 구현예에서, 브라시카케아이 재료는 소비자용 마이크로웨이브 또는 산업용 마이크로웨이브에서 전처리된다. 일 구현예에서, 산업용 마이크로웨이브는 연속 마이크로웨이브 시스템, 이로 제한되지는 않지만, 예를 들어, MIP 11 Industrial Microwave Continuous Cooking Over(Ferrite Microwave Technologies)를 포함한다. 일 구현예에서, 전처리는 브라시카케아이 재료를 마이크로웨이빙하는 것을 포함한다. 일 구현예에서, 브라시카케아이 재료는 약 0.9 내지 약 2.45 GHz에서 마이크로웨이빙된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 30 초 동안 또는 적어도 약 1 분, 또는 적어도 약 2 분, 또는 적어도 3 분 동안 마이크로웨이빙된다.
일 구현예에서, 전처리는 저 주파수에서 중 주파수까지의 초음파에 브라시카케아이 재료를 노출시키는 것을 포함한다. 일 구현예에서, 전처리는 열초음파(약 30℃ 내지 약 60℃의 열과 함께 저 주파수에서 중 주파수까지의 초음파)로 브라시카케아이 재료를 노출시키는 것을 포함한다. 일 구현예에서, 초음파는 산업적 규모 초음파 프로세서로 발생된다. 일 구현예에서, 초음파 프로세서는 연속식 또는 회분식 초음파 프로세서이다. 일 구현예에서, 초음파 프로세서는, 이로 제한되지는 않지만, 예를 들어, UIP500hd 또는 UIP4000(Hielscher, Ultrasound Technology)이다. 일 구현예에서, 초음파는 약 20 kHz 내지 약 600 kHz의 주파수에 있다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 30 초, 또는 적어도 약 1 분, 또는 적어도 약 2 분, 또는 적어도 약 3 분, 또는 약 5 분 동안 음파에 노출된다.
일 구현예에서, 전처리는 브라시카케아이 재료를 펄스 전기장 가공에 노출시키는 것을 포함한다. 펄스 전기장 가공은 짧은 고전압 펄스의 적용을 포함하는 비-열 가공 기술이다. 펄스는 글루코시놀레이트에 대한 미로시나제의 접근을 향상시키도록 브라시카케아이 재료의 세포의 전기천공을 유도한다. 일 구현예에서, 펄스 전기장 가공은 브라시카케아이 재료를 약 40 내지 약 70℃의 온도로 가열한다. 일 구현예에서, 펄스 전기장 가공은 브라시카케아이 재료를 약 50℃ 내지 약 70℃의 온도로 가열한다. 일 구현예에서, 펄스 전기장 가공은 브라시카케아이 재료를 약 60℃ 내지 약 70℃의 온도로 가열한다. 일 구현예에서, 펄스 전기장 가공은 브라시카케아이 재료를 약 20 kV 내지 약 80 kV의 전압 펄스로 처리하는 것을 포함한다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 10% 내지 약 90%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 20% 내지 약 80%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 30% 내지 약 70%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 40% 내지 약 60%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 10%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 20%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 30%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 40%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 50%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 60%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 70%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 80%를 이소티오시아네이트로 전환시킨다. 일 구현예에서, 전처리는 글루코시놀레이트의 약 90%를 이소티오시아네이트로 전환시킨다.
발효
당업자는 본원에 기재된 바와 같은 발효 방법이 임의의 젖산균의 사용을 포함할 수 있다는 것을 인지할 것이다. 본원에서 사용되는 "발효"는 젖산균에 의한 브라시카케아이 재료의 생화학적 분해를 지칭한다. 일 구현예에서, 젖산균으로의 발효는 외인성 젖산균의 첨가를 이용하여 수행된다. 본원에서 사용되는 "락틱 박테리아(lactic bacteria)" 또는 "젖산균(lactic acid bacteria)"은 탄수화물 발효의 최종 산물로서 젖산을 생산하는 세균이고, 락토바실러스(Lactobacillus), 류코노스톡(Leuconostoc), 페디오코쿠스(Pediococcus), 락토코쿠스(Lactococcus), 스트렙토코쿠스(Streptococcus), 아이로코쿠스(Aerococcus), 카르노박테리움(Carnobacterium), 엔테로코쿠스(Enterococcus), 오에노코쿠스(Oenococcus), 스포로락토바실러스(Sporolactobacillus), 테트라게노코쿠스(Tetragenococcus), 바고코쿠스(Vagococcus) 및 바이셀라(Weissella) 속으로부터의 세균을 포함할 수 있지만 이로 제한되지 않는다. 일 구현예에서, 젖산균은 미로시나제 활성을 포함한다. 일 구현예에서, 젖산균은 류코노스톡 속으로부터 유래된다. 일 구현예에서, 젖산균은 락토바실러스 속으로부터 유래된다.
일 구현예에서, 젖산균은 락토바실러스 플란타룸(Lactobacillus plantarum), 류코노스톡 메센테로이데스(Leuconostoc mesenteroides), 락토바실러스 람노서스(Lactobacillus rhamnosus), 락토바실러스 펜토서스(Lactobacillus pentosus), 락토바실러스 브레비스(Lactobacillus brevis), 락토코쿠스 락티스(Lactococus lactis), 페디오코쿠스 펜토사세우스( Pediococcus pentosaceus) 및 페디코쿠스 아키딜라크티( Pedicoccus acidilacti) 중 하나 이상으로부터 선택된다.
일 구현예에서, 젖산균은 브라시카케아이로부터 단리되었다. 일 구현예에서, 젖산균은 브라시카 올레라케아로부터 단리되었다. 일 구현예에서, 젖산균은 브로콜리로부터 단리되었다. 일 구현예에서, 젖산균은 브로콜리 잎으로부터 단리되었다. 일 구현예에서, 젖산균은 브로콜리 줄기로부터 단리되었다. 일 구현예에서, 젖산균은 브로콜리 퓨레로부터 단리되었다. 일 구현예에서, 젖산균은 호주산 브로콜리로부터 단리되었다.
일 구현예에서, 젖산균에는 미로시나제 활성이 결여된다.
일 구현예에서, 젖산균은 락토바실러스이다.
일 구현예에서, 젖산균은 i) 류코노스톡 메센테로이데스; ii) 락토바실러스 플란타룸; iii) 락토바실러스 펜토서스; iv) 락토바실러스 람노서스; v) i) 및 ii)의 조합; vi) i), ii) 및 iii)의 조합; 및 vii) i), ii) 및 iv)의 조합으로부터 선택된다.
일 구현예에서, 젖산균은 류코노스톡 메센테로이데스이다. 일 구현예에서, 류코노스톡 메센테로이데스는 ATCC8293이다. 일 구현예에서, 류코노스톡 메센테로이데스는 BF1 및/또는 BF2이다. 일 구현예에서, 류코노스톡 메센테로이데스에는 미로시나제 활성이 결여된다.
일 구현예에서, 젖산균은 락토바실러스 플란타룸이다. 일 구현예에서, 락토바실러스 플란타룸에는 미로시나제 활성이 결여된다.
일 구현예에서, 젖산균의 약 50%는 류코노스톡 메센테로이데스이고, 젖산균의 약 50%는 락토바실러스 종이다.
일 구현예에서, 젖산균의 약 50%는 류코노스톡 메센테로이데스이고, 젖산균의 약 50%는 락토바실러스 플란타룸이다.
일 구현예에서, 락토바실러스 플란타룸은 B1, B2, B3, B4 및 B5 중 하나 이상 또는 이들 모두로부터 선택된다. 일 구현예에서, 락토바실러스 플란타룸은 B1이다. 일 구현예에서, 락토바실러스 플란타룸은 B2이다. 일 구현예에서, 락토바실러스 플란타룸은 B3이다. 일 구현예에서, 락토바실러스 플란타룸은 B4이다. 일 구현예에서, 락토바실러스 플란타룸은 B5이다.
일 구현예에서, 발효는 BF1, BF2, B1, B2, B3, B4 및 B5로부터 선택된 젖산균의 적어도 2 개, 또는 적어도 3 개, 또는 적어도 4 개, 또는 적어도 5 개, 또는 적어도 6 개의 균주의 존재에서 일어난다.
일 구현예에서, 젖산균은 변형이 없는 대조 세균에 비해 고수준의 미로시나제 활성을 생성시키도록 변형된 재조합 세균이다. 당업자는 재조합 젖산균이 당업자에게 공지된 임의의 기술에 의해 생산된다는 것을 인지할 것이다.
일 구현예에서, 젖산균은, 이로 제한되지는 않지만, 예를 들어, 열 스트레스, 저온 스트레스, 치사량 아래의 초음파, 예를 들어, 약 20 MHz 내지 약 2000 MHz, 고압, 동적 고압 또는 펄스-전기장으로 스트레스를 받아, 스트레스를 받지 않은 대조 젖산균에 비해 미로시나제 활성 및 다당류 분해 효소의 활성을 증가시킨다. 일 구현예에서, 열 스트레스는 약 40℃ 내지 약 75℃ 초과로 세균을 가열하는 것을 포함한다. 일 구현예에서, 열 스트레스는 약 45℃ 내지 약 65℃ 초과로 세균을 가열하는 것을 포함한다. 일 구현예에서, 열 스트레스는 약 45℃ 내지 약 55℃ 초과로 세균을 가열하는 것을 포함한다. 일 구현예에서, 저온 스트레스는 약 0℃ 내지 약 8℃의 온도로 세균을 감소시키는 것을 포함한다. 일 구현예에서, 저온 스트레스는 약 2℃ 내지 약 6℃의 온도로 세균을 감소시키는 것을 포함한다. 일 구현예에서, 저온 스트레스는 약 4℃의 온도로 세균을 감소시키는 것을 포함한다.
일 구현예에서, 브라시카케아이 재료는 적어도 약 105 CFU/g의 본원에 기재된 바와 같은 젖산균으로 접종된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 106 CFU/g의 본원에 기재된 바와 같은 젖산균으로 접종된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 107 CFU/g의 본원에 기재된 바와 같은 젖산균으로 접종된다. 일 구현예에서, 브라시카케아이 재료는 적어도 약 108 CFU/g의 본원에 기재된 바와 같은 젖산균으로 접종된다. 일 구현예에서, 브라시카케아이 재료는 전처리되었다.
일 구현예에서, 발효는 약 20℃ 내지 약 34℃에서이다. 일 구현예에서, 발효는 약 22℃ 내지 약 34℃에서이다. 일 구현예에서, 발효는 약 24℃ 내지 약 34℃에서이다. 일 구현예에서, 발효는 약 24℃ 내지 약 30℃에서이다. 일 구현예에서, 발효는 약 34℃ 내지 약 34℃에서이다. 일 구현예에서, 발효는 약 25℃에서이다. 일 구현예에서, 발효는 약 30℃에서이다. 일 구현예에서, 발효는 약 34℃에서이다.
일 구현예에서, 발효는 약 8 시간 내지 약 17 일 동안이다. 일 구현예에서, 발효는 약 8 시간 내지 약 14 일 동안이다. 일 구현예에서, 발효는 약 8 시간 내지 약 7 일 동안이다. 일 구현예에서, 발효는 약 8 시간 내지 약 5 일 동안이다. 일 구현예에서, 발효는 약 8 시간 내지 약 4 일 동안이다. 일 구현예에서, 발효는 약 8 시간 내지 약 3 일 동안이다. 일 구현예에서, 발효는 약 8 시간 내지 약 30 시간 동안이다. 일 구현예에서, 발효는 약 8 내지 약 24 시간 동안이다. 일 구현예에서, 발효는 약 10 시간 내지 약 24 시간 동안이다. 일 구현예에서, 발효는 약 10 일 동안이다. 일 구현예에서, 발효는 약 9 일 동안이다. 일 구현예에서, 발효는 약 8 일 동안이다. 일 구현예에서, 발효는 약 7 일 동안이다. 일 구현예에서, 발효는 약 4 일 동안이다. 일 구현예에서, 발효는 약 6 일 동안이다. 일 구현예에서, 발효는 약 5 일 동안이다. 일 구현예에서, 발효는 약 72 시간 동안이다. 일 구현예에서, 발효는 약 60 시간 동안이다. 일 구현예에서, 발효는 약 45 시간 동안이다. 일 구현예에서, 발효는 약 30 시간 동안이다. 일 구현예에서, 발효는 약 24 시간 동안이다. 일 구현예에서, 발효는 약 20 시간 동안이다. 일 구현예에서, 발효는 약 18 일 동안이다. 일 구현예에서, 발효는 약 15 시간 동안이다. 일 구현예에서, 발효는 약 16 시간 동안이다. 일 구현예에서, 발효는 약 14 시간 동안이다. 일 구현예에서, 발효는 약 12 시간 동안이다. 일 구현예에서, 발효는 약 10 시간 동안이다. 일 구현예에서, 발효는 약 8 시간 동안이다. 일 구현예에서, 발효 배양물은 교반된다. 일 구현예에서, 교반은 간헐적이다. 일 구현예에서, 교반은 연속적이다. 특히 바람직한 구현예에서, 발효는 간헐적 교반과 함께 15 시간 동안이다. 특히 바람직한 구현예에서, 발효는 간헐적 교반과 함께 24 시간 동안이다.
일 구현예에서, 발효 반응은 조성물이 약 4.5 내지 약 3.8의 pH에 도달할 때 완료된다. 일 구현예에서, 발효 반응은 조성물이 약 4.5 내지 약 3.6의 pH에 도달할 때 완료된다. 일 구현예에서, 발효 반응은 조성물이 약 4.5 내지 약 4.04의 pH에 도달할 때 완료된다. 일 구현예에서, 발효 반응은 조성물이 약 4.3 내지 약 4.04의 pH에 도달할 때 완료된다. 일 구현예에서, 발효 반응은 조성물이 4.5 이하, 또는 4.4 이하, 또는 4.3 이하, 또는 4.04 이하, 또는 3.8 이하의 pH에 도달할 때 완료된다. 일 구현예에서, 발효 반응은 조성물이 4.5 이하의 pH에 도달할 때 완료된다. 일 구현예에서, 발효 반응은 조성물이 4.4 이하의 pH에 도달할 때 완료된다.
일 구현예에서, 존재 시, 발효는 다음 중 하나 이상 또는 이들 모두의 수를 감소시킨다: E. 콜라이, 살모넬라 및 리스테리아. 일 구현예에서, 존재 시, 발효는 다음 중 하나 이상 또는 이들 모두의 CFU/g를 감소시킨다: E. 콜라이, 살모넬라 및 리스테리아.
일 구현예에서, 염은 발효 배양물에 첨가되지 않는다.
일 구현예에서, 발효는 전처리 브라시카케아이 재료에서 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 증가시킨다. 일 구현예에서, 발효는 브라시카케아이 재료에서 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 증가시킨다. 일 구현예에서, 발효는 브라시카케아이 재료에서 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 약 100% 내지 약 500% 증가시킨다. 일 구현예에서, 발효는 브라시카케아이 재료에서 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 약 200% 내지 약 450% 증가시킨다. 일 구현예에서, 발효는 브라시카케아이 재료에서 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 약 250% 내지 약 450% 증가시킨다. 일 구현예에서, 발효는 브라시카케아이 재료에서 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 약 300% 내지 약 400% 증가시킨다. 일 구현예에서, 발효는 브라시카케아이 재료에서 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 약 300% 증가시킨다. 일 구현예에서, 발효는 브라시카케아이 재료에서 추출 가능한 글루코시놀레이트 함량에 비해 추출 가능한 글루코시놀레이트 함량을 약 400% 증가시킨다. 일 구현예에서, 글루코시놀레이트는 글루코라파닌이다.
산성화
전처리된 재료는, 프로덕트의 미생물 안전성 및 안정성(미생물 분해에 대한 민감성)을 개선하고, 프로덕트에서 이소티오시아네이트의 안정성을 증가시키기 위해 산성화될 수 있다. 산성화는 유기산, 이로 제한되지는 않지만, 예컨대, 젖산, 아세트산, 아스코르브산 및 시트르산의 첨가에 의해 달성될 수 있다. 구현예에서, 산성화는 글루코노-델타-락톤의 첨가로 달성될 수 있다. 일 구현예에서, 산성화는 pH를 약 4.4 내지 약 3.4의 pH로 감소시키는 것을 포함한다. 일 구현예에서, 산성화는 pH를 4.5, 또는 4.4, 또는 4.2, 또는 4, 또는 3.8, 또는 3.6, 또는 3.4 이하의 pH로 감소시키는 것을 포함한다. 일 구현예에서, 산성화는 pH를 4.4 이하의 pH로 감소시키는 것을 포함한다.
브라시카케아이
로부터의 이소티오시아네이트 함유 프로덕트
본원에 기재된 바와 같은 브라시카케아이로부터의 이소티오시아네이트 함유 프로덕트는 본원에 기재된 바와 같은 방법에 의해 생산될 수 있다. 본원에 기재된 바와 같은 방법을 이용하여 생산된 이소티오시아네이트 함유 프로덕트가 브라시카케아이 재료 또는 단독의 발효(본원에 기재된 바와 같은 전처리 없이)에 주어진 브라시카케아이 재료보다 높은 수준의 이소티오시아네이트, 예를 들어, 설포라판을 함유한다는 것이 당업자에게 인지될 것이다. 예를 들어, 상업적 브로콜리 재배종으로부터의 침연 브로콜리는 약 800 μmol/Kg dw(약 149.8 mg/Kg dw)의 설포라판 농도를 갖고, 발효 침연 브로콜리는 약 1600 μmol/Kg dw(약 278.8 mg/Kg dw)의 설포라판 농도를 갖고, 본원에 기재된 바와 같은 방법을 이용하여 생산된 전처리 및 발효 브로콜리는 약 13100 μmol/Kg dw(약 2318.7 mg/Kg dw)의 설포라판 농도를 갖는다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 적어도 약 4 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 적어도 약 6 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 적어도 약 8 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 적어도 약 10 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 적어도 약 12 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 적어도 약 14 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 적어도 약 16 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 적어도 약 17 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 약 4 배 내지 약 17 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 약 4 배 내지 약 16 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 약 8 배 내지 약 16 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 약 10 배 내지 약 16 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 약 12 배 내지 약 16 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 침연된 브라시카케아이 재료보다 약 14 배 내지 약 16 배 더 많은 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트는 설포라판이다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트에 존재하는 이소티오시아네이트의 수준은 브라시카케아이 재료의 추출 가능한 글루코시놀레이트 함량으로부터 예상되는 것보다 높다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 적어도 약 1 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 적어도 약 2 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 적어도 약 3 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 적어도 약 3.8 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 적어도 약 4 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 약 1 배 내지 약 4 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 약 1 배 내지 약 3.8 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 약 2 배 내지 약 3.8 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코시놀레이트 함량에 기초한 이소티오시아네이트의 예상 최대 수율의 약 2 배 내지 약 3 배를 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트에 존재하는 설포라판의 수준은 브라시카케아이 재료의 추출 가능한 글루코라파닌 함량으로부터 예상되는 것보다 높다. 일 구현예에서, 이소시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 적어도 약 1 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 적어도 약 2 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 적어도 약 3 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 적어도 약 3.8 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 적어도 약 4 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 약 1 배 내지 약 4 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 약 1 배 내지 약 3.8 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 약 1 배 내지 약 3 배를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 추출 가능한 글루코라파닌 함량에 기초한 설포라판의 예상 최대 수율의 약 2 배 내지 약 3 배를 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 100 mg/kg dw 내지 약 7000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 500 mg/kg dw 내지 약 7000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 1000 mg/kg dw 내지 약 7000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 1600 mg/kg dw 내지 약 4000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 1600 mg/kg dw 내지 약 3000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 2000 mg/kg dw 내지 약 4000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 2000 mg/kg dw 내지 약 7000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 3000 mg/kg dw의 이소티오시아네이트 내지 약 7000 mg/kg의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 2300 mg/kg dw의 이소티오시아네이트를 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 100 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 200 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 250 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 300 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 350 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 400 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 450 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 500 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 550 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 600 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 650 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 700 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 1000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 2000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 3000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 4000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 5000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 6000 mg/kg dw의 이소티오시아네이트를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 7000 mg/kg dw의 이소티오시아네이트를 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 100 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 150 mg/kg의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 200 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 250 mg/kg의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 300 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 350 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 400 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 450 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 500 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 550 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 600 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 650 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 700 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 1000 mg/kg의 설포라판 dw을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 2000 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 3000 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 4000 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 5000 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 6000 mg/kg dw의 설포라판을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 7000 mg/kg dw의 설포라판을 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 5% 더 많은 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 10% 더 많은 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 15% 더 많은 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 20% 더 많은 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 4% 더 많은 단백질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 6% 더 많은 단백질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 8% 더 많은 단백질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 10% 더 많은 단백질을 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 10% 더 적은 탄수화물을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 20% 더 적은 탄수화물을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 30% 더 적은 탄수화물을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 40% 더 적은 탄수화물을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 45% 더 적은 탄수화물을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 적어도 약 48% 더 적은 탄수화물을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료보다 약 10% 내지 약 48% 더 적은 탄수화물을 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료에 비해 증가된 수준의 다가페놀 글리코시드를 포함한다. 일 구현예에서, 다가페놀 글리코시드는 안토시아닌 글리코시드이다. 일 구현예에서, 다가페놀 글리코시드는 페놀산 글리코시드이다. 일 구현예에서, 다가페놀 글리코시드는 페놀산이다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료에 비해 증가된 수준의 글루코시놀레이트를 포함한다. 일 구현예에서, 글루코시놀레이트는 글루코라파닌이다. 일 구현예에서, 글루코라파닌는 적어도 약 25 배 증가된다. 일 구현예에서, 글루코시놀레이트는 글루코브라시신이다. 일 구현예에서, 글루코브라시신은 26 배 증가된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 인돌-3-카비놀을 포함한다. 일 구현예에서, 인돌-3카비놀은 침연된 브라시카케아이 재료에 비해 이소티오시아네이트 함유 프로덕트에서 적어도 약 2 배 증가된다. 일 구현예에서, 인돌-3-카비놀은 침연된 브라시카케아이 재료에 비해 이소티오시아네이트 함유 프로덕트에서 적어도 약 3 배 증가된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 아스코르비겐을 포함한다. 일 구현예에서, 아스코르비겐은 침연된 브라시카케아이 재료에 비해 이소티오시아네이트 함유 프로덕트에서 적어도 약 2 배 증가된다. 일 구현예에서, 아스코르비겐은 침연된 브라시카케아이 재료에 비해 이소티오시아네이트 함유 프로덕트에서 적어도 약 3 배 증가된다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료에 비해 증가된 수준의 페룰산, 시링산, 페닐락트산, 클로로겐산루틴, 시나프산, 메틸 시린게이트, 헤스페레틴, 퀘르세틴 및 캠페롤 중 하나 이상을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료에 비해 증가된 수준의 클로로겐산을 포함한다. 일 구현예에서, 클로로겐산은 약 6.6 배 증가된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료에 비해 증가된 수준의 시나프산을 포함한다. 일 구현예에서, 시나프산은 약 23.8 배 증가된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료에 비해 증가된 수준의 캠페롤을 포함한다. 일 구현예에서, 시나프산은 약 10.5 배 증가된다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브라시카케아이 재료에 비해 감소된 수준의 프로토카테츄산, 갈산, 4,하이드록시벤조산, 바닐산, 2,3디하이드록시벤조산, p-쿠마르산, 신남산, 카테킨, 로즈마린산, 카페산 중 하나 이상을 포함한다.
일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 40%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 50%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 60%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 70%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 80%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 90%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 95%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 97%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 98%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 99%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 100%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 40% 내지 약 100%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다. 일 구현예에서, 브라시카케아이 재료에 존재하는 글루코시놀레이트의 약 40% 내지 약 80%는 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트로 전환된다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트는 약 4℃ 내지 약 25℃에서 저장될 때 적어도 1 주 동안, 또는 적어도 2 주 동안, 또는 적어도 3 주 동안, 또는 적어도 4 주 동안, 또는 적어도 6 주 동안, 또는 적어도 8 주 동안, 또는 적어도 10 주 동안, 또는 적어도 12 주 동안, 또는 적어도 14 주 동안 안정하다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트는 약 4℃ 내지 약 25℃에서 저장될 때 적어도 4 주 동안 안정하다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트는 약 4℃ 내지 약 25℃에서 저장될 때 적어도 8 주 동안 안정하다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트에서 이소티오시아네이트는 약 4℃ 내지 약 25℃에서 저장될 때 적어도 12 주 동안 안정하다.
본원에서 사용되는 "안정한"은 4℃에서 6 주 동안 저장될 때 이소티오시아네이트 농도의 비감소 또는 단지 약간의 감소를 지칭한다. 일 구현예에서, 약간의 감소는 약 1% 내지 약 30%의 이소티오시아네이트 농도의 감소를 지칭한다. 일 구현예에서, 약간의 감소는 약 5% 이하의 이소티오시아네이트 농도의 감소를 지칭한다. 일 구현예에서, 약간의 감소는 약 10% 이하의 이소티오시아네이트 농도의 감소를 지칭한다. 일 구현예에서, 약간의 감소는 약 15% 이하의 이소티오시아네이트 농도의 감소를 지칭한다. 일 구현예에서, 약간의 감소는 약 20% 이하의 이소티오시아네이트 농도의 감소를 지칭한다. 일 구현예에서, 약간의 감소는 약 30% 이하의 이소티오시아네이트 농도의 감소를 지칭한다. 이소티오시아네이트 분석은 당업계에 공지된 임의의 방법에 의해, 그리고, 예를 들어, 설포라판에 대하여 실시예 1에 나타나 있는 바와 같이 수행될 수 있다.
일 구현예에서, 이소티오시아네이트는 설포라판이다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 4℃ 내지 약 25℃에서 저장될 때 적어도 1 주 동안, 또는 적어도 2 주 동안, 또는 적어도 3 주 동안, 또는 적어도 4 주 동안, 또는 적어도 6 주 동안, 또는 적어도 8 주 동안, 또는 적어도 10 주 동안, 또는 적어도 12 주 동안, 또는 적어도 14 주 동안 효모, 곰팡이 및/또는 대장균형 성장에 대해 저항성이다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 4℃ 내지 약 25℃에서 저장될 때 적어도 4 주 동안 효모, 곰팡이 및/또는 대장균형 성장에 대해 저항성이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 4℃ 내지 약 25℃에서 저장될 때 적어도 8 주 동안 효모, 곰팡이 및/또는 대장균형 성장에 대해 저항성이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 4℃ 내지 약 25℃에서 저장될 때 적어도 12 주 동안 효모, 곰팡이 및/또는 대장균형 성장에 대해 저항성이다.
본원에서 사용되는 효모, 곰팡이 및/또는 대장균형 성장에 대해 "저항성"은 실시예 1에 기재된 방법을 이용하여 상기 열거된 기간 후에 샘플에서 <1 Log CFU/g의 효모, 곰팡이 및/또는 대장균형이 검출 가능하다는 것을 의미한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 20 g/100 gdw 내지 약 32 g/100 gdw의 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 20 g/100 gdw의 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 25 g/100 gdw의 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 28 g/100 gdw의 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 29 g/100 gdw의 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 30 g/100 gdw의 총 섬유질을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 32 g/100 gdw의 총 섬유질을 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 14000 μmol TE/100 gdw 내지 약 19000 μmol TE/100 gdw의 ORAC 항산화능을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 14000 μmol TE/100 gdw의 ORAC 항산화능을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 15000 μmol TE/100 gdw의 ORAC 항산화능을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 16000 μmol TE/100 gdw의 ORAC 항산화능을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 17000 μmol TE/100 gdw의 ORAC 항산화능을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 18000 μmol TE/100 gdw의 ORAC 항산화능을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 18695 μmol TE/100 gdw의 ORAC 항산화능을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 19000 μmol TE/100 gdw의 ORAC 항산화능을 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 1750 mg GAE/100 gdw 내지 약 2600 mg GAE/100 gdw의 총 다가페놀 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 1750 mg GAE/100 gdw의 총 다가페놀 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 2000 mg GAE/100 gdw의 총 다가페놀 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 2100 mg GAE/100 gdw의 총 다가페놀 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 2200 mg GAE/100 gdw의 총 다가페놀 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 2300 mg GAE/100 gdw의 총 다가페놀 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 2360 mg GAE/100 gdw의 총 다가페놀 함량을 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 0.9% 내지 약 1.1%의 젖산 당량의 총 적정 산도를 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 1.1%의 젖산 당량의 총 적정 산도를 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 23 mg GAE/100 gdw 내지 약 39 mg GAE/100 gdw의 총 단백질 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 23 mg GAE/100 gdw 내지 약 30 mg GAE/100 gdw의 총 단백질 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 25 mg GAE/100 gdw의 총 단백질 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 27 mg GAE/100 gdw의 총 단백질 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 28 mg GAE/100 gdw의 총 단백질 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 29 mg GAE/100 gdw의 총 단백질 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 30 mg GAE/100 gdw의 총 단백질 함량을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약 32 mg GAE/100 gdw의 총 단백질 함량을 포함한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 100 mg/kg dw의 이소티오시아네이트 및 하기 중 하나 이상 또는 이들 모두를 포함한다:
i) 약 29 내지 약 36g/100 gdw의 총 섬유질;
ii) 약 15000 내지 약 18695 μmol TE/100 gdw의 ORAC 항산화능;
iii) 약 2310 내지 약 2600 mg GAE/100 gdw의 총 다가페놀 함량;
iv) 약 0.9 내지 약 1.1%의 젖산 당량의 총 적정 산도;
v) 약 27 내지 약 39 g/100 gdw의 총 단백질 함량; 및
vi) 류코노스톡 메센테로이데스 및/또는 락토바실러스 플란타룸.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 브로콜리로부터 생산된다.
본원에 기재된 바와 같은 브라시카케아이 프로덕트는 대상체에서 글루코시놀레이트 함유 프로덕트의 소화 동안 이소티오시아네이트 함유 프로덕트에 존재하는 글루코시놀레이트의 이소티아시아네이트로의 전환을 도울 수 있는 생 젖산균을 포함할 수 있다(즉, 이들은 프로바이오틱으로 작용함). 일 구현예에서, 젖산균은 류코노스톡 메센테로이데이다. 일 구현예에서, 젖산균은 락토바실러스 종이다. 일 구현예에서, 젖산균은 락토바실러스 플란타룸이다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 102 CFU/g의 농도의 젖산균을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 102 CFU/g의 농도의 젖산균을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 105 CFU/g의 농도의 젖산균을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 106 CFU/g의 농도의 젖산균을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 107 CFU/g의 농도의 젖산균을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 108 CFU/g의 농도의 젖산균을 포함한다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 적어도 약 109 CFU/g의 농도의 젖산균을 포함한다.
일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 10 일 동안 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 20 일 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 30 일 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 40 일 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 50 일 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 60 일 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 70 일 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 80 일 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 85 일 존재한다. 일 구현예에서, 생 젖산균은 약 4℃ 내지 약 25℃에서 저장될 때 이소티오시아네이트 함유 프로덕트에 적어도 90 일 존재한다.
일 구현예에서, 젖산균은 락토바실러스 종이다. 일 구현예에서, 젖산균은 락토바실러스 플란타룸이다. 일 구현예에서, 젖산균은 류코노스톡 메센테로이데스이다. 일 구현예에서, 젖산균은 적어도 약 107 CFU/g의 농도로 존재한다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 젖산균에 의해 생산된 하나 이상의 박테리오신/들을 포함한다. 일 구현예에서, 박테리오신은 클래스 I 박테리오신이다. 일 구현예에서, 박테리오신은 클래스 II 박테리오신이다. 일 구현예에서, 박테리오신은 클래스 III 박테리오신이다. 젖산균에 의해 생산되는 박테리오신의 예는 문헌[Alvarez-Sieiro et al. (2016)]에서 찾아볼 수 있다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 식품이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 영양약제이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 보충제이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 식품 성분이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 프로바이오틱이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 동물 사료이다. 동물은 수중 동물, 예컨대, 어류 또는 가축일 수 있다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 구충제이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 약용화장품이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트튼 국소용으로 제형화된다.
일 구현예에서, 이소티오시아네이트 함유 프로덕트는 고체, 액체, 퓨레 또는 분말이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 발효 후 분말로 건조된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 발효 후 냉동 건조된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 발효 후 WO2005030229호에 기재된 바와 같이 마이크로캡슐화된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 알약으로 제형화된다.
후처리
일 구현예에서, 발효 또는 산성화 후, 이소티오시아네이트 함유 프로덕트는, 예를 들어, 소비되는 경우, 프로덕트의 손상 또는 병원성에 기여하는 미생물을 불활성화시키도록 후처리될 수 있다.
본원에서 사용되는 "후처리" 또는 "후처리하는"은, 미생물을 불활성화시키기 위한, 발효 후 본원에 기재된 바와 같은 이소티오시아네이트 함유 프로덕트의 처리를 지칭한다. 본원에서 사용되는 "미생물"은 이소티오시아네이트 함유 프로덕트의 손상 또는 부패를 초래할 수 있는 세균, 바이러스, 진균 또는 진핵생물 활성을 지칭한다. 본원에서 사용되는 미생물을 "불활성화시키다" 또는 미생물의 "불활성화"는 생존 가능한 미생물을 약 1 내지 약 7 log 감소시키는 것을 지칭한다. 일 구현예에서, 생존 가능한 미생물은 약 1 내지 6 log 감소된다. 일 구현예에서, 생존 가능한 미생물은 약 2 내지 6 log 감소된다. 일 구현예에서, 생존 가능한 미생물은 약 3 내지 6 log 감소된다.
당업자는, 후처리가, 예를 들어, 열 처리, UV 처리, 초음파 가공, 펄스 전기장 가공 또는 고압 가공을 포함하는, 미생물을 불활성화시키는 임의의 방법일 수 있다는 것을 인지할 것이다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 열 가공으로 후처리된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 고압 가공으로 후처리된다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 후 처리 동안 밀봉된 패키지에 있다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 고압 가공 동안 밀봉된 패키지에 있다. 일 구현예에서, 이소티오시아네이트 함유 프로덕트는 열 처리 동안 밀봉된 패키지에 있다. 일 구현예에서, 고압 가공은 약 300 MPa 내지 약 600 MPa에서 등압으로 이소티오시아네이트 함유 프로덕트를 처리하는 것을 포함한다. 일 구현예에서, 고압 가공은 약 350 MPa 내지 약 550 MPa에서 등압으로 이소티오시아네이트 함유 프로덕트를 처리하는 것을 포함한다. 일 구현예에서, 고압 가공은 약 300 MPa 내지 약 400 MPa에서 등압으로 이소티오시아네이트 함유 프로덕트를 처리하는 것을 포함한다. 일 구현예에서, 열 처리는 약 60℃ 내지 약 121℃의 온도로 샘플을 가열하는 것을 포함한다. 일 구현예에서, 열 처리는 약 65℃ 내지 약 100℃의 온도로 샘플을 가열하는 것을 포함한다. 일 구현예에서, 열 처리는 약 65℃ 내지 약 80℃의 온도로 샘플을 가열하는 것을 포함한다. 일 구현예에서, 열 처리는 약 65℃ 내지 약 75℃의 온도로 샘플을 가열하는 것을 포함한다.
단리된 균주 및 스타터 배양물
일 구현예에서, 본 발명은 본원에 기재된 바와 같은 방법 및 프로덕트에서 사용하기에 적합한 젖산균의 단리된 균주를 제공한다.
일 구현예에서, 본 발명은
i) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021729 하에 기탁된 BF1;
ii) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021730 하에 기탁된 BF2;
iii) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021731 하에 기탁된 B1;
iv) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021732 하에 기탁된 B2;
v) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021733 하에 기탁된 B3;
vi) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021734 하에 기탁된 B4; 및
vii) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021735 하에 기탁된 B5로부터 선택된 젖산균의 단리된 균주를 제공한다.
일 구현예에서, 본 발명은 SmaI 및/또는 NotI으로 절단될 때 BF1 또는 BF2와 동일한 SmaI 및/또는 NotI 핑거프린트를 생성시키는 게놈 DNA를 포함하는 류코노스톡 메센테로이데스의 단리된 균주를 제공한다. BF1 및 BF2에 대한 SmaI 및 NotI 핑거프린트는 도 13에 나타나 있다.
일 구현예에서, 본 발명은 SmaI 및/또는 NotI로 절단될 때 B1, B2, B3, B4 또는 B5와 동일한 SmaI 및/또는 NotI 핑거프린트를 생성시키는 게놈 DNA를 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다.
일 구현예에서, 본 발명은 ATCC8293와 상이한 표 18 또는 표 19에 열거된 다형성 중 하나 이상 또는 이들 모두를 포함하는 류코노스톡 메센테로이데스의 단리된 균주를 제공한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 18 또는 표 19에 열거된 다형성 중 5 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 18 또는 표 19에 열거된 다형성 중 10 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 18 또는 표 19에 열거된 다형성 중 15 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 18 또는 표 19에 열거된 다형성 중 19 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 20 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 30 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 50 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 80 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 100 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 150 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 200 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 300 개 이상을 포함한다. 일 구현예에서, 류코노스톡 메센테로이데스의 단리된 균주는 ATCC8293와 상이한 표 19에 열거된 다형성 중 400 개 이상을 포함한다.
일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 하나 이상 또는 이들 모두를 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다. 일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 5 개 이상을 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다. 일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 10 개 이상을 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다. 일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 15 개 이상을 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다. 일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 20 개 이상을 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다. 일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 25 개 이상을 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다. 일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 30 개 이상을 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다. 일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 35 개 이상을 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다. 일 구현예에서, 본 발명은 ATCC8014와 상이한 표 13, 표 14, 표 15, 표 16 또는 표 17에 열거된 다형성 중 40 개 이상을 포함하는 락토바실러스 플란타룸의 단리된 균주를 제공한다.
일 구현예에서, 본 발명은 본원에 기재된 바와 같은 단리된 균주 중 하나 이상을 포함하는 젖산균을 포함하는 이소티오시아네이트 함유 프로덕트 또는 프로바이오틱을 생산하기 위한 스타터 배양물을 제공한다. 본원에서 사용되는 "스타터 배양물"은 발효를 위한 살아있는 미생물의 배양물이다. 일 구현예에서, 본 발명은 하기 중 하나 이상 또는 이들 모두로부터 선택된 젖산균을 포함하는 이소티오시아네이트 함유 프로덕트 또는 프로바이오틱을 생산하기 위한 스타터 배양물을 제공한다:
i) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021729 하에 기탁된 BF1;
ii) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021730 하에 기탁된 BF2;
iii) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021731 하에 기탁된 B1;
iv) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021732 하에 기탁된 B2;
v) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021733 하에 기탁된 B3;
vi) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021734 하에 기탁된 B4; 및
vii) 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021735 하에 기탁된 B5.
일 구현예에서, 브라시카케아이 재료에는 본원에 기재된 바와 같은 스타터 배양물이 적어도 약 105 CFU/g 접종된다. 일 구현예에서, 브라시카케아이 재료에는 본원에 기재된 바와 같은 스타터 배양물이 적어도 약 106 CFU/g 접종된다. 일 구현예에서, 브라시카케아이 재료에는 본원에 기재된 바와 같은 스타터 배양물이 적어도 약 107 CFU/g 접종된다. 일 구현예에서, 브라시카케아이 재료에는 본원에 기재된 바와 같은 스타터 배양물이 적어도 약 108 CFU/g 접종된다. 일 구현예에서, 브라시카케아이 재료에는 본원에 기재된 바와 같은 스타터 배양물이 적어도 약 1010 CFU/g 접종된다. 일 구현예에서, 브라시카케아이 재료에는 본원에 기재된 바와 같은 스타터 배양물이 약 105 CFU/g 내지 약 1010 CFU/g 접종된다.
프로바이오틱
일 구현예에서, 본 발명은 브라시카케아이로부터 단리된 젖산균 중 하나 이상을 포함하는 프로바이오틱을 제공한다. 본원에서 사용되는 "프로바이오틱"은 적당한 양으로 투여될 때 숙주에게 유익한 건강을 주는 살아있는 미생물을 지칭한다. 일 구현예에서, 젖산균은 브라시카 올레라케아로부터 단리되었다. 일 구현예에서, 젖산균은 브로콜리로부터 단리되었다. 일 구현예에서, 젖산균은 호주산 브로콜리로부터 단리되었다. 일 구현예에서, 젖산균은 i) 류코노스톡 메센테로이데스; ii) 락토바실러스 플란타룸; iii) 락토바실러스 펜토서스; iv) 락토바실러스 람노서스; v) i) 및 ii)의 조합; vi) i), ii) 및 iii)의 조합; 및 vii) i), ii) 및 iv)의 조합으로부터 선택된다. 일 구현예에서, 젖산균은 BF1, BF2, B1, B2, B3, B4 및 B5 중 하나 이상 또는 이들 모두로부터 선택된다. 일 구현예에서, 젖산균은 B1이다. 일 구현예에서, 젖산균은 B2이다. 일 구현예에서, 젖산균은 B3이다. 일 구현예에서, 젖산균은 B4이다. 일 구현예에서, 젖산균은 B5이다. 일 구현예에서, 프로바이오틱은 캡슐, 정제, 분말 또는 액체이다. 일 구현예에서, 프로바이오틱은 WO 2005030229호에 기재된 바와 같이 마이크로캡슐화된다.
실시예
실시예 1 - 방법
화학물질 및 시약
HPLC 등급 메탄올, 소듐 디하이드로젠 포스페이트, 소듐 하이드록사이드(NaOH) 및 염산(HCl)을 Merck(Damstadt, 독일)로부터 구매하였다. 폴린-시오칼토 시약(Folin-Ciocalteu's reagent), 소듐 카보네이트(Na2CO3), 갈산, 플루오레세인 소듐 염 및 이염기성-포타슘 포스페이트를 Sigma Aldrich(St. Louis, MO, USA)로부터 구매하였다. 소듐 디하이드로젠 포스페이트, 6-하이드록시-2,5,7,8-테트라메틸크로만-2-카복실산(trolox), 2,20-아조비스(2-메틸프로피온아미딘) 디하이드로클로라이드(AAPH)를 Sapphire Bioscience(Redfern, NSW, 호주)로부터 구매하였다.
젖산균
발효 동안 사용한 젖산균은
LP: 락토바실러스 플란타룸 ATCC8014;
LGG: 락토바실러스 람노서스 ATCC53103;
B1: 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021731 하에 기탁된 브로콜리로부터 단리된 락토바실러스 플란타룸;
B2: 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021732 하에 기탁된 브로콜리로부터 단리된 락토바실러스 플란타룸;
B3: 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021733 하에 기탁된 브로콜리로부터 단리된 락토바실러스 플란타룸;
B4: 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021734 하에 기탁된 브로콜리로부터 단리된 락토바실러스 플란타룸;
B5: 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021735 하에 기탁된 브로콜리로부터 단리된 락토바실러스 플란타룸;
BF1: 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021729 하에 기탁된 브로콜리 퓨레로부터 단리된 류코노스톡 메센테로이데스;
BF2: 호주 국립 측정 기관에서 2017년 9월 25일에 V17/021730 하에 기탁된 브로콜리 퓨레 BF2로부터 단리된 류코노스톡 메센테로이데스;
BP: 합한 BF1, BF2; 및
LAB: 합한 B1, B2, B3, B4 및 B5 중 하나 이상으로부터 선택되었다.
BF1 및 BF2는 16s-RNA 서열(Australian Genome Research Facility; 데이터 미도시)을 통해 류코노스톡 메센테로이데스로 확인되었다. B1 내지 B5는 16S-RNA 서열을 기초로 락토바실러스 플란타룸으로 확인되었다. 모든 단리물의 동일성을 전장 게놈 서열 분석에 의해 확인하였다.
브로콜리 및 브로콜리 퓨레로부터 젖산균의 단리
상기 락토바실러스 플란타룸 B1, B2, B3, B4 및 B5를 브로콜리 잎 및 줄기로부터 단리시켰다. 잎 및 줄기를 물로 세척하고, 스토마커(stomacher)를 사용하여 펩톤 염수의 첨가로 균질화시켰다. 침지 용액을 연속 희석하고, 드만, 로고사 및 샤르페(De Man, Rogosa and Sharpe: MRS) 한천 상에 평판 도말하였다. 추정 중온성 젖산균을 단리시키기 위해 평판을 37℃에서 48 시간 내지 72 시간 동안 혐기성 조건 하에 인큐베이션하였다. MRS 평판 상의 상이한 집락 형태를 기초로 하여, 콜로니를 단리시키고, MRS 브로스에서 배양하고, 염색 및 생화학적 특성규명 기법을 이용하여 스크리닝하고, -80℃에서 글리세롤과 계속 냉동시켰다. 단리물을 AGRF에서 16s RNA 시퀀싱을 이용하여 종 수준에서 확인하였다.
류코노스톡 메센테로이데스 BF1 및 BF2의 단리를 위해, 브로콜리 잎으로부터의 단리를 위한 상술된 현탁 대신에 브로콜리 꽃송이 퓨레를 연속 희석 후에 사용하였다.
스타터 배양물의 제조
젖산균 균주, 류코노스톡 메센테로이데스 및 락토바실러스 플란타룸을 브로콜리로부터 단리시키고, Australian Genome Research Facility Ltd.에 의해 확인하였다. 일차 배양물을 얻기 위해, -80℃에서 저장된 젖산균 배양물을 10 mL의 MRS 브로스(Oxoid, 빅토리아, 호주)에 접종하고, 30℃에서 24 시간 동안 인큐베이션하여 8 log 집락-형성 단위/밀리리터(CFU/mL)의 초기 바이오매스를 수득하였다. 2 mL의 각 일차 접종원을 200 mL의 MRS 브로스에 접종하고, 30℃에서 24 시간 동안 인큐베이션하였다. 배양물을 4℃에서 15 분 동안 2000g으로 원심분리에 의해 수거하고, 멸균 인산염 완충 식염수(PBS)로 2회 세척하고, 모든 락토바실러스 플란타룸 배양물을 함께 혼합하고, 모든 류코노스톡 메센테로이데스 배양물을 함께 혼합하였다. 두 개의 배양 현탁물을 10 log CFU/ml로 희석하고, 동일한 부피 비율로 혼합하고, 브로콜리 발효를 위한 혼합된 스타터 배양물로서 사용할 때까지 -80℃에서 글리세롤과 저장하였다.
발효 방법
브로콜리(브라시카 올레라케아 L. 아종, 이탈릭; 30 kg) 꽃송이를 크라운(crown)으로부터 대략 2 cm로 자르고, 더 작은 조각으로 잘게 썰고, 매직 불렛 블렌더(magic bullet blender)를 사용하여 3:2 비로 1 분 동안 밀리-Q 워터(Milli-Q water)로 침연시켰다. 브로콜리 슬러리를 잘 혼합하고, 나사형 뚜껑이 있는 멸균 플라스틱 병(200 mL)에 넣었다. 브로콜리 퓨레의 각 병(200 mL)에 8 log CFU/g의 초기 농도로 제조된 스타터 배양물을 접종하였다. 발효 실험을 약 4.0의 pH 값에 도달할 때까지(4 일) 30℃에서 평행하게 48 개의 병에서 수행하였다. 발효 단계가 완료된 후, 3 개의 샘플을 0 일 저장 샘플로 취하고, 다른 샘플들을 저장 실험을 위한 두 개의 로트로 분리하였다: 하나의 로트는 냉장고(4℃)에 저장하고, 또 다른 것은 25℃에서 온도 조절되는 공간에 저장하였다. 미생물학적, 물리화학적 및 식물화학적 분석을 위해 샘플들을 12 주에 걸쳐 주기적으로 취하였다. 발효된 브로콜리 퓨레를, 균질화 후 -20℃에서 저장된 미가공 브로콜리 퓨레 및 LAB에 의한 접종이 없는 발효 샘플과 동일한 기간 동안 인큐베이션된 퓨레 샘플과 비교하였다.
샘플링
시간 경과 실험을 위해, 각각 25℃ 및 4℃에서 저장된 샘플에 대해 10 일, 20 일, 30 일, 40 일, 50 일, 60 일, 70 일, 80 일, 및 90 일에, 그리고 14 일, 28 일, 42 일, 56 일, 70 일 및 84 일에 샘플링을 수행하였다. 샘플링을 표면 상에서 측정된 색으로 3 회 수행하고, 발효 병을 연 직후에 pH를 측정하였다. 그 후에, 미생물학적 분석 및 적정 산도 분석을 위해 샘플을 취하였다. 나머지 재료를 두 개의 분획으로 분리하고, 제1 분획을 냉동시키고, 냉동 건조시키고, 미세 분말로 분쇄하고, 추가 분석을 위해 데시케이터에 저장하고, 제2 분획을 글리코라파닌 및 설포라판 분석까지 -20℃에서 냉동시키고 유지시켰다.
미생물학적 분석
미생물 분석을 위해, 3 가지 상이한 배지를 사용하여 상이한 미생물의 브로콜리 퓨레 g 당 CFU를 측정하였다: 드만-로고사-샤르프(MRS) 한천에 대해 총 젖산균, 바이올렛 레드 담즙 글루코스 한천(violet red bile glucose agar: VRBGA)에 대해 총 장내세균, 및 포테이토 덱스트로스 한천(potato dextrose agar: PDA)에 대해 효모 및 곰팡이의 평판 계수. 각 샘플에 대하여, 멸균된 펩톤 염수 희석제에서 브로콜리 현탁물의 연속 희석을 수행하고, 0.1 mL의 희석물을 한천 평판 상에 2 회 플레이팅하였다. 각각 25℃에서 72 시간 동안 호기성 인큐베이션(PDA), 37℃에서 24 시간 동안(VRBGA), 및 30℃에서 72 시간 동안 혐기성 인큐베이션(MRS) 후, CFU를 계수하였다.
pH 및 적정 산도의 결정
pH 값을 pH 미터(PHM240, MeterLab)에 의해 브로콜리 퓨레를 함유하는 발효 병에서 직접적으로 결정하였다. 브로콜리 샘플의 적정 산도(TA)를 자동 적정기(Titralab 854 적정 매니저, Radiometric Analytical, 프랑스)로 측정하였다. 간략히 말해서, 희석된 브로콜리 퓨레(10 mL)를 종말점 pH = 8.1까지 0.1 M NaOH를 사용하여 적정하고, 얻어진 결과는 하기 식에 따라 샘플의 리터 당 젖산의 그램 당량으로 표현하였다.
상기 식에서, v는 NaOH의 역가 부피이다. 젖산에 대한 산 팩터는 0.009였다.
총 단백질 및 색 분석
브로콜리 샘플의 총 단백질 함량을 총 질소 함량에 6.25를 곱해서 결정하였다. 브로콜리의 총 질소 함량을 LECO TruMac 장치(LECO Corporation, 미시간, USA)로 뒤마 연소법(Dumas combustion method)을 이용하여 분석하였다. 발효 브로콜리 샘플의 색 지수(L, a, b)를 크로마미터 CR-200 삼색 비색계(Minolta, 오사카, 일본)를 사용하여 결정하였다. 얻어진 색 값은 밝음/어두움(L*로서), 적색도/녹색도(a*) 및 황색도/청색도(b*)로 표현되었다. 총 색차(△E)를 하기 식에 따라 계산하였다:
상기 식에서, L0, a0, b0는 신선한 비발효 브로콜리의 색 값이다.
총 다가페놀 함량의 결정
총 페놀 함량(TPC)을 변형으로 폴린-시오칼토 비색법[Singleton and Rossi, 1965]을 이용하여 분광 광도계로 측정하였다. 간략히 말해서, 50 mg의 브로콜리 분말을 10 mL의 산성화(1% HCl) 메탄올/물(70:30, v/v) 용액에 현탁시키고, 8 분 동안 초음파 배쓰(IDK technology Pty Ltd, VIC, 호주)에서 추출하였다. 추출물을 4℃에서 16 시간 동안 유지하고, 0.2 μM 필터로 여과하고, 분석까지 4℃에서 저장하였다. 1 mL의 0.2 N 폴린-시오칼토 시약, 800 μL의 소듐 카보네이트 용액(7.5 % p/v) 및 180 μL의 밀리-Q 등급 물을 추출물(20 μL)에 첨가하였다. 37℃의 암흑에서 1 시간의 인큐베이션 후, 분광 광도계(UV-1700 Pharma Spec, SHIMADZU)를 사용하여 흡광도를 765 nm에서 3 회 측정하였다. 갈산을 표준물로 사용하고, 갈산의 공지된 농도를 이용하여 생성된 표준 곡선을 기초로 생중량 100 g 당 갈산의 당량 mg(GAE)(mg GAE/100 g FW)으로 TPC를 표현하였다.
산소 라디칼 흡수능 검정
냉동-건조된 브로콜리 분말(10 mg)을 10 mL의 메탄올/물(80:20, v/v) 추출 용매에 현탁시켰다. 슬러리를 실온에서 1 시간 동안 Heidolph Multi-Reax(John Morris Scientific, NSW, 호주)로 650 rpm에서 추출하였다. 그 후에, 이를 4℃에서 15 분 동안 25,000 g으로 원심분리하고, 상청액을 수거하고, 75 mM의 포타슘 포스페이트 완충액(pH 7.4)으로 100× 희석 후 분석을 위한 준비를 하였다. 소수 변형으로 문헌[Huang et al. (2002)]에 의해 보고된 절차에 따라 ORAC 분석을 실시하였다. 불투명 96-웰 플레이트(dark optical bottom, 월섬, MA, USA)에서 검정을 수행하였다. 검정 반응물에는 81.6 nM의 플루오레세인, 153 mM의 AAPH, 상이한 농도(100 μM, 50 μM, 25 μM, 12.5 μM, 및 6.25 μM)의 Trolox 표준물, 및 블랭크로서 75 mM의 포스페이트 완충액이 포함되었다. 반응물을 다음 순서로 첨가하였다: 25 μL의 희석된 샘플; 어느 하나의 25 μL의 75 mM 포스페이트 완충액, 25 μL의 Trolox 표준물 및 150 μL의 플루오레세인. 플루오레세인을 첨가한 후, 플레이트를 37℃에서 10 분 동안 인큐베이션한 다음, AAPH(25 μL)를 첨가하였다. AAPH의 첨가 직후, 플레이트를 형광 플레이트 리더(BMG Labtech ClarioStar, 독일)에 넣고, 형광을, 원래 형광의 5% 미만으로 감소될 때까지, 3 분 마다 측정하였다. ORAC 값을 곡선하 면적(AUC)으로 계산하고, 브로콜리의 건조 중량 그램 당 trolox 당량(TE)의 마이크로몰(μmol TE/g DW)로 표현하였다. 각 샘플을 3 회 검정하였다.
설포라판 분석
브로콜리 기질로부터의 설포라판의 추출을 일부 변형으로 문헌[Li et al. (2012)]의 방법에 따라 실시하였다. 간략히 말해서, 냉동 브로콜리(2g)를 2 mL의 밀리-Q 워터와 혼합하고, 1 분 동안 볼텍싱하였다. 그 후에, 20 mL의 에틸 아세테이트를 슬러리에 첨가하고, 이어서, 5 분 동안 소니케이션(sonication) 및 4℃에서 20 분 동안 진탕을 수행하였다. 슬러리를 이후 10 분 동안 15,000 g으로 원심분리하고, 상청액을 수거하였다. 그 후에, 또 다른 15 mL의 에틸 아세테이트를 침전물에 첨가하여 2차 추출을 수행하였다. 각 샘플로부터 합한 추출물을 실온에서 진공 스핀 건조기(SC250EXP, Thermo Fisher Scientific, CA, USA)에 의해 건조 상태로 증발시키고, 분석까지 -20℃에서 저장하였다. 설포라판의 농도를 이성분 용매 전달 매니저 및 샘플 통이 장착된 Acquity™ 초고성능 LC 시스템(Waters Corporation, 밀퍼드, MA, USA)을 이용하여 결정하였다. 크로마토그래피 분리를 2.1 × 50 mm, Acquity BEH C18 크로마토그래피 컬럼에서 수행하였다. 이동상 A 및 B는 각각 밀리큐 워터 중 0.1% 포름산 및 아세토니트릴 중 0.1% 포름산이었다. 구배 용리 시스템은 이동상 A(밀리큐 워터 중 0.1% 포름산) 및 B(아세토니트릴 중 0.1% 포름산)로 이루어졌고, 다음 구배를 이용하여 분리를 달성하였다: 0-2 분, 10% B; 2-5 분, 20% B; 5-10 분, 10% B. 컬럼 온도를 30℃에서 일정하게 유지하였다. 유량은 0.350 mL/분이고, 주입 부피는 5 μL였다.
분석 전, 모든 샘플을 1 mL의 30% 아세토니트릴에 용해시키고, 0.22 μm 멤브레인 필터(Merk Millipore, 빌레리카, MA, USA)를 통해 여과하였다. 각 피크의 확인을 확실한 표준물의 머무름 시간 및 크로마토그래피를 기초로 하였다. 각 화합물의 농도를 표준 곡선에 따라 계산하고, 결과를 브로콜리의 DW 킬로그램 당 마이크로몰(μmol/kg DW)로 표현하였다.
글루코라파닌 분석
미가공 또는 발효 브로콜리로부터 글루코라파닌의 추출을 일부 변형으로 문헌[Cai and Wang (2016)]의 방법에 따라 수행하였다. 이에 따라서, 2 g의 냉동 브로콜리 퓨레에 10 mL의 끓는 밀리-Q 워터를 첨가하고, 혼합물을 끓는 워터 배쓰에서 5 분 동안 인큐베이션하였다. 이를 이후 냉각시키고, 15 분 동안 15000×g으로 원심분리하고, 상청액을 수거하였다. 침전물을 8 mL의 끓는 물로 1 회 넘게 추출하였다. 각 샘플로부터 합한 추출물을 3℃에서 진공 스핀 건조기(Speedvac SC250EXP, Thermo Fisher Scientific, CA, USA)에 의해 건조 상태로 증발시키고, 분석까지 -20℃에서 저장하였다. Photo Diode Array Detector 2998이 장착된 Alliance HPLC 기기(Waters Corporation, 밀퍼드, MA, USA)를 사용하여 글루코라파닌의 농도를 정량화하였다. HPLC 컬럼 - Luna® 3 μM 친수성 상호작용 액체 크로마토그래피(HILIC) 200° A(100×4.6 mm; Phenomenex, 토런스, CA, USA)를 25℃의 컬럼 온도에서 분석을 위해 사용하였다. 이동상은 다음 등용매 흐름 프로그램으로 30 mM의 암모늄 포메이트(용액 A) 및 아세토니트릴(용액 B)과 아세토니트릴/물(85:15, v/v)로 이루어졌다: 용액 A 70%; 용액 B 30%. 다른 크로마토그래피 조건에는 2.0 mL/ min의 일정한 유량, 100 μL의 주입 부피, 8 min의 이동 시간, 및 235 nm의 검출 파장이 포함되었다. 분석 전, 모든 샘플을 1 mL의 용매 A에 용해시키고, 0.22 μm 멤브레인 필터(Merk Millipore, 빌레리카, MA, USA)를 통해 여과하였다. 각 피크의 확인은 확실한 글루코라파닌 표준물의 머무름 시간 및 크로마토그래피를 기초로 하였다. 글루코라파닌의 농도를 표준 곡선을 이용하여 계산하고, 결과를 브로콜리의 DW 킬로그램 당 글루코라파닌의 마이크로몰(μmol/kg DW)로 표현하였다.
통계 분석
모든 실험을 3 회 실시하고, 결과를 평균 값으로 표현하였다. 0.05 유의 수준(p<0.05)에서 평균 값 간의 차이의 유의성을 평가하기 위해 일원 분산 분석(one-way analyses of variance: ANOVA)을 적용하였다. Windows용 통계 소프트웨어 SPSS 16.0(SPSS Inc., 시카고, IL, USA)을 사용하여 통계 분석을 실시하였다.
실시예 2 - 젖산균 발효 브로콜리 꽃송이의 미생물 분석
브로콜리 퓨레의 발효를 실시예 1의 발효 부문에 기재된 바와 같이 수행하였다. 총 젖산균의 수는 표 1에 나타나 있는 바와 같이 접종 브로콜리에 비해 미가공 브로콜리의 경우에 더 낮았다. 4 일의 발효 후, 샘플의 pH는 4.04에 도달하였고, 발효를 중지시키고, 저장 실험 전 발효 샘플을 0 일 샘플로 취하였다. 표 1 및 도 1c로부터, 0 일 샘플의 총 젖산균의 수는 미가공 브로콜리에 비해 유의하게 증가되었다(8 log CFU/g)는 것이 분명하다. 첫 2 주간의 저장 동안, 총 젖산균의 생존수는 25℃와 4℃ 둘 모두에서 저장된 샘플의 경우에 9 log CFU/g의 가장 높은 값으로 증가되었다(표 1 및 표 2). 25℃에서 저장 동안, 총 젖산균 수는 10 일에 9 log CFU/g로 증가되었고, 50 일까지 5 log CFU/g로 저장 동안 서서히 감소되고, 70 일 후에는 거의 검출 불능 수준으로 더 감소되었다. 대조적으로, 4℃에서 저장된 샘플에서 LAB 수는 84 일 동안의 저장 후에도 여전히 높았다(6 log CFU/g).
[표 1]
실온(25℃)에서의 저장 동안 발효 브로콜리의 미생물학적 및 물리화학적 변화
[표 2]
4℃에서의 저장 동안 발효 브로콜리의 미생물학적 및 물리화학적 변화
미가공 브로콜리 샘플에서 효모 및 곰팡이의 총 수는 2 log CFU/g이었다. 미가공 브로콜리에서 장내세균은 3 log CFU/g로 계수되었다. 진균, 곰팡이 및 장내세균은 발효 후에 또는 둘 모두의 온도 조건에서 저장 후 발효 샘플에서 검출되지 않았다. 발효 후 및 저장 동안 병원성 및 부패 유기체는 검출되지 않았다. 결과는, 발효 공정에 의해서, 검출 불능 수준의 잠재적으로 병원성인 장내세균과 및 부패 효모 및 곰팡이를 갖고, 4℃에서 저장될 때 고수준의 총 젖산균을 유지하는, 안전하고 안정한 프로덕트가 생성되었다는 것을 지시한다. 4℃에서 약 3 개월 후에는 약 106 CFU/g의 젖산균이 존재했다.
실시예 3 - 젖산균 발효 브로콜리 꽃송이의 저장 후 pH 및 적정 산도의 평가
미가공 브로콜리, 발효 브로콜리 및 25℃ 및 4℃에서 저장 후의 발효 브로콜리의 pH 및 적정 산도(TA)를 실시예 1에 기재된 바와 같이 분석하였다. TA의 결정을 이용하여 발효 동안 젖산균에 의해 생산되는 주요 산들인 젖산 및 아세트산의 양을 추정하였다. 발효 동안, 젖산균에 의해 생산된 산은 샘플의 pH를 감소시킨다. 표 1에 나타나 있는 바와 같이, TA는 0 일 샘플에서 10.7 g/L로 증가되었다. 25℃에서 저장될 때, 14.4 g/L에 도달한 유의하게 증가된 TA 값과 함께, pH는 10 일 후 저장 동안 3.87로 감소되었다(p<0.05; 표 1 참조). 결과는 저장 초기에 젖산균이 소비하는 기질이 여전히 존재하고 산을 추가로 생산한다는 것을 지시한다. pH 값이나 TA 값 어느 것도 남은 저장 기간 동안 유의하게 변하지 않았다(표 1).
4℃까지 온도를 감소시키는 것은 더 낮은 온도에서 젖산균의 활성이 감소하므로 저장된 샘플에서 pH 및 TA의 감소율을 저하시켰다(표 2 참조). 4℃에서 거의 3 개월간의 저장 후, pH는 3.85이고, TA 값은 13.7 g/L이었다.
실시예 4 - 설포라판으로의 글루코라파닌 전환에 대한 브로콜리 침연 및 발효의 평가
브로콜리 꽃송이를 작은 조각으로 자르고, 3:2의 브로콜리:물 비로 물과 혼합하고, 혼합물을 블렌더를 사용하여 퓨레로 침연시켰다. 퓨레 샘플(200 gm)을 멸균 플라스틱 병에 분취하였다. 샘플에 호주 브로콜리로부터 단리된 젖산균(류코노스톡 메센테로이데스 및 락토바실러스 플란타룸)의 합한 배양물을 108 CFU/gm 접종하였다. 샘플을 pH가 약 4.0으로 떨어질 때까지(4 일의 발효 후 달성됨) 30℃에서 유지되는 워터 배쓰에 인큐베이션하였다. 대조용 비-접종 샘플을 침연 후 즉시 냉동시켰다. 미생물 성장을 억제하기 위해 소듐 벤조에이트가 첨가된 두 번째 비-접종 대조 샘플 세트를 접종 샘플과 함께, 접종 샘플의 발효가 완료될 때까지, 30℃에서 4 일 동안 인큐베이션하였다. 실험을 3 회 실시하였다. 모든 샘플을 설포라판 및 글루코라파닌 분석까지 계속 냉동시켰다. 도 1b 및 표 3에 나타나 있는 바와 같이, 침연 그리고 이어서 발효는 단지 침연만 및 인큐베이션 단독에 비해 설포라판 수율을 증가시켰다.
[표 3]
브로콜리 퓨레에서 설포라판 함량에 대한 침연 및 발효의 효과
실시예 5 - 젖산균 발효된 브로콜리 꽃송이의 저장 후 총 단백질 함량 및 색의 평가
발효 후 젖산 발효된 브로콜리 꽃송이의 총 단백질 함량 및 색을 방법 부문에서 상술된 바와 같이 평가하였다. 미가공 브로콜리에 비해(26.9±0.03), 발효 브로콜리의 총 단백질 함량은 유의하게 증가되었다(29.6±0.8 mg/g; p<0.05). 이는 발효 동안 샘플에 접종된 많은 수의 젖산균 및 성장 및 젖산균에 의한 단백질 합성에 기인한 것일 수 있다. 총 단백질 함량은 샘플 간 유의한 차이 없이 25℃와 4℃ 둘 모두의 저장 동안 안정하게 유지되었다(표 1 및 표 2).
브로콜리 샘플의 색 값(L, a, b) 및 총 색 차이(△E)는 표 1 및 표 2에 요약되어 있다. 표 1 및 표 2에 제시된 바와 같이, 미가공 샘플과 발효 샘플 간의 색 파라미터 및 총 색 차이 값(△E)에서 유의한 차이가 기록되었다. L* 값(밝음)은 유의하게 변하지 않은 반면, a*(녹색도) 및 b*(황색도) 값은 브로콜리 퓨레의 발효 후에 감소되었다. a* 및 b* 값의 감소는 저 pH 하에 페오피틴으로 전환될 클로로필과 같은 유색 화합물의 분해에 기인할 수 있다. 0 일 샘플의 높은 △E 값(12.5)은 브로콜리 퓨레의 색이 발효 후에, 육안으로 눈에 띄게, 유의하게 변했다는 것을 지시한다. 저장 동안(표 1 및 표 2), 25℃ 샘플이나 4℃ 샘플 어느 것도 △E 값에 유의한 변화가 있지 않았다.
LAB+BP(브로콜리로부터 단리된 락토바실러스 플란타룸 B1, B2, B3, B4, B5 및 류코노스톡 메센테로이데스 BF1, BF2)로의 발효 후 브로콜리는 LAB(브로콜리로부터 단리된 락토바실러스 플라타룸(B1, B2, B3, B4, B5))로만 발효된 브로콜리에 비해 미가공 침연 브로콜리와 더 유사한 색으로 더 밝고 더 강렬한 녹색을 가졌다.
실시예 6 - 발효된 브로콜리 꽃송이에서 젖산균의 총 페놀 함량 및 항산화 활성의 변화
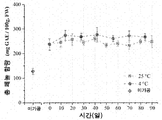
발효 후 젖산 발효된 브로콜리 꽃송이의 총 페놀 함량(TPC) 및 항산화 활성을 방법 부문에서 상술된 바와 같이 평가하였다. 미가공 브로콜리의 TPC는 127.6±12.4 mg GAE/100 g 생중량이었다(도 3a). 0 일에 TPC의 값은 미가공 브로콜리에 비해 236.9±23.4 mg GAE/100 g(p<0.05)로 유의하게 증가되었다. 저장 후 TPC는 25℃ 및 4℃에 저장된 샘플들 간에 유의한 차이가 없었다(도 3a). 25℃에서 저장될 때, 발효 브로콜리에서 TPC의 값은 10 일에 246.2±19.3 mg GAE/100 g, 및 90 일에 248.1±25.0 mg GAE/100 g이었다. 4℃에서 저장될 때, TPC의 값은 각각 14 일 및 84 일에 274.1±20.2 및 267.2±3.3 mg GAE/100 g이었다.
ORAC 값으로 표현된 샘플의 항산화 활성은 도 3b에 나타나 있다. 미가공 샘플의 ORAC 값은 110.1±0.05 μmol TE/g이었다. 발효는 미가공 브로콜리와 비교할 때 ORAC 값을 186.9±3.3 μmol TE/g로 약 70% 유의하게 증가시켰다. 이러한 결과는 항산화 화합물이 발효 동안 증가될 수 있다는 것을 시사했고, 발효 후 TPC의 변화와 일치했다.
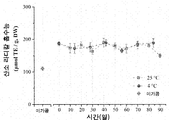
저장 동안, 발효 브로콜리의 항산화 활성은 유의하게 변하지 않았다. 도 3b에 나타나 있는 바와 같이, 25℃에서 저장될 때, 10 일 및 90 일에 ORAC의 값은 각각 173.0±14.4 및 150±5.5 μmol TE/g이었다. 유사한 결과가 4℃에서 저장된 샘플에 대하여 얻어졌다. ORAC 값은 저장 초기에 172.0±15.5 μmol TE/g이었는데, 이는 저장 후에 최대 값(188.7±12.9 μmol TE/g)으로 증가되었다.
실시예 7 - 상이한 조합의 젖산균에 대한 발효 시간의 평가
3:2의 브로콜리 대 물 비 및 1 분의 침연 시간으로 방법 부문에서 상술된 바와 같이 침연 브로콜리를 제조하였다. 브로콜리 재료에 LGG, LAB(호주 브로콜리로부터 단리된 락토바실러스 플란타룸(B1, B2, B3, B4, B5)), LAB+LP(브로콜리로부터 단리된 락토바실러스 플란타룸 및 락토바실러스 종 ATCC 8014), BP(브로콜리로부터 단리된 류코노스톡 메센테로이데스), LAB+BP(방법 부문에 기재된 바와 같은 두 그룹의 혼합) 중 하나를 107 CFU/g 또는 108 CFU/g 접종하고, 25℃, 30℃ 또는 34℃에서 4.4의 표적 pH에 도달하도록 발효시켰다. 도 4에 나타나 있는 바와 같이, 브로콜리 및/또는 브로콜리 퓨레로부터 단리된 젖산균의 첨가는 약 4 일 동안의 발효 후 4.4의 pH에 도달하여 LAB+BP의 조합으로 발효에 소요되는 시간을 유의하게 단축하였다. 발효 브로콜리 프로덕트의 예시적인 조성은 표 4에 나타나 있다.
[표 4]
발효 브로콜리 프로덕트의 조성.
실시예 8: 발효 브로콜리의 설포라판 함량에 대한 저장 효과
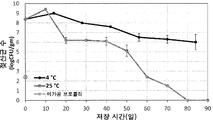
도 2a는 발효 브로콜리 퓨레의 설포라판 함량에 대한 4℃ 내지 25℃에서의 저장 효과를 나타낸 것이다. 도 2a에서 알 수 있는 바와 같이, 25℃에서 저장된 샘플의 설포라판 함량은 20 일 저장 후에 770.7±34.9 μmol/kg(52% 손실)로 급격히 감소되고, 이어서, 나머지 저장 기간 동안 더 서서히 하락하여, 총 69.5%의 손실에 이르렀다. 흥미롭게도, 4℃에서 발효된 브로콜리 샘플의 첫 2 주간의 저장 동안 설포라판 함량의 통계적으로 유의한 변화가 관찰되지 않았다. 약 23.7 %의 유의한 감소는 그 이후 2 주 동안 일어났고, 이어서 나머지 저장 기간 동안 서서히 저하되었다. 저장 말미에(84 일), 설포라판 함량은 4℃에서 저장된 샘플에서 1012.9±57.6 μmol/kg이라서, 총 설포라판 손실이 0 일 샘플에 비해 약 37.4 %가 되었다. 첫 2 주간의 저장 동안 설포라판 함량은 유지되었는데, 이는 아마도, 동일한 기간에 걸쳐 4℃ 저장 샘플에서 글루코라파닌 함량의 일부 감소가 관찰되었으므로 설포라판의 동시 생산 및 저하로 인한 것이다.
실시예 9: 글루코라파닌 함량에 대한 발효 및 저장의 효과
도 7은 4℃ 및 25℃에서의 저장 동안 글루코라파닌 함량 및 이의 안정성에 대한 침연 및 발효의 효과를 나타낸 것이다. 미가공 브로콜리의 글루코라파닌 함량은 3423.7±39.7 μmol/kg이었다(도 7). 발효 후, 글루코라파닌 함량은 712.4±64.2 μmol/kg로 급격히 감소되었다(0 일 샘플). 글루코라파닌은 온전한 조직에서 비교적 안정하고, 이러한 경우에 저하는 발효 동안 침연된 조직에서 증가된 효소-기질 상호작용으로 인해 미로시나제 촉매작용 가수분해에 기인할 수 있다. 글루코라파닌에서 급격한 감소의 기간은 발효 기간과 일치하였다.
25℃ 및 4℃에서의 저장 동안 발효 샘플에서는 글루코라파닌 함량에 유의한 변화가 관찰되지 않았다. 그러나, 25℃에서 저장된 샘플에서 약간 더 높은 글루코라파닌 함량이 관찰되었다. 이는 4℃에서 저장된 샘플(두 번째 시점에 pH 4.04)에 비해 25℃에서 저장된 샘플의 더 빠른 pH 하락(두 번째 시점에 pH 3.87)과 관련이 있을 수 있다. 글루코라파닌의 미로시나제 촉매작용 가수분해를 위한 최적의 pH는 5 내지 6의 범위이고, pH 3.0에서 최저치로 감소한다[Dosz & Jeffery, 2013]. 4℃에서 저장된 샘플의 비교적 더 높은 pH는 25℃에 비해 4℃에서의 저장 동안 글루코라파닌의 약간 더 큰 저하에 기여했을 수 있다.
실시예 10 - 브로콜리 기질에서 설포라판으로의 글루코라파닌 전환을 최대화하기 위한 열 처리 조건의 평가
레토르트 파우치에 패킹된 브로콜리 꽃송이를 60℃ 내지 80℃ 범위의 온도 및 0 분 내지 5 분의 처리 시간으로 열 가공에 주어지게 하였다. 이러한 처리에는 실험 온도보다 5℃ 더 높게 유지되는 워터 배쓰 내 실험 온도로의 예열, 그리고 이어서 실험 온도에서 유지되는 제2 워터 배쓰에서의 인큐베이션을 수반하였다. 열 처리 후, 샘플을 얼음-물에서 냉각시키고, 상술된 바와 같이 2:3의 물 대 브로콜리 비로 첨가된 물로 침연시켰다. 침연 샘플을 30℃에서 1 시간 동안 인큐베이션하고, 설포라판 분석까지 계속 냉동시켰다. 결과는 도 2b 및 표 5에 나타나 있다. 표 5에 나타나 있는 바와 같이, 60℃, 65℃ 또는 80℃에서의 샘플 예열 및 이어서 침연은 예열 없이 침연된 미가공 브로콜리 꽃송이에 비해 설포라판 수율을 증가시켰다.
[표 5]
브로콜리 기질에서 설포라판 생산에 대한 열 처리 효과
실시예 11 - 브로콜리의 설포라판 함량에 대한 젖산균 발효 전 예열의 평가
이 연구는 브로콜리 퓨레의 설포라판 함량에 대한 젖산균과 조합된 에피티오스페시피어 단백질(ESP)을 불활성화시키는 브로콜리 꽃송이의 약한 예열 처리의 영향을 평가하는 것이었다.
재료
브로콜리(재배종 '바이페르(Viper)')를 현지 슈퍼마켓(Coles, 남부 웨리비, VIC, 호주)으로부터 구매하였다. 드만-로고사-샤르프(MRS) 브로스(1823477, CM0359, Oxoid)를 Thermo Fisher Scientific(호주)으로부터 구매하였다. DL-설포라판을 Sigma-Aldrich(St. Louis, 미주리, USA)로부터 구매하였다. 모든 다른 화학물질 및 생화학 시약은 분석용 등급 이상이었고, 현지 화학물질 벤더로부터 구매하였다.
설포라판 수율을 최대화하도록 약한 예열 조건을 최적화시키는 실험
브로콜리 꽃송이를 헤드(head) 아래로 대략 2 cm 자르고, 각각 30g의 무작위 혼합된 브로콜리 꽃송이를 예열 실험에서 사용하였다. 두 유형의 예열 실험을 실시하였다: 인-팩 가공(in-pack processing) 및 직접적인 물 블랜칭(direct water blanching). 인-팩 실험의 경우에, 브로콜리 꽃송이를 레토르트 파우치(Caspak Australia, 멜버른)에 패킹하고, 밀봉하고, 60℃, 65℃ 및 80℃에서 유지되는 온도 조절 워터 배치에서 다양한 시점에 예열하였다. 온도계를 사용함으로써 가장 느린 가열점에서 브로콜리 샘플의 온도를 측정하였다. 시간 0는 심부 온도가 지정된 실험 온도에 도달하는 시간으로 규정되었다. 처리 시간은 60℃ 및 65℃의 경우 0 분, 1 분, 3 분, 및 5 분이고, 80℃의 경우 0 분, 1 분, 2 분, 3 분이었다. 직접적인 물-블랜칭 실험의 경우, 브로콜리 꽃송이를 온도 조절 워터-배쓰에서 가열된 유리 비커 안의 밀리-Q 워터에 담갔다. 직접적인 물 블랜칭 실험을 60℃ 및 65℃에서 실시하였다. 브로콜리 샘플의 온도를 온도계를 사용하여 연속적으로 측정하고, 가장 느린 가열점에서의 온도가 상술된 바와 같이 지정된 실험 온도에 이르면 타이밍을 시작하였다. 모든 열 처리 실험을 3 회 수행하였다. 비가열 브로콜리 꽃송이를 대조로서 사용하였다. 열 처리 직후, 샘플을 얼음 물에서 냉각시키고, 주방 저울 매직 불렛 블렌더(Nutribullet pro 900 시리즈, LLC, USA)를 사용하여 밀리-Q 워터로 3 부의 브로콜리 대 2 부의 물의 비로 1 분 동안 균질화시켰다. 글루코라파닌의 효소 가수분해가 가능하도록 균질화된 샘플을 25℃에서 4 시간 동안 암흑에서 인큐베이션하였다. 인큐베이션 후, 모든 샘플을 설포라판 분석까지 -20℃에서 냉동시켰다.
스타터 배양물의 제조
실시예 1의 방법에 기재된 바와 같이 브로콜리로부터 단리된 류코노스톡 메센테로이데스(BF1, BF2) 및 락토바실러스 플란타룸(B1, B2, B3, B4, B5)의 합한 배양물을 발효 실험에 사용하였다. -80℃에서 저장된 젖산균 저장 배양물을 10 mL의 MRS 브로스(Oxoid, 빅토리아, 호주)에 접종하고, 30℃에서 24 시간 동안 인큐베이션함으로써 활성화시켜 일차 접종원을 수득하였다. 2 mL의 일차 배양물을 200 mL의 MRS 브로스에 접종하여 이차 배양물을 수득하였다. 24 시간 접종 후, 6 개의 이차 배양물을 원심분리하고, 멸균 완충 식염수(PBS)로 2회 세척하고, 각각의 배양물을 밀리-Q 워터에 10 log 집락-형성 단위/밀리리터(CFU/mL)의 농도로 재현탁시켜 100 gm의 브로콜리 퓨레 샘플에서 8 log CFU/mL의 초기 바이오매스를 수득하였다. L. 플란타룸 배양물을 L. 메센테로이데스 배양물과 1:1 비율로 혼합한 후, 브로콜리 퓨레 샘플에 접종하였다.
샘플 제조
브로콜리 꽃송이를 크라운 아래로 대략 2 cm 자르고, 두 개의 로트로 분리하였다: 열 처리 및 비-처리. 상술된 바와 같은 실험의 결과를 기초로 하여 선택된 최적의 조건에서의 열 처리 후, 샘플을 얼음-물에 냉각시키고, 잘게 썰고, 주방 저울 매직 불렛 블렌더(Nutribullet pro 900 시리즈, LLC, USA)를 사용하여 밀리-Q 워터와 3:2의 비로 1 분 동안 균질화시켰다. 비-처리 브로콜리를 또한 유사한 방식으로 균질화시켰다. 브로콜리 퓨레를, 잘 혼합한 후, 추가 실험을 위해 나사형 뚜껑(Technoplast Australia)이 있는 멸균 플라스틱 용기(100 mL)에 분취하였다.
발효
브로콜리 퓨레 샘플(예열 및 비처리)에 본 실시예에 상술된 바와 같이 제조된 LAB 배양물을 접종하였다. 브로콜리 꽃송이의 예열을 예열 조건을 최적화시키는 실험 결과에 기초하여 65℃에서 3 분 동안 인-팩으로 실시하였다. 설포라판으로의 글루코라파닌 전환에 대한 발효가 없는 산성화의 영향을 평가하기 위해, 발효 브로콜리 퓨레의 pH에 이르도록 글루코노-델타-락톤(GDL)을 사용하여 예열 및 비처리 브로콜리 퓨레에 대하여 산성화 실험을 실시하였다. 예열 브로콜리 퓨레 및 추가 처리가 없는 비처리 브로콜리 퓨레를 대조로서 사용하였다.
발효 실험을 위해, 각 브로콜리 퓨레 샘플에 8 log CFU/g의 초기 수준으로 제조된 스타터 배양물을 접종하였다. 15 시간의 인큐베이션 후 pH가 약 4.0에 도달할 때까지 30℃에서 발효 실험을 수행하였다. 발효가 완료되면, 각 발효 그룹의 3 개의 샘플(0 일 샘플)을 취하고, 분석까지 -20℃에서 저장하였다. 나머지 발효물을 저장 시험을 위한 두 개의 로트로 무작위 분리하였다: 14 일 저장 후 샘플의 설포라판 안정성의 평가를 위해, 하나의 로트는 냉장 조건(4℃) 하에 저장하고, 두 번째 로트는 25℃에서 저장하였다. 유사하게, 비처리 브로콜리 퓨레, 예열 브로콜리 퓨레 및 예열-GDL 처리 브로콜리 퓨레를 또한 시간 0(zero)에 샘플링하고, 14 일 저장 시험을 위해 25 및 4℃에서 저장하였다. 14 일 저장 후, 설포라판 분석까지 모든 샘플을 냉동시키고, -20℃에서 유지하였다.
설포라판 분석 및 통계 분석
실시예 1에 기재된 바와 같이 수행하였다.
설포라판 수율을 개선하기 위한 열 처리 조건의 최적화
세 개의 상이한 온도(60℃, 65℃ 및 80℃)에서 다양한 가공 시간(60℃ 또는 65℃의 경우 0 분, 1 분, 3 분 및 5 분; 80℃의 경우 0 분, 1 분, 2 분 및 3 분) 동안 가열-인-팩 브로콜리 꽃송이의 설포라판 형성에 대한 열 처리의 영향은 도 5a에 나타나 있다. 결과는 미가공 브로콜리에 비해 모든 열 처리 샘플에서 설포라판 수율이 증가되었다는 것을 나타냈다. 시간 0은 이들의 심부가 실험 온도에 도달할 때까지 가열된 샘플을 지정한다.
도 5a에 나타나 있는 바와 같이, 패킹된 브로콜리 샘플을 60℃에서 0 분, 1 분, 3 분, 및 5 분 동안 가열할 때 설포라판 수율의 증가가 일어났다. 이러한 샘플들에서 설포라판의 농도는 각각 2343.5±124.1, 2661.5±10.9, 2780.9±270.8, 및 3147.7±148.0 μmol/kg DW였다. 다른 한 편으로, 브로콜리를 65℃에서 가공했을 때, 설포라판 수율은 초기에 가공 시간에 따라 3585.9±119.2(0 분)에서 3983.4±30.5 μmol/kg DW(3 분)의 최고치로 증가했다. 처리 시간의 추가 증가는 5 분 처리 시간 후 관찰된 3620.1±240.7 μmol/kg의 최저치로 더 낮은 수율을 초래하였다. 60 및 65℃에서의 처리와 대조적으로, 80℃에서 가공한 샘플의 경우, 각각 0 분, 1 분, 2 분 및 3 분 처리 후에 1451.5±43.5, 1446.8±17.5, 1043.1±94.2, 및 981.2±35.1 μmol/kg DW의 설포라판 함량으로, 처리 시간이 길어짐에 따라 설포라판 수율의 지속적인 감소가 관찰되었다. 전체적으로, 브로콜리의 인-팩 처리에 대한 설포라판의 최고 수율(3983.4 ± 30.5 μmol/kg)은 65℃에서 3 분 동안 예열된 샘플에 대해 얻어졌으며, 이는 미가공 브로콜리(817.5 ± 9.3 μmol/kg DW)보다 약 5 배 더 높았다. 대조적으로, 물에서 직접적으로 브로콜리를 가열하는 것은, 일반적으로 도 5b에 나타나 있는 바와 같이, 인-팩 가공에 비해 더 낮은 설포라판 수율을 초래하였다. 60℃에서의 직접적인 물 블랜칭을 위해, 설포라판 수율은 처리 시간에 따라 1698.00±121.9 μmol/kg DW(0 분)에서부터 2833.3±118.6 μmol/kg DW(1 분)로 증가되고, 그 후에 60℃에서 5 분 처리의 경우 2345.8±57.7 μmol/kg DW의 최저치로 지속적으로 감소되었다. 60℃에 비해 설포라판 수율 급격한 하락은 샘플을 65℃에서 블랜칭할 때 관찰되었다. 설포라판 수율은 65℃에서 5 분 열 처리 후 503.7±23.8 μmol/kg DW의 브로콜리였는데, 이는 미가공 브로콜리에 대해 얻어진 값보다 훨씬 더 낮은 것이었다. 그 이유는 블랜칭 물로의 글루코라파닌 침출이 설포라판의 낮은 수율을 초래했기 때문일 수 있다. 직접적인 물 블랜칭을 위해, 설포라판 수율을 최적화시키기 위한 최적의 처리 온도는 인-팩 가공의 경우 65℃에 비해 60℃였다.
이러한 연구에서, 설포라판의 최고 수율은 65℃에서 3 분 동안 인-팩 가공된 브로콜리 꽃송이에 대해 얻어졌는데, 이는 상기 조건이 충분한 미로시나제 활성을 유지하면서 ESP의 불활성을 큰 정도로 유리하게 하여 설포라판으로의 최적의 수율을 야기한다는 것을 지시한다. 이러한 조건 하에, 브로콜리 샘플의 글루코라파닌 함량은 3423.7±39.7 μmol/kg DW인 것으로 결정되었으므로, 대부분의 추출 가능한 글루코라파닌이 설포라판으로 전환된 것으로 보여 1 대 1 전환이 추정된다.
발효에 대한 열-처리된 브로콜리의 노출이 미가공 브로콜리로부터 추출 가능한 글루코라파닌의 수준에서 예측되는 것보다 높은 수준의 설포라판을 야기한다는 관찰은 열-처리가 미로시나제에 대한 글루코라파닌의 접근성을 증가시켜 비처리 브로콜리에 존재하는 글루코라파닌의 정량화 가능한 양을 기준으로 예상되는 수율보다 높은 설포라판 수율을 야기한다는 것을 시사한다.
더 낮은 설포라판 수율은 물에서 직접적으로 블랜칭된 브로콜리 꽃송이의 경우에 얻어졌는데, 이는 가장 개연성 있게는, 글루코라파닌이 물에 가용성이라 블랜칭 물로 침출하기 때문이다. 또한, 설포라판의 최대량이, 브로콜리 꽃송이의 열 처리를 인-팩으로 수행할 때 65℃에서 3 분에 비해, 브로콜리 꽃송이를 물에서 직접적으로 가열할 때 60℃에서 1 분 동안 가열함으로써 얻어졌다는 것을 주지하는 것도 흥미롭다. 이는 65℃에서의 더 높은 수준의 ESP 불활성화 효과를 상쇄한 65℃에서의 블랜칭 물로의 더 높은 침출율 때문일 수 있다.
설포라판 수율에 대한 LAB 발효 및 화학적 산성화의 효과
브로콜리 꽃송이를 상기에서 선택된 가장 우수한 처리 조건에서 인-팩으로 예열하였다(65℃, 3 분). 샘플을 이후 젖산균에 의해 발효시키거나 산미료(GDL)를 사용하여 산성화시켰다. 전처리 실험과 일치하여, 브로콜리의 설포라판 값은, 각각 미가공 및 예열 브로콜리의 경우에 806.2±7.0 μmol/kg DW 및 3536.0±136.9 μmol/kg DW의 설포라판 수율로, 열 처리 후에 유의하게 증가되었다(p<0.05). 발효 전 예열된 이러한 개별 브로콜리 배치로 얻어진 3536 μmol/kg DW의 값은 상이한 브로콜리 배치를 이용할 때 얻어진 것과 동일한 순서였고, 여기서 3983 μmol/kg DW가 얻어져 배치마다 약간의 편차를 나타냈다.
표 6에 나타나 있는 바와 같이, 발효 후, 브로콜리 샘플의 설포라판 함량은 발효 전 브로콜리의 처리에 좌우하여 달라졌다. 발효 후 미가공 브로콜리 퓨레의 설포라판 함량(1617.4±10.2 μmol/kg DW)은 미가공 브로콜리 퓨레의 설포라판 함량의 대략 두 배였다. 퓨레화 전 브로콜리의 예열은 발효 후 설포라판 함량의 훨씬 더 높은 증가를 야기하였다. 예열-발효 브로콜리의 설포라판 함량(13121.3±440.8 μmol/kg DW)은 미가공-발효 브로콜리 퓨레의 약 8 배였다. 조합된 예열-발효 처리 후 관찰된 설포라판 수율은 미가공 브로콜리 샘플에서 글루코라파닌의 정량화 가능한 양(3423.7±39.7 μmol/kg)을 기초로 예상되는 것보다 훨씬 더 높았다. 조합된 예열 및 발효 공정은 예열 공정에 의한 ESP의 불활성화에 더하여 전환을 위한 글루코라파닌의 방출 및 접근성을 향상시키는 것으로 보인다. 미생물 세포 벽 분해 효소와 결합된 예열 공정은, 미가공 브로콜리에서 추출 가능하거나 접근 가능하지 않았던, 기질에 결합된 글루코시놀레이트의 방출 및 세포 구획의 파괴를 향상시켰을 수 있다. 일부 젖산 균주는 세포 벽 구조를 분해하고 벽 결합 성분의 방출을 향상시키는 것이 가능한 셀룰라제 및 펙티나제와 같은 다당류 분해 효소를 생산한다.
대조적으로, GDL에 의한 예열 브로콜리 퓨레의 화학적 산성화는 예열 및 예열-발효 샘플에 비해 유의하게 더 낮은(p<0.05) 설포라판 함량을 초래하였다(표 6). GDL 산성화 샘플의 설포라판 함량은 예열 브로콜리 샘플(3536.0±136.9 μmol/kg DW)보다 40% 더 낮은 2169.4±176.0 μmol/kg DW였다(P<0.05). 산성화 동안 pH 4.04로의 빠른 감소가 GDL 샘플에서 설포라판으로의 글루코라파닌 전환을 감소시켰을 수 있는 것으로 보인다. 글루코시놀레이트의 전환이 pH에 매우 의존적이며, 산성 pH가 니트릴로의 전환을 유리하게 한다는 것은 잘 알려져 있다[Latte et al., 2011].
예열 발효 샘플의 경우에, 산성화는 > 15 시간의 기간에 걸쳐 점진적으로 일어나서 주로 설포라판으로의 글루코라파닌 전환을 가능하게 하는데, 그 이유는 ESP의 활성이 65℃에서 3 분 동안 예열한 후 유의하게 감소될 것으로 예상되기 때문이다.
저장 동안 설포라판 함량의 변화
모든 샘플의 설포라판의 농도는 25℃에서 14 일 저장 후에 하락하였다(표 6 및 도 6 참조). 흥미롭게도, 설포라판 함량의 증가가 4℃에서 14 일 저장 동안 발효 샘플을 제외한 모든 샘플에서 관찰되었다. 미가공 퓨레의 설포라판 함량은 4℃에서의 저장 동안 거의 두 배가 되었다. 유사하게, 예열 샘플의 설포라판 함량은 약 2.6 배 증가된 반면, 예열 GDL 샘플의 설포라판 함량은 약 2.3 배 증가되었는데, 이는 저장 동안 기질로부터 글루코라파닌의 연속 방출이 설포라판으로의 추가 전환 및 농도 증가를 가능하게 하여 저장 동안 설포라판 저하의 결과를 상쇄시킨다는 것을 시사한다. 예열-발효 샘플과 관련하여, 설포라판 함량의 감소가 둘 모두의 온도에서의 저장 동안 관찰되었다. 모든 접근 가능한 글루코라파닌은 발효 동안 너무 많이 설포라판으로 전환되어 저장 동안 추가 전환이 일어나지 않은 것이 아니라, 오히려 온도에 좌우하여 상이한 정도로 차이가 있지만 저하된 것일 수 있다. 이와 같이, 4℃에서의 저장 동안 단지 약간의 하락(약 6%)만이 관찰된 반면, 25℃에서 저장 동안 하락은 약 70%였다.
이러한 연구는 젖산균 발효와 결합된 예열이 브로콜리 기반 프로덕트의 설포라판 함량을 상당히 향상시킨다는 것을 보여주었다. 65℃에서 3 분 동안 브로콜리 꽃송이의 인-팩 예열 처리, 그리고 이어서 침연 및 발효는 미가공 브로콜리 퓨레에 비해 약 16 배 만큼 더 높은 설포라판 수율을 야기하였다. 이러한 조건 하에 예열은 브로콜리 퓨레에서 설포라판 수율을 비처리 브로콜리에서의 806 μmol/KgDW(건조 중량)에서부터 3536 μmol/KgDW로 증가시켰는데, 이는 처리가 설포라판으로의 글루코라파닌 전환에 충분한 미로시나제 활성을 유지하면서 ESP를 상당히 억제한다는 것을 지시한다. 직접적인 물 블랜칭 동안 가장 우수한 예열 조건은 60℃에서 1 분이었고, 2833 μmol/KgDW의 설포라판 수율을 야기하였다. 직접 블랜칭 동안 더 낮은 수율은 블랜칭 매질로의 수용성 글로코라파닌 침출에 기인할 수 있다. 젖산균 발효와 조합된 브로콜리 꽃송이 인-팩의 예열(65℃/3 분)은 13121 μmol/KgDW로 설포라판 함량을 더 향상시켰는데, 이는 미가공 브로콜리에 비해 약 16 배 증가한 것이다. 글루코노-델타-락톤에 의한 브로콜리 퓨레의 산성화와 조합된 인-팩 예열(65℃, 3 분)의 화학적 산성화는, 단독의 예열보다 낮은, 2169 μmol/KgDW의 설포라판 수율을 초래하였다. 예열-발효 퓨레의 설포라판 함량은 4℃에서 2 주 저장 동안 안정하게 유지되었다(약 94% 보유).
[표 6]
가공 전 및 후 브로콜리의 설포라판 수율(μmol/Kg DW)
실시예 12 - 브로콜리의 다가페놀 프로파일에 대한 젖산균 발효의 효과
브로콜리 샘플의 다가페놀 대사물에 대한 발효의 효과를 알아내기 위해, 미가공 및 발효 브로콜리 퓨레 샘플의 표적화 액체 크로마토그래피-질량 분석법(LC-MS) 기반 대사체 분석을 실시하였다. 얻어진 다변수 데이터를 Metaboanalyst 소프트웨어(Metaboanalyst 3.0, Xia and Wishart, 2016)를 사용하여 분석하였다. 발효는 브로콜리 샘플의 대사물 프로파일에 유의한 변화를 야기하였다. 데이터의 부분 최소 제곱 판별 분석(partial least square discriminant analysis: PLS-DA)은 발효 및 비-발효 샘플의 다가페놀 프로파일 간의 분명한 차이를 보여준다(도 8).
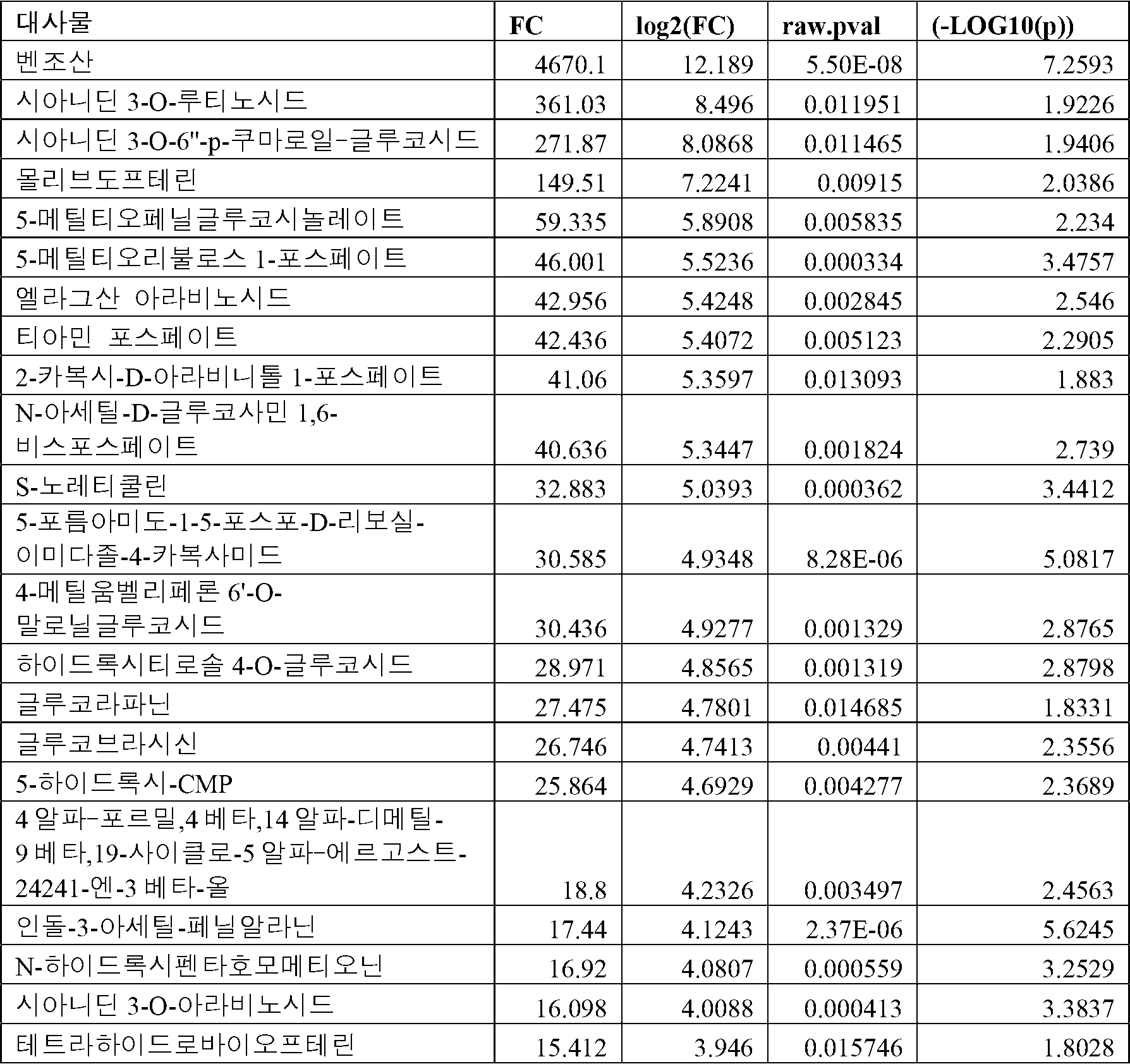
두 그룹 간 차이에 원인이 되는 것으로 확인된 상위 15 개의 대사물은 도 9에 나타나 있다. 이들은 페놀산 및 페놀 아글리콘이었는데, 생체활성 및 생체이용률이 이들의 페놀산 에스테르 및 페놀산 글리코시드 전구체에 비해 더 높았다. 대부분의 이러한 대사물들의 농도가 발효 후 상당한 증가를 나타냈는데, 이는 브로콜리 퓨레의 다가페놀 프로파일에 대한 발효의 유익한 효과를 지시하는 것이다. 일부 대사물에 대한 배수 변화는 표 7에 나타나 있다.
각각 24 배 및 16 배의 시나프산 및 캠페롤에서 상당한 증가가 발효 후에 관찰되었다. 유사하게, 발효는 클로로겐산 및 페닐락트산에서 8 배 증가를 유도하였다. 헤스페레틴, 퀘르세틴, 메틸 시린게이트 및 시링산의 농도가 또한 발효 후에 상당히 증가되었다. 캠페롤, 헤스페레틴 및 퀘르세틴과 같은 아글리콘의 농도 증가는 미생물 글리코시다제의 활성에 의해 이들의 글리코시드 전구체의 전환에 기여할 수 있다. 시나프산과 같은 페놀산의 농도 증가는 미생물 에스테라제의 활성에 의한 브로콜리에서 페놀산 에스테르의 전환으로 인한 것일 수 있다. 카페산 및 갈산에서 약간의 감소가 발효 후에 관찰되었다. 미생물 데카복실라제의 활성은 카페산을 상응하는 비닐 카테콜로 및 갈산을 피르갈롤로 전환시키는데, 이는 이들의 농도 감소에 원인이 될 수 있다[Filanino et al., 2015; Guzman-Lopez et al., 2009].
[표 7]
발효 및 비-발효 브로콜리 퓨레 간 차이의 원인이 되는 상위 13 개의 다가페놀에서의 배수 변화
실시예 13 - 샘플의 표적화 및 비표적화 LC_MS 분석에 의한 브로콜리의 젖산균 발효로 생산된 대사물의 확인
발효 및 비-발효 브로콜리 퓨레 샘플을 냉동시키고, 냉동 건조시켰다. 내부 표준물로서 100 mg/ml의 카페인을 포함하여, 1 ml의 얼음-냉각 메탄올 및 밀리-Q 워터(50:50, v:v)를 사용함으로써 샘플(각각 100 mg의 냉동 건조된 분말)을 추출하였다. 샘플을 이후 2 분 동안 볼텍싱한 다음, 30 분 동안 소니케이션(40 Hz)하였다. 그 후에, 샘플을 4℃에서 30 분 동안 20,000 rpm으로 원심분리하고, 상청액을 청결한 실란화된 LC-MS 바이알로 옮겼다. 샘플을 Agilent 6410 LC-QQQ HPLC(Agilent Technologies, 산타클라라, 캘리포니아, USA)에 1.4 μl 주입함으로써 분석하였다. 30℃의 컬럼 온도 및 0.3 ml/min의 유량으로 역상 Agilent Zorbax Eclipse Plus C18, 고속 분해 HD, 2.1 × 50 mm, 1.8 um(Agilent Technologies, 산타클라라, 캘리포니아, USA)를 사용하여 분석을 수행하였다. 이동상을 등용매로 1 분 동안 95:5(A:B) 이후 추가 12 분 동안 1:99(A:B)로 바꾼 후, 다시 추가 2 분 동안 95:5(A:B)로 되돌려 작동시켜, 15 분의 총 이동 시간을 제공하였다. 이동상 'A'는 100%의 H2O 및 0.1%의 포름산으로 이루어졌고, 이동상 'B'는 75%의 아세토니트릴, 25%의 이소프로판올 및 0.1%의 포름산을 함유하였다. MS는 50 m/z 내지 1000 m/z의 질량 범위에서 데이터를 수집하였다. Massbank 및 METLIN을 포함하여 여러 온라인 LC-MS 대사체 데이터베이스를 사용하여 대사체학 표준 계획(Metabolomics Standard Initiative: MSI) 화학 분석 워크그룹에 따라 화합물의 정성 확인을 수행하였다. 전반적으로, 기기 조건은 양성 전기분무(+ESI)와 음성 전기분무(-ESI) 모드 둘 모두에 대하여 유사했다. 스캔 시간은 500이고, 소스 온도는 350℃에서 유지되고, 가스 유량은 12 L/min이고, 네뷸라이져 압력은 35 psi였다.
비표적화 분석에서 화합물의 확인을 위해, 기준을 >90%의 일치율로 설정하였다. 일치율이 70% 내지 89%로 떨어지는 경우, 화합물을 괄호로 식별하였다(예를 들어, 화합물이 70% 내지 89%인 경우, 이들을 "<name>"으로 하여 주석을 달았다). 70% 미만의 임의의 일치율을 제거하였다. 총, 약 1000 개 내지 1500 개의 파쳐(fature)가 확인되었다: 다수가 잘 일치하지 않거나(그래서 제거됨) 기준으로부터 10 × S/N 비율 미만이었다. 이와 같이, 사용된 화합물/피크는 실제 피크였고, ID는 상당히 강했다(즉, >70%).
비표적화 LC-MS 대사체학 연구는 안토시아닌 글리코시드, 페놀산 글리코시드, 페놀산을 포함하여 특정 다가페놀 글루코시드에서 2 배 내지 360 배 증가, 글루코라파닌의 27 배 증가와 함께 일부 글루코시놀레이트에서 5 배 내지 60 배 증가 및 인돌-3카비놀 및 아스코르비겐에서 약 3 배 내지 4 배 증가를 나타냈다. 결과는 표 8에 요약되어 있고, 도 10에 그리고 도 11의 볼카노 플롯에 나타나 있다. 발효 후 증가된 상위 50 개의 대사물은 여러 다가페놀 글리코시드 및 글루코시놀레이트를 포함하는데, 이는 공정이 이들의 추출성 및 생체접근성을 향상시킨다는 것을 지시한다.
[표 8]
비표적화 LC-MS 분석에 기초한 발효 브로콜리 퓨레와 비-발효 브로콜리 퓨레 간 상이한 대사물의 배수 변화
브로콜리 샘플의 다가페놀 대사물에 대한 발효의 효과를 알아내기 위해, 미가공 및 발효 브로콜리 퓨레 샘플의 표적화 액체 크로마토그래피-질량 분석법(LC-MS) 기반 대사체 분석을 실시하였다. 통계 분석을 전처리 없이 수행하였다. 발효는 브로콜리 샘플의 대사물 프로파일에 유의한 변화를 야기하였다.
표적화 LC-MS 분석에서, 다가페놀 표준물을 대사물의 확인 및 정량화에 사용하였다. 클로로겐산, 페룰산, 시링산, 페닐락트산, 루틴, 시나프산, 메틸 시린게이트, 헤스페레틴, 퀘르세틴 및 캠페롤의 증가를 발효 브로콜리에서 확인하였다(도 12). 프로토카테츄산, 갈산, 4,하이드록시벤조산, 바닐산, 2,3디하이드록시벤조산, p-쿠마르산, 신남산, 카테킨, 로즈마린산, 카페산의 감소를 발효 브로콜리에서 확인하였다(도 12). 주목할 것은 클로로겐산에서 6.6 배수 변화(2.4 μg/mg에서 15.8 μg/mg으로), 시나프산에서 23.8 배수 증가(3.6 μg/mg에서 86.6 μg/mg으로), 캠페롤에서 10.5 증가(12.7 μg/mg에서 134.6 μg/mg으로) 및 p-쿠마르산에서 0.48 배수 변화가 발효 샘플에서 일어났다는 것이다(도 12).
실시예 14 - 의도적으로 도입된 미생물의 성장을 억제하는 브로콜리 발효 배양물의 평가
식품 제조에서 흔히 관찰되고 우려되는 의도적으로 도입된 미생물의 성장을 억제하는 브로콜리 발효 배양물의 능력을 평가하기 위해 과제 연구를 실시하였다.
실험실 배양/스타터 배양
B1, B2, B3, B4, B5, BF1 및 BF2를 포함하는 1010 cfu/mL의 접종원 10 ml로 발효물에서 108 CFU/gm의 샘플을 달성하였다.
병원체 배양
E. 콜라이 단리물 FSAW 1310, FSAW 1311, FSAW 1312, FSAW 1313 및 FSAW 1314를 정치시켜 37℃에서 밤새 NB(영양 액체배지)에서 1 내지 4 × 108 cfu/mL로 개별적으로 성장시켰다. 배양물을 합하고(각각 1 mL), 합한 배양물을 처음 두 희석을 위해 MRD(최대 회수 희석제) 및 마지막 두 희석을 위해 물에 의해 104로 희석하였다.
살모넬라 균주 S. 인판티스 1023, S. 신가포레 1234, S. 타이피무리움 1657 (PT135), S. 타이피무리움 1013 (PT9) 및 S. 비르초우 1563을 정치시켜 37℃에서 밤새 NB에서 1 내지 4 × 108 cfu/mL로 개별적으로 성장시켰다. 배양물을 합하고(각각 1 mL), 합한 배양물을 처음 두 희석을 위해 MRD 및 마지막 두 희석을 위해 물에 의해 104로 희석하였다.
리스테리아 단리물 Lm2987(7497), Lm2965(7475), Lm2939(7449), Lm2994(7537) 및 Lm2619(7514)를 교반 하에 37℃에서 밤새 10 mL의 BHI(뇌심장즙 액체배지(brain heart infusion broth))에서 개별적으로 성장시켰다. 모든 배양물을 이후 합하고(각각 1 mL), 이러한 칵테일을 처음 두 (1/10) 희석을 위해 MRD 및 마지막 두 희석을 위해 멸균 탈이온수를 사용하여 희석하였다.
B. 세루스 포자 채취물을 단리물 B3078, B2603, 2601, 7571 및 7626으로부터 제조하였다.
방법
접종원을 제조하기 전에, 3:2의 브로콜리:멸균 수돗물(900 g의 브로콜리:600 g의 물)로 브로콜리 퓨레를 제조하였다. 브로콜리 헤드를 수돗물에서 헹구고, 80% 에탄올로 살균된 절단 보드 상에서 멸균 칼로 브로콜리에서 줄기를 잘랐다. 브로콜리 꽃송이(900 g)를 작은 조각으로 잘랐다. 450 g의 브로콜리 조각을 총 600 g의 물이 있는 써모믹스(Thermomix) 볼에 넣었다. 반투명 써모믹스 컵/뚜껑을 80% 에탄올로 살균하고, 뚜껑 구멍 위에 두었다. 브로콜리를 1 분 동안 속도 4에서 다졌다. 두 번째 450 g의 브로콜리 조각을 써모믹스 볼에 첨가하고, 1 분 동안 속도 4로 다졌다. 내용물을 속도 10(최대)에서 추가 5 분 동안 다졌다. 퓨레가 실제로 충분히 부드러워졌는지 확인한 후, 써모믹스 볼을 서늘한 공간에 두어 내용물을 30 분 동안 냉각시켰다. 이후, 볼을 인큐베이터에 넣고, 30℃로 평형화시켰다. 그 동안, 스타터 배양물 및 병원체 배양물(E. 콜라이, B. 세레우스, 살모넬라, 리스테리아 모노사이토게네스)을 제조하였다. 10 mL의 LAB 배양물 및 7.5 mL의 10-4-희석 과제 미생물 칵테일(물 중의 104 cfu/mL의 배양물)을 브로콜리 퓨레(105의 B. 세레우스)에 첨가하였다. 배양물을 혼합하기 전에 써모믹스 뚜껑의 큰 구멍 위에 호일을 고정시켰다. 배양물을 최대 속도로 1 분 동안 퓨레에 혼합하였다. 써모믹스에 대한 열 설정을 오프(off)로 바꾸고, 써모믹스를 30℃ 인큐베이터 안에 넣고, 발효를 10:45 am에 시작하였다. 4.5의 속도로 1 분 동안 퓨레를 혼합한 후, pH 및 온도 측정을 7 시간(작업 종료 시간)까지 1 시간마다 수행하였다. pH 미터를 보정하고, 80% 에탄올을 사용하여 살균하였다. 온도 프로브도 또한 80% 에탄올로 측정 전에 살균하였다.
과제 미생물의 성장을, 발효 전(T0) 및 발효 시작 후 4 시간째(T4) 및 22 시간째(T22)에, 미가공 브로콜리의 선별 배지 MRS, DRBX 및 NA +S에서 성장에 대한 계수에 의해 평가하였다.
결과
효모 및 곰팡이는 4 시간째까지 유의하게 감소되었고, 발효 종료 시(T22)에 검출되지 않았다. E. 콜라이 및 살모넬라는 발효 종료 시(T22)에 전혀 검출되지 않았다. 리스테리아는, 103 cfu/mL를 갓 넘는 출발 접종원으로, 발효 종료 시에 소수 검출되었다. B. 세레우스 포자는 일반적으로 발효에 의해 영향을 받지 않았지만, 발아하지 않았다. 과제 연구의 결과는 본원에서 브로콜리로부터 단리된 젖산균 균주가 살모넬라 및 E. 콜라이를 완전히 불활성화시키고, 대부분의 내산성 리스테리아 균주의 성장을 억제할 수 있다는 것을 지시한다. 이들은 또한 B. 세루스 포자의 포자형성을 억제할 수 있다.
[표 9]
E. 콜라이 로의 미생물 과제 연구의 예. 발효 스타터(B1, B2, B3, B4, B5, BF1, BF2의 조합)가 E. 콜라이의 성장을 억제하는지를 평가하기 위해 침연 브로콜리(3:2 브로콜리-물 비) 발효물에 E. 콜라이(5 개의 E. 콜라이 균주 EC1605, EC1606, EC1607, EC1608의 혼합)를 접종하였다(2.2 × 102 CFU/gm). 실험을 3 회 반복하였다. 4.0 미만의 pH로 30℃에서 22 시간 동안 발효를 실시하였다.
[표 10]
살모넬라 로의 미생물 과제 연구의 예. 발효 스타터(B1, B2, B3, B4, B5, BF1, BF2의 조합)가 살모넬라의 성장을 억제하는지를 평가하기 위해 침연 브로콜리(3:2 브로콜리-물 비) 발효물에 살모넬라(5 개의 균주 S. 인판티스 1023, S. 신가포레 1234, S. 타이피무리움 1657 (PT135), S. 타이피무리움 1013 (PT9), S. 비르효 1623의 혼합)를 접종하였다(1.1 × 103). 실험을 3 회 반복하였다. 4.0 미만의 pH로 30℃에서 22 시간 동안 발효를 실시하였다.
[표 11]
리스테리아 모노사이토게네스 로의 미생물 과제 연구의 예. 발효 스타터(B1, B2, B3, B4, B5, BF1, BF2의 조합)가 내산성 리스테리아의 성장을 억제하는지를 평가하기 위해 침연 브로콜리(3:2 브로콜리-물 비) 발효물에 리스테리아 모노사이토게네스(5 개의 균주 Lm2987 (7497), Lm2965 (7475), Lm2939 (7449), Lm2994 (7537), Lm2919 (7514)의 혼합)를 접종하였다(1.9 × 103). 실험을 3 회 반복하였고, 발효 종료 시 최종 리스테리아 수는 <10(비검출) 내지 1.1 × 102 CFU/gm의 범위였다. 4.0 미만의 pH로 30℃에서 22 시간 동안 발효를 실시하였다.
[표 12]
바실러스 세레우스 의 미생물 과제 연구의 예. 발효 스타터(B1, B2, B3, B4, B5, BF1, BF2의 조합)가 내산성 리스테리아의 성장을 억제하는지를 평가하기 위해 침연 브로콜리(3:2 브로콜리-물 비) 발효물에 바실러스 세레우스(5 개의 균주 B3078, B2603, B2601, B7571, B7626의 혼합)를 접종하였다(1.9 × 103). 실험을 3 회 반복하였다. 4.0 미만의 pH로 30℃에서 22 시간 동안 발효를 실시하였다.
실시예 15 -
류코노스톡 메센테로이데스
단리물의 펄스 파일링 겔 전기영동
채소류로부터의 류코노스톡 메센테로이데스를 변형으로 문헌[Chat and Dalmasso (2015)]에 기재된 바와 같이 펄스 파일링 겔 전기영동에 의해 SmaI 및 NotI 제한 효소 분해로 평가하였다.
방법:
1 일
평가된 단리물을 10 mL의 MRS 브로스에 접종하고, 인큐베이터에서 30℃로 밤새 인큐베이션하였다(16 시간).
2 일
단리물을 10 분 동안 3500 g으로 원심분리하고, 상청액을 버렸다. 펠릿을 혼합하고, 5 mL의 탈이온수로 세척하고, 10 분 동안 3500 g으로 원심분리하고, 상청액을 버렸다. 펠릿을 5 mL의 TES(1 mM EDTA, 10 mM 트리스-HCl, 0.5 M 사카로스)와 혼합하고, 볼텍싱하였다. 그 다음, 샘플을 15 분 동안 3500 g으로 원심분리하고, 상청액을 버렸다. 700 μL의 용해액(10 mg/mL의 리소자임과 TE 완충액(1 mM EDTA, 10 mM 트리스-HCl, pH 8.0, 정상적으로 살균))을 펠릿에 첨가하고, 혼합하고, 56 ℃에서 2 시간 동안 인큐베이션하여 세균을 용해시켰다. 그 다음, 700 μL의 아가로스(50 μL의 EDTA/100 mL와 1% SeaChem Gold 아가로스)를 세포 혼합물에 첨가하고, 혼합하고, 플러그 모울드에 분배하고, 튜브에 넣어진 하나의 샘플에 대한 모든 플러그에 2 mL의 제단백 용액(660 μL의 프로테이나제 K 완충액, 11 μL의 프로테이나제 K)을 첨가하고, 55℃에서 밤새 인큐베이션하였다.
3 일
그 다음, 플러그를 100 mL의 멸균 탈이온수에서 55℃까지 가열하고, 제단백 용액을 제거하고, 플러그를 15 mL 원심분리 튜브로 옮기고, 4 mL의 멸균 탈이온수로 세척하고, 실온에서 10 분 동안 55℃까지 가열하고, 이어서, 실온에서 10 분 동안 4 mL의 TE 완충액으로 4 회 세척하였다.
제한 분해
플러그에서 2mm 슬라이스를 100 μL의 1X 제한 완충액과 함께 에펜도르프 튜브에 넣고, 실온에서 20 분 동안 인큐베이션하고, 제한 완충액을 제거하고, 제한 완충액에서 40 μL 내지 100 μL의 SmaI(20 U) 또는 NotI로 교체하고, 최적 온도(25℃)에서 4 시간 동안 인큐베이션하였다.
4 일
제한 단편의 분리
각 튜브에 1 mL의 0.5X TBE 완충액을 첨가하고, 적어도 15 분 동안 정치시켜 반응을 중지시키고, 박테리오파지 λ DNA 사다리(New England Biolab)를 TBE 완충액에서 인큐베이션하였다. 완충액을 제거하고, 슬라이스를 모두 5 개의 레인에서 사다리와 함께 콤(comb) 위에 로딩시켰다. 1.0%의 초순수 DNA 등급 아가로스(펄스장 인증 아가로스)를 0.5X TBE 러닝 완충액에서 제조하였다.
전기영동 조건
완충액을 14℃에서 유지하였다(모델 1000 미니-칠러, BioRad). BioRad "Chef Mapper™"의 2 상태 프로그램(자동 알고리즘 아님)을 선택하였다. 펄스 시간을 2 초에서 25 초까지 선형으로 상승시켰다("a"가 나타나면 엔터(enter)를 누름). 구배 6 V/cm(전압), 끼인각 120°, 24 시간의 이동 시간.
5 일
GelRed에서 약 30 분 겔 염색, 변색, 가시화
결과
BF1에 대한 제한 핑거프린트는 구획되었지만, 당근으로부터 단리된 류코노스톡 메센테로이데스와 유사했다(도 13). BF2에 대한 제한 핑거프린트는 평가된 모든 류코노스톡 메센테로이데스 균주로부터 구획되었다(도 13).
실시예 16 -
류코노스톡 메센테로이데스
및
락토바실러스 플란타룸
단리물의 변이 분석
락토바실러스 플란타룸 단리물(B1 내지 B5)의 SNP 분석을 위해, B1 Prokka gbk를 Snippy SNP 분석(표준 방법)에 대한 참조로서 사용하였다. 각 균주에 대한 판독 데이터를 사용하여 단일 비교를 수행하였다. B1 판독을 대조로서 작용시켰다.
예시적인 명령은 하기와 같았다:
snippy --cpus 24 --outdir B5 --ref B1_S1mod.gbk --pe1
B5_S17_L001_R1_001.fastq.gz --pe2 B5_S17_L001_R2_001.fastq.gz
계산된 개별 비교 및 참조로서 B1 gbk를 사용한 코어
snippy-core --prefix core B1 B2 B3 B4 B5
락토바실러스 플란타룸 ATCC 8014(SRR1552613)에 대한 SRA로부터 다운로드한 참조 균주 판독 데이터와 B1 간의 비교를 또한 수행하였다. sratoolkit.2.9.2-win64를 이용하여 prefetch 및 fastq로의 변환으로 표준 방법을 이용하여 다운로드를 수행하였다. 참조로서 류코노스톡 메센테로이데스 ATCC 8293과 류코노스톡 메센테로이데스 단리물 BF1 및 BF2의 비교를 위해 유사한 접근을 이용하였다.
결과
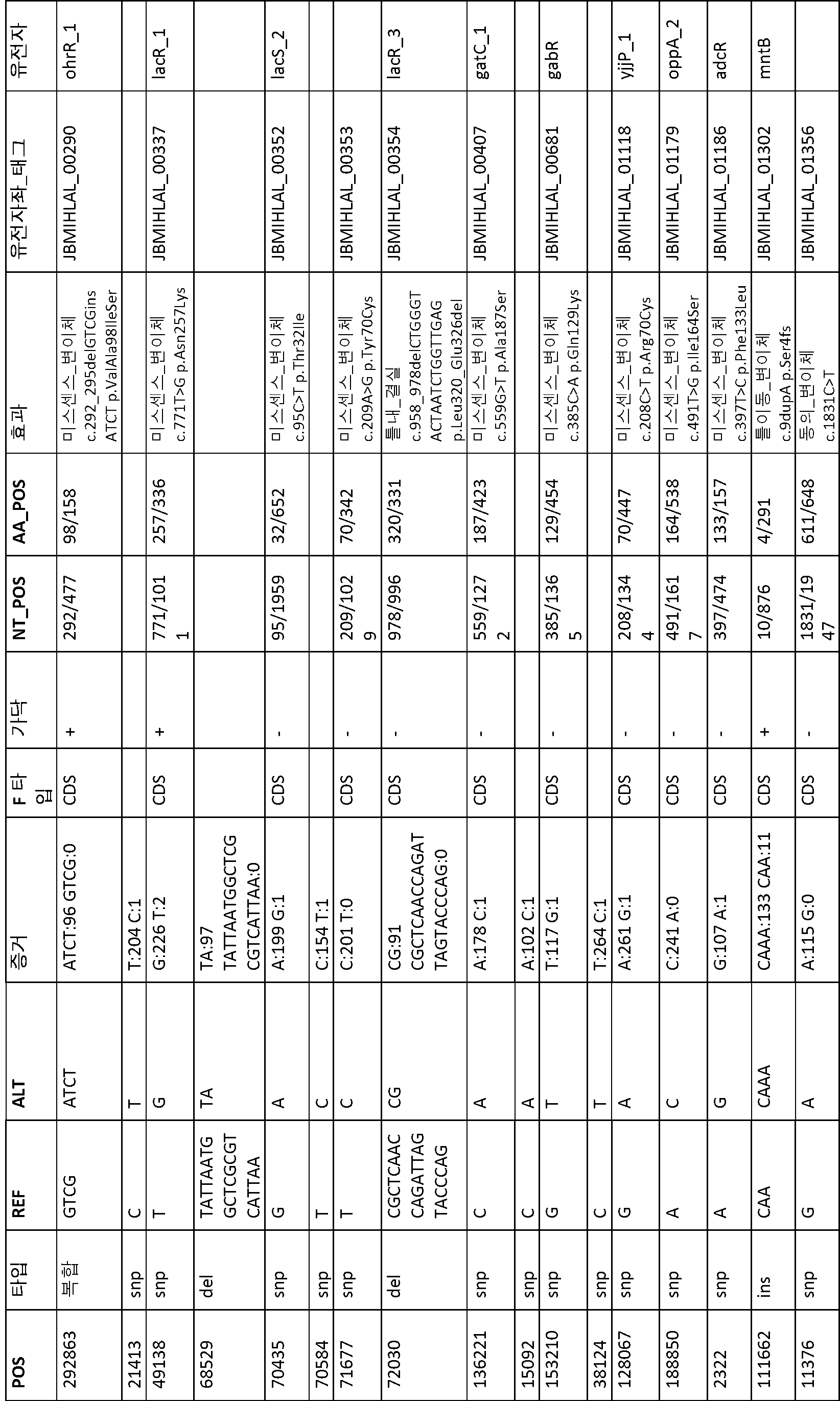
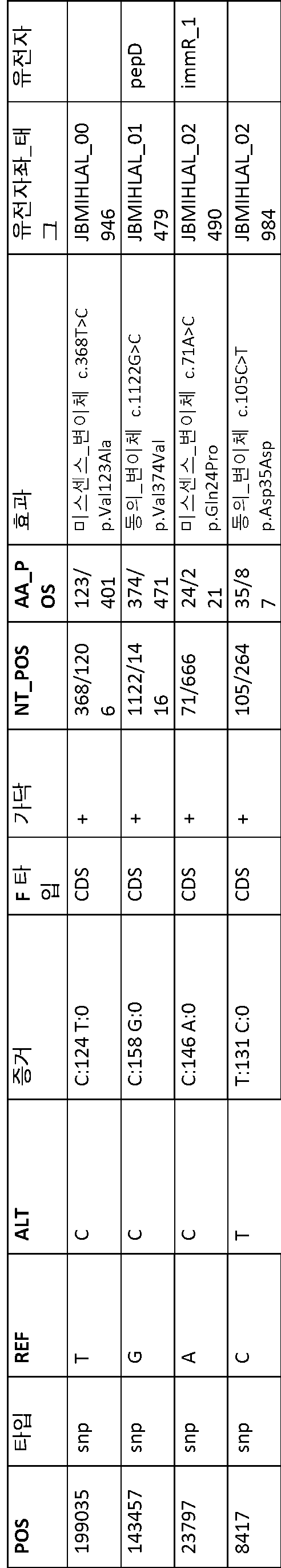
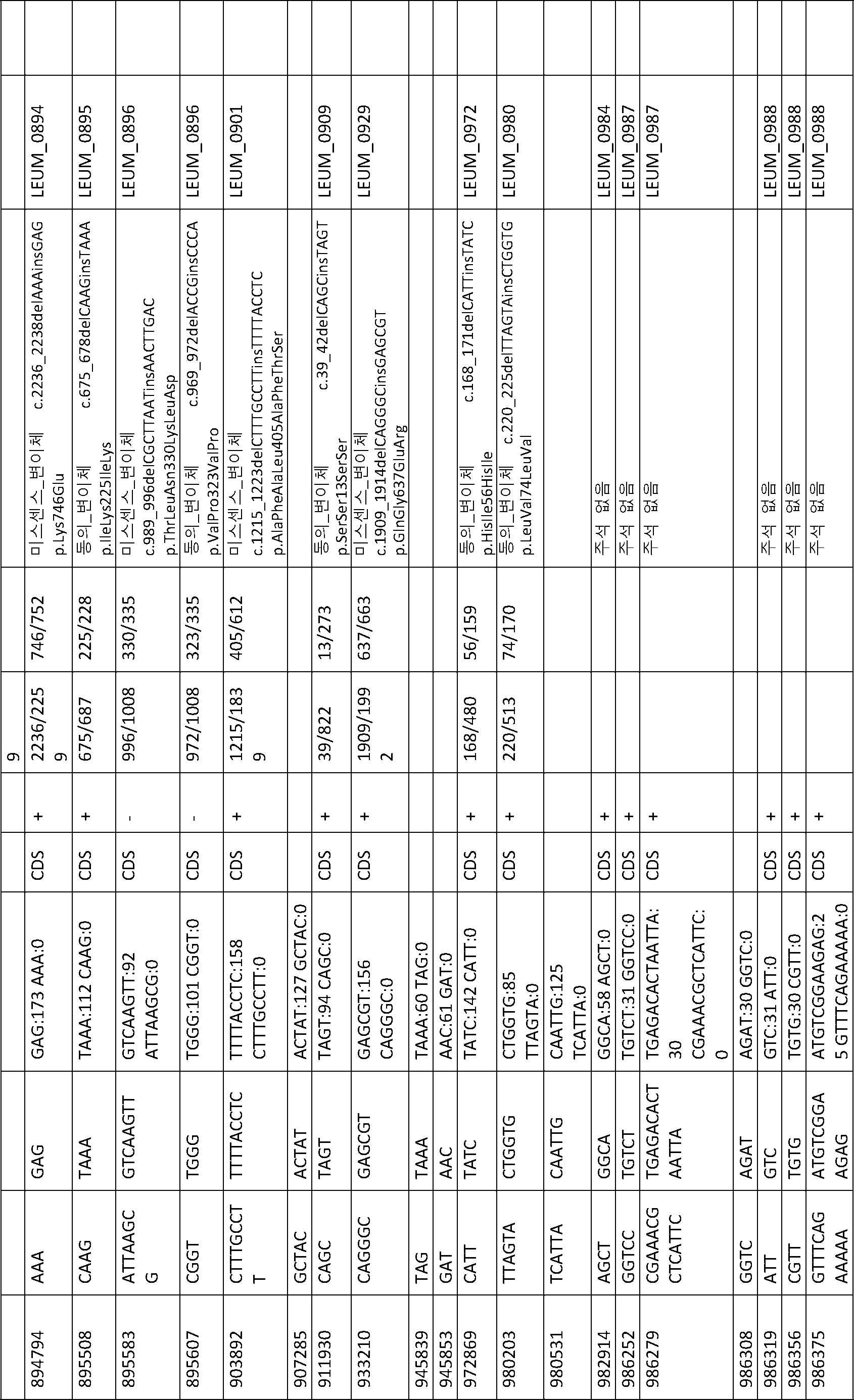
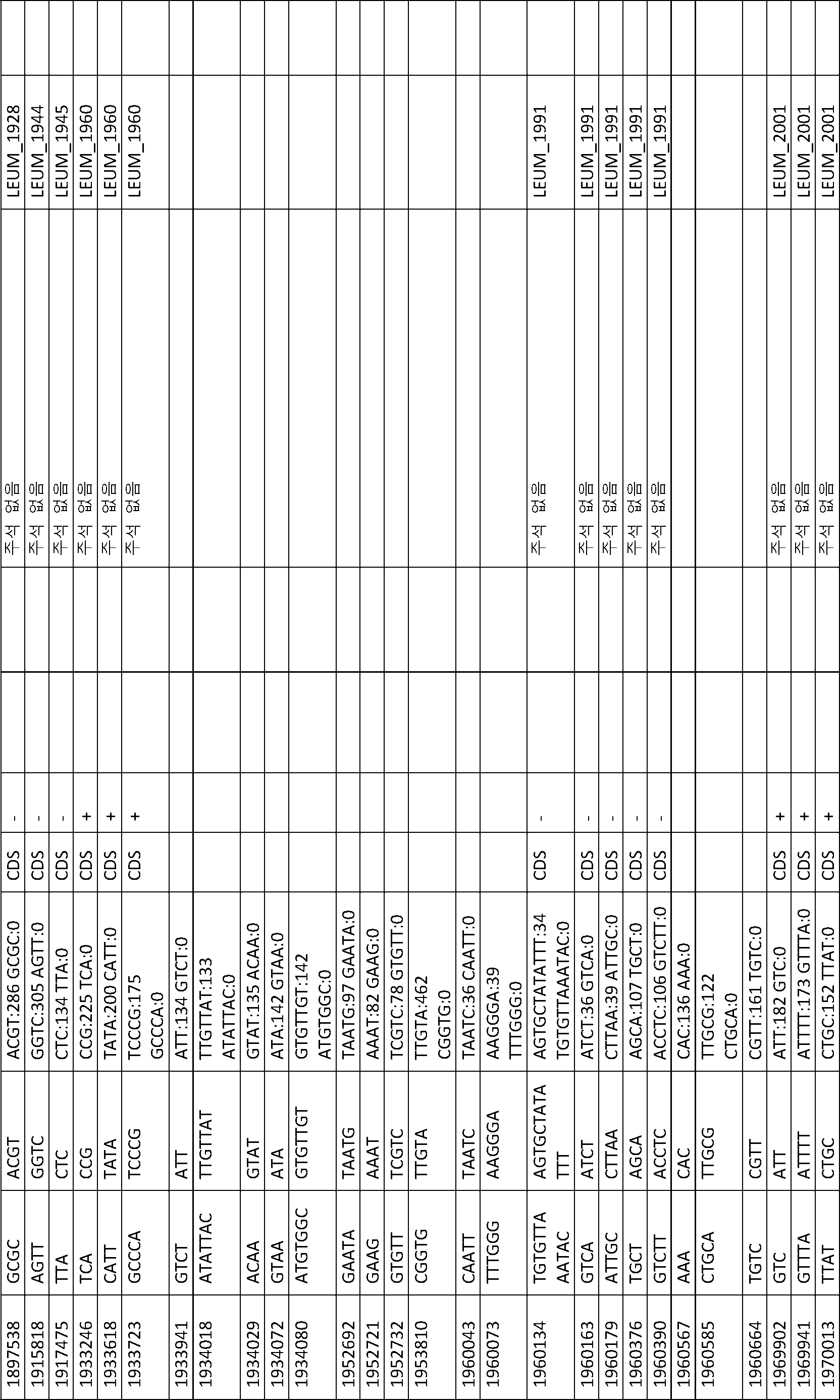
B1과 ATCC 8014 간에 변이(41 개)가 관찰되었다(표 13). B1과 다른 B 단리물 B2, B3, B4 및 B5 간에 변이(1 개 내지 4 개)가 관찰되었다(표 14 내지 표 17). BF1 및 BF2는 서로 매우 상이했다. BF1과 ATCC 8293 간에 변이(19 개)가 관찰되었다(표 18). BF2와 ATCC 8293 간에 변이(약 7000 개)가 관찰되었다. 표 19에 요약된 BF2와 ATCC8293 간에 459 개의 복합 변이가 확인되었다.
[표 13]
ATCC8014와 비교한 B1 변이 분석에 의해 확인된 다형성.
[표 14]
B1과 비교한 B2 변이 분석에 의해 확인된 다형성.
[표 15]
B1과 비교한 B3 변이 분석에 의해 확인된 다형성.
[표 16]
B1과 비교한 B4 변이 분석에 의해 확인된 다형성.
[표 17]
B1과 비교한 B5 변이 분석에 의해 확인된 다형성.
[표 18]
ATCC8293과 비교한 BF1 변이 분석에 의해 확인된 다형성.
[표 19]
ATCC8293과 비교한 BF2 변이 분석에 의해 확인된 다형성.
광범위하게 기재된 바와 같은 본 발명의 사상 또는 범위로부터 벗어남 없이 특정 구현예에 나타나 있는 바와 같은 본 발명에 대해 다수 변화 및/또는 변형이 이루어질 수 있다는 것이 당업자에 의해 인지될 것이다. 따라서, 본 발명의 구현예는 모든 점에서 한정적인 것이 아니라 예시적인 것으로 여겨져야 한다.
본 출원은 "Isothiocyanate containing Brassicaceae products and method of preparation thereof"라는 명칭으로 2017년 9월 28일에 출원된 호주 가출원 번호 제2017903944호의 우선권을 주장하고, 상기 출원의 전체 내용은 본원에 참조로 포함된다.
본원에서 논의되고/논의되거나 참조된 모든 간행물은 그 전체가 본원에 포함된다.
본 명세서에 포함된 문헌, 행위, 재료, 장치, 또는 물품 등의 임의의 논의는 오로지 본 발명에 대한 맥락을 제공하려는 목적을 위한 것이다. 임의의 또는 모든 이러한 물질들은 종래 기술 기반의 일부를 형성하거나 본 출원의 각 청구항의 우선일 전에 존재하는 본 발명과 관련된 분야에서 흔한 일반 지식이었다는 허용으로 받아들여서는 안된다.
참조문헌